oulu 2010 d 1071 university of oulu p.o.b. 7500 fi...
TRANSCRIPT

ABCDEFG
UNIVERS ITY OF OULU P.O.B . 7500 F I -90014 UNIVERS ITY OF OULU F INLAND
A C T A U N I V E R S I T A T I S O U L U E N S I S
S E R I E S E D I T O R S
SCIENTIAE RERUM NATURALIUM
HUMANIORA
TECHNICA
MEDICA
SCIENTIAE RERUM SOCIALIUM
SCRIPTA ACADEMICA
OECONOMICA
EDITOR IN CHIEF
PUBLICATIONS EDITOR
Professor Mikko Siponen
University Lecturer Elise Kärkkäinen
Professor Hannu Heusala
Professor Olli Vuolteenaho
Senior Researcher Eila Estola
Information officer Tiina Pistokoski
University Lecturer Seppo Eriksson
Professor Olli Vuolteenaho
Publications Editor Kirsti Nurkkala
ISBN 978-951-42-6298-2 (Paperback)ISBN 978-951-42-6299-9 (PDF)ISSN 0355-3221 (Print)ISSN 1796-2234 (Online)
U N I V E R S I TAT I S O U L U E N S I S
MEDICA
ACTAD
D 1071
ACTA
Khaled M
. Abass
OULU 2010
D 1071
Khaled M. Abass
METABOLISM AND INTERACTIONS OF PESTICIDES IN HUMANAND ANIMAL IN VITRO HEPATIC MODELS
FACULTY OF MEDICINE,INSTITUTE OF BIOMEDICINE,DEPARTMENT OF PHARMACOLOGY AND TOXICOLOGYFACULTY OF SCIENCE,DEPARTMENT OF CHEMISTRY,UNIVERSITY OF OULU


A C T A U N I V E R S I T A T I S O U L U E N S I SD M e d i c a 1 0 7 1
KHALED M. ABASS
METABOLISM AND INTERACTIONS OF PESTICIDES IN HUMAN AND ANIMAL IN VITRO HEPATIC MODELS
Academic dissertation to be presented with the assent ofthe Faculty of Medicine of the University of Oulu forpublic defence in the Auditorium of the Department ofPharmacology and Toxicology (Aapistie 5 B), on 26November 2010, at 12 noon
UNIVERSITY OF OULU, OULU 2010

Copyright © 2010Acta Univ. Oul. D 1071, 2010
Supervised byProfessor Olavi PelkonenDocent Miia Turpeinen
Reviewed byProfessor Alan R. BoobisDoctor Emanuela Testai
ISBN 978-951-42-6298-2 (Paperback)ISBN 978-951-42-6299-9 (PDF)http://herkules.oulu.fi/isbn9789514262999/ISSN 0355-3221 (Printed)ISSN 1796-2234 (Online)http://herkules.oulu.fi/issn03553221/
Cover designRaimo Ahonen
JUVENES PRINTTAMPERE 2010

Abass, Khaled M., Metabolism and interactions of pesticides in human and animalin vitro hepatic models. Faculty of Medicine, Institute of Biomedicine, Department of Pharmacology and Toxicology,University of Oulu, P.O.Box 5000, FI-90014 University of Oulu, Finland; Faculty of Science,Department of Chemistry, P.O.Box 3000, FI-90014 University of Oulu, FinlandActa Univ. Oul. D 1071, 2010Oulu, Finland
Abstract
Risk assessment of chemicals needs reliable scientific information and one source of informationis the characterization of the metabolic fate and toxicokinetics of a chemical. Metabolism is oftenthe most important factor contributing to toxicokinetics. Cytochrome P450 (CYP) enzymes are asuperfamily of microsomal proteins playing a pivotal role in xenobiotic metabolism.
In the present study, pesticides were used as representative xenobiotics since exposure topesticides is a global challenge to risk assessment. Human and animal in vitro hepatic models wereapplied with the advantage of novel analytical techniques (LC/TOF-MS and LC/MS-MS) toelucidate the in vitro metabolism and interaction of selected pesticides.
The results of these studies demonstrate that CYP enzymes catalyze the bioactivation ofprofenofos, diuron and carbosulfan into their more toxic metabolites desthiopropylprofenofos, N-demethyldiuron and carbofuran, respectively. The suspected carcinogenic metabolite ofmetalaxyl, 2,6-dimethylaniline, was not detected. CYP3A4 and CYP2C19 activities may beimportant in determining the toxicity arising from exposure to profenofos and carbosulfan.Individuals with high CYP1A2 and CYP2C19 activities might be more susceptible to diurontoxicity.
Qualitative results of in vitro metabolism were generally in agreement with the results obtainedfrom the published in vivo data, at least for the active chemical moiety and major metabolites.Considerable differences in the quantities of the metabolites produced within the species, as wellas in the ratios of the metabolites among the species, were observed.
These findings illustrate that in vitro screening of qualitative and quantitative differences areneeded to provide a firm basis for interspecies and in vitro-in vivo extrapolations. Based on ourfindings, in vitro-in vivo extrapolation based on the elucidation of the in vitro metabolic pattern ofpesticides in human and animal hepatic models could be a good model for understanding andextending the results of pesticides metabolism studies to human health risk assessment.
Keywords: cytochrome P450, in vitro-in vivo extrapolation, interindividual variability,interspecies differences, pesticide metabolism


5
Acknowledgements
This work was carried out at the Department of Pharmacology and Toxicology,
Institute of Biomedicine, Faculty of Medicine, and at the Department of
Chemistry, Faculty of Natural Sciences, University of Oulu, Finland, during the
years 2005–2010. First of all, I owe my deepest respect and sincerest gratitude to
my supervisor, Professor Olavi Pelkonen, for his professional, patient and friendly
guidance during this work. I appreciate his continuous optimism and
encouragement. I am deeply grateful to him for offering me an opportunity to be
involved in his projects, for guiding me during the lab work as well as the writing
of scientific papers, and for his invaluable support during these years. Docent
Miia Turpeinen deserves my deep gratitude for her great help and tutoring, and
particularly for her encouraging support in the beginning of my work.
I am grateful to the official reviewers of this thesis, Professor Alan R. Boobis
from Imperial College London and Dr. Emanuela Testai from the Italian National
Institute for Health, for their careful review and expert comments. I thank Sandra
Hänninen for revising the English language of this thesis.
I would like to express my gratitude to my co-authors, Professor Jorma
Jalonen, Docent Sampo Mattila and scientist Petri Reponen, for their fruitful
collaboration and expert knowledge. I am also grateful to Ritva Tauriainen, Päivi
Tyni, and Päivi Joensuu; their excellent technical assistance has been of great help
in this work.
My sincere thanks go to all the people in the Department of Pharmacology
and Toxicology who have helped me in my work. I am grateful to the whole
ClubMed team: Professor Jukka Hakkola, Docent Päivi Myllynen and all the
members are acknowledged.
Especially, my warm thanks belong to Petri Reponen for our close
cooperation and friendship during those numerous hours conducting the
laboratory measurements. I am also thankful to Dr. Pirkko Viitala, Professor Kirsi
Vähäkangas, Professor Arja Rautio, Professor Hannu Raunio, Professor Markku
Pasanen and Dr. Sohvi Hörkkö. My special thanks go to Raija Hanni for her
expert secretarial work and to Esa Kerttula and Marja Räinä for their invaluable
technical service. I wish to extend my sincere appreciation to all my colleagues,
Virpi Lämsä, Larissa Tursas, Sanna-Mari Järvenpää, Marcin Buler, Satu
Arpiainen, Reka Skoumal, and David Vicente for their friendship and advice. I
am grateful to all members of the Pesticides Department, Menoufia University for
their support.

6
My warmest thanks go to my loving parents, my brother and my sisters, who
have always believed in me and supported me in my decisions. Finally, I want to
thank my dear wife Dalia for her never-failing support, encouragement,
understanding and reminding me what is important in life. I am also indebted to
my daughter Nada and my son Amr for being part of my life. I also warmly thank
all other people closely related in my life, especially my parents-in-law, who have
been like second parents to me.
This work was funded by a Ministry of Education-supported position from
the Finnish Graduate School in Toxicology (ToxGS) and supported by grants
from the Center for International Mobility (CIMO), the Agricultural Foundation
of the Economic Association in the Oulu province, Oulu University Scholarship
Foundation and Orion-Pharma Research Foundation.
Oulu, November 2010 Khaled Megahed Abass

7
Abbreviations
AChE acetylcholinesterase
ADI acceptable daily intake
ADME absorption, distribution, metabolism, and excretion
ADUF uncertainty factor for animal to human differences in
toxicodynamics
AKAF adjustment factors for interspecies differences in toxicokinetics
AKUF uncertainty factor for animal to human differences in
toxicokinetics
AOEL acceptable operator exposure level
ARfD acute reference dose
CLH hepatic clearance
CLint, LM in vitro intrinsic clearance
CLint, whole liver hepatic intrinsic clearance
CSAFs chemical-specific adjustment factor
CYP cytochrome P450
CUF composite uncertainty factor
DMSO dimethyl sulphoxide
DogLM dog liver microsomes
EPA Environmental Protection Agency
ESI electrospray ionization
GEMS food global environment monitoring system
GSH L-glutathione (reduced form)
HDUF uncertainty factor for human variability in toxicodynamics
HKAF adjustment factors for human variability in toxicokinetics
HKUF uncertainty factor for human variability in toxicokinetics
HLM human liver microsomes
HPLC high-performance liquid chromatography
IARC International Agency for Research on Cancer
IC50 concentration of inhibitor corresponding to a 50% decrease in
reaction
IEDIs international estimated dietary intakes
IPCS The International Program on Chemical Safety Km Michaelis-Menten constant for a substrate
LC liquid chromatography
MonLM monkey liver microsomes

8
MouseLM mouse liver microsomes
MPPGL microsomal protein per gram of liver weight
MRL maximum residue level
MRM multiple reaction monitoring
MS mass spectrometry
NADPH Nicotinamide adenine dinucleotide phosphate (reduced form)
NOAEL no observed adverse effect level
OP organophosphate PAPS 3′-phosphoadenosine 5′-phosphosulfate
PigLM minipig liver microsomes
PM poor metaboliser
QH liver blood flow
RabLM rabbit liver microsomes
RatLM rat liver microsomes
TLC thin layer chromatography
TOF time-of-flight
UDPGA uridine 5′-diphosphoglucuronic acid
UF uncertainty factor
UM ultra-rapid metabolizer
UNEP United Nations Environment Program
US EPA Uunited States Environmental Protection Agency
Vmax the maximal velocity of the reaction
WHO World Health Organization

9
List of original publications
This thesis is based on the following articles, which are referred to in the text by
Roman numerals I to V:
I Abass K, Reponen P, Jalonen J & Pelkonen O (2007) In vitro metabolism and interactions of the fungicide metalaxyl in human liver preparations. Environ Toxicol Pharmacol 23(1): 39–47.
II Abass K, Reponen P, Jalonen J & Pelkonen O (2007) In vitro metabolism and interaction of profenofos by human, mouse and rat liver preparations. Pestic Biochem Physiol. 87(3): 238–247.
III Abass K, Reponen P, Turpeinen M, Jalonen J & Pelkonen O (2007) Characterization of diuron N-demethylation by mammalian hepatic microsomes and cDNA-expressed human cytochrome P450 enzymes. Drug Metab Dispos. 35(9): 1634–1641.
IV Abass K, Reponen P, Mattila S & Pelkonen O (2009) Metabolism of carbosulfan. I. Species differences in the in vitro biotransformation by mammalian hepatic microsomes including human. Chem Biol Interact. 181(2): 210–219.
V Abass K, Reponen P, Mattila S. & Pelkonen O (2010) Metabolism of carbosulfan. II. Human interindividual variability in its in vitro hepatic biotransformation and the identification of the cytochrome P450 isoforms involved. Chem Biol Interact. 185(3): 163–173.

10

11
Contents
Abstract Acknowledgements 5 Abbreviations 7 List of original publications 9 Contents 11 1 Introduction 15 2 Review of the literature 17
2.1 Xenobiotic biotransformation ................................................................. 17 2.2 CYPs – the human xenobiotic- metabolizing enzymes ........................... 17
2.2.1 CYP1A subfamily ......................................................................... 20 2.2.2 CYP2A subfamily ......................................................................... 21 2.2.3 CYP2B subfamily ......................................................................... 22 2.2.4 CYP2C subfamily ......................................................................... 23 2.2.5 CYP2D subfamily ........................................................................ 25 2.2.6 CYP2E subfamily ......................................................................... 25 2.2.7 CYP2F subfamily ......................................................................... 26 2.2.8 CYP2S subfamily ......................................................................... 27 2.2.9 CYP3A subfamily ......................................................................... 27
2.3 In vitro and human-derived techniques for testing xenobiotic
metabolism .............................................................................................. 29 2.3.1 Subcellular fractions (liver microsomes and homogenates) ......... 31 2.3.2 cDNA-expressed CYPs (single-enzyme systems) ........................ 32 2.3.3 Hepatocytes .................................................................................. 32 2.3.4 Liver slices ................................................................................... 33 2.3.5 Immortalized cell lines ................................................................. 33
2.4 In vitro characterization of the metabolism and metabolic
interactions of xenobiotics ...................................................................... 34 2.4.1 Metabolic stability of the compound ............................................ 34 2.4.2 Identification of metabolites and metabolic routes ....................... 35 2.4.3 Identification of the metabolizing enzymes .................................. 36
cDNA-expressed enzymes 36 Correlation studies 37 Inhibition studies with CYP-selective chemical inhibitors and specific
antibodies 37 Inhibition of CYP enzymes 38

12
2.5 Health risk assessment of chemicals ....................................................... 38 2.5.1 General considerations ................................................................. 38 2.5.2 Interspecies differences and interindividual variability in
toxicokinetics ................................................................................ 41 2.5.3 Considerations regarding in vitro data .......................................... 42
2.6 Pesticides as representative xenobiotics .................................................. 42 2.6.1 General information ...................................................................... 42 2.6.2 Individual pesticides studied ........................................................ 44
Metalaxyl 44 Profenofos 45 Diuron 46 Carbosulfan 47
3 Aims of the present study 49 4 Materials and Methods 51
4.1 Chemicals ................................................................................................ 51 4.2 Human and animal liver samples ............................................................ 51 4.3 Recombinant enzymes ............................................................................ 52 4.4 In vitro assay of metabolites and analytical methods .............................. 52 4.5 Determination of enzyme kinetic parameters .......................................... 53 4.6 Correlation analysis ................................................................................. 53 4.7 Enzymatic and inhibition assays ............................................................. 53 4.8 In vitro - in vivo extrapolations ............................................................... 53
5 Results 55 5.1 Metabolism of pesticides in mammalian hepatic preparations ................ 55
5.1.1 Identification of metabolites produced in vitro by
mammalian hepatic preparations .................................................. 55 5.1.2 Studies of in vitro metabolism by mammalian hepatic
preparations .................................................................................. 56 5.1.3 Interspecies differences in in vitro metabolism ............................ 57 5.1.4 Interindividual variability in in vitro metabolism ......................... 57
5.2 Identification of CYP isoenzymes metabolizing the studied
pesticides in human liver in vitro ............................................................ 59 5.2.1 Kinetic analysis of recombinant CYP enzymes ............................ 59 5.2.2 Analysis of the relative contributions of different CYP
enzymes ........................................................................................ 59 5.2.3 Relationship between CYP activities and metabolite
formation ...................................................................................... 60

13
5.2.4 Inhibition of in vitro metabolism of pesticides by CYP-
selective inhibitors (V) ................................................................. 62 5.3 Evaluation of the CYP inhibition potential of the studied
pesticides in pooled human hepatic microsomes .................................... 62 5.4 In vitro - in vivo extrapolation ................................................................. 64
6 Discussion 67 6.1 Methodological aspects of the in vitro studies ........................................ 67 6.2 Differences in pesticide metabolites formation ....................................... 68
6.2.1 In vitro vs in vivo qualitative differences ...................................... 68 6.2.2 Qualitative interspecies differences in the in vitro
metabolism of profenofos, diuron and carbosulfan ...................... 70 6.2.3 Quantitative interspecies differences in the in vitro
metabolism of profenofos, diuron and carbosulfan ...................... 70 6.2.4 Interindividual variability in in vitro metabolism ......................... 72
6.3 Identification of the P450 isoform(s) involved in metabolism ................ 74 6.4 CYP-pesticides inhibition potential ........................................................ 75 6.5 Quantitative prediction of in vivo metabolic clearance and the
calculation of CSAFs for the studied compounds. .................................. 76 6.5.1 Interspecies differences and the calculation of AKAF ................... 76 6.5.2 Interindividual variability and the calculation of the HKAF
factor for carbosulfan ................................................................... 78 6.5.3 Calculation of the composite uncertainty factor (CUF),
carbosulfan as an example ............................................................ 78 7 Conclusions 81 References 83 Original Publications 113

14

15
1 Introduction
Protecting public health is the primary reason why risk assessment of xenobiotics,
be they pharmaceuticals, agrochemicals, or industrial chemicals, is of utmost
importance. Exposure to pesticides is a global challenge to risk assessment
(Alavanja et al. 2004, Maroni et al. 2006). Chronic exposure to low levels of
pesticides can cause mutations and/ or carcinogenicity (Bull et al. 2006, IARC
1991, Karabay & Gunnehir 2005). On a world-wide basis, acute pesticide
poisoning is an important cause of morbidity and mortality. In an extrapolation,
WHO/UNEP estimated that more than 3 million people were hospitalized for
pesticide poisoning every year and that 220 000 died; it particularly noted that
two-thirds of hospitalizations and the majority of deaths were attributable to
intentional self-poisoning rather than to occupational or accidental poisoning
(Konradsen et al. 2005, WHO/UNEP 1990). Humans are inevitably exposed to
pesticides in a variety of ways: at different dose levels and for varying periods of
time (Boobis et al. 2008, Ellenhorns et al. 1997). This is why pesticides are used
as representative xenobiotics in this research program.
Risk assessment needs reliable scientific information and one source of
information is the characterization of the metabolic fate and toxicokinetics of
compounds. Toxicokinetics refers to the movement of a xenobiotic into, through,
and out of the body and is divided into several processes including absorption,
distribution, metabolism, and excretion, ADME. Metabolism is one of the most
important factors that can affect the overall toxic profile of a pesticide. During
metabolism, the chemical is first biotransformed by phase I enzymes, usually by
the cytochrome P450 (CYP) enzyme system, and then conjugated to a more
soluble and excretable form by phase II conjugating enzyme systems (Guengerich
& Shimada 1991). In general, these enzymatic reactions are beneficial in that they
help eliminate foreign compounds. Sometimes, however, these enzymes
transform an otherwise harmless substance into a reactive form – a phenomenon
known as metabolic activation (Guengerich 2001).
The cytochromes P450 (CYP) comprise a large multigene family of
hemethiolate proteins which are of considerable importance in the metabolism of
xenobiotics and endobiotics. Hepatic clearance is a principal way of elimination
for xenobiotics, including drugs. CYP enzymes in humans as well as in other
species have been intensively studied during recent years (Pelkonen et al. 2008,
Pelkonen & Turpeinen 2007). It is now possible to characterize metabolic
reactions and routes, metabolic interactions, and to assign which CYP is involved

16
in the metabolism of a certain xenobiotic by different in vitro approaches
(Hodgson & Rose 2007a, Pelkonen & Raunio 2005, Pelkonen et al. 2005).
The aim of this research was to the elucidate metabolic factors and
interactions of four pesticides in human and animal in vitro hepatic models and to
overall improve the process of human risk assessment.

17
2 Review of the literature
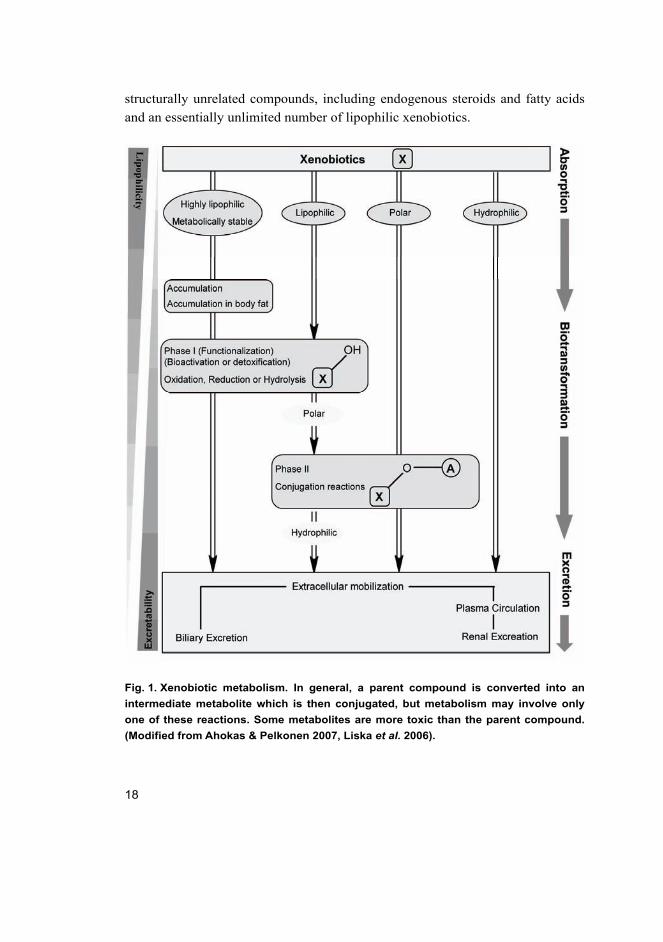
2.1 Xenobiotic biotransformation
Xenobiotic biotransformation is the process by which lipophilic foreign
compounds are metabolized through enzymatic catalysis to hydrophilic
metabolites that are eliminated directly or after conjugation with endogenous
cofactors via renal or biliary excretion. These metabolic enzymes are divided into
two groups, Phase I and Phase II enzymes (Oesch et al. 2000, Rendic & Di Carlo
1997) (Figure 1).
Phase I reactions, considered as functionalization reactions, add or uncover
functional groups on xenobiotics with increasing polarity or nucleophilicity that
create a suitable substrate for Phase II metabolism. Phase I reactions generally
involve oxidation, reduction and hydrolytic reactions as well as other rarer
miscellaneous reactions. These reactions are mediated primarily by the
cytochrome P450 family of enzymes, but other enzymes (e.g. flavin
monooxygenases, peroxidases, amine oxidases, dehydrogenases, xanthine
oxidases) also catalyze the oxidation of certain functional groups. In addition to
the oxidative reactions there are different types of hydrolytic reactions catalyzed
by enzymes like carboxylesterases and epoxide hydrolases (Hodgson & Goldstein
2001, Low 1998, Parkinson 2001).
Phase I products are not usually eliminated rapidly, but undergo a subsequent
reaction in which an endogenous substrate such as glucuronic acid, sulfuric acid,
acetic acid, or an amino acid combines with the newly established functional
group to form a highly polar conjugate to make them more easily excreted.
Sulfation, glucuronidation and glutathione conjugation are the most prevalent
classes of phase II metabolism, which may occur directly on the parent
compounds that contain appropriate structural motifs, or, as is usually the case, on
functional groups added or exposed by Phase I oxidation (LeBlanc & Dauterman
2001, Rose & Hodgson 2004, Zamek-Gliszczynski et al. 2006).
2.2 CYPs – the human xenobiotic- metabolizing enzymes
The P450 enzymes are hemoproteins with approximate molecular weights of
50,000 Da. These enzymes catalyze the monooxidation of a wide variety of
18
structurally unrelated compounds, including endogenous steroids and fatty acids
and an essentially unlimited number of lipophilic xenobiotics.
Fig. 1. Xenobiotic metabolism. In general, a parent compound is converted into an
intermediate metabolite which is then conjugated, but metabolism may involve only
one of these reactions. Some metabolites are more toxic than the parent compound.
(Modified from Ahokas & Pelkonen 2007, Liska et al. 2006).

19
CYP oxidation reactions involve a complex series of steps. The initial step
involves the binding of a substrate to oxidized CYP, followed by a one-electron
reduction catalyzed by NADPH cytochrome P450 reductase to form a reduced
cytochrome-substrate complex. The next several steps involve interaction with
molecular oxygen, the acceptance of the second electron from NADPH
cytochrome b5 reductase, followed by subsequent release of water and the
oxygenated product of the reaction. This reaction sequence results in the addition
of one oxygen atom to the substrate, while the other atom is reduced to water
(Guengerich 2001, Parkinson 2001, Rose & Hodgson 2004).
CYPs are found in high concentrations in the liver, but are present in a variety
of other tissues, including lung (Lawton et al. 1990), kidney (Hjelle et al. 1986,
Tremaine et al. 1985), the gastrointestinal tract (Dutcher & Boyd 1979, Peters &
Kremers 1989), nasal mucosa (Adams et al. 1991, Eriksson & Brittebo 1991),
skin (Khan et al. 1989) and brain (Bergh & Strobel 1992, Dhawan et al. 1990).
P450 enzymes are categorized into families and subfamilies by their sequence
similarities. Humans have 18 families of cytochrome P450 genes and 44
subfamilies. The enzymes are thus identified by a number denoting the family, a
letter denoting the subfamily and a number identifying the specific member of the
subfamily. The example given below explains the system of nomenclature
followed (Figure 2)
Fig. 2. An example of the nomenclature of the cytochrome P450 enzymes (modified
from Wijnen et al. 2007).
The website http://drnelson.utmem.edu/CytochromeP450.html contains more
detailed classification related to the cytochrome P450 metabolizing enzymes. The
CYP enzymes in families 1-3 (Figure 3) are active in the metabolism of a wide
variety of xenobiotics including drugs (Pelkonen et al. 2008, Pelkonen & Raunio
2005, Rendic & Di Carlo 1997) as well as pesticides (Figure 4).

20
Fig. 3. Relative abundance of individual CYP forms in the liver (modified from
Pelkonen et al. 2008).
Fig. 4. Involvement of human recombinant P450 isoforms in pesticides metabolism (n
= 451 reactions based on literature survey, see the literature review).
2.2.1 CYP1A subfamily
CYP1A1 and CYP1A2 are members of the CYP1A subfamily. CYP1A1 is found
mainly in extrahepatic tissues under exposure to inducing agents and its level of
expression in human liver is very low (Ding & Kaminsky 2003, Edwards et al. 1998, Guengerich & Shimada 1991, McKinnon et al. 1991, Pasanen & Pelkonen
1994, Pelkonen et al. 1998, Raunio et al. 1995). However, CYP1A1 is a major
enzyme in the metabolism of a number of pesticides such as atrazine and
ametryne (Lang et al. 1997), carbaryl (Tang et al. 2002), diuron (Abass et al.

21
2007) and carbosulfan (Abass et al. 2010) as well as in the activation of
polycyclic aromatic hydrocarbons into DNA-binding forms (Shimada et al. 1996).
In humans, CYP1A2 is expressed mainly in liver and in lower levels in lung
along with CYP1A1 (Liu et al. 2003, Wei et al. 2001, Wei et al. 2002). CYP1A2
represents about 10% of total CYP enzymes in the human liver (Pelkonen &
Breimer 1994, Shimada et al. 1994). CYP1A2 enzyme levels in the human liver
display some variability between individuals (Shimada et al. 1994). The catalytic
activities of CYP1A2 have been reviewed by Pelkonen et al. (Pelkonen et al. 2008). CYP1A2 has a major role in the metabolism of many important chemicals
such as caffeine (Butler et al. 1989, Tassaneeyakul et al. 1992), phenacetin
(Sesardic et al. 1990, Venkatakrishnan et al. 1998b), theophylline (Sarkar &
Jackson 1994, Tjia et al. 1996), clozapine (Fang et al. 1998), melatonin (Facciolá et al. 2001, von Bahr et al. 2000), and tizanidine (Granfors et al. 2004a, Granfors et al. 2004b). O-deethylation of ethoxyresorufin, the specific in vitro substrate, is
among the CYP1A2-catalytic reactions (Burke et al. 1977). Furafylline and
fluvoxamine have been widely used as in vitro diagnostic inhibitors (Sesardic et al. 1990).
CYP1A2 is known to be responsible for the metabolism of some pesticides.
CYP1A2 mediates atrazine, terbuthylazine, ametryne and terbutryne dealkylation
(Lang et al. 1997), carbaryl methyl hydroxylation and aromatic hydroxylation
(Tang et al. 2002), Furametpyr and diuron N-demethylation (Abass et al. 2007,
Nagahori et al. 2000), methoxychlor O-demethylation (Stresser & Kupfer 1998),
parathion dearylation (Foxenberg et al. 2007, Mutch & Williams 2006), and
bioresmethrin, cypermethrin and trans-permethrin oxidative metabolism (Scollon et al. 2009). Azinphos-methyl, chlorpyrifos, diazinon, parathion and dimethoate
are mainly desulfurated by CYP1A2 at low pesticide concentrations relevant to
human exposure (Buratti et al. 2003, Buratti et al. 2007).
2.2.2 CYP2A subfamily
The human CYP2A subfamily contains three genes i.e. CYP2A6, CYP2A7, and
CYP2A13, and two pseudogenes (Hoffman et al. 1995, Honkakoski & Negishi
1997, Pedro et al. 1995). CYP2A6 represents 10% of the total CYP content in
liver (Pelkonen et al. 2008, Yun et al. 1991). CYP2A6 enzyme activity in the
human liver displays a relatively large variability between individuals, and some
Japanese are known to lack the functional protein completely (Pelkonen et al. 2008, Pelkonen et al. 2000, Shimada et al. 1996). It has been shown that CYP2A6

22
has a major role in the metabolism of nicotine in vitro and in vivo (Kitagawa et al. 1999, Messina et al. 1997, Nakajima et al. 1996a, Nakajima et al. 1996b,
Yamazaki et al. 1999) and in the activation of several nitrosamines (Hecht 1998,
Tiano et al. 1994, Yamazaki et al. 1992) as well as aflatoxin B1 (Salonpää et al. 1993, Yun et al. 1991). The most common in vitro substrate is coumarine, which
is known to be solely catalyzed by CYP2A6 (Pelkonen et al. 2000). A number of
potent inhibitors with variable selectivity against CYP2A6 have been
characterized. The most used in vitro inhibitors include tranylcypromine (Draper et al. 1997, Taavitsainen et al. 2001) and methoxsalen (Draper et al. 1997). More
substrates and inhibitors currently known to be metabolized by or to interact with
CYP2A6 in vitro and in vivo have been summarized by Pelkonen et al. (Pelkonen et al. 2008, Pelkonen et al. 2000).
CYP2A6 participates in the metabolism of quite a few pesticides such as
carbaryl aromatic hydroxylation and methyl oxidation (Tang et al. 2002),
imidacloprid oxidation (Schulz-Jander & Casida 2002), DEET and diuron N-
dealkylation (Abass et al. 2007, Usmani et al. 2002) and carbosulfan N-S
cleavage (Abass et al. 2010).
2.2.3 CYP2B subfamily
CYP2B6 represents approximately 1–10% of the total hepatic CYPs. A notable
interindividual variability in the expression of CYP2B6 has been reported (Code et al. 1997, Faucette et al. 2000, Lang et al. 2001, Stresser & Kupfer 1999,
Yamano et al. 1989). CYP2B6 has a high polymorphic expression and it is
affected by genotype and gender.
CYP2B6 is known to metabolize a large number of substrates including drugs,
pesticides and environmental chemicals, many of which have been described in
detail in reviews by (Ekins & Wrighton 1999, Hodgson & Rose 2007b, Turpeinen et al. 2006b). Clinically used drugs such as cyclophosphamide, tamoxifen, S-
mephenytoin, diazepam, ifosamide and efavirenz are metabolized in part by
CYP2B6 (Granvil et al. 1999, Haas et al. 2004, Huang et al. 2000, Jinno et al. 2003, Roy et al. 1999a, Roy et al. 1999b). CYP2B6 appears to activate and
detoxify a number of precarcinogens (Code et al. 1997, Smith et al. 2003). Ekins
and Wrighton (Ekins & Wrighton 1999) have described the advantages and
disadvantages of several probe substrates for CYP2B6 activity. As a reference
inhibitor for CYP2B6, orphenadrine has been used in drug metabolism studies
even though its selectivity towards this isoform is questionable (Chang et al. 1993,

23
Ekins et al. 1997, Guo et al. 1997, Murray & Reidy 1990). Recently thioTEPA,
an anticancer agent, has been shown to be a specific in vitro inhibitor of CYP2B6
at relatively low concentrations (Harleton et al. 2004, Rae et al. 2002).
CYP2B6 plays a major role in pesticides metabolism. CYP2B6 mediates
alachlor, butachlor, acetachlor and metachlor N-dealkoxylation (Coleman et al. 2000); chlorpyrifos, parathion, malathion and fenthion desulfuration (Buratti et al. 2005, Foxenberg et al. 2007, Leoni et al. 2008, Mutch & Williams 2006, Sams et al. 2000, Tang et al. 2001); endosulfan-α and carbosulfan sulfoxidation (Abass et al. 2010, Casabar et al. 2006, Lee et al. 2006a); metalaxyl O-demethylation and
lactone formation (Abass et al. 2007b); as well as profenofos hydroxylation and
desthiopropylation (Abass et al. 2007a).
2.2.4 CYP2C subfamily
The CYP2C subfamily has four active members, namely 2C8, 2C9, 2C18 and
2C19. CYP2Cs are the second most abundant CYP proteins in human liver and
the CYP2C subfamily consists of three members, comprising about 20% of the
total P450 enzymes. In humans, CYP2C9 is the main CYP2C, followed by
CYP2C8 and CYP2C19, while CYP2C18 is not expressed in liver (Edwards et al. 1998, Gray et al. 1995, Pelkonen et al. 2008, Richardson et al. 1997, Shimada et al. 1994).
CYP2C8 meditates amodiaquine N-deethylation, which is the selective
marker activity, paclitaxel 6α-hydroxylation and cerivastatin demethylation (Li et al. 2002, Rahman et al. 1994). Quercetin, montelukast, zafirlukast and
trimethoprim are potent and specific inhibitors of the CYP2C8 enzyme (Kim et al. 2005, Sousa & Marletta 1985, Walsky et al. 2005a, Walsky et al. 2005b, Wen et al. 2002). A few insecticides are mainly metabolized by CYP2C8. CYP2C8
mediates parathion dearylation and desulfuration (Mutch et al. 2003, Mutch &
Williams 2006); and the oxidative metabolism of deltamethrin, esfenvalerate, and
β-cyfluthrin (Godin et al. 2007, Scollon et al. 2009).
CYP2C9 is a major CYP2C isoform in the human liver, and it is one of
several CYP2C genes clustered in a 500kb region on the proximal 10q24
chromosomal region (Goldstein & de Morais 1994, Gray et al. 1995). In
Caucasian populations, the frequencies of the two variant alleles, CYP2C9*2 and
CYP2C9*3, range from 7% to 19% (Furuya et al. 1995, Ingelman-Sundberg et al. 1999, Miners & Birkett 1998, Stubbins et al. 1996, Sullivan-Klose et al. 1996,
Yasar et al. 1999). CYP2C9 is responsible for the metabolism of the S-isomer of

24
warfarin (Rettie et al. 1992). CYP2C9 also metabolizes tolbutamide, the selective
marker, glipizide, fluvastatin, phenytoin, several non-steroidal anti-inflammatory
agents and many other drug groups (Doecke et al. 1991, Kirchheiner &
Brockmoller 2005, Miners & Birkett 1998, Rettie & Jones 2005, Veenstra et al. 2005). CYP2C9 is inhibited by amiodarone, gemfibrozil, sulphaphenazole and
others (Baldwin et al. 1995, Chen et al. 2004, Heimark et al. 1992, Kumar et al. 2006, Wen et al. 2001).
CYP2C9 is found to be involved in the oxidative metabolism of cis-
permethrin, resmethrin and esfenvalerate insecticides (Godin et al. 2007, Scollon et al. 2009), triphenyltin dearylation and tributyltin dealkylatin (Ohhira et al. 2006), as well as the sulfoxidation of fenthion, phorate sulprofos, disulfoton and
methiocarb insecticides (Leoni et al. 2008, Usmani et al. 2004b).
CYP2C19, another member of the CYP2C enzyme family, represents
approximately 5% of the total hepatic CYPs and metabolizes drugs that are
amides or weak bases with two hydrogen bond acceptors (Lewis 2004, Musana &
Wilke 2005, Pelkonen et al. 2008). Poor metabolizers with low CYP2C19 activity
represent 3 to 5% of Caucasians and African-Americans, and 12 to 23% of most
Asian populations (Goldstein 2001). CYP2C19 participates in the metabolism of
many commonly used drugs including the antiepileptics phenytoin and
mephenytoin (Bajpai et al. 1996, Komatsu et al. 2000, Tsao et al. 2001, Wrighton et al. 1993), selective serotonin receptor inhibitors citalopram and sertraline
(Kobayashi et al. 1997, von Moltke et al. 2001), the psychoactive drug
amitriptyline (Jiang et al. 2002, Venkatakrishnan et al. 1998a) and diazepam,
among others (Jung et al. 1997). Selective marker activities include S-
mephenytoin 4α-hydroxylation and omeprazole 5-hydroxylation (Goldstein et al. 1994, Pelkonen et al. 2008, Wrighton et al. 1993). CYP2C19 inhibitors, such as
tranylcypromine, omeprazole, nootkatone, and ticlopidine are not completely
selective (Jae Wook Ko et al. 2000, Ko et al. 1997, Tassaneeyakul et al. 2000).
(+)-N-3-benzyl-nirvanol and (-)-N-3-benzyl-phenobarbital were found to be
highly potent competitive inhibitors of CYP2C19 (Cai et al. 2004, Suzuki et al. 2002).
Among the substrates of CYP2C19 are several widely used pesticides such as
the phosphorothioate insecticides diazinon, azinophos methyl, chlorpyrifos,
parathion, diazinon, fenthion, phorate, sulprofos, disulfoton and methiocarb
(Buratti et al. 2002, Foxenberg et al. 2007, Kappers et al. 2001, Leoni et al. 2008,
Mutch & Williams 2006, Tang et al. 2001, Usmani et al. 2004b), as well as the
pyrethroid insecticides bifenthrin, s-bioallethrin, bioresmethrin, β-cyfluthrin, λ-

25
cyhalothrin, cypermethrin, cis-permethrin, trans-permethrin, resmethrin,
deltamethrin and esfenvalerate (Godin et al. 2007, Scollon et al. 2009). CYP2C19
mediates the dealkylation of the herbicide diuron and fungicides myclobutanil and
triadimefon (Abass et al. 2007b, Barton et al. 2006).
2.2.5 CYP2D subfamily
CYP2D6 represents 1 to 5% of the total CYP, and approximately 3.5 and 5–10%
of the Caucasian population are ultra-rapid and poor metabolize’s for this enzyme,
respectively. The CYP2D6 gene is clearly the most polymorphic of all known
cytochrome P450s; more than 75 polymorphisms have been identified. Four
alleles account for > 95% of the functional variation observed in the general
population (Al, Omari, A., & Murry 2007, Ingelman-Sundberg 2004, Musana &
Wilke 2005, Pelkonen & Breimer 1994, Pelkonen et al. 2008, Shimada et al. 1994,
Zanger et al. 2004). CYP2D6 metabolizes approximately 20% of all commonly
prescribed drugs in vivo (Brockmöller et al. 2000). For example, CYP2D6
contributes to the metabolism of betablockers metoprolol and timolol (Johnson &
Burlew 1996, Volotinen et al. 2007) and the psychotropic agents amitriptyline and
haloperidol (Coutts et al. 1997, Fang et al. 1997, Fang et al. 2001, Halling et al. 2008, Someya et al. 2003). Dextromethrophan O-demethylation is the most used
in vitro model reaction for CYP2D6 activity (Kronbach et al. 1987, Park et al. 1984). Quinidine is widely used in drug metabolism studies as a highly selective
and specific potent inhibitor of CYP2D6 (Bourrie et al. 1996, Broly et al. 1989,
Newton et al. 1995, Speirs et al. 1986).
The importance of CYP2D6 for pesticide metabolism has been elucidated.
Known pesticide substrates for CYP2D6 include phosphorothioate insecticides
such as diazinon, parathion, chlorpyrifos, sulprofos, disulfoton and methiocarb
(Mutch et al. 2003, Mutch & Williams 2006, Sams et al. 2000, Usmani et al. 2004b) as well as carbamate insecticide (Tang et al. 2002). CYP2D6 is also
involved in the N-dealkylation of the atrazine and diuron herbicides (Abass et al. 2007, Lang et al. 1997).
2.2.6 CYP2E subfamily
Only one gene belonging to this subfamily, namely CYP2E1, has been identified
(Nelson et al. 1996, Nelson et al. 2004). CYP2E1 is one of the most abundant

26
hepatic CYPs, represents 15% of the total P450 and it is also expressed in lung
and brain (Pelkonen et al. 2008, Raunio et al. 1995).
Interest in the role of CYP2E1 as a mediator of toxicity comes from two of its
actions. First, it is known to be important in the metabolic activation and/or
detoxification of a number of carcinogens and hepatotoxins (Kessova &
Cederbaum 2003, Lieber 2004). Second, it may have an important role in free
radical production and oxidative stress. For example, hepatic CYP2E1 generates
reactive oxygen species (ROS), which are linked to Parkinson’s disease
(Gonzalez 2005, Montoliu et al. 1995, Tindberg & Ingelman-Sundberg 1989).
The metabolism of very few clinically important drugs such as paracetamol,
caffine, acetaminophen, enflurane and halothane seems to be mediated to some
extent by CYP2E1 (Gu et al. 1992, Lee et al. 1996, Raucy et al. 1993, Thummel et al. 1993). Chlorzoxazone is probably the most used in vitro model substrate for
CYP2E1 activity (Peter et al. 1990). Pyridine is widely used as a specific in vitro
inhibitor of CYP2E1 (Hargreaves et al. 1994, Pelkonen et al. 2008).
Few pesticides have been reported to be metabolized at least in part by human
CYP2E1 such as atrazine, carbaryl, parathion, imidacloprid and diuron (Abass et al. 2007, Lang et al. 1997, Mutch et al. 2003, Mutch & Williams 2006, Schulz-
Jander & Casida 2002, Tang et al. 2002). One possible explanation for the small
number of pesticides metabolized by CYP2E1 could be the potent inhibitory
effect of DMSO, used often as a solvent to dissolve the pesticides tested, on
CYP2E1.
2.2.7 CYP2F subfamily
CYP2F subfamily consists of one functional gene in mammalian species; in
humans and other primates CYP2F pseudogene is also present. CYP2F genes are
preferentially expressed in the respiratory tract. The best known substrates for
CYP2F enzymes are naphthalene and 3-methylindole (Zhang & Ding 2008). The
role of CYP2F enzymes in catalyzing naphthalene toxicity has been elucidated
(Buckpitt et al. 2002). The recombinant enzyme is active in the dealkylation of
ethoxycoumarin, propoxycoumarin and pentoxyresorufin (Nhamburo et al. 1990).
Moreover, the recombinant CYP2F1 was the most active P450 isoforms in the
conversion of styrene, a lung toxicant, to styrene glycol (Nakajima et al. 1994).
There have no reports of any endogenous compounds that CYP2F enzymes
metabolise. In addition nothing is known about any inducers of CYP2F genes
(Zhang & Ding 2008).

27
2.2.8 CYP2S subfamily
CYP2S1 has been identified in human by Rylander et al. (2001). The human
CYP2S1 gene is located at the proximal end of a cluster of CYP2 family members
on chromosome 19q, which includes the CYP genes CYP2A6, CYP2A13, CYP2B6,
and CYP2F1 (Hoffman et al. 2001). CYP2S1 is expressed in epithelial cells of a
wide variety of extrahepatic tissues. The highest expression levels have been
observed in the epithelial tissues frequently exposed to xenobiotics, e.g., the
respiratory, gastrointestinal, and urinary tracts, and in the skin (Saarikoski et al. 2005a). CYP2S1 is inducible by dioxin and coal tar (Saarikoski et al. 2005b).
CYP2S1 metabolizes naphthalene, a toxic and potentially carcinogenic PAH
(Preuss et al. 2003). CYP2S1 has been described in detail in a review by
Saarikoski et al. (2005b).
2.2.9 CYP3A subfamily
In humans, the CYP3A subfamily contains three functional proteins, CYP3A4,
CYP3A5, and CYP3A7, and one pseudoprotein, CYP3A34. The human CYP3
family constitutes approximately 30% of total hepatic P450 and is estimated to
mediate the metabolism of around 50% of prescribed drugs as well as a variety of
environmental chemicals and other xenobiotics. Because of the large number of
drugs metabolized by CYP3A4, it frequently plays a role in a number of drug-
drug interactions (Bertz & Granneman 1997, Domanski et al. 2001, Imaoka et al. 1996, Musana & Wilke 2005, Pelkonen & Breimer 1994, Pelkonen et al. 2008,
Rostami-Hodjegan & Tucker 2007, Shimada et al. 1994).
CYP3A4 is the major form of P450 expressed in human liver. It is also the
major P450 expressed in human gastrointestinal tract, and intestinal metabolism
of CYP3A4 substrate can contribute significantly to first-pass elimination of
orally ingested xenobiotics (Guengerich 1995, Guengerich 1999). X-ray
crystallography studies demonstrated that CYP3A4 has a very large and flexible
active site, allowing it to oxidize either large substrates such as erythromycin and
cyclosporine or multiple smaller ligands (Scott & Halpert 2005, Tang & Stearns
2001). CYP3A4 participates in the metabolism of several clinically important
drugs such as triazolam, simvastatin, atorvastatin, and quinidine (Bertz &
Granneman 1997, Rendic & Di Carlo 1997). Detailed characteristics of several
CYP3A4 substrates and inhibitors were summarized recently by Liu et al. (Liu et al. 2007). However, testosterone has been shown to be the most commonly used

28
in vitro CYP3A4 probe (50% of reported studies) in contrast to midazolam (15–
20% of in vitro estimates of CYP3A4 activity), whereas nifedipine, felodipine,
and erythromycin were used in less than 10% of studies (Yuan et al. 2002).
The importance of CYP3A4 for pesticide metabolism has been elucidated.
The known pesticides mainly metabolized by CYP3A4 belong to several
chemical groups such as phosphorothioate insecticides (Buratti et al. 2002,
Buratti et al. 2003, Buratti et al. 2005, Buratti & Testai 2007, Butler & Murray
1997, Mutch et al. 2003, Mutch & Williams 2006, Sams et al. 2000, Tang et al. 2001), carbamate insecticides (Abass et al. 2010, Tang et al. 2002, Usmani et al. 2004a), chlorinated cyclodiene insecticides (Casabar et al. 2006, Lee et al. 2006a),
chloroacetamide herbicides (Coleman et al. 2000), conazoles, acylalanine and
other fungicides (Abass et al. 2009, Abass et al. 2007b, Mazur et al. 2007),
neonicotinoid insecticides (Schulz-Jander & Casida 2002), organotin biocide
(Ohhira et al. 2006), phenyl pyrazole insecticide (Tang et al. 2004) and phenyl
urea herbicide (Abass et al. 2007).
CYP3A5 is a minor polymorphic CYP isoform in human liver in addition to
the intestine (Lin et al. 2002, Paine et al. 1997) and kidney (Haehner et al. 1996).
Functional CYP3A5 is expressed in approximately 20% of Caucasians and about
67% of African-Americans (Kuehl et al. 2001). CYP3A5 may have a significant
role in drug metabolism particularly in populations expressing high levels of
CYP3A5 and/or on co-medications known to inhibit CYP3A4 (Soars et al. 2006).
The metabolism of a number of pesticides has been reported to be mediated
by CYP3A5. Parathion, diazinon, chlorpyrifos, malathion and fenthion
desulfuration and/or dearylation (Mutch et al. 2003, Mutch & Williams 2006),
besides deltamethrin, esfenvalerate and carbosulfan oxidative metabolism, are
mainly mediated by CYP3A5 (Abass et al. 2010, Godin et al. 2007).
Expression of CYP3A7 protein is mainly confined to fetal and newborn livers,
although in rare cases CYP3A7 mRNA has been detected in adults (Hakkola et al. 2001, Kitada & Kamataki 1994, Schuetz et al. 1994). CYP3A7 is responsible for
the metabolism of endogenous substrates such as testosterone and DHEA-S in
fetuses (Kitada et al. 1987, Kitada et al. 1987) as well as a few pesticides (Abass et al. 2010, Casabar et al. 2006, Foxenberg et al. 2007). The catalytic activity of
CYP3A7 towards chlorpyrifos, parathion, malathion and fenthion has been
examined (Buratti et al. 2006). Foetal CYP3A7 was able to desulfurate
chlorpyrifos, parathion and malathion but not fenthion. At low chlorpyrifos
concentrations the dearylation reaction, resulting in a non-toxic metabolite, was
highly favored in the foetus.

29
Midazolam, alprazolam and mifepristone are metabolized to an equal or
greater extent by CYP3A5 than by CYP3A4 (Christopher Gorski et al. 1994,
Galetin et al. 2004, Hirota et al. 2001, Huang et al. 2004, Khan et al. 2002,
Williams et al. 2002). Alprazolam has been suggested as a selective probe for
CYP3A5 (Galetin et al. 2004). CYP3A7 has similar catalytic properties compared
with other CYP3A enzymes, including testosterone 6β-hydroxylation (Kitada et al. 1987, Kitada et al. 1985, Kitada et al. 1991). Ketoconazole and itraconazole
are potent inhibitors of CYP3A enzymes but ketoconazole is a relatively selective
inhibitor of CYP3A4 at 1 μM concentration (Baldwin et al. 1995, Newton et al. 1995). Gestodene has been long known as a mechanism-based CYP3A inhibitor
(Pelkonen et al. 1998).
2.3 In vitro and human-derived techniques for testing xenobiotic metabolism
In order to study the metabolism and interactions of pesticides in humans we have
to rely upon in vitro and human-derived techniques. In vitro systems have become
an integral part of drug metabolism studies as well as throughout the drug
discovery process and in academic research (Lin & Lu 1997, Pelkonen & Raunio
2005). In vitro approaches to predict human clearance have become more
frequent with the increase in the availability of human-derived materials (Skett et al. 1995). All models have certain advantages and disadvantages, but the common
advantage to these approaches is the reduction of the complexity of the study
system. However, the use of in vitro models is always a compromise between
convience and relevance (Brandon et al. 2003, Pelkonen & Turpeinen 2007,
Pelkonen & Raunio 2005, Pelkonen et al. 2005). An overview of different in vitro
models and their advantages and disadvantages are collected in Table 1 and
discussed below.

30
Table 1. An overview of different in vitro models and their advantages and
disadvantages (modified from Brandon et al. 2003, Pelkonen & Turpeinen 2007,
Pelkonen & Raunio 2005, Pelkonen et al. 2005).
Enzyme sources Availability Advantages Disadvantages
Liver homogenatesa Relatively good.
Commercially available.
Human liver samples
obtained under proper
ethical permission.
Contains basically all
hepatic enzymes.
Liver architecture lost.
Cofactors are necessary.
Microsomesa Relatively good,
from transplantations or
commercial sources.
Contains important rate-
limiting enzymes.
Inexpensive technique.
Easy storage. Study of
individual, gender-, and
species-specific
biotransformation.
Contains only CYP and
UGTs.
Requires strictly specific
substrates and inhibitors
or antibodies.
Cofactor addition
necessary.
cDNA-expressed
individual CYPsb
Commercially available The role of individual
CYPs in the metabolism
can be easily studied.
Different genotypes.
High enzyme activities.
The effect of only one
enzyme at a time can be
evaluated.
Problems in extrapolation
to HLM and in vivo.
Primary hepatocytesc,d Difficult to obtain,
relatively healthy tissue
needed. Commercially
available
Contains the whole
complement of CYPs
cellularly integrated.
The induction effect can
be studied.
Well established and
characterized.
Transporters still present
and operational.
Requires specific
techniques and well
established procedures.
The levels of many CYPs
decrease rapidly during
cultivation.
Cell damage during
isolation.
Liver slicese Difficult to obtain, fresh
tissue needed.
Contains the whole
complement of CYPs and
cell-cell connections. The
induction effect can be
studied. Morphological
studies possible.
Interindividual variation
can be studied.
Requires specific
techniques and well
established procedures

31
Enzyme sources Availability Advantages Disadvantages
Immortalized cell linesf Available upon request,
not many characterized
cell lines exist.
Non-limited source of
enzymes. Easy to
culture.
Relatively stable enzyme
expression level. The
induction effect can be
studied.
The expression of most
CYPs is poor.
a (Kremers 1999); b (Rodrigues 1999); c (Guillouzo 1995); d (Gomez-Lechon et al. 2004); e (Olinga et al.
1998); f (Allen et al. 2005).
2.3.1 Subcellular fractions (liver microsomes and homogenates)
Subcellular fractions including microsomes are the most widely used in vitro
systems in drug metabolism studies of new drug candidates (Ekins et al. 2000,
McGinnity & Riley 2001). Microsomes can be prepared easily from frozen liver
tissue, and enzymatic activities are stable during prolonged storage (Beaune et al. 1986, Pearce et al. 1996, Yamazaki et al. 1997). Microsomes consist of vesicles
of the hepatocyte endoplasmic reticulum and are prepared by standard differential
ultracentrifugation (Pelkonen et al. 1974). Liver homogenates centrifuged at
9000g results in a supernatant (S9 fraction) rich in drug-metabolizing enzymes
and a pellet containing predominately nuclei, lysosomes, peroxisomes and
mitochondria. Centrifuging the S9 fraction at 100,000g results in a supernatant
(cytosol) and pellet (microsomes). Liver preparations, other than from fresh
human liver, can also be used (e.g., liver slices, liver cell lines, and primary
hepatocytes) for the preparation of microsomes (Olsen et al. 1997, Skaanild &
Friis 2000).
Hepatic microsomes are useful for evaluation of metabolic stability,
metabolite identification and quantification, and enzyme inhibition. Microsomes
have many advantages including easy adaptation to higher throughput assays,
easy preparation and use, good stability during storage, high CYP concentration
and high rate of metabolite turnover. On the other hand, microsomes contain only
phase I enzymes and UGTs, while hepatic homogenates contain basically all
hepatic enzymes. The NADPH-regenerating system is required for CYP enzymes,
while the addition of appropriate cofactors is necessary (e.g UDPGH, GSH and
PAPS) for the catalytic activity of phase II enzymes (Brandon et al. 2003, Coecke et al. 2006, Ekins et al. 1999, Ekins et al. 2000, Pelkonen et al. 2005).

32
2.3.2 cDNA-expressed CYPs (single-enzyme systems)
Recombinant xenobiotic metabolizing enzymes are well established and
commercially available e.g. SupersomesTM. These enzymes are expressed in
bacterial (Fisher et al. 1992), yeast (Peyronneau et al. 1992), and mammalian cell
lines (Guengerich 1995) and human lymphoblast or baculovirus-infected insect
cells (Asseffa et al. 1989). A NADPH-regenerating system (which consists of β-
NADP, glucose 6-phosphate, and glucose 6-phosphate dehydrogenase) or
NADPH is required to supply the energy demand of the CYPs (Taavitsainen et al. 2000).
The presence of a mixture of CYPs and the wide variability in enzyme
profiles between individual samples of liver microsomes makes them difficult to
use for establishing the role of a specific enzyme in the metabolism of a
compound. To overcome these problems, recombinant enzymes are easily applied
for higher throughput screening and they can be used to study the role of
individual CYPs in the metabolism or to produce usable amounts of metabolites.
The disadvantage is that the effect of only one enzyme at a time can be evaluated
(McGinnity & Riley 2001, Moody et al. 1999, Pelkonen et al. 2005).
2.3.3 Hepatocytes
Primary cultures of hepatocytes have shown good in vitro - in vivo correlations in
the metabolic activity of a number of drugs and they allow a comprehensive view
of whole cell metabolism since drug transporters and both phase I and II enzymes
are present. Cultured human hepatocytes can be used to study CYP-mediated
metabolism and induction only when maintained under optimal conditions.
Hepatocytes are used to evaluate the metabolic stability of the compounds and
identify the metabolizing enzymes and enzyme inhibition, much like the use of
microsomes with the advantage of a broader complement of phase I and II
enzymes and transporters (Ferrini et al. 1997, Gomez-Lechon et al. 2004, Hewitt et al. 2007, Kostrubsky et al. 1999, LeCluyse 2001, Li et al. 1997, Pelkonen &
Turpeinen 2007). Hepatocytes in suspension are technically the easiest of all
hepatocytes systems. The major limitation of use of suspensions is that
hepatocytes in suspensions stay viable for a short period (up to 4 h), which
prevents investigation of slowly metabolizing compounds. However, interspecies
comparison seems to be more reliable with hepatocytes in suspensions than in
culture due to potential damage of cytotoxic substances and variations in the

33
expression levels of xenobiotic metabolizing enzymes in cultures (Gebhardt et al. 2003).
2.3.4 Liver slices
Like the hepatocytes, as whole cell systems, liver slices contain the whole
complement of xenobiotic metabolizing enzymes and offer a more reliable in vivo/in vitro correlation than subcellular systems (Gomez-Lechon et al. 2004,
Pelkonen et al. 2005). Liver slices have begun to slowly fall out of use in the
prediction of pharmacokinetic properties and drug metabolism due predominantly
to issues associated with drug movement into and out of the slices, lower enzyme
activities and the increased use of hepatocytes to study similar reactions (Carlile et al. 1999, Olinga et al. 1998, Worboys et al. 1996, Worboys et al. 1997).
However, liver slices are still a powerful tool to study biotransformation in vitro
(Brandon et al. 2003, Ekins et al. 2000).
2.3.5 Immortalized cell lines
Different human and animal cell lines are available (http://www.lgcpromochem-
atcc.com). Because of interspecies differences, animal cell lines cannot predict
human biotransformation. Human liver cell lines can be isolated from primary
tumors of the liver parenchyma, developed after chronic hepatitis or cirrhosis
(Crommelin & Sindelar 1997). The most common human cell lines for
biotransformation studies are probably HepG2 (hepatocellular carcinoma)
(Galijatovic et al. 1999, Urani et al. 1998, Walle et al. 2000, Yoshitomi et al. 2001), BC2 (hepatoma) (Gomez-Lechon et al. 2001) and the lung-derived line
A549 (Hukkanen et al. 2000). HepG2 is also an excellent tool to detect genotoxic
properties of environmental and dietary chemicals (Knasmüller et al. 2004).
Recently a new human hepatoma cell line, HepaRG, was derived from a
hepatocellular carcinoma. The HepaRG cells express a large panel of liver-
specific genes including several cytochrome P450 enzymes such as CYP1A2,
CYP2B6, CYP2C9, CYP2E1 and CYP3A4, which is in contrast to the other
hepatoma cell lines such as HepG2. In addition to P450 enzymes, HepaRG cells
have a stable expression of phase II enzymes, transporters and nuclear
transcription factors over a time period of six weeks in culture (Aninat et al. 2006,
Anthérieu et al. 2010, Kanebratt & Andersson 2008, Turpeinen et al. 2009).

34
2.4 In vitro characterization of the metabolism and metabolic interactions of xenobiotics
The aim of in vitro characterization is to produce relevant and useful information
on metabolism and interactions to anticipate, and even to predict, what happens in vivo in man. Each in vitro model has its own set of advantages and disadvantages
as they range from simple to more complex systems: individual enzymes,
subcellular fractions, cellular systems, liver slices and whole organ, respectively
(Brandon et al. 2003, Pelkonen & Raunio 2005, Pelkonen et al. 2005). To
understand some of the factors related to xenobiotic metabolism that can
influence the achievement of these aims, there are several important points to
consider such as (Hodgson & Rose 2005, Pelkonen & Turpeinen 2007, Pelkonen
& Raunio 2005, Pelkonen et al. 2005):
– Determination of the metabolic stability of the compound
– Identification of reactive metabolites
– Evaluation of the variation between species
– Identification of human CYPs and their isoforms involved in the activation or
detoxification
– Evaluation of the variation between individuals
– Identification of individuals and subpopulations at increased risk
– Overall improvement of the process of human risk assessment
An overview of different in vitro studies for the characterization of
metabolism and metabolic interactions of xenobiotics are collected in Table 2 and
discussed below.
2.4.1 Metabolic stability of the compound
Several in vitro test systems (microsomes, homogenates, hepatocytes or
recombinant proteins) are available to determine metabolic stability. In vitro
methods for the study of stability are probably the most well established of drug
metabolism systems. As a result, if a new chemical entity is rapidly metabolized
in vitro, its bioavailability and persistence in vivo are most probably too low for it
to be a drug candidate. There are two main components to any in vitro screen for
metabolic stability; namely, a system for metabolizing the compounds and a
means of analyzing the metabolic reaction (Eddershaw & Dickins 1999, Tolonen et al. 2009). Similar studies performed in human and test species give valuable
35
information for the selection of test species for pharmacokinetic and toxicological
in vivo studies. Moreover, the determination of metabolic stability of human in vitro incubation systems results in the possibility of extrapolation to the in vivo
situation (Pelkonen & Turpeinen 2007, Riley et al. 2005, Tolonen et al. 2009).
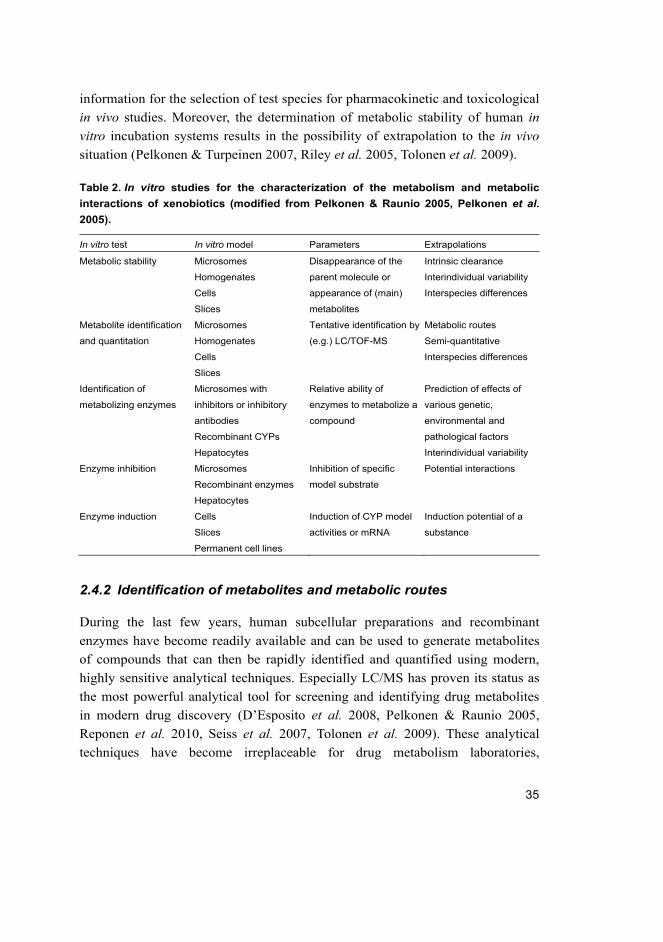
Table 2. In vitro studies for the characterization of the metabolism and metabolic
interactions of xenobiotics (modified from Pelkonen & Raunio 2005, Pelkonen et al. 2005).
In vitro test In vitro model Parameters Extrapolations
Metabolic stability Microsomes
Homogenates
Cells
Slices
Disappearance of the
parent molecule or
appearance of (main)
metabolites
Intrinsic clearance
Interindividual variability
Interspecies differences
Metabolite identification
and quantitation
Microsomes
Homogenates
Cells
Slices
Tentative identification by
(e.g.) LC/TOF-MS
Metabolic routes
Semi-quantitative
Interspecies differences
Identification of
metabolizing enzymes
Microsomes with
inhibitors or inhibitory
antibodies
Recombinant CYPs
Hepatocytes
Relative ability of
enzymes to metabolize a
compound
Prediction of effects of
various genetic,
environmental and
pathological factors
Interindividual variability
Enzyme inhibition Microsomes
Recombinant enzymes
Hepatocytes
Inhibition of specific
model substrate
Potential interactions
Enzyme induction Cells
Slices
Permanent cell lines
Induction of CYP model
activities or mRNA
Induction potential of a
substance
2.4.2 Identification of metabolites and metabolic routes
During the last few years, human subcellular preparations and recombinant
enzymes have become readily available and can be used to generate metabolites
of compounds that can then be rapidly identified and quantified using modern,
highly sensitive analytical techniques. Especially LC/MS has proven its status as
the most powerful analytical tool for screening and identifying drug metabolites
in modern drug discovery (D’Esposito et al. 2008, Pelkonen & Raunio 2005,
Reponen et al. 2010, Seiss et al. 2007, Tolonen et al. 2009). These analytical
techniques have become irreplaceable for drug metabolism laboratories,

36
providing large amounts of information from a wide variety of samples. It has to
be stressed that mass spectrometers are not all the same and choosing the wrong
LC–MS system may lead to compromised data quality or biased results as well as
wasted time and effort (Tolonen et al. 2009).
2.4.3 Identification of the metabolizing enzymes
If the results of the metabolic stability and metabolic routes in human in vitro
systems indicate that CYP enzymes contribute significantly to the metabolism of
a xenobiotic, then identification of the individual CYP enzyme(s) involved in the
metabolism of a xenobiotic is necessary for in vitro – in vivo extrapolation and
prediction. Due to the broad substrate specificity of CYP enzymes, it is possible
for more than one enzyme to be involved in the metabolism of a single compound.
In vitro methods have been established to determine which CYP isoform(s) is (are)
involved in the metabolism of a xenobiotic (Pelkonen & Raunio 2005, Pelkonen et al. 2005). The approaches employed for this purpose will be presented below.
cDNA-expressed enzymes
The availability of a full panel of recombinant enzymes covering the major
human liver CYPs allows a direct approach for assaying the metabolism of a
compound by incubation with the isolated isoforms. This can be done by
following substrate consumption or product formation by each isoform using the
same analytical methods as for HLM-based assays (Reponen et al. 2010). The
biotransformation of a xenobiotic by a single CYP does not necessarily mean its
participation in the reaction in vivo. The relative roles of individual CYPs cannot
be quantitatively estimated using this approach due to the interindividual variation
in the levels of individual active CYPs in the liver (Guengerich 1999, Guengerich
1995). However, cDNA-expressed CYPs are well suited for isozyme
identification in a high-throughput screening format (White 2000). The relative
importance of individual isoform to in vivo clearance is dependent upon the
relative abundance of each isoforms. When taking into account the average
composition of human hepatic CYPs, an approximate prediction of the
participation of any CYP enzyme in the whole liver activity can be achieved
(Rodrigues 1999, Rostami-Hodjegan & Tucker 2007).

37
Correlation studies
Using a bank of “phenotyped” liver microsomes, correlation analysis could be
performed. Correlation analysis involves measuring the rate of xenobiotic
metabolism by several liver samples from individual humans and correlating
reaction rates with the level of activity of the individual CYP enzymes in the
same microsomal samples. If there are a sufficient number of individual samples
(at least ten), the correlation plot would give the information needed for the
evaluation of the participating CYPs. The higher the correlation between the
activities, the larger the probability that the respective CYP enzyme is responsible
for the metabolism of the xenobiotic. Another approach is to correlate the levels
of an individual CYP determined by Western blot analysis against the metabolic
activity (Beaune et al. 1986, Berthou et al. 1994, Brandon et al. 2003, Guengerich
1995, Jacolot et al. 1991, Wolkers et al. 1998).
Inhibition studies with CYP-selective chemical inhibitors and specific antibodies
Pooled human liver microsomes or individual liver microsomal samples should
be used to examine the effect of CYP-selective chemical inhibitors or selective
inhibitory antibodies. Antibody inhibition involves an evaluation of the effects of
inhibitory antibodies against selective CYP enzymes on the metabolism of a
xenobiotic in human liver microsomes. Chemical inhibition involves an
evaluation of the effects of known CYP enzyme inhibitors on the metabolism of a
xenobiotic. Several compounds have been characterized for their inhibitory
potency against different CYPs; for example, furafylline is perhaps the most
potent and selective inhibitor of CYP1A2, tranylccypromine of CYP2A6, thio-
tepa and ticlopidine of CYP2B6, trimethoprim and sulfamethoxazole are selective
inhibitors of CYP2C8 and CYP2C9, respectively, fluconazole of CYP2C19,
quinidine is a commonly used in vitro diagnostic inhibitor of CYP2D6 activity,
pyridine and disulfiram of CYP2E1, and ketoconazole and itraconazole are
among many potent and relatively selective inhibitors of CYP3A4 often used in vitro and in vivo as diagnostic inhibitors (Bourrie et al. 1996, Clarke et al. 1994,
Nebert & Russell 2002, Pelkonen et al. 2008, Pelkonen et al. 2005, Rendic & Di
Carlo 1997, Schmider et al. 1995, Sesardic et al. 1990).

38
Inhibition of CYP enzymes
Testing the inhibitory interactions of a xenobiotic on CYP-specific model activity
in human liver microsomes in vitro provides information about the affinity of the
compound for CYP enzymes (Pelkonen & Raunio 2005). The type of CYP
inhibition can be either irreversible (mechanism-based inhibition) or reversible.
Irreversible inhibition requires biotransformation of the inhibitor, while reversible
inhibition can take place directly, without metabolism. Reversible inhibition is the
most common type of enzyme inhibition and can be further divided into
competitive, noncompetitive, uncompetitive, and mixed-type inhibition (Pelkonen et al. 2008). The inhibitory interactions of a xenobiotic on CYP enzymes can be
tested by co-incubating a series of dilutions of a xenobiotic with a reaction
mixture containing single or multiple substrates. In the single substrate assay,
traditionally CYP interaction studies are performed using specific assays for each
CYP isoform. A decrease in probe metabolite formation produced by inhibition is
usually analyzed by LC-UV, LC-MS or fluorometry. In the cocktail assay, several
CYP-selective probes are incubated with human liver microsomes and analyzed
by LC-MS-MS (Tolonen et al. 2007, Turpeinen et al. 2005, Turpeinen et al. 2006a).
2.5 Health risk assessment of chemicals
2.5.1 General considerations
In the health risk assessment of chemicals, the determination of a NOAEL (No
Observed Adverse Effect Level) is often based on data only from animal
experiments. The safety or uncertainty factor 100 is used to convert NOAEL from
an animal toxicity study to an ADI (Acceptable Daily Intake) value for human
intake, ADI = NOAEL/100. Historically, the assessment factor of 100 intended to
cover the interspecies (animal-to-human) and interindividual (human-to-human)
variations has often been used as a default (Bigwood 1973, Lu 1979, Vettorazzi
1977). Based on factor 100, Renwick (Renwick 1991, Renwick 1993) has
attempted to provide a scientific basis for the default values of 10 for interspecies
and 10 for interindividual variability. Renwick also proposed a division of each of
these factors into subfactors to allow for separate evaluations of differences in
toxicokinetic and toxicodynamic based on the relative magnitude of
toxicokinetics and toxicodynamics variation between and within species. He

39
proposed that the 10-fold factors for inter- and intraspecies variation should be
subdivided into factors of 4 for toxicokinetics and 2.5 for toxicodynamics.
WHO/IPCS (WHO/IPCS 1994) has adapted the Renwick approach with one
deviation. While the uncertainty factor (UF) for interspecies extrapolation should
be divided into default values of 4 for toxicokinetics and 2.5 toxicodynamics, the
UF for interindividual variation should be divided into 3.16- fold for both
toxicokinetics and toxicodynamics. The reason for this deviation from Renwick’s
proposal was that the WHO/IPCS considered the slightly greater variability in the
kinetics in humans compared with dynamics was not sufficient to warrant an
unequal subdivision of the 10-fold factor into a toxicokinetic factor of 4 and a
toxicodynamic factor of 2.5 (Figure 5).
Fig. 5. The subdivision of the 100-fold default uncertainty factor (modified from
WHO/IPCS 2005).
The Swedish Chemical Agency, KEMI, (KEMI 2003) has recommended the
inclusion of assessment factors in risk assessment, when justifiable, since the
scientific background for default assessment (uncertainty) factors in general
remains unsatisfactory. WHO/IPCS, (WHO/IPCS 2005) has proposed the use of

40
chemical-specific toxicological data instead of default assessment factors,
whenever possible. Consequently, Chemical-Specific Adjustment Factors (CSAFs)
have been introduced to provide a method for the incorporation of quantitative
data on interspecies differences or interindividual variability in either
toxicokinetics or toxicodynamics into the risk assessment procedure, by
modifying the relevant default UF of 10. The basic idea is to replace one or more
of these subfactors with chemical-specific factors that can be derived from
quantitative chemical-specific data. The assessment factor may increase or
decrease and the differences may be considerable (Figure 6) (Dorne et al. 2005,
WHO/IPCS 2005).
The European Union procedures of pesticides marketing, use, residues in
food and their risk assessment associated to the regulatory approval of
commercialisation are regulated by Council Directive 91/414/EEC and regulation
(EC) No 396/2005 (http://ec.europa.eu/food/plant/protection/index_en.htm).
Fig. 6. Chemical specific adjustment factors can replace the default uncertainty
factors (WHO/IPCS 2005), modified from (Dorne et al. 2005).

41
2.5.2 Interspecies differences and interindividual variability in toxicokinetics
Due to data limitation for most substances, an assessment factor is usually applied
for extrapolation. Species-specific differences, based on chemical-specific data,
between animals and humans should be taken into consideration in the
extrapolation of data from animal studies to humans (WHO/IPCS 2005). The
interspecies assessment factor is generally recognized as providing an
extrapolation from an average animal to an average human being, assuming that
humans are 10-fold more sensitive than experimental animals. The assessment of
species differences in glucuronidation and the metabolism of human CYP1A2
substrates supported the need to replace the default factor by compound-specific
quantitative data (Walton et al. 2001a, Walton et al. 2001b). KEMI (KEMI 2003)
and Falk-Filipsson et al. (Falk-Filipsson et al. 2007) suggested a species-specific
default factor, if no chemical-specific data are available. This factor should be
based on differences in caloric demand and is thus 4 for extrapolation from rats, 7
from mice, 3 from guinea pigs, 2.4 from rabbits and 1.4 from dogs.
Human susceptibility to chemicals may depend on a number of determinants,
including for example the genotype and gender of the individual, age, growth and
development, health status and lifestyle factors (Falk-Filipsson et al. 2007).
Laboratory animals are initially healthy, of similar age and are fed with the same
feed. Thus, interindividual variation in sensitivity towards toxic agents can be
expected to be much higher within the human population than in inbred strains of
laboratory animals. It has been suggested by KEMI (KEMI 2003) that an
interindividual factor of 3-5, with differences in sensitivity due to enzymatic
polymorphisms not included in this estimate, might be sufficient to reflect the
toxicokinetic variability between healthy adults. Falk-Filipsson et al. (Falk-
Filipsson et al. 2007) concluded based on several analyses (Dorne et al. 2001a,
Dorne et al. 2001b, Dorne et al. 2002, Dorne et al. 2003a, Dorne et al. 2003b,
Dorne et al. 2004a, Dorne et al. 2004b, Ginsberg et al. 2002, Hattis et al. 1987,
Hattis et al. 2003, Kalberlah & Schneider 1998, Renwick 1993) that an inter-
individual factor of 3.2 might not be appropriate to reflect the toxicokinetic
variability between individuals, and they suggested a factor of 4.5 as a minimum.
However, neonates and some genetic subgroups, such as CYP2D6 and CYP2C19
poor metabolizers (PM), may need much higher factors. Pregnant women, infants,
elderly and patients with kidney and liver diseases are other sensitive subgroups.

42
2.5.3 Considerations regarding in vitro data
Risk assessment needs reliable scientific information and one source of
information is the characterization of metabolic fate and toxicokinetics. Human
data are needed in order to obtain quantitative toxicokinetic data for comparisons
between individuals or between animals and humans (Falk-Filipsson et al. 2007).
Because humans cannot be used as study subjects with pesticides except in
occasional circumstances, we have to rely upon in vitro and animal experiments.
In vitro metabolic data can be used as a sufficient data basis to derive a chemical-
specific adjustment factor (Gundert-Remy & Sonich-Mullin 2002, KEMI 2003)
taking into account that metabolism is usually the most important factor
contributing to interindividual and interspecies differences in toxicokinetics.
Using in vitro-derived techniques such as microsomes, the kinetic parameters
Vmax (the maximal velocity of the reaction) and Km (the substrate concentration
that gives one-half Vmax) values and thus, the intrinsic clearance (CLint, LM =
Vmax/Km), can be determined experimentally. A comprehensive discussion of
assumptions behind these measures can be found in an excellent report by
Houston (Brian Houston 1994). However, in vitro chemical specific quantitative
data scaled to determine and compare the efficiency of catalysis CLint, LM between
species, e.g. the clearance in each tested mammalian microsome, is divided by the
clearance in humans and between the individuals. The MPPGL, milligrams of
microsomal protein per gram of liver, as well as the size of the liver are needed
for the extrapolation (Barter et al. 2007, Johnson et al. 2005). For a complete
discussion and extrapolation, see the excellent report by Pelkonen and Turpeinen
(Pelkonen & Turpeinen 2007) and results and discussion sections below.
2.6 Pesticides as representative xenobiotics
2.6.1 General information
The use of pesticides is a hotly debated topic (Steerenberg et al. 2008). While
they can be extremely useful in terms of protecting plants and public health, most
pesticides create risk or harm, by their very nature. Pesticides can cause harm to
humans, animals, or the environment because they are designed to kill or
otherwise adversely affect living organisms. Since pesticides can provide such a
benefit to humans, exposure to pesticides is a global challenge to risk assessment.
This is why pesticides are used as representative xenobiotics in this application.

43
The criteria of selecting the study compounds were the extent of use, lack of
detailed knowledge and prospective benefits for risk assessment. The chemical
structures of the study compounds are shown in Figure 7 and their hazard
characterization is collected in Table 3.
There are more than 1500 pesticides listed in the compendium of pesticides
common names. Each group of pesticides is subdivided into chemical or other
classes. Pesticides can be classified by target organism, chemical structure,
physical state (American Medical Association 1997), or the WHO recommended
classification of pesticides by hazards (IPCS 2005). In the 1940s manufacturers
began to produce large amounts of synthetic pesticides and their use became
widespread (Daly et al. 1998). In 2001 over five billion pounds of pesticides were
used worldwide (USEPA 2000–2001).
Fig. 7. Chemical structures of the study compounds.

44
Table 3. Hazard characterization of the study compounds (Data obtained from
European Commission pesticides database (http://ec.europa.eu/food/plant/protection/-
-evaluation/database_act_subs_en.htm).
Pesticide ADI (mg/kg bw) ARfD (mg/kg bw) AOEL (mg/kg bw) MRL (mg/kg)
Metalaxyl 0.08 0.5 0.08 0.05 – 2.0
Profenofos 0.03 1.0 -- 0.05 – 0.2
Diuron 0.007 0.016 0.007 0.05 – 0.2
Carbosulfan 0.01 0.01 -- 0.05 – 0.1
- ADI, acceptable daily intake, the amount of a chemical that can be consumed every day for a lifetime
in the practical certainty, on the basis of all known facts, that no harm will result
- ARfD, acute reference dose, the amount of a chemical that can be consumed at one meal or on one
day in the practical certainty, on the basis of all known facts, that no harm will result
- AOEL, acceptable operator exposure level, a level of daily exposure that would not cause adverse
effects in operators who work with a pesticide regularly over a period of days, weeks or months.
- MRL, maximum residue level, the maximum concentration of pesticide residue likely to occur in or on
food and feeding stuffs after the use of pesticides according to Good Agricultural Practice (GAP).
2.6.2 Individual pesticides studied
Metalaxyl
Metalaxyl is a highly effective systemic acylanilide fungicide. It is registered for
use in many countries worldwide, including the United States, Europe and
Australia, on a variety of fruits and vegetable crops for control of plant diseases
caused by Phythophthora infestans and Pythiumultimum (Sukul & Spiteller 2000).
The mode of action is to inhibit protein synthesis in fungi by interfering with the
synthesis of ribosomal RNA (Tomlin 2000). Metalaxyl is tolerant to a wide range
of pH, light, and temperature and this leads to its continued use in agriculture. It
has a tendency to migrate to deeper soil horizons with a potential to contaminate
groundwater, particularly in soils with low organic matter and clay content (Sukul
& Spiteller 2000).
Metalaxyl can induce bradycardia in rats and affect the activity of
monoamine oxidase in the rat heart (Naidu & Radhakrishnamurty 1988, Naidu &
Radhakrishnamurty 1989, Naidu 1989). Metalaxyl interacts with the RNA
polymerase-I-template complex inhibiting incorporation of ribonucleotide
triphosphates into ribosomal RNA (Hassall 1990). No evidence of a carcinogenic
or genotoxic potential of metalaxyl has been mentioned in a FAO/WHO report
(2002). Meanwhile, some data indicate a cocarcinogenic potential of metalaxyl,

45
which could be sufficient to induce critical DNA damage either directly or
indirectly (Paolini et al. 1996) and can cause chromosomal changes in humans
and animals (Hrelia et al. 1996).
A few studies on the metabolism of metalaxyl in mammals, rats and goats,
have been reported (FAO/WHO 2002). The metabolic fate of 14C metalaxyl in rats
after administration of a single oral dose of 28 mg/kg and 48 h collection of urine
and faeces was studied. Altogether more than 30 different metabolites were
detected in mammals, some of which were identified by gas chromatography, MS
and NMR, and the metabolism was rapid with more than 60% of the administered
dose excreted within 24 h. Metabolism proceeds through several steps, leading to
glucuronic acid conjugates via methyl ester cleavage and benzyl methyl oxidation.
Didemethyl metalaxyl metabolite was predominant (39%) followed by hydroxy
aromatic methyl (14%) and demethyl ester (4.1%). Minor amounts of phenyl ring
hydroxylated (0.3%), ether group demethylated (0.9%) and lactone metabolite
(0.6%) were also detected. The potential N-dealkylation metabolite of metalaxyl
2,6-dimethylaniline has been proven to be a rat nasal carcinogen (Kornreich &
Montgomery 1990, Tyden et al. 2004).
Profenofos
Profenofos is an organophosphate (OP) non-systemic foliar insecticide and
acaricide with contact and stomach actions used widely for agricultural and
household purposes. Profenofos is moderately toxic to birds and has low
mammalian toxicity (Tomlin 2000).
The insecticidal and nematicidal activities of profenofos are attributed to
inhibition of the enzyme acetylcholinesterase (AChE), which plays an important
role in neurotransmission at cholinergic synapses by rapidly hydrolyzing the
neurotransmitter acetylcholine to choline and acetate (Fulton & Key 2001, Ozmen et al. 1999). Profenofos is activated to desthiopropylprofenofos through the
oxidation of the sulphur in the P-S-n-propyl bond to the more potent AChE
inhibitor (Wing et al. 1983, Wing et al. 1984). The primary effect of profenofos in
studies of acute, short, and long-term toxicity was inhibition of
acetylcholinesterase activity. However, the inhibitory effects of OP insecticides
are dependent on their binding capacity to the enzyme active site as well as their
rate of oxidation (Jokanovic 2001). Profenofos is a moderately severe toxicant,
causing relatively delayed respiratory failure and death (Eddleston et al. 2009).
Profenofos downregulates hepatic and testicular CYP2C11 and CYP3A2 mRNA

46
and protein expression levels as well as decreases the testicular aromatase
(CYP19A) mRNA in treated rats (Moustafa et al. 2008). An evaluation of the
CYPs inhibition potential of selected pesticides in human hepatic microsomes
illustrated that organophosphorus insecticides are potent and extensive inhibitors
of CYP1A1/2, CYP2B6, CYP2C8, CYP2C9, CYP2D6 and CYP3A4 (Abass et al. 2009).
Only a few studies on profenofos metabolism in mammals have been
reported (FAO/WHO 2007). After single and repeated doses of [Phenyl-14C]
profenofos to rats, unchanged profenofos was detected in the feces, but the
amount was very small (approximately 1–2% of the administered dose), and this
was probably the proportion of the dose that was not absorbed. Four major
metabolites were present in urine and no unchanged profenofos was detected. The
major metabolites were the sulfate and glucuronide conjugates of 4-bromo-2-
chlorophenol that were formed by hydrolysis of the aryloxy–phosphorus bond
followed by conjugation with sulfate or glucuronic acid. The other two
metabolites were formed by cleavage of the phosphorus– sulfur bond either by
loss of the propyl group or hydrolysis. The 4-bromo-2-chloro-phenol was
detected in some urine samples, but probably arose as a result of hydrolysis of the
conjugates after excretion.
Diuron
Diuron, one of the most commonly used herbicides, belongs to the family of
halogenophenylureas that represents an important class of contact herbicides
applied in pre- and post-emergence to control broadleaf weeds in a wide variety
of annual and perennial broadleaf and grass weeds (Field et al. 1997, Gooddy et al. 2002). Diuron has been classified as slightly hazardous (toxicity class III)
pesticide by WHO. Diuron is relatively persistent in the environment (with a half-
life of over 300 days). As a result, it is often detected in groundwater and surface
water (Blanchoud et al. 2004, Giacomazzi & Cochet 2004, Lapworth & Gooddy
2006, Malato et al. 2003). Diuron has been detected in marinas and coastal areas
in the United Kingdom, Germany and Japan (Lamoree et al. 2002, Okamura et al. 2003, Thomas et al. 2001). Diuron is included in the list of priority pollutants of
the EU Water Framework Directive (European Commission 2001). Also, the
United States Environmental Protection Agency reported diuron on the list of
substances considered contaminants of drinking water since 1998 (USEPA 2005).

47
The United States Environmental Protection Agency (USEPA) evaluated
diuron as a “known/likely” carcinogen to humans. Meanwhile, no evaluation has
been made for diuron carcinogenicity by FAO/WHO so far. In a study conducted
on male and female Wistar rats and NMRI mice at dietary levels of 25, 250 and
2500 ppm of diuron, diuron showed carcinogenic activity in the highest dose-
treated animals (i.e., increased incidences of urinary bladder and renal pelvis
tumors in both genders of Wistar rats and of mammary tumors in female NMRI
mice) (USEPA 2004). Moreover, diuron also showed in vivo mutagenic activity in
Swiss mice by the bone marrow micronucleus test (Agrawal et al. 1996). Only a
few studies on diuron metabolism in mammals have been published. In metabolic
studies of rats and dogs, N-(3,4-dichlorophenyl)urea was the predominant
metabolite in the urine. Small amounts of N-(3,4-dichlorophenyl)-N-methylurea,
3,4-dichloroaniline, 3,4-dichlorophenol and unchanged diuron were detected
(Hodge et al. 1967). In a human postmortem case, diuron and its demethylation,
didemethylation and hydroxylation metabolites were identified in plasma and
urine (Verheij et al. 1989). On the other hand, in a hospitalized case, diuron was
completely metabolized mainly via demethylation and didemethylation. In
addition urine extracts contained hydroxyphenyldiuron and 3,4-dichloroaniline at
minor concentrations (Van Boven et al. 1990).
Carbosulfan
Carbosulfan belongs to the carbamates class of pesticides and to carbofuran
sulfonylated derivatives. Carbosulfan is a systemic insecticide with contact and
stomach actions, extensively used in agriculture for broad spectrum control of
insect pests of crops. Carbosulfan toxicological evaluation was compiled by
FAO/WHO (2003).
Carbosulfan has been classified as a moderately hazardous (toxicity class II)
pesticide by WHO with a low ADI of 0.0 – 0.01 mg/kg body weight (IPCS 2005).
Carbosulfan, as other carbamates, is extremely toxic to mammals and its toxicity
is mediated through inhibition of AChE in neural tissue, resulting in the
accumulation of ACh in the synaptic cleft. Subsequently, the overstimulation of
ACh receptors leads to neurotoxic symptoms (Renzi & Krieger 1986). The
genotoxicity of carbosulfan was investigated in a wide range of tests. Primarily
negative results were obtained, while positive effects were observed in a few tests.
Carbosulfan has been reported to induce micronucleus formation, sister chromatid

48
exchange and chromosomal aberrations in human peripheral blood lymphocytes
and bone marrow cells of rats (Giri et al. 2003).
There are reports about the metabolism of carbosulfan in the environment
(Chaudhry et al. 2002, de Melo Plese et al. 2005) and in plants (Soler et al. 2006).
In mammals three primary metabolites, 3-hydroxycarbofuran, 3-keto-7-
phenolcarbofuran, and dibutylamine, were detected by TLC in rat in vivo
(Marsden et al. 1982). Moreover, carbofuran and polysulfide derivatives of
carbosulfan were detected in rat stomach by TLC (Umetsu & Fukuto 1982). In
male and female rats in vivo, ten metabolites were identified by TLC and HPLC
and major metabolites were confirmed by GC-MS (FAO/WHO 2003). We have
recently described the in vitro metabolic pathways of carbosulfan in microsomal
hepatic preparations from seven mammalian species including humans. The
primary metabolic pathways in these in vitro studies were the initial oxidation of
sulfur to carbosulfan sulfinamide (‘sulfur oxidation pathway’) and the cleavage of
the nitrogen sulfur bond (N-S) to give carbofuran and dibutylamine (‘carbofuran
pathway’). Carbofuran was oxidized to 3-hydroxycarbofuran and/or 7-
phenolcarbofuran, which were further oxidized to 3-ketocarbofuran or 3-hydroxy-
7-phenolcarbofuran, respectively, and finally to 3-keto-7-phenolcarbofuran
(Abass et al. 2009). It has been shown that, carbosulfan is metabolized in the
environment, plants, and mammals to carbofuran, 3-hydroxycarbofuran, and 3-
ketocarbofuran. While all are actually toxic compounds, carbofuran is found to be
more persistent and toxic than carbosulfan itself. Carbosulfan (LD50 250 and 185
mg kg−1 for male and female rats, respectively) is transformed into a more toxic
one, carbofuran (LD50 8 mgkg−1 for male and female rats), (Iesce et al. 2006,
Tomlin 2000, Trevisan et al. 2004).

49
3 Aims of the present study
Since human data are needed to improve human risk assessment, performing
investigations with human-derived in vitro model systems to predict metabolic
fate and toxicokinetics are of utmost importance. The general aim of this study
was to elucidate the metabolism and interactions of pesticides in animal and
human hepatic preparations. The study focused on characterization of the
metabolic factors of four pesticides, metalaxyl, profenofos, diuron, and
carbosulfan, in in vitro model systems. The specific goals were:
1. To identify and quantify pesticide metabolites formed in vitro by liver
preparations from humans and other mammalian species.
2. To identify the human CYP enzymes responsible for the metabolism by c-
DNA expressed human CYPs and other relevant approaches.
3. To quantify pesticide metabolism in vitro by individual human liver
microsomes.
4. To elucidate the inhibitory interactions with major human liver CYPs.
5. Overall improvement of the process of human risk assessment

50

51
4 Materials and Methods
4.1 Chemicals
Carbosulfan, carbofuran, 3-hydroxycarbofuran, 3-ketocarbofuran, 3-keto-7-
phenolcarbofuran, 3-hydroxy-7-phenolcarbofuran, 7-phenolcarbofuran 3,4-
dichloroaniline, bis(p-chlorophenyl)urea and 3-(4-chlorophenyl) methyl urea
were purchased from ChemService (West Chester, PA). Dibutylamine was
purchased from Sigma-Aldrich (Heidenheim, Germany). Diuron, N-
demethyldiuron, and N-dimethyldiuron were purchased from Dr. Ehrenstorfer
(Augsburg, Germany). Metalaxyl, profenofos and carbaryl were a kind gift from
Agrochem (Alexandria, Egypt). Midazolam was a kind gift from F. Hoffman-La
Roche (Basel, Switzerland), and omeprazole was a gift from Astra Zeneca
(Mölndal, Sweden). HPLC-grade solvents were obtained from Rathburn
(Walkerburn, UK) and Labscan (Dublin, Ireland). All other chemicals used were
from the Sigma Chemical Company (St. Louis, MO) and were of the highest
purity available. Water was freshly prepared in-house with the Simplicity 185
(Millipore S.A., Molsheim, France) water purification system and was UP-grade
(ultra pure, 18.2 MΩ).
4.2 Human and animal liver samples
Human liver samples used in this study were obtained from the University
Hospital of Oulu as surplus from organ donors. The collection of surplus tissue
was approved by the Ethics Committee of the Medical Faculty of the University
of Oulu, Finland. All liver samples were of Caucasian race including 4 female and
6 male between the ages of 21 and 62. Intracerebral hemorrhage was the primary
cause of death. Characteristics of the liver samples are presented in the original
publication (II). The livers were transferred to ice immediately after the surgical
excision and cut into pieces, snap-frozen in liquid nitrogen, and stored at –80° C.
Male DBA/2 mouse, Sprague-Dawley rat, Beagle dog, Cynomolgus monkey,
Göttingen minipig, and New Zealand white rabbit liver samples were left-over
tissue from untreated male animals used as controls in other studies. The Animal
Use and Care Committee of original sites of research had approved the
experimental design of primary studies. All microsomes were prepared by
standard differential ultracentrifugation (Pelkonen et al. 1974).

52
4.3 Recombinant enzymes
Baculovirus insect cell-expressed human CYPs (CYP1A1, 1A2, 1B1, 2A6, 2B6,
2C8, 2C9, 2C18, 2C19, 2D6, 2E1, 3A4, 3A5, 3A7 and 4A11) were purchased
from BD Biosciences Discovery Labware (Bedford, MA) and were used
according to the manufacturer’s instructions.
4.4 In vitro assay of metabolites and analytical methods
The standard incubation mixture contained serial concentrations of the studied
compound; microsomal protein and 1 mM NADPH in a final volume of 200 µl
phosphate buffer (pH 7.4). The compound was first dissolved in DMSO. After a 2
min incubation at +37 °C in a shaking incubator block, the reaction was started by
adding NADPH. The mixture was incubated at +37 °C for different time points
and the reaction was stopped with ice cold acetonitrile. After centrifugation at
10000 × g for 15 min, the supernatant was collected and stored at -20 °C until
analyzed. To measure the production of potential metabolites with human liver
homogenates, incubations were prepared containing serial concentrations of the
studied compound. In addition to human liver homogenates, the other components
in the homogenations incubate were 5 mM UDPGA, 1.2 mM PAPS, 1 mM GSH
and 1 mM NADPH in a final volume of 200 µl phosphate buffer (pH 7.4). The
incubations and analytical method were similar to the microsomal incubations. To
measure the main metabolites of recombinantly expressed CYP enzymes, the
standard incubation mixture contained phosphate buffer (pH 7.4), NADPH,
recombinantly expressed CYP enzymes and the studied compound. Incubations
were carried out according to the manufacturer’s instructions.
Chromatographic separation of metabolites was carried out with the Waters
Alliance 2690 HPLC system (Waters Corp., Milford, MA). The column used was
Waters XTerra MS C18 (2.1 mm x 50 mm, particle size 3.5 µm) (I, II, III) and
Waters Atlantis T3 (2.1 mm x 100 mm, particle size of 3 µm) (IV, V) together
with a Phenomenex C18 2.0 mm x 4.0 mm precolumn (Phenomenex, Torrance,
CA). The initial screening of the compounds present and accurate mass
measurements was carried out using a Micromass LCT (Micromass, Altrincham,
UK) time of flight (TOF) mass spectrometer equipped with a Z-Spray ionization
source. The mass spectrometer and HPLC system were operated under
Micromass MassLynx 3.4 software. The quantification (multiple reaction
monitoring, MRM) and fragmentation measurements were performed with a

53
Micromass Quattro II triple quadrupole instrument equipped with a Z-spray
ionization source.
The lower limit of quantification was 0.1 µM for profenofos and diuron, 0.5
µM for carbosulfan and its metabolites, and 1.0 µM for metalaxyl and 2,6-
dimethylaniline.
4.5 Determination of enzyme kinetic parameters
To measure the enzyme kinetic parameters in both the microsomal samples and
recombinantly expressed CYP enzymes, the standard incubation mixture
contained a series of concentrations of the studied compound. Incubation
mixtures and methods were the same as mentioned above. Samples were analyzed
by LC-MS-MS. The kinetic parameters Vmax and Km were calculated using Prism
5.0 (GraphPad Software, Inc., San Diego, CA) by nonlinear regression. These
values were used to calculate the intrinsic clearance value (Vmax/Km) and to
compare the efficiency of catalysis between species.
4.6 Correlation analysis
Correlations were performed between metabolite formations and the activities of
each CYP model across the human liver bank. Model substrate reactions used for
correlations were the same as those used in the inhibition studies below. The
software program Prism 5.0 (GraphPad Software, Inc., San Diego, CA) was used
for data analysis (II, III, IV, and V).
4.7 Enzymatic and inhibition assays
The incubation and analysis conditions used in this work have been presented in
detail in the original publication (III).
4.8 In vitro - in vivo extrapolations
In vivo hepatic clearance for the metabolism of profenofos, diuron and
carbosulfan were extrapolated from in vitro data obtained from the incubation
with various liver microsomes. In vivo extrapolations were made based on actual
measurements of microsomal protein per gram of liver weight (MPPGL) and
other published values (Boxenbaum 1980, Pelkonen & Turpeinen 2007) (Table 4).

54
Table 4. Average values of body weights, liver weights, hepatic blood flows and
MPPGL.
Species Human Dog Rabbit Rat Mouse Minipig Monkey
Body weight (kg) 70 17 2.9 0.24 0.03 19.4 4.12
liver weight (gm) 1500 480.2 131.5 10.1 1.54 382a 157
liver blood flow QH (L/min) 1.45 0.68 0.12 0.02 0.003 0.85a 0.25
MPPGLb 18.2 24.5 19.5 23.4 20.6 22 12.1 a Estimated values based on pig values b Experimentally determined values
In addition, in vivo hepatic clearance for the metabolism of carbosulfan was
extrapolated from experimentally determined in vitro intrinsic clearance (CLLM)
and actual measurements of microsomal protein per gram of liver weight
(MPPGL) (Table 5).
Table 5. Experimentally determined MPPGL in ten donors
Individual HLM20 HLM21 HLM22 HLM23 HLM24 HLM28 HLM29 HLM30 HLM31 HLM32
MPPGL 19.6 16.3 17.1 16.1 27.2 26.0 17.6 11.8 20.7 9.3
To estimate the hepatic clearance CLH of study compounds, in vitro metabolic
clearance (intrinsic clearance, CLint, LM) was calculated on the basis of liver
microsomal incubations. The reported pharmacokinetic parameters were
calculated as following:
– In vitro intrinsic clearance
CLint, LM [ml/min/mg protein] = Vmax/Km
– Hepatic intrinsic clearance
CLint, whole liver [ml/min] = CLint, LM [ml/min/mg protein]*MPPGL*liver weight
– Hepatic clearance
CLH [L/min] = CLint, whole liver [L/min] * QH / (CLint, whole liver [L/min] + QH)

55
5 Results
5.1 Metabolism of pesticides in mammalian hepatic preparations
5.1.1 Identification of metabolites produced in vitro by mammalian hepatic preparations
Incubations of pooled human liver homogenates and microsomes with various
concentrations of the studied compounds were analyzed by LC-TOF-MS.
Metalaxyl (I) metabolites were tentatively identified on the basis of exact mass
determined by TOF-MS, since the identification of the metabolites from
fragmentation data was not possible due to a lack of reference data. The
metabolites detected were two hydroxymetalaxyl derivatives or atropisomers of
one of the derivatives, demethyl and didemethylmetalaxyl, and a lactone
metabolite, while the suspected carcinogenic metabolite, 2,6-dimethylaniline was
not found. Detection limit was 1.0 µM.
In the case of profenofos, (II), two metabolites, namely
desthiopropylprofenofos and hydroxyprofenofos, were identified from the
extracted mass chromatograms and fragmentation data. The hydroxyprofenofos
fragmentation pattern illustrated that in the first fragmentation step ethanol is
released and in the second step the hydroxypropyl group is released. According to
this pattern it is evident that the hydroxylation has to be in the S-propyl moiety.
Because none of the metabolites were available, they were quantitated using the
calibration curve of profenofos, assuming the signals of the parent and the
metabolites were approximately equal. External standards were measured in the
beginning, middle and end of the experiment to ensure quality of analysis.
Diuron (III) and carbosulfan (IV, V) metabolites were detected from the
extracted mass chromatograms and in general identified with the help of reference
standards. N-demethyldiuron was the only metabolite detected in diuron
incubations and no other potential metabolites were observed even with the help
of the standards. Of eight detected carbosulfan metabolites, seven were identified
with the help of the reference standard. These metabolites were carbofuran, 3-
hydroxycarbofuran, 3-hydroxy-7- phenolcarbofuran, 3-ketocarbofuran, 3-keto-7-
phenolcarbofuran, 7-phenolcarbofuran and dibutylamine. We have assumed that
the unidentified metabolite is carbosulfan sulfinamide based on published reports
(FAO/WHO 2003, Marsden et al. 1982).

56
5.1.2 Studies of in vitro metabolism by mammalian hepatic preparations
Metabolite formations as a function of incubation time were studied using human
and mammalian hepatic fractions. Only human data are reported for metalaxyl,
whereas comparison with other species is done for other pesticides. In most cases,
the amounts of metabolites were increased as a function of incubation time in
correspondence to the disappearance of the parent compounds. Higher amounts of
metabolites were observed in human hepatic homogenates than in microsomes
probably due to the fact that the homogenates contained higher amounts of
microsomal protein than microsomal preparations when added to the incubation
mixtures, even if the same amount of liver tissue was as a starting material. It is
also possible that the homogenate provides more optimal conditions for the
metabolism of selected pesticides than the microsomal preparation.
Metalaxyl (I) incubations indicated that the hydroxymetalaxyl metabolites
were major products, whereas moderate levels of didemethylmetalaxyl and minor
levels of demethylmetalaxyl and lactone metabolite were produced. However, the
didemethylation step was more active in the homogenates than in microsomal
incubations.
The bioactivation of profenofos (II) via CYP-mediated desthiopropylation
was the predominant pathway. Minor amounts of hydroxyprofenofos were found
in all incubations.
Diuron N-demethylathion (III) was the only metabolic pathway detected in
human and seven mammalian hepatic microsomes. It is of interest to note that a
very fast disappearance of both diuron and its metabolite was detected in dog
liver microsomes at the lower diuron concentrations.
3-hydroxycarbofuran was the main carbosulfan (IV,V) metabolite in all
species and individuals, but otherwise there were some qualitative interspecies
differences in carbofuran pathway metabolites. Only rabbit liver microsomes
were able to metabolize carbofuran via hydroxylation to 7-phenolcarbofuran.
Carbofuran was not detected in dog liver microsomes due to the rapid further
metabolism. In general, liver microsomes from all seven species and individuals
produced more toxic products (carbofuran, 3-hydroxy-carbofuran, 3-
ketocarbofuran) more rapidly than the detoxification product (carbosulfan
sulfinamide).

57
5.1.3 Interspecies differences in in vitro metabolism
The metabolic pathways of the three substrates (II, III, and IV) were very similar
in all species, with a similar qualitative, but not quantitative, agreement in the
types of metabolites produced, with the exception of a more distal carbosulfan
metabolite, 7-phenolcarbofuran. On the basis of the data obtained, it is apparent
that there are species specific differences in the ratios and rates of metabolites
formed from the various substrates. Differences in kinetics between the lowest
and highest species are shown in Table 6.
Table 6. The largest differences in kinetic parameters for the formation of metabolites
of profenofos, diuron and carbosulfan between different mammalian liver
microsomesa.
Metabolite Km Vmax CLint, LM
Profenofos
Desthiopropylprofenofos 2.6 6.9 4.3
Hydroxyprofenofos 2.5 3.1 9.0
Diuron
N-demethyldiuron 4.4 5.5 5.4
Carbosulfan
Carbofuran metabolic pathway 5.5 5.1 2.1
Carbosulfan sulfinamide 3.5 4.6 4.3
Dibutylamine 7.7 4.4 6.3 a Differences between the maximum and minimum observed values.
Biotransformation of these pesticides in mammalian liver microsomes occurred
via NADPH-dependent metabolic pathways. There is a relatively moderate
variability (in the level of CLint, LM values) based on animal to human efficiency
comparisons. It is, however, necessary to emphasize that maximum differences
between the lowest and highest rates were not conspicuously large. These
findings illustrate that in vitro screening of metabolite profiles and metabolic
activities for the proper prediction of animal model studies is desirable since
differences in metabolism and disposition could affect the accuracy of the
extrapolation approach.
5.1.4 Interindividual variability in in vitro metabolism
The interindividual variability in the percentage of metabolite formation rates for
the tested compounds is shown in Figure 8. In the present work, a large degree of

58
interindividual variability in the rates of metabolite production (2.6- to 19.6-fold)
was observed as differences between the maximum and minimum observed
values in ten donors. The up to 19-fold interindividual variability could be a
significant determinant in toxicity. There could be substantial differences in toxic
metabolite formation even if different individuals are exposed to the same
quantity of compound. It is an established fact that CYP activities display a large
interindividual variation (Pelkonen et al. 2008). The relatively large
interindividual variability in metabolite formation seems to be due to variable
levels of catalytic activities of CYP enzymes (see below for further discussion).
The uncertainty factor for in vitro human variability in toxicokinetics was up
to 3-fold, as defined by Renwick and Lazarus (Renwick & Lazarus 1998), as a
variation between the mean and the highest value. Thus, a tentative suggestion is
that interindividual variability in toxicokinetics of the studied three pesticides,
even if restricted here to in vitro hepatic metabolism and not to the overall
toxicokinetics, might be within the standard applied factor for interindividual
variability in toxicokinetics.
Fig. 8. The interindividual variability for the percentage of metabolites formation
obtained with individual human liver microsomes. A bank of 10 livers was used to
assess the metabolite formation in individual livers. The values represented are the
minimum, 25th percentile, median, 75th percentile, and the maximum.

59
5.2 Identification of CYP isoenzymes metabolizing the studied pesticides in human liver in vitro
5.2.1 Kinetic analysis of recombinant CYP enzymes
Detailed kinetic analyses were performed based on preliminary screening assays
to determine which enzymes were responsible for the formation of various
metabolites of the studied pesticides. According to the obtained kinetic values
CYP3A4, CYP2B6 and CYP2C19 were identified as profenofos-oxidizing
enzymes (II). The rank order of CYPs in profenofos desthiopropylation was
CYP3A4>CYP2B6>CYP2C19, whereas it was the opposite for profenofos
hydroxylation. All tested recombinant CYPs catalyzed diuron N-demethylation
(III) and the highest activities were possessed by CYP1A1, CYP1A2, CYP2C19,
and CYP2D6. CYP3A5 and CYP2B6 had the greatest efficiency to form
carbosulfan sulfinamide (V), while CYP3A4 and CYP3A5 were the most
efficient in the generation of the carbofuran metabolic pathway. Precise
calculation of the enzyme kinetic parameters for metalaxyl (I) metabolite
formation by recombinant CYPs was not possible. Much higher concentrations of
metalaxyl in the incubation mixture would have been needed for reliable
estimation, but this was not possible because of dissolution and precipitation
problems at higher metalaxyl concentrations. Only CYP3A4 was responsible for
metalaxyl hydroxylation, while CYP2B6 was the major isoform responsible for
(di)demethylation and lactone formation.
5.2.2 Analysis of the relative contributions of different CYP enzymes
The importance of individual isoforms in in vivo clearance is dependent upon the
relative abundance of each isoform. Thus, intrinsic clearance values measured
with individual cDNA-expressed enzymes were scaled to liver microsomes by
employing human hepatic microsomal P450 isoenzyme concentrations based on
the average human hepatic microsomal protein amounts of the main P450s
(Rostami-Hodjegan & Tucker 2007). The contribution of CYP3A4 to profenofos
(II) desthiopropylation and to both of the carbosulfan (V) metabolic pathways
was the highest because of both high intrinsic clearance of the pathway and
average amount of the isoenzyme. CYP2C19 was estimated to contribute 55% to
profenofos hydroxylation, whereas CYP1A2 contributes 60% to diuron N-
demethylation (III) (Figure 9).

60
Fig. 9. The relative contribution of each CYP isoform based on the actual intrinsic
clearance values and the average human hepatic microsomal protein amounts of
P450s.
5.2.3 Relationship between CYP activities and metabolite formation
A correlation analysis was performed between the formation of metabolites and
each CYP activity across the human liver bank. High and significant correlations
between either desthiopropylprofenofos (II) or carbofuran metabolic pathway (V)
and CYP marker activities were seen with CYP3A4 and CYP2C19, whereas a
high correlation was observed between N-demethyldiuron formation (III) and
CYP1A2 marker activity (Figure 10). No correlations were observed between the

61
production of carbosulfan sulfinamide or hydroxyprofenofos metabolites and the
specific activities of P450 isoform-catalyzed reactions. However, exclusion of
two potential outliers resulted in relatively weak correlations except for
carbofuran metabolic pathway and CYP3A4, omeprazole sulfoxidation, (R2 =
0.81). The reason for the potential outliers comes from individuals HLM22 and
HLM28 who had the highest level of both CYP3A4 and CYP2C19.
Fig. 10. Correlation of the metabolite formation rates of profenofos, diuron and
carbosulfan with CYP-selective probe drug activities measured in a panel of ten
human liver microsomes. CYP marker activities (range of values and mean ± SD), on
the X axis, are expressed as pmol/(mg protein*min) and metabolite formation rates are
on the Y axis.

62
5.2.4 Inhibition of in vitro metabolism of pesticides by CYP-selective inhibitors (V)
The inhibitory effects of known CYP isoform-selective inhibitors on metabolites
formation was studied only with carbosulfan (V). Ketoconazole, a CYP3A4-
selective inhibitor, inhibited formation rates of the carbofuran pathway and to a
lesser extent carbosulfan sulfinamide formation. The results suggest that CYP3A
subfamily member has a major role in carbosulfan metabolism (Figure 11).
Fig. 11. Effect of the CYP3A isoform-selective inhibitor, ketoconazole, on the
carbofuran metabolic pathway and carbosulfan sulfinamide formation by HLM.
5.3 Evaluation of the CYP inhibition potential of the studied pesticides in pooled human hepatic microsomes
The ability of the studied pesticides to inhibit selective model activities for all
major CYPs was studied. It has to be stressed that the evaluation does not
consider the mechanisms of inhibition. However, there were quite considerable
differences in the potency and extent of inhibition between tested pesticides.
Generally profenofos (II) and diuron (III) were the most potent and extensive
inhibitors especially towards CYP1A1/2 (IC50 = 2.9 and 4.1 μM, respectively).
Metalaxyl (I) inhibits CYP2B6 moderately with an IC50 value of 41.7 μM.
Carbosulfan (V) inhibits CYP3A4 moderately with IC50s of 11.2 and 23.8 μM for
midazolam 1’-hydroxylation and omeprazole sulfoxidation, respectively, whereas
even higher values of 58.3 and 61.9 μM were observed for CYP1A1/2 and
CYP2C19, respectively (Figure 12).

63
Fig. 12. Observed IC50 values of tested pesticides on CYP-model activities in pooled
human hepatic microsomes.

64
5.4 In vitro - in vivo extrapolation
The data we have presented show both differences and similarities in the in vitro
metabolism of pesticides in liver microsomes of human and experimental animals,
and this may be important in extrapolating data from in vitro to in vivo.
Pharmacokinetic variables for the metabolism of selected pesticides in the human
and animal body were extrapolated from in vitro data. The in vitro intrinsic
clearance (CLint, LM) was determined experimentally and the value obtained used
to estimate hepatic clearances (CLH) per kg body weight (Pelkonen & Turpeinen
2007). The calculated in vivo hepatic clearances based on in vitro intrinsic
clearances (CLint, LM) were considerably different in different species (Table 7) but
not individuals (Table 8).
Table 7. Calculated in vivo hepatic clearance of study compounds extrapolated from
incubation in human and mammalian liver microsomes.
Pharmacokinetic parameters HLM DogLM RabLM RatLM MouseLM PigLM MonLM
Desthiopropylprofenofos
CLint, LM [ml/min/mg protein] 27.9 - - 8.8 37.5 - -
CLint, whole liver [ml/min] 760 - - 2.07 1.19 - -
CLH [L/min] 1.447 - - 0.017 0.003 - -
CLH / kg B.W [ml/min/kg] 20.7 70.3 85.8
Hydroxyprofenofos
CLint, LM [ml/min/mg protein] 0.3 - - 1.9 2.7 - -
CLint, whole liver [ml/min] 8.18 - - 0.45 0.09 - -
CLH [L/min] 1.232 - - 0.016 0.003 - -
CLH / kg B.W [ml/min/kg] 17.6 - - 68.2 83.5 - -
N-demethyldiuron
CLint, LM [ml/min/mg protein] 0.17 0.40 0.31 0.08 0.21 0.16 0.33
CLint, whole liver [ml/min] 4.75 4.71 0.81 0.09 0.01 1.34 0.62
CLH [L/min] 1.111 0.591 0.106 0.009 0.002 0.520 0.178
CLH / kg B.W [ml/min/kg] 15.9 34.8 37.2 36.0 62.1 26.8 43.3
Carbofuran metabolic pathway
CLint, LM [ml/min/mg protein] 0.45 0.22 0.34 0.33 0.25 0.47 0.45
CLint, whole liver [ml/min] 12.4 2.63 0.86 0.08 0.01 3.96 0.86
CLH [L/min] 1.298 0.538 0.107 0.014 0.002 0.699 0.194
CLH / kg B.W [ml/min/kg] 18.5 31.6 37.5 58.0 64.9 36.0 47.0
Carbosulfan sulfinamide
CLint, LM [ml/min/mg protein] 0.02 0.05 0.04 0.01 0.03 0.03 0.03
CLint, whole liver [ml/min] 0.510 0.601 0.098 0.003 0.001 0.212 0.052
CLH [L/min] 0.377 0.318 0.054 0.002 0.001 0.170 0.043

65
Pharmacokinetic parameters HLM DogLM RabLM RatLM MouseLM PigLM MonLM
CLH / kg B.W [ml/min/kg] 5.4 18.7 19.1 10.0 24.6 8.7 10.4
Dibutylamine
CLint, LM [ml/min/mg protein] 0.063 0.087 0.018 0.115 0.080 0.057 0.047
CLint, whole liver [ml/min] 1.723 1.018 0.047 0.027 0.003 0.483 0.090
CLH [L/min] 0.787 0.406 0.034 0.010 0.001 0.308 0.066
CLH / kg B.W [ml/min/kg] 11.2 23.9 11.9 43.4 42.4 15.9 16.1
Table 8. Calculated in vivo hepatic clearance of the carbosulfan active moiety, the
carbofuran metabolic pathway, extrapolated from incubation in ten donor liver
microsomes.
Pharmacokinetic
parameters HLM20 HLM21 HLM22 HLM23 HLM24 HLM28 HLM29 HLM30 HLM31 HLM32
CLint, LM [ml/min/mg
protein] 0.391 0.264 0.365 0.381 0.671 0.420 0.394 0.473 0.101 0.354
CLint, whole liver [ml/min] 11.51 6.46 9.35 9.18 27.39 16.40 10.42 8.36 3.14 4.91
CLH [L/min] 1.288 1.184 1.255 1.252 1.377 1.332 1.273 1.236 0.992 1.119
CLH / kg B.W
[ml/min/kg] 18.40 16.92 17.93 17.89 19.67 19.03 18.18 17.65 14.18 15.99

66

67
6 Discussion
6.1 Methodological aspects of the in vitro studies
Risk assessment of chemicals needs reliable scientific information, and one
source of information is the characterization of metabolic fate and toxicokinetics.
Pesticides were used as representative xenobiotics in this study since exposure to
pesticides is a global challenge to risk assessment. Because humans cannot be
used as study subjects except in occasional circumstances, we have to rely upon
in vitro model systems and animal studies. In vitro metabolism and interactions of
pesticides by human and animal hepatic preparations together with the advantages
of novel analytical techniques (LC/TOF-MS and LC/MS-MS) for metabolite
identifications and quantification provide information for the prediction of in vivo
pharmacokinetics of selected pesticides.
Mass spectrometry is the preferred technique because of its good sensitivity.
Mass spectrometry is also a very robust technique and typically only protein
precipitation is needed as a sample preparation method before analysis. LC/TOF-
MS is of use in the initial screening of the compounds’ present and accurate mass
measurements because of its ability to detect all ionized compounds
simultaneously. Another benefit of TOF-MS is its ability to provide accurate mass
measurements, which helps in identification by providing the elemental
composition of the metabolites. LC/MS-MS is of use in the quantification and
fragmentation measurements because of its excellent selectivity and sensitivity.
In most studies, human liver homogenates and microsomes were used as a
pool of ten individual donors. These pooled preparations provide average
metabolic activities. Moreover, the metabolism has been studied in different
individuals in order to illustrate the interindividual variability. It has to be stressed
here that although we measured only the in vitro hepatic metabolism of selected
pesticides, metabolism is usually the most important factor contributing to
interindividual and interspecies differences in toxicokinetics.
In vitro-in vivo extrapolations are considered a relatively reliable and useful
tool for predicting pharmacokinetics. Simple models for in vitro-in vivo
extrapolation of hepatic clearance have been presented previously (see (Pelkonen
& Turpeinen 2007)) and used for the prediction of in vivo pharmacokinetics of the
selected pesticides and consequently interspecies comparison in this study. In vitro-in vivo extrapolations were made based on the actual measurements of

68
microsomal protein content in liver samples (MPPGL) from humans and animals.
The reason for the use of actual values and not the consensus values is that
reliable consensus values are available only for human and perhaps rat liver
(Barter et al. 2007). Average values of liver weights and hepatic blood flows in
various mammalian species were obtained from the published literature
(Boxenbaum 1980, Pelkonen & Turpeinen 2007).
6.2 Differences in pesticide metabolites formation
6.2.1 In vitro vs in vivo qualitative differences
Metalaxyl metabolites were identified in the light of published reports
(FAO/WHO 2002) because of metabolite fragmentation before the analyzer and a
lack of reference data. The biotransformation reactions were hydroxylation,
(di)demethylation and lactone formation. Metalaxyl principal metabolites
detected in vitro in human liver preparations were the same as in in vivo
metabolism in rats (FAO/WHO 2002). Quantitative differences are probably due
to well-recognized potential differences regarding species and in vitro-in vivo
differences.
Profenofos metabolites were analyzed by LC-TOF-MS and metabolites were
identified from the extracted mass chromatograms in our in vitro studies. A high
concentration of desthiopropylprofenofos, via profenofos oxidative desulfuration,
and a small amount of hydroxyprofenofos were obtained. The corresponding
oxidative desulfuration metabolic pathway has been detected as a major pathway
in the case of phosphorothiolate compounds (Kono et al. 1983, Lee et al. 2006b,
Wing et al. 1983). In a fatal human poisoning case, profenofos metabolites were
detected by GC-FPD in the acid extracts of blood and urine after methylation with
diazomethane. Desthiopropylprofenofos was detected as a major metabolite,
while the minor metabolite, hydroxyprofenofos, was not detected in vivo (Gotoh et al. 2001). Since hydroxyprofenofos was detected only in in vitro studies as a
minor metabolite (Abass et al. 2007a, Wing et al. 1983), the in vitro – in vivo
qualitative difference could be partly explained by further metabolism in humans
in vivo keeping in mind that the lapse of time after death was estimated to be 27
hours at the time of autopsy and the profenofos dose was very high.
Diuron demethylation at the terminal nitrogen atom was the only metabolic
pathway detected in human liver homogenates and seven mammalian liver

69
microsomes including human. In vitro-in vivo qualitative differences were
observed. Diuron has been reported to be metabolized in vivo in rats, dogs (Hodge et al. 1967), and humans (Van Boven et al. 1990, Verheij et al. 1989) via
demethylation, didemethylation, and hydroxylation. It is possible that other
potential metabolic pathways reside in extrahepatic tissues or need much longer
incubation times or different incubation conditions than those we used with
hepatic preparations. Also, diuron doses in human poisoning cases, in which other
human metabolites were observed, were very high. A rough estimate of the total
concentration of diuron plus metabolites in plasma was found to be near 100
mg/L, resulting in an estimated intake of at least several grams (Verheij et al. 1989). Moreover, in the electrospray ionization of the MS analysis, all of the
compound would not necessarily ionize, and consequently, some metabolites
would not be detected.
From eight carbosulfan metabolites detected in vitro in human hepatic
preparations, seven were identified. These metabolites were detected in male and
female rats in vivo and by TLC and HPLC, with the exception of carbosulfan
sulfinamide. Carbosulfan sulfone was detected instead of carbosulfan sulfinamide
in addition to the carbofuran and polysulfide derivatives of carbosulfan
(FAO/WHO 2003, Marsden et al. 1982, Umetsu & Fukuto 1982). Most
metabolites that had been identified in vivo in rat were also present in our in vitro
study with hepatic microsomes. Concerning some discrepancies, the
sulfate/glucuronide conjugation was not detected in vitro. However, it should be
kept in mind that the metabolism in rats in vivo was determined after repeated oral
doses for 14 days followed by 14C carbosulfan single oral dose on day 15 whereas
the maximum incubation period in vitro was 60 min.
Qualitative interspecies differences were observed in the carbofuran distal
metabolite, 7-phenolcarbofuran. 7-Phenolcarbofuran was a rabbit-specific
metabolite, not detected in any other tested species including rat liver microsomes.
However, 7-phenolcarbofuran has been detected as a carbosulfan metabolite in vivo in rat urine and feces and in goat’s milk, liver, and kidney (FAO/WHO 2003).
The probable reason could be the short duration of in vitro incubations, which did
not allow enough time for the formation of minor metabolites.
In vitro metabolism study results were generally in agreement with the results
obtained from the in vivo studies, at least for the active chemical moiety and
major metabolites. With respect to minor metabolites, qualitative differences
could be explained by different incubation conditions and well-recognized
potential in vitro – in vivo differences.

70
6.2.2 Qualitative interspecies differences in the in vitro metabolism of profenofos, diuron and carbosulfan
It has been repeatedly observed that differences in metabolism, both qualitative
and quantitative, between different species even in the case of closely related
species can be considerable (Turpeinen et al. 2007). In our studies, no qualitative
interspecies differences were observed in the in vitro metabolism of profenofos.
The diuron N-demethylation pathway was observed and quantified in all tested
liver preparations. On the other hand, further metabolism could have happened in
mouse, dog, monkey liver microsomes and human liver homogenates, although
we did not detect any metabolites, since the concentrations of diuron and its N-
demethylation metabolite were decreasing especially at the lowest diuron
concentration. Despite extensive screening of the potential metabolites by TOF-
LC-MS, we did not detect any other metabolites. Potential metabolites should be
detected since they contain nitrogen and therefore should be easy to detect with
electrospray ionization, but of course there may be some exceptions. It has to be
mentioned that, in electrospray ionization, all of the compound would not
necessarily ionize, and, consequently, these metabolites would not be detected.
Qualitative interspecies differences were observed in the carbosulfan in vitro
metabolism profile. 7-Phenolcarbofuran was detected only in rabbit liver
microsomes. 3-Keto-7-phenolcarbofuran was not detected in rat, while 3-
hydroxy-7-phenolcarbofuran was not detected in either rat or mouse liver
microsomes. Carbofuran was not detected in dog liver microsomes due to
extensive metabolism into more distal metabolites. Qualitative differences in the
more distal metabolic pathway might be due to slight differences in the
expression or the active site of the enzymes responsible for these reactions,
keeping in mind that in the standard experimental conditions used for carbosulfan
metabolites, reaction rates were linear at least up to 0.15 mg of microsomal
protein/ml and 60 min of incubation time.
6.2.3 Quantitative interspecies differences in the in vitro metabolism of profenofos, diuron and carbosulfan
Animal to human quantitative differences based on intrinsic clearance values are
collected in Table 9. MouseLM catalyzed profenofos desthiopropylation and
hydroxylation more efficiently than human. The rapid disappearances of both
diuron and N-demethyldiuron, at least in dog, could be partially explained by the

71
formation of undetectable metabolites. DogLM was also the most active
mammalian microsomes in metabolite formation over the time and concentration
used. Also the kinetics data showed that DogLM had the highest capacity and
efficiency for metabolite formation. DogLM metabolized diuron 2.3-fold more
than HLM, followed by MonLM and RabLM, whereas RatLM had the smallest
value (0.43-fold). In the case of carbosulfan, there is a relatively small variability
in the level of species differences. MinipigLM, monkeyLM, and humanLM
metabolized carbosulfan to carbofuran almost equally, while dogLM and
mouseLM had the smallest values. On the other hand, dogLM was the most
efficient in carbosulfan sulfinamide and dibutylamine formation (2.7- and 1.4-
fold to human, respectively). Differences in efficiencies between rat, minipig, and
human liver microsomes in the metabolism of triazine herbicides such as atrazine
and ametryn have been reported by Lang et al. (Lang et al. 1996). Also, with
other pesticides such as chlorpyrifos (organophosphorus insecticide) and
carbofuran (carbamate insecticide), human liver microsomes showed lower rates
of metabolism than mouse and rat liver microsomes (Tang et al. 2001, Usmani et al. 2004a).
Profenofos was more readily activated than detoxified by human, mouse and
rat hepatic microsomes, as demonstrated by the greater CLint, LM rates for
desthiopropylation compared with the CLint, LM rates for hydroxylation. The ratios
between activation/detoxification intrinsic clearance values for profenofos were
81.9 for human, followed by 13.9 for mouse, and 4.7 for rat liver microsomes.
A similar trend was observed for the biotransformation of carbosulfan by
several mammalian hepatic microsomes including human. The
bioactivation/detoxification ratios (CLint, LM carbofuran metabolic pathway / CLint, LM carbosulfan
sulfonamide) were rat (27.4) and human (24.4) followed by minipig (18.7) and
monkey liver microsomes (16.5). Rabbit (8.8), mouse (7.7) and dog (4.4) liver
microsomes were proportionally the least active in carbosulfan activation.
It is apparent that there are considerable differences in the quantities of the
metabolites produced within the species, as well as in the ratios of the metabolites
among the species. In general, the types of metabolites formed are identical in all
species, and there is no metabolite produced only by one species, except the more
distal carbosulfan metabolite 7-phenolcarbofuran that was rabbitt-specific.
Therefore, the enzymes of these species responsible for the metabolism are
similar in that they catalyze the same types of reactions. Nevertheless, these
enzymes are to some extent different in their selectivity and efficiency, as is
apparent from the characteristic metabolic pattern of each species. These

72
differences could be related to the variability in the constitutive expressions of
CYP enzymes in the liver of investigated animal species (Turpeinen et al. 2007).
These findings illustrate that in vitro screening of qualitative and quantitative
differences are needed to provide a firm basis for interspecies and in vitro- in vivo
extrapolations.
Table 9. Animal to human fold differences based on in vitro intrinsic clearance values
(CLint, LM).
Metabolite HLM DogLM RabLM RatLM MOUSELM PigLM MonLM
Profenofos
Desthiopropylprofoenofos 1 - - 0.32 1.34 - -
Hydroxyprofenofos 1 - - 6.30 9.00 - -
Diuron
N-demethyldiuron 1 2.30 1.80 0.43 1.23 0.92 1.88
Carbosulfan
Carbofuran metabolic pathway 1 0.49 0.74 0.72 0.56 1.03 0.99
Carbosulfan sulfinamide 1 2.74 2.04 0.64 1.76 1.35 1.46
Dibutylamine 1 1.37 0.29 1.81 1.27 0.91 0.75
6.2.4 Interindividual variability in in vitro metabolism
A bank of ten individual human liver microsomal samples was used to
characterize CYP enzymes activities. The enzyme activities were determined with
appropriate marker substrates and experimental conditions (see the materials and
methods section and Table 10). There are large interindividual differences in the
expression levels and catalytic activities of CYP enzymes in humans, and these
variations sometimes lead to different susceptibilities of humans to the
pharmacological and toxicological actions of drugs, toxic chemicals, and
carcinogens (Pelkonen et al. 2008).
To study interindividual variability in the in vitro metabolism of profenofos
and diuron, metabolite formation rates were measured in hepatic microsomes
from ten donors. Up to 7.8-fold interindividual variability in the rate of
profenofos activation and 19.6-fold for diuron activation, as determined by the
difference between the lowest and highest value, were detected. Interindividual
variability in the biotransformation kinetics for carbosulfan was investigated in
hepatic microsomes from ten donors for the carbofuran metabolic pathway, since
the active chemical moiety is the most relevant for chemical risk assessment.
Human liver microsomes display quite large interindividual differences in all

73
carbosulfan metabolic pathways. Differences between the maximum and
minimum observed values in ten individual liver microsomes indicated 5.9-, 7.0-,
and 6.6-fold variability in the km, Vmax and CLint, LM values for the carbofuran
metabolic pathway, respectively. The up to 7-fold interindividual variability in the
rate of carbosulfan activation, as determined by the difference between the lowest
and highest kinetic values, could be the significant determinant of the toxicity
effects. There will be substantial differences in toxic metabolite formation even if
different individuals are exposed to the same quantity of carbosulfan. High
metabolizers might be at higher risk for carbosulfan-related toxicity.
Table 10. Characterizations of CYP activities in hepatic microsomes from ten donors.
Enzyme Substrate Activity
Values range
pmol/(mg
protein*min)
Mean ± S.D Fold
variation
CYP1A1/2 7-Ethoxyresorufin O-deethylation 3.8–31.7 50.0 ± 31.7 8.3
CYP2A6 Coumarin 7-hydroxylation 95.8–549.2 281.6 ± 167.3 5.7
CYP2B6 Bupropion Hydroxylation 4.5–220.5 73.1 ± 61.9 49
CYP2C8 Amodiaquine N-deethylation 152.6–362.4 238.8 ± 65.4 2.4
CYP2C9 Tolbutamide Methylhydroxylation 79.8–269.3 148.6 ± 55.9 3.4
CYP2C19 Omeprazole 5-hydroxylation 21.5–124.1 55.8 ± 35.8 5.8
CYP2D6 Dextromethorphan O-demethylation 67.9–229.6 156.9 ± 58.9 3.4
CYP2E1 Chlorzoxazone 6-hydroxylation 136.0–773.5 397.4 ± 202.2 5.7
CYP3A4 Midazolam 1`-Hydroxylation 511.6–4568 1499 ± 1336 8.9
CYP3A4 Omeprazole Sulfoxidation 12.9–130.5 49.6 ± 40.8 10.1
There are large interindividual differences in the expression levels and catalytic
activities of CYP enzymes in humans, and these variations sometimes lead to
different susceptibilities of humans to the pharmacological and toxicological
actions of drugs, toxic chemicals, and carcinogens (Pelkonen et al. 2008).
Considerable interindividual variability was evident in the metabolism of the
tested compound as ten individuals progressed through the entire study.
Individuals HLM22 and HLM28 with the highest level of both CYP3A4 and
CYP2C19 exhibited the highest desthiopropylprofenofos and carbofuran
metabolic pathway formation rates. Individual HLM28 exhibited the highest N-
demethyldiuron formation corresponding to highest levels of both CYP1A2 and
CYP2C19, and the reverse was true for individual HLM31, who had the lowest
levels of both isoforms.

74
6.3 Identification of the P450 isoform(s) involved in metabolism
Recently, a number of papers have been published on the activity of human P450s
involved in the metabolism of pesticides (see the literature review) and these
activities may result in activation and/or detoxification reactions. Therefore, in
order to improve risk assessment of chemicals, by improving the interpretation of
toxicological studies in experimental animals and interpretation of
epidemiological studies in exposed individuals, it is essential to identify the CYP
enzymes involved in the metabolism of a given pesticide. The identification of
CYPs gives essential knowledge about the potential interactions with other
chemicals and drugs that are metabolized by the same isoforms, the
interindividual variability and the predicting of the in vivo situation based on its
relative contribution.
Different established approaches were used to identify the CYP isoform(s)
involved in the metabolism of the investigated pesticides using human liver
microsomes and cDNA-expressed P450 isoforms. CYP1A2, CYP2B6, CYP2C19,
and CYP3A4 catalyze the metabolism of the investigated pesticides. In general,
CYP3A4 play a crucial role and CYP2B6 a lesser role in the metabolism of
metalaxyl, profenofos and carbosulfan. CYP1A2, CYP2C19 and to lesser extent
CYP3A4 were the most important CYPs catalyzing diuron metabolism.
In the inhibition study of in vitro metabolism of carbosulfan by ketoconazole,
a relatively high concentration of 100 µM was used. Although ketoconazole is a
selective inhibitor of CYP3A4 at 1 μM concentration (Baldwin et al. 1995,
Newton et al. 1995), there is a risk of having some loss of specificity at a higher
concentration. However, on the basis of comprehensive evidence it seems that
CYP3A4 is the most significant CYP enzyme in carbosulfan metabolism. The
role of other members of the CYP3A subfamily, CYP3A5, and CYP3A7, is
probably negligible. Because CYP3A5 exists in only 10–20% of Caucasians
(Rodrigues et al. 1999) and the amount of CYP3A7 is very low in adult human
livers and is mainly expressed in embryonic, fetal, and newborn livers, where it is
the predominant CYP form (Hakkola et al. 2001, Kitamura et al. 1992). Thus,
CYP3A4 is probably the most important CYP enzyme (Liu et al. 2007, Pelkonen
et al. 2008, Rendic & Di Carlo 1997, Riley et al. 2001) and it has been shown that
it has a prominent role in pesticide metabolism, as demonstrated here for
carbosulfan, metalaxyl and profenofos and furthermore it is involved in the
metabolism of diuron.

75
The known pesticides metabolized mainly by CYP3A4 belong to several
chemical groups due to its very large and flexible active site, allowing it to
oxidize either large substrates or multiple smaller ligands (Scott & Halpert 2005,
Tang & Stearns 2001). Phosphorothioate, carbamate, chlorinated cyclodiene,
neonicotinoid, and phenyl pyrazole insecticides; conazoles, acylalanine and other
fungicides; and chloroacetamide and phenyl urea herbicides are mainly
metabolized by CYP3A4. CYP2B6 can catalyze the oxidation of a number of
structurally diverse classes of xenobiotics, many of which have been described in
detail in reviews by Ekins & Wrighton 1999, Hodgson & Rose 2007b, Turpeinen et al. 2006b. CYP2B6 plays a major role in pesticide metabolism such as
chloroacetamide herbicides, phosphorothioate and carbamate insecticides, and
acylaniline fungicides. Similarly, xenobiotics mainly metabolized by CYP2C19
and CYP1A2 belong to several chemical group and we did not find a correlation
between a specific CYP and specific pesticide chemical group. Based on the
literature survey, the involvement of human recombinant CYPs for metabolism of
pesticides was 24%, 15%, 12% and 12% for CYP3A4, CYP2C19, CYP2B6 and
CYP1A2, respectively (see the literature review and Figure 4).
Data have been presented for the biotransformation of three structurally
different compounds in hepatic microsomes from ten donors. Differences have
been demonstrated in the profile of CYP enzymes catalyzing the bioactivation of
these compounds. Knowledge of the enzymes responsible for the
biotransformation can contribute to the risk assessment of these compounds. In
particular, an understanding of the preference of individual enzymes to
bioactivate or detoxify these compounds can help our knowledge of how
susceptibility to these compounds may be affected by differential expression of
CYPs. Consequently, the activity of CYP3A4 and CYP2C19 may be important in
determining the toxicity arising from exposure to profenofos and carbosulfan.
Thus, an individual with high CYP3A4 and CYP2C19 activities (e.g, ultra-rapid
metabolizers (UMs)) might be particularly susceptible to the toxic effects of this
compound. Similarly, individuals with high CYP1A2 and CYP2C19 activities,
UMs, might be more susceptible to diuron toxicity.
6.4 CYP-pesticides inhibition potential
Human hepatic CYP-associated marker activities were inhibited to a variable
extent by the tested pesticides. Generally, CYPs were more sensitive to
organophosphorus insecticides, e.g. profenofos, than to other pesticide groups. In

76
a screening study to elucidate the ability of 18 pesticides to inhibit selective
model activities for all major CYPs, organophosphorus compounds were the most
potent and extensive inhibitors especially towards CYP1A1/2, CYP2B6, CYP2C8,
CYP2C9, CYP2D6 and CYP3A4. Otherwise there were quite considerable
differences in potency and extent of inhibition between other pesticide groups
(Abass et al. 2009).
In the study of the inhibitory potential of carbosulfan similar inhibition
constants were obtained with both cocktail and single enzyme assays for CYP1A2,
CYP2C19, and CYP3A4. Moderate inhibition of CYP2C9 and CYP2D6 was
observed only with the cocktail assay. In the cocktail assay, possibility of
interference between probe substrates has been reported, for example,
amodiaquine and bupropion have moderate inhibitory potencies towards CYP2D6
(Bapiro et al. 2001, Hesse et al. 2000).
In vivo concentrations of pesticides under different exposure conditions
(occupational, dietary) would be helpful in interpreting in vitro studies, but are
not usually available and not easily calculable. For example, the International
Estimated Dietary Intakes (IEDIs) of profenofos were calculated for the 13
GEMS/Food cluster diets (Food Global Environment Monitoring System–Food
Contamination Monitoring and Assessment Programme). The calculated IEDIs
were 3 µg/kg b.w, representing 1–10% of the maximum ADI (0.03 mg/kg bw)
(FAO/WHO, 2007). Profenofos was the most potent and extensive inhibitor
especially towards CYP1A1/2 and CYP2A6 with IC50 ~ 3 µM, representing 0.11
mg/ml. Because appropriate pharmacokinetic parameters, i.e. volume of
distribution etc, are not known, it is difficult to extrapolate either from in vivo
exposure "down" or from in vitro concentration "up". However, it seems unlikely
that profenofos would cause interactions after dietary exposures, whereas
inhibitory interactions could become relevantafter intense occupational exposures.
6.5 Quantitative prediction of in vivo metabolic clearance and the calculation of CSAFs for the studied compounds.
6.5.1 Interspecies differences and the calculation of AKAF
In the chemical risk assessment, traditional default values are used to account for
interspecies differences and for human variability. Modern approaches aim to
replace the default factors by data-derived values. The basic physiological

77
parameters, such as body weight, liver weight and liver blood flow, vary between
laboratory animal species and humans. There are large differences in
physiological parameters in mice compared to humans (Boxenbaum 1980,
Calabrese et al. 1992). This is the reason why hepatic clearances, CLH, per kg
body weight were used for interspecies comparison in toxicokinetics. In these
calculations CLH, per kg body weight in experimental animals, for the active
chemical moiety, were divided by the clearance in humans (Table 11). The highest
AKAF are 4.2 for dethiopropylprofenofos, 3.9 for N-demethyldiuron and 3.5 for
the carbofuran metabolic pathway, i.e., there would be higher concentrations in
humans than in animals, mouse in this case, for the same dose per kg body weight
because of the lower clearance in humans. It must be stressed here that we
measured only the hepatic metabolism, assuming that there is a negligable
contribution of extrahepatic metabolism; however, metabolism is often the most
important factor contributing to interspecies differences in toxicokinetics.
Table 11. Calculated AKAF based on in vivo hepatic clearance (CLH) for the active
chemical moieties of the study compounds.
Pharmacokinetic parameters HLM DogLM RabLM RatLM MouseLM PigLM MonLM
Desthiopropylprofenofos
CLH / kg B.W [ml/min/kg] 20.7 70.3 85.8
AKUF 3.4 4.2
N-demethyldiuron
CLH / kg B.W [ml/min/kg] 15.9 34.8 37.2 36.0 62.1 26.8 43.3
AKUF 2.2 2.3 2.3 3.9 1.7 2.7
Carbofuran metabolic pathway
CLH / kg B.W [ml/min/kg] 18.5 31.6 37.5 58.0 64.9 36.0 47.0
AKUF 1.7 2.0 3.1 3.5 1.9 2.5
Overall, the results of the present study have demonstrated that the in vitro
metabolic pattern of human microsomes resembled that of minipig and to a lesser
extent that of monkey liver microsomes. It is, therefore, reasonable to assume that
the minipig hepatic microsomes are qualitatively and quantitatively suitable as in
vitro model for exploring and predicting human in vivo metabolism of pesticides,
and the metabolic similarity must be taken into consideration in toxicological
research on carbamates and organophosphates pesticides in humans. Indeed, the
metabolism plays a critical role in the toxicity of carbamate and organophosphate
pesticides toxicities (Buratti et al. 2005, Buratti & Testai 2007, Leoni et al. 2008,
Mutch et al. 2003, Mutch & Williams 2006, Tang et al. 2001, Tang et al. 2002).

78
The concept of CSAFs has been introduced to provide a method for the
incorporation of quantitative data on interspecies differences or human variability
in either toxicokinetics or dynamics into the risk assessment procedure. The AKAF
should be used for the development of the CSAF of studied pesticides rather than
the default values because they are based on quantitative chemical-specific data.
It has to be stressed here that the prediction of in vivo metabolic clearance
and the development of CSAFs for the studied compound is restricted to in vitro
hepatic metabolic characteristics, and all other contributing factors such as the
contribution of extrahepatic metabolism should be either assumed or studied
separately.
6.5.2 Interindividual variability and the calculation of the HKAF factor for carbosulfan
Hepatic clearances were calculated for carbosulfan in ten donors used for HKAF
extrapolation. Since interindividual variability addresses the potential differences
in the response of a sensitive individual compared to an average individual, the
HKAF is 1.1 as the variation between the mean and the highest value as defined by
Renwick and Lazarus (Renwick & Lazarus 1998). Our studies with carbosulfan
suggested that in vivo extrapolated interindividual variability in toxicokinetics is
within the standard applied factors for interindividual extrapolation in
toxicokinetics restricted to in vitro hepatic metabolism in the Caucasian race.
6.5.3 Calculation of the composite uncertainty factor (CUF), carbosulfan as an example
The use of chemical-specific data has the effect of replacing a default factor for
an area of uncertainty with chemical specific data, thereby reducing the overall
uncertainty. CUF is the composite value obtained on multiplying the CSAF, used
to replace a default subfactor, by the remaining default subfactors for which
suitable data were not available. The CUF could be either greater than or less than
the usual default, 100, depending on the quantitative scientific data that have been
introduced to reduce the default uncertainty factor (WHO/IPCS, 2005). The CUF
is generally shown as:
CUF = [AKAF or AKUF] × [ADAF or ADUF] × [HKAF or HKUF] × [HDAF or HDUF].
In the case of carbosulfan, the CUF is a composite of adjustment factors for
interspecies differences and human variability in toxicokinetics and uncertainty

79
factors for interspecies differences and human variability in toxicodynamics. CUF
= AKAF × ADUF × HKAF × HDUF, Therefore, CUF = 3.5 × 2.5 × 1.1 × 3.16 = 30.
On the basis of Cmax, the uncertainty factor for some pesticides has been
changed. FAO/WHO (2002) established an ARfD for the carbamate insecticide,
carbaryl, of 0.2 mg/kg bw based on a NOAEL of 3.8 mg/kg bw per day identified
on the basis of inhibition of cholinesterase activity observed at 10 mg/kg bw per
day in a 5-week dietary study in dogs with the application of a 25-fold CUF. On
the other hand, in the Australian evaluation of carbaryl, the current Australian
ARfD of 0.01 mg/kg bw is based on a NOEL of 1 mg/kg bw per day identified on
the basis of behavioural indications of autonomic neurotoxicity, and inhibition of
brain and plasma erythrocyte cholinesterase activity in a study of developmental
toxicity and a 13-week study of neurotoxicity in rats, and using a 100-fold safety
factor (FAO/WHO 2005).
FAO/WHO (2002, 2008, 2009) has established safety factors of 50 for the
organophosphate insecticide acephate and 25 for triazole fungicide fenpyroximate;
pyrethroid insecticide cyfluthrin, beta-cyfluthrin and lambada cyhalothrin; and
conazole fungicides triadimenol and triadimefon. On the other hand, higher safety
factor was established for other pesticides, e.g. pyrazole acaricide fenpyroximate,
safety factor of 200.
FAO/WHO (2002, 2008), established an ARfD of 0.009 mg/kg bw for
carbofuran, the active chemical moiety in carbosulfan metabolic pathway, on the
basis of the NOAEL of 0.22 mg/kg bw per day with the application of a CUF of
25. This, together with our CUF value of 30 provides strong support for the use of
In vitro chemical-specific quantitative toxicokinetics data and quantitative
prediction of in vivo metabolic clearance for the calculation of the composite
uncertainty factor (CUF) for xenobiotics for the overall improvement of the
process of human risk assessment.
Finally when estimating the composite uncertainty factor from in vitro-in
vivo extrapolation data, one should keep in mind some limitations. Results
obtained from in vitro test systems are highly dependent on several technical
factors including the microsomal protein amount, incubation time, and initial
velocity conditions used in the test system etc (Pelkonen et al. 2008). The best
predictive value is usually obtained when the substrate concentration used is
within the linear part of the time and protein concentration curves for substrate
depletion or metabolite formation (Lin & Lu 1998, Mäenpää et al. 1998, Yuan et al. 1999) (cited from Pelkonen et al. 2008). The prediction of in vivo metabolic
clearance is based on many scaling factors and physiological measures, which

80
should either be assumed or measured (Pelkonen & Turpeinen 2007). These
include for example microsomal yield (if microsomes are used), liver weight,
liver blood flow, extra-hepatic metabolism and clearances via other routes than
hepatic metabolism. However, in the context of this study it is of importance to
note that although the measurements are restricted to in vitro hepatic metabolism
of selected pesticides, metabolism is probably in these cases the most important
factor contributing to interindividual and interspecies differences in toxicokinetics.

81
7 Conclusions
1. Metalaxyl was metabolized in human hepatic microsomes mainly via
hydroxylation, demethylation and to a lesser extent via lactone formation.
Diuron was metabolized by N-demethylation. Profenofos and carbosulfan
were mainly activated to desthiopropylprofenofos and carbofuran,
respectively, in human and mammalian hepatic preparations.
2. In general, the types of metabolites formed are identical in all species, except
for the more distal carbosulfan metabolites. Considerable differences in the
quantities of the metabolites produced within the species, as well as in the
ratios of the metabolites among the species, were observed.
3. CYP1A2, CYP2B6, CYP2C19, and CYP3A4 in particular catalyze the
metabolism of the investigated pesticides. In general, CYP3A4 plays a crucial,
and CYP2B6 a lesser, role in the metabolism of metalaxyl, profenofos, and
carbosulfan. CYP1A2, CYP2C19 and, to a lesser extent CYP3A4, were the
most important CYPs catalyzing diuron metabolism.
4. Individuals with high CYP3A4 and CYP2C19 activities might be particularly
susceptible to the toxic effects of profenofos and carbosulfan, while
individuals with high CYP1A2 and CYP2C19 activities might be more
susceptible to diuron toxicity.
5. Profenofos and diuron were potent inhibitors of CYP1A1/2. Metalaxyl
inhibited CYP2B6 and carbosulfan moderately inhibited CYP3A4,
CYP1A1/2, and CYP2C19.
6. Based on in vivo hepatic clearance, the highest AKAF were 4.2 for
dethiopropylprofenofos, 3.9 for N-demethyldiuron, and 3.5 for the carbofuran
metabolic pathway and the HKAF was 1.1 for carbofuran metabolic pathway.
The AKAF and HKAF, restricted to metabolism studies, should be used for the
development of the CSAF of studied pesticides rather than the default values
because they are based on quantitative chemical-specific data.
7. The information provided is important in that in enables a more rational,
data-based evaluation of species and interindividual differences in the
toxicokinetics of pesticides. This information is of value in refining the risk
assessment of the pesticides in question.

82

83
References
Abass K, Reponen P & Pelkonen O (2009) Metabolic and interactions properties of selected fungicides. In De Costa P & Bezerra P (eds) Fungicides: chemistry, environmental impact and health effects. New York, Nova Science Publisher: 25–62.
Abass K, Reponen P, Jalonen J & Pelkonen O (2007a) In vitro metabolism and interaction of profenofos by human, mouse and rat liver preparations. Pestic Biochem Physiol 87: 238–247.
Abass K, Reponen P, Jalonen J & Pelkonen O (2007b) In vitro metabolism and interactions of the fungicide metalaxyl in human liver preparations. Environ Toxicol Pharmacol 23(1): 39–47.
Abass K, Reponen P, Mattila S & Pelkonen O (2009) Metabolism of carbosulfan. I. Species differences in the in vitro biotransformation by mammalian hepatic microsomes including human. Chem Biol Interact 181(2): 210–219.
Abass K, Reponen P, Mattila S & Pelkonen O (2010) Metabolism of carbosulfan II. Human interindividual variability in its in vitro hepatic biotransformation and the identification of the cytochrome P450 isoforms involved. Chem Biol Interact 185(3): 163–173.
Abass K, Reponen P, Turpeinen M, Jalonen J & Pelkonen O (2007) Characterization of diuron N-demethylation by mammalian hepatic microsomes and cDNA-expressed human cytochrome P450 enzymes. Drug Metab Dispos 35(9): 1634–1641.
Abass K, Turpeinen M & Pelkonen O (2009) An evaluation of the cytochrome P450 inhibition potential of selected pesticides in human hepatic microsomes. J Environ Sci Health, Part B: Pestic, Food Contam , Agric Wastes 44(6): 553.
Adams DR, Jones AM, Plopper CG, Serabjit-Singh CJ & Philpot RM (1991) Distribution of cytochrome P-450 monoxygenase enzymes in the nasal mucosa of hamster and rat. Am J Anat 190(3): 291–298.
Agrawal RC, Kumar S & Mehrotra NK (1996) Micronucleus induction by diuron in mouse bone marrow. Toxicol Lett 89(1): 1–4.
Ahokas J & Pelkonen O (2007) Pharmacokinetics: How Does The Body Handle Drugs? In UNESCO (ed) Pharmacology, Encyclopedia of Life Support Systems (EOLSS). Oxford, UK, UNESCO Publishing-Eolss Publishers.
Al, Omari, A., & Murry D, J. (2007) Pharmacogenetics of the cytochrome P450 enzyme system: review of current knowledge and clinical significance. J Pharm Pract 20(3): 206–218.
Alavanja MCR, Hoppin JA & Kamel F (2004) Health effects of chronic pesticide exposure: cancer and neurotoxicity. Annu Rev Public Health 25(1): 155–197.
Allen DD, Caviedes R, Cárdenas AM, Shimahara T, Segura-Aguilar J & Caviedes PA (2005) Cell lines as in vitro models for drug screening and toxicity studies. Drug Dev Ind Pharm 31(8): 757.
American Medical Association (1997) Educational and informational strategies to reduce pesticide risks. Prev Med 26(2): 191–200.

84
Aninat C, Piton A, Glaise D, Le Charpentier T, Langouet S, Morel F, Guguen-Guillouzo C & Guillouzo A (2006) Expression of cytochromes P450, conjugating enzymes and nuclear receptors in human hepatoma HepaRG cells. Drug Metab Dispos 34(1): 75–83.
Anthérieu S, Chesné C, Li R, Camus S, Lahoz A, Picazo L, Turpeinen M, Tolonen A, Uusitalo J, Guguen-Guillouzo C & Guillouzo A (2010) Stable expression, activity, and inducibility of cytochromes P450 in differentiated HepaRG cells. Drug Metab Dispos 38(3): 516–525.
Asseffa A, Smith SJ, Nagata K, Gillette J, Gelboin HV & Gonzalez FJ (1989) Novel exogenous heme-dependent expression of mammalian cytochrome P450 using baculovirus. Arch Biochem Biophys 274(2): 481–490.
Bajpai M, Roskos L, Shen D & Levy R (1996) Roles of cytochrome P4502C9 and cytochrome P4502C19 in the stereoselective metabolism of phenytoin to its major metabolite. Drug Metab Dispos 24(12): 1401–1403.
Baldwin SJ, Bloomer JC, Smith GJ, Ayrton AD, Clarke SE & Chenery RJ (1995) Ketoconazole and sulphaphenazole as the respective selective inhibitors of P4503A and 2C9. Xenobiotica 25(3): 261–270.
Bapiro TE, Egnell A, Hasler JA & Masimirembwa CM (2001) Application of higher throughput screening (HTS) inhibition assays to evaluate the interaction of antiparasitic drugs with cytochrome P450s. Drug Metab Dispos 29(1): 30–35.
Barter ZE, Bayliss MK, Beaune PH, Boobis AR, Carlile DJ, Edwards RJ, Houston JB, Lake BG, Lipscomb JC, Pelkonen OR, Tucke GT & Rostami-Hodjegan A (2007) Scaling factors for the extrapolation of in vivo metabolic drug clearance from in vitro data: reaching a consensus on values of human micro-somal protein and hepatocellularity per gram of liver. Curr Drug Metab 8(1): 33–45.
Barton HA, Tang J, Sey YM, Stanko JP, Murrell RN, Rockett JC & Dix DJ (2006) Metabolism of myclobutanil and triadimefon by human and rat cytochrome P450 enzymes and liver microsomes. Xenobiotica 36(9): 793–806.
Beaune PH, Kremers PG, Kaminsky LS, De Graeve J, Albert A & Guengerich FP (1986) Comparison of monooxygenase activities and cytochrome P-450 isozyme concentrations in human liver microsomes. Drug Metab Dispos 14(4): 437–442.
Bergh AF & Strobel HW (1992) Reconstitution of the brain mixed function oxidase system: Purification of NADPH-cytochrome P450 reductase and partial purification of cytochrome P450 from whole rat brain. J Neurochem 59(2): 575–581.
Berthou F, Dreano Y, Belloc C, Kangas L, Gautier J & Beaune P (1994) Involvement of cytochrome P450 3A enzyme family in the major metabolic pathways of toremifene in human liver microsomes. Biochem Pharmacol 47(10): 1883–1895.
Bertz RJ & Granneman GR (1997) Use of in Vitro and in vivo data to estimate the likelihood of metabolic pharmacokinetic interactions. Clin Pharmacokinet 32(3): 210–258.
Bigwood EJ (1973) The acceptable daily intake of food additives. CRC Crit Rev Toxicol 2: 41–93.
Blanchoud H, Farrugia F & Mouchel JM (2004) Pesticide uses and transfers in urbanised catchments. Chemosphere 55(6): 905–913.

85
Boobis AR, Ossendorp BC, Banasiak U, Hamey PY, Sebestyen I & Moretto A (2008) Cumulative risk assessment of pesticide residues in food. Toxicol Lett 180(2): 137–150.
Bourrie M, Meunier V, Berger Y & Fabre G (1996) Cytochrome P450 isoform inhibitors as a tool for the investigation of metabolic reactions catalyzed by human liver microsomes. J Pharmacol Exp Ther 277(1): 321–332.
Boxenbaum H (1980) Interspecies variation in liver weight, hepatic blood flow, and antipyrine intrinsic clearance: Extrapolation of data to benzodiazepines and phenytoin. J Pharmacokinet Pharmacodyn 8(2): 165–176.
Brandon EFA, Raap CD, Meijerman I, Beijnen JH & Schellens JHM (2003) An update on in vitro test methods in human hepatic drug biotransformation research: pros and cons. Toxicol Appl Pharmacol 189(3): 233–246.
Brian Houston J (1994) Utility of in vitro drug metabolism data in predicting in vivo metabolic clearance. Biochem Pharmacol 47(9): 1469–1479.
Brockmöller J, Kirchheiner J, Meisel C & Roots I (2000) Pharmacogenetic diagnostics of cytochrome P450 polymorphisms in clinical drug development and in drug treatment. Pharmacogenomics 1(2): 125–151.
Broly F, Libersa C, L’Hermitte M, Becthel P & Dupuis B (1989) Effect of quinidine on the dextromethorphan O-demethylase activity of microsomal fractions from human liver. Br J Clin Pharmacol 28: 29–36.
Buckpitt A, Boland B, Isbell M, Morin D, Shultz M, Baldwin R, Chan K, Karlsson A, Lin C, Taff A, West J, Fanucchi M, Van Winkle L & Plopper C (2002) Naphthalene-induced respiratory tract toxicity: metabolic mechanisms of toxicity. Drug Metab Rev 34(4): 791–820.
Bull S, Fletcher K, Boobis AR & Battershill JM (2006) Evidence for genotoxicity of pesticides in pesticide applicators: a review. Mutagenesis 21(2): 93–103.
Buratti FM, D'Aniello A, Volpe MT, Meneguz A & Testai E (2005) Malathion bioactivation in the human liver liver: The contribution of different cytochrome P450 isoform. Drug Metab Dispos 33(3): 295–302.
Buratti FM, Leoni C & Testai E (2006) Foetal and adult human CYP3A isoforms in the bioactivation of organophosphorothionate insecticides. Toxicol Lett 167(3): 245–255.
Buratti FM & Testai E (2007) Evidences for CYP3A4 autoactivation in the desulfuration of dimethoate by the human liver. Toxicology 241(1-2): 33–46.
Buratti FM, Volpe MT, Fabrizi L, Meneguz A, Vittozzi L & Testai E (2002) Kinetic parameters of OPT pesticide desulfuration by c-DNA expressed human CYPs. Environ Toxicol Pharmacol 11(3-4): 181–190.
Buratti FM, Volpe MT, Meneguz A, Vittozzi L & Testai E (2003) CYP-specific bioactivation of four organophosphorothioate pesticides by human liver microsomes. Toxicol Appl Pharmacol 186(3): 143–154.
Burke MD, Prough RA & Mayer RT (1977) Characteristics of a microsomal cytochrome P-448-mediated reaction. Ethoxyresorufin O-de-ethylation. Drug Metab Dispos 5(1): 1–8.

86
Butler AM & Murray M (1997) Biotransformation of parathion in human liver: Participation of CYP3A4 and its inactivation during microsomal parathion oxidation. J Pharmacol Exp Ther 280: 966–973.
Butler MA, Iwasaki M, Guengerich FP & Kadlubar FF (1989) Human cytochrome P-450PA (P-450IA2), the phenacetin O-deethylase, is primarily responsible for the hepatic 3-demethylation of caffeine and N-oxidation of carcinogenic arylamines. Proc Natl Acad Sci U S A 86(20): 7696–7700.
Cai X, Wang RW, Edom RW, Evans DC, Shou M, Rodrigues AD, Liu W, Dean DC & Baillie TA (2004) Validation of (-)-N-3-benzyl-phenobarbital as a selective inhibitor of CYP2C19 in human liver microsomes. Drug Metab Dispos 32(6): 584–586.
Calabrese EJ, Beck BD & Chappell WR (1992) Does the animal-to-human uncertainty factor incorporate interspecies differences in surface area? Regul Toxicol Pharm 15(2 I): 172–179.
Carlile DJ, Hakooz N & Houston JB (1999) Kinetics of drug metabolism in rat liver slices: IV. Comparison of ethoxycoumarin clearance by liver slices, isolated hepatocytes, and hepatic microsomes from rats pretreated with known modifiers of cytochrome P-450 activity. Drug Metab Dispos 27(4): 526–532.
Casabar RCT, Wallace AD, Hodgson E & Rose RL (2006) Metabolism of endosulfan-alpha by human liver microsomes and its utility as a simultaneous in vitro probe for CYP2B6 and CYP3A4. Drug Metab Dispos 34(10): 1779–1785.
Chang TKH, Weber GF, Crespi CL & Waxman DJ (1993) Differential activation of cyclophosphamide and ifosphamide by cytochromes P-450 2B and 3A in human liver microsomes. Cancer Res 53(23): 5629–5637.
Chaudhry GR, Mateen A, Kaskar B, Sardessai M, Bloda M, Bhatti AR & Walia SK (2002) Induction of carbofuran oxidation to 4-hydroxycarbofuran by Pseudomonas sp. 50432. FEMS Microbiol Ecol 214(2): 171–176.
Chen Y, Ferguson SS, Negishi M & Goldstein JA (2004) Induction of human CYP2C9 by rifampicin, hyperforin, and phenobarbital is mediated by the pregnane X receptor. J Pharmacol Exp Ther 308(2): 495–501.
Christopher Gorski J, Hall SD, Jones DR & VandenBranden M (1994) Regioselective biotransformation of midazolam by members of the human cytochrome P450 3A (CYP3A) subfamily. Biochem Pharmacol 47(9): 1643–1653.
Clarke SE, Ayrton AD & Chenery RJ (1994) Characterization of the inhibition of P4501A2 by furafylline. Xenobiotica 24(6): 517.
Code EL, Crespi CL, Penman BW, Gonzalez FJ, Chang TKH & Waxman DJ (1997) Human cytochrome P4502B6. interindividual hepatic expression, substrate specificity, and role in procarcinogen activation. Drug Metab Dispos 25(8): 985–993.

87
Coecke S, Ahr H, Blaauboer BJ, Bremer S, Casati S, Castell J, Combes R, Corvi R, Crespi CL, Cunningham ML, Elaut G, Eletti B, Freidig A, Gennari A, Ghersi-Egea JF, Guillouzo A, Hartung T, Hoet P, Ingelman-Sundberg M, Munn S, Janssens W, Ladstetter B, Leahy D, Long A, Meneguz A, Monshouwer M, Morath S, Nagelkerke F, Pelkonen O, Ponti J, Prieto P, Richert L, Sabbioni E, Schaack B, Steiling W, Testai E, Vericat JA & Worth A (2006) Metabolism: a bottle neck in in vitro toxicological test development. The report and recommendations of ECVAM workshop 54. Altern Lab Anim 34: 49–84.
Coleman S, Linderman R, Hodgson E & Rose RL (2000) Comparative metabolism of chloroacetamide herbicides and selected metabolites in human and rat liver microsomes. Environ Health Perspect 108(12): 1151.
Coutts RT, Bach MV & Baker GB (1997) Metabolism of amitriptyline with CYP2D6 expressed in a human cell line. Xenobiotica 27(1): 33–47.
Crommelin DJA & Sindelar RD (1997) Pharmaceutical biotechnology: an introduction for pharmacists and pharmaceutical scientists. Australia; United Kingdom, Harwood Academic.
D’Esposito F, Tattam BN, Ramzan I & Murray M (2008) A liquid chromatography/electrospray ionization mass spectrometry (LC–MS/MS) assay for the determination of irinotecan (CPT-11) and its two major metabolites in human liver microsomal incubations and human plasma samples. J Chromatogr B 875(2): 522–530.
Daly H, Doyen JT & Purcell AH (eds) (1998) Introduction to insect biology and diversity. New York, Oxford University Press.
de Melo Plese LP, Paraiba LC, Foloni LL & Pimentel Trevizan LR (2005) Kinetics of carbosulfan hydrolysis to carbofuran and the subsequent degradation of this last compound in irrigated rice fields. Chemosphere 60(2): 149–156.
Dhawan A, Parmar D, Das M & Seth PK (1990) Cytochrome P-450 dependent monooxygenases in neuronal and glial cells: Inducibility and specificity. Biochem Biophys Res Commun 170(2): 441–447.
Ding X & Kaminsky LS (2003) Human extrahepatic cytochromes P450: Function in xenobiotic metabolism and tissue-selective chemical toxicity in the respiratory and gastrointestinal tracts. Annu Rev Pharmacol Toxicol 43(1): 149–173.
Doecke DJ, Veronese ME, Pond SM, Miners JO, Birkett DJ, Sansom LM & McManus ME (1991) Relationship between phenytoin and tolbutamide hydroxylations in human liver microsomes. Br J Clin Pharmacol 31: 125–130.
Domanski TL, Finta C, Halpert JR & Zaphiropoulos PG (2001) cDNA cloning and initial characterization of CYP3A43, a novel human cytochrome P450. Mol Pharmacol 59(2): 386–392.
Dorne JLCM, Dorne JLCM, Walton K & Renwick AG (2005) Human variability in xenobiotic metabolism and pathway-related uncertainty factors for chemical risk assessment: a review. Food Chem Toxicol 43(2): 203.
Dorne JLCM, Walton K & Renwick AG (2001a) Human variability in glucuronidation in relation to uncertainty factors for risk assessment. Food Chem Toxicol 39(12): 1153–1173.

88
Dorne JLCM, Walton K & Renwick AG (2001b) Uncertainty factors for chemical risk assessment: human variability in the pharmacokinetics of CYP1A2 probe substrates. Food Chem Toxicol 39(7): 681–696.
Dorne JLCM, Walton K & Renwick AG (2003a) Human variability in CYP3A4 metabolism and CYP3A4-related uncertainty factors for risk assessment. Food Chem Toxicol 41(2): 201–224.
Dorne JLCM, Walton K & Renwick AG (2003b) Polymorphic CYP2C19 and N-acetylation: human variability in kinetics and pathway-related uncertainty factors. Food Chem Toxicol 41(2): 225–245.
Dorne JLCM, Walton K & Renwick AG (2004a) Human variability for metabolic pathways with limited data (CYP2A6, CYP2C9, CYP2E1, ADH, esterases, glycine and sulphate conjugation). Food Chem Toxicol 42(3): 397–421.
Dorne JLCM, Walton K & Renwick AG (2004b) Human variability in the renal elimination of foreign compounds and renal excretion-related uncertainty factors for risk assessment. Food Chem Toxicol 42(2): 275–298.
Dorne JLCM, Walton K, Slob W & Renwick AG (2002) Human variability in polymorphic CYP2D6 metabolism: is the kinetic default uncertainty factor adequate? Food Chem Toxicol 40(11): 1633–1656.
Draper AJ, Madan A & Parkinson A (1997) Inhibition of Coumarin 7-Hydroxylase Activity in Human Liver Microsomes. Arch Biochem Biophys 341(1): 47–61.
Dutcher JS & Boyd MR (1979) Species and strain differences in target organ alkylation and toxicity by 4-ipomeanol predictive value of covalent binding in studies of target organ toxicities by reactive metabolites. Biochemical Pharmacology 28(23): 3367–3372.
Eddershaw PJ & Dickins M (1999) Advances in in vitro drug metabolism screening. Pharm Sci Technol To 2(1): 13–19.
Eddleston M, Worek F, Eyer P, Thiermann H, Von Meyer L, Jeganathan K, Sheriff MHR, Dawson AH & Buckley NA (2009) Poisoning with the S-Alkyl organophosphorus insecticides profenofos and prothiofos. QJM .
Edwards RJ, Adams DA, Watts PS, Davies DS & Boobis AR (1998) Development of a comprehensive panel of antibodies against the major xenobiotic metabolising forms of cytochrome P450 in humans. Biochem Pharmacol 56(3): 377–387.
Ekins S, Mäenpää J & Wrighton SA (1999) In vitro metabolism: subcellular fractions. In Woolf TF (ed) Handbook of drug metabolism. New York, Marcel Dekker: 369–399.
Ekins S, Ring BJ, Grace J, McRobie-Belle DJ & Wrighton SA (2000) Present and future in vitro approaches for drug metabolism. J Pharmacol Toxicol Methods 44(1): 313–324.
Ekins S, VandenBranden M, Ring BJ & Wrighton SA (1997) Examination of purported probes of human CYP2B6. Pharmacogenetics 7(3): 165–179.
Ekins S & Wrighton SA (1999) The role of CYP2B6 in human xenobiotic metabolism. Drug Metab Rev 31(3): 719–754.
Ellenhorns MJ, Schonwald S, Ordog G & Wasserberger J (1997) Ellenhorn’s medical toxicology: diagnosis and treatment of human poisoning. Maryland, Williams & Wilkins.

89
Eriksson C & Brittebo EB (1991) Metabolic activation of the herbicide dichlobenil in the olfactory mucosa of mice and rats. Chem Biol Interact 79(2): 165–177.
European Commission (2001) Decision No. 2455/2001/EC, Directive 2000/60/EC. Facciolá G, Hidestrand M, Von Bahr C & Tybring G (2001) Cytochrome P450 isoforms
involved in melatonin metabolism in human liver microsomes. Eur J Clin Pharmacol 56(12): 881–888.
Falk-Filipsson A, Hanberg A, Victorin K, Warholm M & Wallén M (2007) Assessment factors—Applications in health risk assessment of chemicals. Environ Res 104(1): 108–127.
Fang J, Coutts RT, McKenna KF & Baker GB (1998) Elucidation of individual cytochrome P450 enzymes involved in the metabolism of clozapine. Naunyn Schmiedebergs Arch Pharmacol 358(5): 592–599.
Fang J, Baker GB, Silverstone PH & Coutts RT (1997) Involvement of CYP3A4 and CYP2D6 in the metabolism of haloperidol. Cell Mol Neurobiol 17(2): 227–233.
Fang J, McKay G, Song J, Remillrd A, Li X & Midha K (2001) In vitro characterization of the metabolism of haloperidol using recombinant cytochrome P450 enzymes and human liver microsomes. Drug Metab Dispos 29(12): 1638–1643.
FAO/WHO (2002) Pesticide residues in food — 2002. Report of the joint meeting of the FAO panel of experts on pesticide residues in food and the environment and the WHO core assessment group on pesticide residues, Rome, Italy, 16–25 September 2002. Rome, World Health Organization and Food and Agriculture Organization of the United Nations.
FAO/WHO (2003) Pesticide residues in food — 2003. Report of the joint meeting of the FAO panel of experts on pesticide residues in food and the environment and the WHO core assessment group on pesticide residues, Geneva, Switzerland, 15–24 September 2003. Rome, World Health Organization and Food and Agriculture Organization of the United Nations (FAO Plant Production and Protection Paper, No. 176).
FAO/WHO (2005) Pesticide residues in food — 2005. Report of the joint meeting of the FAO panel of experts on pesticide residues in food and the environment and the WHO core assessment group on pesticide residues, Geneva, Switzerland, 20–29 September 2005. Rome, World Health Organization and Food and Agriculture Organization of the United Nations (FAO Plant Production and Protection Paper, No. 183).
FAO/WHO (2007) Pesticide residues in food — 2007. Report of the joint meeting of the FAO panel of experts on pesticide residues in food and the environment and the WHO core assessment group on pesticide residues, Geneva, Switzerland, 18–27 September 2007. Rome, World Health Organization and Food and Agriculture Organization of the United Nations (FAO Plant Production and Protection Paper, No. 191).
FAO/WHO (2008) Pesticide residues in food — 2008. Report of the joint meeting of the FAO panel of experts on pesticide residues in food and the environment and the WHO core assessment group on pesticide residues, Rome, Italy, 9–18 September 2008. Rome, World Health Organization and Food and Agriculture Organization of the United Nations (FAO Plant Production and Protection Paper, No. 193).

90
FAO/WHO (2009) Pesticide residues in food — 2009. Report of the joint meeting of the FAO panel of experts on pesticide residues in food and the environment and the WHO core assessment group on pesticide residues, Geneva, Switzerland, 16–25 September 2009. Rome, World Health Organization and Food and Agriculture Organization of the United Nations (FAO Plant Production and Protection Paper, No. 196).
Faucette SR, Hawke RL, Lecluyse EL, Shord SS, Yan B, Laethem RM & Lindley CM (2000) Validation of bupropion hydroxylation as a selective marker of human cytochrome P450 2B6 catalytic activity. Drug Metab Dispos 28(10): 1222–1230.
Ferrini J, Pichard L, Domergue J & Maurel P (1997) Long-term primary cultures of adult human hepatocytes. Chem Biol Interact 107(1-2): 31–45.
Field JA, Reed RL, Sawyer TE & Martinez M (1997) Diuron and its metabolites in surface water and ground water by solid phase extraction and in-vial elution. J Agric Food Chem 45(10): 3897–3902.
Fisher CW, Shet MS, Caudle DL, Martin-Wixtrom CA & Estabrook RW (1992) High-level expression in Escherichia coli of enzymatically active fusion proteins containing the domains of mammalian cytochromes P450 and NADPH-P450 reductase flavoprotein. Proc Natl Acad Sci USA 89(22): 10817–10821.
Foxenberg RJ, McGarrigle BP, Knaak JB, Kostyniak PJ & Olson JR (2007) Human hepatic cytochrome P450-specific metabolism of parathion and chlorpyrifos. Drug Metab Dispos 35(2): 189–193.
Fulton MH & Key PB (2001) Acetylcholinesterase inhibition in estuarine fish and invertebrates as an indicator of organophosphorus insecticide exposure and effects. Environ Toxicol Chem 20(1): 37–45.
Furuya H, FernandezSalguero P, Gregory W, Taber H, Steward A, Gonzalez FJ & Idle JR (1995) Genetic polymorphism of CYP2C9 and its effect on warfarin maintenance dose requirement in patients undergoing anticoagulation therapy. Pharmacogenetics 5(6): 389–392.
Galetin A, Brown C, Hallifax D, Ito K & Houston JB (2004) Utility of recombinant enzyme kinetics in prediction of human clearance: impact of variability, CYP3A5, and CYP2C19 on CYP3A4 probe substrates. Drug Metab Dispos 32(12): 1411–1420.
Galijatovic A, Otake Y, Walle UK & Walle T (1999) Extensive metabolism of the flavonoid chrysin by human Caco-2 and Hep G2 cells. Xenobiotica 29(12): 1241–1256.
Gebhardt R, Hengstler J, Müller D, Glöckner R, Buenning P, Laube B, Schmelzer E, Ullrich M, Utesch D, Hewitt N, Ringel M, Hilz B, Bader A, Langsch A, Koose T, Burger H, Maas J & Oesch F (2003) New Hepatocyte In Vitro Systems for Drug Metabolism: Metabolic Capacity and Recommendations for Application in Basic Research and Drug Development, Standard Operation Procedures. Drug Metab Rev 35(2): 145–213.
Giacomazzi S & Cochet N (2004) Environmental impact of diuron transformation: a review. Chemosphere 56(11): 1021–1032.

91
Ginsberg G, Hattis D, Sonawane B, Russ A, Banati P, Kozlak M, Smolenski S & Goble R (2002) Evaluation of child/adult pharmacokinetic differences from a database derived from the therapeutic drug literature. Toxicol Sci 66(2): 185–200.
Giri S, Giri A, Sharma GD & Prasad SB (2003) Induction of sister chromatid exchanges by cypermethrin and carbosulfan in bone marrow cells of mice in vivo. Mutagenesis 18(1): 53–58.
Godin SJ, Crow JA, Scollon EJ, Hughes MF, DeVito MJ & Ross MK (2007) Identification of rat and human cytochrome P450 isoforms and a rat serum esterase that metabolize the pyrethroid insecticides deltamethrin and esfenvalerate. Drug Metab Dispos 35(9): 1664–1671.
Goldstein JA, Faletto MB, Romkes-Sparks M, Sullivan T, Kitareewan S, Raucy JL, Lasker JM & Ghanayem BI (1994) Evidence that CYP2C19 is the major (S)-mephenytoin 4'-hydroxylase in humans. Biochemistry 33(7): 1743–1752.
Goldstein JA (2001) Clinical relevance of genetic polymorphisms in the human CYP2C subfamily. Br J Clin Pharmacol 52(4): 349–355.
Goldstein JA & de Morais SMF (1994) Biochemistry and molecular biology of the human CYP2C subfamily. Pharmacogenetics 4(6): 285–300.
Gomez-Lechon MJ, Donato MT, Castell JV & Jover R (2004) Human hepatocytes in primary culture: the choice to investigate drug metabolism in man. Curr Drug Metab 5: 443–462.
Gomez-Lechon MJ, Donato T, Jover R, Rodriguez C, Ponsoda X, Glaise D, Castell JV & Guguen-Guillouzo C (2001) Expression and induction of a large set of drug-metabolizing enzymes by the highly differentiated human hepatoma cell line BC2. Eu J Biochem 268(5): 1448–1459.
Gonzalez FJ (2005) Role of cytochromes P450 in chemical toxicity and oxidative stress: studies with CYP2E1. Mutat Res 569(1-2): 101–110.
Gooddy DC, Chilton PJ & Harrison I (2002) A field study to assess the degradation and transport of diuron and its metabolites in a calcareous soil. Sci Total Environ 297(1-3): 67–83.
Gotoh M, Sakata M, Endo T, Hayashi H, Seno H & Suzuki O (2001) Profenofos metabolites in human poisoning. Forensic Sci Int 116(2-3): 221–226.
Granfors MT, Backman JT, Neuvonen M, Ahonen J & Neuvonen PJ (2004a) Fluvoxamine drastically increases concentrations and effects of tizanidine: a potentially hazardous interaction. Clin Pharmacol Ther 75: 331–341.
Granfors MT, Backman JT, Laitila J & Neuvonen PJ (2004b) Tizanidine is mainly metabolized by cytochrome P450 1A2 in vitro. Br J Clin Pharmacol 57(3): 349–353.
Granvil CP, Madan A, Sharkawi M, Parkinson A & Wainer IW (1999) Role of CYP2B6 and CYP3A4 in the in vitro N-dechloroethylation of (R)- and (S)-ifosfamide in human liver microsomes. Drug Metab Dispos 27(4): 533–541.
Gray IC, Nobile C, Muresu R, Ford S & Spurr NK (1995) A 2.4-megabase physical map spanning the CYP2C gene cluster on chromosome 10q24. Genomics 28(2): 328–332.

92
Gu L, Gonzalez FJ, Kalow W & Tang BK (1992) Biotransformation of caffeine, paraxanthine, theobromine and theophylline by cDNA-expressed human CYP1A2 and CYP2E1. Pharmacogenetics 2(2): 73–77.
Guengerich FP (1995) Cytochromes P450 of human liver. Classification and activity profiles of the major enzymes. In Pacifici GM & Fracchia GN (eds) Advances in drug metabolism in man. Luxembourg, Office for the Official Publications of the European Communities: 179–231.
Guengerich FP (1999) Cytochrome P-450 3A4: regulation and role in drug metabolism. Annu Rev Pharmacol Toxicol 39(1): 1–17.
Guengerich FP (1995) Cytochromes P450 of human liver. Classification and activity profiles of the major enzymes. In Pacifici GM & Fracchia GN (eds) Advances in drug metabolism in man. Office for the Official Publications of the European Communities. Luxembourg: 179–231.
Guengerich FP (2001) Common and uncommon cytochrome P450 reactions related to metabolism and chemical toxicity. Chem Res Toxicol 14(6): 611–650.
Guengerich FP & Shimada T (1991) Oxidation of toxic and carcinogenic chemicals by human cytochrome P-450 enzymes. Chem Res Toxicol 4(4): 391–407.
Guillouzo A (1995) Acquisition and use of humanin vitro liver preparations. Cell Biol Toxicol 11(3): 141–145.
Gundert-Remy U & Sonich-Mullin C (2002) The use of toxicokinetic and toxicodynamic data in risk assessment: an international perspective. Sci Total Environ 288(1-2): 3–11.
Guo Z, Raeissi S, White RB & Stevens JC (1997) Orphenadrine and Methimazole Inhibit Multiple Cytochrome P450 Enzymes in Human Liver Microsomes. Drug Metab Dispos 25(3): 390–393.
Haas DW, Ribaudo HJ, Kim RB, Tierney C, Wilkinson GR, Gulick RM, Clifford DB, Hulgan T, Marzolini C & Acosta EP (2004) Pharmacogenetics of efavirenz and central nervous system side effects: an adult AIDS clinical trials group study. AIDS 18(18): 2391–2400.
Haehner B, Gorski J, Vandenbranden M, Wrighton S, Janardan S, Watkins P & Hall S (1996) Bimodal distribution of renal cytochrome P450 3A activity in humans. Mol Pharmacol 50(1): 52–59.
Hakkola J, Raunio H, Purkunen R, Saarikoski S, Vähäkangas K, Pelkonen O, Edwards RJ, Boobis AR & Pasanen M (2001) Cytochrome P450 3A expression in the human fetal liver: evidence that CYP3A5 is expressed in only a limited number of fetal livers. Biol Neonate 80: 193–201.
Halling J, Weihe P & Brosen K (2008) The CYP2D6 polymorphism in relation to the metabolism of amitriptyline and nortriptyline in the Faroese population. Br J Clin Pharmacol 65(1): 134–138.
Hargreaves M, Jones B, Smith D & Gescher A (1994) Inhibition of p-nitrophenol hydroxylase in rat liver microsomes by small aromatic and heterocyclic molecules. Drug Metab Dispos 22(5): 806–810.

93
Harleton E, Webster M, Bumpus NN, Kent UM, Rae JM & Hollenberg PF (2004) Metabolism of N,N',N''-triethylenethiophosphoramide by CYP2B1 and CYP2B6 results in the inactivation of both isoforms by two distinct mechanisms. J Pharmacol Exp Ther 310(3): 1011–1019.
Hassall KA (1990) The biochemistry and uses of pesticides: structure metabolism, mode of action and uses in crop protection. Weinheim, Germany, Cambridge.
Hattis D, Erdreich L & Ballew M (1987) Human variability in susceptibility to toxic chemicals— A preliminary analysis of pharmacokinetic data from normal volunteers. Risk Anal 7(4): 415–426.
Hattis D, Ginsberg G, Sonawane B, Smolenski S, Russ A, Kozlak M & Goble R (2003) Differences in pharmacokinetics between children and adults—II. Children's variability in drug elimination half-lives and in some parameters needed for physiologically-based pharmacokinetic modeling. Risk Anal 23(1): 117–142.
Hecht SS (1998) Biochemistry, biology, and carcinogenicity of tobacco-specific N-nitrosamines. Chem Res Toxicol 11(6): 559–603.
Heimark LD, Wienkers L, Kunze K, Gibaldi M, Eddy AC, Trager WF, O'Reilly R,A. & Goulart DA (1992) The mechanism of the interaction between amiodarone and warfarin in humans. Clin Pharm Ther 51(4): 398–407.
Hesse LM, Venkatakrishnan K, Court MH, von Moltke LL, Duan SX, Shader RI & Greenblatt DJ (2000) CYP2B6 mediates the in vitro hydroxylation of bupropion: potential drug interactions with other antidepressants. Drug Metab Dispos 28(10): 1176–1183.
Hewitt NJ, Lechón MJG, Houston JB, Hallifax D, Brown HS, Maurel P, Kenna JG, Gustavsson L, Lohmann C, Skonberg C, Guillouzo A, Tuschl G, Li AP, LeCluyse E, Groothuis GMM & Hengstler JG (2007) Primary hepatocytes: current understanding of the regulation of metabolic enzymes and transporter proteins, and pharmaceutical practice for the use of hepatocytes in metabolism, enzyme induction, transporter, clearance, and hepatotoxicity studies. Drug Metab Rev 39(1): 159.
Hirota N, Ito K, Iwatsubo T, Green CE, Tyson CA, Shimada N, Suzuki H & Sugiyama Y (2001) In Vitro/in Vivo scaling of alprazolam metabolism by CYP3A4 and CYP3A5 in humans. Biopharm Drug Dispos 22(2): 53–71.
Hjelle J, Hazelton G, Klaassen C & Hjelle J (1986) Glucuronidation and sulfation in rabbit kidney. J Pharmacol Exp Ther 236(1): 150–156.
Hodge HC, Downs WL, Panner BS, Smith DW & Maynard EA (1967) Oral toxicity and metabolism of diuron (N-(3,4-dichlorophenyl)-N',N'-dimethylurea) in rats and dogs. Food Cosmet Toxicol 5(4): 513–31.
Hodgson E & Goldstein JA (2001) Metabolism of toxicants: phase I reactions and pharmacogenetics. In Hodgson E & Smart RC (eds) Introduction to Biochemical Toxicology. New York, Wiely: 67–113.
Hodgson E & Rose RL (2007a) Human metabolic interactions of environmental chemicals. J Biochem Mol Toxicol 21(4): 182–186.
Hodgson E & Rose RL (2007b) The importance of cytochrome P450 2B6 in the human metabolism of environmental chemicals. Pharm Thera 113(2): 420–428.

94
Hodgson E & Rose RL (2005) Human metabolism and metabolic interactions of deployment-related chemicals. Drug Metab Rev 37(1): 1.
Hoffman SMG, Fernandez-Salguero P, Gonzalez FJ & Mohrenweiser HW (1995) Organization and evolution of the cytochrome P450 CYP2A-2B-2F subfamily gene cluster on human chromosome 19. J Mol Evol 41(6): 894–900.
Hoffman SMG, Nelson DR & Keeney DS (2001) Organization, structure and evolution of the CYP2 gene cluster on human chromosome 19. Pharmacogenetics 11(8): 687–698.
Honkakoski P & Negishi M (1997) Characterization of a phenobarbital-responsive enhancer module in mouse P450 cyp2b10 gene. J Biol Chem 272(23): 14943–14949.
Hrelia P, Maffei F, Fimognari C, Vigagni F & Cantelli-Forti G (1996) Cytogenetic effects of metalaxyl on human and animal chromosomes. Mutat Res 369(1): 81–86.
Huang W, Lin YS, McConn DJ,II, Calamia JC, Totah RA, Isoherranen N, Glodowski M & Thummel KE (2004) Evidence of significant contribution from CYP3A5 to hepatic drug metabolism. Drug Metab Dispos 32(12): 1434–1445.
Huang Z, Roy P & Waxman DJ (2000) Role of human liver microsomal CYP3A4 and CYP2B6 in catalyzing N-dechloroethylation of cyclophosphamide and ifosfamide. Biochem Pharmacol 59(8): 961–972.
Hukkanen J, Lassila A, Paivarinta K, Valanne S, Sarpo S, Hakkola J, Pelkonen O & Raunio H (2000) Induction and regulation of xenobiotic-metabolizing cytochrome P450s in the human A549 lung adenocarcinoma cell line. Am J Respir Cell Mol Biol 22(3): 360–366.
IARC (1991) Monographs on the evaluation of carcinogenic risks to humans. Occupational Exposures in Insecticide Application and Some Pesticides 53: 33–586.
Iesce MR, della Greca M, Cermola F, Rubino M, Isidori M & Pascarella L (2006) Transformation and ecotoxicity in carbamic pesticides in water. Environ Sci Pollut Res Int 13(2): 105–109.
Imaoka S, Yamada T, Hiroi T, Hayashi K, Sakaki T, Yabusaki Y & Funae Y (1996) Multiple forms of human P450 expressed in Saccharomyces cerevisiae: systematic characterization and comparison with those of the rat. Biochem Pharmacol 51(8): 1041–1050.
Ingelman-Sundberg M (2004) Genetic polymorphisms of cytochrome P450 2D6 (CYP2D6): clinical consequences, evolutionary aspects and functional diversity. Pharmacogenomics J 5(1): 6–13.
Ingelman-Sundberg M, Oscarson M & McLellan RA (1999) Polymorphic human cytochrome P450 enzymes: an opportunity for individualized drug treatment. Trends Pharmacol Sci 20(8): 342–349.
IPCS (2005) The WHO recommended classification of pesticides by hazard and guidelines to classification 2004.
Jacolot F, Simon I, Dreano Y, Beaune P, Riche C & Berthou F (1991) Identification of the cytochrome P450 IIIA family as the enzymes involved in the N-demethylation of tamoxifen in human liver microsomes. Biochem Pharmacol 41(12): 1911–1919.

95
Jae Wook Ko , Zeruesenay Desta , Nadia V Soukhova , Tracy T & Flockhart DA (2000) In vitro inhibition of the cytochrome P450 (CYP450) system by the antiplatelet drug ticlopidine: potent effect on CYP2C19 and CYP2D6. Br J Clin Pharmacol 49(4): 343–351.
Jiang Z, Shu Y, Chen X, Huang S, Zhu R, Wang W, He N & Zhou H (2002) The role of CYP2C19 in amitriptyline N-demethylation in Chinese subjects. Eur J Clin Pharmacol 58(2): 109–113.
Jinno H, Tanaka-Kagawa T, Ohno A, Makino Y, Matsushima E, Hanioka N & Ando M (2003) Functional characterization of cytochrome P450 2B6 allelic variants. Drug Metab Dispos 31(4): 398–403.
Johnson JA & Burlew BS (1996) Metoprolol metabolism via cytochrome P4502D6 in ethnic populations. Drug Metab Dispos 24(3): 350–355.
Johnson TN, Tucker GT, Tanner MS & Rostami-Hodjegan A (2005) Changes in liver volume from birth to adulthood: A meta-analysis. Liver Transplantation 11(12): 1481–1493.
Jokanovic M (2001) Biotransformation of organophosphorus compounds. Toxicology 166(3): 139–160.
Jung F, Richardson TH, Raucy JL & Johnson EF (1997) Diazepam metabolism by cDNA-expressed human 2C P450s. Identification of P4502C18 and P4502C19 as low Km diazepam N-demethylases. Drug Metab Dispos 25(2): 133–139.
Kalberlah F & Schneider K (1998) Quantification of extrapolation factors. Final report of the research project No 116 06 113 of the Federal Environmental Agency, Schriftenreihe der Bundesanstalt fur Arbeitsschutz und Arbeitsmedizin-Forschung-FB797, Germany.
Kanebratt KP & Andersson TB (2008) Evaluation of HepaRG cells as an in vitro model for human drug metabolism studies. Drug Metab Dispos 36(7): 1444–1452.
Kappers WA, Edwards RJ, Murray S & Boobis AR (2001) Diazinon Is Activated by CYP2C19 in Human Liver. Toxicol Appl Pharmacol 177(1): 68–76.
Karabay NU & Gunnehir OM (2005) Cytogenetic and genotoxic effects of the insecticides, imidacloprid and methamidophos. Genet Mol Res 4(4): 653–662.
KEMI (2003) Human health risk assessment. Proposals for the use of assessment (uncertainty) factors Application to risk assessment for plant protection products, industrial chemicals and biocidal products within the European Union. Stockholm, Sweden, Swedish National Chemicals Inspectorate.
Kessova I & Cederbaum AI (2003) CYP2E1: biochemistry, toxicology, regulation and function in ethanol-induced liver injury. Curr Mol Med 5: 509–518.
Khan KK, He YQ, Correia MA & Halpert JR (2002) Differential oxidation of mifepristone by cytochromes P450 3A4 and 3A5: selective inactivation of P450 3A4. Drug Metab Dispos 30(9): 985–990.
Khan WA, Park SS, Gelboin HV, Bickers DR & Mukhtar H (1989) Monoclonal antibodies directed characterization of epidermal and hepatic cytochrome P-450 isozymes induced by skin application of therapeutic crude coal tar. J Invest Dermatol 93(1): 40–45.

96
Kim K, Park P, Kim H, Ha J & Park J (2005) Effect of quercetin on the pharmacokinetics of rosiglitazone, a CYP2C8 substrate, in healthy subjects. J Clin Pharmacol 45(8): 941–946.
Kirchheiner J & Brockmoller J (2005) Clinical consequences of cytochrome P450 2C9 polymorphisms. Clin Pharmacol Ther 77(1): 1–16.
Kitada M, Kamataki T & Itahashi K (1987) P-450 HFLa, a form of cytochrome P-450 purified from human fetal livers, is the 16α-hydroxylase of dehydroepiandrosterone 3-sulfate. J Biol Chem 262(28): 13534–13537.
Kitada M & Kamataki T (1994) Cytochrome P450 in human fetal liver: significance and fetal-specific expresson. Drug Metab Rev 26(1-2): 305–323.
Kitada M, Kamataki T, Itahashi K, Rikihisa T & Kanakubo Y (1987) Significance of cytochrome P-450 (P-450 HFLa) of human fetal livers in the steroid and drug oxidations. Biochem Pharmacol 36(4): 453–456.
Kitada M, Kamataki T, Itahashi K, Rikihisa T, Kato R & Kanakubo Y (1985) Purification and properties of cytochrome P-450 from homogenates of human fetal livers. Arch Biochem Biophys 241(1): 275–280.
Kitada M, Taneda M, Itahashi K & Kamataki T (1991) Four forms of cytochrome P-450 in human fetal liver: purification and their capacity to activate promutagens. Cancer Sci 82(4): 426–432.
Kitagawa K, Kunugita N, Katoh T, Yang M & Kawamoto T (1999) The significance of the homozygous CYP2A6 deletion on nicotine metabolism: a new genotyping method of CYP2A6 using a single PCR–RFLP. Biochem Biophys Res Commun 262(1): 146–151.
Kitamura R, Sato K, Sawada M, Itoh S, Kitada M, Komori M & Kamataki T (1992) Stable expression of cytochrome P450IIIA7 cDNA in human breast cancer cell line MCF-7 and its application to cytotoxicity testing. Arch Biochem Biophys 292(1): 136–140.
Knasmüller S, Mersch-Sundermann V, Kevekordes S, Darroudi F, Huber WW, Hoelzl C, Bichler J & Majer BJ (2004) Use of human-derived liver cell lines for the detection of environmental and dietary genotoxicants; current state of knowledge. Toxicology 198(1-3): 315–328.
Ko J, Sukhova N, Thacker D, Chen P & Flockhart DA (1997) Evaluation of omeprazole and lansoprazole as inhibitors of cytochrome P450 isoforms. Drug Metab Dispos 25(7): 853–862.
Kobayashi K, Chiba K, Yagi T, Shimada N, Taniguchi T, Horie T, Tani M, Yamamoto T, Ishizaki T & Kuroiwa Y (1997) Identification of cytochrome P450 isoforms involved in citalopram N-demethylation by human liver microsomes. J Pharmacol Exp Ther 280(2): 927–933.
Komatsu T, Yamazaki H, Asahi S, Gillam EMJ, Guengerich FP, Nakajima M & Yokoi T (2000) Formation of a dihydroxy metabolite of phenytoin in human liver microsomes/cytosol: roles of cytochromes P450 2c9, 2c19, and 3a4. Drug Metab Dispos 28(11): 1361–1368.

97
Kono Y, Sato Y & Okada Y (1983) Activation of an O-ethyl S-n-propyl phosphorothiolate, TIA-230, in the central nerve of Spodoptera larvae. Pestic Biochem Physiol 20(2): 225–231.
Konradsen F, van der Hoek W, Gunnell D & Eddleston M (2005) Missing deaths from pesticide self-poisoning at the IFCS Forum IV. Bull World Health Organ 83(2): 157–158.
Kornreich M & Montgomery CA (1990) Toxicology and carcinogenesis studies of 2,6-xylidine (2,6-dimethylaniline) in Charles River CD rats. National Toxicology Program (NTP) NIH Publication No. 90–2534: 1–138.
Kostrubsky VE, Ramachandran V, Venkataramanan R, Dorko K, Esplen JE, Zhang S, Sinclair JF, Wrighton SA & Strom SC (1999) The use of human hepatocyte cultures to study the induction of cytochrome P-450. Drug Metab Dispos 27(8): 887–894.
Kremers P (1999) Liver microsomes: a convenient tool for metabolism studies but ... In Boobis AR, Kremers P, Pelkonen O & Pithan K (eds) European symposium on the prediction of drug metabolism in man: progress and problems. , Office for Official Publications of the European Communities: 38–52.
Kronbach T, Mathys D, Gut J, Catin T & Meyer UA (1987) High-performance liquid chromatographic assays for bufuralol 1'-hydroxylase, debrisoquine 4-hydroxylase, and dextromethorphan O-demethylase in microsomes and purified cytochrome P-450 isozymes of human liver. Anal Biochem 162(1): 24–32.
Kuehl P, Zhang J, Lin Y, Lamba J, Assem M, Schuetz J, Watkins PB, Daly A, Wrighton SA, Hall SD, Maurel P, Relling M, Brimer C, Yasuda K, Venkataramanan R, Strom S, Thummel K, Boguski MS & Schuetz E (2001) Sequence diversity in CYP3A promoters and characterization of the genetic basis of polymorphic CYP3A5 expression. Nat Genet 27(4): 383.
Kumar V, Wahlstrom JL, Rock DA, Warren CJ, Gorman LA & Tracy TS (2006) CYP2C9 inhibition: impact of probe selection and pharmacogenetics on in vitro inhibition profiles. Drug Metab Dispos 34: 1966–1975.
Lamoree MH, Swart CP, van der Horst A & van Hattum B (2002) Determination of diuron and the antifouling paint biocide Irgarol 1051 in Dutch marinas and coastal waters. J Chromatogr A 970(1-2): 183–190.
Lang D, Criegee D, Grothusen A, Saalfrank R & Bocker R (1996) In vitro metabolism of atrazine, terbuthylazine, ametryne, and terbutryne in rats, pigs, and humans. Drug Metab Dispos 24(8): 859–865.
Lang DH, Rettie AE & Bocker RH (1997) Identification of enzymes involved in the metabolism of atrazine, terbuthylazine, ametryne, and terbutryne in human liver microsomes. Chem Res Toxicol 10(9): 1037–1044.
Lang T, Klein K, Fischer J, Nussler AK, Neuhaus P, Hofmann U, Eichelbaum M, Schwab M & Zanger UM (2001) Extensive genetic polymorphism in the human CYP2B6 gene with impact on expression and function in human liver. Pharmacogenetics 11(5): 399–415.
Lapworth DJ & Gooddy DC (2006) Source and persistence of pesticides in a semi-confined chalk aquifer of southeast England. Environ Pollut 144(3): 1031–1044.

98
Lawton M, Gasser R, Tynes R, Hodgson E & Philpot R (1990) The flavin-containing monooxygenase enzymes expressed in rabbit liver and lung are products of related but distinctly different genes. J Biol Chem 265(10): 5855–5861.
LeBlanc GA & Dauterman WC (2001) Conjugation and elimination of toxicants. In Hodgson E & Smart RC (eds) Introduction to Biochemical Toxicology. New York, Wiley.
LeCluyse EL (2001) Human hepatocyte culture systems for the in vitro evaluation of cytochrome P450 expression and regulation. Eur J Pharm Sci 13(4): 343–368.
Lee H, Moon J, Chang C, Choi H, Park H, Park B, Lee H, Hwang E, Lee Y, Liu K & Kim J (2006a) Stereoselective metabolism of endosulfan by human liver microsomes and human cytochrome P450 isoforms. Drug Metab Dispos 34(7): 1090–1095.
Lee S, Ohta K, Tashiro S & Shono T (2006b) Metabolic resistance mechanisms of the housefly (Musca domestica) resistant to pyraclofos. Pestic Biochem Physiol 85(2): 76–83.
Lee SST, Buters JTM, Pineau T, Fernandez-Salguero P & Gonzalez FJ (1996) Role of CYP2E1 in the hepatotoxicity of acetaminophen. J Biol Chem 271(20): 12063–12067.
Leoni C, Buratti FM & Testai E (2008) The participation of human hepatic P450 isoforms, flavin-containing monooxygenases and aldehyde oxidase in the biotransformation of the insecticide fenthion. Toxicol Appl Pharmacol 233(2): 343–352.
Lewis DFV (2004) 57 Varieties: the human cytochromes P450. Pharmacogenomics 5(3): 305–318.
Li AP, Maurel P, Gomez-Lechon MJ, Cheng LC & Jurima-Romet M (1997) Preclinical evaluation of drug—drug interaction potential: present status of the application of primary human hepatocytes in the evaluation of cytochrome P450 induction. Chem Biol Interact 107(1-2): 5–16.
Li X, Bjorkman A, Andersson TB, Ridderstrom M & Masimirembwa CM (2002) Amodiaquine clearance and its metabolism to N-desethylamodiaquine is mediated by CYP2C8: A new high affinity and turnover enzyme-specific probe substrate. J Pharmacol Exp Ther 300(2): 399–407.
Lieber CS (2004) The Discovery of the microsomal ethanol oxidizing system and its physiologic and pathologic role. Drug Metab Rev 36(3-4): 511–529.
Lin JH & Lu AYH (1997) Role of pharmacokinetics and metabolism in drug discovery and development. Pharmacol Rev 49(4): 403–449.
Lin JH & Lu AYH (1998) Inhibition and induction of cytochrome P450 and the clinical implications. Clin Pharmacokinet 35(5): 361–390.
Lin YS, Dowling ALS, Quigley SD, Farin FM, Zhang J, Lamba J, Schuetz EG & Thummel KE (2002) Co-regulation of CYP3A4 and CYP3A5 and contribution to hepatic and intestinal midazolam metabolism. Mol Pharmacol 62(1): 162–172.
Liska D, Lyon M & Jones DS (2006) Detoxification and biotransformational imbalances. J Sci Heal 2(2): 122–140.
Liu C, Russell RM & Wang XD (2003) Exposing ferrets to cigarette smoke and a pharmacological dose of beta-carotene supplementation enhance in vitro retinoic acid catabolism in lungs via induction of cytochrome P450 enzymes. J Nutr 133: 173–179.

99
Liu Y, Hao H, Liu C, Wang G & Xie H (2007) Drugs as CYP3A probes, inducers, and inhibitors. Drug Metab Rev 39(4): 699.
Low LK (1998) Metabolic changes of drugs and related organic compounds. In Delgado JN & Remers WA (eds) Wilson and Gisvold's textbook of organic medicinal and pharmaceutical chemistry. Philadelphia, Lippincott-Raven: 43–122.
Lu FC (1979) Assessment at an international level of health hazards to man of chemicals shown to be carcinogenic in laboratory animals. In Coulston EF (ed) Regulatory aspects of Carcinogenesis. New York, Academic Press: 315–328.
Mäenpää J, Hall SD, Ring BJ, Strom SC & Wrighton SA (1998) Human cytochrome P450 3A (CYP3A) mediated midazolam metabolism: the effect of assay conditions and regioselective stimulation by alpha-naphthoflavone, terfenadine and testosterone. Pharmacogenetics 8(2): 137–155.
Malato S, Caceres J, Fernandez-Alba A, Piedra L, Hernando MD, Aguera A & Vial J (2003) Photocatalytic treatment of diuron by solar photocatalysis: evaluation of main intermediates and toxicity. Environ Sci Technol 37(11): 2516–2524.
Maroni M, Fanetti AC & Metruccio F (2006) Risk assessment and management of occupational exposure to pesticides in agriculture. Med Lav : 430–437.
Marsden PJ, Kuwano E & Fukuto TR (1982) Metabolism of carbosulfan [2,3-dihydro-2,2-dimethylbenzofuran-7-yl (di-n-butylaminothio)methylcarbamate] in the rat and house fly. Pestic Biochem Physiol 18(1): 38–48.
Mazur CS, Kenneke JF, Tebes-Stevens C, Okino MS & Lipscomb JC (2007) In vitro metabolism of the fungicide and environmental contaminant trans-bromuconazole and implications for risk assessment. J Toxicol Environ Health, A: Curr Iss 70(14): 1241–1250.
McGinnity DF & Riley RJ (2001) Predicting drug pharmacokinetics in humans from in vitro metabolism studies. Biochem Soc Trans 29(Pt 2): 135–139.
McKinnon RA, Hall PM, Quattrochi LC, Tukey RH & McManus ME (1991) Localisation of CYP1A1 and CYP1A2 messenger RNA in normal human liver and in hepatocellular carcinoma by in situ hybridization. Hepatology 14: 848–856.
Messina ES, Tyndale RF & Sellers EM (1997) A major role for CYP2A6 in nicotine C-oxidation by human liver microsomes. J Pharmacol Exp Ther 282(3): 1608–1614.
Miners JO & Birkett DJ (1998) Cytochrome P4502C9: an enzyme of major importance in human drug metabolism. Br J Clin Pharmacol 45(6): 525–538.
Montoliu C, Sancho-Tello M, Azorin I, Burgal M, Vallés S & Renau-Piqueras J (1995) Ethanol increases cytochrome P4502E1 and induces oxidative stress in astrocytes. J Neurochem 65(6): 2561–2570.
Moody GC, Griffin SJ, Mather AN, Mcginnity DF & Riley RJ (1999) Fully automated analysis of activities catalysed by the major human liver cytochrome P450 (CYP) enzymes: assessment of human CYP inhibition potential. Xenobiotica 29(1): 53–75.
Moustafa GG, Ibrahim ZS, Ahmed MM, Ghoneim MH, Sakamoto KQ, Ishizuka M & Fujita S (2008) Downregulation of male-specific cytochrome P450 by profenofos. Jpn J Vet Res 56(2): 109–118.

100
Murray M & Reidy G (1990) Selectivity in the inhibition of mammalian cytochromes P-450 by chemical agents. Pharmacol Rev 42(2): 85–101.
Musana AK & Wilke RA (2005) Gene-based drug prescribing: clinical implications of the cytochrome P450 genes. WMJ 104(6): 61–66.
Mutch E, Daly AK, Leathart JB, Blain PG & Williams FM (2003) Do multiple cytochrome P450 isoforms contribute to parathion metabolism in man? Arch Toxicol 77: 313–320.
Mutch E & Williams FM (2006) Diazinon, chlorpyrifos and parathion are metabolised by multiple cytochromes P450 in human liver. Toxicology 224(1-2): 22–32.
Nagahori H, Yoshino H, Tomigahara Y, Isobe N, Kaneko H & Nakatsuka I (2000) Metabolism of furametpyr. 1. identification of metabolites and in vitro biotransformation in rats and humans. J Agric Food Chem 48(11): 5754–5759.
Naidu KA (1989) Inhibition of monoamine oxidase by the fungicide metalaxyl. J Toxicol Environ Health, A 27(3): 395.
Naidu KA & Radhakrishnamurty R (1988) Metalaxyl-induced bradycardia in rats: Mediated by α-adrenoreceptors. J Toxicol Environ Health, A 23(4): 495.
Naidu KA & Radhakrishnamurty R (1989) Effect of metalaxyl on heart rate in rats: Role of α1-adrenoreceptors. J Toxicol Environ Health, A 26(2): 203.
Nakajima M, Yamamoto T, Nunoya K, Yokoi T, Nagashima K, Inoue K, Funae Y, Shimada N, Kamataki T & Kuroiwa Y (1996a) Characterization of CYP2A6 involved in 3'-hydroxylation of cotinine in human liver microsomes. J Pharmacol Exp Ther 277(2): 1010–1015.
Nakajima M, Yamamoto T, Nunoya K, Yokoi T, Nagashima K, Inoue K, Funae Y, Shimada N, Kamataki T & Kuroiwa Y (1996b) Role of human cytochrome P4502A6 in C-oxidation of nicotine. Drug Metab Dispos 24(11): 1212–1217.
Nakajima T, Elovaara E, Gonzalez FJ, Gelboin HV, Raunio H, Pelkonen O, Vainio H & Aoyama T (1994) Styrene metabolism by cDNA-expressed human hepatic and pulmonary cytochromes P450. Chem Res Toxicol 7(6): 891–896.
Nebert DW & Russell DW (2002) Clinical importance of the cytochromes P450. Lancet 360(9340): 1155–1162.
Nelson DR, Koymans L, Kamataki T, Stegeman JJ, Feyereisen R, Waxman DJ, Waterman MR, Gotoh O, Coon MJ, Estabrook RW, Gunsalus IC & Nebert DW (1996) P450 superfamily: update on new sequences, gene mapping, accession numbers and nomenclature. Pharmacogenetics 6(1): 1–42.
Nelson DR, Zeldin DC, Hoffman SMG, Maltais LJ, Wain HM & Nebert DW (2004) Comparison of cytochrome P450 (CYP) genes from the mouse and human genomes, including nomenclature recommendations for genes, pseudogenes and alternative-splice variants. Pharmacogenetics 14(1): 1–18.
Newton DJ, Wang RW & Lu AY (1995) Cytochrome P450 inhibitors. Evaluation of specificities in the in vitro metabolism of therapeutic agents by human liver microsomes. Drug Metab Dispos 23(1): 154–158.

101
Nhamburo P, Kimura S, McBride O, Kozak C, Gelboin H & Gonzalez F (1990) The human CYP2F gene subfamily: identification of a cDNA encoding a new cytochrome P450, cDNA-directed expression, and chromosome mapping. Biochemistry 29(23): 5491–5499.
Oesch F, Herrero ME, Hengstler JG, Lohmann M & Arand M (2000) Metabolic Detoxification: Implications for Thresholds. Toxicol Pathol 28(3): 382–387.
Ohhira S, Enomoto M & Matsui H (2006) In vitro metabolism of tributyltin and triphenyltin by human cytochrome P-450 isoforms. Toxicology 228(2-3): 171–177.
Okamura H, Aoyama I, Ono Y & Nishida T (2003) Antifouling herbicides in the coastal waters of western Japan. Mar Pollut Bull 47(1-6): 59–67.
Olinga P, Meijer DKF, Slooff MJH & Groothuis GMM (1998) Liver slices in in vitro pharmacotoxicology with special reference to the use of human liver tissue. Toxicol In Vitro 12(1): 77–100.
Olsen AK, Hansen KT & Friis C (1997) Pig hepatocytes as an in vitro model to study the regulation of human CYP3A4: Prediction of drug-drug interactions with 17α-ethynylestradiol. Chem Biol Interact 107(1-2): 93–108.
Ozmen M, Sener S, Mete A & Kucukbay H (1999) In vitro and in vivo acetylcholinesterase-inhibiting effect of new classes of organophosphorus compounds. Environ Toxicol Chem 18(2): 241–246.
Paine MF, Khalighi M, Fisher JM, Shen DD, Kunze KL, Marsh CL, Perkins JD & Thummel KE (1997) Characterization of interintestinal and intraintestinal variations in human CYP3A-dependent metabolism. J Pharmacol Exp Ther 283(3): 1552–1562.
Paolini M, Mesirca R, Pozzetti L, Sapone A & Cantelli-Forti G (1996) Biomarkers of effect in evaluating metalaxyl cocarcinogenesis. Selective induction of murine CYP 3A isoform. Mutat Res 361(2-3): 157–164.
Park YH, Kullberg MP & Hinsvark ON (1984) Quantitative determination of dextromethorphan and three metabolites in urine by reverse-phase high-performance liquid chromatography. J Pharm Sci 73(1): 24–29.
Parkinson A (2001) Biotransformation of xenobiotics. In Klaassen CD (ed) Casarett and Doull's toxicology : the basic science of poisons. New York ; London, McGraw-Hill Medical Pub. Division: 113–186.
Pasanen M & Pelkonen O (1994) The expression and environmental regulation of P450 enzymes in human placenta. Crit Rev Toxicol 24(3): 211–229.
Pearce RE, McIntyre CJ, Madan A, Sanzgiri U, Draper AJ, Bullock PL, Cook DC, Burton LA, Latham J, Nevins C & Parkinson A (1996) Effects of freezing, thawing, and storing human liver microsomes on cytochrome P450 activity. Arch Biochem Biophys 331(2): 145–169.
Pedro F, Susan H, Suzanne C, Harvey M, Hannu R, Arja R, Olavi P, Jin-ding H, William E, Jeffrey I & Frank G (1995) A genetic polymorphism in coumarin 7-hydroxylation: sequence of the human CYP2A genes and identification of variant CYP2A6 alleles. Am J Hum Genet 57(3): 651–660.

102
Pelkonen O & Breimer DD (1994) Role of environmental factors in the pharmacokinetics of drugs: Considerations with respect to animal models, P-450 enzymes, and probe drugs. In Welling PG & Balant LP (eds) Handbook of Experimental Pharmacology. Berlin, Germany, Springer-Verlag: 289–332.
Pelkonen O, Turpeinen M, Hakkola J, Honkakoski P, Hukkanen J & Raunio H (2008) Inhibition and induction of human cytochrome P450 enzymes: current status. Arch Toxicol 82(10): 667–715.
Pelkonen O, Kaltiala EH, Larmi TKI & Karki NT (1974) Cytochrome P 450 linked monooxygenase system and drug induced spectral interactions in human liver microsomes. Chem Biol Interact 9(3): 205–216.
Pelkonen O, Maenpaa J, Taavitsainen P, Rautio A & Raunio H (1998) Inhibition and induction of human cytochrome P450 (CYP) enzymes. Xenobiotica 28(12): 1203–1253.
Pelkonen O & Turpeinen M (2007) In vitro–in vivo extrapolation of hepatic clearance: Biological tools, scaling factors, model assumptions and correct concentrations. Xenobiotica 37(10): 1066–1089.
Pelkonen O & Raunio H (2005) In vitro screening of drug metabolism during drug development: can we trust the predictions? Expert Opin Drug Metab Toxicol 1(1): 49–59.
Pelkonen O, Rautio A, Raunio H & Pasanen M (2000) CYP2A6: a human coumarin 7-hydroxylase. Toxicology 144(1-3): 139–147.
Pelkonen O, Turpeinen M, Uusitalo J, Rautio A & Raunio H (2005) Prediction of drug metabolism and interactions on the basis of in vitro investigations. Basic Clin Pharmacol Toxicol 96(3): 167–175.
Peter R, Bocker R, Beaune PH, Iwasaki M, Guengerich FP & Yang CS (1990) Hydroxylation of chlorzoxazone as a specific probe for human liver cytochrome P-450IIE1. Chem Res Toxicol 3(6): 566–573.
Peters WHM & Kremers PG (1989) Cytochromes P-450 in the intestinal mucosa of man. Biochemical Pharmacology 38(9): 1535–1538.
Peyronneau M, Renaud J, Truan G, Urban P, Pompon D & Mansuy D (1992) Optimization of yeast-expressed human liver cytochrome P450 3A4 catalytic activities by coexpressing NADPH-cytochrome P450 reductase and cytochrome b5. Eu J Biochem 207(1): 109–116.
Preuss R, Angerer J & Drexler H (2003) Naphthalene - An environmental and occupational toxicant. Int Arch Occup Environ Health 76(8): 556–576.
Rae JM, Soukhova NV, Flockhart DA & Desta Z (2002) Triethylenethiophosphoramide is a specific inhibitor of cytochrome P450 2B6: implications for cyclophosphamide metabolism. Drug Metab Dispos 30(5): 525–530.
Rahman A, Korzekwa KR, Grogan J, Gonzalez FJ & Harris JW (1994) Selective biotransformation of taxol to 6alpha-hydroxytaxol by human cytochrome P450 2C8. Cancer Res 54(21): 5543–5546.
Raucy JL, Kraner JC & Lasker JM (1993) Bioactivation of halogenated hydrocarbons by cytochrome P4502E1. Crit Rev Toxicol 23(1): 1.

103
Raunio H, Pasanen M, Mäenpää J, Hakkola J & Pelkonen O (1995) Expression of extrahepatic cytochrome P450 in humans. In Pacifici GM & Fracchia GN (eds) Advances in drug mrtablism in man. Luxembourg: 234–287.
Rendic S & Di Carlo FJ (1997) Human cytochrome P450 enzymes: A status report summarizing their reactions, substrates, inducers, and inhibitors. Drug Metab Rev 29(1-2): 413–580.
Renwick AG (1991) Safety factors and establishment of acceptable daily intakes. Food Addit Contam , Part A 8(2): 135.
Renwick AG (1993) Data-derived safety factors for the evaluation of food additives and environmental contaminants. Food Addit Contam , Part A 10(3): 275.
Renwick AG & Lazarus NR (1998) Human variability and noncancer risk assessment--an analysis of the default uncertainty factor. Regul Toxicol Pharm 27(1): 3–20.
Renzi BE & Krieger RI (1986) Sublethal acute toxicity of carbosulfan [2,3-dihydro-2,2-dimethyl-7-benzofuranyl(di-n-butylaminosulfenyl)(methyl)carbamate] in the rat after intravenous and oral exposures. Toxicol Sci 6(1): 7–15.
Reponen P, Abass K, Mattila S & Pelkonen O (2010) Overview of the metabolism and interactions of pesticides in hepatic in vitro systems. Int J Environ Anal Chem 90(3): 429.
Rettie AE & Jones JP (2005) Clinical and toxicological relevance of CYP2C9: drug-drug interactions and pharmacogenetics. Annu Rev Pharmacol Toxicol 45(1): 477–494.
Rettie AE, Korzekwa KR, Kunze KL, Lawrence RF, Eddy AC, Aoyama T, Gelboin HV, Gonzalez FJ & Trager WF (1992) Hydroxylation of warfarin by human cDNA-expressed cytochrome P-450: a role for P-4502C9 in the etiology of (S)-warfarin-drug interactions. Chem Res Toxicol 5(1): 54–59.
Richardson TH, Griffin KJ, Jung F, Raucy JL & Johnson EF (1997) Targeted antipeptide antibodies to cytochrome P450 2C18 based on epitope mapping of an inhibitory monoclonal antibody to P450 2C51. Arch Biochem Biophys 338(2): 157–164.
Riley RJ, McGinnity DF & Austin RP (2005) A unified model for predicting human hepatic, metabolic clearance from in vitro intrinsic clearance data in hepatocytes and microsomes. Drug Metab Dispos 33(9): 1304–1311.
Riley RJ, Parker AJ, Trigg S & Manners CN (2001) Development of a generalized, quantitative physicochemical model of CYP3A4 inhibition for use in early drug discovery. Pharm Res 18(5): 652–655.
Rodrigues AD (1999) Integrated cytochrome P450 reaction phenotyping. Attempting to bridge the gap between cDNA-expressed cytochromes P450 and native human liver microsomes. Biochem Pharmacol 57(5): 465–480.
Rose RL & Hodgson E (2004) Metabolism of toxicants. In Hodgson E (ed) Text Book of Modern Toxicology. New York, Wiley: 111–148.
Rostami-Hodjegan A & Tucker GT (2007) Simulation and prediction of in vivo drug metabolism in human populations from in vitro data. Nat Rev Drug Discov 6: 140–148.

104
Roy P, Tretyakov O, Wright J & Waxman DJ (1999a) Stereoselective metabolism of ifosfamide by human P-450s 3A4 and 2B6. Favorable metabolic properties of R-enantiomer. Drug Metab Dispos 27(11): 1309–1318.
Roy P, Yu LJ, Crespi CL & Waxman DJ (1999b) Development of a substrate-activity based approach to identify the major human liver P-450 catalysts of cyclophosphamide and ifosfamide activation based on cDNA-expressed activities and liver microsomal P-450 profiles. Drug Metab Dispos 27(6): 655–666.
Rylander T, Neve EPA, Ingelman-Sundberg M & Oscarson M (2001) Identification and tissue distribution of the novel human cytochrome P450 2S1 (CYP2S1). Biochem Biophys Res Commun 281(2): 529–535.
Saarikoski ST, Rivera SP, Hankinson O & Husgafvel-Pursiainen K (2005b) CYP2S1: A short review. Toxicol Appl Pharmacol 207(2, Supplement 1): 62–69.
Saarikoski ST, Wikman HA-, Smith G, Wolff CHJ & Husgafvel-Pursiainen K (2005a) Localization of cytochrome P450 CYP2S1 expression in human tissues by in situ hybridization and immunohistochemistry. J Histochem Cytochem 53(5): 549–556.
Salonpää P, Hakkola J, Pasanen M, Pelkonen O, Vähäkangas K, Battula N, Nouso K & Raunio H (1993) Retrovirus-mediated stable expression of human CYP2A6 in mammalian cells. Eur J Pharmcol 248: 95–102.
Sams C, Mason HJ & Rawbone R (2000) Evidence for the activation of organophosphate pesticides by cytochromes P450 3A4 and 2D6 in human liver microsomes. Toxicol Lett 116(3): 217–221.
Sarkar MA & Jackson BJ (1994) Theophylline N-demethylations as probes for P4501A1 and P4501A2. Drug Metab Dispos 22(6): 827–834.
Schmider J, Greenblatt D, von Moltke L, Harmatz J & Shader R (1995) N-demethylation of amitriptyline in vitro: role of cytochrome P-450 3A (CYP3A) isoforms and effect of metabolic inhibitors. J Pharmacol Exp Ther 275(2): 592–597.
Schuetz JD, Beach DL & Guzelian PS (1994) Selective expression of cytochrome P450 CYP3A mRNAs in embryonic and adult human liver. Pharmacogenetics 4(1): 11–20.
Schulz-Jander DA & Casida JE (2002) Imidacloprid insecticide metabolism: human cytochrome P450 isozymes differ in selectivity for imidazolidine oxidation versus nitroimine reduction. Toxicol Lett 132(1): 65–70.
Scollon EJ, Starr JM, Godin SJ, DeVito MJ & Hughes MF (2009) In vitro metabolism of pyrethroid pesticides by rat and human hepatic microsomes and cytochrome P450 isoforms. Drug Metab Dispos 37(1): 221–228.
Scott EE & Halpert JR (2005) Structures of cytochrome P450 3A4. Trends Biochem Sci 30(1): 5–7.
Seiss M, Nitz S, Kleinsasser N, Buters JTM, Behrendt H, Hickel R & Reichl FX (2007) Identification of 2,3-epoxymethacrylic acid as an intermediate in the metabolism of dental materials in human liver microsomes. Dent 23(1): 9–16.
Sesardic D, Boobis AR, Murray BP, Murray S, Segura J, de la Torre R & Davis DS (1990) Furafylline is a potent and selective inhibitor of cytochrome P4501A2 in man. Br J Clin Pharmacol 29: 651–663.

105
Shimada T, Yamazaki H & Guengerich FP (1996) Ethnic-related differences in coumarin 7-hydroxylation activities catalyzed by cytochrome P4502A6 in liver microsomes of Japanese and Caucasian populations. Xenobiotica 26: 395–403.
Shimada T, Yamazaki H, Mimura M, Inui Y & Guengerich FP (1994) Interindividual variations in human liver cytochrome P-450 enzymes involved in the oxidation of drugs, carcinogens and toxic chemicals: studies with liver microsomes of 30 Japanese and 30 Caucasians. J Pharmacol Exp Ther 270(1): 414–423.
Shimada T, Hayes CL, Yamazaki H, Amin S, Hecht SS, Guengerich FP & Sutter TR (1996) Activation of chemically diverse procarcinogens by human cytochrome P-450 1B1. Cancer Res 56(13): 2979–2984.
Skaanild MT & Friis C (2000) Expression changes of CYP2A and CYP3A in microsomes from pig liver and cultured hepatocytes. Pharmacol Toxicol 87(4): 174–178.
Skett P, Tyson C, Guillouzo A & Maier P (1995) Report on the international workshop on the use of human in vitro liver preparations to study drug metabolism in drug development. Biochem Pharmacol 50(2): 280–285.
Smith GBJ, Bend JR, Bedard LL, Reid KR, Petsikas D & Massey TE (2003) Biotransformation of 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK) in peripheral human lung microsomes. Drug Metab Dispos 31(9): 1134–1141.
Soars MG, Grime K & Riley RJ (2006) Comparative analysis of substrate and inhibitor interactions with CYP3A4 and CYP3A5. Xenobiotica 36(4): 287–299.
Soler C, Mañes J & Picó Y (2006) Determination of carbosulfan and its metabolites in oranges by liquid chromatography ion-trap triple-stage mass spectrometry. J Chromatogr A 1109(2): 228–241.
Someya T, Shimoda K, Suzuki Y, Sato S, Kawashima Y, Hirokane G, Morita S, Yokono A & Takahashi S (2003) Effect of CYP2D6 genotypes on the metabolism of haloperidol in a Japanese psychiatric population. Neuropsychopharmacology 28(8): 1501–1505.
Sousa RL & Marletta MA (1985) Inhibition of cytochrome P-450 activity in rat liver microsomes by the naturally occurring flavonoid, quercetin. Arch Biochem Biophys 240(1): 345–357.
Speirs CJ, Murray S, Boobis AR, Seddon CE & Davies DS (1986) Quinidine and the identification of drugs whose elimination is impaired in subjects classified as poor metabolizers of debrisoquine. Br J Clin Pharmacol 22: 739–743.
Steerenberg P, van Amelsvoort L, Colosio C, Corsini E, Fustinoni S, Vergieva T, Zaikov C, Pennanen S, Liesivuori J & Van Loveren H (2008) Toxicological evaluation of the immune function of pesticide workers, a European wide assessment. Hum Exp Toxicol 27(9): 701–707.
Stresser DM & Kupfer D (1998) Human cytochrome P450-catalyzed conversion of the proestrogenic pesticide methoxychlor into an estrogen. Role of CYP2C19 and CYP1A2 in O-demethylation. Drug Metab Dispos 26(9): 868–874.
Stresser DM & Kupfer D (1999) Monospecific Antipeptide Antibody to Cytochrome P-450 2B6. Drug Metab Dispos 27(4): 517–525.
Stubbins MJ, Harries LW, Smith G, Tarbit MH & Wolf CR (1996) Genetic analysis of the human cytochrome P450 CYP2C9 locus. Pharmacogenetics 6(5): 429–439.

106
Sukul P & Spiteller M (2000) Metalaxyl: persistence, degradation, metabolism, and analytical methods. Rev Environ Contam Toxicol 164: 1–26.
Sullivan-Klose TH, Ghanayem BI, Bell DA, Zhang ZY, Kaminsky LS, Shenfield GM, Miners JO, Birkett DJ & Goldstein JA (1996) The role of the CYP2C9-Leu359 allelic variant in the tolbutamide polymorphism. Pharmacogenetics 6(4): 341–349.
Suzuki H, Kneller MB, Haining RL, Trager WF & Rettie AE (2002) (+)-N-3-benzyl-nirvanol and (-)-N-3-benzyl-phenobarbital: new potent and selective in vitro inhibitors of CYP2C19. Drug Metab Dispos 30(3): 235–239.
Taavitsainen P, Anttila M, Nyman L, Karnani H, Salonen JS & Pelkonen O (2000) Selegiline metabolism and cytochrome P450 enzymes: In vitro study in human liver microsomes. Pharmacol Toxicol 86(5): 215–221.
Taavitsainen P, Juvonen R & Pelkonen O (2001) In vitro inhibition of cytochrome P450 enzymes in human liver microsomes by a potent CYP2A6 inhibitor, trans-2-phenylcyclopropylamine (tranylcypromine), and its nonamine analog, cyclopropylbenzene. Drug Metab Dispos 29(3): 217–222.
Tang J, Cao Y, Rose RL, Brimfield AA, Dai D, Goldstein JA & Hodgson E (2001) Metabolism of chlorpyrifos by human cytochrome P450 isoforms and human, mouse, and rat liver microsomes. Drug Metab Dispos 29(9): 1201–1204.
Tang J, Cao Y, Rose RL & Hodgson E (2002) In vitro metabolism of carbaryl by human cytochrome P450 and its inhibition by chlorpyrifos. Chem Biol interact 141(3): 229–241.
Tang J, Tang J, Amin Usmani K, Hodgson E & Rose RL (2004) In vitro metabolism of fipronil by human and rat cytochrome P450 and its interactions with testosterone and diazepam. Chem Biol Interact 147(3): 319–329.
Tang W & Stearns RA (2001) Heterotropic cooperativity of cytochrome P450 3A4 and potential drug-drug interactions. Curr Drug Metab 2(2): 185.
Tassaneeyakul W, Mohamed Z, Birkett DJ, McManus ME, Veronese ME, Tukey RH, Quattrochi LC, Gonzalez FJ & Miners JO (1992) Caffeine as a probe for human cytochromes P450: validation using cDNA-expression, immunoinhibition and microsomal kinetic and inhibitor techniques. Pharmacogenetics 2: 173–183.
Tassaneeyakul W, Guo L, Fukuda K, Ohta T & Yamazoe Y (2000) Inhibition selectivity of grapefruit juice components on human cytochromes P450. Arch Biochem Biophys 378(2): 356–363.
Thomas KV, Hurst MR, Matthiessen P, Sheahan D & Williams RJ (2001) Toxicity characterisation of organic contaminants in stormwaters from an agricultural headwater stream in South East England. Water Res 35(10): 2411–2416.
Thummel K, Kharasch E, Podoll T & Kunze K (1993) Human liver microsomal enflurane defluorination catalyzed by cytochrome P-450 2E1. Drug Metab Dispos 21(2): 350–357.
Tiano HF, Wang R, Hosokawa M, Crespi C, Tindall KR & Langenbach R (1994) Human CYP2A6 activation of 4-(methylnitrosamino)-l-(3-pyridyl)-1-butanone (NNK): mutational specificity in the gpt gene of AS52 cells. Carcinogenesis 15(12): 2859–2866.

107
Tindberg N & Ingelman-Sundberg M (1989) Cytochrome P 450 and oxygen toxicity. Oxygen-dependent induction of ethanol-inducible cytochrome P 450 (IIE1) in rat liver and lung. Biochemistry (N Y ) 28(10): 4499–4504.
Tjia JF, Colbert J & Back DJ (1996) Theophylline metabolism in human liver microsomes: inhibition studies. J Pharmacol Exp Ther 276(3): 912–917.
Tolonen A, Petsalo A, Turpeinen M, Uusitalo J & Pelkonen O (2007) In vitro interaction cocktail assay for nine major cytochrome P450 enzymes with 13 probe reactions and a single LC/MSMS run: analytical validation and testing with monoclonal anti-CYP antibodies. J Mass Spec 42(7): 960–966.
Tolonen A, Turpeinen M & Pelkonen O (2009) Liquid chromatography–mass spectrometry in in vitro drug metabolite screening. Drug Discov Today 14(3-4): 120–133.
Tomlin C (2000) The Pesticide Manual, 12th ed. Berkshire, UK, BCPC Publications. Tremaine LM, Diamond GL & Quebbemann AJ (1985) Quantitative determination of
organ contribution to excretory metabolism. Journal of Pharmacological Methods 13(1): 9–35.
Trevisan MJ, Trevizan LRP, de Baptista GC & Franco AA (2004) Metabolism of carbosulfan to carbofuran and to 3-hydroxy-carbofuran in orange leaves. Rev Bras Toxicol 17(1): 17–22.
Tsao C, Wester MR, Ghanayem B, Coulter SJ, Chanas B, Johnson EF & Goldstein JA (2001) Identification of human CYP2C19 residues that confer S-mephenytoin 4'-hydroxylation activity to CYP2C9. Biochemistry 40(7): 1937–1944.
Turpeinen M, Ghiciuc C, Opritoui M, Tursas L, Pelkonen O & Pasanen M (2007) Predictive value of animal models for human cytochrome P450 (CYP)-mediated metabolism: A comparative study in vitro. Xenobiotica 37(12): 1367–1377.
Turpeinen M, Korhonen LE, Tolonen A, Uusitalo J, Juvonen R, Raunio H & Pelkonen O (2006a) Cytochrome P450 (CYP) inhibition screening: Comparison of three tests. Euro J Pharma Sci 29(2): 130–138.
Turpeinen M, Raunio H & Pelkonen O (2006b) The functional role of CYP2B6 in human drug metabolism: substrates and inhibitors in vitro, in vivo and in silico. Curr Drug Metab 7(7): 705–714.
Turpeinen M, Tolonen A, Chesne C, Guillouzo A, Uusitalo J & Pelkonen O (2009) Functional expression, inhibition and induction of CYP enzymes in HepaRG cells. Toxicol In Vitro 23(4): 748–753.
Turpeinen M, Uusitalo J, Jalonen J & Pelkonen O (2005) Multiple P450 substrates in a single run: rapid and comprehensive in vitro interaction assay. Eur J Pharm Sci 24(1): 123–132.
Tyden E, Tjalve H & Larsson P (2004) Metabolic activation of 2,6-Xylidine in the nasal olfactory mucosa and the mucosa of the upper alimentary and respiratory tracts in rats. Toxicol Sci 81(2): 263–272.
Umetsu N & Fukuto TR (1982) Alteration of carbosulfan [2,3-dihydro-2,2-dimethyl-7-benzofuranyl(di-n-butylaminosulfenyl)methylcarbamate] in the rat stomach. J Agric Food Chem 30(3): 555–557.

108
Urani C, Doldi M, Crippa S & Camatini M (1998) Human-derived cell lines to study xenobiotic metabolism. Chemosphere, 37(14-15): 2785–2795.
USEPA (2000–2001) Pesticide Market Estimates: Usage, http://www.epa.gov/oppbead1/pestsales/01pestsales/usage2001.htm#3_1. viewed May 1, 2010. .
USEPA (2004) Chemicals evaluated for carcinogenic potential. In Anonymous , U.S. Environmental Protection Agency, Health Effects Division, Office of Pesticide Programs. Science information management Branch, p. 22.
USEPA (2005) Fact Sheet: The Drinking Water Contaminant Candidate List - The Source of Priority Contaminants for the Drinking Water Program. Washington DC, U.S. Environmental Protection Agency, Office of Water.
Usmani KA, Hodgson E & Rose RL (2004a) In vitro metabolism of carbofuran by human, mouse, and rat cytochrome P450 and interactions with chlorpyrifos, testosterone, and estradiol. Chem Biol Interact 150(3): 221–332.
Usmani KA, Karoly ED, Hodgson E & Rose RL (2004b) In vitro sulfoxidation of thioether compounds by human cytochrome P450 and flavin-containing monooxygenase isoforms with particular reference to the CYP2C subfamily. Drug Metab Dispos 32(3): 333–339.
Usmani KA, Rose RL, Goldstein JA, Taylor WG, Brimfield AA & Hodgson E (2002) In vitro human metabolism and interactions of repellent N,N-diethyl-m-toluamide. Drug Metab Dispos 30(3): 289–294.
Van Boven M, Laruelle L & Daenens P (1990) HPLC analysis of diuron and metabolites in blood and urine. J Anal Toxicol 14: 231–234.
Veenstra DL, Blough DK, Higashi MK, Farin FM, Srinouanprachan S, Rieder MJ & Rettie AE (2005) CYP2C9 haplotype structure in european american warfarin patients and association with clinical outcomes. Clin Pharmacol Ther 77(5): 353–364.
Venkatakrishnan K, Greenblatt D, von Moltke L, Schmider J, Harmatz J & Shader R (1998a) Five distinct human cytochromes mediate amitriptyline N-demethylation in vitro: dominance of CYP 2C19 and 3A4. J Clin Pharmacol 38(2): 112–121.
Venkatakrishnan K, Moltke LLV & Greenblatt DJ (1998b) Human cytochromes P450 mediating phenacetin O-deethylation in vitro: validation of the high affinity component as an index of CYP1A2 activity. J Pharm Sci 87(12): 1502–1507.
Verheij ER, van der Greef J, La Vos GF, van der Pol W & Niessen WM (1989) Identification of diuron and four of its metabolites in human postmortem plasma and urine by LC/MS with a moving-belt interface. J Anal Toxicol 13(1): 8–12.
Vettorazzi G (1977) Safety factors and their application in the toxicological evaluation. In Hunter WJ & Smeets JGPM (eds) The evaluation of toxicological data for the protection of public health proceedings of the international colloquium Luxembourg, December 1976. Oxford, UK, Pergamon Press: 207–223.
Volotinen M, Turpeinen M, Tolonen A, Uusitalo J, Maenpaa J & Pelkonen O (2007) Timolol metabolism in human liver microsomes is mediated principally by CYP2D6. Drug Metab Dispos 35(7): 1135–1141.

109
von Bahr C, Ursing C, Yasui N, Tybring G, Bertilsson L & Röjdmark S (2000) Fluvoxamine but not citalopram increases serum melatonin in healthy subjects – an indication that cytochrome P450 CYP1A2 and CYP2C19 hydroxylate melatonin. Eur J Clin Pharmacol 56(2): 123–127.
von Moltke LL, Greenblatt DJ, Giancarlo GM, Granda BW, Harmatz JS & Shader RI (2001) Escitalopram (S-citalopram) and its metabolites in vitro: cytochromes mediating biotransformation, inhibitory effects, and comparison to R-citalopram. Drug Metab Dispos 29(8): 1102–1109.
Walle T, Otake Y, Galijatovic A, Ritter JK & Walle UK (2000) Induction of UDP-glucuronosyltransferase UGT1A1 by the flavonoid chrysin in the human hepatoma cell line Hep G2. Drug Metabolism and Disposition 28(9): 1077–1082.
Walsky RL, Gaman EA & Obach RS (2005a) Examination of 209 drugs for inhibition of cytochrome P450 2C8. J Clin Pharmacol 45(1): 68–78.
Walsky RL, Obach RS, Gaman EA, Gleeson JR & Proctor WR (2005b) Selective inhibition of human cytochrome P4502C8 by montelukast. Drug Metab Dispos 33(3): 413–418.
Walton K, Dorne JL & Renwick AG (2001a) Uncertainty factors for chemical risk assessment: interspecies differences in glucuronidation. Food Chem Toxicol 39(12): 1175–1190.
Walton K, Dorne JL & Renwick AG (2001b) Uncertainty factors for chemical risk assessment: interspecies differences in the in vivo pharmacokinetics and metabolism of human CYP1A2 substrates. Food Chem Toxicol 39: 667–680.
Wei C, Caccavale RJ, Kehoe JJ, Thomas PE & Iba MM (2001) CYP1A2 is expressed along with CYP1A1 in the human lung. Cancer Lett 171(1): 113–120.
Wei C, Caccavale RJ, Weyand EH, Chen S & Iba MM (2002) Induction of CYP1A1 and CYP1A2 expressions by prototypic and atypical inducers in the human lung. Cancer Lett 178(1): 25–36.
Wen X, Wang J, Backman JT, Kivisto KT & Neuvonen PJ (2001) Gemfibrozil is a potent inhibitor of human cytochrome P450 2C9. Drug Metab Dispos 29(11): 1359–1361.
Wen X, Wang J, Backman JT, Laitila J & Neuvonen PJ (2002) Trimethoprim and sulfamethoxazole are selective inhibitors of CYP2C8 and CYP2C9, respectively. Drug Metab Dispos 30(6): 631–635.
White RE (2000) High-throughput screening in drug metabolism and pharmacokinetic support of drug discovery. Annu Rev Pharmacol Toxicol 40(1): 133–157.
WHO/IPCS (1994) Assessing human health risks of chemicals: derivation of guidance values for health-based exposure limits. Environmental health criteria, 170. Geneva, World Health Organization.
WHO/IPCS (2005) Chemical-specific adjustment factors for interspecies differences and human variability: guidance document for use of data in dose/concentration-response assessment. Geneva, International Program on Chemical Safety, WHO/UNEP/ILO, World Health Organization.
WHO/UNEP (ed) (1990) Public health impact of pesticides used in agriculture. Geneva.

110
Wijnen PAHM, Buijsch RAMOD, Drent M, Kuipers PMJC, Neef C, Bast A, Bekers O & Koek GH (2007) Review article: the prevalence and clinical relevance of cytochrome P450 polymorphisms. Aliment Pharmacol Ther 26(s2): 211–219.
Williams JA, Ring BJ, Cantrell VE, Jones DR, Eckstein J, Ruterbories K, Hamman MA, Hall SD & Wrighton SA (2002) Comparative metabolic capabilities of CYP3A4, CYP3A5, and CYP3A7. Drug Metab Dispos 30(8): 883–891.
Wing KD, Glickman AH & Casida JE (1983) Oxidative bioactivation of S-alkyl phosphorothiolate pesticides: stereospecificity of profenofos insecticide activation. Science 219(4580): 63–65.
Wing KD, Glickman AH & Casida JE (1984) Phosphorothiolate pesticides and related compounds: Oxidative bioactivation and aging of the inhibited acetylcholinesterase. Pestic Biochem Physiol 21(1): 22–30.
Wolkers J, Witkamp RF, Nijmeijer SM, Burkow IC, de Groene EM, Lydersen C, Dahle S & Monshouwer M (1998) Phase I and phase II enzyme activities in Ringed seals (Phoca hispida): characterization of hepatic cytochrome P450 by activity patterns, inhibition studies, mRNA analyses, and western blotting. Aquatic Toxicol 44(1-2): 103–115.
Worboys PD, Bradbury A & Houston JB (1996) Kinetics of drug metabolism in rat liver slices. II. Comparison of clearance by liver slices and freshly isolated hepatocytes. Drug Metab Dispos 24(6): 676–681.
Worboys PD, Bradbury A & Houston JB (1997) Kinetics of drug metabolism in rat liver slices. III. Relationship between metabolic clearance and slice uptake rate. Drug Metab Dispos 25(4): 460–467.
Wrighton SA, Stevens JC, Becker GW & Vandenbranden M (1993) Isolation and characterization of human liver cytochrome P450 2C19: correlation between 2C19 and S-mephenytoin 4′-hydroxylation. Arch Biochem Biophys 306(1): 240–245.
Yamano S, Nhamburo PT, Aoyama T, Meyer UA, Inaba T, Kalow W, Gelboin HV, McBride OW & Gonzalez FJ (1989) cDNA cloning and sequence and cDNA-directed expression of human P450 IIB1: identification of a normal and two variant cDNAs derived from the CYP2B locus on chromosome 19 and differential expression of the IIB mRNAs in human liver. Biochemistry 28(18): 7340–7348.
Yamazaki H, Inoue K, Hashimoto M & Shimada T (1999) Roles of CYP2A6 and CYP2B6 in nicotine C-oxidation by human liver microsomes. Arch Toxicol 73(2): 65–70.
Yamazaki H, Inoue K, Turvy CG, Guengerich FP & Shimada T (1997) Effects of freezing, thawing, and storage of human liver samples on the microsomal contents and activities of cytochrome P450 enzymes. Drug Metab Dispos 25(2): 168–174.
Yamazaki H, Inui Y, Yun C, Guengerich FP & Shimada T (1992) Cytochrome P450 2E1 and 2A6 enzymes as major catalysts for metabolic activation of N-nitrosodialkylamines and tobacco-related nitrosamines in human liver microsomes. Carcinogenesis 13(10): 1789–1794.
Yasar Ü, Eliasson E, Dahl M, Johansson I, Ingelman-Sundberg M & Sjöqvist F (1999) Validation of methods for CYP2C9 genotyping: frequencies of mutant alleles in a swedish population. Biochem Biophys Res Commun 254(3): 628–631.

111
Yoshitomi S, Ikemoto K, Takahashi J, Miki H, Namba M & Asahi S (2001) Establishment of the transformants expressing human cytochrome P450 subtypes in HepG2, and their applications on drug metabolism and toxicology. Toxicology in Vitro 15(3): 245–256.
Yuan R, Madani S, Wei X, Reynolds K & Huang S (2002) Evaluation of cytochrome P450 probe substrates commonly used by the pharmaceutical industry to study in vitro drug interactions. Drug Metab Dispos 30(12): 1311–1319.
Yuan R, Parmelee T, Balian JD, Uppoor RS, Ajayi F, Burnett A, Lesko LJ & Marroum P (1999) In vitro metabolic interaction studies: experience of the food and drug administration. Clin Pharmacol Ther 66(1): 9–15.
Yun CH, Shimada T & Guengerich FP (1991) Purification and characterization of human liver microsomal cytochrome P-450 2A6. Mol Pharmacol 40(5): 679–685.
Zamek-Gliszczynski MJ, Hoffmaster KA, Nezasa K, Tallman MN & Brouwer KLR (2006) Integration of hepatic drug transporters and phase II metabolizing enzymes: mechanisms of hepatic excretion of sulfate, glucuronide, and glutathione metabolites. Eur J Pharm Sci 27(5): 447–486.
Zanger U, Raimundo S & Eichelbaum M (2004) Cytochrome P450 2D6: overview and update on pharmacology, genetics, biochemistry. Naunyn Schmiedebergs Arch Pharmacol 369(1): 23–37.
Zhang Q & Ding D (2008) The CYP2F, CYP2G and CYP2J subfamilies. In Ioannides C (ed) Cytochrome P450: role in the metabolism and toxicity of drugs and other xenobiotics (issues in toxicology). London, RSC.

112

113
Original Publications
I Abass K, Reponen P, Jalonen J & Pelkonen O (2007) In vitro metabolism and interactions of the fungicide metalaxyl in human liver preparations. Environ Toxicol Pharmacol 23(1): 39–47.
II Abass K, Reponen P, Jalonen J & Pelkonen O (2007) In vitro metabolism and interaction of profenofos by human, mouse and rat liver preparations. Pestic Biochem Physiol. 87(3): 238–247.
III Abass K, Reponen P, Turpeinen M, Jalonen J & Pelkonen O (2007) Characterization of diuron N-demethylation by mammalian hepatic microsomes and cDNA-expressed human cytochrome P450 enzymes. Drug Metab Dispos. 35(9): 1634–1641.
IV Abass K, Reponen P, Mattila S & Pelkonen O (2009) Metabolism of carbosulfan. I. Species differences in the in vitro biotransformation by mammalian hepatic microsomes including human. Chem Biol Interact. 181(2): 210–219.
V Abass K, Reponen P, Mattila S. & Pelkonen O (2010) Metabolism of carbosulfan. II. Human interindividual variability in its in vitro hepatic biotransformation and the identification of the cytochrome P450 isoforms involved. Chem Biol Interact. 185(3): 163–173.
Reprinted with permission from Elsevier (I,II,IV,V) and American society for
Pharmacology and Experimetal Therapeutics (III).
Original publications are not included in the electronic version of the dissertation.

114

A C T A U N I V E R S I T A T I S O U L U E N S I S
Book orders:Granum: Virtual book storehttp://granum.uta.fi/granum/
S E R I E S D M E D I C A
1056. Kastarinen, Helena (2010) Renal function and markers of cardiovascular risk
1057. Nevalainen, Jukka (2010) Utilisation of the structure of the retinal nerve fiberlayer and test strategy in visual field examination
1058. Liukkonen, Esa (2010) Radiologisten kuvien katselussa käytettävien näyttöjenlaatu. Näyttöjen laitekanta, suorituskyky ja laadunvalvonta sekä kuvankatselu-olosuhteet radiologisissa yksiköissä ja terveyskeskuksissa
1059. Ollila, Päivi (2010) Assessment of caries risk in toddlers. A longitudinal cohortstudy
1060. Vähäkangas, Pia (2010) Kuntoutumista edistävä hoitajan toiminta ja senjohtaminen pitkäaikaisessa laitoshoidossa
1061. Saari, Anne (2010) Autonomic dysfunction in multiple sclerosis and optic neuritis
1062. Eriksen, Heidi (2010) Carboxyterminal telopeptide structures of type I collagen invarious human tissues
1063. Gäddnäs, Fiia (2010) Insights into healing response in severe sepsis from aconnective tissue perspective. A longitudinal case-control study on woundhealing, collagen synthesis and degradation, and matrix metalloproteinases inpatients with severe sepsis
1064. Alaräisänen, Antti (2010) Risk factors and pathways leading to suicide with specialfocus in schizophrenia. The Northern Finland 1966 Birth Cohort Study
1065. Oja, Paula (2010) Significance of customer feedback. An analysis of customerfeedback data in a university hospital laboratory
1066. Mustonen, Erja (2010) The expression of novel, load-induced extracellular matrixmodulating factors in cardiac remodeling
1067. Mäkinen, Johanna (2010) Systematic search and evaluation of published scientificresearch. Implications for schizophrenia research
1068. Jartti, Pekka (2010) Computed tomography in subarachnoid haemorrhage. Studieson aneurysm localization, hydrocephalus and early rebleeding
1069. Rantala, Aino (2010) Susceptibility to respiratory tract infections in young men:the role of inflammation, mannose-binding lectin, interleukin-6 and their geneticpolymorphisms
1070. Tanskanen, Päivikki (2010) Brain MRI in subjects with schizophrenia and in adultsborn prematurely. The Northern Finland 1966 Birth Cohort Study

ABCDEFG
UNIVERS ITY OF OULU P.O.B . 7500 F I -90014 UNIVERS ITY OF OULU F INLAND
A C T A U N I V E R S I T A T I S O U L U E N S I S
S E R I E S E D I T O R S
SCIENTIAE RERUM NATURALIUM
HUMANIORA
TECHNICA
MEDICA
SCIENTIAE RERUM SOCIALIUM
SCRIPTA ACADEMICA
OECONOMICA
EDITOR IN CHIEF
PUBLICATIONS EDITOR
Professor Mikko Siponen
University Lecturer Elise Kärkkäinen
Professor Hannu Heusala
Professor Olli Vuolteenaho
Senior Researcher Eila Estola
Information officer Tiina Pistokoski
University Lecturer Seppo Eriksson
Professor Olli Vuolteenaho
Publications Editor Kirsti Nurkkala
ISBN 978-951-42-6298-2 (Paperback)ISBN 978-951-42-6299-9 (PDF)ISSN 0355-3221 (Print)ISSN 1796-2234 (Online)
U N I V E R S I TAT I S O U L U E N S I S
MEDICA
ACTAD
D 1071
ACTA
Khaled M
. Abass
OULU 2010
D 1071
Khaled M. Abass
METABOLISM AND INTERACTIONS OF PESTICIDES IN HUMANAND ANIMAL IN VITRO HEPATIC MODELS
FACULTY OF MEDICINE,INSTITUTE OF BIOMEDICINE,DEPARTMENT OF PHARMACOLOGY AND TOXICOLOGYFACULTY OF SCIENCE,DEPARTMENT OF CHEMISTRY,UNIVERSITY OF OULU