adaptatie van skeletspierweefsel - pure - aanmelden · de maximale kracht is afhankelijk van het...
TRANSCRIPT

Adaptatie van skeletspierweefsel
Citation for published version (APA):Ossevoort, L. (1995). Adaptatie van skeletspierweefsel. (DCT rapporten; Vol. 1995.126). Eindhoven: TechnischeUniversiteit Eindhoven.
Document status and date:Gepubliceerd: 01/01/1995
Document Version:Uitgevers PDF, ook bekend als Version of Record
Please check the document version of this publication:
• A submitted manuscript is the version of the article upon submission and before peer-review. There can beimportant differences between the submitted version and the official published version of record. Peopleinterested in the research are advised to contact the author for the final version of the publication, or visit theDOI to the publisher's website.• The final author version and the galley proof are versions of the publication after peer review.• The final published version features the final layout of the paper including the volume, issue and pagenumbers.Link to publication
General rightsCopyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright ownersand it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights.
• Users may download and print one copy of any publication from the public portal for the purpose of private study or research. • You may not further distribute the material or use it for any profit-making activity or commercial gain • You may freely distribute the URL identifying the publication in the public portal.
If the publication is distributed under the terms of Article 25fa of the Dutch Copyright Act, indicated by the “Taverne” license above, pleasefollow below link for the End User Agreement:
www.tue.nl/taverne
Take down policyIf you believe that this document breaches copyright please contact us at:
providing details and we will investigate your claim.
Download date: 25. Feb. 2019

Adaptatie van skeletspierweefsel
stag ever slag Liesbeth Ossevoort
WFW rapportnummer 95.126
Liesbeth Ossevoort Eindhoven, augustus 1995 Techn isc he Universiteit Eind hoven opleiding Werktuigkundige Medische Technologie Vakgroep WFW Begeleiders: Sander Gielen
Peter Bovendeerd

Inhoud
1. Inleiding. 1.1 Doel van de studie.
1.2 Opbouw wan het verslag.
2. Het 'korte-termijn '-gedrag van de skeletspier. 2.1 Anatomie en fysiologie van de skeletspier.
2.1.1 Anatomie van de skeletspier. 2.1.2 Werking van de skeletspier en factoren die de spierkracht beïnvloeden.
2.2.1 Het modelleren van spierweefseleigenschappen. 2.2.2 Een geometrisch model. 2.2.3 Het DM-model.
2.2.3.1 Inleiding. 2.2.3.2 Het glijdende filamentenmodel. 2.2.3.3 Een interpretatie van het model. 2.2.3.4 Het DM-model.
2.2.4.1 Een uitbreiding van het DM-model.
2.2 Spierrnodellen.
2.2.4 Een dynamisch spiermodel.
3. Het 'lange-termijn '-gedrag van de skeletspier: adaptatie. 3.1 Adaptatie en adaptatie-effecten.
3.2 De oorzaak van adaptatie en mechanotransductie.
3.3 Samenvatting.
4. Het modelleren van adaptatie. 4.1 Het adaptatiemodel.
4.1.1 De stimulus. 4.1.2 De groeiwet. 4.1.3 Keuze van de constanten. 4.1.4 De karakteristieke adaptatietijd.
4.2 De beiacting. 4.2.1 Het belastingspatroon. 4.2.2 De karakteristieke belastingstijd. 4.2.3 Verlenging of kracht als belasting.
4.3.1 Numerieke stabiliteit van de groeiwet. 4.3 Numerieke aspecten.
5. Adaptatie in het geometrisch spiiemodd. 5.1 Simulatie-experimenten.
5.2 Resultaten. 5.2.1 Belastingspatroon 1.
2
4 4
4
4 5 5 5
6 6 6 7 7 7 8 9
10 10
11 12
12
14
14 15 15 16 17 18
18 18 19 20
21 21
23 24
25 25

5.2.2 Belastingspatroon 2. 5.2.3 Belastingspatroon 3.
5.3 Discussie.
6. Adaptatie in het dynamisch spiermodel. 6.1 Inbouwen van het adaptatiemechanisme in het dynamisch spiermodel.
6.2 Keuze van de constanten.
6.3 Srmulatie-exper!meraten.
6.4 Resultaten. 6.4.1 Belastingspatroon 1. 6.4.2 Belastingspatroon 2. 6.4.3 Belastingspatroon 3.
6.5 Discussie.
7. Conclusies en aanbevelingen.
BRQNNEN.
Bijlage Matlabfiles voor het berekenen van spieradaptatie-effecten. Stabiliteitsbepaling.
Berekening van het aantal sarcomeren.
Verlenging als belasting.
Bepaling van de kracht van het contractiele deel.
Bepaling van de participatie-parameter a. De kracht als stimulus.
Hoofdprogramma.
26 28
30
30 31
33
34
35 35 36 37
38
39
42
43 43
45
46
47
50
50
51
3

1. Inleiding.
1.1 Doel van de studie. In deze studie wordt geprobeerd de kennis op het gebied van adaptatie (aanpassing) van skeletspierweefsel in kaart te brengen. Hierbij wordt de nadruk gelegd op die kennis die voor modellering noodzakelijk is: het identificeren van een stimulus die aanzet tot adaptatie en de exacte wijze van adaptatie. Op grond van het gevonden adaptatiemechanisme zal een groeiwet opgesteld worden.Dit is in feite het adaptatiemodel. Vervolgens is deze groeiwet in een spiermodel geïntegreerd.
1.2 Opbouw van het verslag. Allereerst zal in het kort de anatomie van de skeletspier worden beschreven, omdat kennis hiervan noodzakelijk is voor een goed begrip van deze studie. Hetzelfde geldt voor de werking van de spier, die daarna aan de orde zal komen. Vervolgens worden er enkele spiermodellen gepresenteerd die aansluiten bij het beschreven spiergedrag. Deze modellen geven dus het korte-termijngedrag van het spierweefsel weer en brengen adaptatie niet in rekening. In hoofdstuk 3 wordt vervolgens het lange-termijngedrag van spierweefsel, adaptatie besproken. Hierbij wordt gezocht naar stimuli die aanleiding geven tot adaptatie en worden alle mogelijke vormen van adaptatie worden op een rijtje gezet. Na deze kwalitatieve beschrijving van adaptatie wordt een modellering hiervan gepresenteerd. Ook de numerieke aspecten van deze modellering zullen aan de orde komen. Tenslotte zullen we de modellering van adaptatie in twee van de spiermodellen inbouwen. Hiermee worden enkele simulaties uitgevoerd. We besluiten met enkele conclusies en aanbevelingen.
4

2. Het ‘korte-termijn’ggedrag van de s ke I ets p i e r.
2.1 Anatomie en fysio/ogie van de skeletspier.
2.1.1 Anatomie van de skeletspier. Een skeletspier bevat naast spiervezels ook bindweefsel, bloed- en lymfevaten en zenuwvezels. De verschillende componenten van de spier worden bij elkaar gehouden door hulzen van bindweefsel. De spiervezels zelf hebben het uiterlijk van dunne (1 0-1 O0 pm) draadjes, waarvan de lengte enkele millimeters tot tientallen centimeters kan bedragen. iedere vezel bevat enkele honderden tot duizenden intracellulaire vezels: dit zijn myofibrillen en zij vormen het contractiele apparaat van de skeletspiervezel. De myofibrillen zijn op hun beurt weer te ontleden in myofilamenten. Er zijn twee soorten filamenten, dunne actinefilamenten en dikke myosinefilamenten. De myosinefilamenten bestaan uit lichte staven met een scharnierende, zware kop. Contractie komt tot stand doordat actinefilamenten aangetrokken worden door myosinefilamenten en daartussen schuiven. Deze beschrijving van spiercontractie op microscopisch niveau noemt men het ‘glijdende-filamentenmodel’ en dit model zal een grote rol spelen in deze studie. In de lengterichting zijn de myofibrillen geordend in sarcomeren. Door deze ordening is een dwarsstreping zichtbaar. Sarcomeren zijn ongeveer 2 micrometer lang en kunnen als functionele eenheid worden beschouwd. [ I ]
spier . - _-r- ‘. _-
__- - - __--- kernen \ vezelbundei
._ -. -_ -. Z - I I J ~ .----
: H-band 1 A -band i -band - -_
myofiiahenten
I . . ... ... . . . .... .a:*:.. ....... ..... .:::::::. ..... ... : . z . : . . . . ... .... .......
actinefilament
myosinefilament
Figuurlr Structuur van een skeletspier en de rongschik- king var7 de dikke en dunnejlanienter7 in een sarcomeer (genmdijceerdnaar Bloom en Fait - cel) I I I
2.1.2 Werking van de skeletspier en factoren die de spierkracht beïnvloeden. Wanneer in een spier aan diverse voorwaarden voldaan is kan een sterke binding ontstaan tussen de actine- en myosinefilamenten. Zo’n binding wordt een cross-bridge genoemd. Door vorming van cros-bridges kan in het materiaal een stijfheidsverandering optreden. Cross-bridges zijn echter ook in staat de actine en myosinefilamenten langs elkaar te bewegen, zodat spiercontracties op kunnen treden. De snelheid waarmee cross-bridges tot stand komen is bepalend voor de maximale snelheid van krachtstoename. De snelheid waarmee spieren kunnen contraheren wordt voornamelijk bepaald door de snelheid waarmee deze cross-
5

bridges weer los kunnen laten. Het loslaten van cross-bridges is noodzakelijk om ervoor te zorgen dat de filamenten weer verder langs elkaar kunnen worden bewogen door nieuwe cross-bridges te vormen. De snelheid van dit proces wordt bepaald door de snelheid waarmee cross-bridges kunnen loslaten omdat dit de traagste factor is. Er zijn twee typen skeletspieren: slow-twitch spieren die voornamelijk een rol spelen bij houdingshandhaving en fast-twitch spieren die snellere bewegingen mogelijk maken. Ye maximale kracht ei: het vermyen va:: een spier hangen van $en aantal factoren af, te weten: [2] 1. Spiergrootte en -lengte.
De maximale kracht is evenredig met de spierdiameter. De kracht is maximaal bij een spierlengte waarbij de overlap tussen de actine en myosinefilamenten maximaal is. Zo kunnen de meeste cross-bridges gevormd worden.
De maximale kracht is afhankelijk van het aantal cross-bridges dat gevormd wordt en de kracht die per cross-bridge geleverd wordt. Uit experimenten blijkt dat de maximale kracht per cross-bridge constant is. Het aantal cross-bridges dat tegelijk gevormd kan worden kan veranderen.
3. De kracht-snelheidsrelatie en de contractie snelheid. De kracht -snelheidSrelatie is een hyperbool, waarbij de maximale snelheid wordt bereikt wanneer de spier geen kracht hoeft te leveren. In het verloop van de hypgrbool kunnen veranderingen optreden, waarschijnlijk ten gevolge van een veranderde samenstelling van het myosine. Ook de contractiesnelheid kan worden beïnvloed.
De concentratie van de [Ca"]-ionen is van invloed op de hoeveelheid cross- bridges die tot stand kan komen.
2. Aantal cross-bridges en kracht per cross-bridge.
4. De [Ca*+]-concentratie.
2.2 Spiermodellen.
2.2.1 Het modelleren van spierweefseleigenschappen. Het maken van een goed model van de eigenschappen van spierweefsel is niet eenvoudig aangezien spierweefsel zeer complexe eigenschappen heeft. Er bestaan al diverse spiermodellen, ieder met hun eigen mate van complexiteit en hun eigen toepassingsgebied. We zullen in het vervolg drie spiermodellen presenteren: het geometrisch spiermodel, het DM-model en een dynamisch spiermodel. Al deze modellen zijn 1 -dimensionaal. Het geometrisch spiermodel is een uiterst eenvoudig model, waarvan de toepassing erg beperkt is. Het DM-model is een geavanceerd spiermodel, wat de eigenschappen van het contractiele spierweefsel goed beschrijft. Het dynamisch spiermodel is een uitbreiding van het DM-model, waarin ook de passieve componenten van spierweefsel gemodelleerd worden.
2.2.2 Een geometrisch model. Het geometrisch model is een erg eenvoudig model dat gebaseerd is op het aantal sarcomeren dat serieel in een spiervezel geschakeld ligt. Sarcomeren
6

kunnen de meeste kracht leveren als hun lengte tussen de 1.9 en 2.2 micrometer bedraagt, omdat bij deze lengte de overlap tussen actine- en myosinefilamenten optimaal is. Dit houdt in dat er bij deze sarcomeerlengte het grootste aantal cross-bridges gemaakt kunnen worden. [l] Op grond van deze gegevens veronderstellen we dat de sarcomeerlengte bij een spier in rust 2 micrometer is. We kunnen nu op grond van de initiële spierlengte het aantal serieel geschakelde sarcomeren bepalen volgens:
zo no = 7 &arc
met: no initieel aantal sarcomeren IO initiële spierlengte IOSWC optimale sarcomeerlengte Ook kunnen we de sarcomeerlengte bepalen als functie van de verlenging van de spier m.b.v.:
met: 'sa, momentane sarcomeerlengte u verlenging
2.2.3 Het DM-model.
2.2.3.1 Inleiding. Het DM-model is een spiermodel dat geformuleerd is op grond van de microstructuur van het spierweefsel en het is gebaseerd op de glijdende- filamenten theorie die door Huxley ontwikkeld werd [8]. In tegenstelling tot het hierboven beschreven geometrisch model brengt het DM-model de contractiele eigenschappen van spierweefsel in rekening. Inmiddels is er veel onderzoek gedaan naar de toepasbaarheid van het DM- model. Gebleken is dat dit model zeer goed in staat is de eigenschappen van het contractiele spierweefsel te beschrijven en voorspellen.
2.2.3.2 Het glijdende filamentenmodel. Het glijdende filamentenmodel werd in 1957 door A.F. Huxley ontwikkeld en het beschrijft het langs elkaar glijden van actine- en myosinefilamenten tijdens spiercontractie. Het is een zeer ingewikkeld kinematisch model, dat niet geschikt is om geometrisch complexe spiewezelbundels mee te beschrijven omdat dit te veel rekenwerk vergt. Wel is het te gebruiken voor berekeningen aan enkele sarcomeerstrengen. Er liggen een aantal aannamen aan dit model ten grondslag [4]: 1. De lengte van de actine- en myosinefilamenten verandert niet bij rek en
2. De cross-bridges zijn onafhankelijke krachtgeneratoren. 3. Elke cross-bridge kan op ieder willekeurig tijdstip slechts aan één actine-
4. Hoewel bekend is dat er diverse biochemische bindingssterkten van cross-
contractie van de spier.
bindingsplaats gehecht zijn.
bridges bestaan, gaan we ervan uit dat er slechts twee opties zijn: gebonden of ongebonden.
7

5. De actine-myosine bindingsreactie is te beschrijven met eerste orde kinetiek. Hierbij is n(x, t) de verdelingsfunctie van het aantal gebonden cross-bridges met verplaatsing x op tijdstip t. Er wordt verondersteld dat de verdelingsfunctie voldoet aan de volgende vergelijking:
Hierbij is v(t) de verkortingssnelheid van een sarcomeer, f(x) de bindingsparameter, g(x) de ontbindingsparameter en Q/Qt is de materide t ijdsafg el eide.
Een zeer belangrijke aanname in de theorie is dat zowel de parameters fals g functies zijn van x, de afstand tot de evenwichtspositie van een cross-bridge. De trekkracht F(x) per cross-bridge als functie van de verplaatsing xvanuit de neutrale positie kan voorgesteld worden door een polynoom:
We gaan ervan uit dat alleen de eerste orde van belang is, dus F(x)=Kx. F ( x ) = Kx+ax2+...
2.2.3.3 Een interpretatie van het model. Vanuit de microstructuur kunnen we vervolgens diverse macroscopische variabelen interpreteren door de momenten van de verdelingsfunctie te bekijken,
volgens: [8] Mk = In(x,t).x’dx
Aangezien n(x,t) te beschouwen is als de fractie gebonden cross-bridges, met afstand x uit hun evenwichtspositie, levert een integraal over al deze cross- bridges vermenigvuldigd met de stijfheid per cross-bridge een waarde voor de momentane stijfheid van de fibril. Wiskundig is dit als volgt te formuleren:
m
-
K ( t ) = aC,Mo m
Mo = jn(x,t)dn -m
met: K(t) momentane stijfheid 0 participatieniveau C, constante Mo Het participatieniveau geeft aan welk deel van de cross-bridges werkelijk gevormd kan worden, met a tussen O, geen cross-bridgevorming mogelijk, en 1, het maximale aantal cross-bridges kan gevormd worden. Wanneer de sarcomeerlengte niet optimaal is kan slechts een klein deel van de cross-bridges gevormd worden, omdat de overlap tussen actine en myosine maar klein is, in dat geval zal a kleiner zijn dan 1. De waarde van a wordt bepaald in de routine alfam. Hiervoor is gebruik gemaakt van het volgend verloop van a als functie van de sarcomeerlengte:
nulde orde moment van n(x,t)
8

O
Figuur 2: het participatieniveau als functie van de sarcomeerlengte. 111
C, is een constante is die afhangt van de lengte van de sarcomeer, het aantal cross-bridges per volume-eenheid en de cross-bridge stijfheid, volgens:
rn-ssk 2.1
C, =-
waar : k Stijfheid van één cross-bridge m aantal cross-bridges per volume-eenheid S lengte van de sarcomeer I afstand tussen actine-bindingsplaatsen Andere macroscopische variabelen kunnen op gelijke wijze berekend worden met behulp van n(x,t). Zo is het eerste moment van de verdelingsfunctie evenredig met de spanning S(t) die doorgeleid wordt:
S(t> = aC,M, W
Mo = /n(x,t)-xdx - W
De laatste macroscopische variabele die we kunnen bepalen is de hoeveelheid opgeslagen mechanische energie. Hiervoor moeten we het tweede moment van de verdelingsfuntie bekijken:
E ( t ) = aC,M2 W
M2 = /n(x,t).x2 - W
2.2.3.4 Het DM-model. De exacte oplossingen van het hierboven beschreven model zijn slechts met zeer veel rekenwerk te verkrijgen. Dit wordt met name veroorzaakt door de ingewikkelde vorm van de verdelingsfunctie n(x,t). Voor de macroscopische variabelen waarin we geïnteresseerd zijn, zijn alleen de momenten van de verdelingsfunctie van belang. Daarvoor integreren we n(x, t), waardoor de exacte vorm van de verdelingsfunctie er niet meer toe doet. We kunnen ook een eenvoudige benadering van de verdelingsfunctie n(x,t) gebruiken, mits deze benadering dezelfde geïntegreerde waarde, dus hetzelfde oppervlak onder de grafiek heeft.
9

Het blijkt dat als benadering van de verdelingsfunctie een Gaussische kromme goed voldoet [6]. Het gebruiken van de Gaussische kromme heeft als voordeel dat de ingewikkelde verdelingsfunctie n(x,t) vervangen wordt door de eenvoudige vorm n ( p , ~ ) , die exact bekend is. De beschrijving van de Gaussische kromme ziet er als volgt uit:
Wanneer we de werkelijke verdelingsfunctie en de benadering uitzetten ziet dat er als volgt uit:
2 3 4 5 6 - 2 - l '%&
Shortening Stretch
Figuur 3: Werkelijke (getrokken) en benaderde (gestippelde) verdelingsfuncties voor verlenging en verkorfing. [8]
Na deze laatste vereenvoudiging hebben we dus een model dat berust op het glijdende filamentenmodel en waarvan we de momenten beschouwen als de macroscopische variabelen K(t),S(t)en E(t ) . Deze momenten berekenen we door de werkelijke verdelingsfunctie te benaderen door een Gaussische verdeling. Dit is het DM-model en dit model kan gebruikt worden om specifieke spierweefsel eigenschappen te betrekken bij de modellering. De ingangsvariabele van het model, dus de belasting die je oplegt, is de verlengingssnelheid van de spier.
2.2.4 Een dynamisch spiermodel.
2.2.4.1 Een uitbreiding van het DM-model. Om een skeletspier te modelleren hebben we meer nodig dan alleen het DM- model. Dit model brengt immers alleen de interactie tussen cross-bridges in rekening, terwijl een spier uit veel meer componenten bestaat. Voor de modelvorming kunnen we uitgaan van het DM-model als beschrijving van de contractiele component, de overige passieve structuren, zoals bloedvaten, lymfevaten en bindweefselvliezen, kunnen weergegeven worden door een element parallel te schakelen. Dit parallel geschakelde element zou zich net zo moeten gedragen als de genoemde structuren, dat wil zeggen: niet-lineair,
10

anisotroop en visco-elastisch. Voor de eenvoud modelleren we dit element echter als een lineaire veer. We gaan uit van het model zoals weergegeven in figuur 4, waarbij de combinatie van een contractiel en een parallel elastisch element als eenheidselement gezien wordt.
4 c
Figuur 4: De vorm van het element dat we gaan modelleren. C stelt het contractiele gedeelte voor, V stelt het parallel elastisch element voor en B(t) geeft de belasting als functie van de tyd weer.

3. Het ‘lange-ternaijn’-gedrag van de skeletspier: adaptatie.
3.1 Adaptatie en adaptatie-effecten. Onder invloed van de belastingen die op een spier of spiercel werken kunnen er veranderingen optreden die het functioneren van de spier beïnvloeden. De aanpassingen die structureel van aard zijn en op langere termijn gerealiseerd worden kunnen we vangen onder de naam adaptatie.
Adaptatie van de spier kan zich op diverse manieren manifesteren, afhankelijk van de soort en duur van de belasting [3]. Allereerst kunnen er veranderingen van anatomische aard optreden. Zo kan de doorsnede van de spier toenemen, waardoor ook de spierkracht toeneemt. De toename van de doorsnede wordt over het algemeen niet veroorzaakt door toename van het aantal spiervezels, maar door toename van het aantal myof i brillen en myof i lamenten. Zolang er alleen maar sprake is van een toename van de spieromvang veroorzaakt door een vergroting van de al bestaande spiervezels, is er sprake van hypertrofie. Met deze hypertrofie neemt inderdaad de maximale kracht van de spier toe, maar de krachtstoename is niet alleen het gevolg van de hypertrofie. De maximale spierkracht neemt sterker toe dan men op grond van de veranderde spieromvang zou verwachten en bovendien wordt nogal eens een krachtstoename gevonden zonder dat een verandering in spierdiameter is opgetreden. Mogelijk berust de krachtstoename voor een deel op veranderingen van de activering van spieren vanuit het centrale zenuwstelsel, waardoor een betere coördinatie ontstaat. Hierdoor zou het mogelijk kunnen zijn dat men meerdere motor-units (groepen spiervezels) tegelijkertijd activeert. Overigens is het mogelijk dat hypertrofie optreedt zonder dat de omtrek van de gehele spier verandert. De toename van de contractiele eiwitten moet dan gelijke tred houden met een afnemen van het in de spier gelegen vet. Ook is waargenomen dat in een spier gestreefd wordt naar het handhaven van een gelijk aantal sarcomeren per lengte-eenheid. Wanneer een spier langdurig blootgesteld wordt aan rek zal deze daarop reageren door meer sarcomeren in de lengterichting van de vezel aan te maken. Dit adaptatiemechanisme zorgt ervoor dat de sarcomeerlengte optimaal blijft (ongeveer 2 pm). Bij deze optimale lengte is de overlap tussen actine- en myosinefilamenten optimaal, zodat de meeste cross-bridges gevormd kunnen worden en dus ook de meeste kracht geleverd kan worden.[l O] Naast deze anatomische veranderingen kan men ook biochemische veranderingen waarnemen. De concentratie van een aantal enzymen, die betrokken zijn bij de vorming en splitsing van energierijke fosfaten, zal toenemen. Daarnaast treedt een toename van het aantal mitochondriën, de energieleveranciers van de cel, op. [l]
3.2 De oorzaak van adaptatie en mechanotransductie. Adaptatie kan diverse oorzaken hebben, zoals verandering van het biochemisch milieu of het optreden van een mechanische stimulus. Voordat een cel op mechanische stimuli kan reageren met een aanpassing in zijn structuur, is het noodzakelijk dat die mechanische impuls vertaald wordt naar
12

het dialect van de cel. Dit houdt over het algemeen in dat de mechanische kracht omgezet moet worden in een biochemisch signaal. Het principe van het omzetten van het signaal wordt “mechanotransductie” genoemd. In cellen zijn al diverse mechanismen ontdekt waarvan men verwacht dat ze zorgen voor het tot stand komen van mechanotransductie. De belangrijkste van deze mechanismen zijn: [3] 1 . Deformatie-afhan kelij ke ion kanalen.
Afhankelijk van de deformatietoestand van het celmembraan wordt dit meer of
ionkanalen. De ionenconcentratie is van invloed op diverse cellulaire processen en op de contractiliteit.
Bij veranderende deformatiegraad verandert de intercellulaire pH en de snelheid van DNA-synthese. Het regulerend mechanisme tussen deze factoren is nog onbekend, maar aangenomen wordt dat dit proces voor mechanotransductie zorgt.
3. Adenylaat-cyclase (CAMP). Door rek en deformatie wordt de aanmaak van cAMP bevorderd. Dit cAMP reguleert de celactiviteit en de eiwitsynthese en katalyseert de ATP-produktie. (zie fig. 5)
minder permeabel voor ionen door een veranderde doorlââ:baaïheid van de
2. Na+/”-uitwisselaar.
A
I
Figuur 5: Mechanotransductie m.b. v CAMP. Onder invloed van de rek die opgelegd wordt, veranderf de omgeving van de receptor. Hierdoor wordt de aanmaak van cAMP bevorderd. Deze stof reguleerf de celactiviteit en de eiwitsynthese en katalyseerf de A TP-produktie.
13

4. Enzymen. Diverse in het cytoskelet ingebouwde enzymen worden geactiveerd door rek van de celmembraan. Deze enzymen stimuleren tal van cellulaire processen.
Verwacht wordt dat het eerst genoemde mechanisme, het deformatie afhankelijke ionkanaal, vooral actief is bij de regulatie van het celvolume. De overige mechanismen - Na+/"-uitwisselaar, CAMP en enzymen - lijken te zijn betrokken bij adaptat ie processen. Opvallend is dat voor deze kandidaat-mechanotransducers een algemene karakteristiek te geven is. Hiiën zijn ais voigt opgebouwd: i3.i
Ze hebben een molekuul dat functioneel deel uitmaakt van de celmembraan, de receptor. Door deformatie verandert de omgeving van de receptor, waardoor het een katalytische functie krijgt of waardoor het messengermolekulen af gaat scheiden. De katalyse-produkten of de messengermolekulen zetten bepaalde celspecifieke processen in gang. Deze processen kunnen bijvoorbeeld leiden tot versnelde groei of afbraak, verhoogde ATP produktie of veranderingen in de eiwitsynthese. Dit is het begin van de adaptatie.
Op grond hiervan zou je verwachten dat deformatie de aanleiding geeft tot adaptatie. Toch kunnen we niet uitsluiten dat adaptatie ook nog andere stimuli kent, zoals bijvoorbeeld de door te leiden spanning of de energie-inhoud van het weefsel. Voor deze mogelijke stimuli zijn echter nog geen sensoren gevonden.
3.3 Samenvatting. Voor het vervolg van de studie, het modelleren van adaptatie, is het van groot belang inzicht te hebben in de diverse adaptatiemechanismen. We zetten ze daarom hier nog eens op een rij: 1. Aanpassingen in de structuur:
Er kan een toename van het aantal myofilamenten optreden. Hierdoor worden meer sarcomeren parallel geschakeld, wat inhoudt dat er meer cross-bridges gevormd worden. Er kan gestreefd worden naar een optimale sarcomeerlengte, zodat de overlap tussen actine- en myosinefilamenten optimaal is. Hierdoor kan het grootst mogelijke aantal cross-briges gevormd worden.
De aansturing kan effectiever worden. Hierdoor kan ten gevolge van training een beweging na verloop van tijd effectiever uitgevoerd worden.
Er kunnen meer enzymen en rnitochondrih in ket weefsel komen. Dit wijst op een verhoogde ATP-produktie, waardoor het ATP dat benodigd is voor de vorming van cross-bridges sneller en in grotere hoeveelheden aanwezig is. Hierdoor kan de vorming van cross-bridges mogelijk sneller verlopen.
2. Aanpassingen in de sturing:
3. Aanpassingen in energieproduktie:
14

4. Wet modelleren van adaptatie.
4.1 Het adaptatiemodel.
4.1.1 De stimulus. Het voert te ver om alle genoemde adaptatie-effecten in ons model op te nemen, het model zou er slechts onoverzichtelijk door worden. We moeten dus de keuze maken welk adaptatiemechanisme we willen bekijken. De aanpassingen in de sturing laten we buiten beschouwing omdat dit een effect is dat op kortere termijn op kan treden. Ook de aanpassingen in de energieproduktie laten we voorlopig buiten beschouwing, omdat we biochemische effecten niet willen modelleren. Er blijven dus nog twee adaptatiemechanismen over die beide betrekking hebben op een aanpassing in de structuur. Het eerste adaptatiemechanisme is het aanpassen van het aantal serieel geschakelde sarcomeren. De stimulus voor dit adaptatieproces is niet bekend, maar met de volgende redenering is het mogelijk een hypothese op te stellen voor een mogelijke stimulus. Uit paragraaf 3.2 weten we dat er deformatie-gevoelige sensoren bekend zijn in spierweefsel. Wanneer deze deformatie bestaat uit een verlenging of verkorting van de spier zullen de sarcomeren niet meer hun optimale lengte hebben. Volgens Goldspink wordt hierop het aantal seriële sarcomeren aangepast [l O]. We veronderstellen op grond hiervan dat verlenging of verkorting de stimulus is die aanzet tot het aanpassen van het aantal serieel geschakelde sarcomeren. Voor het tweede adaptatiemechanisme, het aanpassen van het aantal parallel geschakelde sarcomeren, is het moeilijker een stimulus te vinden. Wellicht is in dit geval niet de deformatie maar de energie die de spier moet leveren de aanleiding tot adaptatie. De hypothese is dan dat een spiervezel bij een bepaalde door te leiden spanning optimaal functioneert. Indien de spanning die doorgeleid moet worden langdurig te hoog is, zal extra energie door de spier geleverd moeten worden om deze spanning op te kunnen brengen. Het aanmaken van extra parallelle sarcomeren kost ook energie, evenals het in stand houden hiervan, maar tegelijkertijd kunnen deze extra sarcomeren ervoor zorgen dat de spanning die doorgeleid moet worden weer in het optimale gebied terecht komt, zodat daarvoor minder energie nodig is. De kosten van het aanmaken en onderhouden van extra sarcomeren moeten dan afgewogen worden tegen de opbrengsten veroorzaakt doordat de door ie ieiaen spanning weer in het optimale gebied ligt. Op grond hiervan kun je veronderstellen dat de energie die een spier moet leveren een stimulus tot adaptatie kan zijn. Helaas zijn er in spierweefsel nog geen energiesensoren bekend die deze hypothese kunnen ondersteunen. We hebben nu dus twee verschillende hypothesen: 1. De stimulus die aanzet tot adaptatie is een verlenging (of verkorting). Het
adaptatiemechanisme dat actief wordt zorgt voor een aanpassing van het aantal serieel geschakelde sarcomeren, zodat de sarcomeerlengte optimaal blijft.
15

2. De stimulus die aanzet tot adaptatie is de energie-inhoud van de spier. Het hierbij geactiveerde adaptatiemechanisme zorgt voor een aanpassing van het aantal parallele sarcomeren.
Bij dit model zullen we de verlenging als stimulus kiezen, omdat er in spierweefsel deformatie-sensoren bekend zijn. We zullen dus een model gaan bekijken waarin het serieel aantal sarcomeren als functie van de belasting kan variëren.
0.0.2 D e groeiwet. In de vorige paragraaf is gekozen de groeiwet te baseren op het adaptatiemechanisme waarin de sarcomeerlengte optimaal gehouden moet worden, zodat een maximaal aantal cross-bridges gevormd kan worden. De groeiwet beschrijft de relatie tussen de snelheid waarmee het aantal sarcomeren verandert, &/at, de lengte van de sarcomeren, I,,,, de optimale sarcomeerlengte, Zo,, en het aantal sarcomeren, n . Dus: &/at = f(Zsarc -ZosUrc,n). Het is duidelijk dat wanneer de sarcomeerlengte optimaal is, de veranderingssnelheid van het aantal sarcomeren nul moet zijn. We nemen aan dat de aanpassingssnelheid, &/at groter moet zijn naarmate de afwijking groter is. Verder kun je veronderstellen dat er een maximum aan de aanpassingssnelheid zal zitten, zodat &/at niet onbeperkt groter kan worden. Dit is biologisch zeer waarschijnlijk omdat het aanmaken of afbreken van sarcomeren tijd kost en dit dus niet oneindig snel kan gaan. Als de deformatie te groot wordt zal het weefsel kapot gaan zodat er helemaal geen adaptatie meer op kan treden. We bouwen dit gegeven niet in in het model, maar bekijken zelf of de sarcomeerlengte niet te groot wordt. We zullen nu eerst een algemene vorm voor de groeiwet formuleren. Dit kan op de volgende wijze:
met: ' a de karakteristieke adaptatietijd Isarc de momentane sarcomeerlengte 'ûsurc de optimale sarcomeerlengte n het momentane aantal sarcomeren Hier geeft z, de karakteristieke adaptatietijd weer, dit is de tijd die nodig is om één nieuw sarcomeer aan te maken of er een af te breken. De inverse van za, die je in de formule aantreft, geeft dan dus het aantal sarcomeren weer dat per tijdseenheid aangemaakt of afgebroken kan worden. De functie g(n) kan aangeven dat het bijmaken van nieuwe sarcomeren eenvoudiger wordt naarmate er initieel meer sarcomeren aanwezig zijn. De functie h(Zsarc -Zo,,,) tenslotte zorgt ervoor dat er alleen adaptatie optreedt als lSa, f Zosarc, dus wanneer de sarcomeerlengte niet optimaal is.
We zullen twee verschillende groeiwetten met elkaar vergelijken, zodat we kunnen zien wat de invloed van de exacte vorm van de groeiwet is op het adaptatiegedrag. Als groeiwetten hebben we gekozen voor een arctangensrelatie en een 3"-machts polynome vorm, waarbij opgemerkt moet
16

worden dat de arctangensrelatie een boven- en ondergrens heeft, waarvan we veronderstelden dat dit ook in werkelijkheid het geval zou zijn. De twee groeiwetten zien er als volgt uit:
met. I I I C L .
pi, , constanten 'ref referentie-aantal sarcomeren Wanneer we deze functies in een grafiek uitzetten geeft dit het volgende resultaat:
vorm van de groeiwetten
-2.5 -2 -1.5 -1 -0.5 O 0.5 1 1.5 2 2.5 Isarc-IOsarc
Figuur 6: Vorm van de groeiwetten. De arctangensrelatie verloopt steil in de oorsprong en vlakt dan af. BO de derde-machtsrelatie is het net andersom: Hg loopt vlak in de oorsprong en neemt daarbuiten in steilheid toe.
Op de horizontale as staat de sarcomeerlengte minus zijn optimum, Is,, -ZOsarc , uit, op de vertikale as de groeisnelheid, dn/dt. Het is uit deze figuur direkt te zien dat de groeisnelheid door de arctangensrelatie beperkt wordt tot een maximum. Bij de polynome functie ontbreekt dit maximum, waardoor de groeisnelheid in het model zeer groot kan worden. Wat betreft het verschil tussen de beide groeiwetten verwachten we dat de arctangensrelatie kleine afwijkingen van het optimum snel wegwerkt, terwijl de derde-machtsrelatie dan erg langzaam is. Wanneer de afwijkingen van het optimum groot zijn, - dit is wanneer de sarcomeerlengte groter is dan ongeveer 3 micrometer-, is het juist omgekeerd: de derde-machtsrelatie zal sneller reageren dan de arctangensrelatie.
4.1.3 Keuze van de constanten. Om numeriek met het model uit de voeten ie kunnen moeten er waarden voor de constanten ingevuld worden in de twee groeiwetten:
17

We willen er bij de arctangensrelatie voor zorgen dat we de maximumgroeisnelheid bereikt hebben wanneer de sarcomeerlengte 3,5 micrometer is. Wanneer de sarcomeerlengte ongeveer 4 micrometer is gaat het
bereikt hebben. Dit kan door een geschikte keuze voor B, te doen. Het blijkt dat p, = 5n/1.5- [m-l] een goede keus is. De factor n: schaalt de arctangens zodanig dat de maximale waarde 1 is. Voor de waarde van de karakteristieke adaptatietijd kiezen we za = 1/10, een keuze die in de volgende paragraaf uitgelegd zal worden. Tenslotte moeten we nog een keuze maken voor p2. Hiervoor eisen we dat beide groeiwetten dezelfde waarde hebben als er 5000 sarcomeren zijn en de sarcomeerlengte 3 micrometer is (zie figuur 5). We vinden dan dat geldt:
Sai-cûmeei namelijk kapot 2:: we willen :'m: die tijd de rn3xixale grceisnelheid
p, = 9 . 7 9 . 1 0 ~ [ ~ - ~ ] .
4.1.4 De karakteristieke adaptatietijd. De karakteristieke adaptatietijd, za, geeft een indicatie van de tijd die nodig is om één sarcomeer te maken of af te breken. Het adaptatieproces, de vorming en/of afbraak van sarcomeren, heeft waarschijnlijk een tijdschaal van enkele uren tot meer dan een week. Hierover zijn nog weinig betrouwbare fysiologische gegvens beschikbaar. We veronderstellen dat het aanmaken c.q. afbreken van sarcomeren minstens enige uren in beslag neemt. Daarom kiezen we zu = 1/10. Dit houdt in dat er per tijdseenheid tien sarcomeren bij kunnen komen of af kunnen gaan. Wanneer we als tijdseenheid één dag kiezen, kunnen we het model zo interpreteren dat we een sarcomeergroeisnelheid van +/- tien sarcomeren per dag hebben.
4.2 De beiasting.
4.2.1 Het belastingspatroon. Het is modelmatig mogelijk diverse belastingen en belastingspatronen op te leggen, zoals zaagtandbelastingen, blokbelastingen of sinusvormige belastingen. Ook ingewikkelder belastingsvormen zijn te modelleren wanneer de rekencapaciteit geen problemen oplevert. Een sinusvormige belasting lijkt erg praktisch voor het modelleren van spierbewegingen, aangezien veel dagelijkse bezigheden een bij benadering sinusvormige inspanning van de spieren vereisen. Eventuele trainingseffecten kunnen in rekening gebracht worden door een trend op het belastingspatroon te zetten. Er kan ook voor gekozen worden niet iedere individuele beweging te modelleren, maar het effect te middelen door bijvoorbeeld een daghacht-ritme te modelleren of een integrator in het model in te bouwen. Bij dit model zullen we een aantal belastingspatronen bekijken, die allemaal zijn geënt op een sinusoïde namelijk:
18

2. b(t)=Asin -t + B (? 1 3. b(t)=Asin -t +Bt (: 1
met : b(t): A: B: constante. 7,: karakteristieke belastingstijd. In het eerste geval betreft het een spier die aan symmetrische wisselbelasting wordt blootgesteld. We verwachten dat er geen adaptatie optreedt aangezien de netto belasting gelijk is aan nul. In het tweede geval zorgt de invloed van de factor B ervoor dat de belasting niet puur symmetrisch is. Hierdoor zal er een netto-belasting ontstaan waardoor adaptatie optreedt. Aangezien de waarde van B constant is verwacht je dat na verloop van tijd het aantal sarcomeren weer optimaal is, waardoor vanaf dat moment geen adaptatie meer op zal treden. Aangezien bij adaptatie volgens beide groeiwetten de groeisnelheid kleiner wordt naarmate het aantal sarcomeren dichter bij het optimale aantal komt, verwacht je dat de curve afvlakt naarmate het aantal sarcomeren dichter bij het optimum komt. Bij het derde belastingspatroon is een trend aan de sinus toegevoegd, een constante stijging of daling van het verlengingsniveau. Je zou dit kunnen vergelijken met de invloed van steeds zwaardere , -of lichtere-, trainingen. Er zal adaptatie optreden die vooral na verloop van tijd bepaald zal worden door Bf.
de belasting als functie van de tijd. de belastingsamplitude van de sinus.
4.2.2 De karakteristieke belastingstijd. De tijdsschaal waarop de belasting varieert is vrij klein vergeleken bij de karakteristieke adaptatietijd, en wel in de orde van enkele seconden. Wanneer je alle afzonderlijke bewegingen wil simuleren zou je voor een realistische simulatie enkele miljoenen bewegingscycli moet simuleren, waarvoor erg veel rekencapaciteit nodig is. Wel kun je proberen inzicht te krijgen in de invloed van bepaalde effecten, zoals bijvoorbeeld de vorm van de groeiwet, door de tijdschalen aan te passen. Als je zorgt dat A/& groter kan worden dan biologisch realiseerbaar is, is het mogelijk snel simulaties uit te voeren die inzicht geven in het proces. Voor een realistische simulatie waarbij ook de tijdsschalen in acht genomen worden, is het echter nodig het belastingsignaal te middelen of te integreren. Biologisch lijkt dit niet vreemd: de sensor die de belasting registreert zou best een integrerende functie kunnen hebben. In dit model zullen we op eenvoudige wijze het netto-effect van de belastingen in rekening brengen. Hiervoor veronderstellen we dat de belasting overdag groter is dan I s nachts, en ‘middelen’ we alle bewegingen over 24 uur. We krijgen nu een repeterend belastingspatroon met een periode van één dag. We kunnen dus voor de karakteristieke belastingstijd z,, die de periode van de sinusoïde aangeeft, z, = 1 kiezen. Het feit dat we niet alle bewegingen afzonderlijk
19

beschouwen heeft geen invloed: aangezien aanpassingen in spierweefsel traag plaats vinden kan het weefsel niet reageren op deze snelle belastingswisselingen. De waarde van de karakteristieke adaptatietijd za zorgt dat ook modelmatig de aanpassing traag verloopt.
4.2.3 Verlenging of kracht als belasting. We hebben twee mogelijkheden om de belasting voor te schrijven, als kracht of als verlenging. Voor het adaptatiemodel, de groeiwet, maakt het niets uit welke van de twee je voorschrijft zolang je het aantai sarcomeren, n, en de momentane sarcomeerlengte, /sart,, kunt berekenen. De vorm van de belasting speelt pas een rol wanneer het adaptatiemodel in een spiermodel ingebouwd wordt. Zo is het geometrisch model geformuleerd met behulp van de verlenging, waardoor het het eenvoudigst is als je in dat geval de belasting een verlenging laat zijn. Bij het DM-model en het dynamisch spiermodel wordt gebruik gemaakt van een verlengingssnelheid als input van het model. Hier geldt dus ook dat het voorschrijven van de belasting als verlenging het eenvoudigst is, aangezien via de verlenging eenvoudig de verlengingssnelheid berekend kan worden. Wanneer we toch graag de kracht willen voorschrijven als belasting voor het spierweefsel stuiten we op een aantal problemen. Ten eerste is het niet direkt mogelijk om te berekenen welk deel van de kracht door het contractiele deel van de spier en welk deel door het parallel elastisch element doorgeleid wordt. Dit komt doordat de stijfheid van het contractiele weefsel steeds verandert. Ten tweede moet je aannamen doen over het dwarsdoorsnede-oppervlak van de spier die gesimuleerd wordt, met name over de verhouding tussen de hoeveelheid contractiel en parallel elastisch weefsel. Het eerste probleem hebben we als volgt opgelost: We schrijven de kracht, Ft, voor op ieder tijdstip. Op grond van de kracht schatten we de verlenging, us. Met deze verlenging als ingangsvariabele wordt het model doorgerekend (dit is eigenlijk niet goed, we zouden hier de verlengingssnelheid hebben moeten berekenen). Uit deze berekening vind je de kracht í s die veroorzaakt wordt of nodig is om een verlenging us te bewerkstelligen. Door een vergelijking van Ft en Fs, de werkelijke en geschatte kracht, kan de schatting voor de verlenging us aangepast worden met behulp van de Newton-Rhapson iteratiemethode. Dit proces gaat net zo lang door totdat de werkelijke en geschatte kracht vrijwel gelijk zijn aan elkaar (+/- een tolerantie). Je kent dan op één bepaald tijdstip de uitwijking ten gevolge van de opgelegde kracht. Voor ieder volgend tijdstip wordt de iteratie herhaald. Zo kan voor ieder willekeurig tijdstip de verlenging als functie van de opgelegde kracht bepaald worden. Het voordeel van deze methode is tevens dat de bestaande programmering voor berekening van het DM-model niet aangepast hoeft te worden, omdat gerekend wordt met een uitwijking als ingangsvariable. In het DM-model wordt gerekend met een uitwijkingssnelheid als ingangsvariabele, maar doordat we met een sinusvormig belastingspatroon werken is de uitwijking slechts een constante afwijkend van de uitwijkingssnelheid. Het tweede probleem is op te lossen door een juiste keuze voor enkele constanten die in het model een rol spelen. We komen hier nog uitgebreid op terug in paragraaf 6.2. We kunnen nu dus zowel de kracht als de verlenging als belasting opleggen.
20

4.3 Numerieke aspecten.
4.3.1 Numerieke stabiliteit van de groeiwet. Wanneer een continu stabiel proces gediscretiseerd wordt, wat bij een numerieke benadering altijd gebeurt, loop je altijd het gevaar dat het proces instabiel wordt. [7] We kunnen op eenvoudige wijze aantonen dat het continue proces glad en monotoon verloopt. Hiervoor kiezen we de verlenging u gelijk aan nul en beginnen we met een niet-optimaal aantal sarcomeren,n=5050. We zien in de onderstaande figuur aai het äaiîtai saïcûiwi'si: i:: de tijd g!sd en monotoon naar de evenwichtssituatie convergeert.
aantal sarcomeren I I
. . . . . . : . . . . . . 1 . . . . . . . . .-
. . . . . . . . . . . . . ...;. . . . . . . .-
. . . . . . . . . . . . . . . . . . . . . . . -
I I I I I I I I I
tijd in dagen 10 20 30 40 50 60 70 80 90 100 50006
Figuur 7: Hef aantal sarcomeren b$ constante belasting, ni=5050. Om te onderzoeken of onze groeiwet ook stabiel gedrag vertoont wanneer hij gediscretiseerd wordt is het nodig de groeiwet te discretiseren en de gelineariseerde vorm ervan te berekenen. Op deze gelineariseerde vorm zijn dan stabiliteitscriteria van toepassing [7]. Voor de Euler-voorwaarts gediscretiseerde vorm van de arctangensrelatie vinden we het volgende:
Met n,+l = F(n) is dit als volgt te lineariseren:
F' (n ) = -- 2 ~t .,,,,,(~i ( l - ~ ~ a r c 'j - (2/nnref)P 1 ln(At/za ) +1 n n r e f ' a P:(z2 - 2 / n ~ ~ ~ ~ ~ ~ +n21&rc)+n2
Aangezien dit een gelineariseerde vorm is, moet voordat er een stabiliteitsanalyse plaatsvindt een werkpüni gekozen worden, met andere woorden: Er kan geen algemene uitspraak over de stabiliteit van het systeem gedaan worden, we kunnen alleen de stabiliteit in bepaalde werkpunten bepalen. De groeiwet gedraagt zich stabiel als de waarde van de gelineariseerde vorm F ' ( n ) tussen de -1 en 1 ligt in de gekozen werkpunten. Door de werkpunten verstandig te kiezen kunnen we uiteindelijk een uitspraak doen over de stabiliteit in een bepaald gebied. Wanneer we uitgaan van een spier met een initiële lengte van 1 centimeter met nref =5000 sarcomeren serieel geschakeld, eisen we dat de routine stabiel is wanneer het aantal sarcomeren tussen de 4000 en 6000 ligt en de totale spierlengte tussen de 0,9 en 1,1 centimeter. Dit houdt in dat we eisen dat de routine zich stabiel gedraagt wanneer we rekken tot 10% opleggen.
21

Om te onderzoeken of dit het geval is, is de routine facn3d.m in matlab geschreven. Uit de analyse blijkt dat de routine op het vereiste gebied maar beperkt stabiel is, zie de onderstaande figuur:
z 6000 L a5500
2 5000-
5 4500-
2 4000-
Eo a m .c. K
Stabiliteit bij I=1,1 cm. Stabiliteit bij 1=0,9 cm.
-
- 1
I A 60 80 3500; 20 40 60 80
deltat 35000 20 40
deltat
Figuur 8: Stabiliteit van de arctangensrelatie als groeiwet. De factor (dnref) is bij de berekening betrokken.
Het blijkt dat de routine bij een spierlengte van 1.1 cm., dus bij een rek van lo%, al instabiel is vóórdat het theoretisch evenwichtspunt, n=5500 sarcomeren bereikt is. Nadere analyse maakt duidelijk dat de instabiliteit in het werkgebied veroorzaakt wordt door de factor n/nref. Wanneer we deze factor weglaten uit de groeiwet, zodat die er als volgt uitziet:
dn 1 TC 1 - = - - arctan( PI (- - zosarc >> at 7" 2 n
u
krijgen we het volgende beeld wat betreft de stabiliteit.
z 6000 1 Stabiliteit bij I=1,1 cm.
r-----l
2 5000
% 5500
2 4000 I a,
E a m 54500 1 c K
5
Stabiliteit bij 1=0,9 cm
r-----7
I 1 40 60 80 3500; 20 40 60 80
deltat 35000 20
deltat
Figuur 9: Stabiliteit van de arctangensrelatie, met (n/nref)= I , als groeiwet.
Uit de figuren blijkt dat de routine in heel het vereiste gebied stabiel is. Het weglaten van n/n, houdt in dat je kiest voor g(n) = 1. We veronderstellen daarmee dat de snelheid waarmee sarcomeren bijgemaakt of afgebroken worden niet beïnvloed wordt door het aantal sarcomeren dat serieel geschakeld is. Dit is geen vreemde veronderstelling, aangezien een sarcomeer niet kan voelen hoeveel sarcomeren er nog achter hem liggen.
22

Wel is het logisch te veronderstellen dat het aantal parallelle sarcomeerstrengen de aanmaaksnelheid beïnvloedt. Deze parallelle strengen kunnen immers de door te leiden spanning onderling zo verdelen dat er één of enkele sarcomeerstrengen ontlast worden. Hierdoor zou het aanmaken van nieuwe sarcomeren eenvoudiger kunnen worden. Wij beschouwen echter het één-dimensionale geval, zodat het aantal parallelle sarcomeerstrengen geen rol speelt. Hierdoor wordt de keuze g(n)= 1 gerechtvaardigd en kunnen we de bovenstaande groeiwet gebruiken.
2 6000.
5500-
2 5000-
3 4500-
z 4000. 3500
5 VI c !=
We willen O& de stabiliteit van de derde-rnachtsgroeirelatie bepalen. De discrete vorm van deze groeiwet is:
Ook hiervan moeten we de gelineariseerde vorm bepalen. Dit is:
Ook van deze routine kunnen we de stabiliteit in een figuur bekijken. Dit levert het volgende resultaat op:
Stabiliteit bij 1=0,9 cm.
f 6000 I 2 5500
A 1 5 -0.25
20 40 60 80 deltat deltat
Figuur 1 O: Stabiliteit van de derde-machtsrelatie als groeiwet. De routine is stabiel voor - 1 <F'(n)< 1.
Uit de figuur blijkt dat de routine altijd stabiel is zolang At kleiner dan 40 blijft. We kunnen deze routine dus ook gebruiken om adaptatie-effecten te berekenen.
23

5. Adaptatie in het geometrisch spiermodel.
5.1 Simulatie-experimenten. Voor de initiële lengte van de spier kiezen we 1 cm. Dit houdt in dat er in het ideale geval 5000 sarcomeren serieel geschakeld zijn. De belasting die we opleggen is een verlenging en deze mag maximaal +/-lo% van de oorspronkelijke spierlengte bedragen, dus maximaal +/-I mm. De adaptatiesnelheid bedraagt +/- 10 sarcomeren per dag, dus paragraaf 4.2.1 . besloten, leggen we de volgende belastingssituaties op:
= 1/10. Zoals in
1. u f t ) = A sin[: t )
2. u(t)=Asin -t + B [: 1 3. u(t)=Asin -t +Bt [Y I
met : u(t): A: B: constante. T~ :
de verlenging als functie van de tijd. de verlengingsamlitude van de sinus.
karakte rist ie ke belast ingst ijd .
De experimenten worden met beide groeiwetten uitgevoerd, zodat de verschillen tussen de groeiwetten zichtbaar worden. Als responsie bekijken we de variabelen die met behulp van de groeiwet aangepast worden, dus de sarcomeerlengte en het aantal sarcomeren. De waarde van de parameters A en B is niet bij alle experimenten hetzelfde. Om het overzicht te bewaren zullen we deze waarden niet heir, maar bij de resultaatbespreking van de experimenten vermelden. We zullen het aantal sarcomeren, n, niet afronden op gehele getallen. Dit heeft drie redenen. Ten eerste voer je een extra discretisatie uit door n af te ronden, waardoor er meer kans op instabiliteiten is. Ten tweede is het vormen van nieuwe sarcomeren een continu proces, waarbij het moeilijk is de grens te trekken tussen een sarcomeer dat af is, en een dat nog bezig is gevormd te worden. Ten derde bestaat een spiervezel uit een heleboel sarcomeerstrengen met allemaal een net verschillende lengte en een verschillend aantal sarcomeren. Het aantal seriële sarcomeren in het model kan dan ook opgevat worden als het gemiddelde aantal sarcomeren per sarcomeerstreng. De berekeningen zullen worden uitgevoerd in Matlab met behulp van de programma’s die in de bijlage vermeld staan. Voor berekeningen met de arctangensrelatie gebruiken we het programma sarc0.m om het aantal sarcomeren als functie van de verlenging te berekenen. In het geval van de derde-rnachtsrelatie gebruiken we hiervoor het praograrnrna sarc.m. In het programma test.m wordt de belasting voorgeschreven en de tijdsduur van de simulatie. Dit programma is het hoofdprogramma voor simulaties in het geometrisch model. Wanneer je het programma draait krijg je de sarcomeerlengte en het aantal sarcomeren als functie van de belasting te zien.
24

5.2 Resultaten. We zullen de responsie op de belasting per belastingspatroon beschouwen.
L a, & 5000,02 E
g m o
5.2.1 Belastingspatroon 1. Allereerst bekijken we de zuiver sinusvormige belasting waarbij het evenwichtspunt van de sinus O is. Hierbij zijn de volgende waarden ingevuld voor de constanten: A = 5 - At = O.l[s] O<t<10 De responsie die de arctangensrelatie leverde zag er als volgt uit:
[m] dit is een rek van 5%
. . . . . . . . . . . . . . . . . . . .:. . . . . . . . . . . . . . . . . . . -
5000-
aantal sarcomeren 5002 1
5001.5 $ E 5001
2 5000.5
5000
g m (u S c
I I 4999.5‘ I
O 5 10
sarcomeerlengte 2.1
a,
c a,
52.05
- z 2 E s VJ 1.95
1.9 O 5 10
tijd in dagen tijd in dagen
Figuur I I: Responsie op zuivere sinusbelasting met arctangensrelatie. De sarcomeerlengte varieert tussen de 1,9 en 2, I micrometer. Dit is dus een verschil van +/- 5% ten opzichte van de uitgangssituatie wat overeenkomt met de rek van +/- 5% die wordt opgelegd. In de figuur ligt de top iets onder de 2, I micrometer. Dit wordt veroorzaakt door de gekozen tijdsstap.
De trend die zichtbaar is op de responsie is een inschakelverschijnsel. De derde-machtsrelatie gaf op deze zelfde belastingssituatie de volgende responsie te zien:
aantal sarcomeren
- 5000.04/
t - 2 4999.98 m m
4999.96}. . . . . . . . . . . . . . . . . . . .j. . . . . . . . . . . . . . . . . . .
O 5 10 n ‘“O 5 10 tijd in dagen tijd in dagen
Figuur 12: Responsie op zuivere sinusvormige belasting met derde- machtsrelatie. Ook hier varieert de sarcomeerlengte tussen de 1,9 en 2,l micrometer.
25
Uit deze figuren blijkt duidelijk dat de arctangensrelatie een snellere aanpassing van het aantal sarcomeren tot gevolg heeft dan wanneer de groeiwet in de vorm van een derde-machtsrelatie geformuleerd wordt. Dit komt overeen met onze verwachtingen: Aangezien de sarcomeerlengte kleiner dan 3 micrometer blijft, dus relatief dicht bij het optimum, is de arctangensrelatie het snelst in de aanpassing van het aantal sarcomeren.
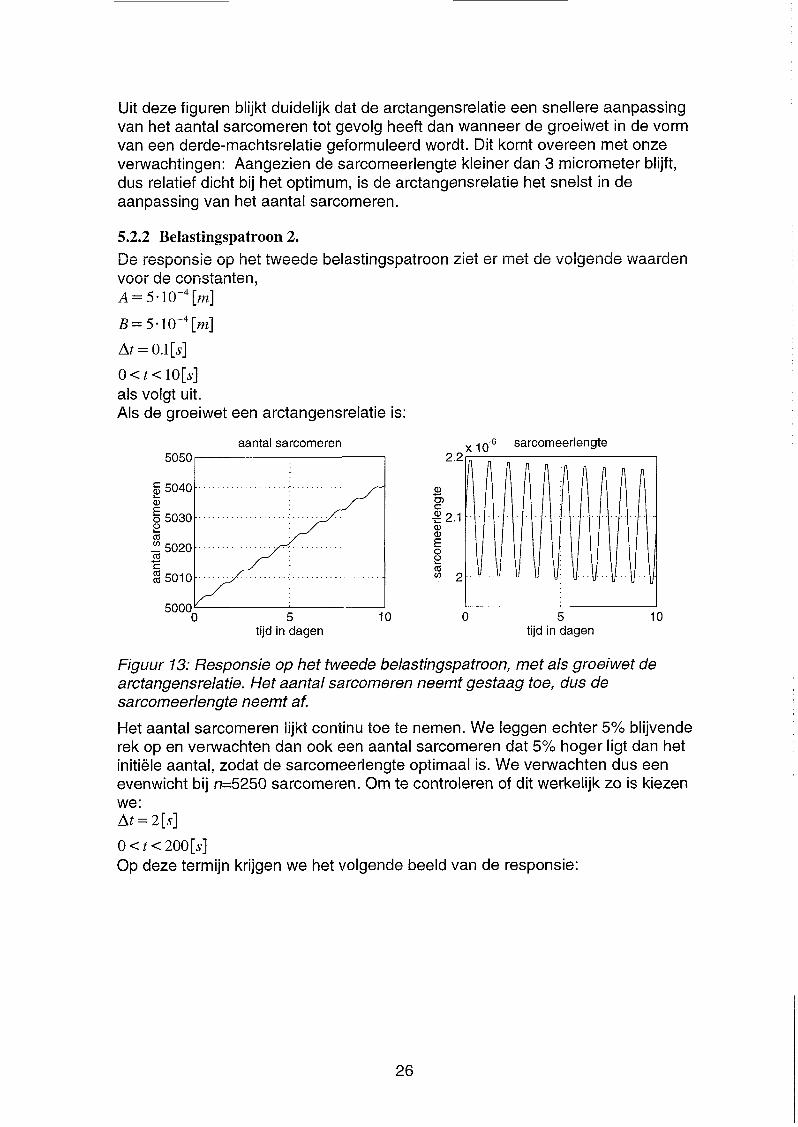
Figuur 13: Responsie op het tweede belastingspatroon, met als groeiwet de arctangensrelatie. Het aantal sarcomeren neemt gestaag toe, dus de sarcomeerlengte neemt af.
Het aantal sarcomeren lijkt continu toe te nemen. We leggen echter 5% blijvende rek op en verwachten dan ook een aantal sarcomeren dat 5% hoger ligt dan het initiële aantal, zodat de sarcomeerlengte optimaal is. We verwachten dus een evenwicht bij n=5250 sarcomeren. Om te controleren of dit werkelijk zo is kiezen we: At = 2[s]
o < t < 200[s] Op deze termijn krijgen we het volgende beeld van de responsie:
26

/
o-6 sarcomeerlengte 2.1
50 100 150 200 tijd in dagen tijd in dagen
Figuur 14: lange termijn responsie op belastingspatroon 2. Bij een blijvende rek van 5% wordt het aantal sarcomeren ook 5% hoger tot n=5250.
Figuur 14 laat zien dat het aantal sarcomeren inderdaad stabiliseert op n=5250. In het geval dat de groeiwet een derde-machtsrelatie is,duurt het veel langer voor evenwicht bereikt is. Dit is onderstaande figuur te zien:
aantal sarcomeren o-6 sarcomeerlengte
. . . . . . ._ . . . . . . . .
O 50 100 150 200 O 50 100 150 200 tijd in dagen tijd in dagen
Figuur 15: Responsie op belastingspatroon 2 met een derde-machtsrelatie.
Uit verdere berekeningen blijkt dat pas bij t=400.000 het evenwichtspunt wordt benaderd. Het aantal sarcomeren is dan namelijk gegroeid tot ongeveer n=5200. De curve is dan echter al zo afgevlakt dat het evenwichtspunt pas veel later bereikt zal worden. Dit is in onderstaande figuren te zien.
27

aantal sarcomeren 5250 I
, o-6 sarcomeerlengte 2.1 I
I I" """V -."- I
21 1 2 3 4 O 1 2 3 4
5000 O
tijd in dagen tijd in dagen
Figuur 16: Met de derde-machtsrelatie wordt het evenwichtspunt slechts langzaam bereikt.
5.2.3 Belastingspatroon 3. Om de laatste belastingssituatie te bekijken hebben we de volgende constanten ingevuld: A = [m]
B= 2.5-10-~
O<t<lO[s]
At = O.l[s]
De waarde van B is met opzet vrij klein gekozen, omdat dit een waarde is die per dag bij de belasting opgeteld wordt. Na twintig dagen is de totale bijdrage van de factor B- ta l 5% rek ten opzichte van de beginsituatie. De responsie die de arctangensrelatie leverde ziet er als volgt uit:
tijd in dagen tijd in dagen
Figuur 17: De responsie op de sinusvormige belasting waarop een trend gesuperponeerd is. Als groeiwet is de arctangensrelatie gebruikt.
De responsie van de derde-machtsrelatie ziet er als volgt uit:
28

aantal sarcomeren 10-6 sarcomeerlengte I I . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
% 5000.02 . . . . . . . . . . . . . . . . .
2 5000 . . . . . . . . . . . . . .
c a,
o (II v) - (II f 4999.98 $
i 4999.96 1 O 5 10
tijd in dagen O 5 10
tijd in dagen
Figuur 18: De responsie op dezelfde belastingssituatie als in figuur 15, nu met de derdemachtsrelatie als groeiwet.
Nadat het inschakelverschijnsel uitgewerkt is gaan de curven inderdaad op B - t lijken. Ook blijkt uit de figuren nog eens dat de arctangensrelatie sneller zorgt dat het optimale aantal sarcomeren gevormd wordt. In geen van bovenstaande gevallen werd de afwijking van het optimale aantal sarcomeren zo groot dat de derde-machtsrelatie getest kon worden op zijn snelheid in het gebied waar de sarcomeerlengte groter is dan 3 micrometer. Daarom leggen we tenslotte nog een verlenging op van 75%. De spierlengte is dan 1,75 cm.. Met een initieel aantal sarcomeren van 5000 levert dit een sarcomeerlengte van 3,5 micrometer, waardoor we in het gebied komen waar de derde-machtsrelatie sneller zou moeten reageren dan de arctangensrelatie. De responsie op de hierboven beschreven belasting ziet er met de arctangensrelatie als groeiwet als volgt uit:
aantal sarcomeren o-6 sarcomeerlengte 51 O0 3.6
5080- a, m c F 8 3.5 %
o 2 3.4
- a,
(II rn
3.3 O 5 10 O 5 i o
tijd in dagen tijd in dagen
Figuur 19: Responsie op een zeer grote uitwijking, u=O. 75 cm. De groeiwet is de
De responsie van de derde-machtsrelatie is inderdaad sneller zoals in de volgende figuur te zien is. Bedenk echter wel dat deze routine zodra de sarcomeerlengte kleiner dan 3 micrometer wordt weer aanzienlijk langzamer wordt dan de arctangensrelatie.
arctangensrela tie.
29

aantal sarcomeren o-6 sarcomeerlengte 5300
c
Figuur 20: Responsie op de grote uitwijking, u=O. 75 cm. Als groeiwet de derde- mach tsrela tie.
5.3 Discussie. Uit de experimenten is gebleken dat de keuze van de groeiwet een vrij grote invloed heeft op het verloop van het aantal sarcomeren ten gevolge van een opgelegde belasting. Het is voor een betrouwbare voorspelling van het aantal sarcomeren dan ook noodzakelijk meer over de werkelijke groeiwet te weten te komen. In het vervolg van deze studie zullen we als groeiwet gebruik maken van de arctangensrelatie, omdat:
de arctangensrelatie een bovengrens heeft voor de groeisnelheid. uit de experimenten is gebleken dat deze groeiwet snel genoeg is om adequaat op belastingswisselingen te reageren, en traag genoeg om te zorgen dat niet op elke beweging gereageerd wordt met adaptatie-effecten.
Ook bij grote afwijkingen heeft de arctangensrelatie de voorkeur. Wanneer er zeer grote deformatie optreedt is het niet waarschijnlijk dat adaptatie snel op gang komt, omdat het weefsel zo beschadigd is dat de normale processen wat langer duren. De adaptatiesnelheid van de arctangensrelatie sluit hierbij aan. Verder duurt het bij de derde-rnachtsrelatie veel te lang voor kleine afwijkingen van de optimum-sarcomeerlengte weggewerkt zijn.
30

6. Adaptatie in het dynamisch spiermodel.
6.1 Inbouwen van het adaptatiemechanisme in het dynamisch spiermodel. In de vorige paragraaf is gebleken dat de arctangensrelatie het best als groeiwet gebruikt kan worden. We willen deze groeiwet dan ook toepassen om adaptatie- effecten te modelleren in het dynamisch spiermodel. We gebruiken dus:
Dit adaptatiemechnisme was gebaseerd op het constant houden van de sarcomeerlengte door het aantal sarcomeren aan te passen. Om dit mechanisme te kunnen toepassen in het dynamisch spiermodel zijn twee dingen noodzakelijk: We moeten de sarcomeerlengte en het aantal sarcomeren kunnen bepalen én we moeten een parameter in het dynamisch spiermodel vinden die verband houdt met de sarcomeerlengte of met het aantal sarcomeren. De sarcomeerlengte en het aantal sarcomeren zijn te bepalen met behulp van de routine sarc0.m die we ook bij het geometrische model gebruikt hebben. In deze routine wordt op grond van de verlenging u, het aantal sarcomeren en de sarcomeerlengte bepaald. Om deze parameters aan het dynamisch spiermodel te koppelen kunnen we de parameter a, het pariicipatieniveau, gebruiken. We zullen in het dynamisch spiermodel gebruik maken van a, die elke tijdstap opnieuw berekend wordt als functie van de sarcomeerlengte.
31

Om het overzicht te bewaren zetten we nu het principe van de modellering nog eens uiteen in een stroomschema:
Niveau 1 Niveau 2 Niveau 3 % Hoofdprogramma
initiële parameters, t=O, a=l
' i 1
F(t) = Ft
schatting voor de - % Estforce verlenging, us.
US =- Yo Moments
berekening van Q, waaruit de kracht te berekenen
Q - valt. I
F, = CS + F p s
F, = a . C,. Q, Fps 1 K p . US
r=,F;-F, met Fc=kracht van het contractiele element,
Fp=kracht van het passieve element en
Fs=de geschatte kracht
Ir1 > tol /jT/ < tol
nieuwe schatting, te verlenging maken mbv is bekend. c2 1 Fs,u usn=us
I de
1
I 1 % Sarco bere ken ing nieuwe sarcomeerlengte, Isarc, en het aantal sarcomeren, n.
Yo Overlap aanpassing van a, het
I participatieniveau.
t<te t=te END
32

6.2 Keuze van de constanten. Zoals in het stroomschema te zien is zijn er drie constanten waarvoor waarden gekozen moeten worden. Dit zijn de parameters C,, C, en K p . Hierbij geeft C, het verband tussen de kracht F ( t ) die een spier levert en het genormeerde eerste orde moment Q,, wat we vinden als uitkomst van de berekening in het programma 'moments', waarmee het DM-model doorgerekend wordt. C, is de parameter die aangeeft hoeveel de schatting van de verlenging aangepast moet worden als functie van het verschil tussen de werkelijke en de geschatte kracht. De parameter K p geeft de stijfheid van het parallelle element weer. De indices die gebruikt worden geven aan over welk deel van het model het gaat: het contractiele element heeft de index c, het passieve element heeft index P. De waarde van C, kunnen we bepalen op grond van literatuurgegevens voor de tetanische kracht van de soleus van een kat. In tetanische contractie levert deze spier een kracht van 2,l N. [6]. ( Zahalak vindt voor deze tetanische kracht een waarde van 2,l kg. Aangenomen is dat hier een kracht van 2,l N wordt bedoeld.) Als we voor de rate-functies de volgende waarden kiezen: f, = 35s-'
g , =7s-l
g, = 200s-'
g, = 30s-' is de waarde van Q, die we uit berekening met momenkm vinden Q, = 0.45177 [- I. Voor de kracht geldt: F ( t ) = aC,Ql [NI. Bij tetanische contractie geldt a=l [-I, waarmee we voor de waarde van C, vinden:
Ook voor de bepaling van de waarden van de andere parameters gaan we ervan uit dat we de soleus van een kat beschouwen. In de literatuur vinden we [9]:
C1=4,65 N.
_ _ - 0.1 _.-- Cl 1
Lo Kp met: C, =4.65[N]
vinden we voor de stijfheid van het parallelle element K p = 4650[N/m2]. Dit geldt echter voor een serieel element, zoals een pees. Doordat het volume van het parallelle element echter veel kleiner is, is ook de stijfheid van dit weefsel veel minder groot. We nemen aan dat dit ongeveer een factor duizend scheelt. We kiezen daarom Kp = 5[N/m2] .
Voor de bepaling van C, is het nodig eerst eens goed te kijken wat deze parameter eigenlijk voorstelt. C, bepaalt de mate van aanpassing van de
33

schatting van de uitwijking als functie van het verschil tussen de werkelijke en de geschatte kracht, AF'. We willen dat deze aanpassing volgens de methode van Newton-Rhapson verloopt. Daarvoor moet gelden:
De totale stijfheid wordt bepaald door de stijfheid van zowel het contractiele als het parallel elastische element volgens:
De stijfheid van het contractiele element veranderi continu door het binden en ontbinden van cross-bridges. Beschouwen we echter de stijfheid bij tetanische contractie dan vind je:
Krot = K, + K p [N/m2]
met [6]: a = i[-]
Lo = 1. [m]
C, = 4.65[ NI
y =O.O15[-]
= 0.7[ -1 Waardoor de waarde voor de stijfheid van het contractiele element, K, = 2.17-104 [ N/m2] wordt. We hadden al beredeneerd dat K p = 5[ iV/m2] zodat we nu de totale stijfheid kunnen bepalen. Hiervoor vinden we: K,, = 2.1705- lo4 [ N / m 2 ] , zodat we uiteindelijk voor de waarde van parameter C,
vinden: C, = 4.6-10-5[m2/N]. We kunnen C, ook bepalen op grond van de momentane waarde van Qo. Dit werkt op dezelfde manier als hierboven en kan in de routine ingebouwd worden. We hebben nu alle noodzakelijke parameters bepaald, zodat we over kunnen gaan naar de experimenten.
6.3 Sim ula tie-experimen ten. We zullen weer dezelfde drie belastingspatronen bekijken als in het geometrische model, maar nu gaan we de kracht voorschrijven in plaats van de verlenging. We beschouwen dus:
1. F ( t ) = A S i i l ( F f )
met: ' b karakteristieke belastingstijd, zb=l A krachtsamplitude
34

B constante We zullen laten zien wat het effect van de aanpassing van a op de verlenging u is. Voor de kracht die maximaal opgelegd wordt kiezen we twee keer de tetanische kracht 2.1 N., dus 4.2 N.
6A Resultaten. We zulien voor de drie beiastingssituaties ue resuitaten hieronaer in enkeie figuren weergeven.
6.4.1 Belastingspatroon 1. De eerste belastingssituatie is:
v- Zb = 1 At = 0.05 0 < t < 2 We vinden dan:
& V
Y 9 Kracht en schatting 0-4Jitwijking met adaptatie
3 10-6 sarcomeeriengte
- i2.0;;
o 2 07 1.95 CG
O 0.5 1 1.5 2 tijd in dagen
alfa
0.5 1 1.5 2 tijd in dagen
1 1.5 2 tijd in dagen
aantal sarcomeren S
a, 9 5001
5 5000.5 % 2 5000
; 4999.5
(u c c
O 0.5 1 1.5 2 tijd in dagen
0-13 verschil in uitwijking
$ 0.5 3
1 1.5 2 tijd in dagen
Figuur 21: Adaptatie op een zuiver sinusvormige belasting.
35

In de figuur is het verloop van volgende parameters weergegeven (van links naar rechts en van boven naar beneden): 1. De werkelijke en de geschatte kracht zijn uitgezet in één figuur. Het is duidelijk
te zien dat de schatting van de kracht vrijwel gelijk is aan de werkelijk opgelegde kracht.
weergegeven. Bij de berekening van de uitwijking is gebruik gemaakt van de steeds aangepaste parameter a.
fundamenteel van het verloop van de sarcomeerlengte in het geometrisch model.
4. Ook het aantal sarcomeren is uitgezet tegen de tijd. Hiervoor geldt ook dat het verloop niet fundamenteel verschilt van dat in het geometrisch model. Dit is overigens ook wat we verwachten, aangezien de berekening van sarcomeerlengte en aantal sarcomeren op exact dezelfde wijze gebeurt als in het geometrisch model.
5. De waarde van de parameter a, berekend op grond van de sarcomeerlengte, is in het vijfde plaatje uitgezet tegen de tijd.
6. We hebben de uitwijking in twee gevallen berekend: In het eerste geval werd parameter a steeds aangepast, we kregen dan de uitwijking u. In het tweede geval werd deze parameter niet aangepast, m.a.w. de invloed van adaptatie werd niet in rekening gebracht. De uitwijking die we in dat geval vonden noemden we UZ. In het laatste plaatje zijn deze twee uitwijkingen van elkaar afgetrokken en uitgezet tegen de tijd. Aangezien bij de berekening van u de waarde van akleiner dan 1 kan zijn, verwacht je dat UZ kleiner is dan u. In de bovenstaande figuur is a steeds 1, zodat u en UZ niet verschillen.
2. De uitwijking ten gevolge van de opgelegde kracht is in het tweede plaatje
3. De sarcomeeriengte staat uitgezet tegen de tijd. Dit veïioop veischilt niet
6.4.2 Belastingspatroon 2. Ook voor de tweede belastingssituatie, waarvoor geldt:
z, = 1 At = 0.05 O<t<2 is een dergelijke figuur gemaakt. In de figuur zijn dezelfde parameters weergegeven als hierboven vermeld is. De amplitude van de sinus is veel groter gekozen omdat uit de vorige figuur bleek dat de sarcomeeriengte niet buiten net voor de parameter a optimale gebied uit kwam.
36

L o Y L Kracht en schatting
o-6 sarcomeerlengte s 2.2
0 2 o L "~ m v) O 0.5 1 1.5 2
tijd in dagen alfa
2r-----l
, 0-4Jitwijking met adaptatie 101 I
-0 0.5 1 1.5 2 tijd in dagen
aantal sarcomeren 5006
5 5004
- 5002
3 5000
al
5 m 8 +
O 0.5 1 1.5 2 tijd in dagen
o-12 verschil in uitwijking
I 0.5 1 1.5 2
O' O
tijd in dagen 1 1.5 2
tijd in dagen
Figuur 22: Adaptatie op een sinusvormige belasting met een constante uitwijking erbij opgeteld.
Uit figuur 23 blijkt dat zelfs bij een kracht van 15 N. de sarcomeerlengte nog niet groter wordt dan 2,2 micrometer. We hebben dus de schatting voor de maximaal op te leggen kracht veel te voorzichtig gedaan. Bij de volgende belastingssituatie zullen we de maximale kracht nog groter kiezen.
6.4.3 &?k3§thg§patl33Qn 3. Als laatste belastingssituatie is gekozen voor:
Zb = 1 At = 0.05
O<t<2 We vinden hierop de volgende responsie:
37

L o Y 9 Kracht en schatting
s -20 Q O 0.5 1 1.5 2 - 5 tijd in dagen s o-6 sarcomeerlengte
L 8 1.8
O 0.5 1 1.5 2 tijd in dagen
alfa
I O 0.5 1 1.5 2
tijd in dagen
0.96'
O-CUitwijking met adaptatie
3 23
I O 0.5 1 1.5 2
tijd in dagen aantal sarcomeren
-1 '
~
E 5010
5 5005
- 5000
a,
3 m c c
O 0.5 1 1.5 2 tijd in dagen
verschil in uitwijking
3 4995
,
tijd in dagen
Figuur 23: Adaptatie op een sinusvormige belasting met een trend.
De belasting die we nu opgelegd hebben heeft tot gevolg dat de sarcomeerlengte buiten zijn optimale gebied komt. Hierdoor wordt de parameter a aangepast. Dit is te zien in de grafiek die a weergeeft als functie van de tijd, maar ook in de grafiek die u-UZ weergeeft.
6.5 Discussie. Gezien de resultaten is het mogelijk een kracht voor te schrijven en daarbij de uitwijking te berekenen met behulp van schatting en iteratie. Nadeel van de methode is dat de rekentijd veel langer wordt. Dit probleem is echter gedeeltelijk op te lossen door een snel convergerende iteratie-methode toe te passen. Uit bovenstaande figuren blijkt dat de waarden die we voor de uitwijking en de sarcomeerlengte vinden realistisch mijn bij de gekozen belasiingssituaties. De uitwijking is in de orde van 10% van de oorspronkelijke spierlengte, de sarcomeerlengte ligt tussen de 1.8 en 2.2 micrometer en het aantal sarcomeren ligt tussen de 4998 en 5100 sarcomeren. Dit is binnen het gebied dat we in paragraaf 4.3.1 hebben onderzocht op stabiliteit. De adaptatieroutine is dus in ieder geval stabiel. We hebben echter de stabiliteit van de tweede iteratie, die voor de bepaling van de uitwijking, niet onderzocht. Op grond van bovenstaande resultaten en de overeenkomst met de resultaten die het geometrisch model leverde, veronderstellen we dat in bovenstaande gevallen ook de tweede iteratie stabiel verliep. Wanneer dit echter niet het geval zal zijn kunnen we niet voorspellen. Gelukkig merk je snel genoeg of de tweede iteratie stabiel is of niet:
38

Als deze iteratie instabiel verloopt treedt geen convergentie op, waardoor de werkelijke en de geschatte kracht nooit gelijk worden aan elkaar. De waarde van u-UZ is zeer klein en positief. Het in rekening brengen van de verandering in overlap tussen de actine- en myosinefilamenten, zoals beschreven met de participatiefactor a , levert in dit geval een relatieve correctie in de uitwijking u op in de orde van lo4. Dat houdt in dat de uitwijking die wordt berekend wanneer de waarde van a steeds aangepast wordt altijd groter is dan de uitwijking die zonder a te veranderen wordt berekend. Dit is in
het aantal cross-bridges dat gevormd kan worden kleiner dan het maximale aantal. Hierdoor zal de uitwijking ten gevolge van een opgelegde kracht groter zijn aangezien er minder cross-bridges zijn om de kracht door te leiden.
uveïeensiemìÏìincj met wat we verwachtten, want wdnNeeÏ ü kleinei- is dan 1, is
39

7. Conclusies en aanbevelingen. De opzet van deze studie was een overzicht te krijgen van het adaptatiegedrag van spieren en op grond daarvan een begin te maken met de modellering. In de eerste twee hoofdstukken is een neerslag van het eerste te zien: een beschrijving van spiergedrag, spiergeometrie en spieradaptatie. De gegevens hierover zijn verkregen uit literatuuronderzoek. Tijdens dit literatuuronderzoek bleken er nog de nodige hiaten te zitten in de kennis over het adaptatiegedrag van de skeletspier. Men is in staat de oorzaken en effecten van adaptatie kwalitatief te beschrijven, maar met name op het gebied van de oorzaken is nog niet voldoende bekend voor een modellering. Zo wordt als oorzaak voor adaptatie vaak ‘training’ of ‘langdurige inactiviteit’ genoemd, terwijl voor een betrouwbare modellering een nauwkeurigere parameter nodig is zoals kracht, spanning, rek of energie-inhoud van het weefsel. Dat gegevens hierover ontbreken lijkt te komen doordat fysiologen, die onderzoek naar adaptatie verrichten, gebruik maken van andere modellen dan technici, waarin andere parameters een rol spelen. Toch hebben we een voorzichtig begin gemaakt met het modelleren van het adaptatiegedrag van de spier, waarbij de nadruk heeft gelegen op de kwalitatieve aspecten en de numerieke stabiliteit van de gekozen groeiwet. Er moeten bij een modellering van zo’n complex materiaal als spierweefsel veel keuzes gemaakt worden, vooral ook doordat er nog veel gegevens ontbreken. We hebben steeds geprobeerd deze keuzes zo goed mogelijk te beredeneren en ook deze redenaties zijn voor een groot deel opgenomen in dit verslag. Zo kan iedereen die op dit terrein werk verricht zijn eigen afwegingen maken, om misschien wel tot heel andere keuzes te komen. In deze studie hebben we op grond van geometrische overwegingen een groeiwet geformuleerd, die na analyse stabiel bleek te zijn op het door ons gewenste gebied.Vervolgens zijn we erin geslaagd deze geometrische groeiwet toe te passen in een dynamisch spiermodel dat geënt is op het DM-model. Het model dat daardoor ontstond moet beschouwd worden als een kwalitatief model aangezien over de werkelijke waarden van diverse parameters nog erg veel onzekerheid bestaat. Wel kunnen we concluderen dat het goed mogelijk is adaptatie in een dynamisch spiermodel te modelleren, zolang we genoegen nemen met een kwalitatieve beschrijving van adaptatiegedrag. Uit de simulaties die met dit model zijn uitgevoerd kunnen we het volgende concluderen:
Het gebruik van een begrensde groeirelatie zoals de arctangensrelatie lijkt het groeigedrag van spierweefsel redelijk goed kwalitatief te beschrijven. Het is mogelijk diverse belastingspatronen voor te schrijven. Bij de belastingspatronen die wij hebben getest vinden we resultaten die we verwachten. We kunnen nu dus ook ingewikkelder belastingspatronen voorschrijven. Het spiermodel waarin de groeiwet ingebouwd is, is niet van belang, zolang er maar een relatie te leggen is tussen de parameters van de groeiwet en het spiermodel. Dit is te zien aan de parameter a, die in het spiermodel het effect van de sarcomeerlengte in rekening brengt. Het is hierdoor waarschijnlijk ook mogelijk de groeiwet in te bouwen in een geavanceerder spiermodel zodat de beschrijving van het spiergedrag nog beter wordt.
40

Ook kunnen nog enkele aanbevelingen voor vervolgonderzoek worden gedaan: Het verdient aanbeveling om het aantal parallelle sarcomeren in de beschouwing op te nemen. In het huidige model zou dit kunnen door uit te gaan van een kracht-belasting die resulteert in een variatie van de sarcomeerlengte rond het optimum. De amplitude dient dan zodanig te zijn dat de participatieparameter a soms kleiner wordt dat 1. Op die momenten zouden er extra sarcomeren parallel gevormd moeten worden. Ook is hei wäârschijniijk moyeiijk een aciaptatiernociei ie baseiein op een energiebeschouwing, zoals in paragraaf 4.1 . I kort is aangegeven. Hiermee denken wij dat het mogelijk is zowel het aantal seriële als het aantal parallelle sarcomeren te berekenen. Wanneer dat goed lukt is het mogelijk beide modellen te vergelijken en eventueel te combineren. Ook lijkt het zinnig in vervolgonderzoek in dit model van één-dimensionale beschouwing over te gaan tot een twee- of driedimensionaal model. Het rekenwerk zal daarmee ingewikkelder worden, maar het is dan ook beter mogelijk het model met de werkelijkheid te vergelijken, doordat je complexere spiergeomet riën kunt beschrijven. De groeiwet die in deze studie gebruikt is, vormt een waarschijnlijk een redelijk goede indicatie van een ‘werkelijke’ groeiwet. Ook wanneer in vervolgonderzoek een andere groeiwet gebruikt wordt is het raadzaam te zorgen dat deze groeiwet een bovengrens heeft aan de groeisnelheid. Zo voorkom je dat sterk gedeformeerd en zelfs zwaar beschadigd weefsel zich in het model supersnel herstelt. Verder mag de groeisnelheid rondom het optimum niet al te klein zijn, omdat gebleken is dat anders het optimum te langzaam bereikt wordt en er pas te laat ingegrepen wordt wanneer de sarcomeerlengte verandert. Verder kan de modellering van het parallelle element beter uitgewerkt worden. Dit element gedraagt zich in werkelijkheid waarschijnlijk meer visco-elastisch dan elastisch en zou ook als zodanig gemodelleerd kunnen worden. Ook kan adaptatie van dit parallelle weefsel optreden, door bijvoorbeeld vezeloriëntatie. Bij een uitgebreid adaptatiemodel zal ook die adaptatie meegenomen moeten worden. Tenslotte is het nog mogelijk naast het parallele element, dat de passieve spierstructuren in rekening brengt, ook de aanhechting aan het bot via een pees in het model op te nemen. Dit kan gebeuren door een serieel element met peeseigenschappen te modelleren. Ook dit element kan aan adaptatie onderhevig zijn.
41

BRONNEN.
Bernards, J.A., Bouman, L.N. (1 988). Fysiologie van de mens. Houten: Bohn, Stafleu Van Loghum.
Fitts, R.H., McDonald, K.S., Schluter, J.M. (1991). The determinants of skeletal muscle force and power: Their adaptability with changes in activity p2m-n. JGüi.i.ial of auiûK€chârilcs, 24, i i i -i22.
Watson, P.A. (1 991). Function follows form: generation of intracellular signals by cell deformation. The FASEB Journal, 5, 2013-201 9.
Fung, Y.C. (1 993). Biomechanics, Mechanical Properties of Livina Tissues. 2"d-edition. Springer-Verlag.
Vandenburgh, H.H., Hatfaludy, S., Karlisch, P., Shansky, J. (1 991). Mechanically induced alterations in cultured skeletal muscle growth. Journal of Biomechanics, 3, 91 -99.
Zahalak, G.I. (1 981). A distribution-moment approximation for kinetic theories of muscular contraction. Mathematical Biosciences, 55, 89-1 14.
Cowin, S.C., Arramon, Y.P., Luo, G.M., Sadegh, A.M. (1993) Chaos in the discrete-time algorithm for bone-density remodelling rate equations. Journal of Biomechanics, 22, 1077-1 089.
Zahalak, G.I. Modelling muscle mechanics (and energetics). In: Winters, J.M., Woo, S.L.-Y. (Eds.) (1 990). Multiple muscle svstems. (hoofdstuk 1). New-York: Springer-Verlag.
Ma, S., Zahalak, G.I. (1 991). A distribution-moment modelof energetics in skeletal muscle. Journal of Biomechanics, 24, 21 -35.
Goldspink,G. Alterations in myofibril size and structure during growth, exercise and changes in environmental temperature. In: Peachy, L.D., Adrian, R.H., Geiger, S.R. (Eds.) (1984). Handbook of Phvsiolouv - Skeletal Muscle , p. 593-654. American Physiological Society.
42

Bijlage Matlabfiles voor het berekenen van spieradaptatie-effecten.
Stabiliteitsbepaling. We hebben diverse routines waarmee we de stabiliteit van de diverse groeiwetten kunnen testen. Deze routines verschillen alleen in de groeiwet die getest wordt.
% faccent
Yo In deze routine wordt de gelineariseerde vorm van de groeiwet, 'faccent', Yo ingevuld voor een bepaald werkpunt. Dit werkpunt is in te stellen door de Yo waarden voor I en n te variëren. Zolang de waarde van 'faccent' tussen de Yo I en -1 ligt is de routine stabiel. Yo Door 'faccent' te laten berekenen als functie van de tijdstap 'deltat' en 'n' Yo is te zien hoe klein de tijdstap gekozen moet worden bij gegeven h ' om % stabiliteit te garanderen. Yo De groeiwet die in deze routine getest wordt is de arctangensrelatie met de % factor (n/nO) erbij.
a=( 2/( pi "5 000)) ; beta=(5*pi)/( 1.5e-6); taua=( 1/1 O); I=0.9e-2; IOs=2e-6;
DELTAT=[O: 1 :80]; N=[3500:50:6500]; [de I tat, n]=mes h do m (D E LTAT, N) ;
Yo De afgeleide van de gelineariseerde vorm van de groeiwet. faccent=a.*(deltat/taua).*atan((beta*(I-n*lOs)./n))-. . .
2*l.*n*lOs+n .A2*IOsA2)+n ./\2)+1; (a* beta" i. * n. * (dek autau a)) ./( betaA2* (P2 -
o/o Als uitvoer krijgen we een plot met hoogtelijnen, zodat te zien is waar f acce n t % tussen de -1 en 1 ligt en dus stabiel is.
cs=contour( DELTAT, N ,faccent,s) cla bel (cs)
s=[I ;.5;0;-.5;-1];
Yo facc
43

YO Ook in dit programma wordt de gelineariseerde vorm van de YO arctangensrelatie ingevuld. Hierbij is echter de factor (n/nO) niet YO meegenomen, omdat wordt verondersteld dat deze factor voor het YO vreemde stabiliteitsverloop zorgt.
beta=(5*pi)/( 1.5e-6); taua=( 1 /I O); I=1 . I e-2; IOs=2e-r;;
N=[3500:50:6500]; DELTAT=[O:I :80]; [de Itat, n]=mes hdom ( D E LTAT, N ) ;
Yo Afgeleide van de gelineariseerde vorm van de groeiwet. fn=l -(deltat.*taua*2*beta*l)./. . .
(pi * ( b et aA2* (IA2 -2. * n. * I * I Os+ n .A2 * I OsA2) +n .A2)) ;
~=[.985; .990; .995; I ] ; cs=contour(DELTAT,N,fn,v); clabel(cs)
Yo fcc
% Ook in dit programma wordt de gelineariseerde vorm van een Yo groeiwet ingevuld. In dit geval bekijken we echter de derde- machtsrelatie. Yo De factor (nhref) wordt gewoon in de beschouwing meegenomen.
clear clf
Yo Parameters beta=9.79e5; taua=( 1 /1 O); I=1.1 e-2; nref=5000; IOs=2e-6;
N=[3500:50:6500]; DELTAT=[O:1:80]; [ delt at, n]=mes h dom (D E LTAT, N) ;
Yo Afgeleide van de gelineariseerde groeiwet. f n =- ( ( bet a)/\3. *(de It at ./( t a u a* n ref)) . * ( ( I - n. *I Os) . A2) . * (2. *I + n. * I Os)) ./( n .A3) ;
SU bplot(22 1 )
cs=con tou r( D E LTAT, N , f n ,v) ; c I a b e I (cs)
~=[-1;-.75;-.5;-.25;0];
44

title('Stabi1iteit bij I=1 , I cm.') xlabel('de1tat') ylabel('aanta1 sarcomeren,n')
Berekening van het aantal sarcomeren. De berekening van het aantal sarcomeren hangt af van de gebruikte groeiwet. We gebruiken twee verschillende groeiwetten, waarvoor we twee routines hebben die weer alleen verschillen in de toegepaste groeiwet. De routine karco' gebruikt de arctangensrelatie a!s groeiwet, de routirie 'sarc' gebruikt de derde- mach ts relat ie.
f u nc t io n [ Isa rc , n n ew] = sarco (u, t , no Id)
Yo Berekening van het aantal sarcomeren.
% In dit programma wordt het aantal sarcomeren Yo als functie van de rek bepaald met behulp van de Yo arctangensrelatie. Yo De initiële parameters zijn:
lospier= 1 e-2; IOsarc = Ze-6; deltat = 1; n0=6000; taua= (1 /I O); beta=(5*pi)/l.5e-6;
o/o Indien de sarcomeerlengte niet in het optimale gebied ligt wordt Yo het aantal sarcomeren aangepast. Hierbij gaan we ervan uit dat per Yo tijdseenheid slechts een beperkt aantal sarcomeren gevormd kan YO worden. De groeifunctie wordt continu beschreven door een arc t an g e ns . Yo We gebruiken: ndot = arctan(((lsarc-2.0e-6)*pi*5)/1.5e-6).
Isarc = (IOspier+u)/nold;
nnew = nold+(deltat/taua)* ... (Z/pi)*atan (beta*( Isarc-IOsarc));
Isarc = (IOspier+u)/nnew;
function[lsarc, nnew] = sarc(u,t,nold)
YO Berekening van het aantal sarcomeren. Yo In dit programma wordt het aantal sarcomeren als functie van Yo als functie var? de rek bepaald met behulp van de derde-machtsrelatie. Yo De initiële parameters zijn:
lospier= 1 e-2; IOsarc = Ze-6;
45

deltat = .I ; taua= (1/1 O); beta=9.79e5; nref=5000;
%O Indien de sarcomeerlengte niet in het optimale gebied ligt wordt % het aantal sarcomeren aangepast. Hierbij gaan we ervan uit dat per O/O tijdseenheid slechts een beperkt aantal sarcomeren gevormd kan
% rnachtsrelatie. % worden. De groeifunctie YIorctt continu heschre\Ier? U m r een derde=
Isarc = (IOspier+u)/nold;
nnew = nold+( deltat/taua)* (nold/n ref) * . . . (( beta*(lsar~-lOsarc))~3);
Isarc = (IOspier+u)/nnew;
Verlenging als belasting. Wanneer we willen kijken wat de responsie is op een bepaalde voorgeschreven verlenging, kunnen we het programma 'test' gebruiken. Met behulp van de parameters A en B kan de belasting veranderd worden. We gebruiken dit programma alleen in het geval we het geometrische model beschouwen.
Yo Test.
o/o Parameters. A=5e-4; B=O; T=l ; t=[0:.01 :IO];
YO Belasting. u=A*sin( (2*pi*t)/T)+B*t; % Initieel aantal sarcomeren. ni=5000;
n=El; Isarc=[];
for i=l :length(t) [ Isa rci , n i]=sarco( u (i), t (i), n i); Isarc=[lsarc Isarci]; n=[n nil;
end
p I o t (t , n) ;tit I e ( 've r I e n g i n g a Is be I ast i n g ') ; xlabel('tijd in dagen'); ylabel('aanta1 sarcomeren'); grid;
46

Bepaling van de kracht van het contractiele deel. Wanneer we een verlenging voorschrijven of schatten, kan de daarbijbehorende kracht van het contractiele deel berekend worden met behulp van de volgende routine 'moments'. In deze routine wordt het DM-model doorgerekend. De input is een verlengingssnelhied, als output krijg je de genormeerde waarden voor de momentane stijfheid van het materiaal, de doorgeleide spanning en de energie- inhoud van het weefsel. Om met moments te kunnen werken zijn nog twee andere programma's nodig: jay.m en adam12.m die ook bijgevoegd zijn.
function Qdot= m ~ m e w t s (t,k);
% Geschreven door Ir. Sander Gielen. Y O Technische Universiteit Eind hoven Y O
% Qdot = moments( t, Q) Y O YO compute distributed moments model % G.I. Zahalak, Math.Biosc.55:89-l14(1981) % YO usage: % [t,Q]=adam 12('rnornents',tO,te,delta t, Q(t=O)); global us
Faculteit der Werktuigbouwkunde , vakgroep WFW.
integrate numerically Q with ode23 or adaml2 as
QO=k( 1 ) ; Q1 =k(2); Q2=k(3) ; Qdot=zeros(3,1); Y O
% parameters fl=35; gl=7; g2=200; g3=30; U=US; if t<O, f l =O; end % % computation p=Ql/QO; q=sq rt(Q2/QO - ((Q 1 /QO) .A2)); poverq=p/q; poverqc=( 1 -p)/q; YO jaypq=jay(-poverq, p,q); jaypqc=jay(poverqc, p, q) ; jayOpq=jawU ); jayOpqc=jaypqc(l ); jay1 pq=jaypq(2); jay1 pqc=jaypqc(2); jay2pq=jaypq(3); jay2~qc=jaypqc(3); jay3pq=jaypq(4); jay3pqc=jaypqc(4); Y O
phi0 = QO*( 92" jay0pq +(fl +gl)*(jayl pqc-jay1 pq) +gl *(p-jay1 pqc) ...
phi1 = QO*( 92" jay1 pq +(fl +gl)*(jay2pqc-jay2pq) +gl *(p*p+q*q -jay2pqc) +g3*(p-jayl pqc -1 +jayOpqc));
... +g3*(p*p+q*q-jay2pqc -P +jay1 pqc));
47

phi2 = QO*( g2* jay2pq +(f 1 +gl)*(jay3pq~-jay3pq)+gI *(p*p*p+3*p*q*q- jay3pqc) ...
% Qdot = [(fl /2 - phi0)
+g3*( p*p*p+3*p*q*q-jay3pqc -(p*p+q*q) +jay2pqc));
( f l /3 - phi1 - u*QO) (f l /4 - phi2 - 2*u*Ql)];
Y O
function J=jay (t , p I q) ; % J = jay(t,p,q) Y O
Yo Geschreven door Ir. Sander Gielen. Yo definition of function J in distribution moment model Yo see \zah81 % t : limit Yo p :mean % q : standard deviation Yo J is returned as: [JO J1 J2 J3] J=ze ros (4,l) ; p h it = m ye tf (t) ; ait= I/sqrt(2*pi)*exp(-t*t/2); J( 1 )=phit; J(2)=p*phit-q*ait; J(3)=p*p*phit - 2*p*q*ait+q*q*(phit-t*ait); J(4)=p*p*p*phit - 3*p*p*q*ait+ 3*p*q*q*(phit-t*ait)-q*q*q*(2+t*t)*ait;
function [tout, yout] = adaml2(FunFcn, to, tfinal, h, yo, trace)
%ADAM1 2 Y O
Y O Yo % Yo % conditions YO. % Yo
Integrate a system of ordinary differential equations using 1 st and 2nd order Adams formulas.
[T,Y] = ADAM12('yprime', TO, Tfinal, dt, YO) integrates the system of ordinary differential equations described by the M-file YPR1ME.M over the interval TO to Tf with step dt and using initial
[T, Y] = ADAM12(F, TO, Tfinal, dt, YO, 1) displays status while the integration proceeds. Y O
- String containing name of user-supplied problem description. Call: yprime = fun(t,y) where F = 'fun'.
Yo INPUT: % F Y O Y O t - Time (scalar). Y O y - Solution column-vector. Y O yprime - Returned derivative column-vector; yprime(i) = dy(i)/dt. % tO - Initial value of t. Yo tfinal- Final value of t. % y0 - Initial value column-vector. Yo lol - The desired accuracy. (Default: tol = I .e-3). Yo trace - If nonzero, each step is printed. (Default: trace = O).
48

% Yo OUTPUT: o/o T - Returned integration time points (column-vector). Yo Y - Returned solution, one solution column-vector per tout-value. % o/o The result can be displayed by: plot(tout, yout).
Yo C.B. Moler, 3-25-87. o/o Copyright (c) 1987 by the MathWorks, Inc. % All rights resewed.
o/o I nit i al i zat ion if nargin < 6, trace = O; end
o/o Initialization ns= (tfinal - tO)/h; ns= round(ns+0.5); t = to; y = yo(:); tout = zeros(ns,l); tout( 1 ) =t; yout = zeros(ns,length(y)); yout( 1 ,:) = y.I; k=l;
if trace
end CIC, t, h, y
% The main loop while (t e tfinal)
if t + h > tfinal, h = tfinal - t; end
Yo Compute the slopes si = feval(FunFcn, t, y); sl = SI (:); s2 = feval(FunFcn, t+h, y+h*sl); s2 = s2(:);
t = t + h ; y = y + h*(sl + ~2)/2; delta = norm( h*(sl - s2)/2, 'inf'); tau = max(norm(y,'inf'),l .O); tol = deltahau; k = k + l ; tout(k) = t; yoüt(k,:) = y.'; if trace
end home, t, h, y, tol
end;
49 ,

Bepaling van de participatie-parameter a. Om te kunnen bepalen hoeveel cross-bridges gevormd kunnen worden hebben we de participatieparameter nodig. Deze wordt bepaald op grond van de sarcomeerlengte.
funct ion[alfa]= overlap( us, nsarc)
% Bepalen van alfa. % We gaan ervm t i t dat dfa rechisiïeeks bepaaid wordt door Yo de mate van overlap tussen de actine en myosinefilamenten. Yo Als de sarcomeerlengte optimaal is, is de waarde van alfa 1. Yo Als de sarcomeerlengte niet optimaal is (dat wil zeggen: niet Yo tussen de 1.9*1 OA(-6) en 2.2*1 OA(-6)) is alfa ook kleiner dan 1
IOsarc=2.Oe-6; Isarc=lOsarc+( us/nsarc);
if Isarc > 2.2e-6
elseif Isarc < 1.9e-6
else alfa = 1; end
alfa = (2.375e-6 - 0.625*lsarc)*I e6;
alfa = (-1.71 e-6 + 1.428*lsarc)*1 e6;
De kracht als stimulus. We kunnen ook de kracht voorschrijven als stimulus. Bij een bepaalde kracht schatten we dan een uitwijking waarmee we de bijbehorende kracht uitrekenen. Het verschil tussen de werkelijke en de geschatte kracht levert dan een nieuwe schatting voor u op.
function[Fs, us, Qs]= estforce(Ft,us,tO,te,QO,alfa)
Yo Estforce % In het programma wordt FCS, de kracht die het contractiele element % levert, geschat door voor us, de geschatte verlenging een schatting te Yo doen en die in de routine ‘moments’ in te vullen. Yo In de routine ‘moments’ wordt Q berekend, waaruit we F weer kunnen bepalen. Yo Door deze schatting voor us telkens te verbeteren vinden we uiteindelijk Yo Fs=Ft+/- tol
% Parameters. tol=.0001; C1=4.65; c2=0.2; K=5 ; deltat=.005; Q10=0.45177;
50

global us; nt=(te-tO)/deltat+l ;
% Bepaling FCS. [t,Qs]=adam12(’moments’,tO,te,deltat,QO); Q1 =Qs(nt,2); QO=Qs(nt,:); Fcs=alfa*Cl *(Q1 -Q1 O); % Bepaling Fps.
O h Bepaling Fs en berekening van het residu, het verschil tussen % Fs en Ft, op grond waarvan de schatting voor us aangepast wordt.
r=Ft-Fs usn=us+r*C2; us=usn
F~s=us*K;
Fs=Fcs+FPs;
w h i I e a bs ( r) >to I [t,Qs]=adaml 2(’moments’,tO,te,deItat,QO); Q1 =Qs(nt,2); Fcs=alfa*Cl *(Qt -Q1 O); FPS=US*K; Fs=Fcs+FPs; r=Ft-Fs usn=us+r*C2; us=usn QO=Qs(nt,:);
end Qs=Qs( nt, :);
Hoofdprogramma. In het hoofdprogramma schrijf je de kracht ‘oor die op de pier aangebracht wordt. Ook wordt vanuit het hoofdpragramma gezorgd dat de sarcomeerlengte steeds opnieuw berekend wordt. Hiermee wordt alfa steeds aangepast. Door het hoofdprogramma te gebruiken kun je de responsie op een kracht bekijken.
YO hoofdprogramma
clear clear global
Yo Tijdsduur van het experiment en de opgelegde belasting. t=[0:.05:1]; F=5*sin(2*pi*t); %taub=l
% Begincondities uo=o; FO=O; lOsarc=2.Oe-6;
51

alfa=I ; nsarc=5000;
u=[];
Is rc=[]; nsrc=[]; alf=[];
Fd=[];
Y k i . 7 .4 31;
Yo Berekening van de responsie op de belasting. for i=l :length(t)
Ft=F(i); if i9l ; tO=t(i-1 ); else
end t e=t (i) ; [Fs,us,Qs]=estforce( Ft,uO,tO,te,QO,alfa);
YO Berekening van de sarcomeerlengte, het aantal % sarcomeren en de waarde van alfa. [lsarc,nsarc]=sarco(us,nsarc); [alfa]=overlap(us,nsarc);
t0=-.05;
QO=Qs; Quit=[Quit;Qs]; uo=us; u=[u us]; Fd=[Fd Fs]; Isrc=[lsrc Isarc]; nsrc=[nsrc nsarc]; alf=[alf alfa];
end
p I ot (t , F, t , Fd) ;tit le (I Kracht en schatting ') ; xlabel('tijd in dagen'); ylabel('werke1ijke en geschatte kracht'); pause plot (t , u) ;tit le( ' Uitwijking '> ; xlabel('tijd in dagen');ylabel('uitwijking in meters'); pause p I o t (t , n s rc) ;tit I e ( ' aa n t a I sa rc o m e ren ' ) ; xlabel('tijd in dagen'); ylabel("aantal sarcomeren'); pause p I o t (t , Is rc) ;tit I e ( ' I engte van de sa rco m e ren ' ) ; xlabel('tijd in dagen'); ylabel('1engte van de sarcomeren'); pause plot(t,alf);title('aIfa'); xlabel('tijd in dagen'); ylabel('a1fa');
52