identification of tyrosine 620 as the major phosphorylation site of
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 269, No. 44, Issue of November 4, pp. 27240-27245, 1994 Printed in U.S.A.
Identification of Tyrosine 620 as the Major Phosphorylation Site of Myelin-associated Glycoprotein and Its Implication in Interacting with Signaling Molecules*
(Received for publication, May 3, 1994, and in revised form, August 22, 1994)
Maria L. JaramilloSO, Daniel E. H. AfarOll, Guillermina AlmazanlI, and John C. Bell** From the Departments of Biochemistry and Medicine, University of Ottawa, Ottawa, K l H 8M5 Ontario, Canada and the IDepartment of Pharmacology and Therapeutics, McGill University, Montreal, H3G lY6 Quebec, Canada
Myelin-associated glycoprotein (MAG) is a myelin-spe- cific cell adhesion molecule of the immunoglobulin su- pergene family and is tyrosine-phosphorylated in the developing brain. To define the role of MAG in signal transduction, the tyrosine phosphorylation sites were analyzed. The major tyrosine phosphorylation residue was identified as "620, which was found to interact specifically with the SH2 domains of phospholipase C (PLCy). This domain may represent a novel protein binding motif that can be regulated by tyrosine phos- phorylation. MAG also specifically bound the Fyn tyro- sine kinase, suggesting that MAG serves as a docking protein that allows the interaction between different signaling molecules.
Protein tyrosine phosphorylation is a tightly controlled proc- ess important in cell regulation. Much research effort has fo- cused on the identification and characterization of protein ty- rosine kinases and their substrates. Myelin-associated glycoprotein (MAG),l a molecule believed to play a critical role in myelination, is an in vivo protein tyrosine kinase substrate (1). It is a type 1 integral membrane protein with an amino- terminal extracellular domain, a single transmembrane do- main, and a cytoplasmic carboxyl terminus (2-5). MAG is a member of the immunoglobulin supergene family and is re- stricted in expression to oligodendrocytes in the central n e w ous system and Schwann cells in the peripheral nervous system (6). MAG is encoded by a single gene as two developmentally regulated alternatively spliced isoforms (2, 3, 5, 7). They are designated L-MAG (for large MAG) and S-MAG (for small MAG) and have molecular masses of 72 and 67 kDa, respec- tively, in their nonglycosylated forms. The MAG isoforms are identical in their extracellular and transmembrane domains. They share a common region in their cytoplasmic domain but are distinct at their carboxyl termini. The unique carboxyl tails
*This work was supported by a grant from the National Cancer Institute of Canada and the Canadian Cancer Society (to J. C. B.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "aduer- tisement" in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ A postdoctoral fellow of the Medical Research Council of Canada. 9 Contributed equally to this work.
University of California, Los Angeles, CA 90024-1662. ll Present address: Dept. of Microbiology and Molecular Genetics,
** An NCI Senior Scientist. To whom correspondence should be ad- dressed: DeDts. of Biochemistrv and Medicine. Universitv of Ottawa. 451 Smyth Rd., Ottawa, K1H 8M5 Ontario, Canada. Tel.: 6i3-787-6698; Fax: 613-787-6779.
The abbreviations used are: MAG, myelin-associated glycoprotein; PLC, phospholipase C; aMEM, a minimum essential medium; FCS, fetal calf serum; PCR, polyerase chain reaction; PAGE, polyacrylamide gel electrophoresis; GST, glutathione S-transferase.
are comprised of 54 amino acids in L-MAG and 10 residues in S-MAG. In vivo and in vitro studies show that MAG is phos- phorylated mainly on serine but also to some extent on threo- nine and tyrosine residues (1, 8). We have previously shown that both L-MAG and S-MAG isoforms are phosphorylated when expressed in heterologous cells in culture (8). Whereas L-MAG is phosphorylated on serine and tyrosine residues, S- MAG is constitutively phosphorylated only on serine. The early appearance of L-MAG at the onset of myelination and its dif- ferential phosphorylation suggest a role for tyrosine phospho- rylation in the initial process of intercellular recognition and signal transduction.
MATERIALS AND METHODS
Cell Lines C5, an L cell fibroblast cell line expressing L-MAG was generated as
previously described (9). Stable S-MAG (SM32) and L-MAG mutant CY620F) expressing L cell lines were generated by transfection of cells with the pECEp67AX1 or mutated pECEp72 construct (see below) us- ing electroporation by following the manufacturer's directions (Bio- Rad). After 24 h, stable transfectants were selected by G418 resistance (600 pg/ml in oMEM, 10% FCS). Cell lines generated were screened by immunoblot analysis. Expression plasmids encoding S-MAG (pECE67AXl) or L-MAG (pECEp72) (10) were a kind gift from M. Tropak and Dr. J. Roder.
Primary rat oligodendrocyte precursors were generated from new- born rat cerebral hemispheres as described by Almazan et al. (11). Cells were expanded in serum-free medium in the presence of 2.5 ng/ml platelet derived growth factor-AA and 2.5 ng/ml basic fibroblast growth factor. After 5 days the precursors were allowed to differentiate in serum-free medium for 12-14 days prior to the experimental treatments.
Cytoplasmic Mutants of MAG Cytoplasmic Deletion Mutants of MAG-A unique XbaI site (under-
lined) was engineered by polymerase chain reaction (PCR) in the pa- rental pECEp72 plasmid with the following primers 5"GGGTCTAGAG- GTACAAATGACGCGGG-3' and 5"GGGTCTAGAAACCTCTACGG- CACC-3'. This allowed subcloning of the 600-bp XbaI-BamHI fragment from the 3'-end of the cDNA into Bluescript (Stratagene) without alter- ing the amino acid sequence (Ser-Arg) at position 490. Carboxyl-termi- nal deletions were generated in the resulting KS/MAGc plasmid by PCR using a common 3'-primer (5'-GGAAGATCl"I'GAGGAAGCTGGGGGC- 3') containing a termination codon (in bold) and the following nested set of primers to amplify truncations of the cytoplasmic domain:MA 539, 5"GGGAGATCTTTTTCTTCTTGTCTGGGT-3'; MA 563, 5"GGGAa- WAATTCGGGGCTGTAC-3'; LMA 576, 5"GGGAGATCTCTTCT- CACTCTCATACTT-3' and LMA 605, 5'-GGGAGATCTTTTCCCCAG- GTCTGAGT-3'. The underlined BglII site was used for intramolecular ligation. After sequencing, the cytoplasmic deletion mutants were subcloned back into the XbaI-BamHI site of the modified pECEp72 construct.
Site-directed Mutants of L-MAG-After subcloning of the XbaI- BamHI fragment into m13 mp18, oligonucleotide-directed mutagenesis was performed following manufacturer's directions (Amersham Corp.). After sequencing, the cytoplasmic site-directed mutants (Y558F and Y620F) were subcloned back into the XbaI-BamHI site of the modified pECEp72 construct.
27240

MAG Tyrosine Phosphorylation
Dansient Expression of MAG Proteins in COS-I Cells pECEp72, pECEIMA539, pECEIMA563, pECEnMA576, and pECE/
LMA605 were transfected into COS-1 monkey kidney cells using the method of Kaufman et al. (12). 18 h before transfection, COS-1 cells were plated a t a density of 1 x lo6 cells per IO-cm tissue culture plate. The cells were maintained in uMEM supplemented with 10% FCS and were washed twice in serum free aMEM immediately prior to treatment with DNA. Each plate of cells was transfected with 8 pg DNA, 0.1 M Tris (pH 7.3), DEAE-dextran a t 0.25 mg/ml in a total volume of 4 ml. After 12 h at 37 "C the plates were washed once in serum free aMEM and treated for 3 min a t room temperature with 2 ml of 10% dimethyl sulfoxide reagent. The cells were then incubated with aMEM, 10% FCS containing 0.1 mM chloroquin. After 2.5 h a t 37 "C, the cells were washed once in uMEM and were maintained for 72 h at 37 "C in aMEM, 10% FCS, at which point the cells were metabolically labeled as de- scribed below.
Phosphorylation of MAG in Cell Culture MAG expressing cell lines were labeled for 4 hours in the presence of
1 mCi of [32Plorthophosphate/ml of phosphate-free and serum free Dul- becco's modified Eagle's medium. Cells were treated with sodium vana- date (500 p ~ ) as previously described (8). Immunoprecipitation and immunoblotting procedures using anti-MAG monoclonal antibodies Gen S1 and Gen S3 were performed as previously documented (8). Samples were analyzed using SDS-PAGE.
Cells were lysed in lysis buffer (10 mM Tris HCI pH 7.5,150 mM NaCl, 1% Triton X-100,2 mM sodium fluoride, 2 mM sodium pyrophosphate, 2 pg/ml aprotinin, 5 pg/ml leupeptin, 200 pg/ml phenylmethylsulfonyl fluoride) containing 500 p~ (NH,), VO,, and lysates were incubated with antibody for 1 h at 4 "C. Subsequently rabbit anti-mouse antibod- ies (Jackson Immunoresearch Laboratory) were added for 1 h prior to addition of protein A-coupled Sepharose beads (Pharmacia Biotech Inc.). After an additional hour incubation the beads were washed five times with ice cold lysis buffer and proteins were eluted using SDS sample buffer. The immunoprecipitated samples were resolved by SDS- PAGE and detected by autoradiography.
Dyptic Digest Mapping Two-dimensional tryptic peptide mapping was essentially performed
as described by Luo et al. (13). Immobilon-P or nitrocellulose membrane slices containing 32P-labeled proteins were soaked in 0.5% polyvinylpyr- rolidone in 100 mM acetic acid for 30 min. After briefly washing in water, the membranes were incubated with 25 pg of trypsin in 50 mM ammo- nium bicarbonate (pH 8.5) a t 37 "C for 2 h. After 2 h, an additional 25 pg of trypsin were added for an overnight incubation. The samples were then centrifuged, and the supernatant was lyophilized. After one wash with 100 pl of water and lyophilization, the samples were oxidized in 100 p1 of performic acid for 1 h at 0 "C. 1 ml of water was then added, and the samples were lyophilized. The dried phosphopeptides were resuspended in 10 pl of chromatography buffer (14), spotted onto Cell Type-100 thin-layer plates, and separated by ascending chromatogra- phy (n-butanol(204 ml), acetic acid (50 ml), pyridine (143 ml), H,O (143 ml)) for 5 h. The chromatogram was dried and rotated 90 "; thin-layer electrophoresis was performed at pH 1.9 (88% formic acid (50 ml), acetic acid (156 ml), H,O (1890 ml)) for 40 min a t 1000 V. Tryptic phosphopep- tides were then visualized by autoradiography. In the case of mixing experiments, approximately equal amounts of radioactivity were com- bined and analyzed.
Phosphoamino Acid Analysis Phosphorylated proteins were immunoprecipitated, resolved on SDS-
PAGE, and transfemed to Immobilon-P membranes (Millipore). Radio- active bands corresponding to the immunoprecipitated proteins were excised from the membranes and hydrolyzed directly in 6 N double- distilled HCl for 75 min a t 110 "C as previously described (15). The hydrolysate was lyophilized, mixed with phosphoamino acid standards, spotted onto cell type-100 thin-layer cellulose plates (Sigma) and re- solved by two dimensional thin-layer electrophoresis as described by Cooper et al. (16). In the first dimension the samples are migrated a t pH 3.5 for 15 min at 1500 volts. The thin-layer is dried, rotated 90" and resolved at pH 1.9 for 30 minutes a t 1500 volts. Where indicated, ,*P- labeled phosphopeptides were recovered by scraping the cellulose off the thin-layer plates and eluting the peptides with 300 pl of pH 1.9 buffer. The samples were lyophilized and acid hydrolyzed as described above. The samples were resolved a t pH 3.5 for 25 min. Phosphoamino acid standards were detected by ninhydrin staining of the thin-layer plates. Radioactive phosphoamino acids were detected by autoradiog- raphy.
a
C
e
27241
b
d
FIG. 1. Tryptic digestion of L- and S-MAG phosphorylated in L cells. L cells expressing LMAG (C5 cells, panels a and b ) or S-MAG (SM32 cells,panels c and d ) were ,,P-labeled and left untreated (panels a and c) or treated with 500 p~ ammonium vanadate (panels b and d) . Phosphorylated MAG was purified and digested with trypsin. The tryp- tic fragments were resolved by thin-layer chromatography (TLC) and electrophoresis (TLE) as described under "Materials and Methods." The resulting thin-layer maps were visualized by autoradiography (3 days of exposure for S-MAG and 18 h of exposure for L-MAG). Phosphopeptides that were affected during vanadate treatment are identified with ar- rows. Panel e, phosphoamino acid analysis of tryptic peptides 1-3 (as indicated) derived from phosphorylated L-MAG. Positions of phospho- serine (PS) , phosphothreonine (PT) and phosphotyrosine (PY) are indicated.
Complex Formation with Fusion Proteins pGEX constructs encoding GST-PLCySH2 (N+C) (provided by G.
Gish) (17) or GST-Fyn(amino acids 1-255) (provided by J. Cambier) (18) were introduced into Escherichia coli DH5a and fusion protein induced with 1 mM isopropyl-1-thio-P-Dollactopyranoside (Boehringer Mann- heim). Expressed glutathione S-transferase (GST) fusion proteins were isolated from bacterial lysates by affinity chromatography with gluta- thione-Sepharose 4B beads (Pharmacia).
Cell lines expressing MAG or isolated mouse brains were treated with vanadate (as indicated) and lysed in lysis buffer (see above) with or without 500 PM Na,VO, (according to treatment). After centrifugation to remove particulate matter, approximately 500 pl of lysate (-500 pg of protein) was incubated with 5-10 pg of the indicated fusion protein overnight at 4 "C. After 3 4 washes with lysis buffer, samples were resuspended in SDS sample buffer and analyzed by SDS-PAGE and Western blotting as previously described (8) and detected by enhanced chemiluminescence (ECL) according to the manufacturer's directions (Amersham Corp.).
RESULTS Analysis of L-MAG and S-MAG Phosphorylation in Fibro-
blasts by Tryptic Peptide Mapping-Two-dimensional tryptic peptide mapping of L " A G phosphorylated in L cell fibroblasts revealed several phosphopeptides (Fig. la). Vanadate, a phos- photyrosine phosphatase inhibitor, stimulated the phosphoryl- ation of peptide 1 (Fig. lb). Phosphoamino acid analysis of the tryptic peptides revealed that peptide 1 contained solely phos- photyrosine whereas peptides 2 and 3 were phosphorylated on serine residues (Fig. le). Although other spots were detected following tryptic digestion of L-MAG expressed in fibroblasts, these phosphopeptides were not reproducibly seen nor observed

27242 MAG Tyrosine Phosphorylation
a 1 2
2oo- plmq e?- 1 M- :
C 1
b
I
‘s 2 ? T
2
2 + L-MAG
t TLC
-Ti. FIG. 2. Phosphorylation of MAG in primary rat oligodendro-
cyte cultures. Primary cultures of rat oligodendrocytes were labeled with [32Plorthophosphate. Panel a, MAG was immunoprecipitated from lysates of cells that were untreated (lane 1 ) or treated with 500 J ~ M vanadate (lane 2 ) and resolved by SDS-PAGE (7.5% polyacrylamide). Molecular mass standards are indicated in kilodaltons. Panel b, phos- phoamino acid analysis of immunoprecipitated MAG from untreated (panel 1 ) uersus vanadate-treated cells (panel 2). S, phosphoserine; T , phosphothreonine; Y , phosphotyrosine. Panel c, tryptic digest analysis
cells. The identities of the phosphopeptides are indicated by the num- of MAG from untreated (panel 1) uersus vanadate-treated (panel 2)
bering used in Fig. 1 and were verified by mixing MAG-derived tryptic digests from vanadate-treated oligodendrocytes with tryptic digests from L-MAG immunopurified from vanadate-treated fibroblasts (panel 2 + L-MAG). The tryptic digests were analyzed by thin-layer chroma- tography (TLC) and electrophoresis (TLE) as described under “Mate- rials and Methods.”
in primary oligodendrocyte cultures (see below) and not studied further,
Phosphorylation of S-MAG occurs primarily on serine resi- dues (8). Tryptic digest analysis of phosphorylated S-MAG re- vealed the presence of one major peptide, peptide b (Fig. IC). Vanadate treatment had no effect on the phosphopeptide pro- file (Fig. Id ). Purified peptide b comigrated with peptide 2 from L-MAG implying that they are identical (data not shown). Thus, while L-MAG and S-MAG have a common serine phos- phorylation site, L-MAG has additional tyrosine and serine phosphorylation sites, presumably in the region of its unique cytoplasmic terminus.
Phosphorylation of MAG in Oligodendrocytes-We wanted to determine if the phosphorylation sites we had identified in MAG expressing fibroblasts resembled the sites phosphoryl- ated during myelination. Rat oligodendrocyte precursors were differentiated for 12-14 days in serum free medium and then incubated in the presence of [32P]orthophosphate. The cultures were comprised of 100% MAG positive cells as determined by immunofluorescent cytochemistry (data not shown). 32P-La- beled MAG was immunoprecipitated from untreated or vana- date treated cells and analyzed by SDS-PAGE (Fig. 2a). Tryptic
digest analysis of L-MAG derived from oligodendrocytes re- vealed the presence of three phosphopeptides with identical mobilities to those of L-MAG expressed in fibroblasts (Fig. 2c, panel 2 + L-MAG; note the increase in intensity of peptide 3). Phosphorylation of MAG in oligodendrocytes was enhanced 2-3 fold in the presence of vanadate. This increase in phosphoryl- ation occurred mainly on the tyrosine residuek) (Fig. 2b, panel 2 ) found in peptide 1 (Fig. 2c, compare panel 1 with 2) . The tryptic map data identify the phosphorylated MAG in oligoden- drocytes as L-MAG. This is confirmed by chemical deglycosyl- ation experiments, which indicate that the MAG isoform ex- pressed in oligodendrocytes at this stage is identical to L-MAG and not S-MAG (data not shown).
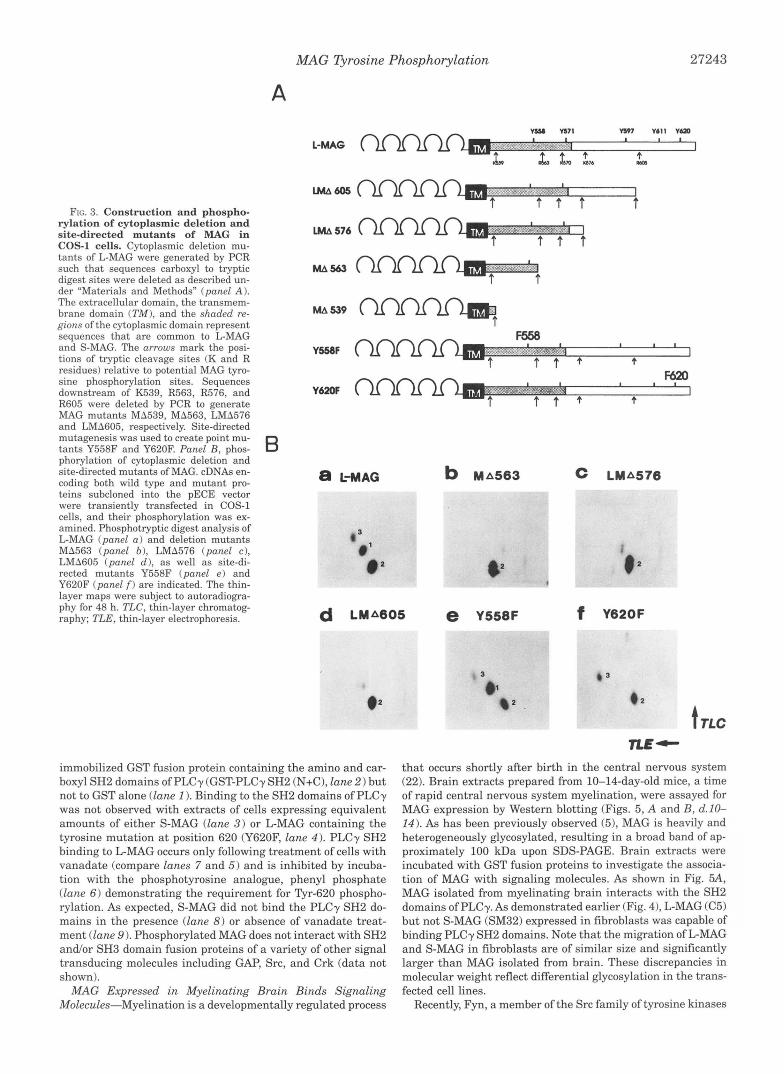
Phosphorylation of Deletion and Site-directed Mutants of MAG in COS-1 Cells-Deletion mutants LMA605, LMA576, MA563 and MA539 were constructed (Fig. 3A) to delineate the region of tyrosine phosphorylation in L-MAG. The mutant pro- teins were transiently expressed in COS-1 cells, and their phos- phorylation status was determined. All cells expressing mutant MAG isoforms were analyzed by Western blotting and immu- nofluorescence to confirm that approximately equivalent amounts of MAG were expressed at the cell surface (data not shown). MA539 had all cytoplasmic residues deleted with the exception of the stop transfer sequence immediately following the transmembrane domain. While this mutant was not phos- phorylated (data not shown), all other MAG mutants were phosphorylated in COS-1 cells (see below). Phosphotryptic di- gest analysis of L-MAG revealed the presence of peptides 1, 2 and 3 (Fig. 3B, panel a), as has been previously demonstrated in L cells and oligodendrocytes. In deletion mutants MA563 (panel b) , LMA576 (panel c ) and LMA605 (panel d), phospho- rylation of peptide 1 was absent while phosphopeptide 2 was unaltered. The presence of a faint phosphopeptide migrating in the vicinity of peptide 1 in deletion mutants LMA576 and LMA605 has also been detected in tryptic peptide maps of S- MAG and most likely represents a minor phosphorylation site common to L- and S-MAG. The presence of phosphopeptide 2 in deletion mutant MA563 but not in MA539 indicates that the juxtamembrane region of MAG, which contains several serine residues, is the site of serine phosphorylation common to L- and S-MAG. Phosphopeptide 3 was absent in all MAG deletion mu- tants demonstrating that the carboxyl-terminal region of L- MAG harbors or is necessary for serine phosphorylation of this site.
The above results suggest that the major tyrosine phospho- rylation site of L-MAG resides within the last 21 amino acids of the carboxyl terminus. Since Tyr-620 is located in a region that shares sequence similarity with one of the epidermal growth factor receptor autophosphorylation sites (191, we reasoned that this residue may be the tyrosine phosphorylation site con- tained within peptide 1. To test this, a cDNA construct of L- MAG in which this amino acid residue was mutated to a phen- ylalanine residue (Y620F) was generated. This mutant cDNA (Y620F) was expressed in COS-1 cells, and its phosphorylation was examined as above. While phosphotryptic digest analysis of another site-directed mutant (Y558F) revealed the presence of peptides 1 , 2 and 3 (Fig. 3B, panel e), phosphopeptide 1 was clearly absent in the Y620F MAG mutant (panel f). This result identifies the major tyrosine phosphorylated residue of L-MAG as Tyr-620.
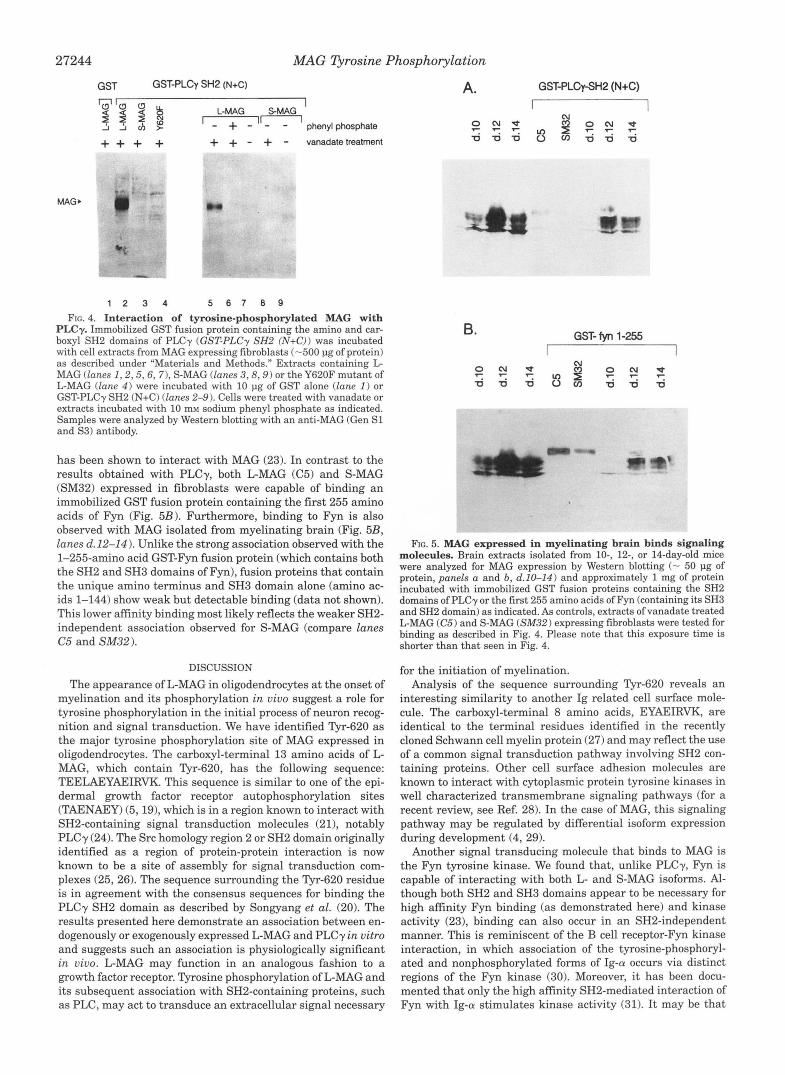
Interaction of MAG with Phospholipase Cy-Tyrosine 620 is contained within a sequence that resembles the binding site for the SH2 domains of certain signal transduction molecules (19- 21). To test for an interaction of MAG with various signaling proteins, an in vitro binding assay was utilized (17,18). As seen in Fig. 4, L-MAG expressed in L cells was able to bind to
MAG Tyrosine Phosphorylation 27243
FIG. 3. Construction and phospho- rylation of cytoplasmic deletion and site-directed mutants of MAG in COS-1 cells. Cytoplasmic deletion mu- tants of L-MAG were generated by PCR such that sequences carboxyl to tryptic digest sites were deleted as described un- der "Materials and Methods" (panel A). The extracellular domain, the transmem- brane domain ( T M ) , and the shaded re- gions of the cytoplasmic domain represent sequences that are common to L-MAG and S-MAG. The arrows mark the posi- tions of tryptic cleavage sites (K and R residues) relative to potential MAG tyro- sine phosphorylation sites. Sequences downstream of K539, R563, R576, and R605 were deleted by PCR to generate MAG mutants "539, MA563, LMA576 and LMA605, respectively. Site-directed mutagenesis was used to create point mu- tants Y558F and Y620F. Panel B, phos- phorylation of cytoplasmic deletion and site-directed mutants of MAG. cDNAs en- coding both wild type and mutant pro- teins subcloned into the pECE vector were transiently transfected in COS-1 cells, and their phosphorylation was ex- amined. Phosphotryptic digest analysis of L-MAG (panel a ) and deletion mutants MA563 (panel b ) , LMA576 (panel c ) , LMA605 (panel c l ) , as well as site-di- rected mutants Y558F (panel e) and Y620F (panel f) are indicated. The thin- layer maps were subject to autoradiogra- phy for 48 h. TLC, thin-layer chromatog- raphy; TLE, thin-layer electrophoresis.
A
MA 539 T , .
Y620F
B a L-AIAG
immobilized GST fusion protein containing the amino and car- boxyl SH2 domains of PLCy (GST-PLCy SH2 (N+C), lane 2) but not to GST alone (lane 1). Binding to the SH2 domains of PLCy was not observed with extracts of cells expressing equivalent amounts of either S-MAG (lane 3 ) or L-MAG containing the tyrosine mutation a t position 620 (Y620F, lane 4). PLCy SH2 binding to L-MAG occurs only following treatment of cells with vanadate (compare lanes 7 and 5) and is inhibited by incuba- tion with the phosphotyrosine analogue, phenyl phosphate (lane 6 ) demonstrating the requirement for Tyr-620 phospho- rylation. As expected, S-MAG did not bind the PLCy SH2 do- mains in the presence (lane 8) or absence of vanadate treat- ment (lane 9). Phosphorylated MAG does not interact with SH2 and/or SH3 domain fusion proteins of a variety of other signal transducing molecules including GAP, Src, and Crk (data not shown).
MAG Expressed in Myelinating Brain Binds Signaling Molecules-Myelination is a developmentally regulated process
b Ma563
e YWF
C
f
LMA576
Y620F
TLE- that occurs shortly after birth in the central nervous system (22). Brain extracts prepared from 10-14-day-old mice, a time of rapid central nervous system myelination, were assayed for MAG expression by Western blotting (Figs. 5, A and B, d.10- 14). As has been previously observed (5), MAG is heavily and heterogeneously glycosylated, resulting in a broad band of ap- proximately 100 kDa upon SDS-PAGE. Brain extracts were incubated with GST fusion proteins to investigate the associa- tion of MAG with signaling molecules. As shown in Fig. 5A, MAG isolated from myelinating brain interacts with the SH2 domains of PLCy. As demonstrated earlier (Fig. 41, L-MAG (C5) but not S-MAG (SM32) expressed in fibroblasts was capable of binding PLCy SH2 domains. Note that the migration of L-MAG and S-MAG in fibroblasts are of similar size and significantly larger than MAG isolated from brain. These discrepancies in molecular weight reflect differential glycosylation in the trans- fected cell lines.
Recently, Fyn, a member of the Src family of tyrosine kinases
27244 MAG nrosine Phosphorylation
GST GST-PLCy SH2 (N+C) - I
‘0”a 0 L-MAG SMAG I
’ - 4- - ’ n phenylphosphate
+ + + + + + - + - vanadate treatment
1 2 3 4 5 6 1 8 9
FIG. 4. Interaction of tyrosine-phosphorylated MAG with PLCy. Immobilized GST fusion protein containing the amino and car- boxyl SH2 domains of PLCy (GST-PLCy SH2 (N+C)) was incubated with cell extracts from MAG expressing fibroblasts (-500 pg of protein) as described under “Materials and Methods.” Extracts containing L- MAG (lanes 1 ,2 ,5 ,6 , 7), S-MAG (lanes 3 , 8 , 9 ) or the Y620F mutant of LMAG (lane 4 ) were incubated with 10 pg of GST alone (lane 1 ) or GST-PLCy SH2 (N+C) (lanes 2-9). Cells were treated with vanadate or extracts incubated with 10 mM sodium phenyl phosphate as indicated. Samples were analyzed by Western blotting with an anti-MAG (Gen S1 and S3) antibody.
has been shown to interact with MAG (23). In contrast to the results obtained with PLCy, both L-MAG ((35) and S-MAG (SM32) expressed in fibroblasts were capable of binding an immobilized GST fusion protein containing the first 255 amino acids of Fyn (Fig. 5B). Furthermore, binding to Fyn is also observed with MAG isolated from myelinating brain (Fig. 5B, lanes (1.12-14). Unlike the strong association observed with the 1-255-amino acid GST-Fyn fusion protein (which contains both the SH2 and SH3 domains of Fyn), fusion proteins that contain the unique amino terminus and SH3 domain alone (amino ac- ids 1-144) show weak but detectable binding (data not shown). This lower affinity binding most likely reflects the weaker SH2- independent association observed for S-MAG (compare lanes C5 and SM32).
DISCUSSION
The appearance of L-MAG in oligodendrocytes at the onset of myelination and its phosphorylation in vivo suggest a role for tyrosine phosphorylation in the initial process of neuron recog- nition and signal transduction. We have identified Tyr-620 as the major tyrosine phosphorylation site of MAG expressed in oligodendrocytes. The carboxyl-terminal 13 amino acids of L- MAG, which contain Tyr-620, has the following sequence: TEELAEYAEIRVK. This sequence is similar to one of the epi- dermal growth factor receptor autophosphorylation sites (TAENAEY) (5,19), which is in a region known to interact with SH2-containing signal transduction molecules (211, notably PLCy (24). The Src homology region 2 or SH2 domain originally identified as a region of protein-protein interaction is now known to be a site of assembly for signal transduction com- plexes (25,26). The sequence surrounding the Tyr-620 residue is in agreement with the consensus sequences for binding the PLCy SH2 domain as described by Songyang et al. (20). The results presented here demonstrate an association between en- dogenously or exogenously expressed L-MAG and PLCy in vitro and suggests such an association is physiologically significant in vivo. L-MAG may function in an analogous fashion to a growth factor receptor. Tyrosine phosphorylation of L-MAG and its subsequent association with SH2-containing proteins, such as PLC, may act to transduce an extracellular signal necessary
A.
e u
GST-PLQ-SHP (N+C)
I I
B. GST- fyn 1-255 I I
FIG. 5. MAG expressed in myelinating brain binds signaling molecules. Brain extracts isolated from IO-, 12-, or 14-day-old mice were analyzed for MAG expression by Western blotting (- 50 pg of protein, panels a and b, d.10-14) and approximately 1 mg of protein incubated with immobilized GST fusion proteins containing the SH2 domains of PLCy or the first 255 amino acids of Fyn (containing its SH3 and SH2 domain) as indicated. As controls, extracts of vanadate treated L-MAG (C5) and S-MAG (SM32) expressing fibroblasts were tested for binding as described in Fig. 4. Please note that this exposure time is shorter than that seen in Fig. 4.
for the initiation of myelination. Analysis of the sequence surrounding Tyr-620 reveals an
interesting similarity to another Ig related cell surface mole- cule. The carboxyl-terminal 8 amino acids, EYAEIRVK, are identical to the terminal residues identified in the recently cloned Schwann cell myelin protein (27) and may reflect the use of a common signal transduction pathway involving SH2 con- taining proteins. Other cell surface adhesion molecules are known to interact with cytoplasmic protein tyrosine kinases in well characterized transmembrane signaling pathways (for a recent review, see Ref. 28). In the case of MAG, this signaling pathway may be regulated by differential isoform expression during development (4, 29).
Another signal transducing molecule that binds to MAG is the Fyn tyrosine kinase. We found that, unlike PLCy, Fyn is capable of interacting with both L- and S-MAG isoforms. Al- though both SH2 and SH3 domains appear to be necessary for high affinity Fyn binding (as demonstrated here) and kinase activity (23), binding can also occur in an SH2-independent manner. This is reminiscent of the B cell receptor-Fyn kinase interaction, in which association of the tyrosine-phosphoryl- ated and nonphosphorylated forms of Ig-a occurs via distinct regions of the Fyn kinase (30). Moreover, it has been docu- mented that only the high affinity SH2-mediated interaction of Fyn with Ig-a stimulates kinase activity (31). It may be that

MAG Tyrosine Phosphorylation 27245
although Fyn can bind both MAG isoforms, it becomes acti- vated only when complexed to L-MAG. Consistent with this is the observation that antibody cross-linking of L-MAG but not S-MAG causes an increase in Fyn kinase activity (23).
The results presented here demonstrate that MAG interacts with PLCy and Fyn through different sites. Taken together, these observations as well as those of others (231, suggest a model wherein Fyn constitutively binds to MAG in a latent form. Ligand stimulation of L-MAG would result in activation of Fyn kinase and phosphorylation of Tyr-620. Binding and activation of PLC y through this phosphotyrosine residue would contribute to the signaling pathway involved in the regulation of myelination.
Acknowledgments-We gratefully acknowledge the excellent techni- cal assistance of David Joyce during the course of this work and thank members of the laboratory as well as Drs. Douglas Gray and Xingquan Liu for helpful discussions and comments on the manuscript. We thank
GST-PLCy SH2 fusion proteins, Christopher Pleiman and John Cam- Gerald Gish and Tony Pawson for providing bacterial clones expressing
bier for pGEX-gst fyn constructs, and Mike Tropak and John Roder for providing MAG expression constructs.
REFERENCES 1. Edwards, A. M., Arquint, M., Braun, P. E., Roder, J. C., Dunn, R. J., Pawson,
T., and Bell, J. C. (1988) Mol. Cell. Biol. 8, 2655-2658 2. Arquint, M., Roder, J., Chia, L.-S., Down, J., Wilkinson, D., Bayley, H., Braun,
P. E., and Dunn, R. (1987) Proc. Natl. Acad. Sci. U. S. A. 84, 600-604 3. Lai, C., Brow, M. A,, Nave, K.-A., Noronha, A. B., Quarles, R. H., Bloom, F. E.,
Milner, R. J., and Sutcliffe, J. G. (1987) Proc. Natl. Acad. Sci. U. S. A. 84, 4337-4341
4. Pedraza, L., Frey, A. B., Hempstead, B. L., Colman, D. R., and Salzer, J. L.
5. Salzer, J. L., Holmes, W. P., and Colman, D. R. (1987) J. Cell Biol. 104,957-965 (1991) J. Neurosci. Res. 29, 141-148
6. Higgins, G. A,, Schmale, H., Bloom, F. E., Wilson, M. C., and Milner, R. J.
7. DEustachio, P., Colman, D. R., and Salzer, J. L. (1988) J. Neurochem. 50,
8. Afar, D. E. H., Salzer, J. L., Roder, J., Braun, P. E., and Bell, J. C. (1990) J.
(1989) Proc. Nutl. Acad. Sci. U. S. A. 86, 2074-2078
589-593
9. Afar, D. E. H., Manus, R. M., Salzer, J. L., Stanners, C. P., Braun, P. E., and Neurochem. 55, 1418-1426
10. Johnson, P. W., Abramow-Newerly, W., Seilheimer, B., Sadoul, R., Tropak, M. Bell, J. C. (1991) J. Neurosci. Res. 29, 429-436
B., Arquint, M., Dunn, R. J., Schachner, M., and Roder, J. C. (1989) Neuron 3,377-385
11. Almazan, G., Afar, D. E. H., and Bell, J. C. (1993) J. Neurosci. Res. 36,163-172 12. Kaufman, R. J., Davies, M. V., Pathak, V. K., and Hersey, J. W. B. (1989) Mol.
14. Brown, E. G., and Prevec, L. (1978) Virology 89, 7-21 13. Luo, K., Hurley, T. R., and Sefion, B. M. (1990) Oncogene 5,921-923
15. Kamps, M. P., and Sefton, B. M. (1989) Anal. Biochem. 176,22-27 16. Cooper, J. A., Sefton, B. M., and Hunter, T. (1983) Methods Enzymol. 99,
387-405 17. Pen, K. G., Gervais, F. G., Weil, R., Davidson, D., Gish, G. D., and Veillette, A.
(1993) Oncogene 8,2765-2772 18. Pleiman, C. M., Clark, M. R., Timson Gauen, L. K., Winitz, S., Coggeshall, K.
M., Johnson, G. L., Shaw,A. S., andcambier, J. C. (1993)Mol. Cell. Biol. 13, 5877-5887
19. Downward, J., Parker, P., and Waterfield, M. D. (1984) Nature 311,483-485 20. Songyang, Z., Shoelson, S. E., Chaudhuri, M., Gish, G., Pawson, T., Haser, W.
G., King, F., Roberts, T., Ratnofsky, S., Lechleider, R. J., Neel, B. G., Birge, R. B., Fajardo, J. E., Chou, M. M., Hanafusa, H., Schaffhausen, B., and Cantley, L. (1993) Cell 72, 767-778
21. Moran, M. E, Koch, C. A., Anderson, D., Ellis, C., England, L., Martin, G. S., and Pawson, T. (1990) Proc. Natl. Acad. Sci. U. S. A. 87, 8622-8626
22. Banik, N. L., and Smith, M. E. (1977) Biochem. J. 162, 247-255 23. Umemori, H., Satm, S., Yagi, T., Aizawa, S., and Yamamoto, T. (1994) Nature
367, 572676 24. Rotin, D., Margolis, B., Mohammadi, M., Daly, R. J., Daum, G., Li, N., Fischer,
E. H., Burgess, W. H., Ullrich, A., and Schlessinger, J. (1992) EMBO J. 11, 559-567
25. Sadowski, I., Stone, J. C., and Pawson, T. (1986) Mol. Cell. Biol. 6,4396-4408 26. Koch, C. A,, Anderson, D., Moran, M. F., Ellis, C., and Pawson, T. (1991)
27. Dulac, C., Tropak, M. B., Cameron-Curry, P., Rosier, J., Marshak, D. R., Science 252,668-674
28. Rudd, C. E., Janssen, O., Prasad, K. V. S., Raab, M., da Silva, A,, Telfer, J. C., Roder, J., and Le Douarin, N. M. (1992) Neuron 8,323-334
29. Tropak, M. B., Johnson, P. W., Dunn, R. J., and Roder, J. C. (1988) Mol. Brain and Yamamoto, M. (1993) Biochim. Biophys. Acta 1155, 239-266
30. Pleiman, C. M.,Abrams, C., Timson Gauen, L., Bedzyk, W., Jongstra, J., Shaw, Res. 4, 143-155
A. S., and Cambier, J. C. (1994) Proc. Natl. Acad. Sci. U. S. A. 91, 4268- 4272
31. Clark, M. R., Johnson, S.A., and Cambier, J. C. (1994)EMBO J. 13,1911-1919
Cell. Biol. 9, 946-958