cabyr, a novel calcium-binding tyrosine ... - core.ac.uk · cabyr, a novel calcium-binding tyrosine...
TRANSCRIPT

Developmental Biology 242, 236–254 (2002)doi:10.1006/dbio.2001.0527, available online at http://www.idealibrary.com on
CABYR, a Novel Calcium-Binding TyrosinePhosphorylation-Regulated Fibrous SheathProtein Involved in Capacitation
Soren Naaby-Hansen,* ,1 Arabinda Mandal,1 Michael J. Wolkowicz,Buer Sen, V. Anne Westbrook, Jagathpala Shetty, Scott A. Coonrod,Kenneth L. Klotz, Young-Howan Kim, Leigh Ann Bush,Charles J. Flickinger, and John C. Herr2
*Ludwig Institute for Cancer Research, Royal Free and University College School of Medicine,London W1P 8BT, United Kingdom; and Department of Cell Biology,University of Virginia, Charlottesville, Virginia 22908
To reach fertilization competence, sperm undergo an incompletely understood series of morphological and molecularmaturational processes, termed capacitation, involving, among other processes, protein tyrosine phosphorylation andincreased intracellular calcium. Hyperactivated motility and an ability to undergo the acrosome reaction serve asphysiological end points to assess successful capacitation. We report here that acidic (pI 4.0) 86-kDa isoforms of a novel,polymorphic, testis-specific protein, designated calcium-binding tyrosine phosphorylation-regulated protein (CABYR), weretyrosine phosphorylated during in vitro capacitation and bound 45Ca on 2D gels. Acidic 86-kDa calcium-binding forms ofCABYR increased during in vitro capacitation, and calcium binding to these acidic forms was abolished by dephosphory-lation with alkaline phosphatase. Six variants of CABYR containing two coding regions (CR-A and CR-B) were cloned fromhuman testis cDNA libraries, including five variants with alternative splice deletions. A motif homologous to the RIIdimerization domain of PK-A was present in the N-terminus of CR-A in four CABYR variants. A single putative EF handlikemotif was noted in CR-A at aas 197–209, while seven potential tyrosine phosphorylation-like sites were noted in CR-A andfour in CR-B. Pro-X-X-Pro (PXXP) modules were identified in the N- and C-termini of CR-A and CR-B. CABYR localizes tothe principal piece of the human sperm flagellum in association with the fibrous sheath and is the first demonstration of asperm protein that gains calcium-binding capacity when phosphorylated during capacitation. © 2002 Elsevier Science (USA)
Key Words: CABYR; spermatozoa; capacitation; calcium-binding protein; fibrous sheath; tyrosine phosphorylation;postmeiotic expression.
INTRODUCTION
Fertilization capacity is acquired by spermatozoa onlyafter residence in the distinct microenvironments of theuterus or oviduct (depending on the species) for a finiteperiod of time. The necessary series of changes, termedcapacitation, was first described independently by Chang(1951, 1955) and Austin (1951). Capacitation involves mo-
1 These authors contributed equally to this work.2 To whom correspondence should be addressed at Department
of Cell Biology, P.O. Box 800732, University of Virginia HealthSystem, Charlottesville, VA 22908. Fax: (804) 982-3912. E-mail:
[email protected].236
lecular changes in both the sperm head and tail, whichallow defined physiological end points to occur, such asmotility hyperactivation, a whiplash-like sperm tail mo-tion, and regulated acrosomal exocytosis (Yanagimachi,1994). Hyperactivation is observed when sperm reach theoocyte and increase their flagellar bend amplitude and beatasymmetry (Yanagimachi, 1970; Suarez, 1988), which arethought to enhance the ability of sperm to penetrate the eggvestments by increasing forward progression and lateralflagellar thrust (Suarez et al., 1993).
Our understanding of the molecular mechanisms under-lying capacitation and hyperactivation is rudimentary atpresent but there is evidence that Ca21, cAMP, and protein
tyrosine phosphorylation are involved. Capacitation can be0012-1606/02 $35.00© 2002 Elsevier Science (USA)
All rights reserved.

237CABYR, a Novel Fibrous Sheath Protein
accomplished in vitro using cauda epididymal or ejaculatedsperm incubated in defined media containing a proteinsource such as albumin, NaHCO3, and Ca21, and energysubstrates such as glucose, pyruvate, or lactate. Conditionsconducive to in vitro capacitation lead to increased tyrosinephosphorylation of a subset of proteins in both mouse(Visconti et al., 1995a,b) and human sperm (Leclerc et al.,1996; Osheroff et al., 1999). The removal of albumin,NaHCO3, or Ca21 from capacitation media prevents theoccurrence of both tyrosine phosphorylation and capacita-tion (Visconti et al., 1995a). Two protein substrates for thiscapacitation-related phosphorylation are members of the Akinase anchoring protein family, AKAP4 (originally calledAKAP82 or Fsc1 in mouse) and AKAP3 (originally calledAKAP95T, FSP95, or AKAP110), which are components ofthe fibrous sheath of the sperm tail (Mandal et al., 1999;Fulcher et al., 1995; Turner et al., 1998).
Although little is known about the kinetics of intracel-lular calcium during capacitation (Yanagmachi, 1994), amassive influx of Ca21 occurs during the acrosome reaction(Thomas and Meizel, 1988, 1989), and extracellular calciumis required for sperm hyperactivation. If hyperactivatedsperm are transferred to calcium-free media for 30–60 min,none is hyperactive, but hyperactivation can be restored byaddition of 2 mM calcium (Yanagimachi, 1982). Calciumcan increase flagellar bend amplitude in demembranatedsperm (Lindemann and Goltz, 1988; Okuno and Brokaw,1981). Intracellular calcium [Ca21]in is increased in hyper-activated sperm in both the head and tail, and [Ca21]in
oscillates with each flagellar bend (Suarez et al., 1993),indicating a direct relationship between intracellular cal-cium and hyperactivation.
The cytosolic level of cAMP increases during capacita-tion (Visconti and Tezon, 1989). Pharmacological stimu-lants that elevate intracellular cAMP, such as the phospho-diesterase inhibitors, caffeine, and pentoxifylline, enhancesperm hyperactivated motility (Yovich et al., 1990), en-hance penetration of cervical mucus, increase tight bindingto homologous zona pellucida (Nassar et al., 1999a,b), andincrease fertilization (Johnston et al., 1994). Calcium/calmodulin is an activator of both mammalian spermadenylate cyclase (AC) (Gross et al., 1987; Hyne and Gar-bers, 1979) and cyclic nucleotide phosphodiesterase (Wascoand Orr, 1984), and sperm AC is stimulated by HCO3
2
anions (Garty and Salomon, 1987; Visconti and Kopf, 1998).A soluble testicular adenylate cyclase has recently beencloned (Buck et al., 1999) and shown to be sensitive tobicarbonate (Chen et al., 2000). Sperm protein tyrosinephosphorylation is accelerated by cAMP agonists, whileantagonists of PKA inhibit tyrosine phosphorylation andcapacitation (Galantino-Homer et al., 1997). These andother observations (Leclerc et al., 1996; Vijayaraghavan etal., 1999; Carrera et al., 1996) suggest that sperm proteintyrosine phosphorylation and capacitation are under theregulation of a cAMP/PKA pathway, which is activated byelevated cytosolic levels of calcium and HCO2 anions.
3Mammalian sperm contain all three subtypes of the gua-
© 2002 Elsevier Science (USA
nine nucleotide-binding regulatory proteins Gi (Glassner etal., 1991), and G proteins have been localized in particularto the sperm tail (Garty et al., 1988; Hinsch et al., 1992)where protein kinases A and C (Parisel et al., 1984; Rotemet al., 1990) have also been reported. A cyclic nucleotide-gated Ca21 channel in mammalian sperm plasma mem-branes has been reported (Wiesner et al., 1998; Weyand etal., 1998), and N- and R-type Ca21 channels have beendefined in mouse sperm (Wennemuth et al., 2000).
We have initiated a proteomic-based strategy to identifytargets at the intersection between the calcium and proteintyrosine kinase signal transduction pathways in humanspermatozoa using two-dimensional (2D) gel analysis toidentify sperm calcium-binding proteins and proteins phos-phorylated by tyrosine kinases. The present report describesthe sequencing, cloning, expression, and characterization ofa novel testis and sperm-specific, calcium-binding protein,CABYR, that is expressed postmeiotically and localized inthe sperm flagellum. This protein shows increases in acidic86-kDa isoforms following capacitation, exhibits increasedtyrosine phosphorylation during in vitro capacitation, andbinds calcium specifically to acidic 86-kDa phosphorylatedisoforms that are sensitive to dephosphorylation.
MATERIALS AND METHODS
Solubilization and Electrophoresis of HumanSpermatozoal Proteins
Preparation of semen specimens and solubilization of spermproteins were performed as previously described (Naaby-Hansen etal., 1997a). For analytical two-dimensional (2D) electrophoresis thedetergent/urea-extracted proteins in the presence of protease in-hibitors were separated by isoelectric focusing (IEF) in acrylamidetube gels prior to 2D gel electrophoresis (SDS–PAGE), which wasperformed in a Protean II xi Multi-Cell apparatus (Bio-Rad, Rich-mond, CA) or on large-format (23 3 23 cm) gels (Investigator 2-DElectrophoresis System; ESA), which were also employed for pre-parative 2D gel electrophoresis. Electrotransfer to nitrocellulosemembranes and subsequent visualizing of the proteins by goldstaining was accomplished as previously described (Naaby-Hansenet al., 1997a), while electrotransfer to PVDF membranes (0.2 mmpore size; Pierce) was carried out as described by Henzel et al.(1993) using the transfer buffer composition of Matsudaira (1987)(10 mM 3-[cyclohexylamino]-1-propanesulfonic acid, 10% metha-nol, pH 11). The immobilized proteins were visualized by stainingin a solution containing 0.1% Coomassie R250, 40% methanol,and 0.1% acetic acid for 1 min, followed by destaining in a solutionof 10% acetic acid and 50% methanol three times for 3 min each.
In Vitro Capacitation
Motile sperm were harvested by the swim-up method of Bronsonand Fusi (1990). A control sample was removed and snap frozen(270°C), while the remaining sperm were resuspended in one of thefollowing media: Dulbecco’s PBS, BWW, human tubal fluid (HTF;Irvine Scientific, Santa Ana, CA) plus HSA (30 mg/ml), HTF plusHSA plus 2, 20, or 100 mM progesterone, HTF plus HSA plus 100
mM progesterone plus either 100, 200, or 400 mM of genistein or). All rights reserved.

238 Naaby-Hansen et al.
daidzein (Akiyama et al., 1987). Capacitation was achieved byincubating the samples at 37°C in 5% CO2 with sperm removed atvarious time points and isolated by centrifugation.
Detection of Calcium-Binding Proteins
Calcium-binding proteins were demonstrated using a 45Ca over-lay assay modified from that described by Maruyama et al. (1984).The experiment was replicated four times. In brief, the 2D gel-separated proteins were transferred to PVDF membranes (Jeth-malani et al., 1994), and the membranes were washed three timesfor 20 min each in a washing buffer (10 mM imidazole HCl, 60 mMKCl, and 5 mM MgCl2, pH 6.8) and incubated with 2 mCi/ml of45CaCl2 in washing buffer for 30 min at room temperature. Themembranes were subsequently rinsed for 2 min in distilled H2Ofollowed by a 30-s rinsing in 50% ethanol and were air dried onfilter paper for 15–20 min. The membranes were then dried by hotair from a hair dryer and exposed on phosphorimaging screens(Molecular Dynamics, Sunnyvale, CA) for 10 days. The use ofPVDF, shortening of the final wash steps, and employment ofphosphorimaging detection increased the signal-to-noise ratiocompared to that achieved with the procedure originally proposedby Maruyama et al. (1984). Some of the PVDF membranes weresubsequently stained with Coomassie to localize the calcium-binding proteins within the total 2D protein pattern, while othermembranes were used for Western blot analysis as described below.Computerized pattern analysis and densitometry of the autoradio-grams and the stained membranes were performed employing the2D Analyzer software (BioImage 2000), as previously described(Naaby-Hansen et al., 1997a,b).
Generation of Antiserum against Gel-PurifiedCABYR
The 86-kDa Coomassie-stained protein spot was cored fromthree 1.5-mm-thick 2D SDS–PAGE gels of human sperm extracts.The gel cylinders were minced into a slurry in 1 ml of PBS andemulsified with an equal volume of complete Freund’s adjuvant. A600 ml sample of this emulsion was intradermally injected into aNew Zealand white rabbit, followed by two monthly subcutaneousbooster injections of similarly prepared antigen with incompleteFreund’s adjuvant. Serum was collected 10 days after each boosterinjection.
Dephosphorylation of Sperm Proteins
To examine the relationship between phosphorylation andcalcium-binding capacity of the 86-kDa CABYR form, sperm fromfour individuals were capacitated for 5 h in HTF plus albumin, andthe sperm were extracted in NP-40/urea and the extracts pooled.The lysate was divided and one aliquot was treated with 2 U/mlcalf intestinal alkaline phosphatase (Boehringer Manheim) for 0.5 hat 37°C, while the other aliquot remained untreated.
Microsequencing of the 86-kDa Calcium-BindingTyrosine-Phosphorylated Protein
The 86-kDa Coomassie-stained protein spot was cored from a1.5-mm-thick 2D SDS–polyacrylamide gel and fragmented intosmaller pieces. The protein was destained in methanol, reduced in
10 mM dithiothreitol, and alkylated in 50 mM iodoacetamide in© 2002 Elsevier Science (USA
0.1 M ammonium bicarbonate. After removing the reagents, the gelpieces were incubated with 12.5 ng/ml trypsin in 50 mM ammo-nium bicarbonate overnight at 37°C. Peptides were extracted fromthe gel pieces in 50% acetonitrile in 5% formic acid and micro-sequenced by tandem mass spectrometry and by Edman degrada-tion at the Biomolecular Research Facility of the University ofVirginia. Five peptide sequences boxed in Fig. 3 were obtained bymass spectrometry. Differentiation of leucine and isoleucine in thesequences was determined by Edman sequencing of HPLC isolatedpeptides.
Cloning, Sequencing, and Analysis of cDNAs
A degenerate deoxyinosine containing sense primer (59-GGI-CAG-CCI-GAG-GTI-CCI-GCI-CAA/G-C/TT-39) was designedfrom peptide GQPEVPAQL and obtained from GIBCO BRL (LifeTechnologies, Gaithersburg, MD). Using this forward primer andan adapter primer (AP1), a 39-RACE (rapid amplification of cDNAends) PCR was performed with 0.25 ng of human testicularMarathon-ready cDNA (Clontech, Palo Alto, CA) in a 25 ml assaysystem for 40 cycles. Thermal cycling was done in an MJ Research(Watertown, MA) thermal cycler (PTC-200 DNA engine) using aprogram of one 3-min cycle at 94°C followed by 40 cycles ofdenaturation, annealing, and elongation at 94°C for 30 s, 60°C for 1min, and 68°C for 2 min. PCR products were separated on a 1.7%NuSieve (FMC BioProducts, Rockland, ME) agarose gel and aunique 1.0-kb DNA fragment was reamplified, cloned into the pCR2.1-TOPO vector (Invitrogen, San Diego, CA), and sequenced on aPerkin–Elmer Applied Biosystems DNA sequencer using BigDyefluorescence dye terminator chemistry with Taq DNA polymerase(Perkin–Elmer, Branchburg, NJ). The 39 clone contained 1001 bp,including a portion of coding region A (CR-A) and all of codingregion B (CR-B). The 59 end of the cDNA was also amplified by 59RACE PCR from the same template using an adapter primer (AP1)and an antisense 39 gene-specific primer (59-TTA-TTC-AGC-TGT-TGA-TTC-CCC-TTC-TGG-TTC-AAT-TTC-TGG-39), which was263 bp downstream from the 59 end of the 1.0-kb 39 clone. Aproduct of 1530 bp was obtained and cloned into the pCR 2.1-TOPO vector. The 59 clone revealed a 48-bp untranslated regionand an open reading frame of 1479 bp. The cDNA clones weresequenced in both directions using vector-derived and insert-specific primers. The nucleotide and amino acid sequence datawere assembled. The PRINTS (Attwood et al., 1998) databasewas accessed at http://www.biochem.ucl.ac.uk/bsm/dbbrowser/PRINTS/PRINTS.html and the BLOCKS (Henikoff and Henikoff,1994) analysis was carried out on the BLOCKS server at http://www.blocks.fhcrc.org/. Psi-BLAST (Altschul et al., 1997) was car-ried out at the BLAST server http://www/ncbi.nlm.nih.gov/cgi-bin/BLAST/nph-psi_blast. FASTA (Pearson and Lipman, 1988) wascarried out on a local server. Determination of potential phosphor-ylation and EF hand motifs were analyzed using 70% similarityat the http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page5npsa_prosite.html web site.
Cloning of alternatively spliced forms of the transcript wasperformed by probing a 59-Stretch lDR2 human testis cDNAlibrary (Clontech) according to the manufacturer’s instructionswith the full-length 32P-labeled cDNA obtained through the RACEprotocol. Purified tertiary plaques were converted to their plasmidforms, plated, and grown in LB broth, and the plasmid DNA was
isolated by Qiagen Mini-Kit columns (Qiagen, Valencia, CA) beforesequencing with both plasmid and gene-specific primers.). All rights reserved.

239CABYR, a Novel Fibrous Sheath Protein
Northern and Dot-Blot Analyses
A Northern blot containing 2 mg of poly(A)1 RNA from eightselected human tissues and a normalized RNA dot-blot containing89–514 ng of mRNA from 50 different human tissues were ob-tained from Clontech. The Northern blot was probed with a32P-labeled 1479-bp DNA corresponding to bps 49–1527 of CR-A.Probes were prepared by random oligonucleotide primer labeling(Feinberg and Vogelstein, 1983). Hybridization was performed inExpressHyb solution (Clontech) at 68°C for 1 h, followed by threewashes in 23 SSC, 0.05% SDS at room temperature, and twowashes in 0.13 SSC, 0.1% SDS for 20 min at 50°C. The blot wasexposed to X-ray film at 270°C for 60 h with two intensifyingscreens. The dot-blot was probed with the same 32P-labeled cDNAcorresponding to coding region A. The blot was hybridized inExpressHyb solution (Clontech) containing salmon sperm DNAand human placental Cot-1 DNA overnight at 65°C. The blot wasthen washed three times in 23 SSC, 1% SDS at 65°C, followed bytwo additional washes in 0.13 SSC, 0.5% SDS at 55°C, and finallywashed twice at 65°C in this solution for 20 min before exposingthe filter to X-ray film for 24, 48, and 72 h at 270°C with twointensifying screens.
Expression and Purification of the RecombinantProtein and Antibody Production
The cDNA encoding CR-A of CABYR was amplified by poly-merase chain reaction from human testicular Marathon-readycDNA (Clontech). Primers were designed to create a NcoI site atthe 59 end and a NotI site at the 39 end of the polymerase chainreaction product. The amplified cDNA was cloned into the NcoI–NotI sites of the pET28b expression vector (Novagen, Madison, WI)and Escherichia coli strain BL21(DE3) was transformed with theplasmid construct. The resulting construct appended six residues ofhistidine on the C-terminus of the protein. The expression plasmidconstruct was sequenced at the 59 and 39 ends to verify the readingframe of the construct.
A single positive colony was used to inoculate 10 L of LB brothwith 30 mg/ml kanamycin and grown at 37°C until the A600
reached 0.6. The recombinant protein expression was induced for3 h with 1.0 mM IPTG (isopropyl-1-thio-b-D-galactopyranoside).The cells were then transferred to 13 binding buffer (20 mMTris–HCl, pH 7.9, 0.5 M NaCl, 5 mM immidazole) containing 0.1%NP-40 (Sigma, St. Louis, MO) and 0.1 mg/ml lysozyme on ice for 30min, and sonicated briefly. The insoluble pellet resulting fromcentrifugation at 15,000g for 15 min was dissolved in 6 M urea in13 binding buffer for 1 h on ice. After recentrifugation at 15,000gfor 15 min the urea soluble fraction was loaded onto a Ni21-activated His-binding resin column (Novagen) following the manu-facturer’s protocol, and the recombinant protein was eluted with300 mM imidazole in 13 binding buffer containing 6 M urea. Theaffinity-purified recombinant protein was used for immunizationof female Lewis rats (200 mg/rat) in Freund’s complete adjuvant.Animals were boosted twice at an interval of 14 days with 200 mgof recombinant protein in incomplete Freund’s adjuvant and serumwas collected 7 days after each boost.
Immunoblotting
Western blotting was performed as described previously (Mandalet al., 1999), employing a 1:3500 dilution of the rabbit antiserum
raised against gel-purified CABYR antigen and a 1:2500 dilution of© 2002 Elsevier Science (USA
the rat antiserum to rCABYR. Sperm proteins phosphorylated ontyrosine residues were identified by immunoblotting with horse-radish peroxidase-conjugated anti-phosphotyrosine monoclonal an-tibody RC-20 (Transduction Laboratories, Lexington, KY) at a1:2500 dilution in 10 mM Tris (pH 7.5), 0.1 M NaCl, and 0.05%Tween 20 for 20 min at 37°C (Ruff-Jamisson et al., 1993).
Localization of CABYR in the SeminiferousEpithelium of Human Testis
Testes were obtained from three patients undergoing electiveorchiectomies. Testes were sliced once with a razor blade andimmersed in neutral buffered formalin (4%) solution (Sigma) for1 h. The tissue was then minced and placed into fresh fixativeovernight. The tissue was dehydrated in a graded series of ethanols,cleared in xylene, and embedded in paraffin. Sections (2.5 mm thick)were cut, mounted onto slides, deparaffinized, rehydrated, andpermeabilized with 100% methanol. Sections were incubated inblocking solution containing 10% normal goat serum (NGS) inPBS, incubated with anti-rCABYR antiserum or preimmune serum(1:200) in PBS containing 1% NGS (PBS–NGS), washed, incubatedwith FITC-labeled goat anti-rat IgG (1:400; Jackson Immuno-Research, West Grove, PA) in PBS–NGS, washed, and mountedwith Slow Fade (Molecular Probes, Eugene, OR) containing DAPI IIcounterstain (Vysis, Downers Grove, IL). Sections were observed byepifluorescence microscopy using a Zeiss microscope. Individualblue and green fluorescent images were obtained using a digitalcamera (Hamamatsu) and compiled using Openlab software (Im-provision, Inc., Boston, MA).
Indirect Immunofluorescence of Human Sperm
For immunofluorescence studies fresh human sperm were har-vested over a discontinuous 55%/80% Percoll gradient and subse-quently washed three times with Ham’s F-10 media. The spermwere counted using a hemocytometer and diluted to a concentra-tion of 1 3 106 sperm/ml. A 20 ml aliquot of the sperm suspensionwas added per well (2 3 105 sperm) onto poly-L-lysine-coated slides.The slides were dried at 40°C and then methanol fixed for 10 min.In some experiments no fixation was performed and the spermwere simply air dried onto the slide. After washing three times for5 min each in PBS, the slides were frozen at 270°C for 1 week. Allsubsequent incubations were done in a humid chamber. Thepreparations were blocked in 10% NGS in PBS with 0.05% Tween20 (PBS-tw) for 30 min. The primary antiserum, either rabbitanti-CABYR antiserum or rat anti-recombinant CABYR and theirpreimmune controls, was diluted 400-fold with 10% NGS inPBS-tw and incubated with the specimen overnight at 4°C. Theslides were then washed three times for 5 min each in PBS-tw, andthe secondary antibody, goat anti-rabbit IgG FITC conjugated(Jackson ImmunoResearch) or goat anti-rat IgG FITC conjugated(Jackson ImmunoResearch), were applied at 1:200 dilutions in 10%NGS in PBS-tw for 1 h at 37°C. The slides were washed three times for5 min each in PBS-tw, and Slow Fade-Light Antifade Kit (MolecularProbes) was used to reduce the fading rate of the fluorescein.
Electron Microscopic Localization
Sperm from four donors were pooled and washed twice bycentrifugation at 550g in wash buffer [Ham’s F10 Nutrient Mixture(Gibco/BRL) with 3% sucrose]. The washed sperm were resus-
pended in fixative consisting of 4% paraformaldehyde and 0.2%). All rights reserved.
240 Naaby-Hansen et al.
glutaraldehyde in wash buffer for 15 min at room temperature.After removing fixative by centrifugation and washing three timeswith wash buffer, the sperm were dehydrated through a gradedseries of ethanols, from 40% to 100%. The cells were infiltratedwith and embedded in Lowicryl K4M (EM Science, Fort Washing-ton, PA) according to the manufacturer’s recommendations. Theblocks were polymerized with UV light for 72 h at 220°C andultrathin sections of 100 nm thickness were cut.
Nonspecific sperm–antibody interactions were blocked by incu-bating the sections in undiluted normal goat serum for 15 min atroom temperature and washing once with wash buffer. Rat anti-serum to rCABYR and preimmune serum were diluted 1:50 in washbuffer with 1% normal goat serum, 1% bovine serum albumin, and0.05% Tween 20. Lowicryl sections were incubated with dilutedanti-rCABYR or wash buffer alone at 4°C for 16 h. After washingfour times in wash buffer, they were incubated for 1.5 h at roomtemperature with 5 nm gold-conjugated secondary antibody, goatanti-rat IgG (Goldmark Biologicals, Phillipsburg, NJ) diluted 1:35 inwash buffer. The sections were washed with distilled water andstained with uranyl acetate before examination with a JEOL 100CXelectron microscope (JEOL, Peabody, MA).
RESULTS
Identification and Characterization of Calcium-Binding Proteins (CBPs) in Human Spermatozoa
The 45Ca overlay technique of Maruyama et al. (1984) was
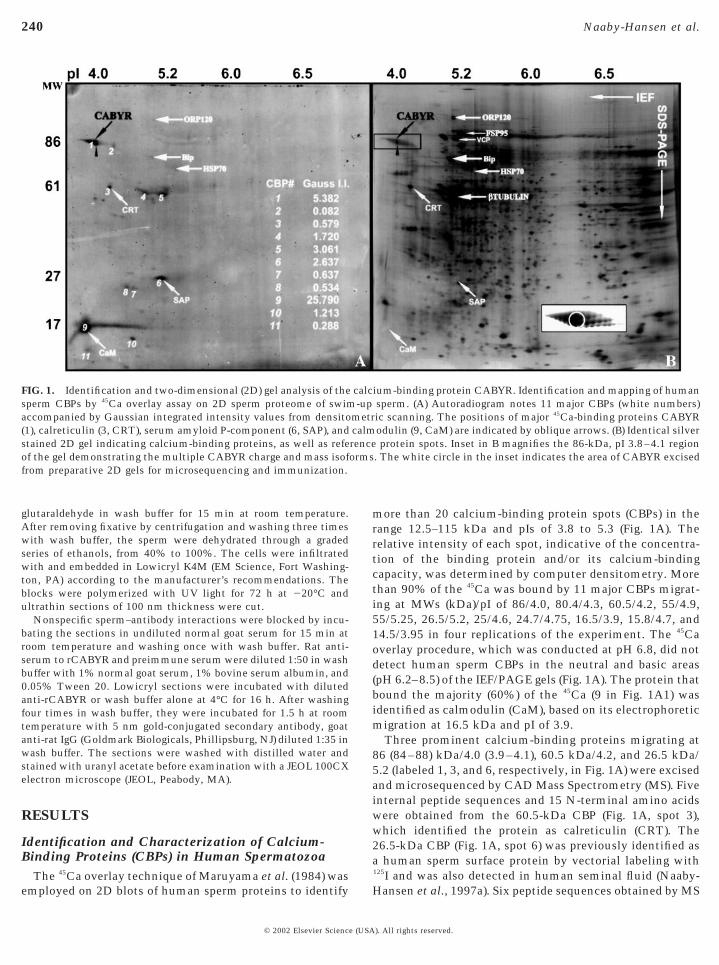
FIG. 1. Identification and two-dimensional (2D) gel analysis of thesperm CBPs by 45Ca overlay assay on 2D sperm proteome of swimaccompanied by Gaussian integrated intensity values from densito(1), calreticulin (3, CRT), serum amyloid P-component (6, SAP), andstained 2D gel indicating calcium-binding proteins, as well as refeof the gel demonstrating the multiple CABYR charge and mass isoffrom preparative 2D gels for microsequencing and immunization.
employed on 2D blots of human sperm proteins to identify
© 2002 Elsevier Science (USA
more than 20 calcium-binding protein spots (CBPs) in therange 12.5–115 kDa and pIs of 3.8 to 5.3 (Fig. 1A). Therelative intensity of each spot, indicative of the concentra-tion of the binding protein and/or its calcium-bindingcapacity, was determined by computer densitometry. Morethan 90% of the 45Ca was bound by 11 major CBPs migrat-ing at MWs (kDa)/pI of 86/4.0, 80.4/4.3, 60.5/4.2, 55/4.9,55/5.25, 26.5/5.2, 25/4.6, 24.7/4.75, 16.5/3.9, 15.8/4.7, and14.5/3.95 in four replications of the experiment. The 45Caoverlay procedure, which was conducted at pH 6.8, did notdetect human sperm CBPs in the neutral and basic areas(pH 6.2–8.5) of the IEF/PAGE gels (Fig. 1A). The protein thatbound the majority (60%) of the 45Ca (9 in Fig. 1A1) wasidentified as calmodulin (CaM), based on its electrophoreticmigration at 16.5 kDa and pI of 3.9.
Three prominent calcium-binding proteins migrating at86 (84–88) kDa/4.0 (3.9–4.1), 60.5 kDa/4.2, and 26.5 kDa/5.2 (labeled 1, 3, and 6, respectively, in Fig. 1A) were excisedand microsequenced by CAD Mass Spectrometry (MS). Fiveinternal peptide sequences and 15 N-terminal amino acidswere obtained from the 60.5-kDa CBP (Fig. 1A, spot 3),which identified the protein as calreticulin (CRT). The26.5-kDa CBP (Fig. 1A, spot 6) was previously identified asa human sperm surface protein by vectorial labeling with125I and was also detected in human seminal fluid (Naaby-
ium-binding protein CABYR. Identification and mapping of humansperm. (A) Autoradiogram notes 11 major CBPs (white numbers)ic scanning. The positions of major 45Ca-binding proteins CABYRodulin (9, CaM) are indicated by oblique arrows. (B) Identical silverprotein spots. Inset in B magnifies the 86-kDa, pI 3.8–4.1 region
. The white circle in the inset indicates the area of CABYR excised
calc-up
metrcalmrenceorms
Hansen et al., 1997a). Six peptide sequences obtained by MS
). All rights reserved.

241CABYR, a Novel Fibrous Sheath Protein
and 22 N-terminal amino acids obtained by Edman degra-dation identified the 26.5-kDa CBP as serum amyloidP-component precursor (SAP). Calcium binding to CaM andSAP resides within EF hand motifs, while CRT’s calciumbinding occurs in repeated, polyacidic C-terminal domains.The ability of these proteins to bind 45Ca validated thesensitivity and specificity of the 45Ca overlay procedure on2D gels.
The train of silver-stained protein spots at ; 86 kDa, pI4.0 (box in Fig. 1B) was the second most intense stainingregion on the autoradiogram after calmodulin (Fig. 1A).Silver staining (Fig. 1B) showed several isoforms, varyingslightly in mass and charge (insert in Fig. 1B). It is thiscluster of proteins to which we have given the nameCABYR, for calcium-binding tyrosine phosphorylation-regulated protein. The acidic isoforms of CABYR boundmore calcium than that bound by the more basic isoforms,even though the two differentially charged groups ofCABYR showed similar staining with silver nitrate (blackreference arrowheads in Figs. 1A and 1B). The center of thispeptide cluster was cored for microsequencing and fivepeptides obtained after tryptic digestion of the excised86-kDa spot did not match any known peptide sequences inany protein or gene database. These peptides obtained bymicrosequencing are boxed in Fig. 3. Later, the acidic regionof 86-kDa form was also microsequenced, producing ninepeptides (including six new peptides), all of which exactlymatched regions within the protein deduced from codingregion A (CR-A; new peptides not marked in Fig. 3).
CABYR Undergoes Tyrosine Phosphorylationduring in Vitro Capacitation
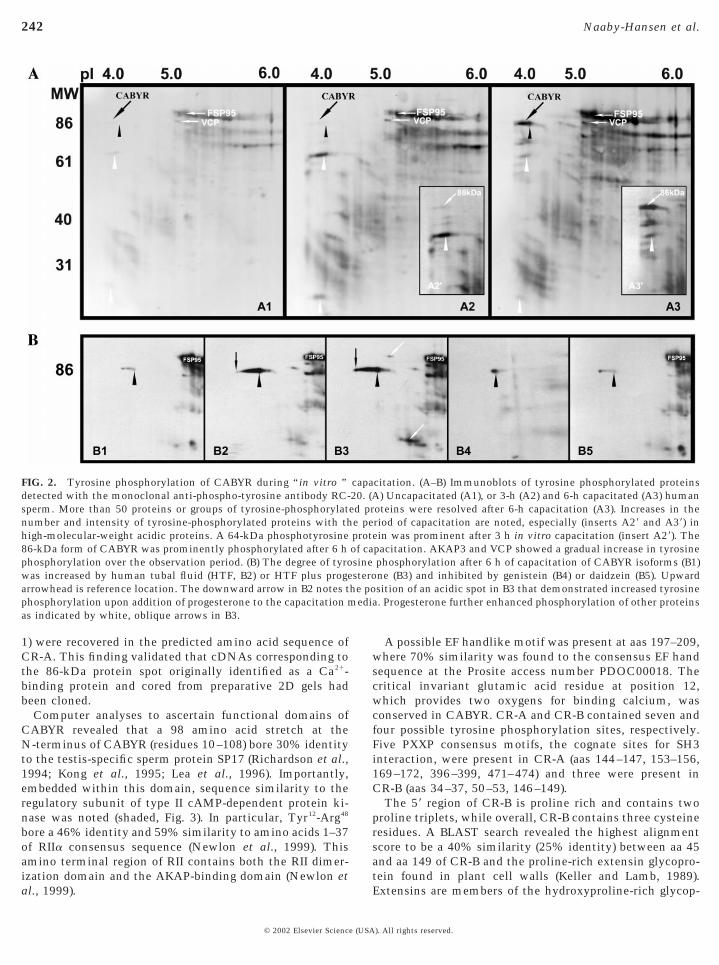
Proteins phosphorylated on tyrosine residues during invitro capacitation in albumin-containing capacitation me-dia were identified on 2D immunoblots of freshly ejacu-lated sperm or from sperm capacitated for 3 or 6 h bystaining with the monoclonal anti-phosphotyrosine anti-body RC-20. After 3 and 6 h of in vitro capacitation asignificant increase was observed in tyrosine phosphoryla-tion of several sperm proteins, including AKAP3 (fibroussheath protein 95) and the 86-kDa forms of CABYR (Fig.2A3) when compared to the uncapacitated samples. Follow-ing 3 h of capacitation the major acidic (pI ; 4.0) tyrosinephosphorylated component was a 64-kDa protein (upwardwhite arrowhead in A2 and A29). However, after 6 h ofcapacitation the intensity of the 64-kDa protein had dimin-ished and the dominant tyrosine phosphoprotein in theacidic region of the gel was the 86-kDa form of CABYR(compare A29 with A39).
Tyrosine phosphorylation of the 86-kDa form of CABYRvaried with the composition of the capacitation medium(Fig. 2B). Tyrosine phosphorylation of the 86-kDa form ofCABYR in human tubal fluid plus albumin (Fig. 2B2) washigher than that observed in Dulbecco’s PBS (Fig. 2B1).Interestingly, addition of 100 mM progesterone to the HTF
1 albumin-containing capacitation media further enhanced© 2002 Elsevier Science (USA
the phosphorylation of the most acidic CABYR isoforms(Fig. 2B3). Capacitation-induced tyrosine phosphorylationof the 86-kDa CABYR isoforms was inhibited in aconcentration-dependent manner by treatment with thetyrosine kinase inhibitor, genistein (Fig. 2B4), while similarconcentrations of the analogue, daidzein (Akiyama et al.,1987) had an inhibitory effect on phosphorylation ofCABYR but not FSP95 (AKAP3; Fig. 2B5).
Cloning of CABYR cDNAsA degenerate inosine-containing forward primer, de-
signed from the peptide GQPEVPAQL (Fig. 3), was em-ployed to amplify a 1.0-kb region of cDNA by 39-RACE PCRfrom human testicular Marathon-ready cDNA (Clontech, A1530-bp 59-cDNA fragment, including a 48-bp untranslatedregion, was similarly amplified and cloned using standard59-RACE PCR with an antisense 39 reverse primer generatedto a sequence 263 bp downstream from the 59 end of the1.0-kb 39 clone. A nucleotide sequence for a composite2228-bp CABYR cDNA (Fig. 3) was obtained by sequencingthe two PCR fragments in both directions. This 2228-bpcDNA (Fig. 3) was the longest CABYR cDNA obtained andwas submitted to the GenBank database under accessionnumber AF088868. This cDNA was 32P-labeled and em-ployed to screen a human testicular lDR2 59-Stretch cDNAlibrary (Clontech). Phage isolates were digested with re-striction endonuclease, grouped according to restrictionfragment sizes, and sequenced to yield several cDNAs (also of2228 bp) as well as five alternative splice variants, which weresubmitted to GenBank under accession numbers AF295037,AF295038, AF295039, AF329634, and AY007205 (Fig. 4).
Analysis of these cDNAs led to the conclusion that alter-native splicing occurs and the CABYR sequence may bediscussed in terms of two coding regions, CR-A and CR-B.CR-A began at bp 49 and ended at bp 1527 (codons 1–494)with a stop codon TAA at bps 1528–30, yielding a predictedprotein of 493 amino acids with a mass of 52.8 kDa and pI of4.5. Fifteen in-frame nucleotides (1531–1545) then separateCR-A from the splice junction that initiates CR-B. CR-B(nucleotides 1546–2145) encodes a peptide that entirely or inpart serves as the carboxy terminus on three CABYR variants(AF295037, AF295038, and AF295039). Splice variants weresequenced containing alternative start codons at bps 49–51(AF088868, AF295037, AF295039, AF329634), bps 343–345(AF295038), or bps 652–654 (AY007205). These splice variantscontained deletions of a small N-terminal region of CR-A(AF329634, AF295038), a major portion of CR-A (AF295037,AF295038), and a large domain spanning CR-A and B(AF295039). These cDNAs predict CABYR proteins of 24, 30,31, 41, 51, and 53 kDa (Fig. 4).
Motif Analysis of Splice Variants Revealed RIIDimerization, Potential EF Hand, Pro-X-X-Pro(PXXP), and Extensin Domains
All five of the tryptic peptides microsequenced by MS
from the original 86-kDa spot excised from the 2D gel (Fig.). All rights reserved.
he poedi
242 Naaby-Hansen et al.
1) were recovered in the predicted amino acid sequence ofCR-A. This finding validated that cDNAs corresponding tothe 86-kDa protein spot originally identified as a Ca21-binding protein and cored from preparative 2D gels hadbeen cloned.
Computer analyses to ascertain functional domains ofCABYR revealed that a 98 amino acid stretch at theN-terminus of CABYR (residues 10–108) bore 30% identityto the testis-specific sperm protein SP17 (Richardson et al.,1994; Kong et al., 1995; Lea et al., 1996). Importantly,embedded within this domain, sequence similarity to theregulatory subunit of type II cAMP-dependent protein ki-nase was noted (shaded, Fig. 3). In particular, Tyr12-Arg48
bore a 46% identity and 59% similarity to amino acids 1–37of RIIa consensus sequence (Newlon et al., 1999). Thisamino terminal region of RII contains both the RII dimer-ization domain and the AKAP-binding domain (Newlon et
FIG. 2. Tyrosine phosphorylation of CABYR during “in vitro ”detected with the monoclonal anti-phospho-tyrosine antibody RC-sperm. More than 50 proteins or groups of tyrosine-phosphorylatnumber and intensity of tyrosine-phosphorylated proteins with thhigh-molecular-weight acidic proteins. A 64-kDa phosphotyrosine86-kDa form of CABYR was prominently phosphorylated after 6 hphosphorylation over the observation period. (B) The degree of tyrowas increased by human tubal fluid (HTF, B2) or HTF plus progearrowhead is reference location. The downward arrow in B2 notes tphosphorylation upon addition of progesterone to the capacitation mas indicated by white, oblique arrows in B3.
al., 1999).
© 2002 Elsevier Science (USA
A possible EF handlike motif was present at aas 197–209,where 70% similarity was found to the consensus EF handsequence at the Prosite access number PDOC00018. Thecritical invariant glutamic acid residue at position 12,which provides two oxygens for binding calcium, wasconserved in CABYR. CR-A and CR-B contained seven andfour possible tyrosine phosphorylation sites, respectively.Five PXXP consensus motifs, the cognate sites for SH3interaction, were present in CR-A (aas 144–147, 153–156,169–172, 396–399, 471–474) and three were present inCR-B (aas 34–37, 50–53, 146–149).
The 59 region of CR-B is proline rich and contains twoproline triplets, while overall, CR-B contains three cysteineresidues. A BLAST search revealed the highest alignmentscore to be a 40% similarity (25% identity) between aa 45and aa 149 of CR-B and the proline-rich extensin glycopro-tein found in plant cell walls (Keller and Lamb, 1989).
citation. (A–B) Immunoblots of tyrosine phosphorylated proteins) Uncapacitated (A1), or 3-h (A2) and 6-h capacitated (A3) human
oteins were resolved after 6-h capacitation (A3). Increases in theriod of capacitation are noted, especially (inserts A29 and A39) inein was prominent after 3 h in vitro capacitation (insert A29). Thepacitation. AKAP3 and VCP showed a gradual increase in tyrosinephosphorylation after 6 h of capacitation of CABYR isoforms (B1)ne (B3) and inhibited by genistein (B4) or daidzein (B5). Upwardsition of an acidic spot in B3 that demonstrated increased tyrosine
a. Progesterone further enhanced phosphorylation of other proteins
capa20. (Aed pre peprotof casinestero
Extensins are members of the hydroxyproline-rich glycop-
). All rights reserved.

calcium-binding EF handlike domain is located at aas 197–209 inCR-A. PXXP motifs are bold and underlined. The locations of the
243CABYR, a Novel Fibrous Sheath Protein
© 2002 Elsevier Science (USA
rotein family (HRGPs) and contain a characteristic pen-tapeptide repeat Ser-Pro4 (Chen and Varner, 1985), which inCR-B may be represented by a modified Ser-Pro3 domain ataas 32–35. Interestingly, a similar Ser-Pro3 motif is presentin CR-A at position 155–158.
Genomic Locus of CABYR
A search of the human genome databases located theCABYR gene on chromosome 18 at 18q12.2 (humangenomic contig sequence accession number NT_011044.5).There are five CABYR exons spanning 18.5 kb of genomicDNA.
CABYR Transcripts Are Testis Specific
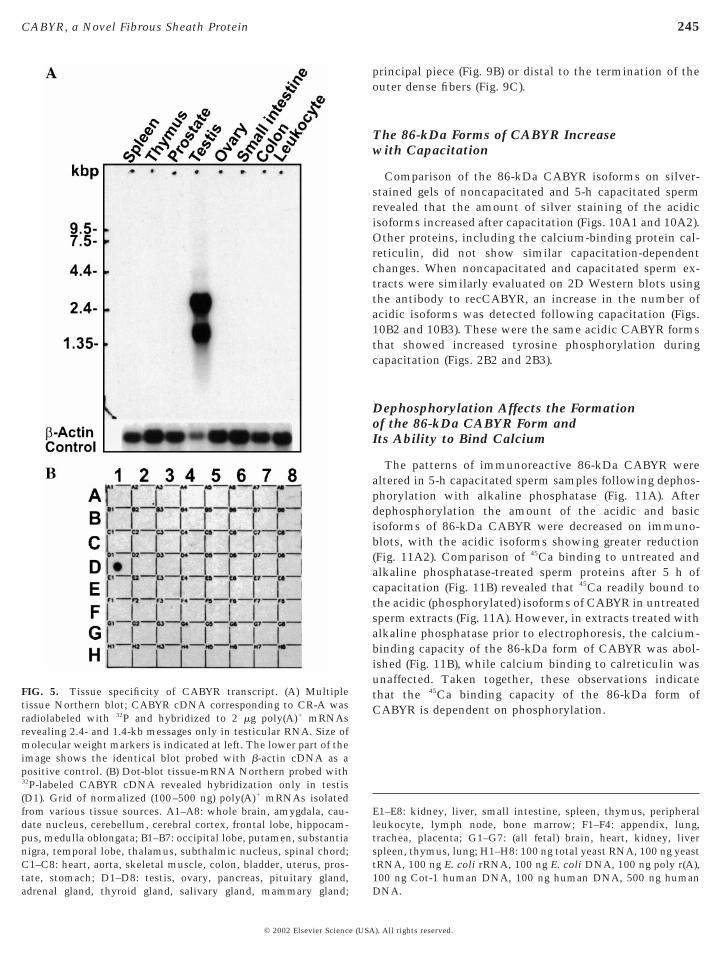
A 32P-labeled cDNA probe corresponding to CR-A wasemployed for Northern analysis of mRNA from severaltissues (Fig. 5A) and a dot-blot (Fig. 5B) containing mRNAsfrom 50 distinct human tissues (Clontech). Interestingly,two broad bands of approximately 2.4 and 1.4 kb were notedin the testicular mRNA (Fig. 5A), indicating that severalCABYR messages of different sizes were expressed in thehuman testis, a finding in concert with the cloning andsequencing of six cDNAs, including five splice variantsnoted above. The broad 2.4-kb transcript (Fig. 5A) detectedin pooled human testicular mRNAs may be accounted forby the largest cDNA (AF088868) and AF329634, whichdeletes a small N-terminal region of CR-A, while the 1.4-kbtranscript may be accounted for by the other splice variantswith their deletions. Importantly, CABYR transcripts wereexpressed in testis (Fig. 5A and Fig. 5B, spot D1) but not inother human tissues.
Western Analyses with Antiserum to RecombinantCABYR Indicate CABYR Protein Polymorphism
The cDNA corresponding to the entire CR-A was clonedinto the bacterial expression vector pET28b and introducedinto BL21(DE3) cells. The expressed recombinant protein(apparent Mr ; 78 kDa) was purified by immobilized metalaffinity chromatography using Ni21-Sepharose column (Fig.6A). Antiserum against purified rCABYR was subsequentlyraised in female rats and demonstrated to recognize therecombinant immunogen on Western blot (Fig. 6B). Whenreacted with human sperm proteins (Fig. 6C), this mono-specific rat antiserum to rCABYR recognized multipleprotein spots on 2D Western blots. This 2D Western blotwas repeated using a rabbit antisera to gel-purified CABYR
five embedded microsequences obtained by mass spectrometryfrom the original 2D isolated spot are indicated in boxes. Potentialtyrosine phosphorylation sites are encircled. Bold arrows indicate
FIG. 3. Nucleotide and amino acid sequences of CABYR. Upperline indicates cDNA sequence (numbered at left) with codingregions in uppercase and UTRs in italics. Starting codon is bold andtermination codons for each coding region are starred. The pre-dicted polyadenylation signal is underlined. Lower line presentsamino acid sequences with coding region A from 1 to 493 andcoding region B from 1 to 199, numbered at right. The regionhomologous to the amino terminus of the RII subunit of PK-A isshaded, beginning at aa 12 in CR-A, while the region homologousto extensins is underlined, beginning at aa 45 in CR-B. A potential
the positions of spliced junctions.
). All rights reserved.

244 Naaby-Hansen et al.
and the results were identical (data not shown). Immuno-reactive species migrated in five major groups based on size:(1) 27–38 kDa; (2) 38–42 kDa; (3) 50–56 kDa; (4) 63–72 kDa;and (5) 81–87 kDa. The finding of similar patterns ofCABYR isoforms on 2D gels probed with antisera to boththe gel-purified and recombinant CABYR confirmed thatalternative splice variants identified as cDNAs during clon-ing were expressed at the protein level, resulting in consid-erable CABYR heterogeneity.
Localization of CABYR in the SeminiferousEpithelium
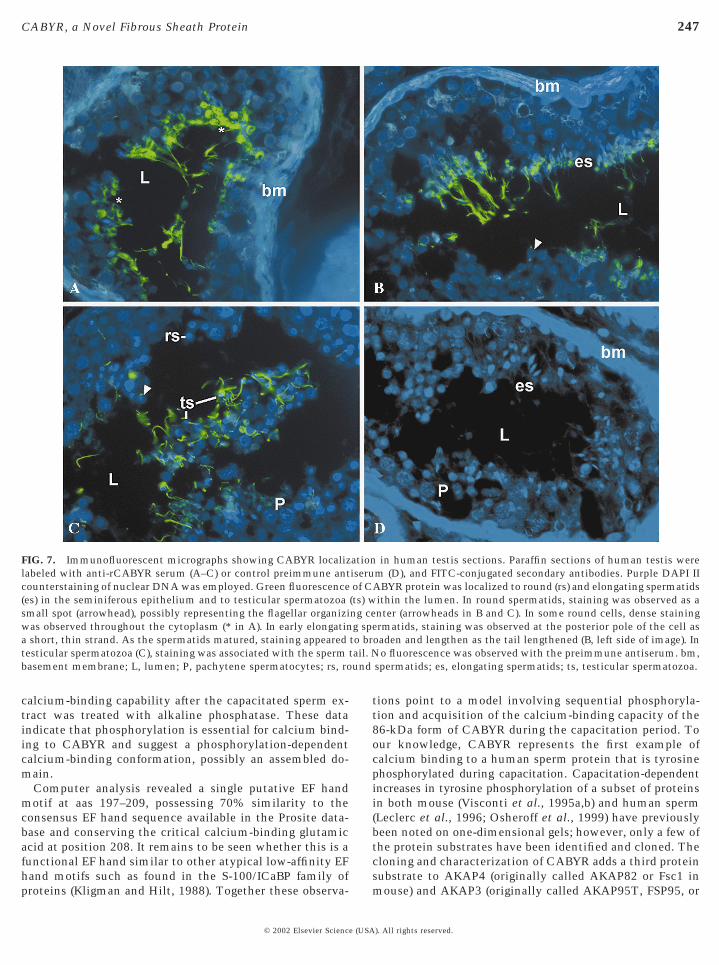
Immunofluorescent localization of CABYR in the humantestes using the antibody to recombinant CABYR showedstaining of round and elongating spermatids in the seminif-erous epithelium and testicular spermatozoa within thelumen of the tubules, indicative of a postmeiotic pattern ofexpression of the CABYR gene. The staining patterns sug-gested a gradual migration of the CABYR protein from adiffuse cytoplasmic localization in round spermatids to theposterior pole of early spermatids and then to the flagellumas the tail formed (Fig. 7). The preimmune serum from theidentical animal did not stain the testis. Testes from three
FIG. 4. Diagrams of homologous motifs and alternative splicedmotifs relative to the two coding regions (shaded) that are comparindicated with nucleotide numbers as well as amino acid sequencepredicted mass, and pI are listed. Splice variant V lacks the RII dimdomain and forms I and II lack the extensin motif.
patients showed identical localization patterns.
© 2002 Elsevier Science (USA
Localization of CABYR to the Principal Pieceof the Mature Human Sperm Flagellumby Immunofluorescence andImmunoelectron Microscopy
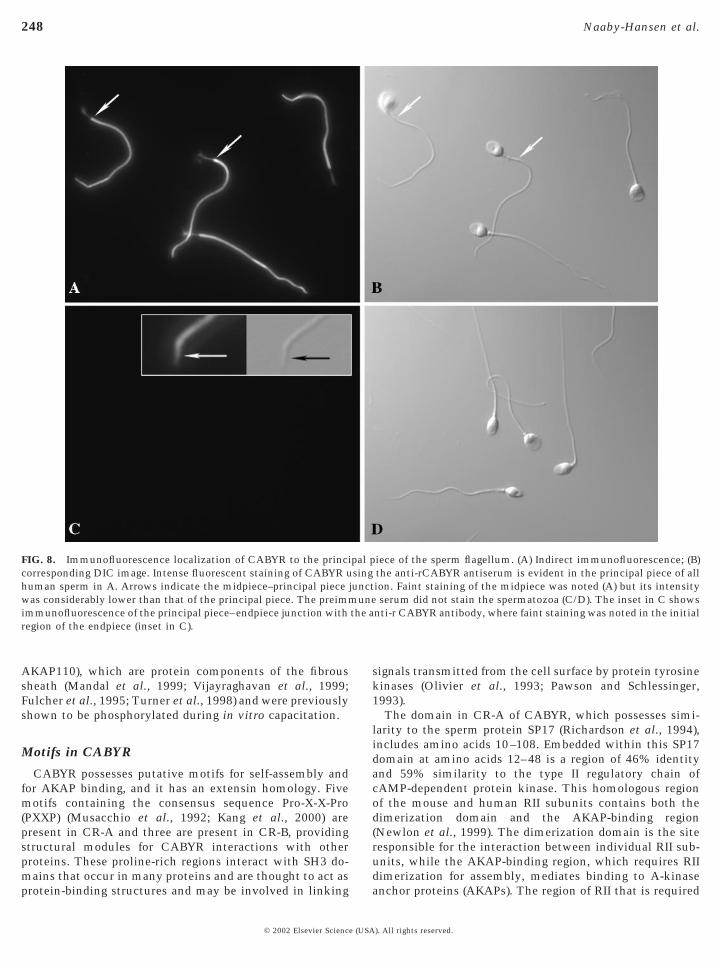
Antibodies raised against rCABYR recognized the entirelength of the principal piece of ejaculated methanol-fixedspermatozoa with an intense signal by indirect immunoflu-orescence microscopy, while both the midpiece (Fig. 8A1)and the endpiece (Fig. 8C, insert) exhibited much fainterstaining patterns (Fig. 8A). No CABYR immunofluores-cence was noted in the human sperm head in these nonca-pacitated sperm. Importantly, no immunofluorescencestaining was observed on live motile sperm, indicating thatCABYR epitopes were not accessible on the plasma mem-brane. A similar staining pattern was achieved with theantiserum raised against gel excised CABYR (data notshown).
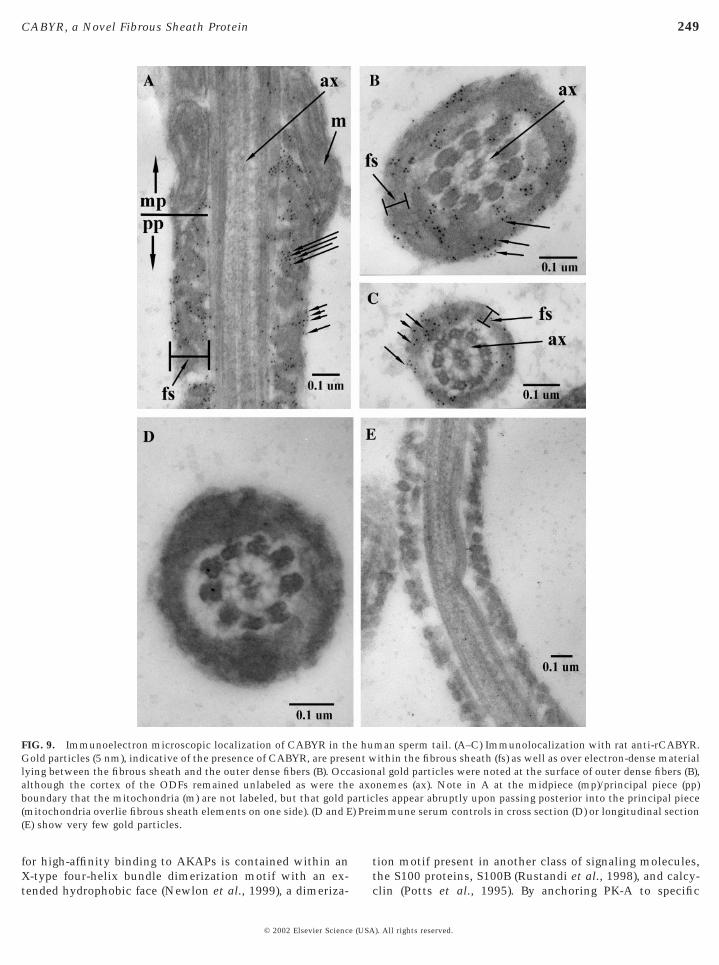
When the distribution of CABYR in freshly ejaculatedhuman sperm was examined by electron microscopic im-munocytochemical staining, gold particles were distributedover the fibrous sheath compartment, including the surfaceof the longitudinal columns and ribs. Smaller numbers ofgold particles were present in the periaxonemal space (Figs.9A–9C). CABYR was not detected in the annular ring ormitochondrial sheath (Fig. 9A). There was no evidence for
nts of CABYR. Top diagram summarizes the location of CABYRthe six splice variants of CABYR below. The splice junctions arenBank accession numbers for each variant as well as amino acids,ation motif, while forms III, V, and VI lack the potential EF-hand
variaed tos. Geeriz
CABYR localization within the axoneme in either the
). All rights reserved.
adrenal gland, thyroid gland, salivary gland, mammary gland;
245CABYR, a Novel Fibrous Sheath Protein
© 2002 Elsevier Science (USA
principal piece (Fig. 9B) or distal to the termination of theouter dense fibers (Fig. 9C).
The 86-kDa Forms of CABYR Increasewith Capacitation
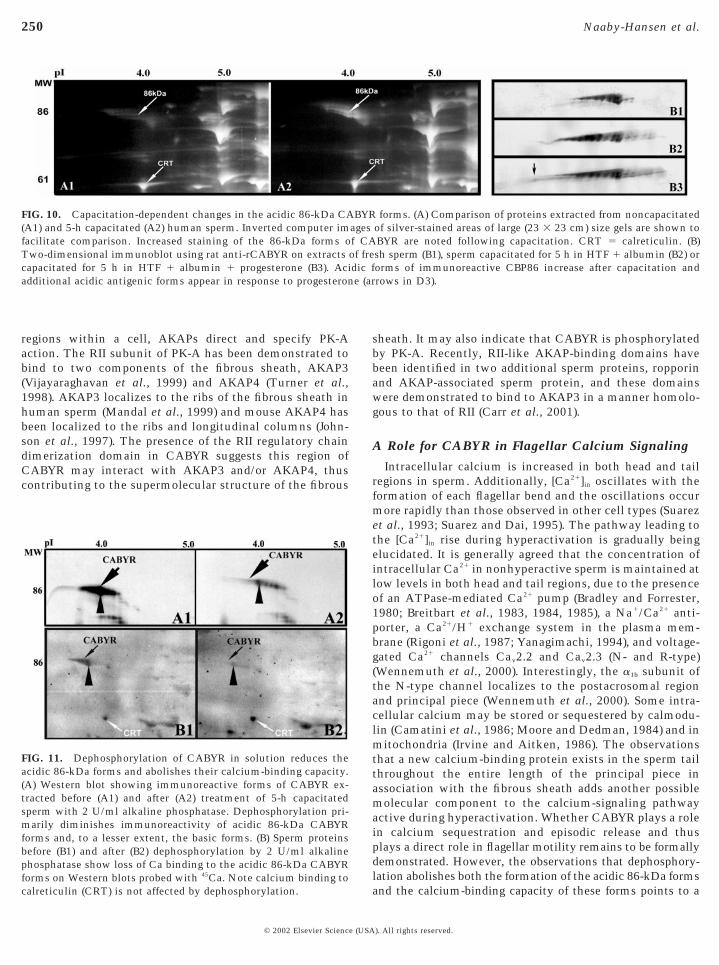
Comparison of the 86-kDa CABYR isoforms on silver-stained gels of noncapacitated and 5-h capacitated spermrevealed that the amount of silver staining of the acidicisoforms increased after capacitation (Figs. 10A1 and 10A2).Other proteins, including the calcium-binding protein cal-reticulin, did not show similar capacitation-dependentchanges. When noncapacitated and capacitated sperm ex-tracts were similarly evaluated on 2D Western blots usingthe antibody to recCABYR, an increase in the number ofacidic isoforms was detected following capacitation (Figs.10B2 and 10B3). These were the same acidic CABYR formsthat showed increased tyrosine phosphorylation duringcapacitation (Figs. 2B2 and 2B3).
Dephosphorylation Affects the Formationof the 86-kDa CABYR Form andIts Ability to Bind Calcium
The patterns of immunoreactive 86-kDa CABYR werealtered in 5-h capacitated sperm samples following dephos-phorylation with alkaline phosphatase (Fig. 11A). Afterdephosphorylation the amount of the acidic and basicisoforms of 86-kDa CABYR were decreased on immuno-blots, with the acidic isoforms showing greater reduction(Fig. 11A2). Comparison of 45Ca binding to untreated andalkaline phosphatase-treated sperm proteins after 5 h ofcapacitation (Fig. 11B) revealed that 45Ca readily bound tothe acidic (phosphorylated) isoforms of CABYR in untreatedsperm extracts (Fig. 11A). However, in extracts treated withalkaline phosphatase prior to electrophoresis, the calcium-binding capacity of the 86-kDa form of CABYR was abol-ished (Fig. 11B), while calcium binding to calreticulin wasunaffected. Taken together, these observations indicatethat the 45Ca binding capacity of the 86-kDa form ofCABYR is dependent on phosphorylation.
E1–E8: kidney, liver, small intestine, spleen, thymus, peripheralleukocyte, lymph node, bone marrow; F1–F4: appendix, lung,trachea, placenta; G1–G7: (all fetal) brain, heart, kidney, liverspleen, thymus, lung; H1–H8: 100 ng total yeast RNA, 100 ng yeasttRNA, 100 ng E. coli rRNA, 100 ng E. coli DNA, 100 ng poly r(A),100 ng Cot-1 human DNA, 100 ng human DNA, 500 ng human
FIG. 5. Tissue specificity of CABYR transcript. (A) Multipletissue Northern blot; CABYR cDNA corresponding to CR-A wasradiolabeled with 32P and hybridized to 2 mg poly(A)1 mRNAsrevealing 2.4- and 1.4-kb messages only in testicular RNA. Size ofmolecular weight markers is indicated at left. The lower part of theimage shows the identical blot probed with b-actin cDNA as apositive control. (B) Dot-blot tissue-mRNA Northern probed with32P-labeled CABYR cDNA revealed hybridization only in testis(D1). Grid of normalized (100–500 ng) poly(A)1 mRNAs isolatedfrom various tissue sources. A1–A8: whole brain, amygdala, cau-date nucleus, cerebellum, cerebral cortex, frontal lobe, hippocam-pus, medulla oblongata; B1–B7: occipital lobe, putamen, substantianigra, temporal lobe, thalamus, subthalmic nucleus, spinal chord;C1–C8: heart, aorta, skeletal muscle, colon, bladder, uterus, pros-tate, stomach; D1–D8: testis, ovary, pancreas, pituitary gland,
DNA.
). All rights reserved.

manisofo
246 Naaby-Hansen et al.
DISCUSSION
The Acidic 86-kDa Isoforms of CABYR Representa New Sperm-Specific Calcium-Binding Protein
When proteins from swim-up human sperm were reactedwith 45Ca on 2D gel autoradiograms, only calmodulin gavea stronger calcium signal than the 86-kDa phosphorylatedisoforms of CABYR. 45Ca bound to protein spots thatyielded mass spec microsequences for known calcium-binding proteins such as calreticulin and serum amyloidP-component precursor, verifying the sensitivity and speci-ficity of the calcium overlay procedure and suggesting thatthe robust calcium-binding capacity of CABYR was authen-tic. Although other immunoreactive forms of CABYR evi-dence a range of masses and charges on 2D immunoblots,the calcium-binding property appears to be unique to theacidic phosphorylated 86-kDa isoforms with pI of approxi-mately 3.8–4.0. The testis specificity of the CABYR mes-sages by Northern and dot-blot analyses coupled to theimmunolocalization of CABYR protein to spermatids in thetestis and to the flagella of ejaculated sperm indicate thatCABYR is a calcium-binding protein unique to the sper-matogenic lineage.
Five lines of evidence currently indicate that the 86-kDaprotein represents a monomeric form of CR-A that migrateshigher than its predicted size (53 kDa). (1) The 86-kDa spotwas microsequenced on two occasions, obtaining five andnine peptides, respectively, all of which matched exactlywith peptides contained only in CR-A. (2) Expression of the53-kDa CR-A in E. coli (Fig. 6A) showed migration at ;78
FIG. 6. Isolation of recCABYR and immunoreactivity of immuneaffinity-purified recombinant CABYR (Lane 3, 10 mg) expressed in B2) rat sera to recCABYR. (C) Two-dimensional Western blots of husera to CABYR, demonstrating the microheterogeneity of CABYR
kDa, indicating that a full-length CR-A recombinant pro-
© 2002 Elsevier Science (USA
tein shows higher apparent Mr, suggesting the higher Mr isintrinsic to this protein. (3) The authenticity of the stopcodon at the end of CR-A was confirmed by PCR-amplifiedfragments, multiple cDNA library clones (see Fig. 4), andalso by genomic sequence analysis. (4) No translationalreadthrough at this stop codon was observed either in E. colior in in vitro rabbit reticulocyte lysate systems, despiteexpression studies (data not shown). (5) Finally, the deducedpI of CR-A is 4.5. This pI is slightly higher than thatobserved in sperm (pI 4.0) but is close enough to suggest the86-kDa form may be accounted for by CR-A. The veryacidic nature of CR-A of CABYR may contribute to thehigher apparent Mr. Forms III and VI have deduced pIs of 8.7and 7.7, respectively.
Tyrosine Phosphorylation of the Acidic 86-kDaForms of CABYR Occurs during in VitroCapacitation
Increased tyrosine phosphorylation of acidic 86-kDa iso-forms of CABYR occurred during capacitation. After 6 h ofcapacitation the 86-kDa isoforms were prominently phos-phorylated. This was accompanied by an increase in thesilver staining of the acidic 86-kDa isoforms, suggestingeither the amount or argyrophilicity of 86-kDa proteinincreased during capacitation. The acidic isoforms ofCABYR (pI of 4.0) were also the forms that bound 45Ca, andcalcium binding to these isoforms was dramatically re-duced when the capacitated sperm sample was dephospho-rylated with alkaline phosphatase prior to electrophoreticseparation. It was notable that calreticulin served as a
. (A) Coomassie stain of uninduced (lane 1), induced (lane 2), andE3). (B) Western blot of preimmune (lane 1) and postimmune (lane
sperm extract reacted with preimmune (C1) and postimmune (C2)rms.
seraL21(D
positive control in this experiment since it did not lose
). All rights reserved.
to brtail. Nund
247CABYR, a Novel Fibrous Sheath Protein
calcium-binding capability after the capacitated sperm ex-tract was treated with alkaline phosphatase. These dataindicate that phosphorylation is essential for calcium bind-ing to CABYR and suggest a phosphorylation-dependentcalcium-binding conformation, possibly an assembled do-main.
Computer analysis revealed a single putative EF handmotif at aas 197–209, possessing 70% similarity to theconsensus EF hand sequence available in the Prosite data-base and conserving the critical calcium-binding glutamicacid at position 208. It remains to be seen whether this is afunctional EF hand similar to other atypical low-affinity EFhand motifs such as found in the S-100/ICaBP family of
FIG. 7. Immunofluorescent micrographs showing CABYR localizlabeled with anti-rCABYR serum (A–C) or control preimmune antcounterstaining of nuclear DNA was employed. Green fluorescence(es) in the seminiferous epithelium and to testicular spermatozoasmall spot (arrowhead), possibly representing the flagellar organiziwas observed throughout the cytoplasm (* in A). In early elongatina short, thin strand. As the spermatids matured, staining appearedtesticular spermatozoa (C), staining was associated with the spermbasement membrane; L, lumen; P, pachytene spermatocytes; rs, ro
proteins (Kligman and Hilt, 1988). Together these observa-
© 2002 Elsevier Science (USA
tions point to a model involving sequential phosphoryla-tion and acquisition of the calcium-binding capacity of the86-kDa form of CABYR during the capacitation period. Toour knowledge, CABYR represents the first example ofcalcium binding to a human sperm protein that is tyrosinephosphorylated during capacitation. Capacitation-dependentincreases in tyrosine phosphorylation of a subset of proteinsin both mouse (Visconti et al., 1995a,b) and human sperm(Leclerc et al., 1996; Osheroff et al., 1999) have previouslybeen noted on one-dimensional gels; however, only a few ofthe protein substrates have been identified and cloned. Thecloning and characterization of CABYR adds a third proteinsubstrate to AKAP4 (originally called AKAP82 or Fsc1 in
in human testis sections. Paraffin sections of human testis werem (D), and FITC-conjugated secondary antibodies. Purple DAPI IIABYR protein was localized to round (rs) and elongating spermatidsithin the lumen. In round spermatids, staining was observed as anter (arrowheads in B and C). In some round cells, dense stainingrmatids, staining was observed at the posterior pole of the cell as
oaden and lengthen as the tail lengthened (B, left side of image). Ino fluorescence was observed with the preimmune antiserum. bm,
spermatids; es, elongating spermatids; ts, testicular spermatozoa.
ationiseruof C(ts) wng ceg spe
mouse) and AKAP3 (originally called AKAP95T, FSP95, or
). All rights reserved.
unethe a
248 Naaby-Hansen et al.
AKAP110), which are protein components of the fibroussheath (Mandal et al., 1999; Vijayraghavan et al., 1999;Fulcher et al., 1995; Turner et al., 1998) and were previouslyshown to be phosphorylated during in vitro capacitation.
Motifs in CABYR
CABYR possesses putative motifs for self-assembly andfor AKAP binding, and it has an extensin homology. Fivemotifs containing the consensus sequence Pro-X-X-Pro(PXXP) (Musacchio et al., 1992; Kang et al., 2000) arepresent in CR-A and three are present in CR-B, providingstructural modules for CABYR interactions with otherproteins. These proline-rich regions interact with SH3 do-mains that occur in many proteins and are thought to act as
FIG. 8. Immunofluorescence localization of CABYR to the princicorresponding DIC image. Intense fluorescent staining of CABYR uhuman sperm in A. Arrows indicate the midpiece–principal piecewas considerably lower than that of the principal piece. The preimmimmunofluorescence of the principal piece–endpiece junction withregion of the endpiece (inset in C).
protein-binding structures and may be involved in linking
© 2002 Elsevier Science (USA
signals transmitted from the cell surface by protein tyrosinekinases (Olivier et al., 1993; Pawson and Schlessinger,1993).
The domain in CR-A of CABYR, which possesses simi-larity to the sperm protein SP17 (Richardson et al., 1994),includes amino acids 10–108. Embedded within this SP17domain at amino acids 12–48 is a region of 46% identityand 59% similarity to the type II regulatory chain ofcAMP-dependent protein kinase. This homologous regionof the mouse and human RII subunits contains both thedimerization domain and the AKAP-binding region(Newlon et al., 1999). The dimerization domain is the siteresponsible for the interaction between individual RII sub-units, while the AKAP-binding region, which requires RIIdimerization for assembly, mediates binding to A-kinase
iece of the sperm flagellum. (A) Indirect immunofluorescence; (B)the anti-rCABYR antiserum is evident in the principal piece of allion. Faint staining of the midpiece was noted (A) but its intensityserum did not stain the spermatozoa (C/D). The inset in C shows
nti-r CABYR antibody, where faint staining was noted in the initial
pal psing
junct
anchor proteins (AKAPs). The region of RII that is required
). All rights reserved.
artic) Pre
249CABYR, a Novel Fibrous Sheath Protein
for high-affinity binding to AKAPs is contained within anX-type four-helix bundle dimerization motif with an ex-
FIG. 9. Immunoelectron microscopic localization of CABYR in thGold particles (5 nm), indicative of the presence of CABYR, are preslying between the fibrous sheath and the outer dense fibers (B). Occalthough the cortex of the ODFs remained unlabeled as were thboundary that the mitochondria (m) are not labeled, but that gold p(mitochondria overlie fibrous sheath elements on one side). (D and E(E) show very few gold particles.
tended hydrophobic face (Newlon et al., 1999), a dimeriza-
© 2002 Elsevier Science (USA
tion motif present in another class of signaling molecules,the S100 proteins, S100B (Rustandi et al., 1998), and calcy-
man sperm tail. (A–C) Immunolocalization with rat anti-rCABYR.ithin the fibrous sheath (fs) as well as over electron-dense materialal gold particles were noted at the surface of outer dense fibers (B),nemes (ax). Note in A at the midpiece (mp)/principal piece (pp)les appear abruptly upon passing posterior into the principal pieceimmune serum controls in cross section (D) or longitudinal section
e huent wasione axo
clin (Potts et al., 1995). By anchoring PK-A to specific
). All rights reserved.
dic fe (ar
250 Naaby-Hansen et al.
regions within a cell, AKAPs direct and specify PK-Aaction. The RII subunit of PK-A has been demonstrated tobind to two components of the fibrous sheath, AKAP3(Vijayaraghavan et al., 1999) and AKAP4 (Turner et al.,1998). AKAP3 localizes to the ribs of the fibrous sheath inhuman sperm (Mandal et al., 1999) and mouse AKAP4 hasbeen localized to the ribs and longitudinal columns (John-son et al., 1997). The presence of the RII regulatory chaindimerization domain in CABYR suggests this region ofCABYR may interact with AKAP3 and/or AKAP4, thuscontributing to the supermolecular structure of the fibrous
FIG. 10. Capacitation-dependent changes in the acidic 86-kDa CA(A1) and 5-h capacitated (A2) human sperm. Inverted computer imfacilitate comparison. Increased staining of the 86-kDa forms oTwo-dimensional immunoblot using rat anti-rCABYR on extractscapacitated for 5 h in HTF 1 albumin 1 progesterone (B3). Aciadditional acidic antigenic forms appear in response to progesteron
FIG. 11. Dephosphorylation of CABYR in solution reduces theacidic 86-kDa forms and abolishes their calcium-binding capacity.(A) Western blot showing immunoreactive forms of CABYR ex-tracted before (A1) and after (A2) treatment of 5-h capacitatedsperm with 2 U/ml alkaline phosphatase. Dephosphorylation pri-marily diminishes immunoreactivity of acidic 86-kDa CABYRforms and, to a lesser extent, the basic forms. (B) Sperm proteinsbefore (B1) and after (B2) dephosphorylation by 2 U/ml alkalinephosphatase show loss of Ca binding to the acidic 86-kDa CABYRforms on Western blots probed with 45
Ca. Note calcium binding tocalreticulin (CRT) is not affected by dephosphorylation.© 2002 Elsevier Science (USA
sheath. It may also indicate that CABYR is phosphorylatedby PK-A. Recently, RII-like AKAP-binding domains havebeen identified in two additional sperm proteins, ropporinand AKAP-associated sperm protein, and these domainswere demonstrated to bind to AKAP3 in a manner homolo-gous to that of RII (Carr et al., 2001).
A Role for CABYR in Flagellar Calcium Signaling
Intracellular calcium is increased in both head and tailregions in sperm. Additionally, [Ca21]in oscillates with theformation of each flagellar bend and the oscillations occurmore rapidly than those observed in other cell types (Suarezet al., 1993; Suarez and Dai, 1995). The pathway leading tothe [Ca21]in rise during hyperactivation is gradually beingelucidated. It is generally agreed that the concentration ofintracellular Ca21 in nonhyperactive sperm is maintained atlow levels in both head and tail regions, due to the presenceof an ATPase-mediated Ca21 pump (Bradley and Forrester,1980; Breitbart et al., 1983, 1984, 1985), a Na1/Ca21 anti-porter, a Ca21/H1 exchange system in the plasma mem-brane (Rigoni et al., 1987; Yanagimachi, 1994), and voltage-gated Ca21 channels Cav2.2 and Cav2.3 (N- and R-type)(Wennemuth et al., 2000). Interestingly, the a1b subunit ofthe N-type channel localizes to the postacrosomal regionand principal piece (Wennemuth et al., 2000). Some intra-cellular calcium may be stored or sequestered by calmodu-lin (Camatini et al., 1986; Moore and Dedman, 1984) and inmitochondria (Irvine and Aitken, 1986). The observationsthat a new calcium-binding protein exists in the sperm tailthroughout the entire length of the principal piece inassociation with the fibrous sheath adds another possiblemolecular component to the calcium-signaling pathwayactive during hyperactivation. Whether CABYR plays a rolein calcium sequestration and episodic release and thusplays a direct role in flagellar motility remains to be formallydemonstrated. However, the observations that dephosphory-lation abolishes both the formation of the acidic 86-kDa forms
forms. (A) Comparison of proteins extracted from noncapacitatedof silver-stained areas of large (23 3 23 cm) size gels are shown toBYR are noted following capacitation. CRT 5 calreticulin. (B)sh sperm (B1), sperm capacitated for 5 h in HTF 1 albumin (B2) ororms of immunoreactive CBP86 increase after capacitation androws in D3).
BYRagesf CAof fre
and the calcium-binding capacity of these forms points to a
). All rights reserved.

251CABYR, a Novel Fibrous Sheath Protein
role for capacitation-dependent phosphorylation in calciumsignaling.
A Structural Model for the Fibrous SheathInvolving CABYR Interaction withTestis-Specific AKAPs
The cloning and localization of CABYR expands the bodyof information on the protein composition of the fibroussheath, a cytoskeletal structure that consists of two longi-tudinal columns connected by semicircular transverse ribsand is unique to the principal piece of the mammaliansperm flagellum. The longitudinal columns replace outerdense fibers 3 and 8 and appear to attach to two minuteridges that project radially from the wall of microtubuledoublets 3 and 8 (Fawcett, 1970). Ultrastructurally, thetransverse ribs are composed of a central medulla and a thinlimiting cortex (Fawcett, 1970, 1975). Initial interpretationsof the fibrous sheath proposed that the opposite locations ofthe longitudinal columns affected the axis of flagellar beat,although subsequent observations of flagellar motion sug-gest tail movements are not constrained to a single plane.The hypotheses that the ribs and intervening spaces conferflexibility during flagellar motion and that the sheathoverall serves as a protective girdle, providing mechanicalresistance during microtubule sliding, remain extant. Theextensin motif in CR-B may be particularly germane to thisstructural function.
Biochemically, the fibrous sheath contains a complex ofproteins. One-dimensional SDS–PAGE of isolated fibroussheaths of mice (Eddy et al., 1991), rabbits (Kim et al., 1997),rats (Olson et al., 1976; Oko and Clermont, 1989; Brito etal., 1989; Kim et al., 1995), and humans (Kim et al., 1997;Jassim et al., 1992) have revealed, in the case of humansperm, as many as 16 silver-stainable protein bands, rangingfrom 148 to 14.5 kDa (Kim et al., 1997). Only a few proteinshave thus far been characterized and localized to the fibroussheath, although these proteins have often proven to beencoded by genes showing a testis-specific pattern of ex-pression. The testis-specific A-kinase anchor proteinsAKAP3 (Mandal et al., 1999; Vijayaraghavan et al., 1999)and AKAP4 (Carrera et al., 1994; Fulcher et al., 1995;Turner et al., 1998) have been localized to the ribs andlongitudinal columns (mouse AKAP4) (Johnson et al., 1997)and the circumferential ribs (human AKAP3) (Mandal et al.,1999). In addition to postulated structural roles, AKAP3 andAKAP4 have both been shown (Turner et al., 1998; Vija-yaraghavan et al., 1999) to bind the regulatory (RII) subunitof cAMP-dependent protein kinase A (PK-A).
Ropporin, another sperm-specific protein that bindsrhophilin (Nakamura et al., 1999) has been localized to theinner and outer surfaces of the ribs of the mouse fibroussheath (Fujita et al., 2000). Interestingly, ropporin’s bindingpartner, rophilin, binds the GTP-bound form of the GTPaseRho and localizes to the outer surface of the outer densefibers (Fujita et al., 2000). Ropporin, like CABYR, has a 40
amino acid sequence at its amino terminus that shows high© 2002 Elsevier Science (USA
homology to that of the regulatory subunit of type IIcAMP-dependent protein kinase (Fujita et al., 2000).
The findings of RII homology domains in CABYR as wellas ropporin suggest that the testis-specific AKAPs mayserve as a scaffold for integrating PK-A, Rho, and calciumsignaling. Much work remains to confirm this hypothesis,including demonstration of protein–protein interactionsbetween AKAPs and CABYR and/or ropporin and the finestructural localization of these proteins in complexes.Nonetheless, the idea that the testis-specific AKAPs orga-nize and order calcium and GTPase activities in addition tobeing sites for PKA action may provide a stimulus to newstudies to expand our view of the function of the testis-specific AKAPs and the fibrous sheath.
Capacitation and the Temporal Aspects of CABYRPhosphorylation and Calcium Binding
Almost 50 years have elapsed since the independentdiscoveries of capacitation by Chang and Austin, but mo-lecular mechanisms to explain this process are not yet fullyunderstood. Our studies of CABYR provide an added dimen-sion to the understanding of capacitation-related molecularevents in the flagellum. Although the time course forcapacitation in vitro differs from species to species, amedian time for in vitro capacitation of human sperm is 3to 6 h. This time course is similar to that which we haveobserved for CABYR phosphorylation, leading to the hy-pothesis that a tyrosine phosphorylation-dependent eventresulting in the calcium-binding capacity of CABYR mayrepresent one molecular mechanism underlying the tempo-ral requirements for capacitation and hyperactivation.
ACKNOWLEDGMENTS
This work was supported by NIH grants U54 29099, P30 28934,and D43 HD 00654 from the Fogarty International Center, TheAndrew W. Mellon Foundation, and Schering AG.
REFERENCES
Akiyama, T., Ishida, J., Nakagawa, S., Ogawara, H., Watanabe, S.,Itoh, N., Shibuya, M., and Fukami, Y. (1987). Genistein, a specificinhibitor of tyrosine-specific protein kinases. J. Biol. Chem. 262,5592–5595.
Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z.,Miller, W., and Lipman, D. J. (1997). Gapped BLAST and Psi-BLAST: A new generation of protein database search programs.Nucleic Acids Res. 25, 3389–3402.
Attwood, T. K., Beck, M. E., Flower, D. R., Scordis, P., and Selley,J. (1998). The PRINTS protein fingerprint database in its fifthyear. Nucleic Acids Res. 26, 304–308.
Austin, C. R. (1951). Observations on the penetration of the sperminto the mammalian egg. Aust. J. Sci. Res. 4, 581–596.
Bradley, M. P., and Forrester, I. T. (1980). A sodium-calciumexchange mechanism in plasma membrane vesicles isolated
from ram sperm flagella. FEBS Lett. 121, 15–18.). All rights reserved.

252 Naaby-Hansen et al.
Breitbart, H., Darshan, R., and Rubinstein, S. (1984). Evidence forthe presence of ATP-dependent calcium pump and ATPaseactivities in bull sperm head membranes. Biochem. Biophys.Res. Commun. 122, 479–484.
Breitbart, H., Rubinstein, S., and Nass-Arden, L. (1985). The role ofcalcium and Ca21-ATPase in maintaining motility in ram sper-matozoa. J. Biol. Chem. 260, 11548–11553.
Breitbart, H., Stern, B., and Rubinstein, S. (1983). Calcium transportand Ca21-ATPase activity in ram spermatozoa plasma membranevesicles. Biochim. Biophys. Acta 728, 349–355.
Brito, M., Figueroa, J., Maldonado, E. U., Herr, J. C., and Burzio,L. O. (1989). The major component of the rat sperm fibroussheath is a phosphoprotein. Gamete Res. 22, 205–217.
Bronson, R. A., and Fusi, F. (1990). Sperm-oolemmal interaction:Role of the Arg-Gly-Asp (RGD) adhesion peptide. Fertil. Steril.54, 527–529.
Buck, J., Sinclair, M. L., Schapal, L., Cann, M. J., and Levin, L. R.(1999). Cytosolic adenylate cyclase defines a unique signalingmolecule in mammals. Proc. Natl. Acad. Sci. USA 96, 79–84.
Camatini, M., Anelli, G., and Casale, A. (1986). Immunocyto-chemical localization of calmodulin in intact and acrosome-reacted boar sperm. Eur. J. Cell Biol. 41, 89–96.
Carr, D. W., Fujita, A., Stentz, C. L., Liberty, G. A., Olsen, G. E.,and Narumiya, S. (2001). Identification of sperm specific proteinsthat intract with A-kinase anchoring proteins in a mannersimilar to the type II regulatory subunit of PKA. J. Biol. Chem.276, 17332–17338.
Carrera, A., Gerton, G. L., and Moss, S. B. (1994). The major fibroussheath polypeptide of mouse sperm: Structural and functionalsimilarities to the A-kinase anchoring proteins. Dev. Biol. 165,272–284.
Carrera, A., Moos, J., Ning, X. P., Gerton, G. L.,Tesarik, J., Kopf,G. S., and Moss, S. B. (1996). Regulation of protein tyrosinephosphorylation in human sperm by a calcium/calmodulin-dependent mechanism: Identification of A kinase anchor pro-teins as major substrates for tyrosine phosphorylation. Dev. Biol.180, 284–296.
Chang, M. C. (1951). Fertilizing capacity of spermatozoa depositedinto the fallopian tubes. Nature 168, 697–698.
Chang, M. C. (1955). Development of fertilizing capacity of rabbitspermatozoa in the uterus. Nature 175, 1036–1037.
Chen, J., and Varner, J. E. (1985). An extracellular matrix protein inplants: Characterization of a genomic clone for carrot extensin.EMBO J. 4, 2145–2151.
Chen, Y., Cann, M. J., Litvin, T. N., Iourgenko, V., Sinclair, M. L.,Levin, L., and Buck, J. (2000). Soluble adenylyl cyclase as aevolutionary conserved bicarbonate sensor. Science 289, 625–628.
Eddy, E. M., O’Brien, D. A., Fenderson, B. A., and Welch, J. E.(1991). Intermediate filament-like proteins in the fibrous sheathof the mouse sperm flagellum. Ann. N. Y. Acad. Sci. 637,224–239.
Fawcett, D. W. (1970). A comparative view of sperm ultrastructure.Biol. Reprod. 2(Suppl.), 90–126.
Fawcett, D. W. (1975). The mammalian spermatozoa. Dev. Biol. 44,394–436.
Feinberg, A. P., and Vogelstein, B. (1983). A technique for radiola-beling DNA restriction endonuclease fragments to high specificactivity. Anal. Biochem. 132, 6–13.
Fujita, A., Nakamura, K., Kato, T., Watanabe, N., Ishizaki, T.,
Kimura, K., Mizoguchi, A., and Narumiya, S. (2000). Ropporin, a© 2002 Elsevier Science (USA
sperm-specific binding protein of rhophilin, that is localized inthe fibrous sheath of sperm flagella. J. Cell Sci. 113, 103–112.
Fulcher, K. D., Mori, C., Welch, J. E., O’Brien, D. A., Klapper, D. G.,and Eddy, E. M. (1995). Characterization of Fsc1 cDNA for amouse sperm fibrous sheath component. Biol. Reprod. 52, 41–49.
Galantino-Homer, H. L., Visconti, P. E., and Kopf, G. S. (1997).Regulation of protein tyrosine phosphorylation during bovinesperm capacitation by a cyclic adenosine 3959-monophosphate-dependent pathway. Biol. Reprod. 56, 707–719.
Garty, N. B., Galiani, D., Aharonheim, A., Ho, Y. K., Phillips,D. M., Dekel, N., and Salomon, Y. (1988). G-proteins in mam-malian gametes: An immunocytochemical study. J. Cell Sci.91(Pt 1), 21–31.
Garty, N. B., and Salomon, Y. (1987). Stimulation of partiallypurified adenylate cyclase from bull sperm by bicarbonate. FEBSLett. 218, 148–152.
Glassner, M., Jones, J., Kligman, I., Woolkalis, M. J., Gerton, G. L.,and Kopf, G. S. (1991). Immunocytochemical and biochemicalcharacterization of guanine nucleotide-binding regulatory pro-teins in mammalian spermatozoa. Dev. Biol. 146, 438–450.
Gross, M. K., Toscano, D. G., and Toscano, W. A. (1987). Cal-modulin-mediated adenylate cyclase from mammalian sperm.J. Biol. Chem. 262, 8672–8676.
Henikoff, S., and Henikoff, J. G. (1994). Protein family classifica-tion based on searching a database of blocks. Genomics 19,97–107.
Henzel, W. J., Billeci, T. M., Stults, J. T., Wong, S. C., Grimley, C.,and Watanabe, C. (1993). Identifying proteins from two-dimensional gels by molecular mass searching of peptide frag-ments in protein sequence databases. Proc. Natl. Acad. Sci. USA90, 5011–5015.
Hinsch, K. D., Hinsch, E., Aumuller, G.,Tychowiecka, I., Schultz,G., and Schill, W. B. (1992). Immunological identification of Gprotein alpha- and beta-subunits in tail membranes of bovinespermatozoa. Biol. Reprod. 47, 337–346.
Hyne, R. V., and Garbers, D. L. (1979). Regulation of guinea pigsperm adenylate cyclase by calcium. Biol. Reprod. 21, 1135–1142.
Irvine, D. S., and Aitkin, R. J. (1986). Measurement of intracellularcalcium in human spermatozoa. Gamete Res. 15, 57–71.
Jassim, A., Gillott, D. J., and Al-Zuhdi, Y. (1992). Isolation andbiochemical characterization of the human sperm tail fibroussheath. Hum. Reprod. 7, 86–94.
Jethmalani, S. M., Henle, K. J., and Kaushal, G. P. (1994). Heatshock induced prompt glycosylation. J. Biol. Chem. 269, 23603–23609.
Johnson, L., Foster, J. A., Haig-Ladewig, L., Vanscoy, H., Moss,S. M., and Gerton, G. L. (1997). Assembly of AKAP82, a proteinkinase A anchor protein, into the fibrous sheath of mouse sperm.Dev. Biol. 192, 340–350.
Johnston, R. C., Mbizvo, M. T., Summerbell, D., Kovacs, G. T., andBaker, H. W. (1994). Relationship between stimulated hyperacti-vated motility of human spermatozoa and pregnancy rate indonor insemination: A preliminary report. Hum. Reprod. 9,1684–1687.
Kang, H., Freund, C., Duke-Cohan, J. S., Musacchio, A., Wagner,G., and Rudd, C. E. (2000). SH3 domain recognition of a proline-independent tyrosine-based RKxxYxxY motif in immune adaptorSKAP55. EMBO J. 19, 2889–2899.
Keller, B., and Lamb, C. J. (1989). Specific expression of a novel cellwall hydroxyproline-rich glycoprotein gene in lateral root initia-
tion. Genes Dev. 3, 1639–1646.). All rights reserved.

253CABYR, a Novel Fibrous Sheath Protein
Kim, Y. H., de Kretser, D. M, Temple-Smith, P. D., Hearn,M. T. W., and McFarlane, J. R. (1997). Isolation and characteriza-tion of human and rabbit sperm tail fibrous sheath. Mol. Hum.Reprod. 3, 307–313.
Kim, Y. H., McFarlane, J. R., O’Bryan, M. K., Almahbobi, G.,Stanton, P. G., Temple-Smith, P. D., de Kretser, D. M. (1995).Isolation and partial characterization of rat sperm tail fibroussheath proteins and comparison with rabbit and human sperma-tozoa using a polyclonal antiserum. J. Reprod. Fertil. 104, 107–114.
Kligman, D., and Hilt, D. C. (1988). The S100 protein family.Trends Biochem. Sci. 13, 437–443.
Kong, M., Richardson, R. T., Widgren, E. E., and O’Rand, M. G.(1995). Sequence and localization of the mouse sperm autoanti-genic protein, Sp17. Biol. Reprod. 53, 579–590.
Lea, I. A., Richardson, R. T., Widgren, E. E., and O’Rand, M. G.(1996). Cloning and sequencing of cDNAs encoding the humansperm protein, Sp17. Biochim. Biophys. Acta 1307, 263–266.
Leclerc, P., de Lamirande, E., and Gagnon, C. (1996). Cyclicadenosine 39,59monophosphate-dependent regulation of proteintyrosine phosphorylation in relation to human sperm capacita-tion and motility. Biol. Reprod. 55, 684–692.
Lindemann, C. B., and Goltz, J. S. (1988). Calcium regulation offlagellar curvature and swimming pattern in Triton X-100–extracted rat sperm. Cell Motil. Cytoskeleton 10, 420–431.
Mandal, A., Naaby-Hansen, S., Wolkowicz, M. J., Klotz, K., Shetty,J., Retief, J. D., Coonrod, S. A., Kinter, M., Sherman, N., Cesar, F.,Flickinger, C. J., and Herr, J. C. (1999). A testis-specific 95kilodalton fibrous sheath antigen that undergoes tyrosine phos-phorylation in capacitated human spermatozoa. Biol. Reprod. 61,1184–1197.
Maruyama, K., Mikawa, T., and Ebashi, S. (1984). Detection ofcalcium binding proteins by 45Ca autoradiography on nitrocellu-lose membrane after sodium dodecyl sulfate gel electrophoresis.J. Biochem. 95, 511–519.
Matsudaira, P. (1987). Sequence from picomole quantities of pro-teins electroblotted onto polyvinylidene difluoride membranes.J. Biol. Chem. 262, 10035–10038.
Moore, P. B., and Dedham, J. R. (1984). Calmodulin, a calmodulinacceptor protein, and calcimedins: Unique antibody localizationin hamster sperm. J. Cell. Biochem. 25, 99–107.
Musacchio, A., Nobel, M., Pauptit, R., Wierenga, R., and Saraste,M. (1992). Crystal structure of an src-homology 3 (SH3) domain.Nature 359, 851–855.
Naaby-Hansen, S., Flickinger, C. J., and Herr, J. C. (1997a). Two-dimensional gel electrophoretic analysis of vectorially labeledsurface proteins of human spermatozoa. Biol. Reprod. 56, 771–787.
Naaby-Hansen, S., Roof, R. W., Flickinger, C. J., Grafer, C. M.,Parsons, S. J., and Herr, J. C. (1997b). A novel negative imagingtechnique for accurate localization of stainable proteins oncomplex two-dimensional autoradiograms. Electrophoresis 18,2065–2070.
Nakamura, K., Fujita, A., Murata, M., Watanabe, G., Mori, C.,Fujita, J., Watanabi, N., Ishizaki, T., Yoshida, O., and Narumiya,S. (1999). Rhophilin, a small GTPase Rho-binding protein, isabundantly expressed in the mouse testis and localized in theprincipal piece of the sperm tail. FEBS Lett. 445, 9–13.
Nassar, A., Mahony, M., Morshedi, M., Lin, M. H., Srisombut, C.,and Oehninger, S. (1999a). Modulation of sperm tail proteintyrosine phosphorylation by pentoxifylline and its correlation
with hyperactivated motility. Fertil. Steril. 71, 919–923.© 2002 Elsevier Science (USA
Nassar, A., Morshedi, M., Mahony, M., Srisombut, C., Lin, M. H.,and Oehninger, S. (1999b). Pentoxifylline stimulates varioussperm motion parameters and cervical mucus penetrability inpatients with asthenozoospermia. Andrologia 31, 9–15.
Newlon, M. G., Roy, M., Morikis, D., Hausken, Z. E., Coghlan, V.,Scott, J. D., and Jennings, P. A. (1999). The molecular basis forprotein kinase A anchoring revealed by solution NMR. Nat.Struct. Biol. 6, 222–227.
Oko, R., and Clermont, Y. (1989). Light microscopic immunocyto-chemical study of fibrous sheath and outer dense fiber formationin the rat spermatid. Anat. Rec. 225, 46–55.
Okuno, M., and Brokaw, C. J. (1981). Calcium-induced change inform of demembrinated sea urchin sperm flagella immobilizedby vanadate. Cell Motil. Cystoskeleton 1, 349–362.
Olivier, J. P., Raabe, T., Henkemeyer, M., Dickson, B., Mbamalu,G., Margolis, B., Schlessinger, J., Hafen, E., and Pawson, T. (1993).A Drosophila SH2-SH3 adaptor protein implicated in couplingthe sevenless tyrosine kinase to an activator of Ras guaninenucleotide exchange, Sos. Cell 73, 179–191.
Olson, G. E., Hamilton, D. W., and Fawcett, D. W. (1976). Isolationand characterization of the fibrous sheath of rat epididymalspermatozoa. Biol. Reprod. 14, 517–530.
Osheroff, J. E., Visconti, P. E., Valenzuela, J. P., Travis, A. J.,Alvarez, J., and Kopf, G. S. (1999). Regulation of human spermcapacitation by a cholesterol efflux-stimulated signal transduc-tion pathway leading to protein kinase A-mediated up-regulationof protein tyrosine phosphorylation. Mol. Hum. Reprod. 5, 1017–1026.
Parisel, C. C., Weinman, J. S., Escaig, F. T., Guyot, M. Y., Iftode,F. C., Weinmam, S. J., and Damaille, J. G. (1984). Analyticalsubcellular distribution of cAMP-dependent protein kinase ac-tivity in bull spermatozoa. Gamete Res. 10, 433–444.
Pawson, T., and Schlessinger, J. (1993). SH2 and SH3 domains.Curr. Biol. 3, 434–442.
Pearson, W. R., and Lipman, D. J. (1988). Improved tools forbiological sequence comparison. Proc. Natl. Acad. Sci. USA 85,2444–2448.
Potts, B. C. M., Smith, J., Akke, M., Macke, T. J., Okazaki, K.,Hidaka, H., Case, D., and Chazin, W. J. (1995). The structure ofcalcyclin reveals a novel homodimeric fold for S100 Ca21-bindingproteins. Nat. Struct. Biol. 2, 790–796.
Richardson, R. T., Yamasaki, N., and O’Rand, M. G. (1994).Sequence of a rabbit sperm zona pellucida binding protein andlocalization during the acrosome reaction. Dev. Biol. 165, 688–701.
Rigoni, F., Dell’Antone, P., and Deana, R. (1987). Evidence for adelta pH-driven Ca21 uptake in EGTA-treated bovine spermato-zoa. Eur. J. Biochem. 169, 417–422.
Rotem, R., Paz, G. F., Homonnai, Z. T., Kalina, M., and Naor, Z.(1990). Protein kinase C is present in human sperm: Possible rolein flagellar motility. Proc. Natl. Acad. Sci. USA 87, 7305–7308.
Ruff-Jamisson, S., Chen, K., and Cohen, S. (1993). Induction by EGFand interferon-g of tyrosine phosphorylated DNA binding pro-teins in mouse liver nuclei. Science 261, 1733–1736.
Rustandi, R. R., Drohat, A. C., Baldisseri, D. M., Wilder, P. T., andWeber, D. (1998). The Ca21-dependent interaction of S100B(BB)with a peptide derived from p53. Biochemistry 37, 1951–1960.
Suarez, S. S. (1988). Hamster sperm motility transformation duringdevelopment of hyperactivation in vitro and epididymal matura-
tion. Gamete Res. 19, 51–65.). All rights reserved.

254 Naaby-Hansen et al.
Suarez, S. S., and Dai, X. (1995). Intracellular calcium reachesdifferent levels of elevation in hyperactivated and acrosome-reacted hamster sperm. Mol. Reprod. Dev. 42, 325–333.
Suarez, S. S., Varosi, S. M., and Dai, X. (1993). Intracellular calciumincreases with hyperactivation in intact, moving hamster spermand oscillates with the flagellar beat cycle. Proc. Natl. Acad. Sci.USA 90, 4660–4664.
Thomas, P., and Meizel, S. (1988). An influx of extracellularcalcium is required for initiation of the human sperm acrosomereaction induced by human follicular fluid. Gamete Res. 20,397–411.
Thomas, P., and Meizel, S. (1989). Phosphatidylinositol 4,5-biphosphate hydrolysis in human sperm stimulated with follicu-lar fluid or progesterone is dependent upon Ca21 influx. Biochem.J. 264, 539–564.
Turner, R. M., Johnson, L. J., Haig-Ladewig, L., Gerton, G. L., andMoss, S. B. (1998). An X-linked gene encodes a major humansperm fibrous sheath protein, hAKAP82: Genomic organization,protein kinase A-RII binding, and distribution of the precursor inthe sperm tail. J. Biol. Chem. 273, 32135–32141.
Vijayaraghavan, S., Liberty, G. A., Mohan, J., Winfrey, V. P., Olson,G. E., and Carr, D. W. (1999). Isolation and molecular character-ization of AKAP110, a novel, sperm specific protein kinaseA-anchoring protein. Mol. Endocrinol. 13, 705–717.
Visconti, P. E., Bailey, J. L., Moore, J. D., Pan, D., Olds-Clarke, P.,and Kopf, G. S. (1995a). Capacitation in mouse spermatozoa. I.Correlation between the capacitation state and protein tyrosinephosphorylation. Development 121, 1129–1137.
Visconti, P. E., and Kopf, G. S. (1998). Regulation of proteinphosphorylation during sperm capacitation. Biol. Reprod. 59,1–6.
Visconti, P. E., Moore, G. D., Bailey, J. L., Leclerc, P., Connors,S. A., Pan, D., Olds-Clarke, P., and Kopf, G. S. (1995b). Capaci-tation of mouse spermatozoa. II. Protein tyrosine phosphoryla-
tion and capacitation are regulated by a cAMP-dependent path-way. Development 121, 1139–1150.© 2002 Elsevier Science (USA
Visconti, P. E., and Tezon, J. G. (1989). Phorbol esters stimulatecyclic adenosine 39, 59-monophosphate accumulation in hamsterspermatozoa during in vitro capacitation. Biol. Reprod. 40,223–231.
Wasco, W. M., and Orr, G. A. (1984). Function of calmodulin inmammalian sperm: Presence of a calmodulin-dependent cyclicnucleotide phosphodiesterase associated with demembranatedrat caudal epididymal sperm. Biochem. Biophys. Res. Commun.118, 636–642.
Wennemuth, G., Westenbroek, R. E., Xu, T., Hille, B., and Babcock,D. F. (2000). Cav2.2 and Cav2.3 (N- and R-type) Ca21 channels indepolarization-evoked entry of Ca21 into mouse sperm. J. Biol.Chem. 275, 21210–21217.
Weyand, I., Gade, M., Frings, S., Weiner, J., Muller, F., Altenhafen,W., Hatt, H., and Kaupp, U. B. (1998). Cloning and functionalexpression of a cyclic-nucleotide-gated channel from mamma-lian sperm. Nature 368, 859–863.
Wiesner, B., Weiner, J., Middendorff, R., Hagen, V., Kaupp, U. B.,and Weyand, I. (1998). Cyclic nucleotide-gated channels on theflagellum control Ca21 entry into sperm. J. Cell Biol. 142,473–484.
Yanagimachi, R. (1970). The movement of golden hamster sperma-tozoa before and after capacitation. J. Reprod. Fertil. 23, 193–196.
Yanagimachi, R. (1982). Requirement of extracellular calcium ionsfor various stages of fertilization and fertilization related phe-nomena in the hamster. Gamete Res. 5, 323–344.
Yanagimachi, R. (1994). Mammalian fertilization. In “The Physi-ology of Reproduction, 2nd ed.” (E. Knobil and J. D. Neill, Eds.),pp. 189–317. Raven Press, New York.
Yovich, J. M., Edirisinghe, W. R., Cummins, J. M., and Yovich, J. L.(1990). Influence of pentoxifylline in severe male factor infertil-ity. Fertil. Steril. 53, 715–722.
Received for publication January 8, 2001Revised October 31, 2001
Accepted October 31, 2001Published online January 15, 2002
). All rights reserved.