universidad de salamanca - repositorio digital...
TRANSCRIPT

UNIVERSIDAD DE SALAMANCA
Centro Hispano Luso de Investigaciones Agrarias
Facultad de Biología
Producción de silimarina en cultivos celulares de
Cardo mariano (Silybum marianum): efecto de
distintos elicitores
Lourdes Ortega Maldonado
Tutora: Dra. Purificación Corchete Sánchez
Salamanca
Junio 2012

Índice
1. Introducción ...........................................................................................................1
2. Antecedentes y Objetivos ....................................................................................10
3. Materiales y Métodos ..........................................................................................13
3.1. Material vegetal ........................................................................................... 13
3.2. Tratamientos ................................................................................................ 13
3.3. Análisis de flavonolignanos ........................................................................ 13
3.4. Extracción y cuantificación de RNA .......................................................... 14
3.5. Síntesis de cDNA ........................................................................................ 14
3.6. Cuantificación de fragmento de DNA ......................................................... 16
4. Resultados y Discusión .......................................................................................17
4.1. Efecto de distintos elicitores en el crecimiento celular y la producción de
silimarina ................................................................................................................ 17
4.1.1. Efecto en el crecimiento ...................................................................... 17
4.1.2. Efecto en la producción de silimarina .................................................. 17
4.1.2.1. Efectos del tiempo de contacto .......................................................... 17
4.1.2.2. Efecto de distintas dosis de elicitores en producción de silimarina ... 23
4.1.2.3. Efectos de la adición de precursores a cultivos elicitados ................. 26
4.2. Efecto de la elicitación en la expresión génica de una proteína con
homología a proteínas transportadoras del tipo MDR-ABC .................................. 29
5. Conclusiones .......................................................................................................31
6. Agradecimientos ..................................................................................................32
7. Bibliografía ..........................................................................................................33

1
1. Introducción
El metabolismo secundario se puede definir como la biosíntesis, la
transformación y la degradación de los compuestos endógenos mediante proteínas de
especialización, las cuales se han formado como resultado de los procesos de
diferenciación y se clasifican según su significación biológica y función en la célula
productora (Valdés y Balbín, 2000).
Según la definición en el marco ecológico propuesta por Stransburger y col.,
(1994), estos compuestos son sustancias ecológicamente eficaces, frente a
compuestos primarios que serían sustancias fisiológicamente eficaces. Una de las
principales diferencias que presentan los metabolitos secundarios con relación a los
primarios es su distribución limitada en el reino vegetal; mientras que los
compuestos primarios se encuentran en todo el reino y las diferencias entre especies
solo son de índole cuantitativa (Ramos y col., 1998).
Estos compuestos químicos se presentan típicamente en sólo una especie o un
grupo de plantas taxonómicamente relacionadas (Gabriela y col., 1997), por lo que
muchos de ellos se consideran marcadores taxonómicos de familias y géneros
(García, 2004). El metabolismo secundario está estrechamente ligado al primario ya
que este proporciona un gran número de moléculas simples, como el ácido
siquímico, el acetato y los aminoácidos, los cuales constituyen los materiales de
partida para las rutas biosintéticas. La figura 1 describe la interconexión de los
principales grupos de metabolitos secundarios y su relación con la fotosíntesis. La
biosíntesis de las estructuras fenólicas se efectúa por dos rutas metabólicas
esenciales: la del ácido siquímico y la del ácido malónico (conocida como la de los
policétidos); la primera es mayoritaria en las plantas superiores y la segunda es
favorecida en los microorganismos. La fenilalanina y la tirosina son intermediarios
metabólicos en la biosíntesis de numerosos compuestos fenólicos, y el triptófano es
el precursor de hormonas como el ácido indolacético.

2
Figura. 1. Vías de síntesis de los principales grupos de metabolitos secundarios y su
relación con la fotosíntesis.
La síntesis específica de cada compuesto suele estar restringida a estados
específicos del desarrollo respectivo de cada tipo de organismo, células
especializadas y períodos de estrés causados por la deficiencia de nutrientes o por el
ataque de microorganismos. Este fenómeno se debe a la formación, dependiente de
fase, de la correspondiente enzima, lo que significa que la expresión del metabolismo
secundario se basa en un proceso de diferenciación (Azcón y Talón, 2000).
Cada tipo de compuesto secundario está estrechamente relacionado con una o
varias funciones específicas en la planta que lo contiene. En algunos casos
intervienen en las relaciones de competencia con otras plantas, actuando como
agentes alelopáticos. Muchos de ellos son efectivos contra invasiones de hongos,
bacterias y virus (Harborne, 1993) y en muchos casos intervienen en la defensa
causando toxicidad (Sepúlveda y col., 2004); actúan en relaciones de mutualismo, en
la atracción de polinizadores y en la dispersión de semillas (Ramos y col., 1998),
como protección contra la radiación ultravioleta y la desecación (Ghasempour y col.

3
1998), como reserva de material nitrogenado, así como en la fijación del N2
atmosférico, la formación de nódulos y la relación simbiótica en las raíces de las
leguminosas (Stafford, 1997).
Los metabolitos secundarios son además útiles para el hombre. Una de las
áreas principales de aplicación es la fitomedicina, donde varios miles de plantas están
siendo usadas en todo el mundo para el tratamiento de enfermedades humanas. Se
emplean también como estimulantes (cafeína y nicotina), para la obtención de
fragancias (aceites esenciales), saborizantes (aceites esenciales, capsaicina, piperina),
colorantes naturales, venenos (estricnina) y alucinógenos (morfina, heroína, cocaína,
tetrahidrocannabinol) por poner sólo algunos ejemplos. Muchos de estos productos
que tienen propiedad insecticida, fungicida y fitotóxicas, pueden ser utilizados en
agricultura como protectores naturales de plantas dando una ventaja ecológica, ya
que se degradan fácilmente en las plantas y en el suelo, a diferencia de los pesticidas
sintéticos que son más resistentes y persistentes. Por lo tanto, es un desafío para
biotecnólogos encontrar maneras de producir estos compuestos en la cantidad y
calidad suficiente (Wildi y Wink, 2002). La manera principal y tradicional es hacer
crecer las plantas respectivas en el campo o en invernaderos para extraer el producto
acumulado.
Para la obtención de estos compuestos de interés, el cultivo de células
vegetales ha surgido como una alternativa. En estos sistemas, la producción es
independiente de factores externos como disponibilidad de tierra, clima y
condiciones geopolíticas; además es posible controlar las condiciones de cultivo,
alcanzando mayor rendimiento y calidad. Sin embargo, para lograr la aplicación
industrial de esta estrategia es necesario resolver algunos problemas como la baja
productividad, el crecimiento lento y la inestabilidad genética (Sajc y col., 2000).
Los cultivos de células en suspensión se caracterizan por proporcionar células
individuales distribuidas en forma homogénea en el medio de cultivo, facilitándose
así la transferencia de nutrientes y oxígeno hacia el citoplasma. Este tipo de cultivo
presenta la ventaja de permitir el control relativamente sencillo de variables como
temperatura, pH y oxígeno disuelto; sin embargo, pueden verse modificadas algunas

4
características de las células presentes en las plantas como su diferenciación y la
comunicación intercelular (Schlatmann y col., 1996), lo que implica en muchos casos
la disminución en la producción de metabolitos secundarios, pues se ha reportado
que en algunas especies la síntesis de ciertos metabolitos requiere la coexistencia de
diferentes tipos celulares o la compartimentalización intracelular (Trejo y Rodríguez,
2007).
Las suspensiones celulares finas están constituidas principalmente por células
meristemáticas indiferenciadas, más débiles e inestables en comparación con su
estado en el ambiente natural (Yeoman y Yeoman, 1996), pero con respecto a los
cultivos de células diferenciadas (raíces y brotes), presentan las ventajas de poder ser
cultivadas a mayores concentraciones celulares y a mayores velocidades de
crecimiento (Trejo y Rodríguez, 2007).
En el cultivo en suspensión el estado que más se aproxima a las condiciones
naturales es la fase estacionaria, en la que diversos autores han reportado un cierto
grado de diferenciación fisiológica (Bougard, y col., 2001), observándose que la
acumulación de metabolitos secundarios se presenta principalmente durante este
periodo; sin embargo, en caso de metabolitos asociados al crecimiento, su
acumulación puede ser proporcional a la producción de biomasa y se ha observado
durante la fase exponencial en ciertas líneas celulares, tales son los casos de la
producción de betalaínas, carotenoides y azadiractina (Aria y col., 2009).
Varias son las causas de la baja productividad en los cultivos. Para iniciarlos,
el material más adecuado es una masa de células indiferenciada (callo) y para
obtenerla, las hormonas necesarias, como el ácido 2,4 diclorofenoxiacético y los
ciclos sucesivos de división celular pueden determinar la aparición de mutaciones y
por tanto de variación somaclonal, lo que aumenta la heterogeneidad del cultivo. En
otros casos, al ser los metabolitos secundarios productos de la diferenciación celular,
las rutas biosintéticas no se activan en cultivos celulares indiferenciados (Verpoorte,
2002). Otra de las causas de la baja producción puede ser debida a que muchos de
esos productos son tóxicos para las células que los producen, aunque de forma
natural en la planta completa se han desarrollado mecanismos de resistencia a estos

5
compuestos. Estos mecanismos pueden ser: excreción extracelular, secuestración
vacuolar, transporte vesicular, biosíntesis extracelular y acumulación del metabolito
en una forma no tóxica (Sirikantaramas y col., 2008). En la figura 2 aparecen los
principales procesos implicados en la síntesis de un metabolito dado y su destino
final, que puede ser el de la acumulación en su forma activa en un compartimento, su
exportación, su degradación o su modificación hacia una forma no tóxica para la
célula.
Figura 2. Síntesis y acumulación de metabolitos secundarios (vías pre-biosintética,
biosintética, y post-biosíntesis).
El diseño de estrategias para aumentar la productividad de los cultivos
celulares vegetales radica en dos puntos: el primero es la intervención en los
mecanismos de regulación a la que están sometidas las complejas vías biosintéticas
(vías pre- y biosintéticas) y el segundo es la intervención en los mecanismos post-
biosintéticos, y más concretamente en aquellos que conducen a la liberación de
producto al medio extracelular (Wei y col., 2004).
Dentro de las estrategias más utilizadas y que se han mostrado relevantes para
solventar la baja producción de metabolitos secundarios en cultivos celulares
vegetales, destaca la elicitación (Arias y col., 2009).

6
La elicitación es un estímulo o factor de índole biológico o abiótico que
produce una perturbación en un sistema vivo. Dichos factores son llamados
elicitores y se clasifican, de acuerdo a su naturaleza, en bióticos (quitosano,
jasmonato de metilo (MJa), alginato, extractos fúngicos) (Dong y Zhong, 2002;
Conceicao y col., 2006; Ignatov y col., 1999; Wang y Zhong, 2002) y abióticos
(metales pesados, estrés térmico, estrés osmótico) (Yu y col., 2005; Wu y col., 2001)
o, de acuerdo a la interacción planta-elicitor, en generales (jasmonato de metilo,
ácido salicílico), si desencadenan una respuesta de defensa en cualquier planta, y
específicos (extractos fúngicos), cuando sólo actúan sobre una especie en particular
como se explica en la tabla 1 (Vasconsuelo y Boland, 2007).
El término elicitor surgió del conocimiento de las relaciones planta-patógeno
y su aplicación a la estimulación de rutas del metabolismo secundario vegetal deriva
del hecho de que muchas plantas, en respuesta a un ataque microbiano o al daño
químico o mecánico, sintetizan sustancias tóxicas de índole química diversa
conocidas como fitoalexinas (Zhao y col., 2005).
Tabla 1. Clasificación de factores usados como elicitores.
Naturaleza Interacción planta-elicitor
Bióticos Abióticos Generales Específicos
Quitosano Metales pesados MJa Extractos fúngicos
MJa Estrés térmico Acido salicílico
Alginato Estrés osmótico
Extractos fúngicos
De manera general, la secuencia de eventos en la respuesta de defensa
impulsada por un elicitor comprende los siguientes pasos: reconocimiento,
fosforilación y desfosforilación de proteínas de membrana y citoplasma, aumento en
la concentración de iones Ca2+
en el citoplasma, despolarización de la membrana,
flujo de iones (Cl-, K
+ y H
+), alcalinización extracelular, acidificación
citoplasmática, activación de proteínas quinasas activadas por mitógeno (MAPK),
activación de la NADPH oxidasa, producción de formas reactivas de O2, expresión
de genes de defensa tempranos, producción de etileno y jasmonato, expresión de

7
genes de defensa tardíos y finalmente acumulación de metabolitos secundarios
(Vasconsuelo y Boland, 2007; Zhao y col., 2005).
Uno de rutas más universalmente implicadas en la activación de la biosíntesis
de metabolitos secundarios es la de los compuestos lipídicos de la familia de los
octadecanoides que dan origen a la síntesis de ácidos jasmónico. Su derivado
metilado, el jasmonato de metilo (MJa) ha sido utilizado con éxito en la elicitación
de metabolitos secundarios en diversos cultivos celulares vegetales y se ha visto que
su aplicación aumenta la producción de alcaloides, terpenos, flavonoides y una gran
variedad de compuestos fenólicos (revisión en Namdeo, 2007).
La elicitación de una especie para la obtención de un metabolito particular en
cultivos de células vegetales, requiere la determinación de la dosis óptima y el
tiempo adecuado de aplicación del elicitor, así como la respuesta en el tiempo del
cultivo y los efectos combinados de distintos elicitores. En general los elicitores
inhiben el crecimiento, debido probablemente a que en su presencia las células
activan mecanismos de defensa desviando los nutrientes y la energía hacia el
metabolismo secundario (Thanh y col., 2005; Zhao y col., 2004). En cuanto al efecto
sobre la producción, se ha encontrado que hay un incremento en ésta a medida que
aumenta la concentración del elicitor hasta un valor máximo, después del cual
decrece nuevamente, probablemente por la saturación de los receptores o por que
dicha molécula puede ser tóxica para la célula. Estos efectos han sido estudiados en
diferentes especies, en las que se muestra un incremento en la producción como
resultado del proceso de elicitación (tabla 2).
El tiempo de aplicación del elicitor al cultivo es importante, ya que los
efectos pueden variar dependiendo de la fase de crecimiento al momento de aplicar el
tratamiento (Zhao y col., 2005). Además es importante estudiar el comportamiento
del cultivo con el tiempo, ya que el incremento en la productividad se observa cierto
tiempo después de la adición del elicitor (Gala y col., 2005; Sánchez y col., 2005). El
acoplamiento de un tipo de elicitor con otros factores aumenta la producción,
generándose un efecto sinérgico (Qu y col., 2006). Es importante tener en cuenta el
incremento en el costo del proceso a la hora de elegir el elicitor.

8
Tabla 2. Efecto de la elicitación en el crecimiento y la acumulación de metabolitos
secundarios en cultivos en suspensión de diferentes especies.
Especie Producto Elicitor T (°C)
Contenido
producto
(%)
Biomasa/l
(%)
Taxus
yunnanesis Paclitaxel Lantano 12 312 -8
Taxus
chinensis
Taxuyunanina
C MJa 7 46 13,5
Poligonum
tinctorium Indirubina
Rhizoctonia
solani 0 190 -
Cistanche
desertícola Feniletanoides
Fusarium
solanum 15 51 -7
Saussurea
medusa Jaseosioina Glutatión 5 100 -
Quizá uno de los aspectos menos estudiados en relación a la producción de
metabolitos secundarios en cultivos celulares, ha sido el de su liberación y
acumulación en el medio extracelular. Este hecho reviste un doble interés, ya que,
por una parte, su excreción puede aliviar el posible efecto tóxico sobre la biomasa
celular y, por otra, ya de índole biotecnológica, la recuperación del producto se ve
facilitada pudiéndose reutilizar la biomasa de forma repetida.
Uno de los agentes solubilizantes más utilizados son las ciclodextrinas (Cx),
compuestos cíclicos que se obtienen durante la degradación enzimática del almidón.
La cavidad interior de las ciclodextrinas es hidrófoba, por lo que estos compuestos
son capaces de albergar moléculas hidrófobas más pequeñas para formar complejos
“anfitrión-huésped“, en los que la molécula huésped queda encapsulada por la Cx.
En consecuencia, moléculas insolubles en agua pueden llegar a ser completamente
solubles mediante un tratamiento con disoluciones acuosas de Cx, sin que se
produzca modificación química alguna en la molécula huésped, ya que no se origina
ningún enlace covalente durante la interacción entre la Cx y la molécula insoluble en
agua (Martínez y Gómez, 2007).
Las Cx se parecen químicamente a oligosacáridos pépticos liberados de
paredes celulares durante el ataque por hongos y se han usado para aumentar la

9
biosíntesis de metabolitos secundarios y su liberación al medio extracelular (Bru y
col., 2006). Recientemente, Zamboni y col. (2009), han determinado que las Cx
desencadenan una cascada de señalización que activa diferentes familias de factores
de transcripción en cultivos de vid, induciendo el cese de la división celular, el
refuerzo de paredes celulares y la biosíntesis de trans-resveratrol y proteínas
relacionadas con la defensa. Loa autores también observaron la inducción de genes
relacionados con transportadores de metabolitos secundarios tales como los
pertenecientes a la familia de transportadores ABC (ATP-binding casette), lo que
explicaría la acumulación de resveratrol en el medio de cultivo.

10
2. Antecedentes y Objetivos
El cardo mariano, Silybum marianum L. (Gaertn), es una planta herbácea
anual o bianual de uso medicinal que pertenece a la familia de las Asteráceas
(Compositae). Originaria de una zona estrecha del Mediterráneo, se ha cultivado
desde hace siglos en toda Europa. También crece en la India, China, África y
Australia, fue llevada a América del Norte por los colonizadores europeos durante el
siglo 19 y ahora se encuentra establecida en los Estados Unidos y América del Sur.
S. marianum posee un tallo de 20-150 cm de alto, ligeramente velloso, recto y
ramificado en la parte superior. Las hojas son alternas, grandes, veteadas en blanco, y
con bordes fuertemente espinosos. Las inflorescencias son grandes y en capítulos
redondos, situadas de forma solitaria en el ápice del tallo o de sus ramas y rodeadas
por brácteas espinosas. Las flores son hermafroditas, con forma tubular y de corola
morado púrpura. Los frutos son aquenios de cutícula dura de 6 a 8 mm de largo,
brillantes y generalmente de color marrón, con un vilano blanco y sedoso en el ápice
(Kuromori y Shinozaki, 2010). Cada inflorescencia produce aproximadamente 100
semillas, y cada planta produce entre 10 y 50 capítulos por planta, con una media de
3.000 semillas por planta, las cuales pueden permanecen viables durante 9 años o
más (Liu, Miao y Zhang, 2011).
El cardo mariano tiene una gran variedad de compuestos flavonoides como la
apigenina, crysoeriol, eriodictiol, kaempferol, naringenina, quercetina y taxifolina,
pero sus propiedades medicinales se encuentran en las paredes de los frutos maduros
(aquenios), y se atribuyen a la silimarina que es un grupo de compuestos de
biogénesis mixta denominados flavonolignanos. Sus constituyentes se forman por
una reacción de acoplamiento oxidativo entre el flavonoide taxifolina y un
fenilpropanoide, generalmente el alcohol coniferílico (figura 3) (revisión en
Corchete, 2008). Estas estructuras derivan de dos de las principales rutas del
metabolismo primario, la ruta del ácido siquímico y la ruta del acetato (figura 4).
El complejo de la silimarina está formado por una mezcla de regio- y
estereoisómeros: silidianina, silicristina A y B, silibina A y B e isosilibina A y B.

11
Otros flavolignanos que se han detectado en menor proporción son silimonina,
silandrina, silihermina y neosilihermins A y B.
Figura 3. Síntesis de silibina por acoplamiento oxidativo entre taxifolina y alcohol
coniferílico.
Figura 4. Ruta de síntesis de taxifolina y alcohol coniferílico en la síntesis de
silimarina.
Taxifolina Alcohol Coniferílico
Silibina
(par de diasteroisómeros)
Oxidación Oxidación
Acoplamiento
oxidativo
Ataque
nucleofílico
Taxifolina Alcohol Coniferílico
Silibina
(par de diasteroisómeros)
Oxidación Oxidación
Acoplamiento
oxidativo
Ataque
nucleofílico
Ác. p-Cumárico
PALFenilalanina Ác. Cinámico
p-CumaroilCoA
CHS
Tetrahidrochalcona
Alcohol Coniferílico
Dihidrocaempferol
Taxifolina
Silidianina Silicristina Silibina
C4H
4CL
Acetil CoA
ACoAC
SS?
Malonil CoAx3
N3D
Ruta del Siquimato
Ruta del Acetato
??
+
PALFenilalanina Ác. Cinámico
p-Cumaroil CoA
COMT
Ác. Ferúlico
Alcohol Coniferílico
C4H
C3H
Ác. p-Cumárico
Cafeoil CoA
Feruloil CoA
Coniferil Aldehído
Ác. Cafeico
CCoA-3H
COMTCCoAOMT
4CL
4CL
4CL
CCR
CADSAD
Ruta del Siquimato

12
Los frutos del cardo mariano se han empleado en la medicina popular para el
tratamiento de trastornos del hígado. Los mecanismos de acción de silimarina son
variados y se ha descrito que es un potente antioxidante, actúa sobre las membranas
de hepatocitos y permeabilidad celular, se une y compite con receptores de toxinas y
fármacos, protegiendo a la célula de efectos tóxicos; estimula la síntesis de proteínas,
inhibición de la proliferación celular en la fibrosis hepática y actividad
antiinflamatoria (revisión en Corchete, 2008).
La producción de silimarina en cultivos celulares derivados de Silybum
marianum es muy baja (revisión en Corchete, 2008) y emula a lo que ocurre de
forma natural en el fruto, en el sentido de que es posible detectar estos compuestos
de forma constitutiva en el medio extracelular, que, técnicamente, puede considerarse
una extensión del apoplasto.
Entre las estrategias diseñadas para aumentar la producción, la elicitación, es
la que ha ofrecido hasta el momento los mejores resultados y así, en trabajos previos
se ha determinado que la aplicación de MJa, extracto de levadura y/o Cx aumenta la
producción de silimarina y su liberación al medio, incluida la de los precursores
alcohol coniferílico (AC) y taxifolina (Tx) (Madrid y Corchete, 2010; Sánchez y col.,
2005).
El objetivo del presente trabajo ha sido comprobar si la estrategia de la
elicitación es efectiva en la producción de silimarina en otras líneas celulares de
cardo mariano. Para ello se ha determinado la acción de MJa, y Cx, solos o en
combinación, en la acumulación de silimarina en cultivos derivados de una línea de
callo de cotiledón establecida en junio de 2011. Se ha comprobado de igual forma si
la coronatina (Cr), análogo estructural del MJa, es también un análogo funcional y,
por último, y teniendo en cuenta la presencia de silimarina en el medio extracelular,
se ha determinado el efecto de MJa y Cx en la expresión de un fragmento
secuenciado con anterioridad con homología al de una proteína transportadora del
tipo ABC.

13
3. Materiales y Métodos
3.1. Material vegetal
Los callos de los que derivó la línea celular empleada en este estudio se
establecieron a partir de explantes de cotiledón obtenidos de plántulas de Silybum
marianum germinadas en oscuridad. Las suspensiones se iniciaron en septiembre de
2011 en medio líquido de Murashige y Skoog (Murashige y Skoog, 1962),
suplementado con 30 g/l de sacarosa, 4,5 mmol/l de ácido 2,4-diclorofenoxiacético y
2,2 mmol/l de bencil adenina a pH 5,6. Los cultivos celulares se incubaron en
oscuridad a 25 ºC con una agitación de 90 rpm, siendo subcultivados cada 2 semanas.
Los cultivos se usaron para realización de los experimentos después de un periodo de
estabilización de 2 meses (8 subcultivos). Para ello, las células del subcultivo previo
se filtraron por gravedad a través de un tamiz estéril, inoculándose 3 gramos de peso
húmedo en matraces Erlenmeyer de 100 ml con 20 ml de medio de cultivo.
3.2. Tratamientos
Cultivos crecidos durante tres días fueron tratados con MJa 100 µM (solución
etanólica diluida), Cr 1 µM (solución acuosa), heptakis (2,6-di-O-metil)-β
ciclodextrina (Cx) 30 mM (solución acuosa) y la combinación de MJa 100 µM y Cx
30 mM, valorándose la producción de metabolitos secundarios a distintos tiempos de
tratamiento. Los controles recibieron la misma cantidad de solvente. Todos los
experimentos se realizaron por duplicado.
3.3. Análisis de flavonolignanos
El medio de cultivo se separó de la biomasa por filtración a vacio,
analizándose el contenido en silimarina separadamente en las células y en el medio.
El medio de cultivos se extrajo con 2 volúmenes de acetato de etilo (x2). Los
extractos combinados se secaron a vacío por debajo de 50 ºC y se resuspendieron en

14
0,5ml de metanol. Las muestras recibieron 50 mg de naringenina como patrón
interno antes de la extracción.
La biomasa se secó hasta peso constante en estufa entre 40-50 ºC. Cien mg de
peso seco se homogenizaron en metanol al 80 % y se extrajeron a 60 ºC durante 8
horas. Los extractos se filtraron, secaron a vacio y se resuspendieron en 1 ml de
metanol. Como en el caso anterior, antes de la extracción se añadieron 100 mg de
naringenina como patrón interno.
El análisis se efectuó por HPLC en una columna de fase reversa (Spherisorb
ODS-2 (5µm), 4,6 x 250 mm). Como fase móvil se usó una mezcla de 34
volúmenes de metanol y 66 volúmenes de ácido acético: agua (5:55 v/v) a 1 ml/min y
detección a 277 nm. La identificación de precursores (AC y Tx) y de los
componentes de la silimarina se llevó a cabo por comparación con estándares
comerciales.
3.4. Extracción y cuantificación de RNA
El RNA se extrajo de células separadas del medio de cultivo por filtración y
congeladas en nitrógeno líquido. Seguidamente se trituraron con un molino
mezclador MM 400 (Retsh, Alemania) en eppendorf, añadiendo 5 bolitas de acero de
3 mm de diámetro (Retsh, Alemania), durante 20 segundos a una frecuencia de 30
oscilaciones por segundo. El RNA se purificó mediante el reactivo Tri-Reagent®
siguiendo las indicaciones de la casa comercial (Ambion).
La concentración de ácidos nucléicos se cuantificó de manera automática
utilizando un espectrofotómetro ultrasensible (Nanodrop Technologies Inc).
3.5. Síntesis de cDNA
El cDNA se sintetizó a partir de 1 μg de RNA mediante el kit Super Script™
de Invitrogen. El cDNA se utilizó como molde para cuantificar por qRT PCR la
expresión de un fragmento de 103 pares de bases cuya secuencia presentó homología

15
a la de proteínas transportadoras del tipo MDR- ABC (proteínas ABC de resistencia
múltiple a drogas) (Prieto, 2011).
Los primers utilizados fueron:
Forward 5’-3’: GGGAGGTTCTTGGGAAGTGTCAGC
Reverse 5’-3’: AGTCGTTGCCCCATGCTCCAA
En las PCR de comprobación rutinaria se emplearon las siguientes condiciones.
Tabla 3. Condiciones de amplificación.
Procesos Temperatura ( ºC) Tiempo (min) Ciclos
Inicio 94 5 1
Desnaturalización 94 1
35 Anillamiento 59 1
Extensión 72 1
Elongación final 72 10 1
Conservación 4 indefinido 1
Tabla 4. Composición medio PCR
Reactivo Volumen
cDNA 1 µl
10X PCR Buffer 5 µl
dNTP Mix 1 µl
Forward primer (25µM) 1 µl
Reverse primer (25 µM) 1 µl
Agua estéril 49.5 µl
Taq Polimerasa (5U/µl) 0,5 µl

16
Los productos de PCR se separaron mediante electroforesis en gel de agarosa
al 2% con bromuro de etidio y se observaron las bandas en un transiluminador.
3.6. Cuantificación de fragmento de DNA
El estudio se realizó en las instalaciones de Cenit Support Systems
(laboratorio 4) del Edificio Incubadora, Villamayor (Salamanca). El método utilizado
para el análisis de la expresión génica fue una cuantificación basada en el 2-ΔΔCT
,
utilizando como referencia un fragmento de 93 bp para la proteína actina de
Arabidopsis thaliana. Todo el estudio se realizó utilizando cuatro réplicas técnicas
por muestra.
Los primers empleados para la actina fueron:
Forward 5’-3’: GCAGGGATCCACGAGACCACC
Reverse 5’-3’: CCCACCACTGAGCACAATGTTCC

17
4. Resultados y Discusión
4.1. Efecto de distintos elicitores en el crecimiento celular y la
producción de silimarina
4.1.1. Efecto en el crecimiento
Como se observa en la figura 5, el crecimiento de la biomasa celular respecto
a la aplicación de MJa, Cr, Cx y Cx+MJa, elicitados durante 7, 24, 48 y 72 horas, no
mostró diferencias significativas, ya que todos tienden al crecimiento celular de
forma progresiva respecto al tiempo en cultivos celulares de S. marianum.
Figura 5. Efecto de diferentes elicitores en el crecimiento de cultivos en suspensión
de S. marianum
4.1.2. Efecto en la producción de silimarina
4.1.2.1. Efectos del tiempo de contacto
Producción en medios extracelulares
La elicitación de cultivos celulares con distintos compuestos ejerció un
efecto positivo en la producción de silimarina, aumentando su acumulación
extracelular a lo largo del tiempo de contacto (figuras 6,7,8,9).

18
Los tratamientos con Cx y Cx+MJa fueron los más efectivos alcanzándose
valores de más de 0,30 mg por cultivo con la aplicación conjunta de Cx y MJa, lo
que supone un incremento de más de 6 veces comparado con el control.
La elicitación, en todos los casos, aumentó la liberación al medio de cultivo
de los precursores Tx y AC, aunque en este estadio de estudio se ignora si la
formación de silimarina por acoplamiento de los precursores ocurre de forma
extracelular o si la silimarina se ha formado intracelularmente y su salida acompaña
también a la de los precursores.
Figura 6. Acumulación de silimarina y sus precursores Tx y AC en el medio
extracelular de cultivos en suspensión de Silybum marianum elicitados durante 7 h
con MJa, Cr, Cx y Cx+MJa. (C) Control.

19
Figura 7. Acumulación de silimarina y sus precursores Tx y AC en el medio
extracelular de cultivos en suspensión de Silybum marianum elicitados durante 24 h
con MJa, Cr, Cx y Cx+MJa. (C) Control.
Figura 8. Acumulación de silimarina y sus precursores Tx y AC en el medio
extracelular de cultivos en suspensión de Silybum marianum elicitados durante 48 h
con MJa, Cr, Cx y Cx+MJa. (C) Control.

20
Figura 9. Acumulación de silimarina y sus precursores Tx y AC en el medio
extracelular de cultivos en suspensión de Silybum marianum elicitados durante 72
horas con MJa, Cr, Cx y Cx+MJa. (C) Control.
Producción en la biomasa celular
El análisis de precursores y silimarina en la biomasa celular reveló que en
este compartimento de cultivo es donde se acumulan mayoritariamente este tipo de
metabolitos secundarios (mg vs µg). Los distintos tratamientos con elicitores también
provocaron un incremento en la producción, aunque en este caso afectó más a la
producción de precursores que a la de silimarina (figuras 10,11,12,13), dependiendo
la intensidad de la respuesta del tipo de elicitor y del tiempo de contacto. La mayor
producción de Tx y AC se observó en el tratamiento con Cx+MJa a las 72 horas.
La producción de silimarina presenta una variación mínima frente al control
en todos los tratamientos, presentando el mayor aumento en tratamientos con Cx y
Cx+MJa.

21
Figura 10. Producción de silimarina y sus precursores Tx y AC en la biomasa celular
de cultivos en suspensión de Silybum marianum elicitados durante 7 h con MJa, Cr,
Cx y Cx+MJa. (C) Control.
Figura 11. Producción de silimarina y sus precursores Tx y AC en la biomasa celular
de cultivos en suspensión de Silybum marianum elicitados durante 24 h con MJa, Cr,
Cx y Cx+MJa. (C) Control.

22
Figura 12. Producción de silimarina y sus precursores Tx y AC en la biomasa celular
de cultivos en suspensión de Silybum marianum elicitados durante 48 h con MJa, Cr,
Cx y Cx+MJa. (C) Control.
Figura 13. Producción de silimarina y sus precursores Tx y AC en la biomasa celular
de cultivos en suspensión de Silybum marianum elicitados durante 72 h con MJa, Cr,
Cx y Cx+MJa. (C) Control.

23
4.1.2.2. Efecto de distintas dosis de elicitores en la producción de
silimarina
Producción en medios extracelulares
Como se muestra en la figura 14, los cultivos celulares de S. marianum
elicitados con diferentes concentraciones de MJa, Cr y Cx, presentaron variación en
la producción de silimarina, mostrando la mayoría de los tratamientos variaciones
progresivas. En los tratamientos con MJa, se pudo observar claramente un descenso
en la producción a dosis mayores, lo que podría deberse a una posible toxicidad del
compuesto. En los tratamientos con Cx, se apreció un aumento significativo en la
producción de silimarina en el medio extracelular. De igual forma, un aumento en la
concentración de Cr incrementó la producción; con 5 µM de Cr el sistema se satura y
no se observaron modificaciones.
Figura 14. Producción de silimarina y sus precursores Tx y AC en medios
extracelulares de cultivos en suspensión de Silybum marianum elicitados durante 48h
con diferentes concentraciones de MJa, Cr, y Cx. (C) Control.

24
Producción en la biomasa celular
La aplicación de distintas dosis de elicitores no modificó la producción de
silimarina y sus precursores con la misma tendencia que en los medios
extracelulares. Como se observa en la figura 15, MJa aumentó la producción de
precursores al aumentar la dosis; sin embargo, la producción de los componentes de
la silimarina descendió a una concentración de 200 µM. El aumento de la
concentración de Cr aumentó también la producción. Sin embargo, las dos dosis de
Cx probadas en este experimento no indujeron variaciones significativas.
Figura 15. Producción de silimarina y sus precursores Tx y AC en la biomasa de
cultivos en suspensión de Silybum marianum elicitados durante 48 h con diferentes
concentraciones de MJa, Cr, y Cx. (C) Control.
La valoración de la biomasa celular en los experimentos tratados con varias
dosis de elicitores (figura 16) mostró el grado de estrés que ejercieron los distintos
compuestos. En tratamientos con MJa se observó una disminución del peso a
200µM, a diferencia de los tratamientos con coronatina que a dosis superiores a

25
2,5µM no afectaron al crecimiento. Las concentraciones de 30 y 60 mM de Cx
tampoco afectaron al peso de la biomasa celular con respecto al control.
Figura 16. Efecto de diferentes dosis de MJa, Cr, y Cx en el crecimiento celular.
El efecto elicitor de MJa en cultivos de S. marianum evidencia el papel
señalizador que este agente ejerce en el metabolismo secundario vegetal. Estos
resultados se suman a los demostrados para la producción de taxol (Wu y Lin, 2003),
ginsenósidos (Thanh y col., 2005), alcaloides indólicos (Lee y col., 2004) y
silimarina en cultivos celulares de S. marianum derivados de hipocotilo (Sánchez
Sampedro y col., 2005). En este estudio se ha observado que una concentración de
100μM es adecuada para la producción; sin embargo cantidades mayores redujeron
el crecimiento de la biomasa.
Se sabe que Cr es una análogo funcional y estructural del ácido jasmónico y
compuestos señalizadores relacionados, como el MJA o el precursor de ambos ácido
12-oxo-fitodienoico (Weiler, 1994). El efecto elicitor de Cr en cultivos fue similar al
de MJA, lo que apoya las observaciones anteriores. Desde el punto de vista
biotecnológico, su menor precio y la menor concentración requerida, hace de la
toxina un agente elicitor más eficaz que MJa.
Los valores de producción de silimarina en tratamientos con Cx a diferentes
tiempos de elicitación fueron más elevados que con MJa, mencionando además que

26
la viabilidad celular no difirió de la de los controles durante el período de
experimentación. Los resultados fueron aun mejores en el tratamiento combinado
(Cx+MJa), lo que está de acuerdo con lo observado por Almagro y col. (2010), para
la producción del alcaloide ajmalicina en cultivos de Catharanthus roseus y para la
producción de resveratrol en cultivos de vid (Pedreño y col., 2009).
4.1.2.3. Efectos de la adición de precursores a cultivos elicitados
Los resultados anteriores muestran que los elicitores afectan tanto a la
producción de silimarina como a la de sus precursores. Con el fin de determinar si la
producción está limitada por al biosíntesis de Tx o AC y si la elicitación altera la
disponibilidad de los precursores para su incorporación en silimarina, a cultivos
control o tratados con MJa, Cx o Cx+MJa durante 24 h se les añadió 3 mM de Tx y
3mM de CA, sólos o en combinación, valorándose la producción extracelular de
silimarina a las 24 h de tratamiento.
Al analizar la producción en términos cuantitativos, se observó que tanto en
cultivos control como en los elicitados con MJa, la incorporación de Tx en los
cultivos estimuló la acumulación de silimarina; sin embargo, la aplicación de CA de
forma aislada la redujo a valores inferiores a la de los cultivos sin adicionar, lo que
indica que este compuesto, a la dosis suministrada de 3 mM, es tóxico para el
metabolismo (figuras 17 y 18). La aplicación conjunta mejoró el resultado, pero
nunca se alcanzaron valores superiores a los obtenidos en presencia de Tx sóla.
La adición de precursores, sin embargo, no mejoró la formación de producto
en cultivos tratados con Cx y aumentó escasamente con Cx+MJa (figuras 19 y 20).
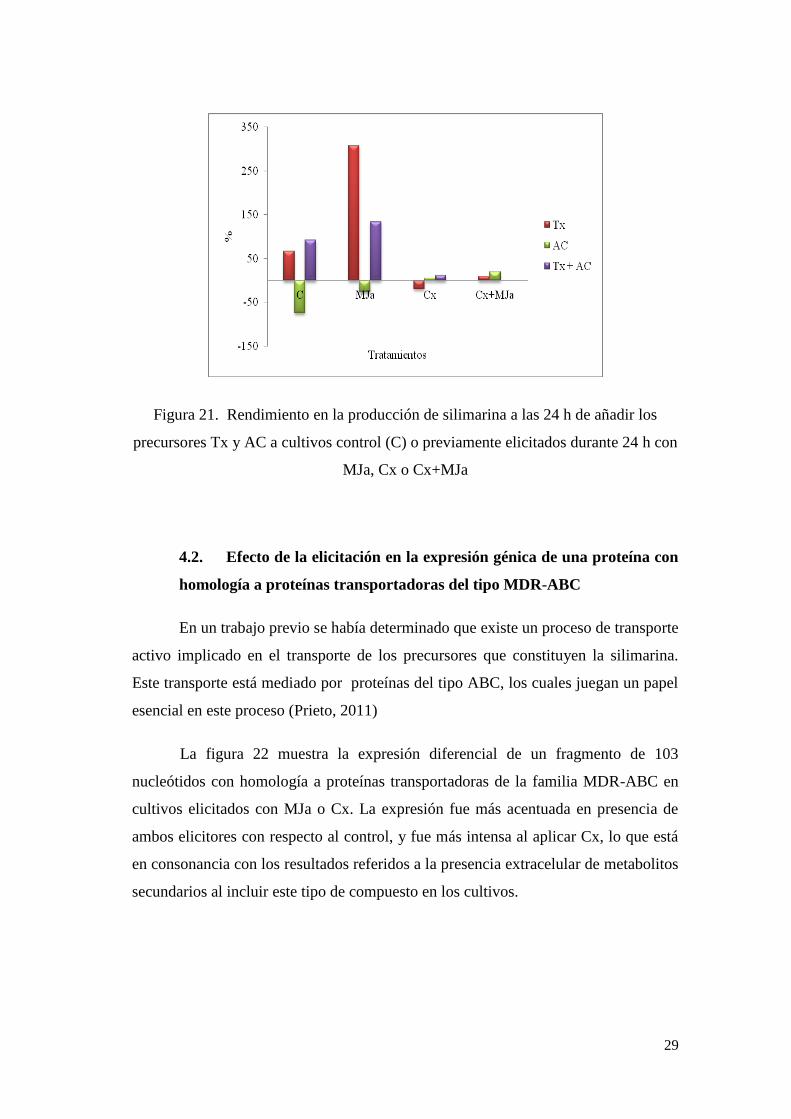
En la figura 21 se expresa el rendimiento en la producción obtenido al incorporar
precursores. De estos resultados se desprende que tanto en cultivos control como en
los elicitados con MJa, Tx parece ser limitante. El efecto tóxico que ejerce AC de
forma aislada en las células se atenúa en presencia de Tx debido a su secuestro por
el acoplamiento entre ambos compuestos. En presencia de Cx, la capacidad del
sistema para formar silimarina parece estar saturada, y la adición de cualquiera de los
precursores es inefectiva. La presencia de Cx además reduce el efecto tóxico de AC
debido a la propia acción que como aductos ejercen estos compuestos; es decir, al

27
tener una estructura cónica con superficie hidrofílica e interior lipofílico, la cavidad
presenta una hidrofobicidad comparable a la de una solución etanólica, permitiendo
así formar cuerpos de inclusión con AC y proteger a las células de su toxicidad.
Figura 17. Efecto de la adición de precursores en la producción de silimarina en
cultivos celulares de Silybum marianum. La producción en el medio extracelular se
evaluó a las 24 h de añadir a cultivos control 3 mM de Tx, 3 mM de AC, o 3+3 mM
de Tx+AC .
Figura 18. Efecto de la adición de precursores en la producción de silimarina en
cultivos celulares de Silybum marianum elicitados con MJa. La producción en el
medio extracelular se evaluó a las 24 h de añadir a cultivos previamente elicitados
con MJa durante 24 h, 3 mM de Tx, 3 mM de AC o 3+ 3 mM de Tx+AC.

28
Figura 19. Efecto de la adición de precursores en la producción de silimarina en
cultivos celulares de Silybum marianum elicitados con Cx. La producción en el
medio extracelular se evaluó a las 24 h de añadir a cultivos previamente elicitados
con Cx durante 24 h, 3 mM de Tx, 3 mM de AC o 3+3 mM de Tx+AC.
Figura 20. Efecto de la adición de precursores en la producción de silimarina en
cultivos celulares de Silybum marianum elicitados con la combinación de Cx y MJa.
La producción en el medio extracelular se evaluó a las 24 h de añadir a cultivos
previamente elicitados con Cx+MJa durante 24 h, 3 mM de Tx, 3 mM de AC o
3+3mM de Tx+AC.
29
Figura 21. Rendimiento en la producción de silimarina a las 24 h de añadir los
precursores Tx y AC a cultivos control (C) o previamente elicitados durante 24 h con
MJa, Cx o Cx+MJa
4.2. Efecto de la elicitación en la expresión génica de una proteína con
homología a proteínas transportadoras del tipo MDR-ABC
En un trabajo previo se había determinado que existe un proceso de transporte
activo implicado en el transporte de los precursores que constituyen la silimarina.
Este transporte está mediado por proteínas del tipo ABC, los cuales juegan un papel
esencial en este proceso (Prieto, 2011)
La figura 22 muestra la expresión diferencial de un fragmento de 103
nucleótidos con homología a proteínas transportadoras de la familia MDR-ABC en
cultivos elicitados con MJa o Cx. La expresión fue más acentuada en presencia de
ambos elicitores con respecto al control, y fue más intensa al aplicar Cx, lo que está
en consonancia con los resultados referidos a la presencia extracelular de metabolitos
secundarios al incluir este tipo de compuesto en los cultivos.

30
Figura 22. Expresión génica de proteína transportadora ABC, en cultivos elicitados
con Cx y MJa.

31
5. Conclusiones
1. La aplicación de elicitores a cultivos celulares de Silybum marianum es una
estrategia efectiva para la producción de silimarina.
2. Los análogos estructurales jasmonato de metilo y coronatina incrementan la
producción de metabolitos de forma similar, lo que indica la analogía
funcional de ambos tipos de compuestos.
3. La aplicación de β-ciclodextrinas, sólas o en combinación con MJa constituye
la mejor estrategia de elicitación, con la ventaja de que se produce una gran
acumulación de silimarina en el medio de cultivo, facilitándose así la
recuperación del producto sin dañar la biomasa celular.
4. La disponibilidad de precursores constituye un factor limitante en los cultivos
crecidos en condiciones control; no se descarta, sin embargo, una limitación
en la producción de silimarina causada por un efecto tóxico del precursor
alcohol coniferílico.
5. La elicitación, tanto con MJa como con Cx, va asociada a un aumento en la
expresión génica de un fragmento con homología a proteínas transportadoras
del tipo MDR-ABC. Se desconoce si esa proteína participa en el proceso de
liberación de silimarina o sus precursores al medio de cultivo.

32
6. Agradecimientos
La realización del presente trabajo fin de Máster es fruto de orientaciones,
sugerencias y estímulos de la Dra. Purificación Corchete Sánchez, quien a lo largo de
estos meses me ha conducido de la forma más sencilla y generosa, mostrando a cada
momento su disposición ante las dudas, contribuyendo importantes observaciones
que siempre direccionaron esta investigación, por lo cual expreso mis más sinceros
agradecimientos y estima hacia ella.
De forma general agradezco a todos los profesores del Máster de
Agrobiotecnología de la Universidad de Salamanca, por compartir sus conocimientos
aplicando siempre la exigencia con el único objetivo de formar excelentes
investigadores. Y en una forma muy especial a la Dra. Berta Dopico que siempre
estuvo presta a colaborar con los trámites requeridos considerando mi situación de
extranjera.
Deseo agradecer la oportunidad que la SENESCYT - Ecuador brindó a través
de la beca obtenida en la “Convocatoria abierta 2011” para realizar mis estudios de
postgrado, además agradecer el apoyo constante de los miembros de la UPSE –
Ecuador, que siempre estuvieron pendientes de mis logros profesionales.
Por supuesto a mis padres que son pilar fundamental en mi vida, cada logro
está dedicado a ellos y son quienes guían mi camino, a mis hermanos y mi esposo
quienes han estado pendiente de mis pasos y a mis amig@s que a pesar de la
distancia estuvieron presentes con sus mensajes positivos.
Y por último, pero no menos importante, a Dios, que siempre me ha
permitido iniciar nuevos retos, capacidad para tomar las decisiones correctas y
fuerzas para no decaer ante los problemas.

33
7. Bibliografía
Arias M., Angarita M., Aguirre A., Restrepo J., Montoya C., 2009.
Estrategias para incrementar la producción de metabolitos secundarios en cultivos de
celulas vegetales. Revista Facultad Nacional de Agronomía, Medellín. 62(1): 4881-
4895 p.
Arias M., Aguirre A., Angarita M., Montoya C., Restrepo J., 2009. Aspectos
ingenieriles del cultivo in vitro de células vegetales para la producción de
metabolitos secundarios. Dyna. 157: 109-121 p.
Azcón-Bieto, J., Talón, M., 2000. Fundamentos de fisiología vegetal.
Interamericana Mc Graw-Hill. 555 p.
Bougard F, Gravot A, Milesi S, Gontier E., 2001. Production of secondary
metabolites: a historical perspective. Plant Science. 161(5): 839-85 p.
Bru R., Selles S., Casado-Vela J., Belchi-Navarro S., Pedreno M., 2006.
Modified cyclodextrins are chemically defined glucan inducers of defence responces
in grapevine cell cultures. J. Agric. Food Chem. 54, 65-71 p.
Chattopadhyay S, Srivastava A, Bhojwani S, Bisaria V., 2002. Production of
podophyllotoxin by plant cell cultures of Podophyllum hexandrum in bioreactor.
Journal of Bioscience and Bioengineering. 93(2):215-220 p.
Conceicao L., Ferreres F, Tavares R., Dias A., 2006. Induction of phenolic
compounds in Hypericum perforatum L. cells by Colletotrichum gloeosporioides
elicitation. Phytochemistry 67(2): 149-155 p.
Corchete P., 2008. Silybum marianum (L.) gaertn: The source of silymarin.
Bioactive Molecules and Medicinal Plants. 2008:123-148.
Dong H., Zhong J., 2002. Enhanced taxane productivity in bioreactor
cultivation of Taxus chinensis cells by combining elicitation sucrose feeding and
ethylene incorporation. Enzyme and Microbial Technology 31(1): 116-121 p.

34
Edahiro J., Nakamura M., Seki M., Furusaki S., 2005. Enhanced
accumulation of anthocyanin in cultured strawberry cells by repetitive feeding of L-
Phenylalanine into the medium. Journal of Biosciences and Bioengineering 99(1):
43-47p.
Edahiro J., Ski M., 2006. Phenylpropanoid metabolite supports cell aggregate
formation in strawberry cell suspension culture. Journal of Bioscience and
Bioengineering. 102(1):8-13 p.
Gala R., Mita G., Caretto S., 2005. Improving a-tocopherol production in
plant cell cultures. Journal of Plant Physiology 162(7): 782-784 p.
Ghasempour H., Anderson E., Gianello R., Gaff D., 1998. Growth inhibitor
effects on protoplasmic drught tolerance and protein synthesis in leaf cells of the
resurrection grass Sporobolus stapfianus. In: Plant growth regulation. Kluwer
Academic Publishers, Netherlands. p. 179 Poulton, J.E. 1990. Cyanogenesis in
plants. Plant Physiol. 94:401 p.
Gabriela C., Dean K., Wiley D., 1997. Phytoestrogens and floral development
indioecious Maclura pomifera (Raf.) Schoneid. and Morus rubra (L.) (Moraceae).
Plant Science. 130:27 p.
García D., 2004. Los metabolitos secundarios de las especies vegetales.
Pastos y Forrajes. 27(1):1-12 p.
Harborne J., 1993. Introduction to ecological biochemistry. Academic Press,
USA. 232p.
Honda H, Hiraoda K, Nagamori E, Omote M, Kato Y, y col., 2002. Enhanced
anthocyanin production from grape callus in air lift type bioreactor using viscous
additive supplemented medium. Journal of Bioscience and Bioengineering. 94
(2):135-139 p.
Hoopen H., Vinke J., Moreno P., Verpoorte R., Heijnen J., 2002. Influence of
temperature on growth and ajmalicine production by Catharanthus roseus
suspension cultures. Enzyme and Microbial Technology 30(1): 56-65 p.

35
Ignatov A., Clark G., Cline S., Psenak M., Krueger R., Coscia C., 1999.
Elicitation of dihydrobenzophenanthridine oxidase in Sanguinaria canadensis cell
cultures. Phytochemistry 43(6): 1141-1144 p.
Karban R., Baldwin I., Baxter K., Laue G., 2000. Communication between
plants: induced resistance in wild tobacco plants following clipping of neighboring
sagebrush. Oecologia 125, 66-7 p.
Kuromori T., Shinozaki K., 2010. ABA transport factors found in
Arabidopsis ABC transporters. Plant Signaling & Behavior 5, 1124–1126 p.
Lee C., Ertürk S., Tengtrakool J., 2004. Enhancement of ajmalicine
production in Catharanthus roseus cell cultures with methyl jasmonate is dependent
on timing and dosage of elicitation. Biotechnol. Lett. 0, 1595–1599 p.
Liu C., Miao Y., Zhang, K., 2011. Sequestration and transport of lignin
monomeric precursors. Molecules 16, 710–727 p.
Madrid E., Corchete P., 2010. Silymarin secretion and its elicitation by
methyl jasmonate in cell cultures of Silybum marianum is mediated by
phospholipase D-phosphatidic acid. Journal of Experimental Botany 61(3), 747-
754p.
Martínez G., Gómez M., 2007. Ciclodextrinas: complejos de inclusión con
polímeros. Revista Iberoamericana de Polímeros. Vol 8:4. 301-313 p.
Murashige T., Skoog F., 1962. A revised medium for rapid growth and
bioassays with tobacco tissue culture. Physiologia Plantarum 15, 473-497 p.
Navarro S., Pedreño M., Corchete P., 2011. Methyl jasmonate increases
silymarin production in Silybum marianum (L.) Gaernt cell cultures treated with β-
cyclodextrins. Biotechnology Letters 33, 179–184 p.
Namdeo, 2007. Plant cell elicitation for production of secondary metabolites:
A review. 1, 1:69-79 p.

36
Pedreño M., Belchí S., Almagro L., Bru R., 2009. Uso combinado de
jasmonato de metilo y ciclodextrinas para la producción de resveratrol.
Prakash G., Srivastava A., 2005. Statistical media optimization for cell
growth and azadirachtin production in Azadirachta indica (A. Jyss) suspension
cultures. Process Biochemistry. 40(12): 3795-3800.
Prakash G, Srivastava A., 2006. Modeling of azadirachtin production by
Azadirachta indica and its use for feed forward optimization studies. Biochemical
Engineering Journal. 29(1-2):62-68 p.
Prieto D., Corchete P., 2011. Estudio de mecanismos de señalización en la
secreción de flavonolignanos en cultivos de Silybum marianum elicitados con
ciclodextrinas.
Qu J., Zhang W., Jin M., Yu. X., 2006. Effect of homogeneity on cell growth
and anthocyanin biosynthesis in suspension cultures of Vitis vinifera. Chinese
Journal of Biotechnology 22(5): 805-810 p.
Ramos G., Frutos P., Giráldes F., Mantecón A., 1998. Los compuestos
secundarios de las plantas en la nutrición de los herbívoros. Arch. Zootec. 47
(180):59p.
Raskin I., 1992. Role of salicylic acid in plants.Annu. Rev. Plant Physiol.
Plant Mol. Biol. 43:439 p.
Sajc L., Grubisic D., Vunjaknovakovic G., 2000. Bioreactors for plant
engineering: an outlook for further research. Biochemical Engineering Journal.
4(2):89-99 p.
Sánchez S., Fernandez T., Corchete P., 2005. Yeast extract and methyl
jasmonate-induced silymarin production in cell cultures of Silybum marianum (L.)
Gaertn. Journal of Biotechnology 119(1): 60–69 p.
Sánchez M., Fernández J., Corchete P., 2005. Some common signal
transduction events are not necessary for the elicitor-induced accumulation of

37
silymarin in cell cultures of Silybum marianum. Journal of Plant Physiology
165(14), 1466-1473 p.
Schlatmann J., Hoopen H., Heijnen J., 1996. Large-scale production of
secondary metabolites by plant cell cultures. En: DiCosmo F, Misawa M, Editores.
Plant cell culture secondary metabolism: toward industrial application. New Cork:
CRS Press. 11-52 p.
Sepúlveda G., Porta. H, Rocha M., 2004. La participación de los metabolitos
secundarios en la defensa de las plantas. Rev Mex Fitopatol 21: 355–363 p.
Sirikantaramas S., Yamazaki M., Saito K., 2008. Mechanisms of resistance to
self-produced toxic secondary metabolites in plants. Phytochemical Review 7, 467–
477p.
Stafford H., 1997. Roles of flavonoids in symbiotic and defense functions in
legumes roots. The Botanical Review. 63 (1):27 p.
Stransburger E., Noll F., Schenk H., Schimper A., 1994. Tratado de Botánica.
Marín. Barcelona, España. 520 p.
Thanh N., Murthy H., Yu K., Hahn E. y Paek K., 2005. Methyl jasmonate
elicitation enhanced synthesis of ginsenoside by cell suspension cultures of Panax
ginseng in 5-l balloon type bubble bioreactors. Applied Microbial and Biotechnology
67(2): 197–201 p.
Trejo T., Rodriguez M., 2007. La agregación celular en la producción de
metabolitos secundarios en cultivos vegetales in-vitro. Interciencia.32(10):669-674 p.
Valdés R., Balbín M., 2000. Curso de fisiología y bioquímica vegetal.
UNAH. La Habana, Cuba. 89 p.
Vasconsuelo A., Boland R., 2007. Molecular aspects of the early stages of
elicitation of secondary metabolites in plants. Plant Science 172(5):861-875 p.

38
Verpoorte R., Contin A., Memelink J., 2002. Biotechnology for the
production of plant secondary metabolites. Phytochemistry Reviews Vol. 1, No. 1,
13–25 p.
Wang Z., Zhong J., 2002. Combination of conditioned medium and elicitation
enhances taxoid production in bioreactor cultures of Taxus chinensis cells.
Biochemical Engineering Journal 12(2): 93–97 p.
Wei Z., Chris F., Chris C., Simon C., 2004. To Stretch the Boundary of
SecondaryMetabolite Production in Plant Cell-Based Bioprocessing: Anthocyanin as
a Case Study. Journal of Biomedicine and Biotechnology 5: 264–271 p.
Weiler E., Kutchan T., Gorba T., Brodschelm W., Niesel U., Bublitz F., 1994.
The Pseudomonas phytotoxin coronatine mimics octadecanoid signalling molecules
of higher plants. FEBS Lett. 345, 9–13 p.
Wildi E., Wink M., 2002. Biotechnology potential of hairy root culture, in
Recent Progress in Medicinal plants. Biotechnology and Genetic Engineering. Sci
Tech Pub., USA. Vol 4.441–54 p.
Wu J., Wang C., Mei X., 2001. Stimulation of taxol production and excretion
in Taxus spp cell cultures by rare earth chemical lanthanum. Journal of
Biotechnology 85(1): 67-73 p.
Wu J., Lin L., 2003. Enhancement of taxol production and release in Taxus
chinensis cell cultures by ultrasound, methyl jasmonate and in situ solvent extraction.
Applied Microbiology and Biotechnology. Vol. 62. 2-3: 151-155 p.
Yeoman M., Yeoman C., 1996. Manipulating secondary metabolism in
cultured plant cells. New Phytologist 134(4): 553-569 p.
Yu K., Murthy H., Jeong C., Hahn E., Paek K., 2005. Organic germanium
stimulates the growth of ginseng adventitious roots and ginsenoside production.
Process Biochemistry 40(9): 2959-2961 p.

39
Zamboni A., Gatto, P., Cestaro A., Pilati S., Viola R., Mattivi F., Moser C. y
Velasco R., 2009. Grapevine cell early activation of specific responses to DIMEB, a
resveratrol elicitor. BMC Genomics. Vol 10:363: 1-13 p.
Zhang W., Curtin C., Franco C., 2002. Towards manipulation of post-
biosynthetic events in secondary metabolism of plant cell cultures. Enzyme and
Microbial Technology 30, 688–696 p.
Zhao Z., Xu Y., Qian Z., Tian W., Qian X., Zhong J., 2004. Novel fluoro-and
hydroxyl-containing jasmonate derivatives as highly efficient elicitors in suspension
cultures of Taxus chinensis. Bioorganic & Medicinal Chemistry Letters 14(18):
4755–4758 p.
Zhao J., Davis L., Verpoorte R., 2005. Elicitor signal transduction leading to
production of plant secondary metabolites. Biotechnology Advances 23(4): 283–
333p.
Zhao D., Fu C., Han Y., Lu D., 2005. Effects of elicitation of jaceosidin and
hispidulin production in cell suspension cultures of Saussurea medusa. Process
Biochemistry; 40(2): 739-745 p.