tyrosine phosphorylation: from discovery to the kinome …super7/30011-31001/30941.pdf · tyrosine...
TRANSCRIPT

The 2007 Herbert Tabor -Journal of Biological Chemistry Lecture
Tony Hunter
Tyrosine phosphorylation: from discovery to the kinome and beyond
ASBMB Annual Meeting
April 28, 2007


So far 43 JBC papers and one submitted!


The History of Protein Phosphorylation
Protein kinase
Protein P.Protein
ATP ADP
P
Protein phosphatase

The History of Protein Phosphorylation
Phospho-proteindiscovery1906
Phospho-serine inproteins1932
Phospho-tyrosinesynthesis1933
Protein kinaseactivity1954
Phosph-orylasekinase1959
cAMPdepen-dent PK1968
Srctyrosine kinase1979
Phospho-tyrosinein fly eggs1964
Gleevecapprovedfor CML2001
1910 1920 1930 1940 1950 1970 1980 1990 200019601900

The History of Protein Phosphorylation
Phospho-proteindiscovery1906
Phospho-serine inproteins1932
Phospho-tyrosinesynthesis1933
Protein kinaseactivity1954
Phosph-orylasekinase1959
cAMPdepen-dent PK1968
Srctyrosine kinase1979
Phospho-tyrosinein fly eggs1964
Gleevecapprovedfor CML2001
1910 1920 1930 1940 1950 1970 1980 1990 200019601900
J. Biol. Chem. 2:127 (1906)

The History of Protein Phosphorylation
Phospho-proteindiscovery1906
Phospho-serine inproteins1932
Phospho-tyrosinesynthesis1933
Protein kinaseactivity1954
Phosph-orylasekinase1959
cAMPdepen-dent PK1968
Srctyrosine kinase1979
Phospho-tyrosinein fly eggs1964
Gleevecapprovedfor CML2001
1910 1920 1930 1940 1950 1970 1980 1990 200019601900
J. Biol. Chem. 98:109 (1932)

The History of Protein Phosphorylation
Phospho-proteindiscovery1906
Phospho-serine inproteins1932
Phospho-tyrosinesynthesis1933
Protein kinaseactivity1954
Phosph-orylasekinase1959
cAMPdepen-dent PK1968
Srctyrosine kinase1979
Phospho-tyrosinein fly eggs1964
Gleevecapprovedfor CML2001
1910 1920 1930 1940 1950 1970 1980 1990 200019601900
J. Biol. Chem. 100:583 (1933)

The History of Protein Phosphorylation
Phospho-proteindiscovery1906
Phospho-serine inproteins1932
Phospho-tyrosinesynthesis1933
Protein kinaseactivity1954
Phosph-orylasekinase1959
cAMPdepen-dent PK1968
Srctyrosine kinase1979
Phospho-tyrosinein fly eggs1964
Gleevecapprovedfor CML2001
1910 1920 1930 1940 1950 1970 1980 1990 200019601900
J. Biol. Chem. 211:969 (1954)

The History of Protein Phosphorylation
Phospho-proteindiscovery1906
Phospho-serine inproteins1932
Phospho-tyrosinesynthesis1933
Protein kinaseactivity1954
Phosph-orylasekinase1959
cAMPdepen-dent PK1968
Srctyrosine kinase1979
Phospho-tyrosinein fly eggs1964
Gleevecapprovedfor CML2001
1910 1920 1930 1940 1950 1970 1980 1990 200019601900
Burnett and Kennedy, J. Biol. Chem. 211:969 (1954)

The History of Protein Phosphorylation
Phospho-proteindiscovery1906
Phospho-serine inproteins1932
Phospho-tyrosinesynthesis1933
Protein kinaseactivity1954
Phosph-orylasekinase1959
cAMPdepen-dent PK1968
Srctyrosine kinase1979
Phospho-tyrosinein fly eggs1964
Gleevecapprovedfor CML2001
1910 1920 1930 1940 1950 1970 1980 1990 200019601900

Historic moments in the discovery of phosphotyrosine
P.SER
P.THR
Pi+
_
|mTacid
|mTprotease
|IgH/Src
P.TYR
Pi
14/6/79
18/9/79
“X”
|mTacid
|LTin vivo
P.SER
P.THR
pH “1.9”

v-Src increases P.Tyr levels in transformed chick cells
uninfected v-Src-transformed
+
pH 1.9
Hunter and Sefton, PNAS 77:1311 (1980)
+32P-labeled control and RSV-transformed chick fibroblasts
pH 3.5

The History of Protein Phosphorylation
Phospho-proteindiscovery1906
Phospho-serine inproteins1932
Phospho-tyrosinesynthesis1933
Protein kinaseactivity1954
Phosph-orylasekinase1959
cAMPdepen-dent PK1968
Srctyrosine kinase1979
Phospho-tyrosinein fly eggs1964
Gleevecapprovedfor CML2001
1910 1920 1930 1940 1950 1970 1980 1990 200019601900

The History of Protein Phosphorylation
Phospho-proteindiscovery1906
Phospho-serine inproteins1932
Phospho-tyrosinesynthesis1933
Protein kinase activity1954
Phosph-orylasekinase1959
cAMPdepen-dent PK1968
Srctyrosine kinase1979
Phospho-tyrosinein fly eggs1964
Gleevecapprovedfor CML2001
1910 1920 1930 1940 1950 1970 1980 1990 200019601900
More than 29,000 papers on tyrosine kinases have been published since 1979!

How many tyrosine kinases are there?1. The finding that v-Src and c-Src phosphorylate tyrosine
gave us the first tyrosine kinase in 1979
2. By the end of 1980 four tyrosine kinases were known (Src, Abl, EGF receptor, Fps/Fes)
3. By the end of 1990 over 50 tyrosine kinases had been identified in vertebrates and equal numbers of tyrosine kinases and serine kinases were known, leading to theprediction that there might be several 100 tyrosine kinases in a vertebrate genome and a total of over 1000 protein kinases
4. The complete human genome sequence reported in 2001 reveals that there are 90 tyrosine kinases (all the tyrosine kinases had been found by other means before the sequence was completed), out of a total of 518 protein kinases

The History of Tyrosine Phosphorylation
c-Srcgene
discovery1976
v-Erb isderived
from EGFR1984
Abl + EGFR
are PTKs1980
PTP1Bis first
PTPase1988
Bcr-Abl fusion
in CML1985
Cdc2 isinhibited by P.Tyr
1989
PTP1Band IRKstructures
1994
Gleevecapprovedfor CML
2001
MT + Src tyrosine kinase1979
SH2domain
binds P.Tyr1990
YersiniaencodesPTPase1990
PI3 kinaseassociateswith MT
1987
SH2domain
structure1992
PLCγas RTKsubstrate
1989
STATsare PTK
substrates1992
Inactivec-Src
structure1997
PTBdomain
binds pY1995
Humankinome has
90 PTKs2002
SH2domain
identified1986
v-Src ∆C activation
mechanism1987
Grb2SH2/SH3adaptor1992
v-Srckinase
sequence1980
v-Src protein +
kinase1977/8
1975 1980 1990 20001985 1995 2005
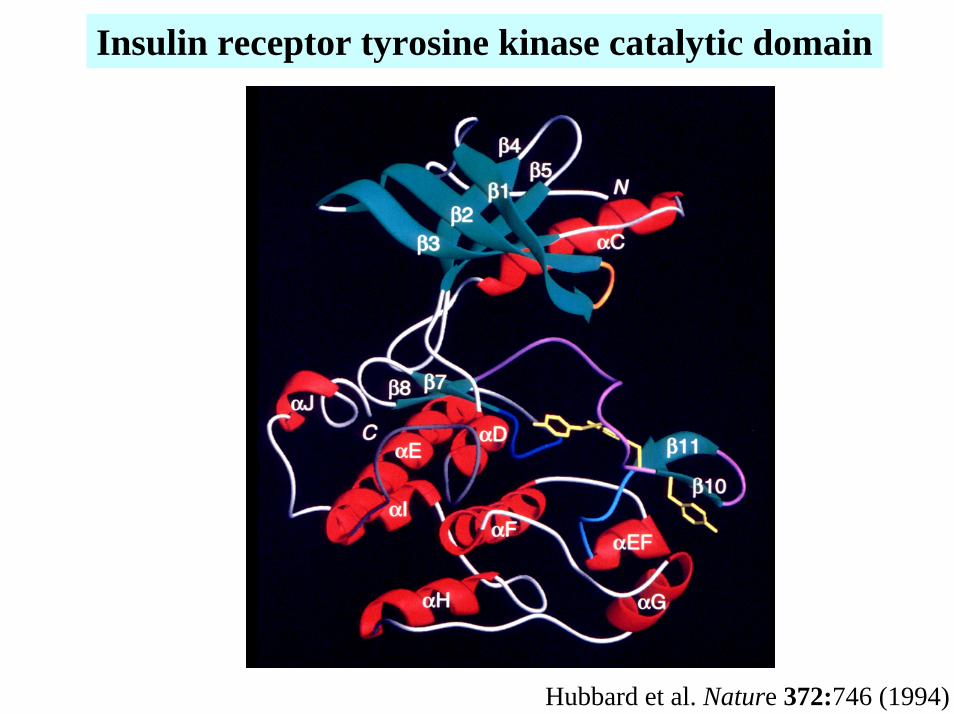
Insulin receptor tyrosine kinase catalytic domain
Hubbard et al. Nature 372:746 (1994)

Receptor tyrosine kinases
F59F3.1F59F3.5F40G9.13
EGL-15DAF-2 CAM-1C16B8.1
VAB-1
F11D5.3C25F6.4
C16D9.2KIN15KIN16TKR-1C08H9.8F59F5.3M01B2.1R09D1.12R09D1.13
T01G5.1T17A3.8W04G5.6NW04G5.6CY50D4B-4ZK938.5B0198.3F54F7.5
LET-23
CCK4FLT1FLK1FLT4
PDGFRαPDGFRβCSF1RKITFLK2
FGFR1FGFR2FGFR3FGFR4
IGF1RINSR
IRR
EGFRHER2HER3HER4
METRON
TRKATRKBTRKC
AXLMERSKY
TIETEK
EphA1EphA2EphA3EphA4EphA5EphA6EphA7EphA8EphB1EphB2EphB3EphB4EphB5EphB6
RYK ROSRET LTKALK
DDR1DDR2
RTK106ROR1ROR2
MuSK
LMR1LMR2LMR3
?
Human RTKs58 (20 classes)
Worm RTKs29 (11 classes)11 Unclassified
T10H9.2
(B0252.1, F11E6.8, F40A3.5, R151.4, T148.1, T22B11.3Y38H6C.20, C24G6.2A, F08F1.1, F09A5.2, F09G2.1)

Y DNA BD Actin BDAbl
YFes/Fe r
YKinase-like domainJak
YSyk/Zap70
YPH domain
Tec
Y Cdc42-bindingAck
YIntegrin-binding/JEF domain Focal adhesion-binding
Fak
YYSrc
CskY
Srm
Y YRak/Fr k
YYBrk/Si k
Myr
(Myr)
SH3 domain
SH2 domain
PTK catalytic domain
Nonreceptor protein-tyrosine kinase s
JEF domai n
SH3 binding regio n
Nonreceptor tyrosine kinases

What is tyrosine phosphorylation used for?
1. Growth factor signaling (and oncogenesis)
2. Cell adhesion, spreading, migration and shape
3. Cell differentiation in development
4. Cell cycle control
5. Gene regulation and transcription
6. Endocytosis and exocytosis
7. Insulin stimulation of glucose uptake
8. Angiogenesis (formation of new blood vessels)
9. Regulation of ion channels in nerve transmission

Transmembrane receptorprotein-tyrosine kinase
Plasma membrane
Ligand binding domain
Bimolecular receptor protein-tyrosine kinase
Out
In
Catalytic domain
NH 2 NH 2
NH 2
EGF receptor PDGF receptor
Interferon receptor T cell receptor
Transmembrane signaling by tyrosine phosphorylationTransmembrane signaling by tyrosine phosphorylation

Schlessinger, Cell 103:211 (2000)
Receptor tyrosine kinase (RTK) signaling through SH2 and PTB P.Tyr-binding domain proteins

Manual alignment - March 1985 (BB)
Stone age bioinformatics!
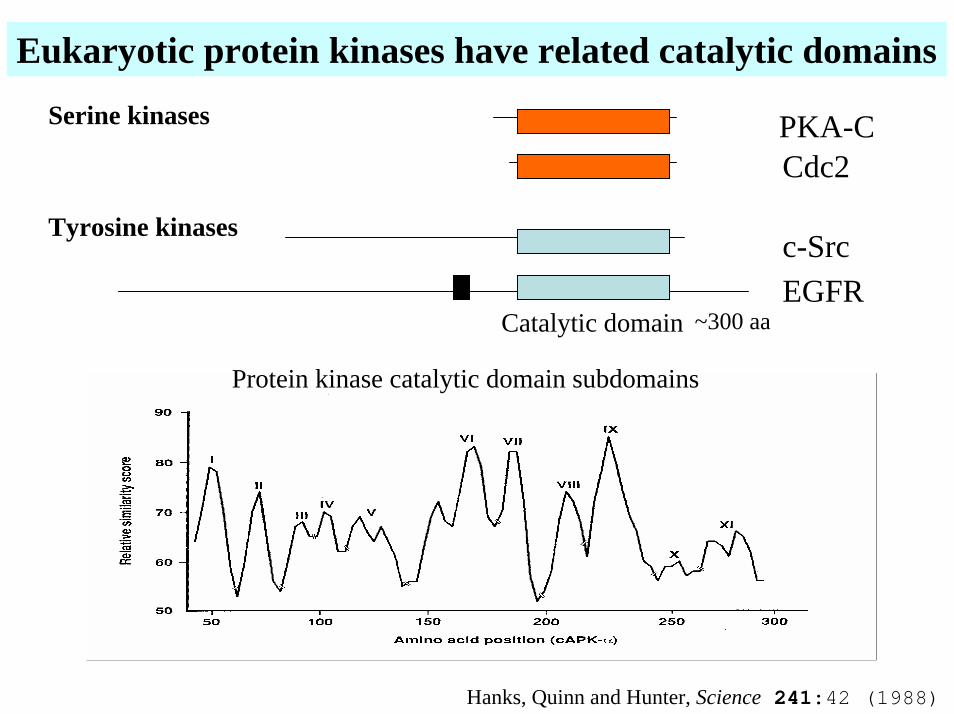
Eukaryotic protein kinases have related catalytic domains
Serine kinases PKA-CCdc2
Tyrosine kinasesc-SrcEGFR
Catalytic domain ~300 aa
Protein kinase catalytic domain subdomains
Hanks, Quinn and Hunter, Science 241:42 (1988)

Structure of PKA catalytic subunit bound to PKI (5-24) and ATP
pT197
N-lobe
C-lobe
catalyticcleft
Knighton et al. Science 253:414 (1991)

Hunter, Cell 50:823 (1987)
The birth of the kinome: a thousand and one protein kinases
Phos K PKA
Src

The first kinome
Hunter and Plowman, TiBS 22:18 (1997)

The first kinome tree
Hunter and Plowman, TiBS 22:18 (1997)

How many protein kinases are there?• S. cerevisiae (6217 genes) 130 (116) protein kinases (2.1%) but no
bona fide tyrosine kinases
• S. pombe (4624 genes) 128 (114) protein kinases (2.8%) but no TKs
• C. elegans (19100 genes) 454 (434) protein kinases (2.4%) including 90 tyrosine kinases (20%)
• D. melanogaster (13600 genes) 239 (223) protein kinases (1.8%) including 32 tyrosine kinases (14%)
• H. sapiens (23,000 genes) 518 (478) protein kinases (2.2%) including90 tyrosine kinases (16%) (chimpanzee kinomeis essentially identical)
• A. thaliana (26,800 genes) 1055 protein kinases (3.8%) but no tyrosine kinases (>630 RLK)
M. brevicolli (unicellular choanoflagellate) has bona fide tyrosine kinases, SH2 domains and protein-tyrosine phosphatases (PTPs); the yeasts have PTPs, but no tyrosine kinases or SH2 domains
~2% of all genes in eukaryotes encode protein kinases

• Tetrahymena (27424 genes) 1069 PKs (3.8%) - no true TKs, but has TKLs and 83 two-component HisK. Has 630 PKs not assignable to known families or subfamilies, with 37 novel classes and 100s of unique PKs
• Dictyostelium (12500 genes) 285 PKs (2.3%) 246 ePKs, includingTKLs but no true TKs, plus 26 aPKs and 14 HisKs
• Sea urchin (24000 genes) 353 PKs (1.5%) 329 ePKs, 24 aPKs (Strongylocentrotus) and 53 TKs, but no HisK. Lacks only
4/187 human kinase families
• Mouse kinome is ~99% identical to human kinome - 540 mouse proteinkinase genes - 510 are orthologous to human protein kinases
• One conclusion is that the tyrosine kinase-like kinases (TKLs) evolved in unicellular organisms, perhaps serving as tyrosine (TK) progenitors, and were secondarily eliminated from yeast. HisKs were lost during evolution of metazoans, apparently replaced by TKLs and TKs
Some more recent kinomes
(Eisen et al. PLoS Biol 4:e286; Goldberg et al. PLoS Genet 2:e38; Bradham et al. Dev Biol 300:180)

Vertebrates
Sea urchin
Drosophila
+4
-11
+15
+9
53
C. elegans+15 / -7
+3
S. cerevisiae
Dictyostelium
Tetrahymena
+6 / -28
+76/-3
+25/-4
+6/-1
+37
Unikon
t
Metazo
an
Opistho
kont
+27
Coelom
ateDeu
teros
tome
80
85
158
173
182
The evolution of the kinome
Gerard Manning
TKLs
TKs

How many human protein kinases and phosphatases?Protein kinases (the kinome)518 protein kinases including: (not quite a 1001!) 478 conventional protein kinases (ePKs)
(16 have tandem catalytic domains)388 protein-serine/threonine kinases 90 protein-tyrosine kinases58 receptor protein-tyrosine kinases32 non-receptor protein-tyrosine kinases
(~50 may lack catalytic activity; ~106 pseudogenes) 40 atypical protein kinases in 7 families (e.g. alpha kinases)
Manning et al. Science 208:1912 (2002) (http://www.kinase.com)
Protein phosphatases (the phosphatome)

Nature has invented several ways to remove phosphate from proteins
1. Ser/Thr - phosphatases Metal-containing enzymes
DxH…DxxD…..N PP1, PP2A, PP2B (PPP); PP2C (PPM)
2. Protein-tyrosine phosphatases Protein-tyrosine phosphatases
C(X)5R Dual-specificity phosphatases
Low molecular weight phosphatases
Cdc25
3. RNA pol CTD phosphatase family Haloacid dehalogenase-related enzymes
DxDxT….GDxxxE Eyes absent
Many others

How many human protein kinases and phosphatases?
Manning et al. Science 208:1912 (2002) (http://www.kinase.com)
Protein phosphatases (the phosphatome)~140 protein phosphatases including:
38 protein-tyrosine phosphatases 38 serine/threonine phosphatases
(18 PP1/2A (PPP); 20 PP2C (PPM))62 DSPs (e.g. MKPs, PTEN); 8 HADs (EyA/FCP)
Protein kinases (the kinome)518 protein kinases including: (not quite a 1001!) 478 conventional protein kinases (ePKs)
(16 have tandem catalytic domains)388 protein-serine/threonine kinases 90 protein-tyrosine kinases58 receptor protein-tyrosine kinases32 non-receptor protein-tyrosine kinases
(~50 may lack catalytic activity; ~106 pseudogenes) 40 atypical protein kinases in 7 families (e.g. alpha kinases)
~2.5% genes directly devoted to protein phosphorylation and dephosphorylation, and possibly up to 5%, if regulatory subunits,
inhibitors and scaffolding/anchoring proteins are included.
There are as many tyrosine phosphatases as tyrosine kinases

Manning, Whyte, Martinez,Hunter and SudarsanamScience 208:1912 (2002)

A 1001 hairs?Marc Bitensky (March, 1990)
The beard kinome

Human tyrosinekinases
(90)

Alternative transcripts of mouse protein kinases and phosphatases
Products Number per locus
Transcripts 6.7
Polypeptides 3.7
Domain combinations 1.6
5’ exons 1.8
3’ exons 1.6
Most of the alternative coding products differ outside the catalytic domain, but a significant number of kinases have variant catalytic domains
Forrest et al. Genome Biol 7:R5 (2006) (variant.imb.uq.edu.au)

Global dynamics of protein phosphorylation
• 2,002 phosphorylation sites identified on 967 HeLa nuclear proteins (Beausoleil, Gygi, PNAS 101:12130, 2004)
• 5,635 phosphorylation sites identified on 2,328 proteins from mouse liver(Villen, Gygi, PNAS 104:1488, 2007)
• 6,600 phosphorylation sites identified on 2,244 proteins in HeLa cells; ~14% of these change >2 fold within 20 min of EGF (Olsen, Mann, Cell127:635, 2006)
• CST Phosphosite database has >48,400 phosphorylation sites, including 6,200 human proteins (N.B. Bodenmiller, Aebersold, Nat Meth 4:231, 2007)
Analysis of global phosphorylation events using enrichment ofphosphopeptides by IMAC, TiO2, PAC or phosphoantibodies derived by proteolytic digestion of cellular fractions followed by tandem MS identification of phosphopeptides and phosphorylation sites
These data suggest that the majority of intracellular proteins are phosphorylated at one or more sites under the appropriate condition -occupancy of many of these sites can change in response to stimuli

Functional categories of EGF-regulated phosphoproteins
QuickTime™ and aTIFF (Uncompressed) decompressor
are needed to see this picture.
Olsen, Mann, Cell 127:635 (2006)

The human kinome
• The completion of the human genome sequence has permitted a complete cataloguing of all human protein kinases
• There are 518 protein kinase genes, of which about 450 have kinase activity and 90 are tyrosine kinases. This represents about 2% of all human genes
• There are more than 140 protein phosphatases, and a large number of additional types of protein that recognize proteins once they are phosphorylated through phosphobinding domains
• Most of the proteins in a cell can be phosphorylated at one or often multiple sites under the right conditions
• Protein phosphorylation is a major mechanism of signal transduction in eukaryotic cells and perturbation in phosphorylation-driven processes can lead to disease

Protein kinases are implicated in diverse diseases
• >150/518 (~30%) of human protein kinases reportedly implicated in various diseases
• Many more are likely to follow from expression, sequencing, and SNP analyses, genetics and functional genomics
• Kinases are tractable drug targets with several approved drugs and huge development efforts
Gerard Manning (CST “Kinase and Signaling Reference Guide”)(www.cellsignal.com/reference/kinase disease.asp)

Human diseases caused by mutations in proteinkinases
DISEASE KINASE
• MYOTONIC MUSCULAR DYSTROPHY MYOTONIN PROTEIN KINASE• X-LINKED AGAMMAGLOBULINEMIA BRUTON TYROSINE KINASE (BTK)• HIRSCHSPRUNG DISEASE, MEN RET• CD8 DEFICIENCY FORM OF SCID ZAP70• X-LINKED SCID JAK3• CRANIOSYNOSTOSIS FGF RECEPTOR KINASES• PEUTZ-JEGHERS SYNDROME LKB1• COFFIN-LOWRY SYNDROME RSK2• ATAXIA-TELANGIELTASIA ATM• LI-FRAUMENI SYNDROME CHK2• WILLIAMS SYNDROME LIMK1• LEPRECHAUNISM, DIABETES INSULIN RECEPTOR• WOLFF-PARKINSON-WHITE SYNDROME AMP-ACTIVATED KINASE (AMPK)• GORDON HYPERTENSION SYNDROME WNK1 AND WNK4• WOLCOTT-RALLISON SYNDROME eIF2AK3/PEK• HEREDITARY EARLY-ONSET PARKINSON’S DISEASE PINK1 • HEREDITARY EARLY-ONSET PARKINSON’S DISEASE LRRK2• PULMONARY HYPERTENSION TGFβ FAMILY RECEPTOR BMPR-II• FAMILIAL ADVANCED SLEEP PHASE SYNDROME CKIδ• RETT SYNDROME (NEURODEVELOPMENT DISORDER) CDKL5• HYPORESPONSIVENESS TO BACTERIAL INFECTION IRAK4
• POLYCYTHEMIA VERA JAK2• MELANOMA & OTHER SPORADIC CANCERSB-RAF• PAPILLARY RENAL CANCER MET RECEPTOR KINASE• CHRONIC MYELOGENOUS LEUKEMIA TEL-PDGF RECEPTOR KINASE• CHRONIC MYELOGENOUS LEUKEMIA BCR-ABL• NON-HODGKINS LYMPHOMA ALK• 5-10% NONSMALL CELL LUNG CANCERS EGF RECEPTOR KINASE• COLON, BREAST & OTHER SPORADIC CANCERS PI-3 KINASE (P110α SUBUNIT)

Cell Signaling Technology, 2005
PKs associated with human diseases
Protein kinases with disease connections

• 355 cancer genes implicated by mutation - ~1% of all human genes
• 90% cancer genes show somatic mutations in cancer, 20% show germ line mutations and 10% show both
• The most common mutation class among cancer genes are chromosomal translocations that create chimeric genes or appose a gene next a regulatory element of another gene
• More cancer genes have been found in leukemias, lymphomas and sarcomas than in other types of cancer despite the fact that they represent only 10% all cancers
• The protein kinase catalytic domain is the commonest domain among cancer genes (30). Domains involved in DNA binding and transcriptional regulation are also common
Human cancer genes
Futreal et al. Nat Rev Cancer 4:177 (2004)

Cancer and the kinomeKinases control cancer pathways
• 120 kinases strongly implicated in cancer; more to come
• Kinases control complex pathways and signal transduction, implicated in all major steps/pathways of transformation
Somatic mutations and kinases• Kinases as the test case for large scale resequencing of
tumor genes: efforts at Sanger Center, JHU, Venter, Broad, elsewhere
• Results to date are a mixture of stunning (BRAF in melanoma), impressive (high frequency of PI3-kinase pathway gene mutations in colon cancer - RTKs, IRS2, PIK3CA, PTEN, PDK1, AKT2, PAK4 - as well as other tyrosine kinase and PTP genes), and underwhelming (breast cancer kinome) (Davies et al. Nature 417:949; Bardelli et al. Science 300:949; Wang et al. Science304:1164; Davies et al. Cancer Res 65:7591; Stephens et al. Nature Genet 37:590; Parsons et al. Nature 436:792; Rand et al. PNAS 102:1434; Sjoblom et al. Science 314:268; Bignell et al Genes Chr Cancer 45:42; Human Cancer Genome Project)

Protein Kinases/Phosphatases and Cancer
• Over half of the 90 tyrosine kinases are implicated in human cancer either through gain of function mutations (e.g. Bcr-Abl), gene amplification (e.g. EGF receptor) or overexpression (e.g. c-Src) or as tumor suppressors (e.g. Syk, c-Fes, Csk, EphB2, EphB3, EphB4)
• Many serine kinases are also implicated in cancer through activating mutations (e.g. B-Raf), overexpression (e.g. Aurora A), or loss of function mutations (e.g. Lkb1)
• 164 protein kinase genes map to amplicons found in tumors• 80 protein kinase genes map to chromosomal disease loci and these
are candidate genes for the causative mutation in hereditary disease (e.g. activating mutations in the Ret and Met RTKs in predisposition to cancer)
• Inactivating and activating mutations in tyrosine/lipid and serine phosphatases have also been implicated in cancer (e.g. PTEN, SHP-2, PRL-3; RPTPβ, PTP-BAS, PEZ, RPTPγ, LAR, PTPH; Pr65/PP2A)

Protein kinases as drug targets in disease therapy• The involvement of protein kinases and altered phosphorylation in
cancer and other diseases has been well established
• Protein kinases are often mutated or overexpressed in cancer. More than 25 protein kinase genes are known to be mutated in human cancer, and ~120 protein kinases are implicated in cancer. It seems likely that additional currently uncharacterized protein kinases will prove to play a role in cancer, and all these protein kinases make potential drug targets for cancer therapy
• Enzymes (e.g. protein kinases) generally make good drug targets
• Initial concerns that it would not be possible to make specific inhibitors because most known kinase inhibitors bind to the conserved ATP binding site and because high intracellular ATP concentrations would compete for binding have proved groundless
• Selective protein kinase inhibitors have been developed and are proving effective in cancer therapy - GleevecTM, TarcevaTM, SutentTM

Cancer drugs that act against tyrosine kinases
DRUG CANCER TARGETSmall molecule drugs
GleevecTM (imatinib) leukemia (CML) Bcr-Abl tyrosine kinaseIressaTM (gefitinib) lung cancer EGF receptor TKTarcevaTM (erlotinib) lung cancer EGF receptor TKSutentTM (sunitinib) GI stromal tumor/RCC Kit receptor TKSprycelTM (dasatinib) leukemia (CML) Bcr-Abl tyrosine kinaseTykerbTM (lapatinib) breast cancer ErbB2 RTK
Many in trials several types/AML angiogenesis/Flt3 RTKs
Monoclonal antibody drugsHerceptinTM (trastuzumab) breast cancer ErbB2 RTKErbituxTM (cetuximab) breast/renal cancer EGF receptor TK AvastinTM (cevacizumab) colon cancer VEGF
>70 protein kinase inhibitors are in cancer clinical trials, including several directed against serine/threonine kinases implicated in cancer (Mark Via, Cambridge Healthtech). The Raf inhibitor sorafenib (NexavarTM) has recently been approved for treatment of renal carcinoma. Rapamycin, an mTOR inhibitor, and analogues are also in clinical trials for several cancers

The long road to the GLEEVECTM cancer drug
1845CML
1960Chr 22∆Ph Chr
1973t(9:22)translocation
1982Bcr-Abl(22:9)
1984Bcr-Abl
PTKactivity
1980v-Abltyrosinekinase
1988First PTKinhibitorsreported(TKI)
1992CGP57148PDGFR/Kit/Abl TKI(Novartis)
1998STI571in CMLpatients
1990Bcr-Abl causesCML in mice
1970Ab-MuLV
1978v-Ablprotein
2000STI571/Ablstructure
1911RSV
1970v-srcgene
1975c-srcgene
1977v-Srcprotein
1978v-SrcPK
1979v-Srctyrosinekinase
1927W mutantmouse
1988W isc-kit
1986HZ4-FeSV
1986v-KitPTK
1998c-kit gofGISTmutants
1996c-kit gofleukemiamutants
2001NEJMpapersreportingSTI571efficacyin CMLand GIST
2000STI571in GISTpatients
1996STI571 inhibitsCML cell growthand v-Abl tumors
1977mTantigen
1952polyomavirus
1979mTassociatedPTK activity
1983mTassociatedc-Src
GleevecTM approved by the FDA May 10, 2001

Some outstanding questions
• How/when did tyrosine kinases and phosphatases evolve?
• Are there additional functions for tyrosine phosphorylation?
• Are there additional types of P.Tyr-binding domain?
• How large is the P.Tyr phosphoproteome?
• How are the actions of the tyrosine kinases and phosphatases coordinated and do they act in combinations? How fast do phosphates turn over at a particular site?
• Where do critical tyrosine phosphorylation and dephosphorylation events occur in the cell?
• Can we develop more specific inhibitors or inhibitors that target a desired set of tyrosine kinases or phosphatases, or block P.Tyr interactions for disease therapy?

KinomicsGerard Manning (Sugen/Salk)
Greg PlowmanSucha SudarsanamSean Caenepeel
Science 208:1912TiBS 27:514PNAS 101:11707
http://www.kinase.com

Walter EckhartMary Anne Hutchinson
Bart SeftonJon CooperKaren Beemon
THANKS GO TO
AND OF COURSE THE OLD BUFFER!