cancer research ancca/atad2 overexpression...
TRANSCRIPT

Tum
ANCPatandCon
EkateCliffor
Abst
Intro
Funcanceoutcomstandimolecestrogselvesrespon
AuthorMedicin2UC DaDavis, S
Note:Resear
E.V. Ka
Correstive MeHutchiadboroCenter/CA 95hwzche
doi: 10
©2010
Cance9402
Dow
Published OnlineFirst September 23, 2010; DOI: 10.1158/0008-5472.CAN-10-1199
Canceresearch
or and Stem Cell Biology
CA/ATAD2 Overexpression Identifies Breast Cancerients with Poor Prognosis, Acting to Drive ProliferationSurvival of Triple-Negative Cells through
R
trol of B-Myb and EZH2
rina V. Kalashnikova1,2, Alexey S. Revenko1,2, Abigael T. Gemo1,2, Nicolas P. Andrews1,2,
d G. Tepper1,2, June X. Zou1,2, Robert D. Cardiff1, Alexander D. Borowsky1, and Hong-Wu Chen1,2ractChr
ATPascruciaIn thiscorrelaoveralthat reof brestrongthat Atrols talong
as moresive to en
s' Affiliatioe, Internalvis Canceacramento
Supplemench Online (h
lashnikova
ponding Audicine, Unson Drive,wsky@ucdBasic Scie817. Phonn@ucdavis.
.1158/0008-
American A
r Res; 70(
nloaded
omatin coregulators are important factors in tumorigenesis and cancer progression. ANCCA is an AAA+e and a bromodomain-containing nuclear coactivator for the estrogen and androgen receptors that isl for assembly of chromatin-modifying complexes and proliferation of hormone-responsive cancer cells.study, we show that ANCCA is overexpressed in >70% of breast tumors and that its high protein leveltes well with tumor histologic grades (P < 0.0001), highlighting ANCCA as a prognostic factor for poorl survival and disease recurrence. Strikingly, high-level ANCCA correlated with triple-negative tumorspresent highly aggressive disease. Analysis of ANCCA transcript levels in multiple expression profilesast cancer identified ANCCA as a common signature gene, indicating that elevated transcripts alsoly correlate with tumor metastasis and poor survival. Biological and mechanistic investigations revealedNCCA is crucial for proliferation and survival of triple-negative/basal-like cancer cells and that it con-he expression of B-Myb, histone methyltransferase EZH2, and an Rb-E2F core program for proliferation,with a subset of key mitotic kinesins and cell survival genes (IRS2, VEGF, and Akt1). In particular, ANCCApression correlated strongly with EZH2 in tumors. Our results suggest that ANCCA may integrate mul-
overextiple oncogenic programs in breast cancer, serving in particular as a prognostic marker and a therapeutictarget for triple-negative cancers. Cancer Res; 70(22); 9402–12. ©2010 AACR.
man eoftening thgester(TNBCmenttors (1most
duction
ctional genomics and gene-profiling studies of breastr continue to improve our prediction of clinicale and selection of therapeutics as well as our under-
ng of tumor biology, by subtyping the tumors on theirular profiles. Among the subtypes, the luminal/en receptor α (ERα)–positive tumors present them-
differentiated, less aggressive, and highlydocrine therapy. Tumors overexpressing hu-
contathoseone stliferatsubtypderpintreatmthe murgentreatmChr
histonrecen(4–6).gene pcancethe prrefs. 7
ns: 1Departments of Biochemistry and MolecularMedicine, and Pathology, School of Medicine, andr Center/Basic Sciences, University of California at, California
tary data for this article are available at Cancerttp://cancerres.aacrjournals.org/).
and A.S. Revenko contributed equally to this work.
thors: Alexander D. Borowsky, Center for Compara-iversity of California, Davis, County Road 98 andDavis, CA 96616. Phone: 530-754-8182; E-mail:avis.edu or Hong-Wu Chen, UC Davis Cancernces, University of California at Davis, Sacramento,e: 916-734-7743; Fax: 916-734-0190; E-mail:edu.
5472.CAN-10-1199
ssociation for Cancer Research.
22) November 15, 2010
on July 28, 2018. © 2cancerres.aacrjournals.org from
pidermal growth factor receptor 2 (HER2), althoughmore aggressive, are responsive to therapeutics target-e growth factor receptors. The triple-negative [ER, pro-one receptor (PR), and HER2 negative] breast cancers), however, remain a major challenge for the develop-of effective therapeutics and identification of risk fac-). Recent expression array studies (2, 3) characterizeTNBCs as being basal-like due to an expression profileining high levels of basal cell cytokeratins, similar toof myoepithelial cells. Although poorly understood,riking feature of TNBCs is the high expression of pro-ion signature genes. Unlike ER-positive or HER2-highes, no molecular markers have been defined that un-TNBC development or effectively guide their clinicalent. As a result, conventional chemotherapy remainsainstay treatment for them. Therefore, there is ant need for the identification of risk factors and newent options for triple-negative tumors.omatin coregulators, particularly in the forms ofe-modifying and chromatin-remodeling enzymes, havetly emerged as important players in tumorigenesisWe recently identified a previously uncharacterizedroduct dubbed ANCCA (for AAA+ nuclear coregulatorr associated) as both a direct target and an activator of
oto-oncogene AIB1 (also known as ACTR and SRC-3;, 8). ANCCA, a novel member of the AAA+ ATPase010 American Association for Cancer Research.

familylevelscancerstrongtion (8of ERandroinvolvthe AANCCtors (8nismhistonIn t
proteibreastpresseANCCrecurrANCCand thmultipand su
Mate
Cell cimmuand WHum
Amer2008. Tity bygrowthmonesing thfamilyPCR (with tbreastcells wing. Rprevioplatedusingturer'swere uand thtion (sequenMater
Cell pformaHCC
well ofor 24ANCC
cells wplatesdescriafter tcounttion inwell p(LonzColonAldric
TermdUTPCel
later aon gladeoxying (T(Rochcol. SMediaOlymp10 raand a
TumoArc
from 2ted atinformry Tabwereretrievmicrofor 30BR100Inc., bstainibeenwas butes atempe(at 1:3(AC22raisedGST-AEsche(IHC)tumorincubwashebodieDH-bia diamcountusing
ANCCA Is a New Prognostic Factor for Breast Cancer
www.a
Dow
Published OnlineFirst September 23, 2010; DOI: 10.1158/0008-5472.CAN-10-1199
, also possesses a bromodomain. We found that highof ANCCA are expressed in breast cancer and prostatecells and that RNAi-mediated knockdown of ANCCAly inhibits hormone-dependent cancer cell prolifera-, 9). We also showed that ANCCA acts as a coactivatorα and androgen receptor to mediate estrogen- orgen-induced expression of specific subsets of genesed in proliferation and survival of cancer cells. BothAA+ ATPase and bromodomains are required forA to serve as a transcriptional coregulator of the recep-). Our further study indicated that one major mecha-of ANCCA function is to facilitate the assembly of ae-modifying protein complex at the chromatin (8).his study, we investigated ANCCA expression at bothn and transcript levels in multiple sets of humancancer specimens and found that ANCCA is overex-d in the majority of the tumors. High levels ofA directly correlate with poor survival and diseaseence in the patients. We also present results that highA is strongly associated with triple-negative tumorsat aberrant ANCCA expression in TNBC cells controlsle oncogenic pathways for cancer cell proliferationrvival.
rials and Methods
ulture, siRNA transfection, chromatinnoprecipitation assay, RNA analysis,estern blottingan breast cancer cell lines were obtained from the
ican Type Culture Collection (ATCC) in 2002 andhey were tested recently (in 2009–2010) for authentic-monitoring cell morphology under a microscope;rate through cell counting; response to factors or hor-for growth; and expression of genes/proteins, includ-
e hormone receptors and members of the HER2/ErbB2, through Western blotting and reverse transcriptase-RT-PCR). These cell lines were confirmed to be in linehe ATCC descriptions and with the literature. Humanepithelial cells (HMEC) were from Lonza/Clonetics. Allere used within 4 months after initial receipt or thaw-NA analysis and Western blotting were performed asusly described (10). For transfection of siRNAs, cellsat 2.0 × 105 per well in six-well plates were transfectedDharmafect 1 (Dharmacon) according to the manufac-instructions. siRNA targeting ANCCA or control siRNAsed at 100 nmol/L concentration. Details of cell lineseir culture conditions, chromatin immunoprecipita-ChIP), antibodies used for Western blotting, andces for PCR primers are described in Supplementaryials and Methods.
roliferation and soft-agar colonytion assays1937 cells were seeded at a density of 1.5 × 105 per
f a six-well plate and maintained in full growth medium
hours before being infected with equal titers of adeno-A-HA or adenovector adenoviruses (8). MDA-MB 468The pfollow
acrjournals.org
on July 28, 2018. © 2cancerres.aacrjournals.org nloaded from
ere plated at a density of 2.0 × 105 per well in six-well, and were transfected with siRNA 24 hours later asbed above. The medium was changed every other dayransfection, and cell proliferation was measured by celling of coded samples in triplicates. For colony forma-soft agar, 5,000 cells were seeded in each well of six-lates in medium containing 0.4% SeaPlaque Agarosea) on top of a base medium containing 0.8% agarose.ies were stained 4 to 5 weeks later with MTT (Sigmah) and counted using a light microscope.
inal deoxynucleotidyl transferase–mediatednick end labeling assayls were plated and transfected with siRNA 24 hourss described above. Cells were plated 72 hours laterss chamber slides (Nunc) and processed for terminalnucleotidyl transferase–mediated dUTP nick end label-UNEL) assay using the In Situ Cell Death Detection kite Diagnostics) according to the manufacturer's proto-lides were mounted with VECTASHIELD Mounting(Vector Laboratories). Images were acquired usingus BX6. For quantification of TUNEL-positive cells,ndom fields of the same condition were countedveraged.
r specimens and immunohistochemistryhival paraffin-embedded primary tumor samples were25 women diagnosed with breast carcinoma and trea-UC Davis Medical Center during 1998 to 2004. Otheration on the patients and tumors are in Supplementa-le S1. Five-micrometer sections of the tumor blocksfirst subjected to deparaffinization and then antigenal in 0.01 mmol/L sodium citrate buffer (pH 6.0) in awave oven at 1,000 W for 5 minutes and then at 100 Wminutes. Breast tumor tissue microarrays (TMA;2, BR961, and BR208) were obtained from US BioMax,aked at 60°C for 2 hours, and processed for immuno-ng. TMA for EZH2 and ANCCA correlation study hasdescribed (7). Nonspecific immunoglobulin bindinglocked using 10% fetal bovine serum in PBS for 30 min-t room temperature. Slides were then incubated at roomrature for 30 minutes with anti-ANCCA antibody00), anti–Ki-67 (at 1:1,000; NeoMarker), and anti-EZH2; at 1:50; Cell Signaling). Anti-ANCCA antibody wasin rabbit (Covance) and affinity-purified by usingNCCA (amino acids 2–264) expressed and purified fromrichia coli. Its specificity for immunohistochemistrywas determined using a panel of cell lines and xenografts (9) and is also shown in Supplementary Fig. S1. Afteration with the primary antibody, the sections wered and incubated with biotin-conjugated secondary anti-s for 30 minutes followed by incubation with avidinotinylated horseradish peroxidase and developed usinginobenzidine substrate kit (Vector Laboratories) and
erstained using Gill's hematoxylin. Images were acquiredan Olympus microscope with DPController software.
ercentage of positively stained nuclei was scored ass: 0% to 10%, score 0; 11% to 25%, score 1; 26% to 50%,Cancer Res; 70(22) November 15, 2010 9403
010 American Association for Cancer Research.
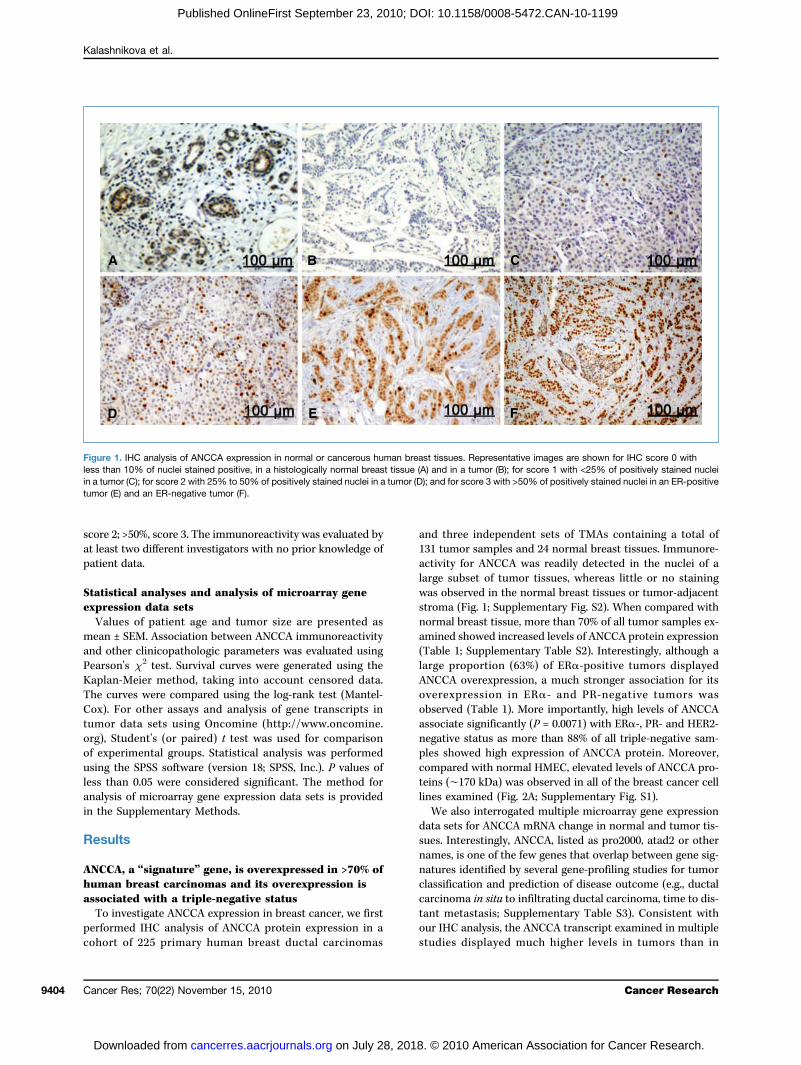
score 2at leaspatien
StatisexpreVal
meanand otPearsoKaplaThe cuCox).tumororg), Sof expusingless thanalysin the
Resu
ANCChumaassocTo i
perforcohor
and th131 tuactivitlargewas ostromnormaamine(TablelargeANCCovereobservassocinegatiples scompateins (lines eWe
data ssues. Inamesnatureclassifcarcintant m
Figureless thain a tumor (C); for score 2 with 25% to 50% of positively stained nuclei in a tumor (D); and for score 3 with >50% of positively stained nuclei in an ER-positivetumor (
Kalashnikova et al.
Cance9404
Dow
Published OnlineFirst September 23, 2010; DOI: 10.1158/0008-5472.CAN-10-1199
; >50%, score 3. The immunoreactivity was evaluated byt two different investigators with no prior knowledge oft data.
tical analyses and analysis of microarray genession data setsues of patient age and tumor size are presented as± SEM. Association between ANCCA immunoreactivityher clinicopathologic parameters was evaluated usingn's χ2 test. Survival curves were generated using then-Meier method, taking into account censored data.rves were compared using the log-rank test (Mantel-For other assays and analysis of gene transcripts indata sets using Oncomine (http://www.oncomine.tudent's (or paired) t test was used for comparisonerimental groups. Statistical analysis was performedthe SPSS software (version 18; SPSS, Inc.). P values ofan 0.05 were considered significant. The method foris of microarray gene expression data sets is providedSupplementary Methods.
lts
A, a “signature” gene, is overexpressed in >70% ofn breast carcinomas and its overexpression isiated with a triple-negative statusnvestigate ANCCA expression in breast cancer, we first
E) and an ER-negative tumor (F).
med IHC analysis of ANCCA protein expression in at of 225 primary human breast ductal carcinomas
our IHstudie
r Res; 70(22) November 15, 2010
on July 28, 2018. © 2cancerres.aacrjournals.org nloaded from
ree independent sets of TMAs containing a total ofmor samples and 24 normal breast tissues. Immunore-y for ANCCA was readily detected in the nuclei of asubset of tumor tissues, whereas little or no stainingbserved in the normal breast tissues or tumor-adjacenta (Fig. 1; Supplementary Fig. S2). When compared withl breast tissue, more than 70% of all tumor samples ex-d showed increased levels of ANCCA protein expression1; Supplementary Table S2). Interestingly, although aproportion (63%) of ERα-positive tumors displayedA overexpression, a much stronger association for itsxpression in ERα- and PR-negative tumors wased (Table 1). More importantly, high levels of ANCCAate significantly (P = 0.0071) with ERα-, PR- and HER2-ve status as more than 88% of all triple-negative sam-howed high expression of ANCCA protein. Moreover,red with normal HMEC, elevated levels of ANCCA pro-∼170 kDa) was observed in all of the breast cancer cellxamined (Fig. 2A; Supplementary Fig. S1).also interrogated multiple microarray gene expressionets for ANCCA mRNA change in normal and tumor tis-nterestingly, ANCCA, listed as pro2000, atad2 or other, is one of the few genes that overlap between gene sig-s identified by several gene-profiling studies for tumorication and prediction of disease outcome (e.g., ductaloma in situ to infiltrating ductal carcinoma, time to dis-etastasis; Supplementary Table S3). Consistent with
1. IHC analysis of ANCCA expression in normal or cancerous human breast tissues. Representative images are shown for IHC score 0 withn 10% of nuclei stained positive, in a histologically normal breast tissue (A) and in a tumor (B); for score 1 with <25% of positively stained nuclei
C analysis, the ANCCA transcript examined in multiples displayed much higher levels in tumors than in
Cancer Research
010 American Association for Cancer Research.
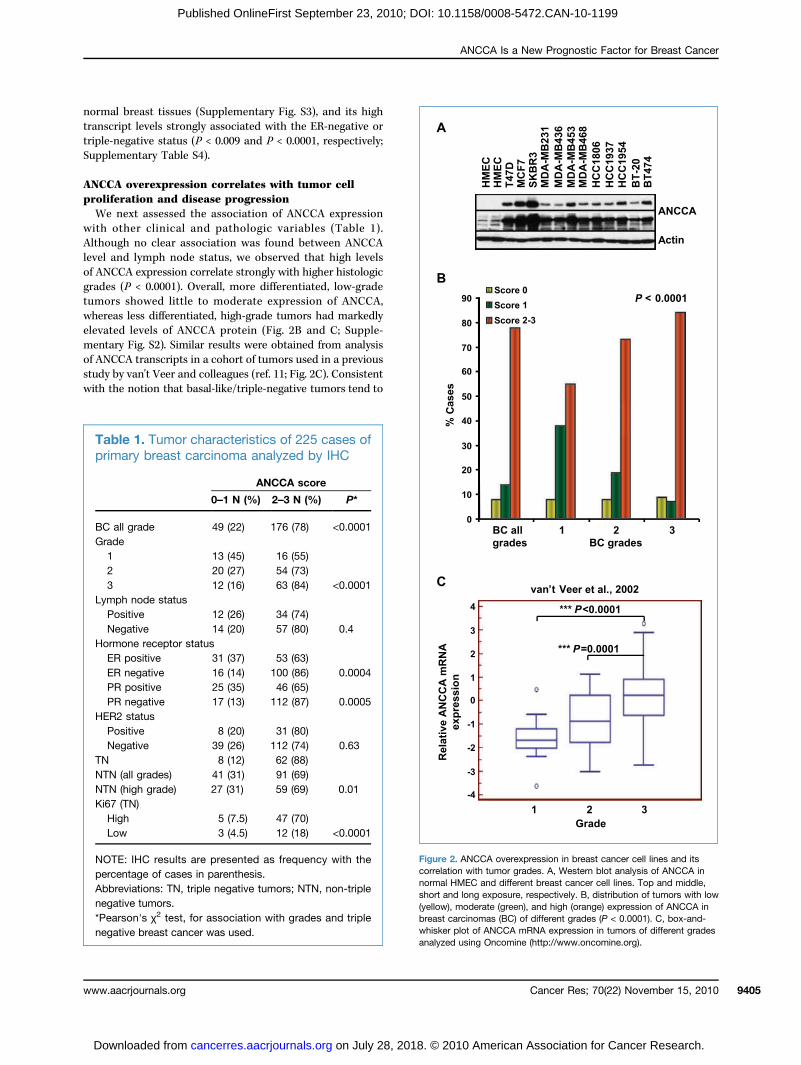
normatransctriple-Supple
ANCCprolifWe
withAlthoulevel aof ANCgradestumorwhereelevatmentaof ANCstudywith t
Tabprim
BCGra
123
LymPN
HorEEPP
HERPN
TNNTNNTNKi67
HL
NOTpercAbbneg*Peneg
ANCCA Is a New Prognostic Factor for Breast Cancer
www.a
Dow
Published OnlineFirst September 23, 2010; DOI: 10.1158/0008-5472.CAN-10-1199
l breast tissues (Supplementary Fig. S3), and its highript levels strongly associated with the ER-negative ornegative status (P < 0.009 and P < 0.0001, respectively;mentary Table S4).
A overexpression correlates with tumor celleration and disease progressionnext assessed the association of ANCCA expressionother clinical and pathologic variables (Table 1).gh no clear association was found between ANCCAnd lymph node status, we observed that high levelsCA expression correlate strongly with higher histologic(P < 0.0001). Overall, more differentiated, low-grades showed little to moderate expression of ANCCA,as less differentiated, high-grade tumors had markedlyed levels of ANCCA protein (Fig. 2B and C; Supple-ry Fig. S2). Similar results were obtained from analysisCA transcripts in a cohort of tumors used in a previous
by van't Veer and colleagues (ref. 11; Fig. 2C). Consistenthe notion that basal-like/triple-negative tumors tend toFigurecorrelanormalshort a(yellowbreastwhiskeanalyze
leary breast carcinom
s
all 8 0de
534 0
phos 4eg 0moR 3R 6 0R 5R 7 02os 0eg 4
(a(h 9
ative breast cancer was used.
acrjournals.org
on July 28, 2018. © 2cancerres.aacrjournals.org nloaded from
2. ANCCA overexpression in breast cancer cell lines and itstion with tumor grades. A, Western blot analysis of ANCCA inHMEC and different breast cancer cell lines. Top and middle,nd long exposure, respectively. B, distribution of tumors with low), moderate (green), and high (orange) expression of ANCCA incarcinomas (BC) of different grades (P < 0.0001). C, box-and-
1. Tumor characteris
tics of 225 cases ofa analyzed by IHC0–1 N (%
ANCCA
) 2–3 N
core
(%) P*
grade 49 (22)
176 (7 ) <0.00 113 (45)
16 (5 ) 20 (27) 54 (7 ) 12 (16) 63 (8 ) <0.00 1node status
itive 12 (26) 34 (7 ) ative 14 (20) 57 (8 ) 0.4 ne receptor status positive 31 (37) 53 (6 ) negative 16 (14) 100 (8 ) 0.00 4 positive 25 (35) 46 (6 ) negative 17 (13) 112 (8 ) 0.00 5 status itive 8 (20) 31 (8 ) ative 39 (26) 112 (7 ) 0.638 (12)
62 (8 ) 8ll grades) 41 (31) 91 (69) igh grade) 27 (31) 59 (6 ) 0.01 N) (Tigh 5 (7.5) 47 (70)ow 3 (4.5) 12 (18) <0.0001
E: IHC results are presented as frequency with theentage of cases in parenthesis.reviations: TN, triple negative tumors; NTN, non-tripleative tumors.arson's χ2 test, for association with grades and triple
r plot of ANCCA mRNA expression in tumors of different gradesd using Oncomine (http://www.oncomine.org).
Cancer Res; 70(22) November 15, 2010 9405
010 American Association for Cancer Research.

displaANCChigh Kand Kalso revidual
HighlsurvivWe
dictivetientswereANCCFig. 3A
sion rwhensionfor hi5 yeaand sconsidbe obSuppleof ANcomp(Fig. 3the da
Figure(A) andcolleag(11). Th
Kalashnikova et al.
Cance9406
Dow
Published OnlineFirst September 23, 2010; DOI: 10.1158/0008-5472.CAN-10-1199
y high proliferation index (1), the majority (70%) ofA-overexpressing triple-negative tumors also showedi-67 staining (P < 0.0001; Table 1). IHC analysis of ANCCAi-67 expression in adjacent sections of tumor specimensvealed a high accordance in their expression in indi-tumors (Supplementary Fig. S4).
y elevated ANCCA predicts poor overallal and disease recurrencealso examinedwhether strong ANCCA expression is pre-of disease progression by analysis of a cohort of 185 pa-with up to 11 years of follow-up information. Tumorsdivided into two groups (low ANCCA and high
A) according to ANCCA IHC scores. As shown in that hues (26), and associates with metastasis at 5 years after diagnosis in tumor data se data sets were analyzed for association by using Oncomine (http://www.oncom
r Res; 70(22) November 15, 2010
on July 28, 2018. © 2cancerres.aacrjournals.org nloaded from
evealed a significantly shorter overall survival periodcompared with those with no or weak ANCCA expres-(P = 0.006 by Cox-Mantel log rank test; 85% survivalgh ANCCA group, 2.5 years, and for low ANCCA,rs). When the relationship between ANCCA levelsurvival was assessed with ER status being taken intoeration, their association with poor survival can stillserved in patients with ER-negative tumors (P = 0.035;mentary Fig. S5). Likewise, patients with high levelsCCA protein had an earlier time to disease recurrenceared with patients with low ANCCA expressionB). Moreover, examination of ANCCA transcripts inta sets of several “gene signature” studies revealed
igh ANCCA mRNA levels associate with high probabi-, patients whose tumors showed high ANCCA expres- lity of death and tumor metastasis (Fig. 3C).
3. ANCCA overexpression correlates with poor outcomes. Kaplan-Meier analysis of ANCCA protein expression and disease-free survivaldisease recurrence (B). C, high level of ANCCA transcripts correlates with poor disease-free survival in tumor data sets from Ivshina and
ets from Wang and colleagues (19) and van't Veer and colleaguesine.org).
Cancer Research
010 American Association for Cancer Research.

ANCCand asurvivBec
triple-ANCCovereHMECMB-46(Suppon theor in adown(SuppHCC1virussignifiinfecteWe
survivfectedin Fig.trol siTUNEshoweSimilaMB-46providmotes
ANCCprolifincludGiv
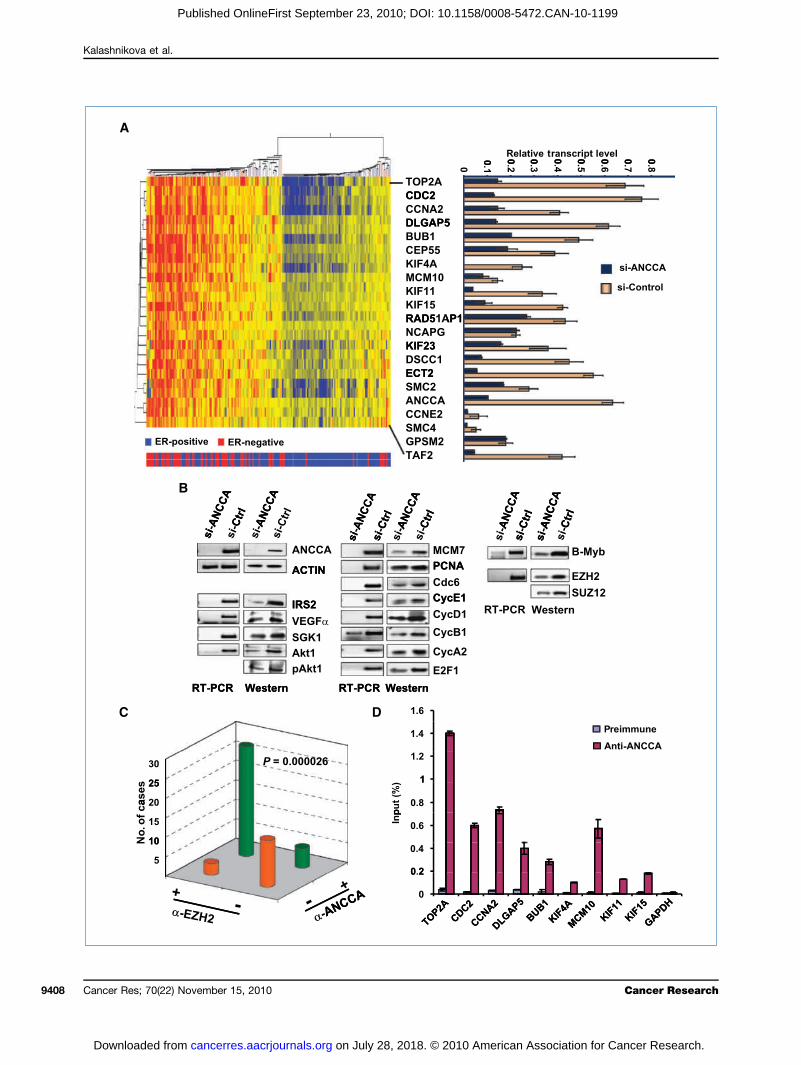
shownBIG CANCCing abANCChighlywith tin mitDSCC(MCMplemestudy,geneER-newherered bANCCthat mbecaupressiFurthDNAcyclinantige
Figureof TNBadeno-targetincell prolater (fowere tragar fowith MTand thecells wday 3 after siRNA transfection. Paired-samples t test (A and B) orindependent-samples t test (C and D) was used.
ANCCA Is a New Prognostic Factor for Breast Cancer
www.aacrjournals.org
on July 28, 2018. © 2cancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 23, 2010; DOI: 10.1158/0008-5472.CAN-10-1199
A overexpression promotes anchorage-dependentnchorage-independent proliferation andal of TNBC cellsause expression of ANCCA was particularly high innegative tumors, we examined the role of overexpressedA in TNBC cells. As shown in Fig. 2A, ANCCA is highlyxpressed in the breast cancer cell lines relative tos. Knockdown of ANCCA through one siRNA in MDA-8 (Fig. 4A), MDA-MB-231 (Fig. 4C), and MDA-MB-453lementary Fig. S6) cells showed strong inhibitory effectsir proliferation either in regular two-dimensional culturesoft-agar, anchorage-independent growth assay. Knock-of ANCCA by using a different siRNA gave a similar effectlementary Fig. S7A and B). In the BRCA1-defective937 cells that express amoderate level of ANCCA, adeno-vector–mediated, ectopic expression of ANCCA led tocantly increased proliferation when compared with cellsd with the adenovector control (Fig. 4B).next examined whether ANCCA was also important foral of triple-negative cells. MDA-MB-231 cells were trans-with siRNA and processed for TUNEL assay. As shown4D and Supplementary Fig. S8, whereas mock- or con-RNA–transfected cells had very few cells positive forL staining (<3%), cultures treated with ANCCA siRNAd a marked increase (∼25%) of TUNEL-positive cells.r results were obtained with MDA-MB-453 and MDA-8 cells (data not shown). Taken together, these resultse strong evidence that ANCCA overexpression pro-both proliferation and survival of TNBC cells.
A controls important regulators of celleration and survival pathways,ing EZH2 and B-Myb
en the transcriptional coactivator function of ANCCAin our previous study (7–9), we analyzed the TRANS-onsortium gene expression data set (12), in whichA serves as a prognostic signature gene, for genes hav-errant coexpression associated with overexpressedA. Strikingly, the analysis identified a group of genesenriched in control of cell proliferation and survivalhe top 20 genes (25 gene probes) functioning primarilyosis (DLGAP5, BUB1, CEP55, KIF4A, KIF11, KIF15, KIF23,1, ECT2, SMC2, SMC4, GPSM2), DNA replication10 and Top2A), and cell cycle progression (Fig. 5A; Sup-ntary Fig. S9). Intriguingly, consistent with our IHChierarchical clustering showed that high levels of thetranscripts, including ANCCA, cluster primarily ingative tumors (Fig. 5A, at the left side of the clustertumors are primarily ER-negative indicated by the
ars at the bottom). More importantly, results fromA knockdown in two different TNBC cells showedost of these genes are indeed controlled by ANCCAse their expression is downregulated upon ANCCA sup-on (Fig. 5A, right; Supplementary Fig. S10A and B).er analysis showed that additional key cell cycle andreplication regulators, including cyclin E1, cyclin B1,
A2, E2F1, Cdc6, MCM7, and proliferating cell nuclear4. Elevated ANCCA is required for proliferation and survivalC cells. A and B, cells were either infected with equal titers ofANCCA or adeno-control (HCC1937) or transfected with siRNAg ANCCA or control sequence (MDA-MB 468). Western blot andliferation assays were performed with cells harvested 48 hoursr Western blot) or at the indicated days. C, MDA-MB 231 cellsansfected with siRNA-ANCCA or siRNA-control and plated in softr colony formation or Western blotting. Colonies were stainedT 4 weeks later and counted. Representative images of coloniesir numbers from different treatments are shown. D, MDA-MB 231ere processed for TUNEL assay (left) or Western blotting, at
n, are also regulated by ANCCA (Fig. 5B, middle;
Cancer Res; 70(22) November 15, 2010 9407
010 American Association for Cancer Research.
Kalashnikova et al.
Cance9408
Dow
Published OnlineFirst September 23, 2010; DOI: 10.1158/0008-5472.CAN-10-1199
r Res; 70(22) November 15, 2010 Cancer Research
on July 28, 2018. © 2010 American Association for Cancer Research.cancerres.aacrjournals.org nloaded from

Suppleical fuexpresvascuphospknockBec
EZH2high pwhething ofsion a(Fig. 5Suz12complB-MybdiseasIHC thinvasiwith ppresseFig. S1correla(P < 0may ctodeteany ofPCR pgion omoterKif-4Atent whyde-3any siGAPD468 cethat Acell pr
Discu
TNBprofileation(1). A(e.g.,pRb-E
prolifemal fuor areidentifand asthe exsurvivANCCbreastHowepoorlypressimensassociInterewithresultlevelsand sTum
in advisms ucanceof clinmotheto beby varas profew getures.rise toof sho76 genpositivwas laConsicohorpredicconstihistoloANCCexprecompoing ththem
FigureincludinSpearmTumor Ewere tratranscrianalyzethe epitbetweeMDA-M
ANCCA Is a New Prognostic Factor for Breast Cancer
www.a
Dow
Published OnlineFirst September 23, 2010; DOI: 10.1158/0008-5472.CAN-10-1199
mentary Fig. S7C). Moreover, consistent with the crit-nction of ANCCA in TNBC cell survival, we found thatsion of survival pathway proteins such as IRS2, SGK1,lar endothelial growth factor α (VEGFα), Akt, andhorylated Akt was also significantly affected by ANCCAdown (Fig. 5B, left).ause overexpression of polycomb group protein (PcG)and mitotic regulator B-Myb seems to correlate withroliferation of basal-like tumors (13, 14), we examineder ANCCA also controls their expression. Indeed, silenc-ANCCA markedly inhibited EZH2 and B-Myb expres-t both the RNA and protein levels in TNBC cellsB, right), and the inhibition was also observed forprotein, a critical component of the polycomb-repressiveex 2 (PRC2). Therefore, these data show that EZH2 andare both downstream targets of ANCCA. To address thee relevance of ANCCA control of EZH2, we examined bye expression of ANCCA and EZH2 in a cohort of 48
ve breast carcinoma specimens on a TMA. Consistentrevious findings (15), we found that EZH2 is overex-d in 62% of the invasive tumors (Fig. 5C; Supplementary1). Importantly, ANCCA overexpression was significantlyted to the expression of EZH2 in the primary tumors.000026). These results suggest that deregulated ANCCAontribute to EZH2 overexpression in the tumors. Finally,rminewhether ANCCAdirectly controls the expression ofthe 20 genes identified, we performed ChIP assays. Withrimers that can readily amplify the proximal promoter re-f indicated genes, we found that ANCCA occupies the pro-region of Top2A, cdc2, cyclin A2, DLGAP5, Bub1, MCM10,, Kif-11, and Kif-15 inMDA-MB-436 cells (Fig. 5D). Consis-ith our data that ANCCA does not regulate glyceralde--phosphate dehydrogenase (GAPDH), we did not detectgnificant anti-ANCCA enrichment at the promoter ofH. Similar ChIP results were obtained from MDA-MB-lls (Supplementary Fig. S10C). Together, our data indicateNCCA is directly involved in control of key components ofoliferation and survival pathways in TNBC cells.
ssion
Cs, particularly those with basal-like gene expressions, tend to show highly elevated mitotic and cell prolifer-indices and poor prognosis for long-term survivalberrations of several key transcriptional regulators
B-Myb, EZH2 or c-Myc overexpression as well as2F deficiency) may underlie the elevated expression oftumorcore s
B-436 cells. ChIP DNA was analyzed by real-time PCR. Anti-ANCCA enrichments
acrjournals.org
on July 28, 2018. © 2cancerres.aacrjournals.org nloaded from
ration-associated genes (13, 16). Whether their abnor-nctions represent independent events in different tumorsintegrated by other mechanisms is unclear. We initiallyiedANCCAas a direct target of the oncogeneAIB1/ACTRa transcriptional coregulator for ERα and AR to promotepression of genes driving cancer cell proliferation andal (8, 9). Recent studies from us and others suggest thatA is overexpressed in many human cancers, includingcancer, and may act as a coactivator of c-Myc (8, 9, 17).ver, its role in breast cancer progression has beenunderstood. In this study, we examined ANCCA ex-
on in several cohorts of human breast cancer speci-and found that its high levels were significantlyated with poor overall survival and disease recurrence.stingly, ANCCA overexpression correlated stronglythe triple-negative subtype of breast cancer. Thes of in vitro experiments further showed that highof ANCCA were required to maintain proliferationurvival of TNBC cells.or gene expression signatures have been instrumentalancing our understanding of the molecular mechan-nderlying the diverse biological phenotypes of breastrs and now have become a proven utility for predictionical outcomes, such as tumor responsiveness to che-rapy (2, 11, 18–23). However, only a few genes seemshared across all of the different signatures identifiedious studies. Interestingly, ANCCA, which was listed2000, ATAD2, or other undefined names, is one of thenes that overlap frequently between different signa-For instance, ANCCA is one of the 231 genes that gavethe 70-gene signature that predicts a clinical outcomert interval to distant metastasis (11). It is also one of thees identified for prediction of distant metastasis of ER-e, lymph node–negative, primary breast cancer, whichter validated in a multicenter study (12, 19, 24, 25).stent with our IHC results from an independentt of tumors showing that ANCCA protein levels alonet tumor grades, a high level of ANCCA transcript is atuent of the genetic grading signature that can reclassifygic grades of breast cancer (26–28). We also found thatA overexpression strongly correlated with elevatedssion of proliferation-associated genes that are oftennents of the different gene signatures, thereby suggest-at overexpressed ANCCA may either collaborate withor act upstream to stimulate their expression in the
s. Intriguingly, ANCCA is not a signature gene for theerum response signature, which largely overlaps with5. ANCCA controls key regulators of cell proliferation and survival pathways including the oncogene EZH2. A, the top 20 genes (25 probe sets, notg ANCCA) found to be coexpressed with ANCCA in a set of 198 patients by unsupervised hierarchical clustering (12). Correlation coefficients (byan's test) for the similarity of the expression of each gene to that of ANCCA expression and other information are indicated in Supplementary Fig. S9.R status is indicated below by blue bars for ER-positive tumors (a total of 134) or red bars for ER-negative tumors (a total of 64). Right,MDA-MB468 cellsnsfected with siRNA targeting ANCCA (dark blue) or control sequence (yellow) and harvested 48 hours later for real-time RT-PCR analysis. Relativept levels were obtained by normalization of expression units for each gene with that of GAPDH. B, MDA-MB 468 cells were treated as above andd by RT-PCR or Western blotting. C, the anti-ANCCA and anti-EZH2 immunoreactivity of a core in the TMA section was scored negative if <10% ofhelial cells displayed any staining, and scored positive if >10% of the epithelial cells displayed staining with moderate to high intensity. The associationn the different antibody staining was analyzed using Pearson's χ2 test. D, ChIP with control serum and anti-ANCCA antibody was performed with
are presented as percentage of input.
Cancer Res; 70(22) November 15, 2010 9409
010 American Association for Cancer Research.

cell prnositidelevatesurvivproproANCCa partas a dgenes.ANCCIt is
examiovereximpormajoror othand cyin thisgenickinesiwith Akinesiassemfunctipeuticpromogethersurvivthat ofor intgramsthe mof thehighlyDes
induceproteithat hthe ERfrom sover, osets pthe ovprimalatedResulANCCand osanismfurthenegatiand fuderegutrol ismay nANCCgenes
Sevinvolvimplicas ACTbreastrevealand Pstimuand hdependistincoverexsion, hthe tra diffecent sexpreEZH2compcancecancenotypuniquof expB-MybANCCgramscoregunismof mu(thisthat Aactivifunctimay rstructexpresies areas a n
Discl
No p
Ackn
WeYang fo
Grant
NIHis a recPredoctof a NIH
Theof pageaccorda
Kalashnikova et al.
Cance9410
Dow
Published OnlineFirst September 23, 2010; DOI: 10.1158/0008-5472.CAN-10-1199
oliferation genes (29), but instead is part of the phosphoi-e 3-kinase signature (30), suggesting that, in some cases,d ANCCAmay function primarily to promote cancer cellal. Our data from cell culture studies indicate that bothliferation and prosurvival genes are indeed controlled byA. Thus, ANCCA overexpression may not only constituteof the gene signature with prognostic value, but also actriving force for the altered expression of the signatureAdditional studies are needed to determine whetherA coordinates the tumor signature gene programs.worth noting that in one of the microarray data setsned (12), the 20 genes most tightly associated withpressed ANCCA (i.e., based on correlation tests) playtant roles in mitosis and/or cell proliferation, theity of which were found overexpressed in breast cancerer types of cancers. When deregulated, Top2A, cdc2,clin E, as well as the other targets of ANCCA identifiedstudy (e.g., cdc6, B-Myb and EZH2), can display onco-activities. Intriguingly, four members of the humann family (a total of 45 genes) were co-overexpressedNCCA and were validated as ANCCA targets. Thesens function in different stages of mitosis for spindlebly and chromosome segregation (31, 32). The aberrantons of kinesins in cancer render them attractive thera-targets (31, 33), or in certain circumstances, mayte cancer cell resistance to taxane-based drugs (34). To-with the role of ANCCA in mediating expression of pro-al genes (VEGF, IRS2, SGK, and Akt), our data suggestverexpressed ANCCA may function as a dominant nodeegrating and/or eliciting multiple gene expression pro-to promote breast cancer progression. Deregulation ofultiple pathways identified here could constitute a partmolecular basis for the observed association betweenelevated ANCCA protein and poor patient outcome.pite our initial identification of ANCCA as a hormone-d gene (8, 9), we show here that high levels of ANCCAn associate most strongly with TNBCs. We also showigh transcript levels of ANCCA tend to associate withα-negative or triple-negative status in several data setstudies of multiple cohorts of tumors (19, 26, 35). More-ur unbiased analysis of tumor gene expression dataoints to the association of high levels of ANCCA witherexpression of other proliferation genes occurringrily in ERα-negative tumors. How ANCCA is deregu-in ERα-negative breast cancers is currently unclear.ts obtained by Ciro and colleagues reported thatA is regulated by the pRb-E2F pathway in fibroblaststeosarcoma cells (17). Whether this is the major mech-for ANCCA overexpression in breast cancer awaitsr analysis. Given that one salient feature of triple-ve/basal-like tumors is their high proliferation indexnctional loss of the pRb-E2F pathway (1, 16, 35, 36),lation of ANCCA through loss of pRb-mediated con-an attractive possibility. On the other hand, ANCCAot be merely a downstream target of pRb-E2F because
A itself mediates expression of multiple Rb-E2F targetcritical for TNBC cell proliferation.ReceOnlineF
r Res; 70(22) November 15, 2010
on July 28, 2018. © 2cancerres.aacrjournals.org nloaded from
eral transcriptional coregulators, including the onesed in steroid hormone signaling, have been stronglyated in human malignancies (37–42). AIB1 (also knownR/SRC-3) was initially identified as a gene amplified incancer and a coactivator for ERα (43). Later studiesed that its overexpression does not correlate with ERR status and that its aberrant function may includelation of tumor growth of both hormone-dependentormone-independent breast cancers through ER-dent and ER-independent pathways (44–48). Onetion between ACTR and ANCCA is that whereas ACTRpression correlates strongly with high HER2 expres-igh levels of ANCCA do not, but instead associate withiple-negative status. Although such distinction impliesrent mechanism for their aberrant expression, our re-tudies suggest that ANCCA and ACTR may regulate thession of each other in certain circumstances (7, 8)., the histone methyltransferase subunit of the PRC2lex, is frequently overexpressed in multiple types ofr (15, 49). Like ANCCA, EZH2 overexpression in breastr correlates with high proliferation and basal-like phe-e and tumor invasiveness (14). ANCCA, however, ise in that it seems to play a critical role in the controlression of proproliferation genes including EZH2 andin triple-negative cancers. Thus, our data suggest thatA may act as an integrator of several oncogenic pro-. Given our finding that ANCCA is a transcriptionallator, it is reasonable to predict that a major mecha-of overexpressed ANCCA in breast cancer is alterationltiple gene networks, including those of EZH2, pRb-E2Fstudy), or c-Myc (17). Our previous study showedNCCA is an AAA+ ATPase protein and that its ATPasety is required for its transcriptional stimulationon (8). ANCCA also possesses a bromodomain thatecognize a distinct histone modification. Given theseure-function features of ANCCA and its aberrantsion in multiple types of human cancers, further stud-warranted to exploit the potential of aberrant ANCCAew prognostic marker and a therapeutic target.
osure of Potential Conflicts of Interest
otential conflicts of interest were disclosed.
owledgments
thank Dr. Laurel Beckett for advice on statistical analysis, and Pingr technical assistance on IHC analysis.
Support
grants R01CA113860 and R01DK060019 (H.W. Chen). E.V. Kalashnikovaipient of a Department of Defense Breast Cancer Research Programoral Fellowship Award (W81XWH-0810689). N.P. Andrews is a recipientT32 training grant.
costs of publication of this article were defrayed in part by the paymentcharges. This article must therefore be hereby marked advertisement innce with 18 U.S.C. Section 1734 solely to indicate this fact.
ived 04/06/2010; revised 07/29/2010; accepted 08/24/2010; publishedirst 09/23/2010.
Cancer Research
010 American Association for Cancer Research.

Refe1. Sc
can80
2. Pebre
3. Soterwit10
4. Lahandcan
5. O'Mbio
6. Xuthe200
7. HsHWonRe
8. ZoestforAc
9. ZoANcan
10. Locelau200
11. vaning530
12. DegeninCa
13. ThInOn
14. Gogroreq
15. Klesivbre116
16. Heretlike10:
17. CirMYRe
18. vannat347
19. WadisLan
20. Bilhum353
21. Dedirde61
ANCCA Is a New Prognostic Factor for Breast Cancer
www.a
Dow
Published OnlineFirst September 23, 2010; DOI: 10.1158/0008-5472.CAN-10-1199
22. BothatheLa
23. Licoca
24. Foexpri
25. Hanomi
26. IvshisCa
27. Mahu10
28. TeproGe
29. Chanpre10
30. Saasssu75
31. Huas19
32. TanMeass
33. TaproCa
34. DesinRe
35. Gaceev47
36. Bomo20
37. QinlatCa
38. Ohtor81
39. Shga20
40. ToanAIB
41. Reesenbr20
renceshneider BP, Winer EP, Foulkes WD, et al. Triple-negative breastcer: risk factors to potential targets. Clin Cancer Res 2008;14:
10–8.rou CM, Sorlie T, Eisen MB, et al. Molecular portraits of humanast tumours. Nature 2000;406:747–52.rlie T, Perou CM, Tibshirani R, et al. Gene expression patns of breast carcinomas distinguish tumor subclassesh clinical implications. Proc Natl Acad Sci U S A 2001;98:869–74.usen T, Henke RT, Kagan BL, Wellstein A, Riegel AT. The roleregulation of the nuclear receptor co-activator AIB1 in breastcer. Breast Cancer Res Treat 2009;116:225–37.alley BW, Kumar R. Nuclear receptor coregulators in cancer
logy. Cancer Res 2009;69:8217–22.J, Wu RC, O'Malley BW. Normal and cancer-related functions ofp160 steroid receptor co-activator (SRC) family. Nat Rev Cancer9;9:615–30.ia EY, Kalashnikova EV, Revenko AS, Zou JX, Borowsky AD, Chen. Deregulated E2F and the AAA+ coregulator ANCCA drive proto-
cogene ACTR/AIB1 overexpression in breast cancer. Mol Cancers 2010;8:183–93.u JX, Revenko AS, Li LB, Gemo AT, Chen HW. ANCCA, anrogen-regulated AAA+ ATPase coactivator for ERα, is requiredcoregulator occupancy and chromatin modification. Proc Natlad Sci U S A 2007;104:18067–72.u JX, Guo L, Revenko AS, et al. Androgen-induced coactivatorCCA mediates specific androgen receptor signaling in prostatecer. Cancer Res 2009;69:3339–46.
uie MC, Revenko AS, Zou JX, Yao J, Chen HW. Direct control ofl cycle gene expression by proto-oncogene product ACTR, itstoregulation underlies its transforming activity. Mol Cell Biol6;26:3810–23.'t Veer LJ, Dai H, van de Vijver MJ, et al. Gene expression profil-predicts clinical outcome of breast cancer. Nature 2002;415:–6.smedt C, Piette F, Loi S, et al. Strong time dependence of the 76-e prognostic signature for node-negative breast cancer patientsthe TRANSBIG multicenter independent validation series. Clinncer Res 2007;13:3207–14.orner AR, Hoadley KA, Parker JS, Winkel S, Millikan RC, Perou CM.vitro and in vivo analysis of B-Myb in basal-like breast cancer.cogene 2009;28:742–51.nzalez ME, Li X, Toy K, et al. Downregulation of EZH2 decreaseswth of estrogen receptor-negative invasive breast carcinoma anduires BRCA1. Oncogene 2009;28:843–53.er CG, Cao Q, Varambally S, et al. EZH2 is a marker of aggres-e breast cancer and promotes neoplastic transformation ofast epithelial cells. Proc Natl Acad Sci U S A 2003;100:06–11.rschkowitz JI, He X, Fan C, Perou CM. The functional loss of theinoblastoma tumour suppressor is a common event in basal-and luminal B breast carcinomas. Breast Cancer Res 2008;
R75.o M, Prosperini E, Quarto M, et al. ATAD2 is a novel cofactor forC, overexpressed and amplified in aggressive tumors. Cancers 2009;69:8491–8.de Vijver MJ, He YD, van't Veer LJ, et al. A gene-expression sig-ure as a predictor of survival in breast cancer. N Engl J Med 2002;:1999–2009.ng Y, Klijn JG, Zhang Y, et al. Gene-expression profiles to predicttant metastasis of lymph-node-negative primary breast cancer.cet 2005;365:671–9.
d AH, Yao G, Chang JT, et al. Oncogenic pathway signatures inan cancers as a guide to targeted therapies. Nature 2006;439:–7.blois G, Hall JA, Perry MC, et al. Genome-wide identification of
ect target genes implicates estrogen-related receptor α as aterminant of breast cancer heterogeneity. Cancer Res 2009;69:49–57.42. HsSRBio
acrjournals.org
on July 28, 2018. © 2cancerres.aacrjournals.org nloaded from
nnefoi H, Potti A, Delorenzi M, et al. Validation of gene signaturest predict the response of breast cancer to neoadjuvant chemo-rapy: a substudy of the EORTC 10994/BIG 00-01 clinical trial.ncet Oncol 2007;8:1071–8.Y, Zou L, Li Q, et al. Amplification of LAPTM4B and YWHAZntributes to chemotherapy resistance and recurrence of breastncer. Nat Med 2010;16:214–8.ekens JA, Atkins D, Zhang Y, et al. Multicenter validation of a genepression-based prognostic signature in lymph node-negativemary breast cancer. J Clin Oncol 2006;24:1665–71.ibe-Kains B, Desmedt C, Piette F, et al. Comparison of prog-stic gene expression signatures for breast cancer. BMC Geno-cs 2008;21:394.hina AV, George J, Senko O, et al. Genetic reclassification oftologic grade delineates new clinical subtypes of breast cancer.ncer Res 2006;66:10292–301.XJ, Salunga R, Tuggle JT, et al. Gene expression profiles of
man breast cancer progression. Proc Natl Acad Sci U S A 2003;0:5974–9.schendorff AE, Naderi A, Barbosa-Morais NL, et al. A consensusgnostic gene expression classifier for ER positive breast cancer.nome Biol 2006;7:R101.ang HY, Nuyten DS, Sneddon JB, et al. Robustness, scalability,d integration of a wound-response gene expression signature indicting breast cancer survival. Proc Natl Acad Sci U S A 2005;2:3738–43.al LH, Johansson P, Holm K, et al. Poor prognosis in carcinoma isociated with a gene expression signature of aberrant PTEN tumorppressor pathway activity. Proc Natl Acad Sci U S A 2007;104:64–9.szar D, Theoclitou ME, Skolnik J, Herbst R. Kinesin motor proteinstargets for cancer therapy. Cancer Metastasis Rev 2009;28:7–208.enbaumME,Macurek L, JanssenA,Geers EF,Alvarez-FernandezM,dema RH. Kif15 cooperates with eg5 to promote bipolar spindleembly. Curr Biol 2009;19:1703–11.niwaki M, Takano A, Ishikawa N, et al. Activation of KIF4A as agnostic biomarker and therapeutic target for lung cancer. Clinncer Res 2007;13:6624–31.S, Cipriano R, Jackson MW, Stark GR. Overexpression of kine-s mediates docetaxel resistance in breast cancer cells. Cancers 2009;69:8035–42.uthier ML, Berman HK, Miller C, et al. Abrogated response tollular stress identifies DCIS associated with subsequent tumorents and defines basal-like breast tumors. Cancer Cell 2007;12:9–91.sco EE, Wang Y, Xu H, et al. The retinoblastoma tumor suppressordifies the therapeutic response of breast cancer. J Clin Invest07;117:218–28.L, Liu Z, Chen H, Xu J. The steroid receptor coactivator-1 regu-
es twist expression and promotes breast cancer metastasis.ncer Res 2009;69:3819–27.shiro K, Rayala SK, Kondo S, et al. Identifying the estrogen recep-coactivator PELP1 in autophagosomes. Cancer Res 2007;67:
64–71.en H, Powers N, Saini N, et al. The SWI/SNF ATPase Brm is atekeeper of proliferative control in prostate cancer. Cancer Res08;68:10154–62.rres-Arzayus MI, de Mora JF, Yuan J, et al. High tumor incidenced activation of the PI3K/AKT pathway in transgenic mice define1 as an oncogene. Cancer Cell 2004;6:263–74.dmond AM, Bane FT, Stafford AT, et al. Coassociation oftrogen receptor and p160 proteins predicts resistance todocrine treatment; SRC-1 is an independent predictor ofeast cancer recurrence. Cl in Cancer Res 2009;15:98–106.
ia E, Zou JX, Chen HW. The roles and action mechanisms of p160/C coactivators and the ANCCA coregulator in cancer. Prog Moll Transl Sci 2009;87:261–98.Cancer Res; 70(22) November 15, 2010 9411
010 American Association for Cancer Research.

43. Anco27
44. Yaprotur20
45. Qincanpro593
46. Lotio
pro51
47. Heexco51
48. FetoracRe
Kalashnikova et al.
Cance9412
Dow
Published OnlineFirst September 23, 2010; DOI: 10.1158/0008-5472.CAN-10-1199
zick SL, Kononen J, Walker RL, et al. AIB1, a steroid receptoractivator amplified in breast and ovarian cancer. Science 1997;7:965–8.n J, Erdem H, Li R, et al. Steroid receptor coactivator-3/AIB1motes cell migration and invasiveness through focal adhesionnover and matrix metalloproteinase expression. Cancer Res08;68:5460–8.L, Liao L, Redmond A, et al. The AIB1 oncogene promotes breastcer metastasis by activation of PEA3-mediated matrix metallo-teinase 2 (MMP2) and MMP9 expression. Mol Cell Biol 2008;28:
7–50.uie MC, Zou JX, Rabinovich A, Chen HW. ACTR/AIB1 func-ns as an E2F1 coactivator to promote breast cancer cell49. Bradoam
r Res; 70(22) November 15, 2010
on July 28, 2018. © 2cancerres.aacrjournals.org nloaded from
liferation and antiestrogen resistance. Mol Cell Biol 2004;24:57–71.ck S, Rom J, Thewes V, et al. Estrogen-related receptor αpression and function is associated with the transcriptionalregulator AIB1 in breast carcinoma. Cancer Res 2009;69:86–93.reshteh MP, Tilli MT, Kim SE, et al. The nuclear receptor coactiva-amplified in breast cancer-1 is required for Neu (ErbB2/HER2)
tivation, signaling, and mammary tumorigenesis in mice. Cancers 2008;68:3697–706.cken AP, Pasini D, Capra M, Prosperini E, Colli E, Helin K. EZH2 is
wnstream of the pRB-E2F pathway, essential for proliferation andplified in cancer. EMBO J 2003;22:5323–35.Cancer Research
010 American Association for Cancer Research.

2010;70:9402-9412. Published OnlineFirst September 23, 2010.Cancer Res Ekaterina V. Kalashnikova, Alexey S. Revenko, Abigael T. Gemo, et al. EZH2Survival of Triple-Negative Cells through Control of B-Myb andPatients with Poor Prognosis, Acting to Drive Proliferation and ANCCA/ATAD2 Overexpression Identifies Breast Cancer
Updated version
10.1158/0008-5472.CAN-10-1199doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2010/09/24/0008-5472.CAN-10-1199.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/70/22/9402.full#ref-list-1
This article cites 49 articles, 29 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/70/22/9402.full#related-urls
This article has been cited by 12 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/70/22/9402To request permission to re-use all or part of this article, use this link
on July 28, 2018. © 2010 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 23, 2010; DOI: 10.1158/0008-5472.CAN-10-1199