aac accepts, published online ahead of print on 2 may...
TRANSCRIPT

Induction of efflux-mediated macrolide resistance in Streptococcus pneumoniae 1
2
Scott Chancey1,3
, Xiaoliu Zhou1,3§
, Dorothea Zähner1,3
, David S. Stephens1,2,3
* 3
4
5
1 Division of Infectious Diseases, Department of Medicine 6
2 Department of Microbiology and Immunology, Emory University School of Medicine, 7
Atlanta, GA 30322 8
3 Department of Veterans Affairs Medical Center Atlanta, GA 30033 9
10
11
12
13
Running title: Inducers of macrolide resistance in S. pneumoniae 14
15
16
§ Present address: Centers for Disease Control and Prevention, 4770 Buford Highway, Chamblee, GA 17
30341, USA 18
19
20
21
Copyright © 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Antimicrob. Agents Chemother. doi:10.1128/AAC.00060-11 AAC Accepts, published online ahead of print on 2 May 2011
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

Abstract 22
The antimicrobial efflux system encoded by the operon mefE/mel on the mobile genetic element 23
MEGA in Streptococcus pneumoniae and other Gram-positive bacteria is inducible by macrolide 24
antibiotics and antimicrobial peptides. Induction may affect the clinical response to the use of 25
macrolides. We developed mefE reporter constructs and a disk diffusion induction and resistance assay 26
to determine the kinetics and basis of mefE/mel induction. Induction occurred rapidly, >15-fold increase 27
in transcription within one hour of exposure to sub-inhibitory concentrations of erythromycin. A 28
spectrum of environmental conditions including competence and non-macrolide antibiotics with distinct 29
cellular targets did not induce mefE. Using sixteen different structurally-defined macrolides, induction 30
was correlated with the amino sugar attached to C5 of the macrolide lactone ring, not with the size (e.g., 31
14-, 15- or 16-member) of the ring or with the presence of the neutral sugar cladinose at C3. Macrolides 32
with a monosaccharide attached to C5, known to block exit of the nascent peptide from the ribosome 33
after the incorporation of up to eight amino acids, induced mefE expression. Macrolides with a C5 34
disaccharide, which extends the macrolide into the ribosomal exit tunnel disrupting peptidyl transferase 35
activity, did not induce. The induction of mefE did not require macrolide efflux, but the affinity of 36
macrolides for the ribosome determined availability for efflux and pneumococcal susceptibility. The 37
induction of mefE/mel expression by inducing macrolides appears to be based on specific interactions of 38
the macrolide C5 saccharide with the ribosome alleviating transcriptional attenuation of mefE/mel. 39
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

Introduction 40
Macrolides are broad spectrum antibiotics with complex macrocyclic 14-, 15- or 16-member 41
lactone rings that bind in the peptide exit tunnel of bacterial ribosomes and inhibit protein synthesis. 42
Macrolides are often recommended as the empirical first-line treatment of upper respiratory bacterial 43
infections including pneumococcal infections and community acquired pneumonia. However, bacterial 44
resistance to macrolides is expanding worldwide in Gram-positive bacteria and is now present in almost 45
a third of all invasive Streptococcus pneumoniae isolates (14). The two most common mechanisms of 46
macrolide resistance in bacterial pathogens are modification of the bacterial ribosome, either by 47
methylation or mutation, and extrusion of the drugs from the bacterial cell by an efflux pump. Genes of 48
the erm (erythromycin ribosomal methylases) family of rRNA methylases confer high-level resistance to 49
lincosamides and streptogramins as well as macrolides (the MLSB phenotype) and can be constitutive or 50
inducible. In the inducible form resistance develops only after exposure of the bacterium to the 51
macrolide. 52
The best-studied mechanism of inducible MLSB resistance involves the ermC gene found in S. 53
aureus and other Gram-positive pathogens (4, 32, 33). Translation of ermC is attenuated in the absence 54
of inducers due to secondary structures that render the ribosomal binding site inaccessible. Inducer-55
bound ribosomes stall during translation of a 19-amino acid leader peptide upstream of ermC resulting 56
in the refolding of the transcript to make available the RBS thus promoting translation (32). A later 57
comparison of inducers for ermC and ermSV in Streptomyces viridochromogenes showed that the 58
specific sequence of leader peptide determines the range of inducers (24). Macrolides with 14-59
membered rings and C3 cladinose induced ermC, and 16-membered macrolides induced ermSV (24). 60
The lincosamide celesticetin induced both ermC and ermSV demonstrating a common inducer between 61
the otherwise disparately induced genes (24). Attenuation of erm genes can also occur at the level of 62
transcription such as with ermK of Bacillus licheniformis (28, 29). Transcriptional attenuation occurs 63
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

when secondary structures in the transcript function like rho-independent terminators to terminate 64
transcription short of the erm gene coding region. 65
Efflux-mediated macrolide-resistance was first reported in Staphylococcus epidermidis and later 66
in Streptococcus pyogenes and S. pneumoniae (30, 40, 45). In S. epidermidis macrolide efflux is 67
conferred by msrA and is induced by the 14- and 15-membered macrolides clarithromycin and 68
azithromycin and the ketolide telithromycin, but not by streptogramin B, even though the latter is a 69
substrate for MsrA-mediated efflux. In streptococci, macrolide efflux is conferred by proteins encoded 70
by mefE, mefA or less commonly mefI, that share >90% identity. The mef genes are transcribed as an 71
operon with the msrA homolog mel, comprising a dual efflux pump (3, 12). In S. pneumoniae, mefE/mel 72
is carried on the small mobile element MEGA, the predominant macrolide efflux determinant in the 73
pneumococcus (15, 43). MefE/Mel confers in vitro moderate-level resistance (1-32 µg/ml) to 14- and 74
15-membered macrolides, but reportedly not 16-membered macrolides, lincosamides, or streptogramins 75
(M phenotype) (45, 47). MefE/Mel-mediated resistance has been shown to be induced by the 14- and 76
15-membered macrolides erythromycin, clarithromycin and azithromycin, but not by 16-membered 77
macrolides (3, 50). Ketolides, such as telithromycin, are considered poor inducers of mefE/mel and 78
retain antimicrobial activity against pneumococcal strains carrying these genes (34, 50). 79
In vivo exposure to macrolides or other potential inducers may result in resistance higher than 80
predicted by in vitro determined MIC’s. We recently reported that certain antimicrobial peptides 81
(AMP), including the human AMP LL-37, activated transcription of mefE/mel and resulted in induced 82
resistance to the macrolide erythromycin (51). This suggests that the MefE/Mel efflux pump can be 83
induced by human host defenses and therefore be primed prior to clinically administered macrolides. 84
The goals of this study were to define the range of compounds, macrolides as well as other antibiotics 85
and other conditions, that induce MefE/Mel-mediated resistance; to elucidate the kinetics of induction; 86
to determine the structural features of macrolides required for induction and to develop a model for the 87
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

mechanism of induction. To accomplish these goals, mefE-lacZ reporter fusion constructs and a disk 88
diffusion-based assay were developed that allowed assessment of both the induction of mefE/mel and of 89
efflux-mediated macrolide resistance. 90
Methods 91
Bacterial strains. Construction of the mefE-lacZ reporter strain XZ7042 and the MEGA element 92
deletion derivative XZ8004 from the erythromycin resistant, MEGA-containing S. pneumoniae clinical 93
isolate GA17457 was as previously described (51). The negative control strains XZ7049 was generated 94
by insertion of the promoterless lacZ of pPP2 (18) into bgaA of GA17457. Likewise, XZ7067 was 95
generated by insertion of the comC-lacZ transcriptional fusion on plasmid pPC2 (18) into GA17457. 96
Quantitative β-galactosidase assays. The rate of induction of mefE by erythromycin (ERY) was 97
determined by adding 0.1 µg/ml erythromycin to mid-log phase cultures (OD600~0.3-0.4). Test strains 98
were incubated at 37°C in parallel with uninduced cultures. At 30 minute intervals, cultures were 99
sampled for assessment of β-galactosidase activity as described previously (35). To determine a 100
transcriptional dose response of mefE-lacZ, parallel cultures of XZ7042 and XZ8004 were grown to 101
mid-log phase (OD600 ~0.3-0.4) and exposed to concentrations of erythromycin varying by four orders 102
of magnitude or more. Each subculture was harvested one hour after induction for assessment of β-103
galactosidase activity. All experiments were performed in duplicate and assay readings were taken in 104
triplicate. 105
Antibiotics. Antibiotic susceptibility disks were either obtained commercially or prepared by 106
application of stock solutions to sterile blank disks, followed by drying for fifteen minutes in a laminar 107
flow hood. Susceptibility disks containing the macrolides azithromycin, clarithromycin, erythromycin, 108
telithromycin and tilmicosin were purchased from Becton, Dickinson and Company (Franklin Lakes, 109
NJ). Disks were prepared for the macrolides dirithromycin, josamycin, midecamycin, roxithromycin, 110
and spiramycin (Sigma-Aldrich, St. Louis, MO); and kitasamycin (MP Biomedicals, Solon, OH), 111
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

oleandomycin (Crescent Chemical Co., Inc., Islandia, NY), troleandomycin (Enzo Life Sciences, 112
Plymouth Meeting, PA) and tylosin (Wako Pure Chemical Industries, Ltd, Osaka, Japan). The 113
macrolide tulathromycin was obtained as the animal health product Draxxin® from Pfizer Animal 114
Health (Kalamazoo, MI). Commercially prepared antibiotic disks were obtained containing bacitracin, 115
clindamycin, colistin, levofloxacin, lincomycin, polymyxin B, and trimethoprim/sulfmethoxazole 116
(Becton, Dickinson and Company, Franklin Lakes, NJ) and quinupristin/dalfopristin (Remel, Lenexa, 117
KS). Disks were prepared as described above for the non-macrolide antibiotics amoxicillin, ampicillin, 118
penicillin, cefotaxime, ceftriaxone, cefuroxime, ciprofloxacin, chloramphenicol, tetracycline, 119
gentamycin, kanamycin, spectinomycin, rifampin and vancomycin (Sigma-Aldrich, St. Louis, MO), 120
mupirocin (AppliChem GmbH, Darmstadt, Germany). Cyclopentadecanolide was obtained from Acros 121
Organics (Geel, Belgium). 122
Susceptibility assays. The minimum inhibitory concentration (MIC) of erythromycin was determined 123
by Etest as per manufacturer recommendations (AB bioMerieux, Solna, Sweden). Briefly, strains 124
incubated overnight at 37ºC in 5% CO2 on trypticase soy agar with 5% sheep blood (TSA-SB) (Becton, 125
Dickinson and Company, Franklin Lakes, NJ) were suspended in Mueller-Hinton broth to a 0.5 126
McFarland Standard and spread onto Mueller-Hinton agar with 5% sheep blood (MH-SB) (Becton, 127
Dickinson and Company, Franklin Lakes, NJ). E-test strips were placed on the plate surface and the 128
plates were incubated overnight at 37ºC in 5% CO2. Susceptibility to other macrolides was determined 129
by disk diffusion in accordance with the standards described by the Clinical and Laboratory Standards 130
Institute (11). To determine the effects of erythromycin induction on resistance, MICs were determined 131
by E-test as described above except strains were incubated on TSA-SB supplemented with 1/10 MIC 132
erythromycin prior to suspension and plating on MH-SB. Susceptibility to other macrolides was 133
determined by disk diffusion in accordance with the standards described by the Clinical and Laboratory 134
Standards Institute (11). Susceptibility disks were purchased or prepared as described above. All 135
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

susceptibility data represents the mean ± standard deviation of at least five independent replications and 136
were analyzed by two-tailed, paired T-tests. p values > 0.05 were considered significant. 137
Disk diffusion assay for mefE-lacZ induction. The mefE-lacZ reporter strain XZ7042 was grown to 138
mid-log phase in Todd-Hewitt broth supplemented with 0.5% yeast extract. The liquid culture was 139
swabbed onto trypticase soy agar (TSA) supplemented with 300 U/ml catalase (Sigma-Aldridge, St. 140
Louis, MO) and 0.032 % X-gal. Susceptibility disks infused with test compounds were placed 141
immediately on the plates and incubated at 37ºC, 5 % CO2 for 24 h. Induction by synthetic competence-142
stimulating peptide-1 (CSP-1) (20) was tested by spotting 15 µl of a 100 ng/ml solution directly to the 143
center of indicator plate swabbed with the indicated strain. CSP-1 was synthesized by the Emory 144
University Microchemical Facility. 145
Results 146
Characterization of erythromycin-induced expression of mefE/mel. The kinetics and dose-147
dependence of mefE/mel induction were determined using the prototype macrolide erythromycin and a 148
series of isogenic reporter strains that contained a mefE-lacZ transcriptional fusion in the S. pneumoniae 149
strain GA17457 [Table 1, (51)]. The “wild type” reporter strain XZ7042 contained a functional MEGA 150
element encoding the MefE/Mel efflux pump that confers resistance to erythromycin (MIC 12 µg/ml), 151
while the MEGA-deletion derivative, XZ8004, was erythromycin-susceptible (MIC 0.1 µg/ml) (51). A 152
promoterless lacZ-reporter strain, XZ7049, which was otherwise identical to XZ7042, served as a 153
negative control. The induction of mefE transcription in XZ7042, expressed as β-galactosidase (β-gal) 154
activity, was measured over time following the addition of subinhibitory concentrations of erythromycin 155
(1/10 of the MIC). β-gal activity increased rapidly upon addition of erythromycin to more than 7-fold 156
within the first 30 minutes post-induction (Fig. 1a). Maximal induction (>15-fold increase) was 157
obtained at 1 hour (Fig. 1a). β-gal activities of XZ7042 cultures without induction were equivalent to 158
those of the induced and uninduced promoterless negative control, indicating a very low level of 159
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

constitutive expression from the mefE-promoter in the absence of an inducer (Fig.1a). 160
The role of a functional MEGA element in mefE induction was assessed by measuring the 161
erythromycin dose-response in XZ7042 and the MEGA deletion derivative XZ8004. The strains were 162
incubated with erythromycin concentrations beginning with 0.0012 µg/ml and increasing by one-half or 163
one-quarter log intervals (Figure 1b). In XZ8004 mefE was induced at lower erythromycin 164
concentrations than in XZ7042 (0.012 and 0.04 µg/ml, respectively) and with higher maximal 165
expression (205.3 MU and 106.9 MU, respectively, Fig. 1b). However, peak expression of mefE in 166
XZ8004 occurred at a concentration equivalent to the erythromycin MIC (0.12 µg/ml) of the strain, 167
while in XZ7042 peak expression was observed at a concentration (0.4 µg/ml) 30-fold less than the 168
strains erythromycin MIC (12 µg/ml) (Fig. 1b). Expression in XZ8004 decreased rapidly with 169
increasing erythromycin concentrations above the MIC (Fig. 1b), consistent with an inhibitory effect of 170
erythromycin. The mefE-lacZ induction in XZ8004 indicated that a functional MEGA element was not 171
required for the induction of mefE/mel; however, strain susceptibility to the inducer and the ability to 172
modify intracellular concentrations of the inducer contributed to the magnitude and duration of 173
mefE/mel induction. 174
To evaluate mefE/mel expression over a range of inducers and inducer concentrations, a disk 175
diffusion assay using the XZ7042 reporter strain was developed (42). In contrast to the quantitative β-176
gal assays, this approach provided a concentration gradient, allowing the detection of mefE-lacZ 177
induction without a priori knowledge of the MIC of the test compound or the time-dependent induction 178
kinetics and was suitable for study of a broad range of potential inducers. Fig. 1c shows the results of 179
the disk diffusion assays using erythromycin as the test compound with the “wild type” reporter 180
XZ7042, the MEGA-deletion strain XZ8004, and the promoter-less control strain XZ7049. Strain 181
XZ7042 showed a characteristic large blue halo, indicative of the induction of mefE-lacZ, surrounding a 182
narrow zone of inhibited growth (Fig. 1c). As anticipated, the MEGA deletion strain XZ8004 showed a 183
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

narrow range of induction, surrounding a large zone of inhibition (Fig. 1c). This result was consistent 184
with the quantitative assay (Fig. 1b), showing a wide range of inducing concentrations for XZ7042 and a 185
strong but narrow range of futile induction of mefE in strain XZ8004 in the absence of the resistance 186
determinant MEGA and the efflux pump (Fig. 1b). The promoterless-lacZ strain XZ7049 showed the 187
same zone of inhibition as XZ7042, as expected due to a functional MEGA element; however, no β-gal 188
activity (i.e., blue halo) was detected (Fig. 1c), confirming that the mefE-lacZ-expression in the reporter 189
strain XZ7042 exclusively depended on mefE. 190
Environmental or antibiotic (non-macrolide) stress and competence did not induce mefE-191
lacZ. To assess the spectrum of inducers of mefE/mel expression, different environmental conditions 192
and antibiotic compounds were analyzed in the disk diffusion assay using XZ7042. Non-macrolide 193
antibiotics were chosen to include representatives of major antimicrobial classes with diverse cellular 194
targets, thus providing a wide range of antibiotic stresses. Also tested were environmental conditions 195
involved in regulation of other genetic systems, as well as known substrates for multi-drug efflux 196
systems. None of the tested compounds and conditions inducted mefE-lacZ expression, with the 197
exception of the previously described antimicrobial peptides LL-37, CRAMP38, CRAMP39 (51). These 198
results demonstrated that mefE/mel induction is restricted to a narrow range of compounds, suggesting 199
one or more well-defined mechanism(s) of induction. 200
In addition, competence was tested as potentially inducing condition of mefE induction. 201
Competence induction in S. pneumoniae has been known to affect expression of a broad range of genes 202
and is mediated by the com regulon (1, 2, 21, 31). Prudhomme et al. (39) showed that the com regulon 203
was induced in response to antibiotic stress. Since it was unknown whether the conditions of the disk 204
diffusion assay supported the development of natural competence, synthetic competence-stimulating 205
peptide (CSP-1) was used to induce competence and monitor the effect on mefE-lacZ induction in the 206
lacZ-disk diffusion assay. Figure 2 shows that erythromycin, but not CSP-1, induced mefE-lacZ in 207
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

XZ7042. As a control for competence development, a reporter strain, XZ7067, which carried the lacZ 208
gene under the control of the promoter for the early competence gene comC, was constructed (Table 1). 209
XZ7067 was induced by CSP-1 confirming competence induction under the assay conditions. 210
Erythromycin did not induce comC (Fig. 2), which is consistent with the previous observation (36) that 211
erythromycin does not induce competence. Thus, general environmental and antibiotic stresses, as well 212
as the development of competence did not induce mefE/mel expression. 213
Induction of mefE by macrolides. To understand the relationship between macrolide structure 214
and mefE/mel induction, a panel of sixteen macrolides composed of 14-, 15- and 16-membered 215
compounds were studied using the disk diffusion assay. The 14-membered cladinosolides, compounds 216
with the lactone ring substituted with the sugar cladinose at C3 and the monosaccharide amino sugar 217
desosamine at C5 (Fig. 3a) (e.g., erythromycin, clarithromycin, dirithromycin and roxithromycin), each 218
strongly induced β-gal activity over a wide diffusion zone (i.e. concentration gradient) (Fig. 3a). 219
Induction was correlated with decreased susceptibility to these compounds in the MEGA-containing 220
reporter strain, but not in the susceptible MEGA-deletion strain (Fig. 3a and Table 2, p <0.001). In 221
addition, preincubation of the reporter strains with a subinhibitory concentration of erythromycin (1/10 222
MIC, 1.2 µg/ml) further increased the resistance to these compounds for the wild type MEGA 223
containing reporter strain (p <0.05, Table 2) background, but not for the MEGA-deletion strain. Thus, 224
resistance in the wild type strain required the efflux-encoding MEGA element and suggested that 225
erythromycin, clarithromycin, clarithromycin, and roxithromycin are substrates for MefE/Mel-mediated 226
efflux. These findings were consistent with earlier reports for 14- and 15-membered macrolides (3, 34, 227
50). 228
Two 14-membered macrolides oleandomycin and troleandomycin with substitution of oleandrose 229
for the cladinose in the lactone ring were tested (Fig. 3c). Induction of mefE-lacZ by oleandomycin 230
resembled the strong induction observed with the cladinosolides; however, troleandomycin was a poor 231
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

inducer of mefE (Fig. 3c). Troleandomycin has three acetate substitutions of the hydroxyl groups of 232
oleandomycin (Fig. 3c), suggesting that one or more of these acetate groups interfered with mefE 233
induction. The wild type reporter strain XZ7042 was significantly more susceptible to troleandomycin 234
than to oleandomycin (p <0.001, Table 2). To determine whether the increased susceptibility of XZ7042 235
for troleandomycin compared to oleandomycin was due to the weak mefE induction by troleandomycin, 236
the reporter strain XZ7042 was preincubated with a subinhibitory concentration of erythromycin (0.12 237
µg/ml) to achieve high induction of mefE/mel prior to susceptibility testing with troleandomycin. Under 238
these conditions, the resistance to troleandomycin increased to the levels of oleandomycin indicating 239
that troleandomycin was a substrate for efflux, despite being a poor mefE/mel inducer. 240
The ketolide telithromycin is a 14-membered macrolide that belongs to the ketolide subclass due 241
to a C3 ketone substitution instead of cladinose (Fig. 3b). Telithromycin retains excellent antimicrobial 242
activity of against MefE/Mel-containing pneumococci (12, 50). However, more detailed reports showed 243
that the efflux pump conferred small increases in pneumococcal MIC’s of telithromycin after 244
preincubation with subinhibitory concentrations of telithromycin (12, 50) or in comparison of clinical 245
isolates with and without mefE/mel (46). To determine whether the lack of TEL resistance was due to 246
an inefficient induction of mefE/mel expression or poor efflux of telithromycin by MefE/Mel, the 247
inducing ability of telithromycin was tested in the disk diffusion assay. Telithromycin strongly induced 248
mefE and the observed induction pattern was quite similar to that observed for the MEGA deletion 249
mutant XZ8004 with erythromycin (Fig. 1c), i.e., a narrow zone of strong mefE-induction surrounding a 250
large zone of inhibition of growth, which indicated that the wild type reporter strain remained 251
susceptible to telithromycin despite the strong induction of mefE-lacZ. This was confirmed by 252
telithromycin resistance (Table 2). As expected, preincubation with erythromycin had no significant 253
impact on telithromycin resistance although, the MEGA mutant was more susceptible to telithromycin 254
than the MEGA-containing strain (Table 2, p <0.001). These data suggest that telithromycin, while a 255
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

strong inducer of mefE/mel, was not sufficiently extruded by the MefE/Mel efflux pump to allow for 256
resistance to develop. 257
The two 15-membered macrolides, azithromycin and tulathromycin (Table 2, Fig. 3d), were 258
studied. Tulathromycin differs from azithromycin by a demethylation of N10 and by a propylamino side 259
chain substitution on the C3 cladinose (Fig. 3d). However, while mefE induction by azithromycin was 260
comparable to that of erythromycin, induction by tulathromycin was weaker. XZ7042 was slightly more 261
susceptible to tulathromycin than to azithromycin, suggesting that the increase in antimicrobial activity 262
of tulathromycin could be due to the weaker induction of mefE/mel. To test this, XZ7042 was 263
preincubated with erythromycin (1.2 µg/ml). Increased resistance to tulathromycin to the level of 264
azithromycin was noted. However, the weaker induction by TUL was still sufficient to render the 265
MEGA-containing strain XZ7042 less susceptible than the MEGA deletion strain XZ8004 (Table 2). 266
16-membered macrolides can induce mefE/mel expression. A current paradigm is that the 267
MefE/Mel efflux pump does not confer resistance to 16-membered macrolides (34). The inability of 16-268
membered macrolides to induce the efflux pump, structural feature(s) of the compounds that preclude 269
them as substrates for the efflux pump, or sequestration, e.g., at the ribosome, making these compounds 270
unavailable for efflux are possible explanations. Seven 16-membered macrolides, josamycin, 271
kitasamycin, midecamycin, rosamicin, spiramycin, tilmicosin and tylosin, were studied for induction of 272
mefE-lacZ (Figs. 4). Josamycin, kitasamycin, midecamycin, spiramycin and tylosin did not induce 273
mefE-lacZ expression (Fig. 4). Surprisingly, two 16-membered macrolides, tilmicosin and rosamicin 274
induced the mefE-lacZ reporter (Fig. 4b). All the16-membered macrolides lacked a cladinose at C3. 275
While the cladinose has been suggested as an important feature of inducing 14- and 15-membered 276
macrolides (8), the above noted 14- or 15- membered macrolides with modifications of the cladinose 277
(oleandomycin and troleandomycin) or with a ketone substitution (telithromycin) at C3 were also 278
inducers. In contrast, the common feature of the16-membered inducers (tilmicosin and rosamicin) and 279
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

the inducing 14- and 15-membered macrolides including telithromycin was the presence of a 280
monosaccharide at C5 (Fig. 4b). All non-inducing macrolides possessed a disaccharide moiety at this 281
position. The requirement of a monosaccharide at C5 was supported by the data with tylosin, a non- 282
inducing 16-membered macrolide structurally closely related to tilmicosin and rosamicin, but 283
distinguished by the disaccharide in the C5 position (Fig. 4b). Thus, the ability to induce mefE/mel 284
transcription was not dependent upon the size (e.g., 14-, 15- or 16-membered) of the lactone ring or the 285
presence of a cladinose at C3. Rather, mefE/mel expression was linked to a monosaccharide amino 286
sugar attached at C5 of the macrolide lactone ring. 287
Differential MefE/Mel-mediated resistance to 16-membered macrolides: further evidence 288
of uncoupling of induction and resistance. As observed for troleandomycin, which did not induce 289
mefE-lacZ but was readily effluxed upon preincubation with erythromycin, the susceptibility of 290
pneumococci to the non-inducing 16-membered macrolides could result from a lack of induction of the 291
efflux pump. Therefore, the wild type reporter XZ7042 was preincubated with erythromycin and 292
susceptibility to the non-inducing 16-membered compounds were determined (Table 2). The results did 293
not show increased resistance towards these 16-membered macrolides suggesting that they were not 294
subjected to efflux in the presence of the MefE/Mel pump. This was confirmed by double disk diffusion 295
assays for induction in which no D-zone of clearing was produced by the non-inducing macrolides in the 296
presence of erythromycin (data not shown). 297
Still remaining was the question of whether the induction of mefE observed with the inducing 16-298
membered macrolides rosamicin and tilmicosin translated into increased resistance to these compounds. 299
The strong, narrow induction ring observed in the disk-diffusion induction assay with tilmicosin 300
indicated a narrow range of inducing concentrations; however, the small zone of inhibition indicated 301
pneumococcal resistance to tilmicosin (Fig. 4b). Preincubation of XZ7042 with erythromycin resulted 302
in further resistance of tilmicosin compared to the uninduced control (Table 2), indicating that tilmicosin 303
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

was subjected to MefE/Mel-mediated efflux. Conversely, the induction of mefE by rosamicin (Fig. 4b), 304
resembled that observed with telithromycin (Fig. 3b) and the MEGA deletion strain XZ8004 (Fig. 1c), 305
and XZ7042 remained a susceptible phenotype despite induction (Table 2). Indeed, there was no change 306
in susceptibility to rosamicin between the MEGA-deletion strain XZ8004 and the wild type strain 307
XZ7042 despite the induction of mefE/mel (Table 2). 308
These data further demonstrate the uncoupling of macrolide induction of mefE/mel expression, and 309
macrolide efflux. The results with 16-membered macrolides further demonstrated that for induction of 310
mefE/mel a monosaccharide in position C5 of the macrolide was required but this was not sufficient to 311
confer MefE/Mel-mediated resistance. Thus, MEGA-containing isolates of S. pneumoniae remained 312
susceptible to most 16-membered macrolides, independent of the capacity to induce mefE/mel. 313
Discussion 314
Mechanisms of S. pneumoniae resistance to macrolides include modification of macrolide target 315
sites on the 23S ribosomal subunit, either by mutation or erm methylases, and macrolide efflux by the 316
MefE/Mel efflux pump encoded on the MEGA element (16, 23, 44, 45, 47, 52). In North America and 317
many other parts of the world, MefE/Mel is the predominant mechanism of pneumococcal macrolide 318
resistance (16, 43). Approximately half of the macrolide resistant pneumococci isolated in the United 319
States in 2006 contained mefE/mel as the sole macrolide resistance determinant and an additional 25% 320
contained both mefE/mel and ermB (22). Resistance levels conferred by MefE/Mel in vitro 321
(erythromycin 1-32 µg/ml) are lower compared to levels conferred by ribosome modification; 322
nevertheless, these levels are correlated with macrolide treatment failures (25, 26). Exposure in vivo to 323
inducers of the efflux pump may lead to resistance levels at the site of infection higher than those 324
predicted by in vitro assays. MefE/Mel resistance can be induced by up to a 200-fold increase in MICs 325
(3, 49), and therefore could be of significant clinical impact. This background led us to further study 326
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

induction of macrolide efflux, to identify structural feature that differentiate inducing and non-inducing 327
macrolides and to determine the relationship between induction and resistance. 328
A range of compounds and conditions were studied for the ability to induce expression of the 329
mefE-lacZ reporter using the disk diffusion assay. This assay allowed the efficient screening of a broad 330
range of inducer concentrations and conditions. Non-macrolide antibiotics or general environmental 331
stresses and conditions (e.g., competence induction), except for certain antimicrobial peptides (51) and 332
macrolides, did not induce expression of mefE/mel. The non-macrolide antibiotics studied represented 333
different classes with distinct cellular targets including ribosomal binding sites on 23S rRNA 334
overlapping the macrolide binding site (chloramphenicol, lincosamides, linezolid and streptogramins) 335
and 16S rRNA binding drugs (e.g., aminoglycosides and tetracycline), suggesting that disruption of 336
protein synthesis per se does not influence mefE/mel expression. Likewise, drugs that disrupt DNA 337
synthesis, cell wall synthesis, or membrane integrity, and various environmental conditions did not 338
induce expression, indicating that the range of inducers for mefE/mel expression is quite specific. 339
The current paradigm for the MefE/Mel efflux pump is that it confers resistance to 14- and 15-340
membered, but not to the 16-membered macrolides (5, 9, 10, 17, 34, 36); and 14- and 15-membered 341
induce MefE/Mel-mediated resistance (3, 36, 50). Our data confirm strong induction of mefE/mel 342
expression by erythromycin, clarithromycin and azithromycin, as previously reported (3, 50), and 343
showed that the most commonly studied 16-membered macrolides indeed did not induce mefE/mel 344
expression. However, the use a broader spectrum of structurally diverse macrolides revealed 14- 345
membered and 15-membered macrolides with reduced ability to induce mefE/mel expression and 16-346
membered macrolides that induced mefE/mel. In addition, the ketolide telithromycin, despite retaining 347
excellent activity against mefE-containing pneumococci, strongly induced mefE expression. 348
Telithromycin has not previously been demonstrated to induce mefE/mel. 349
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

The induction of mefE/mel by 14- and 15-membered macrolides, but not common 16-membered 350
macrolides coincides with differences in macrolide binding to the ribosome. Most 14- and 15-351
membered macrolides contain the monosaccharide desosamine at position C5 of the lactone ring, 352
whereas 16-membered macrolide typically have a C5 disaccharide. C5 substituents form extensive and 353
distinct bonds with the ribosome (Figs 3-8). The disaccharides in the 16-membered macrolides extend 354
deep into the exit tunnel of the ribosome (19, 38), allowing the mycarose (Fig. 4) to interact with G2505 355
and U2506 of the 23S rRNA (E. coli numbering, Fig. 4), interfering with peptidyl transferase activity 356
(19, 37, 38) and resulting in truncated peptides (19). Macrolides with C5 monosaccharides do not 357
interfere with peptidyl transferase activity, instead block egress of the nascent peptide from the exit 358
tunnel leading to peptides up to eight amino acids long (13, 19, 27, 38, 42). 359
We found that the inducing 16-membered macrolides tilmicosin and rosamicin like inducing 14- 360
and 15-membered macrolides have a monosaccharide at C5 and therefore are expected to have a similar 361
affect on peptide synthesis. The weak inducing 14-membered macrolide troleandomycin has 362
acetylations of the C3 and C5 sugars that force a substantially altered conformation when bound to the 363
ribosome resulting in a location of the macrolide farther down the exit tunnel than inducing macrolides 364
(6). The pivotal role of the C5 substituent for induction over the lactone-ring size or the C3 substituent is 365
also supported by our first identification of inducing 16-membered macrolides, which in addition lack 366
the C3 cladinose that is typically found in the common 14- and 15-membered macrolides. Taken 367
together, the induction mechanism involves specific binding of the macrolide to the ribosome and in 368
particular the C5 substituent of the macrolide. 369
Specific macrolide binding to the ribosome is also required for the induction of macrolide 370
resistance by many erm methylases (24, 28, 33, 41, 48), in mechanisms that are controlled by 371
transcriptional or translational attenuation. Weisblum (48) found that translational attenuation of ermC 372
in Staphylococcus aureus is alleviated by 14- but not 16-membered macrolides (48) and proposed that 373
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

when bound by a 14-membered macrolides, ribosomes stall due to constriction of the exit tunnel after a 374
short peptide is synthesized. Ribosome stalling at the end of the leader peptide synthesis promotes 375
refolding of the transcript into an anti-attenuator structure (48). Conversely, C5 disaccharides of many 376
16-membered macrolides disrupt ribosyltransferase activity preventing synthesis of the leader peptide so 377
that the erm transcript remains folded in the attenuator structure (19). 378
We propose a similar mechanism for induction of mefE/mel and efflux-mediated macrolide 379
resistance in pneumococci. Macrolides with C5 disaccharides may prevent synthesis of an unidentified 380
regulatory leader peptide and thus may not induce. Macrolides with a monosaccharide at C5 promote 381
stalling at the precise residue of the leader peptide due to the length of the nascent peptide that is 382
synthesized before being blocked in the ribosomal exit tunnel. This is independent of lactone ring size 383
or the C3 sugar substituent. 384
The MefE/Mel efflux pump does not confer resistance to common 16-membered macrolides. 385
Induction of the efflux pump with erythromycin prior to exposure to the 16-membered macrolides, 386
demonstrated that pneumococcal susceptibility to these macrolides was not due to a failure to induce 387
MefE/Mel. The lack of efflux of 16-membered macrolides could result either from an unavailability of 388
the compounds for export by the efflux pump due to high affinity ribosomal binding or that the 389
macrolides are not recognized as substrates. Most of 16-membered macrolides have a C6 390
acetylaldehyde that can form a covalent bond with the rRNA, resulting in high affinity binding to the 391
ribosome (19). In contrast, the 14- and 15-membered macrolides, which are known substrates for the 392
efflux pump, bind ribosomes through hydrophobic interactions and hydrogen bonding and are easily 393
dissociated from the ribosome (7, 19, 37, 42). Of the two16-membered macrolides, rosamicin and 394
tilmicosin that induced mefE/mel expression, only tilmicosin had reduced activity against MEGA-395
containing pneumococci, suggesting it was subject to efflux. The primary structural difference between 396
these two compounds is the C6 acetylaldehyde of rosamicin (but not tilmicosin), with the potential to 397
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

form the covalent bond effecting efflux by MefE/Mel. Further, telithromycin with 700-fold higher 398
affinity for the ribosome than erythromycin is predicted to have limited availability for export by 399
MefE/Mel. Despite strong induction of the efflux pump by telithromycin, pneumococci remain 400
susceptible. Thus, because of high affinity ribosomal binding most 16-membered macrolides and the 14-401
membered ketolides even if mefE/mel is induced are not effectively available for efflux, and 402
pneumococcal isolates remain susceptible to these compounds. 403
To summarize, induction of the mefE/mel was shown to be limited to a narrow range of 404
macrolides and antimicrobial peptides. The model indicates induction is mediated by ribosomal binding 405
and that inducing 14-, 15- and 16-membered macrolides contain a monosaccharide in position C5 of the 406
lactone ring. The C5 monosaccharide forms a characteristic macrolide-ribosome-mRNA complex 407
consistent with the induction mechanism described for methylase-mediated macrolide resistance, an 408
anti-attenuation-based mechanism for mefE/mel. Most16-membered macrolides and the 14-membered 409
ketolide have high affinity binding to the ribosome and this strong ribosomal binding limits the 410
availability for efflux from the pneumococcus, even in the presence of MefE/Mel induction. The 411
antimicrobial peptides such LL-37 and CRAMP that induce are not expected to bind ribosomes, thus 412
induction by these agents is anticipated to occur by an alternate pathway. Investigations are currently 413
underway to confirm and define the proposed macrolide transcriptional attenuator mechanism and to 414
study the basis for induction by antimicrobial peptides. 415
Acknowledgments 416
This work was supported by grants from the National Institutes of Health (5R01AI070829) and from the 417
Medical Research Service of the Department of Veterans Affairs (to DSS). 418
References 419
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

1. Alloing, G., C. Granadel, D. A. Morrison, and J.-P. Claverys. 1996. Competence pheromone, 420
oligopeptide permease, and induction of competence in Streptococcus pneumoniae. Molecular 421
Microbiology 21:471-478. 422
2. Alloing, G., B. Martin, C. Granadel, and J.-P. Claverys. 1998. Development of competence in 423
Streptococcus pneumoniae: pheromone autoinduction and control of quorum sensing by the 424
oligopeptide permease. Molecular Microbiology 29:75-83. 425
3. Ambrose, K. D., R. Nisbet, and D. S. Stephens. 2005. Macrolide efflux in Streptococcus 426
pneumoniae is mediated by a dual efflux pump (mel and mef) and is erythromycin inducible. 427
Antimicrobial Agents and Chemotherapy 49:4203-9. 428
4. Bailey, M., T. Chettiath, and A. S. Mankin. 2008. Induction of erm(C) Expression by 429
Noninducing Antibiotics. Antimicrob. Agents Chemother. 52:866-874. 430
5. Bejuk, D. 2004. Differentiation of resistance phenotypes among erythromycin-resistant 431
streptococci. Acta Med Croatica 58:301-6. 432
6. Berisio, R., J. Harms, F. Schluenzen, R. Zarivach, H. A. S. Hansen, P. Fucini, and A. 433
Yonath. 2003. Structural Insight into the Antibiotic Action of Telithromycin against Resistant 434
Mutants. J. Bacteriol. 185:4276-4279. 435
7. Berisio, R., F. Schluenzen, J. Harms, A. Bashan, T. Auerbach, D. Baram, and A. Yonath. 436
2003. Structural insight into the role of the ribosomal tunnel in cellular regulation. Nature 437
Structural Biology 10:366. 438
8. Bonnefoy, A., A. Girard, C. Agouridas, and J. Chantot. 1997. Ketolides lack inducibility 439
properties of MLS(B) resistance phenotype. J. Antimicrob. Chemother. 40:85-90. 440
9. Braga, P. C. 2002. Rokitamycin: bacterial resistance to a 16-membered ring macrolide differs 441
from that to 14- and 15-membered ring macrolides. J Chemother 14:115-31. 442
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

10. Clancy, J., J. Petitpas, F. Dib-Hajj, W. Yuan, M. Cronan, A. V. Kamath, J. Bergeron, and 443
J. A. Retsema. 1996. Molecular cloning and functional analysis of a novel macrolide-resistance 444
determinant, mefA, from Streptococcus pyogenes. Mol Microbiol 22:867-79. 445
11. CLSI. 2010. Performance Standards for Antimicrobial Susceptibility Testing; Twentieth 446
Informational Supplement. M100-S20, Wayne, PA. 447
12. Daly, M. M., S. Doktor, R. Flamm, and D. Shortridge. 2004. Characterization and prevalence 448
of MefA, MefE, and the associated msr(D) gene in Streptococcus pneumoniae clinical isolates. J 449
Clin Microbiol 42:3570-4. 450
13. Dinos, G. P., S. R. Connell, K. H. Nierhaus, and D. L. Kalpaxis. 2003. Erythromycin, 451
Roxithromycin, and Clarithromycin: Use of Slow-Binding Kinetics to Compare Their in Vitro 452
Interaction with a Bacterial Ribosomal Complex Active in Peptide Bond Formation. Mol 453
Pharmacol 63:617-623. 454
14. Farrell, D. J., S. G. Jenkins, S. D. Brown, M. Patel, B. S. Lavin, and K. P. Klugman. 2005. 455
Emergence and spread of Streptococcus pneumoniae with erm(B) and mef(A) resistance. Emerg 456
Infect Dis 11:851-8. 457
15. Gay, K., W. Baughman, Y. Miller, D. Jackson, C. G. Whitney, A. Schuchat, M. M. Farley, 458
F. Tenover, and D. S. Stephens. 2000. The emergence of Streptococcus pneumoniae resistant to 459
macrolide antimicrobial agents: a 6-year population-based assessment. J Infect Dis 182:1417-24. 460
16. Gay, K., and D. S. Stephens. 2001. Structure and dissemination of a chromosomal insertion 461
element encoding macrolide efflux in Streptococcus pneumoniae. J Infect Dis 184:56-65. 462
17. Giovanetti, E., M. P. Montanari, F. Marchetti, and P. E. Varaldo. 2000. In vitro activity of 463
ketolides telithromycin and HMR 3004 against Italian isolates of Streptococcus pyogenes and 464
Streptococcus pneumoniae with different erythromycin susceptibility. Journal of Antimicrobial 465
Chemotherapy 46:905-908. 466
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

18. Halfmann, A., R. Hakenbeck, and R. Bruckner. 2007. A new integrative reporter plasmid for 467
Streptococcus pneumoniae. FEMS Microbiology Letters 268:217-224. 468
19. Hansen, J. L., J. A. Ippolito, N. Ban, P. Nissen, P. B. Moore, and T. A. Steitz. 2002. The 469
Structures of Four Macrolide Antibiotics Bound to the Large Ribosomal Subunit. Molecular Cell 470
10:117-128. 471
20. Havarstein, L. S., G. Coomaraswamy, and D. A. Morrison. 1995. An unmodified 472
heptadecapeptide pheromone induces competence for genetic transformation in Streptococcus 473
pneumoniae. Proc Natl Acad Sci U S A 92:11140-11144. 474
21. Hui, F. M., and D. A. Morrison. 1991. Genetic transformation in Streptococcus pneumoniae: 475
nucleotide sequence analysis shows comA, a gene required for competence induction, to be a 476
member of the bacterial ATP-dependent transport protein family. J. Bacteriol. 173:372-381. 477
22. Jenkins, S., and D. Farrell. 2009. Increase in pneumococcus macrolide resistance, United 478
States. . Emerg Infect Dis [serial on the Internet] 15:1260-1264. Available from 479
http://www.cdc.gov/EID/content/15/8/1260.htm. 480
23. Johnston, N. J., J. C. de Azavedo, J. D. Kellner, and D. E. Low. 1998. Prevalence and 481
characterization of the mechanisms of macrolide, lincosamide, and streptogramin resistance in 482
isolates of Streptococcus pneumoniae. Antimicrob. Agents Chemother. 42:2425-2426. 483
24. Kamimiya, S., and B. Weisblum. 1997. Induction of ermSV by 16-membered-ring macrolide 484
antibiotics. Antimicrob. Agents Chemother. 41:530-534. 485
25. Klugman, K., and J. Lonks. 2005. Hidden epidemic of macrolide-resistant pneumococci. 486
Emerg Infect Dis 11:802-807. 487
26. Klugman, K. P. 2002. Bacteriological evidence of antibiotic failure in pneumococcal lower 488
respiratory tract infections. Eur Respir J 20:3S-8. 489
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

27. Korshunova, G., N. Sumbatyan, N. Fedorova, I. Kuznetsova, A. Shishkina, and A. 490
Bogdanov. 2007. Peptide derivatives of tylosin-related macrolides. Russian Journal of 491
Bioorganic Chemistry 33:218-226. 492
28. Kwak, J. H., E. C. Choi, and B. Weisblum. 1991. Transcriptional attenuation control of ermK, 493
a macrolide-lincosamide-streptogramin B resistance determinant from Bacillus licheniformis. J. 494
Bacteriol. 173:4725-4735. 495
29. Kwon, A. R., Y. H. Min, E. J. Yoon, J. A. Kim, M. J. Shim, and E. C. Choi. 2006. ErmK 496
leader peptide: amino acid sequence critical for induction by erythromycin. Arch Pharm Res 497
29:1154-7. 498
30. Lampson, B. C., W. von David, and J. T. Parisi. 1986. Novel mechanism for plasmid-499
mediated erythromycin resistance by pNE24 from Staphylococcus epidermidis. Antimicrob. 500
Agents Chemother. 30:653-658. 501
31. Martin, B., M. Prudhomme, G. Alloing, C. Granadel, and J.-P. Claverys. 2000. Cross-502
regulation of competence pheromone production and export in the early control of 503
transformation in Streptococcus pneumoniae. Molecular Microbiology 38:867-878. 504
32. Mayford, M., and B. Weisblum. 1989. ermC leader peptide. Amino acid sequence critical for 505
induction by translational attenuation. J Mol Biol 206:69-79. 506
33. Mayford, M., and B. Weisblum. 1990. The ermC leader peptide: amino acid alterations leading 507
to differential efficiency of induction by macrolide-lincosamide-streptogramin B antibiotics. J 508
Bacteriol 172:3772-9. 509
34. Mazzariol, A., R. Koncan, L. A. Vitali, and G. Cornaglia. 2007. Activities of 16-membered 510
ring macrolides and telithromycin against different genotypes of erythromycin-susceptible and 511
erythromycin-resistant Streptococcus pyogenes and Streptococcus pneumoniae. J. Antimicrob. 512
Chemother. 59:1171-1176. 513
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

35. Miller, J. H. 1972. Experiments in molecular genetics. A laboratory manual and handbook for 514
Escherichia coli and related bacteria. Cold Springs Harbor Laboratory Press, Cold Springs 515
Harbor, New York. 516
36. Nishijima, T., Y. Saito, A. Aoki, M. Toriya, Y. Toyonaga, and R. Fujii. 1999. Distribution of 517
mefE and ermB genes in macrolide-resistant strains of Streptococcus pneumoniae and their 518
variable susceptibility to various antibiotics. J. Antimicrob. Chemother. 43:637-643. 519
37. Pfister, P., S. Jenni, J. Poehlsgaard, A. Thomas, S. Douthwaite, N. Ban, and E. C. Bottger. 520
2004. The Structural Basis of Macrolide-Ribosome Binding Assessed Using Mutagenesis of 23 S 521
rRNA Positions 2058 and 2059. Journal of Molecular Biology 342:1569-1581. 522
38. Poulsen, S. M., C. Kofoed, and B. Vester. 2000. Inhibition of the ribosomal peptidyl 523
transferase reaction by the mycarose moiety of the antibiotics carbomycin, spiramycin and 524
tylosin. Journal of Molecular Biology 304:471-481. 525
39. Prudhomme, M., L. Attaiech, G. Sanchez, B. Martin, and J. P. Claverys. 2006. Antibiotic 526
stress induces genetic transformability in the human pathogen Streptococcus pneumoniae. 527
Science 313:89-92. 528
40. Ross, J. I., E. A. Eady, J. H. Cove, W. J. Cunliffe, S. Baumberg, and J. C. Wootton. 1990. 529
Inducible erythromycin resistance in staphylococci is encoded by a member of the ATP-binding 530
transport super-gene family. Molecular Microbiology 4:1207-1214. 531
41. Sandler, P., and B. Weisblum. 1989. Erythromycin-induced ribosome stall in the ermA leader: 532
a barricade to 5'-to-3' nucleolytic cleavage of the ermA transcript. J. Bacteriol. 171:6680-6688. 533
42. Schlunzen, F., R. Zarivach, J. Harms, A. Bashan, A. Tocilj, R. Albrecht, A. Yonath, and F. 534
Franceschi. 2001. Structural basis for the interaction of antibiotics with the peptidyl transferase 535
centre in. Nature 413:814. 536
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

43. Stephens, D. S., S. M. Zughaier, C. G. Whitney, W. S. Baughman, L. Barker, K. Gay, D. 537
Jackson, W. A. Orenstein, K. Arnold, A. Schuchat, and M. M. Farley. 2005. Incidence of 538
macrolide resistance in Streptococcus pneumoniae after introduction of the pneumococcal 539
conjugate vaccine: population-based assessment. The Lancet 365:855-863. 540
44. Sutcliffe, J., T. Grebe, A. Tait-Kamradt, and L. Wondrack. 1996. Detection of erythromycin-541
resistant determinants by PCR. Antimicrobial Agents and Chemotherapy 40:2562-6. 542
45. Sutcliffe, J., A. Tait-Kamradt, and L. Wondrack. 1996. Streptococcus pneumoniae and 543
Streptococcus pyogenes resistant to macrolides but sensitive to clindamycin: a common 544
resistance pattern mediated by an efflux system. Antimicrobial Agents and Chemotherapy 545
40:1817-24. 546
46. Syrogiannopoulos, G. A., I. N. Grivea, L. M. Ednie, B. Bozdogan, G. D. Katopodis, N. G. 547
Beratis, T. A. Davies, and P. C. Appelbaum. 2003. Antimicrobial susceptibility and macrolide 548
resistance inducibility of Streptococcus pneumoniae carrying erm(A), erm(B), or mef(A). 549
Antimicrobial Agents and Chemotherapy 47:2699-702. 550
47. Tait-Kamradt, A., J. Clancy, M. Cronan, F. Dib-Hajj, L. Wondrack, W. Yuan, and J. 551
Sutcliffe. 1997. mefE is necessary for the erythromycin-resistant M phenotype in Streptococcus 552
pneumoniae. Antimicrobial Agents and Chemotherapy 41:2251-5. 553
48. Weisblum, B. 1995. Insights into erythromycin action from studies of its activity as inducer of 554
resistance. Antimicrob. Agents Chemother. 39:797-805. 555
49. Wierzbowski, A. K., D. Boyd, M. Mulvey, D. J. Hoban, and G. G. Zhanel. 2005. Expression 556
of the mef(E) gene encoding the macrolide efflux pump protein increases in Streptococcus 557
pneumoniae with increasing resistance to macrolides. Antimicrob Agents Chemother 49:4635-558
40. 559
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

50. Wierzbowski, A. K., D. Boyd, M. Mulvey, D. J. Hoban, and G. G. Zhanel. 2005. Expression 560
of the mef(E) gene encoding the macrolide efflux pump protein increases in Streptococcus 561
pneumoniae with increasing resistance to macrolides. Antimicrobial Agents and Chemotherapy 562
49:4635-40. 563
51. Zähner, D., X. Zhou, S. T. Chancey, J. Pohl, W. M. Shafer, and D. S. Stephens. 2010. 564
Human antimicrobial peptide LL-37 induces mefE/mel-mediated macrolide resistance in 565
Streptococcus pneumoniae. Antimicrob. Agents Chemother. 54 3516-3519 566
52. Zhanel, G. G., and J. A. Karlowsky. 1999. Ribosomal Resistance: Emerging Problems and 567
Potential Solutions. Curr Infect Dis Rep 1:458-463. 568
569
570
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

Figure 1. Induction of mefE-lacZ in isogenic S. pneumoniae strains. All strains were derived from the 1
MEGA-containing, invasive pneumococcal isolate GA17457 and include XZ7042, the MEGA deletion 2
mutant XZ8004 and the promoterless lacZ negative control XZ7049. (a) β-galactosidase expression by 3
XZ7042 (solid lines, circles) and XZ7049 (dashed lines, squares). Strains were untreated (open symbols) or 4
exposed to 0.1 µg/ml erythromycin (closed symbols) at 0 hours and cells were harvested and assayed at 5
half-hour intervals. Error bars indicate standard deviation of duplicate replications. (b) Erythromycin dose 6
response curves of the XZ7042 (solid lines, open squares) and XZ8004 (dashed lines, closed triangles). 7
Arrows indicate the erythromycin MIC of each strain (solid, XZ7042; dashed, XZ8004). Error bars indicate 8
standard deviation of duplicate replications. (c) Disk diffusion assay for induction of mefE-lacZ by 9
erythromycin. Disks containing 15 µg/ml erythromycin were placed on plates swabbed with XZ7042 (first 10
panel), XZ8004 (middle panel) and XZ7049 (last panel). Expression of the reporter is indicated as a blue 11
halo surrounding a zone of inhibition. 12
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

Figure 2. Competence stimulating peptide 1 (CSP-1) did not induced mefE-lacZ. Disk diffusion 1
induction assay of the mefE-lacZ (XZ7042) and comC-lacZ (XZ7067) reporter strains by 2
erythromycin (ERY) and CSP-1. CSP-1 was spotted directly onto THY indicator plates streaked 3
with XZ7042 or XZ7067. As controls, diffusion disks containing erythromycin (15 µg) were 4
placed onto TSA indicator plates streaked with each strain. A blue spot or blue halo indicated 5
comC-lacZ or mefE-lacZ induction, respectively. 6
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

Figure 3. 14- and 15-membered macrolides. Structure and disk diffusion induction assays for the 14-
and 15-membered macrolides used in this study. Discs containing 15 µg/ml of each macrolide were
placed onto TSA indicator plates spread with XZ7042. (a) Cladinosolides. Erythromycin and
derivatives each contain a C3 cladinose. 1, erythromycin; 2, clarithromycin; 3, dirithromycin; 4,
roxithromycin. (b) Telithromycin. The C11/C12 carbamate with a aryl-akyl side chain is responsible for
increased ribosomal affinity and compensates for the absence of cladinose at C3 (19). 1, erythromycin;
2, telithromycin. (c) Oleandomycins. Oleandomycin and troleandomycin differ only by acetylation of
hydroxyl groups at three positions in troleandomycin (R1, R2, and R3). The dashed arrow indicates
interaction of acetyl group of the oleandrose of troleandomycin and the 23 rRNA nucleotide U790 (E.
coli numbering). 1, erythromycin; 2, troleandomycin; 3, oleandomycin. (d). 15-membered azalides.
Previously described interaction between azithromycin cladinose hydroxyl group and the 23S rRNA
nucleotide G2505 (E. coli numbering, dashed arrow) (19). C5 desosamine is indicated in red. 1,
erythromycin; 2, tulathromycin; 3, azithromycin.
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

Figure 4. Structures and disk diffusion induction assays for all 16-membered macrolides used in this study. 1
(a) Common non-inducing 16-membered macrolides. Positions of the carbon atoms in the lactone ring are 2
indicated and the C5 disaccharide composed of the amino sugar mycaminose and neutral sugar mycarose is 3
shown in red. Previously described points of interactions between 16-membered macrolides and 23S rRNA 4
nucleotides (E. coli numbering) are indicated by dashed arrows (43, 47). The C6 acetylaldehyde and the 5
reversible covalent bond (wavy line) formed with the nucleotide A2062 (solid arrow) are indicated in blue 6
(19). 1, josamycin; 2, kitasamycin; 3, midecamycin; 4, spiramycin; center disk, erythromycin control. Abbr. 7
foro, forosamide. (b) Tylosin and C5 monosaccharide-containing derivatives rosamicin and tilmicosin. The 8
C5 monosaccharides desosamine and mycaminose are indicated in red. 1, rosamicin; 2. erythromycin control; 9
3, tilmicosin; 4, tylosin. Discs containing 15 µg/ml of each macrolide were placed onto TSA indicator plates 10
spread with XZ7042. 11
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

Table 1. Strains and plasmids used in this work
Strains or
plasmid Description
Reference
or Source
S. pneumoniae
GA17457 Wild type, parent; MEGA; ERYR (51)
XZ7042 GA17457-derivative; bgaA::mefE-lacZ; MEGA; ERYR (51)
XZ8004 GA17457-derivative; bgaA::PmefE-lacZ; MEGA::aad9; ERYS (51)
XZ7067 GA17457-derivative; bgaA::PcomC-lacZ; MEGA; ERYR This study
XZ7049 GA17457-derivative; bgaA::lacZ (promoterless); MEGA; ERYR This study
Plasmids
pPP2 pPP1 derivative, lacZ reporter gene with the htrA ribosomal binding site (18)
pPC2 pPP2 derivative; carries PcomC-lacZ fusion (18)
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
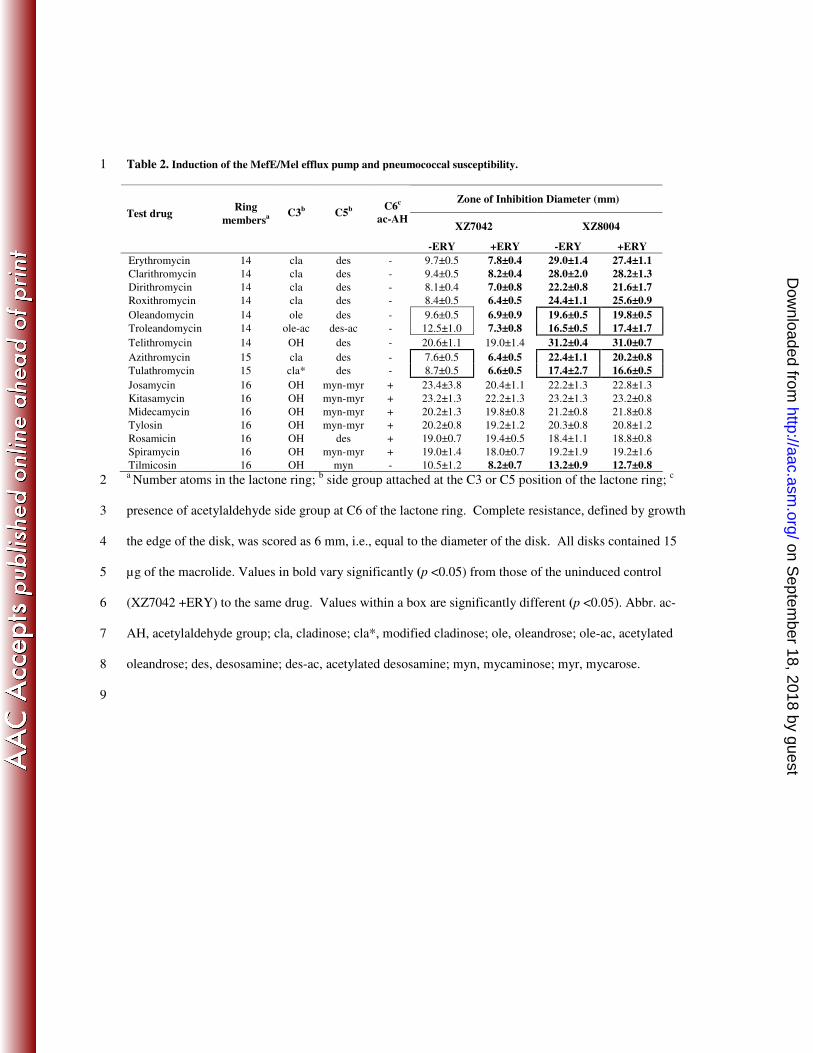
Table 2. Induction of the MefE/Mel efflux pump and pneumococcal susceptibility. 1
Zone of Inhibition Diameter (mm)
XZ7042 XZ8004
Test drug Ring
membersa
C3b C5
b
C6c
ac-AH
-ERY +ERY -ERY +ERY
Erythromycin 14 cla des - 9.7±0.5 7.8±0.4 29.0±1.4 27.4±1.1
Clarithromycin 14 cla des - 9.4±0.5 8.2±0.4 28.0±2.0 28.2±1.3
Dirithromycin 14 cla des - 8.1±0.4 7.0±0.8 22.2±0.8 21.6±1.7
Roxithromycin 14 cla des - 8.4±0.5 6.4±0.5 24.4±1.1 25.6±0.9
Oleandomycin 14 ole des - 9.6±0.5 6.9±0.9 19.6±0.5 19.8±0.5
Troleandomycin 14 ole-ac des-ac - 12.5±1.0 7.3±0.8 16.5±0.5 17.4±1.7
Telithromycin 14 OH des - 20.6±1.1 19.0±1.4 31.2±0.4 31.0±0.7
Azithromycin 15 cla des - 7.6±0.5 6.4±0.5 22.4±1.1 20.2±0.8
Tulathromycin 15 cla* des - 8.7±0.5 6.6±0.5 17.4±2.7 16.6±0.5
Josamycin 16 OH myn-myr + 23.4±3.8 20.4±1.1 22.2±1.3 22.8±1.3
Kitasamycin 16 OH myn-myr + 23.2±1.3 22.2±1.3 23.2±1.3 23.2±0.8
Midecamycin 16 OH myn-myr + 20.2±1.3 19.8±0.8 21.2±0.8 21.8±0.8
Tylosin 16 OH myn-myr + 20.2±0.8 19.2±1.2 20.3±0.8 20.8±1.2
Rosamicin 16 OH des + 19.0±0.7 19.4±0.5 18.4±1.1 18.8±0.8
Spiramycin 16 OH myn-myr + 19.0±1.4 18.0±0.7 19.2±1.9 19.2±1.6
Tilmicosin 16 OH myn - 10.5±1.2 8.2±0.7 13.2±0.9 12.7±0.8 a Number atoms in the lactone ring;
b side group attached at the C3 or C5 position of the lactone ring;
c 2
presence of acetylaldehyde side group at C6 of the lactone ring. Complete resistance, defined by growth 3
the edge of the disk, was scored as 6 mm, i.e., equal to the diameter of the disk. All disks contained 15 4
µg of the macrolide. Values in bold vary significantly (p <0.05) from those of the uninduced control 5
(XZ7042 +ERY) to the same drug. Values within a box are significantly different (p <0.05). Abbr. ac-6
AH, acetylaldehyde group; cla, cladinose; cla*, modified cladinose; ole, oleandrose; ole-ac, acetylated 7
oleandrose; des, desosamine; des-ac, acetylated desosamine; myn, mycaminose; myr, mycarose. 8
9
on Septem
ber 18, 2018 by guesthttp://aac.asm
.org/D
ownloaded from



