unep eeap report
TRANSCRIPT

Photochemical &Photobiological SciencesAn international journalwww.rsc.org/pps
ISSN 1474-905X
Themed Bar text in here xxxxxxxxxxxxxxxxxxxxxxxxxxxxxxThemed issue: Environmental effects of ozone depletion: 2014 assessment
Volume 14 Number 1 January 2015 Pages 1–186

Photochemical &Photobiological Sciences
EDITORIAL
Cite this: Photochem. Photobiol. Sci.,2015, 14, 7
DOI: 10.1039/c4pp90039a
www.rsc.org/pps
Preface
The assessments presented in the sevenpapers published in this journal dealwith the effects of ozone depletion onhuman health and the environmentand the consequences of interactionsbetween ozone depletion and globalclimate change. This report is producedin the first instance as a limited editionfor governments, via the United NationsEnvironment Programme. The report isnow made more readily available to thescientific community by publication inthe scientific literature. It was written bythe 42 members and co-authors of thePanel, reviewed by 45 scientists from anumber of countries, and is based onthe research work by many more col-leagues and scientists in many fields.
The assessment is presented here infull. We hope that publication in thisscientific journal will give it a wide reader-ship. As editors and panel members,we also hope that it will help keep scien-tists aware of their involvement in theprotection of the environment for allforms of life on Earth.
Keith R. SolomonJanet F. Bornman
AcknowledgmentsCover illustration
We thank Dr Cedric Fichot for supplyingthe cover for this issue.
The cover illustrates terrestrial inputsof dissolved organic carbon into theArctic Ocean (browner colours) as deter-mined by an optical metric based on the
spectral slope between 275–295 nm.This ultraviolet radiation (UV) basedmetric can be linked to remote sensinginstruments on satellites, and is aneffective tracer of chromophoric dis-solved organic matter (CDOM) that canreduce the UV transparency of inlandand coastal waters. Solar UV disinfectionof parasites and pathogens is corre-spondingly reduced in regions with highCDOM. See Häder et al., Photochem.Photobiol. Sci., 2015, DOI: 10.1039/c4pp90035a.
Financial support
Publication of this special issue by thejournal, Photochemical & PhotobiologicalSciences, was made possible by financialsupport from:• Ecosystems Research Division,National Exposure Research Laboratory,US Environmental Protection Agency,USA• Institute for Conservation Biology andFaculty of Science, Medicine and Health,University of Wollongong, Australia• International Institute of Agri-FoodSecurity, Curtin University, Australia• Laboratory of Atmospheric Physics,Aristotle University of Thessaloniki,Greece• Loyola University, J.H. MullahyEndowed Chair in Environmental Bio-logy, USA• Miami University, Eminent Scholar inEcosystem Ecology, USA• National Centre for Epidemiology andPopulation Health, Research School ofPopulation Health, The AustralianNational University
• State Environmental Protection Com-monwealth Project of China (201309073)• Telethon Kids Institute, Australia• The Japanese Ministry of the Environ-ment, Japan• The National Center for AtmosphericResearch, sponsored by the NationalScience Foundation, USA
Our thanks to the reviewers
This assessment would not have beenpossible without the contributions of thefollowing reviewers (in alphabeticalorder), all of whom made written and/orverbal comments on the content of thepapers in this assessment. We thankthem for their generous contribution oftime and knowledge:
Dr Marcos Andrade, Prof. ManuelAnton, Dr Pedro J. Aphalo, Dr MarianneBerwick, Prof. Lars Olof Björn, Dr MarioBlumthaler, Dr Natalia Chubarova,Dr Thomas P. Coohill, Dr SandraL. Cooke, Dr Raul Cordero, Dr RoseCory, Prof. Andy Davis, Dr Tad Day,Prof. Edward DeFabo, Dr Susana Diaz,Prof. Nils Ekelund, Dr Dylan Gwynn-Jones, Prof. Prue Hart, Prof. MichaelaHegglin, Prof. Wen-Che Hou, Dr GarethJenkins, Dr Stelios Kazadzis, Prof. JenniferY. King, Dr Helena Korpelainen,Dr Shaoshan Li, Dr Masaji Ono,Dr Madan Pal, Dr Ruben Piacentini,Prof. Henning Rodhe, Prof. AndrewL. Rose, Prof. Gunther Seckmeyer,Dr Jonathan Shanklin, Dr Anna MariaSiani, Dr J. Richard Soulen, Dr JohannaTamminen, Prof. Hugh R. Taylor, Dr FionaTummon, Prof. Roland von Glasow,
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 7–8 | 7

Dr Jason Wargent, Dr Ann R. Webb,Prof. Cheryl A. Wilson, Dr CaradeeWright, Prof. Huixiang Xie, Prof.Prodromos Zanis, and Dr FrancescoZaratti.
Review meetingThe review meeting of the Panel washeld in Zhengzhou, Henan Province,China. We thank the University ofZhengzhou and the University of Peking
for providing logistical support and assist-ance. The Yuda Palace Hotel, Zhengzhou,provided excellent facilities for themeeting and we thank the staff for excep-tional service and attention to detail.
Editorial Photochemical & Photobiological Sciences
8 | Photochem. Photobiol. Sci., 2015, 14, 7–8 This journal is © The Royal Society of Chemistry and Owner Societies 2015
Photochemical & Photobiological SciencesThe official journal of the European Photochemistry Association, the European Society forPhotobiology, the Asia and Oceania Society for Photobiology and the Korean Society ofPhotoscience
www.rsc.org/ppsThe Royal Society of Chemistry is the world's leading chemistry community. Through our high impact journals and publications weconnect the world with the chemical sciences and invest the profits back into the chemistry community.
IN THIS ISSUE
ISSN 1474-905X CODEN PPSHCB 14(1) 1–186 (2015)
CoverThe cover illustrates terrestrialinputs of dissolved organiccarbon into the Arctic Ocean(browner colours) asdetermined by an opticalmetric based on the spectralslope between 275-295 nm.See Donat-P. Häder et al.,pp. 108–126 for a fulldescription.
Image reproduced bypermission of Cedric Fichot,Photochem. Photobiol. Sci.,2015, 14, 108.
EDITORIALS
7
Preface
9
Environmental effects of ozone depletion and itsinteractions with climate change: 2014 assessmentIntroduction
10
Abbreviations and glossary
14
Environmental effects of ozone depletion and itsinteractions with climate change: 2014 assessmentExecutive summary
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 3–6 | 3

Editorial staffExecutive editorSarah Ruthven
Deputy executive editorGuy Jones
Development editorSimon Neil
Editorial production managerAndrew Shore
Publishing editorsRachel Cooper, Debora Giovanelli, Caroline Knapp, Helen Lunn, Anisha Ratan
Publishing assistantsCharlotte Rowley, Catherine Smith
PublisherJamie Humphrey
Honorary editorsGiulio Jori, Frank Wilkinson
For queries about submitted articles please contact Andrew Shore, Editorial production manager, in the first instance. E-mail [email protected]
For pre-submission queries please contact Sarah Ruthven, Executive editor. E-mail [email protected]
Photochemical & Photobiological Sciences (print: ISSN 1474-905X; electronic: ISSN 1474-9092) is published 12 times a year by the Royal Society of Chemistry, Thomas GrahamHouse, Science Park, Milton Road, Cambridge, CB4 0WF, UK on behalf of the European Society for Photobiology and the European Photochemistry Association.
All orders, with cheques made payable to the Royal Society of Chemistry, should be sent to RSC Order Department, Royal Society of Chemistry, Thomas Graham House, Science Park, Milton Road, Cambridge, CB4 0WF, UK Tel +44 (0)1223 432398; E-mail [email protected]
2015 Annual (print + electronic) subscription price: £1783; US$3329. 2015 Annual (electronic) subscription price: £1694; US$3162 Customers in Canada will be subject to a surcharge to cover GST. Customers in the EU subscribing to the electronic version only will be charged VAT.
If you take an institutional subscription to any RSC journal you are entitled to free, site-wide web access to that journal. You can arrange access via Internet Protocol (IP) address at www.rsc.org/ip. Customers should make payments by cheque in sterling payable on a UK clearing bank or in US dollars payable on a US clearing bank.
Advertisement sales: Tel +44 (0) 1223 432246; Fax +44 (0) 1223 426017; E-mail [email protected]
For marketing opportunities relating to this journal, contact [email protected]
Photochemical & Photobiological SciencesThe official journal of the European Photochemistry Association, the European Society for Photobiology, the Asia and Oceania Society for Photobiology and the Korean Society of Photoscience
www.rsc.org/ppsPhotochemical & Photobiological Sciences contains articles on all aspects of photochemistry and photobiology, including elemental photochemical and photophysical processes, the interaction of light with living systems, environmental photochemistry, environmental photobiology, the use of light as a reagent, how light affects health, the use of light as a diagnostic tool and for curative purposes and areas in which light is a cost-effective catalyst.
Editorial board
Editors-in-chiefRex Tyrell, University of Bath, UKDario Bassani, Université Bordeaux
1, France
Deputy editors-in-chiefSanti Nonell, University Ramon Llull,
SpainJacob Wirz, University of Basel,
Switzerland
Associate editorsPatrizia Agostinis, Katholieke
Universiteit Leuven, BelgiumNicola Armaroli, Istituto ISOF/CNR,
ItalyKristian Berg, The Norwegian Radium
Hospital, NorwayCornelia Bohne, University of
Victoria, CanadaJanet Bornman, Curtin University,
AustraliaLuisa De Cola, University of
Strasbourg, FrancePaolo Di Mascio, Universidade de São
Paulo, BrazilHermenegildo García, Universidad
Politécnica de Valencia, SpainKen Ghiggino, University of
Melbourne, AustraliaMasahiro Irie, Rikkyo University, JapanYunbao Jiang, Xiamen University,
ChinaChoon-Hwan Lee, Pusan National
University, Korea
Fred Lewis, Northwestern University, USA
Aba Losi, University of Parma, ItalyTim Maisch, University Hospital
Regensburg, GermanySixto Malato, CIEMAT, SpainJochen Mattay, Universitat Bielefeld,
GermanyRichard McKenzie, National Institute
of Water & Atomspheric Research (NIWA), New Zealand
Hiroshi Miyasaka, Osaka University, Japan
Sandra Monti, ISOF/CNR, ItalyPatrick Neale, Smithsonian
Environmental Research Center, USA
Frank Quina, University of São Paulo, Brazil
Vivienne Reeve, University of Sydney, Australia
Evelyne Sage, Centre Universitaire, France
Masako Sasakai, Tokai University, Japan
Paola Taroni, Politecnico di Milano, Italy
Franz Trautinger, Karl Landsteiner Institute of Dermatological Research, Austria
Cristiano Viappiani, Universita di Parma, Italy
Vivian WW Yam, University of Hong Kong, Hong Kong
Minjoong Yoon, Chungnam National University, Korea
Honorary editorsGiulio Jorii, University of Padova, ItalyFrank Wilkinson, Loughborough
University, UK
Ownership boardDario Bassani, Université Bordeaux
1, FranceWerner Nau, Jacobs University,
GermanyJacques Piette, Université de Liège,
Belgium
Lesley Rhodes, University of Manchester, UK
Frans De Schryver, Universiteit Leuven, Belgium
Rex Tyrrell, University of Bath, UKEric Vauthey, University of Geneva,
Switzerland
Associate membersChoon-Hwan Lee, Korean Society of
PhotoscienceSarah Ruthven, Royal Society of
Chemistry, Cambridge, UKJamie Humphrey, Royal Society of
Chemistry, Cambridge, UK
Information for authors
Full details of how to submit material for publication in Photochemical & Photobiological Sciences are given in the Instructions for Authors (available from http://www.rsc.org/authors). Submissions should be made via the journal’s homepage: http://www.rsc.org/pps
Authors may reproduce/republish portions of their published contribution without seeking permission from the RSC, provided that any such republication is accompanied by an acknowledgement in the form: (Original Citation)—Reproduced by permission of the Royal Society of Chemistry, the European Society for Photobiology and the European Photochemistry Association.
©The Royal Society of Chemistry and Owner Societies 2015. Apart from fair dealing for the purposes of research or private study for non-commercial purposes or criticism or review, as permitted under the Copyright, Designs and Patents Act 1988 and the Copyright and Related Rights Regulations 2003, this publication may only be reproduced, stored or transmitted, in any form or by any means, with the prior permission in writing of the Publisher or in the case of reprographic reproduction in accordance with the terms of licences issued by the Copyright Licensing Agency in the UK. US copyright law is applicable to users in the USA.
The Royal Society of Chemistry takes reasonable care in the preparation of this publication but does not accept liability for the consequences of any errors or omissions.
∞ The paper used in this publication meets the requirements of ANSI/NISO Z39.48–1992 (Permanence of Paper).

PERSPECTIVES
19
Ozone depletion and climate change: impacts onUV radiation
A. F. Bais,* R. L. McKenzie, G. Bernhard, P. J. Aucamp,M. Ilyas, S. Madronich and K. Tourpali
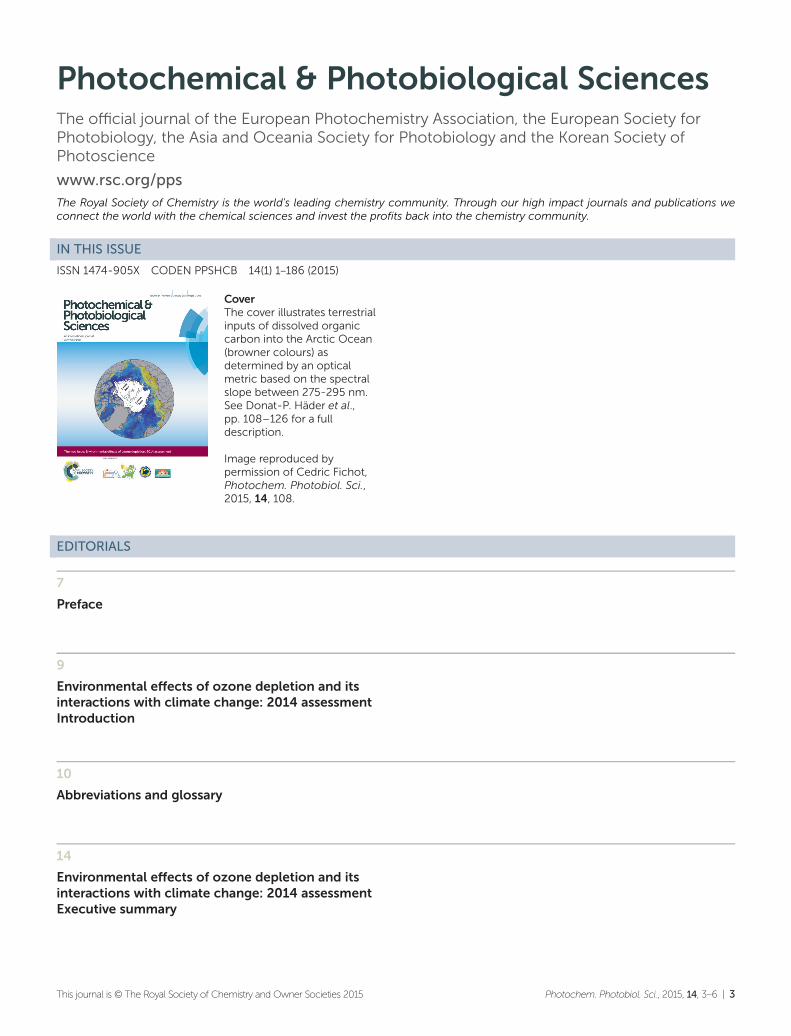
Percentage changes in the UV Index (UVI) for 2090 relativeto 2015 due to changes in ozone (left) and aerosols (right)only. Large decreases are projected over Antarctica due tostratospheric ozone recovery. Increases are projected forparts of Asia due to decreases in aerosols, partly reversingthe possible large reductions in UVI after the 1950s.
53
The consequences for human health ofstratospheric ozone depletion in association withother environmental factors
R. M. Lucas,* M. Norval, R. E. Neale, A. R. Young,F. R. de Gruijl, Y. Takizawa and J. C. van der Leun
Ozone depletion, climate and human health.
88
Solar ultraviolet radiation and ozone depletion-driven climate change: effects on terrestrialecosystems
J. F. Bornman,* P. W. Barnes, S. A. Robinson,C. L. Ballaré, S. D. Flint and M. M. Caldwell
We summarise advances in our knowledge of how UV-Bradiation (280–315 nm) together with other climatechange factors interact in their influence on terrestrialorganisms and ecosystems.
108
Effects of UV radiation on aquatic ecosystems andinteractions with other environmental factors
Donat-P. Häder,* Craig E. Williamson,Sten-Åke Wängberg, Milla Rautio, Kevin C. Rose,Kunshan Gao, E. Walter Helbling, Rajeshwar P. Sinhaand Robert Worrest
Combined effects of anthropogenic changes in theenvironmental condition in marine ecosystems, includingUV, CO2 and temperature.
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 3–6 | 5

PERSPECTIVES
127
Effects of stratospheric ozone depletion, solar UVradiation, and climate change on biogeochemicalcycling: interactions and feedbacks
David J. Erickson III,* Barbara Sulzberger,Richard G. Zepp and Amy T. Austin
Solar UV radiation and climate change interact to influenceand determine the environmental conditions for humanson planet Earth.
149
Changes in air quality and troposphericcomposition due to depletion of stratosphericozone and interactions with changing climate:implications for human and environmental health
S. Madronich,* M. Shao, S. R. Wilson, K. R. Solomon,J. D. Longstreth and X. Y. Tang
UV radiation is an essential driver for the formation ofphotochemical smog, which includes ground-level ozoneand particulate matter (PM).
170
Consequences of stratospheric ozone depletionand climate change on the use of materials
A. L. Andrady,* A. Torikai, H. H. Redhwi, K. K. Pandey andP. Gies
Materials used in the exterior of buildings and inconstruction are routinely exposed to solar UV radiation.Especially in the case of wood and plastic buildingmaterials, the service life is determined by theirweather-induced deterioration.
6 | Photochem. Photobiol. Sci., 2015, 14, 3–6 This journal is © The Royal Society of Chemistry and Owner Societies 2015

Photochemical &Photobiological Sciences
EDITORIAL
Cite this: Photochem. Photobiol. Sci.,2015, 14, 9
DOI: 10.1039/c4pp90040e
www.rsc.org/pps
Environmental effects of ozone depletionand its interactions with climate change:2014 assessment
Introduction
The Montreal Protocol on Substancesthat Deplete the Ozone Layer is an inter-national agreement on the protection ofthe stratospheric ozone layer, now rati-fied by 196 countries. Three AssessmentPanels have continuously contributedinformation for facilitating informeddecisions on the science of ozonedepletion, environmental effects andclimate change interactions, and techno-logical and economic implications of
ozone depleting substances and theirsubstitutes. This current assessment bythe Environmental Effects AssessmentPanel (EEAP) reports on key findingson the environment and health sincethe last full assessment of 2010 (Photo-chem. Photobiol. Sci., 2011, 10 (1)), payingattention to the interactions betweenozone depletion and climate change.
The publication of this assessment inthe scientific literature aims to inform
the scientific community how their data,modeling and interpretations are playinga role in information dissemination tothe Parties to the Montreal Protocol,other policymakers, and fellow scientists.
Janet F. BornmanNigel PaulMin ShaoCo-chairs of the Environmental Effects
Assessment Panel
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 9 | 9

Photochemical &Photobiological Sciences
EDITORIAL
Cite this: Photochem. Photobiol. Sci.,2015, 14, 10
DOI: 10.1039/c4pp90041c
www.rsc.org/pps
Abbreviations and glossary
1,25(OH)-2D
1,25-Dihydroxyvitamin D
25(OH)D 25-Hydroxyvitamin D
Airshed Airshed; an area where themovement of air (and there-fore air pollutants) can be hin-dered by local geographicalfeatures such as mountains
AK Actinic keratosis
AMD Age-related maculardegeneration
AMP Anti-microbial peptide
ANSI American National StandardsInstitute
AO Arctic oscillation; a large-scale variation in Arctic windpatterns
AOD Aerosol optical depth
APase Alkaline phosphatase
APC Antigen presenting cell
ASL Above sea level
BCC Basal cell carcinoma(s)
Br Bromine (an ozone depletingchemical)
BRAF B-Rapidly Accelerated Fibro-sarcoma; a gene that is com-monly mutated in melanoma
BrO Bromine monoxide
BSWF Biological spectral weightingfunctions
BWF Biological weighting function
CAS Chemical Abstracts Service
CAT Catalase
CC Cortical cataract(s)
CCl4 Carbon tetrachloride (anozone depleting gas)
CCM Chemistry-climate model(used to predict futurechanges in atmosphericcomposition)
CDFA Chlorodifluoroacetic acid
CDK Climatic droplet keratopathy
CDKN2A Cyclin-dependent kinaseinhibitor 2A; a gene com-monly mutated in melanoma
CDK4 Cyclin-dependent kinase 4;mutations in the gene arefound in melanoma
CDOC Coloured dissolved organiccarbon
CDOM Coloured (or chromophoric)dissolved organic matter
CDR Carbon dioxide reduction
CFC Chlorofluorocarbon; ozonedepleting substance (e.g.CFC12, dichlorodifluoro-methane: CCl2F2), now con-trolled under the MontrealProtocol
CH Contact hypersensitivity
CH4 Methane (a greenhouse gas)
CHCl3 Chloroform (an ozone deplet-ing gas)
CIE Commission Internationale del’Eclairage (InternationalCommission on Illumination)
Cl Chlorine (an ozone depletingsubstance)
CMF Cloud modification factor
CMM Cutaneous melanoma
CO Carbon monoxide
CO2 Carbon dioxide (a green-house gas)
COS Carbonyl sulfide
COT Cloud optical depth
CPD Cyclobutane pyrimidinedimer
Cu Copper (Cu(I) and Cu(II)being different oxidationstates)
DIC Dissolved inorganic carbon
DMS Dimethylsulfide
DMSP Dimethylsulfoniopropionate
DNA Deoxyribonucleic acid
DOC Dissolved organic carbon
DOM Dissolved organic matter
DON Dissolved organic nitrogen
DSB Double strand break
DTH Delayed type hypersensitivity
DU Dobson unit; used for themeasurement of totalcolumn ozone (1 DU = 2.69 ×1016 molecule cm−2)
DVM Daily vertical migration
EAE Experimental allergicencephalitis
10 | Photochem. Photobiol. Sci., 2015, 14, 10–13 This journal is © The Royal Society of Chemistry and Owner Societies 2015

EDUCE European Database for Ultra-violet Radiation Climatologyand Evaluation
EESC Equivalent Effective Strato-spheric Chlorine; a term usedto represent the total chlorineconcentration in the strato-sphere from all sources ofozone depleting substances(including CFCs, HCl, Cl2,ClONO2, etc.) and a scaledcontribution from other halo-carbons and bromine, takingits ODP into account
ENSO El Niño Southern Oscillation;a large-scale climate variabil-ity in the Pacific region
EP Earth Probe (a NASA satellite)
EPA Environmental ProtectionAgency
EV Epidermodysplasia verrucifor-mis, a rare recessive genetichereditary skin disorderassociated with a high risk ofcarcinoma of the skin
Fe Iron (Fe(II) and Fe(III) beingdifferent oxidation states)
FMI Finnish MeteorologicalInstitute
GHG Greenhouse gas
Glu I A pathogenesis-related (PR)protein
GNA11 Guanine nucleotide-bindingprotein subunit alpha-11; agene coding for proteinsinvolved in various trans-membrane signaling systems
GNAQ Guanine nucleotide-bindingprotein G(q)
GST Glutathione-S-transferase
GWP Global warming potential; ameasure of the warmingeffectiveness of a gas com-pared with CO2
HALS Hindered amine light stabilizer
HCFC Hydrochlorofluorocarbon;interim replacements forCFCs with small ozone
depletion potential (e.g. R22:chlorodifluoromethane,CHClF2) to be phased out
HFC Hydrofluorocarbon; long-term replacements for CFCs
HFO Hydrofluoro-olefin; replace-ments for CFCs. An exampleis 2,3,3,3-tetrafluoropropene(HFO-1234-yf )
Hg Mercury (Hg(0)aq and Hg(II)being different oxidationstates)
HIV Human immunodeficiencyvirus
H2O2 Hydrogen peroxide
HNV Hematopoietic necrosis virus
HONO Nitrous acid
HPV Human papillomavirus
HSV Herpes simplex virus
HY5 Transcription factor HY5;which is a key downstreameffector of the UVR8 (UV-regulatory protein) pathway
IBD Inflammatory bowel disease
Ig Immunoglobulin
IL Interleukin
Ink4a Murine inhibitor of kinase 4aprotein (gene when in italics)
IPCC Intergovernmental Panel onClimate Change
IPF Immune protection factor
kDa Kilodalton
KNMI Dutch National Institute forWeather, Climate and Seis-mology (Netherlands)
L• Lipid radical
LER Lambertian equivalent reflecti-vity
MAAs Mycosporine-like aminoacids
Mb Megabase; equal to 1 millionbase pairs
MC1R Melanocortin 1 receptor; agene coding for proteins
involved in regulating colourof mammalian skin and hair
MDD Minimal vitamin-D dose
MED Minimal erythemal dose
MCC Merkel cell carcinoma
MHC Major histocompatibilitycomplex
MS Multiple sclerosis
mtDNA Mitochondrial DNA
N2O Nitrous oxide (a greenhousegas that is also a source ofNO2)
NAO North Atlantic Oscillation; alarge-scale variation andredistribution of atmosphericmass in the Atlantic regionproducing large changes inthe Northern hemispheredynamics
NASA National Aeronautic andSpace Administration (USA)
NaTFA Sodium trifluoroacetate
NC Nuclear cataract(s)
NCAR National Centre for Atmos-pheric Research (USA)
NH Northern Hemisphere
NIMBUS-7 A NASA satellite
NIVR Netherlands Agency for Aero-space Programmes
NMHCs Non-methane hydrocarbons
NMSC Non-melanoma skin cancer
NO Nitric oxide (an ozone deplet-ing gas)
NO2 Nitrogen dioxide (an ozonedepleting gas)
NOAA National Oceanic and Atmos-pheric Administration (USA)
NOAEL No observed adverse effectlevel; a level of exposurebelow which no adverseeffects are observed in a testorganism, similar to NOAEC
NOEC No observed effect concen-tration; a concentration ofexposure below which no
Photochemical & Photobiological Sciences Editorial
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 10–13 | 11

effects of any kind areobserved in a test organism
NOEL No observed effect level; alevel of exposure belowwhich no effects of any kindare observed in a test organ-ism, similar to NOEC
NOX Nitrogen oxides
NP Nanoparticle
O3 Ozone
OA Organic aerosol
OCA Oculocutaneous albinism
OCS Carbonyl sulfide
ODP Ozone depletion potential;the ratio of the impact onozone of a chemical com-pared to the impact of asimilar mass of CFC-11.Thus, the ODP of CFC-11 isdefined to be 1.0
ODS Ozone depleting substance(s)(e.g. CFCs)
•OH Hydroxyl radical (an impor-tant atmospheric cleaningagent)
OMI Ozone Monitoring Instru-ment (on board the Aurasatellite)
OTR Organ transplant recipient
P Phosphorous
PAH Polycyclic Aromatic Hydro-carbon
PAM Pulse amplitude modulated(fluorescence); a measure ofthe efficiency of photo-synthesis
PAR Photosynthetically ActiveRadiation; 400–700 nmwaveband
PAUR II Photochemical Activity andsolar Ultraviolet Radiationcampaign 2
pCO2 Partial pressure of carbondioxide
PEC Predicted environmentalconcentration
PER Photoenzymatic repair
PFBI Perfluoro-n-butyl iodide; asubstitute for CFCs used as asolvent for cleaning duringthe maintenance of aircrafts
Pg Peta gram (1 × 1012 grams)
PHR1 The gene encoding CPDphotolyase
PLE Polymorphic light eruption
PM Particulate matter (aerosolsin the atmosphere)
PM2.5 Particulate matter in air thatis smaller than 2.5 μm and isinhaled deeper into thelungs than larger particles(PM10)
PM10 Particulate matter in air thatis smaller than 10 μm
PNEC Predicted no effectconcentration
POC Particulate organic carbon
POM Particulate organic matter
POP Persistent organic pollutant
ppm(ppb)
Parts per million; a mixingratio of 1 molecule of a sub-stance per million moleculesof air. Similarly, ppb is partsper billion, one molecule perbillion molecules of air
PR Pathogenesis-related protein
PS Polysulphone; a thermoplasticthat contains sulfur and isresistant to high temperatures
PSC Posterior subcapsular catar-act(s)
PSC Polar stratospheric cloud (icecrystals which form at highaltitudes in Polar regionswhen the temperature isbelow a critical threshold)
PSI Photosystem I
PSII Photosystem II
Ptc Murine patch protein (genewhen in italics)
PTCH Human patch protein (genewhen in italics)
QBO Quasi biennial oscillation (ashift in wind patterns –
especially over the tropics –
with a period of approxi-mately 2.2 years)
RA Rheumatoid arthritis
RAC1 Ras-related C3 botulinumtoxin substrate (gene when initalics); mutations of thegene are found in melanoma
Radiativeforcing
A measure of the influence afactor (e.g. GHGs, ice albedo,tropospheric aerosols, etc.)has in altering the balance ofincoming solar and outgoinginfrared irradiance (W m−2)in the Earth–atmospheresystem. It is an index of theimportance of the factor as apotential climate changemechanism. Radiativeforcing is approximately pro-portional to temperaturechanges at the Earth’ssurface, so a positive radia-tive forcing is associated withheating in the troposphere
RAF Radiation amplificationfactor (a measure of sensi-tivity to ozone change)
RCP Representative concentrationpathways; scenarios forfuture climate resulting fromdifferent combinations ofeconomic, technological,demographic, policy, andinstitutional futures, definedby their total radiativeforcing (cumulative measureof human emissions ofGHGs from all sourcesexpressed in W m−2) pathwayand level by 2100
ROS Reactive oxygen species (•OHfor example)
RR Relative risk; usually ofincreased risk above back-ground of contracting adisease. The RR of backgroundincidence of the disease is 1
RT Radiative transfer
Editorial Photochemical & Photobiological Sciences
12 | Photochem. Photobiol. Sci., 2015, 14, 10–13 This journal is © The Royal Society of Chemistry and Owner Societies 2015

SAGE Stratospheric Aerosol andGas Experiment; a satellite-based instrument
SCC Squamous cell carcinoma
SCCC Squamous cell carcinomaof the cornea andconjunctiva
SD (SE) Standard Deviation; ameasure of the variance of avalue. Standard Error is asimilar term
SED Standard erythemal dose
SH Southern Hemisphere
SPF Sun protection factor
SOD Superoxide dismutase
SODIS Solar disinfection
SOX Oxides of sulfur
SSA Single scattering albedo;quantifies the absorptionefficiency of aerosols
STAT Signal transducer and activa-tor of transcription; a genethat regulates many aspectsof growth, survival and differ-entiation in cells
SZA Solar zenith angle in degrees(= 90°, the solar elevationangle from the horizontal)
TB Tuberculosis
TFA Trifluoroacetic acid
Th1 T-helper 1
Th2 T-helper 2
TiO2 Titanium dioxide
TOC Total ozone column
TOMS Total Ozone MappingSpectrometer; a satellite-based instrument
Treg cell T-regulatory cell
Tropos-phere
Lowest part of the Earth’satmosphere (0–16 km)
UCA Urocanic acid
UML Upper mixed layer; of waterin lakes or the ocean
UNEP United Nations EnvironmentProgramme
UV Ultraviolet; wavelengths from100 nm to 400 nm. Ozoneand other atmospheric gasesprogressively absorb moreand more of the radiation atwavelengths less than320 nm. Only those greaterthan 290 nm are transmittedto the Earth’s surface
UV-A Electromagnetic radiation ofwavelengths in the 315 to400 nm range (weaklyabsorbed by ozone)
UV-B Electromagnetic radiation ofwavelengths in the 280 to315 nm range (stronglyabsorbed by ozone)
UV-C Electromagnetic radiation ofwavelengths in the 100 to280 nm range (solar UV-C isnot transmitted to theEarth’s surface)
UVeff UV irradiance weighted bythe spectral sensitivity of aneffect, integrated overwavelength
UVery Erythemally-weighted UVirradiance; where the irradi-ance is weighted by theerythemal action spectrum
UVI UV index; a measure oferythemally-weighted UV forproviding information to thepublic. UVI values greaterthan 10 are considered“extreme” by the WHO. IfUVery is specified in units ofW m−2, then UVI = 40 ×UVery)
UVR Ultraviolet radiation
UVR8 UV-regulatory protein
VDR Vitamin D receptor
VOC Volatile organic compound(s)
WHO World Health Organization
WMO World MeteorologicalOrganization
WOUDC World Ozone and UV DataCentre
XP Xeroderma pigmentosum;recessive genetic disorder ofDNA repair in which theability to repair damagecaused by UV radiation iscompromised
ZnO Zinc oxide
Photochemical & Photobiological Sciences Editorial
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 10–13 | 13

Photochemical &Photobiological Sciences
EDITORIAL
Cite this: Photochem. Photobiol. Sci.,2015, 14, 14
DOI: 10.1039/c4pp90042a
www.rsc.org/pps
Environmental effects of ozone depletion and itsinteractions with climate change: 2014 assessment
Executive summary†‡
Ozone depletion andclimate change• The Montreal protocol continuesto be effective
The Scientific Assessment Panel of theMontreal Protocol on Substances thatDeplete the Ozone Layer concludes thatthe atmospheric abundance of most con-trolled ozone-depleting substances(ODSs) is decreasing. There are severalindications that the global ozone layer isbeginning to recover from ODS-induceddepletion. However, the variability of theatmosphere and the influence of climatechange have hindered a definitive attri-bution of the observed global ozoneincreases since 2000 to the concomitantODS decreases. In Antarctica, largeozone depletion continues to occureach year. In the Arctic, ozone depletionis generally less pronounced than in
Antarctica but more variable: the veryhigh stratospheric ozone concentrationsobserved in the spring of 2010 werefollowed by record-low concentrations inspring 2011.
• As a result of the success of theMontreal Protocol in limiting ozonedepletion, changes in UV-B irradiancemeasured at many sites since the mid-1990s are due largely to factors otherthan ozone
Increases in UV-B irradiance (280–315 nm)ranging from 5 to 10% per decade havebeen reported for several northern mid-latitude sites, caused predominantly byreductions in cloudiness and aerosols.However, at some northern high latitudesites, UV-B irradiance has decreasedduring that period mainly due toreduction in snow- or ice-cover. Becauseof the large natural variability, anyresponses of UV-B irradiance to stabili-sation of the concentrations of strato-spheric ozone and possible beginning ofa recovery are not yet detectable in themeasurements.
• Large short-term increases in UV-Birradiance have been measured at somelocations in response to episodicdecreases of ozone at high latitudes
For example, the low ozone in spring2011 in the Arctic increased the erythe-mal (sunburning) dose averaged over theduration of the low-ozone period by40–50% at several Arctic and Scandina-vian sites. Corresponding increases overCentral Europe were estimated to beabout 25%.
• Future levels of UV-B irradiance at highlatitudes will be determined by therecovery of stratospheric ozone and bychanges in clouds and reflectivity of theEarth’s surface
In Antarctica, reductions of up to 40% inmean noontime UV Index (UVI) are pro-jected for 2100 due to the continuingrecovery of ozone. These reductions arecomparable in magnitude with theincreases in UVI that occurred in thepast due to ozone depletion. Because ofthe anticipated increases in cloud cover,the UVI is projected to decrease byup to 7% at northern high latitudes.Reductions in surface reflectivity due toice-melt will continue to contribute toreductions in UVI by up to 3% in themargins of the Antarctic continent andby up to 10% in the Arctic, but confi-dence in the magnitude of these effectsis low.
• With continued effectiveimplementation of the MontrealProtocol, future changes in UV-Birradiance outside the Polar regions willlikely be dominated by changes infactors other than ozone
By the end of the 21st century, the effectof the recovery of ozone on UV-B irradi-ance will be very small, leading todecreases in UVI of between 0 and 5%.Additional decreases of up to 3% in theUVI are projected due to the anticipatedincreases in cloud cover. Future changesin UVI would be likely dominated bydecreases in aerosols, resulting inincreases in the UVI, particularly indensely populated areas. For example,
†Electronic supplementary information (ESI) avail-able. Questions and answers about the Environ-mental Effects of the Ozone Layer Depletion andClimate Change: 2014 update. See DOI: 10.1039/c4pp90042a‡List of contributing authors in alphabetical order.A. L. Andrady, P. J. Aucamp, A. Austin, A. F. Bais,C. L. Ballaré, P. W. Barnes, G. H. Bernhard,J. F. Bornman (Co-Chair), M. M. Caldwell, F. R. deGruijl, D. J. Erickson III, S. D. Flint, K. Gao, P. Gies,D.-P. Häder, M. Ilyas, J. Longstreth, R. Lucas,S. Madronich, R. L. McKenzie, R. Neale, M. Norval,K. K. Pandy, N. D. Paul (Co-Chair), M. Rautio, H. HRedhwi, S. A. Robinson, K. Rose, M. Shao (Co-Chair), R. P. Sinha, K. R. Solomon (Secretary),B. Sulzberger, Y. Takizawa, X. Tang, A. Torikai,K. Tourpali, J. C. van der Leun, S.-Å. Wängberg,C. E. Williamson, S. R. Wilson, R. C. Worrest,A. R. Young, and R. G. Zepp.
14 | Photochem. Photobiol. Sci., 2015, 14, 14–18 This journal is © The Royal Society of Chemistry and Owner Societies 2015

increases in the UVI of up to 40% areprojected for parts of Asia, reversing thelarge reductions in UVI that have prob-ably occurred in this region during thesecond half of the 20th century. The con-fidence in these effects of aerosols isvery low due to uncertainties in the pro-jected amounts and optical properties ofaerosols, as well as in future policy onemission controls.
Human health• Changing behaviour with regard to sunexposure by many fair-skinnedpopulations has probably had a moresignificant effect on human health thanincreasing UV-B irradiance due to ozonedepletion
The increase in holiday travel to sunnyclimates, wearing clothing that coversless of the body, and the desire for a tanare all likely to have contributed tohigher personal levels of exposure toUV-B radiation than in previous decades.Such changes in behaviour have bothadverse and beneficial consequences forhealth.
• Immediate adverse effects of excessiveUV-B irradiation are sunburn of theskin and inflammation of the eye(photoconjunctivitis or photokeratitis).Long-term regular low-dose or repeatedhigh-dose exposure to the sun causesmelanoma and non-melanoma (basaland squamous cell) carcinomas of theskin and cataract and pterygium(a growth on the conjunctiva) of the eye
The incidence of each of these skincancers has risen significantly since the1960s in fair-skinned populations, buthas stabilised in recent years in youngerage groups in several countries, perhapsdue to effective public health campaigns.Cataract is the leading cause of blind-ness worldwide.
• The major known beneficial effectof exposure of the skin to solar UVradiation is the synthesis of vitamin D
Vitamin D is critical in maintainingblood calcium levels and is required forstrong bones. People vary in howefficiently their skin makes vitamin D
from sun exposure and perhaps in theirphysiological needs for this vitamin.Vitamin D deficiency might increasethe risk of an array of diseases suchas cancers, autoimmune diseases andinfections. At present it is not clear if thelow level of vitamin D is a cause of thesediseases, occurs as a consequence ofthem, or is a marker of other factors thatpredispose to ill-health.
• Strategies to avoid over-exposure tosolar UV radiation include stayingindoors, seeking shade, wearingprotective clothing, brimmed hats andsunglasses, and applying sunscreens.These methods should aim to balancethe harmful and beneficial effects of sunexposure
Such a balance may be difficult toachieve in practice as the recommendedtime outdoors will differ between indi-viduals, depending on personal factorssuch as skin colour, age, and clothing aswell as on environmental factors such aslocation, time of day, and season of year.Current uncertainties centre on definingan optimal level of vitamin D and theamount and pattern of sun exposurerequired to achieve the optimum indifferent individuals. Thus, devisingappropriate health messages for thepublic at the present time is notstraightforward.
• Climate change may affect personalsun-exposure behaviour, but the impactis likely to vary according to season andlocation
For example, increasing temperaturesmay lead to decreased time outdoors inclimates where it is already hot, butmore time outdoors in cooler climates.
Terrestrial ecosystems• The effects of UV-B radiation on plantsare influenced by various abiotic andbiotic factors in ways that can have bothpositive and negative consequences onplant productivity and functioning ofecosystems
Ozone depletion, increased exposureto ultraviolet-B radiation, and climatechange affect biological systems, result-
ing in intricate feedbacks and com-plexity. In mid-high latitudes of theSouthern Hemisphere plant productivityhas likely decreased slightly due to theincreased UV radiation as a result of theozone depletion. On the other hand,exposure to UV-B radiation can promoteplant hardiness, and enhance plantresistance to herbivores and pathogens.It can also improve the quality, andincrease or decrease the yields of agri-cultural and horticultural products,with subsequent implications for foodsecurity.
• Exposure to UV-B radiation canincrease or decrease rates ofdecomposition of dead plant matter(litter), depending on prevailing climateand the chemistry and structure ofthe litter
In arid and semi-arid ecosystems (grass-lands, savannas and deserts), photo-degradation generally increases rates ofdecay of plant litter and is now beingconsidered as an important driver ofdecomposition, although uncertaintyexists in quantifying its regional andglobal biogeochemical significance.Changes in the decomposition of plantlitter from exposure to UV-B and alsoUV-A (315–400 nm) and visible radiationhave potential consequences for thecycling and storage of carbon and othernutrients.
• Solar UV radiation has the potential tocontribute to climate change via itsstimulation of emissions of carbonmonoxide, carbon dioxide, methane,and other volatile organic compoundsfrom plants, plant litter and soil surfaces
Mechanisms and sources of emissionsof trace gases have been identified inplants and ecosystems. UV radiationtogether with other abiotic factors, inparticular temperature, stimulates theseemissions. The magnitude, rates andspatial patterns of the emissions remainhighly uncertain at present. These UVradiation processes could increase emis-sions of trace gases that affect the atmos-pheric radiation budget (radiativeforcing) and hence changes in climate.
Photochemical & Photobiological Sciences Editorial
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 14–18 | 15

• While UV-B radiation does notpenetrate into soil to any significantdepth, it can affect a number ofbelowground processes throughalterations in aboveground plant parts,microorganisms, and plant litter
These include modifications of the inter-actions between plant roots, microbes,soil animals and neighbouring plants,with potential consequences for soil fer-tility, carbon storage, plant productivityand species composition.
• Terrestrial ecosystems in the SouthernHemisphere are being affected by theAntarctic ozone ‘hole’
Resultant changes in precipitation pat-terns have been correlated with ecosys-tem changes such as increased treegrowth in Eastern New Zealand andexpansion of agriculture in South-eastern South America. Conversely, inPatagonia and East Antarctica, decliningtree and moss bed growth have beenlinked to reduced availability of water. Afull understanding of the effects ofozone depletion on terrestrial ecosys-tems in these regions should thereforeconsider both UV radiation and climatechange.
Aquatic ecosystems• Climate change and UV radiation affectphytoplankton productivity and speciescomposition of marine ecosystems
Phytoplankton (primary producers) aredecreasing along the West side of theAntarctic Peninsula due to increasedsolar UV-B radiation and rapid regionalclimate change. Changes in ice pheno-logy as well as light and nutrient avail-ability may affect species composition.Organisms mitigate UV-B radiation-induced damage by repair mechanismsor by producing UV-absorbing compounds.
• Interactions between climate changeand UV radiation are having strongeffects on aquatic ecosystems that willchange in the future due to feedbacksbetween temperature, UV radiation andgreenhouse gas concentrations
Higher air temperatures are increasingthe surface water temperatures of
numerous lakes and oceans, with manylarge lakes warming at twice the rate ofair temperatures in some regions.Species composition and distribution ofmany marine ecosystems may changewith warmer oceans. For others, such ascorals, the warming may alter their toler-ance to other stressors. This warmingcan also shift the thermal niche oforganisms towards the pole and causeschanges in community structure.
• Warming of the ocean results instronger stratification that decreases thedepth of the upper mixed layer
The decrease in the depth of the uppermixed layer exposes organisms thatdwell in it to greater amounts of solarvisible and UV radiation which may over-whelm their capability for protectionand repair. Enhanced stratification alsoreduces upward transport of nutrientsacross the thermocline from deeperlayers. In the polar waters, increasingtemperature results in explosions of phyto-plankton growth under the ice andaround the ice edges.
• Increased concentrations ofatmospheric CO2 are continuing tocause acidification of the ocean, whichalters marine chemical environmentswith consequences for marineorganisms
Acidification interferes with the calcifica-tion process by which organisms, suchas phytoplankton, macroalgae and manyanimals including mollusks, zooplank-ton and corals, produce exoskeletonsprotecting themselves from predatorsand solar UV radiation. Consequently,they become more sensitive to UV radi-ation, so that they calcify even less anddecrease their production of biomass.
• Climate change-induced increases inconcentrations of dissolved organicmatter (DOM) in inland and coastalwaters reduce the depth of penetrationof UV radiation
Increased extreme precipitation eventsand enhanced growth of terrestrial vege-tation produce greater fluxes of UV-absorbing DOM from the landscape.This creates a refuge for UV-sensitiveorganisms including some invasive
species. Decreased penetration of UVradiation also reduces the natural dis-infection of surface water containingviruses, pathogens, and parasites.
Biogeochemical cycles• Climate change modulates the effectsof solar UV radiation on biogeochemicalcycles in terrestrial and aquaticecosystems resulting in UV-mediatedpositive or negative feedbacks on climate
For example, where photochemicalpriming plays an important role,changes in continental runoff and icemelting, due to climate change, arelikely to result in enhanced UV-inducedand microbial degradation of dissolvedorganic matter (DOM) and the release ofcarbon dioxide (CO2). Such positive feed-backs are particularly pronounced in theArctic resulting in Arctic amplification ofthe release of CO2 (see next point).
• Solar UV radiation is driving theproduction of substantial amounts ofcarbon dioxide from Arctic waters
The production is enhanced by thechanges in rainfall, and melting of ice,snow and permafrost, which lead tomore organic material being washedfrom the land into Arctic rivers, lakesand coastal oceans. Solar UV radiationdegrades this organic material, whichstimulates CO2 and CO emissions fromthe water bodies, both directly and byenhanced microbial decomposition.New results indicate that up to 40% ofthe emissions of CO2 from the Arcticmay come from this source, much largerthan earlier estimates.
• The changes in climate associated withthe Antarctic ozone ‘hole’ includechanges to wind patterns, temperatureand precipitation across the SouthernHemisphere
More intense winds lead to enhancedwind-driven upwelling of carbon-richdeep water and less uptake of atmos-pheric CO2 by the Southern Ocean, redu-cing the oceans’ potential to act as acarbon sink (less sequestering of carbon).These winds also transport more dustfrom drying areas of South America
Editorial Photochemical & Photobiological Sciences
16 | Photochem. Photobiol. Sci., 2015, 14, 14–18 This journal is © The Royal Society of Chemistry and Owner Societies 2015

into the oceans and onto the Antarcticcontinent. In the oceans this canenhance iron fertilisation resulting inmore plankton and increased numbersof krill. On the continent the dust maycontain spores of novel microbes thatincrease the risk of invasion of non-indigenous species. The ozone ‘hole’ hasalso helped to keep East Antarctica cold,but conversely has helped to make theMaritime Antarctic region one of thefastest warming regions on the planet.These climate-related impacts of ozonedepletion on ecosystems may also interactwith changing UV radiation, leading totipping points.
• The carbon cycle is strongly influencedby interactions between droughts andthe intensity of UV-radiation at theEarth’s surface
Increased aridity due to climate changeand severity of droughts will change theamount of plant cover, thereby increasingUV-induced decomposition of dead plantmatter (plant litter). These increasedlosses could have large impacts on terres-trial carbon cycling in arid ecosystems.
• Lignin present in all terrestrialvegetation plays a significant role in thecarbon cycle, sequestering atmosphericcarbon into the tissues of perennialvegetation
Although it is well known that lignin isone of the components of dead vege-tation most resistant to biotic decompo-sition, new results have shown thatlignin is readily decomposed withexposure to solar UV radiation. Conse-quently, UV-induced degradation ofplant litter is correlated with its lignincontent, reducing long-term storage ofcarbon in terrestrial systems.
Air quality• UV radiation is an essential driver forthe formation of photochemical smog,which consists mainly of ground-levelozone and particulate matter. Recentanalyses support earlier work showingthat poor outdoor air quality is a majorenvironmental hazard
Greater exposures to these pollutantshave been linked to increased risks of
cardiovascular and respiratory diseasesin humans and are associated globallywith several million premature deathsper year. Ozone also has adverse effectson yields of crops, leading to loss ofbillions of US dollars each year. Thesedetrimental effects may also alter bio-logical diversity and affect the functionof natural ecosystems.
• Future air quality will depend mostlyon changes in emission of pollutantsand their precursors; changes in UVradiation and climate will alsocontribute
Significant reductions in emissions,mainly from the energy and transpor-tation sectors, have led to improved airquality in many locations. Air qualitywill continue to improve in thosecities/states that can afford controls,and worsen where the regulatory infra-structure is not available. Futurechanges in UV radiation and climatewill alter the rates of formation ofground-level ozone and some particu-late matter and must be considered inpredictions of air quality and conse-quences for human and environmentalhealth.
• Decreases in UV radiation associatedwith the recovery of stratospheric ozonewill, according to recent globalatmospheric model simulations, lead toincreases in ground-level ozone overlarge geographic scales
If correct, this would add significantly tofuture ground-level ozone trends.However, the spatial resolution of thesemodels is insufficient to inform policy,especially for urban areas.
• UV radiation affects the atmosphericconcentration of hydroxyl radicals, •OH,which are responsible for the self-cleaning of the atmosphere
Recent measurements confirm that on alocal scale, •OH radicals respond rapidlyto changes in UV radiation. However, onlarge (global) scales, models differ intheir predictions by nearly a factor oftwo, with consequent uncertainties forestimating the atmospheric lifetimeand concentrations of greenhouse gasesand key air pollutants. Projections of
future climate need to consider theseuncertainties.
• No new negative environmental effectsof the substitutes for ozone depletingsubstances or their breakdown-productshave been identified
However, some substitutes for ozonedepleting substances will continue tocontribute to global climate change ifconcentrations rise above current levels.
Materials• A trend towards environmentallysustainable materials in building hasincreased the use of wood andwood–plastic composites
Despite this trend, the use of rigid PVC,the most-used plastic in building, willcontinue to be popular at least in themedium term. Improvements are beingdeveloped that make PVC easier toprocess and environmentally friendly. Theeffects of solar UV radiation and climatechange on the lifetime of PVC buildingproducts continue to be a concern.
• The role of solar UV radiation increating microplastics debris in theoceans from the weathering of plasticlitter on beaches is an emergingenvironmental issue
These microplastic particles concentratetoxic chemicals dissolved in seawaterand are ingested by zooplankton, thusproviding a potential mechanism for thetransfer of pollutants into the marinefood web. While the process has notbeen studied in any great detail, the pro-duction of microplastics will likelyincrease at high solar UV-B radiationlevels and/or elevated temperatures.
• Nanoscale inorganic fillers can providesuperior stability against solar UVirradiation relative to conventional fillersin coatings and plastics
Nanoparticle fillers in coatings, especiallythose in clear-coatings on wood or fibre-coatings of textiles, also provideenhanced stability. With nanoparticlesthat absorb UV radiation, such as themineral rutile, the stabiliser effect isparticularly evident. The benefits of
Photochemical & Photobiological Sciences Editorial
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 14–18 | 17

nanofillers in bulk plastics, however, areless clear and more information isneeded to assess their efficacy. Nanofil-lers may provide a low-cost means of sta-bilising some polymer and wood-basedproducts and help increase service life-times in the face of variations in UV radi-ation or climate change.
• Clothing and glass can provideprotection against exposure to solarUV radiationTextile fabrics block personal exposureto solar UV radiation, whereas glassusually blocks mainly UV-B radiation.The effectiveness of specific fabricsdepends on the weave characteristics but
can be further improved by surface-treat-ing the fibres with a UV absorber.Glazing for windows is being developedto further improve their thermal pro-perties and also results in increased fil-tering of the UV radiation with benefitsto the health of humans and indoorcomponents of buildings and artwork.
Editorial Photochemical & Photobiological Sciences
18 | Photochem. Photobiol. Sci., 2015, 14, 14–18 This journal is © The Royal Society of Chemistry and Owner Societies 2015

Photochemical &Photobiological Sciences
PERSPECTIVE
Cite this: Photochem. Photobiol. Sci.,2015, 14, 19
Received 20th October 2014,Accepted 20th October 2014
DOI: 10.1039/c4pp90032d
www.rsc.org/pps
Ozone depletion and climate change: impacts onUV radiation
A. F. Bais,*a R. L. McKenzie,b G. Bernhard,c P. J. Aucamp,d M. Ilyas,e S. Madronichf
and K. Tourpalia
We assess the importance of factors that determine the intensity of UV radiation at the Earth’s surface.
Among these, atmospheric ozone, which absorbs UV radiation, is of considerable importance, but other
constituents of the atmosphere, as well as certain consequences of climate change, can also be major
influences. Further, we assess the variations of UV radiation observed in the past and present, and provide
projections for the future. Of particular interest are methods to measure or estimate UV radiation at the
Earth’s surface. These are needed for scientific understanding and, when they are sufficiently sensitive,
they can serve as monitors of the effectiveness of the Montreal Protocol and its amendments. Also
assessed are several aspects of UV radiation related to biological effects and health. The implications for
ozone and UV radiation from two types of geoengineering methods that have been proposed to combat
climate change are also discussed. In addition to ozone effects, the UV changes in the last two decades,
derived from measurements, have been influenced by changes in aerosols, clouds, surface reflectivity,
and, possibly, by solar activity. The positive trends of UV radiation observed after the mid-1990s over
northern mid-latitudes are mainly due to decreases in clouds and aerosols. Despite some indications
from measurements at a few stations, no statistically significant decreases in UV-B radiation attributable to
the beginning of the ozone recovery have yet been detected. Projections for erythemal irradiance (UVery)
suggest the following changes by the end of the 21st century (2090–2100) relative to the present time
(2010–2020): (1) Ozone recovery (due to decreasing ozone-depleting substances and increasing green-
house gases) would cause decreases in UVery, which will be highest (up to 40%) over Antarctica.
Decreases would be small (less than 10%) outside the southern Polar Regions. A possible decline of solar
activity during the 21st century might affect UV-B radiation at the surface indirectly through changes
induced in stratospheric ozone. (2) The projected changes in cloud cover would lead to relatively small
effects (less than 3%), except at northern high latitudes where increases in cloud cover could lead to
decreases in UVery by up to 7%. (3) Reductions in reflectivity due to the melting of sea-ice in the Arctic
would lead to decreases of UVery by up to 10%, while at the margins of the Antarctic the decreases would
be smaller (2–3%). The melting of the sea-ice would expose the ocean surface formerly covered by ice to
UV-B radiation up to 10 times stronger than before. (4) The expected improvement of air-quality and
reductions of aerosols over the most populated areas of the northern hemisphere may result in 10–20%
increases in UVery, except over China where even larger increases are projected. The projected aerosol
effect for the southern hemisphere is generally very small. Aerosols are possibly the most important factor
for future UV levels over heavily populated areas, but their projected effects are the most uncertain.
Introduction
For the purposes of the current assessment (2010–2014),which addresses the negative and positive effects of solar UVradiation on humans, terrestrial and aquatic ecosystems,materials, and air quality (see companion papers), we assessthe short- and long-term changes in ambient UV radiation atthe Earth’s surface resulting from changes in atmosphericozone and climate. The effects of ozone on climate andclimate on ozone are also discussed. Absorption by ozone is
aLaboratory of Atmospheric Physics, Aristotle University of Thessaloniki,
Campus Box 149, 54124 Thessaloniki, Greece. E-mail: [email protected] Institute of Water and Atmospheric Research, NIWA Lauder,
PB 50061 Omakau, Central Otago, New ZealandcBiospherical Instruments Inc., 5340 Riley Street, San Diego, California, 92110-2621,
USAdPtersa Environmental Management Consultants, PO Box 915751, Faerie Glen, 0043,
South AfricaeSchool of Environmental Engineering, University Malaysia Perlis, Kangar, MalaysiafNational Center for Atmospheric Research, Boulder, Colorado, 80307-3000, USA
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 19

the dominant factor controlling the levels of surface UV-B(280–315 nm) radiation for cloud-free and low-aerosol con-ditions. With the continuing success of the amended andadjusted Montreal Protocol in reducing the concentrations ofozone depleting substances (ODSs), the focus is now on thedetection of possible decreases in UV-B radiation in responseto the first signs of recovery of the ozone layer. Changes in theclimate caused by the increasing concentrations of greenhousegases may also affect the UV radiation at the Earth’s surfaceindirectly, as detailed below.
Current status of atmospheric ozone
Since the last assessment of ozone depletion1 efforts to quan-tify the geographic and temporal variability of ozone have con-tinued through ground- and satellite-based measurements.This extension of the observation of ozone by four years hasincreased the statistical confidence in the estimated long-termchanges in total ozone column (TOC). There are indicationsthat the global ozone layer is beginning to recover fromthe depletion caused by ODSs. However, the variability ofthe atmosphere, the uncertainty of measurements, and theinfluence of climate change prevent unequivocal attributionof the observed increases in ozone since 2000 to decreasesin ODSs.2,3
Ozone at mid-latitudes and the tropics
The present (2008–2012) mean values of ozone relative to the1964–1980 mean values are smaller by ∼3.5% in the NorthernHemisphere mid-latitudes (35° N–60° N) and by ∼6% in thesouthern hemisphere mid-latitudes (35° S–60° S). In thetropics (20° S–20° N), no significant changes have occurred intotal ozone over this period. The observed average changes intotal ozone over time, relative to the 1998–2008 mean values,in different latitude bands are shown in Fig. 1.
Following the decline in total ozone between the 1960s and1990s, the levels of total ozone outside the polar regions havestopped decreasing since the late 1990s, consistent with theslow decline of ODSs over the same period.4 Several datasetsindicate that total ozone has increased by ∼1% since 2000 inthe latitude band 60° S–60° N in response to stratosphericozone recovery. However, there is disagreement about the mag-nitude and statistical significance of this increase.2 Presum-ably any increase in ozone would have resulted in acorresponding decrease in the surface UV-B radiation at theEarth’s surface, by analogy with the increases in UV-B radi-ation observed for the ozone decline.5
The amounts of total ozone are subject to large year-to-yearvariability caused by variations in atmospheric circulation.Examples include the unusually high values in 2010 and low
Fig. 1 Total column ozone annual mean deviations relative to the 1998–2008 mean for different latitude bands as derived from different ground-and space-based datasets. Adapted from WMO 2015.2
Perspective Photochemical & Photobiological Sciences
20 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

values in 2011 in the northern hemisphere mid and high lati-tudes. Because of these large variations, the relatively smallincreases in total ozone expected after the recent decline ofODSs are still not statistically significant.6,7 Without theremoval of these circulation effects, the attribution of ozonerecovery to decreases in ODSs would not be detectable even inAntarctica before the period of 2017–2021.8,9 The separation ofrecent changes of ozone into the contributions by ODSs, green-house gases (GHG), and natural low-frequency variabilityremains challenging. For example, after removing the vari-ations caused by the solar variability from satellite data fortotal ozone, the residuals averaged over the band 60° S–60° Nshow a decline of about 5% between 1980 and 2000, followedby a partial recovery after about 2000.10 However, significantsporadic reductions in total ozone have been observed in geo-graphically localised areas at high latitudes in South America11
and in northern high- and mid-latitudes,12 showing that theozone depletion problem is not yet fully solved.
Analysis of the variability of extreme values in the longesttime series of total ozone (Arosa, Switzerland, starting in the1920s) has revealed an increasing frequency of both low-ozoneand high-ozone events, which dominate trends in the 1970sand 1980s. After the removal of the extreme events from theoriginal time series, the overall downward trend in the period1970–1990 is reduced from 2.4% per decade to 0.9% perdecade.13,14 However, since the extremes are relative to thelong-term climatology, the values that were removed alsoinclude small values that have been caused by the increase inODSs during that period. A similar study for Sonnblick,Austria, over a shorter period (1994–2011), revealed a declinein the frequency of low-ozone events and an increase in high-ozone events.15
In the northern hemisphere, the increase in troposphericozone from precursors (CO, NOx, and hydrocarbons) since pre-industrial times nearly equals the decrease in stratosphericozone from increases in ODSs. In the southern hemisphere,the decrease in stratospheric ozone dominates the total ozonecolumn. Currently, the impact of increasing N2O, a source gasthat leads to the formation of reactive nitrogen in the strato-sphere, on ozone almost cancels the globally averagedincreases from climate change effects16 (see discussion below).
Our present understanding is that the present levels ofglobal total ozone are still less than the mean levels in theperiod 1960–1980. Over most latitudes, total ozone values havestabilized after the mid-1990s, but the year-to-year variabilityhas increased relative to the period before the mid-1990s, pre-cluding the unequivocal detection of possible increasesexpected from the observed decline of ODSs concentrations.
Ozone at high latitudes. Over the high latitudes (63°–90°) ofboth hemispheres, ozone depletion continues to occur duringwinter and spring. Compared to the average values before1980, the 2010–2013 mean total ozone is lower by ∼27% in thesouthern hemisphere in October and by ∼10% in the northernhemisphere in March.2
The Antarctic ozone hole has continued to appear eachspring. The evolution of total ozone in Antarctica over the last
decade has been significantly affected by variations in strato-spheric temperature and circulation, which have masked theeffect of the decreases in ODSs since the early 2000s. With anaccurate account of circulation-induced changes, a small(3–8%) increase in total ozone over Antarctica during the lastdecade is now apparent.17–19 Even without accounting for thesecirculation effects, reductions in the severity of ozone depletionin Antarctica since the 1990s are now becoming clear.2
However, uncertainties in methods and measurements precludea definite conclusion that the recent increases of ozone inAntarctica are due to declining concentrations of ODSs alone.2
Any reversal of total ozone trends is not yet apparent in theArctic spring, where the largest ever ozone loss was observedin 2011.20,21 The concentration of ozone in the Arctic strato-sphere during the spring of 2011 was the lowest since satelliterecords began in 1979. The minimum daily average columnozone (297 Dobson Units (DU)) was observed in March 2011.22
This value is 18 DU below the previous record-low observed inMarch 2000, and 100 DU (25%) below the average for1979–1988. At some locations and times, the amounts of totalozone observed between February and April 2011 were morethan 50% below the climatological mean.23 The fraction of theArctic vortex area with total ozone below 275 DU is typicallynear zero for March, but reached nearly 45% in March 2011.12
In that year, the minimum total ozone in spring was continu-ously below 250 DU for about 27 days, and values between 220and 230 DU were observed for about one week in late March,12
leading to increases in UV-B radiation, as discussed later.This large chemically-mediated loss of ozone in the Arctic
was the result of an unusually prolonged cold period in thelower stratosphere and an anomalously strong Arctic vortex,which weakened the transport of ozone from middle lati-tudes24 and facilitated the formation of polar stratosphericclouds (PSCs). These clouds provide surfaces for the hetero-geneous reactions that activate stratospheric chlorine, whichin turn destroys ozone in catalytic cycles. Temperatures belowthe threshold for the formation of PSCs of about −77° Coccurred between December 2010 and early April 2011. Over80% of the ozone present in January from about 18 to 20 kmaltitude had been chemically destroyed by late March, which isroughly twice that in the previous record-setting winters of1996 and 2005.12 The anomalies for 2011 due to chemistry andtransport stand out as extreme events, greater than 2σ (stan-dard deviation), while the total anomaly was nearly 3σ.24 Theamount of ozone loss and the chemistry of the Arctic strato-sphere in the spring of 2011 was remarkably similar to thatcommonly observed in Antarctica, justifying the conclusionthat there was an Arctic ozone hole in 2011.25
There are indications that this Arctic ozone depletion eventcontributed to the smaller total ozone values recorded at mid-latitude locations. Measurements at 34 European stationsrevealed that the total ozone over Western Europe from lateMarch to late April 2011 was 15%–25% less than the meanvalue for this period over the last decade.26
There is no indication that the extreme meteorological con-ditions that led to the loss of ozone in the Arctic in 2011 were
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 21

driven by climate change.21 Severe ozone depletion, such asoccurred in 2011, or even worse, could possibly happen overthe next decades under similar conditions of long-lasting coldstratospheric temperatures.27 The effect of these largereductions of ozone on surface UV-B radiation is discussedlater.
Effects of depletion and recovery of ozone on climate
Changes in stratospheric ozone influence the climate bothdirectly through radiative effects and indirectly by affectingstratospheric and tropospheric circulation.2 Ozone depletionwas the dominant driver of the globally averaged cooling thatoccurred in the lower stratosphere during the last part of the20th century,28 but no statistically significant temperaturechange has occurred there since the mid-1990s.29,30 Episodicwarming over this period has occurred due to aerosols aftermajor volcanic eruptions.30 Cooling of the stratosphere due toozone depletion over Antarctica is, in turn, the dominantdriver of circulation changes in the southern hemisphere tro-posphere during summer (see also ref. 31 and 32). Accordingto model simulations, these changes have led to changes insurface wind patterns, pole-ward shifting of the midlatitudemaximum of precipitation33 and increases of moisture inthe subtropics.33–38 Opposite effects for the southern hemi-sphere circulation and climate would be expected for thefuture from the projected recovery of stratospheric ozone.However, increases in GHGs would compensate partly for theseozone recovery-induced effects on climate.39–44 For a moredetailed discussion on the effects of ozone depletion and recov-ery on climate see the WMO Scientific Assessment of OzoneDepletion: 2014.2
Indirect effects of climate change on surface UV radiation
Climate change may have indirectly influenced the levels of UVradiation in the past by altering the amounts of ozone, UV-absorbing tropospheric gases, aerosols, and clouds in theatmosphere. These influences will likely continue into thefuture.2,45 Future changes in the reflectivity of the Earth’ssurface, either due to the melting of sea-ice and ice-caps athigh latitudes46 or due to reduced snow-cover, may also beimportant. Cooling of the stratosphere resulting fromincreased concentrations of CO2 and other GHGs will lead togreater concentrations of ozone in the future because thedestruction rates of ozone in the cooler middle and upper strato-sphere, outside the Polar Regions, will decrease. However, athigh latitudes, where temperatures in the lower stratospheremay drop below the threshold for the formation of PSCs, het-erogeneous chemistry on the surfaces of these clouds in thepresence of chlorine can potentially lead to a rapid loss ofozone. An example of these processes is the annually recurringspringtime Antarctic ozone-hole. For the Arctic, chemistry-climate models (CCM) suggest that while in the near futurethere is a chance of low springtime ozone in individual years,there is no indication of a formation of regular Arctic ozoneholes.47 Although a much wider area may be susceptible to
heterogeneous processes later this century, the projectedsmaller concentrations of chlorine by that time are expected tomoderate the potential for loss of ozone.
Increasing concentrations of GHGs will increase thestrength of the primary large-scale transport and overturningof the upper atmosphere (the Brewer–Dobson circulation),leading to decreases of ozone in the tropics and increasesoutside the tropics. Emissions of CH4 and N2O would alsoaffect the evolution of global stratospheric ozone, particularlyin the second half of the 21st century, when concentrations ofODSs are expected to be small.2 The increases of ozone outsidethe tropics caused by rising concentrations of GHGs will bepartly offset by additional chemical destruction arising fromanthropogenic emissions of N2O.
48
Clouds respond to climate-forcing mechanisms in multipleways, and the feedback of clouds can be positive or negative.Climate change is projected to reduce the amount of clouds inthe future over most of the tropics and mid-latitudes, withmostly reductions in the amount of low clouds.45 Changes inclouds in the marine boundary layer are most uncertain. Overhigher latitudes (>50°), increases in the fraction of cloud coverand optical depth are projected. This would increase theamount of solar UV radiation scattered back to space and,therefore, reduce the UV radiation reaching Earth’s surface.Furthermore, clouds play a critical role in the climate system,since they can increase the planetary albedo, thereby counter-acting global warming, but they can also contribute towarming of the troposphere through absorption of infraredradiation emitted from the surface.
Reductions in the fraction of ice and snow cover, as well aschanges in their characteristics (e.g., thickness of ice, depth ofsnow) may influence the exposure of ecosystems to solar UVradiation. This is mainly through: (a) less UV radiation reach-ing the Earth’s surface due to reduced surface reflectivity (see“Surface reflectivity”, below) leading to less exposure; and/or(b) greater exposure to UV radiation for systems formerlyunder the ice or snow if that protective cover diminishes. Thecomplete removal of ice would lead to a much greater exposureto UV-B radiation, because the transmittance of UV-B radiationthrough the existing snow-covered ice is much smaller than1%.49 Recent observations in the Arctic suggest that thesummer melt season starts earlier, the winter freeze occurslater, the area of the ice has decreased, and more ice is failingto last through the summer.50–53 Under such conditions, it hasbeen estimated that over the course of one melt season nearly40% more solar radiation would enter the ocean system.54 Inrecent years the extent of the northern ocean’s ice cover hasdeclined, with large interannual variability;55 while in Antarc-tica the sea-ice has been expanding since the 1980s.51
The combined direct or indirect effects of these climatechange-related factors are likely to influence the levels of solarUV radiation in the future and modulate the effects of the pro-jected recovery of ozone. This interaction, which depends onlatitude and on the emissions of GHGs, increases the complex-ity of assessing the future levels of solar UV radiation at theEarth’s surface. Projections for these factors by climate models
Perspective Photochemical & Photobiological Sciences
22 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

can be used to estimate the UV radiation in the future;however, with large uncertainty, as discussed later.
Other factors affecting UV radiation
As UV radiation propagates through the atmosphere, inaddition to being affected by ozone, it is modified throughabsorption and scattering by atmospheric constituents, includ-ing aerosols and clouds, and by reflections on the Earth’ssurface. The effects of the most important factors are dis-cussed in the following sections based on established knowl-edge and on new findings. The effects are discussed in orderto assess the relative importance of these factors on the UVirradiance that reaches the surface in the context of ozone andclimate changes. In addition to UV-B irradiance, the erythe-mally weighted irradiance (UVery) and the UV Index (UVI), bothdefined below in “Biological effects of UV radiation”, are dis-cussed in the following as these quantities appear frequentlyin the cited literature.
Aerosols
Aerosols (particles suspended in the atmosphere) interactwith solar photons and thus can have a significant effect onthe atmospheric transmission of solar radiation (see alsoMadronich et al.56). These particles may be natural (e.g., wind-generated dust and sea salt), anthropogenic (e.g., sulfate,soot, and organic particles), or a mixture of both. The par-ticles scatter and absorb sunlight, with relative probabilitiesthat are complex functions of their size, shape, and chemicalcomposition. They have important effects on air quality andclimate, and a considerable body of knowledge has been deve-loped on their sources, properties, and sinks.57,58 Obser-vational methods include the evaluation of trends invisibility,59–64 in situ determination of size-resolved chemicaland thermodynamic properties,65–68 and remote global-scaledetection from ground-based networks and satellite plat-forms. The AERONET network provides total (scattering +absorption) aerosol optical depth, τ, at a wavelength of340 nm as well as several visible wavelengths; but absorptionoptical depths, τabs, are only available at wavelengths of440 nm and longer.69,70 Satellite-based instruments measur-ing aerosols include the MISR, MODIS and CALIOP.71–73
Global climatologies of aerosols have been developed basedlargely on satellite observations.74,75
Many observations have documented reductions in ground-level UV irradiance in the presence of aerosols.5,76–78
Reductions range from a few percent or less in non-pollutedlocations, such as New Zealand,79,80 to over 50% in pollutedcities, such as Mexico City,81 and can be more than 90% forbiomass burning aerosols, such as in Russia in 2010.82 Thereductions are typically greater at UV than at visible wave-lengths, implying that the aerosol optical depth (AOD) is largerat these wavelengths as well. Quantitative effects depend onaerosol type, and enhancements compared with clear-skiesmay even occur in some conditions, such as in bright scatter-
ing hazes.83 Extrapolation from visible wavelengths is oftenbased on a simple power model for AOD:
τ / λ�α;
where α, the Ångström exponent, parameterizes the strength ofthe wavelength dependence and has typical values between 0.5and 2.0 at visible wavelengths.69 Extrapolation to UV wave-lengths is often a reasonable approximation for scattering butless so for absorption, which is more dependent on chemicalcomposition. Some aerosols (e.g., sea salt, sulfate, and nitrate)have negligible absorption at visible as well as at UV-A(315–400 nm) and UV-B wavelengths. For dust and soot, theabsorption spectrum is sufficiently broad that UV propertiescan be estimated by extrapolation from visible wavelengths.However, for organic aerosols the state of knowledge is extre-mely poor, as these particles, depending on their origin andenvironmental conditions (e.g., humidity), have highly variablechemical composition. So called “brown carbon,” which ismostly composed of combustion-derived organic aerosols, isnow recognized as a significant contributor to climate radiativeforcing due to its absorption of solar radiation at visible wave-lengths,84 and is likely to have even larger effects on UVspectra. Measurements on organic particles derived from theburning of biomass show Ångström exponents for absorptionas high as 6–7 when extrapolated into the UV range,85,86 andUV mass absorption coefficients in the range 1–10 m2 g−1,with the latter value approaching that of black carbon.
The total AOD includes both scattering and absorption, butit is predominately the absorption that is most important inreducing the intensity of UV irradiance at the Earth’s surface.For example, decreases in AOD account for 4.2% of the UV-Airradiance increase at Thessaloniki during 1998–2006, whilethe additional 2% increase can only be explained if the absorp-tion efficiency of aerosols has also decreased over thatperiod.87 The relative importance of scattering is defined bythe single scattering albedo (SSA),
ωo ¼ τscaτsca þ τabs
¼ τscaτ
;
so that the scattering component is ωο × τ while the absorptioncomponent is (1 − ωο) × τ. Based on radiative transfer modelcalculations (TUV, http://acd.ucar.edu/TUV) with typical inputparameters (TOC: 300 DU, solar zenith angle (SZA): 20°,surface reflectivity: 0.05), Fig. 2 illustrates how clear-skysurface erythemal irradiance (UVery) depends on the totalAOD and on SSA, for different target orientations. With non-absorbing aerosols (ωο = 1), reductions are small and enhance-ments can even occur for vertically oriented cylindrical sur-faces. In contrast, strongly absorbing aerosols (ωο = 0.6) causesignificant reductions in radiation regardless of the targetgeometry.
Direct measurements of ωο at UV wavelengths are difficultbecause the absorption by aerosols co-occurs with scattering,and with absorption by gases, especially ozone, but also NO2,or SO2 at UV-B wavelengths. Fig. 3 presents a summary of suchstudies. Values below 400 nm cluster near ωο ≈ 0.8–0.9, which
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 23

is below the typical visible values of about 0.9 or larger. Thedifferent results are almost certainly due to different aerosoltypes present, and more recent studies noted the strongabsorption of UV radiation in biomass-burning aerosols.82,85,88
A likely explanation for this enhanced absorption of UVradiation is the presence of organic material in the particles. Amajor new insight of the past decade is that organic aerosols(OA) are more abundant than previously thought, often exceed-ing the concentrations of sulfate aerosols.65,100,101 They haveboth natural and anthropogenic sources, the largest being thephoto-oxidation of hydrocarbons emitted by vegetation, butother major sources include the combustion of biomass, andthe production and use of fossil fuels. Many UV-absorbingorganic chromophores have been identified in collectedaerosol and rain samples, including conjugated carbonyls and
nitrates,102 nitroaromatics,103,104 and organic peroxides.105
Laboratory-generated OA, such as from smog chamber simu-lations of the oxidation of biogenic hydrocarbons, are found toabsorb below 400 nm but not necessarily at visible wave-lengths.106 The atmospheric aging of OA also appears toincrease UV absorption,107 although photo-bleaching has alsobeen reported.108 Therefore, OA have the potential to inducelarge variations in surface UV radiation, but quantification isstill very uncertain and caution should be used when estimat-ing changes in UV radiation in regions where large concen-trations of these organic particles are found.
Trends in aerosols over the past few decades have beenderived from the analysis of surface radiation network dataand satellite-based observations. Decreases in aerosols haveoccurred over the US and much of Europe109,110 and theassociated brightening has been detected at visible and UVwavelengths.111–113 However, increases in aerosols haveoccurred in South and East Asia.74,114 In some cases, the massconcentrations of aerosols have decreased but the AOD hasstill increased due to a shift in the size distribution of theaerosol particles.115 Historical (1850–2000) reconstructions ofanthropogenic and biomass burning aerosols have beenderived summarising the known historical changes.116
The future trends of aerosols are of great interest to climatestudies, and scenarios spanning a large range of uncertaintieshave been developed.117,118 Global emissions of sulfate mayalready have peaked two decades ago and may now be decreas-ing, while those of fossil fuel black and organic carbon areexpected to peak in the next few decades. The evaluation of theeffects on UV radiation is relatively straightforward for sulfateand black carbon, but is highly uncertain for organics,especially if these absorb UV radiation as discussed above.
In many cases, it is difficult to separate anthropogenicfrom natural influences, for example, changes in dust or seasalt from winds affected by climate change, or changes in bio-genic aerosols following deforestation or other major changes
Fig. 3 Summary of available measurements of aerosol single scattering albedo, ωο, at ultraviolet wavelengths from various sources: w98;89 p03;90
w03;91 g05;92 b05;93 b08;94 c09;95 i09;96 g11;97 b11;98 m12;99 k05;280 k12;281 n13.282
Fig. 2 Variation of UVery with aerosol optical depth, for different valuesof the single scattering albedo, ωο, and different receiver geometries(horizontal, spherical, and vertical cylinder).
Perspective Photochemical & Photobiological Sciences
24 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

in land use. This adds uncertainty to the future projec-tions because of the challenges in modeling the complexinteractions.
Clouds
Clouds play an important role in modifying the solar UV radi-ation that is received at the Earth’s surface, generally leadingto attenuation, but in some conditions to enhancement. Arecent example of the latter is from measurements in Spain,which have shown that clouds can enhance UVery by up to22%.119 Reductions of irradiance depend on cloud cover,depth and type (water/ice), and can be moderated by surfacereflectivity, particularly when the latter is high (e.g., snow, ice),leading to increased irradiance at the surface through multiplescattering of radiation (see the discussion in the next section).The effects of clouds are more pronounced in the visible thanin UV wavelengths. Even under skies covered completely withclouds, the UVI can still be high: maximum UVI values close to10 and average values of about 3 were measured at an urbanlocation in Brazil (19.9° S; 858 m altitude) under overcast con-ditions, predominantly of cumulus clouds.120 Typical noonUVI values at this location under cloud-free skies are about8.3. Cirrus clouds are formed of relatively large ice crystals,which efficiently scatter the solar radiation towards theground, with only small losses compared with the clear-skycase. According to model calculations based on the global esti-mates of the optical depth of cirrus clouds averaged over theperiod of 1984–2007, surface UV-B radiation has been attenu-ated on average by up to ∼2% compared to clear skies.121
In the Arctic region, clouds could be affected by the loss ofsea-ice. Based on the satellite data for the period of 2000–2010,a 1% decrease in sea-ice cover leads to a 0.36–0.47% increasein cloud cover, suggesting that a further decline in sea-icecover would result in an even cloudier Arctic.122 Due to thecomplexity of the processes involved, it is difficult to simulatethe impact on UV radiation from the combined reduction insea-ice and increase in clouds.
The spatial and temporal inhomogeneity of clouds makes itdifficult to accurately assess and quantify their effects on radi-ation. Thus, empirical parameterizations are often used todescribe their effect, such as the cloud modification factor(CMF), which is defined as the ratio between the measuredsurface irradiance to the corresponding clear-sky value. Itdescribes the average effect of clouds, implicitly taking intoaccount the optical thickness of clouds (COT). A clear exponen-tial dependence between the CMF derived from UV measure-ments and the COT from a Cimel sun-photometer was found;for COT between 10 and 50 the CMF ranged from 0.7 to0.25.123
Since our last assessment124 there has been little new infor-mation on how clouds affect the solar UV radiation received atthe Earth’s surface. Experimental evidence is constrained bythe uncertainty in the measurements and by the complexity ofthe cloud characteristics. Climate change is projected to alterthe amount of clouds over particular areas and, as the ozonelayer recovers, clouds will have an important role in controlling
the future levels of solar UV radiation that would be availablefor the ecosystems.
Surface reflectivity
The reflectivity of Earth’s surface, usually referred to as“surface albedo”, is defined as the ratio of the reflected to theincident amount of radiation. The radiation reflected upwardsfrom the surface undergoes subsequent scattering by air mole-cules and particles, resulting in enhancement of the irradianceat the surface. These effects are greater when the surface iscovered by a highly reflecting material, such as snow or ice.Most materials have low UV reflectivity, with only snow and icehaving UV reflectivity greater than 0.5, whereas many surfaceshave relatively high reflectivity in the visible. Consequently, theeffects of ice and snow are more readily apparent for radiationin the UV than in the visible region. The sea-surface alsoreflects radiation impinging on the water, but less effectivelycompared to snow and ice. The roughness of surfaces alsoaffects their reflectivity. The fraction of reflected radiation gen-erally increases with the angle of incidence of the photons;hence it depends on season and latitude. In the Arctic and Ant-arctica, this effect becomes more important due to the largerSZAs and the prevalence of diffuse radiation in these regions.Spectral measurements of UV irradiance at Ny Ålesund, Sval-bard, (79° N) revealed an enhancement in clear-sky irradianceat 320 nm of about 15% between two sites affected differentlyby the reflectivity of the snow-covered surface and the partiallyice-covered ocean. The effect was doubled under overcast con-ditions.125 The deposition of black carbon aerosols on snow orice may substantially reduce the reflectivity, resulting inweaker solar irradiance at the surface. For example, during thesnow-melt period the reflectivity at Sodankylä, Finland,decreased from 0.65 to 0.45 at 330 nm and from 0.72 to 0.53 at450 nm, partly due to the deposition of black and organiccarbon.126
In mountainous regions, the reflection of radiation mayoccur both on the surface (usually covered by snow) and fromthe top of clouds located below the altitude of the site, whilemultiple reflections may occur between snow-covered surfacesand the base of the clouds. These complex processes canresult in considerable enhancement of the incident irradiance.At Sonnblick, Austria, clouds below the observatory increasedthe average reflectivity by 0.28 ± 0.15, leading to an increase inthe irradiance of between 2% and 14% in most (∼75%) of theinvestigated cases. Compared to snow-free conditions, anenhancement of 22% in irradiance at 305 nm arose from amean reflectivity of 0.68 under clear-sky conditions. The analy-sis of total sky images revealed that an enhancement couldalso be observed when the solar disk was obstructed by cloudsor under overcast skies.127
In the context of climate change, reductions in the surfacereflectivity due to melting snow or ice would result in areduction of irradiance over land, but to an enhancement ofthe irradiance received at and under the sea-surface in regionswhere sea-ice disappears. This situation may occur, forexample, in the Arctic during the summer period in the
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 25

2030s.46 Model simulations for several scenarios suggest thatthe snow depth in April on Arctic sea-ice will decrease over the21st century. This is due mainly to the loss of sea-ice area inautumn and, to a lesser extent, in winter,128 which results in asmaller snow accumulation. When the snow depth becomesshallower, the reflectivity is reduced and, in turn, the UV radi-ation at the surface is also reduced, while more radiation istransmitted through the ice below the snow.
The sea-ice cover of the Chukchi and Beaufort Seas in theArctic is currently undergoing a fundamental shift from multi-year ice to first-year ice, which is generally thinner and morespatially heterogeneous with a more complex pattern of reflec-tion and transmission of solar radiation.54,129 When theannual Arctic sea-ice starts melting, it becomes less reflectivethan old ice, leading to a reduction in UV irradiance at thesurface. In Antarctica, the sea ice is generally expanding ratherthan shrinking. This expansion is not uniform, but variesregionally. Glaciers in western Antarctica have becomethinner130 and the loss of ice sheets is projected for the futurein this region.131,132
Reductions in surface reflectivity are expected to play animportant role for the levels of UV radiation in the future overareas that, in the past, were covered by ice or snow, such as thehigh and polar latitudes and the high mountains. Implicationscan be expected for ecosystems in these areas, either from thereduced (mainly terrestrial systems) or from the enhanced(mainly aquatic systems) exposure to UV radiation. Comparedto other regions, except for smaller, high-reflecting areas (e.g.,salt lakes affected by dust deposition), the changes in reflectiv-ity are likely to be small and thus unlikely to have a significanteffect on UV.
Solar activity
Solar activity, particularly the 11-year solar cycle, influencesUV-B radiation that penetrates to the surface of the Earth,mainly through changes induced in stratospheric ozone,rather than directly due to increased solar emission. In theupper stratosphere, solar activity affects the photochemicalproduction of ozone by UV-C (200–280 nm), while in the lowerstratosphere it affects ozone predominantly by changing theatmospheric circulation.133 This latter effect is the most impor-tant for UV radiation at the Earth’s surface because it occurs inthe layer where ozone is abundant.
Recent observations from the SIM and SOLSTICE instru-ments onboard the SORCE satellite revealed an ‘exceptionally’low minimum in the solar activity, with larger reductions inthe emitted UV-C radiation during the declining phase of the11-year solar cycle (2004–2008) than at the same phase of pre-vious solar cycles.134 These reductions were about 8 timeslarger than expected by semi-empirical models.135,136 Theinconsistencies of these observations with the perception ofvariations in solar irradiance from earlier measurements andmodels have been assessed recently,137 along with the relevantimplications for the variability of stratospheric and totalozone. The inclusion of these new observations of solar varia-bility in photochemical models and in CCMs revealed
decreases of ozone in the upper stratosphere and mesosphere(related to photochemical processes). This would allow thepenetration of more UV-B radiation to lower altitudes in thestratosphere and below. However, it has been suggested thatthe penetration of UV-C in the lower stratosphere will enhancethe ozone production there (self-healing effect) and, in turn,reduce UV-B radiation penetration to the Earth’s surface.136,138
The net effect on UV-B has not yet been quantified.Recent studies139–143 suggest that solar activity may evolve
into a declining phase in the course of the 21st century result-ing in weaker emission of solar UV radiation. CCM simulationsfor the future showed that such strong reductions in UV-C radi-ation would lead to a significant decrease in the production ofstratospheric ozone from the photolysis of oxygen. This wouldslow down the recovery of stratospheric ozone by more than 10years or even cancel it,144 leading to greater levels of UV-B radi-ation at the ground for as long as the concentrations of strato-spheric ozone remain small. These effects are most pronouncedin the region between about 40° S and 40° N, where UV-B radi-ation is already high, and would be likely reinforced by the pro-jected strengthening of the Brewer–Dobson circulation.
The detection of the effects of solar activity on the surfaceUV radiation measurements is difficult, as they are masked bystronger natural variations due to other factors. An averagedecrease of about 1.8 ± 1.0% in the ground-level irradiancefrom solar maximum to solar minimum for the UV-A and 2.4 ±1.9% for the 400–600 nm spectral band was reported by corre-lating 17 years of spectral solar irradiance measurements atthe South Pole with the 10.7 cm solar radio flux (indicative ofthe 11-year solar activity).145 As the effects appear to be toolarge to originate directly from differences in the radiationemitted by the Sun, it was suggested that these decreases arepartly due to a small variation in atmospheric attenuation withthe solar cycle, with the greatest attenuation occurring at thesolar minimum. However, there is no experimental proof forthis suggestion.
Although the direct influence of solar activity on the UVradiation at the surface is small, the indirect effects, throughchanges in the production of ozone, could be more important.If a substantial solar minimum occurs in the future, it mayinfluence global climate and the ozone layer, and could lead toincreases in UV-B radiation at the surface.
UV radiation changes and trendsderived from measurementsMeasurements of UV radiation
Ground-based. With a few exceptions,146,147 the coordinatedmeasurements of UV radiation from the ground started in thelate 1980s after the discovery of the ozone hole.148 It is there-fore not possible to directly assess the changes in UV radiationfor the entire period between the 1960s (i.e. the time beforeconcentrations of ODSs in the atmosphere became important)and the present. Several ground-based networks now providedata records in excess of 20 years with instruments deployed
Perspective Photochemical & Photobiological Sciences
26 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

in the U.S.,149,150 Canada,151,152 South America,153,154
Europe,113,155–158 New Zealand,80,159 Australia,160 the Arctic,161
and Antarctica.162 Relatively few measurements have been per-formed historically in Africa, the Middle East, and Asia, butseveral programs have recently been established in Nepal,163
Thailand,164,165 and China.166 By the mid-1990s, the technol-ogy of UV radiation measurements with spectroradiometershad already reached a level of accuracy that would allow thedetection of changes in UV of a few percent at stations withappropriate quality control protocols.167 A European projecthas been completed that aimed to provide traceable solar UVirradiance measurements with an uncertainty of less than 2%(http://projects.pmodwrc.ch/env03/). However, the methodsdeveloped in this project have not yet been implemented inoperational UV monitoring.
Recently, array spectrometers have been adapted for spec-tral irradiance measurements in the UV. These instrumentshave been used to quantify the effects of solar UV-B radiationin terrestrial ecosystems (see Bornman et al.31). However,being single monochromators, they are susceptible to straylight problems in the UV-B.168,169 Efforts have been made todetermine the uncertainty of this type of instrument.170,171
Because of their large uncertainties compared to scanningspectroradiometers, these instruments are not yet widely usedin UV monitoring programs.
Spectroradiometric measurements of UV irradiance on aflat, horizontal surface are the most common and are generallyconsidered the most accurate method to quantify UV radiation.However, this geometry is not the most appropriate to gaugethe exposure levels of humans and most animals because theanatomical distribution of UV exposure is highly hetero-geneous, poorly correlated to surface irradiance, and, in thecase of humans, influenced by factors such as posture, orien-tation to the sun (see Fig. 2), skin complexion, clothing, andother sun-related protective behavior.172 In principle, the mostaccurate method to quantify human exposure is to measurethe incident solar spectral radiance distribution (which mayoriginate from the Sun, the sky, or radiation scattered upwardfrom the ground) and integrate this distribution over allexposed parts of the human body.173 The development of asystem capable of measuring sky radiance at different zenithand azimuth angles within seconds rather than minutesenables new possibilities to study the spectral influence of fastchanging cloud conditions without the disadvantages of scan-ning instruments.174 This method is still under developmentand has not yet been reported for exposure studies. In analternative approach,172 measured global-horizontal, direct-normal, diffuse-horizontal and upwelling irradiance were com-bined with a three-dimensional numerical model of thehuman body to calculate exposure. An important conclusionof this study, as well as of Seckmeyer et al.,173 was that the con-tribution of diffuse UV radiation to total sun exposure is largerthan commonly expected, explaining almost 80% of the cumu-lative annual exposure dose.
Satellite based. The estimation of UV irradiance at theground has been repeatedly undertaken from various satellite-
borne sensors.175–177 As these products are mainly derived bymodels fed with measured or estimated radiation-related para-meters, they are associated with relatively large uncertainties,which vary according to location, season, atmospheric situ-ation and the characteristics of the satellite instrument.Known sources of errors that affect the accuracy of the derivedsurface UV irradiance include: absorption and scattering bytropospheric aerosols, inhomogeneities of clouds, assump-tions or estimations of the surface reflectivity, variability ofaltitude within the sub-satellite pixel, various modeling para-meterizations, and the inability of current satellites to dis-tinguish between clouds and surfaces covered with snow and/or ice. It is important to note that ground-based observationsare point measurements while satellite observations are repre-sentative for a pixel of several square kilometers. This differ-ence must be taken into account when comparing satellite-and ground-based measurements; particularly when theground-based instrument is located in a non-homogenousarea (e.g., mountains).
In the previous assessment124 it was noted that, althoughsatellites have the advantage of near global coverage, satellite-borne instruments cannot adequately probe the boundarylayer (approximately the lowest 1–2 km) of the atmosphere.Therefore, they tend to overestimate UV radiation whenabsorbing aerosols are present,176 particularly under clearskies. In a recent study in Santiago, Chile, a city with heavy airpollution and complex surrounding topography, this effect wasquantified, reporting an average overestimation of UVI byabout 46% from the Total Ozone Mapping Spectrometer(TOMS) for the period 1995–2007, and by about 47% from theOzone Monitoring Instrument (OMI) for the period2005–2007.153 These results were qualitatively confirmed bytwo other studies.177,178 Similar results were found for fourlocations in Thailand,164 with average biases between 40% and60%. Smaller biases were reported for locations with smalleraerosol concentrations. In France, UVI derived from OMI andthe Global Ozone Monitoring Experiment, (GOME-2) wasfound to be larger by about 6% during 2008–2009,179 while inSouthern Spain, for the period of 2004–2008, the average biaswas about 12%, rising to 19% for days with large aerosoloptical depth (>0.25 at 440 nm).180 The UVI derived fromTOMS has been also compared with ground-based data at 27stations of the USDA network, showing results consistent withthe above studies, with an average positive bias in the satelliteestimates of the order of 15% over all sites. Under clear skies,the biases can be either negative (up to 3.4%) or positive (upto ∼24%), depending on the amounts of tropospheric aerosolsand UV-absorbing air pollutants.181
A comparison of the satellite retrieval of UV irradiance fromOMI with UV spectra measured at six Austrian sites with alti-tudes ranging between ∼600 and ∼3100 m concluded that thesatellite estimates were significantly smaller (average ratio0.89, range 0.6–1.35) for most stations due to erroneous correc-tion for the effects of clouds.182 In contrast, under cloud-freeconditions, the satellite data were closer to the ground-basedmeasurements, but they cannot distinguish between mountain
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 27

and valley sites due to the large variability in altitude within ashort horizontal distance, smaller than the size of a pixel. Themain deficiencies in the satellite retrieval algorithm arosefrom the incorrect determination of the effective surface alti-tude and albedo due to the complex topography.
Such deficiencies have been taken into consideration in theSemi-Analytical Cloud Retrieval Algorithm (SACURA), in whichthe background spectral albedo is properly specified and cloudparameters are derived from the infrared sensors of the satel-lite.183 Comparisons of estimates based on satellite data andradiative transfer modelling with observations at two locationsin Belgium revealed a good agreement, with correlation coeffi-cients of 0.88 and 0.91 for UV-B and UV-A irradiance,respectively.
Variations of UV radiation in time and space
UV radiation at the Earth’s surface varies with the season, timeof day, latitude, and altitude. It is also affected by the absorp-tion and scattering processes from atmospheric constituents,as discussed above.
Latitudinal variations in annual doses of UV-B and UV-Ahave been assessed with high-resolution measurements fromground-based spectroradiometers that comply with the qualitystandards of the Network for the Detection of AtmosphericComposition Change (NDACC)184 (Fig. 4). For all sites, theannual averages were derived from at least 10 years overperiods where trends in irradiance were small. As expected,doses of UV-B and UV-A are generally largest close to theequator and smallest at high latitudes. Doses at high-altitudesites (South Pole, Mauna Loa, Boulder and Summit) are largerthan for sites located at similar latitudes but at sea level.5 Thisis most obvious when comparing data from Barrow (71.3° N;8 m altitude) and Summit (72.6° N; 3202 m altitude), wherethe annual doses of UV-B and UV-A at the latter site are about58% and 83% larger due to higher elevation and differentsurface reflectivity, respectively. Surface reflectivity in the order
of 0.98 also contributes to the relatively large doses at theSouth Pole and Summit, while attenuation of UV radiation byaerosols is responsible for the relatively low dose at Tokyo.Latitudinal gradients are stronger in the UV-B than the UV-Aregion, partly because photons travel a longer path throughthe atmosphere for the lower solar elevations prevailing athigher latitudes, allowing a greater absorption of UV-B radi-ation by ozone. Another factor contributing to the differencesin gradients of UV-B and UV-A is the relatively small ozonecolumn in the tropics. As a consequence, the ratio of theannual dose of UV-B/UV-A is roughly 0.03 close to the equator,0.02 at mid-latitude sites and less than 0.02 at high latitudes(Fig. 4, lower panel). It is interesting to note that the UV-B/UV-A ratios are not very different in polluted locations, such asTokyo, compared with clean-air sites, suggesting that theoptical depth of aerosols in the UV-B is not very different fromthat in the UV-A region, and/or that the wavelength depen-dence of the single scattering albedo throughout the UVregion is small (see Fig. 3).
It has long been known that mid-latitude UV levels in thesummer are larger in the southern than in the northern hemi-sphere.185 Factors contributing to this disparity include thesmaller Sun-Earth distance during the southern-hemispheresummer, plus smaller ozone columns and less attenuation byaerosols in southern latitudes. However, for annual doses, thehemispherical differences are relatively small (Fig. 4, upperpanel).
Satellite observations indicate that the greatest UV levels atthe surface of the Earth occur in the Altiplano region of Peru,Bolivia, Chile, and Argentina, where the UVI in summer mayexceed 20.186 The large UV levels in this region can be attribu-ted to small SZA, overhead Sun, small total ozone, highelevation (hence less aerosol and unrestricted horizon), andminimum Earth-Sun separation in the austral summer. Thesefindings have recently been confirmed by ground-based spec-troradiometric measurements at the Chajnantor Plateau (23° S,67° W; 5100 m altitude) of the Atacama Desert in NorthernChile.187 The measured UVI peaked at a value of 20 underbroken cloud conditions and was 18 for clear skies. Very highUVI values were reported for locations in Tibet in2008–2010.188 The measured monthly mean UVI in July was14.5 in Tingri (28.7° N; 4335 m altitude) and 12.9 in Lhasa(29.7° N; 3683 m altitude), with a peak value of 20.6 in Tingri.Extremely high UV levels (UVI of up to 19 for clear sky and upto 22 under broken cloud conditions) were also measured atsea-level in the tropical Pacific (3.6° S, 85° W), when totalozone was 234 DU.189 These values are more than doublethose that are common at northern mid-latitudes insummer.190 More recently, UVI values higher than 40 havebeen reported at Licancabur, Bolivia (5916 m altitude).191
However, these data are inconsistent with the satellite-derivedozone columns. Further work is needed to verify the result.
During the Arctic spring of 2011, when total ozone wasextremely small, greatly increased levels of UV radiation wererecorded at thirteen Arctic and sub-Arctic ground stations.Measurements of the noontime UVI during the low-ozone
Fig. 4 Latitudinal variation of UV-A (315–400 nm) and UV-B(280–315 nm) annual dose (top) and the ratio of UV-B/UV-A dose(bottom).
Perspective Photochemical & Photobiological Sciences
28 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

episode exceeded the climatological mean by up to 77% atlocations in Alaska, Canada, and Greenland, and by up to161% in Scandinavia.161 The cumulative UV dose integratedover the duration of the low-ozone period increased by 40–50%at several sites in the Arctic and Scandinavia, and exceeded theclimatological mean by more than 3σ at seven sites and bymore than 4σ at two sites. Despite these large relative enhance-ments, absolute UV anomalies remained small (less than 0.5UVI units at the western-hemisphere sites) or moderate (1.0and 2.2 UVI units at the Scandinavian sites) because the low-ozone episode occurred at a time when the Sun was still low inthe sky. In Alaska, Canada, and Greenland, the UV increasescan be explained by low ozone, but at the Scandinavian sitesthey were caused by a combination of low ozone and theabsence of clouds. Despite the low absolute levels of UV radi-ation at northern polar latitudes, biological systems duringthis Arctic event may have been exposed to greater UV thanusually experienced.
The small ozone values observed in 2011 in the Arctic pro-pagated to midlatitudes and, as a consequence, ozonecolumns over Western Europe were 15–25% below the long-term mean between late March and late April 2011.26 Modelassessments suggest that noontime erythemal doses on clear-sky days were larger than usual by about 25% during theaffected period. However, such increases have not been con-firmed by measurement, so far.
Furthermore, it was found from ground-based and satelliteobservations that the interannual variability in springtimeozone in the Arctic was correlated with ozone in the summerand explained 20–40% of the summer UV variability at somelocations.192 Particularly for spring 2011, it was estimated thatthe massive ozone depletion in the Arctic increased theMarch–August cumulative erythemal clear-sky UV dose in thenorthern hemisphere outside the tropics by 3–4% compared tothe climatological mean, with about 75% of the increase accu-mulated after the breakup of the polar vortex.
Unusually large UV levels were observed between 11 and30 November 2009 over the southern tip of South America(∼55° S) when the center of the Antarctic vortex became stag-nant just south of South America for a three-week period,leading to ozone columns continuously more than 2σ belowaverage. Ground-based measurements for three stationslocated in this region showed UVI values of 10 to 14, which,for clear-skies, typically only occur at latitudes lower than 40°in the northern hemisphere.11 The analysis of 30 years of satel-lite observations revealed that this event was unique for thelatitude belt of 52° to 56° S.
In August 2011, southern Australia was affected by ozone-poorair originating from tropical latitudes, resulting in measuredUVery levels of up to 40% greater than normal.160 This is anexample where meteorological factors have produced an anoma-lous reduction of ozone on an almost continental scale and for alonger duration than previously observed.
The combined effect of all the factors discussed previouslymay result in very high levels of UV radiation at mid and lowlatitudes, primarily at high-altitude locations. These high-UV
episodes will continue to occur in the future during low-ozoneperiods, irrespective of the recovery of ozone.
Observed long-term changes in UV radiation
In view of the expected rebound of stratospheric ozonedepletion and recovery to levels before the 1980s, an importantquestion is whether this change is reflected in the trends ofUV radiation measurements. At most locations, any trends arecurrently still below the detection threshold imposed by instru-ment uncertainties and variabilities due to factors other thanozone, such as changes in aerosols, clouds and surface reflec-tivity. For example, over the period of the peak ozone depletionbetween the 1980s and 1990s, ozone and cloud effects contrib-uted equally to the UVI increases over populated areas of thenorthern mid-latitudes.193
Long-term changes in UV radiation can be estimated bothfrom space and the ground. Satellite observations have largeuncertainties as discussed above. Changes in UV radiationat different spectral bands over the period of 1979 to2008 have been derived from a series of polar orbiting satelliteinstruments for the latitude range of 55° S to 55° N,194 andthe results were summarized in the last assessment.124,195
A similar study for the time period of 1997–2010 based on themeasurements of three satellites (TOMS/Nimbus 7, TOMS/Earth Probe and OMI)196 has qualitatively confirmed theearlier work by Herman.194 Over this later time period, thederived linear trends in erythemal irradiance (same for UVI)ranged between 0 and +5% per decade between 50° S and 50°N. These positive trends are significant at the 95% level withthe exception of the equatorial zone and winter months ofboth hemispheres. The largest increases were observed dur-ing the spring and summer at mid-latitudes of the SouthernHemisphere, where the largest decrease of ozone was observed(Fig. 5). However, most of the UVery changes due to ozone
Fig. 5 Monthly linear trends (%/decade) of erythemal irradiance derivedfrom TOMS and OMI data from 1979 to 2010 according to ref. 196. Alltrends are positive. The data are zonally averaged for every 5° of latitudefrom 50° S to 50° N and mostly represent changes that occurred in thefirst half of this period, when ozone depletion was progressing. Adaptedfrom Ialongo et al., 2011.196
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 29

occurred during the 1980s and the early 1990s; therefore, thecalculated linear trends do not necessarily reflect a tendencyin ozone that would continue into the future. At high-latitudes,the satellite-based estimates of surface UV radiation can be toolow by up to 50%, when high albedo from snow and ice coveris misinterpreted as clouds.197 These systematic errors canalso affect UV trend assessments. For example, it is difficult toquantify the changes in UV radiation from space measure-ments over high-latitude locations that are affected by sea-icevariability.194 For these reasons the trends discussed abovewere derived only for latitudes lower than 55°.
The combined effect of the surface reflectivity (RS), clouds,water haze, and aerosols on UV-A radiation, which is notaffected by ozone, can be inferred by the, so-called, “Lamber-tian equivalent reflectivity” (LER). This represents the equi-valent scene reflectivity, as seen from space, after the removalof Rayleigh scattering effects.198 The atmospheric transparencyT is approximately T = (100 − LER)/(100 − RS), where LER andRS are expressed in percent. Because the reflectivity of mostsurfaces is small (typically 2–4% over land), a decrease of LERwill lead to an increase in T by approximately the sameamount. The LER at 340 nm during the past 33 years(1979–2011) has recently been analyzed globally199 andchanges in the surface irradiance at 340 nm, but withoutaccounting for the effects of local air pollution sources, can beinferred from this study. Between 1979 and 2011, most of thedecreases in LER (resulting in an increase of surface radiation)occurred over land, with the largest wide-spread decreasestaking place over the US (0.97% per decade), Brazil (0.9% perdecade), and Europe (1.4–1.9% per decade). Over India,southern China, and Indochina, LER has increased by 1–1.5%per decade (Fig. 6). A trend that is twice as large was observed
on the west coast of South America, but there was almost nochange over most of Australia. These trends can be translatedinto downward trends in UV-A radiation, caused only bychanges in cloud cover and, partly, in aerosols. In the oceanregion near the Antarctic Peninsula (160° W–50° W), LER hasdecreased strongly (>2% per decade), probably due to changesin clouds and sea-ice. Neither of these studies took intoaccount the effects of absorbing aerosols in the estimates ofirradiance. Therefore, these trend estimates may not accuratelyreflect the changes for regions where the concentrations ofabsorbing aerosols and air pollutants have changed over time,such as in urban areas.
The variability of the solar UV irradiance at 305 and 325 nmbetween 1990 and 2011 has been assessed from ground-basedmeasurements at twelve sites in Canada, Europe, and Japan(latitudes between 25° N to 60° N).200 For these sites at least,this period can be divided into three sub-periods that arecharacterised by different physical processes. UV radiationdecreased during the first period (1991–1994), greatly affectedby stratospheric aerosols from the Pinatubo volcanic eruptionin 1991. The volcanic stratospheric aerosol layer had twoeffects: it induced the destruction of ozone through hetero-geneous chemical reactions201 and reduced the path-length ofUV-B solar radiation through the ozone layer for small solarelevations due to increased scattering by the aerosols.202 Bothmechanisms led to an increase in UV-B at the surface after theeruption in 1991, which became smaller in the next years asthe aerosol effect decayed. The second period (1995–2006) wascharacterized by a 1.4% per decade increase in total ozone,coinciding with a significant decline of the aerosol opticaldepth over the regions of study. This “brightening” effect(increase of the atmospheric transparency T ) more than offsetthe effect of the increase in ozone, resulting in positive UVtrends of 9.4% per decade at 305 nm and 8.8% per decade at325 nm. The third period (2007–2011) showed statistically sig-nificant evidence of a slowdown or turning point in theupward trends in UV-B radiation over Canada, Europe, andJapan. These results are consistent with the decrease in LERseen from satellites (Fig. 6) and observations of surface short-wave (300–3000 nm) solar irradiance, which indicate that thebrightening effect, which started in the late 1980s and ismostly attributed to changes in cloud cover and aerosols, hasslowed down during the last few years or is no longerprogressing.45,203–205 However, since some of the sites dis-cussed above200 are located in urban areas that are affected byair pollution, the results cannot be simply applied to a globalscale or to sites located at higher altitudes, where aerosols mayevolve differently or remain constant.206
Several recent studies that have reported positive trends inUV radiation at European sites caused by a reduction in cloudcover are consistent with satellite observations (Fig. 6). A stat-istically significant increasing trend of 6.6% per decade indaily maximum UVI from 1993 to 2008 was found for Reading,United Kingdom, presumably caused by a reduction in middayclouds, since ozone remained constant.207 An increase in theerythemal dose of 5.5 ± 1.0% (1σ) per decade was found for
Fig. 6 Trends in LER (combined reflectivity from clouds, aerosols, andthe surface) as seen from space. Trends in LER smaller than 0.3% perdecade are not statistically significant from zero. Negative trends in LERlead to positive trends in UV-A by approximately the same amount.Trends in LER outside the Polar Regions are caused by clouds and notaerosols. Adapted from Herman et al., 2013.199
Perspective Photochemical & Photobiological Sciences
30 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

April–October in the period of 1976–2008 from ground-basedUV measurements at Belsk, Poland (51° N).155 At bothlocations, the total ozone levels had stopped declining in themid-1990s, and the observed increases in UV radiation wereattributed to decreasing attenuation by clouds. In suchlocations, which are dominated by cloudy weather, any effectsfrom changing aerosols cannot be detected as the opticaldepth of clouds is much greater than that of aerosols. Simi-larly, the trend of increasing spectral UV irradiance at HoherSonnblick (Austrian Alps, 3106 m altitude) was also attributedto decreasing attenuation by clouds (5.8% per decade duringsummer), which was confirmed by the synoptic observationsof clouds and measurements of the duration of sunshine.Spectral irradiance at 315 nm was found to increase between1997 and 2011 from 9.3% per decade at SZA = 45° (spring–summer) to 14.2% per decade at SZA = 65° (whole year).208
Because ozone has been increasing by 1.9 ± 1.3% per decadeover this period, the increase of irradiance at 305 nm wassmaller (between 5.1% and 7.9%) and not statisticallysignificant.
Trends in the monthly average UVI at Barrow, Alaska (71.3° N),calculated from spectral UV measurements, between 1991and 2011 were not statistically significant, except for October(−14% per decade).209 This large trend was attributed todecreasing surface reflectivity as the onset of snow cover inautumn has been delayed at this site with a statistically signifi-cant trend of 13.6 days per decade. This study emphasizes theimportance of climate factors on long-term changes in UVradiation.
After removing the annual variability, the UV irradiance at305 nm was found to decrease with an average rate of 3.9% perdecade for 1991–2011 over four northern hemisphere high-lati-tude stations (Barrow, Sodankylä, Jokioinen, and Churchill),whereas no significant change was found for irradiance at325 nm, which is only slightly affected by ozone.210 For thethree southern stations examined (Ushuaia, Palmer, andSyowa) no significant changes for either wavelength werefound.
The above studies indicate that factors other than ozonehave dominated the changes in UV radiation during the lasttwo decades at many sites. They also indicate that UV-B irradi-ance has stopped increasing at mid-latitude locations inresponse to the slowdown of the ozone decline.
Simulations of historic changes in UV radiation
As mentioned in our previous assessment,124,195 changes inUV radiation over timescales of centuries to a few decades canbe estimated using various proxies or simulated variations offactors that may directly or indirectly affect the solar UV radi-ation at the Earth’s surface (see also Bornman et al.31).Although such estimates have large uncertainties, they areuseful in assessing qualitatively the causes of the variations inUV radiation that may have occurred in the past.
A modelling study211 suggests that levels of UV-B radiationin the year 2000 were 2–8% lower than in 1850 over the north-ern hemisphere and the tropics, and higher by 4% and 30%,
respectively, over the mid- and high-latitudes of the southernhemisphere. At most locations outside the tropics, the UV-Bchanges were caused by changes in tropospheric ozone, exceptfor the northern hemisphere mid-latitudes where changes intropospheric ozone and aerosols are equally important. Theseincreases in tropospheric ozone in the northern midlatitudescounteracted the increase of UV-B radiation due to strato-spheric ozone depletion in the 1900s.
Artificial neural networks trained with measured erythemalirradiance, duration of sunshine, and a combination ofmeasured and modeled total column ozone, were used toreconstruct the daily erythemal dose for Potsdam, Germanyfrom 1901 to 1999 (Fig. 7).212 A positive, statistically significanttrend was found for the first half of the 20th century, in linewith the observed negative trend in cloud cover. Since 1950,the trend in annual UVery was negative until the mid-1980s,when it turned positive again. However, for both these latterperiods, the trends were not statistically significant. These esti-mates do not include potential effects from aerosols, whichdecreased substantially in the second part of the 20th century(as discussed above), and would likely result in further increas-ing of UVery in the 1980s and 1990s. Furthermore, the uncer-tainty in the ozone data before 1949, and implicitly in UVery, ishigher because it has been obtained by a CCM.
In a study for Australia, the UVI under cloud-free conditionswas calculated with a radiative transfer model over a 50-yearperiod (1959–2009) based on measured meteorological para-meters.213 After the 1990s, an overall increase in annual meanUVI of 2–6% relative to the 1970–1980 levels was reported forall latitudes in that country.
UV radiation under the water
The amount of UV radiation under the water surface of theocean, lakes, and rivers depend on the available radiation fieldreaching the water surface and additionally on the transpar-ency of the water body. The first is determined by the absorp-tion and scattering processes of solar radiation in theatmosphere, as discussed above, and, at high latitudes, on theamount of ice over the water and the amount of snow over theice. The transparency of water depends on the concentrations
Fig. 7 Reconstructed annual total of UVery time series (blue line, left-hand scale, lower graph, “ER”), mean annual total ozone in DU (gray line,right-hand scale) and mean annual sunshine duration (SD) in minutesper day (red line, left-hand scale, upper graph) for Potsdam, Germany,1901 until 1999 (thin lines, annual values, thick lines 7-year runningaverages). Adapted from Junk et al., 2012.212
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 31

of dissolved and particulate material in the water, with chro-mophoric dissolved organic matter (CDOM) being the domi-nant attenuator of solar UV radiation.214,215,32 The attenuationof solar UV radiation penetrating into the water column (quan-tified with the diffuse attenuation coefficient Kd, m
−1) can bemeasured directly by radiometers or spectroradiometers, butsuch monitoring programs are sparse.215,216 Recently,measurements of the spectral shape of the surface reflectancefrom the MODIS satellite have been used successfully to deriveglobally the diffuse attenuation of coastal waters.216
In some areas, the penetration depth of solar radiation intothe water is large, such as in the South Pacific Gyre, where theirradiance at 305, 325, 340 and 380 nm was reduced to 10% ofthe initial values at 28, 42, 59 and 110 m of depth, respect-ively.217 These constitute the greatest depths of penetrationever reported for oceanic waters and are comparable withthose measured in the clearest fresh waters.218 In contrast, alarge attenuation of the solar irradiance was measured in 2004in the Mackenzie Delta Lakes, Canada, with different gradientsin renewal rate of water, concentration of dissolved organiccarbon (DOC), and composition of dissolved organic matter(DOM). However, because these lakes are shallow, UV-B andUV-A radiation is still able to penetrate the top 19% and 31%of water columns, respectively.219 Thus, the climate changeeffects on the composition of DOM may significantly alter theUV radiation environment in such circumpolar delta lakes.The influence of inputs of DOM from rivers into the ArcticOcean can now be inferred from the analysis of satellite data(see Erickson et al.32).
In Polar Regions, sea-ice prevents a large fraction of UVradiation from reaching the (liquid) ocean surface, while a fewcentimeters of snow over the ice almost completely blocks thetransmission of UV-B radiation.49,220 Solar radiation under“first-year” ice in the upper ocean is spatially heterogeneousand depends on wavelength, thickness of ice, and the area andgeometric distribution of melt ponds and bare ice surfaces.129
Although there is an exponential decay in transmission or radi-ation through the ice sheet, it was reported that the trans-mission of radiation in the water under the ice can increasewith depth when bare and melt-pond sea-ice surfaces areinterspersed close to the observation site.129 Projectionsbased on earth-system models and radiative transfer calcu-lations suggest that, compared to the 1950s, up to 10 timesmore UV-B radiation will enter large parts of the Arctic Oceanby 2100, mainly because of the partial disappearance ofsea-ice.221
The future evolution of sea-ice and its snow cover is linkedto changes in climate and will likely lead to increases in UV-Bradiation reaching the ocean surface beneath the ice. Thecomplex radiation field beneath the first-year sea-ice duringthe melt-season has significant implications for biological pro-duction, biogeochemical processes, and the heat balance ofsea-ice and under-ice ocean waters. The effects of UV pene-tration into the water column, as well as the modification ofthe ratio UV-B/UV-A by CDOM in the water column, are dis-cussed further in Erickson et al.32
Projections of UV radiation: causesand health effectsProjected changes back to the 1960s and out to the 2090srelative to the present
Surface UV radiation in the future will be influenced by:increases in stratospheric ozone due to reduction in ODSs;changes in ozone and cloud cover induced by increasing con-centrations of GHGs; changes in tropospheric UV-absorbingaerosols; and decreases in surface reflectivity at high latitudesand high altitudes. Simulations of these UV radiation levelsare usually derived from radiative transfer model calculationsthat use input parameters estimated by climate models.
In our previous assessment,124,195 we reported estimatesfrom model projections222 suggesting that, by about 2050, theUVery would decrease relative to that in 1980 by 2–10% at mid-latitudes, and by up to 20% at northern and 50% at southernhigh latitudes, mainly due to the recovery of stratosphericozone and to changes in cloud cover. In the tropics, UVery wasprojected to be higher by less than 2%. We also comparedmodel projections between 2100 and 1960 to estimate theeffects of climate change on surface UV, because the ozonedepletion started after 1960 and ozone recovery would havebeen completed by 2100. By the end of the 21st century UVerywas projected to: (a) remain below 1960 levels due to changesin clouds and GHG-induced transport of ozone at mid-lati-tudes, (b) decrease at high latitudes (particularly in the Arctic)by 5–10% due to changes in clouds, and (c) increase in thetropics by 3–8% due to decreases in clouds and ozone,induced by GHGs.
However, these projections did not consider changes inaerosols and surface reflectivity. Similar results were reportedby another simulation223 that accounted also for the effects ofchanging albedo and cloud cover. By assuming typical aerosoloptical depth and single scattering albedo values over Europe,in addition to projections of ozone, small reductions in erythe-mal and the vitamin D-effective daily doses were predicted for2006–2100,224 and were attributed to the recovery of strato-spheric ozone and partially to a reduction in the optical depthof aerosols. A recent modelling study focussing on the ArcticOcean221 projected reductions in UV-B irradiance by the end ofthe 21st century relative to the levels in the 1950s over a largefraction of the area. Under clear skies, UV-A irradiance is pro-jected to decrease on average by 4–7% (depending on scenarioand season), entirely driven by decreases in surface reflectivity,while UV-B is projected to decrease on average by 10–18%,mainly due to the projected ozone recovery. Under all skies,these effects are modulated by clouds, leading to changes inthe monthly mean noontime UVI from +15% to −38%,depending on the location and season. Increases in irradiancewere found only during August for the latitude band 55–65° N,caused by the projected decrease in cloud cover.
In this assessment, we provide updated estimates of theprojections of the previous assessment,124 taking into con-sideration effects from most factors affecting the UV radiationat the Earth’s surface. This analysis is based on recent projections
Perspective Photochemical & Photobiological Sciences
32 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

of cloud cover, ozone, surface reflectivity, and aerosols for theperiod of 1955–2100 by different Earth-System models thatwere included in the fifth phase of the Climate Model Inter-comparison Project (CMIP-5),225 and for the RepresentativeConcentration Pathways (RCP) emissions scenario 4.5. For theozone projections, the ensemble mean of the CESM1(WACCM)model,226,227 which includes interactive chemistry, was used.These projections have also been used in the fifth IPCC Assess-
ment Report.45 Changes in annually averaged noon UVI due tochanges in these factors are shown separately in Fig. 8between the past (1955–1965 mean) and the present(2010–2020 mean) and between the present and the future(2085–2095 mean).
Separating the effects of changes in surface reflectivity,aerosols and clouds on UV radiation is potentially challengingbecause of the interactive influence of these factors on irradi-
Fig. 8 Simulated annually averaged percentage changes in noontime UVI (or erythemally-weighted UV irradiance) relative to the “present” (i.e.2010–2020). The left column shows simulated changes since 1955–1965. The right column shows the simulated changes expected from thepresent to the period 2085–2095. Effects of aerosols, surface reflectivity, cloud cover and total ozone on UVI are shown in each row, with ourassessment of the confidence in UVI projections. Note the two different color-scales.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 33

ance. Despite these inter-connected effects, the largestchanges in reflectivity are projected for high and polar lati-tudes due to the melting of ice or snow, while the largestchanges in aerosols are projected for mid-latitudes and thetropics, particularly over regions with strong anthropogenicactivities.
Effects of changes in aerosols. The future evolution of aero-sols and their radiative effects depend on emissions scenariosthat may differ from the actual development, both in terms ofamount and composition of aerosols. Current trends in airpollution (hence in aerosols) show large regional differ-ences;228 at some regions decreasing (Europe and NorthAmerica) and at others increasing (Asia). For all emissionsscenarios associated with the RCP, aerosols are expected todecrease significantly in the second half of the 21st centuryglobally,229 and particularly over Asia as a result of measuresfor improvement of air quality,117,118 even though the air pol-lution there is presently increasing.228
However, even if these scenarios were realistic, there wouldstill be large uncertainties in the simulations of UV radiationdue to poor knowledge of the spectral absorption efficiency ofaerosols (i.e. of SSA) and its wavelength dependence. Mostclimate models use input parameters that are appropriate forless absorbing aerosols, which result in an underestimation oftheir effect on UV radiation. The UVI simulations shown hereare based on projected aerosol optical depth values from theearth-system models. Because of the lack of specific SSA pre-dictions, climatological values for SSA75 were used, assumingno change with time, which may also be unrealistic. Finally,over areas dominated by clouds, the projected effects on UVIfrom changes in aerosols are more uncertain, particularly forlarge SZAs, for highly absorbing aerosols, and for highly reflec-tive surfaces.
High levels of air pollution in the 1950s and 1960s oversome urban areas in Eastern Europe should have resulted inless UV radiation at that time; the simulations show that esti-mated improvement of air quality since then110 yieldedincreases in the UVI of up to 40% in the 2010s. Unfortunately,no high-quality direct measurements of UV radiation exist forthat period, and neither have the reconstructed data series(see previous discussion) been forced with aerosol data toshow this effect. However, measurements in Moscow revealedan ∼4% increase in UV-A radiation from 1981 to 2003 due tothe reduction of aerosols.5 In contrast to central Europe, UVradiation may have decreased in eastern Asia (by ∼25%), dueto increases in air pollution.230 The decreases in UV (by ∼15%)over north-west Africa are not statistically significant and areprobably caused by the large differences among the model pro-jections. By the end of the 21st century, the improvement ofair-quality over most of the populated areas of the northernhemisphere may result in small increases in UVI compared tothe 2010s by 10–20%, except over China where much largerincreases are projected. The projected aerosol effect in thesouthern hemisphere is generally very small, because sourcesof aerosols are weaker compared to the northern hemisphere,and aerosols originate mostly from the ocean, while the
fraction of land with important anthropogenic activities is verysmall. Consequently, the assumed changes in aerosol amountwith time are generally very small there.110
In our assessment, estimates of the characteristics of aero-sols in the past and understanding the effects of changes ofaerosols in the future are highly uncertain at present. Althoughwe have tools to carry out the UV calculations, knowledge ofthe input parameters to the RT models, and a complete under-standing of the interactions between the various effects is stilllacking. Therefore, the simulated changes in UV radiationshown in Fig. 8 are associated with significant uncertainties,and particularly for the potential aerosol effects, are onlyillustrative. Despite the uncertainties, it is likely that, outsidethe polar regions, changes in aerosols and their properties inthe future will be more important for the levels of UV at thesurface than those from changes in ozone.
Effects of changes in surface reflectivity. As discussed pre-viously, surface reflectivity is projected to decrease between the1960s and the end of the 21st century over areas that werecovered by sea-ice and snow earlier in this period, whereas inother areas the expected effects, mainly from changes in land-use, would be much smaller. The projected effects on UV radi-ation are therefore significant only over high and polarlatitudes.
Over the Arctic, large reductions in reflectivity due to sea-ice melting have already occurred; hence the simulated UVI is∼5% lower than in the 1960s, over and close to areas coveredby sea-ice. These effects are most pronounced in the summerand autumn, when the sea-ice disappears over large areas.This phenomenon is projected to continue through the end ofthe 21st century,54 resulting in decreases in UVI with respect tothe present by up to ∼10%. In Antarctica, ice cover has gener-ally increased slightly (as discussed above), while a small frac-tion of sea-ice has been lost in localized regions (mainly overthe Weddell Sea), leading to small decreases in annual averageUVI of less than ∼2%. Small decreases are projected also forthe future since Antarctica will be still covered by snow and iceby the end of the 21st century. However, other factors (e.g.,ozone, clouds, and aerosols) that might be different from thepresent would likely modulate the reflectivity effect.
Effects of changes in clouds. The effects of clouds onclimate are significant and complex; and their representationin climate models continues to be a challenge. Many cloudprocesses, including aerosol-cloud processes, occur at scalessmaller than those resolved in large-scale climate models.Therefore, general circulation models typically use parameteri-zations to represent a range of cloud properties. A recentassessment reports considerable improvements in the abilityof models to account for effects from clouds.231
For the UV projections presented here, the effect of cloudson UVI was estimated through the CMF calculated from projec-tions of all-sky and clear-sky total solar radiation by the Earthsystem models. Extrapolation of the CMF from visible to ultra-violet wavelengths is based on empirical relationships.Changes in the CMF between the two periods shown aredirectly translated into changes in UVI due to clouds. The
Perspective Photochemical & Photobiological Sciences
34 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

modification of solar radiation by clouds depends on aerosolsin the underlying layers and on surface reflectivity, as bothlead to increasing multiple scattering of radiation. Thesefactors are implicitly taken into account in the total radiationprojections, but their effect on the derived CMF is small.
The projected changes in noontime UVI due to clouds aremostly negative. Cloud cover is projected to increase over theArctic Ocean due to increased evaporation as sea-ice declines.Therefore, the highest decreases in UVI are simulated for lati-tudes north of 60° N, and are up to 4% from the 1960s to the2010s, and about double this between the 2010s and the2090s. Pole-ward of ∼60° S, reductions of up to 3% in the UVIhave been projected for both periods, mainly over the ocean.At all other latitudes the projected changes are very small,ranging between −2% and +2%.
Effects of changes in ozone. Depletion of ozone led toincreases in UV-B radiation during the 1980s and 1990s, andthe recovery of ozone will likely lead to reductions of UV-B rela-tive to present levels. According to state-of-the-art simulationsby CCMs, it is likely that, by the early-2030s, total ozonecolumns at mid-latitudes will exceed 1980 values.2 The pro-jected increases in total ozone are due to declining concen-trations of ODSs and increases in the concentration ofgreenhouse and other source gases. Declining ODSs, strato-spheric cooling, the possible strengthening of the Brewer–Dobson circulation and other factors are likely to result in a“super-recovery” of mid-latitude ozone columns, after 2040 to2060, i.e. to levels greater than observed in the 1960s, leadingto smaller levels of UV-B.
The future levels of ozone will greatly depend on futureemissions of GHGs into the atmosphere, but also on influ-ences from possible volcanic eruptions. Simulations indicatethat the differences between GHG scenarios become importantonly in the latter half of the 21st century, and are largest in thenorthern mid-latitudes.2 Improved understanding of theeffects of the Mt. Pinatubo eruption on stratospheric ozonesuggests that a major volcanic eruption in the near future(while atmospheric chlorine levels from ODSs remain elevated)would result in lower levels of stratospheric ozone over muchof the globe that would persist for several years.232 In theArctic, the evolution of springtime ozone in the future isuncertain because it is still debated whether changes in ozonewill be driven by increases in PSCs from stratospheric coolingor by decreases in PCSs from stratospheric warming due toincreases in planetary wave activity.47
Despite these open issues, the simulations and predictionsof total ozone are more certain than the evolution of the otherfactors discussed above. Compared to the levels in the 1960sthe UVI levels in the 2010s are higher only at southern highand polar latitudes where the ozone hole continues to formduring the austral spring, and forces the annually averagedUVI to be up to 70% higher. Increases in UVI everywhere elseare very small or close to zero. The pattern for the futureis a near-complete reversal, as UVI is projected to decreaseover Antarctica in the 2090s by up to 40% compared to the2010s. Decreases in the UVI are projected for the rest of the
mid-latitude areas ranging between 5% and 10%. In thetropics, the changes are very small (±2–4%).
Overall effects. From the above discussion of individualfactors that will affect the levels of UV radiation by 2100, itappears that the ozone will continue to be the dominant factorover Antarctica, while clouds and surface reflectivity will domi-nate the changes over the Arctic. The effects of the aerosols,although highly uncertain, are potentially very important, andwill probably dominate future changes in both the UV-B andUV-A radiation in highly populated regions. Because thelargest potential effects are also the most uncertain, we do notattempt to combine the four panels to show an overall effect.
In our last assessment, it was projected, on the basis ofmodels available at that time, that there would be increases inUV at low latitudes by 2100 (where the UV is already high). Thepresent assessment does not support that general statement;Fig. 8 shows a more complex picture and any projectedincreases (due mainly to reductions in cloud) are smaller thanprovisionally projected.
Effect of the Montreal Protocol on UV radiation
The amended and adjusted Montreal Protocol continues to besuccessful in reducing emissions and atmospheric abun-dances of most controlled ODSs, and has been hailed as themost effective environmental treaty ever. As a result of itssuccess, the concentrations of most of the man-made chemi-cals that led to ozone depletion are declining, and ozone isjudged to be on a path towards recovery.2 Despite their longatmospheric lifetimes, by 2012, the total combined abundanceof anthropogenic ODSs in the troposphere had decreased bynearly 10% from the peak value in 1993. New estimates of thecontributions of specific substances or groups of substances tothe decline in tropospheric chlorine and bromine are nowavailable.2
Several attempts have been made to quantify the success ofthe amended and adjusted Montreal Protocol by comparingthe environmental implications of ozone differences in thefuture “world expected” with those in the future “worldavoided” by its successful implementation. A recent modelsimulation, that included the effects of coupling with the deepocean,233 showed that without the Montreal Protocol, ozoneconcentrations would have continued to decline, with an accel-eration of that decline in the latter part of this century. In2070, the stratospheric ozone layer would have collapsed toless than 100 DU worldwide and the peak UVI would havereached values greater than 35 in the tropics; at the sunlitnorthern polar cap UVI values would have been in the range5–15, which are similar to or larger than the values found inthe subtropics and tropics in 2000. Such an enhancement ofUV irradiance at the surface is beyond anything that modernecosystems have presumably experienced.
Another simulation223 that accounted also for the effects ofclimate change (e.g., changing albedo and cloud cover),reported the geographical distribution of changes in UVerythat would occur without the implementation of the MontrealProtocol. For the no-Montreal Protocol simulation, dramatic
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 35
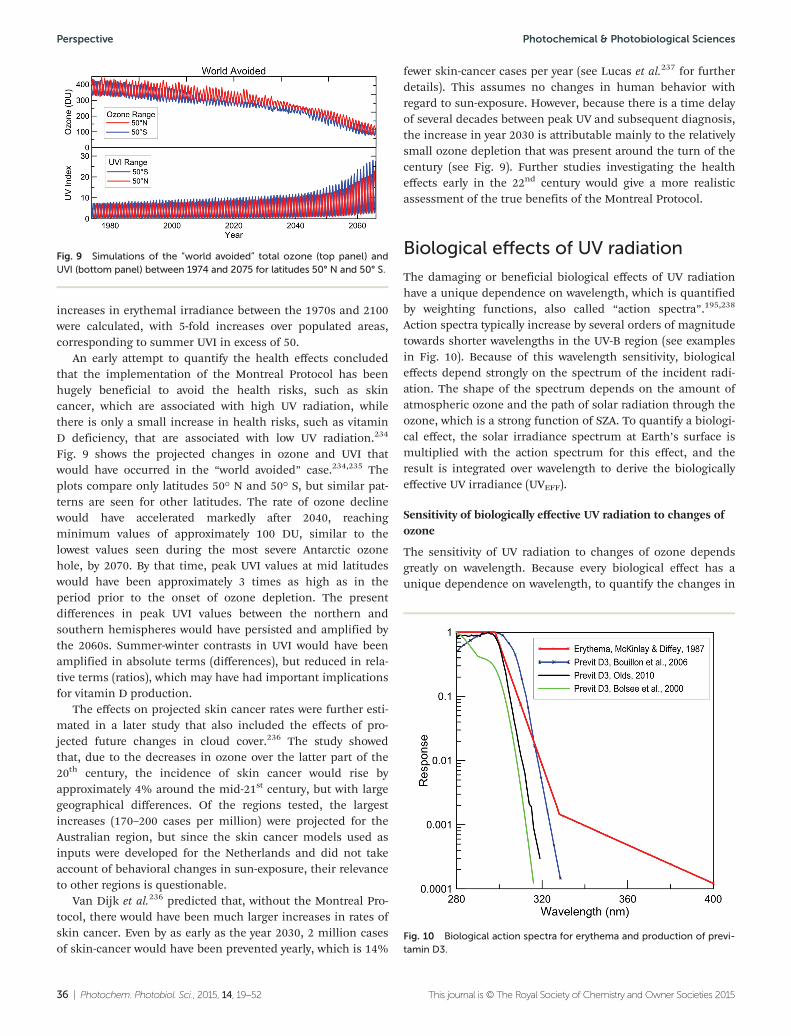
increases in erythemal irradiance between the 1970s and 2100were calculated, with 5-fold increases over populated areas,corresponding to summer UVI in excess of 50.
An early attempt to quantify the health effects concludedthat the implementation of the Montreal Protocol has beenhugely beneficial to avoid the health risks, such as skincancer, which are associated with high UV radiation, whilethere is only a small increase in health risks, such as vitaminD deficiency, that are associated with low UV radiation.234
Fig. 9 shows the projected changes in ozone and UVI thatwould have occurred in the “world avoided” case.234,235 Theplots compare only latitudes 50° N and 50° S, but similar pat-terns are seen for other latitudes. The rate of ozone declinewould have accelerated markedly after 2040, reachingminimum values of approximately 100 DU, similar to thelowest values seen during the most severe Antarctic ozonehole, by 2070. By that time, peak UVI values at mid latitudeswould have been approximately 3 times as high as in theperiod prior to the onset of ozone depletion. The presentdifferences in peak UVI values between the northern andsouthern hemispheres would have persisted and amplified bythe 2060s. Summer-winter contrasts in UVI would have beenamplified in absolute terms (differences), but reduced in rela-tive terms (ratios), which may have had important implicationsfor vitamin D production.
The effects on projected skin cancer rates were further esti-mated in a later study that also included the effects of pro-jected future changes in cloud cover.236 The study showedthat, due to the decreases in ozone over the latter part of the20th century, the incidence of skin cancer would rise byapproximately 4% around the mid-21st century, but with largegeographical differences. Of the regions tested, the largestincreases (170–200 cases per million) were projected for theAustralian region, but since the skin cancer models used asinputs were developed for the Netherlands and did not takeaccount of behavioral changes in sun-exposure, their relevanceto other regions is questionable.
Van Dijk et al.236 predicted that, without the Montreal Pro-tocol, there would have been much larger increases in rates ofskin cancer. Even by as early as the year 2030, 2 million casesof skin-cancer would have been prevented yearly, which is 14%
fewer skin-cancer cases per year (see Lucas et al.237 for furtherdetails). This assumes no changes in human behavior withregard to sun-exposure. However, because there is a time delayof several decades between peak UV and subsequent diagnosis,the increase in year 2030 is attributable mainly to the relativelysmall ozone depletion that was present around the turn of thecentury (see Fig. 9). Further studies investigating the healtheffects early in the 22nd century would give a more realisticassessment of the true benefits of the Montreal Protocol.
Biological effects of UV radiation
The damaging or beneficial biological effects of UV radiationhave a unique dependence on wavelength, which is quantifiedby weighting functions, also called “action spectra”.195,238
Action spectra typically increase by several orders of magnitudetowards shorter wavelengths in the UV-B region (see examplesin Fig. 10). Because of this wavelength sensitivity, biologicaleffects depend strongly on the spectrum of the incident radi-ation. The shape of the spectrum depends on the amount ofatmospheric ozone and the path of solar radiation through theozone, which is a strong function of SZA. To quantify a biologi-cal effect, the solar irradiance spectrum at Earth’s surface ismultiplied with the action spectrum for this effect, and theresult is integrated over wavelength to derive the biologicallyeffective UV irradiance (UVEFF).
Sensitivity of biologically effective UV radiation to changes ofozone
The sensitivity of UV radiation to changes of ozone dependsgreatly on wavelength. Because every biological effect has aunique dependence on wavelength, to quantify the changes in
Fig. 9 Simulations of the “world avoided” total ozone (top panel) andUVI (bottom panel) between 1974 and 2075 for latitudes 50° N and 50° S.
Fig. 10 Biological action spectra for erythema and production of previ-tamin D3.
Perspective Photochemical & Photobiological Sciences
36 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

biologically effective UV radiation due to changes in ozone,these wavelength dependencies should be taken into account.The relationship between change in total ozone column (TOC)and change in biologically effective UV irradiance can be quan-tified in terms of the Radiation Amplification Factor (RAF). Forsmall (<10%) changes in ozone the RAF is simply the relativefractional change in effective UV irradiance with fractionalchange in total column ozone:
RAF ¼ �ðΔUVEFF=UVEFFÞ=ðΔTOC=TOCÞ;
where ΔUVEFF and ΔTOC are the respective changes in UVEFF
and TOC. For example, RAF = 1.5 means that a 1% decrease inozone will lead to a 1.5% increase in biologically effective UVradiation. For larger (>10%) changes in ozone, the powerform239 is more appropriate:
UVEFFþ=UVEFF� ¼ ðTOC�=TOCþÞRAF;
where the subscripts (+ and −) refer to the cases with larger orsmaller values of ozone, respectively.
Biological effects that are dominated by UV-B wavelengthshave larger RAFs than effects where the contribution from thelonger wavelengths is significant. As an example, we illustratethis for the case of vitamin D production by UV radiation,because this topic has received prominence in recent years,and the action spectrum for previtamin D3 production is con-troversial.240 Table 1 shows the RAFs for three suggestedaction spectra for previtamin D3 production (Fig. 10) com-pared with the RAF for erythema for typical conditions inJanuary and July. The previtamin D3-related action spectra arefor illustrative purposes only and although the accuracy of thecurrently-accepted spectrum241 has been called into ques-tion,240 we are not in a position to advocate any change to it. Acomprehensive list of RAFs and action spectra for a largevariety of biological effects was included in the previousassessment report.124
RAFs also depend to some degree on factors other than SZAand ozone that alter the shape of the solar spectrum, such asextinction by aerosols or the vertical distribution of ozone inthe atmosphere.247 Limitations of the RAF and its applicationto other action spectra have been discussed in more detailelsewhere.239
New numerical parameterizations have recently beendeveloped to calculate the biologically effective irradiance as a
function of TOC and solar elevation angle for commonly usedaction spectra.194 Furthermore, a new method has beendevised to calculate RAFs from measurements of UVery andTOC during times when UVery is also affected by clouds andaerosols.248
While the basic understanding of the sensitivity of UV radi-ation to changes in ozone has not changed since the lastassessment, additional studies are now available that corrobo-rate the magnitude of this sensitivity and allow refinement ofthe values of action spectra.
The action spectrum for erythema
There have been slight variations in the definition of theaction spectrum for erythema since it was first introduced.243
The action spectrum for erythema was standardized by theCommission Internationale de l’Éclairage (CIE) in 1987249 andupdated in 1998.242 Deviations in erythemal irradiance result-ing from the two versions of the action spectrum are less than0.5% for SZAs <40° and increase to around 2% at 85° SZA244
and the RAF changes by less than 0.02 (Fig. 11). Even thoughthe differences are small, this change is important due to thelarge number of studies and time series that have been basedon the CIE 1987 action spectrum. In accordance with Webbet al.,244 we recommend that the standard action spectrum forerythema recommended by CIE242 should be used in thefuture.
Measurements of personal exposure to UV radiation
To date, the most widely used method for personal UVexposure studies is to equip volunteers with small dosimetersattached to various parts of the body. Dosimeters may bebased on photoresponsive films, which change their trans-mission upon exposure to UV radiation,251–253 DNA moleculesin cultures of immobilized spores with a spectral responsecorresponding to erythema,189,254,255 and photodetectors thatconvert UV radiation into signals of voltage or current.256–258
Table 1 RAFs for action spectra calculated on the basis of daily inte-grals for latitude 30° N. This is an update of Table 1 in McKenzie et al.195
EffectRAF Jan(290 DU)
RAF July(305 DU) Reference
Erythema(CIE, standardreference)
1.1 1.2 CIE;242 McKinley andDiffey;243 Webb et al.244
Previtamin D3 (CIE) 1.7 1.4 Bouillon et al.241
Previtamin D3 (CIEtruncated to 315 nm)
1.8 1.5 Bouillon et al.241
Previtamin D3 1.7 1.4 Olds245
Previtamin D3 2.6 2.2 Bolsee et al.246
Fig. 11 Ratio of radiation amplification factors (RAF) calculated for theCIE 1998 and CIE 1987 erythema action spectra. The figure is based onmodel spectra that were calculated for different solar zenith angles andtotal column ozone250 and subsequently weighted with either the CIE1998 or the CIE 1987 erythema action spectrum.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 37

The ability to convert doses absorbed by polysulphone (PS)badges into biologically effective solar UV exposure wasassessed,259 taking as an example two relevant effects forhuman skin: induction of erythema and production of previta-min D3. Comparisons of doses derived from PS badges posi-tioned horizontally and at different inclination angles (tosimulate various anatomic sites of the human body), revealedlarger deviations at large solar zenith angles and/or for highlyreflective surfaces.260
More recently, electronic dosimeters have become available,and their use is becoming more widespread.261–263 The accu-racy of measurements of two types of personal dosimeters,namely PS films and electronic ultraviolet (EUV) dosimetersusing an aluminum gallium nitride (Al27Ga73N) photodetector,have recently been assessed.262 PS dosimeters showed meanabsolute deviations of 26% relative to a reference spectro-radiometer, with a maximum deviation of 44%. Since thePS doses were derived using a single calibration curve, furtherexperimental investigation of these dosimeters is neededto better assess their accuracy. The calibrated EUV dosimetersshowed mean absolute deviations of 15% (maximum 33%),which were partly caused by small, but significant sensitivitiesto visible radiation (i.e. stray light). It was concluded that cali-brating UV sensors by direct comparison with a referenceinstrument leads to reliable results and that these simpledevices are useful to estimate personal UV exposures. Theyshould not be used, however, as an inexpensive replacementfor meteorological grade instruments.
Inexpensive, non-scientific instruments for UVI measure-ments have recently become available. These sensors are partof watches, portable weather stations, and handheld UVmeters. The accuracy of several of these devices has beenassessed.264 While the measurements of some test devicesagreed with those of a reference spectroradiometer to within20%, some instruments overestimated the UVI by up to afactor of three and hence did not provide trustworthy results.
Health-related exposure to UV radiation
Exposure to sunlight, specifically the UV radiation, has bothpositive and negative health effects, as discussed in Lucaset al.237 Despite the positive effects, it is excessive sun-exposurethat has been of greater concern, because of its adverse effectsto humans, terrestrial and aquatic ecosystems, materials, andair quality (see companion papers31,32,56,237,265,266). Quantifi-cation of exposure of humans to UV radiation is complex asmany factors are involved, including the natural variations ofradiation, the orientation of the exposed parts in conjunctionwith the time and location, behavioral aspects, clothing, aswell as effects of reflections on the surroundings.253
This complexity is confirmed by recent exposure studiesusing personal dosimeters, which include: seafarers of mer-chant vessels,189 farmers at a mid-latitude site,256 urban dwell-ers engaging in typical outdoor activities such as shopping,walking, cycling, and sightseeing,257 professional cyclists,254
young (age 9–12) skiers,255 vineyard workers,252 and peopleengaged in activities such as walking, sitting, and lying.258
From measurements with personal electronic UV dosimetersin New Zealand over a few weeks (outside the peak summerperiod),261,267 it was shown that cumulative doses received bydosimeters worn on the wrist were typically less than 2% of theavailable ambient doses, and that the equivalent full bodyexposure is less than 1% of the ambient. This implies that thepeople wearing the dosimeters were probably indoors for about95% of the time. These studies suggest that personal exposure toUV radiation is better approximated by dosimeters and diariesthan by measurements of the ambient UV irradiance. Further-more, the translation of traditional global-horizontal irradiancemeasurements into exposure levels relevant to humans thereforedepends critically on information on behavior and location.
Maximizing the benefits while minimizing the damage is amultifaceted problem in which many of the elements areimportant and need to be quantified.268 A recent attempt hasbeen made to quantify the available ambient UV doses eachmonth, including both beneficial and detrimental effects, inNorthern Eurasia.269
As discussed above, problems arise when one uses UVmeasurements on a horizontal surface to perform risk-benefitassessments because they do not yield the actual doses peopleget while they are outdoors, as different parts of the body areexposed at different angles. More realistic UV doses for peoplewho are outdoors engaged in a variety of different activitiescan be estimated from simple geometrical parameterizations.270
As part of further refinements, the importance of includingthe effects of clouds and aerosols in parameterizations toderive vitamin D-effective irradiance from erythemal irradiancewas highlighted by Feister et al.190 They showed, from 4 yearsof measurements in Germany, that optically thick clouds canstrongly modify the ratio between erythemal and vitaminD-effective irradiance, suggesting that the parameterizationsderived for cloud-free conditions are not always applicable.
Similarly, the role of shade has been emphasised by Turn-bull and Parisi271 who measured the spectral dependence ofthe ratio of diffuse to global UV radiation, and showed thatunder clear skies this ratio decreases with wavelength through-out most of the UV region for wavelengths greater than300 nm. For example, for unpolluted conditions at SZA = 40°,the diffuse to global ratio decreases from ∼0.6 at 300 nm to∼0.3 at 400 nm. This means that in the UV-B region, the pro-tection offered is less than one would estimate from our per-ception of the visible solar radiation. To get protectionsufficient for most purposes (e.g., Solar Protection Factor >30)from UV-B radiation in the shade, it is important to ensuremost of the diffuse sky radiation is also blocked. Interestingly,because the wavelengths for previtamin D production areslightly shorter than for erythema, this implies that exposureto diffuse light, such as in the shade of trees or buildings, mayslightly favor the production of vitamin D while minimizingthe risk of erythema in non-covered skin.
The field of atmospheric UV research is plagued withdifficulties in nomenclature. A recent report272 highlightssome of the issues, taking vitamin-D synthesis, a beneficialeffect, as a specific example. Terminologies for the standard
Perspective Photochemical & Photobiological Sciences
38 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

vitamin-D dose (SDD) and the minimum vitamin-D dose – fordaily sufficiency – (MDD) are proposed, analogous to the stan-dard erythema dose (SED) and minimal erythema dose (MED)that are in common use for erythema. Note that the quantitativevalue of the MDD is not yet known; nor is there agreement onthe recommended minimum levels of the status of vitamin-D(see Chapter 2). In the present literature, the SDD has confus-ingly been defined in terms of a physiological response. In rec-ognition of the fact that currently accepted action spectra maybe revised if new data become available, the continuation ofspectrally resolved irradiance measurements will allow reproces-sing of biologically effective irradiances and doses in the future.
Effects of geoengineering on ozoneand UV radiation
Geoengineering – or “climate engineering” – refers to a broadset of methods and technologies that could be used to deliber-ately alter the climate system in order to alleviate the impactsof climate change. Solar Radiation Management (SRM) hasbeen suggested as a means to counteract the warming fromincreasing GHG by reducing the amount of solar radiationabsorbed by the Earth’s surface. Carbon Dioxide Reduction(CDR) aims at reducing the future concentrations of CO2 byaccelerating the natural removal of atmospheric CO2 orincreasing the storage of carbon in reservoirs.45
Of those two geoengineering methods, only the SRM woulddirectly influence the amount of UV radiation received atEarth’s surface. Space reflectors, injection of aerosols in thestratosphere, or seeding of marine clouds would reduce theamount of UV radiation reaching the surface. In contrast,increasing of surface reflectivity by creating micro bubbles atthe ocean surface, growing more reflective crops, or paintingroofs and other built structures in light colors may lead toincreased surface UV radiation through scattering of reflectedradiation towards the ground.
The injection of sulfur dioxide into the stratosphere273 isone of the methods suggested to reduce the amount of solarradiation reaching Earth’s surface, through increased scatter-ing of solar radiation to space. However, it is known that stra-tospheric sulfate aerosols from volcanic eruptions and naturalemissions deplete the stratospheric ozone, and similar effectsshould be expected from stratospheric aerosols introduced forSRM, leading ultimately to increases in the amount of UV-Bradiation reaching the surface, which are larger than thereduction achieved from the SRM.
This is further supported by recent modelling studies274–276
suggesting that such interventions would lead to a generaldecrease in stratospheric ozone concentrations, mainly viachanges in photolysis rates, tropical upwelling of ozone-poorair, and an increase in available surfaces for heterogeneouschemistry. Considering the role of very short-lived halogens inthe stratosphere (e.g., Bry and Cly), increases in annual averageUVery of up to 5% in mid and high latitudes were simulated forthe 2040s due to the impact of stratospheric sulfur on
ozone.275 Recently, it was projected that the increase in UV-Bradiation at the surface due to ozone depletion could be offsetin the 2040s by the screening due to the SRM aerosols in thetropics and mid-latitudes, while in polar regions the UV-B radi-ation would increase by 5% on average, with 12% peakincreases during springtime.277
Other potential UV-related impacts of geoengineering havebeen investigated with models, revealing effects on cloudcover278 and rainfall patterns,279 both of which ultimately leadto changes in UV radiation at the Earth’s surface. Model simu-lations showed that SRM, to counteract a 1% annual rise inatmospheric CO2, suppresses the increases in precipitationthat would otherwise accompany the rising GHG, had geoengi-neering never been used. However, in some of these sensitivitystudies extreme and perhaps unlikely scenarios279 have beenassumed. For example, the effect of geoengineering withsulfate aerosols was investigated in a world with a differentclimate by first setting global CO2 levels at an extremely largelevel of 1120 parts per million (ppm) or four times the pre-industrial level. They found that global precipitation rateswould increase by approximately 7% compared with pre-indus-trial times, but with high spatial and temporal variability.However, when they re-ran the models, with SRM geoengineer-ing included, they found a 4.5% reduction in global precipi-tation. Again, there was a high degree of variability, butnotably, decreases in precipitation rates on land and in oceanswere much more similar.
The interactions and feedback of the suggested geoengineer-ing methods with the natural variability of the atmospheric andsurface reflectivity are not yet fully explored. The atmosphere isa complex system and any deliberate interventions should betreated with great care as they may have unanticipated adverseeffects. Moreover, with the current observing systems, it wouldnot be feasible to assess whether the small intended changesin solar radiation at Earth’s surface from the implementationof geo-engineering would have actually occurred.
Gaps in knowledge
Simulations of surface UV radiation for the future are limitedin accuracy due to difficulties in assessing the combinedeffects of clouds and aerosols that are expected to change.Over ice- and or snow-covered areas, these effects are evenmore complicated. Additional uncertainties for UV projectionsarise from the scenarios describing the evolution of the atmos-phere in the future.
Significant changes in aerosol concentrations are expectedin the future (both positive and negative, depending on theregion). The effect of these aerosols on surface UV irradiancewill depend strongly on their single scattering albedo.However, even for the aerosols that are present in the currentatmosphere, this parameter is not well quantified in the UV-Bregion because of the confounding effects of ozone absorption.Better quantification of the SSA in the UV-B region over a widerange of aerosol types (possibly involving laboratory studies to
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 39

avoid the effect of ozone) would increase our ability to modelthe effects of aerosol on the UV radiation at the Earth’s surface.
To our current understanding, variations in solar activitylead to decreases in total ozone of up to 3% between themaximum and the minimum of the 11-year solar cycle.However, the relative contributions of solar activity-inducedradiative and dynamical effects on ozone are not yet fullyresolved. Moreover, there are still uncertainties in the measure-ments of solar spectral irradiance outside Earth’s atmosphere.
Improvement in the understanding of these processes andthe availability of higher quality information of the UV-relatedfactors would strengthen our ability to effectively interpretongoing changes and predict future changes in UV radiation.
Instruments at the ground measure UV irradiance directlyand the results are therefore more accurate than the inversionresults from satellite data, but the spatial coverage of surfaceobservations is sparse and vast regions of the Earth (e.g.,Africa, Siberia, the global oceans, particularly in the southernhemisphere) are not being monitored from the ground. Inlight of these limitations, robust assessments of long termchanges in UV radiation must be based both on observationsfrom space and from the ground.
Assessment of the long-term benefits of the Montreal Proto-col requires input from both the atmospheric sciences and thehealth communities. To date, the health costs of non-implementation of the Montreal Protocol have been calculatedonly up to the year 2030. Because of the lag between UVexposure and the onset of diseases this is representative onlyof the changes due to ozone depletion up to the year 2000.This severely underestimates the true benefits of the MontrealProtocol because ozone changes are likely to have becomemuch larger in the latter half of the 21st century (see Fig. 9).
There is incomplete characterisation of many action spectraof interest. Examples are the action spectrum for the formationof previtamin D3 from sunlight and the action spectrum formelanoma in human skin. These are required for quantitativeassessments of the environmental effects of future changes inUV radiation, particularly those due to changes in ozone.
Acknowledgements
We acknowledge the contribution of Ilias Fountoulakis, PhDstudent at the Aristotle University of Thessaloniki, in the model-ling of UV radiation projections. GB’s contribution was sup-ported by the U.S. National Science Foundation (grantARC-1203250). KT’s contribution was supported by the projectFP7-SPACE-2012–313188. Richard McKenzie’s participation wassponsored by the New Zealand Government’s Ministry for theEnvironment, and supported through the Ministry of Business,Innovation and Employment’s research contract C01X1008.
References
1 WMO, (World Meteorological Organization): ScientificAssessment of Ozone Depletion: 2010, Global Ozone
Research and Monitoring Project, Report No. 52, Geneva,Switzerland 2011, p. 438.
2 WMO, (World Meteorological Organization): ScientificAssessment of Ozone Depletion: 2014, Global OzoneResearch and Monitoring Project, Report No. 55, Geneva,Switzerland 2015.
3 S. Tilmes, J. Fasullo, J.-F. Lamarque, D. R. Marsh,M. Mills, K. Alterskjær, H. Muri, J. E. Kristjánsson,O. Boucher, M. Schulz, J. N. S. Cole, C. L. Curry, A. Jones,J. Haywood, P. J. Irvine, D. Ji, J. C. Moore, D. B. Karam,B. Kravitz, P. J. Rasch, B. Singh, J.-H. Yoon, U. Niemeier,H. Schmidt, A. Robock, S. Yang and S. Watanabe, Thehydrological impact of geoengineering in the Geoengi-neering Model Intercomparison Project (GeoMIP),J. Geophys. Res., [Atmos.], 2013, 118, 11,036–11,058.
4 R. S. Stolarski, A. R. Douglass, E. E. Remsberg,N. J. Livesey and J. C. Gille, Ozone temperature corre-lations in the upper stratosphere as a measure of chlorinecontent, J. Geophys. Res., [Atmos.], 2012, 117, D10305.
5 WMO, (World Meteorological Organization): ScientificAssessment of Ozone Depletion: 2006, Global ozoneResearch and Monitoring Project, Report No. 50, Geneva,Switzerland 2007, p. 572.
6 W. Chehade, M. Weber and J. P. Burrows, Total ozonetrends and variability during 1979–2012 from mergeddata sets of various satellites, Atmos. Chem. Phys., 2014,14, 7059–7074.
7 M. Coldewey-Egbers, D. G. Loyola R, P. Braesicke,M. Dameris, M. van Roozendael, C. Lerot and W. Zimmer,A new health check of the ozone layer at global andregional scales, Geophys. Res. Lett., 2014, 41, 4363–4372.
8 B. Hassler, J. S. Daniel, B. J. Johnson, S. Solomon andS. J. Oltmans, An assessment of changing ozone loss ratesat South Pole: Twenty-five years of ozonesonde measure-ments, J. Geophys. Res., [Atmos.], 2011, 116, D22301.
9 B. Hassler, G. E. Bodeker, S. Solomon and P. J. Young,Changes in the polar vortex: Effects on Antarctic totalozone observations at various stations, Geophys. Res. Lett.,2011, 38, L01805.
10 J. R. Ziemke and S. Chandra, Development of a climaterecord of tropospheric and stratospheric column ozonefrom satellite remote sensing: evidence of an early recov-ery of global stratospheric ozone, Atmos. Chem. Phys.,2012, 12, 5737–5753.
11 A. T. J. de Laat, R. J. van der A, M. A. F. Allaart, M. vanWeele, G. C. Benitez, C. Casiccia, N. M. Paes Leme,E. Quel, J. Salvador and E. Wolfram, Extreme sunbathing:Three weeks of small total O3 columns and high UV radi-ation over the southern tip of South America during the2009 Antarctic O3 hole season, Geophys. Res. Lett., 2010,37, L14805.
12 G. L. Manney, M. L. Santee, M. Rex, N. J. Livesey,M. C. Pitts, P. Veefkind, E. R. Nash, I. Wohltmann,R. Lehmann, L. Froidevaux, L. R. Poole, M. R. Schoeberl,D. P. Haffner, J. Davies, V. Dorokhov, H. Gernandt,B. Johnson, R. Kivi, E. Kyro, N. Larsen, P. F. Levelt,
Perspective Photochemical & Photobiological Sciences
40 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

A. Makshtas, C. T. McElroy, H. Nakajima, M. C. Parrondo,D. W. Tarasick, P. von der Gathen, K. A. Walker andN. S. Zinoviev, Unprecedented Arctic ozone loss in 2011,Nature, 2011, 478, 469–475.
13 H. E. Rieder, J. Staehelin, J. A. Maeder, T. Peter,M. Ribatet, A. C. Davison, R. Stübi, P. Weihs andF. Holawe, Extreme events in total ozone over Arosa - Part2: Fingerprints of atmospheric dynamics and chemistryand effects on mean values and long-term changes,Atmos. Chem. Phys., 2010, 10, 10033–10045.
14 H. E. Rieder, J. Staehelin, J. A. Maeder, T. Peter,M. Ribatet, A. C. Davison, R. Stübi, P. Weihs andF. Holawe, Extreme events in total ozone over Arosa - Part1: Application of extreme value theory, Atmos. Chem. Phys.,2010, 10, 10021–10031.
15 M. Fitzka, J. Hadzimustafic and S. Simic, Total ozone andUmkehr observations at Hoher Sonnblick 1994–2011: Cli-matology and extreme events, J. Geophys. Res., [Atmos.],2014, 119, 739–752.
16 M. C. Reader, D. A. Plummer, J. F. Scinocca andT. G. Shepherd, Contributions to twentieth century totalcolumn ozone change from halocarbons, troposphericozone precursors, and climate change, Geophys. Res. Lett.,2013, 40, 6276–6281.
17 M. Salby, E. A. Titova and L. Deschamps, Changes of theAntarctic ozone hole: Controlling mechanisms, seasonalpredictability, and evolution, J. Geophys. Res., [Atmos.],2012, 117, D10111.
18 M. Salby, E. Titova and L. Deschamps, Rebound of Antarcticozone, Geophys. Res. Lett., 2011, 38, L09702.
19 J. Kuttippurath, F. Lefèvre, J. P. Pommereau, H. K. Roscoe,F. Goutail, A. Pazmiño and J. D. Shanklin, Antarctic ozoneloss in 1979–2010: first sign of ozone recovery, Atmos.Chem. Phys., 2013, 13, 1625–1635.
20 J. Kuttippurath, S. Godin-Beekmann, F. Lefèvre, G. Nikulin,M. L. Santee and L. Froidevaux, Record-breaking ozoneloss in the Arctic winter 2010/2011: comparison with 1996/1997, Atmos. Chem. Phys., 2012, 12, 7073–7085.
21 J. P. Pommereau, F. Goutail, F. Lefèvre, A. Pazmino,C. Adams, V. Dorokhov, P. Eriksen, R. Kivi, K. Stebel,X. Zhao and M. van Roozendael, Why unprecedentedozone loss in the Arctic in 2011? Is it related to climatechange?, Atmos. Chem. Phys., 2013, 13, 5299–5308.
22 G. Bernhard, G. Manney, V. Fioletov, J.-U. Grooß,A. Heikkilä, B. Johnsen, T. Koskela, K. Lakkala, R. Müller,C. L. Myhre and M. Rex, Ozone and UV radiation, in: Stateof the Climate in 2011, Bull. Am. Meteorol. Soc., 2012, 93,S129–S132.
23 D. Balis, I. S. A. Isaksen, C. Zerefos, I. Zyrichidou,K. Eleftheratos, K. Tourpali, R. Bojkov, B. Rognerud,F. Stordal, O. A. Søvde and Y. Orsolini, Observed and mod-elled record ozone decline over the Arctic during winter/spring 2011, Geophys. Res. Lett., 2011, 38, L23801.
24 I. S. A. Isaksen, C. Zerefos, W. C. Wang, D. Balis,K. Eleftheratos, B. Rognerud, F. Stordal, T. K. Berntsen,J. H. LaCasce, O. A. Søvde, D. Olivié, Y. J. Orsolini,
I. Zyrichidou, M. Prather and O. N. E. Tuinder, Attributionof the Arctic ozone column deficit in March 2011,Geophys. Res. Lett., 2012, 39, L24810.
25 R. R. Garcia, Atmospheric science: An Arctic ozone hole?,Nature, 2011, 478, 462–463.
26 B. Petkov, V. Vitale, C. Tomasi, A. M. Siani, G. Seckmeyer,A. Webb, A. R. D. Smedley, G. R. Casale, R. Werner,C. Lanconelli, M. Mazzola, A. Lupi, M. Busetto,H. Diémoz, F. Goutail, U. Köhler, B. T. Mendeva,W. Josefsson, D. Moore, M. L. Bartolomé, J. R. MoretaGonzález, O. Mišaga, A. Dahlback, Z. Tóth, S. Varghese,H. De Backer, R. Stübi and K. Vaníček, Response of theozone column over Europe to the 2011 Arctic ozonedepletion event according to ground-based observationsand assessment of the consequent variations in surfaceUV irradiance, Atmos. Environ., 2014, 85, 169–178.
27 B. M. Sinnhuber, G. Stiller, R. Ruhnke, T. von Clarmann,S. Kellmann and J. Aschmann, Arctic winter 2010/2011 atthe brink of an ozone hole, Geophys. Res. Lett., 2011, 38,L24814.
28 D. L. Hartmann, A. M. G. K. Tank, M. Rusticucci,L. V. Alexander, S. Brönnimann, Y. Charabi, F. J. Dentener,E. J. Dlugokencky, D. R. Easterling, A. Kaplan, B. J. Soden,P. W. Thorne, M. Wild and P. M. Zhai, Observations: Atmos-phere and Surface, in Climate Change 2013: The PhysicalScience Basis. Contribution of Working Group I to the FifthAssessment Report of the Intergovernmental Panel on ClimateChange, ed. T. F. Stocker, D. Qin, G.-K. Plattner, M. Tignor,S. K. Allen, J. Boschung, A. Nauels, Y. Xia and V. Bex and P.M. Midgley, Cambridge University Press, Cambridge,United Kingdom and New York, NY, U.S.A., 2013.
29 D. W. J. Thompson, D. J. Seidel, W. J. Randel, C.-Z. Zou,A. H. Butler, C. Mears, A. Osso, C. Long and R. Lin, Themystery of recent stratospheric temperature trends,Nature, 2012, 491, 692–697.
30 D. J. Seidel, N. P. Gillett, J. R. Lanzante, K. P. Shine andP. W. Thorne, Stratospheric temperature trends: our evol-ving understanding, Wiley Interdiscip. Rev. Climate Change,2011, 2, 592–616.
31 J. F. Bornman, P. W. Barnes, S. A. Robinson, C. L. Ballaré,S. D. Flint and M. M. Caldwell, Solar ultraviolet radiationand ozone depletion-driven climate change: effects onterrestrial ecosystems, Photochem. Photobiol. Sci., 2015, 14,DOI: 10.1039/c4pp90034k, this issue.
32 D. J. Erickson III, B. Sulzberger, R. Zepp, A. T. Austin andN. Paul, Effects of solar UV radiation and climate changeon biogeochemical cycling: Interactions and feedbacks,Photochem. Photobiol. Sci., 2015, 14, DOI: 10.1039/c4pp90036g, this issue.
33 S. M. Kang, L. M. Polvani, J. C. Fyfe, S. W. Son,M. Sigmond and G. J. P. Correa, Modeling evidence thatozone depletion has impacted extreme precipitation inthe austral summer, Geophys. Res. Lett., 2013, 40, 4054–4059.
34 S. M. Kang, L. M. Polvani, J. C. Fyfe and M. Sigmond, Impactof polar ozone depletion on subtropical precipitation,Science, 2011, 332, 951–954.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 41

35 R. D. Hudson, Measurements of the movement of the jetstreams at mid-latitudes, in the Northern and SouthernHemispheres, 1979 to 2010, Atmos. Chem. Phys., 2012, 12,7797–7808.
36 S.-W. Son, E. P. Gerber, J. Perlwitz, L. M. Polvani, N. Gillett,K.-H. Seo, V. Eyring, T. G. Shepherd, D. Waugh, H. Akiyoshi,J. Austin, A. Baumgaertner, S. Bekki, P. Braesicke, C. Brühl,N. Butchart, M. Chipperfield, D. Cugnet, M. Dameris,S. Dhomse, S. Frith, H. Garny, R. Garcia, S. C. Hardiman,P. Jöckel, J.-F. Lamarque, E. Mancini, M. Marchand,M. Michou, T. Nakamura, O. Morgenstern, G. Pitari,D. A. Plummer, J. Pyle, E. Rozanov, J. F. Scinocca,K. Shibata, D. Smale, H. Teyssèdre, W. Tian andY. Yamashita, The impact of stratospheric ozone on theSouthern Hemisphere circulation change: A multimodelassessment, J. Geophys. Res., [Atmos.], 2010, 115, D014271.
37 D. W. Waugh, F. Primeau, T. DeVries and M. Holzer,Recent Changes in the Ventilation of the SouthernOceans, Science, 2013, 339, 568–570.
38 S. A. Robinson and D. J. Erickson III, Not just aboutsunburn – the ozone hole’s profound effect on climate hassignificant implications for Southern Hemisphere ecosys-tems, Global Change Biol., 2014, DOI: 10.1111/gcb.12739.
39 J. M. Arblaster, G. A. Meehl and D. J. Karoly, Futureclimate change in the Southern Hemisphere: Competingeffects of ozone and greenhouse gases, Geophys. Res. Lett.,2011, 38, L02701.
40 A. Y. Karpechko, N. P. Gillett, L. J. Gray and M. Dall’Amico,Influence of ozone recovery and greenhouse gas increaseson Southern Hemisphere circulation, J. Geophys. Res.,[Atmos.], 2010, 115, D22117.
41 L. M. Polvani, M. Previdi and C. Deser, Large cancellation,due to ozone recovery, of future Southern Hemisphereatmospheric circulation trends, Geophys. Res. Lett., 2011,38, L04707.
42 M. Sigmond, M. C. Reader, J. C. Fyfe and N. P. Gillett,Drivers of past and future Southern Ocean change: Strato-spheric ozone versus greenhouse gas impacts, Geophys.Res. Lett., 2011, 38, L12601.
43 K. L. Smith, L. M. Polvani and D. R. Marsh, Mitigation of21st century Antarctic sea ice loss by stratospheric ozonerecovery, Geophys. Res. Lett., 2012, 39, L20701.
44 C. McLandress, T. G. Shepherd, J. F. Scinocca, D. A. Plummer,M. Sigmond, A. I. Jonsson and M. C. Reader, Separatingthe Dynamical Effects of Climate Change and OzoneDepletion. Part II: Southern Hemisphere Troposphere,J. Clim., 2011, 24, 1850–1868.
45 IPCC, (Intergovernmental Panel on Climate Change):Working Group I Contribution to the IPCC Fifth Assess-ment Report Climate Change 2013: The Physical ScienceBasis Cambridge University Press, Cambridge, UnitedKingdom and New York, NY, U.S.A., Cambridge, UK, andNew York, NY, 2013.
46 M. Wang and J. E. Overland, A sea ice free summer Arcticwithin 30 years: An update from CMIP5 models, Geophys.Res. Lett., 2012, 39, L18501.
47 U. Langematz, S. Meul, K. Grunow, E. Romanowsky,S. Oberländer, J. Abalichin and A. Kubin, Future arctictemperature and ozone: The role of stratospheric compo-sition changes, J. Geophys. Res., [Atmos.], 2014, 119, 2092–2112.
48 R. W. Portmann, J. S. Daniel and A. R. Ravishankara, Stra-tospheric ozone depletion due to nitrous oxide: influencesof other gases, Philos. Trans. R. Soc. London., Ser B, 2012,367, 1256–1264.
49 R. Lei, Z. Zhang, I. Matero, B. Cheng, Q. Li and W. Huang,Reflection and transmission of irradiance by snow andsea ice in the central Arctic Ocean in summer 2010, PolarRes., 2012, 31, 17325.
50 J. C. Comiso, C. L. Parkinson, R. Gersten and L. Stock,Accelerated decline in the Arctic sea ice cover, Geophys.Res. Lett., 2008, 35, L01703.
51 C. L. Parkinson and D. J. Cavalieri, Antarctic sea ice varia-bility and trends, 1979–2010, Cryosphere, 2012, 6, 871–880.
52 R. Kwok and D. A. Rothrock, Decline in Arctic sea icethickness from submarine and ICESat records: 1958–2008,Geophys. Res. Lett., 2009, 36, L15501.
53 T. Markus, J. C. Stroeve and J. Miller, Recent changes inArctic sea ice melt onset, freezeup, and melt seasonlength, J. Geophys. Res.: Oceans, 2009, 114, C12024.
54 D. K. Perovich and C. Polashenski, Albedo evolution ofseasonal Arctic sea ice, Geophys. Res. Lett., 2012, 39,L08501.
55 J. Blunden and D. S. Arndt, State of the Climate in 2013,Bull. Am. Meteorol. Soc., 2014, 95, S1–S257.
56 S. Madronich, M. Shao, S. R. Wilson, K. R. Solomon,J. Longstrethe and X. Tang, Changes in air quality and tro-pospheric composition due to depletion of stratosphericozone and interactions with changing climate: Impli-cations for human and environmental health, Photochem.Photobiol. Sci., 2015, 14, DOI: 10.1039/c4pp90037e, this issue.
57 IPCC, (Intergovernmental Panel on Climate Change):Climate Change 2007: The Physical Science Basis. Contri-bution of Working Group I to the Fourth AssessmentReport of the Intergovernmental Panel on ClimateChange, Cambridge University Press, Cambridge, UnitedKingdom and New York, NY, USA, Cambridge, UK, andNew York, NY, 2007.
58 NARSTO, Particulate Matter Science for Policy Makers: ANARSTO Assessment, Cambridge, England, 2004.
59 Y. Chen and S. Xie, Temporal and spatial visibility trendsin the Sichuan Basin, China, 1973 to 2010, Atmos. Res.,2012, 112, 25–34.
60 J. Wu, X. Zhang, J. Hu and D. Zhao, Probability ofdifferent visibility grades in China over a 50-year period,Meteorol. Atmos. Phys., 2013, 122, 115–123.
61 S. Sabetghadam, F. Ahmadi-Givi and Y. Golestani,Visibility trends in Tehran during 1958–2008, Atmos.Environ., 2012, 62, 512–520.
62 K. Wang, R. E. Dickinson and S. Liang, Clear Sky VisibilityHas Decreased over Land Globally from 1973 to 2007,Science, 2009, 323, 1468–1470.
Perspective Photochemical & Photobiological Sciences
42 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

63 M. Doyle and S. Dorling, Visibility trends in the UK1950–1997, Atmos. Environ., 2002, 36, 3161–3172.
64 C. Ruckstuhl, R. Philipona, K. Behrens, M. Collaud Coen,B. Dürr, A. Heimo, C. Mätzler, S. Nyeki, A. Ohmura,L. Vuilleumier, M. Weller, C. Wehrli and A. Zelenka,Aerosol and cloud effects on solar brightening and therecent rapid warming, Geophys. Res. Lett., 2008, 35,L12708.
65 J. L. Jimenez, M. R. Canagaratna, N. M. Donahue,A. S. H. Prevot, Q. Zhang, J. H. Kroll, P. F. DeCarlo,J. D. Allan, H. Coe, N. L. Ng, A. C. Aiken, K. S. Docherty,I. M. Ulbrich, A. P. Grieshop, A. L. Robinson, J. Duplissy,J. D. Smith, K. R. Wilson, V. A. Lanz, C. Hueglin, Y. L. Sun,J. Tian, A. Laaksonen, T. Raatikainen, J. Rautiainen,P. Vaattovaara, M. Ehn, M. Kulmala, J. M. Tomlinson,D. R. Collins, M. J. Cubison, E. J. Dunlea, J. A. Huffman,T. B. Onasch, M. R. Alfarra, P. I. Williams, K. Bower,Y. Kondo, J. Schneider, F. Drewnick, S. Borrmann,S. Weimer, K. Demerjian, D. Salcedo, L. Cottrell,R. Griffin, A. Takami, T. Miyoshi, S. Hatakeyama,A. Shimono, J. Y. Sun, Y. M. Zhang, K. Dzepina,J. R. Kimmel, D. Sueper, J. T. Jayne, S. C. Herndon,A. M. Trimborn, L. R. Williams, E. C. Wood,A. M. Middlebrook, C. E. Kolb, U. Baltensperger andD. R. Worsnop, Evolution of organic aerosols in theatmosphere, Science, 2009, 326, 1525–1529.
66 A. Laskin, J. Laskin and S. A. Nizkorodov, Mass spectro-metric approaches for chemical characterisation of atmos-pheric aerosols: critical review of the most recentadvances, Environ. Chem., 2012, 9, 163–189.
67 M. Collaud Coen, E. Andrews, A. Asmi, U. Baltensperger,N. Bukowiecki, D. Day, M. Fiebig, A. M. Fjaeraa,H. Flentje, A. Hyvärinen, A. Jefferson, S. G. Jennings,G. Kouvarakis, H. Lihavainen, C. Lund Myhre, W. C. Malm,N. Mihapopoulos, J. V. Molenar, C. O’Dowd, J. A. Ogren,B. A. Schichtel, P. Sheridan, A. Virkkula, E. Weingartner,R. Weller and P. Laj, Aerosol decadal trends – Part 1:In situ optical measurements at GAW and IMPROVEstations, Atmos. Chem. Phys., 2013, 13, 869–894.
68 A. Cazorla, R. Bahadur, K. J. Suski, J. F. Cahill, D. Chand,B. Schmid, V. Ramanathan and K. A. Prather, Relatingaerosol absorption due to soot, organic carbon, and dustto emission sources determined from in situ chemicalmeasurements, Atmos. Chem. Phys., 2013, 13, 9337–9350.
69 T. F. Eck, B. N. Holben, A. Sinyuk, R. T. Pinker, P. Goloub,H. Chen, B. Chatenet, Z. Li, R. P. Singh, S. N. Tripathi,J. S. Reid, D. M. Giles, O. Dubovik, N. T. O’Neill,A. Smirnov, P. Wang and X. Xia, Climatological aspects ofthe optical properties of fine/coarse mode aerosol mix-tures, J. Geophys. Res., [Atmos.], 2010, 115, D19205.
70 D. M. Giles, B. N. Holben, T. F. Eck, A. Sinyuk, A. Smirnov,I. Slutsker, R. R. Dickerson, A. M. Thompson andJ. S. Schafer, An analysis of AERONET aerosol absorptionproperties and classifications representative of aerosolsource regions, J. Geophys. Res., [Atmos.], 2012, 117,D17203.
71 R. A. Kahn, D. L. Nelson, M. J. Garay, R. C. Levy,M. A. Bull, D. J. Diner, J. V. Martonchik, S. R. Paradise,E. G. Hansen and L. A. Remer, MISR aerosol product attri-butes and statistical comparisons with MODIS, Geosci.Remote Sens. IEEE Transact., 2009, 47, 4095–4114.
72 A. H. Omar, D. M. Winker, M. A. Vaughan, Y. Hu,C. R. Trepte, R. A. Ferrare, K.-P. Lee, C. A. Hostetler,C. Kittaka, R. R. Rogers, R. E. Kuehn and Z. Liu, TheCALIPSO Automated Aerosol Classification and LidarRatio Selection Algorithm, J. Atmos. Ocean. Technol., 2009,26, 1994–2014.
73 L. A. Remer, S. Mattoo, R. C. Levy and L. A. Munchak,MODIS 3 km aerosol product: algorithm and global per-spective, Atmos. Meas. Tech., 2013, 6, 1829–1844.
74 J. Zhang and J. S. Reid, A decadal regional and globaltrend analysis of the aerosol optical depth using a data-assimilation grade over-water MODIS and Level 2 MISRaerosol products, Atmos. Chem. Phys., 2010, 10, 10949–10963.
75 S. Kinne, D. O’Donnel, P. Stier, S. Kloster, K. Zhang,H. Schmidt, S. Rast, M. Giorgetta, T. F. Eck andB. Stevens, MAC-v1: A new global aerosol climatology forclimate studies, J. Adv. Model. Earth Syst., 2013, 5, 704–740.
76 V. E. Cachorro, A. Berjón, C. Toledano, S. Mogo, N. Prats,A. M. de Frutos, J. M. Vilaplana, M. Sorribas, B. A. De LaMorena, J. Gröbner and N. Laulainen, Detailed AerosolOptical Depth Intercomparison between Brewer and Li-Cor 1800 Spectroradiometers and a Cimel Sun Photo-meter, J. Atmos. Ocean. Technol., 2009, 26, 1558–1571.
77 E. Gerasopoulos, V. Amiridis, S. Kazadzis, P. Kokkalis,K. Eleftheratos, M. O. Andreae, T. W. Andreae, H. El-Askaryand C. S. Zerefos, Three-year ground based measurementsof aerosol optical depth over the Eastern Mediterranean:the urban environment of Athens, Atmos. Chem. Phys.,2011, 11, 2145–2159.
78 W. Kumharn, J. S. Rimmer, A. R. D. Smedley, T. Y. Yingand A. R. Webb, Aerosol optical depth and the globalBrewer network: A study using U.K.- and Malaysia-basedBrewer spectrophotometers, J. Atmos. Ocean. Technol.,2012, 29, 857–866.
79 J. B. Liley and B. W. Forgan, Aerosol optical depth overLauder, New Zealand, Geophys. Res. Lett., 2009, 36,L07811.
80 R. L. McKenzie, C. Weinreis, P. V. Johnston, B. Liley,H. Shiona, M. Kotkamp, D. Smale, N. Takegawa andY. Kondo, Effects of urban pollution on UV spectral irra-diances, Atmos. Chem. Phys., 2008, 8, 5683–5697.
81 G. G. Palancar, B. L. Lefer, S. R. Hall, W. J. Shaw,C. A. Corr, S. C. Herndon, J. R. Slusser and S. Madronich,Effect of aerosols and NO2 concentration on ultravioletactinic flux near Mexico City during MILAGRO: measure-ments and model calculations, Atmos. Chem. Phys., 2013,13, 1011–1022.
82 N. Chubarova, Y. Nezval, I. Sviridenkov, A. Smirnov andI. Slutsker, Smoke aerosol and its radiative effects during
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 43

extreme fire event over Central Russia in summer 2010,Atmos. Meas. Tech., 2012, 5, 557–568.
83 R. R. Dickerson, S. K. Ondragunta, G. Stenchikov,K. L. Civerolo, B. G. Doddridge and B. N. Holben, Theimpact of aerosols on solar ultraviolet radiation andphotochemical smog, Science, 1997, 278, 827–830.
84 Y. Feng, V. Ramanathan and V. R. Kotamarthi, Browncarbon: a significant atmospheric absorber of solar radi-ation?, Atmos. Chem. Phys., 2013, 13, 8607–8621.
85 A. Hoffer, A. Gelencsér, P. Guyon, G. Kiss, O. Schmid,G. P. Frank, P. Artaxo and M. O. Andreae, Optical pro-perties of humic-like substances (HULIS) in biomass-burning aerosols, Atmos. Chem. Phys., 2006, 6, 3563–3570.
86 T. W. Kirchstetter and T. L. Thatcher, Contribution oforganic carbon to wood smoke particulate matter absorp-tion of solar radiation, Atmos. Chem. Phys., 2012, 12,6067–6072.
87 S. Kazadzis, N. Kouremeti, A. Bais, A. Kazantzidis andC. Meleti, Aerosol forcing efficiency in the UVA regionfrom spectral solar irradiance measurements at an urbanenvironment, Ann. Geophys., 2009, 27, 2515–2522.
88 C. A. Corr, S. R. Hall, K. Ullmann, B. E. Anderson,A. J. Beyersdorf, K. L. Thornhill, M. J. Cubison,J. L. Jimenez, A. Wisthaler and J. E. Dibb, Spectral absorp-tion of biomass burning aerosol determined fromretrieved single scattering albedo during ARCTAS, Atmos.Chem. Phys., 2012, 12, 10505–10518.
89 B. N. Wenny, J. S. Schafer, J. J. DeLuisi, V. K. Saxena,W. F. Barnard, I. V. Petropavlovskikh and A. J. Vergamini,A study of regional aerosol radiative properties and effectson ultraviolet-B radiation, J. Geophys. Res., [Atmos.], 1998,103, 17083–17097.
90 J. L. Petters, V. K. Saxena, J. R. Slusser, B. N. Wenny andS. Madronich, Aerosol single scattering albedo retrievedfrom measurements of surface UV irradiance and a radia-tive transfer model, J. Geophys. Res., [Atmos.], 2003, 108,4288.
91 M. A. Wetzel, G. E. Shaw, J. R. Slusser, R. D. Borys andC. F. Cahill, Physical, chemical, and ultraviolet radiativecharacteristics of aerosol in central Alaska, J. Geophys.Res., [Atmos.], 2003, 108, 4418.
92 C. D. Goering, T. S. L’Ecuyer, G. L. Stephens, J. R. Slusser,G. Scott, J. Davis, J. C. Barnard and S. Madronich, Simul-taneous retrievals of column ozone and aerosol opticalproperties from direct and diffuse solar irradiancemeasurements, J. Geophys. Res., [Atmos.], 2005, 110, D05204.
93 A. F. Bais, A. Kazantzidis, S. Kazadzis, D. S. Balis,C. S. Zerefos and C. Meleti, Deriving an effective aerosolsingle scattering albedo from spectral surface UV irradi-ance measurements, Atmos. Environ., 2005, 39, 1093–1102.
94 J. C. Barnard, R. Volkamer and E. I. Kassianov, Estimationof the mass absorption cross section of the organiccarbon component of aerosols in the Mexico City Metro-politan Area, Atmos. Chem. Phys., 2008, 8, 6665–6679.
95 C. A. Corr, N. Krotkov, S. Madronich, J. R. Slusser,B. Holben, W. Gao, J. Flynn, B. Lefer and S. M. Kreidenweis,
Retrieval of aerosol single scattering albedo at ultravioletwavelengths at the T1 site during MILAGRO, Atmos. Chem.Phys., 2009, 9, 5813–5827.
96 I. Ialongo, V. Buchard, C. Brogniez, G. R. Casale andA. M. Siani, Aerosol single scattering albedo retrieval inthe UV range: an application to OMI satellite validation,Atmos. Chem. Phys., 2010, 10, 331–340.
97 M. Gyawali, W. P. Arnott, R. A. Zaveri, C. Song,H. Moosmüller, L. Liu, M. I. Mishchenko, L. W. A. Chen,M. C. Green, J. G. Watson and J. C. Chow, Photoacousticoptical properties at UV, VIS, and near IR wavelengths forlaboratory generated and winter time ambient urban aero-sols, Atmos. Chem. Phys., 2012, 12, 2587–2601.
98 V. Buchard, C. Brogniez, F. Auriol and B. Bonnel, Aerosolsingle scattering albedo retrieved from ground-basedmeasurements in the UV and visible region, Atmos. Meas.Tech., 2011, 4, 1–7.
99 R. Medina, R. M. Fitzgerald and Q. Min, Retrieval of thesingle scattering albedo in the El Paso-Juarez Airshedusing the TUV model and a UV-MFRSR radiometer, Atmos.Environ., 2012, 46, 430–440.
100 Q. Zhang, J. L. Jimenez, M. R. Canagaratna, J. D. Allan,H. Coe, I. Ulbrich, M. R. Alfarra, A. Takami,A. M. Middlebrook, Y. L. Sun, K. Dzepina, E. Dunlea,K. Docherty, P. F. DeCarlo, D. Salcedo, T. Onasch,J. T. Jayne, T. Miyoshi, A. Shimono, S. Hatakeyama,N. Takegawa, Y. Kondo, J. Schneider, F. Drewnick,S. Borrmann, S. Weimer, K. Demerjian, P. Williams,K. Bower, R. Bahreini, L. Cottrell, R. J. Griffin,J. Rautiainen, J. Y. Sun and Y. M. Zhang, Ubiquity anddominance of oxygenated species in organic aerosols inanthropogenically-influenced Northern Hemisphere mid-latitudes, Geophys. Res. Lett., 2007, 34, L13801.
101 C. L. Heald, H. Coe, J. L. Jimenez, R. J. Weber,R. Bahreini, A. M. Middlebrook, L. M. Russell, M. Jolleys,T. M. Fu, J. D. Allan, K. N. Bower, G. Capes, J. Crosier,W. T. Morgan, N. H. Robinson, P. I. Williams,M. J. Cubison, P. F. DeCarlo and E. J. Dunlea, Exploringthe vertical profileof atmospheric organic aerosol: com-paring 17 aircraft field campaigns with a global model,Atmos. Chem. Phys., 2011, 11, 12673–12696.
102 D. L. Bones, D. K. Henricksen, S. A. Mang, M. Gonsior,A. P. Bateman, T. B. Nguyen, W. J. Cooper andS. A. Nizkorodov, Appearance of strong absorbers andfluorophores in limonene-O3 secondary organic aerosoldue to NH4+-mediated chemical aging over long timescales, J. Geophys. Res., [Atmos.], 2010, 115, D05203.
103 X. Zhang, Y.-H. Lin, J. D. Surratt, P. Zotter, A. S. H. Prévôtand R. J. Weber, Light-absorbing soluble organic aerosolin Los Angeles and Atlanta: A contrast in secondaryorganic aerosol, Geophys. Res. Lett., 2011, 38, L21810.
104 Y. Desyaterik, Y. Sun, X. Shen, T. Lee, X. Wang, T. Wangand J. L. Collett, Speciation of “brown” carbon in cloudwater impacted by agricultural biomass burning ineastern China, J. Geophys. Res., [Atmos.], 2013, 118,7389–7399.
Perspective Photochemical & Photobiological Sciences
44 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

105 S. A. Epstein, E. Tapavicza, F. Furche and S. A. Nizkorodov,Direct photolysis of carbonyl compounds dissolved in cloudand fog-droplets, Atmos. Chem. Phys., 2013, 13, 9461–9477.
106 A. T. Lambe, C. D. Cappa, P. Massoli, T. B. Onasch,S. D. Forestieri, A. T. Martin, M. J. Cummings,D. R. Croasdale, W. H. Brune, D. R. Worsnop andP. Davidovits, Relationship between Oxidation Level andOptical Properties of Secondary Organic Aerosol, Environ.Sci. Technol., 2013, 47, 6349–6357.
107 K. M. Updyke, T. B. Nguyen and S. A. Nizkorodov, For-mation of brown carbon via reactions of ammonia withsecondary organic aerosols from biogenic and anthropo-genic precursors, Atmos. Environ., 2012, 63, 22–31.
108 A. G. Rincón, M. I. Guzmán, M. R. Hoffmann andA. J. Colussi, Thermochromism of Model Organic AerosolMatter, J. Phys. Chem. Lett., 2009, 1, 368–373.
109 D. G. Streets, F. Yan, M. Chin, T. Diehl, N. Mahowald,M. Schultz, M. Wild, Y. Wu and C. Yu, Anthropogenic andnatural contributions to regional trends in aerosol opticaldepth, 1980–2006, J. Geophys. Res., [Atmos.], 2009, 114,D00D18.
110 M. Chin, T. Diehl, Q. Tan, J. M. Prospero, R. A. Kahn,L. A. Remer, H. Yu, A. M. Sayer, H. Bian, I. V. Geogdzhayev,B. N. Holben, S. G. Howell, B. J. Huebert, N. C. Hsu,D. Kim, T. L. Kucsera, R. C. Levy, M. I. Mishchenko,X. Pan, P. K. Quinn, G. L. Schuster, D. G. Streets,S. A. Strode, O. Torres and X. P. Zhao, Multi-decadalaerosol variations from 1980 to 2009: a perspective fromobservations and a global model, Atmos. Chem. Phys.,2014, 14, 3657–3690.
111 P. N. den Outer, H. Slaper, J. Kaurola, A. Lindfors,A. Kazantzidis, A. F. Bais, U. Feister, J. Junk, M. Janouchand W. Josefsson, Reconstructing of erythemal ultravioletradiation levels in Europe for the past 4 decades,J. Geophys. Res., [Atmos.], 2010, 115, 10102.
112 C. S. Zerefos, K. Eleftheratos, C. Meleti, S. Kazadzis,A. Romanou, C. Ichoku, G. Tselioudis and A. Bais, Solardimming and brightening over Thessaloniki, Greece, andBeijing, China, Tellus Ser. B, 2009, 61, 657–665.
113 C. Meleti, A. F. Bais, S. Kazadzis, N. Kouremeti, K. Garaneand C. Zerefos, Factors affecting solar ultraviolet irradi-ance measuredsince 1990 at Thessaloniki, Greece,Int. J. Remote Sens., 2009, 30, 4167–4179.
114 J. Yoon, W. von Hoyningen-Huene, M. Vountas andJ. P. Burrows, Analysis of linear long-term trend of aerosoloptical thickness derived from SeaWiFS using BAER overEurope and South China, Atmos. Chem. Phys., 2011, 11,12149–12167.
115 Z. B. Wang, M. Hu, Z. J. Wu, D. L. Yue, L. Y. He,X. F. Huang, X. G. Liu and A. Wiedensohler, Long-termmeasurements of particle number size distributions andthe relationships with air mass history and source appor-tionment in the summer of Beijing, Atmos. Chem. Phys.,2013, 13, 10159–10170.
116 J. F. Lamarque, T. C. Bond, V. Eyring, C. Granier, A. Heil,Z. Klimont, D. Lee, C. Liousse, A. Mieville, B. Owen,
M. G. Schultz, D. Shindell, S. J. Smith, E. Stehfest, J. VanAardenne, O. R. Cooper, M. Kainuma, N. Mahowald,J. R. McConnell, V. Naik, K. Riahi and D. P. van Vuuren,Historical (1850–2000) gridded anthropogenic andbiomass burning emissions of reactive gases and aerosols:methodology and application, Atmos. Chem. Phys., 2010,10, 7017–7039.
117 S. J. Smith and T. C. Bond, Two hundred fifty years ofaerosols and climate: the end of the age of aerosols,Atmos. Chem. Phys., 2014, 14, 537–549.
118 N. Bellouin, J. Rae, A. Jones, C. Johnson, J. Haywood andO. Boucher, Aerosol forcing in the Climate Model Inter-comparison Project (CMIP5) simulations by HadGEM2-ESand the role of ammonium nitrate, J. Geophys. Res.,[Atmos.], 2011, 116, D20206.
119 A. de Miguel, R. Román, J. Bilbao and D. Mateos, Evol-ution of erythemal and total shortwave solar radiation inValladolid, Spain: Effects of atmospheric factors, J. Atmos.Sol.–Terr. Phys., 2011, 73, 578–586.
120 A. A. Silva, Erythemal dose rate under noon overcast skies,Photochem. Photobiol. Sci., 2013, 12, 777–786.
121 A. Kazantzidis, K. Eleftheratos and C. S. Zerefos, Effects ofcirrus cloudiness on solar irradiance in four spectralbands, Atmos. Res., 2011, 102, 452–459.
122 Y. Liu, J. R. Key, Z. Liu, X. Wang and S. J. Vavrus, A clou-dier Arctic expected with diminishing sea ice, Geophys.Res. Lett., 2012, 39, L05705.
123 M. Antón, L. Alados-Arboledas, J. L. Guerrero-Rascado,M. J. Costa, J. C. Chiu and F. J. Olmo, Experimental andmodeled UV erythemal irradiance under overcast con-ditions: the role of cloud optical depth, Atmos. Chem.Phys., 2012, 12, 11723–11732.
124 UNEP, Environmental Effects of Ozone Depletion and itsInteraction with Climate Change: 2010 Assessment, UnitedNations Environment Programme (UNEP), Nairobi, 2010.
125 A. Kreuter, M. Blumthaler, A. R. Webb, A. F. Bais, R. Kiftand N. Kouremeti, Effects of Albedo on Solar Irradiance,in Advances in Meteorology, Climatology and AtmosphericPhysics, Springer, Berlin, Heidelberg, Germany, 2013,pp. 1089–1095.
126 O. Meinander, S. Kazadzis, A. Arola, A. Riihelä,P. Räisänen, R. Kivi, A. Kontu, R. Kouznetsov, M. Sofiev,J. Svensson, H. Suokanerva, V. Aaltonen, T. Manninen,J. L. Roujean and O. Hautecoeur, Spectral albedo of seaso-nal snow during intensive melt period at Sodankylä,beyond the Arctic Circle, Atmos. Chem. Phys., 2013, 13,3793–3810.
127 S. Simic, M. Fitzka, A. Schmalwieser, P. Weihs andJ. Hadzimustafic, Factors affecting UV irradiance atselected wavelengths at Hoher Sonnblick, Atmos. Res.,2011, 101, 869–878.
128 P. J. Hezel, X. Zhang, C. M. Bitz, B. P. Kelly andF. Massonnet, Projected decline in spring snow depth onArctic sea ice caused by progressively later autumn openocean freeze-up this century, Geophys. Res. Lett., 2012, 39,L17505.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 45

129 K. E. Frey, D. K. Perovich and B. Light, The spatial distri-bution of solar radiation under a melting Arctic sea icecover, Geophys. Res. Lett., 2011, 38, L22501.
130 E. J. Steig, Q. Ding, D. S. Battisti and A. Jenkins, Tropicalforcing of Circumpolar Deep Water Inflow and outletglacier thinning in the Amundsen Sea Embayment, WestAntarctica, Ann. Glaciol., 2012, 53, 19–28.
131 E. Rignot, J. Mouginot, M. Morlighem, H. Seroussi andB. Scheuchl, Widespread, rapid grounding line retreat ofPine Island, Thwaites, Smith, and Kohler glaciers, WestAntarctica, from 1992 to 2011, Geophys. Res. Lett., 2014,41, 3502–3509.
132 I. Joughin, B. E. Smith and B. Medley, Marine ice sheetcollapse potentially under way for the Thwaites GlacierBasin, West Antarctica, Science, 2014, 344, 735–738.
133 L. L. Hood and B. E. Soukharev, The Lower-StratosphericResponse to 11-Yr Solar Forcing: Coupling to the Tropo-sphere–Ocean Response, J. Atmos. Sci., 2012, 69, 1841–1864.
134 M. Lockwood, C. Bell, T. Woollings, R. G. Harrison,L. J. Gray and J. D. Haigh, Top-down solar modulation ofclimate: evidence for centennial-scale change, Environ.Res. Lett., 2010, 5, 034008.
135 J. W. Harder, J. M. Fontenla, P. Pilewskie, E. C. Richardand T. N. Woods, Trends in solar spectral irradiance varia-bility in the visible and infrared, Geophys. Res. Lett., 2009,36, L07801.
136 J. D. Haigh, A. R. Winning, R. Toumi and J. W. Harder, Aninfluence of solar spectral variations on radiative forcingof climate, Nature, 2010, 467, 696–699.
137 I. Ermolli, K. Matthes, T. Dudok de Wit, N. A. Krivova,K. Tourpali, M. Weber, Y. C. Unruh, L. Gray, U. Langematz,P. Pilewskie, E. Rozanov, W. Schmutz, A. Shapiro,S. K. Solanki and T. N. Woods, Recent variability of thesolar spectral irradiance and its impact on climate model-ling, Atmos. Chem. Phys., 2013, 13, 3945–3977.
138 W. H. Swartz, R. S. Stolarski, L. D. Oman, E. L. Flemingand C. H. Jackman, Middle atmosphere response todifferent descriptions of the 11-yr solar cycle in spectralirradiance in a chemistry-climate model, Atmos. Chem.Phys., 2012, 12, 5937–5948.
139 M. Lockwood, M. J. Owens, L. Barnard, C. J. Davis andF. Steinhilber, The persistence of solar activity indicatorsand the descent of the Sun into Maunder Minimum con-ditions, Geophys. Res. Lett., 2011, 38, L22105.
140 M. Lockwood, A. P. Rouillard and I. D. Finch, The riseand fall of open solar flux during the current grand solarmaximum, Astrophys. J., 2009, 700, 937.
141 F. Steinhilber and J. Beer, Prediction of solar activity forthe next 500 years, J. Geophys. Res.: Space Phys., 2013, 118,1861–1867.
142 M. J. Owens, M. Lockwood, L. Barnard and C. J. Davis,Solar cycle 24: Implications for energetic particles andlong-term space climate change, Geophys. Res. Lett., 2011,38, L19106.
143 J. A. Abreu, J. Beer, F. Steinhilber, S. M. Tobias andN. O. Weiss, For how long will the current grand
maximum of solar activity persist?, Geophys. Res. Lett.,2008, 35, L20109.
144 J. G. Anet, E. V. Rozanov, S. Muthers, T. Peter,S. Brönnimann, F. Arfeuille, J. Beer, A. I. Shapiro,C. C. Raible, F. Steinhilber and W. K. Schmutz, Impact ofa potential 21st century “grand solar minimum” onsurface temperatures and stratospheric ozone, Geophys.Res. Lett., 2013, 40, 4420–4425.
145 J. E. Frederick and A. L. Hodge, Solar irradiance at theearth’s surface: long-term behavior observed at the SouthPole, Atmos. Chem. Phys., 2011, 11, 1177–1189.
146 D. S. Berger and F. Urbach, A climatology of sunburningultraviolet radiation, Photochem. Photobiol., 1982, 35, 187–192.
147 N. Y. Chubarova and Y. I. Nezval, Thirty year variability ofUV irradiance in Moscow, J. Geophys. Res., [Atmos.], 2000,105, 12529–12539.
148 J. C. Farman, B. G. Gardiner and J. D. Shanklin, Largelosses of total ozone in Antarctica reveal seasonal ClOx/NOx interaction, Nature, 1985, 315, 207–210.
149 Z. Gao, N.-B. Chang and W. Gao, Comparative analyses ofthe ultraviolet-B flux over the continental United Statebased on the NASA total ozone mapping spectrometerdata and USDA ground-based measurements, J. Appl.Remote Sens., 2010, 4, 043547.
150 M. G. Kimlin, K. A. Schallhorn, R. S. Meltzer, J. R. Slusserand K. Lantz, Comparison of ultraviolet data from colo-cated instruments from the U.S. EPA Brewer Spectropho-tometer Network and the U.S. Department of AgricultureUV-B Monitoring and Research Program, Opt. Eng., 2005,44, 1–8.
151 V. E. Fioletov, L. J. McArthur, T. W. Mathews andL. Marrett, Estimated ultraviolet exposure levels for asufficient vitamin D status in North America, J. Photo-chem. Photobiol., B, 2010, 100, 57–66.
152 H. He, V. E. Fioletov, D. W. Tarasick, T. W. Mathews andC. Long, Validation of Environment Canada and NOAAUV Index Forecasts with Brewer Measurements fromCanada, J. Appl. Meteorol. Climatol., 2013, 52, 1477–1489.
153 S. Cabrera, A. Ipiña, A. Damiani, R. R. Cordero andR. D. Piacentini, UV index values and trends in Santiago,Chile (33.5°S) based on ground and satellite data, J. Photo-chem. Photobiol., B, 2012, 115, 73–84.
154 S. Diaz, M. Vernet, A. Paladini, H. Fuenzalida, G. Deferrari,C. R. Booth, S. Cabrera, C. Casiccia, M. Dieguez,C. Lovengreen, J. Pedroni, A. Rosales and J. Vrsalovic,Availability of vitamin D photoconversion weighted UVradiation in southern South America, Photochem. Photo-biol. Sci., 2011, 10, 1854–1867.
155 J. Krzyścin, P. Sobolewski, J. Jarosławski, J. Podgórski andB. Rajewska-Więch, Erythemal UV observations at Belsk,Poland, in the period 1976–2008: Data homogenization,climatology, and trends, Acta Geophys., 2011, 59, 155–182.
156 G. Seckmeyer, D. Pissulla, M. Glandorf, D. Henriques,B. Johnsen, A. Webb, A. M. Siani, A. Bais, B. Kjeldstad,C. Brogniez, J. Lenoble, B. Gardiner, P. Kirsch, T. Koskela,
Perspective Photochemical & Photobiological Sciences
46 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

J. Kaurola, B. Uhlmann, H. Slaper, P. den Outer,M. Janouch, P. Werle, J. Grobner, B. Mayer, A. de la Casi-niere, S. Simic and F. Carvalho, Variability of UV irradi-ance in Europe, Photochem. Photobiol., 2008, 84, 172–179.
157 M. P. Utrillas, M. J. Marín, A. R. Esteve, V. Estellés,S. Gandía, J. A. Núnez and J. A. Martínez-Lozano, Tenyears of measured UV Index from the Spanish UVB Radio-metric Network, J. Photochem. Photobiol., B, 2013, 125,1–7.
158 A. M. Siani, S. Modesti, G. R. Casale, H. Diemoz andA. Colosimo, Biologically effective surface UV climatologyat Rome and Aosta, Italy, AIP Conf. Proc., 2013, 1531, 903–906.
159 R. L. McKenzie, J. B. Liley and L. O. Björn, UV Radiation:Balancing Risks and Benefits, Photochem. Photobiol., 2009,85, 88–98.
160 P. Gies, A. Klekociuk, M. Tully, S. Henderson,J. Javorniczky, K. King, L. Lemus-Deschamps andJ. Makin, Low Ozone Over Southern Australia in August2011 and its Impact on Solar Ultraviolet Radiation Levels,Photochem. Photobiol., 2013, 89, 984–994.
161 G. Bernhard, A. Dahlback, V. Fioletov, A. Heikkilä,B. Johnsen, T. Koskela, K. Lakkala and T. Svendby, Highlevels of ultraviolet radiation observed by ground-basedinstruments below the 2011 Arctic ozone hole, Atmos.Chem. Phys., 2013, 13, 10573–10590.
162 G. Bernhard, C. Booth and J. Ehramjian, Climatology ofUltraviolet Radiation at High Latitudes Derived fromMeasurements of the National Science Foundation’s Ultra-violet Spectral Irradiance Monitoring Network, in UV Radi-ation in Global Climate Change, ed. W. Gao, J. Slusserand D. Schmoldt, Springer, Berlin, Heidelberg, 2010,pp. 48–72.
163 N. P. Sharma, B. K. Bhattarai, B. Sapkota and B. Kjeldstad,Satellite estimation and ground measurements of solarUV index and influence of aerosol in ground based UVdata in Kathmandu, Nepal, Int. J. Environ. Sci., 2013, 3,1362–1372.
164 S. Buntoung and A. R. Webb, Comparison of erythemalUV irradiances from Ozone Monitoring Instrument (OMI)and ground-based data at four Thai stations, J. Geophys.Res., [Atmos.], 2010, 115, D18215.
165 S. Janjai, S. Wisitsirikun, S. Buntoung, S. Pattarapanitchai,R. Wattan, I. Masiri and B. K. Bhattarai, Comparison of UVindex from Ozone Monitoring Instrument (OMI) withmulti-channel filter radiometers at four sites in the tropics:effects of aerosols and clouds, Int. J. Climatol., 2013, 34,453–461.
166 B. Hu and Y. S. Wang, Ultraviolet radiation in the back-ground region over Northeast China, J. Atmos. Chem.,2013, 70, 283–296.
167 A. Webb, B. G. Gardiner, K. Leszczynski, V. A. Mohnen,P. Johnston, N. Harrison and D. Bigelow, Quality Assur-ance in Monitoring Solar Ultraviolet Radiation: The Stateof the Art, World Meteorological Organisation, GlobalAtmospheric Watch, Report No. 146, Geneva, 2003.
168 G. Seckmeyer, A. Bais, G. Bernhard, M. Blumthaler,S. Drüke, P. Kiedron, K. Lantz, R. L. McKenzie andS. Riechelmann, Instruments to Measure Solar UltravioletRadiation Part 4: Array Spectroradiometers, World Meteoro-logical Organization, Global Atmospheric Watch, ReportNo. 191, Geneva, 2010, p. 43.
169 A. Kreuter, R. Buras, B. Mayer, A. Webb, R. Kift, A. Bais,N. Kouremeti and M. Blumthaler, Solar irradiance in theheterogeneous albedo environment of the Arctic coast:measurements and a 3-D model study, Atmos. Chem.Phys., 2014, 14, 5989–6002.
170 R. R. Cordero, G. Seckmeyer, S. Riechelmann, A. Damianiand F. Labbe, Monte Carlo-based uncertainty analysis ofUV array spectroradiometers, Metrologia, 2012, 49, 745.
171 R. R. Cordero, G. Seckmeyer, A. Damiani, F. Labbe andD. Laroze, Monte Carlo-based uncertainties of surface UVestimates from models and from spectroradiometers,Metrologia, 2013, 50, DOI: 10.1088/0026-1394/50/5/L1.
172 D. Vernez, A. Milon, L. Vuilleumier and J. L. Bulliard, Ana-tomical exposure patterns of skin to sunlight: relative con-tributions of direct, diffuse and reflected ultravioletradiation, Br. J. Dermatol., 2012, 167, 383–390.
173 G. Seckmeyer, M. Schrempf, A. Wieczorek, S. Riechelmann,K. Graw, S. Seckmeyer and M. Zankl, A novel method tocalculate solar UV exposure relevant to vitamin D pro-duction in humans, Photochem. Photobiol., 2013, 89, 974–983.
174 S. Riechelmann, M. Schrempf and G. Seckmeyer, Simul-taneous measurement of spectral sky radiance by a non-scanning multidirectional spectroradiometer (MUDIS),Meas. Sci. Technol., 2013, 24, 125501.
175 A. Lindfors, A. Tanskanen, A. Arola, R. van der A, A. Bais,U. Feister, M. Janouch, W. Josefsson, T. Koskela,K. Lakkala, P. N. den Outer, A. R. D. Smedley, H. Slaperand A. R. Webb, The PROMOTE UV record: Toward aglobal satellite-based climatology of surface ultravioletirradiance, IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens.,2009, 2, 207–212.
176 A. Arola, S. Kazadzis, A. Lindfors, N. Krotkov, J. Kujanpää,J. Tamminen, A. Bais, A. d. Sarra, J. M. Villaplana,C. Brogniez, A. M. Siani, M. Janouch, P. Weihs, A. Webb,T. Koskela, N. Kouremeti, D. Meloni, V. Buchard,F. Auriol, I. Ialongo, M. Staneck, S. Simic, A. Smedley andS. Kinne, A new approach to correct for absorbing aerosolsin OMI UV, Geophys. Res. Lett., 2009, 36, L22805.
177 A. Damiani, S. Cabrera, R. C. Muñoz, R. R. Cordero andF. Labbe, Satellite-derived UV irradiance for a regionwith complex morphology and meteorology: comparisonagainst ground measurements in Santiago de Chile,Int. J. Remote Sens., 2013, 34, 5812–5833.
178 R. R. Cordero, G. Seckmeyer, A. Damiani, J. Jorquera,J. Carrasco, R. Muñoz, L. Da Silva, F. Labbe and D. Laroze,Aerosol effects on the UV irradiance in Santiago de Chile,Atmos. Res., 2014, 149, 282–291.
179 F. Jégou, S. Godin-Beekman, M. P. Corrêa, C. Brogniez,F. Auriol, V. H. Peuch, M. Haeffelin, A. Pazmino, P. Saiag,
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 47

F. Goutail and E. Mahé, Validity of satellite measurementsused for the monitoring of UV radiation risk on health,Atmos. Chem. Phys., 2011, 11, 13377–13394.
180 M. Antón, V. E. Cachorro, J. M. Vilaplana, C. Toledano,N. A. Krotkov, A. Arola, A. Serrano and B. de la Morena,Comparison of UV irradiances from Aura/Ozone Monitor-ing Instrument (OMI) with Brewer measurements at ElArenosillo (Spain) – Part 1: Analysis of parameter influ-ence, Atmos. Chem. Phys., 2010, 10, 5979–5989.
181 M. Xu, X. Z. Liang, W. Gao and N. Krotkov, Comparison ofTOMS retrievals and UVMRP measurements of surfacespectral UV radiation in the United States, Atmos. Chem.Phys., 2010, 10, 8669–8683.
182 J. E. Wagner, F. Angelini, A. Arola, M. Blumthaler, M. Fitzka,G. P. Gobbi, R. Kift, A. Kreuter, H. E. Rieder, S. Simic,A. N. N. Webb and P. Weihs, Comparison of surface UVirradiance in mountainous regions derived from satelliteobservations and model calculations with ground-basedmeasurements, Metero. Zeits., 2010, 19, 481–490.
183 P. Pandey, K. De Ridder, D. Gillotay and N. P. M. vanLipzig, Estimating cloud optical thickness and associatedsurface UV irradiance from SEVIRI by implementing asemi-analytical cloud retrieval algorithm, Atmos. Chem.Phys., 2012, 12, 7961–7975.
184 R. L. McKenzie, P. V. Johnston and G. Seckmeyer,UV spectro-radiometry in the network for the detectionof stratospheric change (NDSC), in Solar UltravioletRadiation. Modelling, Measurements and Effects, ed.C. S. Zerefos and A. F. Bais, Springer-Verlag, Berlin, 1997,vol. 1.52, pp. 279–287.
185 G. Seckmeyer and R. L. McKenzie, Increased ultraviolet-radiation in New-Zealand (45-degrees-S) relative toGermany (48-degrees-N), Nature, 1992, 359, 135–137.
186 F. Zaratti, R. D. Piacentini, H. A. Guillen, S. Cabrera,B. Liley and R. McKenzie, Proposal for a modification ofthe UVI Risk Scale, Photochem. Photobiol. Sci., 2014, 13,980–985.
187 R. R. Cordero, G. Seckmeyer, A. Damiani, S. Riechelmann,J. Rayas, F. Labbe and D. Laroze, The world’s highestlevels of surface UV, Photochem. Photobiol. Sci., 2014, 13,70–81.
188 G. Norsang, Y.-C. Chen, N. Pingcuo, A. Dahlback,Ø. Frette, B. Kjeldstad, B. Hamre, K. Stamnes andJ. J. Stamnes, Comparison of ground-based measurementsof solar UV radiation at four sites on the Tibetan Plateau,Appl. Opt., 2014, 53, 736–747.
189 U. Feister, G. Meyer and U. Kirst, Solar UV exposure of sea-farers along subtropical and tropical shipping routes,Photochem. Photobiol., 2013, 89, 1497–1506.
190 U. Feister, G. Laschewski and R.-D. Grewe, UV index fore-casts and measurements of health-effective radiation,J. Photochem. Photobiol., B, 2011, 102, 55–68.
191 N. A. Cabrol, U. Feister, D.-P. Häder, H. Piazena,E. A. Grin and A. Klein, Record solar UV irradiance in thetropical Andes, Front. Environ. Sci., 2014, 2, DOI: 10.3389/fenvs.2014.00019.
192 A. Y. Karpechko, L. Backman, L. Thölix, I. Ialongo,M. Andersson, V. Fioletov, A. Heikkilä, B. Johnsen,T. Koskela, E. Kyrölä, K. Lakkala, C. L. Myhre, M. Rex,V. F. Sofieva, J. Tamminen and I. Wohltmann, The linkbetween springtime total ozone and summer UV radiationin Northern Hemisphere extratropics, J. Geophys. Res.,[Atmos.], 2013, 118, 8649–8661.
193 J. Lee-Taylor, S. Madronich, C. Fischer and B. Mayer, A Cli-matology of UV Radiation, 1979–2000, 65S–65N, in UVRadiation in Global Climate Change, ed. W. Gao, J. Slusserand D. Schmoldt, Springer, Berlin, Heidelberg, 2010,pp. 1–20.
194 J. R. Herman, Use of an improved radiation amplificationfactor to estimate the effect of total ozone changes onaction spectrum weighted irradiances and an instrumentresponse function, J. Geophys. Res., [Atmos.], 2010, 115,D23119.
195 R. L. McKenzie, P. J. Aucamp, A. F. Bais, L. O. Björn,M. Ilyas and S. Madronich, Ozone depletion and climatechange: Impacts on UV radiation, Photochem. Photobiol.Sci., 2011, 10, 182–198.
196 I. Ialongo, A. Arola, J. Kujanpää and J. Tamminen, Use ofsatellite erythemal UV products in analysing the global UVchanges, Atmos. Chem. Phys., 2011, 11, 9649–9658.
197 A. Tanskanen, A. Lindfors, A. Maatta, N. Krotkov,J. Herman, J. Kaurola, T. Koskela, K. Lakkala, V. Fioletov,G. Bernhard, R. McKenzie, Y. Kondo, M. O’Neill,H. Slaper, P. den Outer, A. F. Bais and J. Tamminen, Vali-dation of daily erythemal doses from ozone monitoringinstrument with ground-based UV measurement data,J. Geophys. Res., [Atmos.], 2007, 112, D24S44.
198 G. J. Labow, J. R. Herman, L.-K. Huang, S. A. Lloyd,M. T. DeLand, W. Qin, J. Mao and D. E. Larko, Diurnalvariation of 340 nm Lambertian equivalent reflectivity dueto clouds and aerosols over land and oceans, J. Geophys.Res., [Atmos.], 2011, 116, D11202.
199 J. R. Herman, M. T. DeLand, L. K. Huang, G. Labow,D. Larko, S. A. Lloyd, J. Mao, W. Qin and C. Weaver, A netdecrease in the Earth’s cloud, aerosol, and surface340 nm reflectivity during the past 33 year (1979–2011),Atmos. Chem. Phys., 2013, 13, 8505–8524.
200 C. S. Zerefos, K. Tourpali, K. Eleftheratos, S. Kazadzis,C. Meleti, U. Feister, T. Koskela and A. Heikkilä, Evidenceof a possible turning point in solar UV-B over Canada,Europe and Japan, Atmos. Chem. Phys., 2012, 12, 2469–2477.
201 A. M. Vogelmann, T. P. Ackerman and R. P. Turco,Enhancements in biologically effective ultraviolet radi-ation following volcanic eruptions, Nature, 1992, 359, 47.
202 G. Bernhard, C. R. Booth, J. C. Ehramjian, R. Stone andE. G. Dutton, Ultraviolet and visible radiation at Barrow,Alaska: Climatology and influencing factors on the basisof version 2 National Science Foundation network data,J. Geophys. Res., [Atmos.], 2007, 112, D09101.
203 A. F. Bais, T. Drosoglou, C. Meleti, K. Tourpali andN. Kouremeti, Changes in surface shortwave solar
Perspective Photochemical & Photobiological Sciences
48 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

irradiance from 1993 to 2011 at Thessaloniki (Greece),Int. J. Climatol., 2013, 33, 2871–2876.
204 R. Philipona, K. Behrens and C. Ruckstuhl, How decliningaerosols and rising greenhouse gases forced rapidwarming in Europe since the 1980s, Geophys. Res. Lett.,2009, 36, L02806.
205 A. Sanchez-Lorenzo, J. Calbó and M. Wild, Global anddiffuse solar radiation in Spain: Building a homogeneousdataset and assessing their trends, Global Planet. Change,2013, 100, 343–352.
206 S. Nyeki, C. H. Halios, W. Baum, K. Eleftheriadis,H. Flentje, J. Gröbner, L. Vuilleumier and C. Wehrli,Ground-based aerosol optical depth trends at three high-altitude sites in Switzerland and southern Germany from1995 to 2010, J. Geophys. Res., [Atmos.], 2012, 117, D18202.
207 A. R. D. Smedley, J. S. Rimmer, D. Moore, R. Toumi andA. R. Webb, Total ozone and surface UV trends in theUnited Kingdom: 1979–2008, Int. J. Climatol., 2012, 32,338–346.
208 M. Fitzka, S. Simic and J. Hadzimustafic, Trends in spec-tral UV radiation from long-term measurements at HoherSonnblick, Austria, Theotet. Appl. Climatol., 2012, 110,585–593.
209 G. Bernhard, Trends of solar ultraviolet irradiance at Barrow,Alaska, and the effect of measurement uncertainties ontrend detection, Atmos. Chem. Phys., 2011, 11, 13029–13045.
210 K. Eleftheratos, S. Kazadzis, C. S. Zerefos, K. Tourpali,C. Meleti, D. Balis, I. Zyrichidou, K. Lakkala, U. Feister,T. Koskela, A. Heikkilä and J. M. Karhu, Ozone andspectroradiometric UV changes in the past 20 yearsover high latitudes, Atmos.–Ocean, 2014, DOI: 10.1080/07055900.2014.919897.
211 S. Watanabe, T. Takemura, K. Sudo, T. Yokohata andH. Kawase, Anthropogenic changes in the surface all-skyUV-B radiation through 1850–2005 simulated by an Earthsystem model, Atmos. Chem. Phys., 2012, 12, 5249–5257.
212 J. Junk, U. Feister, A. Helbig, K. Görgen, E. Rozanov,J. W. Krzyścin and L. Hoffmann, The benefit of modeledozone data for the reconstruction of a 99-yearUV radiationtime series, J. Geophys. Res., [Atmos.], 2012, 117, D16102.
213 L. Lemus-Deschamps and J. K. Makin, Fifty years ofchanges in UV Index and implications for skin cancer inAustralia, Int. J. Biometeorol., 2012, 56, 727–735.
214 J. Para, B. Charrière, A. Matsuoka, W. L. Miller,J. F. Rontani and R. Sempéré, UV/PAR radiation and DOMproperties in surface coastal waters of the Canadian shelfof the Beaufort Sea during summer 2009, Biogeosciences.,2013, 10, 2761–2774.
215 R. G. Zepp, G. C. Shank, E. Stabenau, K. W. Patterson,M. Cyterski, W. Fisher, E. Bartels and S. L. Anderson,Spatial and temporal variability of solar ultravioletexposure of coral assemblages in the Florida Keys: Impor-tance of colored dissolved organic matter, Limnol. Ocea-nogr., 2008, 53, 1909–1922.
216 B. B. Barnes, C. Hu, J. P. Cannizzaro, S. E. Craig,P. Hallock, D. L. Jones, J. C. Lehrter, N. Melo,
B. A. Schaeffer and R. Zepp, Estimation of diffuse attenu-ation of ultraviolet light in optically shallow Florida Keyswaters from MODIS measurements, Rem. Sens. Environ.,2014, 140, 519–532.
217 M. Tedetti, R. Sempéré, A. Vasilkov, B. Charrière,D. Nérini, W. L. Miller, K. Kawamura and P. Raimbault,High penetration of ultraviolet radiation in the south eastPacific waters, Geophys. Res. Lett., 2007, 34, L12610.
218 M. P. Gall, R. J. Davies-Colley and R. A. Merrilees, Excep-tional visual clarity and optical purity in a sub-alpine lake,Limnol. Oceanogr., 2013, 58, 443–451.
219 J. A. L. Gareis, L. F. W. Lesack and M. L. Bothwell, Attenu-ation of in situ UV radiation in Mackenzie Delta lakeswith varying dissolved organic matter compositions,Water Resou. Res., 2010, 46, W09516.
220 D. K. Perovich, The interaction of ultraviolet light withArctic sea ice during SHEBA, Ann. Glaciol., 2006, 44,47–52.
221 I. Fountoulakis, A. F. Bais, K. Tourpali, K. Fragkos andS. Misios, Projected changes in solar UV radiation inthe Arctic and sub-Arctic Ocean: Effects from changesin reflectivity, ice transmittance, clouds, and ozone,J. Geophys. Res., [Atmos.], 2014, 119, 8073–8090.
222 A. F. Bais, K. Tourpali, A. Kazantzidis, H. Akiyoshi,S. Bekki, P. Braesicke, M. P. Chipperfield, M. Dameris,V. Eyring, H. Garny, D. Iachetti, P. Jöckel, A. Kubin,U. Langematz, E. Mancini, M. Michou, O. Morgenstern,T. Nakamura, P. A. Newman, G. Pitari, D. A. Plummer,E. Rozanov, T. G. Shepherd, K. Shibata, W. Tian andY. Yamashita, Projections of UV radiation changes in the21st century: impact of ozone recovery and cloud effects,Atmos. Chem. Phys., 2011, 11, 7533–7545.
223 T. Egorova, E. Rozanov, J. Gröbner, M. Hauser andW. Schmutz, Montreal Protocol Benefits simulatedwith CCM SOCOL, Atmos. Chem. Phys., 2013, 13, 3811–3823.
224 M. d. P. Correa, S. Godin-Beekmann, M. Haeffelin,S. Bekki, P. Saiag, J. Badosa, F. Jegou, A. Pazmino andE. Mahe, Projected changes in clear-sky erythemal andvitamin D effective UV doses for Europe over the period2006 to 2100, Photochem. Photobiol. Sci., 2013, 12, 1053–1064.
225 K. E. Taylor, R. J. Stouffer and G. A. Meehl, An overview ofCMIP5 and the experiment design, Bull. Am. Meteorol.Soc., 2011, 93, 485–498.
226 V. Eyring, J. M. Arblaster, I. Cionni, J. Sedláček, J. Perlwitz,P. J. Young, S. Bekki, D. Bergmann, P. Cameron-Smith,W. J. Collins, G. Faluvegi, K. D. Gottschaldt,L. W. Horowitz, D. E. Kinnison, J. F. Lamarque,D. R. Marsh, D. Saint-Martin, D. T. Shindell, K. Sudo,S. Szopa and S. Watanabe, Long-term ozone changes andassociated climate impacts in CMIP5 simulations,J. Geophys. Res., [Atmos.], 2013, 118, 5029–5060.
227 P. R. Gent, G. Danabasoglu, L. J. Donner, M. M. Holland,E. C. Hunke, S. R. Jayne, D. M. Lawrence, R. B. Neale,P. J. Rasch, M. Vertenstein, P. H. Worley, Z.-L. Yang and
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 49

M. Zhang, The Community Climate System Model Version4, J. Clim., 2011, 24, 4973–4991.
228 A. Hilboll, A. Richter and J. P. Burrows, Long-termchanges of tropospheric NO2 over megacities derived frommultiple satellite instruments, Atmos. Chem. Phys., 2013,13, 4145–4169.
229 J.-F. Lamarque, G. P. Kyle, M. Meinshausen, K. Riahi,S. J. Smith, D. P. van Vuuren, A. J. Conley and F. Vitt,Global and regional evolution of short-lived radiatively-active gases and aerosols in the Representative Concen-tration Pathways, Clim. Change, 2011, 109, 191–212.
230 Y. W. Wang and Y. H. Yang, China’s dimming and bright-ening: evidence, causes and hydrological implications,Ann. Geophys., 2014, 32, 41–55.
231 D. A. Randall, Beyond deadlock, Geophys. Res. Lett., 2013,40, 5970–5976.
232 V. Aquila, L. D. Oman, R. Stolarski, A. R. Douglass andP. A. Newman, The response of ozone and nitrogendioxide to the eruption of Mt. Pinatubo at southern andnorthern midlatitudes, J. Atmos. Sci., 2012, 70, 894–900.
233 R. R. Garcia, D. E. Kinnison and D. R. Marsh, “Worldavoided” simulations with the Whole Atmosphere Com-munity Climate Model, J. Geophys. Res., [Atmos.], 2012,117, D23303.
234 P. A. Newman and R. L. McKenzie, UV impacts avoided bythe Montreal Protocol, Photochem. Photobiol. Sci., 2011,10, 1152–1160.
235 P. A. Newman, L. D. Oman, A. R. Douglass, E. L. Fleming,S. M. Frith, M. M. Hurwitz, S. R. Kawa, C. H. Jackman,N. A. Krotkov, E. R. Nash, J. E. Nielsen, S. Pawson,R. S. Stolarski and G. J. M. Velders, What would have hap-pened to the ozone layer if chlorofluorocarbons (CFCs)had not been regulated?, Atmos. Chem. Phys., 2009, 9,2113–2128.
236 A. van Dijk, H. Slaper, P. N. d. Outer, O. Morgenstern,P. Braesicke, J. A. Pyle, H. Garny, A. Stenke, M. Dameris,A. Kazantzidis, K. Tourpali and A. F. Bais, Skin cancerrisks avoided by the Montreal Protocol – worldwide mod-elling integrating coupled climate-chemistry models witha risk model for UV, Photochem. Photobiol. Sci., 2013, 89,234–246.
237 R. M. Lucas, M. Norval, R. E. Neale, A. R. Young,F. R. d. Gruijl, Y. Takizawa and J. C. v. d. Leun, The conse-quences for human health of stratospheric ozonedepletion in association with other environmental factors,Photochem. Photobiol. Sci., 2015, 14, DOI: 10.1039/c4pp90033b, this issue.
238 A. W. Schmalwieser, S. Wallisch and B. Diffey, A library ofaction spectra for erythema and pigmentation, Photochem.Photobiol. Sci., 2012, 11, 251–268.
239 M. I. Micheletti, R. D. Piacentini and S. Madronich, Sensi-tivity of biologically active UV radiation to stratosphericozone changes: Effects of action spectrum shape andwavelength range, Photochem. Photobiol., 2003, 78, 456–461.
240 M. Norval, L. O. Björn and F. R. d. Gruijl, Is the actionspectrum for the UV-induced production of previtamin
D3 in human skin correct?, Photochem. Photobiol. Sci.,2009, 9, 11–17.
241 R. Bouillon, J. Eisman, M. Garabedian, M. Holick,J. Kleinschmidt, T. Suda, I. Terenetskaya and A. Webb,Action Spectrum for the Production of Previtamin D3 inHuman Skin, CIE Report No 174, Vol. 174:2006, Commis-sion Internationale de L’Eclairage, Vienna, 2006.
242 CIE, Erythema Reference Action Spectrum and StandardErythema Dose, CIE DS 007.1/E-1998, Commission Interna-tionale de l’Eclairage, Vienna, Austria, 1998.
243 A. F. McKinlay and B. L. Diffey, A Reference Action Spec-trum for Ultraviolet Induced Erythema in Human Skin, inHuman Exposure to Ultraviolet Radiation: Risks and Regu-lations, ed. W. F. Passchier and B. F. M. Bosnajakovic,Elsevier, Amsterdam, 1987, pp. 83–87.
244 A. R. Webb, H. Slaper, P. Koepke and A. W. Schmalwieser,Know your standard: Clarifying the CIE erythema actionspectrum, Photochem. Photobiol., 2011, 87, 483–486.
245 W. Olds, Elucidating the Links Between UV Radiation andVitamin D Synthesis: Using an In Vitro Model, PhD Thesis,Queensland University of Technology, Brisbane, 2010.
246 D. Bolsee, A. R. Webb, D. Gillotay, B. Dorschel,P. Knuschke, A. Krins and I. Terenetskaya, Laboratoryfacilities and recommendations for the characterization ofbiological ultraviolet dosimeters, Appl. Opt., 2000, 39,2813–2822.
247 V. Vitale, B. Petkov, F. Goutail, C. Lanconelli, A. Lupi,M. Mazzola, M. Busetto, A. Pazmino, R. Schioppo,L. Genoni and C. Tomasi, Variations of UV irradiance atAntarctic station Concordia during the springs of 2008and 2009, Antarct. Sci., 2011, 23, 389–398.
248 B. Petkov, V. Vitale, J. Gröbner, G. Hülsen, S. De Simone,V. Gallo, C. Tomasi, M. Busetto, V. L. Barth, C. Lanconelliand M. Mazzola, Short-term variations in surface UV-Birradiance and total ozone column at Ny-Ålesund duringthe QAARC campaign, Atmos. Res., 2012, 108, 9–18.
249 CIE, A Reference Action Spectrum for Ultraviolet InducedErythemal in Human Skin, Commission Internationale del’Eclairage, Vienna, Austria, 1987, vol. 6(1).
250 G. Seckmeyer, A. Bais, G. Bernhard, M. Blumthaler,C. R. Booth, K. Lantz and R. L. McKenzie, Instruments tomeasure solar ultraviolet irradiance. Part 2: Broadbandinstruments measuring erythemally weighted solar irradi-ance, World Meteorological Organisation, Global Atmos-pheric Watch, Report No. 164, Geneva, 2005.
251 E. T. Butson, T. Cheung, P. K. N. Yu and M. J. Butson,Measuring solar UV radiation with EBT radiochromicfilm, Phys. Med. Biol., 2010, 55, N487–N493.
252 A. M. Siani, G. R. Casale, R. Sisto, A. Colosimo, C. A. Langand M. G. Kimlin, Occupational Exposures to Solar Ultra-violet Radiation of Vineyard Workers in Tuscany (Italy),Photochem. Photobiol., 2011, 87, 925–934.
253 J. Turner and A. V. Parisi, Influence of reflected UV irradi-ance on occupational exposure from combinations ofreflective wall surfaces, Photochem. Photobiol. Sci., 2013,12, 1589–1595.
Perspective Photochemical & Photobiological Sciences
50 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

254 M.-A. Serrano, J. Canada and J. C. Moreno, Erythemalultraviolet exposure of cyclists in Valencia, Spain, Photo-chem. Photobiol., 2010, 86, 716–721.
255 M.-A. Serrano, J. Canada, J. C. Moreno and Membersof the Research Group of Solar Radiation of Valencia,Erythemal ultraviolet solar radiation doses received byyoung skiers, Photochem. Photobiol. Sci., 2013, 12, 1976–1983.
256 A. W. Schmalwieser, A. Cabaj, G. Schauberger, H. Rohn,B. Maier and H. Maier, Facial solar UV exposure of Aus-trian farmers during occupation, Photochem. Photobiol.,2010, 86, 1404–1413.
257 A. W. Schmalwieser, C. Enzi, S. Wallisch, F. Holawe,B. Maier and P. Weihs, UV exposition during typical life-style behavior in an urban environment, Photochem.Photobiol., 2010, 86, 711–715.
258 P. Weihs, A. Schmalwieser, C. Reinisch, E. Meraner,S. Walisch and M. Harald, Measurements of personal UVexposure on different parts of the body during variousactivities, Photochem. Photobiol., 2013, 89, 1004–1007.
259 A. M. Siani, G. R. Casale, S. Modesti, A. V. Parisi andA. Colosimo, Investigation on the capability of polysul-phone for measuring biologically effective solar UVexposures, Photochem. Photobiol. Sci., 2014, 13, 521–530.
260 G. R. Casale, A. M. Siani, H. Diémoz, M. G. Kimlin andA. Colosimo, Applicability of the Polysulphone HorizontalCalibration to Differently Inclined Dosimeters, Photochem.Photobiol., 2012, 88, 207–214.
261 R. L. McKenzie, B. Liley, P. Johnston, R. Scragg, A. Stewart,A. I. Reeder and M. W. Allen, Small doses from artificialUV sources elucidate the photo-production of vitamin D,Photochem. Photobiol. Sci., 2013, 12, 1726–1737.
262 G. Seckmeyer, M. Klingebiel, S. Riechelmann, I. Lohse,R. L. McKenzie, J. Ben Liley, M. W. Allen, A.-M. Siani andG. R. Casale, A critical assessment of two types of per-sonal UV dosimeters, Photochem. Photobiol., 2012, 88,215–222.
263 P. Datta, M. K. Bogh, P. Olsen, P. Eriksen, A. V. Schmedes,M. M.-L. Grage, P. A. Philipsen and H. C. Wulf, Increasein serum 25-hydroxyvitamin-D3 in humans after solarexposure under natural conditions compared to artificialUVB exposure of hands and face, Photochem. Photobiol.Sci., 2012, 11, 1817–1824.
264 M. d. P. Correa, S. Godin-Beekmann, M. Haeffelin,C. Brogniez, F. Verschaeve, P. Saiag, A. Pazmino andE. Mahe, Comparison between UV index measurementsperformed by research-grade and consumer-productsinstruments, Photochem. Photobiol. Sci., 2010, 9, 459–463.
265 D.-P. Häder, C. E. Williamson, S.-Å. Wängberg, M. Rautio,K. C. Rose, K. Gao, E. W. Helbling, R. P. Sinha andR. Worrest, Effects of UV radiation on aquatic ecosystems andinteractions with other environmental factors, Photochem.Photobiol. Sci., 2015, 14, DOI: 10.1039/c4pp90035a, this issue.
266 A. L. Andrady, H. H. Redhwi, AyakoTorikai, K. K. Pandeyand P. Gies, Consequences of stratospheric ozonedepletion and climate change on the use of materials,
Photochem. Photobiol. Sci., 2015, 14, DOI: 10.1039/c4pp90038c, this issue.
267 R. L. McKenzie, R. Scragg, B. Liley, P. Johnston,J. Wishart, A. Stewart and R. Prematunga, Serum 25-hydroxyvitamin-D responses to multiple UV exposuresfrom solaria: inferences for exposure to sunlight, Photo-chem. Photobiol. Sci., 2012, 11, 1174–1185.
268 A. R. Webb, R. Kift, J. L. Berry and L. E. Rhodes, Thevitamin D debate: Translating controlled experiments intoreality for human sun exposure times, Photochem. Photo-biol., 2011, 87, 741–745.
269 N. Chubarova and Y. Zhdanova, Ultraviolet resources overNorthern Eurasia, J. Photochem. Photobiol., B, 2013, 127,38–51.
270 S. J. Pope and D. E. Godar, Solar UV Geometric Conver-sion Factors: Horizontal Plane to Cylinder Model dagger,Photochem. Photobiol., 2010, 86, 457–466.
271 D. J. Turnbull and A. V. Parisi, Latitudinal variations overAustralia of the solar UV-radiation exposures for vitaminD(3) in shade compared to full sun, Radiat. Res., 2010,173, 373–379.
272 CIE, Rationalizing Nomenclature for UV Doses and Effectson Humans, CIE 209:2014/WMO/GAW report No. 211,Commission Internationale de l’Eclairage, Vienna,Austria, 2014.
273 P. J. Crutzen, Albedo enhancement by stratospheric sulfurinjections: A contribution to resolve a policy dilemma?,Clim. Change, 2006, 77, 211–220.
274 P. Heckendorn, D. Weisenstein, S. Fueglistaler, B. P. Luo,E. Rozanov, M. Schraner, L. W. Thomason and T. Peter,The impact of geoengineering aerosols on stratospherictemperature and ozone, Environ. Res. Lett., 2009, 4, 045108.
275 S. Tilmes, D. E. Kinnison, R. R. Garcia, R. Salawitch,T. Canty, J. Lee-Taylor, S. Madronich and K. Chance,Impact of very short-lived halogens on stratospheric ozoneabundance and UV radiation in a geo-engineered atmos-phere, Atmos. Chem. Phys., 2012, 12, 10945–10955.
276 F. D. Pope, P. Braesicke, R. G. Grainger, M. Kalberer,I. M. Watson, P. J. Davidson and R. A. Cox, Stratosphericaerosol particles and solar-radiation management, Nat.Clim. Change, 2012, 2, 713–719.
277 G. Pitari, V. Aquila, B. Kravitz, A. Robock, S. Watanabe,I. Cionni, N. D. Luca, G. D. Genova, E. Mancini andS. Tilmes, Stratospheric ozone response to sulfate geo-engineering: Results from the Geoengineering ModelIntercomparison Project (GeoMIP), J. Geophys. Res., [Atmos.],2014, 119, 2629–2653.
278 M. Kuebbeler, U. Lohmann and J. Feichter, Effects ofstratospheric sulfate aerosol geo-engineering on cirrusclouds, Geophys. Res. Lett., 2012, 39, L23803.
279 A. J. Ferraro, E. J. Highwood and A. J. Charlton-Perez,Weakened tropical circulation and reduced precipitationin response to geoengineering, Environ. Res. Lett., 2014, 9,DOI: 10.1088/1748-9326/9/1/014001.
280 N. A. Krotkov, P. K. Bhartia, J. R. Herman, J. R. Slusser, G.R. Scott, G. J. Labow, A. P. Vasilkov, T. Eck, O. Doubovik
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 19–52 | 51

and B. N. Holben, Aerosol ultraviolet absorption exper-iment (2002 to 2004), part 2: absorption optical thickness,refractive index, and single scattering albedo, Opt. Eng.,2005, 44, 1–17.
281 S. Kazadzis, V. Amiridis and N. Kouremeti, The effect ofaerosol absorption in solar UV radiation, in Advances inMeteorology, Climatology and Atmospheric Physics, ed.
C. G. Helmis and P. T. Nastos, Springer, Berlin, Heidelberg,2013, pp. 1041–1047.
282 E. Nikitidou, A. Kazantzidis, V. De Bock and H. De Backer,The aerosol forcing efficiency in the UV region andthe estimation of single scattering albedo at a typicalWest European site, Atmos. Environ., 2013, 69, 313–320.
Perspective Photochemical & Photobiological Sciences
52 | Photochem. Photobiol. Sci., 2015, 14, 19–52 This journal is © The Royal Society of Chemistry and Owner Societies 2015

Photochemical &Photobiological Sciences
PERSPECTIVE
Cite this: Photochem. Photobiol. Sci.,2015, 14, 53
Received 20th October 2014,Accepted 20th October 2014
DOI: 10.1039/c4pp90033b
www.rsc.org/pps
The consequences for human health ofstratospheric ozone depletion in association withother environmental factors
R. M. Lucas,*a,b M. Norval,c R. E. Neale,d A. R. Young,e F. R. de Gruijl,f Y. Takizawag,h
and J. C. van der Leuni
Due to the implementation of the Montreal Protocol, which has limited, and is now probably reversing,
the depletion of the stratospheric ozone layer, only modest increases in solar UV-B radiation at the
surface of the Earth have occurred. For many fair-skinned populations, changing behaviour with regard to
exposure to the sun over the past half century – more time in the sun, less clothing cover (more skin
exposed), and preference for a tan – has probably contributed more to greater levels of exposure to UV-B
radiation than ozone depletion. Exposure to UV-B radiation has both adverse and beneficial effects on
human health. This report focuses on an assessment of the evidence regarding these outcomes that has
been published since our previous report in 2010. The skin and eyes are the organs exposed to solar UV
radiation. Excessive solar irradiation causes skin cancer, including cutaneous malignant melanoma and
the non-melanoma skin cancers, basal cell carcinoma and squamous cell carcinoma, and contributes to
the development of other rare skin cancers such as Merkel cell carcinoma. Although the incidence of
melanoma continues to increase in many countries, in some locations, primarily those with strong sun
protection programmes, incidence has stabilised or decreased over the past 5 years, particularly in
younger age-groups. However, the incidence of non-melanoma skin cancers is still increasing in most
locations. Exposure of the skin to the sun also induces systemic immune suppression that may have
adverse effects on health, such as through the reactivation of latent viral infections, but also beneficial
effects through suppression of autoimmune reactivity. Solar UV-B radiation damages the eyes, causing
cataracts and pterygium. UV-B irradiation of the skin is the main source of vitamin D in many geographic
locations. Vitamin D plays a critical role in the maintenance of calcium homeostasis in the body; severe
deficiency causes the bone diseases, rickets in children and osteomalacia in adults. Although many
studies have implicated vitamin D deficiency in a wide range of diseases, such as cancer and cardio-
vascular disease, more recent evidence is less compelling, with meta-analyses of supplementation trials
failing to show a beneficial effect on the health outcomes that have been tested. It continues to be
difficult to provide public health messages to guide safe exposure to the sun that are accurate, simple,
and can be used by people with different skin types, in different locations, and for different times of the
year or day. There is increasing interest in relating sun protection messages to the UV Index. Current sun
protection strategies are outlined and assessed. Climatic factors affect the amount of UV radiation
aNational Centre for Epidemiology and Population Health, The Australian National
University, Canberra 2601, Australia. E-mail: [email protected] Kids Institute, University of Western Australia, Perth 6008, AustraliacBiomedical Sciences, University of Edinburgh Medical School, Edinburgh EH8 9AG,
Scotland, UKdQIMR Berghofer Medical Research Institute, Brisbane 4029, AustraliaeKing’s College London (KCL), St John’s Institute of Dermatology, London SE1 9RT, UKfDepartment of Dermatology, Leiden University Medical Centre, P.O. Box 9600,
NL-2300 RC Leiden, The NetherlandsgAkita University Graduate School of Medicine, Akita-shi, Akita Prefecture, JapanhNational Institute for Minamata Diseases, Minamata-sh, Kumamoto Prefecture,
JapaniEcofys, Kanaalweg 16G, NL-3526 KL Utrecht, The Netherlands
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 53

received by the skin and eyes, separately from the effect of ozone depletion. For example, cloud cover
can decrease or increase the intensity of UV radiation at Earth’s surface and warmer temperatures and
changes in precipitation patterns may alter the amount of time people spend outdoors and their choice
of clothing. The combination of changes in climate and UV radiation may affect the number of patho-
genic microorganisms in surface waters, and could have an impact on food security through effects on
plant and aquatic systems. It remains difficult to quantify these effects and their possible importance for
human health.
Introduction
Stratospheric ozone limits the amount of biologically damag-ing UV radiation in the UV-B waveband (280–315 nm) thatreaches the Earth’s surface – a 1000-fold reduction in muta-genic UV radiation.1 Depletion of the ozone layer results ingreater potential exposure to UV-B radiation, while, forhumans, actual exposure also depends on their behaviour,such as time spent outdoors, use of shade, and wearing of sunprotective clothing. Global climate change is altering the recov-ery of the stratospheric ozone layer and, through effects oncloud cover, will modify the levels of UV radiation at Earth’ssurface (for detail, see Bais et al.2). Loss of snow cover willdecrease albedo in mountain regions, possibly reducing UVradiation incident on body surfaces. Changing climate mayalso alter human behaviour with regard to exposure to thesun. Warmer temperatures may accelerate the genesis of skincancer and vitamin D production. In the following sections, weassess the health risks associated with ozone depletion,focussing on effects related to UV radiation in the UV-B wave-lengths. In addition, the risks and benefits of changing per-sonal exposure to UV radiation under the combined effects ofozone depletion and global climate change are considered.
Effects of solar UV radiation on theskin
Human skin comprises an outer thin epidermis of about 10cell layers and typically <100 μm deep and an inner layer, thedermis, that consists mainly of connective tissue and givesskin its mechanical properties. Epidermal cells are mostlykeratinocytes, with melanocytes in the basal layer producingmelanin that determines pigmentation of the skin. UV radi-ation is absorbed in the skin by specific molecules called chro-mophores, with the ensuing chemical changes initiatingmultiple biological processes. UV-A radiation (315–400 nm)penetrates the skin to a greater depth (into the dermis) thanUV-B radiation. Exposure to solar UV radiation may cause skincancer and photo-ageing, but also initiates the synthesis ofvitamin D, which is critical for human health.
Skin cancer
Solar UV radiation is the major environmental risk factor forboth melanoma and non-melanoma skin cancers (NMSCs).3
Cutaneous malignant melanoma (CMM) is the least common
of the skin cancers, but accounts for most deaths due to skincancer. NMSCs include basal cell carcinoma (BCC), squamouscell carcinoma (SCC), and other rarer skin cancers. BCC occursapproximately 3–4 times more frequently than SCC. It has thelowest mortality rate but can cause significant ill-health due toextensive local invasion, particularly when it arises on the face.
It is difficult to assess whether, or to what level, alterationsin ozone or climate have contributed to the rising incidence ofskin cancer globally. Over the 20th century, increasing travel tosunny locations and changed styles of clothing, at least insome populations, have led to higher personal exposure to thesun. This has probably been a more important driver of theincreasing incidence of skin cancer than changes in ozone orclimate.4 Changes in the diagnostic criteria for skin cancers,including the use of dermoscopy or epiluminescentmicroscopy and molecular classification,5 a lack of accuraterecording of lesions, and a more general growing awareness ofskin cancer and conservatism (erring on the safe side) in diag-noses,6 may also have introduced temporal biases in someinstances.
Cutaneous malignant melanoma
Geographic variation and temporal trends in incidence andmortality. World-wide, it is estimated that there were around230 000 new cases of CMM and 55 000 deaths in 2012.7 Theincidence varies widely from country to country (Table 1), withthe highest age-standardised annual incidence rates (∼35 per100 000 population in 2012) in Australia and New Zealand.7
CMM is rare in darker skinned populations, such as in SouthAsia.8
The incidence of CMM in fair-skinned populations hasapproximately doubled every 10–20 years since the 1960s andthis trend is projected to continue for at least 20 more years9
(Table 1).The rise in incidence of CMM has been attributed to
changes in recreational behaviour leading to increasedexposure to the sun.12 The incidence rate has stabilised or isdecreasing in younger age groups (<44 years) in somecountries, such as Australia,9 but continues to increase else-where.13,14 Public health campaigns to encourage protectionfrom the sun, beginning in the 1980s, have probably contribu-ted to the decrease in incidence in younger age groups inAustralia.15
Although CMM accounts for only 4% of all skin cancercases, it is responsible for about 80% of deaths from skincancer.9 Mortality due to CMM is increasing in Southern and
Perspective Photochemical & Photobiological Sciences
54 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015
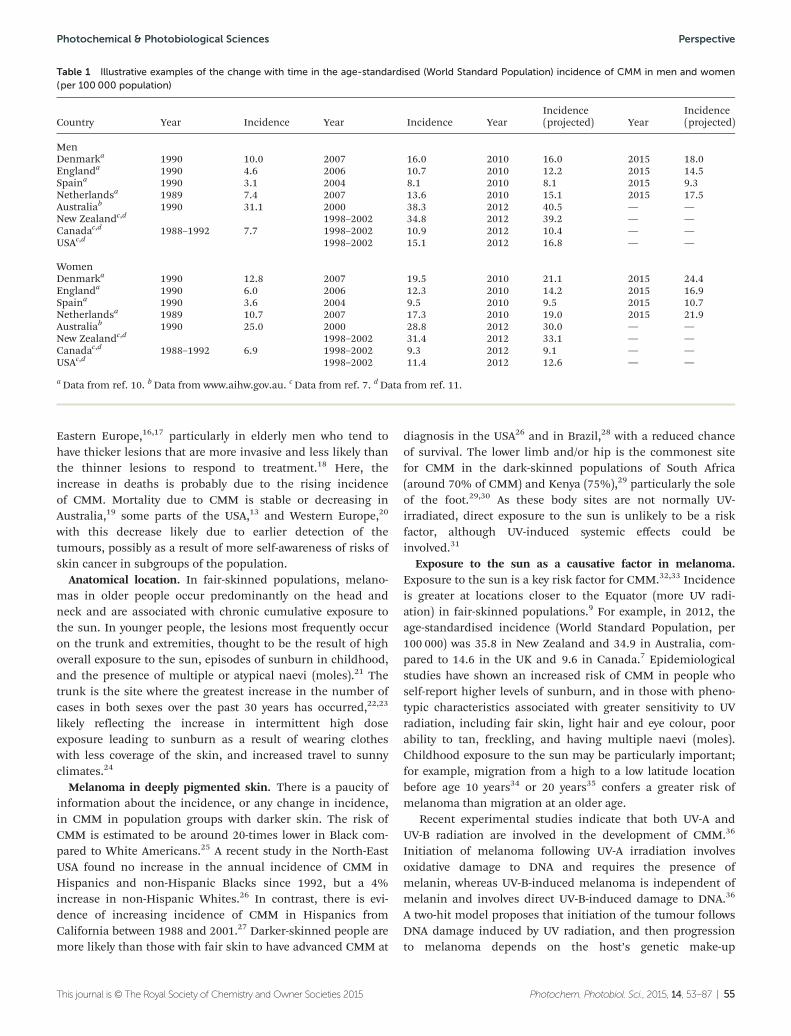
Eastern Europe,16,17 particularly in elderly men who tend tohave thicker lesions that are more invasive and less likely thanthe thinner lesions to respond to treatment.18 Here, theincrease in deaths is probably due to the rising incidenceof CMM. Mortality due to CMM is stable or decreasing inAustralia,19 some parts of the USA,13 and Western Europe,20
with this decrease likely due to earlier detection of thetumours, possibly as a result of more self-awareness of risks ofskin cancer in subgroups of the population.
Anatomical location. In fair-skinned populations, melano-mas in older people occur predominantly on the head andneck and are associated with chronic cumulative exposure tothe sun. In younger people, the lesions most frequently occuron the trunk and extremities, thought to be the result of highoverall exposure to the sun, episodes of sunburn in childhood,and the presence of multiple or atypical naevi (moles).21 Thetrunk is the site where the greatest increase in the number ofcases in both sexes over the past 30 years has occurred,22,23
likely reflecting the increase in intermittent high doseexposure leading to sunburn as a result of wearing clotheswith less coverage of the skin, and increased travel to sunnyclimates.24
Melanoma in deeply pigmented skin. There is a paucity ofinformation about the incidence, or any change in incidence,in CMM in population groups with darker skin. The risk ofCMM is estimated to be around 20-times lower in Black com-pared to White Americans.25 A recent study in the North-EastUSA found no increase in the annual incidence of CMM inHispanics and non-Hispanic Blacks since 1992, but a 4%increase in non-Hispanic Whites.26 In contrast, there is evi-dence of increasing incidence of CMM in Hispanics fromCalifornia between 1988 and 2001.27 Darker-skinned people aremore likely than those with fair skin to have advanced CMM at
diagnosis in the USA26 and in Brazil,28 with a reduced chanceof survival. The lower limb and/or hip is the commonest sitefor CMM in the dark-skinned populations of South Africa(around 70% of CMM) and Kenya (75%),29 particularly the soleof the foot.29,30 As these body sites are not normally UV-irradiated, direct exposure to the sun is unlikely to be a riskfactor, although UV-induced systemic effects could beinvolved.31
Exposure to the sun as a causative factor in melanoma.Exposure to the sun is a key risk factor for CMM.32,33 Incidenceis greater at locations closer to the Equator (more UV radi-ation) in fair-skinned populations.9 For example, in 2012, theage-standardised incidence (World Standard Population, per100 000) was 35.8 in New Zealand and 34.9 in Australia, com-pared to 14.6 in the UK and 9.6 in Canada.7 Epidemiologicalstudies have shown an increased risk of CMM in people whoself-report higher levels of sunburn, and in those with pheno-typic characteristics associated with greater sensitivity to UVradiation, including fair skin, light hair and eye colour, poorability to tan, freckling, and having multiple naevi (moles).Childhood exposure to the sun may be particularly important;for example, migration from a high to a low latitude locationbefore age 10 years34 or 20 years35 confers a greater risk ofmelanoma than migration at an older age.
Recent experimental studies indicate that both UV-A andUV-B radiation are involved in the development of CMM.36
Initiation of melanoma following UV-A irradiation involvesoxidative damage to DNA and requires the presence ofmelanin, whereas UV-B-induced melanoma is independent ofmelanin and involves direct UV-B-induced damage to DNA.36
A two-hit model proposes that initiation of the tumour followsDNA damage induced by UV radiation, and then progressionto melanoma depends on the host’s genetic make-up
Table 1 Illustrative examples of the change with time in the age-standardised (World Standard Population) incidence of CMM in men and women(per 100 000 population)
Country Year Incidence Year Incidence YearIncidence(projected) Year
Incidence(projected)
MenDenmarka 1990 10.0 2007 16.0 2010 16.0 2015 18.0Englanda 1990 4.6 2006 10.7 2010 12.2 2015 14.5Spaina 1990 3.1 2004 8.1 2010 8.1 2015 9.3Netherlandsa 1989 7.4 2007 13.6 2010 15.1 2015 17.5Australiab 1990 31.1 2000 38.3 2012 40.5 — —New Zealandc,d 1998–2002 34.8 2012 39.2 — —Canadac,d 1988–1992 7.7 1998–2002 10.9 2012 10.4 — —USAc,d 1998–2002 15.1 2012 16.8 — —
WomenDenmarka 1990 12.8 2007 19.5 2010 21.1 2015 24.4Englanda 1990 6.0 2006 12.3 2010 14.2 2015 16.9Spaina 1990 3.6 2004 9.5 2010 9.5 2015 10.7Netherlandsa 1989 10.7 2007 17.3 2010 19.0 2015 21.9Australiab 1990 25.0 2000 28.8 2012 30.0 — —New Zealandc,d 1998–2002 31.4 2012 33.1 — —Canadac,d 1988–1992 6.9 1998–2002 9.3 2012 9.1 — —USAc,d 1998–2002 11.4 2012 12.6 — —
aData from ref. 10. bData from www.aihw.gov.au. cData from ref. 7. dData from ref. 11.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 55

(particularly for melanoma of the trunk) and/or tumour pro-motion by ongoing exposure to both UV-A and UV-Bradiation.37
There is growing interest in the effects of exposures in earlylife on the risks of diseases that develop later in life. One indi-cation that exposure early in life is important for melanomacomes from observations that, in predominantly fair-skinnedpopulations, people with the disease are more likely to havebeen born in particular months of the year, compared with thegeneral population. In two studies, young women (aged 15–24years) with CMM from northern England38 and Sweden39 weresignificantly more likely to have been born in March (earlySpring). The data are consistent with the two-hit modelsuggested in animal studies37 – early exposure to UV radiationduring the months soon after birth initiates the first event,then later exposure causes progression to CMM. It is interest-ing to note that, in the past, exposure of infants to sunlightwas encouraged in some countries, a practice that has not con-tinued in more recent years.40–43
The higher incidence of small (<2 mm) melanomas duringsummer compared to winter in Northern Ireland (1984–2006)is consistent with exposure to UV radiation having a short-term promotional effect on melanocytes.44
Genetics of cutaneous melanoma. Susceptibility to CMM ispartially determined by genetic factors (reviewed in Eggermontet al.5 and Nikolaou & Stratigos12). A family history of CMMconfers a two-fold increase in risk.45 Alternatively, in approxi-mately 10% of people with CMM, there is a strong familyhistory,46 associated with specific mutations in genes involvedin control of the cell cycle (e.g., CDKN2A, CDK4). Increased riskin association with polymorphisms in other genes, includingthose associated with fair-skin phenotype (MC1R)46 andcharacteristic UV-induced cytosine to thymine (C → T)mutations in the tumour-control pathways,47 provide strongevidence of a causal role for UV radiation in the developmentof CMM. Very recently, further experimental corroboration forthe involvement of UV radiation in accelerated development ofmelanocytes has been obtained.48 Most strikingly, recentstudies show that melanomas (and cell lines thereof) havemore mutations in their genomes than most other tumourtypes, e.g., more than 30 000 point mutations per cell, andhundreds of mutations in protein-coding genes. Tumoursfrom sun-exposed skin have the greatest number of mutations,most of which are characteristic of those induced by exposureto UV radiation.49,50 It is difficult to distinguish causative fromincidental mutations, but special intron–exon comparativeanalyses provide evidence of likely causative mutations that arerelated to UV radiation.47
Non-melanoma skin cancer
Geographic variation and temporal trends in incidence.BCC and SCC are the most frequently occurring cancers infair-skinned populations. However, establishing accurate inci-dence data, and comparing incidence rates between countriesor regions, or over time, is challenging for several reasons.First, these cancers may be treated using destructive therapies
without prior biopsy, and such clinically-diagnosed lesions,particularly BCC, are often not included in estimates of inci-dence.51 Differences in therapeutic approaches betweencountries or changes over time can therefore have a consider-able influence on comparisons and trends. Secondly, there arevery few regions that require mandatory reporting of NMSCs tocancer registries and population-based studies are rare.Thirdly, most reports are person-based rather than lesion-based so they do not account for the multiple lesions oftenobserved in people living at lower latitudes; and lastly, variabi-lity in the population used to age-standardise incidence ratesmakes reported results difficult to compare.
Despite these difficulties, there is a strong associationbetween intensity of ambient UV radiation and incidence ofboth BCC and SCC.52 In a recent review, the highest annualincidence rates were in Australia (>1000 per 100 000 populationfor BCC) and the lowest in Africa (<1 per 100 000 population).53
Data for the latter are sparse and the low incidence masks rela-tively high incidence in some sub-populations, for exampleamong people with oculocutaneous albinism (OCA) [seesection below], and in Caucasians in South Africa, where BCCand SCC are typically among the top 5 or 10 cancers reported(depending on year).54
There have been substantial increases in the incidence ofNMSC over the past several decades. Across Europe, theannual incidence of BCC was estimated to be around 50 per100 000 persons in 1980.53 It has more than doubled sincethen in many parts of this region, and has quadrupled in theNetherlands.55 The incidence of SCC was approximately 10 per100 000 persons in Europe in 198053 with an increase toabout 25 per 100 000 by 2000.53 There are no estimates of SCCand BCC separately for the United States, but a study of astudy of data from national Medicare claims suggests that theage-adjusted rate of procedures for skin cancer increased by77% between 1992 and 2006.56
A recent quantitative review of data published between 1979and 2012 showed that, in fair-skinned populations worldwide,after adjustment for age, sex, and the levels of ambient UVradiation, the average annual increases in SCC and BCC inci-dence were 4% and 1%, respectively.52 The incidence of SCCincreased over time in both the older (≥60 years) and younger(<60 years) age groups, but only in the older age group forBCC.
Exposure to the sun as a risk factor for non-melanoma skincancer. As the studies showing latitudinal variation suggest,exposure to solar UV radiation is the primary cause of BCCand SCC; almost 40% of the variability in incidence in SCCand BCC in populations of predominantly European ancestrycould be explained by differences in the average daily levels ofambient UV radiation alone.52 Nevertheless, evidence suggeststhat the timing and patterns of exposure to the sun that giverise to the two tumour types are different. SCC appears to bestrongly associated with cumulative exposure to the sun. Infair-skinned people, SCC is rare on parts of the body that arenot routinely exposed to the sun57–59 and the presence ofactinic keratoses, which are a marker of cumulative exposure
Perspective Photochemical & Photobiological Sciences
56 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015
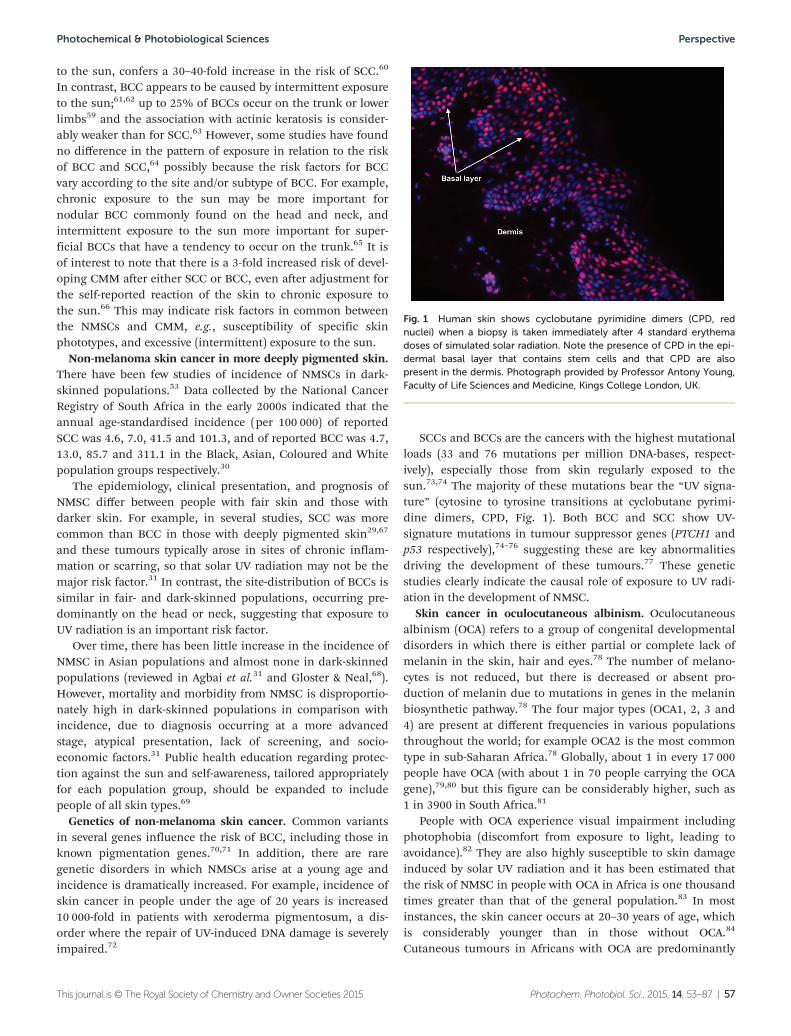
to the sun, confers a 30–40-fold increase in the risk of SCC.60
In contrast, BCC appears to be caused by intermittent exposureto the sun;61,62 up to 25% of BCCs occur on the trunk or lowerlimbs59 and the association with actinic keratosis is consider-ably weaker than for SCC.63 However, some studies have foundno difference in the pattern of exposure in relation to the riskof BCC and SCC,64 possibly because the risk factors for BCCvary according to the site and/or subtype of BCC. For example,chronic exposure to the sun may be more important fornodular BCC commonly found on the head and neck, andintermittent exposure to the sun more important for super-ficial BCCs that have a tendency to occur on the trunk.65 It isof interest to note that there is a 3-fold increased risk of devel-oping CMM after either SCC or BCC, even after adjustment forthe self-reported reaction of the skin to chronic exposure tothe sun.66 This may indicate risk factors in common betweenthe NMSCs and CMM, e.g., susceptibility of specific skinphototypes, and excessive (intermittent) exposure to the sun.
Non-melanoma skin cancer in more deeply pigmented skin.There have been few studies of incidence of NMSCs in dark-skinned populations.53 Data collected by the National CancerRegistry of South Africa in the early 2000s indicated that theannual age-standardised incidence (per 100 000) of reportedSCC was 4.6, 7.0, 41.5 and 101.3, and of reported BCC was 4.7,13.0, 85.7 and 311.1 in the Black, Asian, Coloured and Whitepopulation groups respectively.30
The epidemiology, clinical presentation, and prognosis ofNMSC differ between people with fair skin and those withdarker skin. For example, in several studies, SCC was morecommon than BCC in those with deeply pigmented skin29,67
and these tumours typically arose in sites of chronic inflam-mation or scarring, so that solar UV radiation may not be themajor risk factor.31 In contrast, the site-distribution of BCCs issimilar in fair- and dark-skinned populations, occurring pre-dominantly on the head or neck, suggesting that exposure toUV radiation is an important risk factor.
Over time, there has been little increase in the incidence ofNMSC in Asian populations and almost none in dark-skinnedpopulations (reviewed in Agbai et al.31 and Gloster & Neal,68).However, mortality and morbidity from NMSC is disproportio-nately high in dark-skinned populations in comparison withincidence, due to diagnosis occurring at a more advancedstage, atypical presentation, lack of screening, and socio-economic factors.31 Public health education regarding protec-tion against the sun and self-awareness, tailored appropriatelyfor each population group, should be expanded to includepeople of all skin types.69
Genetics of non-melanoma skin cancer. Common variantsin several genes influence the risk of BCC, including those inknown pigmentation genes.70,71 In addition, there are raregenetic disorders in which NMSCs arise at a young age andincidence is dramatically increased. For example, incidence ofskin cancer in people under the age of 20 years is increased10 000-fold in patients with xeroderma pigmentosum, a dis-order where the repair of UV-induced DNA damage is severelyimpaired.72
SCCs and BCCs are the cancers with the highest mutationalloads (33 and 76 mutations per million DNA-bases, respect-ively), especially those from skin regularly exposed to thesun.73,74 The majority of these mutations bear the “UV signa-ture” (cytosine to tyrosine transitions at cyclobutane pyrimi-dine dimers, CPD, Fig. 1). Both BCC and SCC show UV-signature mutations in tumour suppressor genes (PTCH1 andp53 respectively),74–76 suggesting these are key abnormalitiesdriving the development of these tumours.77 These geneticstudies clearly indicate the causal role of exposure to UV radi-ation in the development of NMSC.
Skin cancer in oculocutaneous albinism. Oculocutaneousalbinism (OCA) refers to a group of congenital developmentaldisorders in which there is either partial or complete lack ofmelanin in the skin, hair and eyes.78 The number of melano-cytes is not reduced, but there is decreased or absent pro-duction of melanin due to mutations in genes in the melaninbiosynthetic pathway.78 The four major types (OCA1, 2, 3 and4) are present at different frequencies in various populationsthroughout the world; for example OCA2 is the most commontype in sub-Saharan Africa.78 Globally, about 1 in every 17 000people have OCA (with about 1 in 70 people carrying the OCAgene),79,80 but this figure can be considerably higher, such as1 in 3900 in South Africa.81
People with OCA experience visual impairment includingphotophobia (discomfort from exposure to light, leading toavoidance).82 They are also highly susceptible to skin damageinduced by solar UV radiation and it has been estimated thatthe risk of NMSC in people with OCA in Africa is one thousandtimes greater than that of the general population.83 In mostinstances, the skin cancer occurs at 20–30 years of age, whichis considerably younger than in those without OCA.84
Cutaneous tumours in Africans with OCA are predominantly
Fig. 1 Human skin shows cyclobutane pyrimidine dimers (CPD, rednuclei) when a biopsy is taken immediately after 4 standard erythemadoses of simulated solar radiation. Note the presence of CPD in the epi-dermal basal layer that contains stem cells and that CPD are alsopresent in the dermis. Photograph provided by Professor Antony Young,Faculty of Life Sciences and Medicine, Kings College London, UK.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 57

SCCs, with BCCs less frequent, and CMMs only occasionallyseen,85,86 although the last may be under-reported as they arenormally amelanotic (non-pigmented).87 Most commonly, theskin cancers in Africans with OCA occur on the head and neckand tend to progress rapidly with metastasis to the cartilage,bone, and muscle, resulting in high mortality.84,88 The key roleof exposure to the sun in the oncogenic process is emphasisedby finding an increased frequency of skin tumours and lowerlife expectancy in people with OCA living in equatorial regionsof sub-Saharan Africa than in parts further from the Equator.84
Viruses and skin cancer
There are two instances (see following sections) where anassociation between certain viruses and skin cancer has beendemonstrated. In both cases, the tumours occur predomi-nantly on body sites most exposed to the sun, suggesting thatexposure to solar UV radiation is likely to play a crucial role inthe carcinogenic process.
Merkel cell carcinoma and polyomavirus. Merkel cell carci-noma (MCC) is a rare, aggressive, neuroendocrine skin cancerwith high rates of recurrence, metastasis, and mortality.89 Itmainly affects fair-skinned, elderly people (peak age around 75years) or those who are immunosuppressed, particularly if theylive in a sunny location.89 The tumours occur most frequentlyon sun-damaged skin on the head and neck.90 There is a posi-tive correlation between ambient levels of UV radiation andage-adjusted incidence of MCC across the USA.91 The inci-dence has risen in recent years, for example, from 0.15 cases to0.44 cases per 100 000 between 1986 and 2001 in the USA.92
This could be explained by increasing life expectancy, greaterexposure to the sun and the rising number of immunosup-pressed people in the general population as a result of anincreasing number and range of organ transplants93 and infec-tion with HIV.89
Merkel cell polyomavirus is present in around 80% ofMCCs. The viral DNA is integrated into the host DNA and isthought to cause cancer after genomic mutations that elimi-nate its ability to replicate but maintain its oncogenic function(reviewed in Arora et al.94). Exposure to UV radiation may playa role in the integration or mutagenic processes (for example,Demetriou et al.95 and Mogha et al.95,96) or in suppression ofthe immune response to the virus.97
Squamous cell carcinoma and papillomaviruses. Humanpapilloma viruses (HPVs) can infect the squamous epitheliumof the skin and may play a causative role in the development ofcutaneous SCC. There are many types of HPVs; phylogeneticanalysis describes 120 different types across 5 genera.98 Severalstudies have found that the presence in serum of antibodies tothe beta or gamma HPV types is associated with an increasedrisk of SCC.99–101 Furthermore, people with SCC have higherlevels of beta HPV DNA in hair follicles of the eyebrow (used asa marker of infection) than controls without SCC.102 The betaand gamma HPV types code for proteins that affect the normalcontrols of the cell cycle and may also subvert the normalimmune response. The mechanism by which various types ofHPV might influence the risk of cutaneous SCC is unclear, but
is most likely through potentiating the effects of exposure toUV radiation.103
Photoageing
Chronic exposure to solar UV radiation results in photoagedskin, which is wrinkled, leathery, shows loss of elasticity, andis often associated with the development of SCC. Photoageingresults from the UV-induced degradation of proteins such ascollagen and elastin in the extracellular matrix of the dermis.UV-A radiation may be primarily responsible for chronic photo-ageing, given its greater depth of penetration.104 In addition, arole for UV-B radiation is indicated. First, UV-B radiation candirectly degrade some proteins, such as fibrillin and fibronec-tin, involved in maintaining the structure of the dermis.105
Second, the action spectrum for the induction of matrix metal-loproteinase (an enzyme that degrades collagen) in humanskin is similar to that for erythema (reddening of the skin,inflammation), suggesting that this is primarily an effect ofexposure to UV-B radiation.106 While not a risk to healthper se, photoageing of the skin incurs considerable coststhrough the use of cosmetic and hydrating agents to improvethe appearance and feel of the skin.
Melasma
Melasma appears as dark, macular, pigmented patches on thebrow, cheek, upper lip and jaw, and is due to a localisedincrease in melanin production.107 It can result in profoundemotional and psychological stress, significantly reducingquality of life.108 Melasma is particularly common in adultwomen living in tropical areas of the world and occurs morefrequently in individuals with skin types of intermediate pig-mentation (i.e., types III–V) than in those with fair (skin types Iand II) or very dark skin (skin type VI).107,109 Its prevalence hasbeen estimated as 3.4% in the general population in Beirut,Lebanon, 10.1% in Cuzco, Peru,107 34% in adult women inBotucatu, Brazil,110 and 40% in adult women and 20% inadult men in South-East Asia.111
The precise pathophysiology of melasma is unclear, but isknown to be complex. There is a genetic predisposition andseveral environmental triggers, one of which is exposure tosunlight. For example, a report from Tunisia indicated that51% of patients recognised exposure to the sun as a triggeringfactor and 84% as an aggravating factor, with high lifetimeexposure to the sun increasing the risk of severe melasmathree-fold.112 A case-control study in Brazil found that patientswith melasma had a greater number of years of seaside orrural residence and greater exposure to the sun at work orduring leisure than the controls; there was a lack of associationwith sunburn, implying that cumulative exposure to the sunmay be more important than acute exposure.109 Solar UV radi-ation can induce proliferation and migration of melanocytes113
and the production of several cytokines that increase the pro-duction of melanin. In addition, inflammatory cells, especiallymast cells, which produce a variety of potent pro-inflammatorysubstances, are likely to play key roles.114
Perspective Photochemical & Photobiological Sciences
58 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015

Effects of solar UV radiation on the eye
The eye is partly protected from direct UV radiation by thebrow ridges. This means that reflected radiation is likely tocontribute more to the total UV radiation received than occursfor the skin. Higher intensity UV radiation induces the mostdamage, although less intense exposure over a long periodalso increases the risk of disease. Transmission of UV radi-ation through the eye (Fig. 2) generally decreases with increas-ing age, but there is wide inter-individual variability.115 Thecornea filters out wavelengths less than about 280 nm,although this is relevant for artificial sources only, as sunlightat the Earth’s surface is confined to wavelengths >290 nm.There is further absorption in the aqueous humour (Fig. 2).The lens of a young child (e.g., <5 years) transmits close to100% of the visible light spectrum (wavelength >400 nm), butabsorbs UV radiation, except for a small window at 320 nm.116
The lens of an older person (e.g., 60+ years) commonly filtersout even some of the short blue visible light (400–500 nm) sothat this, and UV radiation of shorter wavelengths, do notreach the retina.
Exposure to UV radiation increases the risk of a number ofocular conditions, with the strongest evidence of a specificeffect of UV-B radiation for photokeratitis, pterygium, andcataract.
Photokeratitis and photoconjunctivitis
Exposure of the eye to high-dose UV radiation from the suncan result in inflammation of the cornea (photokeratitis) and/or conjunctiva (photoconjunctivitis) (Fig. 2). The maximumsensitivity is to the UV-B wavelengths.117 The damage is prob-ably caused by oxidative stress,118–120 with the squamous cellsof the epithelium of the cornea, the keratocytes of the stromallayer of the cornea, and the endothelial cells lining the back ofthe cornea, being affected.
Transmission of UV radiation decreases across the corneafrom the centre to the periphery, due to scattering and absorp-tion.121 The centrally located endothelial cells receive a higherdose of UV radiation, and show evidence of higher oxidativestress than cells in the periphery. It is the damage to these
central corneal endothelial cells that particularly causescorneal swelling and temporary loss of vision in UV-inducedphotokeratitis.121
Pterygium
Pterygium, a wing-shaped invasive growth of the conjunctiva(Fig. 3), is common in adults living in environments with highUV radiation. For example, it affects at least one eye in approxi-mately 10% of: adults (≥15 years) on Norfolk Island,Australia;122 south Indians (≥40 years) in Chennai, India;123
and indigenous people (≥40 years) in Central Australia.124
Some recent studies show that the prevalence of pterygiumincreases with increasing age and is more common in menthan women,123 but the increase with age is not consistentlyfound.125 Key risk factors for pterygium are greater time out-doors (including sports with high ocular exposure to UV radi-ation, such as surfing126), rural residence, having a skin typethat tans122 and non-use of spectacles.123 Of note, the onlystudy that has examined the risk of pterygium in associationwith wavelengths of solar radiation other than those in the UVrange (using retrospectively reconstructed exposure behaviourduring working hours) found a stronger association withvisible light, and a weaker but still significant association withUV-B radiation.127 Recent evidence suggests that a pterygium
Fig. 2 A schematic drawing of a section through the human eye, with an enlarged schematic of the lens to the right.
Fig. 3 Pterygium of the eye; photograph provided by Dr David Mackey,University of Western Australia, Perth, Australia.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 59

is not always benign: there is histological evidence of inflam-matory and dysplastic changes in the epithelium and under-lying connective tissue,128 and of neoplasia in 2–10% ofexcised pterygia.129,130
Squamous cell carcinoma of the cornea and conjunctiva
Squamous cell carcinoma of the cornea and conjunctiva(SCCC) is rare; the annual incidence in the USA is estimated tobe 0.84 per 100 000 population.131 The incidence is higher inmen, and in association with older age, residence at lower lati-tude, infection with HIV, and high exposure to UV radiation.131
Infection with HPV and exposure to UV radiation may becommon risk factors for pterygium and SCCC.132
Cataract
In 2010, cataract was the leading cause of blindness world-wide.133 Of the three main types of age-related cataract – corti-cal, nuclear, and posterior subcapsular (Fig. 2) – UV radiationis primarily linked to an increased risk of cortical cataract.134
The action spectrum for acute/short term cataract develop-ment is relatively consistent across animal models, with UV-Bradiation of shorter wavelengths being most damaging.135
However, these models have limited relevance for cataractformation in humans because of wide inter-species variationin the dose of UV irradiation required,136 and the typical useof a single very high dose, rather than repeated lower doses.Melanin in pigmented irises may absorb UV radiation, leadingto fewer cataracts in those with more pigmented, thanless pigmented, irises.137 Lenses in older individuals may bemore susceptible to UV-induced cataracts due to poorerdefence against oxidative damage and decreased repairmechanisms.138–140
In the large Salisbury Eye Evaluation Study with partici-pants aged 65–84 years, the incidence and progression of corti-cal cataract (but not nuclear cataract) were associated withhigher levels of estimated exposure to UV-B radiation (calcu-lated based on an empirical model incorporating self-reportedtime outdoors and use of protection from the sun).141 In con-trast, in the 15 year follow-up of participants in the BeaverDam Eye Study,142 there was no association between exposureto the sun (as measured by residential history) and cumulativeincidence of any type of age-related cataract, after controllingfor age and sex. However, the combined use of sun-sensitisingmedications and high exposure to the sun led to a significantlyincreased risk of cortical cataract.142
Ocular malignancies
There is strong evidence to support exposure to UV radiationas a risk factor for tumours of the eyelid and weaker evidencefor ocular melanomas (reviewed in Yam and Kwok143). Over90% of the malignancies of the eyelid are BCCs, particularlyaffecting the lower lid (50–65%), but also the medial canthus(inner corner of the eye, 25–30%), upper eyelid (15%) andlateral canthus (outer corner of the eye, 5%).143 SCC accountsfor most of the remainder of the periocular cutaneoustumours.143
Melanomas of the eye can involve the surface (i.e., theeyelid or conjunctiva), or occur at an intra-ocular location,affecting the elements of the uvea (i.e., the iris, ciliary body,and choroid; Fig. 2). Uveal melanoma is the most commonprimary intraocular malignancy (>90%) and the leading causeof death from intraocular cancer.144,145 The reported annualincidence varies from 0.53 to 1.09 cases per 100 000 popu-lation, and is stable or decreasing.144,145 Uveal melanoma isprimarily a disease of white populations; light-coloured irises,blond hair, and fair skin are risk factors.146 There is mixed evi-dence implicating UV radiation as a risk factor for uveal mela-noma: latitudinal variation in incidence is not consistentlyfound;145 occupational exposure to UV radiation may have aprotective effect, but intermittent exposure may increase risk(reviewed in Mallet et al.146). People with the disease xero-derma pigmentosum, in which there is impaired ability torepair UV-induced DNA damage (see discussion above), have a58-fold increased risk of uveal melanoma.146,147 Geneticstudies show that the mutation patterns of the most frequentlymutated genes in CMM and uveal melanoma (i.e., BRAF vs.GNAQ, GNA11) are similar,146 and a mutation recently identi-fied in CMM (RAC1) is also found in 20% of uveal melanomacell lines.146 Given the strong evidence supporting a role forUV radiation as a cause of CMM, these studies also providesome evidence that exposure to UV radiation is a risk factor foruveal melanoma.
Age-related macular degeneration
Age-related macular degeneration (AMD) was the cause of 7%of blindness worldwide in 2010,133 and was the most frequentcause of blindness in older (50–75 years) white populations inEurope.148 UV radiation had been discounted previously as arisk factor in AMD as it does not reach the retina. However,recognition of the considerable individual variability in thetransmission of longer UV-B/shorter UV-A wavelengths(<320 nm) in older adults (60+ years) has led to reconsidera-tion of a potential role of UV radiation. Possible mechanismsinclude: UV-induced oxidative damage to mitochondrial DNA,particularly in the macular region of the neural retina and theretinal pigment epithelium;149 and/or upregulation of inflam-matory cytokines (e.g., IL-6) and transcription factors (e.g.,STAT3). Higher vitamin D status is associated with lower riskof AMD in women <75 years, but a higher risk in women≥75.150
In a systematic review and meta-analysis of case-control andcross-sectional studies, higher exposure to the sun was associ-ated with a 38% increase in the odds of having AMD.151
However, blue light (400–500 nm) may be more importantthan UV radiation as a risk factor for AMD.143,152,153
Other possible effects on the eye
There is a well-established association between spending lesstime outdoors and an increased risk of developing myopia inchildhood.154–157 While early hypotheses focused on theimportance of variation in focal length with a mix of indoor andoutdoor activities, more recent work suggests the importance
Perspective Photochemical & Photobiological Sciences
60 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015

of exposure to light, possibly through increased secretion ofdopamine in the retina, with effects on the growth of theeye.158 The wavelength dependence of this effect, and whetherthe pathways are mediated by vitamin D159 or not,157 are cur-rently unknown.
Evidence from animal studies suggests that UV irradiationof the eye can cause systemic immunosuppression,160,161 butthe relevance of these findings for human health is unclear atpresent.
Effects of solar UV radiation onimmune function and consequencesfor diseaseMechanisms
UV photons penetrate the epidermis and upper dermis162 andare absorbed by chromophores (Table 2), which then initiate acascade leading to changes in immune responses.
While much of this information has been gathered fromstudies in vitro or in rodent models, less is known abouthumans. However, an action spectrum for the UV-induced sup-pression of the human immune response to a previously-encountered antigen (termed memory or recall immuneresponses) has been constructed: it has two peaks, one withinthe UV-B waveband at 300 nm and one at 370 nm in the UV-Awaveband.164,165 There is also evidence from studies in bothhumans and mice that interactive and additive effects betweenwavebands can occur.166–168
Briefly, exposure to UV radiation causes up-regulation ofsome innate immune responses, and down-regulation of someacquired primary and memory immune responses, mainlythrough effects on T cell activity (reviewed in Gibbs &Norval,163 Schwarz & Schwarz,169 and Ullrich & Byrne170). Theup-regulation includes the production of several antimi-crobial peptides (AMPs) in the epidermis,171,172 possiblythrough a vitamin D pathway (see below). The AMPs provideimmediate protection against a variety of pathogens (bacteria,fungi, and viruses having a viral envelope) and they are alsoinvolved in the promotion of cell growth, healing, and angio-
genesis. In contrast to these stimulatory functions, exposure toUV radiation induces T regulatory cells (Tregs) and other celltypes which contribute to immunosuppression and help torestore cutaneous homeostasis.172,173 Mediators such as plate-let-activating factor, prostaglandin E2, histamine, and tumournecrosis factor-α are produced locally at the irradiated site.These alter the migration patterns and functions of variouspopulations of immune cells. The end result is the generationof cell subsets with suppressive activity which are thought toremain for the life-time of the individual.174,175
The UV-induced alterations in the normal immuneresponse can be beneficial for some human diseases and detri-mental for others. Vitamin D, synthesised following exposureof the skin to UV-B radiation, also has positive and negativeeffects on immune-related diseases. Indeed, it is difficult todistinguish between immunoregulation by vitamin D andother mediators induced by UV radiation,176–180 since thedownstream effects on immune parameters are similar. Forclarity, the effects of UV radiation and those of vitamin D havebeen assessed separately in the sections below. We first focuson the effects of UV radiation on immunity, and addressvitamin D-related effects on immune function in the sectionspecifically on vitamin D.
Polymorphic light eruption
Polymorphic light eruption (PLE) is the commonest of thephotodermatoses, with a prevalence of up to 20%.181 PLEmanifests as an intermittent itchy red skin eruption whichresolves without scarring after a few days to weeks. It occurs2–3 times more frequently in women than in men, with onsettypically in the first three decades of life,181 and is found pre-dominantly in those with fair skin, although all skin types canbe affected.181 A recent study of Indian patients with dark skinphototypes (IV and V) who suffered from various photoderma-toses revealed that PLE was the commonest of these, affecting60% of the group.182 The lesions occur most often in thespring and early summer or during a sunny holiday, followingthe first exposure to a large dose of sunlight. After repeatedexposures, the lesions are less likely to occur. This process,called photohardening, is used therapeutically with goodresults. Recent investigations indicate that key events in photo-
Table 2 Cutaneous chromophores involved in the initiation of UV-induced changes in immune function (reviewed in ref. 163)
Chromophore Change in structure following irradiation Effect on immune function
DNA Cyclobutane pyrimidine dimers and reactive oxygenspecies-induced base oxidation after exposure toradiation in both the UV-A and UV-B wavelengths
Oxidative stress; up-regulation of severalimmunosuppressive mediators and down-regulation ofsome immunostimulatory mediators
trans-Urocanic acid cis-Urocanic acid (peak effectiveness of isomerisationabout 300 nm)
Oxidative damage; up-regulation of severalimmunosuppressive mediators; stimulation ofneuropeptides; mast cell degranulation; cell growth arrest
Membranephospholipids
Oxidative stress and lipid peroxidation Clustering of receptors; activation of transcription factors;release of immune mediators
7-Dehydrocholesterol Previtamin D after UV-B irradiation, leading to vitamin D,then 25-hydroxyvitamin D and finally the active form,1,25-dihydroxyvitamin D
Up-regulation of some antimicrobial responses and DNArepair; down-regulation of most acquired immune responses
Tryptophan Activation of the arylhydrocarbon receptor followingexposure to UV-B radiation
Clustering and internalisation of growth factor receptors
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 61

hardening include a decrease in the number of Langerhanscells in the epidermis and recruitment of mast cells into thedermis,183 together with changes in systemic cytokine levels.184
PLE is immunologically-mediated as a result of a failure toestablish the normal suppression of immune responses follow-ing exposure to UV radiation. The antigen involved has notbeen identified but is likely to be novel, induced by the DNAdamaging properties of UV radiation. Various abnormalities inthe cutaneous immune response following UV radiation havebeen demonstrated in people with PLE compared with con-trols.185,186 This disease therefore illustrates the positive evo-lutionary advantage of UV-induced immunosuppression inindividuals who are not susceptible to PLE and what canhappen if it is absent.
Asthma
Asthma comprises a group of diseases that evidence as wheeze,chest tightness, or shortness of breath, occurring as a result ofobstruction of the airways and restriction of airflow that isusually reversible. The level of severity, frequency of symptoms,age of onset, main inflammatory phenotypes, and triggers andpathways are variable. This heterogeneity may explain thecurrent lack of consistency in results from studies examiningthe relationship between UV radiation and the risk of asthma.
There are anecdotal accounts that sunny holidays or livingat high altitude decrease asthma symptoms. The prevalence ofasthma was inversely associated with the intensity of UV radi-ation,187 or past personal exposure to solar UV radiation.188
However, in a study where different sub-types of asthma wereconsidered, residence at latitudes closer to the equator (andwith greater intensity of UV-B radiation) was associated withan increased risk of having asthma in atopic participants (witha history of allergic responses to specific antigens) but adecreased risk in those without atopy.189 These findings high-light the importance of differentiating between subtypes ofasthma in examining associations with exposure to UV radi-ation. Nevertheless, individual-level exposure to UV radiationwas not measured (only latitude and ambient UV radiation), sothe results could reflect exposure to other latitude-associatedfactors such as temperature and indoor heating.
Infection and vaccination
Studies over the past 20 years have shown that exposure tosolar UV radiation suppresses microbe-specific acquiredimmune responses in animal models of infection. This modu-lation can lead to an increased microbial load, reactivationfrom latency, and more severe symptoms, including death(reviewed in Norval et al.190). A recent study showed thatspending 8 or more hours outdoors per week when the UVIndex was ≥4 was associated with an increased risk of ocularrecurrence of herpes simplex virus (HSV) infection resulting ineruptive lesions.191 UV radiation prior to vaccination causes aless effective immune response in several mouse models(reviewed in Norval & Woods192), but whether exposure to UVradiation adversely affects the course of infections and theefficacy of vaccination in humans remains an open question.
Despite the paucity of new information, there remains thepossibility that UV-induced immunosuppression could convertan asymptomatic infection into a symptomatic one, reactivatea range of persistent infections, increase the oncogenic poten-tial of microbes, and reduce the memory immune response,for example after vaccination, so that it is no longer protective.
Autoimmune diseases
Many autoimmune diseases are considered to have bothenvironmental and genetic risk factors. Evidence to supportthe importance of environmental exposures comes from geo-graphical variation (changing incidence with changing lati-tude), temporal patterns (such as variations in incidence withseason or season-of-birth) and results from observational epi-demiological studies. Several studies show an inverse associ-ation between exposure to UV radiation and immune-mediateddiseases, suggesting that the UV may be protective. In manycases, the assumed pathway has been through enhanced syn-thesis of vitamin D (see section on Vitamin D below). However,this evidence is now being re-evaluated in light of possiblealternative pathways, including UV-induced immune modu-lation and altered susceptibility to relevant viral infections,and non-UV pathways such as changes in the secretion ofmelatonin (reviewed in Hart et al.193). While there have beensuggestions that exposure to UV radiation may be importantfor conditions such as inflammatory bowel disease (forexample, Nerich et al.194), type 1 diabetes,195 and rheumaticdiseases (including rheumatoid arthritis, systemic lupuserythematosus, dermatomyositis, and others),196 the strongestevidence is for multiple sclerosis.
Multiple sclerosis. Many studies (but not all) have shownthat the prevalence, incidence, or mortality from multiplesclerosis (MS) increases with increasing latitude and decreas-ing altitude or intensity of ambient UV radiation, in predomi-nantly fair-skinned populations (reviewed in Hewer et al.197).In the US Nurses Health Studies, a latitudinal gradient presentin a cohort of female nurses born before 1946 was not appar-ent in a similar cohort born after 1946.198 The findingsreflected an increase in incidence in the south in the latercohort (rather than a decrease in the north). One explanationgiven to explain this change was that increasing sun-protectivebehaviours in the south had reduced the difference in per-sonal dose of UV between the north and south.199 Studiesfrom the northern200 and southern201 hemispheres show that,compared to the general population, people with MS weremore likely to have been born in late spring and less likely tohave been born in late autumn. This timing would be consist-ent with a hypothesis that exposure of the mother to more UVradiation during the late first trimester, when the foetalnervous system is developing and maturing, is protective forthe development of MS in later life.201 Alternatively, it is alsopossible that exposures early in infancy, rather than in preg-nancy, influence risk, or other factors that vary seasonallycould be important. Animal studies suggest that UV-Birradiation can prevent the onset of experimental autoimmune
Perspective Photochemical & Photobiological Sciences
62 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015

encephalomyelitis, used as a model for MS,202 and there issupportive evidence from recent studies in humans.203,204
The role of UV-induced immune suppression in skin cancer
Cutaneous malignant melanoma. Evidence that theimmune response is important for the development of CMMis clearly shown by the increase in incidence following organtransplantation that requires ongoing treatment with immuno-suppressive medications.205 UV radiation, particularly UV-B,can cause suppression of many aspects of cell-mediated immu-nity but, until recently, how it influenced the initiation ofCMM was unknown. In a transgenic mouse model, the recruit-ment of macrophages to the skin following UV-B irradiationand their subsequent proliferation were shown to be critical inthe survival of melanocytes, including those with UV-inducedDNA damage.206–208 In addition, inflammation induced by UVradiation increased metastasis of melanoma, with neutrophilsbeing the main drivers of the inflammatory process.209 Con-sistent with these reports from animal models, in patientswith metastatic melanoma there was a shorter survival time ifmetastases contained a high proportion of macrophages.210
Non-melanoma skin cancer. Tumours induced by UV radi-ation are highly antigenic. UV-induced immune suppressionplays a critical role in the development of NMSC as evidencedby the dramatically increased incidence in immunosuppressedpeople, for example, following organ transplantation.211 Thisis especially shown for SCCs in organ transplant recipientsreceiving immunosuppressive drugs that suppress T cellactivity, suggesting that effector T cells are of particular impor-
tance in the control of SCC.212 Furthermore, Tregs induced byUV irradiation infiltrate SCCs and surround BCCs. Pharmaco-logically blocking steps in the pathway of UV-induced immu-nosuppression may be effective in preventing the developmentof skin cancers and actinic keratoses.212–214
UV-induced vitamin D and its effecton healthMetabolism of vitamin D
Vitamin D can be synthesised in the skin or ingested in thediet or as a supplement. The pathway by which vitamin D isproduced in the skin and metabolised to its active form, 1,25-dihydroxyvitamin D (1,25(OH)2D) is shown in Fig. 4. Synthesisis initiated by absorption of UV-B radiation by 7-dehydrocho-lesterol. The enzymatic steps converting vitamin D to 1,25(OH)2D occur predominantly in the liver and the kidney butalso in other tissues, including the skin.
Both pre-vitamin D3 and vitamin D3 can be converted toinactive photoproducts by continued UV-A or UV-B irradiation(discussed in Galkin & Terenetskaya215 and Norval et al.216).Once pre-vitamin D has formed and isomerization to vitaminD has occurred, there is preferential degradation of vitamin Dcompared with synthesis of pre-vitamin D at wavelengths ofUV radiation between 300–330 nm.217 This may explain whyvitamin D toxicity from exposure to solar UV radiation doesnot occur and may be of importance to public health messagesabout safe exposure to the sun – recurrent shorter periods of
Fig. 4 Synthesis of vitamin D and the vitamin D metabolic pathway. VDBP: vitamin D binding protein; 25(OH)D: 25-hydroxyvitamin D; 1,25(OH)2D:1,25-dihydroxyvitamin D; T1/2: half-life.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 63

exposure to the sun are preferable to prolonged exposure toachieve vitamin D production while minimizing UV-induceddamage of DNA.218
Vitamin D status – assessment and geographic variability
Vitamin D status is assessed by measuring the concentrationof the intermediate metabolite, 25-hydroxyvitamin D(25(OH)D) (Fig. 4) in serum. Vitamin D deficiency is reportedlywidespread globally, but it is important to note that there areconsiderable problems with the accuracy and reproducibilityof vitamin D assays219 as well as a lack of consensus on theconcentration of 25(OH)D that denotes deficient, insufficient,sufficient, or optimal vitamin D status.220 Common cut-offpoints are provided in Table 3.
There is a striking lack of data on the vitamin D status ofinfants, children, and adolescents and an almost completelack of data from Africa and South America.225–227
Variation in vitamin D status according to latitude is appar-ent when results arise from a single assay with good agree-ment across batches of samples,228 and is stronger whereblood is taken in winter compared to summer.229 Comparisonsacross studies may not show a latitudinal gradient due to theanalytical challenges and lack of standardisation of the seasonof blood collection.225–227 In addition, contributions tovitamin D status from dietary intake and sunny holidays mayobscure latitudinal gradients that would otherwise occur. Forexample, a lower latitude holiday in the previous year with thepurpose of sun-bathing was associated with higher 25(OH)Dlevels by 20–30 nmol L−1 in the following winter months inresidents of Uppsala, Sweden.230
Skin pigmentation and vitamin D status
Within a given location, people with darker skin commonlyhave lower concentrations of 25(OH)D than those with fairerskin231,232 and this is usually attributed to the photoprotectiveproperties of melanin. However, recent work highlights theimportance of cultural practices, or personal preferences,leading to avoidance of the sun.233 In a recent study, however,black Americans (n = 2085) had lower concentrations of 25(OH)D, but also lower concentrations of vitamin D bindingprotein, than white Americans.231 The consequence of this wassimilar (calculated) levels of “free” (not bound to vitamin Dbinding protein) or loosely bound (to albumin), i.e., bioavail-able, 25(OH)D. This observation may explain why black peoplewith low total 25(OH)D have higher bone mineral density thanwhite people with similar 25(OH)D concentrations. Further-
more, it may mean that vitamin D status should be definedby the concentration of bioavailable, rather than total,25(OH)D.234,235
Several recent experimental studies have examined theimpact of pigmentation of the skin on synthesis of vitaminD. Most of these,236,237 although not all,238 show that for aspecific dose of simulated solar UV irradiation there is agreater increase in the concentration of 25(OH)D in fairer-skinned than in darker-skinned participants. The lack of effectin the latter study has been attributed to the short wavelengthUV-B output from the source lamp, resulting in penetrationonly into the superficial layers of the epidermis above themain concentration of melanin.239
Exposure to the sun and vitamin D status
In general, laboratory studies show an initial linear dose–response between exposure to UV radiation and change in con-centration of 25(OH)D,240 but a plateau with continuingexposures over a longer period of time.218,241–243 Resultssuggest that the shape of the dose–response curve depends onthe baseline concentration of 25(OH)D, with a greater responseto UV irradiation238 and no plateau effect in those with a lowerstarting concentration (<50 nmol L−1),244 although conflictingresults have also been obtained.245
Understanding the relationship between the dose of UVradiation, the surface area of skin exposed and the productionof vitamin D is important for the development of public healthmessages. Results from population-based epidemiologicalstudies and experimental studies using artificial irradiationdemonstrate that exposing a larger area of skin to UVradiation results in a greater increase in the concentration of25(OH)D.246,247 Field studies have shown that increases in theconcentrations of 25(OH)D are positively associated with DNAdamage (assessed by concentration of CPDs in urine),suggesting that improved vitamin D status is always associatedwith some potentially mutagenic damage to DNA.248 However,production of vitamin D may be optimised, and skin DNAdamage minimised, by increasing the body surface areaexposed, and decreasing the UV-B-dose per unit area.
The World Health Organization’s INTERSUN programmerecommends sun protection when the UV Index is ≥3 andthere have been concerns that there may be little or no vitaminD production when the UV Index is <3. However, recent com-putational work suggests that vitamin D could be synthesisedat these lower levels of UV radiation, albeit more slowly.249
Webb and colleagues have shown that for the white-skinned
Table 3 Commonly used cut-off points for vitamin D status based on serum concentrations of 25(OH)D in nmol L−1
Author Year
Concentration of 25(OH)D in nmol L−1
Deficiency Insufficiency Sufficiency Optimal
Hollis221 2005 >80U.S. Institute of Medicine222 2010 <30 30–50 ≥50Pearce and Cheetham223 2010 <25 25–50 50–75 >75US Endocrine Society224 2011 <50 52.5–72.5 ≥75
Perspective Photochemical & Photobiological Sciences
64 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015

population of Manchester, UK, a normal lifestyle with relativelyshort, regular exposures to summer sunlight in northernmid-latitudes could increase vitamin D enough at the endof summer to maintain sufficiency levels (>50 nmol L−1 or20 ng ml−1) throughout the winter.218,250
Evidence of associations betweenvitamin D and human disease
The vital role of 1,25(OH)2D in maintaining the concentrationof calcium in the blood within a narrow range is well-estab-lished; vitamin D deficiency causes rickets in children andosteomalacia in adults. In recent years, many protective func-tions have been attributed to vitamin D. However, two recentsystematic reviews of observational and intervention studiescast doubt on the importance of vitamin D in decreasing therisk of many of these diseases.251,252
Immune function, infections, autoimmune diseases andcancer
Many of the cell types involved in immune function are able toconvert 25(OH)D to the active form, 1,25(OH)2D.
253 Theactions of 1,25(OH)2D are mediated through ligation with anuclear vitamin D receptor (VDR) that regulates gene transcrip-tion, or via rapid-response membrane receptors. The VDR isexpressed in many human cells, including those with immunefunctions. Polymorphisms in the VDR can affect the effective-ness of gene transcription, altering the action of the activehormone. Immunostimulatory and immunosuppressive path-ways are induced by 1,25(OH)2D (reviewed in Hewison254,255
and Christakos et al.255). Immunostimulatory effects includethe production of AMPs such as cathelicidin by macrophages,neutrophils, and epithelial cells, and the maturation of macro-phages. Immunosuppressive effects include the inhibition ofproinflammatory cytokines and the differentiation and matu-ration of dendritic cells and their ability to present antigens.Furthermore, 1,25(OH)2D can inhibit the differentiation andproliferation of B cells and their production of antibodies, andactivate Treg cells that have suppressor activity. These multipleeffects make it difficult to determine what role, if any, vitaminD has in protection against disorders of immunity. Studies ofinfectious diseases (using tuberculosis and respiratory viralinfections as examples), autoimmune diseases (using multiplesclerosis as an example), and the risk of cancer incidence andprogression are discussed below.
Vitamin D and tuberculosis. Tuberculosis (TB), a diseasecaused by infection with the bacterium Mycobacterium tubercu-losis, is a massive global health burden, with an estimated9 million new cases and 1.7 million deaths each year. Infectioncan lead to symptomatic active disease or, more commonly, toa latent infection which can reactivate later in a small pro-portion of cases. Recent studies suggest an associationbetween low vitamin D status (or lower ambient UV radiationas a presumed proxy) and the prevalence of TB (reviewed inRalph et al.256), but it is not clear whether low vitamin D status
increases the risk of symptomatic TB or vice versa. A beneficialeffect of vitamin D is plausible through its immune properties,such as the macrophage-induced death of the M. tubercu-losis.256 However, vitamin D supplementation as an adjuvantto standard antimicrobial therapy has shown no clinicalbenefit in most studies.257–259 In a trial in Mongolian children,supplementation with vitamin D stimulated innate immunityagainst M. tuberculosis, which could be sufficient to preventthe infectious process.260 Further research using optimaldoses of vitamin D supplementation and with exploration ofthe effect of host determinants, such as VDR genotype, arerequired to establish whether improving vitamin D statuscould aid in the prevention or treatment of TB.
Vitamin D and respiratory infections. Viruses infecting therespiratory tract and causing disease, such as bronchiolitisand pneumonia, are a leading cause of hospitalisations anddeath in young children, and of serious illness and death inthose over 65 years. Examples include influenza virus, respira-tory syncytial virus, and rhinovirus. Viral respiratory infectionsare most common in winter and least common in summer.This seasonal pattern is diminished in the tropics where thereis also relatively little seasonal variation in solar UV-B radi-ation, although there are two peaks of infection in somecountries, perhaps reflecting the rainy seasons.261 It is hypoth-esised that vitamin D-dependent immunoregulation mediatesthese seasonal patterns.262
Recent observational studies typically show that lowerconcentrations of 25(OH)D are associated with greater risk ofhaving a respiratory tract infection (for example263–266).However, in most cases, study participants had disease symp-toms when they were first assessed, so that low vitamin Dstatus could be either a cause or a consequence of the infec-tion. The results from supplementation trials with vitamin Dare inconsistent, as indicated by the following studies. Separ-ate trials in Japanese267 and Mongolian268 children showedreduced incidence of respiratory infection in the sup-plemented groups, and there is some evidence of benefit inpostmenopausal women,269 young Finnish men270 and olderAustralian adults (using antibiotic use as a surrogate for infec-tion).271 In contrast, trials of supplementation in adults fromthe United States272,273 and New Zealand274 failed to show anybeneficial effect of vitamin D.
Reducing the risk of infectious diseases would have signifi-cant impacts on personal morbidity and global economies;however, the role of vitamin D in promoting this is unclear atthis time.
Vitamin D and multiple sclerosis. Observational studiesconsistently show that higher concentrations of 25(OH)D areassociated with lower risk of MS (reviewed in Hewer et al.197),and that vitamin D deficiency is associated with decreasedresponsiveness to MS treatment, and may be a risk factor forhigher MS disease activity and more rapid progression (forexample, Ascherio et al.275).
Most studies, however, cannot distinguish between causeand effect; that is, does low vitamin D status cause the diseaseor does the disease cause the low vitamin D status? Indeed,
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 65

there is evidence that concentrations of 25(OH)D are reducedby inflammation.276 Further, a review of randomised controlledtrials of supplementation with vitamin D in people with MSfound no evidence of improvement in clinical endpoints.251
Overall, the authors hypothesised that either uncontrolled con-founding or reverse causality provided an explanation for thestrong and consistent associations in observational studiesand the lack of effect of supplementation.
It is also important to consider possible heterogeneity inMS, coupled with a lack of understanding of the timing of theonset of disease pathology and thus the most appropriate timeto test vitamin D status or to give vitamin D supplementation.In addition, the risk factors for disease onset may differ fromthose of progression.277
Vitamin D and cancer risk. Vitamin D deficiency mayincrease the risk of developing cancer as 1,25(OH)2D has regu-latory effects on cellular growth, apoptosis and formation ofnew blood vessels. Observational studies consistently showassociations between low circulating 25(OH)D and increasedrisk of colorectal cancer (reviewed in Autier et al.251) but, forother cancers, there have either been too few studies or theresults are inconsistent.251,252 Trials do not show evidence ofbeneficial effects with vitamin D supplementation.278 Inaddition to the reasons discussed above for similar discrepantfindings in relation to MS, the null findings from trials withsupplementation of vitamin D may be due to too low a sup-plement dose, poor compliance, too short a follow-up,inadequate statistical power of the study, or, parsimoniously,suggest that vitamin D has no effect.251
It is particularly difficult to assess the effects of vitamin Don the risk of skin cancer, as UV radiation induces both. Pro-duction of 1,25(OH)2D in the skin enhances the repair of UV-induced DNA damage.279,280 Cohort studies have shown protec-tive281,282 and adverse283 associations between concentrationsof 25(OH)D and risk of NMSC and similar inconsistencies forthe risk of CMM.283,284 Post-hoc analyses of the Women’sHealth Initiative trial did not show a protective effect of sup-plementation with vitamin D on the risk of NMSC or CMM, butin people with a history of NMSC, the incidence of CMM wasreduced in those randomised to 400 IU of vitamin D per day.285
In summary, the evidence that higher vitamin D status isprotective for cancer is weak and inconsistent, except possiblyfor colorectal cancer risk. If vitamin D deficiency is truly a riskfactor, it is low concentrations of 25(OH)D (i.e., <30 nmol L−1)that are associated with increased risk with little additionalbenefit for concentrations >50 nmol L−1.286
Vitamin D and cancer-survival. Higher vitamin D status hasbeen positively associated with survival from a number ofdifferent cancers,251,287 but there is a paucity of trial data, andconfounding by severity of disease or co-morbidities is highlylikely. At this time there is insufficient evidence to draw anyconclusions.
Effects of vitamin D on other health conditions. Severalstudies suggest that maternal vitamin D deficiency is associ-ated with adverse outcomes in the offspring across multipledomains: bone mineral content,288 cognitive function,289
depression and risk of eating disorders,290 and autism.289 Fur-thermore, vitamin D deficiency in early life has been linked toincreased risk of schizophrenia in later life.291 These resultsmay indicate that vitamin D status during pregnancy is impor-tant for foetal development, but further research will berequired to confirm whether these are causal associations ordue to a related factor(s).
Prospective studies have reported moderate to stronginverse correlations between concentrations of 25(OH)D andcardiovascular diseases, concentrations of serum lipids, inflam-mation, disorders of glucose metabolism, weight gain, mooddisorders, declining cognitive function, dementia, Alzheimer’sdisease, impaired physical functioning, and mortality. In con-trast, intervention trials show no effect of vitamin D sup-plementation on these outcomes.251
U-shaped associations between vitamin D metabolites anddisease
Recent research shows that both high and low concentrationsof 25(OH)D are associated with increased disease risk – a so-called U-shaped association. Table 4 provides a summary ofstudies showing this effect.
An additional U-shaped association has been shownbetween concentration of 1,25(OH)2D and viral load in patientswith HIV.306
There is a trend to advocate ever higher cut-offs to denotethe concentration of 25(OH)D concentrations that is optimalto prevent disease.307 It is therefore important to continue toinvestigate possible disease risks at higher concentrations of25(OH)D, especially if the aim is to maintain such levels overthe long-term.
Other effects of solar UV radiation onhuman health
Chronic exposure to UV radiation has been weakly linked to arange of other adverse health outcomes, including: decreased
Table 4 Health outcomes for which a U-shaped association withserum 25(OH)D levels has been described, and the 25(OH)D level oflowest risk
Disease/condition
Concentration (nmol L−1) oflowest risk, i.e. the turningpoint of the U-shaped doseresponse
All-cause mortality292–294 80–100Cardiovascular events295,296 50–100Cancer mortality297,298 100 (men only)Prostate cancer299,[300] 50 [≤55 (highest risk, 91–106)]Pancreatic cancer301 <100Allergen-specific IgE duringchildhood302
50–74.9
Tuberculosis303 76–140Schizophrenia (neonatal vitamin Dstatus)291
47
Small-for-gestational-age birthsamong white women304
60–70
Physical frailty in older women305 50–74.9
Perspective Photochemical & Photobiological Sciences
66 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015

epidermal308 and subcutaneous309 lipid synthesis resulting inweakening of the barrier function of the skin; hearing impair-ment, through oxidative stress pathways;310,311 lactase non-persistence;312 increased risk of prostate cancer;313 andacquired bilateral nevus-of-Ota-like macule, a common pig-mentation disorder in Asian females.314
In contrast, there is evidence of a protective effect of higherUV radiation on development of restless legs syndrome315 andkeloid formation in scars.316 There are a growing number ofstudies examining the apparently beneficial effects of exposureto the sun. A recent study has shown that UV-A irradiation iseffective in lowering blood pressure, possibly through UV-A-induced nitric oxide bioactivity.317 A reduced risk of cancerand particularly breast cancer has been reported in associationwith greater exposure to the sun in early life,318 as well as adecreased risk of myocardial infarction, and all-cause mor-tality, and hip fracture in those below age 90, where history ofskin cancer was the measure of past exposure to the sun.319 InChile the most frequent cases of food-related anaphylaxis(severe allergic reaction) occur at higher latitudes where thereis lower solar radiation.320
Personal protection from solar UVradiation
The threat of increasing levels of UV-B radiation due to strato-spheric ozone depletion, and rising skin cancer incidencerates, led to the development of sun protection programs andstrategies. Despite the partial recovery of the ozone layer(reviewed in Bais et al.2), such strategies remain importantbecause changes in climate are likely to alter both ambient UVradiation and behaviour that affects exposure to the sun.
A sustained effort is required to change attitudes and behav-iours in relation to exposure to the sun, especially in youngpeople.321 Several recent surveys indicate that adults,322,323
adolescents,324 and children,325,326 still commonly reporthaving been sunburned in the previous year, and this appliesto both fair- and dark-skinned populations.327
While in some locations a majority of children use someform of photoprotection, particularly shade,325,326,328 this isnot true of adults.323 Good knowledge of protection from thesun may not translate into attitudes and practices for reducingexposure.326,328,329 Further, more education about the specificneeds for photoprotection in different situations may berequired. In a sample of young German children, parentalknowledge of appropriate use of shade, clothing, a sunhat,and sunscreen was considered to be adequate for summerholidays at the beach, but not for everyday outdoor activi-ties.330 In a Danish study, travel to a sunny destination wascommon (almost 50% of those aged 15–59 years took such aholiday each year), with a high likelihood of sunburn andintentional tanning.331 There are limited data on strategies forphotoprotection in tropical countries but one survey in adoles-cents living in Bangkok, Thailand, found that sunscreens, sun-protective clothing, and shade were seldom used, particularly
in males, compared with Western countries.332 In a study inPhiladelphia, USA, Hispanic adolescents and young adultswho showed evidence of greater adoption of US culture wereless likely to use sunscreen and more likely to deliberatelyexpose themselves to the sun than those who retained theirtraditional culture.333 Barriers to personal protection from thesun in young Australians include peer pressure, lifestyle,fashion and social norms.334
The ultraviolet index (UVI) is routinely published in themedia in some countries, available online,335 on a mobilephone,336 on mobile phone apps,337 or can be approximatedusing a compact disk as a sundial.338 However, its use to guidebehaviour in relation to exposure to the sun remainslimited.339 Furthermore, the specific guidance in relation tothe UVI varies across different countries. For example, inAustralia, photoprotection is recommended when the UVIndex is ≥3.335 In New Zealand, a “UV Sun protection AlertPeriod” is provided daily, rather than the UVI, and is definedas the period of the day where the forecast clear-sky UVI is>3.340 The United States Environmental Protection Agency pro-vides a UV Alert when the forecast UVI is ≥6, with advice tominimise time in the sun and use protection.341 As noted,exposure to the sun when the UVI is <3 is relatively ineffectivefor vitamin D production, whereas, for some fair-skinned indi-viduals, even short exposures at a UVI of 6 may result inerythema. It is difficult to provide blanket recommendations,even according to the UVI and skin type, as there is wide vari-ation in the minimal erythema dose (MED, the dose of UVradiation required to cause a slight reddening of the skin)even within a specific skin type, and different messages maybe required for different regions.69
It is not generally realised that measures for protectionagainst the sun may be required on cloudy days, in addition toclear days, due to diffuse solar UV radiation,342,343 and thatsome shade devices, such as umbrellas, provide incompleteprotection from UV-B radiation.344
Sunscreens
Public health bodies have long advocated the use of sun-screens as a means of photoprotection. A long-term prospec-tive study in Queensland, Australia, showed that daily use ofsunscreen reduced the incidence of SCC345 with some evi-dence of reduction in the incidence of CMM.346 In addition,such use also reduced photoageing of the skin.347 However, arecent meta-analysis of the effectiveness of interventions pro-moting use of sunscreen in adults and children in recreationalsettings showed no reduction of sunburn in adults and only amodest effect in children.348
Sunscreens are formulated and tested for their ability toprevent erythema in vivo and their index of efficacy is thesun protection factor (SPF).349 The labelled SPF is equal to:[MEDprotected skin]/[MEDunprotected skin], when tested under labo-ratory conditions with simulated solar radiation. Theerythema action spectrum dictates that this is primarily anindex of protection from UV-B radiation, although sunscreensare also required to have a measure of protection against UV-A
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 67

radiation.350 Sunscreens with uniform absorption acrossthe whole UV spectrum (broad-spectrum) provide photoprotec-tion that is similar to shade or some types of clothing fabric351
(for more detail, see Andrady et al.352). Some UV radiationfilters may also have anti-inflammatory properties,353 in whichcase the SPF may be more than a measure of opticalfiltering.354
One requirement of the SPF test is that sunscreen isapplied at a coverage of 2 mg cm−2, but several studies haveshown that people apply much less (for example, Petersenet al.355). The thickness of the application is also dependenton the formulation, with coverage of only 0.22 mg cm−2
achieved by children applying sunscreen with a roll-on.356 Useof a higher SPF sunscreen or two applications of sunscreencan achieve a greater level of protection.357,358 In addition toinadequate thickness of application, failure to apply sunscreento all areas of exposed skin also limits the amount of photo-protection achieved. There are concerns that using sunscreenwill decrease synthesis of vitamin D. Current evidence suggeststhat, if sunscreen is correctly applied, there may be no increasein concentrations of 25(OH)D following exposure to the sun.359
However, with usual applications, there is minimal impair-ment of the synthesis of vitamin D.282,360
Traditional topical sunscreens depend on the filtering orscattering of UV radiation, i.e., “passive” photoprotection.Table 5 summarises some recent developments in compoundsproviding “active” photoprotection at a topical and systemiclevel.361
Overall, people are probably getting much less protectionfrom the harmful effects of UV radiation than they believewhen they use sunscreens, especially if their intention is toprolong their time in the sun. This is a public health issuethat has to be addressed either by encouraging people not togo outdoors when the UVI is high, or to use appropriate cloth-ing (see also Andrady et al.352) for sun protection.335 Alterna-tively, people need to apply sunscreen more thickly, or usesunscreen of a higher SPF to compensate for inadequateapplication.
Clothing and shade
Clothing modifies the skin surface area exposed to solar UVradiation. It offers good protection against sunburn, althoughthis is dependent on the properties of the fabric such ascolour, structure (e.g., woven vs. knit and tightness of theweave for woven fabrics), and wetness (see Andrady et al.352).Clothing typically strongly attenuates transmission of erythe-mally-effective UV radiation (i.e., weighted with the erythemaaction spectrum) to the skin. However, this blocking of UV-Bradiation commonly leads to low vitamin D status in peoplewho wear full body clothing,376 for example for religious or cul-tural reasons.377,378 It may be possible to design and manufac-ture clothing from fabrics that allows synthesis of vitamin Dwhile preventing a visible erythema.379 However, it is likelythat this will not protect from suberythemal damage, such asto the DNA of the epidermal cells.
Shade-seeking is a well-recognised and effective way ofreducing exposure to solar UV radiation, as evidenced by alower level of sunburn380 and lower vitamin D status in adultsfrom the USA who reported frequent use of shade on a sunnyday, compared to those who used shade rarely.360 Provision ofshade over playgrounds, particularly in sunny locations, is arelatively inexpensive method of mass protection from the sun.Nevertheless, a systematic review of 23 publications found noevidence that health promotion interventions had any effect inincreasing shade-seeking in adults or children.348 Shade maybe less effective at reducing exposure to diffuse UV radiationand this may account for about 80% of people’s cumulativeannual erythemal exposure.343 A recent study has used amanikin head (in a fixed position) to measure exposure to UVradiation under different conditions of shade, cloud cover andsolar angle.381 This approach can be used to quantify the protec-tive benefits of shade that are currently not well documented.
Preventing skin cancer versus ensuring adequate vitamin Dstatus
There is no simple message to guide optimal levels of exposureto the sun. There is considerable variation between individuals
Table 5 Newer active sunscreens and evidence of their effectiveness
Class Route and Effectiveness
α-Melanocyte stimulating hormone analogues – stimulatemelanogenesis (tanning), e.g., afamelanotide
Sub-cutaneous; effective in some photosensitive patients362,363
Natural anti-oxidants, e.g., vitamin A (retinol), vitamin C (ascorbic acid),vitamin E (tocopherol), pycnogenol (pine bark extract), carotenoids
Oral; not as effective as topical sunscreens361,364
Nicotinamide (an amide form of vitamin B3) Topical; enhances DNA repair, prevents UV-inducedimmunosuppression, reduces incidence of NMSC365
Resveratrate, found in red wine, grapes, plums and peanuts Topical; application immediately after exposure to the sun protectedagainst erythema and sunburn cell formation366
Flavonoids, e.g., luteolin (a) Protects skin by a combination of UV-absorbing, DNA-protective,antioxidant and anti-inflammatory effects.367
Lycopene, found in tomato paste Oral; ingestion over a 12 week period reduced acute & chronic effectsof photodamage368
Green tea polyphenols Oral; some evidence of modest effect369,370
Diet rich in omega-3 fatty acids Oral; may reduce risk of skin cancer371,372
Liposomes containing natural endonucleases or photolyases Topical; enhance nucleotide excision repair of UV-induced DNAdamage; shown to reduce the incidence of solar keratoses and theseverity of polymorphic light eruption.373–375
Perspective Photochemical & Photobiological Sciences
68 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015

in the doses of UV radiation that cause damage to DNA andinduce synthesis of vitamin D. However, some broad recom-mendations can be made. Repeated short exposures to thesun are more efficient at vitamin D production than a singleprolonged exposure.218 Levels of UV-B radiation are greatest atmidday and vitamin D synthesis is thus most efficient at thistime,382 although it is also the time when sunburn occursmost quickly.383 The amount of time in the sun that is neededfor the synthesis of vitamin D varies according to location,time of day, and time of year. After controlling for the level ofexposure to UV radiation, having more skin not covered byclothing is associated with higher concentrations of 25(OH)Din serum.246,247 As noted above, there are conflicting results onthe effect of darker pigmentation of skin on the UV-inducedproduction of vitamin D. In high latitude locations where UV-Blevels are too low for vitamin D synthesis and a cooler climatemay mean that little skin is exposed to the sun even duringsummer, greater intake of vitamin D may be required to avoidvitamin D deficiency.
Protection of the eye
One of the cheapest and most practical methods of protectingthe eye from exposure to UV-B radiation is wearing a hat with abrim of at least 6–7 cm.335,384 Sunglasses provide variable pro-tection, and standards are more rigorous for UV-B than forUV-A radiation. For UV-B radiation the upper transmissionlimits range from 1.0% to 12.5% depending on the inter-national jurisdiction and the type of use. For UV-A radiation,the limit is either a maximum of 50% of visible transmittanceor is unspecified. The size of the frame and design can influ-ence eye protection and some standards for sunglasses incor-porate a minimum size limit.385 Wrap-around designs aremost protective and are especially important when in highlyreflective conditions, for example when skiing.385 Many ordi-nary eye-glass lenses have UV filters in them.
The American National Standards Institute (ANSI) requiresthat contact lenses absorb at least 95% of UV-B radiation and70% of UV-A radiation for a “UV blocking” claim. A recentsurvey on a selection of lenses showed compliance with claimsfor photoprotection.386
Studies using manikin heads387,388 have shown that,although the ambient UV radiation is greatest at solar noon,the highest dose of UV radiation was received by the eye at4 hours before and after noon, when the solar elevation anglewas lower. Thus, photoprotection of the eyes during outdooractivities is important not only at noon, but also at othertimes, and during winter.389 In some occupations, e.g.,working on a building site, the reflectivity of the buildingmaterials may influence the amount of UV radiation receivedby the eye390 and should be considered when wearing eye-protection.
Protection of the eyes from the sun should reduce the riskof pterygium; UV absorbing contact lenses that cover most ofthe cornea can protect against UV-induced damage.385,391 Eyedrops for the prevention of pterygium and photokeratitis/photoconjunctivitis through anti-oxidant and anti-inflamma-
tory pathways have been effective in animal models, but arenot commonly used in humans.119,392
Effects of interactions between solarUV radiation and the environment
Environmental contaminants may interact synergistically withUV radiation to harm human health. For example, in the pres-ence of UV-B radiation, chrysene,393 a common environmentalcontaminant produced by incomplete burning of fossil fuels,and some pesticides394 have adverse effects on human health,including through damage to DNA.395 Topical corticosteroidsare unstable under UV-B irradiation, possibly causing skindamage as well as loss of therapeutic effect.396 Engineerednanoparticles (NPs) are increasingly incorporated into sportingequipment, sunscreens, clothing and cosmetics (see alsoAndrady et al.352). Concerns have been raised about possiblehealth risks of NPs.397,398 A recent review of the evidence onNPs in sunscreens concluded that “on current evidence,neither TiO2 nor ZnO NPs are likely to cause harm when usedas ingredients in sunscreens”.399 Nevertheless, exposure ofskin to UV radiation may enhance the penetration of engi-neered NPs,400 and the formulations using these particles areoften used around the time of irradiation when skin damagemay occur. Further, UV-induced immunosuppression couldpotentially impair an immune protective response induced byengineered NPs applied to the skin.400
On the positive side, UV radiation is a potent environ-mental disinfectant able to inactivate viruses in clear water(for further discussion see Häder et al.401). This property isused in the SODIS (solar disinfection) technique, whereexposure of surface water within a transparent bottle to sun-light effectively disinfects the water, decreasing the incidenceof diarrhoeal diseases (reviewed in McGuigan et al.402).
Health implications of interactionsbetween ozone depletion and climatechange
Past studies have estimated the health gains in terms of skincancers avoided through the implementation of the MontrealProtocol and its amendments.403 A recent update of this workthat also integrated coupled climate-chemistry models has esti-mated that the world-wide incidence of skin cancer wouldhave been 14% greater (2 million people) by 2030 withoutimplementation of the Montreal Protocol and its amend-ments,404 with the largest effects in the South West USA and inAustralia (see Bais et al.2 for further details).
Model estimates405 suggest that by 2050, any increases inerythemal UV radiation above present levels will be small andconfined to the tropical region (reviewed in Bais et al.2). Theseshould have only a small effect on the incidence of skin
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 69

cancer406 but may impair immune responses to some vacci-nations. However, outside the tropical region, erythemal UVradiation is projected to be lower, especially in winter(reviewed Bais et al.2), which could be detrimental for thevitamin D status of populations in these regions, as well as fordiseases that may be modulated by exposure to UV radiation,such as some of the autoimmune diseases discussed above.Estimates of exposure times for erythema and vitamin D syn-thesis in Europe, taking account of ozone recovery and inter-actions with different concentrations of greenhouse gases(GHG),406 suggest that there will be very little change in theexposure time for both endpoints for Southern Europe.However, an increased exposure time of about 30% for vitaminD production would be required in a worst case scenario inStockholm in spring with high levels of GHGs.406
A major determinant of the received dose of UV radiation isbehaviour in relation to exposure to the sun.407 Ambienttemperature is likely to influence time spent outdoors. Intemperate parts of Australia, the increase in temperature islikely to increase skin cancer incidence because people willspend more time outside with less clothing.408,409 However,with temperature increases in already warm climates, peoplewill be more likely to stay indoors or to seek shade.229 Warmerambient temperatures may have direct physiological effects toaccelerate both skin cancer development410 and vitamin Dproduction.411
In addition to these direct effects of interactions betweenclimate change and ozone depletion and/or UV radiation forhuman health, there are potential indirect effects that maybecome important, but at this stage remain ill-defined.412 Con-current changes in climate and levels of ambient UV radiationwill influence aquatic and terrestrial ecosystems (see alsoHäder et al.401 and Bornman et al.413), which may have conse-quences for food safety, quality and supply (reviewed in412).
Migration of populations, often with dark skin pigmenta-tion, from low-lying tropical regions because of rising sealevels, to higher latitude regions may increase the diseasesassociated with vitamin D deficiency.
Higher temperatures should foster microbial growth insurface waters, and this will be more pronounced in the pres-ence of lower levels of disinfecting UV-B radiation, or whereincreases in colour due to dissolved organic matter limit pene-tration of UV-B radiation (for more detail, see Häder et al.,401
Bornman et al.,413 and Erickson et al.414). However, there hasbeen little research to date that allows prediction or quantifi-cation of the risks to human health that might arise fromthese interactions.
Gaps in our knowledge
Considerable evidence from animal models suggests that UV-induced immunosuppression may increase the risk of someinfections and decrease the protection offered by vaccination(reviewed in Norval & Halliday212). Studies in humans arerequired to determine whether any increased risk is clinically
relevant, for example, requiring changes in vaccination proto-cols. Although some animal studies similarly suggest thatvitamin D status affects the outcomes of vaccination orimmune responses to infection, results from clinical trialsmostly show no effect.415,416
There is currently considerable controversy about whichhealth conditions are influenced by vitamin D. Large-scalevitamin D supplementation trials that are in progress willprovide some answers (see Table 1 in Byrne417). While there isconsensus that vitamin D is important for bone health, thereis lack of agreement about the concentration of 25(OH)Drequired. In addition, there is substantial individual variabilityin the change in concentration in 25(OH)D in response to UVirradiation and vitamin D supplementation, and in the clinicaleffects associated with different concentrations of 25(OH)D.The reasons for this are poorly understood, but are likely todepend on variation in the genes encoding the vitamin Dbinding protein and/or the vitamin D receptor.
Production of vitamin D occurs readily at sub-erythemaldoses, but repeated sub-erythemal exposures can also causeaccumulation of CPDs that repair only slowly (over24–36 hours) and thus may increase the risk of skin cancer.418
Until there is a better understanding of the numbers of CPDsthat accrue during brief sun exposures, and their importancein determining the risk of subsequent skin cancer, it isdifficult to recommend safe exposures that would result insufficient production of vitamin D.
There is a lack of data on the vitamin D status of infants,children, and adolescents and for populations in Africa andSouth America. Further, the lack, until recently, of accurateand precise assays has limited our ability to examine variabilityin vitamin D status across countries (for example those withand without fortification of food) and over time. The advent ofthe standardised vitamin D assays419 means that this is nowpossible. Development of a less invasive sampling method, forexample using saliva, would allow more widespread assess-ment of the vitamin D status of infants and children.
Protection from the sun is currently recommended by theWorld Health Organization when the UVI is ≥3. The corollaryof this message is that photoprotection is not required whenthe UVI is <3. At these low UVI values, there is little UV-Bradiation (and thus little vitamin D synthesis), but, with pro-longed exposure, there may be a relatively high dose of UV-Aradiation. With recognition that UV-A radiation can induceimmune suppression164 and is involved in the initiation ofCMM, but may also have beneficial effects on blood pressure,the doses received and potential health effects need to bebetter defined.
The action spectra for a number of health outcomes havenot been determined. These include cataract from protractedexposure, myopia, carcinoma in deeply pigmented skin, mela-noma and production of pre-vitamin D from a polychromaticsource (the sun) and in both dark and fair skin types.However, the relevance of the currently available animalmodels is uncertain, so obtaining these action spectra withdefinite relevance to humans is very difficult.
Perspective Photochemical & Photobiological Sciences
70 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015

There is emerging evidence that exposure to the sun mayhave beneficial effects independently of vitamin D. The lack ofa strong evidence-base challenges our ability to provide accu-rate guidance to the public regarding exposure to the sun.
As noted above, there is much uncertainty about the effectof the potential indirect interactions of climate change andozone depletion on human health. Effects on disinfection ofsurface water and on supply and security of food could becomeimportant sources of risks to health, particularly in someregions of the world. Modelling now may be able to identifythose areas at greatest risk, and allow forward planning tomitigate the risks.
Acknowledgements
We would like to acknowledge the following people andorganizations for their support in the preparation of thispaper. Prof Robyn Lucas’ participation in the Panel was sup-ported through funding from the Australian Government’sOzone Science Strategy. A/Prof Rachel Neale was supported bythe QIMR Berghofer Institute for Medical Research. Prof YukioTakizawa was sponsored by the Japanese Ministry of theEnvironment. Ms Tammy Gibbs provided support with thefigures in this paper.
References
1 P. Rettberg, U. Eschweiler, K. Strauch, G. Reitz,G. Horneck, H. Wanke, A. Brack and B. Barbier, Survivalof microorganisms in space protected by meteoritematerial: results of the experiment ‘EXOBIOLOGIE’ of thePERSEUS mission, Adv. Space Res., 2002, 30, 1539–1545.
2 A. F. Bais, R. L. McKenzie, P. J. Aucamp, M. Ilyas,S. Madronich, G. Bernhard and K. Tourpali, Ozonedepletion and climate change: Impacts on UV radiation,Photochem. Photobiol. Sci., 2015, 14, DOI: 10.1039/c4pp90032d, this issue.
3 WHO, Environmental Health Criteria 160 - Ultraviolet Radi-ation, World Health Organization, 1994.
4 M. R. Albert and K. G. Ostheimer, The evolution ofcurrent medical and popular attitudes toward ultravioletlight exposure: part 2, J. Am. Acad. Dermatol., 2003, 48,909–918.
5 A. M. Eggermont, A. Spatz and C. Robert, Cutaneous mela-noma, Lancet, 2013, 383, 816–827.
6 J. E. Frangos, L. M. Duncan, A. Piris, R. M. Nazarian,M. C. Mihm, Jr., M. P. Hoang, B. Gleason, T. J. Flotte,H. R. Byers, R. L. Barnhill and A. B. Kimball, Increaseddiagnosis of thin superficial spreading melanomas: A20-year study, J. Am. Acad. Dermatol., 2012, 67, 387–394.
7 IARC, Globocan 2012. Estimated cancer incidence, mortalityand prevalence worldwide in 2012, International Agency forResearch on Cancer, http://globocan.iarc.fr/Pages/summary_table_site_sel.aspx, accessed 28 June 2014.
8 R. Laishram, A. Banerjee, P. Punyabati, L. Chandra andA. Sharma, Pattern of skin malignancies in Manipur,India: A 5-year histopathological review, J. Pak. Assoc.Dermatol., 2010, 20, 128–132.
9 F. Erdmann, J. Lortet-Tieulent, J. Schuz, H. Zeeb,R. Greinert, E. W. Breitbart and F. Bray, Internationaltrends in the incidence of malignant melanoma1953–2008 – are recent generations at higher or lowerrisk?, Int. J. Cancer, 2013, 132, 385–400.
10 M. Arnold, C. Holterhues, L. M. Hollestein, J. W. Coebergh,T. Nijsten, E. Pukkala, B. Holleczek, L. Tryggvadottir,H. Comber, M. J. Bento, S. Diba Ch, R. Micallef, M. Primic-Zakelj, M. I. Izarzugaza, J. Perucha, R. Marcos-Gragera,J. Galceran, E. Ardanaz, R. Schaffar, A. Pring and E. deVries, Trends in incidence and predictions of cutaneousmelanoma across Europe up to 2015, J. Eur. Acad. Derma-tol. Venereol., 2014, 28, 1170–1178.
11 J. Ferlay, F. Bray, P. Pisani and D. Parkin, GLOBOCAN2000: Cancer Incidence, Mortality and Prevalence World-wide, IARC Cancer Base No. 5. Lyon, 2001.
12 V. Nikolaou and A. J. Stratigos, Emerging trends in theepidemiology of melanoma, Br. J. Dermatol., 2014, 170,11–19.
13 A. C. Geller, R. W. Clapp, A. J. Sober, L. Gonsalves,L. Mueller, C. L. Christiansen, W. Shaikh and D. R. Miller,Melanoma epidemic: an analysis of six decades of datafrom the Connecticut Tumor Registry, J. Clin. Oncol.,2013, 31, 4172–4178.
14 P. T. Bradford, W. F. Anderson, M. P. Purdue,A. M. Goldstein and M. A. Tucker, Rising melanoma inci-dence rates of the trunk among younger women in theUnited States, Cancer Epidemiol. Biomarkers Prev., 2010,19, 2401–2406.
15 P. D. Baade, A. C. Green, B. M. Smithers and J. F. Aitken,Trends in melanoma incidence among children: possibleinfluence of sun-protectionprograms, Expert. Rev. Anti-cancer Ther., 2011, 11, 661–664.
16 A. Stang, S. Valiukeviciene, B. Aleknaviciene andJ. Kurtinaitis, Time trends of incidence, mortality, andrelative survival of invasive skin melanoma in Lithuania,Eur. J. Cancer Prev., 2006, 42, 660–667.
17 N. Duschek, H. Skvara, H. Kittler, G. Delir, A. Fink,A. Pinkowicz and T. Waldhor, Melanoma epidemiology ofAustria reveals gender-related differences, Eur. J. Dermatol.,2013, 23, 872–878.
18 L. Tryggvadottir, M. Gislum, T. Hakulinen, A. Klint,G. Engholm, H. H. Storm and F. Bray, Trends in the survi-val of patients diagnosed with malignant melanoma ofthe skin in the Nordic countries 1964–2003 followed up tothe end of 2006, Acta Oncol., 2010, 49, 665–672.
19 M. J. Sneyd and B. Cox, A comparison of trends in mela-noma mortality in New Zealand and Australia: the twocountries with the highest melanoma incidence and mor-tality in the world, BMC Cancer, 2013, 13, 372.
20 A. Katalinic, A. Waldmann, M. A. Weinstock, A. C. Geller,N. Eisemann, R. Greinert, B. Volkmer and E. Breitbart,
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 71

Does skin cancer screening save lives?: an observationalstudy comparing trends in melanoma mortality in regionswith and without screening, Cancer, 2012, 118, 5395–5402.
21 A. E. Cust, M. A. Jenkins, C. Goumas, B. K. Armstrong,H. Schmid, J. F. Aitken, G. G. Giles, R. F. Kefford,J. L. Hopper and G. J. Mann, Early-life sun exposure andrisk of melanoma before age 40 years, Cancer CausesControl, 2011, 22, 885–897.
22 S. C. Wallingford, R. D. Alston, J. M. Birch andA. C. Green, Increases in invasive melanoma in England,1979–2006, by anatomical site, Br. J. Dermatol., 2011, 165,859–864.
23 N. B. Fuglede, U. O. Brinck-Claussen, I. Deltour,E. H. Boesen, S. O. Dalton and C. Johansen, Incidence ofcutaneous malignant melanoma in Denmark, 1978–2007,Br. J. Dermatol., 2011, 165, 349–353.
24 B. Diffey, A behavioral model for estimating populationexposure to solar ultraviolet radiation, Photochem. Photo-biol., 2008, 84, 371–375.
25 P. Rouhani, S. Hu and R. S. Kirsner, Melanoma in Hispa-nic and black Americans, Cancer Control, 2008, 15, 248–253.
26 M. Clairwood, J. Ricketts, J. Grant-Kels and L. Gonsalves,Melanoma in skin of color in Connecticut: an analysisof melanoma incidence and stage at diagnosis in non-Hispanic blacks, non-Hispanic whites, and Hispanics,Int. J. Dermatol., 2013, 53, 425–433.
27 M. G. Cockburn, J. Zadnick and D. Deapen, Developingepidemic of melanoma in the Hispanic population ofCalifornia, Cancer, 2006, 106, 1162–1168.
28 R. A. Schmerling, D. Loria, G. Cinat, W. E. Ramos,A. F. Cardona, J. L. Sanchez, H. Martinez-Said andA. C. Buzaid, Cutaneous melanoma in Latin America: theneed for more data, Rev. Panam. Salud. Pacific., 2011, 30,431–438.
29 P. M. Nthumba, P. C. Cavadas and L. Landin, Primarycutaneous malignancies in sub-Saharan Africa, Ann. Plast.Surg., 2011, 66, 313–320.
30 M. Norval, P. Kellett and C. Y. Wright, The Incidence andBody Site of Skin Cancers in the Population Groups ofSouth Africa, Photodermatol. Photoimmunol. Photomed.,2014, 30(5), 262–265.
31 O. N. Agbai, K. Buster, M. Sanchez, C. Hernandez,R. V. Kundu, M. Chiu, W. E. Roberts, Z. D. Draelos,R. Bhushan, S. C. Taylor and H. W. Lim, Skin cancer andphotoprotection in people of color: a review and recom-mendations for physicians and the public, J. Am. Acad.Dermatol, 2014, 70, 748–762.
32 S. W. Lin, D. C. Wheeler, Y. Park, E. K. Cahoon,A. R. Hollenbeck, D. M. Freedman and C. C. Abnet, Pro-spective study of ultraviolet radiation exposure and risk ofcancer in the United States, Int. J. Cancer, 2012, 131,E1015–E1023.
33 H. E. Kanavy and M. R. Gerstenblith, Ultraviolet radiationand melanoma, Semin. Cutaneous Med. Surg., 2011, 30,222–228.
34 H. Levine, A. Afek, A. Shamiss, E. Derazne, D. Tzur,N. Astman, L. Keinan-Boker, D. Mimouni and J. D. Kark,Country of origin, age at migration and risk of cutaneousmelanoma: a migrant cohort study of 1,100,000 Israelimen, Int. J. Cancer, 2013, 133, 486–494.
35 L. Le Marchand, B. S. Saltzman, J. H. Hankin,L. R. Wilkens, A. A. Franke, S. J. Morris and L. N. Kolonel,Sun exposure, diet, and melanoma in Hawaii Caucasians,Am. J. Epidemiol., 2006, 164, 232–245.
36 F. P. Noonan, M. R. Zaidi, A. Wolnicka-Glubisz,M. R. Anver, J. Bahn, A. Wielgus, J. Cadet, T. Douki,S. Mouret, M. A. Tucker, A. Popratiloff, G. Merlino andE. C. De Fabo, Melanoma induction by ultraviolet A butnot ultraviolet B radiation requires melanin pigment, Nat.Commun., 2012, 3, 884.
37 D. L. Mitchell and A. A. Fernandez, Different types of DNAdamage play different roles in the etiology of sunlight-induced melanoma, Pigm. Cell Melanoma. Res., 2011, 24,119–124.
38 N. O. Basta, P. W. James, A. W. Craft and R. J. McNally,Seasonal variation in the month of birth in teenagers andyoung adults with melanoma suggests the involvement ofearly-life UV exposure, Pigm. Cell Melanoma. Res., 2011,24, 250–253.
39 C. Crump, K. Sundquist, W. Sieh, M. A. Winkleby andJ. Sundquist, Season of birth and other perinatal riskfactors for melanoma, Int. J. Epidemiol., 2014, 43(3), 793–801.
40 S. L. Harrison, P. G. Buettner and R. MacLennan, Why domothers still sun their infants?, J. Pediatr. Child Health,1999, 35, 296–299.
41 S. Harrison, M. Nowak, S. Devine, V. Saunders, A. Smithand P. Buettner, An intervention to discourage Australianmothers from unnecessarily exposing their babies to thesun for therapeutic reasons, J. Trop. Pediatr., 2013, 59,403–406.
42 A. S. Paller, J. L. Hawk, P. Honig, Y. C. Giam, S. Hoath,M. C. Mack and G. N. Stamatas, New insights about infantand toddler skin: implications for sun protection, Pedi-atrics, 2011, 128, 92–102.
43 J. Reichrath, Sunlight, Vitamin D and Skin Cancer, inAdvances in Experimental Medicine and Biology, ed.N. Back, I. Cohen, A. Lajtha, J. Lambris and R. Poaletti,Landes Bioscience/Springer Science+Business media,2014, 2nd edn.
44 I. Chaillol, M. Boniol, R. Middleton, J. F. Dore, P. Autierand A. Gavin, Seasonality of cutaneous melanoma diagno-sis in Northern Ireland with a review, Melanoma Res.,2011, 21, 144–151.
45 C. M. Olsen, H. J. Carroll and D. C. Whiteman, Familialmelanoma: a meta-analysis and estimates of attributablefraction, Cancer Epidemiol. Biomarkers Prev., 2010, 19, 65–73.
46 V. K. Hill, J. J. Gartner, Y. Samuels and A. M. Goldstein,The genetics of melanoma: recent advances, Annu. Rev.Genomics Hum. Genet., 2013, 14, 257–279.
Perspective Photochemical & Photobiological Sciences
72 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015

47 E. Hodis, I. R. Watson, G. V. Kryukov, S. T. Arold,M. Imielinski, J. P. Theurillat, E. Nickerson, D. Auclair,L. Li, C. Place, D. Dicara, A. H. Ramos, M. S. Lawrence,K. Cibulskis, A. Sivachenko, D. Voet, G. Saksena,N. Stransky, R. C. Onofrio, W. Winckler, K. Ardlie,N. Wagle, J. Wargo, K. Chong, D. L. Morton, K. Stemke-Hale, G. Chen, M. Noble, M. Meyerson, J. E. Ladbury,M. A. Davies, J. E. Gershenwald, S. N. Wagner, D. S. Hoon,D. Schadendorf, E. S. Lander, S. B. Gabriel, G. Getz,L. A. Garraway and L. Chin, A landscape of drivermutations in melanoma, Cell, 2012, 150, 251–263.
48 A. Viros, B. Sanchez-Laorden, M. Pedersen, S. J. Furney,J. Rae, K. Hogan, S. Ejiama, M. R. Girotti, M. Cook,N. Dhomen and R. Marais, Ultraviolet radiation accele-rates BRAF-driven melanomagenesis by targeting TP53,Nature, 2014, 511, 478–482.
49 M. F. Berger, E. Hodis, T. P. Heffernan, Y. L. Deribe,M. S. Lawrence, A. Protopopov, E. Ivanova, I. R. Watson,E. Nickerson, P. Ghosh, H. Zhang, R. Zeid, X. Ren,K. Cibulskis, A. Y. Sivachenko, N. Wagle, A. Sucker,C. Sougnez, R. Onofrio, L. Ambrogio, D. Auclair,T. Fennell, S. L. Carter, Y. Drier, P. Stojanov, M. A. Singer,D. Voet, R. Jing, G. Saksena, J. Barretina, A. H. Ramos,T. J. Pugh, N. Stransky, M. Parkin, W. Winckler, S. Mahan,K. Ardlie, J. Baldwin, J. Wargo, D. Schadendorf,M. Meyerson, S. B. Gabriel, T. R. Golub, S. N. Wagner,E. S. Lander, G. Getz, L. Chin and L. A. Garraway,Melanoma genome sequencing reveals frequentPREX2 mutations, Nature, 2012, 485, 502–506.
50 M. Krauthammer, Y. Kong, B. H. Ha, P. Evans,A. Bacchiocchi, J. P. McCusker, E. Cheng, M. J. Davis,G. Goh, M. Choi, S. Ariyan, D. Narayan, K. Dutton-Regester, A. Capatana, E. C. Holman, M. Bosenberg,M. Sznol, H. M. Kluger, D. E. Brash, D. F. Stern,M. A. Materin, R. S. Lo, S. Mane, S. Ma, K. K. Kidd,N. K. Hayward, R. P. Lifton, J. Schlessinger, T. J. Boggonand R. Halaban, Exome sequencing identifies recurrentsomatic RAC1 mutations in melanoma, Nat. Genet., 2012,44, 1006–1014.
51 S. C. Flohil, C. M. Proby, A. D. Forrest, S. van Tiel,O. Saksela, S. Pitkanen, T. Ahti, R. Micallef andE. de Vries, Basal cell carcinomas without histolo-gical confirmation and their treatment: an audit infour European regions, Br. J. Dermatol., 2012, 167(Suppl2), 22–28.
52 F. Xiang, R. Lucas, S. Hales and R. E. Neale, Incidenceof Nonmelanoma Skin Cancer in Relation toAmbient UV Radiation in White Populations, 1978-2012:Empirical Relationships, JAMA Dermatol., 2014, 150(10),1063–71.
53 A. Lomas, J. Leonardi-Bee and F. Bath-Hextall, A systema-tic review of worldwide incidence of nonmelanoma skincancer, Br. J. Dermatol., 2012, 166, 1069–1080.
54 National Cancer Registry of South Africa, Cancer in SouthAfrica, http://www.nioh.ac.za/?page=national_cancer_registry&id=41, accessed 29 July, 2014.
55 S. C. Flohil, I. Seubring, M. M. van Rossum,J. W. Coebergh, E. de Vries and T. Nijsten, Trends in Basalcell carcinoma incidence rates: a 37-year Dutch observa-tional study, J. Invest. Dermatol., 2013, 133, 913–918.
56 H. W. Rogers, M. A. Weinstock, A. R. Harris,M. R. Hinckley, S. R. Feldman, A. B. Fleischer andB. M. Coldiron, Incidence estimate of nonmelanoma skincancer in the United States, 2006, Arch. Dermatol., 2010,146, 283–287.
57 S. Franceschi, F. Levi, L. Randimbison and C. La Vecchia,Site distribution of different types of skin cancer: newaetiological clues, Int. J. Cancer, 1996, 67, 24–28.
58 B. A. Raasch, P. G. Buettner and C. Garbe, Basal cell carci-noma: histological classification and body-site distri-bution, Br. J. Dermatol., 2006, 155, 401–407.
59 M. P. Staples, M. Elwood, R. C. Burton, J. L. Williams,R. Marks and G. G. Giles, Non-melanoma skin cancer inAustralia: the 2002 national survey and trends since 1985,Med. J. Aust., 2006, 184, 6–10.
60 A. Kricker, B. K. Armstrong, D. R. English andP. J. Heenan, Pigmentary and cutaneous risk factors fornon-melanocytic skin cancer–a case-control study,Int. J. Cancer, 1991, 48, 650–662.
61 C. Dessinioti, C. Antoniou, A. Katsambas andA. J. Stratigos, Basal cell carcinoma: what’s new under thesun, Photochem. Photobiol., 2010, 86, 481–491.
62 A. Bauer, T. L. Diepgen and J. Schmitt, Is occupationalsolar ultraviolet irradiation a relevant risk factor for basalcell carcinoma? A systematic review and meta-analysis ofthe epidemiological literature, Br. J. Dermatol., 2011, 165,612–625.
63 M. Khalesi, D. C. Whiteman, S. A. Doi, J. Clark,M. G. Kimlin and R. E. Neale, Cutaneous markers ofphoto-damage and risk of Basal cell carcinoma of theskin: a meta-analysis, Cancer Epidemiol. Biomarkers Prev.,2013, 22, 1483–1489.
64 M. R. Iannacone, W. Wang, H. G. Stockwell, K. O’Rourke,A. R. Giuliano, V. K. Sondak, J. L. Messina,R. G. Roetzheim, B. S. Cherpelis, N. A. Fenske andD. E. Rollison, Patterns and timing of sunlight exposureand risk of basal cell and squamous cell carcinomas ofthe skin–a case-control study, BMC Cancer, 2012, 12, 417.
65 R. E. Neale, M. Davis, N. Pandeya, D. C. Whiteman andA. C. Green, Basal cell carcinoma on the trunk is associ-ated with excessive sun exposure, J. Am. Acad. Dermatol.,2007, 56, 380–386.
66 J. R. Rees, M. S. Zens, J. Gui, M. O. Celaya, B. L. Riddleand M. R. Karagas, Non melanoma skin cancer and sub-sequent cancer risk, PLoS One, 2014, 9, e99674.
67 R. M. Halder and S. Bridgeman-Shah, Skin cancer inAfrican Americans, Cancer, 1995, 75, 667–673.
68 H. M. Gloster, Jr. and K. Neal, Skin cancer in skin ofcolor, J. Am. Acad. Dermatol., 2006, 55, 741–760; quiz761–744.
69 F. Zaratti, R. D. Piacentini, H. A. Guillen, S. H. Cabrera,J. B. Liley and R. L. McKenzie, Proposal for a modification
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 73

of the UVI risk scale, Photochem. Photobiol. Sci., 2014, 13,980–985.
70 H. Nan, M. Xu, P. Kraft, A. A. Qureshi, C. Chen, Q. Guo,F. B. Hu, G. Curhan, C. I. Amos, L. E. Wang, J. E. Lee,Q. Wei, D. J. Hunter and J. Han, Genome-wide associationstudy identifies novel alleles associated with risk ofcutaneous basal cell carcinoma and squamous cell carci-noma, Hum. Mol. Genet., 2011, 20, 3718–3724.
71 S. N. Stacey, D. F. Gudbjartsson, P. Sulem,J. T. Bergthorsson, R. Kumar, G. Thorleifsson,A. Sigurdsson, M. Jakobsdottir, B. Sigurgeirsson,K. R. Benediktsdottir, K. Thorisdottir, R. Ragnarsson,D. Scherer, P. Rudnai, E. Gurzau, K. Koppova, V. Hoiom,R. Botella-Estrada, V. Soriano, P. Juberias, M. Grasa,F. J. Carapeto, P. Tabuenca, Y. Gilaberte, J. Gudmundsson,S. Thorlacius, A. Helgason, T. Thorlacius, A. Jonasdottir,T. Blondal, S. A. Gudjonsson, G. F. Jonsson,J. Saemundsdottir, K. Kristjansson, G. Bjornsdottir,S. G. Sveinsdottir, M. Mouy, F. Geller, E. Nagore,J. I. Mayordomo, J. Hansson, T. Rafnar, A. Kong,J. H. Olafsson, U. Thorsteinsdottir and K. Stefansson,Common variants on 1p36 and 1q42 are associatedwith cutaneous basal cell carcinoma but not with mela-noma or pigmentation traits, Nat. Genet., 2008, 40, 1313–1318.
72 J. J. DiGiovanna and K. H. Kraemer, Shining a light onxeroderma pigmentosum, J. Invest. Dermatol., 2012, 132,785–796.
73 S. Durinck, C. Ho, N. J. Wang, W. Liao, L. R. Jakkula,E. A. Collisson, J. Pons, S. W. Chan, E. T. Lam, C. Chu,K. Park, S. W. Hong, J. S. Hur, N. Huh, I. M. Neuhaus,S. S. Yu, R. C. Grekin, T. M. Mauro, J. E. Cleaver,P. Y. Kwok, P. E. LeBoit, G. Getz, K. Cibulskis, J. C. Aster,H. Huang, E. Purdom, J. Li, L. Bolund, S. T. Arron,J. W. Gray, P. T. Spellman and R. J. Cho, Temporal dissec-tion of tumorigenesis in primary cancers, Cancer Discov-ery, 2011, 1, 137–143.
74 S. S. Jayaraman, D. J. Rayhan, S. Hazany andM. S. Kolodney, Mutational landscape of basal cell carci-nomas by whole-exome sequencing, J. Invest. Dermatol.,2014, 134, 213–220.
75 D. E. Brash, A. Ziegler, A. S. Jonason, J. A. Simon,S. Kunala and D. J. Leffell, Sunlight and sunburn inhuman skin cancer: p53, apoptosis, and tumor pro-motion, J. Invest. Dermatol. Symp. Proc., 1996, 1, 136–142.
76 A. Ziegler, A. S. Jonason, D. J. Leffell, J. A. Simon,H. W. Sharma, J. Kimmelman, L. Remington, T. Jacks andD. E. Brash, Sunburn and p53 in the onset of skin cancer,Nature, 1994, 372, 773–776.
77 E. H. Epstein, Basal cell carcinomas: attack of the hedge-hog, Nat. Rev. Cancer, 2008, 8, 743–754.
78 P. Manga, R. Kerr, M. Ramsay and J. G. Kromberg, Biologyand genetics of oculocutaneous albinism and vitiligo -common pigmentation disorders in southern Africa,S. Afr. Med. J., 2013, 103, 984–988.
79 K. Gronskov, J. Ek and K. Brondum-Nielsen, Oculocuta-neous albinism, Orphanet J. Rare. Dis., 2007, 2, 43.
80 C. V. David, Oculocutaneous albinism, Cutis, 2013, 91,E1–E4.
81 J. G. Kromberg and T. Jenkins, Prevalence of albinismin the South African negro, S. Afr. Med. J., 1982, 61, 383–386.
82 C. G. Summers, Albinism: classification, clinical charac-teristics, and recent findings, Optom. Visual Sci., 2009, 86,659–662.
83 E. S. Hong, H. Zeeb and M. H. Repacholi, Albinism inAfrica as a public health issue, BMC Public Health, 2006,6, 212.
84 J. Luande, C. I. Henschke and N. Mohammed, The Tanza-nian human albino skin. Natural history, Cancer, 1985,55, 1823–1828.
85 J. B. Mabula, P. L. Chalya, M. D. McHembe, H. Jaka,G. Giiti, P. Rambau, N. Masalu, E. Kamugisha, S. Robertand J. M. Gilyoma, Skin cancers among Albinos at aUniversity teaching hospital in Northwestern Tanzania: aretrospective review of 64 cases, BMC Dermatol., 2012,12, 5.
86 S. K. Kiprono, B. M. Chaula and H. Beltraminelli, Histo-logical review of skin cancers in African Albinos: a 10-yearretrospective review, BMC Cancer, 2014, 14, 157.
87 P. K. Perry and N. B. Silverberg, Cutaneous malignancy inalbinism, Cutis, 2001, 67, 427–430.
88 R. Aquaron, Oculocutaneous albinism in Cameroon. A 15-year follow-up study, Ophthalmic. Paediatr. Genet., 1990,11, 255–263.
89 M. P. Hughes, M. E. Hardee, L. A. Cornelius,L. F. Hutchins, J. C. Becker and L. Gao, Merkel Cell Carci-noma: Epidemiology, Target, and Therapy, Curr. Dermatol.Rep., 2014, 3, 46–53.
90 J. Girschik, K. Thorn, T. W. Beer, P. J. Heenan andL. Fritschi, Merkel cell carcinoma in Western Australia: apopulation-based study of incidence and survival,Br. J. Dermatol., 2011, 165, 1051–1057.
91 R. W. Miller and C. S. Rabkin, Merkel cell carcinoma andmelanoma: etiological similarities and differences, CancerEpidemiol. Biomarkers Prev., 1999, 8, 153–158.
92 N. C. Hodgson, Merkel cell carcinoma: changing inci-dence trends, J. Surg. Oncol., 2005, 89, 1–4.
93 R. A. Wolfe, E. C. Roys and R. M. Merion, Trends in organdonation and transplantation in the United States,1999–2008, Am. J. Transplant., 2010, 10, 961–972.
94 R. Arora, Y. Chang and P. S. Moore, MCV and Merkel cellcarcinoma: a molecular success story, Curr. Opin. Virol,2012, 2, 489–498.
95 S. K. Demetriou, K. Ona-Vu, E. M. Sullivan, T. K. Dong,S. W. Hsu and D. H. Oh, Defective DNA repair and cellcycle arrest in cells expressing Merkel cell polyomavirus Tantigen, Int. J. Cancer, 2012, 131, 1818–1827.
96 A. Mogha, A. Fautrel, N. Mouchet, N. Guo, S. Corre,H. Adamski, E. Watier, L. Misery and M. D. Galibert,Merkel cell polyomavirus small T antigen mRNA level is
Perspective Photochemical & Photobiological Sciences
74 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015

increased following in vivo UV-radiation, PLoS One, 2010,5, e11423.
97 S. Bhatia, O. Afanasiev and P. Nghiem, Immunobiology ofMerkel cell carcinoma: implications for immunotherapyof a polyomavirus-associated cancer, Curr. Oncol. Rep.,2011, 13, 488–497.
98 H. U. Bernard, R. D. Burk, Z. Chen, K. van Doorslaer,H. zur Hausen and E. M. de Villiers, Classification ofpapillomaviruses (PVs) based on 189 PV types andproposal of taxonomic amendments, Virology, 2010, 401,70–79.
99 S. F. Farzan, T. Waterboer, J. Gui, H. H. Nelson, Z. Li,K. M. Michael, A. E. Perry, S. K. Spencer, E. Demidenko,A. C. Green, M. Pawlita and M. R. Karagas, Cutaneousalpha, beta and gamma human papillomaviruses inrelation to squamous cell carcinoma of the skin: a popu-lation-based study, Int. J. Cancer, 2013, 133, 1713–1720.
100 C. M. Proby, C. A. Harwood, R. E. Neale, A. C. Green,S. Euvrard, L. Naldi, G. Tessari, M. C. Feltkamp, M. N. deKoning, W. G. Quint, T. Waterboer, M. Pawlita,S. Weissenborn, U. Wieland, H. Pfister, E. Stockfleth,I. Nindl, D. Abeni, J. T. Schegget and J. N. BouwesBavinck, A case-control study of betapapillomavirus infec-tion and cutaneous squamous cell carcinoma in organtransplant recipients, Am. J. Transplant., 2011, 11, 1498–1508.
101 K. Andersson, K. M. Michael, T. Luostarinen,T. Waterboer, R. Gislefoss, T. Hakulinen, O. Forslund,M. Pawlita and J. Dillner, Prospective study of humanpapillomavirus seropositivity and risk of nonmelanomaskin cancer, Am. J. Epidemiol., 2012, 175, 685–695.
102 R. E. Neale, S. Weissenborn, D. Abeni, J. N. Bavinck,S. Euvrard, M. C. Feltkamp, A. C. Green, C. Harwood,M. de Koning, L. Naldi, I. Nindl, M. Pawlita, C. Proby,W. G. Quint, T. Waterboer, U. Wieland and H. Pfister,Human papillomavirus load in eyebrow hair follicles andrisk of cutaneous squamous cell carcinoma, Cancer Epide-miol. Biomarkers Prev., 2013, 22, 719–727.
103 J. N. Bouwes Bavinck, M. Feltkamp, L. Struijk and J. terSchegget, Human papillomavirus infection and skincancer risk in organ transplant recipients, J. Invest. Derma-tol. Symp. Proc., 2001, 6, 207–211.
104 N. J. Lowe, D. P. Meyers, J. M. Wieder, D. Luftman, T. Borget,M. D. Lehman, A. W. Johnson and I. R. Scott, Low doses ofrepetitive ultraviolet A induce morphologic changes inhuman skin, J. Invest. Dermatol., 1995, 105, 739–743.
105 E. C. Naylor, R. E. Watson and M. J. Sherratt, Molecularaspects of skin ageing, Maturitas, 2011, 69, 249–256.
106 A. Tewari, C. Lahmann, R. Sarkany, J. Bergemann andA. R. Young, Human erythema and matrix metalloprotei-nase-1 mRNA induction, in vivo, share an action spectrumwhich suggests common chromophores, Photochem.Photobiol. Sci., 2012, 11, 216–223.
107 V. M. Sheth and A. G. Pandya, Melasma: a comprehensiveupdate: part I, J. Am. Acad. Dermatol, 2011, 65, 689–697;quiz 698.
108 R. Jobanputra and M. Bachmann, The effect of skin dis-eases on quality of life in patients from different socialand ethnic groups in Cape Town, South Africa,Int. J. Cancer, 2000, 39, 826–831.
109 A. C. Handel, P. B. Lima, V. M. Tonolli, L. D. Miot andH. A. Miot, Risk factors for facial melasma in women: acase-control study, Br. J. Dermatol., 2014, 171(3), 588–94.
110 M. de Paula Correa and L. C. Pires, Doses of erythemalultraviolet radiation observed in Brazil, Int. J. Dermatol.,2013, 52, 966–973.
111 A. Sivayathorn, Melasma in Orientals, Clin. Drug Invest.,1995, 10, 34–40.
112 C. Guinot, S. Cheffai, J. Latreille, M. A. Dhaoui, S. Youssef,K. Jaber, O. Nageotte and N. Doss, Aggravating factors formelasma: a prospective study in 197 Tunisian patients,J. Eur. Acad. Dermatol. Venereol., 2010, 24, 1060–1069.
113 W. C. Chou, M. Takeo, P. Rabbani, H. Hu, W. Lee,Y. R. Chung, J. Carucci, P. Overbeek and M. Ito, Directmigration of follicular melanocyte stem cells to the epi-dermis after wounding or UVB irradiation is dependenton Mc1r signaling, Nat. Med., 2013, 19, 924–929.
114 R. Hernandez-Barrera, B. Torres-Alvarez, J. P. Castanedo-Cazares, C. Oros-Ovalle and B. Moncada, Solar elastosisand presence of mast cells as key features in the patho-genesis of melasma, Clin. Exp. Dermatol., 2008, 33, 305–308.
115 M. B. Rozanowska, Light-induced damage to the retina:current understanding of the mechanisms and unresolvedquestions: a symposium-in-print, Photochem. Photobiol.,2012, 88, 1303–1308.
116 J. M. Artigas, A. Felipe, A. Navea, A. Fandino andC. Artigas, Spectral transmission of the human crystallinelens in adult and elderly persons: color and total trans-mission of visible light, Invest. Ophthalmol. Visual. Sci.,2012, 53, 4076–4084.
117 A. P. Cullen, Photokeratitis and other phototoxic effectson the cornea and conjunctiva, Int. J. Toxicol., 2002, 21,455–464.
118 C. Cejka, J. Luyckx and J. Cejkova, Central corneal thick-ness considered an index of corneal hydration of the UVBirradiated rabbit cornea as influenced by UVB absorber,Physiol. Res., 2012, 61, 299–306.
119 J. Cejkova, C. Cejka and J. Luyckx, Trehalose treatmentaccelerates the healing of UVB-irradiated corneas. Com-parative immunohistochemical studies on corneal cryostatsections and corneal impression cytology, Histol. Histo-pathol., 2012, 27, 1029–1040.
120 A. T. Black, M. K. Gordon, D. E. Heck, M. A. Gallo,D. L. Laskin and J. D. Laskin, UVB light regulatesexpression of antioxidants and inflammatory mediators inhuman corneal epithelial cells, Biochem. Pharmacol., 2011,81, 873–880.
121 J. J. Doutch, A. J. Quantock, N. C. Joyce and K. M. Meek,Ultraviolet light transmission through the human cornealstroma is reduced in the periphery, Biophys. J., 2012, 102,1258–1264.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 75

122 J. C. Sherwin, A. W. Hewitt, L. S. Kearns, L. R. Griffiths,D. A. Mackey and M. T. Coroneo, The association betweenpterygium and conjunctival ultraviolet autofluorescence:The Norfolk Island Eye Study, Acta Ophthalmol., 2013,91(4), 363–370.
123 R. Asokan, R. S. Venkatasubbu, L. Velumuri, V. Lingamand R. George, Prevalence and associated factors forpterygium and pinguecula in a South Indian population,Ophthalmic Physiol. Opt., 2012, 32, 39–44.
124 J. Landers, T. Henderson and J. Craig, Prevalence of ptery-gium in indigenous Australians within central Australia:the Central Australian Ocular Health Study, Clin. Exp.Ophthalmol., 2011, 39, 604–606.
125 J. C. Sherwin, A. W. Hewitt, L. S. Kearns, L. R. Griffiths,D. A. Mackey and M. T. Coroneo, The association betweenpterygium and conjunctival ultraviolet autofluorescence:the Norfolk Island Eye Study, Acta Ophthalmol., 2013, 91,363–370.
126 D. M. Taylor, D. Bennett, M. Carter, D. Garewal andC. F. Finch, Acute injury and chronic disability resultingfrom surfboard riding, J. Sci. Med. Sport., 2004, 7, 429–437.
127 H. R. Taylor, S. West, B. Munoz, F. S. Rosenthal,S. B. Bressler and N. M. Bressler, The long-term effects ofvisible light on the eye, Arch. Ophthalmol., 1992, 110, 99–104.
128 T. Golu, L. Mogoanta, C. T. Streba, D. N. Pirici,D. Malaescu, G. O. Mateescu and G. Mutiu, Pterygium:histological and immunohistochemical aspects,Rom. J. Morphol. Embryol., 2011, 52, 153–158.
129 P. Artornsombudh, A. Sanpavat, U. Tinnungwattana,V. Tongkhomsai, L. Sansopha and W. Tulvatana, Preva-lence and clinicopathologic findings of conjunctival epi-thelial neoplasia in pterygia, Ophthalmol., 2013, 120,1337–1340.
130 P. Oellers, C. L. Karp, A. Sheth, A. A. Kao, A. Abdelaziz,J. L. Matthews, S. R. Dubovy and A. Galor, Prevalence,treatment, and outcomes of coexistent ocular surfacesquamous neoplasia and pterygium, Ophthalmol., 2013,120, 445–450.
131 B. Emmanuel, E. Ruder, S. W. Lin, C. Abnet,A. Hollenbeck and S. Mbulaiteye, Incidence of squamous-cell carcinoma of the conjunctiva and other eye cancers inthe NIH-AARP Diet and Health Study, Ecancermedi-calscience, 2012, 6, 254.
132 N. Di Girolamo, Association of human papilloma viruswith pterygia and ocular-surface squamous neoplasia,Eye, 2012, 26, 202–211.
133 R. Bourne, G. A. Stevens, R. White, J. Smith, S. Flaxman,H. Price, J. B. Jonas, J. Keeffe, J. Leasher, K. Naidoo,K. Pesudovs, S. Resnikoff, H. Taylor and Vision LossExpert Group, Causes of vision loss worldwide,1990–2010: a systematic analysis, Lancet Global. Health,2013, 1, e339–e349.
134 H. R. Taylor, S. K. West, F. S. Rosenthal, B. Munoz,H. S. Newland, H. Abbey and E. A. Emmett, Effect of ultra-violet radiation on cataract formation, N. Engl. J. Med.,1988, 319, 1429–1433.
135 T. Okuno, T. Nakanishi-Ueda, T. Ueda, H. Yasuhara andR. Koide, Ultraviolet action spectrum for cell killing ofprimary porcine lens epithelial cells, J. Occup. Health,2012, 54, 181–186.
136 V. C. Mody, Jr., M. Kakar, P. G. Soderberg and S. Lofgren,High lenticular tolerance to ultraviolet radiation-B bypigmented guinea-pig; application of a safety limit strat-egy for UVR-induced cataract, Acta Ophthalmol., 2012, 90,226–230.
137 S. Lofgren, R. Michael and P. G. Soderberg, Impact of irispigment and pupil size in ultraviolet radiation cataract inrat, Acta Ophthalmol., 2012, 90, 44–48.
138 Y. Ishikawa, K. Hashizume, S. Kishimoto, Y. Tezuka,H. Nishigori, N. Yamamoto, Y. Kondo, N. Maruyama,A. Ishigami and D. Kurosaka, Effect of vitamin Cdepletion on UVR-B induced cataract in SMP30/GNLknockout mice, Exp. Eye Res., 2012, 94, 85–89.
139 M. Kronschlager, K. Galichanin, J. Ekstrom, M. F. Lou andP. G. Soderberg, Protective effect of the thioltransferasegene on in vivo UVR-300 nm-induced cataract, Invest.Ophthalmol. Visual Sci., 2012, 53, 248–252.
140 J. Zhang, H. Yan, S. Lofgren, X. Tian and M. F. Lou, Ultra-violet radiation-induced cataract in mice: the effect of ageand the potential biochemical mechanism, Invest.Ophthalmol. Visual Sci., 2012, 53, 7276–7285.
141 P. Storey, B. Munoz, D. Friedman and S. West, Racialdifferences in lens opacity incidence and progression: theSalisbury Eye Evaluation (SEE) study, Invest. Ophthalmol.Visual Sci., 2013, 54, 3010–3018.
142 B. E. Klein, K. E. Lee, L. G. Danforth, T. M. Schaich,K. J. Cruickshanks and R. Klein, Selected sun-sensitizingmedications and incident cataract, Arch. Ophthalmol.,2010, 128, 959–963.
143 J. C. Yam and A. K. Kwok, Ultraviolet light and ocular dis-eases, Int. Ophthalmol., 2014, 34, 383–400.
144 I. Franco-Lie, T. Iversen, T. E. Robsahm andM. Abdelnoor, Incidence trends of melanoma of the skincompared with other localisations, in the Norwegianpopulation, 1956–2005, Ann. Oncol., 2011, 22, 1443–1450.
145 V. P. Papastefanou and V. M. Cohen, Uveal melanoma,J. Skin Cancer, 2011, 2011, 573974.
146 J. D. Mallet, S. P. Gendron, M. C. Drigeard Desgarnier andP. J. Rochette, Implication of ultraviolet light in the etio-logy of uveal melanoma: a review, Photochem. Photobiol.,2014, 90(1), 15–21.
147 B. P. Brooks, A. H. Thompson, R. J. Bishop, J. A. Clayton,C. C. Chan, E. T. Tsilou, W. M. Zein, D. Tamura,S. G. Khan, T. Ueda, J. Boyle, K. S. Oh, K. Imoto, H. Inui,S. Moriwaki, S. Emmert, N. T. Iliff, P. Bradford,J. J. Digiovanna and K. H. Kraemer, Ocular Manifestationsof Xeroderma Pigmentosum: Long-Term Follow-up High-lights the Role of DNA Repair in Protection from SunDamage, Ophthalmol., 2013, 120, 1324–1336.
148 E. Prokofyeva and E. Zrenner, Epidemiology of major eyediseases leading to blindness in Europe: a literaturereview, Ophthalmic. Res., 2012, 47, 171–188.
Perspective Photochemical & Photobiological Sciences
76 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015

149 S. P. Gendron, N. Bastien, J. D. Mallet and P. J. Rochette,The 3895-bp mitochondrial DNA deletion in the humaneye: a potential involvement in corneal ageing andmacular degeneration, Mutagenesis, 2013, 28, 197–204.
150 A. E. Millen, R. Voland, S. A. Sondel, N. Parekh,R. L. Horst, R. B. Wallace, G. S. Hageman, R. Chappell,B. A. Blodi, M. L. Klein, K. M. Gehrs, G. E. Sarto andJ. A. Mares, Vitamin D status and early age-relatedmacular degeneration in postmenopausal women, Arch.Ophthalmol., 2011, 129, 481–489.
151 G. Y. Sui, G. C. Liu, G. Y. Liu, Y. Y. Gao, Y. Deng,W. Y. Wang, S. H. Tong and L. Wang, Is sunlight exposurea risk factor for age-related macular degeneration? A sys-tematic review and meta-analysis, Br. J. Opthalmol., 2013,97, 389–394.
152 T. H. Margrain, M. Boulton, J. Marshall and D. H. Sliney,Do blue light filters confer protection against age-relatedmacular degeneration?, Prog. Retinal Eye Res., 2004, 23,523–531.
153 S. K. West, F. S. Rosenthal, N. M. Bressler, S. B. Bressler,B. Munoz, S. L. Fine and H. R. Taylor, Exposure to sun-light and other risk factors for age-related maculardegeneration, Arch. Ophthalmol., 1989, 107, 875–879.
154 J. A. Guggenheim, K. Northstone, G. McMahon, A. R. Ness,K. Deere, C. Mattocks, B. S. Pourcain and C. Williams,Time outdoors and physical activity as predictors of inci-dent myopia in childhood: a prospective cohort study,Invest. Ophthalmol. Visual Sci., 2012, 53, 2856–2865.
155 Y. Guo, L. J. Liu, L. Xu, P. Tang, Y. Y. Lv, Y. Feng, M. Mengand J. B. Jonas, Myopic Shift and Outdoor Activity amongPrimary School Children: One-Year Follow-Up Study inBeijing, PLoS One, 2013, 8, e75260.
156 Y. Guo, L. J. Liu, L. Xu, Y. Y. Lv, P. Tang, Y. Feng, M. Mengand J. B. Jonas, Outdoor activity and myopia amongprimary students in rural and urban regions of Beijing,Ophthalmol., 2013, 120, 277–283.
157 J. C. Sherwin, A. W. Hewitt, M. T. Coroneo, L. S. Kearns,L. R. Griffiths and D. A. Mackey, The association betweentime spent outdoors and myopia using a novel biomarkerof outdoor light exposure, Invest. Ophthalmol. Visual Sci.,2012, 53, 4363–4370.
158 M. Nebbioso, A. M. Plateroti, B. Pucci and N. Pescosolido,Role of the Dopaminergic System in the Development ofMyopia in Children and Adolescents, J. Child Neurol., 2014,pii: 0883073814538666.
159 S. Yazar, A. W. Hewitt, L. J. Black, C. M. McKnight,J. A. Mountain, J. C. Sherwin, W. H. Oddy, M. T. Coroneo,R. M. Lucas and D. A. Mackey, Myopia is associated withlower vitamin D status in young adults, Invest. Ophthalmol.Visual Sci., 2014, 55, 4552–4559.
160 K. Hiramoto, Y. Yamate, H. Kobayashi and M. Ishii, Long-term ultraviolet A irradiation of the eye induces photo-aging of the skin in mice, Arch. Dermatol. Res., 2012, 304,39–45.
161 K. Hiramoto, Y. Yamate, H. Kobayashi, M. Ishii, E. F. Satoand M. Inoue, Ultraviolet B irradiation of the mouse eye
induces pigmentation of the skin more strongly than doesstress loading, by increasing the levels of prohormoneconvertase 2 and alpha-melanocyte-stimulating hormone,Clin. Exp. Dermatol., 2013, 38, 71–76.
162 A. Tewari, M. M. Grage, G. I. Harrison, R. Sarkany andA. R. Young, UVA1 is skin deep: molecular and clinicalimplications, Photochem. Photobiol. Sci., 2013, 12, 95–103.
163 N. K. Gibbs and M. Norval, Photoimmunosuppression: abrief overview, Photodermatol. Photoimmunol. Photomed.,2013, 29, 57–64.
164 D. L. Damian, Y. J. Matthews, T. A. Phan andG. M. Halliday, An action spectrum for ultravioletradiation-induced immunosuppression in humans,Br. J. Dermatol., 2011, 164, 657–659.
165 Y. J. Matthews, G. M. Halliday, T. A. Phan andD. L. Damian, Wavelength dependency for UVA-inducedsuppression of recall immunity in humans, J. Dermatol.Sci., 2010, 59, 192–197.
166 T. S. Poon, R. S. Barnetson and G. M. Halliday, Sunlight-induced immunosuppression in humans is initiallybecause of UVB, then UVA, followed by interactive effects,J. Invest. Dermatol., 2005, 125, 840–846.
167 Y. Ibuki, M. Allanson, K. M. Dixon and V. E. Reeve, Radi-ation sources providing increased UVA/UVB ratios attenu-ate the apoptotic effects of the UVB waveband UVA-dose-dependently in hairless mouse skin, J. Invest. Dermatol.,2007, 127, 2236–2244.
168 V. E. Reeve, D. Domanski and M. Slater, Radiation sourcesproviding increased UVA/UVB ratios induce photoprotec-tion dependent on the UVA dose in hairless mice, Photo-chem. Photobiol., 2006, 82, 406–411.
169 T. Schwarz and A. Schwarz, Molecular mechanisms ofultraviolet radiation-induced immunosuppression,Eur. J. Cell Biol., 2011, 90, 560–564.
170 S. E. Ullrich and S. N. Byrne, The immunologic revolution:photoimmunology, J. Invest. Dermatol., 2012, 132, 896–905.
171 R. Glaser, F. Navid, W. Schuller, C. Jantschitsch, J. Harder,J. M. Schroder, A. Schwarz and T. Schwarz, UV-B radiationinduces the expression of antimicrobial peptides inhuman keratinocytes in vitro and in vivo, Invest. Ophthal-mol. Visual Sci., 2009, 123, 1117–1123.
172 M. K. Crispin, J. Fuentes-Duculan, N. Gulati,L. M. Johnson-Huang, T. Lentini, M. Sullivan-Whalen,P. Gilleaudeau, I. Cueto, M. Suarez-Farinas, M. A. Lowesand J. G. Krueger, Gene profiling of narrowband UVB-induced skin injury defines cellular and molecular innateimmune responses, J. Invest. Dermatol., 2013, 133, 692–701.
173 F. Navid, M. Boniotto, C. Walker, K. Ahrens, E. Proksch,T. Sparwasser, W. Muller, T. Schwarz and A. Schwarz,Induction of regulatory T cells by a murine beta-defensin,J. Immunol., 2012, 188, 735–743.
174 R. L. Ng, N. M. Scott, D. H. Strickland, S. Gorman,M. A. Grimbaldeston, M. Norval, J. Waithman andP. H. Hart, Altered immunityand dendritic cell activity
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 77

in the periphery of mice after long-term engraftmentwith bone marrow from ultraviolet-irradiated mice,J. Immunol., 2013, 190, 5471–5484.
175 H. M. McGee, R. C. Malley, H. K. Muller andG. M. Woods, Neonatal exposure to UVR alters skinimmune system development, and suppresses immunityin adulthood, Immunol. Cell. Biol., 2011, 89, 767–776.
176 P. H. Hart and S. Gorman, Exposure to UV Wavelengths inSunlight Suppresses Immunity. To What Extent is UV-induced Vitamin D3 the Mediator Responsible?, Clin.Biochem. Rev., 2013, 34, 3–13.
177 A. Schwarz, F. Navid, T. Sparwasser, B. E. Clausen andT. Schwarz, 1,25-dihydroxyvitamin D exerts similar immu-nosuppressive effects as UVR but is dispensable for localUVR-induced immunosuppression, J. Invest. Dermatol.,2012, 132, 2762–2769.
178 M. Baarnhielm, A. K. Hedstrom, I. Kockum, E. Sundqvist,S. A. Gustafsson, J. Hillert, T. Olsson and L. Alfredsson,Sunlight is associated with decreased multiple sclerosisrisk: no interaction with human leukocyte antigen-DRB1*15, Eur. J. Neurol., 2012, 19, 955–962.
179 H. van der Rhee, J. W. Coebergh and E. de Vries, Is pre-vention of cancer by sun exposure more than just theeffect of vitamin D? A systematic review of epidemiologi-cal studies, Eur. J. Cancer. Prev., 2013, 49, 1422–1436.
180 S. V. Milliken, H. Wassall, B. J. Lewis, J. Logie,R. N. Barker, H. Macdonald, M. A. Vickers andA. D. Ormerod, Effects of ultraviolet light on humanserum 25-hydroxyvitamin D and systemic immune func-tion, J. Allergy Clin. Immunol., 2012, 129, 1554–1561.
181 V. K. Sharma, K. Sahni and A. R. Wadhwani, Photoderma-toses in pigmented skin, Photochem. Photobiol. Sci., 2013,12, 65–77.
182 A. R. Wadhwani, V. K. Sharma, M. Ramam andB. K. Khaitan, A clinical study of the spectrum of photo-dermatoses in dark-skinned populations, Clin. Exp. Der-matol., 2013, 38, 823–829.
183 P. Wolf, A. Gruber-Wackernagel, I. Bambach,U. Schmidbauer, G. Mayer, M. Absenger, E. Frohlich andS. N. Byrne, Photohardening of polymorphic lighteruption patients decreases baseline epidermal Langer-hans cell density while increasing mast cell numbersin the papillary dermis, Exp. Dermatol., 2014, 23, 428–430.
184 P. Wolf, A. Gruber-Wackernagel, B. Rinner, A. Griesbacher,K. Eberhard, A. Groselj-Strele, G. Mayer, R. E. Stauber andS. N. Byrne, Phototherapeutic hardening modulates sys-temic cytokine levels in patients with polymorphic lighteruption, Photochem. Photobiol. Sci., 2013, 12, 166–173.
185 T. Gambichler, S. Terras, P. Kampilafkos, A. Kreuter andM. Skrygan, T regulatory cells and related immunoregula-tory factors in polymorphic light eruption following ultra-violet A1 challenge, Br. J. Dermatol., 2013, 169, 1288–1294.
186 A. Gruber-Wackernagel, S. N. Byrne and P. Wolf, Polymor-phous light eruption: clinic aspects and pathogenesis,Dermatol. Clin., 2014, 32, 315–334.
187 G. Krstic, Asthma prevalence associated with geogra-phical latitude and regional insolation in the UnitedStates of America and Australia, PLoS One, 2011, 6,e18492.
188 A. M. Hughes, R. M. Lucas, A. L. Ponsonby, C. Chapman,A. Coulthard, K. Dear, T. Dwyer, T. J. Kilpatrick,A. J. McMichael, M. P. Pender, B. V. Taylor, P. Valery,I. A. van der Mei and D. Williams, The role of latitude,ultraviolet radiation exposure and vitamin D in childhoodasthma and hayfever: an Australian multicenter study,Pediatr. Allergy Immunol., 2011, 22, 327–333.
189 V. Oktaria, S. C. Dharmage, J. A. Burgess, J. A. Simpson,S. Morrison, G. G. Giles, M. J. Abramson, E. H. Waltersand M. C. Matheson, Association between latitude andallergic diseases: a longitudinal study from childhoodto middle-age, Ann. Allergy Asthma Immunol., 2013, 110,80–85.e1.
190 M. Norval, J. Garssen, H. Van Loveren and A. A. el-Ghorr,UV-induced changes in the immune response tomicrobial infections in human subjects and animalmodels, J. Epidemiol, 1999, 9, S84–S92.
191 C. Ludema, S. R. Cole, C. Poole, J. S. Smith,V. J. Schoenbach and K. R. Wilhelmus, AssociationBetween Unprotected Ultraviolet Radiation Exposureand Recurrence of Ocular Herpes Simplex Virus,Am. J. Epidemiol., 2013, 179(2), 208–215.
192 M. Norval and G. M. Woods, UV-induced immunosup-pression and the efficacy of vaccination, Photochem. Photo-biol. Sci., 2011, 10, 1267–1274.
193 P. H. Hart, S. Gorman and J. J. Finlay-Jones, Modulationof the immune system by UV radiation: more than just theeffects of vitamin D?, Nat. Rev. Immunol., 2011, 11, 584–596.
194 V. Nerich, P. Jantchou, M. C. Boutron-Ruault, E. Monnet,A. Weill, V. Vanbockstael, G. R. Auleley, C. Balaire,P. Dubost, S. Rican, H. Allemand and F. Carbonnel, Lowexposure to sunlight is a risk factor for Crohn’s disease,Aliment. Pharmacol. Ther., 2011, 33, 940–945.
195 A. L. Ponsonby, R. M. Lucas and I. A. van der Mei, UVR,Vitamin D and three autoimmune diseases - MultipleSclerosis, Type 1 Diabetes, Rheumatoid Arthritis, Photo-chem. Photobiol., 2005, 81, 1267–1275.
196 P. Gatenby, R. Lucas and A. Swaminathan, Vitamin Ddeficiency and risk for rheumatic diseases: an update,Curr. Opin. Rheumatol., 2013, 25, 184–191.
197 S. Hewer, R. Lucas, I. van der Mei and B. V. Taylor,Vitamin D and multiple sclerosis, J. Clin. Neurosci., 2013,20, 634–641.
198 M. A. Hernan, M. J. Olek and A. Ascherio, Geographic vari-ation of MS incidence in two prospective studies of USwomen, Neurology, 1999, 53, 1711–1718.
199 A. Ascherio and K. L. Munger, Environmental risk factorsfor multiple sclerosis. Part I: the role of infection, Ann.Neurol., 2007, 61, 288–299.
200 C. J. Willer, D. A. Dyment, A. D. Sadovnick,P. M. Rothwell, T. J. Murray and G. C. Ebers, Timing of
Perspective Photochemical & Photobiological Sciences
78 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015

birth and risk of multiple sclerosis: population basedstudy, Br. Med. J., 2005, 330, 120.
201 J. Staples, A. L. Ponsonby and L. Lim, Low maternalexposure to ultraviolet radiation in pregnancy, month ofbirth, and risk of multiple sclerosis in offspring: longi-tudinal analysis, Br. Med. J., 2010, 340, c1640.
202 B. R. Becklund, K. S. Severson, S. V. Vang andH. F. Deluca, UV radiation suppresses experimental auto-immune encephalomyelitis independent of vitaminD production, Proc. Natl. Acad. Sci. U. S. A., 2010, 107,6418–6423.
203 R. M. Lucas, A. L. Ponsonby, K. Dear, P. C. Valery,M. P. Pender, B. V. Taylor, T. J. Kilpatrick, T. Dwyer,A. Coulthard, C. Chapman, I. van der Mei, D. Williamsand A. J. McMichael, Sun exposure and vitamin D areindependent risk factors for CNS demyelination, Neuro-logy, 2011, 76, 540–548.
204 R. Zivadinov, C. N. Treu, B. Weinstock-Guttman,C. Turner, N. Bergsland, K. O’Connor, M. G. Dwyer,E. Carl, D. P. Ramasamy, J. Qu and M. Ramanathan, Inter-dependence and contributions of sun exposure andvitamin D to MRI measures in multiple sclerosis,J. Neurol. Neurosurg. Psychiatry, 2013.
205 M. T. van Leeuwen, A. C. Webster, M. R. McCredie,J. H. Stewart, S. P. McDonald, J. Amin, J. M. Kaldor,J. R. Chapman, C. M. Vajdic and A. E. Grulich, Effect ofreduced immunosuppression after kidney transplantfailure on risk of cancer: population based retrospectivecohort study, Br. Med. J., 2010, 340, c570.
206 M. R. Zaidi, E. C. De Fabo, F. P. Noonan and G. Merlino,Shedding light on melanocyte pathobiology in vivo,Cancer Res., 2012, 72, 1591–1595.
207 H. Y. Handoko, M. P. Rodero, G. M. Boyle, B. Ferguson,C. Engwerda, G. Hill, H. K. Muller, K. Khosrotehrani andG. J. Walker, UVB-induced melanocyte proliferation inneonatal mice driven by CCR2-independent recruitmentof Ly6c(low)MHCII(hi) macrophages, J. Invest. Dermatol.,2013, 133, 1803–1812.
208 M. R. Zaidi, S. Davis, F. P. Noonan, C. Graff-Cherry,T. S. Hawley, R. L. Walker, L. Feigenbaum, E. Fuchs,L. Lyakh, H. A. Young, T. J. Hornyak, H. Arnheiter,G. Trinchieri, P. S. Meltzer, E. C. De Fabo and G. Merlino,Interferon-gamma links ultraviolet radiation to melano-magenesis in mice, Nature, 2011, 469, 548–553.
209 T. Bald, T. Quast, J. Landsberg, M. Rogava, N. Glodde,D. Lopez-Ramos, J. Kohlmeyer, S. Riesenberg, D. van denBoorn-Konijnenberg, C. Homig-Holzel, R. Reuten,B. Schadow, H. Weighardt, D. Wenzel, I. Helfrich,D. Schadendorf, W. Bloch, M. E. Bianchi, C. Lugassy,R. L. Barnhill, M. Koch, B. K. Fleischmann, I. Forster,W. Kastenmuller, W. Kolanus, M. Holzel, E. Gaffal andT. Tuting, Ultraviolet-radiation-induced inflammation pro-motes angiotropism and metastasis in melanoma, Nature,2014, 507, 109–113.
210 G. Erdag, J. T. Schaefer, M. E. Smolkin, D. H. Deacon,S. M. Shea, L. T. Dengel, J. W. Patterson and
C. L. Slingluff, Jr., Immunotype and immunohistologiccharacteristics of tumor-infiltrating immune cells areassociated with clinical outcome in metastatic melanoma,Cancer Res., 2012, 72, 1070–1080.
211 F. J. Moloney, H. Comber, P. O’Lorcain, P. O’Kelly,P. J. Conlon and G. M. Murphy, A population-basedstudy of skin cancer incidence and prevalence inrenal transplant recipients, Br. J. Dermatol., 2006, 154,498–504.
212 M. Norval and G. M. Halliday, The consequences of UV-induced immunosuppression for human health, Photo-chem. Photobiol., 2011, 87, 965–977.
213 S. M. Thanos, G. M. Halliday and D. L. Damian, Nicotin-amide reduces photodynamic therapy-induced immuno-suppression in humans, Br. J. Dermatol., 2012, 167, 631–636.
214 D. Surjana, G. M. Halliday, A. J. Martin, F. J. Moloney andD. L. Damian, Oral nicotinamide reduces actinic keratosesin phase II double-blinded randomized controlled trials,J. Invest. Dermatol., 2012, 132, 1497–1500.
215 O. N. Galkin and I. P. Terenetskaya, ‘Vitamin D’ biodosi-meter: basic characteristics and potential applications,J. Photochem. Photobiol., B., 1999, 53, 12–19.
216 M. Norval, L. O. Bjorn and F. R. de Gruijl, Is the actionspectrum for the UV-induced production of previtaminD3 in human skin correct?, Photochem. Photobiol. Sci.,2010, 9, 11–17.
217 A. R. Webb, B. R. DeCosta and M. F. Holick, Sunlight regu-lates the cutaneous production of vitamin D3 by causingits photodegradation, J. Clin. Endocrinol. Metab., 1989, 68,882–887.
218 A. R. Webb, R. Kift, J. L. Berry and L. E. Rhodes, Thevitamin D debate: translating controlled experiments intoreality for human sun exposure times, Photochem. Photo-biol., 2011, 87, 741–745.
219 C. T. Sempos, H. Vesper, K. Phinney and L. Thienpont,P. Coates and Vitamin D Standardization Program,Vitamin D status as an international issue: Nationalsurveys and the problem of standardization, Scand. J. Clin.Lab. Invest., 2012, 72, 32–40.
220 C. J. Rosen, S. A. Abrams, J. F. Aloia, P. M. Brannon,S. K. Clinton, R. A. Durazo-Arvizu, J. C. Gallagher,R. L. Gallo, G. Jones, C. S. Kovacs, J. E. Manson,S. T. Mayne, A. C. Ross, S. A. Shapses and C. L. Taylor,IOM committee members respond to Endocrine Societyvitamin D guideline, J. Clin. Endocrinol. Metab., 2012, 97,1146–1152.
221 B. W. Hollis, Circulating 25-hydroxyvitamin D levelsindicative of vitamin D sufficiency: implications for estab-lishing a new effective dietary intake recommendation forvitamin D, J. Nutr., 2005, 135, 317–322.
222 Institute of Medicine, Dietary Reference Intakes forCalcium and Vitamin D, Institute of Medicine of theNational Academies Report No., 2010. http://www.iom.edu/Reports/2010/Dietary-Reference-Intakes-for-Calcium-and-Vitamin-D.aspx.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 79

223 S. H. S. Pearce and T. D. Cheetham, Diagnosis andmanagement of vitamin D deficiency, Br. Med. J., 2010, 340.
224 M. F. Holick, N. C. Binkley, H. A. Bischoff-Ferrari,C. M. Gordon, D. A. Hanley, R. P. Heaney, M. H. Muradand C. M. Weaver, Evaluation, Treatment, and Preventionof Vitamin D Deficiency: an Endocrine Society ClinicalPractice Guideline, J. Clin. Endocrinol. Metab., 2011, 96,1911–1930.
225 C. Palacios and L. Gonzalez, Is vitamin D deficiency amajor global public health problem?, J. Steriod Biochem.Mol. Biol., 2014, 144PA, 138–145.
226 J. Hilger, A. Friedel, R. Herr, T. Rausch, F. Roos,D. A. Wahl, D. D. Pierroz, P. Weber and K. Hoffmann, Asystematic review of vitamin D status in populationsworldwide, Br. J. Nutr., 2014, 111, 23–45.
227 D. A. Wahl, C. Cooper, P. R. Ebeling, M. Eggersdorfer,J. Hilger, K. Hoffmann, R. Josse, J. A. Kanis, A. Mithal,D. D. Pierroz, J. Stenmark, E. Stocklin and B. Dawson-Hughes, A global representation of vitamin D status inhealthy populations, Arch. Osteoporos., 2012, 7, 155–172.
228 R. M. Lucas, A. L. Ponsonby, K. Dear, P. C. Valery,B. Taylor, I. van der Mei, A. J. McMichael, M. P. Pender,C. Chapman, A. Coulthard, T. J. Kilpatrick, J. Stankovich,D. Williams and T. Dwyer, Vitamin D status: multifactorialcontribution of environment, genes and other factors inhealthy Australian adults across a latitude gradient,J. Steriod Biochem. Mol. Biol., 2013, 136, 300–308.
229 R. M. Lucas, P. Valery, I. van der Mei, T. Dwyer,M. P. Pender, B. Taylor, A. L. Ponsonby andG. Ausimmune Investigator, Sun exposure over a lifetimein Australian adults from latitudinally diverse regions,Photochem. Photobiol., 2013, 89, 737–744.
230 A. Bjork, A. Andersson, G. Johansson, K. Bjorkegren,A. Bardel and P. Kristiansson, Evaluation of sun holiday,diet habits, origin and other factors as determinants ofvitamin D status in Swedish primary health care patients:a cross-sectional study with regression analysis of ethnicSwedish and immigrant women, BMC Fam. Pract., 2013,14, 129.
231 C. E. Powe, M. K. Evans, J. Wenger, A. B. Zonderman,A. H. Berg, M. Nalls, H. Tamez, D. Zhang, I. Bhan,S. A. Karumanchi, N. R. Powe and R. Thadhani, VitaminD-binding protein and vitamin D status of black Ameri-cans and white Americans, N. Engl. J. Med., 2013, 369,1991–2000.
232 A. M. Renzaho, J. A. Halliday and C. Nowson, Vitamin D,obesity, and obesity-related chronic disease among ethnicminorities: a systematic review, Nutrition, 2011, 27, 868–879.
233 R. Kift, J. L. Berry, A. Vail, M. T. Durkin, L. E. Rhodes andA. R. Webb, Lifestyle factors including less cutaneous sunexposure contribute to starkly lower vitamin D status inUK South Asians compared to the white Caucasian popu-lation, Br. J. Dermatol., 2013, 169(6), 1272–1278.
234 I. Bhan, C. E. Powe, A. H. Berg, E. Ankers, J. B. Wenger,S. A. Karumanchi and R. I. Thadhani, Bioavailable
vitamin D is more tightly linked to mineral metabolismthan total vitamin D in incident hemodialysis patients,Kidney International, 2012, 82, 84–89.
235 C. E. Powe, C. Ricciardi, A. H. Berg, D. Erdenesanaa,G. Collerone, E. Ankers, J. Wenger, S. A. Karumanchi,R. Thadhani and I. Bhan, Vitamin D-binding proteinmodifies the vitamin D-bone mineral density relationship,J. Bone Miner. Res., 2011, 26, 1609–1616.
236 F. Libon, E. Cavalier and A. F. Nikkels, Skin color is rele-vant to vitamin D synthesis, Dermatol., 2013, 227, 250–254.
237 P. Springbett, S. Buglass and A. R. Young, Photoprotectionand vitamin D status, J. Photochem. Photobiol., B, 2010,101, 160–168.
238 M. K. Bogh, A. V. Schmedes, P. A. Philipsen, E. Thiedenand H. C. Wulf, Vitamin D production after UVB exposuredepends on baseline vitamin D and total cholesterol butnot on skin pigmentation, J. Invest. Dermatol., 2010, 130,546–553.
239 L. O. Bjorn, Vitamin D synthesis may be independent ofskin pigmentation only with UV of short wavelength,J. Invest. Dermatol., 2010, 130, 2848–2850.
240 M. K. Bogh, A. V. Schmedes, P. A. Philipsen, E. Thiedenand H. C. Wulf, Vitamin D production depends on ultra-violet-B dose but not on dose rate: a randomized con-trolled trial, Exp. Dermatol., 2011, 20, 14–18.
241 R. McKenzie, R. Scragg, B. Liley, P. Johnston, J. Wishart,A. Stewart and R. Prematunga, Serum 25-hydroxyvitamin-D responses to multiple UV exposures from solaria: infer-ences for exposure to sunlight, Photochem. Photobiol. Sci.,2012, 11, 1174–1185.
242 L. E. Rhodes, A. R. Webb, H. I. Fraser, R. Kift,M. T. Durkin, D. Allan, S. J. O’Brien, A. Vail andJ. L. Berry, Recommended summer sunlight exposurelevels can produce sufficient (≥20 ng ml−1) but not theproposed optimal (≥32 ng ml−1) 25(OH)D levels at UK lati-tudes°, J. Invest. Dermatol., 2010, 130, 1411–1418.
243 A. C. Porojnicu, O. S. Bruland, L. Aksnes, W. B. Grant andJ. Moan, Sun beds and cod liver oil as vitamin D sources,J. Photochem. Photobiol., B, 2008, 91, 125–131.
244 J. Moan, Z. Lagunova, E. Cicarma, L. Aksnes, A. Dahlback,W. B. Grant and A. C. Porojnicu, Sunbeds as vitamin Dsources, Photochem. Photobiol., 2009, 85, 1474–1479.
245 M. D. Farrar, A. R. Webb, R. Kift, M. T. Durkin, D. Allan,A. Herbert, J. L. Berry and L. E. Rhodes, Efficacy of a doserange of simulated sunlight exposures in raising vitaminD status in South Asian adults: implications for targetedguidance on sun exposure, Am. J. Clin. Nutr., 2013, 97,1210–1216.
246 M. G. Kimlin, R. M. Lucas, S. L. Harrison, I. van der Mei,B. K. Armstrong, D. C. Whiteman, A. Kricker, M. Nowak,A. M. Brodie and J. Sun, The contributions of solar ultra-violet radiation exposure and other determinants toserum 25-hydroxyvitamin D concentrations in Australianadults: The AusD Study, Am. J. Epidemiol., 2014, 179(7),864–874.
Perspective Photochemical & Photobiological Sciences
80 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015

247 M. K. Bogh, A. V. Schmedes, P. A. Philipsen, E. Thiedenand H. C. Wulf, Interdependence between body surfacearea and ultraviolet B dose in vitamin D production: a ran-domized controlled trial, Br. J. Dermatol., 2011, 164, 163–169.
248 B. Petersen, H. C. Wulf, M. Triguero-Mas, P. A. Philipsen,E. Thieden, P. Olsen, J. Heydenreich, P. Dadvand,X. Basagana, T. S. Liljendahl, G. I. Harrison, D. Segerback,A. W. Schmalwieser, A. R. Young and M. J. Nieuwenhuijsen,Sun and ski holidays improve vitamin D status, but areassociated with high levels of DNA damage, J. Invest. Der-matol., 2014, 134(11), 2806–2813.
249 R. McKenzie, B. Liley, P. Johnston, R. Scragg, A. Stewart,A. I. Reeder and M. W. Allen, Small doses from artificialUV sources elucidate the photo-production of vitamin D,Photochem. Photobiol. Sci., 2013, 12, 1726–1737.
250 A. R. Webb, R. Kift, M. T. Durkin, S. J. O’Brien, A. Vail,J. L. Berry and L. E. Rhodes, The role of sunlight exposurein determining the vitamin D status of the U.K. whiteadult population, Br. J. Dermatol., 2010, 163, 1050–1055.
251 P. Autier, M. Boniol, C. Pizot and P. Mullie, Vitamin Dstatus and ill health: a systematic review, Lancet Diab.Endocrinol., 2014, 2, 76–89.
252 E. Theodoratou, I. Tzoulaki, L. Zgaga and J. P. Ioannidis,Vitamin D and multiple health outcomes: umbrella reviewof systematic reviews and meta-analyses of observationalstudies and randomised trials, Br. Med. J., 2014, 348,g2035.
253 J. H. White, Regulation of intracrine production of 1,25-dihydroxyvitamin D and its role in innate immunedefense against infection, Arch. Biochem. Biophys., 2012,523, 58–63.
254 M. Hewison, Vitamin D and immune function: An over-view, Proc. Nutrit. Soc., 2012, 71, 50–61.
255 S. Christakos, M. Hewison, D. G. Gardner, C. L. Wagner,I. N. Sergeev, E. Rutten, A. G. Pittas, R. Boland, L. Ferrucciand D. D. Bikle, Vitamin D: beyond bone, Ann. N. Y. Acad.Sci., 2013, 1287, 45–58.
256 A. P. Ralph, R. M. Lucas and M. Norval, Vitamin D andsolar ultraviolet radiation in the risk and treatment oftuberculosis, Lancet Infect. Dis., 2013, 13, 77–88.
257 C. Wejse, V. F. Gomes, P. Rabna, P. Gustafson, P. Aaby,I. M. Lisse, P. L. Andersen, H. Glerup and M. Sodemann,Vitamin D as supplementary treatment for tuberculosis: adouble-blind, randomized, placebo-controlled trial,Am. J. Respir Crit. Care Med., 2009, 179, 843–850.
258 A. P. Ralph, G. Waramori, G. J. Pontororing,E. Kenangalem, A. Wiguna, E. Tjitra, Sandjaja,D. B. Lolong, T. W. Yeo, M. D. Chatfield, R. K. Soemanto,I. Bastian, R. Lumb, G. P. Maguire, J. Eisman, R. N. Price,P. S. Morris, P. M. Kelly and N. M. Anstey, L-arginine andvitamin D adjunctive therapies in pulmonary tuberculosis:a randomised, double-blind, placebo-controlled trial,PLoS One, 2013, 8, e70032.
259 A. R. Martineau, P. M. Timms, G. H. Bothamley,Y. Hanifa, K. Islam, A. P. Claxton, G. E. Packe, J. C. Moore-
Gillon, M. Darmalingam, R. N. Davidson, H. J. Milburn,L. V. Baker, R. D. Barker, N. J. Woodward, T. R. Venton,K. E. Barnes, C. J. Mullett, A. K. Coussens,C. M. Rutterford, C. A. Mein, G. R. Davies, R. J. Wilkinson,V. Nikolayevskyy, F. A. Drobniewski, S. M. Eldridge andC. J. Griffiths, High-dose vitamin D(3) during intensive-phase antimicrobial treatment of pulmonary tuberculosis:a double-blind randomised controlled trial, Lancet, 2011,377, 242–250.
260 D. Ganmaa, E. Giovannucci, B. R. Bloom, W. Fawzi,W. Burr, D. Batbaatar, N. Sumberzul, M. F. Holick andW. C. Willett, Vitamin D, tuberculin skin test conversion,and latent tuberculosis in Mongolian school-age children:a randomized, double-blind, placebo-controlled feasibilitytrial, Am. J. Clin. Nutr., 2012, 96, 391–396.
261 K. Bloom-Feshbach, W. J. Alonso, V. Charu, J. Tamerius,L. Simonsen, M. A. Miller and C. Viboud, Latitudinal vari-ations in seasonal activity of influenza and respiratory syn-cytial virus (RSV): a global comparative review, PLoS One,2013, 8, e54445.
262 M. Science, J. L. Maguire, M. L. Russell, M. Smieja,S. D. Walter and M. Loeb, Low serum 25-hydroxyvitaminD level and risk of upper respiratory tract infection in chil-dren and adolescents, Clin. Infect. Dis., 2013, 57, 392–397.
263 A. A. Ginde, J. M. Mansbach and C. A. Camargo, Jr.,Association between serum 25-hydroxyvitamin D level andupper respiratory tract infection in the Third NationalHealth and Nutrition Examination Survey, Arch. Intern.Med., 2009, 169, 384–390.
264 D. J. Berry, K. Hesketh, C. Power and E. Hypponen,Vitamin D status has a linear association with seasonalinfections and lung function in British adults, Br. J. Nutr.,2011, 106, 1433–1440.
265 V. Hirani, Associations between vitamin D and self-reported respiratory disease in older people from a nation-ally representative population survey, J. Am. Geriat. Soc.,2013, 61, 969–973.
266 K. J. Bryson, A. A. Nash and M. Norval, Does vitamin Dprotect against respiratory viral infections?, Epidemiol.Infect., 2014, 142, 1789–1801.
267 M. Urashima, T. Segawa, M. Okazaki, M. Kurihara,Y. Wada and H. Ida, Randomized trial of vitamin D sup-plementation to prevent seasonal influenza A in school-children, Am. J. Clin. Nutr., 2010, 91, 1255–1260.
268 C. A. Camargo, Jr., D. Ganmaa, A. L. Frazier,F. F. Kirchberg, J. J. Stuart, K. Kleinman, N. Sumberzuland J. W. Rich-Edwards, Randomized trial of vitamin Dsupplementation and risk of acute respiratory infection inMongolia, Pediatrics, 2012, 130, e561–e567.
269 J. F. Aloia and M. Li-Ng, Re: epidemic influenza andvitamin D, Epidemiol. Infect., 2007, 135, 1095–1096.
270 I. Laaksi, J. P. Ruohola, V. Mattila, A. Auvinen, T. Ylikomiand H. Pihlajamaki, Vitamin D supplementation for theprevention of acute respiratory tract infection: a random-ized, double-blinded trial among young Finnish men,J. Infect. Dis., 2010, 202, 809–814.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 81

271 B. Tran, B. K. Armstrong, P. R. Ebeling, D. R. English,M. G. Kimlin, J. C. van der Pols, A. Venn, V. Gebski,D. C. Whiteman, P. M. Webb and R. E. Neale, Effectof vitamin D supplementation on antibiotic use: a ran-domized controlled trial, Am. J. Clin. Nutr., 2014, 99, 156–161.
272 M. Li-Ng, J. F. Aloia, S. Pollack, B. A. Cunha, M. Mikhail,J. Yeh and N. Berbari, A randomized controlled trialof vitamin D3 supplementation for the prevention ofsymptomatic upper respiratory tract infections, Epidemiol.Infect., 2009, 137, 1396–1404.
273 J. R. Rees, K. Hendricks, E. L. Barry, J. L. Peacock,L. A. Mott, R. S. Sandler, R. S. Bresalier, M. Goodman,R. M. Bostick and J. A. Baron, Vitamin D3 supplemen-tation and upper respiratory tract infections in a random-ized, controlled trial, Clin. Infect. Dis., 2013, 57, 1384–1392.
274 D. R. Murdoch, S. Slow, S. T. Chambers, L. C. Jennings,A. W. Stewart, P. C. Priest, C. M. Florkowski, J. H. Livesey,C. A. Camargo and R. Scragg, Effect of vitamin D3 sup-plementation on upper respiratory tract infections inhealthy adults: the VIDARIS randomized controlled trial,J. Am. Med. Assoc., 2012, 308, 1333–1339.
275 A. Ascherio, K. L. Munger, R. White, K. Kochert,K. C. Simon, C. H. Polman, M. S. Freedman, H. P. Hartung,D. H. Miller, X. Montalban, G. Edan, F. Barkhof,D. Pleimes, E. W. Radu, R. Sandbrink, L. Kappos andC. Pohl, Vitamin D as an Early Predictor of Multiple Scle-rosis Activity and Progression, J. Am. Med. Assoc. Neurol.,2014, 71(3), 306–314.
276 D. Reid, B. J. Toole, S. Knox, D. Talwar, J. Harten,D. S. O’Reilly, S. Blackwell, J. Kinsella, D. C. McMillan andA. M. Wallace, The relation between acute changes in thesystemic inflammatory response and plasma 25-hydroxy-vitamin D concentrations after elective knee arthroplasty,Am. J. Clin. Nutr., 2011, 93, 1006–1011.
277 R. Lucas and B. Taylor, Challenges in exposure andoutcome definition in neuroepidemiology: the case ofvitamin D and multiple sclerosis, Australas. Epidemiol.,2013, 20, 4–8.
278 G. Bjelakovic, L. L. Gluud, D. Nikolova, K. Whitfield,G. Krstic, J. Wetterslev and C. Gluud, Vitamin D sup-plementation for prevention of cancer in adults, CochraneDatabase Syst. Rev., 2014, 6, CD007469.
279 C. Gordon-Thomson, R. Gupta, W. Tongkao-on, A. Ryan,G. M. Halliday and R. S. Mason, 1alpha,25 dihydroxyvita-min D3 enhances cellular defences against UV-inducedoxidative and other forms of DNA damage in skin, Photo-chem. Photobiol. Sci., 2012, 11, 1837–1847.
280 D. L. Damian, Y. J. Kim, K. M. Dixon, G. M. Halliday,A. Javeri and R. S. Mason, Topical calcitriol protectsfrom UV-induced genetic damage but suppressescutaneous immunity in humans, Exp. Dermatol., 2010, 19,e23–e30.
281 J. Y. Tang, N. Parimi, A. Wu, W. J. Boscardin,J. M. Shikany, M. M. Chren, S. R. Cummings,
E. H. Epstein, Jr., D. C. Bauer and G. OsteoporoticFractures in Men Study, Inverse association betweenserum 25(OH) vitamin D levels and non-melanoma skincancer in elderly men, Cancer Causes Control, 2010, 21,387–391.
282 J. C. van der Pols, A. Russell, U. Bauer, R. E. Neale,M. G. Kimlin and A. C. Green, Vitamin D status and skincancer risk independent of time outdoors: 11-year pro-spective study in an Australian community, J. Invest. Der-matol., 2013, 133, 637–641.
283 S. Afzal, B. G. Nordestgaard and S. E. Bojesen, Plasma25-hydroxyvitamin D and risk of non-melanoma andmelanoma skin cancer: a prospective cohort study,J. Invest. Dermatol., 2013, 133, 629–636.
284 J. M. Major, C. Kiruthu, S. J. Weinstein, R. L. Horst,K. Snyder, J. Virtamo and D. Albanes, Pre-diagnostic circu-lating vitamin D and risk of melanoma in men, PLoS One,2012, 7, e35112.
285 J. Y. Tang, T. Fu, E. Leblanc, J. E. Manson, D. Feldman,E. Linos, M. Z. Vitolins, N. C. Zeitouni, J. Larson andM. L. Stefanick, Calcium plus vitamin D supplementationand the risk of nonmelanoma and melanoma skincancer: post hoc analyses of the women’s health initiativerandomized controlled trial, J. Clin. Oncol., 2011, 29,3078–3084.
286 M. Peterlik, Vitamin D insufficiency and chronic diseases:hype and reality, Food Funct., 2012, 3, 784–794.
287 T. D. Shanafelt, M. T. Drake, M. J. Maurer, C. Allmer,K. G. Rabe, S. L. Slager, G. J. Weiner, T. G. Call, B. K. Link,C. S. Zent, N. E. Kay, C. A. Hanson, T. E. Witzig andJ. R. Cerhan, Vitamin D insufficiency and prognosis inchronic lymphocytic leukemia, Blood, 2011, 117, 1492–1498.
288 K. Zhu, A. J. Whitehouse, P. Hart, M. Kusel, J. Mountain,S. Lye, C. Pennell and J. P. Walsh, Maternal Vitamin DStatus During Pregnancy and Bone Mass in Offspring at20 Years of Age: A Prospective Cohort Study, J. Bone Miner.Res., 2013, 29(5), 1088–1095.
289 A. J. Whitehouse, B. J. Holt, M. Serralha, P. G. Holt,M. M. Kusel and P. H. Hart, Maternal serum vitamin Dlevels during pregnancy and offspring neurocognitivedevelopment, Pediatrics, 2012, 129, 485–493.
290 K. L. Allen, S. M. Byrne, M. M. Kusel, P. H. Hart andA. J. Whitehouse, Maternal vitamin D levels during preg-nancy and offspring eating disorder risk in adolescence,Int. J. Eat. Disord., 2013, 46, 669–676.
291 J. J. McGrath, D. W. Eyles, C. B. Pedersen, C. Anderson,P. Ko, T. H. Burne, B. Norgaard-Pedersen, D. M. Hougaardand P. B. Mortensen, Neonatal vitamin D status and riskof schizophrenia: a population-based case-control study,Arch. Gen. Psychiatry, 2010, 67, 889–894.
292 D. Durup, H. L. Jorgensen, J. Christensen, P. Schwarz,A. M. Heegaard and B. Lind, A Reverse J-Shaped Associ-ation of All-Cause Mortality with Serum 25-Hydroxyvita-min D in General Practice, the CopD Study, J. Clin.Endocrinol. Metab., 2012, 97(8), 2644–2652.
Perspective Photochemical & Photobiological Sciences
82 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015

293 M. L. Melamed, E. D. Michos, W. Post and B. Astor, 25-hydroxyvitamin D levels and the risk of mortality in thegeneral population, Arch. Intern. Med., 2008, 168, 1629–1637.
294 C. T. Sempos, R. A. Durazo-Arvizu, B. Dawson-Hughes,E. A. Yetley, A. C. Looker, R. L. Schleicher, G. Cao, V. Burt,H. Kramer, R. L. Bailey, J. T. Dwyer, X. Zhang, J. Gahche,P. R. Thomas, P. M. Coates and M. F. Picciano, Is there aReverse J-shaped Association between 25-HydroxyvitaminD and All-Cause Mortality? Results from the US NationallyRepresentative NHANES, J. Clin. Endocrinol. Metab., 2013,98, 3001–3009.
295 Y. Dror, S. M. Giveon, M. Hoshen, I. Feldhamer,R. D. Balicer and B. S. Feldman, Vitamin D levels for pre-venting acute coronary syndrome and mortality: evidenceof a nonlinear association, J. Clin. Endocrinol. Metab.,2013, 98, 2160–2167.
296 A. Zittermann, J. Kuhn, J. Dreier, C. Knabbe,J. F. Gummert and J. Borgermann, Vitamin D status andthe risk of major adverse cardiac and cerebrovascularevents in cardiac surgery, Eur. Heart J., 2013, 34, 1358–1364.
297 D. M. Freedman, A. C. Looker, C. C. Abnet, M. S. Linetand B. I. Graubard, Serum 25-hydroxyvitamin D andcancer mortality in the NHANES IIIstudy (1988–2006),Cancer Res., 2010, 70, 8587–8597.
298 K. Michaelsson, J. A. Baron, G. Snellman, R. Gedeborg,L. Byberg, J. Sundstrom, L. Berglund, J. Arnlov, P. Hellman,R. Blomhoff, A. Wolk, H. Garmo, L. Holmberg andH. Melhus, Plasma vitamin D and mortality in older men:a community-based prospective cohort study, Am. J. Clin.Nutr., 2010, 92, 841–848.
299 P. Tuohimaa, L. Tenkanen, M. Ahonen, S. Lumme,E. Jellum, G. Hallmans, P. Stattin, S. Harvei, T. Hakulinen,T. Luostarinen, J. Dillner, M. Lehtinen and M. Hakama,Both high and low levels of blood vitamin D are associ-ated with a higher prostate cancer risk: a longitudinal,nested case-control study in the Nordic countries,Int. J. Cancer, 2004, 108, 104–108.
300 J. Brandstedt, M. Almquist, J. Manjer and J. Malm,Vitamin D, PTH, and calcium and the risk of prostatecancer: a prospective nested case-control study, CancerCauses Control, 2012, 23, 1377–1385.
301 R. Z. Stolzenberg-Solomon, E. J. Jacobs, A. A. Arslan,D. Qi, A. V. Patel, K. J. Helzlsouer, S. J. Weinstein,M. L. McCullough, M. P. Purdue, X. O. Shu, K. Snyder,J. Virtamo, L. R. Wilkins, K. Yu, A. Zeleniuch-Jacquotte,W. Zheng, D. Albanes, Q. Cai, C. Harvey, R. Hayes,S. Clipp, R. L. Horst, L. Irish, K. Koenig, L. Le Marchandand L. N. Kolonel, Circulating 25-hydroxyvitamin D andrisk of pancreatic cancer: Cohort Consortium Vitamin DPooling Project of Rarer Cancers, Am. J. Epidemiol., 2010,172, 81–93.
302 J. Rothers, A. L. Wright, D. A. Stern, M. Halonen andC. A. Camargo, Jr., Cord blood 25-hydroxyvitamin D levelsare associated with aeroallergen sensitization in children
from Tucson, Arizona, J. Allergy. Clin. Immunol., 2011, 128,1093–1099.e5.
303 N. O. Nielsen, T. Skifte, M. Andersson, J. Wohlfahrt,B. Soborg, A. Koch, M. Melbye and K. Ladefoged, Bothhigh and low serum vitamin D concentrations are associ-ated with tuberculosis: a case-control study in Greenland,Br. J. Nutr., 2010, 104, 1487–1491.
304 L. M. Bodnar, J. M. Catov, J. M. Zmuda, M. E. Cooper,M. S. Parrott, J. M. Roberts, M. L. Marazita andH. N. Simhan, Maternal serum 25-hydroxyvitamin D con-centrations are associated with small-for-gestational agebirths in white women, J. Nutr., 2010, 140, 999–1006.
305 K. E. Ensrud, S. K. Ewing, L. Fredman, M. C. Hochberg,J. A. Cauley, T. A. Hillier, S. R. Cummings, K. Yaffe andP. M. Cawthon, Circulating 25-hydroxyvitamin D levelsand frailty status in older women, J. Clin. Endocrinol.Metab., 2010, 95, 5266–5273.
306 A. Bearden, C. Abad, R. Gangnon, J. M. Sosman,N. Binkley and N. Safdar, Cross-sectional study of vitaminD levels, immunologic and virologic outcomes in HIV-infected adults, J. Clin. Endocrinol. Metab., 2013, 98, 1726–1733.
307 Vitamin D Council, Johnson Avenue, San Luis Obispo,California, United States, 93401, http://www.vitamind-council.org, accessed 18 October 2011.
308 H. Bak, S. P. Hong, S. K. Jeong, E. H. Choi, S. E. Lee,S. H. Lee and S. K. Ahn, Altered epidermal lipid layersinduced by long-term exposure to suberythemal-doseultraviolet, Int. J. Dermatol., 2011, 50, 832–837.
309 E. J. Kim, Y. K. Kim, J. E. Kim, S. Kim, M. K. Kim,C. H. Park and J. H. Chung, UV modulation of subcu-taneous fat metabolism, J. Invest. Dermatol., 2011, 131,1720–1726.
310 T. Michikawa, Y. Nishiwaki, K. Asakura, G. Hillebrand,K. Miyamoto, M. Ono, Y. Kinjo, S. Akiba andT. Takebayashi, Sunlight Exposure May Be a Risk Factor ofHearing Impairment: A Community-Based Study in Japa-nese Older Men and Women, J. Gerontol. A Biol. Sci. Med.Sci., 2013, 68(1), 96–103.
311 R. A. Miller, D. Dolan, M. Han, W. Kohler and J. Schacht,Resistance of skin fibroblasts to peroxide and UV damagepredicts hearing loss in aging mice, Aging Cell, 2011, 10,362–363.
312 A. Szilagyi, H. Leighton, B. Burstein and I. Shrier, Signifi-cant positive correlation between sunshine and lactasenonpersistence in Europe may implicate both in similarlyaltering risks for some diseases, Nutr. Cancer, 2011, 63,991–999.
313 V. Nair-Shalliker, D. P. Smith, S. Egger, A. M. Hughes,J. M. Kaldor, M. Clements, A. Kricker and B. K. Armstrong,Sun exposure may increase risk of prostate cancer inthe high UV environment of New South Wales, Australia:a case-control study, Int. J. Cancer, 2012, 131, E726–E732.
314 B. Q. Wang, Z. Y. Shen, Y. Fei, H. Li, J. H. Liu, H. Xu,Z. Zhang, X. H. Yu and X. D. Chen, A population-based
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 83

study of acquired bilateral nevus-of-Ota-like macules inShanghai, China, J. Invest. Dermatol., 2011, 131, 358–362.
315 B. B. Koo, Restless legs syndrome: relationship betweenprevalence and latitude, Sleep Breath., 2012, 16(4), 1237–1245.
316 Y. W. Wirohadidjojo, S. Radiono, A. Budiyanto andH. Soebono, Cellular viability, collagen deposition, andtransforming growth factor beta1 production among ultra-violet B-irradiated keloid fibroblasts, Aesthetic. Plast. Surg.,2011, 35, 1050–1055.
317 D. Liu, B. O. Fernandez, A. Hamilton, N. N. Lang,J. M. Gallagher, D. E. Newby, M. Feelisch and R. B. Weller,UVA Irradiation of Human Skin Vasodilates Arterial Vascu-lature and Lowers Blood Pressure Independently of NitricOxide Synthase, J. Invest. Dermatol., 2014, 134(7), 1839–1846.
318 L. Yang, M. B. Veierod, M. Lof, S. Sandin, H. O. Adamiand E. Weiderpass, Prospective study of UV exposure andcancer incidence among Swedish women, Cancer Epide-miol. Biomarkers Prev., 2011, 20, 1358–1367.
319 P. Brondum-Jacobsen, B. G. Nordestgaard, S. F. Nielsenand M. Benn, Skin cancer as a marker of sun exposureassociates with myocardial infarction, hip fracture anddeath from any cause, Int. J. Epidemiol., 2013, 42, 1486–1496.
320 R. Hoyos-Bachiloglu, P. S. Morales, J. Cerda, E. Talesnik,G. Gonzalez, C. A. Camargo, Jr. and A. Borzutzky, Higherlatitude and lower solar radiation influence on anaphyl-axis in Chilean children, Pediatr. Allergy Immunol., 2014,25, 338–343.
321 S. J. Balk, Ultraviolet radiation: a hazard to children andadolescents, Pediatrics, 2011, 127, e791–e817.
322 I. Galan, A. Rodriguez-Laso, L. Diez-Ganan and E. Camara,Prevalence and correlates of skin cancer risk behaviors inMadrid (Spain), Gac. Sanit., 2011, 25, 44–49.
323 D. B. Buller, V. Cokkinides, H. I. Hall, A. M. Hartman,M. Saraiya, E. Miller, L. Paddock and K. Glanz, Prevalenceof sunburn, sun protection, and indoor tanning behaviorsamong Americans: review from national surveys and casestudies of 3 states, J. Am. Acad. Dermatol., 2011, 65, S114–S123.
324 D. Reinau, C. Meier, N. Gerber, G. F. Hofbauer andC. Surber, Sun protective behaviour of primary and secon-dary school students in North-Western Switzerland, SwissMed. Wkly., 2012, 142, w13520.
325 S. W. Dusza, A. C. Halpern, J. M. Satagopan, S. A. Oliveria,M. A. Weinstock, A. Scope, M. Berwick and A. C. Geller, Pro-spective study of sunburn and sun behavior patterns duringadolescence, Pediatrics, 2012, 129, 309–317.
326 S. Dobbinson, M. Wakefield, D. Hill, A. Girgis, J. F. Aitken,K. Beckmann, A. I. Reeder, N. Herd, M. J. Spittal,A. Fairthorne and K. A. Bowles, Children’s sun exposureand sun protection: Prevalence in Australia and relatedparental factors, J. Am. Acad. Dermatol, 2012, 66, 938–947.
327 C. Battie, M. Gohara, M. Verschoore and W. Roberts, Skincancer in skin of color: an update on current facts, trends,
and misconceptions, J. Drugs Dermatol., 2013, 12, 194–198.
328 S. Ergul and E. Ozeren, Sun protection behavior and indi-vidual risk factors of Turkish Primary School Studentsassociated with skin cancer: a questionnaire-based study,Asian Pac. J. Cancer Prev., 2011, 12, 765–770.
329 F. Mousavi, B. Golestan, M. Vaseie, L. Vaseie andR. Khajeh-Kazemi, Knowledge, attitude, and practice ofadults to the protective actions against sun in northwestTehran, Iran, Arch. Iran. Med., 2011, 14, 126–131.
330 J. Li, W. Uter, A. Pfahlberg and O. Gefeller, A comparisonof patterns of sun protection during beach holidays andeveryday outdoor activities in a population sample ofyoung German children, Br. J. Dermatol., 2012, 166, 803–810.
331 B. Koster, C. Thorgaard, A. Philip and I. H. Clemmensen,Vacations to sunny destinations, sunburn, and intentionto tan: a cross-sectional study in Denmark, 2007–2009,Scand. J. Public Health, 2011, 39, 64–69.
332 T. Tempark, S. Chatproedprai and S. Wanankul, Attitudes,knowledge, and behaviours of secondary school adoles-cents regarding protection from sun exposure: a survey inBangkok, Thailand, Photodermatol. Photoimmunol. Photo-med., 2012, 28, 200–206.
333 C. J. Heckman and J. Cohen-Filipic, Brief Report: Ultra-violet Radiation Exposure, Considering AcculturationAmong Hispanics (Project URECAH), J. Cancer Educ.,2012, 27, 342–346.
334 S. Potente, K. Coppa, A. Williams and R. Engels, Legallybrown: using ethnographic methods to understand sunprotection attitudes and behaviours among young Austra-lians ‘I didn’t mean to get burnt–it just happened!’,Health Educ. Res., 2011, 26, 39–52.
335 Cancer Council Victoria, SunSmart, http://www.sunsmart.com.au, accessed 31 July, 2014.
336 Australian Radiation and Nuclear Safety Agency, Austra-lian UV levels for mobile phones, http://www.arpansa.gov.au/uvindex/realtime/cellrt.htm, accessed 31 July, 2014.
337 K. Dunstone and C. Conway, There is an app for that!Communicating UV via the SunSmart app, in UV radiationand its effects: an update 2014, National Institute for Waterand Atmosphere, Auckland, NZ, 2014.
338 G. R. Casale, A. M. Siani and A. Colosimo, A UV IndexSundial on compact disk, Household Person. Care Today,2014, 9, 45–47.
339 J. Morris, T. Laing-Morton, P. Marno and A. Curnow,An investigation into the awareness and understand-ing of the ultraviolet index forecasts in the South Westof England, Photochem. Photobiol. Sci., 2011, 10, 103–108.
340 B. Rajiv and R. Gray, Sun Protection Alert - a simple toolfor a complex issue, in UV radiation and its effects: anupdate 2014, Auckland, NZ, 2014.
341 EPA, UV Alert, United States Environmental ProtectionAgency, http://www2.epa.gov/sunwise/uv-alert, accessed 11May, 2014.
Perspective Photochemical & Photobiological Sciences
84 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015

342 P. A. Andersen, D. B. Buller, B. J. Walkosz, M. D. Scott,J. A. Maloy, G. R. Cutter and M. D. Dignan, Environmentalcues to UV radiation and personal sun protection inoutdoor winter recreation, Arch. Dermatol., 2010, 146, 1241–1247.
343 D. Vernez, A. Milon, L. Vuilleumier and J. L. Bulliard, Ana-tomical exposure patterns of skin to sunlight: relative con-tributions of direct, diffuse and reflected ultravioletradiation, Br. J. Dermatol., 2012, 167(2), 383–390.
344 A. I. Kudish, M. Harari and E. G. Evseev, The solar ultra-violet B radiation protection provided by shading deviceswith regard to its diffuse component, Photodermatol.Photoimmunol. Photomed., 2011, 27, 236–244.
345 J. C. van der Pols, G. M. Williams, N. Pandeya, V. Loganand A. C. Green, Prolonged prevention of squamous cellcarcinoma of the skin by regular sunscreen use, CancerEpidemiol. Biomarkers Prev., 2006, 15, 2546–2548.
346 A. C. Green, G. M. Williams, V. Logan and G. M. Strutton,Reduced melanoma after regular sunscreen use: random-ized trial follow-up, J. Clin. Oncol., 2011, 29, 257–263.
347 M. C. Hughes, G. M. Williams, P. Baker and A. C. Green,Sunscreen and prevention of skin aging: a randomizedtrial, Ann. Intern. Med., 2013, 158, 781–790.
348 A. Rodrigues, F. F. Sniehotta and V. Araujo-Soares, Areinterventions to promote sun-protective behaviors in rec-reational and tourist settings effective? A systematicreview with meta-analysis and moderator analysis, Ann.Behav. Med., 2013, 45, 224–238.
349 R. Jansen, U. Osterwalder, S. Q. Wang, M. Burnett andH. W. Lim, Photoprotection: Part II. Sunscreen: Develop-ment, efficacy, and controversies, J. Am. Acad. Dermatol,2013, 69, 867 e861–867 e814.
350 A. Fourtanier, D. Moyal and S. Seite, UVA filters in sun-protection products: regulatory and biological aspects,Photochem. Photobiol. Sci., 2012, 11, 81–89.
351 B. L. Diffey and M. W. Brown, The ideal spectral profile oftopical sunscreens, Photochem. Photobiol., 2012, 88, 744–747.
352 A. L. Andrady and H. H. Redhwi, AyakoTorikai,K. K. Pandey and P. Gies, Consequences of stratosphericozone depletion and climate change on the use ofmaterials, Photochem. Photobiol. Sci., 2015, 14, this issue.
353 C. Couteau, C. Chauvet, E. Paparis and L. Coiffard, UVfilters, ingredients with a recognized anti-inflammatoryeffect, PLoS One, 2012, 7, e46187.
354 R. M. Sayre, J. C. Dowdy and E. W. Rosenberg, Sun-protec-tion factor confounded by anti-inflammatory activity ofsunscreen agents?, J. Am. Acad. Dermatol, 2013, 69, 481.
355 B. Petersen, P. Datta, P. A. Philipsen and H. C. Wulf,Sunscreen use and failures–on site observations on a sun-holiday, Photochem. Photobiol. Sci., 2013, 12, 190–196.
356 A. Diaz, R. E. Neale, M. G. Kimlin, L. Jones and M. Janda,The children and sunscreen study: a crossover trial inves-tigating children’s sunscreen application thickness andthe influence of age and dispenser type, Arch. Dermatol.,2012, 148, 606–612.
357 M. Pissavini and B. Diffey, The likelihood of sunburnin sunscreen users is disproportionate to the SPF, Photo-dermatol. Photoimmunol. Photomed., 2013, 29, 111–115.
358 T. Teramura, M. Mizuno, H. Asano, N. Naito, K. Arakaneand Y. Miyachi, Relationship between sun-protectionfactor and application thickness in high-performancesunscreen: double application of sunscreen is rec-ommended, Clin. Exp. Dermatol., 2012, 37, 904–908.
359 A. Faurschou, D. M. Beyer, A. Schmedes, M. K. Bogh,P. A. Philipsen and H. C. Wulf, The relation betweensunscreen layer thickness and vitamin D production afterultraviolet B exposure: a randomized clinical trial,Br. J. Dermatol., 2012, 167, 391–395.
360 E. Linos, E. Keiser, M. Kanzler, K. L. Sainani, W. Lee,E. Vittinghoff, M. M. Chren and J. Y. Tang, Sun protectivebehaviors and vitamin D levels in the US population:NHANES 2003–2006, Cancer Causes Control, 2012, 23,133–140.
361 R. Jansen, S. Q. Wang, M. Burnett, U. Osterwalder andH. W. Lim, Photoprotection: Part I. Photoprotection bynaturally occurring, physical, and systemic agents, J. Am.Acad. Dermatol, 2013, 69, 853 e851–853 e812.
362 A. K. Haylett, Z. Nie, M. Brownrigg, R. Taylor andL. E. Rhodes, Systemic photoprotection in solar urticariawith alpha-melanocyte-stimulating hormone analogue[Nle4-D-Phe7]-alpha-MSH, Br. J. Dermatol., 2011, 164, 407–414.
363 E. A. Langan, Z. Nie and L. E. Rhodes, Melanotropic pep-tides: more than just ‘Barbie drugs’ and ‘sun-tan jabs’?,Br. J. Dermatol., 2010, 163, 451–455.
364 L. Chen, J. Y. Hu and S. Q. Wang, The role of antioxidantsin photoprotection: A critical review, J. Am. Acad. Dermatol,2012, 67(5), 1013–1024.
365 A. C. Chen and D. L. Damian, Nicotinamide and the skin,Australas. J. Dermatol., 2014, 55(3), 169–175.
366 Y. Wu, L. L. Jia, Y. N. Zheng, X. G. Xu, Y. J. Luo, B. Wang,J. Z. Chen, X. H. Gao, H. D. Chen, M. Matsui and Y. H. Li,Resveratrate protects human skin from damage due torepetitive ultraviolet irradiation, J. Eur. Acad. Dermatol.Venereol., 2013, 27(3), 345–350.
367 U. Wolfle, P. R. Esser, B. Simon-Haarhaus, S. F. Martin,J. Lademann and C. M. Schempp, UVB-induced DNAdamage, generation of reactive oxygen species, andinflammation are effectively attenuated by the flavonoidluteolin in vitro and in vivo, Free Radicals Biol. Med., 2011,50, 1081–1093.
368 M. Rizwan, I. Rodriguez-Blanco, A. Harbottle, M. A. Birch-Machin, R. E. Watson and L. E. Rhodes, Tomato pasterich in lycopene protects against cutaneous photodamagein humans in vivo: a randomized controlled trial,Br. J. Dermatol., 2011, 164, 154–162.
369 U. Heinrich, C. E. Moore, S. De Spirt, H. Tronnier andW. Stahl, Green tea polyphenols provide photoprotection,increase microcirculation, and modulate skin propertiesof women, J. Nutr., 2011, 141, 1202–1208.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 85

370 L. E. Rhodes, G. Darby, K. A. Massey, K. A. Clarke,T. P. Dew, M. D. Farrar, S. Bennett, R. E. Watson,G. Williamson and A. Nicolaou, Oral green tea catechinmetabolites are incorporated into human skin and protectagainst UV radiation-induced cutaneous inflammation inassociation with reduced production of pro-inflammatoryeicosanoid 12-hydroxyeicosatetraenoic acid, Br. J. Nutr.,2013, 110, 891–900.
371 Y. R. Lou, Q. Y. Peng, T. Li, C. M. Medvecky, Y. Lin,W. J. Shih, A. H. Conney, S. Shapses, G. C. Wagner andY. P. Lu, Effects of high-fat diets rich in either omega-3or omega-6 fatty acids on UVB-induced skin carcino-genesis in SKH-1 mice, Carcinogenesis, 2011, 32, 1078–1084.
372 J. C. van der Pols, C. Xu, G. M. Boyle, M. C. Hughes,S. J. Carr, P. G. Parsons and A. C. Green, Serum omega-3and omega-6 fatty acids and cutaneous p53 expression inan Australian population, Cancer Epidemiol. BiomarkersPrev., 2011, 20, 530–536.
373 T. DeBoyes, D. Kouba, D. Ozog, E. Fincher, L. Moy,K. Iwata and R. Moy, Reduced number of actinic keratoseswith topical application of DNA repair enzyme creams,J. Drugs Dermatol., 2010, 9, 1519–1521.
374 A. Hofer, F. J. Legat, A. Gruber-Wackernagel,F. Quehenberger and P. Wolf, Topical liposomal DNA-repair enzymes in polymorphic light eruption, Photochem.Photobiol. Sci., 2011, 10, 1118–1128.
375 E. Berardesca, M. Bertona, K. Altabas, V. Altabas andE. Emanuele, Reduced ultraviolet-induced DNA damageand apoptosis in human skin with topical application ofa photolyase-containing DNA repair enzyme cream: cluesto skin cancer prevention, Mol. Med. Rep., 2012, 5, 570–574.
376 R. C. Ojah and J. M. Welch, Vitamin D and musculoskele-tal status in Nova Scotian women who wear concealingclothing, Nutrients, 2012, 4, 399–412.
377 S. Hatun, O. Islam, F. Cizmecioglu, B. Kara, K. Babaoglu,F. Berk and A. S. Gokalp, Subclinical vitamin D deficiencyis increased in adolescent girls who wear concealing cloth-ing, J. Nutr., 2005, 135, 218–222.
378 D. Bassil, M. Rahme, M. Hoteit and H. Fuleihan Gel,Hypovitaminosis D in the Middle East and North Africa:Prevalence, risk factors and impact on outcomes,Dermato-endocrinol., 2013, 5, 274–298.
379 P. S. Sobolewski, J. W. Krzyscin, J. Jaroslawski, J. Wink,A. Lesiak and J. Narbutt, Controlling adverse and bene-ficial effects of solar UV radiation by wearing suitableclothes - Spectral transmission of different kinds offabrics, J. Photochem. Photobiol., B, 2014, 140C, 105–110.
380 E. Linos, E. Keiser, T. Fu, G. Colditz, S. Chen and J. Y. Tang,Hat, shade, long sleeves, or sunscreen? Rethinking US sunprotection messages based on their relative effectiveness,Cancer Causes Control, 2011, 22, 1067–1071.
381 A. V. Parisi, R. Eley and N. Downs, Determination of theusage of shade structures via a dosimetry technique,Photochem. Photobiol., 2012, 88, 1012–1015.
382 R. L. McKenzie, J. B. Liley and L. O. Bjorn, UV radiation:balancing risks and benefits, Photochem. Photobiol., 2009,85, 88–98.
383 A. R. Webb and O. Engelsen, Ultraviolet exposure scen-arios: risks of erythema from recommendations oncutaneous vitamin D synthesis, Adv. Exp. Med. Biol., 2008,624, 72–85.
384 F. S. Rosenthal, C. Phoon, A. E. Bakalian and H. R. Taylor,The ocular dose of ultraviolet radiation to outdoorworkers, Invest. Ophthalmol. Visual Sci., 1988, 29, 649–656.
385 D. H. Sliney, Intraocular and crystalline lens protectionfrom ultraviolet damage, Eye Contact Lens, 2011, 37, 250–258.
386 U. L. Osuagwu, K. C. Ogbuehi and T. M. Almubrad,Changes in ultraviolet transmittance of hydrogel and sili-cone-hydrogel contact lenses induced by wear, Eye ContactLens, 2014, 40, 28–36.
387 N. Gao, L. W. Hu, Q. Gao, T. T. Ge, F. Wang, C. Chu,H. Yang and Y. Liu, Diurnal variation of ocular exposureto solar ultraviolet radiation based on data from amanikin head, Photochem. Photobiol., 2012, 88, 736–743.
388 F. Wang, Q. Gao, L. Hu, N. Gao, T. Ge, J. Yu and Y. Liu,Risk of eye damage from the wavelength-dependent bio-logically effective UVB spectrum irradiances, PLoS One,2012, 7, e52259.
389 L. Hu, Q. Gao, N. Gao, G. Liu, Y. Wang, H. Gong andY. Liu, Solar UV exposure at eye is different from environ-mental UV: diurnal monitoring at different rotationangles using a manikin, J. Occup. Environ. Hyg., 2013, 10,17–25.
390 J. Turner and A. V. Parisi, Influence of reflected UV irradi-ance on occupational exposure from combinations ofreflective wall surfaces, Photochem. Photobiol. Sci., 2013,12, 1589–1595.
391 H. Chandler, Ultraviolet absorption by contact lenses andthe significance on the ocular anterior segment, EyeContact Lens, 2011, 37, 259–266.
392 W. Lan, A. Petznick, S. Heryati, M. Rifada and L. Tong,Nuclear Factor-kappaB: central regulator in ocular surfaceinflammation and diseases, Ocul. Surf., 2012, 10, 137–148.
393 D. Ali, A. Verma, F. Mujtaba, A. Dwivedi, R. K. Hans andR. S. Ray, UVB-induced apoptosis and DNA damagingpotential of chrysene via reactive oxygen species inhuman keratinocytes, Toxicol. Lett., 2011, 204, 199–207.
394 M. A. Beketov, A. Speranza and M. Liess, Ultraviolet radi-ation increases sensitivity to pesticides: synergistic effectson populationgrowth rate of Daphnia magna at low concen-trations, Bull. Environ. Contam. Toxicol., 2011, 87, 231–237.
395 B. Epe, DNA damage spectra induced by photosensitiza-tion, Photochem. Photobiol. Sci., 2012, 11, 98–106.
396 G. Miolo, S. Caffieri, D. Dalzoppo, F. Gallocchio, E. Fasaniand G. M. Beyersbergen van Henegouwen, Photoactivationof corticosteroids in UVB-exposed skin, J. Photochem.Photobiol., B, 2011, 103, 35–41.
397 B. Gulson, M. McCall, M. Korsch, L. Gomez, P. Casey,Y. Oytam, A. Taylor, M. McCulloch, J. Trotter, L. Kinsley
Perspective Photochemical & Photobiological Sciences
86 | Photochem. Photobiol. Sci., 2015, 14, 53–87 This journal is © The Royal Society of Chemistry and Owner Societies 2015

and G. Greenoak, Small amounts of zinc from zinc oxideparticles in sunscreens applied outdoors are absorbedthrough human skin, Toxicol. Sci., 2010, 118, 140–149.
398 T. Kunisue, Z. Chen, G. M. Buck Louis, R. Sundaram,M. L. Hediger, L. Sun and K. Kannan, Urinary concen-trations of benzophenone-type UV filters in U.S. womenand their association with endometriosis, Environ. Sci.Technol., 2012, 46, 4624–4632.
399 Therapeutic Goods Administration, Literature review on thesafety of titanium dioxide and zinc oxide nanoparticles insunscreens. Scientific review report, Department of Healthand Ageing, Canberra, 2013, pp. 0–19.
400 S. Jatana and L. A. Delouise, Understanding engineerednanomaterial skin interactions and the modulatoryeffects of ultraviolet radiation skin exposure, Wiley Inter-discip. Rev.: Nanomed. and Nanobiotechnol., 2014, 6,61–79.
401 D.-P. Häder, C. E. Williamson, S.-Å. Wängberg, M. Rautio,K. C. Rose, K. Gao, E. W. Helbling, R. P. Sinha andR. Worrest, Effects of UV radiation on aquatic ecosystemsand interactions with other environmental factors, Photo-chem. Photobiol. Sci., 2015, 14, this issue.
402 K. G. McGuigan, R. M. Conroy, H. J. Mosler, M. du Preez,E. Ubomba-Jaswa and P. Fernandez-Ibanez, Solar waterdisinfection (SODIS): a review from bench-top to roof-top,J. Hazard. Mater., 2012, 235–236, 29–46.
403 H. Slaper, G. J. Velders, J. S. Daniel, F. R. de Gruijl andJ. C. van der Leun, Estimates of ozone depletion and skincancer incidence to examine the Vienna Conventionachievements, Nature, 1996, 384, 256–258.
404 A. van Dijk, H. Slaper, P. N. den Outer, O. Morgenstern,P. Braesicke, J. A. Pyle, H. Garny, A. Stenke, M. Dameris,A. Kazantzidis, K. Tourpali and A. F. Bais, Skin cancerrisks avoided by the Montreal Protocol–worldwide model-ing integrating coupled climate-chemistry models with arisk model for UV, Photochem. Photobiol., 2013, 89, 234–246.
405 A. F. Bais, K. Tourpali, A. Kazantzidis, H. Akiyoshi,S. Bekki, P. Braesicke, M. P. Chipperfield, M. Dameris,V. Eyring, H. Garny, D. Iachetti, P. Jockel, A. Kubin,U. Langematz, E. Mancini, M. Michou, O. Morgenstern,T. Nakamura, P. A. Newman, G. Pitari, D. A. Plummer,E. Rozanov, T. G. Shepherd, K. Shibata, W. Tian andY. Yamashita, Projections of UV radiation changes in the21st century: impact of ozone recovery and cloud effects,Atmos. Chem. Phys., 2011, 11, 7533–7545.
406 P. Correa Mde, S. Godin-Beekmann, M. Haeffelin,S. Bekki, P. Saiag, J. Badosa, F. Jegou, A. Pazmino andE. Mahe, Projected changes in clear-sky erythemal andvitamin D effective UV doses for Europe over the period2006 to 2100, Photochem. Photobiol. Sci., 2013, 12, 1053–1064.
407 B. Petersen, E. Thieden, P. A. Philipsen, J. Heydenreich,H. C. Wulf and A. R. Young, Determinants of personalultraviolet-radiation exposure doses on a sun holiday,Br. J. Dermatol., 2013, 168, 1073–1079.
408 L. Lemus-Deschamps and J. K. Makin, Fifty years ofchanges in UV Index and implications for skin cancer inAustralia, Int. J. Biometeorol., 2012, 56, 727–735.
409 J. Makin, Implications of climate change for skin cancerprevention in Australia, Health Promo. J. Aust., 2011, 22(Spec No), S39–S41.
410 J. C. van der Leun, R. D. Piacentini and F. R. de Gruijl,Climate change and human skin cancer, Photochem.Photobiol. Sci., 2008, 7, 730–733.
411 W. G. Tsiaras and M. A. Weinstock, Factors influencingvitamin D status, Acta Derm. Venereol., 2011, 91, 115–124.
412 P. Thomas, A. Swaminathan and R. M. Lucas, Climatechange and health with an emphasis on interactions withultraviolet radiation: a review, Global. Change Biol., 2012,18, 2392–2405.
413 J. F. Bornman, P. W. Barnes, S. A. Robinson, C. L. Ballaré,S. D. Flinte and M. M. Caldwell, Solar ultraviolet radiationand its interaction with other ozone depletion and associ-ated climate effects on terrestrial ecosystems, Photochem.Photobiol. Sci., 2015, 14, this issue.
414 D. J. Erickson, III, B. Sulzberger, R. Zepp, A. T. Austin andN. Paul, Effects of solar UV radiation and climate changeon biogeochemical cycling: Interactions and feedbacks,Photochem. Photobiol. Sci., 2015, 14, this issue.
415 N. Principi, P. Marchisio, L. Terranova, A. Zampiero,E. Baggi, C. Daleno, S. Tirelli, C. Pelucchi and S. Esposito,Impact of vitamin D administration on immunogenicityof trivalent inactivated influenza vaccine in previouslyunvaccinated children, Hum. Vaccines Immunother., 2013,9, 969–974.
416 S. Esposito, E. Baggi, S. Bianchini, P. Marchisio andN. Principi, Role of vitamin D in children with respiratorytract infection, Int. J. Immunopathol. Pharmacol., 2013, 26,1–13.
417 S. N. Byrne, How much sunlight is enough?, Photochem.Photobiol. Sci., 2014, 13, 840–852.
418 S. Seite, A. Fourtanier, D. Moyal and A. R. Young, Photo-damage to human skin by suberythemal exposure to solarultraviolet radiation can be attenuated by sunscreens: areview, Br. J. Dermatol., 2010, 163, 903–914.
419 K. D. Cashman, M. Kiely, M. Kinsella, R. A. Durazo-Arvizu,L. Tian, Y. Zhang, A. Lucey, A. Flynn, M. J. Gibney,H. W. Vesper, K. W. Phinney, P. M. Coates, M. F. Piccianoand C. T. Sempos, Evaluation of Vitamin D Standardiz-ation Program protocols for standardizing serum 25-hydroxyvitamin D data: a case study of the program’spotential for national nutrition and health surveys,Am. J. Clin. Nutr., 2013, 97, 1235–1242.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 53–87 | 87

Photochemical &Photobiological Sciences
PERSPECTIVE
Cite this: Photochem. Photobiol. Sci.,2015, 14, 88
Received 20th October 2014,Accepted 20th October 2014
DOI: 10.1039/c4pp90034k
www.rsc.org/pps
Solar ultraviolet radiation and ozone depletion-driven climate change: effects on terrestrialecosystems
J. F. Bornman,*a P. W. Barnes,b S. A. Robinson,c C. L. Ballaré,d S. D. Flinte andM. M. Caldwellf
In this assessment we summarise advances in our knowledge of how UV-B radiation (280–315 nm),
together with other climate change factors, influence terrestrial organisms and ecosystems. We identify
key uncertainties and knowledge gaps that limit our ability to fully evaluate the interactive effects of
ozone depletion and climate change on these systems. We also evaluate the biological consequences of
the way in which stratospheric ozone depletion has contributed to climate change in the Southern Hemi-
sphere. Since the last assessment, several new findings or insights have emerged or been strengthened.
These include: (1) the increasing recognition that UV-B radiation has specific regulatory roles in plant
growth and development that in turn can have beneficial consequences for plant productivity via effects
on plant hardiness, enhanced plant resistance to herbivores and pathogens, and improved quality of agri-
cultural products with subsequent implications for food security; (2) UV-B radiation together with UV-A
(315–400 nm) and visible (400–700 nm) radiation are significant drivers of decomposition of plant litter in
globally important arid and semi-arid ecosystems, such as grasslands and deserts. This occurs through
the process of photodegradation, which has implications for nutrient cycling and carbon storage,
although considerable uncertainty exists in quantifying its regional and global biogeochemical signifi-
cance; (3) UV radiation can contribute to climate change via its stimulation of volatile organic compounds
from plants, plant litter and soils, although the magnitude, rates and spatial patterns of these emissions
remain highly uncertain at present. UV-induced release of carbon from plant litter and soils may also con-
tribute to global warming; and (4) depletion of ozone in the Southern Hemisphere modifies climate
directly via effects on seasonal weather patterns (precipitation and wind) and these in turn have been
linked to changes in the growth of plants across the Southern Hemisphere. Such research has broadened
our understanding of the linkages that exist between the effects of ozone depletion, UV-B radiation and
climate change on terrestrial ecosystems.
Introduction
We have focused mainly on recent work in order to highlightthe progress made to date, and to attempt an analysis of the
complexity of both independent and interacting factors on ter-restrial ecosystems in terms of UV radiation and other environ-mental constraints, including emerging evidence of the role ofstratospheric ozone trends in affecting climate.
Ozone depletion, changed exposure to ultraviolet-B (UV-B,280–315 nm) radiation, and climate change exert both indi-vidual and interactive effects on biological systems, with intri-cate feedbacks.1,2 Some of the key factors interacting with UVradiation that affect organism response are water availability,temperature, and nutrient availability. UV radiation has alsobeen implicated as a contributor to global warming throughits stimulation of volatile organic compounds from plants,plant litter and soils. Emission of carbon dioxide (CO2) fromplant litter and soils may also contribute to global warming.3,4
Ozone depletion modifies Southern Hemisphere summerweather through its effect on the Southern Annular Mode(SAM), with consequences for plant growth in South America,
aInternational Institute of Agri-Food Security (IIAFS), Curtin University,
Building 408, PO Box U1987, Perth, WA 6845, Australia.
E-mail: [email protected] of Biological Sciences and Environment Program, Loyola University New
Orleans, Box 169, New Orleans, LA 70118, USAcInstitute for Conservation Biology, School of Biological Sciences, The University of
Wollongong, New South Wales 2522, AustraliadIFEVA Universidad de Buenos Aires and IIB Universidad Nacional de San Martín,
Consejo Nacional de Investigaciones Científicas y Técnicas, Avenida San Martín
4453, C1417DSE Buenos Aires, ArgentinaeDepartment of Forest, Rangeland, and Fire Sciences, University of Idaho, Moscow,
ID 83844-1133, USAf2126 Connecticut Ave., NW #61, Washington, DC 20008, USA
88 | Photochem. Photobiol. Sci., 2015, 14, 88–107 This journal is © The Royal Society of Chemistry and Owner Societies 2015

New Zealand, and Antarctica already reported.5–7 Theseimpacts of ozone depletion on other climate factors (e.g. windpatterns, precipitation, and warming) may result in an increasein the interactive effects of UV radiation with drought andtemperature. Other seasonal weather phenomena need to betaken into account to gain an accurate perspective of thedifferent determinants of UV exposure on terrestrial organi-sms. These include La Niña and El Niño events, which changecloud cover, winds, sea surface temperatures, and atmosphericpressure at sea level. In addition, changes in land-use andvegetation cover, which also feed back to climate systems, haveimplications on the exposure, and thus response, of organismsto UV radiation.
During the course of research on the effects of UV radi-ation, much emphasis has been placed on the potential detri-mental impacts on plants and ecosystems. However, thebalance of recent evidence is shifting to show that while somedetrimental effects do occur, UV radiation is also a key regula-tor of plant morphology and physiological, biochemical andgenetic processes, and is important in animal and plant sig-nalling. Following on from this line of investigation, it has alsobecome apparent that UV radiation and climate variables canbe usefully exploited for value-adding to, e.g., agriculturalcrops.8 The emerging concept, that agricultural2,9 plants canbecome more hardy through exposure to UV radiation, rep-resents a marked shift in perspective.8,9 In addition, certainplants produce more medicinal compounds with exposure toUV radiation.10 The overall objective is to boost the qualityand/or quantity of the yield, usually selectively, e.g. by makingplants less prone to attack by pests and diseases. Conceptssuch as that of “eustress” are also relevant. Eustress is ana-logous to “priming” where a stress is imposed on plants toacclimate them and develop tolerance, which facilitates bettergrowth when exposed to a more severe stress.9,11
Exposure to ecologically-relevant levels of UV radiation isgenerally not deleterious as long as plants are able to accli-matise, although this depends on the environmental con-ditions, including climate variables, latitudinal location12 andplant type (e.g. whether plants are herbaceous or woody). Con-sequently, the direct negative effects of exposure to UV-B radi-ation on plant growth, photosynthesis, and productivity aregenerally minor, or not detectable (summarised in meta-analyses by Searles et al.13 and Newsham and Robinson14).However, indirect effects of exposure to UV radiation are oftenmore pronounced than direct effects and need to be addressedto obtain a holistic perspective of the role of UV radiation as aregulator and modifier of ecosystem and organism response.
In this current assessment, we focus on the way in whichUV radiation, stratospheric ozone trends, climate and otherphenomena affect the biosphere, in order to better understandthe current interactive effects from different stresses and toidentify possible new interactions and their implications. Thiswill allow for an evaluation of the capability of terrestrial eco-systems to adapt to a changing environment in which UV radi-ation plays an integral part in the response. Additionally, anassessment of these interactive processes on organisms and
ecosystems recognises that the effects of UV radiation oftenrepresent a balance of both positive and negative influences.2
Although the role of UV-B radiation is a major consideration inthis paper, other relevant and often interacting factors, suchas stratospheric ozone trends and climate change cannot bemeaningfully separated.
Multiple plant stresses and theirimplications in the response to UVradiation
Evaluation of the effect of different levels of exposure to UV-Bradiation, whether beneficial or not, is complicated by thedependency on many other variables, including the sensitivityof diverse ecosystems and species. This differential sensitivity,which results in distinctive response patterns, reflects the com-plexity of the biosphere as our understanding increases.
UV radiation, temperature and drought
The volume of research concentrating on the impacts of UVradiation and drought is indicative of the increasing awarenessof a changing climate coupled with other interlinking factors.However, there are still several knowledge gaps, e.g. at themechanistic level around drought and UV-B signalling inter-actions and the consequences at the molecular level leading toplant response. Areas such as the Middle East, North Africa,certain regions of Australia and the Mediterranean are particu-larly vulnerable to climate change.15 Research on plants ori-ginating from drought-prone regions (e.g. the Mediterraneanarea), provides insight into current and potential long-termconstraints for growth of agricultural plants and ecosystems byincreased temperature, scarcity of water and enhancedexposure to UV radiation.16 Predicted decreased cloudiness inthe Mediterranean area is likely to enhance exposure to UV-Band UV-A radiation (315–400 nm).17 Other co-occurring factorsmodifying UV radiation at the Earth’s surface include air pol-lution and aerosol load.18
High solar radiation, high temperatures, and drought con-ditions can lead to an array of responses including oxidativestress and physiological and metabolic acclimation.19 The fre-quently observed, although not universal, cross-tolerance todrought stress and UV radiation is in contrast to the increasedsensitivity to UV radiation that can occur in adequatelywatered plants (Caldwell et al.20 and references therein andothers21–23). For example, in many woody Mediterraneanplants, several physiological and biochemical traits (e.g. plantgrowth, net rate of assimilation of CO2, and photochemistry)show little response to changes in UV radiation against a back-ground of high temperatures and drought periods.23
UV radiation and other stress factors can increase allocationof newly assimilated carbon to polyphenols, and in particular,flavonoid compounds, indicative of an energy shift in order toacclimate to stress conditions (Ballaré et al.1 and referencestherein and Guidi et al.19). The lack of a substantial response
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 88–107 | 89

to UV radiation in many of the parameters measured in Medi-terranean plants may also be explained by biochemical acclim-ation induced by the visible part of the solar spectrum, thusleading to acclimation to UV, and ultimately adaptation.19 Thisinvolves effective scavenging of reactive oxygen species andother protective mechanisms such as morphological, physio-logical, and biochemical changes16 that contribute to droughtand high temperature tolerance, as well as positive plant–insect interactions.24 Decreased plant productivity andchanges in crop quality may, however, occur with long-termand multiple stress exposure.
Significance of sequential stress
In the context of interactive stresses, recent work has contin-ued to address the importance of the sequence in whichplants are exposed to the different stresses (reviewed inBandurska et al.25), although further supporting research isneeded in this area. Field studies are also lacking, which maybe a reflection of the difficulties encountered in trying toevaluate sequential stresses under more natural conditions. Agrowth chamber study by Bandurska and Cieslak26 showedthat exposure of plants to UV-B radiation and drought,whether in combination or individually applied, affected meta-bolic processes differently at different locations within theplant. With separate applications of enhanced UV-B radiationand simulated drought conditions, plant growth is oftenretarded and oxidative stress increases. In general, exposure toboth UV-B radiation and drought can elicit a number ofphysiological and biochemical responses that are common toboth abiotic stresses. These involve increases in oxidativestress through production of reactive oxygen species, includinghydrogen peroxide and nitric oxide, growth inhibition and, insome cases, the induction of phenolic compounds (Bandurskaet al.26 and references therein). UV-induced oxidative damage,such as lipid peroxidation, can be reduced by pre-exposingplants to mild drought conditions.
Sequential or simultaneous exposure to stress factors canalso be genotype-specific. For example, as illustrated in agrowth-chamber study, drought-susceptible genotypes weremore adversely affected by simultaneous application ofenhanced UV-B radiation and drought conditions than thedrought-tolerant genotype.27 However, the response may differunder natural environments. When either drought orenhanced UV-B radiation is first supplied singly, the negativeeffects may be lessened when followed by the other stress.27
Similarly, pre-exposure of plants to enhanced levels of UV-Bradiation before exposure to high levels of visible light andhigh temperatures results in an increase in photosynthesisand relative growth.8 Pre-exposure to relatively high tempera-tures can also induce UV tolerance.28
These studies are of relevance particularly for horticulturalpractices, where the vitality of plants can be increased throughexposure to the relevant sequential stress prior to transferfrom a greenhouse/nursery to open field conditions.
Fertiliser application and nitrogen deposition
The degree of complexity of plant and ecosystem responses tomultiple environmental and climatic factors is becomingincreasingly evident as research moves from single-stressexperiments to more natural environmental conditions. This isexemplified by a number of recent field studies where appli-cation of fertiliser to plants exposed to enhanced UV radiationinduced varied responses. Fertiliser can modify the effects ofelevated temperature and UV-B radiation. For example,changes in some secondary metabolites, including certainphenolic compounds and alkaloids, occur, as was shown inNorway spruce (Picea abies L.).29 Furthermore, increased sensi-tivity towards enhanced UV-B radiation may result where ferti-liser is applied in excess of recommended levels.30 This wasmanifested in a shift in biomass towards the shoots of field-grown radish plants, causing a loss in yield compared togrowth with recommended levels of nitrogen (N), phosphorus(P), and potassium (K).30 In contrast, additional but not exces-sive N may alleviate the negative effects of UV-B radiationstress.31,32
Volatile organic compounds
Several environmental stresses, both biotic and abiotic, stimu-late the emission of volatile organic compounds (VOCs) fromplant tissues, partly as a protective mechanism.33,34 While alarge diversity of VOCs are produced from terrestrial ecosys-tems, only a few of these compounds contribute substantiallyto the total emissions.35 So far, only a few studies have focusedon co-occurring stresses33,36 to elucidate the potential interac-tive effects on VOC emissions. These stressors include UV radi-ation, drought, soil moisture, temperature, troposphericozone, and herbivore attack, presenting a challenge in termsof quantifying their individual and combined contributions toemissions of VOCs at a local, regional and global level. Thiscomplexity is further overlaid by the variable nature of emis-sions according to the type of plant. VOCs such as isoprene arepotential contributors to increasing greenhouse gases37 andthus may modify atmospheric composition, including extend-ing the lifetime of methane.38 For example, one of the majorsources of tropospheric carbon monoxide (CO) is oxidation ofisoprene, and the scavenging of OH radicals by CO reduces theoxidation rate of methane, increasing its lifetime.
Isoprene, the most environmentally important VOC emittedfrom terrestrial plants, contributes to ca. 50% of the totalglobal biogenic VOCs.35 While UV-B radiation and temperaturecan have interactive effects on VOC emissions, temperature isthe more significant driver of increasing emissions. Thosefactors that stimulate VOC emissions, including UV radi-ation,39 result in an energy-cost to plants with a potential lossin productivity as a result of allocation of energy and carbonto, for example, the synthesis of isoprene.33,40 The extent towhich UV radiation plays a stimulatory role in VOC emissionsis currently unknown.
VOCs from a range of terrestrial and certain marine systemshave been widely measured, including in extreme environ-
Perspective Photochemical & Photobiological Sciences
90 | Photochem. Photobiol. Sci., 2015, 14, 88–107 This journal is © The Royal Society of Chemistry and Owner Societies 2015

ments such as the Arctic regions and deserts. The effects ofenhanced UV-B radiation on emissions of VOCs from sub-Arctic peatlands are highly variable, even when averaged over agrowing season, ranging from no detectable change to 60%greater compared to ambient UV-B treatments.41–43 Thus,emissions of VOCs from plants and peatlands reflect the pre-vailing environmental conditions, including temperature,availability of water and UV radiation,39,44,45 and can lead tofurther interactions and feedbacks within the biosphere.
Methane, the second most important greenhouse gas aftercarbon dioxide, is produced by both microbial and non-microbial mechanisms as well as geological processes.Methane is emitted from peatlands and wetlands (Fig. 1) aswell as from other vegetation types,45 soils, surface waters,animals, and fungi (Wang et al.46 and references therein).Microbially-produced methane has been well documented(Wang et al.46 and references therein), while methane pro-duction from oxygen-rich environments (non-microbial pro-duction) has only recently been investigated and itsimplications discussed. Recent estimates of emission ofmethane from plants indicate that this may be very low,although these estimates have varied substantially (e.g. contri-buting <0.2% to 40% of all methane released to theatmosphere).47–50 The source of methane emitted from vege-tation is still under investigation, with reports suggesting emis-sion is from an internal, plant structural cell wall component,pectin,51 or from surface waxes of leaves, the production ofwhich may be stimulated by UV radiation.52 Additional factorsleading to the stimulation of methane emissions include pro-duction of reactive oxygen species induced by environmentalstress and injury to plants.53–55
As is the case for many of the non-methane VOCs, the mag-nitude of methane emissions is variable and dependent onlocation and ambient conditions including UV radiation. A
long-term study over 6 years showed that enhanced UV-B radi-ation in a boreal peatland increased methane emissions.45 Acomplementary study on the emission of methane from treesgrowing in natural wetlands showed that emissions from treesexceeded those of the peatland on which they were growing.These studies found that wetland trees mediated emission ofmethane from the peatland.56 In contrast, a study of a wetland(fen) in Northern Finland over three growing seasons did notshow UV-B induced changes in net emission of methane,although enhanced UV-B radiation contributed to a slightincrease in the organic acid precursors for methanogenesis.57
In field experiments specifically studying the response of riceplants and paddy fields to enhanced UV-B radiation, UV-B sup-plementation using lamps significantly increased the emissionof methane from the paddy, especially between the tilleringand heading stages of the rice. At the same time, decreasedtiller number and biomass occurred as a response to theenhanced UV-B radiation levels.58
The important roles played by VOCs in ecosystem function-ing – from inter- and intra-plant communications to plantchemical defence that decreases damage by herbivores – needto be analysed within a dynamic environment of interactingstresses from rapid and frequent climate events and changesin exposure to UV radiation. These factors should be con-sidered when addressing the roles of different VOCs in mediat-ing the diverse range of interactions occurring in ecosystems.In general, further investigations under natural environmentalconditions are required to clarify the overall significance of UVradiation as a contributing factor to VOC emissions fromplants and peatlands.
The ozone ‘hole’ as a driver forSouthern Hemisphere climate andecosystem change
The role of ozone depletion in Southern Hemisphere climateprocesses59 has been largely overlooked in the biological litera-ture and this section highlights some of the ways ozonedepletion, independent of UV radiation, has affected ecosys-tems through climatic change. While these climate pertur-bations are likely to have had a significant impact over thepast few decades they have only recently started to be con-sidered at the ecological level.
The ozone ‘hole’ moderates Southern Hemisphere climate
Ozone depletion and our response to it have had major impli-cations for the Earth’s climate. In the Southern Hemisphere,the ozone ‘hole’ has been a dominant driver5–7 of atmosphericcirculation changes since the 1970s and has shielded Antarc-tica from much of the effects of global warming.6,60–63
The Southern Hemisphere Annular Mode (SAM), or Antarc-tic Oscillation, refers to the difference in atmospheric pressurebetween the mid- and high-latitudes of the Southern Hemi-sphere and thus the position of the polar jet. When the SAM
Fig. 1 Wetland ecosystems, such as in south-eastern Louisiana, USA,are important sources of methane and have been the subject of anumber of studies examining the effects of UV radiation on methaneemissions (photograph: P. Barnes).
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 88–107 | 91

index is positive the polar jet is located towards the South Poleand when it is negative it moves northwards. Since the late1970s, the atmospheric polar jet has shifted south by 1–2° oflatitude and increased in strength by 15–20%.64,65 Theobserved trend in the SAM has been largest during the australsummer and this is believed to be driven primarily by thedevelopment of the Antarctic ozone ‘hole’ (Thompson et al.5
and references therein). From records of ice cores it appearsthat intensification of the westerlies started over a century ago,driven by increases in greenhouse gases, but in recent decadesthe intensification associated with ozone depletion has beenmore pronounced.66 The SAM index is now at its highest levelfor at least 1000 years.67
Climate-related effects of ozone depletion on SouthernHemisphere ecosystems
The changes to circumpolar westerly winds attributed to ozonedepletion are likely to have far-reaching consequences for bio-logical ecosystems, particularly due to the influence of windon the availability of water. The availability of liquid water toorganisms broadly depends on the balance between annualprecipitation and losses by evaporation, sublimation, andfreezing. Increased wind causes evaporation, and in polar andalpine environments, sublimation and scouring of snow.68 InAntarctica, increased wind speeds have been linked todecreased growth rates of plants69 and changes in biodiversityin lakes70 in East Antarctica (Fig. 2). These changes are corre-lated with declining availability of water in East Antarcticcoastal sites associated with increasing wind speeds and, inthe case of moss growth rates, ozone depletion (Fig. 2). Thisresearch thus links ozone depletion to negative biodiversityoutcomes through factors other than increasing UV-B radiation(reviewed in Robinson and Erickson7).
Winds also transport dust (including nutrients, seeds,spores, and other reproductive structures) from lower latitudesinto the Southern Ocean and central West Antarctica (Fig. 2).Changes to either the location or strength of winds can affectthe sources of dust and the quantities transported, with wide-spread implications including changes in productivity of theocean. For example, increased wind-blown iron deposited intothe ocean leads to phytoplankton blooms, indicative ofincreased productivity.71,72
These biological impacts are not restricted to the Antarctic.Significant changes to tree growth across the Southern Hemi-sphere in the last 50 years, relative to the previous 250 years,correlate with ozone ‘hole’-influenced changes to the SAM73
(Fig. 2). This is illustrated by the ca. 50% decline in growthrates of three species of trees (Austrocedrus chilensis, Araucariaaraucana and Nothofagus betuloides) in southernmost SouthAmerica since the 1950s, associated with SAM-induceddecreased precipitation in the Andes. Similarly, changes to cir-culation patterns have increased precipitation over sub-alpineareas of New Zealand resulting in greater than average rates ofgrowth in another tree species (Halocarpus biformis). In thiscase, a third of the growth increase was attributed to changesin the SAM.73
Recent modelling studies suggest that stratospheric ozonelosses since the 1970s have also increased the frequency andintensity of extreme precipitation in austral summer.74,75 Thishas resulted in drying at the southern tip of South Americaand SW Australia, wetter summers in SE Australia, E NewZealand and SE South America and increased precipitation andfreshening (increase in freshwater input) in the Southern Ocean76
(Fig. 2). Given the importance of water availability for all life onEarth, the vital role it plays in human and ecosystem health andfor food security, these findings suggest that ozone depletionhas far greater ecosystem impacts than previously anticipated.In particular, extremes of precipitation (droughts and floods)can be economically and socially devastating (cf. IPCC77).
Changes to the Southern Hemisphere circulation processescan have wider modifying effects than simply on wind speedsand associated water availability. It has been suggested thatthe shift to warmer summers in Southern Africa strongly corre-lates with the large ozone ‘hole’ era (1993–2010).78 Manatsaet al.78 re-analysed satellite data, focusing on October–Decem-ber, to separate the effect of greenhouse gases on the SAMindex from effects attributed to the ozone ‘hole’ (Fig. 2). Whilethis analysis does not specifically link to biological impacts, itillustrates the potential effects of the ozone ‘hole’ in terms ofhuman health, natural ecosystems, and agriculture. It alsoillustrates the need to investigate the role of the ozone ‘hole’ inecological processes and systems other than those directlyrelated to changes in UV radiation. Since ozone depletion isdriving multiple stressors across the Southern hemisphere (e.g.increased UV-B radiation combined with either increaseddrought or increased precipitation), the interactive effectsdescribed above need to be considered.
Recovery of the ozone ‘hole’ over the next century will havewidespread and complex effects on Southern Hemisphereclimate processes with counter forcing from greenhouse gasemissions predicted to play a pivotal role.79 Contrary to expec-tations, some consequences of ozone depletion have beenpositive, such as maintaining Antarctica’s cold temperatures,and their reversal may have negative effects for life on Earth. Amore holistic picture of the true ecological consequences ofozone depletion on terrestrial, aquatic and marine ecosystemsin the Southern Hemisphere is required in order to betterproject future changes and ways of mitigating risk.7
UV-B radiation and litterdecomposition
Decomposition of organic material is a crucial component ofglobal biogeochemical cycles that affects soil fertility, the fateand residence times of carbon and nutrients in organic matterpools, and ultimately plant community composition andproduction.43
Although the activity and make-up of the decomposingmicrobial community (bacteria and fungi) are key determi-nants of decomposition, solar radiation, including the UV-Bcomponent, plays a significant role. Recent studies, including
Perspective Photochemical & Photobiological Sciences
92 | Photochem. Photobiol. Sci., 2015, 14, 88–107 This journal is © The Royal Society of Chemistry and Owner Societies 2015
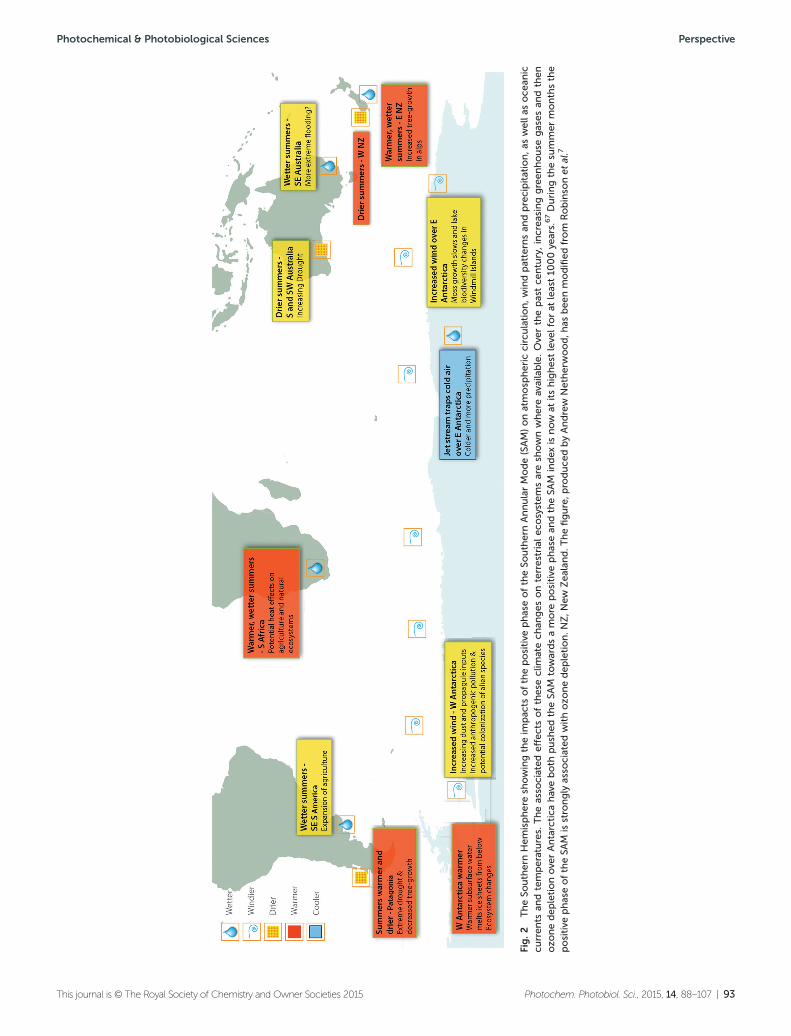
Fig.2
TheSo
uthern
Hemisphere
showingtheim
pac
tsofthepositive
phaseoftheSo
uthern
AnnularMode(SAM)onatmosp
hericcirculation,w
indpatternsan
dprecipitation,a
swellas
oce
anic
currents
andtemperatures.
Theasso
ciatedeffectsofthese
clim
atech
angesonterrestrial
eco
systemsaresh
ownwhere
available.Ove
rthepastce
ntury,increasinggreenhouse
gasesan
dthen
ozo
nedepletionove
rAntarctica
haveboth
push
edtheSA
Mtowardsamore
positive
phasean
dtheSA
Mindex
isnow
atitshighest
leve
lforat
least10
00ye
ars.67Duringthesu
mmermonthsthe
positive
phaseoftheSA
Misstrongly
asso
ciatedwithozo
nedepletion.N
Z,N
ewZealan
d.T
hefigure,p
roduce
dbyAndrew
Netherw
ood,h
asbeenmodifiedfrom
Robinso
net
al.7
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 88–107 | 93

several meta-analyses,80,81 have increased our understandingof the ecological significance of UV-B radiation on decompo-sition, including the fundamental mechanisms by which UV-B(and UV-A) radiation modifies the decomposition process, andthe potential linkages between changes in UV radiation andother climate change factors. These analyses indicate thatUV-B radiation, at ambient or enhanced levels, has complexeffects on decomposition of litter and can either retard oraccelerate rates of decomposition depending on UV-Bexposure, climatic factors (e.g. temperature and precipitation),and litter chemistry and structure. The conditions and ecosys-tems where UV-B radiation is expected to play a significant rolein litter decomposition are described below.
UV exposure of litter and wavelength sensitivity ofdecomposition processes
Ultimately, the effectiveness of incident solar UV radiation onthe decomposition of litter is determined largely by exposure
to UV and the sensitivity of the underlying decomposition pro-cesses to the wavelength (Fig. 3). At present, little is known ofthe precise nature of the dose–response relationships for thevarious mechanisms of UV-driven decomposition and whataction spectra best describe the sensitivity of these processes.This makes it difficult to quantitatively assess the importanceof the effects of ozone depletion and associated changes in UVradiation on the decomposition of litter, storage of carbon byecosystems and the emissions of CO2 and other trace gasesfrom decomposing plant litter. Recent studies have high-lighted the importance of factors such as cloud cover,1,82 veg-etation structure,82 litter depth,83 litter orientation,84 and soilcoverage85 in altering the exposure of litter to UV radiation(Fig. 3 and 4). Also, while several recent studies have shownthat photodegradation can be driven by UV-A and visible radi-ation (photosynthetically active radiation; PAR, 400–700 nm)in addition to the UV-B component,86,87 little progress hasbeen made in developing action spectra for specific biotic
Fig. 3 Conceptual model of the direct and indirect effects of solar UV radiation (290–400 nm) on the decomposition of terrestrial leaf litter, includ-ing potential interactions with other environmental factors. The total exposure of litter to UV radiation will depend on a combination of climatic,landscape/vegetation and species-specific factors. UV radiation generally reduces rates of biotic decomposition, whereas abiotic processes tend toenhance decomposition. Both processes exhibit distinct wavelength sensitivities (as seen from action spectra) depending on the underlying chromo-phores and mechanisms involved. Indirect effects of UV radiation are primarily mediated by those on leaf chemistry and structure, leading, e.g., todecreasing attractiveness of the litter for decomposing microbes. The role played by UV radiation on decomposition can depend also on otherenvironmental factors interacting with biotic and abiotic processes and leaf chemistry and structure. The net effect of solar UV radiation on rates ofdecomposition, nutrient cycling and carbon storage will depend on the combined effects of biotic and abiotic processes and may be positive, nega-tive or neutral. PAR, photosynthetically active radiation (400–700 nm).
Perspective Photochemical & Photobiological Sciences
94 | Photochem. Photobiol. Sci., 2015, 14, 88–107 This journal is © The Royal Society of Chemistry and Owner Societies 2015

(microbial) and abiotic (photodegradation) decompositionprocesses (Barnes et al.82 and references therein; but see Gaoand Garcia-Pichel88).
Effects of UV radiation on decomposer microorganisms
Solar UV-B radiation affects litter decomposition in terrestrialecosystems via several mechanisms including direct effects onmicrobes and abiotic photochemistry (photodegradation), aswell as through indirect effects mediated through alterationsin leaf chemistry (Fig. 3; Barnes et al.82) These mechanismscan interact with one another and multiple pathways ofdecomposition can occur within a given process (see below).These processes are difficult to differentiate under field con-ditions. Hence, the quantitative importance of individualeffects is largely unknown. In general, UV-B radiation tends tohave negative effects on the growth, survival, and reproduction
of microbes, which then retards rates of decomposition (Fig. 3;see also ref. 1, 82, 89). However, microbial species vary in theirsensitivity to UV-B radiation, as evidenced by studies demon-strating insensitivity of certain bacterial and fungal species toUV-B radiation in extreme environments.90,91 Consequently,the overall effect of solar UV radiation (UV-B + UV-A) on thecommunity composition of microbial decomposers iscomplex92,93 and requires a multi-criteria approach.
UV radiation and photodegradation of litter
The abiotic process of photodegradation occurs via photoche-mical mineralisation of photo-reactive compounds, such aslignin, and/or the transformation of compounds as a result ofUV-induced formation of reactive oxygen species and otherintermediates.80,86 In addition, apparent photodegradation isenhanced in the presence of oxygen but can also occur underanoxic conditions,4 indicative of the involvement of multiplechemical pathways.80 Generally, rates of photodegradationtend to increase with increasing moisture content of litter87,94
and air temperature.4
The effects of UV radiation on biotic (microbial) and abiotic(photodegradation) processes are not entirely independentand there is evidence that photodegradation can modify orpartially degrade compounds in ways that enhance or retardsubsequent microbial decomposition of litter (i.e. photoprim-ing95,96) Thus, even when direct photodegradation has a minoreffect on the loss of mass of litter, subsequent biological turn-over can be positively correlated with the length of priorexposure to radiation.95 Photopriming may be of particularimportance in the “conditioning” of litter prior to its detach-ment from living vegetation (i.e. “standing litter”; Fig. 4) andincorporation into the soil.86 However, photopriming canenhance carbon mineralisation from organic matter at the soilsurface.97 In addition, susceptibility to photopriming will varyamong plant species. Future photopriming experiments withmultiple species in field situations are needed to assesswhether this is a frequent or important facet of the photode-gradation processes.
Effects of UV radiation on decomposition mediated by plants
Solar UV-B radiation can alter the chemistry and structure ofliving plant tissue, which then makes subsequent litter lesssuitable for the growth of decomposing microbes (Fig. 3; seealso ref. 82 and 89). These UV-induced changes in plant tissuechemistry generally involve stimulation of the production ofphenolic compounds that function in the protection of plantsagainst UV radiation,13,98 although concentrations of otherchemical constituents (e.g. C, N, P, K, lignin, C : N) can changeas well.89 UV-B-induced changes in leaf chemistry need furtherevaluation for their potential role in photodegradation.
Interactive effects of UV radiation, climate and vegetation ondecomposition
Photodegradation results in the efflux of a number of gases,including CO2,
4,99 CH4,49,51 carbon monoxide (CO),4,87 and
nitrous oxide (N2O).95 Analyses suggest that photodegradation
Fig. 4 Temporal and spatial aspects of litter production and distributionin a semi-desert savanna in the Sonoran Desert, southern Arizona,USA. A. End-of-growing season standing litter of the C4 grass, Hetero-pogon contortus with the winter-deciduous shrub, Prosopis velutinabefore leaf drop. At this time of year there is significant potential forphotodegradation of the standing litter of the grass. B. Spatial variationin bare ground, surface litter accumulation and light conditions underand near a P. velutina canopy after leaf drop and prior to the onset ofthe growing season. Once litter reaches the ground it can be redistribu-ted across the landscape and mixed with soil such that microbial-drivendecomposition increases while photodegradation decreases (photo-graph: S. Archer).
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 88–107 | 95

of surface litter can have measurable effects on landscape-levelrates of the flux of CO2, and ultimately carbon storage.99,100
The potential therefore exists for significant involvement ofUV-driven photodegradation in influencing atmospheric CO2
levels and carbon sequestration, at least in certain ecosystems.The magnitude of these effects, however, is at presentuncertain.
Under field conditions, the effects of UV-B radiation onmicrobial decomposition and photodegradation usually occursimultaneously, such that the net impact of UV-B radiationwill reflect the balance of the biotic and abiotic mecha-nisms82,94 (Fig. 3). Consequently, UV-B radiation can have apositive, negative or neutral effect on decomposition, depend-ing on environmental conditions and quality of the litter.Under dry conditions, exposure to UV-B radiation tends toincrease decomposition via photodegradation. In contrast,where litter is exposed to UV-B radiation under moist con-ditions, rates of decomposition of litter generally decrease.This could reflect the inhibition of microbial activity by theUV-B radiation. Results from a recent meta-analysis of 93 fieldand laboratory studies conducted across six biomes (naturalenvironmental communities), revealed the complex role ofUV-B radiation in the decomposition of litter.81 Investigatorsreported that the direct effects of elevated and ambient UV-Bradiation tended to increase the rate of decomposition (7 to23%). However, the indirect effects (i.e. plant-mediated effects)were variable in magnitude and direction (−7 to 12%) depend-ing on exposure to UV radiation. None of these positive andnegative changes were statistically significant, however.Overall, the effects of UV-B radiation on litter decompositionare influenced by exposure to UV-B radiation, climatic factors(temperature and precipitation), and litter chemistry. Whilethese findings are generally consistent with our understandingof the role of UV-B radiation on decomposition rates, they doindicate that detecting a statistically significant effect of UV-Bradiation on the decomposition of litter may often be difficultwithin the limits of field experiments.
In arid and semi-arid climates (referred to as dryland eco-systems), UV-B radiation has significant and measurableimpacts on decomposition (Fig. 4). These ecosystems typicallyreceive large solar UV radiation fluxes at ground level, hightemperatures, and low and highly variable precipitation-conditions that tend to shift the balance of effects of UV-Bradiation on decomposition in favour of abiotic photodegrada-tion processes. Photodegradation is now being considered asan important process affecting carbon cycling and storage inthese systems. However, most studies in drylands have notexplicitly considered factors that routinely alter the exposure oflitter to UV radiation (e.g., litter depth, soil-litter mixing andlitter movement across the landscape). While such studies mayreasonably determine the decomposition of attached standingplant litter, their extrapolation to decomposition of detachedplant litter on soil surfaces fails to account for these factorsthat can strongly mediate or even negate the abiotic effects.Thus, use of more realistic field conditions in experimentswould serve to increase understanding and allow a more rigo-
rous quantification of the role of solar UV-B and UV-A radi-ation on the decomposition of litter and biogeochemistry. Ingeneral, these findings highlight the linkages betweenchanges in UV-B radiation, climate, and land-cover andsuggest that these may alter the magnitude and even the direc-tion of the effect of UV-B radiation on decomposition in terres-trial ecosystems.
It is evident that the degree of photodegradation varies withplant species and chemical composition of the litter.4,101 Ameta-analysis of data from 50 field studies largely fromdryland ecosystems80 showed that loss of mass due to photo-degradation alone is, on average, 23%, but considerable vari-ation occurs in rates among and within study sites. Thevariation in photodegradation was related to structural andchemical attributes of litter (area : mass and C : N ratios butnot lignin content), precipitation, and exposure to solar radi-ation. This suggests that photodegradation may be an impor-tant but historically overlooked mechanism of decompositionin these systems and may account for the discrepanciesbetween measured rates of decomposition and those predictedfrom ecosystem models based largely on climatic factors(temperature and precipitation) and initial litterquality.80,102–104 However, the role of the UV-B radiation com-ponent may be less effective than that of short wavelengthvisible radiation, as suggested by the meta-analysis of Kinget al.,80 although more evidence is needed to confirm this.
Elevated UV-B radiation and deposition of N also affectdecomposition of plant litter, an important aspect of nutrientcycling.43 A field experiment in China showed that a combi-nation of UV radiation and N deposition markedly increasedthe rate of litter decomposition under bamboo stands.105 Also,loss of carbon and phosphorus, and lignin degradation werepromoted. Nitrogen deposition changes the availability of N insoil as well as the enzymatic activity of soil microbes such thatincreasing N can stimulate decomposition of litter. In contrast,enhanced UV-B radiation may decrease release of N from plantlitter, probably through a direct effect on the microbial decom-posers. Opposite outcomes of the effects of UV-B radiation anddeposition of N have also been documented where the type oflitter and ambient environmental conditions differ.105
UV radiation, soil organisms andbelowground processes
Previous assessments have reported that although UV radi-ation does not penetrate soil to any great extent, a variety ofsoil-dwelling organisms and processes below ground can bemodified indirectly via the effects of UV radiation on plantsand plant litter. This occurs through UV-B-induced alterationsin the chemistry of the leaf, which in turn can affect thephysiology and composition of species of decomposer microor-ganisms (bacteria and fungi) in soil. There is considerable vari-ation in these responses, depending on species, ecosystemtype and developmental stages of the associated plants.1,106,107
Perspective Photochemical & Photobiological Sciences
96 | Photochem. Photobiol. Sci., 2015, 14, 88–107 This journal is © The Royal Society of Chemistry and Owner Societies 2015

While certain compounds in roots, such as flavonoids, canchange in response to aboveground exposure of plants to UV-Bradiation,108 a number of interactions in the soil immediatelyaround root systems (rhizosphere) are mediated by flavonoidspresent in the exudates of roots.109 These include signallingbetween the root and Rhizobium (N-fixing bacteria), mycorrhi-zal infection, and plant–plant allelochemistry (release of bio-chemicals by plants that may be harmful or beneficial, i.e.allelopathy).110 Solar UV radiation has been implicated inplaying a role in altering competitive interactions betweeninvasive and native plant species by increasing the negativeallelopathic effects of exotic species on native species.111 Infield studies,112 exclusion of solar UV radiation significantlyincreased root growth, root nodulation, nitrogenase activity,and the leghaemoglobin and hemechrome contents in rootnodules, suggesting a negative effect on the fixation of nitro-gen in certain species under ambient levels of solar radiation.The potential exists for significant modifications in soil invert-ebrate communities in Antarctic and Arctic ecosystems cur-rently experiencing appreciable ozone depletion and climatechange. Reduction in herbivory by insects under solar UV-Bradiation would be expected to be driven largely by differencesarising from plant biochemistry and composition of vegetationrather than by direct effects of elevated UV-B radiation onthese organisms.113 Although additional studies are clearlyneeded, these findings suggest a role for UV-B radiation inseveral belowground processes, which would have importantconsequences for mineral-nutrition of plants, storage ofcarbon in soil, biogeochemical cycles, and composition ofplant species.
Implications of UV-induced changes inplant defence systems
Exposure of plants to solar UV-B radiation leads to an increasein several secondary metabolites that play a key role in theinteractions of plants with other organisms, including her-bivorous insects and microbial pathogens (Paul et al.,93
reviewed in Ballaré et al.114). Reduction in herbivory by insectsunder solar UV-B radiation is well documented in field-grownplants1 with the majority (>80%) of such studies reportingincreases in plant damage or insect growth when solar UV-Bradiation is experimentally reduced.115,116 Direct avoidance ofUV radiation by many insects partly accounts for the reductionin herbivory.117,118 There are also some indications that, underambient levels of UV-B radiation, infection of plants by patho-gens is reduced.119,120 Pretreatment of plants with UV-B radi-ation before inoculation with a pathogen can also increaseresistance to infection.120
Some of the increased production of secondary metabolitesby UV-B radiation that boost plant defenses against pestsincludes leaf phenolics,120–122 conjugated polyamines,122 diter-penes123 and, in some cases, defense-related proteins such asproteinase inhibitors.122,124,125 On the other hand, phenoliccompounds induced by UV-A radiation may modulate the
responses to UV-B radiation via complex interactions betweenthe UV-B photoreceptor, UVR8126,127 and UV-A/blue light sig-nalling pathways.128 The functional relationships betweenUV-B radiation and the resistance of plants to pests (involvingsalicylic acid (SA) and jasmonic acid (JA) pathways) are not yetwell characterised (for a review, see Ballaré et al.114), althoughthere is increasing evidence that some of the effects of UV-Bradiation on the resistance of plants to herbivory are mediatedby increased JA signalling.122,123 In addition, UV-B radiationcan affect plant defense against herbivores and pathogens viamechanisms that are not mediated by JA. For example, UV-Bradiation, acting through UVR8, increases resistance of certainplants to the necrotrophic fungus Botrytis cinerea, and thiseffect is conserved in JA-insensitive genotypes.120
Protection and acclimation
Apart from plant defense mechanisms that protect againstpests and disease, other UV-acclimative processes involvingthe accumulation of UV-absorbing compounds result inchanges in plant optical properties (primarily epidermal trans-mittance, see Ballaré et al.1 and references therein). This accli-mative response varies according to a wide range of prevailingconditions, including geographical and temporal, as well aswith plant morphology (e.g. Ruhland et al.129) and increasesunderstanding of the capacity of plants to respond to changinglevels of UV radiation. Examples of dynamic acclimationinclude that of mature shade leaves of Populus tremuloides andVicia faba where epidermal transmittance of UV radiationdecreases when plants are transferred to sunny environments.Sun leaves, on the other hand, do not immediately respond bydecreasing UV-absorbing pigments upon transfer to shadyenvironments.130 This ability to increase but not decreasetransmittance of UV radiation rapidly may be associated withthe location of the UV-absorbing compounds. Compoundsbound to cell walls are less likely to be re-mobilised than thoseinside the cell.131,132
Plants may also rapidly adjust their mechanism of UV-pro-tection in response to daily changes in UV irradiances. Protec-tion against UV radiation increases from dawn to midday andthen decreases towards sunset.133 How plants achieve theserapid changes and what the significance is for functionbeyond protection from UV radiation is not yet known.Increased allocation of carbon to UV-absorbing compoundsmay divert carbon from growth and photosynthetic func-tions,134 such that reducing UV protection during times of theday when levels of UV radiation are low could enhance thedaily gain of carbon. In comparison to plants that maintainhigh UV protection throughout the day, plants that exhibitdiurnal changes in epidermal UV transmittance experienceincreased UV radiation exposure to the underlying mesophyllboth early and late in the day, but not at midday. It is possiblethat increased penetration of UV radiation at these times mayprotect leaves from photoinhibition that can occur at midday.8
Since UV-A radiation can also drive photosynthesis,135 the
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 88–107 | 97

increased penetration of UV-A radiation may enhance photo-synthesis at times of the day when leaves are light-limited.Added to this, several of the compounds induced by UV radi-ation (e.g. quercetin, kaempferol) inhibit the plant growthregulator, auxin, and its transport within the plant.136 Thus,maintaining high concentrations of flavonoids could interferewith plant growth during night-time periods. Although theprocesses of acclimation of plants to UV radiation are complex,it is now clear that UV-absorbing acclimation involving UV-absorbers is much more flexible and dynamic than previouslythought and has implications for the evaluation of plant har-diness under different environmental conditions.
Revisiting the potential forreconstructing past variations instratospheric ozone and UV-Bradiation
The relationships between ozone and climate change arecomplex59 and further investigation of the past nature of theserelationships will lead to a better understanding of the physi-cal interactions among solar activity, ozone and climate as wellas more clarity on how ecosystems respond to changing UV-Bradiation.137 The near ubiquitous response of plants toproduce chemically stable UV-absorbing compounds, and thusachieve protection from UV-B radiation, means that planttissues preserved in herbaria and in sedimentary and ice-corearchives can be used to reconstruct past variations (prior tothe instrumental period) in stratospheric ozone and UV-Bradiation.138–140 In particular, UV-absorbing compoundsbound to cell walls of plant tissues are extremely stable69 andmay even persist in fossils.138,139 In addition to tracking pastUV-B radiation, plant material is increasingly being assessedfor its usefulness as a biological proxy for historical availabilityof water,141 temperature, and concentration of CO2 in theatmosphere (reviewed in Jordan142). Spores and pollen arecommon in the fossil record, and thus this type of proxyenables reconstruction of past UV-B radiation for providingspatial and temporal fidelity.140
UV-absorbing compounds (e.g. p-coumaric acid) in cellwalls vary with latitude (and estimated exposure to UV-B radi-ation) in both spores of club mosses and in pine pollen.Greater variation is observed in samples collected from polarregions, where UV-B radiation has varied in recent decadesdue to ozone depletion, than in those collected from thetropics, where UV radiation levels have remained relativelystable.138,143 Similarly, UV-absorbing compounds in thesesame tissues show good temporal correlations with indepen-dent instrumental records and model results, allowing recon-struction of ozone concentration and flux of UV-B radiationover decades138 to millennia.143 However, further validation ofproxies using UV-absorbing compounds requires developmentof UV radiation dose–response curves, as well as confirmation
of the long-term chemical stability of the UV-absorbing com-pounds within paleobiological samples.139
Methodological advances, especially in microspectro-scopy,138,140,143 allow analysis of very small samples (e.g.50 grains of pine pollen). Such micro-scale analysis would alsopermit UV-absorbing compounds to be tracked down thelength of shoots of slow growing individual plants such aspolar mosses.69,141 Proxies that provide concurrent infor-mation on past climate and UV-B radiation, over centuries tomillennia, would be especially valuable for greater understand-ing of past polar environments.
Implications of exposure to UVradiation and climate interactions forfood production and food quality
Given the way in which solar UV radiation, stratosphericozone, and climate variables have multiple and often inter-dependent effects on terrestrial and agricultural systems, it islogical that these effects will modify the development, pro-duction and crop quality of agricultural crops.9,144–146
However, there are currently few studies focusing on the inter-play of climatic variables on crop quality.9,146 In contrast,many studies have reported both negative and positive effectsof UV radiation and other climate variables on crop pro-duction. The type of response in the crop is largely dependentupon species and cultivar, geographical location, geneticdifferences, and the co-occurring environmental conditions(cf. ref. 1, 9, 147). Where UV-B radiation has had a negativeeffect on crop production, this has been manifested mainly assmall decreases in biomass, and reduced leaf area (Ballaréet al.1 and references therein). As noted in previous sections,there are also indirect effects of UV-B radiation on plantgrowth such as decreased herbivory, due to increased second-ary metabolites such as phenols, which is a positive planteffect.24,93,148
Many investigations have documented changes in bio-chemical and regulatory pathways in plants under ambientand enhanced UV-B radiation, although few studies have expli-citly related these to follow-on effects on the quality of foodcrops.9,146 The UV-stimulated biochemical changes also con-tribute to differences in taste and aroma in, e.g., herbs such asmints.149 Several field studies have shown that ambient UV-Bradiation increases concentrations of chemicals, e.g. flavonols,esters and fatty acids, which can enhance the aroma andflavour of wine.150–152 Terpene emission from grapes alsoincreases with enhanced UV-B radiation and may be anothermodifier of quality.153
In general, a number of stress factors, including UV radi-ation, tend to increase the concentrations of proteins and anti-oxidants and reduce those of starch and lipids.144,154 Whilethese biochemical changes often reflect the acclimativeresponse of plants to enhanced levels of UV radiation andother environmental stresses, they may have either positive or
Perspective Photochemical & Photobiological Sciences
98 | Photochem. Photobiol. Sci., 2015, 14, 88–107 This journal is © The Royal Society of Chemistry and Owner Societies 2015

negative consequences for the quality of crops in terms ofnutrition and commercial use for specific products.
The degree to which plants can modulate acclimativeresponse to often rapid changes in UV radiation is also ofinterest for agricultural and horticulture practices. A lag in thisresponse has implications for UV susceptibility of crops thatare propagated in low UV radiation environments (e.g. green-houses) and are then subsequently transplanted to the field.This is illustrated by growth chamber studies with lettuce(Lactuca sativa), where new developing leaves require at least6–8 days to fully acclimate to UV-B radiation and high visiblelight (PAR, photosynthetically active radiation,400–700 nm).8,155
In terms of the beneficial effects of UV radiation on foodcrops, increased exposure to UV radiation through, forexample, land-use changes and climate change, as well asmanipulations of controlled growth conditions can be exploitedto enhance food quality and nutritional value through inducedchanges in the secondary metabolism of plants.
Visual sensitivity to and damage fromUV radiation in terrestrial animals
The range of wavelengths an animal perceives depends on thespectrum available in the environment, the degree to whichthis is transmitted through the ocular media and the visualpigments found in the retina. Visible light represents the spec-trum perceived by humans but other animals often see adifferent range of “colours” due to visual pigments absorbingelsewhere in the spectrum. UV-vision is used extensively by awide range of invertebrates and vertebrates for critical life pro-cesses including mate selection and location of food resourcesin birds, fish, insects, spiders, and other taxa. Some invert-ebrates are specifically able to detect and respond to UV-B radi-ation under natural conditions.118 The recent discovery thatUV-vision (>300 nm) in mammals may be more widespreadthan previously recognised, suggests the need for moreresearch into the ecological significance of this finding(Douglas and Jeffery,156 and references therein).
As with humans, animals can develop UV-related diseases,although research on these topics tends to be concentrated oneconomically important animals such as cattle. The occur-rence of ocular squamous cell carcinoma (OSCC) or “cancereye” has been reported in cattle worldwide, with 10–20% ofanimals in some Australian herds diagnosed with this disease.OSCC is the most common malignant tumour affecting cattlein North America and is responsible for significant economiclosses (estimated at $20 million per annum in the United Statesalone; reviewed in Tsujita and Plummer157). In 2002, OSCC wasthe third-leading cause of carcass condemnation at slaughter-houses inspected by the US Department of Agriculture. Euro-pean breeds of cattle, particularly those with light coloured skinsuch as Herefords and white-faced Holstein breeds, commonlydevelop OSCC, particularly when they are raised in regions withhigh natural levels of UV-B radiation (e.g. Australia, and the
southwestern USA); see also Ballaré et al.1 While a proportion ofthe disease might be related to increasing UV radiation in areasaffected by ozone depletion, the bulk appears to be caused byagricultural practices (e.g. lack of shade) and the movement ofanimals traditionally bred in low UV-B radiation environmentsto latitudes with higher UV-B radiation.
Progress in technical and experimentalissues
Methodological issues in UV radiation supplementationstudies, where filtered fluorescent UV lamps do not have aspectral output that perfectly matches the solar spectrum, werediscussed previously.1 Briefly, Biological Spectral WeightingFunctions (BSWF), dimensionless factors that represent therelative effectiveness of the different wavelengths of UV radi-ation in influencing a particular biological response, are usedto calculate “biologically effective” UV radiation.59 This hasbeen the only way to compare artificial UV radiation levels inexperiments with solar UV radiation.
A new filter, termed the urate anion liquid filter, has beendeveloped that permits fluorescent UV lamps to approximate sun-light much more closely.158 This new filter removes more of theshortwave UV-B radiation (λ ≤ 305 nm) than traditional cellulosediacetate filters and transmits more longwave UV-B (λ ≥ 310 nm).While this filter was developed for laboratory use in algal studies,there is potential for it to be adapted to other systems.
Accurate measurement of UV radiation is an essentialaspect of experimental work on the effects of UV radiation.59
Ideally these measurements are done with a spectroradio-meter, where radiation is quantified at discrete wavelengths.Usually, double-grating spectroradiometers are used for thesemeasurements, where two gratings are linked in tandem torefine the signal sent to the detector. This double-gratingsystem reduces “stray light” in the instrument. For example, asmall amount of stray visible radiation could swamp radiationof a UV-B wavelength being measured. An instrumentationinnovation that is rapidly supplementing the mechanicaldouble-grating spectroradiometer is the diode-array spectro-radiometer. This instrument has no moving parts and canproduce a spectral irradiance measurement almost instan-taneously. In contrast, typical mechanical double-grating unitsrequire several minutes to complete a scan, during whichirradiation conditions can change.
Since diode-array spectroradiometers are much less costlythan double-grating units, they may often replace broadbandUV detectors in experimental studies. These spectral irradiancedata will be much more valuable than the broadband measure-ments. The unit’s small size and portability is also an asset.However, they have limitations that users need to take intoaccount. Stray light is a considerable issue as these are basicallysingle-grating spectroradiometers. Some features of the instru-ment can help minimise this problem, but measurements infull sunlight may still not be accurate. These issues are dis-cussed at length by Aphalo et al.159 and references therein.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 88–107 | 99

Sensitivity of instruments to temperature is also anissue, especially when conducting measurements in thefield. A 15% change in the sensitivity of a double-gratingspectroradiometer over the range of 11.5 to 33.5 °C canoccur.160 Temperature also affects diode-array units as wave-length accuracy, dark current offset, and spectral responsivityare all influenced by temperature.161 It is possible in somecases to develop correction algorithms for particular units (e.g.Baczynska et al.160), although one must consider whetherdifferential heating of the unit in direct intense sunlight canbe adequately simulated in a test chamber. Ideally spectroradi-ometers used in measuring sunlight should be temperaturestabilised.
Some ecological studies require measurements of time-inte-grated biologically effective UV radiation in environmentswhere there is considerable spatial variability in UV irradiance(e.g. within plant canopies, soil surfaces under plant canopiesand others). These applications require deployment of anumber of inexpensive devices to measure the UV radiation.UV-absorbing polymers that were originally developed ashuman UV dosimeters59,162 have been used to characterise theUV radiation environment of individual leaves in canopies,163
as well as to quantify fine-scale differences in exposure ofplants to UV radiation in heterogeneous habitats.164 Whilethese devices are no substitute for spectroradiometers, theycan provide useful and inexpensive estimates of exposure toUV radiation for certain applications.165 Current polymershave wavelength sensitivities similar to common plant biologi-cal weighting functions166,167 and have been developed withthe inclusion of neutral density filters allowing estimates ofexposure to UV radiation over periods of days to weeks.168 Pro-vided these dosimeters are calibrated against spectroradio-meters at the field locations where they are to be deployed,they may have particular utility for decomposition studies andin areas under the ozone ‘hole’ where long-term continuousmeasurement of the incident UV radiation on standing andground level plant litter is needed.
Increased attention needs to be given to developing tech-niques and approaches that allow for the determination of rea-listic exposures to UV radiation of individual plants, naturalecosystems, and agricultural environments. The spectral sensi-tivities and exposures to UV radiation are needed in models thatevaluate the combined effects of changes in UV radiation,climate, and vegetation on terrestrial ecosystems. The numeroustechnical issues involved in conducting experiments with UVradiation can, however, be intimidating to researchers startingwork in this field. A comprehensive guide to all aspects ofexperimental design, implementation, analysis and instrumen-tation by Aphalo et al.159 will help standardize protocols andensure reliable results that can be meaningfully assessed.
Gaps in knowledge
Since the last assessment,1 evidence of the coupling of ozonedepletion effects and those of climate change2 has been
strengthened, and has revealed the complexity of the inter-actions that are occurring. The success of the Montreal Proto-col has been two-fold, viz., the phasing out of ozone depletingsubstances (ODS) and the contribution to decreasing some ofthe load of greenhouse gases, since many of the phased outODS are themselves greenhouse gases. It should also be recog-nised that exposure to changing levels of UV radiation is notonly ozone-dependent, but also reflects changes in land-useand climate-related phenomena, such as projected changes inrainfall, decreased or increased cloud cover in some regions,and snow and ice melting. These events are likely to affect eco-system functioning and food production, all of which calls fora holistic approach to research that encompasses the role ofUV radiation within a rapidly changing environment. From theresearch to date, evidence is accumulating that these interlink-ing factors of UV radiation, changes in ozone, climate, andenvironment are modifying responses of plants and ecosys-tems. Further evaluation of where the potential tipping pointsor beneficial effects are occurring will increase our under-standing and ability to project potential future effects from theinteractions between exposure to UV radiation and othersimultaneously occurring environmental stresses. Currently,our knowledge of the consequences for the biosphere is farfrom comprehensive. What has become clear is that an inte-grative research approach under realistic conditions is essen-tial for future projections of the response of ecosystems. Formany of the processes discussed in this present paper, moreempirical evidence is needed to determine the ecological sig-nificance of the role played by UV-B radiation in the presenceof other environmental stresses.
Apart from the focus on UV radiation over more than 25years of the Montreal Protocol, ozone depletion has beenimplicated in changes to the climate of the Southern Hemi-sphere, highlighting the need to assess the impact of suchchanges on the ecosystems of this region.7 One of the keyuncertainties here is the extent to which climate is affected byozone depletion versus greenhouse gas forcing and particularlythe seasonality of these effects. Resolving this will improveunderstanding of how ozone recovery will feed back on theseclimate processes and is vital to our ability to model futureecosystems across half of the globe.
Acknowledgements
JFB acknowledges support from Curtin University, WesternAustralia; PWB was supported by the U.S. National ScienceFoundation (DEB 0815897) and Loyola University J.H. MullahyEndowment for Environmental Biology; SAR was supported bythe Australian Government, Department of Environment andthe University of Wollongong. CLB was funded by grants fromCONICET (Consejo Nacional de Investigaciones Científicas yTécnicas), ANPCyT (Agencia Nacional de Promoción Científicay Tecnológica) and UBACyT (Universidad de Buenos AiresCiencia y Técnica).
Perspective Photochemical & Photobiological Sciences
100 | Photochem. Photobiol. Sci., 2015, 14, 88–107 This journal is © The Royal Society of Chemistry and Owner Societies 2015

References
1 C. L. Ballaré, M. M. Caldwell, S. D. Flint, S. A. Robinsonand J. F. Bornman, Effects of solar ultraviolet radiation onterrestrial ecosystems. Patterns, mechanisms, and inter-actions with climate change, Photochem. Photobiol. Sci.,2011, 10, 226–241.
2 C. E. Williamson, R. G. Zepp, R. M. Lucas, S. Madronich,A. T. Austin, C. L. Ballaré, M. Norval, B. Sulzberger,A. F. Bais, R. L. McKenzie, S. A. Robinson, D.-P. Häder,N. D. Paul and J. F. Bornman, Solar ultraviolet radiation ina changing climate, Nat. Clim. Change, 2014, 4, 434–441.
3 R. M. Cory, B. C. Crump, J. A. Dobkowski and G. W. Kling,Surface exposure to sunlight stimulates CO2 release frompermafrost soil carbon in the Arctic, Proc. Natl. Acad.Sci. U. S. A., 2013, 110, 3429–3434.
4 H. Lee, T. Rahn and H. L. Throop, An accounting of C-based trace gas release during abiotic plant litter degra-dation, Global Change Biol., 2012, 18, 1185–1195.
5 D. W. J. Thompson, S. Solomon, P. J. Kushner,M. H. England, K. M. Grise and D. J. Karoly, Signatures ofthe Antarctic ozone hole in Southern Hemisphere surfaceclimate change, Nat. Geosci., 2011, 4, 741–749.
6 J. Turner, N. E. Barrand, T. J. Bracegirdle, P. Convey,D. A. Hodgson, M. Jarvis, A. Jenkins, G. Marshall,M. P. Meredith, H. Roscoe, J. Shanklin, J. French,H. Goosse, M. Guglielmin, J. Gutt, S. Jacobs,M. C. Kennicutt Ii, V. Masson-Delmotte, P. Mayewski,F. Navarro, S. Robinson, T. Scambos, M. Sparrow,C. Summerhayes, K. Speer and A. Klepikov, Antarcticclimate change and the environment: An update, PolarRec., 2014, 50, 237–259.
7 S. A. Robinson and D. J. Erickson III, Not just aboutsunburn–the ozone hole’s profound effect on climate hassignificant implications for Southern Hemisphere ecosys-tems, Global Change Biol., 2014, DOI: 10.1111/gcb.12739.
8 J. J. Wargent, E. M. Elfadly, J. P. Moore and N. D. Paul,Increased exposure to UV-B radiation during early deve-lopment leads to enhanced photoprotection and improvedlong-term performance in Lactuca sativa, Plant, CellEnviron., 2011, 34, 1401–1413.
9 J. J. Wargent and B. R. Jordan, From ozone depletion toagriculture: understanding the role of UV radiation in sus-tainable crop production, New Phytol., 2013, 197, 1058–1076.
10 W. J. Zhang and L. O. Björn, The effect of ultraviolet radi-ation on the accumulation of medicinal compounds inplants, Fitoterapia, 2009, 80, 207–218.
11 B. Mauch-Mani and F. Mauch, The role of abscisic acid inplant-pathogen interactions, Curr. Opin. Plant Biol., 2005,8, 409–414.
12 D. Comont, J. Martinez Abaigar, A. Albert, P. Aphalo,D. R. Causton, F. L. Figueroa, A. Gaberscik, L. Llorens,M.-T. Hauser, M. A. K. Jansen, M. Kardefelt, P. de la CobaLuque, S. Neubert, E. Núñez-Olivera, J. Olsen, M. Robson,M. Schreiner, R. Sommaruga, Å. Strid, S. Torre,
M. Turunen, S. Veljovic-Jovanovic, D. Verdaguer,M. Vidovic, J. Wagner, J. B. Winkler, G. Zipoli andD. Gwynn-Jones, UV responses of Lolium perenne raisedalong a latitudinal gradient across Europe: a filtrationstudy, Physiol. Plant., 2012, 145, 604–618.
13 P. S. Searles, S. D. Flint and M. M. Caldwell, A meta-analy-sis of plant field studies simulating stratospheric ozonedepletion, Oecologia, 2001, 127, 1–10.
14 K. K. Newsham and S. A. Robinson, Responses of plantsin polar regions to UVB exposure: a meta-analysis, GlobalChange Biol., 2009, 15, 2574–2589.
15 T. F. Stocker, D. Qin, G.-K. Plattner, M. Tignor, S. K. Allen,J. Boschung, A. Nauels, Y. Xia, V. Bex and P. M. Midgley,IPCC: Summary for Policymakers, in Climate Change 2013:The Physical Science Basis. Contribution of Working Group Ito the Fifth Assessment Report of the IntergovernmentalPanel on Climate Change, Cambridge University Press,Cambridge, UK and New York, NY, 2013.
16 F. Bussotti, F. Ferrini, M. Pollastrini and A. Fini, The chal-lenge of Mediterranean sclerophyllous vegetation underclimate change: From acclimation to adaptation, Environ.Exp. Bot., 2014, 103, 80–98.
17 WMO, Scientific Assessment of Ozone Depletion: 2010,Global Ozone Research and Monitoring Project-ReportNo. 52, Report No., Geneva, Switzerland, 2011, p. 516.
18 S. Madronich, M. Shao, S. R. Wilson, K. R. Solomon,J. Longstrethe and X. Tang, Changes in air quality andtropospheric composition due to depletion of strato-spheric ozone and interactions with changing climate:Implications for human and environmental health, Photo-chem. Photobiol. Sci., 2015, 14, this issue.
19 L. Guidi, E. Degl’Innocenti, D. Remorini, S. Biricolti,A. Fini, F. Ferrini, F. P. Nicese and M. Tattini, The impactof UV-radiation on the physiology and biochemistry ofLigustrum vulgare exposed to different visible-light irradi-ance, Environ. Exp. Bot., 2011, 70, 88–95.
20 M. M. Caldwell, L. O. Björn, J. F. Bornman,S. D. Flint, G. Kulandaivelu, A. H. Teramura andM. Tevini, Effects of increased solar ultraviolet radi-ation on terrestrial ecosystems, J. Photochem. Photo-biol., B, 1998, 46, 40–52.
21 J. D. Turnbull, S. J. Leslie and S. A. Robinson,Desiccation protects two Antarctic mosses from ultra-violet-B induced DNA damage, Funct. Plant Biol., 2009, 36,214–221.
22 D. Verdaguer, L. Llorens, M. Bernal and J. Badosa, Photo-morphogenic effects of UVB and UVA radiation on leavesof six Mediterranean sclerophyllous woody species sub-jected to two different watering regimes at the seedlingstage, Environ. Exp. Bot., 2012, 79, 66–75.
23 M. Bernal, L. Llorens, J. Badosa and D. Verdaguer, Interac-tive effects of UV radiation and water availability on seed-lings of six woody Mediterranean species, Physiol. Plant.,2013, 147, 234–247.
24 C. A. Mazza, P. I. Gimenez, A. G. Kantolic andC. L. Ballaré, Beneficial effects of solar UV-B radiation on
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 88–107 | 101

soybean yield mediated by reduced insect herbivory underfield conditions, Physiol. Plant., 2013, 147, 307–315.
25 H. Bandurska, J. Niedziela and T. Chadzinikolau, Separateand combined responses to water deficit and UV-B radi-ation, Plant Sci., 2013, 213, 98–105.
26 H. Bandurska and M. Cieslak, The interactive effect ofwater deficit and UV-B radiation on salicylic acid accumu-lation in barley roots and leaves, Environ. Exp. Bot., 2013,94, 9–18.
27 L. He, X. Jia, Z. Gao and R. Li, Genotype-dependentresponses of wheat (Triticum aestivum L.) seedlings todrought, UV-B radiation and their combined stresses,Afr. J. Biotechnol., 2011, 10, 4046–4056.
28 E. A. Kravets, L. B. Zelena, E. P. Zabara and Y. B. Blume,Adaptation strategy of barley plants to UV-B radiation,Emirates J. Food Agric., 2012, 24, 632–645.
29 V. Virjamo, S. Sutinen and R. Julkunen-Tiitto, Combinedeffect of elevated UVB, elevated temperature and fertiliza-tion on growth, needle structure and phytochemistry ofyoung Norway spruce (Picea abies) seedlings, GlobalChange Biol., 2014, 20, 2252–2260.
30 S. Singh, R. Kumari, M. Agrawal and S. B. Agrawal, Modifi-cation in growth, biomass and yield of radish under sup-plemental UV-B at different NPK levels, Ecotoxicol.Environ. Saf., 2011, 74, 897–903.
31 T. S. L. Lau, E. Eno, G. Goldstein, C. Smith andD. A. Christopher, Ambient levels of UV-B in Hawaii com-bined with nutrient deficiency decrease photosynthesis innear-isogenic maize lines varying in leaf flavonoids: Flavo-noids decrease photoinhibition in plants exposed toUV-B, Photosynthetica, 2006, 44, 394–403.
32 X.-R. Guo, B.-W. Chang, Y.-G. Zu and Z.-H. Tang, Theimpacts of increased nitrate supply on Catharanthusroseus growth and alkaloid accumulations under ultra-violet-B stress, J. Plant Interact., 2014, 9, 640–646.
33 J. K. Holopainen and J. Gershenzon, Multiple stressfactors and the emission of plant VOCs, Trends Plant Sci.,2010, 15, 176–184.
34 M. Possell and F. Loreto, The role of volatile organic com-pounds in plant resistance to abiotic stresses: responsesand mechanisms, in Biology, controls and models of treevolatile organic compound emissions, ed. Ü. Niinemets andR. K. Monson, Springer, Berlin, 2013, pp. 209–235.
35 A. Guenther, Upscaling biogenic volatile compound emis-sions from leaves to landscapes, in Biology, controls andmodels of tree volatile organic compound emissions, ed.Ü. Niinemets and R. K. Monson, Springer, Berlin, 2013,pp. 391–414.
36 J. Peñuelas and M. Staudt, BVOCs and global change,Trends Plant Sci., 2010, 15, 133–144.
37 M. J. Potosnak, B. M. Baker, L. LeStourgeon, S. M. Disher,K. L. Griffin, M. S. Bret-Harte and G. Starr, Isoprene emis-sions from a tundra ecosystem, Biogeosciences, 2013, 10,871–889.
38 A. T. Archibald, J. G. Levine, N. L. Abraham, M. C. Cooke,P. M. Edwards, D. E. Heard, M. E. Jenkin, A. Karunaharan,
R. C. Pike, P. S. Monks, D. E. Shallcross, P. J. Telford,L. K. Whalley and J. A. Pyle, Impacts of HOx regenerationand recycling in the oxidation of isoprene: Consequencesfor the composition of past, present and future atmos-pheres, Geophys. Res. Lett., 2011, 38, GL046520.
39 J. Llusia, L. Llorens, M. Bernal, D. Verdaguer andJ. Penuelas, Effects of UV radiation and water limitationon the volatile terpene emission rates, photosynthesisrates, and stomatal conductance in four Mediterraneanspecies, Acta Physiol. Plant., 2012, 34, 757–769.
40 T. D. Sharkey, Is it useful to ask why plants emit iso-prene?, Plant, Cell Environ., 2013, 36, 517–520.
41 P. Tiiva, R. Rinnan, P. Faubert, J. Räsänen, T. Holopainen,E. Kyro and J. K. Holopainen, Isoprene emission from asubarctic peatland under enhanced UV-B radiation, NewPhytol., 2007, 176, 346–355.
42 P. Faubert, P. Tiiva, A. Rinnan, J. Räsänen,J. K. Holopainen, T. Holopainen, E. Kyrö and R. Rinnan,Non-methane biogenic volatile organic compound emis-sions from a subarctic peatland under enhanced UV-Bradiation, Ecosystems, 2010, 13, 860–873.
43 D. J. Erickson III, B. Sulzberger, R. Zepp, A. T. Austin andN. Paul, Effects of stratospheric ozone depletion, solar UVradiation, and climate change on biogeochemical cycling:Interactions and feedbacks, Photochem. Photobiol. Sci.,2015, 14, this issue.
44 F. Loreto and J. P. Schnitzler, Abiotic stresses and inducedBVOCs, Trends Plant Sci., 2010, 15, 154–166.
45 R. Rinnan, S. Saarnio, J. K. Haapala, S. K. Mörsky,P. J. Martikainen, J. Silvola and T. Holopainen, Borealpeatland ecosystems under enhanced UV-B radiation andelevated tropospheric ozone concentration, Environ. Exp.Bot., 2013, 90, 43–52.
46 Z. P. Wang, S. X. Chang, H. Chen and X. G. Han, Wide-spread non-microbial methane production by organiccompounds and the impact of environmental stresses,Earth-Sci. Rev., 2013, 127, 193–202.
47 F. Keppler, J. T. G. Hamilton, M. Brass and T. Rockmann,Methane emissions from terrestrial plants under aerobicconditions, Nature, 2006, 439, 187–191.
48 A. McLeod and F. Keppler, Vegetation, in Methane andClimate Change, ed. D. S. Reay, P. Smith and A. vanAmstel, Earthscan, London and Washington, D.C., 2010,pp. 74–96.
49 A. A. Bloom, J. Lee-Taylor, S. Madronich, D. J. Messenger,P. I. Palmer, D. S. Reay and A. R. McLeod, Global methaneemission estimates from ultraviolet irradiation of terres-trial plant foliage, New Phytol., 2010, 187, 417–425.
50 R. G. Zepp, D. J. Erickson, N. D. Paul and B. Sulzberger,Effects of solar UV radiation and climate change on bio-geochemical cycling: interactions and feedbacks, Photo-chem. Photobiol. Sci., 2011, 10, 261–279.
51 A. R. McLeod, S. C. Fry, G. J. Loake, D. J. Messenger,D. S. Reay, K. A. Smith and B.-W. Yun, Ultraviolet radi-ation drives methane emissions from terrestrial plantpectins, New Phytol., 2008, 180, 124–132.
Perspective Photochemical & Photobiological Sciences
102 | Photochem. Photobiol. Sci., 2015, 14, 88–107 This journal is © The Royal Society of Chemistry and Owner Societies 2015

52 D. Bruhn, T. N. Mikkelsen, M. M. M. Rolsted, H. Egsgaardand P. Ambus, Leaf surface wax is a source of plantmethane formation under UV radiation and in the pre-sence of oxygen, Plant Biol., 2014, 16, 512–516.
53 D. Bruhn, I. M. Møller, T. N. Mikkelsen and P. Ambus,Terrestrial plant methane production and emission,Physiol. Plant., 2012, 144, 201–209.
54 F. Althoff, A. Jugold and F. Keppler, Methane formation byoxidation of ascorbic acid using iron minerals and hydro-gen peroxide, Chemosphere, 2010, 80, 286–292.
55 D. J. Messenger, A. R. McLeod and S. C. Fry, The role ofultraviolet radiation, photosensitizers, reactive oxygenspecies and ester groups in mechanisms of methane for-mation from pectin, Plant, Cell Environ., 2009, 32, 1–9.
56 S. R. Pangala, S. Moore, E. R. C. Hornibrook and V. Gauci,Trees are major conduits for methane egress from tropicalforested wetlands, New Phytol., 2013, 197, 524–531.
57 S. K. Mörsky, J. K. Haapala, R. Rinnan, S. Saarnio,H. Suokanerva, K. Latola, E. Kyrö, J. Silvola, T. Holopainenand P. J. Martikainen, Minor long-term effects of ultra-violet-B radiation on methane dynamics of a subarctic fenin Northern Finland, Biogeochemistry, 2012, 108, 233–243.
58 Y. Lou, W. Zhou and L. Ren, Elevated UV-B radiationincreased CH4 emission in transgenic rice from a paddysoil, Agric., Ecosyst. Environ., 2012, 151, 16–20.
59 A. F. Bais, R. L. McKenzie, P. J. Aucamp, M. Ilyas,S. Madronich, G. Bernhard and K. Tourpali, Ozonedepletion and climate change: Impacts on UV radiation,Photochem. Photobiol. Sci., 2015, 14, this issue.
60 Y. Wu, L. M. Polvani and R. Seager, The Importance of theMontreal Protocol in Protecting Earth’s Hydroclimate,J. Clim., 2013, 26, 4049–4068.
61 J. Turner, R. A. Bindschadler, P. Convey, G. Di Prisco,E. Fahrbach, J. Gutt, D. A. Hodgson, P. A. Mayewski andC. P. Summerhayes, Antarctic Climate Change and theEnvironment, SCAR, Cambridge, 2009, http://acce.scar.org/wiki/Antarctic_Climate_Change_and_the_Environment,accessed 1/11/2014.
62 E. R. Thomas, T. J. Bracegirdle, J. Turner and E. W. Wolff,A 308 year record of climate variability in West Antarctica,Geophys. Res. Lett., 2013, 40, 5492–5496.
63 P. Convey, R. Bindschadler, G. di Prisco, E. Fahrbach,J. Gutt, D. A. Hodgson, P. A. Mayewski,C. P. Summerhayes and J. Turner, Antarctic climatechange and the environment, Antarct. Sci., 2009, 21, 541–563.
64 H. Korhonen, K. S. Carslaw, P. M. Forster, S. Mikkonen,N. D. Gordon and H. Kokkola, Aerosol climate feedbackdue to decadal increases in Southern Hemisphere windspeeds, Geophys. Res. Lett., 2010, 37, GL041320.
65 J. Turner and G. J. Marshall, Climate Change in the PolarRegions, Cambridge University Press, Cambridge, 2011.
66 D. A. Dixon, P. A. Mayewski, I. D. Goodwin, G. J. Marshall,R. Freeman, K. A. Maasch and S. B. Sneed, An ice-coreproxy for northerly air mass incursions into West Antarc-tica, Int. J. Climatol., 2012, 32, 1455–1465.
67 N. J. Abram, R. Mulvaney, F. Vimeux, S. J. Phipps,J. Turner and M. H. England, Evolution of the SouthernAnnular Mode during the past millennium, Nat. Clim.Change, 2014, 4, 564–569.
68 P. Convey, S. L. Chown, A. Clarke, D. K. A. Barnes,S. Bokhorst, V. Cummings, H. W. Ducklow, F. Frati,T. G. A. Green, S. Gordon, H. J. Griffiths, C. Howard-Williams, A. H. L. Huiskes, J. Laybourn-Parry, W. B. Lyons,A. McMinn, S. A. Morley, L. S. Peck, A. Quesada,S. A. Robinson, S. Schiaparelli and D. H. Wall, The spatialstructure of Antarctic biodiversity, Ecol. Monogr., 2014, 84,203–244.
69 L. J. Clarke, S. A. Robinson, Q. Hua, D. J. Ayre andD. Fink, Radiocarbon bomb spike reveals biologicaleffects of Antarctic climate change, Global Change Biol.,2012, 18, 301–310.
70 D. A. Hodgson, D. Roberts, A. McMinn, E. Verleyen,B. Terry, C. Corbett and W. Vyverman, Recent rapid sali-nity rise in three East Antarctic lakes, J. Paleolimnol., 2006,36, 385–406.
71 M. Cataldo, H. Evangelista, J. C. Simoes, R. H. M. Godoi,I. Simmonds, M. H. Hollanda, I. Wainer, F. Aquino andR. Van Grieken, Mineral dust variability in central WestAntarctica associated with ozone depletion, Atmos. Chem.Phys., 2013, 13, 2165–2175.
72 J. R. McConnell, A. J. Aristarain, J. R. Banta, P. R. Edwardsand J. C. Simões, 20th-Century doubling in dust archivedin an Antarctic Peninsula ice core parallels climate changeand desertification in South America, Proc. Natl. Acad.Sci. U. S. A., 2007, 104, 5743–5748.
73 R. Villalba, A. Lara, M. H. Masiokas, R. Urrutia,B. H. Luckman, G. J. Marshall, I. A. Mundo, D. A. Christie,E. R. Cook, R. Neukom, K. Allen, P. Fenwick,J. A. Boninsegna, A. M. Srur, M. S. Morales, D. Araneo,J. G. Palmer, E. Cuq, J. C. Aravena, A. Holz andC. LeQuesne, Unusual Southern Hemisphere tree growthpatterns induced by changes in the Southern AnnularMode, Nat. Geosci., 2012, 5, 793–798.
74 C. Böning, A. Dispert, M. Visbeck, R. SR andF. Schwarzkopf, The response of the Antarctic Circum-polar Current to recent climate change, Nat. Geosci., 2008,1, 864–869.
75 A. Purich and S. W. Son, Impact of Antarctic ozonedepletion and recovery on Southern Hemisphere precipi-tation, evaporation, and extreme changes, J. Clim., 2012,25, 3145–3154.
76 S. M. Kang, L. M. Polvani, J. C. Fyfe, S. W. Son,M. Sigmond and G. J. P. Correa, Modeling evidence thatozone depletion has impacted extreme precipitation inthe austral summer, Geophys. Res. Lett., 2013, 40, 4054–4059.
77 IPCC, Climate Change 2014: Impacts, Adaptation, andVulnerability. Contribution of Working Group II to theFifth Assessment Report of the Intergovernmental Panel onClimate Change. WGII AR5 Summary for Policymakers.,Report No., IPCC, Geneva, Switzerland, 2014, p. 44.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 88–107 | 103

78 D. Manatsa, Y. Morioka, S. K. Behera, T. Yamagata andC. H. Matarira, Link between Antarctic ozone depletionand summer warming over southern Africa, Nat. Geosci.,2013, 6, 934–939.
79 J. Perlwitz, ATMOSPHERIC SCIENCE Tug of war on the jetstream, Nat. Clim. Change, 2011, 1, 29–31.
80 J. Y. King, L. A. Brandt and E. C. Adair, Shedding light onplant litter decomposition: advances, implications andnew directions in understanding the role of photodegra-dation, Biogeochemistry, 2012, 111, 57–81.
81 X. Song, C. Peng, H. Jiang, Q. Zhu and W. Wang, Directand indirect effects of UV-B exposure on litter decompo-sition: a meta-analysis, PLoS One, 2013, 8, e68858.
82 P. W. Barnes, H. L. Throop, S. R. Archer, D. D. Breshears,R. L. McCulley and M. A. Tobler, Sunlight and soil-littermixing: Drivers of litter decomposition in drylands, Prog.Bot., 2015, 76, 273–302.
83 H. A. L. Henry, K. Brizgys and C. B. Field, Litter decompo-sition in a California annual grassland: Interactionsbetween photodegradation and litter layer thickness, Eco-systems, 2008, 11, 545–554.
84 Y. Lin and J. Y. King, Effects of UV exposure and litterposition on decomposition in a California grassland, Eco-systems, 2014, 17, 158–168.
85 P. W. Barnes, H. L. Throop, D. B. Hewins, M. L. Abbeneand S. R. Archer, Soil coverage reduces photodegradationand promotes the development of soil-microbial films ondryland leaf litter, Ecosystems, 2012, 15, 311–321.
86 A. T. Austin and C. L. Ballaré, Dual role of lignin in plantlitter decomposition in terrestrial ecosystems, Proc. Natl.Acad. Sci. U. S. A., 2010, 107, 4618–4622.
87 G. W. Schade, R. M. Hofmann and P. J. Crutzen, CO emis-sions from degrading plant matter (I). Measurements,Tellus, Ser. B, 1999, 51, 889–908.
88 Q. Gao and F. Garcia-Pichel, Microbial ultraviolet sun-screens, Nat. Rev. Microbiol., 2011, 9, 791–802.
89 X. Song, H. L. Zhang, H. Jiang, L. A. Donaldson andH. L. Wang, Influence of elevated UV-B radiation onleaf litter chemistry and subsequent decomposition inhumid subtropical China, J. Soils Sediments, 2013, 13,846–853.
90 L. Selbmann, D. Isola, L. Zucconi and S. Onofri, Resist-ance to UV-B induced DNA damage in extreme-tolerantcryptoendolithic Antarctic fungi: detection by PCR assays,Fungal Biol., 2011, 115, 937–944.
91 S. A. Alharbi, How do bacteria survive UV irradiation inthe stratosphere?, J. Food Agric. Environ., 2012, 10, 843–845.
92 H. Kadivar and A. E. Stapleton, Ultraviolet radiation altersmaize phyllosphere bacterial diversity, Microb. Ecol., 2003,45, 353–361.
93 N. D. Paul, J. P. Moore, M. McPherson, C. Lambourne,P. Croft, J. C. Heaton and J. J. Wargent, Ecologicalresponses to UV radiation: interactions between the bio-logical effects of UV on plants and on associated orga-nisms, Physiol. Plant., 2012, 145, 565–581.
94 W. K. Smith, W. Gao, H. Steltzer, M. D. Wallenstein andR. Tree, Moisture availability influences the effect of ultra-violet-B radiation on leaf litter decomposition, GlobalChange Biol., 2010, 16, 484–495.
95 B. Foereid, J. Bellarby, W. Meier-Augenstein and H. Kemp,Does light exposure make plant litter more degradable?,Plant Soil, 2010, 333, 275–285.
96 S. Ma, D. Baldocchi, J. Hatala, J. M. Detto and J. Yuste, Arerain-induced ecosystem respiration pulses enhanced bylegacies of antecedent photodegradation in semi-aridenvironments?, Agric. For. Meteorol., 2012, 154–155, 203–213.
97 L. M. Mayer, K. R. Thornton, L. L. Schick, J. D. Jastrowand J. W. Harden, Photodissolution of soil organicmatter, Geoderma, 2012, 170, 314–321.
98 G. Agati, C. Brunetti, M. Di Ferdinando, F. Ferrini,S. Pollastri and M. Tattini, Functional roles of flavonoidsin photoprotection: New evidence, lessons from the past,Plant Physiol. Biochem., 2013, 72, 35–45.
99 L. A. Brandt, C. Bohnet and J. Y. King, Photochemicallyinduced carbon dioxide production as a mechanism forcarbon loss from plant litter in arid ecosystems,J. Geophys. Res.: Biogeosci., 2009, 114, G02004.
100 S. Rutledge, D. I. Campbell, D. Baldocchi andL. A. Schipper, Photodegradation leads to increasedcarbon dioxide losses from terrestrial organic matter,Global Change Biol., 2010, 16, 3065–3074.
101 S. M. Uselman, K. A. Snyder, R. R. Blank and T. J. Jones,UVB exposure does not accelerate rates of litter decompo-sition in a semi-arid riparian ecosystem, Soil Biol.Biochem., 2011, 43, 1254–1265.
102 W. Parton, W. L. Silver, I. C. Burke, L. Grassens,M. E. Harmon, W. S. Currie, J. Y. King, E. C. Adair,L. A. Brandt, S. C. Hart and B. Fasth, Global-scale simi-larities in nitrogen release patterns during long-termdecomposition, Science, 2007, 315, 361–364.
103 E. C. Adair, W. J. Parton, S. J. Del Grosso, W. L. Silver,M. E. Harmon, S. A. Hall, I. C. Burke and S. C. Hart,Simple three-pool model accurately describes patterns oflong-term litter decomposition in diverse climates, GlobalChange Biol., 2008, 14, 2636–2660.
104 A. T. Austin, Has water limited our imagination for arid-land biogeochemistry?, Trends Ecol. Evol., 2011, 26, 229–235.
105 X. Song, H. Jiang, Z. Zhang, G. Zhou, S. Zhang andC. Peng, Interactive effects of elevated UV-B radiation andN deposition on decomposition of Moso bamboo litter,Soil Biol. Biochem., 2014, 69, 11–16.
106 M. M. Caldwell, J. F. Bornman, C. L. Ballaré, S. D. Flint andG. Kulandaivelu, Terrestrial ecosystems, increased solarultraviolet radiation, and interactions with otherclimate change factors, Photochem. Photobiol. Sci., 2007, 6,252–266.
107 X. Bao, Q. Li, J. Hua, T. Zhao and W. Liang, Interactiveeffects of elevated ozone and UV-B radiation on soil nema-tode diversity, Ecotoxicology, 2014, 23, 11–20.
Perspective Photochemical & Photobiological Sciences
104 | Photochem. Photobiol. Sci., 2015, 14, 88–107 This journal is © The Royal Society of Chemistry and Owner Societies 2015

108 B. Li, A. Krumbein, S. Neugart, L. Li and M. Schreiner,Mixed cropping with maize combined with moderateUV-B radiations lead to enhanced flavonoid productionand root growth in faba bean, J. Plant Interact., 2012, 7,333–340.
109 S. Cesco, G. Neumann, N. Tomasi, R. Pinton andL. Weisskopf, Release of plant-borne flavonoids into therhizosphere and their role in plant nutrition, Plant Soil,2010, 329, 1–25.
110 S. Hassan and U. Mathesius, The role of flavonoids inroot-rhizosphere signalling: opportunities and challengesfor improving plant-microbe interactions, J. Exp. Bot.,2012, 63, 3429–3444.
111 S. Chen, S. Xiao and R. M. Callaway, Light intensity altersthe allelopathic effects of an exotic invader, Plant Ecol.Diversity, 2012, 5, 521–526.
112 S. Sharma and K. N. Guruprasad, Enhancement of rootgrowth and nitrogen fixation in Trigonella by UV-exclusionfrom solar radiation, Plant Physiol. Biochem., 2012, 61, 97–102.
113 U. N. Nielsen and D. H. Wall, The future of soil invert-ebrate communities in polar regions: different climatechange responses in the Arctic and Antarctic?, Ecol. Lett.,2013, 16, 409–419.
114 C. L. Ballaré, Light regulation of plant defense, Annu. Rev.Plant Biol., 2014, 65, 335–363.
115 C. L. Ballaré, C. A. Mazza, A. T. Austin and R. Pierik,Canopy Light and Plant Health, Plant Physiol., 2012, 160,145–155.
116 F. Kuhlmann and C. Müller, Impacts of ultraviolet radi-ation on interactions between plants and herbivorousInsects: A chemo-ecological perspective, Prog. Bot., 2011,72, 305–347.
117 C. A. Mazza, M. M. Izaguirre, J. Zavala, A. L. Scopel andC. L. Ballaré, Insect perception of ambient ultraviolet-Bradiation, Ecol. Lett., 2002, 5, 722–726.
118 C. A. Mazza, M. M. Izaguirre, J. Curiale and C. L. Ballaré,A look into the invisible. Ultraviolet-B sensitivity in aninsect (Caliothrips phaseoli) revealed through a behaviouralaction spectrum, Proc. R. Soc. London, Ser. B, 2010, 277,367–373.
119 T. S. Gunasekera, N. D. Paul and P. G. Ayres, The effects ofultraviolet-B (UV-B: 290–320 nm) radiation on blisterblight disease of tea (Camellia sinensis), Plant Pathol.,1997, 46, 179–185.
120 P. V. Demkura and C. L. Ballaré, UVR8 Mediates UV-B-Induced Arabidopsis Defense Responses against Botrytiscinerea by Controlling Sinapate Accumulation, Mol. Plant,2012, 5, 642–652.
121 M. C. Rousseaux, R. Julkunen-Tiitto, P. S. Searles,A. L. Scopel, P. J. Aphalo and C. L. Ballaré, Solar UV-Bradiation affects leaf quality and insect herbivory in thesouthern beech tree Nothofagus antarctica, Oecologia,2004, 138, 505–512.
122 P. V. Demkura, G. Abdala, I. T. Baldwin and C. L. Ballaré,Jasmonate-dependent and -independent pathways
mediate specific effects of solar ultraviolet-B radiation onleaf phenolics and antiherbivore defense, Plant Physiol.,2010, 152, 1084–1095.
123 S. T. Đinh, I. Gális and I. T. Baldwin, UVB radiation and17-hydroxygeranyllinalool diterpene glycosides providedurable resistance against mirid (Tupiocoris notatus)attack in field-grown Nicotiana attenuata plants, Plant, CellEnviron., 2013, 36, 590–606.
124 J. W. Stratmann, B. A. Stelmach, E. W. Weiler andC. A. Ryan, UVB/UVA radiation activates a 48 kDa myelinbasic protein kinase and potentiates wound signaling intomato leaves, Photochem. Photobiol., 2000, 71, 116–123.
125 M. M. Izaguirre, A. L. Scopel, I. T. Baldwin andC. L. Ballaré, Convergent responses to stress. Solar ultra-violet-B radiation and Manduca sexta herbivory elicit over-lapping transcriptional responses in field-grown plants ofNicotiana longiflora, Plant Physiol., 2003, 132, 1755–1767.
126 L. Rizzini, J. J. Favory, C. Cloix, D. Faggionato, A. O’Hara,E. Kaiserli, R. Baumeister, E. Schafer, F. Nagy,G. I. Jenkins and R. Ulm, Perception of UV-B by the Arabi-dopsis UVR8 Protein, Science, 2011, 332, 103–106.
127 G. I. Jenkins, The UV-B Photoreceptor UVR8: From Struc-ture to Physiology, Plant Cell, 2014, 26, 21–37.
128 L. O. Morales, M. Brosché, J. Vainonen, G. I. Jenkins,J. J. Wargent, N. Sipari, A. Strid, A. V. Lindfors,R. Tegelberg and P. J. Aphalo, Multiple roles for UVRESISTANCE LOCUS8 in regulating gene expression andmetabolite accumulation in Arabidopsis under solar ultra-violet radiation, Plant Physiol., 2013, 161, 744–759.
129 C. T. Ruhland, M. J. Dyslin and J. D. Krenz, Wyoming bigsagebrush screens ultraviolet radiation more effectively athigher elevations, J. Arid. Environ., 2013, 96, 19–22.
130 P. W. Barnes, A. R. Kersting, S. D. Flint, W. Beyschlag andR. J. Ryel, Adjustments in epidermal UV-transmittance ofleaves in sun-shade transitions, Physiol. Plant., 2013, 149,200–213.
131 L. J. Clarke and S. A. Robinson, Cell wall-bound ultra-violet-screening compounds explain the high ultraviolettolerance of the Antarctic moss, Ceratodon purpureus, NewPhytol., 2008, 179, 776–783.
132 S. A. Robinson and M. J. Waterman, Sunsafe bryophytes:photoprotection from excess and damaging solar radi-ation, Adv. Photosynth. Respir., 2014, 37, 113–130.
133 P. W. Barnes, S. D. Flint, J. R. Slusser, W. Gao andR. J. Ryel, Diurnal changes in epidermal UV transmittanceof plants in naturally high UV environments, Physiol.Plant., 2008, 133, 363–372.
134 S. Sumbele, M. N. Fotelli, D. Nikolopoulos, G. Tooulakou,V. Liakoura, G. Liakopoulos, P. Bresta, E. Dotsika,M. A. Adams and G. Karabourniotis, Photosyntheticcapacity is negatively correlated with the concentration ofleaf phenolic compounds across a range of differentspecies, AoB PLANTS, 2012, 2012, DOI: 10.1093/aobpla/pls025.
135 T. L. Turnbull, A. Barlow and M. A. Adams, Photosyntheticbenefits of ultraviolet-A to Pimelea ligustrina, a woody
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 88–107 | 105

shrub of sub-alpine Australia, Oecologia, 2013, 173, 375–385.
136 M. L. Falcone-Ferreyra, S. P. Rius and P. Casati, Flavo-noids: biosynthesis, biological functions, and biotechno-logical applications, Front. Plant Sci., 2012, 3, 222.
137 D. Magri, Past UV-B flux from fossil pollen: prospects forclimate, environment and evolution, New Phytol., 2011,192, 310–312.
138 B. H. Lomax, W. T. Fraser, M. A. Sephton, T. V. Callaghan,S. Self, M. Harfoot, J. A. Pyle, C. H. Wellman andD. J. Beerling, Plant spore walls as a record of long-termchanges in ultraviolet-B radiation, Nat. Geosci., 2008, 1,592–596.
139 J. Rozema, P. Blokker, M. A. Mayoral Fuertes andR. Broekman, UV-B absorbing compounds in present-dayand fossil pollen, spores, cuticles, seed coats and wood:evaluation of a proxy for solar UV radiation, Photochem.Photobiol. Sci., 2009, 8, 1233–1243.
140 B. H. Lomax, W. T. Fraser, G. Harrington, S. Blackmore,M. A. Sephton and N. B. W. Harris, A novel palaeoaltime-try proxy based on spore and pollen wall chemistry, EarthPlanet. Sci. Lett., 2012, 353–354, 22–28.
141 J. Royles, J. Ogée, L. Wingate, D. A. Hodgson, P. Conveyand H. Griffiths, Carbon isotope evidence for recentclimate-related enhancement of CO2 assimilation andpeat accumulation rates in Antarctica, Global Change Biol.,2012, 18, 3112–3124.
142 G. J. Jordan, A critical framework for the assessment ofbiological palaeoproxies: predicting past climate andlevels of atmospheric CO2 from fossil leaves, New Phytol.,2011, 192, 29–44.
143 K. J. Willis, A. Feurdean, H. J. B. Birks, A. E. Bjune,E. Breman, R. Broekman, J. A. Grytnes, M. New,J. S. Singarayer and J. Rozema, Quantification of UV-B fluxthrough time using UV-B-absorbing compounds con-tained in fossil Pinus sporopollenin, New Phytol., 2011, 192,553–560.
144 Y. X. Wang and M. Frei, Stressed food - The impact ofabiotic environmental stresses on crop quality, Agric.,Ecosyst. Environ., 2011, 141, 271–286.
145 D. Gwynn-Jones, A. G. Jones, A. Waterhouse, A. Winters,D. Comont, J. Scullion, R. Gardias, B. J. Graae, J. A. Leeand T. V. Callaghan, Enhanced UV-B and Elevated CO2
Impacts Sub-Arctic Shrub Berry Abundance, Quality andSeed Germination, Ambio, 2012, 41(Suppl. 3), 256–268.
146 M. Schreiner, I. Mewis, S. Huyskens-Keil, M. A. K. Jansen,R. Zrenner, J. B. Winkler, N. O’Brien and A. Krumbein,UV-B-induced secondary plant metabolites - potentialbenefits for plant and human health, Crit. Rev. PlantSci.,2012, 31, 229–240.
147 B. Liu, X.-B. Liu, Y.-S. Li and S. J. Herbert, Effects ofenhanced UV-B radiation on seed growth characteristicsand yield components in soybean, Field Crops Res., 2013,154, 158–163.
148 J. A. Zavala, C. A. Mazza, F. M. Dillon, H. D. Chludil andC. L. Ballaré, Soybean resistance to stink bugs (Nezara viri-
dula and Piezodorus guildinii) increases with exposure tosolar UV-B radiation and correlates with isoflavonoidcontent in pods under field conditions, Plant, CellEnviron., 2014, DOI: 10.1111/pce.12368.
149 Y. Dolzhenko, C. M. Bertea, A. Occhipinti, S. Bossi andM. E. Maffei, UV-B modulates the interplay between terpe-noids and flavonoids in peppermint (Mentha piperita L.),J. Photochem. Photobiol., B, 2010, 100, 67–75.
150 S. M. Gregan, J. J. Wargent, L. Liu, J. Shinkle,R. Hofmann, C. Winefield, M. Trought and B. Jordan,Effects of solar ultraviolet radiation and canopymanipulation on the biochemical composition ofSauvignon Blanc grapes, Aust. J. Grape Wine Res., 2012,18, 227–238.
151 P. Carbonell-Bejerano, M. P. Diago, J. Martinez-Abaigar,J. M. Martinez-Zapater, J. Tardaguila and E. Nunez-Olivera, Solar ultraviolet radiation is necessary to enhancegrapevine fruit ripening transcriptional and phenolicresponses, BMC Plant Biol., 2014, 14, 183.
152 K. Suklje, G. Antalick, Z. Coetzee, L. M. Schmidtke,H. B. Cesnik, J. Brandt, W. J. du Toit, K. Lisjak andA. Deloire, Effect of leaf removal and ultraviolet radiationon the composition and sensory perception of Vitis vini-fera L. cv. Sauvignon Blanc wine, Aust. J. Grape Wine Res.,2014, 20, 223–233.
153 M. Gil, R. Bottini, F. Berli, M. Pontin, M. Fernanda Silvaand P. Piccoli, Volatile organic compounds characterizedfrom grapevine (Vitis vinifera L. cv. Malbec) berriesincrease at pre-harvest and in response to UV-B radiation,Phytochemistry, 2013, 96, 148–157.
154 F. J. Berli, R. Alonso, R. Bressan-Smith and R. Bottini,UV-B impairs growth and gas exchange in grapevinesgrown in high altitude, Physiol. Plant., 2013, 149, 127–140.
155 J. J. Wargent, B. C. W. Nelson, T. K. McGhie andP. W. Barnes, Acclimation to UV-B radiation and visiblelight in Lactuca sativa involves up-regulation of photosyn-thetic performance and orchestration of metabolome-wide responses, Plant, Cell Environ., 2014, DOI: 10.1111/pce.12392.
156 R. H. Douglas and G. Jeffery, The spectral transmission ofocular media suggests ultraviolet sensitivity is widespreadamong mammals, Proc. R. Soc. London, Ser. B, 2014, 281,20132995.
157 H. Tsujita and C. E. Plummer, Bovine ocular squamouscell carcinoma, Vet. Clin. North Am. Food Anim. Pract.,2010, 26, 511–529.
158 P. Sampath-Wiley and L. S. Jahnke, A new filter that accu-rately mimics the solar UV-B spectrum using standard UVlamps: the photochemical properties, stabilization anduse of the urate anion liquid filter, Plant, Cell Environ.,2011, 34, 261–269.
159 P. J. Aphalo, A. Albert, L. O. Björn, A. McLeod,T. M. Robson and E. Rosenqvist, Beyond the visible: Ahandbook of best practice in plant UV photobiology, inCOST Action FA0906 UV4growth, University of Helsinki,Department of Biosciences, Division of Plant Biology,
Perspective Photochemical & Photobiological Sciences
106 | Photochem. Photobiol. Sci., 2015, 14, 88–107 This journal is © The Royal Society of Chemistry and Owner Societies 2015

Helsinki, Finland, 2012, p. 176, https://helda.helsinki.fi/handle/10138/37558.
160 K. Baczynska, J. B. O’Hagan, A. J. Pearson and P. Eriksen,Temperature Correction of UV Spectral Solar Measure-ments for ICEPURE Project, Photochem. Photobiol., 2011,87, 1464–1467.
161 G. Seckmeyer, A. Bais, G. Bernhard, M. Blumthaler,S. Drüke, P. Kiedron, K. Lantz, R. McKenzie andS. Riechelmann, Instruments to Measure Solar UltravioletRadiation WMO/TD-No. 1538, Part 4: Array Spectroradio-meters (vol. No. 191), World Meteorological Organization,Geneva, 2010.
162 R. M. Lucas, M. Norval, R. E. Neale, A. R. Young,F. R. de Gruijl, Y. Takizawa and J. C. van der Leun, Theconsequences for human health of stratospheric ozonedepletion in association with other environmental factors,Photochem. Photobiol. Sci., 2015, 14, this issue.
163 A. V. Parisi, P. Schouten, N. J. Downs and J. Turner, SolarUV exposures measured simultaneously to all arbitrarilyoriented leaves on a plant, J. Photochem. Photobiol., B,2010, 99, 87–92.
164 A. V. Parisi, M. G. Kimlin, J. C. F. Wong and M. Wilson,Solar ultraviolet exposures at ground level in tree shadeduring summer in south east Queensland, Int. J. Environ.Health Res., 2001, 11, 117–127.
165 G. Seckmeyer, M. Klingebiel, S. Riechelmann, I. Lohse,R. L. McKenzie, J. Ben Liley, M. W. Allen, A.-M. Siani andG. R. Casale, A critical assessment of two types of per-sonal UV dosimeters, Photochem. Photobiol., 2012, 88,215–222.
166 J. Turner, A. V. Parisi and D. J. Turnbull, Dosimeter forthe measurement of plant damaging solar UV exposures,Agric. For. Meteorol., 2009, 149, 1301–1306.
167 M. M. Caldwell, Solar UV irradiation and the growth anddevelopment of higher plants, Photophysiology, 1971, 6,131–177.
168 A. V. Parisi, P. Schouten and D. J. Turnbull, UV dosimeterbased on Polyphenylene Oxide for the measurement ofUV exposures to plants and humans over extendedperiods, in NIWA 2010 UV Workshop: UV radiation and itseffects-an update 2010, Queenstown, New Zealand, 2010,pp. 25–26.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 88–107 | 107

Photochemical &Photobiological Sciences
PERSPECTIVE
Cite this: Photochem. Photobiol. Sci.,2015, 14, 108
Received 20th October 2014,Accepted 20th October 2014
DOI: 10.1039/c4pp90035a
www.rsc.org/pps
Effects of UV radiation on aquatic ecosystems andinteractions with other environmental factors
Donat-P. Häder,*a Craig E. Williamson,b Sten-Åke Wängberg,c Milla Rautio,d
Kevin C. Rose,e Kunshan Gao,f E. Walter Helbling,g Rajeshwar P. Sinhah andRobert Worresti
Interactions between climate change and UV radiation are having strong effects on aquatic ecosystems due
to feedback between temperature, UV radiation, and greenhouse gas concentration. Higher air tempera-
tures and incoming solar radiation are increasing the surface water temperatures of lakes and oceans, with
many large lakes warming at twice the rate of regional air temperatures. Warmer oceans are changing habi-
tats and the species composition of many marine ecosystems. For some, such as corals, the temperatures
may become too high. Temperature differences between surface and deep waters are becoming greater.
This increase in thermal stratification makes the surface layers shallower and leads to stronger barriers to
upward mixing of nutrients necessary for photosynthesis. This also results in exposure to higher levels of UV
radiation of surface-dwelling organisms. In polar and alpine regions decreases in the duration and amount
of snow and ice cover on lakes and oceans are also increasing exposure to UV radiation. In contrast, in lakes
and coastal oceans the concentration and colour of UV-absorbing dissolved organic matter (DOM) from
terrestrial ecosystems is increasing with greater runoff from higher precipitation and more frequent extreme
storms. DOM thus creates a refuge from UV radiation that can enable UV-sensitive species to become
established. At the same time, decreased UV radiation in such surface waters reduces the capacity of solar
UV radiation to inactivate viruses and other pathogens and parasites, and increases the difficulty and price
of purifying drinking water for municipal supplies. Solar UV radiation breaks down the DOM, making it more
available for microbial processing, resulting in the release of greenhouse gases into the atmosphere. In
addition to screening solar irradiance, DOM, when sunlit in surface water, can lead to the formation of reac-
tive oxygen species (ROS). Increases in carbon dioxide are in turn acidifying the oceans and inhibiting the
ability of many marine organisms to form UV-absorbing exoskeletons. Many aquatic organisms use adaptive
strategies to mitigate the effects of solar UV-B radiation (280–315 nm), including vertical migration, crust
formation, synthesis of UV-absorbing substances, and enzymatic and non-enzymatic quenching of ROS.
Whether or not genetic adaptation to changes in the abiotic factors plays a role in mitigating stress and
damage has not been determined. This assessment addresses how our knowledge of the interactive effects
of UV radiation and climate change factors on aquatic ecosystems has advanced in the past four years.
Introduction
Interactions between climate change, ozone, and ultraviolet(UV) radiation are altering exposure to UV radiation in aquaticecosystems.1,2 Climate change is causing the average global airtemperature to rise and precipitation patterns to change, withimportant consequences for UV exposure in aquatic eco-systems. On a regional scale, changes in climate are highlyvariable in both space and time, leading to widespread floodsin wetter regions, more severe droughts in drier regions, andincreases in extreme storm events.3 Climate change is reducingannual snow and ice cover, increasing runoff and concen-trations of UV-absorbing dissolved organic matter (DOM) ininland and coastal waters, and increasing the strength of
aEmeritus from Friedrich-Alexander Universität Erlangen-Nürnberg, Dept. Biology,
Neue Str. 9, 91096 Möhrendorf, Germany. E-mail: [email protected] of Biology, Miami University, Oxford, OH 45056-1400, USAcDept. Biological and Environmental Science, University of Gothenburg,
P.O. Box 461, SE-40530 Göteborg, SwedendDépartement des Sciences Fondamentales and Centre for Northern Studies (CEN),
Université du Québec à Chicoutimi, Saguenay, Québec, CanadaeDepartment of Zoology, University of Wisconsin, Madison, 250 North Mills Street,
Madison, WI 53706, USAfState Key Laboratory of Marine Environmental Science, Xiamen University (XiangAn
Campus, ZhouLongQuan A1-211), XiangAn, Xiamen, Fujian 361102, ChinagEstación de Fotobiología Playa Unión, Casilla de Correo 15, (U9103ZAA) Rawson,
Chubut, ArgentinahCentre of Advanced Study in Botany, Banaras Hindu University, Varanasi-221005,
IndiaiCIESIN, Columbia University, 190 Turnbull Road, New Hartford, CT 06057-4139, USA
108 | Photochem. Photobiol. Sci., 2015, 14, 108–126 This journal is © The Royal Society of Chemistry and Owner Societies 2015

thermal stratification in these systems. Rising atmosphericCO2 concentration induces ocean acidification and altersseawater chemistry and consequently changes UV protectionprovided by calcified exoskeletons in many aquatic organismsas well as UV exposure levels in aquatic ecosystems.
The ecosystem services provided by marine and inlandwaters include food and drinking water for a growing humanpopulation, moderating extreme temperature and weather con-ditions, and regulating important greenhouse gas concen-trations such as that of atmospheric CO2. Freshwater is anindispensible requirement for human existence as well as forall wildlife in terrestrial ecosystems and inland waters. Aquaticecosystems generate important regional food supplies as wellas stimulate regional economies. Fisheries and aquacultureproduction have increased faster than the world’s humanpopulation over the last 50 years, constituting an importantsource of animal protein4 and feeding approximately 1 billionpeople in Asia alone,5 but these increases may not be sustain-able. In the next decade, fish production including that frominland and coastal fish farms is expected to exceed that ofother forms of protein.4 Aquatic ecosystems provide otherecosystem services including recreation and tourism, withcoral reefs alone estimated to generate US$9.6 billionannually.6 All of these ecosystem services are being influencedby changes in climate and exposure to changing levels of UVradiation.
Here we present an assessment of the advances inour knowledge over the past four years of how interactiveeffects of climate change and UV radiation are altering aquaticecosystems, and the critical ecosystem services that theyprovide.
Consequences of climate changeon snow, ice, DOM and exposure toUV radiationMelting snow and ice: aquatic productivity under high solarradiation
Over the last few decades, rising temperatures have reducedsea and freshwater ice and snow cover with important con-sequences for underwater exposure to UV radiation. The globalocean temperature has increased by about 1 °C over the last112 years.7 However, the temperature was almost 2 °C abovethe average from 1951–1980 in the Arctic8 and the warming ofthe water along the Antarctic Peninsula has been five timesfaster than the global average over the past 50 years. One ofthe reasons for the large temperature increase and drop in icevolume at the poles is an effective feedback mechanism. Iceand snow reflect most solar radiation back into space. In con-trast, water and soil absorb most of this radiation, whichresults in a substantial warming and increased penetration ofUV radiation into the ocean water. The higher water tempera-tures have reduced the Arctic ice cover by 49% during thesummer compared to the average during the years between
1979 and 2000.9 The total floating ice volume dropped byabout 75% during the same time period. Melting of the ArcticOcean ice now typically starts in April, 50 days earlier thanbefore the warming,10 and freezing starts in October, about1 month later than in the past. In recent years Arcticozone concentrations have decreased, but it is not clear ifthis trend will continue. It is the first time the O3-depletedarea is as large as that in the Antarctic.11 Higher watertemperatures resulting in a thinner mixing layer and longergrowing season together with increased O3 depletion allhave the potential to increase the exposure to UV radiation ofaquatic organisms that live in the upper layers of the watercolumn.
Of major concern is how climate and UV radiation will alterphytoplankton biomass in the open oceans. Oceanic phyto-plankton biomass is important in explaining variations in UVtransparency and constitutes a large sink for atmospheric CO2
by taking up a comparable amount of CO2 to all terrestrial eco-systems. Satellite imaging of chlorophyll data shows that phy-toplankton concentrations are much higher at polar latitudesthan at mid or equatorial regions:12 e.g. chlorophyll a concen-trations can exceed 20 mg m−3 in the Southern Ocean. Meltingice and snow affect phytoplankton, but local weather andmixing dynamics in the water column contribute differently tothe rate and direction of change and are influenced by localweather and mixing dynamics. Judging from 30 years of fieldstudies and satellite chlorophyll fluorescence data, cumulateddensities of phytoplankton have decreased by 12% along theWest side of the Antarctic Peninsula (Bellingshausen Sea),which has been attributed to increased solar UV-B radiation(280–315 nm) and rapid regional climate change.13 In thenorth of the Antarctic Peninsula, however, a lower photo-synthetic biomass production is attributed to denser cloudcover and the resulting decreased PAR (photosynthetic activeradiation, 400–700 nm). In contrast, further south there is lessmixing, fewer clouds and consequently lower phytoplanktonproductivity.13
Increasing cloudiness has been found to limit phyto-plankton productivity in the Arctic in open water.14 However,recent research suggests that thinning of the ice is increasingthe overall primary productivity and algal biomass in theArctic. Before recent Arctic warming, the ice cover was about3 m thick and accumulated over several years, and preventedmost light from penetrating into the water below and limitedphytoplankton production. Currently the summer ice layer isonly about 1 m thick. Pools from meltwater form on thesurface, which function as “skylights”. Reduced snowfallfurther enhances the light availability so that the penetratingsolar radiation amounts to about 50% of that incident on thesurface.15 This increased light availability fosters a largegrowth of ice algae and phytoplankton, which is further sup-ported by nutrients upwelling from below. In 2012 a NASAresearch cruise (ICESCAPE) to the Chukchi Sea off the coastof Alaska reported an unprecedented huge planktonbloom under the Arctic ice extending down to 50 m,16 “asdramatic and unexpected as finding a rainforest in the
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 108–126 | 109

middle of a desert”.17 This high chlorophyll concentrationunder the sea ice was not known before and could notbe detected by satellite-based remote sensing. Consequentlythe phytoplankton concentration was among the highest everrecorded extending down to 50 m. Similarly, it was found thatthese massive blooms occur all over the Arctic Ocean. In theopen waters of the Arctic Ocean satellite data have shown a20% increase in the chlorophyll content between 1998 and2009.18
Very limited information is available on the effects of chan-ging PAR and exposure to UV radiation on the structure of thefood web and total system productivity. In contrast to the dis-covery of high phytoplankton concentrations under the ice,predictions for the future posit a gradual loss of marine icealgae through loss of sea ice, causing a cascade through thehigher trophic levels of the food web. Additionally, meltwaterfrom sea ice and glaciers reduces the salinity, which negativelyaffects primary producers and the upper levels of the foodweb.19
Melting sea ice contains about four times more nitrogenthan bulk water,20 while increasing PAR results in an increasein the carbon to phosphorous ratio in plankton. Thereforereduced sea ice and increased PAR likely mean that phyto-plankton food quality is reduced for herbivorous grazers.Changing ice phenology and light and nutrient availabilitymay also affect species composition. Faster melting of sea iceshifts plankton species toward smaller cell types21 with abetter capacity to absorb solar radiation and take up nutrients,which affects the subsequent food web including fish andmammals.
During a 2010 cruise northwest of Svalbard even tropicalRadiolaria were found in Arctic waters.22,23 Out of the 145 taxaidentified during the cruise, 98 had come from areas muchfarther south and tropical species were reproducing intheir new habitat. Due to the decreased ice cover phyto-plankton productivity has extended further north attractingmore fish. For example, in the past, capelin – an importantprey for Atlantic cod – had a maximal distribution up to 75° N,but capelin were found up to 78° N in 2012 with cod followingthem.24
Increasing dissolved organic matter and exposure to UVradiation
The increased exposure to PAR and UV radiation caused by thesmaller and thinner ice and O3 depletion is partially offset incoastal and inland ecosystems by higher runoff from terrestrialdissolved organic matter (DOM), which decreases water trans-parency. Increases in global temperature and precipitation (insome areas) are accelerating the release of DOM into lakes,rivers, and coastal oceans.25,26 Strong inshore–offshore gradi-ents in DOM are common to distances of 20 km or more fromthe shore in large lakes,27 and tens to hundreds of kilometresin the Arctic Ocean (Fig. 1). Remote sensing of this DOM28 pro-vides mechanistic insights into how DOM is changing UV irra-diance in coastal and inland waters.
The reasons for the increases in DOM in the Arctic appearto be related primarily to a loss of permafrost. In 2012 amajority of North American and Russian regions reported a6–10% increase in the depth of soil that thaws annually rela-tive to the average from previous decades.29 The amount oforganic carbon stored in permafrost soils is more than twicethat in the atmosphere. Thus, when permafrost thaws,large quantities of DOM stored in the soils can be transportedto aquatic ecosystems and outgassed as CO2 and/or CH4
after being acted upon by UV radiation and microbes. ThisDOM is highly photoreactive so exposure to solar UVradiation and visible light accelerate its breakdown andrelease it to the atmosphere as CO2.
30 When permafrost thaws,as much as one third of the organic residues can be convertedto CO2 within two weeks.31 The increases in greenhousegases in turn increase warming and further thawing ofpermafrost with important consequences for the global radia-tive balance.
DOM concentrations have more than doubled in manytemperate inland waters in recent decades, and climatechange has been suggested to play an important role in theseincreases,32 and is expected to lead to a 65% increase in DOM
Fig. 1 Image of Earth over the North Pole showing the extent ofinfluence of terrestrial DOM inputs from rivers using an optical metricbased on UV absorption, called the spectral slope (the slope of thelinear relationship between the natural log of the absorption coefficientand wavelength in the 275–295 nm spectral range, nm−1). Although thesatellite cannot detect these shorter wavelengths, algorithms have beendeveloped that lead to an accurate relationship between spectral slopein this range and thus DOM source. Browner colours around the peri-phery of the Arctic Sea show greater inputs of terrestrial DOM, which isparticularly pronounced above Eurasia (from ref. 28).
Perspective Photochemical & Photobiological Sciences
110 | Photochem. Photobiol. Sci., 2015, 14, 108–126 This journal is © The Royal Society of Chemistry and Owner Societies 2015

in boreal waters in the future.25 Increases in precipitationreduce the time that water spends in lakes, which reduces thedegradation that occurs when DOM is exposed to UV radi-ation.33 The frequency of extreme precipitation events such ashurricanes and summer storms has increased in recent yearsand dramatically increases the export of UV-absorbing terres-trial DOM to aquatic ecosystems. Up to 63% of the annualDOM may come from the top 10% of precipitation events.34
A single hurricane can contribute up to 19% of the annualDOM input to receiving waters35 and increases in DOM associ-ated with a single storm can decrease UV transparency of lakewater by a factor of three.36 Lakes can act as buffers of the sea-sonal variability in DOM input to streams and rivers, thusmoderating DOM increases in downstream regions duringhigh flow periods, and increasing DOM concentrations duringbase flow conditions.37 This buffering of extremes does notseem to influence total annual outflow of DOM from thewatershed.38
At the same time DOM inputs contribute to the role ofcoastal and inland waters in the global carbon cycle. Inlandwaters are net sources of CO2 to the atmosphere, ventingcarbon fixed by terrestrial land plants that have subsequentlydied and decomposed.39 Metabolism and UV-dependent photo-lysis of terrestrial DOM may have important consequenceson greenhouse gas emissions from lakes and for globalclimate. Inland waters emit large amounts of CH4, a green-house gas that is more than 20 times as potent as CO2. Thequantity of methane emitted from the world’s inland waters isestimated to be equivalent to 25% of the global terrestrialcarbon sink40 making inland waters important players in theclimate–UV interactions, despite comprising only a fraction(1%) of the total water on Earth.
In addition to the concentration of DOM, the radiation itabsorbs influences exposure to UV radiation of inland andcoastal waters. Changes in iron (Fe) concentrations, pH, andland-use patterns modify DOM optical characteristics, degra-dation, and absorption of UV radiation. While the precisereasons for the Fe increase are still not completely understood,there is evidence that waters with high concentrations ofsoluble Fe are feeding into the surface waters41 and contribut-ing to higher UV absorbance of DOM. Photodegradation ofDOM is higher at low pH and high Fe concentrations.42 Peat-lands have higher DOM export than do agricultural watershedsand some agricultural landscapes may export more DOM thanforested ecosystems.43 The susceptibility of DOM to degra-dation by UV radiation and visible light (photoreactivity) andmicrobial decomposition (bioreactivity) varies with the sourceof the DOM. The DOM from agricultural watersheds is lessphotoreactive than is DOM from forested lands, but these twotypes of DOM may be similar in their bioreactivity.44 Whenexposed to artificial UV lamps in the lab, DOM in water col-lected from the Chesapeake Bay during base-flow conditions ismore photoreactive than that collected during snowmelt intributaries, but land-use (urban vs. agriculture vs. forested)made little difference.45 These data collectively suggest thatland-use patterns may alter not only the amount of DOM in
aquatic ecosystems, but also its quality, UV-absorptivity, andsubsequent breakdown rates by UV radiation and microbialdecomposition to CO2 and CH4.
Apart from reducing the underwater UV radiation andvisible light zone, increases in terrestrial DOM alter aquaticfood webs via changes in ratios between different basal carbonand nutrient sources. In situ mesocosm studies have demon-strated that increases in nutrient-poor DOM inputs to Arcticlakes will decrease primary productivity and increase hetero-trophy (uptake of organic material, in contrast to autotrophy,light driven photosynthesis) within the lake.46 In contrast,addition of DOM with a higher nutrient content to a nutrient-poor alpine lake can stimulate autotrophy more than hetero-trophy.47 Increasing DOM also traps heat closer to the surfaceof aquatic ecosystems, increasing surface temperatures,decreasing the depth of the surface mixed layer and decreasingtemperatures in deeper waters. These patterns collectively leadto stronger thermal stratification.48 Because mixing of thewater column is reduced, the UV exposure of surface-dwellingorganisms may increase or decrease depending on the relativechanges in mixing depth versus UV transparency. Furtherevidence shows that exposure of DOM to solar radiation canlead to the formation of ROS.49,50 Increases in DOM may alteraquatic community structure by altering the temperature ofinland and coastal waters, decrease exposure to UV radiation,and ameliorate effects of toxic metals and organic pollutantson fish and other aquatic organisms.51
Collectively these data indicate that climate change-inducedinputs of DOM cause severe change in UV transparency andfunctioning of inland and coastal waters. While DOM providesa refuge from damaging UV radiation for many ecologicallyand economically important aquatic organisms, it also has thepotential to increase the survival of pathogens.52 Higher con-centrations of DOM reduce the effectiveness of natural UVradiation on disinfection of drinking water supplies as well asincrease its cost and potential for production of carcinogenicdisinfection byproducts.53 Understanding the role of interac-tive effects of DOM, ROS concentrations, UV radiation andclimate change in aquatic ecosystems will thus be importantfor sustaining structure and function of the aquatic ecosystem,for example, through fisheries production and the potential touse the water as a drinking water resource.
Thermal stratification and exposureto UV radiation
Many aquatic organisms, such as zoo- and phytoplankton, arerestricted to the upper mixed layer (UML), the lower boundaryof which is the thermocline. Temperate latitudes are character-ised by seasonal changes in temperature and irradiance, whichare reflected in seasonal cycles of abundance and speciescomposition.54,55
Tropical waters typically exhibit stable thermal stratifica-tion.56 In contrast to polar waters, where the UML can exceed100 m, in tropical waters it is usually limited to the upper
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 108–126 | 111

10–35 m. Across latitudes, nutrient concentrations are higherin deeper layers, but the transport into the mixing zone islimited. As a consequence, these two clearly separated layersshelter distinctly different organisms. At higher latitudes, theinput of freshwater from melting ice increases stratificationbecause freshwater is less dense than saltwater.57
Global climate change results in ocean warming, whichmakes stratification more pronounced and decreases thedepth of the UML, causing organisms through all trophiclevels to be exposed to increased visible and UV radiation.56,58
In addition, it further limits the transport of nutrients fromdeeper waters because the lower boundary is more stable.59
Changes in wind speeds are also altering the depth of theUML in many water bodies. Higher temperatures mitigate theinhibitory effects of UV-B radiation by enhancing enzyme-mediated photo-repair as well as photosynthetic carbon fix-ation and quantum yield.60,61 The molecular mechanism ofthis enhancement is based on a significantly higher geneexpression and activity at 25 °C compared to 20 °C as well asaugmented enzyme-driven repair. The mitigating effects ofelevated temperatures can reduce the UV stress as has beenshown in the South China Sea where the photosyntheticcarbon fixation was less inhibited by UV-B radiation in thesummer than in the winter.54,62 The respiration index (log ofoxygen to carbon dioxide pressure), which may increase withocean acidification and changes in multiple climate changestressors, could affect photosynthetic production.63 In con-trast, higher temperatures can impair the cell cycle resulting inlower growth rates.64
Vertical mixing in the water column largely reduces theUV-induced inhibition of photosynthesis dependent on themixing frequency and depth, since phytoplankton are con-stantly moved from the surface to the thermocline and back.Being at the bottom of the UML allows organisms to repairdamage that they encounter at the surface, e.g. phytoplanktoncommunities in a coral reef ecosystem where an increasedmixing rate and depth results in less UV-B-induced reductionof photosynthetic carbon incorporation.65 UV-A radiation(315–400 nm) can have positive effects on the growth of largerphytoplankton cells under mixing conditions, since this radi-ation is used by the enzyme photolyase to split UV-B-inducedcyclobutane pyrimidine dimers (CPDs). UV-A radiation alsocontributes to harvesting of photosynthetic energy.62,66
When stratification becomes more pronounced and themixing layer shallower, hypoxic (low oxygen) areas in inlandand coastal waters expand.67 Harmful algal blooms (dinoflagel-lates and cyanobacteria) can increase in intensity and fre-quency in both freshwater and marine habitats due toincreasing nutrient availability from terrestrial runoff, risingtemperatures and increased stratification.68 These organismsare not very sensitive to solar UV-B radiation.
In summary, the increased water temperature due to globalclimate change reduces the depth of the UML and the organ-isms dwelling in this layer are exposed to higher UV radiation(Fig. 2). Damage from UV-B radiation encountered at thesurface is mitigated by repair processes, which are activated
when the organisms are passively transported to the lowerboundary of the UML. Higher temperatures favour enzyme-mediated repair of damage by UV radiation.
Ocean acidification and exposure toUV radiation
The pH of seawater is in the range of 7.5 to 8.4 and isrelatively stable due to its buffering capacity. However, increas-ing atmospheric CO2 concentrations have lowered this valueby about 0.1 units, which corresponds to an increase in the H+
concentration by 30%.69 Assuming increasing CO2 emissions(IPCC A1F1 scenario), an atmospheric concentration of800–1000 ppmv is predicted by 2100, which will correspond toa pH reduction by 0.3–0.4 in the ocean, an increase in H+ ionsin surface waters by 100–150%.56 Ocean acidification inconjunction with UV-B radiation affects enzymatic and otherbiochemical processes in several aquatic organisms, suchas phytoplankton, macroalgae and many animals, such asmollusks and corals.70–72 Phytoplankton may serve as a partialremedy to the problem, since they sequester CO2 throughphotosynthetic carbon fixation.21 While many higher plantsbenefit from increased atmospheric CO2 concentration, thisdoes not support higher growth rates in phytoplankton.56,73
The red tide microalga, Phaeocystis, showed a much lowergrowth rate under elevated UV-B which was more pronounced
Fig. 2 Combined effects of anthropogenic changes in the environ-mental conditions in marine ecosystems. Increasing atmospheric andwater CO2 concentrations reduce the calcifying abilities of many organ-isms. Increasing water temperatures and incoming solar radiationdecrease the depth of the mixing layer (exposing organisms to higherirradiances) and increase the temperature difference between surfaceand deeper layers. This temperature difference limits the exchange ofmaterials such as nutrients between layers (modified from ref. 56).
Perspective Photochemical & Photobiological Sciences
112 | Photochem. Photobiol. Sci., 2015, 14, 108–126 This journal is © The Royal Society of Chemistry and Owner Societies 2015

under elevated CO2, indicating that increasing ocean acidifica-tion and UV-B act synergistically to reduce photochemicalperformance.74
Zooplankton seem to be little affected by water acidifica-tion, although acclimation leads to higher respiration andincreased grazing rates.75 Their shells are mainly composed ofchitin, which is not affected by acidification. In contrast, thoseorganisms with outer skeletons of calcium carbonate areaffected. Increasing acidity affects calcification in phyto-plankton, calcified macroalgae76 and animals with exo- orendoskeletons such as corals, depriving these organisms ofsome defense against solar UV-B radiation.77 UV-B radiationstrongly impairs the photosynthetic apparatus in cocco-lithophorides, while UV-A radiation inhibits calcification.78
The calcified outer scales form a protective exoskeleton.79
Cells grown at high calcium concentrations are more resistantto UV radiation than under limited calcium concentrations.79
In polar regions, dissolution of CO2 from the air intoseawater differs from that in low latitude areas. Low seasurface temperature means that more CO2 is dissolved thanin low latitude waters. Changes in carbonate chemistry ofseawater in the high-latitude oceans are already negativelyaffecting some species. Consequently, it is projected thatwithin decades, large parts of the polar oceans will becomecorrosive to the shells of calcareous marine organisms.80 Theshells of pteropods, small marine snails (sea butterflies),that are key species in the food web are already dissolvingin parts of the Southern Ocean surrounding Antarctica.81
Ocean acidification has effects not only on biological processes
but also on the uptake and availability of iron82 andammonium.55
Degrees of sensitivity of aquaticorganismsMechanisms of UV radiation damage
In the upper photic zone, aquatic organisms are exposed tosolar UV radiation. Although the UV-B irradiance amounts toonly a few percent of the total solar radiation, this wavelengthband can be hazardous since it affects biomolecules and cellu-lar structures (Fig. 3) and may block enzymatic reactions andinterfere with physiological responses such as motility andorientation.69 UV-B radiation can either directly alter bio-molecules or induce the formation of reactive oxygen species(ROS) inside the cell, such as singlet oxygen (1O2).
83,84 For-mation of ROS is augmented by increasing temperatures.85
Photosynthesis is specifically prone to damage by solarUV-B radiation. In addition to other targets, radiation damagesthe D1 protein in the electron transport chain of photosystemII (PS II), and is subsequently removed and during repairreplaced with a newly synthesized protein.86 Higher watertemperatures enhance the repair process, while limited nutri-ent supply impairs the repair mechanisms.87 An unexpectedfinding was that UV-B radiation damages phytoplankton moreby impairing repair mechanisms than by directly damagingthe protein.88 In addition, solar UV-B radiation affects theaccessory pigments that funnel solar energy to the reaction
Fig. 3 Effects of solar UV radiation on biomolecules, cellular components and physiological responses as well as mitigating strategies and repairmechanisms (for details see text).
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 108–126 | 113

centres. The blue pigments, phycobilins, in cyanobacteria andred algae are especially sensitive to damage.89
Another main target of solar UV-B radiation is the DNA inboth prokaryotic (bacteria) and eukaryotic (organisms with acell nucleus) organisms. In addition to single- and doublestrand breaks and the formation of 6-4 photoproducts (andtheir Dewar valence isomers), the most frequent lesioninduced by UV-B radiation is the induction of CPDs.90 Repairmechanisms for DNA damage include several mechanisms(such as excision repair, mismatch repair and SOS response),but above all the photoactivated CPD photolyase is engaged tobreak the dimers using the energy of UV-A radiation or bluelight photons.90
Mechanisms to avoid or moderate UV-B radiation-induceddamage include vertical migration to move out of zones ofexcessive radiation found in zooplankton and phytoplanktonor by mat formation.91 With the exception of very small cells(picoplankton), many aquatic organisms produce UV-absorb-ing compounds to prevent solar UV-B radiation from damagingthe central regions of the cell such as the nucleus. Cyano-bacteria synthesize scytonemin to diminish the impact of UVradiation.92 In addition, phytoplankton and macroalgaeproduce several mycosporine-like amino acids (MAAs).93 UV-Bradiation-induced reactive oxygen species are removed by enzy-matic reactions and non-enzymatic quenchers including caro-tenoids.83,94 Animals such as zooplankton are not capable ofsynthesizing MAAs, but may take up these substances withtheir food and use them for protection from solar UVradiation.
Several factors make accurate assessment and measurementof the effects of UV radiation on organisms in natural eco-systems very challenging. These factors include the wide varia-tion among organisms in the mechanisms of defense againstdamage by UV radiation, the need to allow organisms ade-quate time to adapt to sudden highly elevated UV radiationlevels, and assuring accurate measurements of UV radiation.Laboratory experiments with artificial sources of UV radiationoften use excessive short wavelength UV-B irradiance levelsthat are more damaging and have little ecological significanceto natural solar radiation due to inappropriate balance in thespectral composition (see Bornman et al.95).
UV radiation exposure levels in aquatic ecosystems exhibitstrong gradients over time (daily to annual), depth, and dis-tance from the shore. Different natural populations may varyin their sensitivity to UV radiation over time and may acclimateto the radiation, resulting in some adaptation.
Parasites and pathogens
UV radiation plays an important role in the ecology of manyinfectious diseases of aquatic organisms, particularly whenthere is a pronounced difference in the UV radiation toleranceof the host and pathogen or parasite. For example, Metchnikowiais a fungal parasite that is lethal to the important fresh-water zooplankton grazer, Daphnia. Relative to its host, theparasite is extremely sensitive to UV radiation and longer wave-length sunlight. Thus in more UV radiation transparent water
bodies, outbreaks of this parasite are suppressed and delayeduntil later in the autumn after incident solar UV radiation hassubsided.52 Natural solar radiation is also highly effective atreducing viral infections in some aquatic organisms includingviruses of fish and harmful algal bloom (HAB) species. Someexperiments that have manipulated natural sunlight reveal amillion-fold decrease in the infectious hematopoietic necrosisvirus (HNV) in Atlantic salmon in treatments exposing to sun-light versus dark controls for just 3 hours.96 Viruses may beresponsible for more than half of the mortality of aquatic cya-nobacteria, which has led to interest in using cyanophages tocontrol HABs of toxic cyanobacteria.97 The extreme sensitivityof many viruses to UV radiation damage suggests that knowingmore about the changing underwater UV radiation environ-ment may lead to new insights into the potential role ofviruses in controlling HAB of cyanobacteria.97
Trematodes are some of the most common parasites inintertidal systems. Their larvae are free-living for short periodsof time between hosts, which range from snails to other invert-ebrates and birds. During this short, free-living period in veryshallow aquatic environments the larvae may be exposed tohigh levels of solar UV radiation. Recent UV radiation exposureexperiments in the laboratory showed that UV radiation causedDNA damage and oxidative stress in the larvae and showed noevidence of photoprotective MAAs or photoenzymatic repair.98
Similar experiments also demonstrated negative effects ofUV-B as well as UV-A radiation exposure on the survival oflarvae. In addition, susceptibility to infection of the amphipodsecondary host increased when the host was exposed to UVradiation.99 Further experiments on this trematode parasitesystem revealed significant interaction effects between UV radi-ation and temperature, with greater UV radiation effects at20 °C vs. 30 °C.100
Parasites may also play some role in altering the exposureto UV radiation of infected fish hosts. Three-spined sticklebackundergo daily vertical migration and are generally deeper inthe water column during the day than during the night.However, individuals captured in the surface waters during theday have a higher parasite load than those captured at night,which increases the potential for this parasite to be trans-mitted to its definitive host – fish-eating birds.101 Increasingevidence is accumulating for the direct and indirect effects ofexposure to UV radiation on several marine fish species. Mela-nosis and melanoma skin cancer rates of up to 15% have beenreported in coral trout in the Great Barrier Reef of Australia(Fig. 4).102 The role of UV radiation in the induction of thishigh prevalence is unknown. In shallow, UV-transparentaquatic ecosystems, such as coral reef flats, photobleachingmay lead to destruction of corals and may result in furthernegative effects when UV radiation and interactions withmultiple stressors occurs.103
Bacteria and viruses
Heterotrophic bacteria and viruses are more affected by UVradiation than phytoplankton since they do not synthesize UV-absorbing pigments (but they can repair UV induced damage).
Perspective Photochemical & Photobiological Sciences
114 | Photochem. Photobiol. Sci., 2015, 14, 108–126 This journal is © The Royal Society of Chemistry and Owner Societies 2015

Their small dimensions would require extremely large concen-trations of these substances to effectively protect them fromexcessive short-wavelength radiation.104 These organismsrespond to UV radiation with adaptive processes such asincreased frequency of division, but exposure to UV radiationresults in significant changes in species composition due to avarying UV sensitivity of different bacteria taxa.105 Acclimationto long-term UV exposure has also been found in natural sub-Antarctic phytoplankton communities.
Inland and coastal waters are rich in bacterioplankton,which feed on the high concentrations of humic substances.106
Bacteria are under pressure from UV-B and UV-A radiationcomponents of sunlight, but recover when the solar radiationdecreases during the daily cycle. The highest bacterioplanktonactivity has been found between 5 and 10 m, below the depthsof high solar UV-B radiation exposure. Inhibitory effects of UVradiation on bacterial growth are mitigated by water mixing.107
Different strains have different sensitivity to higher tempera-tures and UV radiation, leading to selection of more resistantstrains and species. Whether or not genetic adaptation mayalso mitigate the effects of altered stress parameters needs tobe examined.108
Phytoplankton
The amount of solar UV and visible radiation determines thespecies composition of phytoplankton blooms. Large-celleddiatoms are less affected by solar UV-B radiation and canutilise high light irradiances.109 In contrast, small-celled
phytoplankton such as the ecologically important cyano-bacteria and small eukaryotes experience more UV-B-induceddamage than larger cells.110
In oligotrophic waters (waters with low nutrient contents)solar radiation induces a higher kill rate in marine picoplank-ton than in larger cyanobacteria and eukaryotic phytoplank-ton.111 The main driver for mortality is UV-B radiation sincefiltering out this short wavelength radiation significantlyenhances survival. Surface samples are more resistant to solarUV-B radiation than samples collected at depth. Exposingphytoplankton samples experimentally at a fixed depthinhibits photosynthesis more than under natural conditions,where the organisms are moved within the mixing layer.112
The mitigation of the UV-induced inhibition of photosyntheticcarbon fixation depends on the mixing rate and depths asshown in tropical coral reef phytoplankton assemblages.65
Phytoplankton of the same taxonomic groups can have sig-nificant different sensitivities toward solar UV-B radiationdepending on their geographical origin, such as tropical, tem-perate and Antarctic habitats. In contrast to UV-B radiation,even the highest applied doses of UV-A radiation did not causegrowth inhibition. After periods of excessive UV radiationsome phytoplankton species, such as the marine diatomPhaeodactylum tricornutum, show a higher growth rate, whichpartially compensates for prior UV-induced growth reductions.Photosynthesis in phytoplankton is damaged by UV-B radi-ation mainly at the D1 protein but with higher temperaturesincreasing the repair rate.87
Primary production in freshwater systems is, in contrast tothat in marine systems, often limited by phosphorus. Becauseof this, the effects of UV radiation on phosphorus metabolismcan be important in freshwater aquatic ecosystems. Forexample, in a high mountain mesocosm experiment, hetero-trophic microorganism biomass (bacteria and flagellates)increased when phosphorus addition was reduced by 80%if UV radiation was excluded.113 Sereda et al. investigatedhow ambient UV radiation affects the phosphorus metabolismof plankton communities from 18 lakes in Ontario andSaskatchewan.114 The turn-over time for phosphorus and thesteady state phosphate concentration increased when theorganisms were exposed to UV radiation.
The concentration of phytoplankton strongly depends onthe pressure by grazers. For example, the seasonal abundanceand feeding patterns of copepods in a pelagic food web in theWhite Sea showed that up to 85% of the daily phytoplanktonbiomass was consumed by calanoid copepods.115 Exposure toUV radiation affects the quality of phytoplankton in terms offood for zooplanktonic grazers.116 Therefore the level of UV-Bradiation is an important modulator of the phytoplanktonstanding crop.
Mitigation of UV-induced damage by UV-absorbing substances
Phytoplankton use a number of effective repair mechanismsas well as UV-absorbing substances (mostly MAAs) to mitigateUV-B-induced damage of DNA and the photosynthetic appar-atus. Samples from phytoplankton blooms under the ozone
Fig. 4 Cancer lesions on coral trout from the Great Barrier Reefobserved in up to 15% the natural population and thought to be due toexposure to solar UV radiation. Shown are individuals with only partial(a) and almost full body (b) lesions as well as close-up photographstaken under a microscope of healthy skin (c) and skin lesions (d). Scalebars are 20 µm (from ref. 102).
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 108–126 | 115

‘hole’ counterintuitively show significantly less inhibition ofthe photosynthetic quantum yield by UV-B radiation thanthose from outside the ozone-depleted areas, indicating theprotective role of their higher MAA concentration.117,118
Another reason for these findings may be that large-size cellsoccur inside the blooms while outside the blooms smallercells prevail, which, due to their small size, cannot take advan-tage of the MAA protection. Ryan et al. experimentallymeasured the effects of UV-B radiation on sea-ice algae inAntarctica119 and concluded that brine channel communitieswere better protected from UV-B radiation. They speculate thatthe high tolerance to UV-B radiation in the brine communitiescould be due to production of MAAs.
Concentrations of UV-absorbing compounds such as MAAsincrease with higher exposure to solar radiation in phyto-plankton and macroalgae and decrease under experimentalconditions without UV.120 In parallel, inhibition by UV radi-ation of growth and photosynthesis is mitigated with increasedcontent of MAAs. Higher levels of nitrate result in higher con-tents of MAAs, while phosphate limitation did not affect theMAA content.121 In addition to their UV-absorbing properties,MAAs serve as antioxidants scavenging ROS.122
MAAs are produced in algal cells and protect against UV-Bradiation. The production depends on species, degree ofimpairment, and locality.123 Large differences have beenfound in MAA production, which correlated with differences inspecies composition and sensitivity to UV-B radiation. Phyto-plankton in inland and coastal waters are generally less toler-ant to UV radiation than open ocean assemblages. This isprobably because they need less protection and therefore havedeveloped lower MAA concentrations as a consequence of thelower transparency of the water.124 UV-B radiation can impairgrowth and development, morphology, photosynthesis andnutrient uptake in coastal phytoplankton species.125
In the tropics UV-absorbing MAAs within phytoplanktoncells show the same concentration year round, while in tem-perate waters MAA concentrations are lower in winter than insummer in surface waters <50 m.93 MAA expression is linearlyrelated with the UV irradiance at the surface. In the tropics,phytoplankton is under considerable UV-B radiation stress onsunny days. On cloudy days microplankton (>20 µm) use UV-Aradiation as an energy source for photosynthesis, while pico-and nanoplankton are impaired.126 Cloud patterns anddensity affect the level of UV radiation, but this has rarely beenstudied in detail. Along a 13 000 km meridional transect (from52° N to 45° S) the highest MAA concentrations were found inthe south (>40° S) and in the north subtropical region.93
Interacting stress factors
Satellite monitoring of the oceans shows that phytoplanktonconcentrations have been declining at about 1% per year overthe past 50 years.127 Laufkoetter et al.128 calculated a differentnumber for the decrease (6.5% during the period 1960–2006),simulating phytoplankton net primary production on a globalscale with large spatial resolution. However, there are largeareas of uncertainty: even though the external factors of temp-
erature, pH, CO2 supply, nutrients, PAR and UV irradiancesand mixing depths are known to be primary variables drivingphotosynthesis and production, their interactions have notbeen thoroughly investigated.56 This interacting web can onlybe disentangled by multifactorial analysis and model-ling.129,130 In order to reveal the effects in nature with its fastchanging temperature, solar radiation and nutrient avail-ability, large scale, long-term studies are needed in the rele-vant ecosystems.56 This is difficult and time-consumingbecause of the vast areas to be covered, the diversity of organ-isms and ecosystems and the low concentrations of cells,especially in open ocean waters. The effects of the many poss-ible feedback mechanisms on marine primary producers arelargely unknown. For example, do higher temperatures of theoceans result in denser cloud cover? This could result in lowerexposure of the phytoplankton to solar UV radiation and PAR,favouring taxa that are more sensitive to UV radiation.131 Feed-back systems have the potential to change the species compo-sition of future assemblages of primary producers with largeconsequences for inland and marine food webs.
Feedback between UV radiation and inland and coastalecosystem organisms also has the potential to moderateclimate. For example, phytoplankton and macroalgae producedimethylsulfoniopropionate (DMSP),132 which acts as anosmolytic substance or as an antioxidant. DMSP is excretedand broken down to dimethylsulfide (DMS), which enters theatmosphere and down-regulates global warming and reducesthe UV-B radiation reaching Earth. In temperate shelf areas,the key phytoplankton species, UV irradiance and nutrientconcentrations determine the seasonal cycle of DMS. Oceanacidification reduces DMS accumulation, enhancing globalwarming, but the role of UV radiation on the production ofDMS has not been investigated.133
Increasing environmental pollution, such as crude oilspills, affects algae and bacteria, especially in the Arcticshallow-water marine habitats. Pyrene is a component of crudeoil and accumulates in the sediment where it exerts a synergis-tically negative effect with increased solar UV-B radiation.134
Pyrene is taken up by and concentrated in the cells where itreduces growth rate. In the Greenland Current and ArcticOcean persistent organic pollutants (POP) accumulate inphytoplankton as documented during the ATOS-ARCTIC cruiseon board the R/V Hespérides.135
Exposure to solar UV radiation alters the fatty acid concen-tration in several phytoplankton groups136 and affects enzymeactivity and nitrogen assimilation in both eukaryotic and pro-karyotic phytoplankton. However, the sensitivity of phyto-plankton species to UV-B radiation is modified by the lighthistory indicating some short-term acclimation to UV-B radi-ation stress.137 Comparable levels of solar UV-B radiationcaused the same degree of growth inhibition in phytoplanktonin coastal and offshore surface waters of the South China Seaunder clear skies by about 28%. In contrast, inhibition byUV-A radiation was higher in open water samples (13%) thanin coastal water samples (4%).66 Due to terrestrial runoff,coastal ecosystems often have higher nutrient concentrations
Perspective Photochemical & Photobiological Sciences
116 | Photochem. Photobiol. Sci., 2015, 14, 108–126 This journal is © The Royal Society of Chemistry and Owner Societies 2015

than open ocean systems. In some taxa, such as dinoflagel-lates, higher nutrient supply (N and P) augments the quantumyield resulting in a different species composition than in openoceanic waters.138 For example, dinoflagellates may out-compete diatoms when rivers deliver larger nutrient loads tocoastal waters.139 High nutrient concentrations in coastalwaters often induce blooms of toxic phytoplankton (e.g. dino-flagellates), which enter the food chain and can be poisonousto humans. However, solar UV-B radiation does not seem toimpair these red tide phytoplankton species.68
The toxicity of many pollutants increases with exposure toUV radiation. Inland and coastal marine environments areunder stress from these interactions between UV radiation andpollutants such as polycyclic aromatic hydrocarbons (PAHs),which form from combustion engines, and water-soluble frac-tions of heavy oil that affect plankton communities. Toxicity ofPAHs is enhanced by solar UV radiation as shown with naturalphytoplankton communities from the Mediterranean Sea, andAtlantic, Arctic and Southern Oceans.140 Natural phyto-plankton communities pre-stressed with UV-B radiation aremore susceptible to pollutants, such as atrazine, tributyltin orcrude oil, which enter coastal waters from terrestrial drainageor maritime traffic, than those grown under UV radiation-freeconditions.
Benthic organisms
Several studies have demonstrated the response of benthic(bottom-dwelling) organisms and communities to solar UVradiation in shallow aquatic habitats. In combined laboratoryand field experiments freshwater snails were found to haveseveral mechanisms to avoid damage from UV radiationincluding behavioural avoidance, photorepair, increased shellthickness, pigmentation and body size.141,142 Juvenile benthicmarine polychaetes showed reduced growth and developmentof tentacles when fed detritus derived from diatoms pre-exposed to artificial UV-B radiation from lamps versus diatomsthat were not pre-exposed to UV-B radiation.143 Ostracods,small crustaceans that thrive in shallow benthic habitats, haveshells that block 60–80% of UV radiation, as opposed to theexoskeleton of more planktonic Daphnia that block only 35%of UV radiation.144 In a four month-long field experiment ontidal flats where the benthic community was followed whenambient UV-B or UVB + UVA radiation was excluded, the onlystructural change was a doubling of ostracod biomass underUV radiation compared to PAR only treatments.
Seaweeds are an important group of benthic organisms forcoastal ecosystems. In addition to being primary producers,seaweeds shape local environments that are important formany animals including fish larvae. Seaweeds are located infixed positions and need sunlight for photosynthesis and thuscannot escape exposure to high UV radiation. A substantialspecies-dependent variation in sensitivity to UV radiation cor-relates with vertical zonation, and smaller and juvenile sea-weeds are generally the most sensitive.145 Seaweeds haveseveral life stages of which some are motile (zoospores andgametes) and important for the expansion of the species to
new locations. These stages may also be sensitive to DNAdamage by solar UV-B radiation as they have only one genecopy.146
The seafloor also harbours benthic communities in theform of invertebrates, bacteria, fungi and microalgae. Anexperimental study examined the effects of different radiationregimes on the development of these benthic communities inSpitsbergen.147 A total of 17 algal and invertebrate taxa wereanalysed. No detrimental effects were found from UV radiation(relative to PAR only), although in some species the abundanceincreased, especially under UV-A radiation. This indicates thatat the community level the effects of exposure to UV radiationare dependent on species composition and successional stage.A recent study on the impact of multiple stressors, includingUV radiation, temperature and ocean acidification on mollus-can development148 showed that the embryos developedsignificantly better at 26 °C than at 22 °C. Mortality was signifi-cantly higher at 22 °C and pH of 7.6. UV radiation had no sig-nificant impact on the embryonic development.
Zooplankton
Changes in species composition resulting from climatechange149 may favour species that have different UV tolerances.Species that routinely experience high levels of UV radiationare better protected than those that are used to low levels ofUV radiation, as was demonstrated by the differences in theUV absorbance of carapaces of Daphnia originating from highUV alpine and low UV boreal lakes.150 In a field survey inArgentina, the relative abundance of more UV-tolerant cope-pods versus less UV-tolerant Daphnia increased with the dis-tance from the turbid input of a glacier, suggesting UVradiation as a possible regulator of zooplankton communitystructure, although a role for other factors that changed alongthe gradient cannot be ruled out.151
The response of species to UV radiation varies and isrelated to the extent of their ability to use various UV avoid-ance or protection strategies. Evidence confirms that exposureto UV radiation plays an important role in stimulating down-ward migration of zooplankton during the day in highly trans-parent waters. These observations have recently beenintegrated with past studies to develop a more comprehensivetheory of daily vertical migration (DVM).152 Some of thestrongest evidence for the importance of UV radiation in DVMcomes from alpine lakes that lack the visually feeding fish thatare often implicated in DVM. For example, in high elevationlakes in Northern Italy153 and Poland,154 crustacean zoo-plankton migrate to deeper depths during the day in spite ofthe lack of fish or other visual predators. In the Italian lakesthe abundance of crustacean zooplankton in the surfacewaters of 13 lakes during the day was found to be similar inthose lakes with and without fish, suggesting that it is not fishpredation that excludes zooplankton from the surface watersof these lakes.155 In temperate lakes of glacial origin in situexperimental manipulation of fish and UV radiation in 15 mdeep mesocosms similarly revealed that UV radiation inducedstronger downward migration than did the presence of fish.156
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 108–126 | 117

In this same study, Daphnia were found to migrate upward inthe water column during daylight following a strong stormevent that reduced water transparency to UV radiation andvisible light. In some marine systems copepods may occurdeep enough in the water column that UV radiation plays littleor no role in DVM.151 Collectively these observations suggestthat changes in water transparency due to climate change arelikely to influence the vertical distribution and abundance ofzooplankton, a critical link in both freshwater and marinefood webs.
The importance of the sublethal effects of UV radiation onboth freshwater and marine zooplankton has become increas-ingly recognized. Sublethal effects of UV radiation on marinecopepods include reduced egg quality and survival of larvae.151
In a series of laboratory experiments marine copepods grazedat higher rates on algae that had previously been exposed toelevated levels of UV radiation.157
Further advances have been made in understandingdefense mechanisms against UV radiation in freshwater andmarine zooplankton, including the trade-offs amongmultiple defenses and pressures from visual predators andother environmental factors. A recent meta-analysis shows thatcopepods from freshwater ecosystems have more carotenoidsthan marine copepods, but that the two groups have similaramounts of MAAs.151 Repair and antioxidant enzymes maysimilarly provide defense against UV radiation and visual pre-dators simultaneously. Following exposure to artificial UV radi-ation, freshwater calanoid copepods rapidly activated enzymesystems that reduce peroxidation, cell death, and damageto neurotransmitters.158 Freshwater copepods challenged bysimultaneous exposure to fish predation and potentially dama-ging UV radiation can exhibit trait compensation wherein theyincrease anti-oxidant enzymes and decrease pigmentation,thus reducing damage by both threats.159 Higher concen-trations of carotenoid photoprotective pigments have beenobserved in copepods in shallow turbid lakes with lower watertransparency, apparently due to the lack of aquatic plants andincrease in wind-driven turbulence that exposes the copepodsto surface UV radiation.160
Fish, amphibians and mammals
Recent evidence indicates that changes in the underwater UVradiation environment may play an important role in regulat-ing invasive fish in cold, clear-water lakes. Studies of twospecies of invasive warm-water fish (bluegill and largemouthbass) and one native (Lahontan redside) fish in Lake Tahoe,California-Nevada, have demonstrated strong differences intolerance to UV radiation between native and invasive species.Warm temperatures in shallow near-shore habitats are necess-ary for the invasive species to breed. High UV transparency inthese near-shore habitats prevents the warm-water invasivespecies from successfully breeding due to the low tolerance toUV radiation of their larvae. Climate change and other disturb-ances that reduce UV transparency of waters in the warmershallow shoreline habitat can open an invasion window thatpermits the invasive warm-water species to become established
and reduces native species population sizes through compe-tition and predation.161 The differences in tolerance to UVradiation between invasive and native species can be used infisheries management to exclude the warm-water invasivespecies by developing minimum UV attainment thresholdsand maintaining high UV transparency of the shoreline breed-ing habitats.162 Histological studies have demonstrated thatspecies of fish native to highly UV transparent lakes are alsomore tolerant of simultaneous exposure to solar UV radiationand pollutants such as polycyclic aromatic hydrocarbons(PAHs) than are invasive warm-water species found in charac-teristically less UV transparent lakes.163 UV radiation and PAHexposure experiments confirmed these results as well as thoseshowing that the native species have more melanin for copingwith high UV radiation levels than do the invasive species.164
Other recent studies have demonstrated both behaviouralavoidance as well as other indirect and direct damaging effectsof UV radiation on fish. Experiments that manipulated naturalUV radiation with filter foils demonstrated that the survival offreshwater yellow perch larvae is more negatively influenced byUV-A radiation and longer wavelength UV-B and UV-A radiationthan by the shorter wavelength UV-B.165 Experimental studiesmanipulated exposure of two species of salmon fry to solarUV-B radiation in outdoor rearing tanks and tagged smoltsusing acoustic transmitters to examine their growth ratesduring rearing as well as subsequent survival rates in themarine environment.166 Exposure to UV-B radiation led to adecrease in early growth of Coho salmon, but had no effectson early survival of either species in the oceans. Exposure ofEuropean sea bass larvae to even low levels of artificial UVradiation in the laboratory led to behavioural avoidance,reduced ability to osmoregulate, as well as increased mor-tality.167 Atlantic cod larvae subjected to prior exposure to arti-ficial UV lamps showed subsequent reductions in the ability toescape suction predators, but not tactile predators;168 feedingrates were also reduced compared to unexposed controls.169
While these studies suggest reductions in feeding will translateto reduced survival under natural conditions,169 this specu-lation is in contrast to a prior study with largemouth basslarvae under more natural conditions in lakes, where the pres-ence of UV radiation stimulated feeding on zooplankton.170 Ananalysis of the relationship between brown trout biomass andDOM in 168 lakes in Southern Norway revealed a unimodalrelationship with a peak in fish biomass at intermediate DOMlevels;171 UV radiation was hypothesized to play a role indecreased biomass at low DOM levels.
In a comprehensive assessment of exposure to UV-B radi-ation in amphibian breeding habitats, UV-B radiation levelswere estimated to be high enough to seriously threaten woodfrogs breeding in northern Minnesota vernal pools.172
Reductions in forest canopy cover due to timber harvest as wellas changes in water transparency or pool depth related toclimate change may further alter exposure to UV radiation inthese amphibian breeding habitats.
Marine mammals are being influenced not only by directdamage from exposure to solar UV radiation, but also
Perspective Photochemical & Photobiological Sciences
118 | Photochem. Photobiol. Sci., 2015, 14, 108–126 This journal is © The Royal Society of Chemistry and Owner Societies 2015

indirectly through pollution of coastal marine habitats byanthropogenic sunscreen chemicals (sun-tan lotions). Theycontribute to the bleaching of corals by promoting viral infec-tions and may change the sex of fish.173 Methods for identify-ing UV-induced skin damage in humans based on real timePCR and mitochondrial DNA (mtDNA) biomarkers have beenrecently modified and applied to whales.174 Individuals ofthree different whale species showed significant variations inmtDNA damage in the skin using these techniques. Differentspecies of whales vary in their strategies for coping withexposure to UV radiation. Blue whales, which have relativelypale skin, vary their melanin production seasonally as UV radi-ation levels vary with their migrations across latitudes; theinverse relationship between melanin and levels of damagefrom UV radiation suggest that melanin is an effective defenseagainst UV radiation in these cetaceans.175 Sperm whales,which spend more time in the high UV radiation environmentsat the surface of the ocean all year long, have more melano-cytes than blue whales, but similar amounts of melanin.175
The widely used human sunscreen compound, octocrylene,has recently been found in the liver of dolphins off the coastof Brazil.176 Thus it could also have effects on humans.
Gaps in knowledge
While the response of aquatic biomass producers to solarUV-B radiation and global climate change have been character-ized to some extent, the effects of interacting stress factors onnatural assemblages and ecosystems needs to be further inves-tigated. How the changes in phytoplankton species compo-sition, due to differential sensitivity of individual speciescaused by UV radiation and interaction with other environ-mental factors such as temperature, will affect the subsequentfood web including fish and mammals also needs to be quan-tified. There are limited records on the dynamics of overalleffects of UV radiation on physical, chemical, and biologicalattributes of oligotrophic biomass changes.
Ocean acidification due to increased atmospheric CO2 con-centrations alters the marine chemical environment, which inturn interferes with UV radiation-protecting calcification inmany aquatic organisms including phytoplankton, macroalgaeand animals such as molluscs and corals. Multifactorialeffects including UV-B radiation and ocean acidification ondiverse organisms as well as ecosystems should to be studiedin order to understand the impacts of future ocean climatechanges.
Aquatic organisms employ several lines of defense mecha-nisms to mitigate the damaging effects of UV radiation.A number of ecologically and economically important organ-isms need to be screened for the presence of photoprotectivecompounds and molecular mechanisms of repair.
There is limited knowledge on the cumulative effects of UVradiation–climate change interactions on the nature and typeof invasive species and its impact on native populations inaquatic ecosystems.
Acknowledgements
Participation by Prof. Donat-P Häder was supported by theBundesministerium für Umwelt Naturschutz und Reaktorsi-cherheit. Participation by Dr Kevin Rose was supported by theWisconsin Department of Natural Resources (WI DNR) and theNorth Temperate Lakes Long Term Ecological Research Site(NTL LTER). Participation by Drs Craig Williamson, andRobert Worrest was supported by the U.S. Global ChangeResearch Program. Participation by Prof. Milla Rautio was sup-ported by Canada Research Chair Program.
References
1 A. F. Bais, R. L. McKenzie, P. J. Aucamp, M. Ilyas,S. Madronich, G. Bernhard and K. Tourpali, Ozonedepletion and climate change: Impacts on UV radiation,Photochem. Photobiol. Sci., 2015, 14, DOI: 10.1039/c4pp90032d, this issue.
2 D. J. Erickson III, B. Sulzberger, R. Zepp, A. T. Austin andN. Paul, Effects of stratospheric ozone depletion, solar UVradiation, and climate change on biogeochemical cycling:Interactions and feedbacks, Photochem. Photobiol. Sci.,2015, 14, DOI: 10.1039/c4pp90036g, this issue.
3 IPCC, Summary for Policymakers: Climate change2013 – The physical science basis, Report No., 2013,pp. 1–38.
4 FAO, The state of world fisheries and aquaculture 2012, FAO,Rome, 2012.
5 UNEP, Marine and coastal ecosystems and human well-being: a synthesis report based on the findings of themillennium ecosystem assessment, UNEP Report, Nairobi,2006.
6 A. V. Parisi and M. G. Kimlin, Personal solar UV exposuremeasurements employing modified polysulphone with anextended dynamic range, Photochem. Photobiol., 2004, 79,411–415.
7 M. Fischetti, Deep heat threatens marine life, Sci. Am.,2013, 308, 92.
8 M. New, D. Liverman, H. Schroder and K. Anderson, Fourdegrees and beyond: the potential for a global tempera-ture increase of four degrees and its implications, Philos.Trans. R. Soc. London, A, 2011, 369, 6–19.
9 J. Zhang, R. Lindsay, A. Schweiger and M. Steele, Theimpact of an intense summer cyclone on 2012 Arctic seaice retreat, Geophys. Res. Lett., 2013, 40, 720–726.
10 M. Kahru, V. Brotas, B. Manzano-Sarabia andB. G. Mitchell, Are phytoplankton blooms occurringearlier in the Arctic?, Global Change Biol., 2010, 17, 1733–1739.
11 G. L. Manney, M. L. Santee, M. Rex, N. J. Livesey,M. C. Pitts, P. Veefkind, E. R. Nash, I. Wohltmann,R. Lehmann, L. Froidevaux, L. R. Poole, M. R. Schoeberl,D. P. Haffner, J. Davies, V. Dorokhov, H. Gernandt,B. Johnson, R. Kivi, E. Kyrö, N. Larsen, P. F. Levelt,
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 108–126 | 119

A. Makshtas, C. T. McElory, H. Nakajima andM. C. Parrondo, Unprecedented Arctic ozone loss in 2011,Nature, 2011, 478, 469–475.
12 W. Parry, Arctic’s Spring Phytoplankton Blooms ArriveEarlier, http://www.livescience.com/13082-arctic-plankton-blooms-ocean-climate-change.html.
13 M. Montes-Hugo, S. C. Doney, H. W. Ducklow, W. Fraser,D. Martinson, S. E. Stammerjohn and O. Schofield,Recent changes in phytoplankton communities associatedwith rapid regional climate change along the WesternAntarctic Peninsula, Science, 2009, 323, 1470–1473.
14 S. Bélanger, M. Babin and J.-E. Tremblay, Increasingcloudiness in Arctic damps the increase in phytoplanktonprimary production due to sea ice receding, Bio-geosciences, 2013, 10, 4087–4101.
15 K. E. Frey, D. K. Perovich and B. Light, The spatial distri-bution of solar radiation under a melting Arctic sea icecover, Geophys. Res. Lett., 2011, 38, L22501.
16 K. R. Arrigo, D. K. Perovich, R. S. Pickart, Z. W. Brown,G. L. van Dijken, K. E. Lowry, M. M. Mills, M. A. Palmer,W. M. Balch, F. Bahr, N. R. Bates, C. Benitez-Nelson,B. Bowler, E. Brownlee, J. K. Ehn, K. E. Frey, R. Garley,S. R. Laney, L. Lubelczyk, J. Mathis, A. Matsuoka,B. G. Mitchell, G. W. K. Moore, E. Ortega-Retuerta, S. Pal,C. M. Polashenski, R. A. Reynolds, B. Schieber,H. M. Sosik, M. Stephens and J. H. Swift, Massive phyto-plankton blooms under Arctic sea ice, Science, 2012, 336,1408.
17 M. J. Viñas, NASA discovers unprecedented blooms ofocean plant life, NASA Sci. News, 2012.
18 K. R. Arrigo and G. L. van Dijken, Secular trends in ArcticOcean net primary production, J. Geophys. Res.: Oceans,2011, 116, 160–168.
19 S. Cui, J. He, P. He, F. Zhang, C. Ling and Y. Ma, Theadaptation of Arctic phytoplankton to low light andsalinity in Kongsfjorden (Spitsbergen), Adv. Polar Sci.,2012, 23, 19–24.
20 Y. Zhuang, H. Jin, J. Chen, B. Wang, H. Li, F. Chen, Y. Liuand J. Xu, Nutrient status and phytoplankton-pigmentsresponse to ice melting in the Arctic Ocean, Adv. PolarSci., 2012, 24, 151–158.
21 P. Coupel, H. Y. Jin, M. Joo, R. Horner, H. A. Bouvet,M.-A. Sicre, J.-C. Gascard, J. F. Chen, V. Garçon andD. Ruiz-Pino, Phytoplankton distribution in unusually lowsea ice cover over the Pacific Arctic, Biogeosciences, 2012,9, 4835–4850.
22 http://earth.columbia.edu, http://earth.columbia.edu/articles/view/2993.
23 K. R. Bjørklund, S. B. Kruglikova and O. R. Anderson,Modern incursions of tropical Radiolaria into the ArcticOcean, J. Micropalaeontol., 2012, 31, 139–158.
24 P. Renaud, J. Berge, Ø. Varpe, O. Lønne, J. Nahrgang,C. Ottesen and I. Hallanger, Is the poleward expansionby Atlantic cod and haddock threatening nativepolar cod, Boreogadus saida?, Polar Biol., 2012, 35, 401–412.
25 S. Larsen, T. Andersen and D. Hessen, Climate changepredicted to cause severe increase of organic carbon inlakes, Global. Change Biol., 2011, 17, 1186–1192.
26 H. F. Wilson, J. E. Saiers, P. A. Raymond andW. V. Sobczak, Hydrologic drivers and seasonality of dis-solved organic carbon concentration, nitrogen content,bioavailability, and export in a forested New Englandstream, Ecosystems, 2013, 16, 604–616.
27 S. A. Bocaniov, D. R. Barton, S. L. Schiff andR. E. H. Smith, Impact of tributary DOM and nutrientinputs on the nearshore ecology of a large, oligotrophiclake (Georgian Bay, Lake Huron, Canada), Aquat. Sci.,2013, 75, 321–332.
28 C. G. Fichot, K. Kaiser, S. B. Hooker, R. M. Amon,M. Babin, S. Bélanger, S. A. Walker and R. Benner,Pan-Arctic distributions of continental runoff in the ArcticOcean, Sci. Rep., 2013, 3, 1053.
29 V. E. Romanovsky, S. L. Smith, H. H. Christiansen,N. I. Shiklomanov, D. A. Streletskiy, D. S. Drozdov,N. G. Oberman, A. L. Kholodov and S. S. Marchenko,Permafrost, NOAA Report No., 2013, pp. 131–136. http://www.arctic.noaa.gov/reportcard.
30 R. M. Cory, B. C. Crump, J. A. Dobkowski and G. W. Kling,Surface exposure to sunlight stimulates CO2 release frompermafrost soil carbon in the Arctic, Proc. Natl. Acad.Sci. U. S. A., 2013, 110, 3429–3434.
31 J. E. Vonk, P. J. Mann, S. Davydov, A. Davydova,R. G. M. Spencer, J. Schade, W. V. Sobczak, N. Zimov,S. Zimov, E. Bulygina, T. J. Eglinton and R. M. Holmes,High biolability of ancient permafrost carbon upon thaw,Geophys. Res. Lett., 2013, 40, 2689–2693.
32 S. Couture, D. Houle and C. Gagnon, Increases ofdissolved organic carbon in temperate and boreal lakesin Quebec, Canada, Environ. Sci. Pollut. Res., 2012, 19,361–371.
33 S. J. Kohler, D. Kothawala, M. N. Futter, O. Liungman andL. Tranvik, In-lake processes offset increased terrestrialinputs of dissolved organic carbon and color to lakes,PLoS One, 2013, 8, e70598.
34 L. B. Knoll, Linking watershed-scale features and processesto carbon, nitrogen, and phosphorus fluxes, PhD thesis,Miami University, Oxford, OH, 2011.
35 G. S. Dhillon and S. Inamdar, Extreme storms and changesin particulate and dissolved organic carbon in runoff:Entering uncharted waters?, Geophys. Res. Lett., 2013, 40.
36 S. Sandro and J. Melack, The effect of an extreme rainevent on the biogeochemistry and ecosystem metabolismof an oligotrophic high-elevation lake, Arct. Antarct. Alp.Res., 2012, 44, 222–231.
37 K. J. Goodman, M. A. Baker and W. A. Wurtsbaugh, Lakesas buffers of stream dissolved organic matter (DOM)variability: Temporal patterns of DOM characteristics inmountain stream-lake systems, J. Geophys. Res.: Biogeosci.,2011, 116, G00N02.
38 N. R. Lottig, I. Buffam and E. H. Stanley, Comparisons ofwetland and drainage lake influences on stream dissolved
Perspective Photochemical & Photobiological Sciences
120 | Photochem. Photobiol. Sci., 2015, 14, 108–126 This journal is © The Royal Society of Chemistry and Owner Societies 2015

carbon concentrations and yields in a north temperatelake-rich region, Aquat. Sci., 2013, 75, 619–630.
39 L. J. Tranvik, J. Downing, J. B. Cotner, S. Loiselle,R. Striegl, T. J. Ballatore, P. Dillon, K. Finlay, K. Fortino,L. B. Knoll, P. L. Korelainen, T. Kutser, S. Larsen,I. Laurion, D. M. Leech, S. L. McCallister, D. McKnight,J. Melack, E. Overholt, J. A. Porter, Y. Prairie, W. Renwick,F. Roland, B. S. Sherman, D. E. Schindler, S. Sobek,A. Tremblay, M. J. Vanni, A. M. Verschoor,E. v. Wachenfeldt and G. A. Weyhenmeyer, Lakes andreservoirs as regulators of carbon cycling and climate,Limnol. Oceanogr., 2009, 54, 2298–2314.
40 D. Bastviken, L. J. Tranvik, J. A. Downing, P. M. Crill andA. A. Enrich-Prast, Freshwater methane emissionsoffset the continental carbon sink, Science, 2011,331, 50.
41 E. S. Kritzberg and S. M. Ekström, Increasing iron concen-trations in surface waters – a factor behind brownifica-tion?, Biogeosciences, 2012, 9, 1465–1478.
42 P. Porcal, P. J. Dillon and L. A. Molot, Interaction of extrin-sic chemical factors affecting photodegradation of dis-solved organic matter in aquatic ecosystems, Photochem.Photobiol. Sci., 2014, 13, 799–812.
43 E. Caverly, J. M. Kaste, G. S. Hancock andR. M. Chambers, Dissolved and particulate organiccarbon fluxes from an agricultural watershed during con-secutive tropical storms, Geophys. Res. Lett., 2013, 40,5147–5152.
44 Y. H. Lu, J. E. Bauer, E. A. Canuel, Y. Yamashita,R. M. Chambers and R. Jaffe, Photochemical andmicrobial alteration of dissolved organic matter in tem-perate headwater streams associated with different landuse, J. Geophys. Res.: Biogeosci., 2013, 118, 566–580.
45 M. J. Macdonald and E. C. Minor, Photochemical degra-dation of dissolved organic matter from streams in thewestern Lake Superior watershed, Aquat. Sci., 2013, 75,509–522.
46 L. Forsstrom, T. Roiha and M. Rautio, Responses ofmicrobial food web to increased allochthonous DOM inan oligotrophic subarctic lake, Aquat. Microb. Ecol., 2013,68, 171–184.
47 C. E. H. Kissman, C. E. Williamson, K. C. Rose andJ. E. Saros, Response of phytoplankton in an alpine laketo inputs of dissolved organic matter through nutrientenrichment and trophic forcing., Limnol. Oceanogr., 2013,58, 867–880.
48 J. S. Read and K. C. Rose, Physical responses of smalltemperate lakes to variation in dissolved organic carbonconcentrations, Limnol. Oceanogr., 2013, 58, 921–931.
49 K. Kadir and K. L. Nelson, Sunlight mediated inactivationmechanisms of Enterococcus faecalis and Escherichia coliin clear water versus waste stabilization pond water, WaterRes., 2014, 50, 307–317.
50 A. Silverman, B. M. Peterson, A. B. Boehm, K. McNeill andK. L. Nelson, Sunlight inactivation of human viruses andbacteriophages in coastal waters containing natural
photosensitizers, Environ. Sci. Technol., 2013, 47, 1870–1878.
51 A. D. Stasko, J. M. Gunn and T. A. Johnston, Role ofambient light in structuring north-temperate fish commu-nities: potential effects of increasing dissolved organiccarbon concentration with a changing climate, Environ.Rev., 2012, 20, 173–190.
52 E. P. Overholt, S. H. Hall, C. E. Williamson, C. K. Meikle,M. A. Duffy and C. E. Cáceres, Solar radiation decreasesparasitism in Daphnia, Ecol. Lett., 2012, 15, 47–54.
53 S. Haaland, D. Hongve, H. Laudon, G. Riise andR. D. Vogt, Quantifying the drivers of the increasingcolored organic matter in boreal surface waters, Environ.Sci. Technol., 2010, 44, 2975–2980.
54 Y. Wu, K. Gao, G. Li and E. W. Helbling, Seasonal impactsof solar UV radiation on photosynthesis of phytoplanktonassemblages in the coastal waters of the South China Sea,Photochem. Photobiol., 2010, 86, 586–592.
55 J. M. Beman, C.-E. Chow, A. L. King, Y. Feng,J. A. Fuhrman, A. Andersson, N. R. Bates, B. N. Popp andD. A. Hutchins, Global declines in oceanic nitrificationrates as a consequence of ocean acidification, Proc. Natl.Acad. Sci. U. S. A., 2011, 108, 208–213.
56 K. Gao, E. W. Helbling, D.-P. Häder and D. A. Hutchins,Responses of marine primary producers to interactionsbetween ocean acidification, solar radiation, andwarming, Mar. Ecol.: Prog. Ser., 2012, 470, 167–189.
57 M. Steinacher, F. Joos, T. L. Froelicher, L. Bopp, P. Cadule,V. Cocco, S. C. Doney, M. Gehlen, K. Lindsay, J. K. Moore,B. Schneider and J. Segschneider, Projected 21st centurydecrease in marine productivity: a multi-model analysis,Biogeosciences, 2010, 7, 979–1005.
58 J. Turner, N. E. Barrand, T. J. Bracegirdle, P. Convey,D. A. Hodgson, M. Jarvis, A. Jenkins, G. Marshall,M. P. Meredith, H. Roscoe, J. Shanklin, J. French,H. Goosse, M. Guglielmin, J. Gutt, S. Jacobs,M. C. Kennicutt II, V. Masson-Delmotte, P. Mayewski!,F. Navarro, S. Robinson, T. Scambos, M. Sparrow,C. Summerhayes, K. Speer and A. Klepikov, Antarcticclimate change and the environment: an update,Cambridge University Press, Report No., Cambridge,2013, pp. 1–23.
59 G. T. Taylor, F. E. Muller-Karger, R. C. Thunell,M. I. Scranton, Y. Astor, R. Varela, L. T. Ghinaglia,L. Lorenzoni, K. A. Fanning and S. Hameed, Ecosystemresponses in the southern Caribbean Sea to global climatechange, Proc. Natl. Acad. Sci. U. S. A., 2012, 109, 19315–19320.
60 E. W. Helbling, A. G. Buma, P. Boelen, H. J. Van der Strate,M. V. Fiorda Giordanino and V. E. Villafañe, Increase inRubisco activity and gene expression due to elevated temp-erature partially counteracts ultraviolet radiation-inducedphotoinhibition in the marine diatom Thalassiosira weiss-flogii, Limnol. Oceanogr., 2011, 56, 1330–1342.
61 S. R. Halac, V. E. Villafañe and E. W. Helbling, Tempera-ture benefits the photosynthetic performance of the
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 108–126 | 121

diatoms Chaetoceros gracilis and Thalassiosira weissflogiiwhen exposed to UVR, J. Photochem. Photobiol., B, 2010,101, 196–205.
62 G. Li and K. Gao, Variation in UV irradiance related tostratospheric ozone levels affects photosynthetic carbonfixation of winter phytoplankton assemblages fromsurface coastal water of the South China Sea, Mar. Biol.Res., 2012, 8, 670–676.
63 K. Gao and D. Campbell, Photophysiological responses ofmarine diatoms to elevated CO2 and decreased pH: areview, Funct. Plant Biol., 2014, 41, 449–459.
64 M. Thyssen, G. Ferreyra, S. Moreau, I. Schloss, M. Denisand S. Demers, The combined effect of ultraviolet Bradiation and temperature increase on phytoplanktondynamics and cell cycle using pulse shape recordingflow cytometry, J. Exp. Mar. Biol. Ecol., 2011, 406, 95–107.
65 G. Li, Z. W. Che and K. Gao, Photosynthetic carbonfixation by tropical coral reef phytoplankton assemblages:a UVR perspective, Algae, 2013, 28, 281–288.
66 G. Li, K. Gao and G. Gao, Differential impacts of solar UVradiation on photosynthetic carbon fixation from thecoastal to offshore surface waters in the South China Sea,Photochem. Photobiol., 2011, 87, 329–334.
67 R. F. Keeling, A. Körtzinger and N. Gruber, Ocean deoxy-genation in a warming world, Ann. Rev. Mar. Sci., 2010, 2,199–229.
68 G. M. Hallegraeff, Ocean climate change, phytoplanktoncommunity responses, and harmful algal blooms: aformidable predictive challenge, J. Phycol., 2010, 46, 220–235.
69 Richa, R. P. Sinha and D.-P. Häder, Phytoplankton pro-ductivity in a changing global climate, in Phytoplankton:Biology, Classification and Environmental Impacts, ed.M. T. Sebastiá, Nova Science Publishers, New York, 2014,pp. 1–35.
70 O. Hoegh-Guldberg, Dangerous shifts in ocean ecosystemfunction?, ISME J., 2010, 4, 1090–1092.
71 R. Terrado, K. Scarcella, M. Thaler, W. F. Vincent andC. Lovejoy, Small phytoplankton in Arctic seas: vulner-ability to climate change, Biodiversity Conserv., 2013, 14,2–18.
72 L. Beaufort, I. Probert, T. De Garidel-Thoron,E. M. Bendif, D. Ruiz-Pino, N. Metzl, C. Goyet, N. Buchet,P. Coupel, M. Grelaud, B. Rost, R. E. M. Rickaby andC. de Vargas, Sensitivity of coccolithophores to carbonatechemistry and ocean acidification, Nature, 2011, 476,80–83.
73 K. Gao, J. Xu, G. Gao, Y. Li, D. A. Hutchins, B. Huang,Y. Zheng, P. Jin, X. Cai, D.-P. Häder, W. Li, K. Xu, N. Liuand U. Riebesell, Rising carbon dioxide and increasinglight exposure act synergistically to reduce marineprimary productivity, Nat. Clim. Change, 2012, 2, 519–523.
74 S. Chen and K. Gao, Solar ultraviolet radiation and CO2-induced ocean acidification interacts to influence thephotosynthetic performance of the red tide alga Phaeo-
cystis globosa (Prymnesiophyceae), Hydrobiologia, 2011,675, 105–117.
75 W. Li and K. S. Gao, A marine secondary producer respiresand feeds more in a high CO2 ocean, Mar. Pollut. Bull.,2012, 64, 699–703.
76 K. Gao and Y. Zheng, Combined effects of ocean acidifica-tion and solar UV radiation on photosynthesis, growth,pigmentation and calcification of the coralline algaCorallina sessilis (Rhodophyta), Global. Change Biol., 2010,16, 2388–2398.
77 K. R. N. Anthony, J. A. Maynard, G. Diaz-Pulido,P. J. Mumby, P. A. Marshall, L. Cao andO. Hoegh-Guldenberg, Ocean acidification and warmingwill lower coral reef resilience, Global. Change Biol., 2011,17, 1798–1808.
78 W. Guan and K. Gao, Impacts of UV radiation on photo-synthesis and growth of the coccolithophore Emilianiahuxleyi (Haptophyceae), Environ. Exp. Bot., 2010, 67, 502–508.
79 K. Xu, K. Gao, V. Villafañe and E. Helbling, Photosyntheticresponses of Emiliania huxleyi to UV radiation and elev-ated temperature: roles of calcified coccoliths, Biogeo-sciences, 2011, 8, 1441–1452.
80 W. Broadgate, U. Riebesell, C. Armstrong, P. Brewer,K. Denman, R. Feely, K. Gao, J.-P. Gattuso, K. Isensee andJ. Kleypas, Ocean acidification summary for policymakers–Third symposium on the ocean in a high-CO2 world,2013.
81 N. Bednaršek, G. A. Tarling, D. C. E. Bakker,Z. S. Fielding, E. M. Jones, H. J. Venables, P. Ward,A. Kuzirian, B. Lézé, R. A. Feely and E. J. Murphy, Exten-sive dissolution of live pteropods in the Southern Ocean,Nat. Geosci., 2012, 5, 881–885.
82 D. Shi, Y. Xu, B. M. Hopkinson and F. M. M. Morel, Effectof ocean acidification on iron availability to marine phyto-plankton, Science, 2010, 327, 676–679.
83 Y. Zhang, H.-B. Jiang and B.-S. Qiu, Effects of UVB radi-ation on competition between the bloom-forming cyano-bacterium Microcystis aeruginosa and the chlorophyceaeChlamydomonas microsphaera, J. Phycol., 2013, 49, 318–328.
84 S. P. Singh, R. P. Rastogi, R. P. Sinha and D.-P. Häder,Temporal dynamics of ROS biogenesis under simulatedsolar radiation in the cyanobacterium Anabaena variabilisPCC 7937, Protoplasma, 2014, 251, 1223–1230.
85 R. Muller, C. Desel, F. S. Steinhoff, C. Wiencke andK. Bischof, UV-radiation and elevated temperaturesinduce formation of reactive oxygen species in gameto-phytes of cold-temperate/Arctic kelps (Laminariales,Phaeophyceae), Phycol. Res., 2012, 60, 27–36.
86 D.-P. Häder, E. W. Helbling, C. E. Williamson andR. C. Worrest, Effects of UV radiation on aquatic eco-systems and interactions with climate change, Photochem.Photobiol. Sci., 2011, 10, 242–260.
87 Y. Li, K. Gao, V. Villafañe and E. Helbling, Ocean acidifica-tion mediates photosynthetic response to UV radiation
Perspective Photochemical & Photobiological Sciences
122 | Photochem. Photobiol. Sci., 2015, 14, 108–126 This journal is © The Royal Society of Chemistry and Owner Societies 2015

and temperature increase in the diatom Phaeodactylumtricornutum, Biogeosci. Discuss., 2012, 9, 7197–7226.
88 D. Wu, Q. Hu, Z. Yan, W. Chen, C. Yan, X. Huang,J. Zhang, P. Yang, H. Deng and J. Wang, Structural basisof ultraviolet-B perception by UVR8, Nature, 2012, 484,214–219.
89 S. P. Singh, D.-P. Häder and R. P. Sinha, Cyanobacteriaand ultraviolet radiation (UVR) stress: mitigation strat-egies, Ageing Res. Rev., 2010, 9, 79–90.
90 Richa, R. P. Sinha and D.-P. Häder, Physiologicalaspects of UV-excitation of DNA, in Photoinduced Phenom-ena in Nucleic Acids, ed. B. Barbatti, A. C. Borin andA. C. Ullrich, Springer, Berlin, Heidelberg, 2014,pp. 1–46.
91 B. B. Barnes, C. Hu, J. P. Cannizzaro, S. E. Craig,P. Hallock, D. L. Jones, J. C. Lehrter, N. Melo,B. A. Schaeffer and R. Zepp, Estimation of diffuse attenu-ation of ultraviolet light in optically shallow Florida Keyswaters from MODIS measurements, Rem. Sens. Environ.,2014, 140, 519–532.
92 R. P. Rastogi, R. P. Sinha and A. Incharoensakdi, Partialcharacterization, UV-induction and photoprotective func-tion of sunscreen pigment, scytonemin from Rivularia sp.HKAR-4, Chemosphere, 2013, 93, 1874–1878.
93 C. A. Llewellyn, D. A. White, V. Martinez-Vincente,G. Tarran and T. J. Smyth, Distribution of mycosporine-like amino acids along a surface water meridional transectof the Atlantic, Micro. Ecol., 2012, 64, 320–333.
94 G. Singh, P. K. Babele, R. P. Sinha, M. B. Tyagi andA. Kumar, Enymatic and non-enzymatic defense mecha-nisms against ultraviolet-B radiation in two Anabaenaspecies, Process Biochem., 2013, 48, 796–802.
95 J. F. Bornman, P. W. Barnes, S. A. Robinson, C. L. Ballaré,S. D. Flinte and M. M. Caldwell, Solar ultraviolet radiationand ozone depletion-driven climate change: effects on ter-restrial ecosystems, Photochem. Photobiol. Sci., 2015, 14,DOI: 10.1039/c4pp90034k, this issue.
96 K. A. Garver, A. A. M. Mahony, D. Stucchi, J. Richard,C. Van Woensel and M. Foreman, Estimation of para-meters influencing waterborne transmission of infectiousHematopoietic Necrosis Virus (IHNV) in Atlantic Salmon(Salmo salar), PLoS One, 2013, 8, e82296.
97 S. A. A. Jassim and R. G. Limoges, Impact of externalforces on cyanophage-host interactions in aquatic eco-systems, World J. Micrbiol. Biotechnol, 2013, 29, 1751–1762.
98 A. Studer, V. M. Cubillos, M. D. Lamare, R. Poulin andD. J. Burritt, Effects of ultraviolet radiation on an interti-dal trematode parasite: An assessment of damage andprotection, Int. J. Parasitol., 2012, 42, 453–461.
99 A. Studer, M. D. Lamare and R. Poulin, Effects ofultraviolet radiation on the transmission process of anintertidal trematode parasite, Parasitology, 2012, 139,537–546.
100 A. Studer and R. Poulin, Cercarial survival in an intertidaltrematode: a multifactorial experiment with temperature,
salinity and ultraviolet radiation, Parasitol. Res., 2013, 112,243–249.
101 T. N. Quinn, N. W. Kendall, H. B. Rich Jr. andB. E. Chasco, Diel vertical movements, and effects ofinfection by the cestode Schistocephalus solidus ondaytime proximity of three-spined sticklebacks Gastero-steus aculeatus to the surface of a large Alaskan lake, Oeco-logia, 2012, 168, 43–51.
102 M. Sweet, N. Kirkham, M. Bendall, L. Currey, J. Bythelland M. Heupel, Evidence of melanoma in wild marinefish populations, PLoS One, 2012, 7, e41989.
103 A. R. Harborne, The ecology, behaviour and physiology offishes on coral reef flats, and the potential impacts ofclimate change, J. Fish Biol., 2013, 83, 417–447.
104 X. Yuan, K. Yin, P. J. Harrison and J. Zhang, Phyto-plankton are more tolerant to UV than bacteria andviruses in the northern South China Sea, Aquat. Microbiol.Ecol., 2011, 65, 117–128.
105 J. M. Manrique, A. Y. Calvo, S. R. Halac, V. E. Villafañe,L. R. Jones and E. W. Helbling, Effects of UV radiation onthe taxonomic composition of natural bacterioplanktoncommunities from Bahía Engaño (Patagonia, Argentina),J. Photochem. Photobiol., B, 2012, 117, 171–178.
106 C. Romera-Castillo, H. Sarmento, X. A. Álvarez-Salgado,J. M. Gasol and C. Marrasé, Net production and con-sumption of fluorescent colored dissolved organic matterby natural bacterial assemblages growing on marinephytoplankton exudates, Appl. Environ. Microbiol., 2011,77, 7490–7498.
107 R. Bertoni, W. H. Jeffrey, M. Pujo-Pay, L. Oriol, P. Conanand F. Joux, Influence of water mixing on the inhibitoryeffect of UV radiation on primary and bacterial productionin Mediterranean coastal water, Aquat. Sci., 2011, 73, 377–387.
108 R. P. Goldman and M. Travisano, Experimental evolutionof ultraviolet radiation resistance in Escherichia coli, Evol-ution, 2011, 65, 3486–3498.
109 T. Key, A. McCarthy, D. A. Campbell, C. Six, S. Roy andZ. V. Finkel, Cell size trade-offs govern light exploitationstrategies in marine phytoplankton, Environ. Microbiol.,2010, 12, 95–104.
110 M. Llabrés, S. Agustí, P. Alonso-Laita and G. Herndl,Synechococcus and Prochlorococcus cell death inducedby UV radiation and the penetration of lethal UVR inthe Mediterranean Sea, Mar. Ecol.: Prog. Ser., 2010, 399,27–37.
111 M. Llabrés, S. Agustí, M. Fernández, A. Canepa,F. Maurin, F. Vidal and C. M. Duarte, Impact of elevatedUVB radiation on marine biota: a meta-analysis, GlobalEcol. Biogeogr., 2013, 22, 131–144.
112 P. Jin, K. Gao, V. Villafañe, D. Campbell and W. Helbling,Ocean acidification alters the photosynthetic responses ofa coccolithophorid to fluctuating ultraviolet and visibleradiation, Plant Physiol., 2013, 162, 2084–2094.
113 M. Medina-Sanchez, J. A. Delgado-Molina, J. Bratbak,G. J. Bullejos, F. Villar-Argaiz and M. Carrillo, Maximum
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 108–126 | 123

in the middle: Nonlinear response of microbial planktonto ultraviolet radiation and phosphorus, PLoS One,2013, 8.
114 J. M. Sereda, D. M. Vandergucht and J. J. Hudson, In situUVA exposure modulates change in the uptake of radio-phosphate in size-fractionated plankton assemblages fol-lowing UVR exposure, Micro. Ecol., 2012, 63, 751–760.
115 D. M. Martynova, N. A. Kazus, U. V. Bathmann, M. Graeveand A. A. Sukhotin, Seasonal abundance and feeding pat-terns of copepods Temora longicornis, Centropages hamatusand Acartia spp. in the White Sea (66°N), Polar Biol., 2011,34, 1175–1195.
116 S. Nahon, F. Charles, F. Lantoine, G. Vétion,K. Escoubeyrou, M. Desmalades and A. M. Pruski,Ultraviolet radiation negatively affects growth and foodquality of the pelagic diatom Skeletonema costatum, J. Exp.Mar. Biol. Ecol., 2010, 383, 164–170.
117 A. U. Bracher and C. Wiencke, Simulation of the effects ofnaturally enhanced UV radiation on photosynthesis ofAntarctic phytoplankton, Mar. Ecol.: Prog. Ser., 2000, 196,127–141.
118 K. Oubelkheir, L. A. Clementson, G. F. Moore andG. H. Tilstone, Production of mycosporine-like aminoacids by phytoplankton under ultraviolet radiationexposure in the Sub-Antarctic Zone south of Tasmania,Mar. Ecol.: Prog. Ser., 2013, 494, 41–63.
119 K. G. Ryan, A. McMinn, E. N. Hegseth and S. K. Davy, Theeffects of ultraviolet radiation on Antarctic sea-ice algae,J. Phycol., 2012, 48, 74–84.
120 S. P. Singh, S.-Y. Ha, R. P. Sinha and D.-P. Häder, Photo-heterotrophic growth unprecedentedly increases the bio-synthesis of mycosporine-like amino acid shinorine inthe cyanobacterium Anabaena sp., isolated from hotsprings of Rajgir (India), Acta Physiol. Plant., 2014, 36,389–397.
121 Z. Xu and K. Gao, Impacts of UV radiation on growthand photosynthetic carbon acquisition in Gracilarialemaneiformis (Rhodophyta) under phosphorus-limitedand replete conditions, Funct. Plant Biol., 2009, 36, 1057–1064.
122 C. A. Llewellyn and R. L. Airs, Distribution and abundanceof MAAs in 33 species of microalgae across 13 classes,Mar. Drugs, 2010, 8, 1273–1291.
123 S. Y. Ha, Y. M. Kim, M. O. Park, S. H. Kang, H. C. Kim andK. H. Shin, Production of mycosporine-like amino acidsof in situ phytoplankton community in Kongsfjorden,Svalbard, Arctic, J. Photochem. Photobiol., B, 2012, 114,1–14.
124 L. M. Ayoub, P. Hallock, P. G. Coble and S. S. Bell,MAA-like absorbing substances in Florida Keys phyto-plankton vary with distance from shore and CDOM: Impli-cations for coral reefs, J. Exp. Mar. Biol. Ecol., 2012, 420,91–98.
125 M. Fiorda Giordanino, S. M. Strauch, V. E. Villafañe andE. W. Helbling, Influence of temperature and UVRon photosynthesis and morphology of four species of
cyanobacteria, J. Photochem. Photobiol., B, 2011, 103,68–77.
126 G. Li and K. Gao, Cell size-dependent effects of solar UVradiation on primary production in coastal waters of theSouth China Sea, Estuaries Coasts, 2013, 36, 728–736.
127 D. G. Boyce, M. R. Lewis and B. Worm, Global phyto-plankton decline over the past century, Nature, 2010, 466,591–596.
128 C. Laufkoetter, M. Vogt and N. Gruber, Trends in marineplankton composition and export production ina CCSM-BEC hindcast (1960–2006), in EGU GeneralAssembly Conference Abstracts, 2013, vol. 15, p. 11917.
129 P. W. Boyd, Beyond ocean acidification, Nat. Geosci., 2011,4, 273–274.
130 D.-P. Häder, Does enhanced solar UV-B radiation affectmarine primary producers in their natural habitats?,Photochem. Photobiol., 2011, 87, 263–266.
131 I. E. Huertas, M. Rouco, V. López-Rodas and E. Costas,Warming will affect phytoplankton differently: evidencethrough a mechanistic approach, Proc. R. Soc. London, B,2011, 278, 3534–3543.
132 C. Ruiz-González, M. Galí, E. Sintes, G. J. Herndl,J. M. Gasol and R. Simó, Sunlight effects on theosmotrophic uptake of DMSP-sulfur and leucine by Polarphytoplankton, PLoS One, 2012, 7, e45545.
133 K. D. Six, S. Kloster, T. Ilyina, S. D. Archer, K. Zhang andE. Maier-Reimer, Global warming amplified by reducedsulphur fluxes as a result of ocean acidification, Nat. Clim.Change, 2013, 3, 975–978.
134 D. G. Petersen and I. Dahllöf, Combined effects of pyreneand UV-light on algae and bacteria in an Arctic sediment,Ecotoxicology, 2007, 16, 371–377.
135 C. J. Galbán-Malagón, A. Cabrerizo, N. Berrojálbiz,M. J. Ojeda and J. Dachs, Air-water exchange andphytoplankton accumulation of persistent organicpollutants in the Greenland current and Arctic Ocean,http://132.246.11.198/2012-ipy/pdf-all/ipy2012arAbstract00801.pdf.
136 M. Piepho, M. T. Arts and A. Wacker, Species-specific vari-ation in fatty acid concentrations of four phytoplanktonspecies: Does phosphorus supply influence the effect oflight intensity or temperature?, J. Phycol., 2012, 48, 64–73.
137 D.-P. Häder, P. Richter, V. E. Villafañe and E. W. Helbling,Influence of light history on the photosynthetic and moti-lity responses of Gymnodinium chlorophorum exposed toUVR and different temperatures, J. Photochem. Photobiol.,B, 2014, in press.
138 S. Arndt, G. Lacroix, N. Gypens, P. Regnier andC. Lancelot, Nutrient dynamics and phytoplankton devel-opment along an estuary–coastal zone continuum: amodel study, J. Mar. Syst., 2011, 84, 49–66.
139 M. P. Skinner, R. J. Lewis and S. Morton, Ecology of theciguatera causing dinoflagellates from the Northern GreatBarrier Reef: Changes in community distribution andcoastal eutrophication, Mar. Pollut. Bull., 2013, 77, 210–219.
Perspective Photochemical & Photobiological Sciences
124 | Photochem. Photobiol. Sci., 2015, 14, 108–126 This journal is © The Royal Society of Chemistry and Owner Societies 2015

140 P. Echeveste, S. Agustí and J. Dachs, Cell size dependenceof additive versus synergetic effects of UV radiation andPAHs on oceanic phytoplankton, Environ. Pollut., 2011,159, 1307–1316.
141 M. H. Olson and N. E. Barbieri, Mechanisms of ultravioletradiation tolerance in the freshwater snail Physa acuta,Freshwater Sci., 2013, 33, 66–72.
142 J. Ahlgren, X. Yang, L.-A. Hansson and C. Brönmark,Camouflaged or tanned: plasticity in freshwater snail pig-mentation, Biol. Lett., 2013, 9, 20130464.
143 S. Nahon, A. M. Pruski, J. C. Duchene, L. Mejanelle,G. Vetion, M. Desmalades and F. Charles, Can UV radi-ation affect benthic deposit-feeders through biochemicalalteration of food resources? An experimental study withjuveniles of the benthic polychaete Eupolymnia nebulosa,Mar. Environ. Res., 2011, 71, 266–274.
144 L. Van Den Broecke, K. Martens, V. Pieri and I. Schön,Ostracod valves as efficient UV protection, J. Limnol., 2012,71.
145 K. Bischof and F. S. Steinhoff, Impacts of stratosphericozone depletion and solar UVB radiation on seaweeds,in Seaweed Biology, Springer, Berlin, Heidelberg, 2012,vol. 219, pp. 433–448.
146 M. Olischläger and C. Wiencke, Seasonal fertility andcombined effects of temperature and UV-radiation onAlaria esculenta and Laminaria digitata (Phaeophyceae)from Spitsbergen, Polar Biol., 2013, 36, 1019–1029.
147 A. Fricke, M. Molis, C. Wiencke, N. Valdivia andA. S. Chapman, Effects of UV radiation on the structureof Arctic macrobenthic communities, Polar Biol., 2011, 34,995–1009.
148 A. R. Davis, D. Coleman, A. Broad, M. Byrne,S. A. Dworjanyn and R. Przeslawski, Complex responses ofintertidal molluscan embroys to a warming and acidifyingocean in the presence of UV radiation, PLoS One, 2013, 8,e55939.
149 C. Vadadi-Fulop, C. Sipkay, G. Meszaros and L. Hufnagel,Climate change and freshwater zooplankton: what does itboil down to?, Aquat. Ecol., 2012, 46, 501–519.
150 L. Nevalainen and M. Rautio, Cladoceran carapace absor-bance as a new method for inferring past UV radiationexposure of aquatic biota, Quat. Sci. Rev., 2013, 84, 109–115.
151 S. Hylander, J. C. Grenvald and T. Kiorboe, Fitness costsand benefits of UVR exposure in marine pelagic copepods,Funct. Ecol., 2014, 28, 149–158.
152 C. E. Williamson, J. M. Fischer, S. M. Bollens,E. P. Overholt and J. K. Breckenridge, Toward a more com-prehensive theory of zooplankton diel vertical migration:Integrating ultraviolet radiation and water transparencyinto the biotic paradigm, Limnol. Oceanogr., 2011, 56,1603–1623.
153 R. Tiberti and M. Barbieri, Evidences of zooplankton verti-cal migration in stocked and never-stocked alpine lakes inGran Paradiso National Park (Italy), Oceanol. Hydrobiol.Stud., 2011, 40, 36–42.
154 B. Pietrzak, A. Bednarska, M. Markowska, M. Rojek,E. Szymanska and M. Slusarczyk, Behavioural and physio-logical mechanisms behind extreme longevity in Daphnia,Hydrobiologia, 2013, 715, 125–134.
155 R. Tiberti and R. Iacobuzio, Does the fish presenceinfluence the diurnal vertical distribution of zooplanktonin high transparency lakes?, Hydrobiologia, 2013, 709,27–39.
156 K. C. Rose, C. E. Williamson, J. M. Fischer, S. J. Connelly,M. Olson, A. J. Tucker and D. A. Noe, The role of UV andfish in regulating the vertical distribution of Daphnia,Limnol. Oceanogr., 2012, 57, 1867–1876.
157 D. M. Fields, C. M. F. Durif, R. M. Bjelland, S. D. Shema,A. B. Skiftesvik and H. I. Browman, Grazing rates ofCalanus finmarchicus on Thalassiosira weissflogii culturedunder different levels of ultraviolet radiation, PLoS One,2011, 6.
158 M. S. Souza, L. A. Hansson, S. Hylander, B. Modenutti andE. Balseiro, Rapid enzymatic response to compensate UVradiation in copepods, PLoS One, 2012, 7, e32046.
159 S. Hylander, M. S. Souza, E. Balseiro, B. Modenutti andL. A. Hansson, Fish-mediated trait compensation in zoo-plankton, Funct. Ecol., 2012, 26, 608–615.
160 T. Schneider, A. Herzig, K. A. Koinig and R. Sommaruga,Copepods in turbid shallow soda lakes accumulateunexpected high levels of carotenoids, PLoS One, 2012, 7,e43063.
161 A. J. Tucker and C. E. Williamson, The invasion window forwarmwater fish in clearwater lakes: the role of ultravioletradiation and temperature, Div. Distrib., 2014, 20, 181–192.
162 A. J. Tucker, C. E. Williamson and J. T. Oris, Developmentand application of a UV attainment threshold for theprevention of warmwater aquatic invasive species, Biol.Invasions, 2012, 14, 2331–2342.
163 A. K. Gevertz, A. J. Tucker, A. M. Bowling,C. E. Williamson and J. T. Oris, Differential tolerance ofnative and nonnative fish exposed to ultraviolet radiationand fluoranthene in Lake Tahoe (California/Nevada), USA,Environ. Toxicol. Chem., 2012, 31, 1129–1135.
164 A. K. Gevertz and J. T. Oris, Microscopic examination ofskin in native and nonnative fish from Lake Tahoeexposed to ultraviolet radiation and fluoranthene, Aquat.Toxicol., 2014, 147, 151–157.
165 V. Boily, A. Bertolo, P. Magnan, M.-G. Martinoli andH.-M. Thérien, The effects of UVR irradiance and spectralcomposition on yellow perch (Perca flavescens) larvae sur-vival, Aquat. Sci., 2011, 73, 345–354.
166 M. C. Melnychuk, C. J. Walters, V. Christensen,M. L. Bothwell and D. W. Welch, Effects of solar ultra-violet radiation exposure on early ocean survival and fry-to-smolt growth of juvenile salmon, Mar. Ecol.: Prog. Ser.,2012, 457, 251–264.
167 E. Sucre, F. Vidussi, B. Mostajir, G. Charmantier andC. Lorin-Nebel, Impact of ultraviolet-B radiation on plank-tonic fish larvae: Alteration of the osmoregulatory func-tion, Aquat. Toxicol., 2012, 109, 194–201.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 108–126 | 125

168 Y. Fukunishi, H. I. Browman, C. M. F. Durif,R. M. Bjelland and A. B. Skiftesvik, Effect of sub-lethalexposure to ultraviolet radiation on the escape perform-ance of atlantic cod larvae (Gadus morhua), PLoS One,2012, 7, e35554.
169 Y. Fukunishi, H. I. Browman, C. M. F. Durif,R. M. Bjelland, S. D. Shema, D. M. Fields andA. B. Skiftesvik, Sub-lethal exposure to ultraviolet radi-ation reduces prey consumption by Atlantic cod larvae(Gadus morhua), Mar. Biol., 2013, 160, 2591–2596.
170 D. M. Leech, W. J. Boeing, S. L. Cooke, C. E. Williamsonand L. Torres, UV-enhanced fish predation and thedifferential migration of zooplankton in response to UVradiation and fish, Limnol. Oceanogr., 2009, 54, 1152–1161.
171 A. G. Finstad, I. P. Helland, O. Ugedal, T. Hesthagen andD. O. Hessen, Unimodal response of fish yield to dis-solved organic carbon, Ecol. Lett., 2014, 17, 36–43.
172 J. H. Olker, L. B. Johnson, R. P. Axler and C. M. Johnson,Factors influencing ultraviolet radiation dose to develop-ing frogs in northern vernal pools, Can. J. Fish. Aquat. Sci.,2013, 70, 1531–1541.
173 R. Danovaro, L. Bongiorni, C. Corinaldesi, D. Giovannelli,E. Damiani, P. Astolfi, L. Greci and A. Pusceddu, Sun-screens cause coral bleaching by promoting viral infec-tions, Environ. Health Perspect., 2008, 116, 441–447.
174 A. Bowman, L. M. Martinez-Levasseur, K. Acevedo-Whitehouse, D. Gendron and M. A. Birch-Machin,The simultaneous detection of mitochondrial DNAdamage from sun-exposed skin of three whale species andits association with UV-induced microscopic lesions andapoptosis, Mitochondrion, 2013, 13, 342–349.
175 L. M. Martinez-Levasseur, M. A. Birch-Machin,A. Bowman, D. Gendron, E. Weatherhead, R. J. Knell andK. Acevedo-Whitehouse, Whales use distinct strategies tocounteracts solar ultraviolet radiation, Sci. Rep., 2013, 3,2386.
176 P. Gago-Ferrero, M. B. Alonso, C. P. Bertozzi, J. Mango,L. Barbosa, M. Cremer, E. R. Secchi, C. Domit, A. Azevedo,J. Lailson-Brito, J. P. M. Torres, O. Malm, E. Eljarrat,M. S. Diaz-Cruz and D. Barcelo, First determination ofUVf filters in marine mammals. Octocrylene levels inFranciscana dolphins, Environ. Sci. Technol., 2013, 47,5619–5625.
Perspective Photochemical & Photobiological Sciences
126 | Photochem. Photobiol. Sci., 2015, 14, 108–126 This journal is © The Royal Society of Chemistry and Owner Societies 2015

Photochemical &Photobiological Sciences
PERSPECTIVE
Cite this: Photochem. Photobiol. Sci.,2015, 14, 127
Received 20th October 2014,Accepted 20th October 2014
DOI: 10.1039/c4pp90036g
www.rsc.org/pps
Effects of stratospheric ozone depletion, solar UVradiation, and climate change on biogeochemicalcycling: interactions and feedbacks
David J. Erickson III,*a Barbara Sulzberger,b Richard G. Zeppc and Amy T. Austind
Climate change modulates the effects of solar UV radiation on biogeochemical cycles in terrestrial and
aquatic ecosystems, particularly for carbon cycling, resulting in UV-mediated positive or negative feed-
backs on climate. Possible positive feedbacks discussed in this assessment include: (i) enhanced UV-
induced mineralisation of above ground litter due to aridification; (ii) enhanced UV-induced mineralisation
of photoreactive dissolved organic matter (DOM) in aquatic ecosystems due to changes in continental
runoff and ice melting; (iii) reduced efficiency of the biological pump due to UV-induced bleaching of
coloured dissolved organic matter (CDOM) in stratified aquatic ecosystems, where CDOM protects phyto-
plankton from the damaging solar UV-B radiation. Mineralisation of organic matter results in the pro-
duction and release of CO2, whereas the biological pump is the main biological process for CO2 removal
by aquatic ecosystems. This paper also assesses the interactive effects of solar UV radiation and climate
change on the biogeochemical cycling of aerosols and trace gases other than CO2, as well as of chemical
and biological contaminants. Interacting effects of solar UV radiation and climate change on biogeo-
chemical cycles are particularly pronounced at terrestrial-aquatic interfaces.
Introduction
The Montreal protocol has been successful in phasing outozone-depleting CFCs and, as a consequence, stratosphericozone concentrations are recovering at low and mid-latitudes1,2
(see Bais et al.3). However, springtime ozone depletion isexpected to continue at polar latitudes for many decades,4,5
largely due to climate change. A major consequence of strato-spheric ozone change is altered intensity of solar UV-B radi-ation which in turn affects the biogeochemical cycling ofcarbon and other chemical elements. Terrestrial and aquaticbiogeochemical cycles are discussed here in the context oftheir possible interactions with UV radiation and climatechange (Fig. 1).
The intensity of solar UV radiation reaching the Earth’ssurface is also controlled by climate-related variables such ascloud cover and aerosols. In addition, the penetration of UV-Bradiation into water bodies largely depends on the concen-tration and the optical properties of chromophoric dissolved
Fig. 1 Conceptual model of aquatic and terrestrial processes that arepossibly influenced by interactions between UV radiation and climatechange. Recent findings on these interactions are discussed in moredetail in this assessment. Reprinted from Williamson et al.,250 with per-mission from Macmillan Publishers Ltd: Nature Climate Change, copy-right 2014.
aComputational Earth Sciences Group Computer Science and Mathematics Division,
Oak Ridge National Laboratory, P.O. Box 2008, MS 6016 Oak Ridge TN 37831-6016,
USA. E-mail: [email protected]: Swiss Federal Institute of Aquatic Science and Technology, Ueberlandstrasse
133, CH-8600 Duebendorf, SwitzerlandcUnited States Environmental Protection Agency, 960 College Station Road Athens,
Georgia 30605-2700, USAdInstituto de Investigaciones Fisiológicas y Ecológicas Vinculadas a la Agricultura
(IFEVA) and Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET),
Universidad de Buenos Aires, Av. San Martín 4453, C1417DSE Buenos Aires,
Argentina
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 127–148 | 127

organic matter (referred to as coloured dissolved organicmatter, CDOM, hereafter). Climate-change related effects onterrestrial and aquatic ecosystems, e.g. desertification, oceanacidification and stratification, as well as land use changeinteract in various ways with solar UV radiation, resulting inUV-mediated feedbacks on climate. Feedbacks, combinedeffects and interactions of solar UV radiation and climatechange on biogeochemical cycles are discussed and assessed,and gaps in knowledge identified.
Interactive effects of stratosphericozone depletion and climate changeon terrestrial and aquatic ecosystems
We include a section that summarises some important issuesof climate change, since climate change modulates the effectsof solar UV radiation of biogeochemical cycles. Climatechange also affects the stratospheric ozone concentration andthus the intensity of solar UV radiation reaching the Earth’ssurface.
Climate change and polar stratospheric ozone depletion
Some chemistry-climate models and observations predict thatan ozone ‘hole’ may still be present in the Antarctic spring in2100.4 A large loss of stratospheric ozone, comparable to thatin the Antarctic ozone ‘hole’, was also observed over the Arcticin spring 20115 due, in part, to the extremely cold strato-spheric Arctic winter 2010/2011.6 In spring 2011 increasedlevels of solar UV radiation were observed at Arctic and sub-Arctic ground stations.7 The Arctic stratosphere is particularlyaffected by radiative cooling due to Arctic amplification(enhanced warming in high northern latitudes relative to thenorthern hemisphere).8 There are several reasons for the fastresponse of the Arctic to global warming, including increasedadvection of water from the Atlantic to the Arctic,9,10 andreduced albedo due to sea ice melting and vegetation shifts.11
Hence, increasing attention is being paid to Arctic tippingpoints.12 The term “tipping point” is used for systems (e.g.Arctic ecosystems), where a small change in forcing couldpotentially cause a large change in future ecosystems.
Further detail on climate change and polar stratosphericozone depletion is given in Bais et al.3
Ice melting: effects on local climates
Global warming has dramatically enhanced melting of gla-ciers, ice caps, and sea ice in both hemispheres, with thelargest present rates of melting in the NorthernHemisphere.13–15 The Arctic sea ice, whose extent usuallyreaches a minimum each year in September, is particularlyaffected13,15 (Fig. 2) (also see Fig. 1 in Häder et al.16). Meltingof Greenland and Antarctic sea ice is mainly due to warmerocean temperatures.17 Increasing meltwater ponds on thesurface of polar ice reduce albedo and allow more solar radi-ation, both UV and photosynthetically active radiation (PAR),
to enter the water column through the ice18 (see Bais et al.3).Screen and Simmonds (2010)8 suggest that positive ice-temp-erature feedbacks occur in the Arctic due to increases inatmospheric water vapour content that enhances warming inthe lower part of the atmosphere, especially in high northernlatitudes. Sea ice melting at high latitudes could also be due tothe transformation of solar UV-B radiation into heat by plank-tonic microorganisms in a cyclic reaction consisting of photo-oxidation of water at 300 nm that yields H2O2 with subsequentdecomposition of H2O2 by catalase.
19
The decline in Arctic sea ice affects local climates, particu-larly in North America, Europe, and East Asia with largeimpacts on biogeochemical cycles. For example, changes inrates of primary production and respiration in terrestrial andaquatic ecosystems may occur as a consequence of droughtsand enhanced runoff of terrestrially derived organic carbon,respectively (see below). These effects on local climates can besummarised as follows: the melting of Arctic sea ice results inincreased air temperatures due to evaporation and conden-sation of surface water, changes in the turbulent heat fluxesand, as a consequence, in increased geopotential height overthe Arctic.20 As a result, the jet stream is transformed intoRossby waves that are characterised by cold air moving to thenorth and warm air moving to the south.21,22 The higher theamplitude of Rossby waves, the slower they move from west toeast, resulting in more frequent episodes of blocking ofweather patterns and therefore in longer lasting extremeweather events such as droughts, flooding, cold spells, andheat.21–25
Further impacts of land and sea ice melting are sea levelrise,13 increases in freshwater input and hence changes inocean circulation, reduced surface albedo26 (see Bais et al.3),and enhanced exposure of organisms in aquatic ecosystemsand on land surfaces such as tundra to solar UV radiation (thisassessment and Häder et al.16). Over the period 1901–2010,global mean sea level rose by 0.19 (0.17 to 0.21) m and theIPCC WGI Report 2013 (summary for policymakers)13 states
Fig. 2 Arctic sea ice loss from 1979 to 2012 as derived from satellitepassive microwave data. The yellow lines (after 2002) are from AMSR-Edata. (Panel (b) of Fig. 4.4 in Chapter 4 “Observations: Cryosphere” ofthe IPCC Report 2013, Working Group I: Climate Change 2013: thePhysical Science Basis).13
Perspective Photochemical & Photobiological Sciences
128 | Photochem. Photobiol. Sci., 2015, 14, 127–148 This journal is © The Royal Society of Chemistry and Owner Societies 2015

with high confidence that the “rate of sea level rise since themid-19th century has been larger than the mean rate duringthe previous two millennia”.
Interacting effects of stratospheric ozone depletion andclimate change on ecosystems
The interplay between stratospheric ozone depletion andclimate change affects atmospheric circulation, the speed anddirection of winds, and, as a consequence, ocean mixing.There is a global trend of increasing values of surface andhigher level wind speed, linked, in part, to stratospheric ozonedepletion.27–29 Causes of this phenomenon have been reportedmore than ten years ago in a key paper by Gillet and Thomp-son (2003).30 Based on both observations and modelingresults, these investigators found falling geopotential heightsat constant pressure (500 hPa) at high southern latitudes from1979–2000 due to Antarctic stratospheric ozone depletion andthus decreasing temperatures. Concomitantly, 500 hPa heightswere rising in the middle southern latitudes, due to increasinggreenhouse gas concentrations and thus increasing tempera-tures. This interplay between Antarctic stratospheric ozonedepletion and climate change results in an upward trend ofthe Southern Annular Mode (SAM).28,31,32 Positive SAM indicesresult in mean high values of the westerly polar vortexes and apoleward shift of the westerly wind belts at the Earth’ssurface.27,33 As a consequence, net uptake of carbon dioxide inthe Southern Ocean, believed to be a major sink for atmo-spheric CO2,
34,35 may be reduced by enhanced wind-drivenupwelling of carbon-rich deepwater.36,37 Paleo-oceanographicstudies suggest that a Southern Ocean CO2 ventilation eventmight have caused glacial-interglacial changes in the atmo-spheric CO2 concentration.38–41 Changes in atmospheric andocean circulation due to the combined effects of Antarcticstratospheric ozone depletion and climate change alterSouthern Hemisphere weather. These alterations potentiallyinclude increased incidence of extreme events, floods,droughts, and wildfires.42
Ocean warming, stratification, acidification, anddeoxygenation
Global warming results in increased sea-surface temperatures(SST) and, in turn, in thermal stratification. Another effect ofincreasing SST is ocean deoxygenation as a consequence ofreduced oxygen solubility. Furthermore, increasing CO2 con-centrations result in ocean acidification.34 Ocean acidificationlowers the saturation state of calcite and aragonite (Fig. 3)and thus the ability of calcifiers such as coccolithophores andcorals to produce and maintain their shells of calcite andaragonite.34,43
Framework building corals are also sensitive to oceanwarming because of the breakdown of the symbiosis with sym-biotic algae (zooxanthellae), resulting in coral bleaching.34,44–47
Algae that no longer live in symbiosis with corals are less pro-tected from solar UV-B radiation. Furthermore, iron stress ofzooxanthellae has been shown to reduce their photosyntheticefficiency and to alter the pigment composition at elevated
temperatures.48 The combined effects of increasing SST, oceanacidification and stratification, and solar UV radiation onmarine ecosystems are likely to reduce the rate of primary pro-duction by phytoplankton (see below). Based on a Biogeo-chemical Elemental Cycling model, Laufkoetter et al. (2013)49
reported a decrease in global net primary production of phyto-plankton with increasing SST from 1960–2006. An excellentreview article on these topics has been published by Gruber(2011).34
Changes in hydroclimatic conditions and continental runoff
The interplay between global warming and stratospheric ozonedepletion affects precipitation patterns in space and time andenhances the intensity of precipitation extremes.33,50–53
Cooling of the Antarctic stratosphere due to ozone depletioncauses circulation cells to shift polewards, resulting in wetterconditions in the southern subtropics in the austral summerand drier conditions around 40° S.33,51 On the other hand,strong El Niño/Southern Oscillation (ENSO) events, potentiallydue to global warming, cause the South Pacific convergencezone to shift towards the Equator with concomitant changes inhydroclimatic conditions resulting in more frequent weatherevents such as droughts, floods, and tropical cyclones.54 In theNorthern Hemisphere, the intensity of heavy precipitationevents has increased over much of the land area due towarming.52,55,56 Changes in hydroclimatic conditions mayresult in an enhanced input of terrestrial organic matter intoaquatic systems57–60 with consequences for the rate of UV-induced mineralisation of organic carbon (see below).
Changes in continental runoff of CDOM have consequencesfor the transmission of solar UV radiation into water bodiesand thus for the protection of phytoplankton against thedamaging UV-B radiation (see below). Remote sensing studieshave provided new information on the extent of CDOM runoffand its impact on the penetration depth of UV radiation in thebiologically-rich waters and coral reefs of South Florida (Fig. 4;Barnes et al. 201461).
Fig. 3 Global map showing oceanic regions of particular vulnerabilityto climate-change effects (see legend above). Reprinted from Gruber,201134 with permission from The Royal Society: Philosophical Trans-actions A, copyright 2011.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 127–148 | 129

Vegetation shifts, droughts, fires, and tipping points
Climate related changes in the spatial distribution andregional intensities of precipitation and temperaturesextremes, among other variables, drive vegetation shifts,droughts and the occurrence and extent of fires. This resultsin changes in the biogeochemical cycles that are initiated byUV interactions with the terrestrial biosphere and fire-scarredcontinental surfaces. Solar UV radiation also impacts the oxi-dative state of the atmosphere, in both the gas and aerosolphases, further influencing greenhouse gases concentrationsand budgets.62 This section seeks to provide heuristic linkagesbetween climate related drivers such as wild fires, droughtsand how UV fluxes, and changes and trends in UV fluxes, inter-act may push terrestrial ecosystems up to and over tippingpoints in ecosystem health.
Intensification of agriculture and the evolution of naturallandscapes and ecosystems to managed ecosystems haveincreased rapidly over the past 50 years.11,63,64 Changes inplant species also occur in regions that are undergoing largeclimate-related ecosystem shifts.65 Climate-related changes insome of the Earth’s largest terrestrial ecosystems are beingdetected.66,67 Droughts, for example, are reducing terrestrialprimary production and are increasing in many regions of theEarth.68–71 UV radiation interacts extensively with the biogeo-chemical cycles that are shifting with changing land surfacecharacteristics. The interactions of vegetation with the chan-ging UV fluxes result in altered trace gas emission to theatmosphere including volatile organic carbon compounds(VOCs) (see below and Bornman et al.72). These UV-inducedtrace gas emissions include both direct emission of VOCs fromplants, thought to be weakly sensitive to changes in UV radi-ation, and UV interactions with burned and/or modified litter.It is, however, difficult to separate changes in ecosystems dueto ecosystem maturation and those due to climate change.73
As ozone, aerosols and trace gases change and evolve inthe atmosphere, the UV radiation at the surface changes, as
well as the characteristics of the surface itself.74 UV radiation-mediated release of trace gases and aerosols from the terres-trial biosphere then changes as a result of the altered make-upof the land surface. Fires have emerged as an important com-ponent of such land surface change.75,76 Not only do firesinput massive amounts of chemicals and aerosols to theatmosphere, altering atmospheric UV radiation and chemistry,but they also leave charred substrate behind. Charcoal canpersist for long periods in terrestrial systems. As a result,increasing amounts of black carbon, an important constituentof charred substrate, are entering rivers and flowing intolakes and the ocean where photochemical transformationsare important sinks.77–80 The intent of this section is to high-light and draw attention to the often ignored interactionsbetween UV radiation, atmospheric chemistry and surfaceland characteristics that are dependent on the extant eco-systems. The way in which these interacting systems arechanging at the same time, often at different rates, is alsoaddressed.
Terrestrial biosphere tipping points occur due to thecomplex interactions between UV radiation, drought-inducedaerosol distributions, anthropogenic pollution and columnozone depletion over specific areas.7,81 UV radiation changescould represent the final source of variability that pushes/pullsthe terrestrial biosphere over tipping points with large impactson global biogeochemical cycles. Even with the generalsuccess of the Montreal protocol, there are still large areaswith high UV radiation reaching the Earth’s surface due tocolumn ozone depletion.7 Tipping points in biogeochemicalcycles may be traversed by strong gradients in surface UV radi-ation. Biogeochemical cycles are occurring over large regionsof similar ecosystem make-up and experience large differencesin UV flux due to atmospheric chemistry and physical climatevariables such as clouds. As in the purely physical climatesystem, extreme events often have a greater impact on biogeo-chemical cycles than a change in the mean state.82,83 There aredirect feedbacks where the processes of afforestation lead tochanges in the general circulation of the atmosphere resultingin changes in precipitation.84 Extreme events, physical climatesystem shifts, fires and droughts all may contribute to UVinitiation of tipping points in both terrestrial and aquatic bio-geochemical cycles. For example, extensive increases inmelting snow and ice cover lead to increases in the exposure ofaquatic ecosystems to UV and PAR,18 which have the potentialto create tipping points – shifts in photosynthetic vs. hetero-trophic organisms where community as well as ecosystemstructure and function are fundamentally altered.85
As ecosystems change due to climate change, fire anddroughts, the chemicals released (i.e. hydrocarbon, CO2, CH4,aerosols) alter atmospheric chemistry. These biogeochemicalchanges have, in turn, implications for the residence times ofgreenhouse gases that are modulated by UV photochemistry.These interactions and biogeochemical cycling aspects of theEarth system often fall outside of traditional boundaries ofscientific specialization and are only now becoming more fullyappreciated.
Fig. 4 Image showing UV attenuation caused by runoff of colored dis-solved organic matter in coastal regions of South Florida. Results werederived from satellite measurements (MODIS/A) on 9 January 2009using a model described by Barnes et al.61 Land is masked in black.White indicates coastline, clouds or algorithm failure. Figure modifiedfrom Barnes et al.61.
Perspective Photochemical & Photobiological Sciences
130 | Photochem. Photobiol. Sci., 2015, 14, 127–148 This journal is © The Royal Society of Chemistry and Owner Societies 2015

Interactive effects of solar UV radiationand climate change on the carboncycleCarbon exchange between ocean and atmosphere
Air-sea CO2 fluxes vary locally and seasonally and depend onthe partial pressure of CO2 and, in turn, on ocean temperature,mixing, and biology.86,87 The main biological process that con-trols the uptake of CO2 by the ocean is the so-called biologicalpump (CO2 binding in photosynthesis by phytoplankton andexport of dead particulate organic matter to the ocean sedi-ment) (Fig. 5).
Rates of primary production depend on various factorsincluding light and nutrient availability, temperature, and thespecies composition of phytoplankton. Also phytoplanktonperformance is important for efficient CO2 uptake and may benegatively affected by solar UV-B radiation.88,89 Evidence hascontinued to mount that the concentration of CDOM controlsthe penetration of UV and short-wavelength visible radiationinto lakes and the coastal ocean.61,88–91 CDOM effectively pro-tects aquatic ecosystems from harmful UV-B radiation whilepermitting PAR to be transmitted into the water column. Thedegradation of CDOM by photochemical and biological pro-cesses leads to a loss of colour and reduced UV absorbance,both in the UV-B and UV-A regions.92–98 As a consequence ofCDOM photobleaching, the penetration depths of solar UVradiation into water bodies increase, thereby increasing theexposure of phytoplankton to damaging solar UV-B radiation(see also Häder et al.16). CDOM photobleaching is particularlyevident in thermally stratified waters bodies.95,98,99 Stratifica-tion results in a shallower mixing depth, which in additionleads to a greater exposure of phytoplankton to UV-B radi-ation.100 Enhanced stratification, due to increasing sea surface
temperatures, occurs mainly in low latitude marine environ-ments where it also hinders the transport of nutrients to theeuphotic zone from deeper water layers and hence negativelyaffects primary production.49,101 Stratification has also beenobserved in ice-free Arctic marine regions due to increasedmeltwater input.102 Cai et al.102 predict that an ice-free ArcticOcean will not become a larger CO2 sink due, in part, to aninefficient biological pump as a consequence of negativeeffects of solar UV radiation on phytoplankton.
CDOM in aquatic systems originates from several sources.In the coastal ocean, continental runoff of CDOM plays animportant role (see also above).90,91,96,103–105 In the open oceanand large lakes, CDOM is a by-product of biological degra-dation of dead phytoplankton.90,97,106 Thus, a reduced concen-tration of phytoplankton will drive decreased CDOMproduction, further increasing transmission of UV radiationinto the ocean. The observed reductions in chlorophyll concen-tration that have been attributed to increasing sea surfacetemperatures101 are probably caused by a combination ofincreased exposure to UV radiation and reduced nutrientupwelling.107
An important part of the biological pump is the export pro-duction, i.e. the formation and sedimentation of particulateorganic matter (POM) stemming from dead phytoplanktonand zooplankton material. The export production hasdecreased by 8% from 1960–2006 globally with strong spatialvariability, based on model calculations.49 One reason for thisdecline may be a global decrease in the biomass of small phy-toplankton and diatoms in the period 1960–2006 (8.5% and3%, respectively49). Ocean acidification may further reduce thebiomass of coccolithophores, small calcifying phytoplankton,and hence the production of CaCO3 “ballast” that enhancessinking rates of carbonate-rich POM.34,49,108,109 Overall, theinteraction of solar UV radiation with climate-change effectssuch as ocean stratification is likely to decrease the efficiencyof the biological pump and to cause UV-mediated, positivefeedbacks on atmospheric CO2 (see also below).
Carbon exchange between terrestrial ecosystems andatmosphere
The uptake of CO2 by terrestrial ecosystems via photosynthesisof plants and the release of CO2 to the atmosphere viadecomposition of senescent plant material (litter) are affectedboth positively and negatively by solar UV radiation andclimate change.89,110 Climate change, and in particular theintensity and frequency of droughts in terrestrial ecosystemscould negatively impact primary production by plants andcrops and thus CO2 uptake by terrestrial ecosystems.13,68,71 Atthe same time, changes in climate that decrease cloudinessand increase UV radiation at the terrestrial surface mayincrease losses of carbon from terrestrial ecosystems due toincreased photochemical degradation of plant material.111
A part of the CO2 fixed by plants may be sequestered inforests or long-lived plant soil components such as woodytissue or peat in soil organic matter, thus increasing its resi-dence time in organic pools. Plant litter on the soil surface
Fig. 5 Schematic representation of the biological pump, indicated bythe green arrows. The efficiency of the biological pump depends on therates of both photosynthesis (primary production) with photosyntheticactive radiation (PAR) and the formation and sedimentation of particu-late organic matter (POM). Numbers in red are from Houghton251 andcorrespond to approximate global mean fluxes in Pg C year−1.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 127–148 | 131

is, however, subject to rapid degradation, which results inemissions of CO2 and other greenhouse gases to theatmosphere.89,112–114 The release of CO2 from terrestrial eco-systems by microbial degradation of soil organic matter isstrongly affected by temperature and moisture.115–117 Inaddition, there is increasing evidence of abiotic decom-position of above ground litter due to UV radiation, particu-larly in arid and semiarid regions with low soil bioticactivity.111,112,114,116–124 Rates of plant litter photodegradationdue to exposure to UV radiation depend on various factors,including: (i) plant cover,111 (Fig. 6), (ii) the intensity of solarUV radiation (altitude, latitude), (iii) exposure of litter mass tosolar radiation due to differences in litter area to mass ratio,and (iv) chemical composition of plant litter118,122 (Fig. 7) (seealso Bornman et al.72).
Recent information regarding the role of litter chemistry indetermining the rate of UV-induced litter degradation hasidentified the importance of lignin in the process of photo-degradation. Several studies have shown a decrease in litterlignin content with exposure to solar or UV-B radiation125,126
and that lignin content of litter is positively correlated withhigher rates of mass loss (Fig. 7, Austin and Ballaré118). Thelatter can be explained in terms of a higher percentage of aro-matic components in lignin and thus a higher rate of lightabsorption in the UV and visible range.118,120,121,127 Althoughabiotic photodegradation increases with the lignin content,the rate of microbial degradation tends to decrease withincreasing lignin content, due to the bio-recalcitrant nature oflignin118 (Fig. 7). Finally, there is some evidence that exposureof plant litter to solar UV-radiation can increase the bioavail-ability of some compounds to soil microorganisms,112,119–121
although the mechanistic basis of this stimulation has notbeen established.
The direct and indirect effects of UV radiation on decompo-sition of plant litter is likely to become a much more signifi-cant global pathway for terrestrial organic matterdecomposition in the future. Aridification due to land useand/or climate change could amplify photodegradative lossesfrom senescent plant litter, with large potential impacts on thecarbon balance of terrestrial ecosystems. Particular attentionneeds to be paid to Arctic terrestrial ecosystems. The Arcticregion is experiencing rapid warming and its land area, includ-ing the tundra, has large stores of carbon that are likely to berapidly converted to GHGs with warming. For example, lossesfrom plant litter may be enhanced due to the thawing of per-mafrost119 and Arctic tundra wildfires,128 (see also above). Allof these global changes have potential to affect plant cover(Fig. 6, Austin111), and hence increase the importance ofphotodegradation in these modified ecosystems.
The terrestrial-aquatic interface
Terrestrial and aquatic ecosystems are largely linked via thebiogeochemical cycling of carbon. Runoff and leaching ofplant litter is a major source of CDOM in aquatic environ-ments, especially in freshwaters. In coastal marine environ-ments, a large fraction of organic carbon originates fromterrestrial ecosystems that is transported to coastal oceans(i) via rivers,59,129–133 and (ii) fluxes from wetlands, particularlyfrom mangrove swamps134 (Fig. 4). Terrestrial organic matter(tDOM), in particular its CDOM fraction, more strongly attenu-ates solar UV radiation than organic matter produced withinlakes and the sea due to its relatively high aromatic
Fig. 6 Hypothesized effect of plant cover on photodegradation in ter-restrial ecosystems. Plant cover is a key factor which will affect thedegree of exposure to UV radiation and impacts of photodegradation asa vector of carbon loss in arid and semiarid ecosystems. As precipitationincreases, primary production and resulting plant cover increase,thereby reducing the relative importance of photodegradation in mesicand humid ecosystems. Global changes such as shrub encroachment,increased fire frequency, invasion of non-native annual species andafforestation will all affect plant cover and consequently the relativeimpact of photodegradation on decomposition in these ecosystems.Red line shows the importance of photodegradation as rainfall increases,the dashed line is changes in plant cover. Green arrows indicate thedirection of change in plant cover with global change. Figure modifiedfrom Austin (2011).111
Fig. 7 Effects of lignin concentration on biotic decomposition andphotodegradation. While biotic degradation of plant litter is inhibited byhigh lignin concentrations, lignin serves as a light absorbing compoundfacilitating photodegradation and carbon release from decomposingplant litter. Green circles indicate material of varying lignin concen-trations which was decomposed in shaded conditions in contact withsoil (biotic decomposition) while red diamonds indicate identicalmaterial of varying lignin concentrations that was isolated from the soilsurface and exposed to full solar radiation. Figure modified from Austinand Ballaré (2010).118
Perspective Photochemical & Photobiological Sciences
132 | Photochem. Photobiol. Sci., 2015, 14, 127–148 This journal is © The Royal Society of Chemistry and Owner Societies 2015
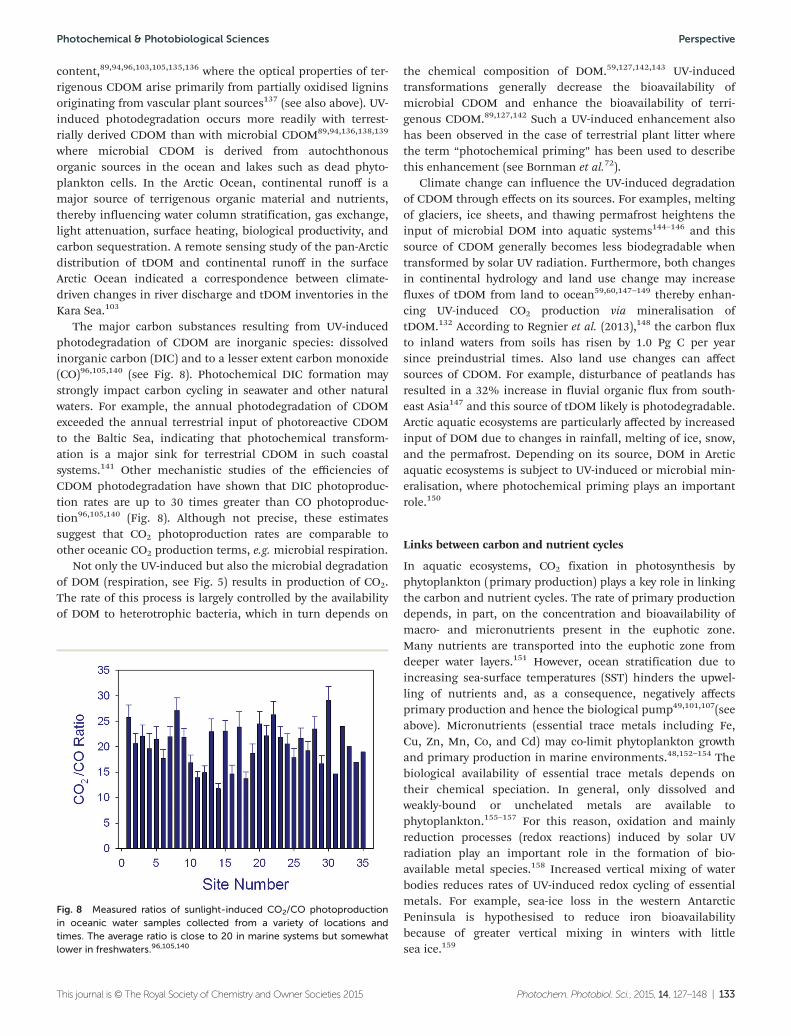
content,89,94,96,103,105,135,136 where the optical properties of ter-rigenous CDOM arise primarily from partially oxidised ligninsoriginating from vascular plant sources137 (see also above). UV-induced photodegradation occurs more readily with terrest-rially derived CDOM than with microbial CDOM89,94,136,138,139
where microbial CDOM is derived from autochthonousorganic sources in the ocean and lakes such as dead phyto-plankton cells. In the Arctic Ocean, continental runoff is amajor source of terrigenous organic material and nutrients,thereby influencing water column stratification, gas exchange,light attenuation, surface heating, biological productivity, andcarbon sequestration. A remote sensing study of the pan-Arcticdistribution of tDOM and continental runoff in the surfaceArctic Ocean indicated a correspondence between climate-driven changes in river discharge and tDOM inventories in theKara Sea.103
The major carbon substances resulting from UV-inducedphotodegradation of CDOM are inorganic species: dissolvedinorganic carbon (DIC) and to a lesser extent carbon monoxide(CO)96,105,140 (see Fig. 8). Photochemical DIC formation maystrongly impact carbon cycling in seawater and other naturalwaters. For example, the annual photodegradation of CDOMexceeded the annual terrestrial input of photoreactive CDOMto the Baltic Sea, indicating that photochemical transform-ation is a major sink for terrestrial CDOM in such coastalsystems.141 Other mechanistic studies of the efficiencies ofCDOM photodegradation have shown that DIC photoproduc-tion rates are up to 30 times greater than CO photoproduc-tion96,105,140 (Fig. 8). Although not precise, these estimatessuggest that CO2 photoproduction rates are comparable toother oceanic CO2 production terms, e.g. microbial respiration.
Not only the UV-induced but also the microbial degradationof DOM (respiration, see Fig. 5) results in production of CO2.The rate of this process is largely controlled by the availabilityof DOM to heterotrophic bacteria, which in turn depends on
the chemical composition of DOM.59,127,142,143 UV-inducedtransformations generally decrease the bioavailability ofmicrobial CDOM and enhance the bioavailability of terri-genous CDOM.89,127,142 Such a UV-induced enhancement alsohas been observed in the case of terrestrial plant litter wherethe term “photochemical priming” has been used to describethis enhancement (see Bornman et al.72).
Climate change can influence the UV-induced degradationof CDOM through effects on its sources. For examples, meltingof glaciers, ice sheets, and thawing permafrost heightens theinput of microbial DOM into aquatic systems144–146 and thissource of CDOM generally becomes less biodegradable whentransformed by solar UV radiation. Furthermore, both changesin continental hydrology and land use change may increasefluxes of tDOM from land to ocean59,60,147–149 thereby enhan-cing UV-induced CO2 production via mineralisation oftDOM.132 According to Regnier et al. (2013),148 the carbon fluxto inland waters from soils has risen by 1.0 Pg C per yearsince preindustrial times. Also land use changes can affectsources of CDOM. For example, disturbance of peatlands hasresulted in a 32% increase in fluvial organic flux from south-east Asia147 and this source of tDOM likely is photodegradable.Arctic aquatic ecosystems are particularly affected by increasedinput of DOM due to changes in rainfall, melting of ice, snow,and the permafrost. Depending on its source, DOM in Arcticaquatic ecosystems is subject to UV-induced or microbial min-eralisation, where photochemical priming plays an importantrole.150
Links between carbon and nutrient cycles
In aquatic ecosystems, CO2 fixation in photosynthesis byphytoplankton (primary production) plays a key role in linkingthe carbon and nutrient cycles. The rate of primary productiondepends, in part, on the concentration and bioavailability ofmacro- and micronutrients present in the euphotic zone.Many nutrients are transported into the euphotic zone fromdeeper water layers.151 However, ocean stratification due toincreasing sea-surface temperatures (SST) hinders the upwel-ling of nutrients and, as a consequence, negatively affectsprimary production and hence the biological pump49,101,107(seeabove). Micronutrients (essential trace metals including Fe,Cu, Zn, Mn, Co, and Cd) may co-limit phytoplankton growthand primary production in marine environments.48,152–154 Thebiological availability of essential trace metals depends ontheir chemical speciation. In general, only dissolved andweakly-bound or unchelated metals are available tophytoplankton.155–157 For this reason, oxidation and mainlyreduction processes (redox reactions) induced by solar UVradiation play an important role in the formation of bio-available metal species.158 Increased vertical mixing of waterbodies reduces rates of UV-induced redox cycling of essentialmetals. For example, sea-ice loss in the western AntarcticPeninsula is hypothesised to reduce iron bioavailabilitybecause of greater vertical mixing in winters with littlesea ice.159
Fig. 8 Measured ratios of sunlight-induced CO2/CO photoproductionin oceanic water samples collected from a variety of locations andtimes. The average ratio is close to 20 in marine systems but somewhatlower in freshwaters.96,105,140
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 127–148 | 133

Iron plays a key role as a micronutrient and co-limits phyto-plankton growth particularly in the so-called high-nutrient-lowchlorophyll (HNLC) oceanic regions.152,153 In oxygenatedmarine environments, Fe(III) is largely present in the form ofsolid Fe(III) phases, with low solubility. In the Southern Ocean,for example, dissolved iron concentrations range between 0.4and 1.5 nmol L−1.160 However, these solid iron phases canundergo UV-induced dissolution yielding dissolved ironspecies161,162 and this photochemical process has been shownto occur at pH-values of marine waters.161 Ocean acidification(see above) can result in reduced availability of essential tracemetals such as Fe, Zn, and Cd to phytoplankton, either viachanges in the chemical speciation of metals or by decreasingthe effectiveness of the enzymatic reduction of metals.157,163
For example, the uptake rate of iron bound to the siderophoredesferrioxamine B by a model diatom was by a factor of ∼2lower in seawater samples with pH 7.8 as compared tosamples with pH 8.7.163 Ocean acidification also slows downnitrogen uptake by phytoplankton under low iron conditionsby lowering the efficiency of N2 fixation.
164
Combined effects of solar UV radiationand climate change on thebiogeochemistry of trace gases andaerosols
Terrestrial and aquatic environments are important atmos-pheric sources and sinks of trace gases and aerosols. Recentresearch on the air–sea exchange of trace gases has providedfurther evidence that these sources are strongly influenced byinteractions between solar UV radiation and climate change.
Methane
Methane is an important greenhouse gas that is stored invarious reservoirs in the terrestrial, aquatic and atmosphericsystems. UV radiation is involved in the release and exchangeof CH4 between these reservoirs as well as atmospheric chemi-cal processes in the troposphere and stratosphere.165–167
Temperature, water vapour, stratospheric ozone, biomassburning, and lightning NOx are the dominant sources of inter-annual changes in methane lifetime.166 UV radiation plays acritical role in determining the effectiveness of each of thesesources in inter-annual changes in methane lifetime which is∼9 year.168 The linkages and inferences of this section revolvearound the UV-mediated atmospheric chemistry, ozone, OHetc. that drive atmospheric methane concentrations, and howCH4 release and uptake from land, aquatic and atmosphericreservoirs is influenced by UV radiation and biogeochemicalcycles in general.
New satellite and airborne data assimilation techniquesallow high-resolution, global estimates of the sources andsinks of atmospheric methane.169 These emerging techno-logies allow quantification and evaluation of the wild-fire pro-duction of atmospheric methane, an increasing source due to
the climate related increased occurrence of wildfires.170,171 UVradiation plays a critical role in the atmospheric chemistry ofmethane as well as modulating its production in soils andother components of the terrestrial biosphere,172,173 althoughMorsky et al.174 found its role in methane production in a sub-arctic fen was more modest. The release of methane to theatmosphere from land surfaces is dependent upon climatechange in the terrestrial biosphere as well as heat and UVsupply to specific methane reservoirs such as permafrostregions.
Methane fluxes from the Arctic Ocean are strong andoccur mainly in areas of open water between sea-ice (leads)and fractional sea-ice up to 82° N.175 Additional sources fromsubmarine permafrost regions have also been reported.176
Such oceanic regions are likely to become increasing sourcesof atmospheric methane in the future due to climate-relatedchanges in sea-ice cover. Warming has also been shown toaffect the emission of methane in Western Siberian lakeswith implications for atmospheric chemistry and UV inter-actions.177 We posit that as climate changes, ocean sea-ice cov-erage changes and often decreases, resulting in new andincreased distributions of air-sea methane flux. The resultantmethane releases will change and will be occurring simul-taneously with the evolving atmospheric UV radiation budget.
Carbon monoxide
Carbon monoxide (CO) is widely produced through inter-actions of solar UV radiation with organic substances inaquatic and terrestrial systems. The average ratio of measuredCO2/CO fluxes from various marine environments is close to20 (Fig. 8). CO participates in chemical reactions that changeair quality and it is one of the most important sinks for atmo-spheric hydroxyl radicals (•OH). Due to its significant effectson •OH, CO also indirectly influences the concentrations ofmethane, halocarbons and other gases in the troposphere (seeabove). Recent research has provided further evidence that UV-induced processes as well as abiotic thermal processes invol-ving tDOM in aquatic systems and plant litter on land are sig-nificant sources of CO to the atmosphere.96,105,122,140,178–182
tDOM generally is a more efficient source of CO (higher appar-ent quantum yields) than microbial CDOM derived frommarine sources such as algal detritus. For example, CO photo-production efficiencies in the Tyne estuary were highest forhigh CDOM riverine samples and almost an order of magni-tude lower for low CDOM coastal seawater samples.180 Oneexception to this general observation is the unusually highefficiency of CO photoproduction from CDOM within floatingSargassum colonies.96 There are also significant CO microbialsinks in aquatic and terrestrial systems and the competitionbetween these sources and sinks often results in diurnal fluc-tuations in the net exchange of CO between biosphere andatmosphere.181–183
An extensive study of the sea to air exchange of CO2 in thesub-arctic estuarine water body, the Canadian St. Lawrenceestuary system, showed that the rates of photoproduction andmicrobial consumption of CO are approximately balanced.182
Perspective Photochemical & Photobiological Sciences
134 | Photochem. Photobiol. Sci., 2015, 14, 127–148 This journal is © The Royal Society of Chemistry and Owner Societies 2015

Other findings are that the photochemical efficiency for COproduction from DOM in aquatic systems decreases withincreasing salinity across the freshwater-marine mixingzone.140,173,179,184
Studies of terrestrial plants have provided additionalevidence that exposure to UV radiation leads to productionof CO from dead plant matter (litter) and that productionrates depend on the litter traits.123 Studies of the effects ofchanging wavelength on CO photoproduction from litter areuncommon but one study showed that UV radiation inducesCO production from Sequoia dendrongiganteum litter. CO emis-sions were strongly reduced in the absence of oxygen as com-pared to air.185 Other studies have shown that living plantleaves also photoproduce CO,186 although the productionefficiency generally is much lower than for senescent or deadvegetation.
Nitrogen compounds
Among nitrogen-containing gases, nitrogen oxides (NOx = NO+ NO2) play an important role in atmospheric chemistry, andmany nitrogen gases have important interactions with UV radi-ation. In the troposphere, NOx enhances the formation of O3
via UV-induced reactions (see Madronich et al.187). In thestratosphere, the opposite is the case, where NOx destroys O3
by the following reaction: NO + O3 → NO2 + O2, where NO2
acts as a catalyst in stratospheric O3 destruction.188 Further-more, the reaction of NO2 with ClO yields ClONO2, a reservoirof reactive chlorine that continues to play an important role inpolar stratospheric ozone depletion in the Arctic and Antarcticspring. Given these important interactions, changes in theatmospheric concentrations of nitrogen oxides due to climatechange or other human activity may indirectly affect ozonerecovery and the amount of UV radiation reaching the Earth’ssurface.
Anthropogenic NOx emission from combustion of fossilfuel, is an important source of NOx to the atmosphere.Additionally, nitric oxide (NO) from natural sources is pro-duced mainly by anaerobic bacteria that reduce nitrate (NO−
3 )to nitrogen gas N2;
189 NO is also a by-product of the oxidationof ammonium (NH4
+) to nitrate by ammonia-oxidising soilbacteria.189,190 Nitrous acid (HONO) emissions from ammonia-oxidising bacteria are comparable with emissions of NO.190,191
Since soil emissions of both NO and HONO depend on thesoil water content,190 increasing soil dryness, due to climatechange, may increase the emission of NO and HONO fromsoils. UV-induced photolysis of HONO is a major source of thehydroxyl radical (HO•) (up to ∼30% of production in the loweratmosphere191). In addition to soil emissions of NOx frombiotic processes, UV-induced photolysis of NO3
− (photodenitri-fication) is an important source of NOx to the atmos-phere.192,193 This abiotic process has been shown recently tooccur on the surface of nitrate-containing snow in theArctic,192 and in sea salt particles containing Cl− and Br−.193
Another key nitrogen-containing gas is nitrous oxide (N2O),an important greenhouse and stratospheric ozone depletinggas. The surface emission of N2O approximately balances the
stratospheric N2O loss188 via formation of the ozone destroyingNO [N2O + O* → 2NO, where O* is produced via photolysis ofozone: O3 + hν → O* + O2]. In terrestrial ecosystems, N2O isformed by denitrifying soil bacteria that reduce NO3
− to N2.189
N2O formation in soils depends on soil water content (SWC)and temperature and occurs at higher SWC than soil emissionof HONO and NO.190,194 Van Groenigen et al.195 predict thatrising levels of CO2 will result in increased emissions of N2Ofrom upland soil due to reduced plant respiration and thushigher SWC. In addition, more frequent precipitation events,due to climate change, that increase SWC could thus enhancethe N2O flux from soils.194
In aquatic ecosystems, ammonia is produced by the UV-induced degradation of DOM184 and N2O by both denitrifyingbacteria and ammonia oxidising archaea.196 N2O formation isparticularly efficient in suboxic and hypoxic oceanic regions(∼10% of the ocean volume).197,198 The area of such regions islikely to expand (Fig. 3) due to the following factors: (i) eutro-phication of coastal waters, (ii) decreasing O2 solubility withwarmer water temperatures, and (iii) shifts in respirationtowards the upper oceanic water column because of reducedsinking velocity of particles in the ocean108,197,199 (see alsoabove). Furthermore, it has been shown that during storms,N2O rich subsurface water is mixed upwards198 suggesting thatmore frequent storm events, due to climate change, may resultin increased emission of N2O from marine environments. Inaddition, sea-ice retreat has been suggested to decrease theresidence time of N2O in the surface water of the Arctic Oceanand thus could enhance the sea-air flux of N2O.
200,201 Geo-engineering via fertilisation of marine regions with iron couldenhance marine N2O production,202 (see also Bais et al.3).
Halogen compounds
Biogenic processes in terrestrial and aquatic ecosystems areimportant sources of halocarbons other than CFC (e.g. themethyl halides CH3Br, CH3Cl, and CH3I) to theatmosphere.203–207 In aquatic ecosystems, phytoplankton isinvolved in the formation of these halocarbons, particularlypico-sized phytoplankton,206 and to a lesser extent bacteriabelonging to Erythrobacter or Pseudomonas.204 The rate of halo-carbon production in aquatic ecosystems also depends on thechemical composition of dissolved organic matter (DOM),with tDOM playing an important role in the formation of halo-carbons such as bromoform (CHBr3) and dibromomethane(CH2Br2).
207 Hence increased fluxes of tDOM from land toocean, due to climate change, may enhance the formation ofhalocarbons, particularly in eutrophic coastal waters. Inaddition to the biotic production of halocarbons, abioticUV-induced processes could also be involved in the formationof CH2ClI, and CH2I2 in seawater.206
In the atmosphere, halocarbons are transformed into reac-tive halogen species (RHS, e.g. the radicals Br• and BrO•),where the hydroxyl radical (HO•, formed in reactionsinduced by solar UV radiation) acts as the oxidant.191,208 Inthe troposphere, RHS react with ozone209,210 and other gases,e.g. gaseous elemental mercury (Hg0)211 (see also below).
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 127–148 | 135

Halocarbons can reach the lowermost stratosphere andundergo UV radiation-induced transformations yielding RHS,particularly BrO•, that participate in stratospheric ozonedepletion.208,212 Models suggest that the loading of the strato-sphere with reactive bromine species will increase in thefuture due to climate change213 and thus the destruction ofozone in the stratosphere by RHS represents a positive feed-back on solar UV radiation. This feedback could be enhancedby climate-change related processes, particularly by increasedinput of tDOM into coastal zones207 and by increases inocean mixing and thus transport of CH3Br to the lowerstratosphere.208
Aerosols
Atmospheric aerosols play an important role in the biogeo-chemistry of greenhouse gases, atmospheric circulation trendsdue to changes in climate and atmospheric chemistry.214 Thesurfaces of atmospheric aerosol particles provide reactivemulti-phase regions for chemical transformations in theatmosphere. Surfaces, as well as the liquid phase portionsinside the aerosols, interact with UV radiation to processchemicals critical to climate and atmospheric chemistry ingeneral. Aerosol distributions and trends in distributions dueto climate change alter UV fluxes to Earth’s surface. This isdue to direct blocking of UV radiation by aerosols in theatmosphere and the fact that the distributions and the radia-tive and chemical make–up of the aerosols are evolving asclimate changes.
Aerosols affect circulation directly via interactions with radi-ation with attendant thermal gradient impacts.215,216 Thismeans that strong feedbacks exist between aerosol distri-butions, UV radiation, and the circulation that influence pre-cipitation and atmospheric transport.217 Changes in thedistribution of atmospheric aerosols are occurring at the sametime as shifts in terrestrial and aquatic and marine eco-systems.218 UV radiation is involved in and affected by biogeo-chemical feedbacks that result from chemical exchanges at theEarth surface, physical circulation changes and human-induced modification of the environment. UV interactionswith the liquid phase of marine aerosol particles result intrace gas production, which alter the oxidation state of theatmosphere. Fe compounds have recently been implicated inthese aerosol multiphase interactions.219 Aerosols not onlyscatter and absorb atmospheric UV radiation they influenceand change atmospheric gas and liquid phase chemistry.
Urbanisation and managed agriculture have resulted inchanging patterns of atmospheric dust distributions and depo-sition.220 These dust aerosols, which affect the transparency ofthe troposphere, have a concurrent impact on UV flux toaquatic and terrestrial ecosystems. Non-Asian sources are a sig-nificant source of dust to the North Pacific221 and will likelyresult in significant changes in the supply of nutrients, asorganically bound Fe and P is transported atmosphericallyfrom continental regions to ocean ecosystems.147,218,222 Infor-mation from studies of the impacts of volcanic supplies of
aerosols are being used to predict the impact of future aerosoldistributions on the carbon cycle.223
There is an emerging realisation that liquid phase UV-driven aerosol chemistry can influence biogeochemical cyclingof atmospheric greenhouse gases.224 This means that not onlydo atmospheric aerosols modulate surface UV fluxes via theirdistribution in the atmospheric column, but also the multi-phase chemical processes that occur on/in aerosols affect theoxidative state of the atmosphere. An example of this multi-phase chemistry is the Fe chemistry in aerosols that influencesatmospheric radicals, oxidative state and lifetimes of green-house gases.219 These newly identified biogeochemical cyclesassist in understanding some of the interactions between UVradiation, aerosol chemistry and atmospheric greenhouse gasdistributions.
Combined effects of solar UV radiationand climate change on chemical andbiological contaminantsOrganics
Global climate change and interactions with solar ultravioletradiation will potentially influence chemical fate and bioaccu-mulation in terrestrial and aquatic environments.225,226
Reviews and modeling studies have particularly focused on theArctic and persistent organic pollutants (POPs)227–229 andpredict the following effects: (1) Increased ambient tempera-tures could lead to increased emissions of chemicalsthrough passive volatilisation from materials and stockpiles;(2) Increased emissions of POPs are predicted as a result ofchanging land-use patterns and changes in global agriculturalpractices, such as pesticide formulations and applicationrates; (3) Changes in energy use will affect use and release ofchemicals; (4) Increases in the frequency of forest fires willincrease the emissions of polycyclic aromatic hydrocarbonsand other combustion by-products such as charcoal and blackcarbon.
Although POPs are chemicals such as chlorinated com-pounds that are resistant to biodegradation and/or to photo-degradation through direct absorption of sunlight, thesechemicals can be transformed by indirect photoreactions thatinvolve natural substances such as tDOM (or related naturalorganic substances that were isolated from soils or naturalwaters), peroxides, nitrate or certain trace metals. Most of therecent examples of indirect photoreactions involve partici-pation of tDOM.228,230–233 Chemicals having a wide variety ofstructural features participate in these indirect photoreactionsincluding chlorinated contaminants,228,233 antibiotics,232 beta-blockers,231 and other drugs.230
One manner through which environmental variables influ-enced by global climate change could affect contaminant toxi-city involves direct effects of the variable on chemicalcharacteristics. The toxicity of polycyclic aromatic hydrocar-bons (PAHs), for example, can be photoactivated by solar UV
Perspective Photochemical & Photobiological Sciences
136 | Photochem. Photobiol. Sci., 2015, 14, 127–148 This journal is © The Royal Society of Chemistry and Owner Societies 2015

radiation.226 The intensity and wavelength distribution of UVradiation, key factors in determining PAH phototoxicity, arelikely to be affected by variables altered by climate change. Forexample, these variables could include decreases in pH thatcan increase water clarity, thereby increasing exposure ofaquatic animals to UV radiation, and increased inputs ofCDOM or particulate carbon to aquatic systems, which wouldeffectively reduce UV penetration. Hence, specific influences ofclimate change on UV intensity in aquatic systems are likely tobe site- and situation-specific.
Although increased UV-B radiation can negatively affect thegrowth and viability of many organisms in aquatic food webs,sensitivity to UV radiation has the beneficial effect of disinfect-ing pathogens.141,234–236 This process is facilitated by climateand UV-induced changes that alter exposure of surface-dwell-ing organisms through increased water transparency and stra-tification, and reduced ice and snow cover. For example,human pathogenic viruses, which are frequently found inrivers, lakes and drinking water, are sensitive to solar UV radi-ation236 and disinfection by solar UV irradiation (SODIS) isbecoming a widely-used tool for purification of drinking waterin developing countries.235 The observed increases in CDOMin aquatic environments can have opposing effects on patho-gen levels. On the one hand, the increases in CDOM reduceUV exposure and thus disinfection by direct photoinactivationof parasites and pathogens. On the other hand, photoinactiva-tion of pathogens can be enhanced through photosensitisationby CDOM.236
Inorganics: mercury
Mercury (Hg) is one of the inorganic priority pollutants and itsbiological availability and thus toxicity is strongly affected bysolar UV radiation and climate change. Mercury is not anessential metal and is toxic at very low concentrations. Mostmercury released to the atmosphere is in the form of gaseouselemental mercury (Hg0), which can be transported over longdistances and is deposited mainly in the oxidised form as di-valent mercury (HgII).237 In the oxidation process, the radicalsBrO• and Br•, that are formed in UV-induced reactions, play akey role.211 In addition, UV-induced re-reduction of HgII
occurs in the troposphere, however, at a lower rate than Hg0
oxidation.237 Following deposition to terrestrial and aquaticecosystems, HgII undergoes methylation yielding methylmer-cury (MeHg), the toxic form of mercury. In sunlit surfacewaters MeHg may undergo UV-induced decomposition, aprocess that is enhanced by DOM.60
Climate change may amplify effects of solar UV radiationon the biogeochemical cycling of mercury. For example, oceanwarming increases the sea-air flux of Hg0.238 As a consequence,the rate of oxidation of Hg0 to HgII (via UV-induced formationof reactive radicals, see above) can be expected to increasesince the oxidation rate depends on the concentration of bothreactive radicals and Hg0. Hence it is likely that the combinedeffects of ocean warming and solar UV radiation on the biogeo-chemical cycling of mercury result in increased formation ofthe toxic form of mercury.
Nanomaterials
In addition to organic contaminants and mercury, new chemi-cals are coming into global markets that require initial assess-ment. Inorganic and organic engineered nanomaterials(ENMs) and their composites with polymers and coatings (seealso Andrady et al.239) are being introduced into the environ-ment through widespread use in consumer products. Con-cerns about their health and safety have stimulated researchon their persistence and toxicity. UV-induced photoreactionsare important environmental processes for ENMs such asnanosilver, nano-titanium dioxide, fullerenes, carbon nano-tubes and graphene oxide. These photoreactions are sensitisedby tDOM in some cases. For example, nanosilver can be readilyformed by the tDOM-induced photoreduction of ionicsilver,240,241 a sunlight-induced process that helps reverse toxi-city associated with oxidation of nanosilver. UV-induced pro-cesses initiate the environmental release of ENMs frompolymer nanocomposites by photodegradation of the polymermatrix.242
Gaps in knowledge
Our understanding of the interacting effects of solar UV radi-ation and climate change on the biogeochemical cycling ofcarbon, nitrogen and halogen compounds, essential and toxicmetals, and on the fate of organic pollutants has increased inthe four years since our last assessment. However, thereremain important gaps in knowledge. For example, climatemodels generally do not include effects of solar UV radiationon biogeochemical cycles. To do this would be important forpredicting trends in the net CO2 sink strength of terrestrialand aquatic ecosystems.
Terrestrial and aquatic ecosystems are currently net CO2
sinks on a global average, where the CO2 uptake correspondsto ca. 30% and 25%, respectively, of anthropogenic CO2
emission.243,244 However, the CO2 sink strength of terrestrialand aquatic ecosystems may decrease due to climatechange49,245,246 and interactions between solar UV radiationand climate change could accelerate this decline. Based on abiogeochemical elemental cycling model (without taking intoaccount effects of solar UV radiation), Laufkoetter et al.49 simu-lated a decrease in phytoplankton net primary production by6.5% within 50 years (1960–2006) on a global average, due tovarious climate-change related factors, including ocean stratifi-cation. A decrease in net primary production by phytoplanktonis likely to reduce the efficiency of the biological pump andthus the CO2 sink strength of aquatic ecosystems. Evenwithout a decrease in the global CO2 sink strength of terrestrialand aquatic ecosystems, their capacity to take up CO2
does not keep pace with increasing atmospheric CO2
concentrations.243,247,248
As discussed earlier, an important effect of solar UV-radi-ation in aquatic ecosystems is photobleaching of CDOM,resulting in increased transmission of the damaging solarUV-B radiation into water bodies with negative effects on
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 127–148 | 137

photosynthetic organisms (organisms such as phytoplanktonthat transform CO2 into organic matter). The enhancement ofUV-induced bleaching of CDOM due to stratification of waterbodies therefore represents a UV-mediated, positive feedbackon climate. In terrestrial ecosystems a positive feedback onclimate is increased UV-induced decomposition and mineralis-ation of above ground litter (under release of CO2) due to aridi-fication and decreased plant cover from human activity.
Important greenhouse gases other than CO2 are nitrousoxide (N2O) and methane (CH4). Climate change, via changingprecipitation and temperature distributions and extremeevents, can have large impacts on the emission of these gases.For example, N2O and CH4 emissions from plants may beincreased through temperature effects, indirectly due to risingCO2 levels. UV-induced inhibition of microbial processes inaquatic systems can also influence trace gas exchange such asCO and N2O to air. Interactions in atmospheric chemistry alsoexist via UV-induced reactions involving N2O resulting in theformation of ozone destroying compounds.
Halocarbons such as methyl bromide (CH3Br), which areemitted by phytoplankton, are other gases that undergo UV-induced transformations to highly reactive species and thusalso have the potential to cause stratospheric ozone depletion.Changes in oceanic phytoplankton distributions and commu-nity structure due to climate change is expected to alter ratesof emission of these trace gases. Highly reactive speciesformed in UV-induced transformations of halocarbons alsoreact with tropospheric pollutants, e.g. gaseous elementalmercury yielding the precursor of the bioavailable and thustoxic form of mercury. While there exists a large body of litera-ture on the effects of climate change or of solar UV radiationon the fate of mercury, the interactive effects of climate changeand solar UV radiation on the biogeochemical cycling ofmercury have, to our knowledge, not yet been extensivelystudied.
Levels of organic pollutants and pathogens in aquaticenvironments are affected by interactions with changingclimate and solar UV radiation. For example, UV radiation caninactivate pathogenic viruses by direct pathways and also byindirect pathways that involve sensitisation by CDOM. CDOM,a climate-sensitive component of most aquatic environments,can also modulate pathogen levels by screening out the UVcomponent of sunlight.
The combined effects of solar UV radiation and climatechange on biogeochemical cycles are likely to be particularlypronounced in the Arctic due to Arctic amplification that alsohinders the recovery of the stratospheric ozone concentrationin Arctic spring. As a consequence of Arctic stratosphericozone depletion, increased levels of solar UV radiation reachArctic terrestrial and aquatic ecosystems. Negative effects ofsolar UV-B radiation on phytoplankton and thus on the bio-logical pump are expected in ice-free Arctic marine regions,particularly in combination with stratification due to increas-ing melt-water input. Based on a radiation transfer model,Fountoulakis et al. (2014)249 predict that up to 10 times higherlevels of solar UV irradiance will enter large parts of the Arctic
Ocean by 2100, compared to the 1950s, mainly due to Arcticsea-ice melting. Furthermore, increased emissions of CO2 viamineralisation of organic matter due to thawing permafrostand Arctic tundra wildfires as well as decreased albedo due tovegetation shifts,11 have large effects on carbon cycling in theArctic150 and together can act as a positive feedback on globalwarming. Therefore, more attention needs to be paid to Arctictipping points.
Acknowledgements
D. E. was supported by the U.S. Global Change ResearchProgram and by the Oak Ridge National Laboratory, which issupported by the Office of Science of the U.S. Department ofEnergy under Contract Number DE-AC05-00OR22725. B. S. wassupported by Eawag: Swiss Federal Institute of Aquatic Scienceand Technology. R. Z. was supported by the National ExposureResearch Laboratory, Ecosystems Research Division, U.S.Environmental Protection Agency, and A. A. was supported bygrants from the National Agency of Science and Technology ofArgentina (ANPCyT).
References
1 R. L. McKenzie, P. J. Aucamp, A. F. Bais, L. O. Björn,M. Ilyas and S. Madronich, Ozone depletion and climatechange: Impacts on UV radiation, Photochem. Photobiol.Sci., 2011, 10, 182–198.
2 Y. Wu, L. M. Polvani and R. Seager, The importance of theMontreal Protocol in protecting Earth’s hydroclimate,J. Climate, 2013, 26, 4049–4068.
3 A. F. Bais, R. L. McKenzie, P. J. Aucamp, M. Ilyas,S. Madronich, G. Bernhard and K. Tourpali, Ozonedepletion and climate change: Impacts on UV radiation,Photochem. Photobiol. Sci., 2015, 14, DOI: 10.1039/c4pp90032d, this issue.
4 J. Austin, H. Struthers, J. Scinocca, D. A. Plummer,H. Akiyoshi, A. J. G. Baumgaertner, S. Bekki,G. E. Bodeker, P. Braesicke, C. Bruehl, N. Butchart,M. P. Chipperfield, D. Cugnet, M. Dameris, S. Dhomse,S. Frith, H. Garny, A. Gettelman, S. C. Hardiman,P. Joeckel, D. Kinnison, A. Kubin, J. F. Lamarque,U. Langematz, E. Mancini, M. Marchand, M. Michou,O. Morgenstern, T. Nakamura, J. E. Nielsen, G. Pitari,J. Pyle, E. Rozanov, T. G. Shepherd, K. Shibata, D. Smale,H. Teyssedre and Y. Yamashita, Chemistry-climate modelsimulations of spring Antarctic ozone, J. Geophys. Res.,[Atmos.], 2010, 115.
5 G. L. Manney, M. L. Santee, M. Rex, N. J. Livesey,M. C. Pitts, P. Veefkind, E. R. Nash, I. Wohltmann,R. Lehmann, L. Froidevaux, L. R. Poole, M. R. Schoeberl,D. P. Haffner, J. Davies, V. Dorokhov, H. Gernandt,B. Johnson, R. Kivi, E. Kyro, N. Larsen, P. F. Levelt,A. Makshtas, C. T. McElroy, H. Nakajima, M. C. Parrondo,
Perspective Photochemical & Photobiological Sciences
138 | Photochem. Photobiol. Sci., 2015, 14, 127–148 This journal is © The Royal Society of Chemistry and Owner Societies 2015

D. W. Tarasick, P. von der Gathen, K. A. Walker andN. S. Zinoviev, Unprecedented Arctic ozone loss in 2011,Nature, 2011, 478, 469–U465.
6 B. M. Sinnhuber, G. Stiller, R. Ruhnke, T. von Clarmann,S. Kellmann and J. Aschmann, Arctic winter 2010/2011at the brink of an ozone hole, Geophys. Res. Lett., 2011,38.
7 G. Bernhard, A. Dahlback, V. Fioletov, A. Heikkila,B. Johnsen, T. Koskela, K. Lakkala and T. Svendby,High levels of ultraviolet radiation observed by ground-based instruments below the 2011 Arctic ozone hole,Atmos. Chem. Phys., 2013, 13, 10573–10590.
8 J. A. Screen and I. Simmonds, The central role of dimin-ishing sea ice in recent Arctic temperature amplification,Nature, 2010, 464, 1334–1337.
9 F. Pithan and T. Mauritsen, Arctic amplification domi-nated by temperature feedbacks in contemporary climatemodels, Nat. Geosci., 2014, 7, 181–184.
10 R. F. Spielhagen, K. Werner, S. A. Sorensen, K. Zamelczyk,E. Kandiano, G. Budeus, K. Husum, T. M. Marchitto andM. Hald, Enhanced modern heat transfer to the arctic bywarm Atlantic water, Science, 2011, 331, 450–453.
11 R. G. Pearson, S. J. Phillips, M. M. Loranty, P. S. A. Beck,T. Damoulas, S. J. Knight and S. J. Goetz, Shifts in Arcticvegetation and associated feedbacks under climatechange, Nat. Clim. Change, 2013, 3, 673–677.
12 T. M. Lenton, Arctic climate tipping points, Ambio, 2012,41, 10–22.
13 IPCC, Climate Change 2013: The Physical Science Basis.Contribution of Working Group I to the Fifth AssessmentReport of the Intergovernmental Panel on ClimateChange, 2013.
14 A. Shepherd, A reconciled estimate of ice-sheet massbalance (vol 57, pg 88, 2010), Science, 2012, 338, 1539–1539.
15 J. E. Walsh, Melting ice what is happening to Arctic SeaIce, and what does it mean for us?, Oceanography, 2013,26, 171–181.
16 D.-P. Häder, C. E. Williamson, S.-Å. Wängberg, M. Rautio,K. C. Rose, K. Gao, E. W. Helbling, R. P. Sinha andR. Worrest, Effects of UV radiation on aquatic ecosystemsand interactions with other environmental factors, Photo-chem. Photobiol. Sci., 2015, 14, DOI: 10.1039/c4pp90035a,this issue.
17 I. Joughin, R. B. Alley and D. M. Holland, Ice-sheetresponse to oceanic forcing, Science, 2012, 338, 1172–1176.
18 K. Frey, D. Perovich and B. Light, The spatial distributionof solar radiation under a melting Arctic sea ice cover,Geophys. Res. Lett., 2011, 38, L22501.
19 C. M. Moreno, Hydrogen peroxide production driven byUV-B in planktonic microorganisms: a photocatalyticfactor in sea warming and ice melting in regions withozone depletion?, Biogeochemistry, 2012, 107, 1–8.
20 J. Sedlacek, R. Knutti, O. Martius and U. Beyerle, Impactof reduced Arctic Sea ice cover on ocean and atmosphericproperties, J. Clim., 2012, 25, 307–319.
21 J. A. Francis, The where and when of wetter and drier: dis-appearing Arctic sea ice plays a role, Environ. Res. Lett.,2013, 8.
22 J. A. Francis and S. J. Vavrus, Evidence linking Arcticamplification to extreme weather in mid-latitudes,Geophys. Res. Lett., 2012, 39.
23 S. Haekkinen, P. B. Rhines and D. L. Worthen, Atmos-pheric blocking and Atlantic multidecadal ocean variabil-ity, Science, 2011, 334, 655–659.
24 J. Liu, J. A. Curry, H. Wang, M. Song and R. M. Horton,Impact of declining Arctic sea ice on winter snowfall(vol 109, pg 4074, 2012), Proc. Natl. Acad. Sci. U. S. A.,2012, 109, 6781–6783.
25 Q. Tang, X. Zhang, X. Yang and J. A. Francis, Cold winterextremes in northern continents linked to Arctic sea iceloss, Environ. Res. Lett., 2013, 8.
26 A. Hu, G. A. Meehl, W. Han, J. Lu and W. G. Strand,Energy balance in a warm world without the ocean con-veyor belt and sea ice, Geophys. Res. Lett., 2013, 40, 6242–6246.
27 S. Lee and S. B. Feldstein, Detecting ozone- and green-house gas-driven wind trends with observational data,Science, 2013, 339, 563–567.
28 H. Paeth and F. Pollinger, Enhanced evidence in climatemodels for changes in extratropical atmospheric circula-tion, Tellus Ser. A, 2010, 62, 647–660.
29 I. R. Young, S. Zieger and A. V. Babanin, Global trendsin wind speed and wave height, Science, 2011, 332, 451–455.
30 N. P. Gillett and D. W. J. Thompson, Simulation of recentSouthern Hemisphere climate change, Science, 2003, 302,273–275.
31 G. J. Marshall, Trends in the southern annular modefrom observations and reanalyses, J. Clim., 2003, 16,4134–4143.
32 G. J. Marshall, S. Di Battista, S. S. Naik and M. Thamban,Analysis of a regional change in the sign of the SAM-temp-erature relationship in Antarctica, Clim. Dyn., 2011, 36,277–287.
33 S. M. Kang, L. M. Polvani, J. C. Fyfe and M. Sigmond,Impact of polar ozone depletion on subtropical precipi-tation, Science, 2011, 332, 951–954.
34 N. Gruber, Warming up, turning sour, losingbreath: ocean biogeochemistry under global change,Philos. Trans. R. Soc. London, Ser. A, 2011, 369, 1980–1996.
35 T. Ito, M. Woloszyn and M. Mazloff, Anthropogeniccarbon dioxide transport in the Southern Ocean driven byEkman flow, Nature, 2010, 463, 80–U85.
36 A. Lenton, F. Codron, L. Bopp, N. Metzl, P. Cadule,A. Tagliabue and J. Le Sommer, Stratospheric ozonedepletion reduces ocean carbon uptake and enhancesocean acidification, Geophys. Res. Lett., 2009, 36.
37 D. W. Waugh, F. Primeau, T. DeVries and M. Holzer,Recent changes in the ventilation of the southern oceans,Science, 2013, 339, 568–570.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 127–148 | 139

38 A. Burke and L. F. Robinson, The southern ocean’s role incarbon exchange during the last deglaciation, Science,2012, 335, 557–561.
39 K. A. Rose, E. L. Sikes, T. P. Guilderson, P. Shane,T. M. Hill, R. Zahn and H. J. Spero, Upper-ocean-to-atmos-phere radiocarbon offsets imply fast deglacial carbondioxide release, Nature, 2010, 466, 1093–1097.
40 D. M. Sigman, M. P. Hain and G. H. Haug, The polarocean and glacial cycles in atmospheric CO2 concen-tration, Nature, 2010, 466, 47–55.
41 T. Tschumi, F. Joos, M. Gehlen and C. Heinze, Deepocean ventilation, carbon isotopes, marine sedimentationand the deglacial CO2 rise, Clim. Past, 2011, 7, 771–800.
42 S. A. Robinson and D. J. Erickson, Not just about sunburn- the ozone hole’s profound effect on climate has signifi-cant implications for Southern Hemisphere ecosystems,Global Change Biol., 2014, DOI: 10.1111/gcb.12739.
43 R. Gangsto, F. Joos and M. Gehlen, Sensitivity of pelagiccalcification to ocean acidification, Biogeosciences, 2011, 8,433–458.
44 N. E. Cantin, A. L. Cohen, K. B. Karnauskas, A. M. Tarrantand D. C. McCorkle, Ocean warming slows coral growth inthe central Red Sea, Science, 2010, 329, 322–325.
45 C. Hauri, N. Gruber, A. M. P. McDonnell and M. Vogt, Theintensity, duration, and severity of low aragonite satur-ation state events on the California continental shelf,Geophys. Res. Lett., 2013, 40, 3424–3428.
46 J. M. Pandolfi, S. R. Connolly, D. J. Marshall andA. L. Cohen, Projecting coral reef futures under globalwarming and ocean acidification, Science, 2011, 333, 418–422.
47 S. Sinutok, R. Hill, M. A. Doblin, R. Wuhrer andP. J. Ralph, Warmer more acidic conditions causedecreased productivity and calcification in subtropicalcoral reef sediment-dwelling calcifiers, Limnol. Oceanogr.,2011, 56, 1200–1212.
48 J. M. Shick, K. Iglic, M. L. Wells, C. G. Trick, J. Doyle andW. C. Dunlap, Responses to iron limitation in two colo-nies of Stylophora pistillata exposed to high temperature:Implications for coral bleaching, Limnol. Oceanogr., 2011,56, 813–828.
49 C. Laufkoetter, M. Vogt and N. Gruber, Long-term trendsin ocean plankton production and particle export between1960–2006, Biogeosciences, 2013, 10, 7373–7393.
50 R. P. Allan, Human influence on rainfall, Nature, 2011,470, 344–341.
51 S. B. Feldstein, Subtropical rainfall and the Antarcticozone hole, Science, 2011, 332, 925–926.
52 S.-K. Min, X. Zhang, F. W. Zwiers and G. C. Hegerl,Human contribution to more-intense precipitationextremes, Nature, 2011, 470, 378–379.
53 P. Pall, T. Aina, D. A. Stone, P. A. Stott, T. Nozawa,A. G. J. Hilberts, D. Lohmann and M. R. Allen, Anthropo-genic greenhouse gas contribution to flood risk inEngland and Wales in autumn 2000, Nature, 2011, 470,382–384.
54 W. Cai, M. Lengaigne, S. Borlace, M. Collins, T. Cowan,M. J. McPhaden, A. Timmermann, S. Power, J. Brown,C. Menkes, A. Ngari, E. M. Vincent and M. J. Widlansky,More extreme swings of the South Pacific convergencezone due to greenhouse warming, Nature, 2012, 488, 365.
55 D. Coumou and S. Rahmstorf, A decade of weatherextremes, Nat. Clim. Change, 2012, 2, 491–496.
56 P. Kopparla, E. M. Fischer, C. Hannay and R. Knutti,Improved simulation of extreme precipitation in a high-resolution atmosphere model, Geophys. Res. Lett., 2013,40, 5803–5808.
57 A. M. Bass, M. I. Bird, M. J. Liddell and P. N. Nelson,Fluvial dynamics of dissolved and particulate organiccarbon during periodic discharge events in a steep tropi-cal rainforest catchment, Limnol. Oceanogr., 2011, 56,2282–2292.
58 S. Haaland, D. Hongve, H. Laudon, G. Riise andR. D. Vogt, Quantifying the drivers of the increasingcolored organic matter in boreal surface waters, Environ.Sci. Technol., 2010, 44, 2975–2980.
59 C. A. Stedmon, R. M. W. Amon, A. J. Rinehart andS. A. Walker, The supply and characteristics of colored dis-solved organic matter (CDOM) in the Arctic Ocean: PanArctic trends and differences, Mar. Chem., 2011, 124, 108–118.
60 J. Zhang, J. Hudson, R. Neal, J. Sereda, T. Clair,M. Turner, D. Jeffries, P. Dillon, L. Molot, K. Somers andR. Hesslein, Long-term patterns of dissolved organiccarbon in lakes across eastern Canada: Evidence of apronounced climate effect, Limnol. Oceanogr., 2010, 55,30–42.
61 B. B. Barnes, C. Hu, J. P. S. E. Cannizzaro, S. E. P. Craig,P. Hallock, D. L. Jones, J. C. N. Lehrter, N. Melo,B. A. Schaeffer and R. Zepp, Estimation of diffuse attenu-ation of ultraviolet light in optically shallow Florida Keyswaters from MODIS measurements, Remote Sens. Environ.,2014, 140, 519–532.
62 S. Bekki, A. Rap, V. Poulain, S. Dhomse, M. Marchand,F. Lefevre, P. M. Forster, S. Szopa and M. P. Chipperfield,Climate impact of stratospheric ozone recovery, Geophys.Res. Lett., 2013, 40, 2796–2800.
63 J. E. Halofsky, M. A. Hemstrom, D. R. Conklin,J. S. Halofsky, B. K. Kerns and D. Bachelet, Assessingpotential climate change effects on vegetation using alinked model approach, Ecol. Modell., 2013, 266, 131–143.
64 T. Hickler, K. Vohland, J. Feehan, P. A. Miller, B. Smith,L. Costa, T. Giesecke, S. Fronzek, T. R. Carter, W. Cramer,I. Kuehn and M. T. Sykes, Projecting the future distri-bution of European potential natural vegetation zoneswith a generalized, tree species-based dynamic vegetationmodel, Global Ecol. Biogeogr., 2012, 21, 50–63.
65 H. W. Polley, V. L. Jin and P. A. Fay, CO2-caused change inplant species composition rivals the shift in vegetationbetween mid-grass and tallgrass prairies, Global ChangeBiol., 2012, 18, 700–710.
Perspective Photochemical & Photobiological Sciences
140 | Photochem. Photobiol. Sci., 2015, 14, 127–148 This journal is © The Royal Society of Chemistry and Owner Societies 2015

66 E. A. Davidson, A. C. de Araujo, P. Artaxo, J. K. Balch,I. F. Brown, M. M. C. Bustamante, M. T. Coe,R. S. DeFries, M. Keller, M. Longo, J. W. Munger,W. Schroeder, B. S. Soares-Filho, C. M. Souza andS. C. Wofsy, The Amazon basin in transition (vol 481, pg321, 2012), Nature, 2012, 483, 232–232.
67 M. Gottfried, H. Pauli, A. Futschik, M. Akhalkatsi,P. Barancok, J. L. Benito Alonso, G. Coldea, J. Dick,B. Erschbamer, M. R. Fernandez Calzado, G. Kazakis,J. Krajci, P. Larsson, M. Mallaun, O. Michelsen,D. Moiseev, P. Moiseev, U. Molau, A. Merzouki, L. Nagy,G. Nakhutsrishvili, B. Pedersen, G. Pelino, M. Puscas,G. Rossi, A. Stanisci, J.-P. Theurillat, M. Tomaselli,L. Villar, P. Vittoz, I. Vogiatzakis and G. Grabherr,Continent-wide response of mountain vegetation toclimate change, Nat. Clim. Change, 2012, 2, 111–115.
68 C. Beer, M. Reichstein, E. Tomelleri, P. Ciais, M. Jung,N. Carvalhais, C. Roedenbeck, M. A. Arain, D. Baldocchi,G. B. Bonan, A. Bondeau, A. Cescatti, G. Lasslop,A. Lindroth, M. Lomas, S. Luyssaert, H. Margolis,K. W. Oleson, O. Roupsard, E. Veenendaal, N. Viovy,C. Williams, F. I. Woodward and D. Papale, TerrestrialGross Carbon Dioxide Uptake: Global Distribution andCovariation with Climate, Science, 2010, 329, 834–838.
69 B. Choat, S. Jansen, T. J. Brodribb, H. Cochard, S. Delzon,R. Bhaskar, S. J. Bucci, T. S. Feild, S. M. Gleason,U. G. Hacke, A. L. Jacobsen, F. Lens, H. Maherali,J. Martinez-Vilalta, S. Mayr, M. Mencuccini, P. J. Mitchell,A. Nardini, J. Pittermann, R. B. Pratt, J. S. Sperry,M. Westoby, I. J. Wright and A. E. Zanne, Global conver-gence in the vulnerability of forests to drought, Nature,2012, 491, 752–755.
70 C. R. Schwalm, C. A. Williams, K. Schaefer, D. Baldocchi,T. A. Black, A. H. Goldstein, B. E. Law, W. C. Oechel,U. Kyaw Tha Paw and R. L. Scott, Reduction in carbonuptake during turn of the century drought in westernNorth America, Nat. Geosci., 2012, 5, 551–556.
71 M. S. Zhao and S. W. Running, Drought-Inducedreduction in global terrestrial net primary productionfrom 2000 through 2009, Science, 2010, 329, 940–943.
72 J. F. Bornman, P. W. Barnes, S. A. Robinson, C. L. Ballaré,S. D. Flinte and M. M. Caldwell, Solar ultraviolet radiationand ozone depletion-driven climate change: effects on ter-restrial ecosystems, Photochem. Photobiol. Sci., 2015, 14,DOI: 10.1039/c4pp90034k, this issue.
73 J. Bodin, V. Badeau, E. Bruno, C. Cluzeau, J.-M. Moisselin,G.-R. Walther and J.-L. Dupouey, Shifts of forest speciesalong an elevational gradient in Southeast France: climatechange or stand maturation?, J. Veg. Sci., 2013, 24, 269–283.
74 J. J. Wargent and B. R. Jordan, From ozone depletion toagriculture: understanding the role of UV radiation in sus-tainable crop production, New Phytol., 2013, 197, 1058–1076.
75 D. S. Ward, S. Kloster, N. M. Mahowald, B. M. Rogers,J. T. Randerson and P. G. Hess, The changing
radiative forcing of fires: global model estimates for past,present and future, Atmos. Chem. Phys., 2012, 12, 10857–10886.
76 D. A. King, D. M. Bachelet and A. J. Symstad, Climatechange and fire effects on a prairie-woodland ecotone:projecting species range shifts with a dynamic global veg-etation model, Ecol. Evol., 2013, 3, 5076–5097.
77 T. Dittmar, C. E. de Rezende, M. Manecki, J. Niggemann,A. R. C. Ovalle, A. Stubbins and M. C. Bernardes, Continu-ous flux of dissolved black carbon from a vanished tropi-cal forest biome, Nat. Geosci., 2012, 5, 618–622.
78 R. Jaffe, Y. Ding, J. Niggemann, A. V. Vahatalo,A. Stubbins, R. G. M. Spencer, J. Campbell and T. Dittmar,Global charcoal mobilization from soils via dissolutionand riverine transport to the oceans, Science, 2013, 340,345–347.
79 C. A. Masiello and P. Louchouarn, Fire in the ocean,Science, 2013, 340, 287–288.
80 A. Stubbins, J. Niggemann and T. Dittmar, Photo-labilityof deep ocean dissolved black carbon, Biogeosciences,2012, 9, 1661–1670.
81 B. W. Brook, E. C. Ellis, M. P. Perring, A. W. Mackay andL. Blomqvist, Does the terrestrial biosphere have planetarytipping points?, Trends Ecol. Evol., 2013, 28, 396–401.
82 M. S. Bret-Harte, M. C. Mack, G. R. Shaver, D. C. Huebner,M. Johnston, C. A. Mojica, C. Pizano and J. A. Reiskind,The response of Arctic vegetation and soils following anunusually severe tundra fire, Philos. Trans. R. Soc. London,B., 2013, 368.
83 F. Lloret, A. Escudero, J. Maria Iriondo, J. Martinez-Vilaltaand F. Valladares, Extreme climatic events and vegetation:the role of stabilizing processes, Global Change Biol., 2012,18, 797–805.
84 A. L. S. Swann, I. Y. Fung and J. C. H. Chiang, Mid-latitudeafforestation shifts general circulation and tropical pre-cipitation, Proc. Natl. Acad. Sci. U. S. A., 2012, 109, 712–716.
85 G. F. Clark, J. S. Stark, E. L. Johnston, J. W. Runcie,P. M. Goldsworthy, B. Raymond and M. J. Riddle, Light-driven tipping points in polar ecosystems, Global ChangeBiol., 2013, 19, 3749–3761.
86 M. Ribas-Ribas, A. Gomez-Parra and J. M. Forja, Air-seaCO2 fluxes in the north-eastern shelf of the Gulf of Cadiz(southwest Iberian Peninsula), Mar. Chem., 2011, 123,56–66.
87 E. H. Shadwick, H. Thomas, K. Azetsu-Scott,B. J. W. Greenan, E. Head and E. Horne, Seasonal variabil-ity of dissolved inorganic carbon and surface water pCO2
in the Scotian Shelf region of the Northwestern Atlantic,Mar. Chem., 2011, 124, 23–37.
88 D.-P. Häder, E. W. Helbling, C. E. Williamson andR. C. Worrest, Effects on aquatic ecosystems and inter-actions with climate change, Photochem. Photobiol. Sci.,2011, 10, 242–260.
89 R. G. Zepp, D. J. Erickson III, N. D. Paul andB. Sulzberger, Effects of solar UV radiation and climate
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 127–148 | 141

change on biogeochemical cycling: interactions andfeedbacks, Photochem. Photobiol. Sci., 2011, 10, 261–279.
90 N. B. Nelson and D. A. Siegel, The global distribution anddynamics of chromophoric dissolved organic matter, inAnnual Review of Marine Science, ed. C. A. Carlson andS. J. Giovannoni, Annual Reviews, Palo Alto, 2013, vol. 5,pp. 447–476.
91 R. G. M. Spencer, G. R. Aiken, M. M. Dornblaser,K. D. Butler, R. M. Holmes, G. Fiske, P. J. Mann andA. Stubbins, Chromophoric dissolved organic matterexport from U.S. rivers, Geophys. Res. Lett., 2013, 40, 1575–1579.
92 A. Bricaud, A. M. Ciotti and B. Gentili, Spatial-temporalvariations in phytoplankton size and colored detritalmatter absorption at global and regional scales, as derivedfrom twelve years of SeaWiFS data (1998–2009), Global Bio-geochem. Cycles, 2012, 26.
93 M. Gonsior, P. Schmitt-Kopplin and D. Bastviken, Depth-dependent molecular composition and photo-reactivity ofdissolved organic matter in a boreal lake under winterand summer conditions, Biogeosciences, 2013, 10, 6945–6956.
94 M. A. Granskog, C. A. Stedmon, P. A. Dodd, R. M. W. Amon,A. K. Pavlov, L. de Steur and E. Hansen, Characteristics ofcolored dissolved organic matter (CDOM) in the Arcticoutflow in the Fram Strait: Assessing the changes and fateof terrigenous CDOM in the Arctic Ocean, J. Geophys. Res.:Oceans, 2012, 117.
95 J. R. Helms, A. Stubbins, E. M. Perdue, N. W. Green,H. Chen and K. Mopper, Photochemical bleaching ofoceanic dissolved organic matter and its effect on absorp-tion spectral slope and fluorescence, Mar. Chem., 2013,155, 81–91.
96 G. C. Shank, R. G. Zepp, A. Vahatalo, R. Lee andE. Bartels, Photobleaching kinetics of chromophoric dis-solved organic matter derived from mangrove leaf litterand floating Sargassum colonies, Mar. Chem., 2010, 119,162–171.
97 C. M. Swan, N. B. Nelson, D. A. Siegel andT. S. Kostadinov, The effect of surface irradiance on theabsorption spectrum of chromophoric dissolved organicmatter in the global ocean, Deep-Sea Res., Part I, 2012, 63,52–64.
98 Y. Yamashita, Y. Nosaka, K. Suzuki, H. Ogawa,K. Takahashi and H. Saito, Photobleaching as a factorcontrolling spectral characteristics of chromophoric dis-solved organic matter in open ocean, Biogeosciences, 2013,10, 7207–7217.
99 L. Bracchini, A. M. Dattilo, V. Hull, S. A. Loiselle,L. Nannicini, M. P. Picchi, M. Ricci, C. Santinelli,A. Seritti, A. Tognazzi and C. Rossi, Spatial and seasonalchanges in optical properties of autochthonous andallochthonous chromophoric dissolved organic matter ina stratified mountain lake, Photochem. Photobiol. Sci.,2010, 9, 304–314.
100 V. E. Villafane, A. T. Banaszak, S. D. Guendulain-Garcia,S. M. Strauch, S. R. Halac and E. W. Helbling, Influence ofseasonal variables associated with climate change onphotochemical diurnal cycles of marine phytoplanktonfrom Patagonia (Argentina), Limnol. Oceanogr., 2013, 58,203–214.
101 D. G. Boyce, M. R. Lewis and B. Worm, Global phytoplank-ton decline over the past century, Nature, 2010, 466,591–596.
102 W. J. Cai, L. Q. Chen, B. S. Chen, Z. Y. Gao, S. H. Lee,J. F. Chen, D. Pierrot, K. Sullivan, Y. C. Wang, X. P. Hu,W. J. Huang, Y. H. Zhang, S. Q. Xu, A. Murata,J. M. Grebmeier, E. P. Jones and H. S. Zhang, Decrease inthe CO2 uptake capacity in an ice-free Arctic Ocean basin,Science, 2010, 329, 556–559.
103 C. G. Fichot, K. Kaiser, S. B. Hooker, R. M. W. Amon,M. Babin, S. Belanger, S. A. Walker and R. Benner, Pan-Arctic distributions of continental runoff in the ArcticOcean, Sci. Rep., 2013, 3.
104 M. A. Granskog, Changes in spectral slopes of coloreddissolved organic matter absorption with mixing andremoval in a terrestrially dominated marine system(Hudson Bay, Canada), Mar. Chem., 2012, 134, 10–17.
105 H. E. Reader and W. L. Miller, Variability of carbon mon-oxide and carbon dioxide apparent quantum yield spectrain three coastal estuaries of the South Atlantic Bight, Bio-geosci. Discuss., 2012, 9, 6927–6985.
106 N. B. Nelson, D. A. Siegel, C. A. Carlson and C. M. Swan,Tracing global biogeochemical cycles and meridionaloverturning circulation using chromophoric dissolvedorganic matter, Geophys. Res. Lett., 2010, 37.
107 P. Falkowski, Ocean Science: The power of plankton,Nature, 2012, 483, S17–S20.
108 M. Hofmann and H.-J. Schellnhuber, Oceanic acidifica-tion affects marine carbon pump and triggers extendedmarine oxygen holes, Proc. Natl. Acad. Sci. U. S. A., 2009,106, 3017–3022.
109 M. Hofmann and H. J. Schellnhuber, Ocean acidification:a millennial challenge, Energy Environ. Sci., 2010, 3, 1883–1896.
110 C. L. Ballaré, M. M. Caldwell, S. D. Flint, A. Robinson andJ. F. Bornman, Effects of solar ultraviolet radiation on ter-restrial ecosystems. Patterns, mechanisms, and inter-actions with climate change, Photochem. Photobiol. Sci.,2011, 10, 226–241.
111 A. T. Austin, Has water limited our imagination for aridland biogeochemistry?, Trends Ecol. Evol., 2011, 26, 229–235.
112 L. A. Brandt, J. Y. King, S. E. Hobbie, D. G. Milchunas andR. L. Sinsabaugh, The Role of Photodegradation inSurface Litter Decomposition Across a Grassland Ecosys-tem Precipitation Gradient, Ecosystems, 2010, 13, 765–781.
113 J. A. Hatala, M. Detto, O. Sonnentag, S. J. Deverel,J. Verfaillie and D. D. Baldocchi, Greenhouse gas (CO2,CH4, H2O) fluxes from drained and flooded agricultural
Perspective Photochemical & Photobiological Sciences
142 | Photochem. Photobiol. Sci., 2015, 14, 127–148 This journal is © The Royal Society of Chemistry and Owner Societies 2015

peatlands in the Sacramento-San Joaquin Delta, Agric.Ecosyst. Environ., 2012, 150, 1–18.
114 S. Rutledge, D. Campbell, D. Baldocchi and L. Schipper,Photodegradation leads to increased carbon dioxidelosses from terrestrial organic matter, Global Change Biol.,2010, 16, 3065–3074.
115 B. Bond-Lamberty and A. Thomson, Temperature-associ-ated increases in the global soil respiration record,Nature, 2010, 464, 579–U132.
116 S. Ma, D. D. Baldocchi, J. A. Hatala, M. Detto and J. CurielYuste, Are rain-induced ecosystem respiration pulsesenhanced by legacies of antecedent photodegradation insemi-arid environments?, Agric. Forest Meteorol., 2012,154, 203–213.
117 W. K. Smith, W. Gao, H. Steltzer, M. D. Wallenstein andR. Tree, Moisture availability influences the effect ofultraviolet-B radiation on leaf litter decomposition, GlobalChange Biol., 2010, 16, 484–495.
118 A. T. Austin and C. L. Ballaré, Dual role of lignin in plantlitter decomposition in terrestrial ecosystems, Proc. Natl.Acad. Sci. U. S. A., 2010, 107, 4618–4622.
119 R. M. Cory, B. C. Crump, J. A. Dobkowski and G. W. Kling,Surface exposure to sunlight stimulates CO2 release frompermafrost soil carbon in the Arctic, Proc. Natl. Acad.Sci. U. S. A., 2013, 110, 3429–3434.
120 X. Feng, K. M. Hills, A. J. Simpson, J. K. Whalen andM. J. Simpson, The role of biodegradation and photo-oxi-dation in the transformation of terrigenous organicmatter, Org. Geochem., 2011, 42, 262–274.
121 J. Frouz, T. Cajthaml and O. Mudrak, The effect of ligninphotodegradation on decomposability of Calamagrostisepigeios grass litter, Biodegradation, 2011, 22, 1247–1254.
122 J. Y. King, L. A. Brandt and E. C. Adair, Shedding light onplant litter decomposition: advances, implications andnew directions in understanding the role of photodegra-dation, Biogeochemistry, 2012, 111, 57–81.
123 H. Lee, T. Rahn and H. L. Throop, An accounting ofC-based trace gas release during abiotic plant litter degra-dation, Global Change Biol., 2012, 18, 1185–1195.
124 L. M. Mayer, K. R. Thornton, L. L. Schick, J. D. Jastrowand J. W. Harden, Photodissolution of soil organicmatter, Geoderma, 2012, 170, 314–321.
125 B. Foereid, J. Bellarby, W. Meier-Augenstein and H. Kemp,Does light exposure make plant litter more degradable?,Plant Soil, 2010, 333, 275–285.
126 H. A. L. Henry, K. Brizgys and C. B. Field, Litter decompo-sition in a California annual grassland: Interactionsbetween photodegradation and litter layer thickness, Eco-systems, 2008, 11, 545–554.
127 B. Sulzberger and E. Durisch-Kaiser, Chemical character-ization of dissolved organic matter (DOM): A prerequisitefor understanding UV-induced changes of DOM absorp-tion properties and bioavailability, Aquat. Sci., 2009, 71,104–126.
128 M. C. Mack, M. S. Bret-Harte, T. N. Hollingsworth,R. R. Jandt, E. A. G. Schuur, G. R. Shaver and
D. L. Verbyla, Carbon loss from an unprecedented Arctictundra wildfire, Nature, 2011, 475, 489–492.
129 M. Fernanda Adame, S. F. Wright, A. Grinham, K. Lobb,C. E. Reymond and C. E. Lovelock, Terrestrial-marine con-nectivity: Patterns of terrestrial soil carbon deposition incoastal sediments determined by analysis of glomalinrelated soil protein, Limnol. Oceanogr., 2012, 57, 1492–1502.
130 C. G. Fichot and R. Benner, The spectral slope coefficientof chromophoric dissolved organic matter (S275–295) as atracer of terrigenous dissolved organic carbon in river-influenced ocean margins, Limnol. Oceanogr., 2012, 57,1453–1466.
131 M. Gonsior, B. M. Peake, W. T. Cooper, D. C. Podgorski,J. D’Andrilli, T. Dittmar and W. J. Cooper, Characteriz-ation of dissolved organic matter across the SubtropicalConvergence off the South Island, New Zealand, Mar.Chem., 2011, 123, 99–110.
132 R. T. Letscher, D. A. Hansell and D. Kadko, Rapidremoval of terrigenous dissolved organic carbon over theEurasian shelves of the Arctic Ocean, Mar. Chem., 2011,123, 78–87.
133 C. J. Mundy, M. Gosselin, M. Starr and C. Michel, Riverineexport and the effects of circulation on dissolved organiccarbon in the Hudson Bay system, Canada, Limnol. Ocea-nogr., 2010, 55, 315–323.
134 B. A. Bergamaschi, D. P. Krabbenhoft, G. R. Aiken,E. Patino, D. G. Rumbold and W. H. Orem, Tidally DrivenExport of Dissolved Organic Carbon, Total Mercury, andMethylmercury from a Mangrove-Dominated Estuary,Environ. Sci. Technol., 2012, 46, 1371–1378.
135 D. Olefeldt, K. J. Devito and M. R. Turetsky, Sources andfate of terrestrial dissolved organic carbon in lakes of aBoreal Plains region recently affected by wildfire, Biogeo-sciences, 2013, 10, 6247–6265.
136 A. Stubbins, R. G. M. Spencer, H. Chen, P. G. Hatcher,K. Mopper, P. J. Hernes, V. L. Mwamba, A. M. Mangangu,J. N. Wabakanghanzi and J. Six, Illuminated darkness:Molecular signatures of Congo River dissolved organicmatter and its photochemical alteration as revealed byultrahigh precision mass spectrometry, Limnol. Oceanogr.,2010, 55, 1467–1477.
137 E. S. Boyle, N. Guerriero, A. Thiallet, R. Del Vecchio andN. V. Blough, Optical properties of humic substances andCDOM: Relation to structure, Environ. Sci. Technol., 2009,43, 2262–2268.
138 L. Jorgensen, C. A. Stedmon, T. Kragh, S. Markager,M. Middelboe and M. Sondergaard, Global trends in thefluorescence characteristics and distribution of marinedissolved organic matter, Mar. Chem., 2011, 126, 139–148.
139 J. F. Rontani, N. Zabeti and S. G. Wakeham, Degradationof particulate organic matter in the equatorial PacificOcean: Biotic or abiotic?, Limnol. Oceanogr., 2011, 56,333–349.
140 E. M. White, D. J. Kieber, J. Sherrard, W. L. Miller andK. Mopper, Carbon dioxide and carbon monoxide photo-
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 127–148 | 143

production quantum yields in the Delaware Estuary, Mar.Chem., 2010, 118, 11–21.
141 H. Aarnos, P. Ylostalo and A. V. Vahatalo, Seasonal photo-transformation of dissolved organic matter toammonium, dissolved inorganic carbon, and labile sub-strates supporting bacterial biomass across the Baltic Sea,J. Geophys. Res., 2012, 117, DOI: 10.1029/2010jg001633.
142 R. M. Cory and L. A. Kaplan, Biological lability of stream-water fluorescent dissolved organic matter, Limnol. Ocea-nogr., 2012, 57, 1347–1360.
143 C. Romera-Castillo, M. Nieto-Cid, C. G. Castro,C. Marrase, J. Largier, E. D. Barton and X. A. Alvarez-Salgado, Fluorescence: Absorption coefficient ratio -Tracing photochemical and microbial degradation pro-cesses affecting coloured dissolved organic matter in acoastal system, Mar. Chem., 2011, 125, 26–38.
144 R. M. DeConto, S. Galeotti, M. Pagani, D. Tracy,K. Schaefer, T. Zhang, D. Pollard and D. J. Beerling, Pastextreme warming events linked to massive carbon releasefrom thawing permafrost (vol 484, pg 87, 2012), Nature,2012, 490, 292–292.
145 J. B. Fellman, E. Hood and R. G. M. Spencer, Fluorescencespectroscopy opens new windows into dissolved organicmatter dynamics in freshwater ecosystems: A review,Limnol. Oceanogr., 2010, 55, 2452–2462.
146 B. G. Pautler, G. C. Woods, A. Dubnick, A. J. Simpson,M. J. Sharp, S. J. Fitzsimons and M. J. Simpson, Molecularcharacterization of dissolved organic matter in glacialice: Coupling natural abundance H-1 NMR and fluo-rescence spectroscopy, Environ. Sci. Technol., 2012, 46,3753–3761.
147 C. M. Moore, M. M. Mills, K. R. Arrigo, I. Berman-Frank,L. Bopp, P. W. Boyd, E. D. Galbraith, R. J. Geider,C. Guieu, S. L. Jaccard, T. D. Jickells, J. La Roche,T. M. Lenton, N. M. Mahowald, E. Maranon, I. Marinov,J. K. Moore, T. Nakatsuka, A. Oschlies, M. A. Saito,T. F. Thingstad, A. Tsuda and O. Ulloa, Processes and pat-terns of oceanic nutrient limitation, Nat. Geosci., 2013, 6,701–710.
148 P. Regnier, P. Friedlingstein, P. Ciais, F. T. Mackenzie,N. Gruber, I. A. Janssens, G. G. Laruelle, R. Lauerwald,S. Luyssaert, A. J. Andersson, S. Arndt, C. Arnosti,A. V. Borges, A. W. Dale, A. Gallego-Sala, Y. Godderis,N. Goossens, J. Hartmann, C. Heinze, T. Ilyina, F. Joos,D. E. LaRowe, J. Leifeld, F. J. R. Meysman, G. Munhoven,P. A. Raymond, R. Spahni, P. Suntharalingam andM. Thullner, Anthropogenic perturbation of the carbonfluxes from land to ocean, Nat. Geosci., 2013, 6, 597–607.
149 I. Valiela, L. Camilli, T. Stone, A. Giblin, J. Crusius, S. Fox,C. Barth-Jensen, R. O. Monteiro, J. Tucker, P. Martinettoand C. Harris, Increased rainfall remarkably freshens estu-arine and coastal waters on the Pacific coast of Panama:Magnitude and likely effects on upwelling and nutrientsupply, Global Planet. Change, 2012, 92–93, 130–137.
150 L. Tranvik, Carbon cycling in the Arctic, Science, 2014,345, 870.
151 K. S. Johnson, S. C. Riser and D. M. Karl, Nitratesupply from deep to near-surface waters of theNorth Pacific subtropical gyre, Nature, 2010, 465, 1062–1065.
152 A. L. King and K. A. Barbeau, Dissolved iron and macro-nutrient distributions in the southern California CurrentSystem, J. Geophys. Res. Oceans, 2011, 116.
153 A. L. King, K. N. Buck and K. A. Barbeau, Quasi-Lagran-gian drifter studies of iron speciation and cycling offPoint Conception, California, Mar. Chem., 2012, 128,1–12.
154 A. Lelong, E. Bucciarelli, H. Hegaret and P. Soudant, Ironand copper limitations differently affect growth rates andphotosynthetic and physiological parameters of themarine diatom Pseudonitzschia delicatissima, Limnol. Ocea-nogr., 2013, 58, 613–623.
155 L. Aristilde, Y. Xu and F. M. M. Morel, Weak organicligands enhance zinc uptake in marine phytoplankton,Environ. Sci. Technol., 2012, 46, 5438–5445.
156 F. M. M. Morel, A. B. Kustka and Y. Shaked, The role ofunchelated Fe in the iron nutrition of phytoplankton,Limnol. Oceanogr., 2008, 53, 400–404.
157 Y. Xu, D. Shi, L. Aristilde and F. M. M. Morel, The effect ofpH on the uptake of zinc and cadmium in marine phyto-plankton: Possible role of weak complexes, Limnol. Ocea-nogr., 2012, 57, 293–304.
158 M. I. Heller and P. L. Croot, Superoxide decay as a probefor speciation changes during dust dissolution in TropicalAtlantic surface waters near Cape Verde, Mar. Chem.,2011, 126, 37–55.
159 H. J. Venables, A. Clarke and M. P. Meredith, Wintertimecontrols on summer stratification and productivity at thewestern Antarctic Peninsula, Limnol. Oceanogr., 2013, 58,1035–1047.
160 C. Schlosser, C. L. De La Rocha, P. Streu and P. L. Croot,Solubility of iron in the southern ocean, Limnol. Ocea-nogr., 2012, 57, 684–697.
161 P. Borer, B. Sulzberger, S. J. Hug, S. M. Kraemer andR. Kretzschmar, Photoreductive Dissolution of Iron(III)(Hydr)oxides in the Absence and Presence of Organicligands: Experimental Studies and Kinetic Modeling,Environ. Sci. Technol., 2009, 43, 1864–1870.
162 K. Kim, W. Choi, M. R. Hoffmann, H.-I. Yoon andB.-K. Park, Photoreductive dissolution of iron oxidestrapped in ice and its environmental implications,Environ. Sci. Technol., 2010, 44, 4142–4148.
163 D. Shi, Y. Xu, B. M. Hopkinson and F. M. M. Morel, Effectof ocean acidification on iron availability to marine phyto-plankton, Science, 2010, 327, 676–679.
164 D. Shi, S. A. Kranz, J.-M. Kim and F. M. M. Morel, Oceanacidification slows nitrogen fixation and growth in thedominant diazotroph Trichodesmium under low-iron con-ditions, Proc. Natl. Acad. Sci. U. S. A., 2012, 109, E3094–E3100.
165 E. J. Dlugokencky, E. G. Nisbet, R. Fisher and D. Lowry,Global atmospheric methane: Budget, changes and
Perspective Photochemical & Photobiological Sciences
144 | Photochem. Photobiol. Sci., 2015, 14, 127–148 This journal is © The Royal Society of Chemistry and Owner Societies 2015

dangers, Philos. Trans. R. Soc. London, Ser. A, 2011, 369,2058–2072.
166 C. D. Holmes, M. J. Prather, O. A. Sovde and G. Myhre,Future methane, hydroxyl, and their uncertainties: keyclimate and emission parameters for future predictions,Atmos. Chem. Phys., 2013, 13, 285–302.
167 V. Naik, A. Voulgarakis, A. M. Fiore, L. W. Horowitz,J. F. Lamarque, M. Lin, M. J. Prather, P. J. Young,D. Bergmann, P. J. Cameron-Smith, I. Cionni,W. J. Collins, S. B. Dalsoren, R. Doherty, V. Eyring,G. Faluvegi, G. A. Folberth, B. Josse, Y. H. Lee,I. A. MacKenzie, T. Nagashima, T. P. C. van Noije,D. A. Plummer, M. Righi, S. T. Rumbold, R. Skeie,D. T. Shindell, D. S. Stevenson, S. Strode, K. Sudo,S. Szopa and G. Zeng, Preindustrial to present-daychanges in tropospheric hydroxyl radical and methanelifetime from the Atmospheric Chemistry and ClimateModel Intercomparison Project (ACCMIP), Atmos. Chem.Phys., 2013, 13, 5277–5298.
168 S. A. Montzka, E. J. Dlugokencky and J. H. Butler, Non-CO2 greenhouse gases and climate change, Nature, 2011,476, 43–50.
169 S. Houweling, M. Krol, P. Bergamaschi, C. Frankenberg,E. J. Dlugokencky, I. Morino, J. Notholt, V. Sherlock,D. Wunch, V. Beck, C. Gerbig, H. Chen, E. A. Kort,T. Röckmann and I. Aben, A multi-year methane inversionusing SCIAMACHY, accounting for systematic errors usingTCCON measurements, Atmos. Chem. Phys. Discuss., 2013,13, 28117–28171.
170 A. N. Ross, M. J. Wooster, H. Boesch and R. Parker, Firstsatellite measurements of carbon dioxide and methaneemission ratios in wildfire plumes, Geophys. Res. Lett.,2013, 40, 4098–4102.
171 A. Vasileva and K. Moiseenko, Methane emissions from2000 to 2011 wildfires in Northeast Eurasia estimatedwith MODIS burned area data, Atmos. Environ., 2013, 71,115–121.
172 J. Worden, Z. Jiang, D. B. A. Jones, M. Alvarado,K. Bowman, C. Frankenberg, E. A. Kort, S. S. Kulawik,M. Lee, J. Liu, V. Payne, K. Wecht and H. Worden,El Niño, the 2006 Indonesian peat fires, and the distri-bution of atmospheric methane, Geophys. Res. Lett., 2013,40, 4938–4943.
173 Y. Zhang, D. Chu, Y. Li, L. Wang and Y. Wu, Effect of elev-ated UV-B radiation on CH4 emissions from the stands ofSpartina alterniflora and Phragmites australis in a coastalsalt marsh, Aquat. Bot., 2013, 111, 150–156.
174 S. K. Morsky, J. K. Haapala, R. Rinnan, S. Saarnio,H. Suokanerva, K. Latola, E. Kyro, J. Silvola, T. Holopainenand P. J. Martikainen, Minor long-term effects ofultraviolet-B radiation on methane dynamics of a subarc-tic fen in Northern Finland, Biogeochemistry, 2012, 108,233–243.
175 E. A. Kort, S. C. Wofsy, B. C. Daube, M. Diao, J. W. Elkins,R. S. Gao, E. J. Hintsa, D. F. Hurst, R. Jimenez,F. L. Moore, J. R. Spackman and M. A. Zondlo,
Atmospheric observations of Arctic Ocean methane emis-sions up to 82o north, Nat. Geosci., 2012, 5, 318–321.
176 N. Shakhova, I. Semiletov, A. Salyuk, V. Yusupov,D. Kosmach and O. Gustafsson, Extensive methaneventing to the atmosphere from sediments of the EastSiberian Arctic Shelf, Science, 2010, 327, 1246–1250.
177 O. S. Pokrovsky, L. S. Shirokova, S. N. Kirpotin,S. P. Kulizhsky and S. N. Vorobiev, Impact of westernSiberia heat wave 2012 on greenhouse gases and tracemetal concentration in thaw lakes of discontinuouspermafrost zone, Biogeosciences, 2013, 10, 5349–5365.
178 C. G. Fichot and W. L. Miller, An approach to quantifydepth-resolved marine photochemical fluxes using remotesensing: Application to carbon monoxide (CO) photo-production, Remote Sens. Environ., 2010, 114, 1363–1377.
179 G. Song, H. Xie, S. Belanger, E. Leymarie and M. Babin,Spectrally resolved efficiencies of carbon monoxide (CO)photoproduction in the western Canadian Arctic: Particlesversus solutes, Biogeosciences, 2013, 10, 3731–3748.
180 A. Stubbins, C. S. Law, G. Uher and R. C. Upstill-Goddard,Carbon monoxide apparent quantum yields and photo-production in the Tyne estuary, Biogeosciences, 2011, 8,703–713.
181 G. P. Yang, C. Y. Ren, X. L. Lu, C. Y. Liu and H. B. Ding,Distribution, flux, and photoproduction of carbon mon-oxide in the East China Sea and Yellow Sea in spring,J. Geophys. Res.: Oceans, 2011, 116.
182 Y. Zhang and H. X. Xie, The sources and sinks of carbonmonoxide in the St. Lawrence estuarine system, Deep-SeaRes., Part II, 2012, 81–84, 114–123.
183 C. E. Schmidt and B. G. Heikes, Aqueous carbon monox-ide cycling in a fjord-like estuary, Estuaries Coasts, 2014,37, 751–762.
184 H. Xie, S. Belanger, G. Song, R. Benner, A. Taalba,M. Blais, J. E. Tremblay and M. Babin, Photoproduction ofammonium in the southeastern Beaufort Sea and its bio-geochemical implications, Biogeosciences, 2012, 9, 3047–3061.
185 L. Derendorp, J. B. Quist, R. Holzinger and T. Rockmann,Emissions of H2 and CO from leaf litter of Sequoiadendrongiganteum, and their dependence on UV radiation andtemperature, Atmos. Environ., 2011, 45, 7520–7524.
186 D. Bruhn, K. R. Albert, T. N. Mikkelsen and P. Ambus,UV-induced carbon monoxide emission from livingvegetation, Biogeosciences, 2013, 10, 7877–7882.
187 S. Madronich, M. Shao, S. R. Wilson, K. R. Solomon,J. Longstrethe and X. Tang, Changes in air quality andtropospheric composition due to depletion of strato-spheric ozone and interactions with changing climate:Implications for human and environmental health, Photo-chem. Photobiol. Sci., 2015, 14, DOI: 10.1039/c4pp90037e,this issue.
188 M. J. Prather and J. Hsu, Coupling of nitrous oxide andmethane by global atmospheric chemistry, Science, 2010,330, 952–954.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 127–148 | 145

189 K. Pilegaard, Processes regulating nitric oxide emissionsfrom soils, Philos. Trans. R. Soc. London, B, 2013, 368.
190 R. Oswald, T. Behrendt, M. Ermel, D. Wu, H. Su,Y. Cheng, C. Breuninger, A. Moravek, E. Mougin,C. Delon, B. Loubet, A. Pommerening-Roeser, M. Soergel,U. Poeschl, T. Hoffmann, M. O. Andreae, F. X. Meixnerand I. Trebs, HONO emissions from soil bacteria as amajor source of atmospheric reactive nitrogen, Science,2013, 341, 1233–1235.
191 H. Su, Y. Cheng, R. Oswald, T. Behrendt, I. Trebs,F. X. Meixner, M. O. Andreae, P. Cheng, Y. Zhang andU. Poeschl, Soil nitrite as a source of atmospheric HONOand OH radicals, Science, 2011, 333, 1616–1618.
192 S. Morin, J. Erbland, J. Savarino, F. Domine, J. Bock,U. Friess, H. W. Jacobi, H. Sihler and J. M. F. Martins, Anisotopic view on the connection between photolytic emis-sions of NOx from the Arctic snowpack and its oxidationby reactive halogens, J. Geophys. Res., [Atmos.], 2012, 117.
193 N. K. Richards and B. J. Finlayson-Pitts, Production of gasphase NO2 and halogens from the photochemical oxi-dation of aqueous mixtures of sea salt and nitrate ions atroom temperature, Environ. Sci. Technol., 2012, 46, 10447–10454.
194 J. R. Brown, J. C. Blankinship, A. Niboyet,K. J. van Groenigen, P. Dijkstra, X. Le Roux, P. W. Leadleyand B. A. Hungate, Effects of multiple global change treat-ments on soil N2O fluxes, Biogeochemistry, 2012, 109,85–100.
195 K. J. van Groenigen, C. W. Osenberg and B. A. Hungate,Increased soil emissions of potent greenhouse gasesunder increased atmospheric CO2, Nature, 2011, 475,214–U121.
196 A. E. Santoro, C. Buchwald, M. R. McIlvin andK. L. Casciotti, Isotopic signature of N2O produced bymarine ammonia-oxidizing Archaea, Science, 2011, 333,1282–1285.
197 L. A. Codispoti, Interesting times for marine N2O, Science,2010, 327, 1339–1340.
198 J. T. Walker, C. A. Stow and C. Geron, Nitrous oxide emis-sions from the Gulf of Mexico hypoxic zone, Environ. Sci.Technol., 2010, 44, 1617–1623.
199 M. Voss, H. W. Bange, J. W. Dippner, J. J. Middelburg,J. P. Montoya and B. Ward, The marine nitrogen cycle:recent discoveries, uncertainties and the potential rele-vance of climate change, Philos. Trans. R. Soc. London, B,2013, 368.
200 V. Kitidis, R. C. Upstill-Goddard and L. G. Anderson,Methane and nitrous oxide in surface water along theNorth-West Passage, Arctic Ocean, Mar. Chem., 2010, 121,80–86.
201 K. Randall, M. Scarratt, M. Levasseur, S. Michaud, H. Xieand M. Gosselin, First measurements of nitrous oxide inArctic sea ice, J. Geophys. Res.: Oceans, 2012, 117.
202 X. Jin and N. Gruber, Offsetting the radiative benefit ofocean iron fertilization by enhancing N2O emissions,Geophys. Res. Lett., 2003, 30.
203 E. Blei and M. R. Heal, Methyl bromide and methyl chlor-ide fluxes from temperate forest litter, Atmos. Environ.,2011, 45, 1543–1547.
204 T. Fujimori, Y. Yoneyama, G. Taniai, M. Kurihara,H. Tamegai and S. Hashimoto, Methyl halide productionby cultures of marine proteobacteria Erythrobacter andPseudomonas and isolated bacteria from brackish water,Limnol. Oceanogr., 2012, 57, 154–162.
205 C. J. Hardacre and M. R. Heal, Characterization of methylbromide and methyl chloride fluxes at temperate fresh-water wetlands, J. Geophys. Res.: Atmos., 2013, 118, 977–991.
206 M. K. Kurihara, M. Kimura, Y. Iwamoto, Y. Narita,A. Ooki, Y. J. Eum, A. Tsuda, K. Suzuki, Y. Tani,Y. Yokouchi, M. Uematsu and S. Hashimoto, Distributionsof short-lived iodocarbons and biogenic trace gases in theopen ocean and atmosphere in the western North Pacific,Mar. Chem., 2010, 118, 156–170.
207 C. Y. Lin and S. L. Manley, Bromoform production fromseawater treated with bromoperoxidase, Limnol. Oceanogr.,2012, 57, 1857–1866.
208 R. Hossaini, M. P. Chipperfield, W. Feng, T. J. Breider,E. Atlas, S. A. Montzka, B. R. Miller, F. Moore andJ. Elkins, The contribution of natural and anthropogenicvery short-lived species to stratospheric bromine, Atmos.Chem. Phys., 2012, 12, 371–380.
209 A. Saiz-Lopez and R. von Glasow, Reactive halogen chem-istry in the troposphere, Chem. Soc. Rev., 2012, 41, 6448–6472.
210 A. Saiz-Lopez, J. F. Lamarque, D. E. Kinnison, S. Tilmes,C. Ordonez, J. J. Orlando, A. J. Conley, J. M. C. Plane,A. S. Mahajan, G. S. Santos, E. L. Atlas, D. R. Blake,S. P. Sander, S. Schauffler, A. M. Thompson andG. Brasseur, Estimating the climate significance ofhalogen-driven ozone loss in the tropical marine tropo-sphere, Atmos. Chem. Phys., 2012, 12, 3939–3949.
211 A. Qureshi, N. J. O’Driscoll, M. MacLeod, Y.-M. Neuholdand K. Hungerbuehler, Photoreactions of mercury insurface ocean water: gross reaction kinetics and possiblepathways, Environ. Sci. Technol., 2010, 44, 644–649.
212 R. Hossaini, H. Mantle, M. P. Chipperfield, S. A. Montzka,P. Hamer, E. Ziska, B. Quack, K. Krueger, S. Tegtmeier,E. Atlas, S. Sala, A. Engel, H. Boenisch, T. Keber, D. Oram,G. Mills, C. Ordonez, A. Saiz-Lopez, N. Warwick, Q. Liang,W. Feng, E. Moore, B. R. Miller, V. Marecal,N. A. D. Richards, M. Dorf and K. Pfeilsticker, Evaluatingglobal emission inventories of biogenic bromocarbons,Atmos. Chem. Phys., 2013, 13, 11819–11838.
213 R. Hossaini, M. P. Chipperfield, S. Dhomse, C. Ordonez,A. Saiz-Lopez, N. L. Abraham, A. Archibald, P. Braesicke,P. Telford, N. Warwick, X. Yang and J. Pyle, Modellingfuture changes to the stratospheric source gas injection ofbiogenic bromocarbons, Geophys. Res. Lett., 2012, 39.
214 N. M. Mahowald, Atmospheric Biogeochemistry, inEncyclopedia of Sustainability Science and Technology,Springer, 2012.
Perspective Photochemical & Photobiological Sciences
146 | Photochem. Photobiol. Sci., 2015, 14, 127–148 This journal is © The Royal Society of Chemistry and Owner Societies 2015

215 F. Lambert, J. S. Kug, R. J. Park, N. Mahowald,G. Winckler, A. Abe-Ouchi, R. O’Ishi, T. Takemura andJ. H. Lee, The role of mineral-dust aerosols in polartemperature amplification, Nat. Clim. Change, 2013, 3,487–491.
216 Y. Ming, V. Ramaswamy and G. Chen, A modelinvestigation of aerosol-induced changes in borealwinter extratropical circulation, J. Clim., 2011, 24, 6077–6091.
217 J. M. Creamean, K. J. Suski, D. Rosenfeld, A. Cazorla,P. J. DeMott, R. C. Sullivan, A. B. White, F. M. Ralph,P. Minnis, J. M. Comstock, J. M. Tomlinson andK. A. Prather, Dust and biological aerosols from theSahara and Asia influence precipitation in the WesternU.S., Science, 2013, 339, 1572–1578.
218 M. Schulz, J. M. Prospero, A. R. Baker, F. Dentener,L. Ickes, P. S. Liss, N. M. Mahowald, S. Nickovic,C. P. Garcia-Pando, S. Rodriguez, M. Sarin, I. Tegen andR. A. Duce, Atmospheric transport and deposition ofmineral dust to the ocean: Implications for researchneeds, Environ. Sci. Technol., 2012, 46, 10390–10404.
219 J. Mao, S. Fan, D. J. Jacob and K. R. Travis, Radical loss inthe atmosphere from Cu-Fe redox coupling in aerosols,Atmos. Chem. Phys., 2013, 13, 509–519.
220 M. Kulmala, T. Nieminen, R. Chellapermal, R. Makkonen,J. Bäck and K. Veli-Matti, Climate feedbacks linking theincreasing atmospheric CO2 concentration, BVOC emis-sions, aerosols and clouds in forest ecosystems, TreePhysiol., 2013, 5, 489–508.
221 S.-C. Hsu, C.-A. Huh, C.-Y. Lin, W.-N. Chen,N. M. Mahowald, S.-C. Liu, C. C. K. Chou, M.-C. Liang,C.-J. Tsai, F.-J. Lin, J.-P. Chen and Y.-T. Huang, Dust trans-port from non-East Asian sources to the North Pacific,Geophys. Res. Lett., 2012, 39.
222 M. Kanakidou, R. A. Duce, J. M. Prospero, A. R. Baker,C. Benitez-Nelson, F. J. Dentener, K. A. Hunter, P. S. Liss,N. Mahowald, G. S. Okin, M. Sarin, K. Tsigaridis,M. Uematsu, L. M. Zamora and T. Zhu, Atmosphericfluxes of organic N and P to the global ocean, Global Bio-geochem. Cycles, 2012, 26.
223 D. Rothenber, N. M. Mahowald, K. Lindsay, D. Scott,K. Moore and P. Thornton, Volcano impacts on climateand biogeochemistry in a coupled carbon-climate model,Earth Syst. Dyn., 2012, 3, 121–136.
224 H. Liang, Z. M. Chen, D. Huang, Y. Zhao and Z. Y. Li,Impacts of aerosols on the chemistry of atmospheric tracegases: a case study of peroxides and HO2 radicals, Atmos.Chem. Phys. Discuss., 2013, 13, 16549–16595.
225 T. Gouin, J. M. Armitage, I. T. Cousins, D. C. G. Muir,C. A. Ng, L. Reid and S. Tao, Influence of global climatechange on chemical fate and bioaccumulation: The roleof multimedia models, Environ. Toxicol. Chem., 2013, 32,20–31.
226 M. J. Hooper, G. T. Ankley, D. A. Cristol, L. A. Maryoung,P. D. Noyes and K. E. Pinkerton, Interactions betweenchemical and climate stressors: A role for mechanistic
toxicology in assessing climate change risks, Environ.Toxicol. Chem., 2013, 32, 32–48.
227 J. Armitage, C. Quinn and F. Wania, Global climatechange and contaminants—an overview of opportunitiesand priorities for modelling the potential implications forlong-term human exposure to organic compounds in theArctic, J. Environ. Monit., 2011, 13, 1352–1546.
228 A. M. Grannas, C. Bogdal, K. J. Hageman, C. Halsall,T. Harner, H. Hung, R. Kallenborn, P. Klan, J. Klanova,R. W. Macdonald, T. Meyer and F. Wania, The role of theglobal cryosphere in the fate of organic contaminants,Atmos. Chem. Phys., 2013, 13, 3271–3305.
229 UNEP, Climate change and POPs: Predicting the impacts.Report of the UNEP/AMAP Expert Group, (Ed.: Secretariatof the Stockholm Convention), United Nations Environ-ment Programme/Arctic Monitoring and Assessmentprogram, Geneva, Switzerland, 2011.
230 L. Carlos, D. O. Martire, M. C. Gonzalez, J. Gomis,A. Bernabeu, A. M. Amat and A. Arques, Photochemicalfate of a mixture of emerging pollutants in the presenceof humic substances, Water Res., 2012, 46, 4732–4740.
231 Y. Chen, H. Li, Z. P. Wang, H. J. Li, T. Tao and Y. G. Zuo,Photodegradation of selected beta-blockers in aqueousfulvic acid solutions: Kinetics, mechanism, and productanalysis, Water Res., 2012, 46, 2965–2972.
232 J. J. Guerard and Y. P. Chin, Photodegradation oformetoprim in aquaculture and stream-derived dissolvedorganic matter, J. Agric. Food Chem., 2012, 60, 9801–9806.
233 G. A. Rowland, A. R. Bausch and A. M. Grannas, Photo-chemical processing of aldrin and dieldrin in frozenaqueous solutions under arctic field conditions, Environ.Pollut., 2011, 159, 1076–1084.
234 M. B. Fisher, D. C. Love, R. Schuech and K. L. Nelson,Simulated sunlight action spectra for inactivation of MS2and PRD1 bacteriophages in clear water, Environ. Sci.Technol., 2011, 45, 9249–9255.
235 K. G. McGuigan, R. M. Conroy, H. J. Mosler, M. du Preez,E. Ubomba-Jaswa and P. Fernandez-Ibanez, Solar waterdisinfection (SODIS): A review from bench-top to roof-top,J. Hazard. Mater., 2012, 235, 29–46.
236 A. I. Silverman, B. M. Peterson, A. B. Boehm, K. McNeilland K. L. Nelson, Sunlight inactivation of human virusesand bacteriophages in coastal waters containing naturalphotosensitizers, Environ. Sci. Technol., 2013, 47, 1870–1878.
237 E. S. Corbitt, D. J. Jacob, C. D. Holmes, D. G. Streets andE. M. Sunderland, Global source-receptor relationshipsfor mercury deposition under present-day and 2050 emis-sions scenarios, Environ. Sci. Technol., 2011, 45, 10477–10484.
238 D. P. Krabbenhoft and E. M. Sunderland, Global changeand mercury, Science, 2013, 341, 1457–1458.
239 A. L. Andrady, A. Torikai, H. H. Redhwi, K. K. Pandey andP. Gies, Consequences of stratospheric ozone depletionand climate change on the use of materials, Photochem.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 127–148 | 147

Photobiol. Sci., 2015, 14, DOI: 10.1039/c4pp90038c, thisissue.
240 N. F. Adegboyega, V. K. Sharma, K. Siskova, R. Zboril,M. Sohn, B. J. Schultz and S. Banerjee, Interactions ofAqueous Ag+ with Fulvic Acids: Mechanisms of SilverNanoparticle Formation and Investigation of Stability,Environ. Sci. Technol., 2013, 47, 757–764.
241 W. C. Hou, B. Stuart, R. Howes and R. G. Zepp, Sunlight-driven reduction of silver ions by natural organic matter:formation and transformation of silver nanoparticles,Environ. Sci. Technol., 2013, 47, 7713–7721.
242 C. Kingston, R. Zepp, A. Andrady, D. Boverhof, R. Fehir,D. Hawkins, J. Roberts, P. Sayre, B. Shelton, Y. Sultan,V. Vejins and W. Wohlleben, Release characteristics ofselected carbon nanotube polymer composites, Carbon,2014, 68, 33–57.
243 A. P. Ballantyne, C. B. Alden, J. B. Miller, P. P. Tans andJ. W. C. White, Increase in observed net carbon dioxideuptake by land and oceans during the past 50 years,Nature, 2012, 488, 70.
244 J. G. Canadell, C. Le Quere, M. R. Raupach, C. B. Field,E. T. Buitenhuis, P. Ciais, T. J. Conway, N. P. Gillett,R. A. Houghton and G. Marland, Contributions to acceler-ating atmospheric CO2 growth from economic activity,carbon intensity, and efficiency of natural sinks, Proc.Natl. Acad. Sci. U. S. A., 2007, 104, 18866–18870.
245 C. Le Quere, T. Takahashi, E. T. Buitenhuis,C. Roedenbeck and S. C. Sutherland, Impact of climatechange and variability on the global oceanic sink of CO2,Global Biogeochem. Cycles, 2010, 24.
246 G. A. McKinley, A. R. Fay, T. Takahashi and N. Metzl, Con-vergence of atmospheric and North Atlantic carbondioxide trends on multidecadal timescales, Nat. Geosci.,2011, 4, 606–610.
247 W. Knorr, Is the airborne fraction of anthropogenic CO2
emissions increasing?, Geophys. Res. Lett., 2009, 36.248 C. L. Sabine and T. Tanhua, Estimation of anthropogenic
CO2 inventories in the ocean, in Annual Review of MarineScience, vol. 2, 2010, pp. 175–198.
249 I. Fountoulakis, A. F. Bais, K. Tourpali, K. Fragkos andS. Misios, Projected changes in solar UV radiation in theArctic and sub-Arctic Oceans: Effects from changes inreflectivity, ice transmittance, clouds, and ozone,J. Geophys. Res., [Atmos.], 2014, 119, 8073–8090.
250 C. E. Williamson, R. G. Zepp, R. M. Lucas, S. Madronich,A. T. Austin, C. L. Ballaré, M. Norval, B. Sulzberger,A. F. Bais, R. L. McKenzie, S. A. Robinson, D. P. Hader,N. D. Paul and J. F. Bornman, Solar UV radiation in achanging climate, Nat. Clim. Change, 2014, 4, 434–441.
251 R. A. Houghton, Balancing the global carbon budget,Annu. Rev. Earth Planet. Sci., 2007, 35, 313–347.
Perspective Photochemical & Photobiological Sciences
148 | Photochem. Photobiol. Sci., 2015, 14, 127–148 This journal is © The Royal Society of Chemistry and Owner Societies 2015

Photochemical &Photobiological Sciences
PERSPECTIVE
Cite this: Photochem. Photobiol. Sci.,2015, 14, 149
Received 20th October 2014,Accepted 20th October 2014
DOI: 10.1039/c4pp90037e
www.rsc.org/pps
Changes in air quality and troposphericcomposition due to depletion of stratosphericozone and interactions with changing climate:implications for human and environmental health
S. Madronich,*a M. Shao,b S. R. Wilson,c K. R. Solomon,d J. D. Longstrethe andX. Y. Tangb
UV radiation is an essential driver for the formation of photochemical smog, which includes ground-level
ozone and particulate matter (PM). Recent analyses support earlier work showing that poor outdoor air
quality is a major environmental hazard as well as quantifying health effects on regional and global scales
more accurately. Greater exposure to these pollutants has been linked to increased risks of cardiovascular
and respiratory diseases in humans and is associated globally with several million premature deaths per
year. Ozone also has adverse effects on yields of crops, leading to loss of billions of US dollars each year.
These detrimental effects also may alter biological diversity and affect the function of natural ecosystems.
Future air quality will depend mostly on changes in emission of pollutants and their precursors, but
changes in UV radiation and climate will contribute as well. Significant reductions in emissions, mainly
from the energy and transportation sectors, have already led to improved air quality in many locations. Air
quality will continue to improve in those cities/states that can afford controls, and worsen where the regu-
latory infrastructure is not available. Future changes in UV radiation and climate will alter the rates of for-
mation of ground-level ozone and photochemically-generated particulate matter and must be considered
in predictions of air quality. The decrease in UV radiation associated with recovery of stratospheric ozone
will, according to recent global atmospheric model simulations, lead to increases in ground-level ozone at
most locations. If correct, this will add significantly to future ground-level ozone trends. However, the
spatial resolution of these global models is insufficient to inform policy at this time, especially for urban
areas. UV radiation affects the atmospheric concentration of hydroxyl radicals, •OH, which are responsible
for the self-cleaning of the atmosphere. Recent measurements confirm that, on a local scale, •OH radicals
respond rapidly to changes in UV radiation. However, on large (global) scales, models differ in their predic-
tions by nearly a factor of two, with consequent uncertainties for estimating the atmospheric lifetime and
concentrations of key greenhouse gases and air pollutants. Projections of future climate need to consider
these uncertainties. No new negative environmental effects of substitutes for ozone depleting substances
or their breakdown-products have been identified. However, some substitutes for the ozone depleting
substances will continue to contribute to global climate change if concentrations rise above current levels.
Introduction
The degradation of air quality is one of the major environ-mental hazards facing modern society. Human activities resultin the emission of many chemicals to the atmosphere, whichare either toxic themselves, or produce noxious compoundswhen exposed to ambient ultraviolet (UV) radiation. UV radi-ation is an essential driver for the generation of ground-levelozone (O3) and some particulate matter (PM, frequently calledaerosol) including sulfate, nitrate, and organic aerosols. Thesepollutants have major health implications for humans and theenvironment. Future changes in tropospheric UV radiation,
aAtmospheric Chemistry Division, National Center for Atmospheric Research,
Boulder, Colorado 80307, USA. E-mail: [email protected] University, College of Environmental Science and Engineering,
Beijing 100871, ChinacSchool of Chemistry, University of Wollongong, NSW, 2522, AustraliadCentre for Toxicology and School of Environmental Sciences, University of Guelph,
ON, N1G 2W1, CanadaeThe Institute for Global Risk Research, LLC, Bethesda, Maryland 20817, USA
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 149–169 | 149

whether from stratospheric ozone changes or other factorssuch as clouds, are likely to contribute to trends in air qualityand associated health effects.
UV radiation makes hydroxyl (•OH) radicals, the so-calledcleaning agents of the troposphere. These radicals limit theatmospheric lifetime of many gases that are important to bothtropospheric and stratospheric chemistry as well as climatechange, including methane (CH4), hydrogen-containing halo-carbons (e.g. hydrofluorocarbons, hydrochlorofluorocarbonsand hydrobromocarbons), and the oxides of sulfur and nitro-gen (SO2 and NOx).
This paper provides an assessment of how UV radiationaffects air quality, particularly ground-level O3 and PM, in thecontext of the growing body of knowledge on their healthimpacts, geographic distributions, and long-term trends. Itshould be recognised that many factors drive these trends,including changes in both natural and anthropogenic emis-sions, as well as climate variability through changes in temp-erature, moisture, and atmospheric circulation patterns.Tropospheric UV radiation is one of these factors, and itseffects can be approximately superimposed onto the effects ofthe other factors, but complex non-linear interactions must beconsidered to obtain reliable estimates.
Since the previous assessment of the interactions betweenozone depletion, climate change, and air-quality in the tropo-sphere,10,11 significant advances are noted in: (i) understandingand quantifying the important consequences of poor air qualityfor human health, separating the effects of O3 and PM; (ii)understanding changes in air pollution on urban and regionalscales, in terms of changes in anthropogenic emissions(increases or decreases, depending on location) as well aslong-range transport; (iii) understanding long-term changes ofkey tropospheric oxidants (O3 and •OH) on continental andglobal scales, as anthropogenic emissions continue in anenvironment where both stratospheric ozone and climate arealso changing.
An equally important advance is a better understanding ofuncertainties inherent in numerical models used to predictthe future chemical composition of the atmosphere. Thesecomputer models endeavor to represent and integrate themany chemical, physical, and biological processes that controlair quality as well as climate. Recent inter-comparisons amongthe models (see below) highlight important differences thatcast some doubt on the reliability of future projections, whilealso indicating a path to model improvements.
Ozone-depleting substances (ODS) could also affect airquality. While the long-lived chlorofluorocarbons (CFCs) breakdown almost exclusively in the stratosphere, the halogenatedreplacements break down in the troposphere. The cycling ofthe halogenated species in the troposphere needs to beassessed to ensure that there are no other significant short-and long-term effects that will result from the replacementsfor the halocarbons. Health effects could result from exposureto these substances, and are therefore included in this assess-ment (previously this was included in the health assessment,which now focuses on UV-mediated effects).
This paper provides summaries of the state of knowledgeon ground-level O3, PM, and •OH radicals, and addresses someof the more complex but still largely unquantified interactionsbetween air quality, climate change, and human activities.An important additional interaction between air quality andstratospheric ozone depletion is the introduction of substitutecompounds for ODSs pursuant to international agreements.The last part of this paper provides an update on selected sub-stitutes whose potential environmental and health impactsshould be considered.
In summary, our assessment updates and reinforces severalkey conclusions. Air pollution is increasingly recognised as amajor environmental hazard and a risk to human health, glob-ally leading to several million premature deaths per year.Air pollution also damages vegetation and reduces agriculturalyields, with associated economic losses estimated as $10–20billion annually. UV radiation is an essential ingredient for theformation of ground-level O3 and some PM, and of •OH radi-cals that control the global self-cleaning capacity of the tropo-sphere. Future trends in UV radiation will modulate futuretrends in air quality. Air quality is sensitive to other changes inthe environment including atmospheric circulation, hydrologi-cal cycles, and temperatures, all of which are likely to changedue to the combined effects of changing stratospheric ozoneand climate. No new negative environmental effects of the sub-stitutes for the ODSs have been identified.
Ground-level ozone
Health effects. Tropospheric O3 has significant effects onhuman morbidity and mortality. Premature mortality has beenestimated in recent studies, which are summarised in Table 1.Ozone and particulate matter (PM) often co-occur in the tropo-sphere and therefore their effects on human health aredifficult to separate. However, there appears to be no inter-action between these in terms of premature mortality. Earlierepidemiological studies (reviewed in ref. 12) have supportedthis conclusion and further studies by the same authors13,14
on the individual components of PM have shown that there isno interaction between these and ozone. Thus for the purposesof protecting human health, PM and O3 can be treated separ-ately. Premature mortality associated with exposure to ground-level ozone, while lower than that from PM, is still substantialwith several hundred thousand people affected globally eachyear (Table 1). Recent studies are broadly consistent on effects
Table 1 Premature mortality from ground-level ozone (O3) andparticulate matter (PM)
Source Year Area
Premature mortality(millions per year)
From O3 From PM
OECD19 2010 Global 0.35 1.42050 Global 0.75 3.6
Lim et al.18 2010 Global 0.05–0.27 2.8–3.6Fang et al.20 2000 Global 0.38 1.5Fann et al.21 2005 US only 0.005 0.05–0.2
Perspective Photochemical & Photobiological Sciences
150 | Photochem. Photobiol. Sci., 2015, 14, 149–169 This journal is © The Royal Society of Chemistry and Owner Societies 2015

of ozone on mortality in humans from cardiovascular andrespiratory diseases in Oporto, Portugal,15 Taipei,16 andPrague.17,18
Ozone-related morbidities manifested as acute and chronicbronchitis, asthma and/or atopic dermatitis,22 appendicitis,23
venous thromboembolic disease, and pulmonary embolisms24
have been reported. Ozone may also interact synergisticallywith viral infections. A study in Hong Kong reported interac-tive effects between viral infections and concentration of O3
that resulted in increased risks for hospitalisation for respi-ratory disease.25 The mechanism for this apparent synergismwas not reported.
A number of studies have extrapolated the effects ofexposure to O3 into the future. Based on the OECD19 study,premature deaths from ground-level ozone will increase toabout 0.75 million per year worldwide by 2050. The greatestincreases are predicted in India (130 premature deaths permillion per decade by 2050) but those in OECD countries willbe almost as large (95 premature deaths per million perdecade by 2050), mostly as a result of greater sensitivity in anageing population.19 Modelling of the interactions betweenconcentrations of tropospheric ozone, precursors of ozone inthe atmosphere, and climate change suggests that, by 2050,concentrations of O3 will increase in developing countries anddecrease in developed countries,26 due to regional differencesin future emissions.
Effects on plants. For plants, the most important air-pollu-tant is O3;
27,28 particulates have not been observed to have sub-stantial direct effects on plants. In our previous assessment,11
we noted that damage to crops by air-pollutants is likely tobecome more severe in the future. Since the last assessment,further studies have reinforced this conclusion. Based on ascenario of a world population of 9.1 billion, concentration ofCO2 of 550 ppm,† a concentration of ozone of 60 ppb (about10 ppb above current), and the climate warmer by ca. 2 °C by2050, Jaggard et al.29 postulated that yields of major crops (e.g.wheat, rice, soy, and maize) would be reduced by about 5%because of O3. However, this may be compensated byan increase in yield for most crops by about 13% because ofthe increased concentrations of CO2, depending on wateravailability.
In a review of studies on the effects of O3 on plants, it wasconcluded that reductions in the yields of 18 to 27% resultfrom exposure to O3 at concentrations of 70 to 100 ppb, at theupper end of typical regional concentrations.30 Not all cropsare equally sensitive to ozone. In a sensitive crop, such assoybean, yields in several cultivars were shown to decrease byas much as a factor of 2 with long-term exposure to 20–30 ppb(24 hour mean) of added ozone.31 Reductions in photosyn-thesis upon exposure to O3 were estimated in three differentforest types and found to be statistically significant for anorange orchard, and observable for ponderosa pine.32 The
potential for damage from O3 in crop plants was assessed inrelation to the IPCC (pessimistic) A2 scenario33 for 2100.1
Changes in gross plant productivity resulting from changes intropospheric O3 were projected to range from −40 to +15%,depending on location (Fig. 1). In another example, the rangesof global crop losses for wheat and soybean in 2030 as esti-mated from the IPCC A2 scenario were 5.4–26% and 15–19%,respectively.34 In this same study, yield reductions in the B1(optimistic) scenario were 4.0–17% for wheat and 9.5–15% forsoybean, with monetised annual losses estimated to rangefrom $12–21 billion (year 2000 dollar equivalents).
Although most of the research on the effects of O3 in eco-systems has been directed toward plants, effects on soil micro-organisms have also been reported.35 The abundance anddiversity of methanogenic bacteria in soils of rice paddies werefound to be reduced after exposure to elevated concentrationsof O3 (60 ppb) in ground-level air.
Overall, productivity and yields of crops will likely bereduced as a result of increases in concentrations of tropo-spheric O3. There is some hope that genetic selection of plantstolerant to O3 will mitigate these adverse effects on productionof food and fibre but other plants in the ecosystem are likely tosuffer greater adverse effects. These are expected to impactdiversity and functions of natural ecosystems. Other effects ofclimate change, e.g., mediated by temperature and precipi-tation, will also affect yields of crops.
Photochemical processes. Atmospheric ozone (O3) is gener-ated primarily in the atmosphere by photochemical reactionsinvolving UV radiation. In the stratosphere, it is made directlyby the photo-dissociation of molecular oxygen (O2) into twooxygen atoms (2O), followed by the association of each of theseatoms with remaining O2 (see Table 2) to make two O3 mole-cules. In the troposphere, this direct formation is notpossible because photons of sufficient energy to dissociate O2
(λ < 240 nm) are nearly completely absorbed by stratosphericO2, and so are not available in the troposphere. Descent ofstratospheric ozone to the troposphere does occur (e.g., duringstratospheric intrusions common during springtime atmid-latitudes) but accounts for only about 1/10th of thetropospheric production in global models,6 and is only
Fig. 1 Global assessment of the projected percentage changes in grossprimary productivity (GPP) due to O3 under the Intergovernmental Panelon Climate Change A2 scenario in 2100 within the World Wildlife Foun-dation Global 200 priority conservation areas. From Ainsworth et al.1
Reproduced with permission of the Royal Society.
†A ppm of X, parts per million, is a mixing ratio of 1 molecule of X per millionmolecules of air. Similarly, ppb is parts per billion.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 149–169 | 151

a minor source of ozone found near the surface in pollutedregions.
Formation of O3 in polluted urban atmospheres has beenrecognised since the 1950s,36 and occurs when mixtures ofvolatile organic compounds (VOCs) and nitrogen oxides (NOx =NO + NO2) are exposed to the UV radiation available in the tropo-sphere. The chemistry leading to tropospheric O3 formationis generally complex because it (i) requires the absorption ofmultiple photons, (ii) is augmented by several catalytic cycles,and (iii) can be fuelled by many different VOCs, includingthose of anthropogenic and biogenic origin. Table 2 provides ahighly simplified schematic of the major chemical pathways.The production of tropospheric O3 is autocatalytic becausephotolysis of an initial amount of O3 (Table 2, reactions 3 and4) results in two •OH radicals, each of which can then continuethrough the reaction sequence to regenerate more O3. Notethat two photons are required for this process (Table 2, reac-tions 3 and 7) with their combined minimum energy beingmore than sufficient to break O2 directly.
Reaction 3 in Table 2, the photolysis of ozone to yieldexcited oxygen atoms O*, is the primary source of radicals(•OH, HO2, and RO2) involved in production of O3. It is alsovery sensitive to the overhead ozone column (see Table 1 ofMcKenzie et al.37). Other sources of radicals include thephotolysis of formaldehyde (CH2O), hydrogen peroxide (H2O2),and nitrous acid (HONO). These latter compounds are typicallythe products of previous chemical reactions initiated by O3
and •OH and so they are sensitive to the production of primaryradicals (Table 2, reaction 3) and therefore to UV-B radiationas affected by changes in stratospheric ozone.
Distributions and trends. Tropospheric ozone has increasedsince preindustrial times, mostly because of increasing anthro-pogenic emissions of the precursor gases, VOCs and NOx.Relative to that, the effects of changing the UV radiationenvironment, e.g., from mid-latitude stratospheric ozonedepletion, have been smaller but remain important due to thevery large number of people living in areas with poor air quality.
Urban ozone trends differ in different cities. Considerableprogress has been made in reducing urban O3 in Europe, theUnited States, and some other locations.2,38–40 Fig. 2 shows thereductions in ground level ozone achieved in Los Angeles andMexico City over the past several decades. In Beijing, from2005 to 2011, ground-based measurements give daytimeaverage O3 increasing at 2.6 ppb (5%) per year.41 In compari-son, increases of 3% per year over 2002–2010 have beenreported for the tropospheric O3 column above Beijing.42
Other Asian cities showing increases include Hong Kong(0.55 ppb per year over 1994–2007), Seoul (about 5 ppbover 1991–2007), and Tokyo with a doubling of days with O3
exceedances (incidences where air quality standards areexceeded).40
Regional ozone (ozone averaged over large areas extendingwell beyond cities) is increasing at some locations, particularlyin densely populated areas, for example 6–7% per decade inthe Indo-Gangetic Plains.43 Regional production of ozone wasalso shown to be a factor limiting air quality improvementsfrom local emission reductions during the Beijing Olympics.44
Background ozone continues to increase at many locations,such as central and northwestern Europe, but is decreasing ineastern and southwestern Europe.38,39 Increases of 0.25 ppbper year have been reported for ground-level ozone atMace Head, Ireland.45 In the western U.S., mid-tropospheric(3–10 km) ozone has increased by 0.6 ppb per year in thespringtime over 1995–2008, and may be indicative of longrange transport.46 Changes in the seasonal cycle observed atvarious mid-latitude locations in the Northern Hemisphere areconsistent with increasing emissions of precursors.47 On theother hand, a review of measurements made with balloon-borne instruments (ozone sondes) and surface observations atremote locations showed that most of the increases in bothhemispheres occurred in the early part of the 20–40 year
Table 2 Atmospheric photochemical reactions
Stratospheric ozone formation: Reaction
O2 + hν (λ < 240 nm) → O + O (1)O + O2 → O3 (2)
Tropospheric ozone and •OH formation:
O3 + hν (λ < 330 nm) → O* + O2 (3)O* + H2O → •OH + •OH (4)•OH + VOC + O2 → HO2 (or organic analog RO2) +other products
(5)
HO2 + NO → NO2 +•OH (6)
NO2 + hν (λ < 420 nm) → NO + O (7)O + O2 → O3 (2)
Secondary tropospheric radical sources:
CH2O + hν (λ < 340 nm) + 2 O2 → HO2 + HO2 + CO (8)H2O2 + hν (λ < 350 nm) → •OH + •OH (9)HONO + hν (λ < 395 nm) → •OH + NO (10)
Fig. 2 Improvements in air quality in Los Angeles and Mexico City.Plotted is the 3-year average of the 4th highest maximum ozone 8-houraverage. (From Parrish et al.2).
Perspective Photochemical & Photobiological Sciences
152 | Photochem. Photobiol. Sci., 2015, 14, 149–169 This journal is © The Royal Society of Chemistry and Owner Societies 2015

record, with more recent changes characterised by little or noincrease.48
Changes in atmospheric circulation are also likely to haveaffected trends of tropospheric ozone. Some increases mayhave been due to changes in the rate of stratosphere–tropo-sphere air exchange.49 Trends at Mauna Loa, Hawaii, wereinfluenced by changes in circulation due to El Niño, transport-ing air masses from different regions of Asia to Hawaii.50
Prediction of tropospheric concentrations of O3 via numeri-cal models remains problematic, with large differences atregional38 and global scales.6,51 This is illustrated in Fig. 3,where several global models, represented by different lines, areseen to differ by as much as 25% for current conditions, andfuture predictions show a wider range depending on thechoice of future scenario as well as model. However, it shouldbe noted that future photochemical ozone formation will inany case depend to a large extent on details of future emis-sions, particularly those associated with different fuel choices(e.g. diesel, gasoline, or biofuels).
The specific response of tropospheric O3 to future changesin stratospheric O3 was modeled by Zeng et al.52 and Zhanget al.53 with both studies showing large-scale increases in tro-pospheric O3, as a net result of slowing both production andloss in response to declining UV levels. However the lowresolution of their models (several degrees latitude × longi-tude) is insufficient to discern urban effects where higherlevels of NOx are expected to maintain an opposite (positive)relationship between UV radiation and ground-level ozone54,55
that would indicate improvement in air quality in response torecovery of stratospheric ozone. Global models also do notagree well with measurements of the background atmosphere(e.g. ref. 56), indicating that there is still significant work to bedone in understanding this chemistry. These uncertaintiesmake it difficult to identify precisely which geographic regionswill experience decreases in tropospheric ozone as strato-spheric ozone recovers, and which ones will suffer increases.
Nevertheless, all models agree that over large regions tropo-spheric ozone will increase.
Nitrogen dioxide. The near-term outlook for troposphericoxidants can also be surmised from satellite measurements ofNO2 (see Fig. 4), a precursor of tropospheric ozone and animportant pollutant in its own right. The geographic distri-bution agrees with the general understanding of major emis-sion sources, particularly over the U.S., Europe, and East Asia.Trends, also derived from satellite-based observations, areshown in the lower panel for specific regions. Notably,decreases in NO2 are seen to have occurred over the U.S. andEurope, in accordance with NOx emission reduction policies,and consistent with the reductions in urban ozone reportedfor these regions. However, positive trends are noted for east-central China, the Middle East, and north-central India. Itseems likely that such recent trends will also continue into thenear-term future, with the associated expectation that ground-level ozone (and other photochemical pollutants) may increasein some areas and decrease in others.
Particulate matter
Particulate matter (PM) in the atmosphere consists of smallsolid or liquid particles suspended in air, also called aerosols.The size of PM is recognised as important for health effects,with PM smaller than 2.5 μm (termed PM2.5) being inhaled
Fig. 3 Global tropospheric ozone burden simulated by differentmodels. Drawn from data in Table 1 of Young et al.6 Thin lines are forindividual model results, thick lines are multi-model averages, for twoscenarios of future emissions, RCP2.6 (green) and RCP8.5 (red), asdefined by the Intergovernmental Panel on Climate Change.9
Fig. 4 Tropospheric vertical column of nitrogen dioxide, a major airpollutant and precursor of tropospheric ozone. Top panel: global distri-bution averaged from May 2004 to April 2005.3 Bottom panel: region-specific trends.8
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 149–169 | 153

deeper into lungs than larger particles, typically measured asall particles below 10 μm (PM10).
Health effects. Particulate matter in the troposphere causessignificant adverse health effects. A large body of literaturespanning decades of research has been reviewed and assessedconfirming this causal relationship (e.g. ref. 57). Table 1 showsrecent estimates of premature deaths from particulate matter.Recent studies have reported PM health effects from differentlocations. In the time-series analysis of hospitalisations forvenous thromboembolic disease in Chile between 2000 and2007 discussed for ozone above, an increase in concentrationof PM2.5 of 20 μg m−3 resulted in a relative risk (RR) of 1.05(95% CI 1.03–1.06) for all hospitalisations.24 A similar RR wasobserved for pulmonary embolism. In a study of 90 cities inChina over 1981–2000, those north of the river Huai had totalsuspended particulates (PM of all sizes) higher by 100 μg m−3
(95% CI = 61–307) than those south of the river, and wereassociated with a decrease in life-expectancy of 3 years (95% CI0.4–5.6).58 Much of the PM2.5 is due to particles generated inthe atmosphere by UV-dependent photochemistry (e.g. sulfate,nitrate, and organics, see below), although other PM sourcescan be extremely important, e.g., biomass burning plumesreaching densely populated urban areas (e.g., van Donkelaaret al.59).
Where control of emissions into the troposphere hasresulted in decreases in PM2.5, fewer health effects have beenobserved. In a study of life expectancy in 545 U.S. counties,reductions in PM2.5 of 10 μg m−3 from 2000 to 2007 wereassociated with an increase in mean life expectancy of 0.35years (SD = 0.16 years).60
Overall, the global relevance of particulates to humanhealth is very large, and substantial changes are expected tooccur in response to changes in climate.61 Future predictionsare uncertain due to limitations of atmospheric models andtheir assumptions62 and, specifically for human health effects,the difficulty to clearly separate effects of O3 and PM2.5.63
Future changes in aerosols are uncertain but may be substan-tial regionally. A multi-model analysis of past and futuretrends in aerosol, described in Fig. 8 of Bais et al.,64 indicateslarge changes in industrialised regions, particularly in China.
Effects on plants. Direct effects of PM on plants appear tobe minor, for example through direct deposition of PM onfoliage.57 However, two important indirect effects should berecognised. The first is an increase in diffuse visible radiationfrom the scattering of solar photons by aerosol particles, alter-ing photosynthetic efficiency within partly shaded canopies.The second is the surface deposition of some aerosol chemi-cals, for example, the heavy metals Cu, Ni, and Zn, with poten-tial effects on soil chemistry, microbial communities, andnutrient cycling.57
Atmospheric processes. Particles in the atmosphere includethose emitted directly, such as wind-blown dust and soil, com-bustion-generated soot (black carbon), and salt from sea-spray,as well as those formed in situ by condensation of vapours,such as sulfates, nitrates, and many organics. The latter,secondary, aerosols depend on UV-initiated reactions of •OH
radicals (see section on Global •OH models), and thus arelikely to be affected by changes in stratospheric ozone.However, we note the absence of specific studies addressinghow changes in UV associated with stratospheric O3 wouldaffect the formation and removal of tropospheric particles.
The formation of sulfate and nitrate aerosols is well under-stood in terms of the •OH oxidation of SO2 and NO2 giving sul-furic and nitric acids, respectively. While the majority of thisproduction occurs in the gas phase, the sulfate and nitratecondense rapidly to form particles, particularly if ammonia ispresent. Chemical reactions in cloud and rain water can alsocontribute.65
Considerable progress has been made recently in under-standing secondary organic aerosols (SOA), which previousobservations had shown to be seriously underestimated bymodels. While many details remain poorly understood,numerous studies support the basic conceptual model thathydrocarbons are oxidised (by •OH and NO3 radicals, and O3)into a myriad of heavier, more functionalised molecules aswell as smaller fragments.66 Molecules with multiple func-tional groups (e.g., alcohols, ketones, aldehydes, organic acids,nitrates and peroxides) typically have lower vapour pressuresand therefore are likely to condense onto particles. However,quantification remains a problem due to the large number ofchemical species contributing to particle mass. Significantadvances in modelling have been made by classifying thesemultifunctional compounds according to relevant properties,such as vapour pressure,67,68 solubility,69 oxidation state,70
atomic ratios (O, C, H, etc.),71,72 and carbon number andpolarity.73 In practice, for ambient aerosols many of these pro-perties are not known and therefore cannot be used to con-strain predictions. However, these modelling frameworks nowallow exploratory sensitivity analyses to help identify the mostimportant processes for more accurate parameterisation.
Removal of aerosols from the atmosphere is poorly under-stood. Ultimately removal from the atmosphere occurs by wetor dry deposition. Incorporation of aerosol particles into rain-drops (wet deposition) leads to lifetimes estimated to rangefrom 0.5 to 2 weeks.74–76 Dry deposition of particles is gener-ally slower.65,77
Distributions and trends. The global distribution of aero-sols is shown in Fig. 5. Satellite observations and models agreeon broad features, including the dust belt extending fromN. Africa to S. Asia, biomass burning evident over tropicalS. America, and high values over E. Asia. These optical depthvalues represent the entire aerosol vertical column and notnecessarily those at ground level. Surface network data areavailable in many countries and have been used to showdetailed geographical and seasonal distributions of majorchemical constituents of collected particles (e.g. ref. 78 forthe U.S.).
Heavily populated urban locations are of special interest,and some recently reported measurements in megacities aresummarised in Table 3. World Health Organization (WHO)guidelines are frequently exceeded by all cities listed.Reductions in PM concentrations are occurring in many cities,
Perspective Photochemical & Photobiological Sciences
154 | Photochem. Photobiol. Sci., 2015, 14, 149–169 This journal is © The Royal Society of Chemistry and Owner Societies 2015

in some cases well-documented by long-term urban monitor-ing networks, and evidently related to emissions-loweringtechnologies for both fixed and mobile sources. However,measurements at many polluted locations are still sparse,making an assessment of trend difficult.
Future concentrations of aerosols are subject to similarscenario assumptions as other pollutants.4,79,80 Globally aver-aged sulfate concentrations have already decreased over thelast two decades and are expected to continue decreasing.Organic and black carbon are expected to continue to increaseover the next few decades globally, but then decrease, withtiming and magnitude depending on the specific scenario.
On smaller geographic scales, aerosol concentrations aresensitive to local and regional emissions, and may improve orworsen depending on regulatory strategies. For example, amulti-model analysis of past and future trends in aerosols,
described in Fig. 8 of Bais et al.64 shows strong regionalreductions of aerosol concentrations by 2090, particularly inChina.
Hydroxyl radicals
Tropospheric self-cleaning capacity. An important role ofUV radiation in the troposphere is the production of •OHradicals by photolysis of tropospheric ozone (Reaction 3 inTable 2) followed by reaction with H2O (Reaction 4, Table 2).The •OH radicals react with many of the gases emitted at theEarth’s surface, including carbon monoxide (CO), methane(CH4) and other volatile organic compounds (VOCs), oxides ofnitrogen and sulfur (NO2 and SO2), and hydrohalocarbons(HFCs and HCFCs). The reactions with •OH determine theatmospheric residence time of these gases, as well as theiramount in the atmosphere since this is directly proportionalto the product of emission rates and lifetime.
Understanding •OH is fundamental to understanding thechemistry of ozone and secondary aerosols as well. Cyclingbetween •OH and HO2 (Reactions 5 and 6, Table 2) is essentialfor tropospheric ozone formation. Notably, •OH itself has alifetime of only seconds, but it affects O3 on the time scale ofhours to days, CO over months, and methane over a decade.For this reason, direct detection of •OH has focused on localshort-term measurements, while longer-term impacts, forexample, on methane lifetimes, have been estimated fromglobal models.
Fig. 5 Annual average aerosol optical depth at 550 nm from the MODISand MISR satellite instruments (top and bottom, for the years2004–2006) and models (middle, for the year 2000). From Schindellet al.4
Table 3 Concentrations of particulate matter in megacitiesa
CityPM10(μg m−3)
PM2.5(μg m−3)
Measurementperiod
WHO guidelines 20 10 Annual mean50 25 24-hour mean
Cairo 90–260 30–220 1999–2002Dakar 30–60 2008–2009Bangkok 40–90 (−) 1995–2008Beijing 150–180 (−) 95–155 1999–2008Delhi 50–300 50–250 2004–2009Dhaka >100 (+) >30 (+) 2002–2006Hong Kong 40–50 (−) 20–40 1998–2008Jakarta 60–100 (−) 2001–2007Manila 40–50 20–30 (−) 2001–2008Seoul 60–80 (−) 1995–2007Shanghai 90–110 (−) 2002–2007Tokyo 15–30 (−) 2001–2008Tehran 65–370 2003Santiago 50–100 (−) 20–30 (−) 2000–2008Sao Paulo 40–70 (−) 1996–2006Los Angeles 40–80 (−) 1998–2008Houston 30–40 (−) 1998–2008New York City 30–70 (−) 1998–2008Mexico City 50–180 (−) 20–25 (−) 1990–2010London 20–35 1994–2004Moscow 35–50 (+) 2006–2008Milan 35–60 (−) 2000–2009Istanbul 45 20 2002–2003
a Positive (+) and negative (−) trends are indicated. Values shown areranges observed over the measurement period. Compiled from datareported by Zhu et al.;40 see original for details of the samplingintervals.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 149–169 | 155

Local measurements of •OH. Direct measurements of •OHare difficult because of its high chemical reactivity. Withinseconds of being produced, •OH reacts with various gases (seeprevious paragraph) limiting its concentration to low values(about 106–107 mol cm−3 during daytime, smaller at night)that are exceedingly difficult to detect and quantify. The highreactivity also implies that •OH has high spatial and temporalfluctuations, being sensitive to variations in production (e.g.,to variations in UV radiation, O3, and H2O) as well as loss (e.g.,via reactions with CO, NOx, or VOCs). For this reason, it isimportant to note that locally measured •OH concentrationscannot be easily integrated spatially or temporally to estimate,for example, an annually averaged global •OH concentration.The main objective of direct local measurements is to evaluatewhether the variations in •OH follow the expected variations insimultaneously measured meteorological (UV radiation,humidity) and chemical variables (O3, NOx, VOCs, etc.).
Several techniques have been developed over the past fewdecades to detect and measure concentrations of •OH.Measurements have been reviewed by Heard and Pilling81 andmore recently by Stone et al.82 Most of the recent measure-ments are consistent with model calculations within afactor of approximately two, (e.g., urban locations includingNew York City,83 Tokyo,84 Mexico City,85,86 and Houston87).Studies in forested locations, specifically West Africa88 andnorthern Michigan89 also show reasonable agreement withmodels.
Measured •OH is much greater, by as much as an order ofmagnitude, than predicted by models in environments con-taining high concentrations of biogenic hydrocarbons (e.g.,isoprene, methyl butenol, and terpenes) and low concen-trations of NOx, including over the tropical forest ofSuriname,90–92 Borneo,93,94 the Pearl River Delta (PRD),95,96
and suburban Beijing during low-NOx episodes.97 This appar-ent underestimation of •OH by models has led to a re-examin-ation of the chemistry of isoprene at low NOx, and to thesuggestion that at least part of the •OH initially lost by reactionwith isoprene is later regenerated by secondary reactions.98,99
Simulations using an environmental smog chamber also indi-cate the need for some recycling of •OH by isoprene chemistryunder very low NOx conditions, although not to an extent thatwould explain the large discrepancies between observationsand models found over tropical Suriname (Fuchs et al.,2013).100
Several inter-comparisons between different •OH instru-ments show good agreement in some circumstances but alsodisagreement in others.82,101–103 The largest discrepanciesappear to occur in environments dominated by biogenichydrocarbons. For example, Mao et al.104 found a factor of twodifference between laser-induced fluorescence and a chemicalanalysis methods at Blodget Forest, California.
Instruments have been developed recently to measure thetotal •OH reactivity, i.e., the rate at which •OH molecules areremoved by reaction with the many constituents of sampledair (e.g., CO, VOCs, and NOx). The reactivity of •OH provides animportant constraint on the budget of •OH since it must essen-
tially balance the rate of production. Measurements show thatthis reactivity is larger than predicted from the simple sum oftypically known constituents, indicating the presence of sub-stantial but unmeasured amounts of other reactive com-pounds, by values ranging from 25–35% in Tokyo,105 about30% in a terpene-rich mid-latitude forest,106 a factor of 2 inthe Pearl River Delta107 and 60–90% in a boreal forest.108,109
The missing compounds are presumed to be a multitude ofpartly oxygenated organic compounds (aldehydes, ketones,etc.) formed during the •OH-initiated photo-degradation ofVOCs.
Despite unresolved differences among various •OH instru-ments and models, some fundamental aspects of the photo-chemistry have been clearly demonstrated. Important in thepresent context is the theoretical expectation that, for relativelyclean conditions, concentrations of •OH should scale more orless linearly with the photolysis of O3 to generate O* (Reaction3, Table 2), j (O3), which in turn is dependent on the amountof UV radiation. This linear correlation has now been re-con-firmed by direct measurements of •OH and j (O3) over a year atMace Head, Ireland110 and is in agreement with earlier obser-vations in the tropical Atlantic111 and in the European Alps.112
Because tropospheric j (O3) values are sensitive to the overheadozone column, with a ∼1.5% increase in j (O3) for each 1%decrease in the O3 column (see Table 1 of McKenzie et al.,37),these studies reaffirm the importance of stratospheric ozone totropospheric •OH and to the photochemistry of the loweratmosphere.
Global •OH models. Estimates of long-term changes inglobal •OH are uncertain and variable. Empirical estimates,based on the measured concentrations of trace gases, such asmethyl chloroform, whose emissions and •OH kinetics are wellknown, are difficult due to large changes in emissions of suit-able gases. An analysis of the decline in concentrations ofmethyl chloroform by Montzka et al.113 concluded that glob-ally averaged •OH varied by less than ±5% during 1997–2007.On the other hand, Monteil et al.114 interpreted measurementsof methane isotopes (13C) to infer that the slowing of CH4
trends in the early 2000s was due to increasing concentrationsof •OH, at about 5% per decade, due to global increases inNOx emissions. Thus, the direction and magnitude of recenttrends in global •OH remains unclear.
Estimates over longer time scales are largely based onmodels, which differ significantly. This was demonstrated bythe recent intercomparison of 16 global chemistry-transportmodels for predictions of the methane lifetime, which islimited by reaction with •OH.5,7 The modelled mean lifetimeof methane was 8.6 ± 1.2 years (range 6.4–11.6 years) for theyear 2000. Pre-industrial (1850) to present day (2000) changesin •OH were either positive or negative (see Fig. 6), dependinglargely on how each model specified relative changes in emis-sions of CO and NOx. Pike and Young115 showed that globalconcentrations of •OH (and therefore the lifetime of CH4) weresensitive to how models represent •OH recycling by isoprene,which remains uncertain, as discussed above. If suchbuffering of •OH by biogenic VOCs is pervasive, it casts doubt
Perspective Photochemical & Photobiological Sciences
156 | Photochem. Photobiol. Sci., 2015, 14, 149–169 This journal is © The Royal Society of Chemistry and Owner Societies 2015
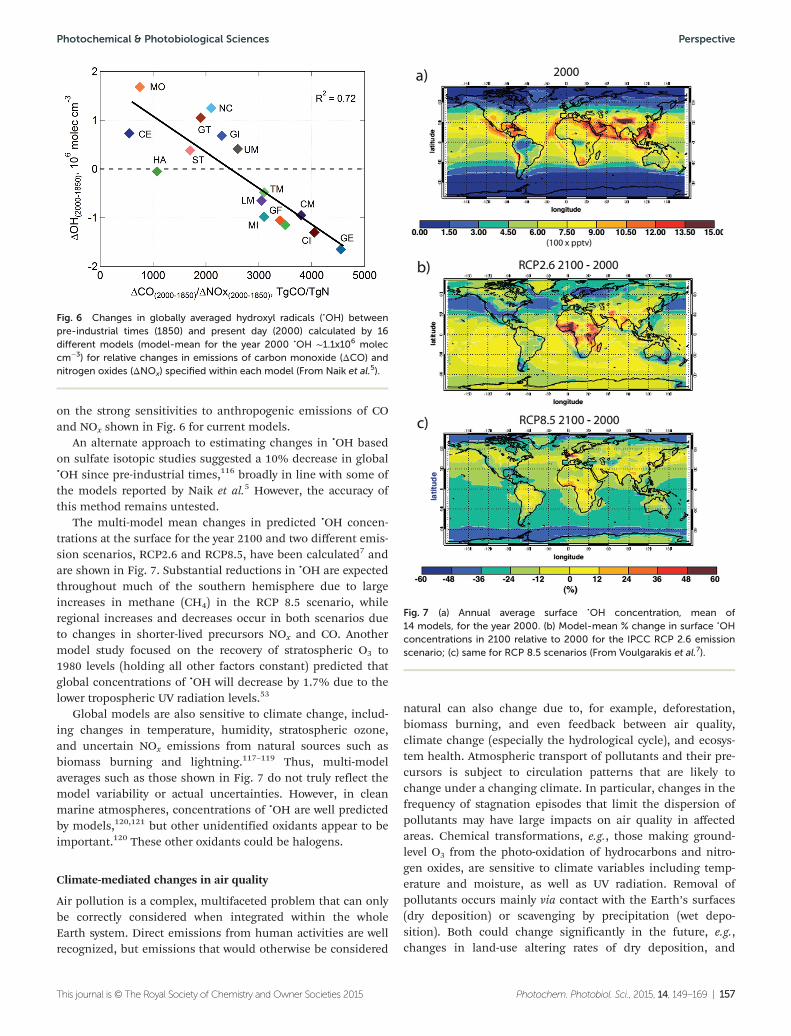
on the strong sensitivities to anthropogenic emissions of COand NOx shown in Fig. 6 for current models.
An alternate approach to estimating changes in •OH basedon sulfate isotopic studies suggested a 10% decrease in global•OH since pre-industrial times,116 broadly in line with some ofthe models reported by Naik et al.5 However, the accuracy ofthis method remains untested.
The multi-model mean changes in predicted •OH concen-trations at the surface for the year 2100 and two different emis-sion scenarios, RCP2.6 and RCP8.5, have been calculated7 andare shown in Fig. 7. Substantial reductions in •OH are expectedthroughout much of the southern hemisphere due to largeincreases in methane (CH4) in the RCP 8.5 scenario, whileregional increases and decreases occur in both scenarios dueto changes in shorter-lived precursors NOx and CO. Anothermodel study focused on the recovery of stratospheric O3 to1980 levels (holding all other factors constant) predicted thatglobal concentrations of •OH will decrease by 1.7% due to thelower tropospheric UV radiation levels.53
Global models are also sensitive to climate change, includ-ing changes in temperature, humidity, stratospheric ozone,and uncertain NOx emissions from natural sources such asbiomass burning and lightning.117–119 Thus, multi-modelaverages such as those shown in Fig. 7 do not truly reflect themodel variability or actual uncertainties. However, in cleanmarine atmospheres, concentrations of •OH are well predictedby models,120,121 but other unidentified oxidants appear to beimportant.120 These other oxidants could be halogens.
Climate-mediated changes in air quality
Air pollution is a complex, multifaceted problem that can onlybe correctly considered when integrated within the wholeEarth system. Direct emissions from human activities are wellrecognized, but emissions that would otherwise be considered
natural can also change due to, for example, deforestation,biomass burning, and even feedback between air quality,climate change (especially the hydrological cycle), and ecosys-tem health. Atmospheric transport of pollutants and their pre-cursors is subject to circulation patterns that are likely tochange under a changing climate. In particular, changes in thefrequency of stagnation episodes that limit the dispersion ofpollutants may have large impacts on air quality in affectedareas. Chemical transformations, e.g., those making ground-level O3 from the photo-oxidation of hydrocarbons and nitro-gen oxides, are sensitive to climate variables including temp-erature and moisture, as well as UV radiation. Removal ofpollutants occurs mainly via contact with the Earth’s surfaces(dry deposition) or scavenging by precipitation (wet depo-sition). Both could change significantly in the future, e.g.,changes in land-use altering rates of dry deposition, and
Fig. 6 Changes in globally averaged hydroxyl radicals (•OH) betweenpre-industrial times (1850) and present day (2000) calculated by 16different models (model-mean for the year 2000 •OH ∼1.1x106 moleccm−3) for relative changes in emissions of carbon monoxide (ΔCO) andnitrogen oxides (ΔNOx) specified within each model (From Naik et al.5).
Fig. 7 (a) Annual average surface •OH concentration, mean of14 models, for the year 2000. (b) Model-mean % change in surface •OHconcentrations in 2100 relative to 2000 for the IPCC RCP 2.6 emissionscenario; (c) same for RCP 8.5 scenarios (From Voulgarakis et al.7).
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 149–169 | 157

changes in precipitation patterns modifying wet depositionrates.
Several potentially major interactions are discussed here.Many other feedback processes are possible and may be plausi-ble but are not fully understood and cannot yet be quantifiedreliably. These are, by and large, outside the scope of thepresent assessment, except to the extent that we recognisetheir existence and therefore provide a cautionary note that thegeneral aspects of our assessment need to be evaluated care-fully for any given location with full consideration of theseadditional factors.
Ozone. Concentrations of ozone in urban environments aredetermined by a number of key factors. Firstly, the amount ofozone in the air entering the urban environment may beimportant. Then, within the urban airshed, reactions involvinga range of emitted chemicals, most notably VOCs and NOx,produce ozone as a result of UV-driven photochemistry. Thesechemicals arise from both anthropogenic and biogenicsources, the latter often being outside normal air quality man-agement. Finally, a number of processes such as dry depo-sition (loss at the surface) might remove ozone from theatmosphere.
Background ozone concentrations can be influenced bylong-range air transport, and this is likely to be an importantfactor in the future, transporting air between continents.122
The background ozone concentrations can also be altered bychanges in stratosphere/troposphere exchange and by thechanges in global atmospheric composition, notably somegreenhouse gases.119 By analysing the natural variability ofstratospheric transport, it has been estimated that changes instratospheric circulation due to climate change will lead toaround a 2% increase in tropospheric ozone in the northernmid-latitudes by the end of this century.123 Although small,the significant geographic extent implies that this might be anadditional factor affecting air quality.
Increased temperatures at ground level are expected toincrease biogenic emissions of reactive organics (e.g., ref. 124and 125). However, emissions may well depend on otherfactors as well, such as water stress on the plants.32 Indeed,changes in the climate, coupled to decreases in air quality, cansubstantially alter biogenic activity in ways that are difficult topredict.
Models suggest that other critical processes are also likelyto be altered by climate change. Variations in cloudiness canalter the rate of photochemical production of ozone. Increasedsurface heating can result in changes in atmospheric move-ment (wind speed, both horizontal and vertical). Cloudinesscan also be altered by human activity, with evidence that con-trols on air pollution have increased solar radiation at somelocations.126 Finally, changes in rainfall patterns and cloudi-ness can alter the rate of removal of both reactive precursorsand ozone itself.26 Estimates predict increasing ozone concen-trations at ground level throughout the 21st century, driven byall of these meteorological factors, which are regionally depen-dent. This can be offset by changes in emissions from humanactivity, which may either augment or reverse the overall trend,
depending on the levels of controls implemented.26,127 Signifi-cant regional air quality changes may result, even if onlyepisodically.128
Particles. The impact of climate change on aerosols remainshighly uncertain. As mentioned above, increasing tempera-tures will increase biogenic gas emissions. The oxidation ofthese compounds will produce aerosols. Additional wild firescould become more important as a source of particulatematter.129 These aerosols can scatter radiation and reducewarming (a negative feedback), and also impact upon cloudproperties.130 Changes in clouds in turn can alter the trans-formation (growth, chemistry) of aerosols in the atmosphere.In polluted environments the changes induced by climate willbe overwhelmed by anthropogenic emissions, but in locationswhere anthropogenic emissions are small these changes couldbe significant. However, the net effect of climate change onaerosols remains unclear.127,129 While there has been a lot ofwork in this area (e.g., on climate/aerosol feedback models131)the level of scientific understanding remains very low.
Biological interactions between air-pollutants and climatechange. Increased ambient temperatures may interact directlyor indirectly to exacerbate the effects of pollutants such as O3
in humans. A study of cardiovascular and respiratory mortalityin 2002 to 2006 in Buenos Aires showed a relative risk of1.0184 (95% confidence interval (CI) = 1.0139–1.0229) on thesame day for each 1 °C increase in temperature.132 Anotherstudy in several large cities in the UK reported that tempera-ture increased mortality from cardiovascular and respiratorydiseases and other non-accidental causes.133 These authorsalso reported that the mean mortality rate ratio for O3 was1.003 (95% CI = 1.001–1.005) per 10 μg m−3 increase in con-centration. On hot days (greater than the whole-year 95th
centile) this increased to 1.006 (95% CI = 1.002–1.009) but wasonly statistically significant for London. A study in older menin 2000 to 2008 in Boston showed that greater ambient temp-erature was associated with decreases in heart rate variabilityvia dysfunction of the autonomic nervous system.134 Thesewarm-season associations were significantly greater whenambient ozone concentrations were above the median butwere not affected by particulates (PM2.5). These studies areconsistent with earlier analyses of the 2003 European heatwave episode135,136 which attributed a significant fraction(20–60%) of excess mortality to the effects of elevated levels ofO3 and PM.
Change in climate also may affect human health indirectly.A study on allergic respiratory diseases and bronchial asthmashowed that while exacerbation is related to air-pollutants,amounts of allergen in the air are also important.137 The pres-ence of allergenic pollens in the atmosphere might be pro-longed by climate change and increase frequency and severityof these diseases.
Halogenated organic and other substitutes in the troposphere
Toxicity and risks of replacements for ozone depletingchemicals to humans and the environment. The United StatesEnvironmental Protection Agency (USEPA) has a regulatory
Perspective Photochemical & Photobiological Sciences
158 | Photochem. Photobiol. Sci., 2015, 14, 149–169 This journal is © The Royal Society of Chemistry and Owner Societies 2015

process for evaluating alternatives for Ozone Depleting Sub-stances (ODS) prior to their wide-spread use in the U.S.Anyone planning to market or produce a new substitute mustprovide 90 days’ advance notice to the Significant New Alterna-tives Policy (SNAP) program at USEPA of their intent as well asproviding health and safety information before introducing itinto interstate commerce in the U.S. Normally the health andsafety information will include information on chemical andphysical properties, flammability and basic toxicological infor-mation, and more recently, global warming potential. TheSNAP program reviews the information in the context of theproposed use, and issues one of 4 decisions: acceptable; accep-table subject to use conditions; acceptable subject to narroweduse limits; and unacceptable. This information on a particularcompound is continually updated so that compounds may beproposed for additional uses or additional information may beadded to the portfolio for a particular use that could changethe initial decision.
Updates on selected halocarbonsBrominated substances. Natural bromo-carbons bromoform
and dibromomethane are emitted from the oceans, and theiremission strengths and role in the atmosphere are becomingbetter understood. These compounds release bromine uponoxidation in the atmosphere that is generally observed asbromine monoxide (BrO). The presence of bromine can lead todepletion of ground level ozone.138 However, observations inother marine locations do not find such events occurring.139
In the tropics, where sources are believed to be the biggest andvertical motion is enhanced, it has been estimated that thesecompounds have a lifetime in the atmosphere of 1 to 3 weeks.However, even with this relatively short lifetime, over 90% ofthe bromine that is transported to the stratosphere comesfrom these species.140
Bromine monoxide (BrO) is also calculated to be a signifi-cant oxidant for dimethylsulfide (20%) in the clean marineenvironment of the southern hemisphere.141 This oxidation isa significant source of aerosol in this environment (nearly20% of the total) and so has direct impacts upon cloud for-mation and light scattering, and hence climate. A large frac-tion of this BrO is derived from sea salt, and thus is animportant part of the natural bromine background to whichanthropogenic brominated organics are added. The overall sig-nificance of changes due to climate, as noted above, is not yetknown.
n-Propyl bromide (C3H7Br), or 1-bromopropane, was intro-duced in the early to mid-1990s as an intermediate in avariety of closed commercial manufacturing processes forproducts such as pesticides, pharmaceuticals, and quaternaryammonium compounds. From the mid-1990s, it began to beused as a less toxic substitute for methylene chloride in openair uses such as vapour and immersion degreasing and clean-ing of electronics and metals.142 In 2003, it was first proposedby the SNAP program as an acceptable alternative for CFC-113and methyl chloroform in a limited number of specific appli-cations where emissions could be tightly controlled for bothenvironmental and exposure concerns. Specifically, these
included use as a solvent in industrial equipment for metals,electronics, and precision cleaning and in aerosol solvents andadhesive end-uses. However, the final rule issued in 2007143
allowed use only as a solvent for industrial equipment; otheruses such as aerosol solvents and adhesives are listed as un-acceptable by the Agency.
The SNAP program decisions were based on health datarelated to reproductive and neurological end-points for whichthey considered a work place standard of an 8-hour time-weight acceptable exposure limit (AEL) of 25 ppm to be accep-table.144 Subsequent to that information, reports have indi-cated that additional adverse effects have been added to thetoxicological dossier for n-propyl bromide including immuno-toxicity (significant decreases in a specific antibody) in rodentsfollowing 10 week inhalation exposures at levels of125–500 ppm (mice) or 1000 ppm (rat)145 and multi-site carci-nogenicity following two-year chronic inhalation exposures at250 or 500 ppm in rats and mice.142
Chlorinated substances. Chloroform (CHCl3) has a numberof poorly known natural sources as well as anthropogenicsources.146 Peat bogs may be a large unrecognized naturalsource of chloroform (10% of the total).147,148 Sources such asthese are likely to be sensitive to both climate and land usechange, and so represent an uncertainty in future predictions.
Production and consumption of carbon tetrachloride (CCl4)are regulated under the Montreal Protocol. Observations showthat atmospheric CCl4 mixing ratios are decreasing at a rateslower than expected from assumed phase-out schedules149,150
The sources and sinks of CCl4, and their uncertainties, arebeyond the scope of this paper, but it is probable that theslower decline is due to yet unidentified emissions. If suchsources involve personal exposure, e.g., in solvent use, poten-tial health effects may be anticipated.151
Chlorofluorinated substances. Historically the first com-pounds developed as replacements for refrigerants, the largestsector where CFCs were used, were saturated hydrochlorofluoro-carbons (HCFCs) and then saturated hydrofluorocarbons(HFCs), both classes of compounds which because of theirhydrogen content, were susceptible to attack from hydroxylradicals in the atmosphere, resulting in shorter atmosphericlifetimes than CFCs. Readers are referred to earlier reports fora review about the human and environmental risks for manyof these replacements.
There is one compound in this category, trans-1-chloro,3,3,3-trifluoropropene (HCFO 1233zd(E)), which is justnow being developed as a foam blowing agent, refrigerant, andsolvent. A recent toxicology study152 reported that HCFO1233zd(E) was not acutely toxic and was not associated withany genetic toxicity in a battery of tests. The compound had anacute 4-hour 50% lethal concentration value (LC50) of120 000 ppm in rats, and a no observed effect level (NOEL) incanine cardiac sensitization studies of 25 000 ppm. The heartwas identified as the apparent target organ on the basis of histo-pathological observations from a 2 week range finding studyin male and female rats exposed levels of 0, 2000, 5700 and20 000 ppm 6 hours per day for 5 days per week, Males at the
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 149–169 | 159

mid and high doses and females at the high dose developedmultifocal mononuclear infiltrates of cardiac tissue. In a12-week study at 4000, 10 000 and 15 000 ppm, 6 hours per dayfor 5 days per week, a NOEL/lowest observed adverse effectlevel for multifocal mononuclear infiltrates of the heart was4000 ppm. In a full inhalation developmental toxicity study atconcentrations of 0, 4000, 10 000 and 150 000 ppm, HCFO1233zd(E) was not associated with any effects on uterine, pla-cental or fetal weights, nor were there any fetal abnormalitiesobserved so that the study resulted in a NOEL for develop-mental toxicity of 15 000 ppm.
Fluorinated substances. Sulfuryl fluoride (SO2F2) is a substi-tute for the ozone-depleting fumigant methyl bromide used oncrops and soils. Several reports on its use in agriculture havebeen published recently153–156 and it will likely become morewidely used in the future. As previously stated,11 its atmo-spheric oxidation lifetime is estimated to be large (>300 years).However, SO2F2 is relatively soluble in water and is expected topartition into cloud water and ultimately rain out of the atmo-sphere with a half-life of about 2 weeks. The major sink is theoceans where the ultimate breakdown products are inorganicsulfate and fluoride. These breakdown products are not ofconcern for environment or health. However, SO2F2 has a largeglobal warming potential (GWP) and its use is likely toincrease in the future, so that monitoring of concentrations inthe atmosphere should continue.
Hydrochlorofluorocarbons and hydrofluorocarbons (HCFCsand HFCs): As has been discussed previously,11,157 several ofthe hydrochlorofluorocarbons (HCFCs) and hydrofluoro-carbons (HFCs) used as substitutes for ozone-depleting CFCsand a new fluorinated olefin (HFO) can break down intotrifluoroacetic acid (TFA). TFA is stable in the environment butis water soluble and accumulates in playas, land-lockedlakes, and the oceans where it combines with cations such assodium, potassium, calcium, and magnesium (Na+, K+, Ca2+,and Mg2+). More than 95% of the salts of TFA found in theoceans are naturally produced. These salts are inert and not oftoxicological or environmental concern in the small concen-trations (≈200 ng L−1) that are present in the oceans.
Perfluoro-n-butyl iodide (PFBI) is a compound being investi-gated as a substitute for CFCs. Its use is being explored as areplacement in aircraft maintenance operations as an alterna-tive cleaner for liquid and gaseous oxygen aerospace systemsand as a wipe cleaner solvent. A recent publication158 summar-ised earlier preliminary toxicity testing results in which anacute inhalation of no observed adverse effect level (NOAEL) of3900 ppm for cardiac sensitisation was established. Based onthe results of a preliminary 4-week inhalation toxicity test, a13-week inhalation study was conducted in which 15 male and10 female rats per group were exposed by inhalation via nose-only exposure for 6 hours per day and 5 days per week tonominal concentrations of 500, 1500, and 5000 ppm PFBI. The13-week inhalation exposure at 1500 ppm was associated withincreases in concentrations of thyroid hormones and thyroidstimulating hormone in serum of male and female rats. Theseconcentrations returned to control values following the 4-week
recovery period without exposure in males (the only animalsgiven a recovery period). The authors noted that, based onpublished information, they assumed that the return tonormal following the recovery period indicated that the hor-monal changes were only transient and not adverse and thatthe changes in female rats would also return to normal. Thusthe NOAEL was selected to be 1500 ppm. The 3900 ppm fromthe earlier work served as an occupational acute exposure limitfor PFBI, while the 1500 ppm NOAEL was converted to anoccupational exposure limit (OEL) by adjusting it to an 8-hour/day time-weighted average exposure and applying uncertaintyfactors for animal to human, inter-human variability andsubchronic to chronic extrapolation to arrive at a final value of40 ppm.
Modelling of susceptibility of the analogous compound(perfluoro-n-propyl iodide) to photolysis suggested a half-lifeof a few hours in sunlight. Based on rapid degradation, PFBIhas zero GWP and is not an ODS. Its atmospheric degradationis likely to produce TFA, analogously to many other fluorinatedorganics, e.g. some HCFCs and HFCs (see above).
Hydrofluoroolefins (HFOs). Among the other new compoundsbeing used as substitutes are those belonging to the classknown as hydrofluoroolefins (HFOs). Two examples currentlybeing developed for use in the refrigeration, foam-blowingand/or aerosol sectors, 1,3,3,3 tetrafluoropropene (HFO1234ze) and 2,3,3,3 tetrafluoropropene (HFO-1234yf) haverecently been characterised toxicologically.159–161 In the case ofHFO 1234ze, the compound was not acutely toxic at levels ashigh as 207 000 ppm following 4 hours of exposure andshowed no activity in a battery of genetic toxicity tests. Inaddition, the compound did not induce cardiac sensitizationat levels as high as 120 000. Following a 2 week range findingstudy at 5000, 20 000 and 50 000 ppm 6 hours per day, 5 daysper week, the liver and heart were identified as the targetorgans; however, in a subsequent 4 week study at 1000, 5000,10 000, and 15 000, and a 90 day study at 1500, 5000, and15 000, only the heart was identified as the target organ. Theassociated finding was multifocal mononuclear cell infiltrateswith no evidence of fibrosis and no evidence of increasedseverity with increased duration of exposure.
No new papers on the relevance of TFA to human healthand the environment have been published in the literaturesince the date of the previous assessment (2010). Thus, pro-jected future increased loadings of TFA to playas, land-lockedlakes, and the oceans due to continued use of HCFCs, HFCs,and replacement products are still judged to present negligiblerisks for aquatic organisms and humans.
Gaps in knowledge
A key air quality constituent is ozone, and any future changesin ozone have significant outcomes for both human andenvironmental health. However, the direction and the magni-tude of change in tropospheric ozone due to recovery of strato-spheric ozone are still under debate, may depend on location,
Perspective Photochemical & Photobiological Sciences
160 | Photochem. Photobiol. Sci., 2015, 14, 149–169 This journal is © The Royal Society of Chemistry and Owner Societies 2015

and need to be better quantified. Modulation by UV radiationwill modify the impacts of climate change and regulation oflocal emissions. This represents a significant gap in our under-standing. Computer modelling now has the potential toaddress this issue, but still requires the development andrefinement of parameterisations for physical and chemicalprocesses, as well as future scenarios under different socio-economic assumptions.
Particulate matter (aerosol) plays a significant role inclimate change and also in air quality. While the role of UVradiation in PM formation is known, the sensitivity of PM pro-perties to changes in UV radiation has not been sufficientlyquantified.
The oxidation capacity of the atmosphere remains poorlycharacterised in a number of environmentally sensitiveregions, with an order of magnitude difference betweenmeasurements and models. Both measurements and ourunderstanding of the key chemical processes have largeuncertainties. One example of this lack of understanding isthe uncertainty in future methane concentrations, withmodels predicting •OH driven lifetimes that differ by a factorof 2.
Acknowledgements
M. Shao is supported by the Natural Science Foundation ofCHINA for Outstanding Young Scholars (Grant No. 41125018).The National Center for Atmospheric Research is sponsored bythe National Science Foundation. S. Madronich was supportedby the U.S. Global Change Research Program. The partici-pation of Associate Professor S. Wilson was supported throughfunding from the Australian Government’s Ozone ScienceStrategy.
References
1 E. A. Ainsworth, C. R. Yendrek, S. Sitch, W. J. Collins andL. D. Emberson, The effects of tropospheric ozone on netprimary productivity and implications for climate change,Annu. Rev. Plant Biol., 2012, 63, 637–661.
2 D. D. Parrish, H. B. Singh, L. Molina and S. Madronich,Air quality progress in North American megacities: Areview, Atmos. Environ., 2011, 45, 7015–7025.
3 R. V. Martin, C. E. Sioris, K. Chance, T. B. Ryerson,T. H. Bertram, P. J. Wooldridge, R. C. Cohen,J. A. Neuman, A. Swanson and F. M. Flocke, Evaluation ofspace-based constraints on global nitrogen oxide emis-sions with regional aircraft measurements over and down-wind of eastern North America, J. Geophys. Res., [Atmos.],2006, 111, D15308.
4 D. T. Shindell, J. F. Lamarque, M. Schulz, M. Flanner,C. Jiao, M. Chin, P. J. Young, Y. H. Lee, L. Rotstayn,N. Mahowald, G. Milly, G. Faluvegi, Y. Balkanski,W. J. Collins, A. J. Conley, S. Dalsoren, R. Easter, S. Ghan,
L. Horowitz, X. Liu, G. Myhre, T. Nagashima, V. Naik,S. T. Rumbold, R. Skeie, K. Sudo, S. Szopa, T. Takemura,A. Voulgarakis, J. H. Yoon and F. Lo, Radiative forcing inthe ACCMIP historical and future climate simulations,Atmos. Chem. Phys., 2013, 13, 2939–2974.
5 V. Naik, A. Voulgarakis, A. M. Fiore, L. W. Horowitz,J. F. Lamarque, M. Lin, M. J. Prather, P. J. Young,D. Bergmann, P. J. Cameron-Smith, I. Cionni,W. J. Collins, S. B. Dalsøren, R. Doherty, V. Eyring,G. Faluvegi, G. A. Folberth, B. Josse, Y. H. Lee,I. A. MacKenzie, T. Nagashima, T. P. C. van Noije,D. A. Plummer, M. Righi, S. T. Rumbold, R. Skeie,D. T. Shindell, D. S. Stevenson, S. Strode, K. Sudo,S. Szopa and G. Zeng, Preindustrial to present daychanges in tropospheric hydroxyl radical and methanelifetime from the Atmospheric Chemistry and ClimateModel Intercomparison Project (ACCMIP), Atmos. Chem.Phys., 2013, 13, 5277–5298.
6 P. J. Young, A. T. Archibald, K. W. Bowman,J. F. Lamarque, V. Naik, D. S. Stevenson, S. Tilmes,A. Voulgarakis, O. Wild, D. Bergmann, P. Cameron-Smith,I. Cionni, W. J. Collins, S. B. Dalsøren, R. M. Doherty,V. Eyring, G. Faluvegi, L. W. Horowitz, B. Josse, Y. H. Lee,I. A. MacKenzie, T. Nagashima, D. A. Plummer, M. Righi,S. T. Rumbold, R. B. Skeie, D. T. Shindell, S. A. Strode,K. Sudo, S. Szopa and G. Zeng, Pre-industrial to end 21st
century projections of tropospheric ozone from theAtmospheric Chemistry and Climate Model Intercompari-son Project (ACCMIP), Atmos. Chem. Phys., 2013, 13, 2063–2090.
7 A. Voulgarakis, V. Naik, J. F. Lamarque, D. T. Shindell,P. J. Young, M. J. Prather, O. Wild, R. D. Field,D. Bergmann, P. Cameron-Smith, I. Cionni, W. J. Collins,S. B. Dalsøren, R. M. Doherty, V. Eyring, G. Faluvegi,G. A. Folberth, L. W. Horowitz, B. Josse, I. A. MacKenzie,T. Nagashima, D. A. Plummer, M. Righi, S. T. Rumbold,D. S. Stevenson, S. A. Strode, K. Sudo, S. Szopa andG. Zeng, Analysis of present day and future OH andmethane lifetime in the ACCMIP simulations, Atmos.Chem. Phys., 2013, 13, 2563–2587.
8 A. Hilboll, A. Richter and J. P. Burrows, Long-termchanges of tropospheric NO2 over megacities derived frommultiple satellite instruments, Atmos. Chem. Phys., 2013,13, 4145–4169.
9 IPCC, Climate Change 2014: Mitigation of ClimateChange. Contribution of Working Group III to the FifthAssessment Report, Intergovernmental Panel on ClimateChange Report No., Geneva, 2014.
10 UNEP, Environmental effects of ozone depletion and itsinteractions with climate change: Progress Report 2009,Photochem. Photobiol. Sci., 2010, 9, 275–294.
11 X. Tang, S. R. Wilson, K. R. Solomon, M. Shao andS. Madronich, Changes in tropospheric composition andair quality due to stratospheric ozone depletion and inter-actions with changes in climate, Photochem. Photobiol.Sci., 2011, 10, 280–291.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 149–169 | 161

12 S. C. Anenberg, J. Schwartz, D. Shindell, M. Amann,G. Faluvegi, Z. Klimont, G. Janssens-Maenhout, L. Pozzoli,R. Van Dingenen, E. Vignati, L. Emberson, N. Z. Muller,J. J. West, M. Williams, V. Demkine, W. K. Hicks,J. Kuylenstierna, F. Raes and V. Ramanathan, Global airquality and health co-benefits of mitigating near-termclimate change through methane and black carbon emis-sion controls, Environ. Health Perspect., 2012, 120, 831–839.
13 S. C. Anenberg, L. W. Horowitz, D. Q. Tong and J. J. West,An estimate of the global burden of anthropogenic ozoneand fine particulate matter on premature human mor-tality using atmospheric modeling, Environ. Health Per-spect., 2010, 118, 1189–1195.
14 S. C. Anenberg, J. J. West, L. W. Horowitz and D. Q. Tong,The global burden of air pollution on mortality: Anenberget al. respond, Environ. Health Perspect., 2011, 119, A158–A159.
15 S. P. de Almeida, E. Casimiro and J. Calheiros, Short-term association between exposure to ozone andmortality in Oporto, Portugal, Environ. Res., 2011, 111,406–410.
16 Y. L. Hsieh, Y. H. Yang, T. N. Wu and C. Y. Yang, Air pol-lution and hospital admissions for myocardial infarctionin a subtropical city: Taipei, Taiwan, J. Toxicol. Environ.Health, Part A, 2010, 73, 757–765.
17 I. Hůnová, M. Malý, J. Řezáčová and M. Braniš, Associ-ation between ambient ozone and health outcomes inPrague, Int. Arch. Occup. Environ. Hlth., 2012, 86, 89–97.
18 S. S. Lim, T. Vos, A. D. Flaxman, G. Danaei, K. Shibuya,H. Adair-Rohani, M. A. AlMazroa, M. Amann,H. R. Anderson, K. G. Andrews, M. Aryee, C. Atkinson,L. J. Bacchus, A. N. Bahalim, K. Balakrishnan, J. Balmes,S. Barker-Collo, A. Baxter, M. L. Bell, J. D. Blore, F. Blyth,C. Bonner, G. Borges, R. Bourne, M. Boussinesq,M. Brauer, P. Brooks, N. G. Bruce, B. Brunekreef, C. Bryan-Hancock, C. Bucello, R. Buchbinder, F. Bull, R. T. Burnett,T. E. Byers, B. Calabria, J. Carapetis, E. Carnahan,Z. Chafe, F. Charlson, H. Chen, J. S. Chen, A. T.-A. Cheng,J. C. Child, A. Cohen, K. E. Colson, B. C. Cowie, S. Darby,S. Darling, A. Davis, L. Degenhardt, F. Dentener, D. C. DesJarlais, K. Devries, M. Dherani, E. L. Ding, E. R. Dorsey,T. Driscoll, K. Edmond, S. E. Ali, R. E. Engell, P. J. Erwin,S. Fahimi, G. Falder, F. Farzadfar, A. Ferrari,M. M. Finucane, S. Flaxman, F. G. R. Fowkes,G. Freedman, M. K. Freeman, E. Gakidou, S. Ghosh,E. Giovannucci, G. Gmel, K. Graham, R. Grainger,B. Grant, D. Gunnell, H. R. Gutierrez, W. Hall,H. W. Hoek, A. Hogan, H. D. Hosgood Iii, D. Hoy, H. Hu,B. J. Hubbell, S. J. Hutchings, S. E. Ibeanusi, G. L. Jacklyn,R. Jasrasaria, J. B. Jonas, H. Kan, J. A. Kanis,N. Kassebaum, N. Kawakami, Y.-H. Khang,S. Khatibzadeh, J.-P. Khoo, C. Kok, et al., A comparativerisk assessment of burden of disease and injury attribu-table to 67 risk factors and risk factor clusters in 21regions, 1990–2010: a systematic analysis for the Global
Burden of Disease Study 2010, Lancet, 2012, 380, 2224–2260.
19 OECD, OECD Environmental Outlook to 2050: The Conse-quences of Inaction: Key Findings on Health and Environ-ment, Organization for Economic Cooperation andDevelopment Report No., Paris, 2012. http://www.oecd.org/environment/outlookto2050.
20 Y. Fang, V. Naik, L. W. Horowitz and D. L. Mauzerall, Airpollution and associated human mortality: the role of airpollutant emissions, climate change and methane con-centration increases from the preindustrial period topresent, Atmos. Chem. Phys., 2013, 13, 1377–1394.
21 N. Fann, A. D. Lamson, S. C. Anenberg, K. Wesson,D. Risley and B. J. Hubbell, Estimating the national publichealth burden associated with exposure to ambient PM2.5and ozone, Risk Anal., 2012, 32, 81–95.
22 J. T. Lee, Y. S. Cho and J. Y. Son, Relationship betweenambient ozone concentrations and daily hospital admis-sions for childhood asthma/atopic dermatitis in two citiesof Korea during 2004-2005, Int. J. Environ. Health Res.,2010, 20, 1–11.
23 G. G. Kaplan, D. Tanyingoh, E. Dixon, M. Johnson,A. J. Wheeler, R. P. Myers, S. Bertazzon, V. Saini,K. Madsen, S. Ghosh and P. J. Villeneuve, Ambient ozoneconcentrations and the risk of perforated and nonperfo-rated appendicitis: a multicity case-crossover study,Environ. Health Perspect., 2013, 121, 939–943.
24 R. E. Dales, S. Cakmak and C. B. Vidal, Air pollution andhospitalization for venous thromboembolic disease inChile, J. Thromb. Haemost., 2010, 8, 669–674.
25 C. M. Wong, T. Q. Thach, P. Y. Chau, E. K. Chan,R. Y. Chung, C. Q. Ou, L. Yang, J. S. Peiris, G. N. Thomas,T. H. Lam, T. W. Wong and A. J. Hedley, Part4. Interaction between air pollution and respiratoryviruses: time-series study of daily mortality and hospitaladmissions in Hong Kong, Research Report, 2010, 283–362.
26 H. Lei, D. J. Wuebbles and X.-Z. Liang, Projected risk ofhigh ozone episodes in 2050, Atmos. Environ., 2012, 59,567–577.
27 Royal Society, Ground-level ozone in the 21st century:future trends, impacts and policy implications, The RoyalSociety Report No. RS Policy document 15/08, London,UK, October 2008 2008, p. 148. http://royalsociety.org/policy/publications/2008/ground-level-ozone/.
28 S. Wilkinson, G. Mills, R. Illidge and W. J. Davies, How isozone pollution reducing our food supply?, J. Exp. Bot.,2012, 63, 527–536.
29 K. W. Jaggard, A. Qi and E. S. Ober, Possible changes toarable crop yields by 2050, Philos. Trans. R. Soc. London,Ser. B, 2010, 365, 2835–2851.
30 C. P. Leisner and E. A. Ainsworth, Quantifying the effectsof ozone on plant reproductive growth and development,Glob. Change Biol., 2012, 18, 606–616.
31 A. M. Betzelberger, C. R. Yendrek, J. Sun, C. P. Leisner,R. L. Nelson, D. R. Ort and E. A. Ainsworth, Ozone
Perspective Photochemical & Photobiological Sciences
162 | Photochem. Photobiol. Sci., 2015, 14, 149–169 This journal is © The Royal Society of Chemistry and Owner Societies 2015

exposure response for U.S. soybean cultivars: linearreductions in photosynthetic potential, biomass, andyield, Plant Physiol., 2012, 160, 1827–1839.
32 S. Fares, R. Vargas, M. Detto, A. H. Goldstein, J. Karlik,E. Paoletti and M. Vitale, Tropospheric ozone reducescarbon assimilation in trees: estimates from analysis ofcontinuous flux measurements, Glob. Change Biol., 2013,19, 2427–2443.
33 IPCC, Special Report on Emissions Scenarios: A SpecialReport of Working Group III of the IntergovernmentalPanel on Climate Change, Intergovernmental Panel onClimate Change Report No., Geneva, 2000.
34 S. Avnery, D. L. Mauzerall, J. Liu and L. W. Horowitz,Global crop yield reductions due to surface ozoneexposure: 2. Year 2030 potential crop production lossesand economic damage under two scenarios of O3 pol-lution, Atmos. Environ., 2011, 45, 2297–2309.
35 Y. Feng, X. Lin, Y. Yu, H. Zhang, H. Chu and J. Zhu,Elevated ground-level O3 negatively influences paddymethanogenic archaeal community, Sci. Rep., 2013, 3,3193.
36 A. J. Haagen-Smit, C. E. Bradley and M. M. Fox, Ozone for-mation in photochemical oxidation of organic substances,Ind. Eng. Chem. Res., 1953, 45, 2086–2089.
37 R. L. McKenzie, P. J. Aucamp, A. F. Bais, L. O. Björn,M. Ilyas and S. Madronich, Ozone depletion and climatechange: Impacts on UV radiation, Photochem. Photobiol.Sci., 2011, 10, 182–198.
38 A. Colette, C. Granier, Ø. Hodnebrog, H. Jakobs,A. Maurizi, A. Nyiri, B. Bessagnet, A. D’Angiola,M. D’Isidoro, M. Gauss, F. Meleux, M. Memmesheimer,A. Mieville, L. Rouïl, F. Russo, S. Solberg, F. Stordal andF. Tampieri, Air quality trends in Europe over the pastdecade: a first multi-model assessment, Atmos. Chem.Phys., 2011, 11, 11657–11678.
39 R. C. Wilson, Z. L. Fleming, P. S. Monks, G. Clain,S. Henne, I. B. Konovalov, S. Szopa and L. Menut, Haveprimary emission reduction measures reduced ozoneacross Europe? An analysis of European rural backgroundozone trends 1996–2005, Atmos. Chem. Phys., 2012, 12,437–454.
40 T. Zhu, M. Melamed, D. Parrish, M. Gauss, L. Gallardo,M. Lawrence, A. Knare and C. Liousse, WMO/IGACImpacts of Megacities on Air Pollution and Climate,World Meteorological Organization Report No. WorldMeteorological Organization (WMO) Global AtmosphereWatch (GAW) Report No. 205, Geneva, January 2013,http://www.wmo.int/pages/prog/arep/gaw/gaw-reports.html.
41 Q. Zhang, B. Yuan, M. Shao, X. Wang, S. Lu, K. Lu,M. Wang, L. Chen, C. C. Chang and S. C. Liu, Variationsof ground-level O3 and its precursors in Beijing in sum-mertime between 2005 and 2011, Atmos. Chem. Phys.,2014, 14, 6089–6101.
42 Y. Wang, P. Konopka, Y. Liu, H. Chen, R. Müller,F. Plöger, M. Riese, Z. Cai and D. Lü, Tropospheric ozone
trend over Beijing from 2002–2010: ozonesonde measure-ments and modeling analysis, Atmos. Chem. Phys., 2012,12, 8389–8399.
43 D. M. Lal, S. D. Ghude, S. D. Patil, S. H. Kulkarni, C. Jena,S. Tiwari and M. K. Srivastava, Tropospheric ozone andaerosol long-term trends over the Indo-Gangetic Plain(IGP), India, Atmos. Res., 2012, 116, 82–92.
44 T. Wang, W. Nie, J. Gao, L. K. Xue, X. M. Gao, X. F. Wang,J. Qiu, C. N. Poon, S. Meinardi, D. Blake, S. L. Wang,A. J. Ding, F. H. Chai, Q. Z. Zhang and W. X. Wang, Airquality during the 2008 Beijing Olympics: secondarypollutants and regional impact, Atmos. Chem. Phys., 2010,10, 7603–7615.
45 R. G. Derwent, A. J. Manning, P. G. Simmonds, T. G. Spainand S. O’Doherty, Analysis and interpretation of 25 yearsof ozone observations at the Mace Head AtmosphericResearch Station on the Atlantic Ocean coast of Irelandfrom 1987 to 2012, Atmos. Environ., 2013, 80, 361–368.
46 O. R. Cooper, D. D. Parrish, A. Stohl, M. Trainer,P. Nedelec, V. Thouret, J. P. Cammas, S. J. Oltmans,B. J. Johnson, D. Tarasick, T. Leblanc, I. S. McDermid,D. Jaffe, R. Gao, J. Stith, T. Ryerson, K. Aikin, T. Campos,A. Weinheimer and M. A. Avery, Increasing springtimeozone mixing ratios in the free troposphere over westernNorth America, Nature, 2010, 463, 344–348.
47 D. D. Parrish, K. S. Law, J. Staehelin, R. Derwent,O. R. Cooper, H. Tanimoto, A. Volz-Thomas, S. Gilge,H. E. Scheel, M. Steinbacher and E. Chan, Lowertropospheric ozone at northern midlatitudes: Changingseasonal cycle, Geophys. Res. Lett., 2013, 40, 1631–1636.
48 S. J. Oltmans, A. S. Lefohn, D. Shadwick, J. M. Harris,H. E. Scheel, I. Galbally, D. W. Tarasick, B. J. Johnson,E. G. Brunke, H. Claude, G. Zeng, S. Nichol, F. Schmidlin,J. Davies, E. Cuevas, A. Redondas, H. Naoe, T. Nakano andT. Kawasato, Recent tropospheric ozone changes – Apattern dominated by slow or no growth, Atmos. Environ.,2013, 67, 331–351.
49 P. G. Hess and R. Zbinden, Stratospheric impact on tropo-spheric ozone variability and trends: 1990-2009, Atmos.Chem. Phys., 2013, 13, 649–674.
50 M. Lin, L. W. Horowitz, S. J. Oltmans, A. M. Fiore andS. Fan, Tropospheric ozone trends at Mauna Loa Observa-tory tied to decadal climate variability, Nat. Geosci., 2014,7, 136–143.
51 D. S. Stevenson, P. J. Young, V. Naik, J. F. Lamarque,D. T. Shindell, A. Voulgarakis, R. B. Skeie, S. B. Dalsoren,G. Myhre, T. K. Berntsen, G. A. Folberth, S. T. Rumbold,W. J. Collins, I. A. MacKenzie, R. M. Doherty, G. Zeng,T. P. C. van Noije, A. Strunk, D. Bergmann, P. Cameron-Smith, D. A. Plummer, S. A. Strode, L. Horowitz, Y. H. Lee,S. Szopa, K. Sudo, T. Nagashima, B. Josse, I. Cionni,M. Righi, V. Eyring, A. Conley, K. W. Bowman, O. Wildand A. Archibald, Tropospheric ozone changes, radiativeforcing and attribution to emissions in the AtmosphericChemistry and Climate Model Intercomparison Project(ACCMIP), Atmos. Chem. Phys., 2013, 13, 3063–3085.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 149–169 | 163

52 G. Zeng, O. Morgenstern, P. Braesicke and J. A. Pyle,Impact of stratospheric ozone recovery on troposphericozone and its budget, Geophys. Res. Lett., 2010, 37.
53 H. Zhang, S. Wu, Y. Huang and Y. Wang, Effects of strato-spheric ozone recovery on photochemistry and ozone airquality in the troposphere, Atmos. Chem. Phys., 2014, 14,4079–4089.
54 S. C. Liu and M. Trainer, Responses of the troposphericozone and odd hydrogen radicals to column ozonechange, J. Atmos. Chem., 1988, 6, 221–233.
55 A. M. Thompson, R. W. Stewart, M. A. Owens andJ. A. Herwehe, Sensitivity of tropospheric oxidants toglobal chemical and climate change, Atmos. Environ.,1989, 23, 519–532.
56 D. D. Parrish, J. F. Lamarque, V. Naik, L. Horowitz,D. T. Shindell, J. Staehelin, R. Derwent, O. R. Cooper,H. Tanimoto, A. Volz-Thomas, S. Gilge, H. E. Scheel,M. Steinbacher and M. Fröhlich, Long-term changes inlower tropospheric baseline ozone concentrations: Com-paring chemistry-climate models and observations atnorthern mid-latitudes, J. Geophys. Res., [Atmos.], 2014,2013JD021435.
57 USEPA, Integrated Science Assessment for ParticulateMatter, U.S. Environmental Protection Agency, Office ofResearch and Development Report No. EPA/600/R-08/139F, Research Triangle Park, NC, 2010, p. 2228.
58 Y. Chen, A. Ebenstein, M. Greenstone and H. Li, Evidenceon the impact of sustained exposure to air pollution onlife expectancy from China’s Huai River policy, Proc. Natl.Acad. Sci. U. S. A., 2013, 110, 12936–12941.
59 A. van Donkelaar, R. Martin, R. C. Levy, A. M. da Silva,M. Krzyzanowski, N. E. Chubarova, E. Semutnikova andA. J. Cohen, Satellite-based estimates of ground-levelfine particulate matter during extreme events: A casestudy of the Moscow fires, Atmos. Environ., 2011, 45, 6225–6232.
60 A. W. Correia, C. A. Pope, 3rd, D. W. Dockery, Y. Wang,M. Ezzati and F. Dominici, Effect of air pollution controlon life expectancy in the United States: an analysis of 545U.S. counties for the period from 2000 to 2007, Epidemiol.,2013, 24, 23–31.
61 J. T. Spickett, H. L. Brown and K. Rumchev, Climatechange and air quality: the potential impact on health,Asia. Pac. J. Public. Health., 2011, 23, 37S–345.
62 E. S. Post, A. Grambsch, C. Weaver, P. Morefield, J. Huang,L. Y. Leung, C. G. Nolte, P. Adams, X. Z. Liang, J. H. Zhuand H. Mahoney, Variation in estimated ozone-relatedhealth Impacts of climate change due to modelingchoices and assumptions, Environ. Health Perspect., 2012,120, 1559–1564.
63 R. L. Prueitt and R. L. Goodman, The global burden ofozone on respiratory mortality: No clear evidence forassociation, Environ. Health Perspect., 2011, 119, A158.
64 A. F. Bais, R. L. McKenzie, P. J. Aucamp, M. Ilyas,S. Madronich, G. Bernhard and K. Tourpali, Ozonedepletion and climate change: Impacts on UV radiation,
Photochem. Photobiol. Sci., 2014, 14, DOI: 10.1039/c1034pp90032d.
65 J. H. Seinfeld and S. N. Pandis, Atmospheric Chemistry andPhysics: From Air Pollution to Climate Change, John Wiley &Sons, Hoboken, NJ, USA, 2006.
66 X. Bi, B. R. Simoneit, G. Sheng, S. Ma and J. Fu, Compo-sition and major sources of organic compounds in urbanaerosols, Atmos. Res., 2008, 88, 256–265.
67 J. L. Jimenez, M. R. Canagaratna, N. M. Donahue,A. S. H. Prevot, Q. Zhang, J. H. Kroll, P. F. DeCarlo,J. D. Allan, H. Coe, N. L. Ng, A. C. Aiken, K. S. Docherty,I. M. Ulbrich, A. P. Grieshop, A. L. Robinson, J. Duplissy,J. D. Smith, K. R. Wilson, V. A. Lanz, C. Hueglin, Y. L. Sun,J. Tian, A. Laaksonen, T. Raatikainen, J. Rautiainen,P. Vaattovaara, M. Ehn, M. Kulmala, J. M. Tomlinson,D. R. Collins, M. J. E. Cubison, J. Dunlea, J. A. Huffman,T. B. Onasch, M. R. Alfarra, P. I. Williams, K. Bower,Y. Kondo, J. Schneider, F. Drewnick, S. Borrmann,S. Weimer, K. Demerjian, D. Salcedo, L. Cottrell,R. Griffin, A. Takami, T. Miyoshi, S. Hatakeyama,A. Shimono, J. Y. Sun, Y. M. Zhang, K. Dzepina,J. R. Kimmel, D. Sueper, J. T. Jayne, S. C. Herndon,A. M. Trimborn, L. R. Williams, E. C. Wood,A. M. Middlebrook, C. E. Kolb, U. Baltensperger andD. R. Worsnop, Evolution of organic aerosols in theatmosphere, Science, 2009, 326, 1525–1529.
68 N. M. Donahue, K. M. Henry, T. F. Mentel, A. Kiendler-Scharr, C. Spindler, B. Bohn, T. Brauers, H. P. Dorn,H. Fuchs, R. Tillmann, A. Wahner, H. Saathoff,K.-H. Naumann, O. Möhler, T. Leisner, L. Müller,M.-C. Reinnig, T. Hoffmann, K. Salo, M. Hallquist,M. Frosch, M. Bilde, T. Tritscher, P. Barmet, A. P. Praplan,P. F. DeCarlo, J. Dommen, A. S. H. Prévôt andU. Baltensperger, Aging of biogenic secondary organicaerosol via gas-phase OH radical reactions, Proc. Natl.Acad. Sci. U. S. A., 2012, 109, 13503–13508.
69 A. Hodzic, S. Madronich, B. Aumont, J. Lee-Taylor, T. Karl,M. Camredon and C. Mouchel-Vallon, Limited influenceof dry deposition of semivolatile organic vapors on sec-ondary organic aerosol formation in the urban plume,Geophys. Res. Lett., 2013, 40, 3302–3307.
70 J. H. Kroll, N. M. Donahue, J. L. Jimenez, S. H. Kessler,M. R. Canagaratna, K. R. Wilson, K. E. Altieri,L. R. Mazzoleni, A. S. Wozniak, H. Bluhm, E. R. Mysak,J. D. Smith, C. E. Kolb and D. R. Worsnop, Carbon oxi-dation state as a metric for describing the chemistry ofatmospheric organic aerosol, Nat. Chem., 2011, 3, 133–139.
71 C. L. Heald, J. H. Kroll, J. L. Jimenez, K. S. Docherty,P. F. DeCarlo, A. C. Aiken, Q. Chen, S. T. Martin,D. K. Farmer and P. Artaxo, A simplified description ofthe evolution of organic aerosol composition in theatmosphere, Geophys. Res. Lett., 2010, 37, L08803.
72 C. D. Cappa and K. R. Wilson, Multi-generation gas-phaseoxidation, equilibrium partitioning, and the formationand evolution of secondary organic aerosol, Atmos. Chem.Phys., 2012, 12, 9505–9528.
Perspective Photochemical & Photobiological Sciences
164 | Photochem. Photobiol. Sci., 2015, 14, 149–169 This journal is © The Royal Society of Chemistry and Owner Societies 2015

73 J. F. Pankow and K. C. Barsanti, The carbon number-polarity grid: A means to manage the complexity of themix of organic compounds when modeling atmosphericorganic particulate matter, Atmos. Environ., 2009, 43,2829–2835.
74 B. Croft, J. R. Pierce, R. V. Martin, C. Hoose andU. Lohmann, Uncertainty associated with convective wetremoval of entrained aerosols in a global climate model,Atmos. Chem. Phys., 2012, 12, 10725–10748.
75 B. Croft, J. R. Pierce and R. V. Martin, Interpreting aerosollifetimes using the GEOS-Chem model and constraintsfrom radionuclide measurements, Atmos. Chem. Phys.,2014, 14, 4313–4325.
76 N. I. Kristiansen, A. Stohl and G. Wotawa, Atmosphericremoval times of the aerosol-bound radionuclides 137Csand 131I measured after the Fukushima Dai-ichi nuclearaccident - a constraint for air quality and climate models,Atmos. Chem. Phys., 2012, 12, 10759–10769.
77 R. Zalakeviciute, M. L. Alexander, E. Allwine, J. L. Jimenez,B. T. Jobson, L. T. Molina, E. Nemitz, S. N. Pressley,T. M. VanReken, I. M. Ulbrich, E. Velasco and B. K. Lamb,Chemically-resolved aerosol eddy covariance flux measure-ments in urban Mexico City during MILAGRO 2006,Atmos. Chem. Phys., 2012, 12, 7809–7823.
78 J. L. Hand, B. A. Schichtel, M. Pitchford, W. C. Malm andN. H. Frank, Seasonal composition of remote and urbanfine particulate matter in the United States, J. Geophys.Res., [Atmos.], 2012, 117, D05209.
79 M. Amann, Z. Klimont and F. Wagner, Regional andGlobal Emissions of Air Pollutants: Recent Trends andFuture Scenarios, Annu. Rev. Environ. Resources, 2013, 38,31–55.
80 S. J. Smith and T. C. Bond, Two hundred fifty years ofaerosols and climate: the end of the age of aerosols,Atmos. Chem. Phys., 2014, 14, 537–549.
81 D. E. Heard and M. J. Pilling, Measurement of OH andHO2 in the troposphere, Chem. Rev., 2003, 103, 5163–5198.
82 D. Stone, L. K. Whalley and D. E. Heard, Tropospheric OHand HO2 radicals: field measurements and model com-parisons, Chem. Soc. Rev., 2012, 41, 6348–6404.
83 X. Ren, W. H. Brune, J. Mao, M. J. Mitchell, R. L. Lesher,J. B. Simpas, A. R. Metcalf, J. J. Schwab, C. Cai and Y. Li,Behavior of OH and HO2 in the winter atmosphere inNew York City, Atmos. Environ., 2006, 40, 252–263.
84 Y. Kanaya, R. Cao, H. Akimoto, M. Fukuda, Y. Komazaki,Y. Yokouchi, M. Koike, H. Tanimoto, N. Takegawa andY. Kondo, Urban photochemistry in central Tokyo:1. Observed and modeled OH and HO2 radical concen-trations during the winter and summer of 2004,J. Geophys. Res., [Atmos.], 2007, 112, DOI: 10.1029/2007JD008670.
85 S. Dusanter, D. Vimal, P. S. Stevens, R. Volkamer,L. T. Molina, A. Baker, S. Meinardi, D. Blake, P. Sheehy,A. Merten, R. Zhang, J. Zheng, E. C. Fortner,W. Junkermann, M. Dubey, T. Rahn, B. Eichinger,
P. Lewandowski, J. Prueger and H. Holder, Measurementsof OH and HO2 concentrations during the MCMA-2006field campaign – Part 2: Model comparison and radicalbudget, Atmos. Chem. Phys., 2009, 9, 6655–6675.
86 G. Li, W. Lei, M. Zavala, R. Volkamer, S. Dusanter,P. Stevens and L. T. Molina, Impacts of HONO sources onthe photochemistry in Mexico City during theMCMA-2006/MILAGO Campaign, Atmos. Chem. Phys.,2010, 10, 6551–6567.
87 S. Chen, X. Ren, J. Mao, Z. Chen, W. H. Brune, B. Lefer,B. Rappenglück, J. Flynn, J. Olson and J. H. Crawford, Acomparison of chemical mechanisms based onTRAMP-2006 field data, Atmos. Environ., 2010, 44, 4116–4125.
88 D. Stone, M. J. Evans, R. Commane, T. Ingham,C. F. A. Floquet, J. B. McQuaid, D. M. Brookes,P. S. Monks, R. Purvis, J. F. Hamilton, J. Hopkins, J. Lee,A. C. Lewis, D. Stewart, J. G. Murphy, G. Mills, D. Oram,C. E. Reeves and D. E. Heard, HOx observations over WestAfrica during AMMA: Impact of isoprene and NOx, Atmos.Chem. Phys., 2010, 10, 9415–9429.
89 S. M. Griffith, R. F. Hansen, S. Dusanter, P. S. Stevens,M. Alaghmand, S. B. Bertman, M. A. Carroll, M. Erickson,M. Galloway, N. Grossberg, J. Hottle, J. Hou, B. T. Jobson,A. Kammrath, F. N. Keutsch, B. L. Lefer, L. H. Mielke,A. O’Brien, P. B. Shepson, M. Thurlow, W. Wallace,N. Zhang and X. L. Zhou, OH and HO2 radical chemistryduring PROPHET 2008 and CABINEX 2009 – Part 1:Measurements and model comparison, Atmos. Chem.Phys., 2013, 13, 5403–5423.
90 D. Kubistin, H. Harder, M. Martinez, M. Rudolf,R. Sander, H. Bozem, G. Eerdekens, H. Fischer, C. Gurk,T. Klüpfel, R. Königstedt, U. Parchatka, C. L. Schiller,A. Stickler, D. Taraborrelli, J. Williams and J. Lelieveld,Hydroxyl radicals in the tropical troposphere over theSuriname rainforest: comparison of measurements withthe box model MECCA, Atmos. Chem. Phys., 2010, 10,9705–9728.
91 J. Lelieveld, T. M. Butler, J. N. Crowley, T. J. Dillon,H. Fischer, L. Ganzeveld, H. Harder, M. G. Lawrence,M. Martinez, D. Taraborrelli and J. Williams, Atmosphericoxidation capacity sustained by a tropical forest, Nature,2008, 452, 737–740.
92 M. Martinez, H. Harder, D. Kubistin, M. Rudolf,H. Bozem, G. Eerdekens, H. Fischer, T. Klüpfel, C. Gurk,R. Königstedt, U. Parchatka, C. L. Schiller, A. Stickler,J. Williams and J. Lelieveld, Hydroxyl radicals in the tropi-cal troposphere over the Suriname rainforest: airbornemeasurements, Atmos. Chem. Phys., 2010, 10, 3759–3773.
93 T. A. M. Pugh, A. R. MacKenzie, C. N. Hewitt, B. Langford,P. M. Edwards, K. L. Furneaux, D. E. Heard, J. R. Hopkins,C. E. Jones, A. Karunaharan, J. Lee, G. Mills, P. Misztal,S. Moller, P. S. Monks and L. K. Whalley, Simulatingatmospheric composition over a South-East Asian tropicalrainforest: performance of a chemistry box model, Atmos.Chem. Phys., 2010, 10, 279–298.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 149–169 | 165

94 L. K. Whalley, P. M. Edwards, K. L. Furneaux, A. Goddard,T. Ingham, M. J. Evans, D. Stone, J. R. Hopkins,C. E. Jones, A. Karunaharan, J. D. Lee, A. C. Lewis,P. S. Monks, S. J. Moller and D. E. Heard, Quantifying themagnitude of a missing hydroxyl radical source in a tropi-cal rainforest, Atmos. Chem. Phys., 2011, 11, 7223–7233.
95 A. Hofzumahaus, F. Rohrer, K. Lu, B. Bohn, T. Brauers,C. C. Chang, H. Fuchs, F. Holland, K. Kita, Y. Kondo,X. Li, S. Lou, M. Shao, L. Zeng, A. Wahner and Y. Zhang,Amplified trace gas removal in the troposphere, Science,2009, 324, 1702–1704.
96 K. D. Lu, F. Rohrer, F. Holland, H. Fuchs, B. Bohn,T. Brauers, C. C. Chang, R. Häseler, M. Hu, K. Kita,Y. Kondo, X. Li, S. R. Lou, S. Nehr, M. Shao, L. M. Zeng,A. Wahner, Y. H. Zhang and A. Hofzumahaus, Observationand modelling of OH and HO2 concentrations in the PearlRiver Delta 2006: a missing OH source in a VOC richatmosphere, Atmos. Chem. Phys., 2012, 12, 1541–1569.
97 K. D. Lu, A. Hofzumahaus, F. Holland, B. Bohn,T. Brauers, H. Fuchs, M. Hu, R. Häseler, K. Kita,Y. Kondo, X. Li, S. R. Lou, A. Oebel, M. Shao, L. M. Zeng,A. Wahner, T. Zhu, Y. H. Zhang and F. Rohrer, MissingOH source in a suburban environment near Beijing:observed and modelled OH and HO2 concentrationsin summer 2006, Atmos. Chem. Phys., 2013, 13, 1057–1080.
98 F. Paulot, J. D. Crounse, H. G. Kjaergaard, J. H. Kroll,J. H. Seinfeld and P. O. Wennberg, Isoprene photooxid-ation: new insights into the production of acids andorganic nitrates, Atmos. Chem. Phys., 2009, 9, 1479–1501.
99 J. Peeters and J. F. Műller, HOx radical regeneration in iso-prene oxidation via peroxy radical isomerisations. II:experimental evidence and global impact, Phys. Chem.Chem. Phys., 2010, 12, 14227–14235.
100 H. Fuchs, A. Hofzumahaus, F. Rohrer, B. Bohn,T. Brauers, H. P. Dorn, R. Haseler, F. Holland,M. Kaminski, X. Li, K. Lu, S. Nehr, R. Tillmann,R. Wegener and A. Wahner, Experimental evidence forefficient hydroxyl radical regeneration in isopreneoxidation, Nat. Geosci., 2013, 6, 1023–1026.
101 H. Fuchs, H. P. Dorn, M. Bachner, B. Bohn, T. Brauers,S. Gomm, A. Hofzumahaus, F. Holland, S. Nehr,F. Rohrer, R. Tillmann and A. Wahner, Comparison of OHconcentration measurements by DOAS and LIF duringSAPHIR chamber experiments at high OH reactivity andlow NO concentration, Atmos. Measure. Technol., 2012, 5,1611–1626.
102 X. Ren, J. Mao, W. H. Brune, C. A. Cantrell, R. L. MauldinIii, R. S. Hornbrook, E. Kosciuch, J. R. Olson,J. H. Crawford, G. Chen and H. B. Singh, Airborne inter-comparison of HOx measurements using laser-inducedfluorescence and chemical ionization mass spectrometryduring ARCTAS, Atmos. Measure. Technol., 2012, 5, 2025–2037.
103 E. Schlosser, T. Brauers, H. P. Dorn, H. Fuchs, R. Häseler,A. Hofzumahaus, F. Holland, A. Wahner, Y. Kanaya,
Y. Kajii, K. Miyamoto, S. Nishida, K. Watanabe,A. Yoshino, D. Kubistin, M. Martinez, M. Rudolf,H. Harder, H. Berresheim, T. Elste, C. Plass-Dülmer,G. Stange and U. Schurath, Technical Note: Formal blindintercomparison of OH measurements: results from theinternational campaign HOxComp, Atmos. Chem. Phys.,2009, 9, 7923–7948.
104 J. Mao, X. Ren, L. Zhang, D. M. Van Duin, R. C. Cohen,J. H. Park, A. H. Goldstein, F. Paulot, M. R. Beaver,J. D. Crounse, P. O. Wennberg, J. P. DiGangi, S. B. Henry,F. N. Keutsch, C. Park, G. W. Schade, G. M. Wolfe,J. A. Thornton and W. H. Brune, Insights into hydroxylmeasurements and atmospheric oxidation in a Californiaforest, Atmos. Chem. Phys., 2012, 12, 8009–8020.
105 A. Yoshino, Y. Nakashima, K. Miyazaki, S. Kato,J. Suthawaree, N. Shimo, S. Matsunaga, S. Chatani,E. Apel, J. Greenberg, A. Guenther, H. Ueno, H. Sasaki,J.-y. Hoshi, H. Yokota, K. Ishii and Y. Kajii, Air qualitydiagnosis from comprehensive observations of total OHreactivity and reactive trace species in urban centralTokyo, Atmos. Environ., 2012, 49, 51–59.
106 Y. Nakashima, S. Kato, J. Greenberg, P. Harley, T. Karl,A. Turnipseed, E. Apel, A. Guenther, J. Smith and Y. Kajii,Total OH reactivity measurements in ambient air in asouthern Rocky mountain ponderosa pine forest duringBEACHON-SRM08 summer campaign, Atmos. Environ.,2014, 85, 1–8.
107 S. Lou, F. Holland, F. Rohrer, K. Lu, B. Bohn, T. Brauers,C. C. Chang, H. Fuchs, R. Häseler, K. Kita, Y. Kondo,X. Li, M. Shao, L. Zeng, A. Wahner, Y. Zhang, W. Wangand A. Hofzumahaus, Atmospheric OH reactivities in thePearl River Delta – China in summer 2006: measurementand model results, Atmos. Chem. Phys., 2010, 10, 11243–11260.
108 A. C. Nölscher, J. Williams, V. Sinha, T. Custer, W. Song,A. M. Johnson, R. Axinte, H. Bozem, H. Fischer,N. Pouvesle, G. Phillips, J. N. Crowley, P. Rantala, J. Rinne,M. Kulmala, D. Gonzales, J. Valverde-Canossa, A. Vogel,T. Hoffmann, H. G. Ouwersloot, J. Vilà-Guerau de Arellanoand J. Lelieveld, Summertime total OH reactivity measure-ments from boreal forest during HUMPPA-COPEC 2010,Atmos. Chem. Phys., 2012, 12, 8257–8270.
109 V. Sinha, J. Williams, J. Lelieveld, T. M. Ruuskanen,M. K. Kajos, J. Patokoski, H. Hellen, H. Hakola,D. Mogensen, M. Boy, J. Rinne and M. Kulmala, OH reac-tivity neasurements within a boreal forest: Evidence forunknown reactive emissions, Environ. Sci. Technol., 2010,44, 6614–6620.
110 H. Berresheim, J. McGrath, M. Adam, R. L. Mauldin,B. Bohn and F. Rohrer, Seasonal measurements of OH:NOx, and J(O1D) at Mace Head, Ireland, Geophys. Res.Lett., 2013, 40, 1659–1663.
111 T. Brauers, M. Hausmann, A. Bister, A. Kraus andH.-P. Dorn, OH radicals in the boundary layer of the Atlan-tic Ocean: 1. Measurements by long-path laser absorptionspectroscopy, J. Geophys. Res., [Atmos.], 2001, 106, 7399.
Perspective Photochemical & Photobiological Sciences
166 | Photochem. Photobiol. Sci., 2015, 14, 149–169 This journal is © The Royal Society of Chemistry and Owner Societies 2015

112 F. Rohrer and H. Berresheim, Strong correlation betweenlevels of tropospheric hydroxyl radicals and solar ultra-violet radiation, Nature, 2006, 442, 184–187.
113 S. A. Montzka, E. J. Dlugokencky and J. H. Butler, Non-CO2 greenhouse gases and climate change, Nature, 2011,476, 43–50.
114 G. Monteil, S. Houweling, E. J. Dlugockenky,G. Maenhout, B. H. Vaughn, J. W. C. White andT. Rockmann, Interpreting methane variations in the pasttwo decades using measurements of CH4 mixing ratio andisotopic composition, Atmos. Chem. Phys., 2011, 11, 9141–9153.
115 R. C. Pike and P. J. Young, How plants can influence tro-pospheric chemistry: the role of isoprene emissions fromthe biosphere, Weather, 2009, 64(12), 332–336.
116 E. D. Sofen, B. Alexander and S. A. Kunasek, The impactof anthropogenic emissions on atmospheric sulfate pro-duction pathways, oxidants, and ice core Δ17O(SO4
2−),Atmos. Chem. Phys., 2011, 11, 3565–3578.
117 C. D. Holmes, M. J. Prather, O. A. Søvde and G. Myhre,Future methane, hydroxyl, and their uncertainties: keyclimate and emission parameters for future predictions,Atmos. Chem. Phys., 2013, 13, 285–302.
118 J. G. John, A. M. Fiore, V. Naik, L. W. Horowitz andJ. P. Dunne, Climate versus emission drivers ofmethane lifetime against loss by tropospheric OH from1860–2100, Atmos. Chem. Phys., 2012, 12, 12021–12036.
119 O. Morgenstern, G. Zeng, N. Luke Abraham, P. J. Telford,P. Braesicke, J. A. Pyle, S. C. Hardiman, F. M. O’Connorand C. E. Johnson, Impacts of climate change, ozonerecovery, and increasing methane on surface ozone andthe tropospheric oxidizing capacity, J. Geophys. Res.,[Atmos.], 2013, 118, 1028–1041.
120 Z. H. Beygi, H. Fischer, H. D. Harder, M. Martinez,R. Sander, J. Williams, D. M. Brookes, P. S. Monks,J. Lelieveld and V. P. N. Ca, Oxidation photochemistry inthe Southern Atlantic boundary layer: unexpected devi-ations of photochemical steady state, Atmos. Chem. Phys.Disc., 2011, 11, 8497–8513.
121 R. Sommariva, A. L. Haggerstone, L. J. Carpenter,N. Carslaw, D. J. Creasey, D. E. Heard, J. D. Lee,A. C. Lewis, M. J. Pilling and J. Zádor, OH and HO2 chem-istry in clean marine air during SOAPEX-2, Atmos. Chem.Phys., 2004, 4, 839–856.
122 H. Lei, D. J. Wuebbles, X.-Z. Liang and S. Olsen, Domesticversus international contributions on 2050 ozone airquality: How much is convertible by regional control?,Atmos. Environ., 2013, 68, 315–325.
123 J. L. Neu, T. Flury, G. L. Manney, M. L. Santee,N. J. Livesey and J. Worden, Tropospheric ozone variationsgoverned by changes in stratospheric circulation, Nat.Geosci., 2014, 7, 340–344.
124 P. Jimenez-Guerrero, J. J. Gomez-Navarro, R. Baro,R. Lorente, N. Ratola and J. P. Montavez, Is there acommon pattern of future gas-phase air pollution in
Europe under diverse climate change scenarios?, Clim.Change, 2013, 121, 661–671.
125 E. Katragkou, P. Zanis, I. Kioutsioukis, I. Tegoulias,D. Melas, B. C. Krüger and E. Coppola, Future climatechange impacts on summer surface ozone from regionalclimate-air quality simulations over Europe, J. Geophys.Res., [Atmos.], 2011, 116.
126 C. M. Gan, J. Pleim, R. Mathur, C. Hogrefe, C. N. Long,J. Xing, S. Roselle and C. Wei, Assessment of the effect ofair pollution controls on trends in shortwave radiationover the United States from 1995 through 2010 from mul-tiple observation networks, Atmos. Chem. Phys., 2014, 14,1701–1715.
127 A. Colette, B. Bessagnet, R. Vautard, S. Szopa, S. Rao,S. Schucht, Z. Klimont, L. Menut, G. Clain, F. Meleux,G. Curci and L. Rouil, European atmosphere in 2050, aregional air quality and climate perspective under CMIP5scenarios, Atmos. Chem. Phys., 2013, 13, 7451–7471.
128 M. Demuzere and N. P. M. van Lipzig, A new method toestimate air-quality levels using a synoptic-regressionapproach. Part II: Future O3 concentrations, Atmos.Environ., 2010, 44, 1356–1366.
129 D. J. Jacob and D. A. Winner, Effect of climate change onair quality, Atmos. Environ., 2009, 43, 51–63.
130 P. Paasonen, A. Asmi, T. Petäjä, M. K. Kajos, M. Äijälä,H. Junninen, T. Holst, J. P. D. Abbatt, A. Arneth,W. Birmili, H. D. van der Gon, A. Hamed, A. Hoffer,L. Laakso, A. Laaksonen, W. Richard Leaitch, C. Plass-Dülmer, S. C. Pryor, P. Räisänen, E. Swietlicki,A. Wiedensohler, D. R. Worsnop, V.-M. Kerminen andM. Kulmala, Warming-induced increase in aerosolnumber concentration likely to moderate climate change,Nat. Geosci., 2013, 6, 438–442.
131 K. S. Carslaw, O. Boucher, D. V. Spracklen, G. W. Mann,J. G. L. Rae, S. Woodward and M. Kulmala, A review ofnatural aerosol interactions and feedbacks withinthe Earth system, Atmos. Chem. Phys., 2010, 10, 1701–1737.
132 R. Abrutzky, Health effects of climate and air pollution inBuenos Aires: A first time series analysis, J. Environ.Protect., 2012, 03, 262–271.
133 S. Pattenden, B. Armstrong, A. Milojevic, M. R. Heal,Z. Chalabi, R. Doherty, B. Barratt, R. S. Kovats andP. Wilkinson, Ozone, heat and mortality: acute effects in15 British conurbations, Occ. Environ. Med., 2010, 67, 699–707.
134 C. Ren, M. S. O’Neill, S. K. Park, D. Sparrow, P. Vokonasand J. Schwartz, Ambient temperature, air pollution, andheart rate variability in an aging population, Am. J. Epid.,2011, 173, 1013–1021.
135 P. H. Fischer, B. Brunekreef and E. Lebret, Air pollutionrelated deaths during the 2003 heat wave in the Nether-lands, Atmos. Environ., 2004, 38, 1083–1085.
136 J. R. Stedman, The predicted number of air pollutionrelated deaths in the UK during the August 2003 heatwave,Atmos. Environ., 2004, 38, 1087–1090.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 149–169 | 167

137 G. D’Amato, L. Cecchi, M. D’Amato and G. Liccardi,Urban air pollution and climate change as environmentalrisk factors of respiratory allergy: an update, J. Investig.Allergol. Clin. Immunol., 2010, 20, 95–102.
138 K. A. Read, A. S. Mahajan, L. J. Carpenter, M. J. Evans,B. V. E. Faria, D. E. Heard, J. R. Hopkins, J. D. Lee,S. J. Moller, A. C. Lewis, L. Mendes, J. B. McQuaid,H. Oetjen, A. Saiz-Lopez, M. J. Pilling and J. M. C. Plane,Extensive halogen-mediated ozone destruction over thetropical Atlantic Ocean, Nature, 2008, 453, 1232–1235.
139 J. C. G. Martin, A. S. Mahajan, T. D. Hay, C. Prados-Roman, C. Ordonez, S. M. MacDonald, J. M. C. Plane,M. Sorribas, M. Gil, J. F. P. Mora, M. V. A. Reyes,D. E. Oram, E. Leedham and A. Saiz-Lopez, Iodinechemistry in the eastern Pacific marine boundary layer,J. Geophys. Res., [Atmos.], 2013, 118, 887–904.
140 R. Hossaini, R. Hossaini, M. P. Chipperfield,B. M. Monge-Sanz, N. A. D. Richards, E. Atlas andD. R. Blake, Bromoform and dibromomethane in thetropics: a 3-D model study of chemistry and transport,Atmos. Chem. Phys., 2010, 10, 719–735.
141 T. J. Breider, M. P. Chipperfield, N. A. D. Richards,K. S. Carslaw, G. W. Mann and D. V. Spracklen, Impact ofBrO on dimethylsulfide in the remote marine boundarylayer, Geophys. Res. Lett., 2010, 37.
142 NTP, Toxicology and Carcinogenesis Studies of 1-Bromo-propane (CAS No. 106-94-5) in F344/N Rats and B6C3F1Mice (Inhalation Studies), National Toxicology ProgramReport No. NTP TR 564, Research Triangle Park, NC,August 2011 2011, p. 195.
143 USEPA, Protection of stratospheric ozone: Listing of sub-stitutes for ozone-depleting substances-n-propyl bromidein solvent cleaning, Fed. Regist., 2007, 72, 30142–30167.
144 USEPA, Q and A: 2007 Final and Proposed Regulations forn-Propyl Bromide (nPB), United States Environmental Pro-tection Agency http://www.epa.gov/ozone/snap/solvents/2007nPBRegsQA.html, accessed March 03, 2014.
145 S. E. Anderson, A. E. Munson, L. F. Butterworth,D. Germolec, D. L. Morgan, J. A. Roycroft, J. Dill andB. J. Meade, Whole-body inhalation exposure to 1-bromo-propane suppresses the IgM response to sheep red bloodcells in female B6C3F1 mice and Fisher 344/N rats, Inhal.Toxicol., 2010, 22, 125–132.
146 D. R. Worton, W. T. Sturges, J. Schwander, R. Mulvaney,J. M. Barnola and J. Chappellaz, 20th century trends andbudget implications of chloroform and related tri-anddihalomethanes inferred from firn air, Atmos. Chem.Phys., 2006, 6, 2847–2863.
147 P. G. Simmonds, R. G. Derwent, A. J. Manning,S. O’Doherty and G. Spain, Natural chloroform emissionsfrom the blanket peat bogs in the vicinity of Mace Head,Ireland over a 14-year period, Atmos. Environ., 2010, 44,1284–1291.
148 M. A. H. Khan, M. E. Whelan and R. C. Rhew, Effects oftemperature and soil moisture on methyl halide and
chloroform fluxes from drained peatland pasture soils,J. Environ. Monit., 2012, 14, 241–249.
149 X. Xiao, R. G. Prinn, P. J. Fraser, R. F. Weiss,P. G. Simmonds, S. O’Doherty, B. R. Miller, P. K. Salameh,C. M. Harth, P. B. Krummel, A. Golombek, L. W. Porter,J. H. Butler, J. W. Elkins, G. S. Dutton, B. D. Hall,L. P. Steele, R. H. J. Wang and D. M. Cunnold, Atmo-spheric three-dimensional inverse modeling of regionalindustrial emissions and global oceanic uptake ofcarbon tetrachloride, Atmos. Chem. Phys., 2010, 10, 10421–10434.
150 Q. Liang, P. A. Newman, J. S. Daniel, S. Reimann,B. D. Hall, G. Dutton and L. J. M. Kuijpers, Constrainingthe carbon tetrachloride (CCl4) budget using its globaltrend and inter-hemispheric gradient, Geophys. Res. Lett.,2014, 41, 2014GL060754.
151 G. Malaguarnera, E. Cataudella, M. Giordano, G. Nunnari,G. Chisari and M. Malaguarnera, Toxic hepatitis in occu-pational exposure to solvents, World J. Gastroenterol.,2012, 18, 2756–2766.
152 A. Tveit, G. M. Rusch, H. Muijser, M. J. van den Hovenand G. M. Hoffman, The acute, genetic, developmentaland inhalation toxicology of trans-1-chloro,3,3,3-trifluoro-propene (HCFO-1233zd(E)), Drug Chem. Toxicol., 2014, 37,83–92.
153 L. F. Bonifacio, E. Sousa, P. Naves, M. L. Inacio,J. Henriques, M. Mota, P. Barbosa, M. J. Drinkall andS. Buckley, Efficacy of sulfuryl fluoride against the pine-wood nematode, Bursaphelenchus xylophilus (Nematoda:Aphelenchidae), in Pinus pinaster boards, Pest Manag. Sci.,2014, 70, 6–13.
154 K. A. Buckman, J. F. Campbell and B. Subramanyam,Tribolium castaneum (Coleoptera: Tenebrionidae) associ-ated with tice mills: Fumigation efficacy and populationrebound, J. Econ. Entomol., 2013, 106, 499–512.
155 A. Cao, M. Guo, D. Yan, L. Mao, Q. Wang, Y. Li, X. Duanand P. Wang, Evaluation of sulfuryl fluoride as a soilfumigant in China, Pest Manag. Sci., 2014, 70, 219–227.
156 J. Susaya, K. H. Kim, Z. H. Shon and R. J. Brown, Demon-stration of long-term increases in tropospheric O3 levels:causes and potential impacts, Chemosphere, 2013, 92,1520–1528.
157 S. R. Wilson, K. R. Solomon and X. Tang, Changes intropospheric composition and air quality due to strato-spheric ozone depletion and climate change, Photochem.Photobiol. Sci., 2007, 6, 301–310.
158 D. R. Mattie, G. M. Hoffman, L. Narayanan, T. R. Sterner,M. J. Wagner and D. E. Dodd, A 13-week nose-only inhala-tion toxicity study for perfluoro-n-butyl iodide (PFBI) inrats with recommended occupational exposure levels,Inhal. Toxicol., 2010, 22, 847–860.
159 G. M. Rusch, A. Tveit, H. Muijser, M. M. Tegelenbosch-Schouten and G. M. Hoffman, The acute, genetic, develop-mental and inhalation toxicology of trans-1,3,3,3-tetra-fluoropropene (HFO-1234ze), Drug Chem. Toxicol., 2013,36, 170–180.
Perspective Photochemical & Photobiological Sciences
168 | Photochem. Photobiol. Sci., 2015, 14, 149–169 This journal is © The Royal Society of Chemistry and Owner Societies 2015

160 T. Schmidt, R. Bertermann, G. M. Rusch, G. M. Hoffmanand W. Dekant, Biotransformation of 2,3,3,3-tetrafluoro-propene (HFO-1234yf) in male, pregnant and non-pregnant female rabbits after single high dose inhalationexposure, Toxicol. Appl. Pharmacol., 2012, 263, 32–38.
161 A. Tveit, G. M. Rusch, H. Muijser andM. M. Tegelenbosch-Schouten, The acute, developmental,genetic and inhalation toxicology of 2,3,3,3-tetrafluoropro-pene (HFO-1234yf), Drug Chem. Toxicol., 2013, 36, 412–420.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 149–169 | 169

Photochemical &Photobiological Sciences
PERSPECTIVE
Cite this: Photochem. Photobiol. Sci.,2015, 14, 170
Received 20th October 2014,Accepted 20th October 2014
DOI: 10.1039/c4pp90038c
www.rsc.org/pps
Consequences of stratospheric ozone depletionand climate change on the use of materials
A. L. Andrady,*a A. Torikai,b H. H. Redhwi,c K. K. Pandeyd and P. Giese
Materials used in the exterior of buildings and in construction are routinely exposed to solar UV radiation.
Especially in the case of wood and plastic building materials, the service life is determined by their
weather-induced deterioration. Any further increase in ground-level solar UV radiation, UV-B radiation in
particular, will therefore reduce the outdoor service life of these products. Any increase in ambient temp-
erature due to climate change will also have the same effect. However, the existing light-stabilizer
technologies are likely to be able to mitigate the additional damaging effects due to increased solar UV
radiation and maintain the outdoor lifetimes of these materials at the present levels. These mitigation
choices invariably increase the lifetime cost of these products. A reliable estimate of what this additional
cost might be for different products is not available at the present time. Personal exposure to UV radiation
is reduced both by clothing fabrics and glass windows used in buildings and automobiles. This assessment
describes how the recent technical advances in degradation and stabilization techniques impact the life-
times of plastics and wood products routinely exposed to solar UV radiation and the protection to
humans offered by materials against solar UV radiation.
Introduction
Wood and plastics are used extensively as constructionmaterials. Of the 280 million tons of plastics produced globally[2011 data], about 23% is used in building construction, thesecond largest market for plastics after packaging appli-cations.1 Rigid poly(vinyl chloride) [rPVC] is the dominantplastic used in building applications. Polymers are also usedextensively in architectural and industrial coatings. Wood isused widely in building construction, often as the principalstructural element. Annually, ≈1.8 billion m3 of industrialroundwood [all industrial wood in the rough] is harvestedworldwide. Materials for construction used in exterior appli-cations require long-term durability that is delivered by stabil-ized plastics and coated or treated wood.
Some of the materials used in agricultural and transpor-tation applications are also regularly exposed to solar radi-ation, rain, and pollution but are expected to have service livesthat span several decades. Those used as coatings on auto-mobiles or on aircraft exteriors in particular are exposed to
high heat and UV radiation fluxes on a routine basis. Theirouter clear-coat layers responsible for finish and gloss is oftencompromised by exposure to UV radiation. Fig. 1 shows themain uses of plastics in outdoor applications.
Plastics and wood absorb solar UV radiation and areinherently susceptible to damage, especially at high ambient
Fig. 1 Uses of plastic materials in outdoor applications. PE = polyethyl-ene; PP = polypropylene; PVC = poly(vinyl chloride); PC =polycarbonate.
aDepartment of Chemical and Biomolecular Engineering, North Carolina State
University, Raleigh, NC 27523, USA. E-mail: [email protected] Life Society of Japan, Chuo-ku, Tokyo, JapancDepartment of Chemical Engineering, King Fahd University of Petroleum &
Minerals, Dhahran, Saudi ArabiadInstitute of Wood Science and Technology, Bangalore, IndiaeAustralian Radiation Protection And Nuclear Safety Agency, Melbourne, Australia
170 | Photochem. Photobiol. Sci., 2015, 14, 170–184 This journal is © The Royal Society of Chemistry and Owner Societies 2015

temperatures. The primary cause of weathering damage towood and plastics is from solar UV-B radiation that is efficien-tly absorbed by chromophores generally present in thesematerials. High humidity, temperature, and pollutants in airoften accelerate light-induced degradation. With plastics, thedamage ranges from uneven discoloration or surface changesto cracking and loss of material strength.2 With wood pro-ducts, weathering degradation also renders the surfacehydrophilic, facilitating subsequent moisture absorption andsurface biodegradation.3
Depletion of the stratospheric ozone layer to any extentincreases the fraction of UV-B (290–315 nm) in the solar radi-ation reaching the Earth’s surface. Potential latitude-depen-dent increases in UV-B radiation can significantly shorten theservice life of wood and plastics used outdoors, especially atlocations where the ambient temperatures are relatively high.Levels of UV radiation are expected to decrease globally in thedecades ahead (see Bais et al.4), but there is still concern aboutits impact on materials due to the interactive effects withclimate change. Climate change is widely expected to resultin an increase in the average global temperature by 1.1–6.4 °Cby the end of the century and there is an international effortto keep the increase under 2 °C.5 Any increase in ambienttemperatures exacerbates the damage as weatheringreactions in both wood and plastics proceed at faster rates atthe higher temperatures.6 Intrinsic factors such as additives orimpurities of trace metals (including pro-oxidant additives tomake the plastic photodegradable)7 also tend to acceleratethe rate of light-induced photo-damage to both classes ofmaterials.
The use of efficient UV stabilizers in compounding of plas-tics and effective surface coatings or treatments of wood,allows them to be successfully used in long-term outdoorapplications such as in cladding, panels, fencing, or decking.Even at concentrations as low as <0.1% by wt, light stabilizers,such as the Hindered–Amine Light Stabilizers [HALS] cancontrol light-induced damage in plastics to yield service livesof several decades. Existing stabilizer and coating technologiesare likely to be able to mitigate additional damaging effectsfrom small potential increases in UV-B radiation in the solarspectrum reaching the Earth.5 Either greater concentrations ofstabilizers in plastics or the use of more efficient surface treat-ments for wood will have to be employed to mitigate theseeffects. Intensive industrial research effort is focused on disco-vering better and lower-cost light stabilizers for specific classesof plastics used in building8 and improved surface coatings forpreserving wood against weathering. New materials, additivesor new variants of existing plastics that are more weather-resistant emerge in the marketplace regularly.
Polymeric materials are also used to protect humansagainst exposure to solar UV radiation. These include clothingmade of synthetic fibers and plastic glazing used in buildings(and vehicles) to filter out the UV radiation. The effective useof these limits exposure and therefore the adverse healthimpacts of UV radiation on humans. This paper assesses therelevant literature published since the last report in 2010.
Plastics use in building
Unlike with biota, that may show a limited capacity to adapt toincreased UV-B radiation levels and where evolution of newtraits is relatively slow, man-made materials can be designedto withstand new, harsher, solar UV radiation environments.Plastics, for instance, can be formulated with either greaterlevels of conventional stabilizers or more efficient novel stabil-izers, to ensure a minimal loss in their service life. A substi-tution of materials with different, more UV-resistant classes ofplastic or non-plastic material is a second strategy. Surfacecoatings and modifications of wood can also be designed forbetter resistance to solar UV radiation. However, invariably, itis the economics of their use that will dictate the specific miti-gation strategy adopted. Regardless of the technology choice, itwill inevitably add to the lifetime cost of using either class ofmaterials.
Durability of materials outdoors is determined primarily bythe dose of solar UV radiation, especially the proportion ofUV-B radiation the material will be exposed to. As the dose ofUV-B is latitude and altitude dependent, the durability of agiven material will vary widely with the geographic location.The certified lifetime for a plastic material assessed at onelocation cannot be assumed to be the same for another with adifferent UV environment. Global UV-A and UV-B radiationmaps (for Nov. 1, 1978 to June 30, 2000) estimated from TOMSdata, corrected for the effect of cloud,9 (Fig. 2) illustrates thewide range of UV-B radiation environments encountered atdifferent locations around the World. However, these must beviewed with caution. Firstly, they are generally based on data
Fig. 2 A global map of the annual un-weighted mean dose of UVA(above) and UVB (below) in units of kJ m−2 day−1. Reproduced with per-mission from Lee-Taylor.9
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 170–184 | 171

from a single satellite overpass each day, and amounts ofcloud at that time may not be representative of a true average.Secondly, satellite estimations of UV tend to be too high underpolluted conditions.4
The main uses of plastics in building construction includeplastic pipes, siding, windows, soffit, fascia, rainwater goodsand decorative panels and a majority of these are made ofrigid PVC (rPVC). In fact, 76% of the PVC produced globally isused in the construction of buildings. The most popular clad-ding in residential housing in North America is rPVC sidingproduced by a profile extrusion process. In this application,the useful lifetime of the product is generally determined byuneven discoloration and loss of impact strength from photo-damage by solar UV radiation.
Polycarbonate [PC] glazing used in architectural windowpanels as well as continuous windows and domes is similar toor better in performance than the conventional glazing inseveral characteristics.10 One important advantage of these istheir lower thermal conductivity, k, relative to glass. The valueof k for PC glazing panels are as low as 1.2–1.9 W m−2 (25 mmthickness), but new technology such as PC/aerogel compo-sites11 can bring this value down to 0.5 W m−2 (25 mmsection). In these products as well, discoloration induced bysolar radiation, determines their useful lifetime.
The present push towards sustainable materials in con-struction has led to some reassessment of the use of PVCmaterials in buildings.12 Despite its dominance in the build-ing sector and excellent performance in construction, PVC isperhaps the worst choice of a plastic in terms of environ-mental merit.13 Not only does the production of PVC result inpotential emission of toxic monomers and precursors to airbut it is also compounded into soft products with phthalateplasticizers that are potent endocrine disruptors. Nevertheless,no imminent move away from its use in buildings is apparent.Alternative plastics that can replace PVC in construction suchas polyolefins are available, but at a greater lifetime cost. Thesusceptibility of these alternatives to solar UV radiation willalso influence their lifetime costs.
PVC in building and construction
Improvements in rigid PVC technology aimed at making itmore environmentally acceptable, better performing, and withlower lifetime costs, are being made on a continuing basis. Inthe face of competition from alternative materials, PVC tech-nology has advanced considerably. The lighter micro-foamedPVC siding14 and the emerging (polystyrene) foam-backed PVCsiding with greater insulation efficiency are examples of suchimprovements.
Most of the PVC products are processed as extruded pro-files. Recent introduction to the market of a new processingaid15 as well as an acrylic impact modifier16 for PVC extrusionproducts (in profile, window, and siding applications) is a sig-nificant improvement in the technology. The new additivesallow PVC to be extruded at a lower temperature with reducedrisk of degradation and atmospheric emissions. On heating,PVC breaks down emitting hazardous corrosive HCl gas
making it difficult to process. Typically, either tin-based orlead-based heat stabilizers are used to allow thermal proces-sing of rPVC. A new thermal stabilizer17 with a lower tincontent (19%) was recently commercialized for use in thesame types of PVC products used in building. Improved pro-cessibility and superior thermal stabilization of PVC areafforded by the use of new stabilizers, which also are claimedto impart superior weatherability to the plastic material. Lowerthermal degradation during processing improves the servicelifetimes of rPVC products as it reduces the buildup of UV-absorbing functionalities in the polymer. No long-term UV-stability data for PVC compounds that incorporate these neweradditives are as yet available. However newer tin-maleate stabil-izers effectively control yellowing due to weathering of thematerial. Other manufacturers18 have also introduced theirown novel PVC additives for the same set of building products.
The solar UV-B-induced degradation of rPVC is catalyzed byZnO used as a filler in the plastic.19 The findings have impli-cations only in the management of plastic waste as outdoorPVC products are generally formulated to be resistant to UVradiation. The wavelength sensitivity and activation spectra fortypical injection molded rPVC formulations are known.Similar data for solvent-cast sheets of PVC, manufactured forgraphic arts applications, were recently published and themerit of the material as a UV dosimeter discussed.20 The dataon wavelength sensitivity as well as activation spectra deter-mined for solvent-cast plastic generally agreed with thosealready published for injection molded rPVC.21,22 This showsthat the wavelength sensitivity of these materials is governedprimarily by the nature of the polymer rather than the addi-tives or processing-related factors. In accelerated weathering ofplastics under laboratory conditions, it is essential to use alight source with a spectrum similar to that of sunlight. Forexample, mercury vapor lamps cannot be used as a substitutefor a xenon source.23 Recent advances in PVC technology havecontributed to improved service lifetimes of the materialexposed to solar UV-B radiation and the anticipated effects ofclimate change.
Wood-plastic composites
The 2010 global market for wood–plastic composites (WPC)was 2.3 million tonnes and, given the short-term projectedgrowth rate of 13.8%, per year this is expected24 to grow to4.6 million tonnes by 2016. WPCs are essentially thermoplasticcomposites (of PE, PP, PVC, and polystyrene (PS)) highly filledwith powdered wood. Often, post-consumer plastics are usedin their manufacture. Embedding wood fibers in a polymermatrix restricts absorption of moisture and hence avoidsfungal growth and biodegradation of the wood fraction.25 Ahigher-grade product results when a single type of plastic suchas virgin PP is used26,27 and the compounding and/or proces-sing operations are carefully controlled28 to obtain good dis-persion of the wood filler.
To improve miscibility of the hydrophilic wood and hydro-phobic plastics in fabricating WPCs, compatibilisers are oftenemployed.29,30 Good adhesion between the phases was
Perspective Photochemical & Photobiological Sciences
172 | Photochem. Photobiol. Sci., 2015, 14, 170–184 This journal is © The Royal Society of Chemistry and Owner Societies 2015

reported, for instance, with maleic anhydride grafted PE andPP.28,31 A continued search for good adhesion-promoters orcompatibilisers for specific wood/plastic systems is critical forfuture growth of this technology. Alternatively, the wood frac-tion can be chemically functionalized (e.g., acetylated;32) forbetter compatibility. Low processing temperatures (<200 °C)have to be used in processing WPCs because the wood com-ponent can otherwise thermally degrade. However, WPCsbased on waste plastics and wood powder can be processedusing conventional equipment into “plastic lumber” that canbe handled and worked on using tools designed for wood.WPC is promoted as a ‘green’ (environmentally friendly)material as it uses waste wood and often post-consumer plas-tics as well.33–35
Lignin in the wood fraction of WPC absorbs solar UV radi-ation and undergoes photo-degradation36 leading to delignifi-cation of the wood particles at the surface of the WPCmaterial.37 Photodegradation is reported to cause discolor-ation and breakdown of the filler in PP/wood38 and PE/woodWPCs.39 These changes render the surfaces hydrophilic,encouraging fungal growth, and biodegradation of the cellu-lose rich tissue.40 Absorption of water by wood particles underfreeze–thaw cycles causes swelling/shrinkage of the fibres thatalso destabilizes WPC creating voids or empty spaces in thematrix. These recent findings on WPC parallel that which isalready known of the mechanisms of degradation for woodspecies. Understanding all the degradation pathways involvedfor WPCs is critical to developing better UV-B stabilizers forthe material. The main advantage of WPC is that hydrophilicwood fibres that absorb water and are prone to biodegradationare embedded in plastic protecting them from solar UV radi-ation and environmental biodegradation.
The color, mechanical properties and durability of WPCsvary with the species of wood fiber used. For instance, inWPCs made of high density polyethylene [HDPE] with DouglasFir and HDPE-hybrid Poplar wood discolored the least (>15%)on exposure to solar-simulated radiation (xenon lamp) com-pared to HDPE composites with wood species such as Whiteoak and Ponderosa pine.39
As expected, the plastic component of WPC also undergoeslight-induced degradation. Of the common thermoplasticsused in WPCs, polystyrene41 is the most susceptible. There issome evidence that the presence of wood in PVC-based WPCsmay also promote photodegradation of the plastic matrix.42
Naturally, the same light stabilizers used with plastics are alsoeffective in protecting WPCs; for instance, the use of HALSstabilizers in wood-HDPE42,43 and light absorber (benzotri-azole type) in wood-PVC44 composites have been successful.The extent of protection afforded depends on the level of thestabilizer and its dispersion in the polymer matrix.45 Whetherthere is an advantage in using a single stabilizer known to beeffective in both wood and plastics, to protect WPC is not clearat this time.
Fig. 3 shows the effect of brown-rot fungi acting on asample of WPC biodegrading the particles of wood exposed atthe surface of the composite. A thin layer of plastic (or a cap
layer) extruded on to the WPC surface may help seal in theseexposed wood particles at the surface, discouraging biodegra-dation.46 With a HDPE cap layer, the rate of discoloration fromexposure to solar UV radiation was reduced by ≈50% andaccess of the wood fibers to moisture was also substantiallyreduced.47 Blending UV absorbers with a HALS in the HDPEcap layer can further enhance photostability.48 While severalstudies have established the effectiveness of cap-layer technol-ogy, its techno-economic feasibility in specific product cat-egories remains to be demonstrated.
Biodegradable and compostable plastics such as poly(lacticacid)49 or poly(3-hydroxybutyrate)50 have been studied for theirpotential use in WPC. The perceived sustainability of thesepotentially fully biodegradable WPCs that return all the carboninto the carbon cycle is attractive. However, the greater cost ofthese specialized plastics will likely preclude commercializa-tion of such WPCs in spite of the claimed but questionableenvironmental advantage of biodegradability.
Emerging issue of weathering and microplastics
Weathering of plastics outdoors produces micro-scale plasticfragments that have serious environmental consequences.51
Microplastics (fragments in the size range of 1 mm to 1 µm)
Fig. 3 Scanning electron micrograph of a WPC (white oak (60%)/HDPE(40%) control sample (A) and after exposure to Gloeophyllum trabeum(Brown rot fungi) for 12 weeks (B). Reproduced with permission fromFabiyi et al.39
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 170–184 | 173

are primarily believed to be derived from weathered, brittleplastic litter and are widely accepted to be a serious ecologicalconcern, especially in the marine environment.52,53 Thesurface cracking of highly weathered plastics is the likelyorigin of microplastics.51 Plastic particles, regardless of size,concentrate persistent organic pollutants (POPs).54 dissolvedin the ocean water at very low concentrations. The distributioncoefficients (K) of a compound between the plastic and waterphase is the ratio of its equilibrium concentration in theplastic and in seawater. For common POPs, values of theK between sea water and plastics are large, ranging from 104
to 105 in favor of plastic. This means the equilibrium con-centration of a POP species can be several orders of magnitudegreater in the plastic debris compared to in seawater. Becauseof their small size, some of the microplastics are ingested byzooplankton and other marine species,55 thus providinga pathway for POPs to enter the marine food web.56 Thespecific compounds of concern include endocrine disruptors(EDs) such as PCBs, BPA, phthalates, and residual polymercatalysts.
Polypropylene (PP) is commonly found in urban litter andis an important generator of microplastics via surface photo-degradation. The wavelength sensitivity of the surface crackingof PP stabilized by HALS has been reported recently.57 Thesame logarithmic dependence of photodamage on the wave-length of exposure, already reported for various measures ofdamage, such as discoloration or mechanical properties, wasalso confirmed for surface cracking. This is the first reportedaction spectrum for crack formation and is relevant as micro-plastics originate from surface cracks. However, quantifyingsurface cracking was indirect and not entirely satisfactory,being based on a correlation between carbonyl index (a spec-troscopic quantity) and the degree of cracking. Extrinsicfluorescence techniques using Rhodamine dye, recently usedfor the first time to study microstructural changes in polymersduring UV degradation,58,59 hold more promise for quantifi-cation of weathering-induced microcracking of plastics.
Higher degrees of crystallinity obtained under extensiveweathering are one of the main reasons for embrittlement. Onweathering of semi-crystalline plastics (such as PE or PP), it isthe amorphous regions that oxidize first because of the lessersolubility and diffusion rates of oxygen in the crystallites.This was recently confirmed for PE60 using dielectric relaxa-tional spectroscopy, a technique not hitherto used for thispurpose. In contrast, Ojeda2 reported that polypropylenesamples underwent a decline (>12%) in crystallinity duringnatural weathering. The latter observation has not beensatisfactorily explained although impurities or accumulationof product during degradation has been suggested as thecause. Research is needed to clarify the relationship betweencrystallinity changes and exposure to UV radiation duringweathering.
Plastics in solar photovoltaic applications
While plastics are also being evaluated as solar thermal absor-bers,61 it is the photovoltaic (P-V) applications that are of
greater commercial interest.62,63 Research emphasis appears tobe divided between the solar UV-damage to light-harvestingsemiconductor polymers in organic P-V devices47,64,65 and thatof plastic encapsulants used to protect silicon P-V modules.
Recent designs of P-V modules utilize plastics extensively.Generally, two plastic protective laminates or sheets in themodules are affected by solar radiation (Fig. 4). Light trans-mitted through the transparent front panel reaches theexposed parts of the front sheet and the front surface of theback sheet, discoloring and weakening these after long dur-ations of exposure. The back sheet is often constructed of amulti-layered laminate designed for the service life of themodule (20–25 years). These sheets are made of a poly(vinylfluoride) (PVF) outer layer, a thicker inner core of poly(ethyleneterephthalate) (PET) with titanium dioxide (for opacity), and aPVF surface layer (or a PE/EVA tie layer). Of the plastics evalu-ated over the years, PVF has been selected as the best suitedfor the application. Long-term exposure can result in discolor-ation and loss of strength in these sheets and, although accel-erated test methods are available64,66,67 they are inadequate toreliably predict their service lifetimes. PE and PET degradeunder solar UV exposure and undergo yellowing and cracking.The back surface of the back sheet is also affected by diffusedUV radiation that, along with high temperatures68 cause initialdamage within ≈5 years of outdoor exposure.69
There is an urgent research need to (a) develop weatherresistant encapsulants as well as accelerated test methods toassess the long-term (>20–30 years) service lifetimes of P-Vmodules, reliably. Any increase in UV-B radiation and/or temp-erature due to climate change will clearly accelerate the degra-dation, but techniques and standards to reliably quantify theeffects reliably are still being developed.70 The lifetime of pro-tective plastics is a determining factor in implementing large-scale economical P-V technology.
Fig. 4 Structure of a P-V module showing the back sheet and frontsheet material placement. Source: Courtesy of DuPont.
Perspective Photochemical & Photobiological Sciences
174 | Photochem. Photobiol. Sci., 2015, 14, 170–184 This journal is © The Royal Society of Chemistry and Owner Societies 2015

Plastics in greenhouse covers
Plastic films and laminates are the leading greenhouse clad-ding materials used worldwide. Thin plastic films are some-times used on conventional glass cladding to modify thespectral quality of light reaching the crops.71 Plastic panelshave different transmission rates for photosynthetically activeradiation (PAR, 400–700 nm) and sometimes the plastics incor-porate dyes or modifiers72 to change the transmitted spectrumof solar radiation. The photostability of the dye is a concern inthis technology and has not been explored as yet in detail.
Rigid plastics (>10 year service life) commonly used ingreenhouses are fibre-reinforced polyester panels, polycarbo-nate (PC), and acrylics. Flexible cladding (3–4 years of servicelife) is generally made from PE, ethylene copolymers, and PVC;although PET is also sometimes used. Both classes of claddingundergo discoloration and loss of PAR transmission withweathering during use.73
Nanofillers have been evaluated in low-density polyethylene(LDPE) and linear low-density polyethylene (LLDPE) green-house films to improve mechanical properties and to reducetransmission of UV. Nanoscale oxides of titanium, zinc oxide,and silica were evaluated for this purpose.74 Nano-zinc oxidefillers improve weatherability of the films via UV-absorption orshielding, without affecting their transparency for PAR wave-lengths. With certain varieties of crops, some degree of trans-parency to UV radiation might be needed to ensure a greaterquality produce (see Bornman et al.75) and plastic films can bedesigned to accommodate this need. The market for durablegreenhouse films is large enough for speciality chemical com-panies (Clariant, Swizerland) to offer tailor-made light stabiliz-ation solutions designed for greenhouse films.76 While thepotential for durable but UV-resistant greenhouse films basedon nano-composites clearly exists the lifetime cost of their usehas not been reported.
Dependence of solar UV radiation damage to greenhousefilms on the dose of solar UV-B radiation is of interest in devel-oping lifetime assessments. Dehbi and colleagues77,78 reporteda U-shaped (nonlinear) dependence of yellow discolorationwith duration of exposure for LDPE greenhouse films, in bothnatural and accelerated weathering. However, this form ofdose–response behaviour is unexpected and atypical and islikely an artifact of the layered structure of the film. Oreskiand colleagues79 investigated the weathering changes in stabil-ized EVA copolymer greenhouse films (200 µm) and showedthat only a thin surface layer was degraded. Even in thinnerfilms (30 µm) of EVA, the mechanical properties were not sig-nificantly affected at 1000 h of accelerated weathering exposureat 60 W m−2 of UV irradiance (300–400 nm) at 40 °C. Present-day stabilizer technology is clearly adequate for greenhousefilm applications. Energy costs of maintaining greenhousetemperatures vary with the choice of cladding. In a new model-ling study, Al-Madhouri and colleagues80 found that the lowemissivity of PVC is a better choice than LDPE in conservingheat in greenhouses. The search continues for a single plasticmaterial that combines long-term resistance to solar UV radi-
ation, transmittance of PAR, and thermal insulation, and isideally suited for greenhouses.
Nanofiller in photostabilisation of plastics
The bulk of reported work on nanofilled polymers involvespolymer-based coatings, especially clear coats used in protect-ing exterior surfaces from solar UV damage (these are assessedbelow).
Redhwi81,82 studied three nano-composites of LDPE (withClay, ZnO, and Silica at 5% (wt)) and found that natural resist-ance of the plastic to weathering was not compromised by thepresence of the nanofillers except in the case of nanosilica. Instudies on nanocomposite systems, surface-modified rutiletitania (40–100 nm) in LDPE83 was reported to control the lossin tensile properties relative to unfilled samples in acceleratedweathering studies. Nanofillers generally need to be eithersurface-treated or a compatibiliser must be used in the formu-lation to ensure that high levels of dispersion are achievedthroughout the material. While some work has been carriedout on this topic, further information is needed across thewide range of filler and/or plastics. The stability of the compa-tibilisers or modifiers to UV radiation also has not beenstudied in sufficient detail.
In general, the inclusion of nanoscale fillers should impartincreased photostability to polymer coatings (including woodtopcoats) and bulk polymers. Because variables, such as theparticle size distribution and dispersion effectiveness areinvolved, quantitative effects are difficult to compare even forthe same nanofillers in identical polymer matrices. Differentcombinations of these have been used in different studies. Itappears that, in some systems at least, nanofillers yield at leastthe same level of protection as conventional fillers, but at alower volume fraction and can therefore be an economicalchoice. However, further work is needed to elucidate the con-ditions under which they act as photostabilizers.
Kingston et al.,84 recently reviewed the potential for releaseof nanofillers from several nano-composites filled with multi-wall carbon nanotubes (MWCNT) and concluded that thepotential for release of MWCNT with typical intended consu-mer use is expected to be small. It is not known if weatheringof the composite surfaces facilitates the release of nanofillerparticles from filled-coatings or composites. Release of nano-scale fillers from composites during their use is an emergingenvironmental concern and is being investigated.
Temperature effects in solar UV-damage of materials
At higher ambient temperatures the light-induced degradationrates of materials accelerate. In the case of wood, heat aloneoften does not result in significant degradation; yet highertemperatures accelerate the photo-degradation process. Inmodelling effects of temperature in weathering studies,ambient temperatures are typically used. However, because ofthe absorption of solar radiation, especially the infrared radi-ation, the surface temperature of the sample is often higher byas much as 10–20 °C (depending on the material) and shouldbe used in more realistic weathering models. A sophisticated
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 170–184 | 175

heat transfer model that allows the estimation of samplesurface temperatures from metrological data (without resortingto expensive field measurements) has been developed and vali-dated.85 Its availability will not only refine the modelling ofdamage estimates but also contribute to the assessment ofstabilizers.
The surface cracking of light-stabilized PP exposed to xenonlamp radiation was recently reported.57 In this first quantitat-ive study of light-induced crack formation, the activationenergy estimated for this process was 20 kJ mol−1. However,models based on the Arrhenius equation86 or the reciprocityrule are inadequate in describing the case of concurrent light-induced and thermal (photothermal) degradation of PP.87 Thelack of a satisfactory general quantitative model is a majordrawback in predicting the rates of degradation and servicelife in plastics exposed to solar UV radiation where heat build-up also increases the temperature of the material. Thisdeficiency is particularly apparent in P-V device technology.
The temperature dependence of UV radiation-inducedphotodegradation was recently demonstrated experimentallyvia the exposure of wood from conifers and deciduous speciesto UV-B.88 However, as a mercury vapor lamp that also emitsUV-C radiation was used as the source in this study, the resultsare not pertinent to natural weathering. Increases in tempera-ture from 30 °C to 80 °C increased the light induced colorchanges by 33–57% in pine, spruce, ash, and poplar wood,89
and the effect is more pronounced at greater humidity.90 Thehigh temperatures used in the laboratory accelerated weather-ing tests reported by Persze and Tolvaj,89 do not correspond torealistic temperatures of storage or use of wood.
Wood as a building material
In North America 25% of the windows are made of woodwith nearly 9 million units made in 2010. The overridingmarket trend for building materials in general is the increas-ing demand for “green” or sustainable materials ofconstruction.12,91–93 Exposed wood products are common inbuildings, and these are generally surface-coated to ensuredurability. A wide range of protective finishes such as paints,varnishes, stains, or water repellents are used for thispurpose.94 Recent life-cycle analyses (LCA) confirm wood to bean environmentally friendly building material.95,96
Photodegradation of wood
Lignin in wood is a potent chromophore that readily absorbsUV radiation and undergoes ready photodegradation. Thisresults in rapid color changes,90,97,98 lignin degradation97,99,100
and loss in microtensile strength.101 As with plastics, the oxi-dative degradation process in wood also increases the carbonylindex (the relative spectral signal from carbonyl groups in itsinfra-red spectrum) as a result of lignin degradation.102 Thefundamental measures in weathering are the efficiency of thephotoprocess, the photodamage/mole of available photonsand the action spectrum.103 The action spectrum for yellowingof mechanical pulp (newsprint paper) under exposure to simu-lated solar radiation was re-investigated recently in a rigorous
laboratory study.104 The action spectrum for yellowing showeda logarithmic dependence of yellowing efficiency on wave-length as previously reported for newsprint paper.103 Theserecent findings are in line with and confirm the known dataon photodamage to wood.
As with plastics, solar UV radiation typically penetrates onlythe surface layer of wood. Photodegradation of surface layersmonitored by microtensile measurements101,105 shows thedamage is limited to about a 250 µm layer for Spruce woodexposed to simulated sunlight. Scanning electron microscopyshowed some cellular damage at greater depths, but this wasnot supported by tensile property changes.
The role of wood extractives in UV stabilization
Extractives are naturally-occurring compounds that can beextracted from wood using common solvents such as acetone,ether and methanol. Natural wood extractives, being goodabsorbers of solar UV radiation, protect the wood from photo-degradation.106 Extractives in wood generally include phenoliccompounds, stilbenes, and flavonoids that act as antioxidantsand light stabilizers. Nzokou and Kamden107 found the extrac-tives to act as antioxidants and a stabilizer against degradationby UV radiation, during artificial weathering of Prunus serotina,Quercus rubra, and Pinus resinosa. Chang and colleagues106
also showed that extractives slow down the rate of wood degra-dation in Cryptomeria japonica and Acacia confuse heartwood.Wood/LLDPE composites prepared with extractive-free wood(extractives removed by solvent extraction) showed poorer UVradiation stability compared to un-extracted wood fiber.3 Incontrast to the above, Sharratt and colleagues;56 however,reported that discoloration of Scots pine (Pinus sylvestris)exposed to simulated sunlight exposure, was unaffected by thepresence of extractives. Consistent with this observation of UV-stabilizer activity of extractives, textile dyes derived from plantshave very good UV-protective properties.108
Drawbacks of extractives as stabilizers, however, are theirwater solubility and ready leachability from wood. While heattreatment, at least in the case of Merbau heartwood, appears tohelp prevent this,109 a general solution to the problem is notavailable. Wood extractives, depending on the species, couldbe a new source of light stabilizer that can be further develo-ped and refined to guard both wood and WPCs againstdamage caused by solar UV radiation.
Stabilizing wood against UV radiation
Improved protection of wood against solar UV radiation isafforded by coating the surface with layers containing lightstabilizers110 such as HALS (or synergistic combinations ofstabilizers111) or inorganic fillers. Coating and surface modifi-cations (as with cap layers already referred to in connectionwith WPC) also reduced moisture pick-up and thereforeincreased the fungal resistance112 of wood.
A wide range of protective surface coverings or finishessuch as paints, varnishes, stains, or water repellents113 arenow commercially available. Transparent coatings that are notUV stabilized merely protect the wood against moisture but are
Perspective Photochemical & Photobiological Sciences
176 | Photochem. Photobiol. Sci., 2015, 14, 170–184 This journal is © The Royal Society of Chemistry and Owner Societies 2015
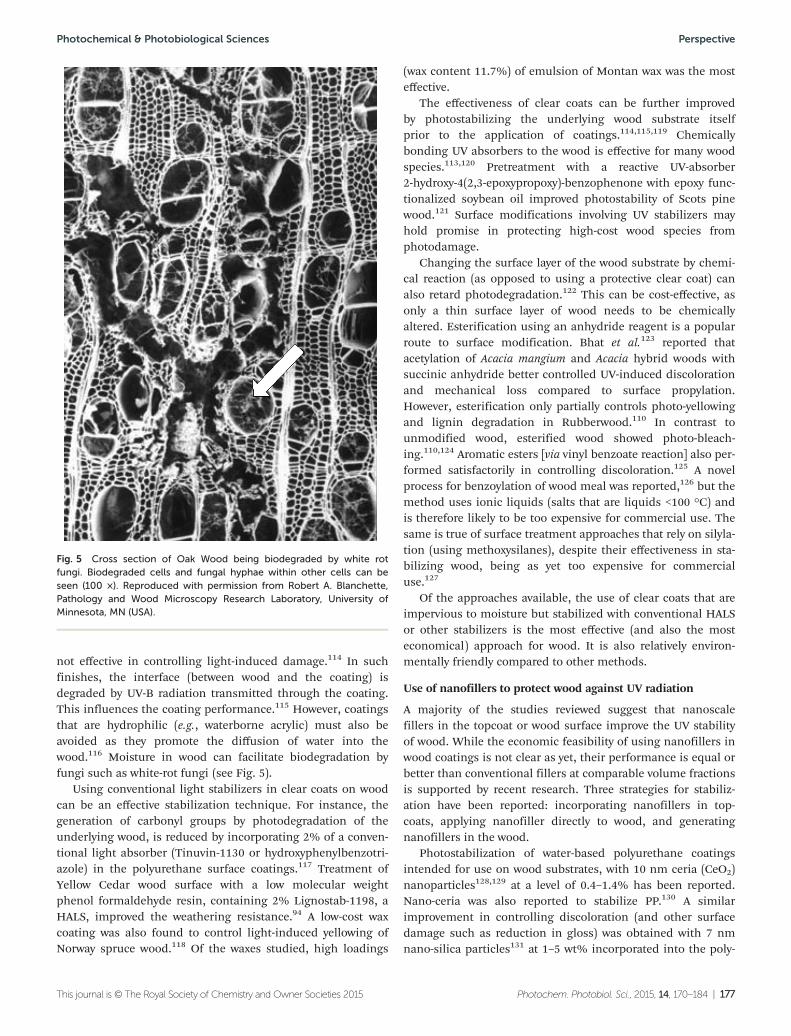
not effective in controlling light-induced damage.114 In suchfinishes, the interface (between wood and the coating) isdegraded by UV-B radiation transmitted through the coating.This influences the coating performance.115 However, coatingsthat are hydrophilic (e.g., waterborne acrylic) must also beavoided as they promote the diffusion of water into thewood.116 Moisture in wood can facilitate biodegradation byfungi such as white-rot fungi (see Fig. 5).
Using conventional light stabilizers in clear coats on woodcan be an effective stabilization technique. For instance, thegeneration of carbonyl groups by photodegradation of theunderlying wood, is reduced by incorporating 2% of a conven-tional light absorber (Tinuvin-1130 or hydroxyphenylbenzotri-azole) in the polyurethane surface coatings.117 Treatment ofYellow Cedar wood surface with a low molecular weightphenol formaldehyde resin, containing 2% Lignostab-1198, aHALS, improved the weathering resistance.94 A low-cost waxcoating was also found to control light-induced yellowing ofNorway spruce wood.118 Of the waxes studied, high loadings
(wax content 11.7%) of emulsion of Montan wax was the mosteffective.
The effectiveness of clear coats can be further improvedby photostabilizing the underlying wood substrate itselfprior to the application of coatings.114,115,119 Chemicallybonding UV absorbers to the wood is effective for many woodspecies.113,120 Pretreatment with a reactive UV-absorber2-hydroxy-4(2,3-epoxypropoxy)-benzophenone with epoxy func-tionalized soybean oil improved photostability of Scots pinewood.121 Surface modifications involving UV stabilizers mayhold promise in protecting high-cost wood species fromphotodamage.
Changing the surface layer of the wood substrate by chemi-cal reaction (as opposed to using a protective clear coat) canalso retard photodegradation.122 This can be cost-effective, asonly a thin surface layer of wood needs to be chemicallyaltered. Esterification using an anhydride reagent is a popularroute to surface modification. Bhat et al.123 reported thatacetylation of Acacia mangium and Acacia hybrid woods withsuccinic anhydride better controlled UV-induced discolorationand mechanical loss compared to surface propylation.However, esterification only partially controls photo-yellowingand lignin degradation in Rubberwood.110 In contrast tounmodified wood, esterified wood showed photo-bleach-ing.110,124 Aromatic esters [via vinyl benzoate reaction] also per-formed satisfactorily in controlling discoloration.125 A novelprocess for benzoylation of wood meal was reported,126 but themethod uses ionic liquids (salts that are liquids <100 °C) andis therefore likely to be too expensive for commercial use. Thesame is true of surface treatment approaches that rely on silyla-tion (using methoxysilanes), despite their effectiveness in sta-bilizing wood, being as yet too expensive for commercialuse.127
Of the approaches available, the use of clear coats that areimpervious to moisture but stabilized with conventional HALSor other stabilizers is the most effective (and also the mosteconomical) approach for wood. It is also relatively environ-mentally friendly compared to other methods.
Use of nanofillers to protect wood against UV radiation
A majority of the studies reviewed suggest that nanoscalefillers in the topcoat or wood surface improve the UV stabilityof wood. While the economic feasibility of using nanofillers inwood coatings is not clear as yet, their performance is equal orbetter than conventional fillers at comparable volume fractionsis supported by recent research. Three strategies for stabiliz-ation have been reported: incorporating nanofillers in top-coats, applying nanofiller directly to wood, and generatingnanofillers in the wood.
Photostabilization of water-based polyurethane coatingsintended for use on wood substrates, with 10 nm ceria (CeO2)nanoparticles128,129 at a level of 0.4–1.4% has been reported.Nano-ceria was also reported to stabilize PP.130 A similarimprovement in controlling discoloration (and other surfacedamage such as reduction in gloss) was obtained with 7 nmnano-silica particles131 at 1–5 wt% incorporated into the poly-
Fig. 5 Cross section of Oak Wood being biodegraded by white rotfungi. Biodegraded cells and fungal hyphae within other cells can beseen (100 ×). Reproduced with permission from Robert A. Blanchette,Pathology and Wood Microscopy Research Laboratory, University ofMinnesota, MN (USA).
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 170–184 | 177

urethane coating. The same was reported for 50 nm nano-ZnO132 in the same class of coating and for a 20 nm grade ofthe nanofiller at 1–2 wt% in an acrylic wood stain.133 However,using 0.5–1.0 wt% of 20 nm rutile, a crystalline form of nano-titania, in similar coatings showed a decrease in photostabilitycompared to base polymer, in terms of discoloration underaccelerated weathering.
Acrylic water-based coatings for wood, containing nano-titania (NT) as well as nano-ZnO133 have also been evaluatedfor control of discoloration, loss of gloss, and carbonyl groupformation indicative of degradation. Fufa et al.134,135 found NTin the topcoat of wood cladding to be only marginally betterthan the base coating (with no NT) in controlling discolorationand carbonyl formation in Norway spruce. However, a very lowfraction of only 1 wt% (based on solids) of filler was used inthat study. A 50 μm thick clear coat with 1.14 wt% of NT fillerafforded a much higher level of protection compared to thatfrom a comparable coating with benzotriazole light absor-bers.136 Direct deposition of rutile nanoparticles on hardwoodsurface (without a coating) is also an option; this is achievedby hydrothermal processing.118 Such coatings are thicker andhigher in concentration of NT particles, yielding excellentresistance against discoloration. Nanoparticles can also bechemically generated in situ on wood and bamboo surfaces118
and this greatly enhances the material’s photostability. Thiswas demonstrated with Chinese hardwood where the in situgeneration was followed by silylation treatment to increasehydrophobicity. Wang and colleagues137 studied NT coatingson Chinese fir wood followed by silylation. However, such two-step processes, especially those involving silylation, are likelyto be too expensive for large-scale commercial use.
Role of fabric in protection against UV radiation
Personal protection afforded by clothing against the damagingeffects of solar UV radiation is considerable and depends onfiber composition, (natural, synthetic or mixed fibers), fabricconstruction (porosity, weave and thickness) and dyeing(natural or synthetic dyes) and their UV-absorbing pro-perties.138 In addition, outdoor uses of fabric such as in tents,awnings, shading and sunshade fabrics also provide a protec-tive role to humans from solar radiation. Clothing is in factone of the best ways of protecting people against solar UV radi-ation139 as even where the fabric materials themselvesundergo limited UV-induced degradation they continue toprovide excellent protection. The effectiveness of fabrics inthis regard is quantified using the “ultraviolet protectionfactor” or UPF of the fabric. UPF is the ratio of the minimumerythemal dose of solar UV radiation for skin protected byfabric to that unprotected.140 The percentage of effective UVradiation transmittance at each UPF value range is given inTable 1. For instance a cotton T-shirt provides a UPF of 3–5(dry) and even lower when wet while denim has a UPF 1700.Dark colored fabrics have a higher UPF compared to light-colored ones.141
Several features of fabric determine its UPF: (a) the porosityof the material or how open the structure is will be inversely
proportional to the UPF; (b) the absorption of UV by the fabricmaterial; (c) dyes, chemical reactants,43 or UV stabilizers142
applied to the fabric; and (d) moisture level of the fabric.143
Of these, the porosity or tightness of the weave is the domi-nant factor in obtaining UV radiation protection, followed bythe ability of fibers to absorb UV radiation. Grifoni et al.144
compared the UV protection afforded by natural-fiber fabrics(cotton, hemp, flax and ramie) and concluded that thickerfabrics with denser weaves (with a cover factor, CF > 94%) suchas drapery fabrics usually showed UPF > 50. Cover factor is theratio of the area covered by the yarns to the whole area of thefabric. Cotton fabrics afforded excellent protection from UVradiation as confirmed in other studies.142 Lighter textile-grade fabrics, however, showed high UPF values only when theporosity was low (Cover factor > 94%). Different natural fibershave about the same UV absorbance, and the difference in theperformance of corresponding fabrics is mostly determined bycover factor and dyeing. Commercial sun-protective garments(UPF 50+ that transmit <1% of UV radiation) perform by redu-cing their porosity or “open areas” in the weave of the fabricand using UV absorbers. Even these, when wet (with waterfilling the spaces) do not scatter but transmit UV, reducingtheir effective UPF value.
The UPF of different fabric materials varies as follows:Cotton > Polyester > Nylon > Elastane138 Chemical modifi-cation of fiber surfaces can increase the UPF ratings. A particu-larly successful approach is the use of inorganic oxides(including nanoscale particles145 or zinc oxide in poly-ester146,147 and in cotton148 and with titania-silica,149 aluminaor nanosilica150 particles in cotton, yielded very high UPFvalues in the hundreds (UPF > +50). While the untreated thick(tightly woven) fabric often has high UPF values to afford excel-lent protection, the use of surface modification technologiesallows even a lighter-weight fabric to deliver the same highlevel of UV protection.
Layer–by–layer (LBL) deposition of nanolayers of threebrightener compounds also increased the UPF of cotton fibers(UPF > 40) and improved durability in laundering as well.137
Optical whiteners absorb UV radiation and re-emit as blueradiation (typically 420–470 nm). LBL has also been used todeposit nano-alumina on cotton.151 Despite the impressiveperformance of LBL technologies, they are expensive and arenot near commercialization at this time. Chitosan metalcomplexes have also been used as organic stabilizer coatingson textile fibers but whilst these improve the antibacterial pro-
Table 1 The relationship between UPF and fraction of erythemallyeffective UV transmitted through the material. (From Australian/NewZealand Standard: AS/NZS 4399 (1996). Sun Protective Clothing: Evalu-ation and classification.)
UPF rangeUV radiationprotection category
Effective UV radiationtransmission (%)
15–24 Good protection 6.7–4.225–39 Very good protection 4.1–2.640–50, 50+ Excellent protection <2.5, <1%
Perspective Photochemical & Photobiological Sciences
178 | Photochem. Photobiol. Sci., 2015, 14, 170–184 This journal is © The Royal Society of Chemistry and Owner Societies 2015

perties significantly they only provide a moderate improvementin UPF.152
Glazing and protection against UV radiation
Window glass (glazing) filters out the solar UV-B radiation butallows some UV-A radiation to pass through; the transmittedfraction depends on the type of glazing used. In buildings andin automobiles where some of the solar radiation is screenedby glass, sunlight reaching into the cabin can still cause dis-coloration of materials and bleaching of dyes. In spite of thelower efficiency of UV-A wavelengths (compared to UV-B) in dis-coloration, the relatively greater amounts of the former in theterrestrial solar spectrum can still affect exposed fabric andother materials. Individuals, however, are protected from theUV-B radiation by almost all the different types of glass usedon residential, commercial and automobile applications.Reported values for transmission of UV-A by different glazingare summarized below (Table 2 153).
Thermal performance of windows can be improved dra-matically by incorporating monolithic or granular aerogel inthe interspace between panes in double-glazed windows.154
According to Buratti11 the rate of heat transfer decreased by23% in aluminium-framed windows using aerogel technology.An associated benefit is that the presence of aerogel also cutsdown the radiation transmitted, especially UV-A and UV-B radi-ation in sunlight.
References
1 Plastics Europe, Plastics Facts 2012, Plastics Europe,http://www.plastics europe.org, accessed August, 2014.
2 T. Ojeda, Degradability of linear polyolefins under naturalweathering, Polym. Degrad. Stab., 2011, 96, 703–707.
3 V. Sharratt, C. A. Hill and D. P. Kint, A study of earlycolour change due to simulated accelerated sunlightexposure in Scots pine (Pinus sylvestris), Polym. Degrad.Stab., 2009, 94, 1589–1594.
4 A. F. Bais, R. L. McKenzie, P. J. Aucamp, M. Ilyas,S. Madronich, G. Bernhard and K. Tourpali, Ozonedepletion and climate change: Impacts on UV radiation,Photochem. Photobiol. Sci., 2015, 14, this issue.
5 IPCC, Intergovernmental Panel on Climate Change WorkingGroup I Contribution to the IPCC Fifth Assessment ReportClimate Change 2013: The Physical Science Basis, ReportNo., University Press, Cambridge, United Kingdom andNew York, NY, USA, 2010.
6 M. C. Celina, Review of polymer oxidation and its relation-ship with materials performance and lifetime prediction,Polym. Degrad. Stab., 2013, 98, 2419–2429.
7 J. L. Pablos, C. Abrusci, I. Marín, J. López-Marín,F. Catalina, E. Espí and T. Corrales, Photodegradation ofpolyethylenes: Comparative effect of Fe and Ca-stearatesas pro-oxidant additives, Polym. Degrad. Stab., 2010, 95,2057–2064.
8 S. M. Al-Salem, Influence of natural and acceleratedweathering on various formulations of linear low densitypolyethylene (LLDPE) films, Mater. Des., 2009, 30, 1729–1736.
9 J. Lee-Taylor, S. Madronich, C. Fischer and B. Mayer, A Cli-matology of UV Radiation, 1979–2000, 65S–65N, in UVRadiation in Global Climate Change: Measurements, Model-ing and Effects on Ecosystems, eds. W. Gao, D. Schmoldtand J. R. Slusser, Springer-Verlag and Tsinghua UniversityPress, 2010, pp. 1–20.
10 E. Moretti, M. Zinzi and E. Belloni, Polycarbonatepanels for buildings: Experimental investigation ofthermal and optical performance, Energy Build., 2014, 70,23–35.
11 C. Buratti and E. Moretti, Glazing systems with silicaaerogel for energy savings in buildings, Appl. Energy, 2012,98, 396–403.
12 S. Kubba, Green Building Materials and Products, inHandbook of Green Building Design and Construction, ed.S. Kubba, Butterworth-Heinemann, Boston, 2012, pp.227–311.
13 G. Akovali, Plastic materials: polyvinyl chloride (PVC), inWoodhead Publishing Series in Civil and Structural Engineer-ing, eds. F. Pacheco-Torgal, S. Jalali and A. Fucic, Wood-head Publishing, 2012, pp. 23–53.
14 FerroCorp, Ferro and PolyOne target LDPE and PVCfoams with K debuts, Addit. Polym., 2014, 1, 4–5.
15 Arkema, PVC processing aid from Arkema Inc. sets newstandards, Addit. Polym., 2011, 2011, 2–3.
16 Arkema, Arkema unveils new impact modifier for windowprofiles, collaborates with Purac, Addit. Polym., 2010,2010, 2–3.
17 Akcros, Akcros USA announces new series of stabilizerproducts for PVC, Addit. Polym., 2010, 10, 1–2.
18 ISP, International Speciality Products reveals new addi-tives family for high-performance PVC applications, Addit.Polym., 2010, 2010, 1–2.
19 D. Sil and S. Chakrabarti, Photocatalytic degradation ofPVC–ZnO composite film under tropical sunlight and arti-
Table 2 Percentage transmission of UV-A radiation by different typesof glazing materials
Type of glazingThicknessa
(mm)Percentagetransmission
Residential windowsDouble glazed clear glass 3.0/3.0 57Double glazed tinted glass 3.0/3.0 20–33Double glazed laminated glass 3.0/6.0 0.5Double glazed UV-blocking glass 3.0/3.0 0.1
Automobile windowsLaminated windshield glass — 2–3Tempered glass in rear and sidewindow
— 33–48
Grey privacy glass — 8Moon-roof glass — 2
a The two numbers refer to the two sheets of glass in double-glazedwindows.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 170–184 | 179

ficial UV radiation: A comparative study, Sol. Energy, 2010,84, 476–485.
20 A. Amar and A. V. Parisi, Spectral response of solvent-castpolyvinyl chloride (PVC) thin film used as a long-term UVdosimeter, J. Photochem. Photobiol., B, 2013, 125, 115–120.
21 A. L. Andrady, Wavelength sensitivity of common poly-mers: A Review, Adv. Polym. Sci., 1996, 128, 45–94.
22 A. L. Andrady and N. D. Searle, Photoyellowing of mechan-ical pulps II Activation spectra for light-Induced yellowingof newsprint paper by polychromatic radiation, Tech.Assoc. Pulp Pap. Ind. J., 1995, 78, 131–138.
23 A. Heikkilä, P. Kärhä, A. Tanskanen, M. Kaunismaa,T. Koskela, J. Kaurola, T. Ture and S. Syrjälä, Characteriz-ing a UV chamber with mercury lamps for assessment ofcomparability tonatural UV conditions, Polym. Test., 2009,28, 57–65.
24 BCC Research Market Forecasting, Global market forwood-plastic composites to pass 46 million metric tonsby 2016, http://bccresearch.blogspot.ca/2011/11/global-market-for-wood-plastic.html. VC6DtxYd7WU, accessedAugust, 2014.
25 S. K. Najafi, Use of recycled plastics in wood plastic com-posites—a review, Waste Manage., 2013, 33, 1898–1905.
26 L. Sobczak, R. W. Lang and A. Haider, Polypropylene com-posites with natural fibers and wood – General mechanicalproperty profiles, Compos. Sci. Technol., 2012, 72, 550–557.
27 C. Homkhiew, T. Ratanawilai and W. Thongruang, Effectsof natural weathering on the properties of recycled poly-propylene composites reinforced with rubberwood flour,Ind. Crops Prod., 2014, 56, 52–59.
28 S. Y. Leu, T. H. Yang, S. F. Lo and T. H. Yang, Optimizedmaterial composition to improve the physical and mech-anical properties of extruded wood–plastic composites(WPCs), Constr. Build. Mater., 2012, 29, 120–127.
29 B. Nörnberg, E. Borchardt, G. A. Luinstra and J. Fromm,Wood plastic composites from poly(propylene carbonate)and poplar wood flour—Mechanical, thermal and mor-phological properties, Eur. Polym. J., 2014, 51, 167–176.
30 C. Guo, L. Li and Q. Wang, Investigation on the com-patibilizing effect of m-isopropenyl-α,α-dimethylbenzylisocyanate grafted polypropylene on polypropylene andwood flour composites, Wood Sci. Technol., 2012, 46, 257–270.
31 Y. Lei and Q. Wu, Wood plastic composites based onmicrofibrillar blends of high density polyethylene/poly-(ethylene terephthalate), Bioresour. Technol., 2010, 101,3665–3671.
32 K.-C. Hung, Y.-L. Chen and J.-H. Wu, Natural weatheringproperties of acetylated bamboo plastic composites,Polym. Degrad. Stab., 2012, 97, 1680–1685.
33 N. Ayrilmis and A. Kaymakci, Fast growing biomass asreinforcing filler in thermoplastic composites: Paulowniaelongata wood, Ind. Crops Prod., 2013, 43, 457–464.
34 M. A. Binhussain and M. M. El-Tonsy, Palm leave andplastic waste wood composite for out-door structures,Constr. Build. Mater., 2013, 47, 1431–1435.
35 M. C. N. Yemele, A. Koubaa, A. Cloutier, P. Souloungangaand M. Wolcott, Growth of ZnO nanofilms on wood withimproved photostability, Holzforschung, 2010, 64, 385–390.
36 S. H. Ghaffar and M. Fan, Structural analysis for lignincharacteristics in biomass straw, Biomass Bioenergy, 2013,57, 264–279.
37 A. Naumann, I. Stephan and M. Noll, Material resistanceof weathered wood-plastic composites against fungaldecay, Int. Biodeterior. Biodegrad., 2012, 75, 28–35.
38 S. Butylina, M. Hyvärinen and T. Kärki, Accelerated weath-ering of wood–polypropylene composites containing min-erals, Composites, Part A, 2012, 43, 2087–2094.
39 J. S. Fabilyi, A. G. McDonald, M. P. Wolcott andP. R. Griffiths, Wood plastic composites weathering:Visual appearance and chemical changes, Polym. Degrad.Stab., 2008, 93, 1405–1414.
40 D. B. Dittenber and H. V. S. Ganga Ra, Critical review ofrecent publications on use of natural composites in infra-structure, Composites, Part A, 2012, 43.
41 C. H. Lee, K. C. Hung and Y. L. Chen, Effects of polymericmatrix on accelerated UV weathering properties of wood-plastic composites, Holzforschung, 2012, 66, 981–987.
42 K. Chaochanchaikul, V. Rosarpitak and N. Sombatsompop,Photodegradation profiles of PVC compound and wood/PVC composites under UV weathering, eXPRESS Polym.Lett., 2013, 7, 146–160.
43 K. Xie, Y. Wang and L. Xu, Modification of cellulose withreactive polyhedral oligomericsilsesquioxane and nano-crosslinking effect on color properties of dyed cellulosematerials, Carbohydr. Polym., 2010, 80, 480–485.
44 K. Chaochanchaikul and N. Sombatsompop, Stabiliz-ations of molecular structures and mechanical propertiesof PVC and wood/PVC composites by Tinuvin and TiO2
stabilizers, Polym. Eng. Sci., 2011, 51, 1354–1365.45 L. K. Krehula, Z. Katančić, A. P. Siročić and Z. Hrnjak-
Murgić, Weathering of high-density polyethylene-woodplastic composites, J. Wood Chem. Technol., 2014, 34,39–54.
46 N. M. Stark and L. M. Matuana, Coextrusion of WPCs witha clear cap layer to improve color stability, in 4th WoodFibre Polymer Composites International Symposium, InstitutTechnologique, Bordeaux, France, 2009.
47 M. Manceau, A. Rivaton, J. L. Gardette, S. Guillerez andN. Lemaître, Light-induced degradation of the P3HT-based solar cells active layer, Sol. Energy Mater. Sol. Cells,2011, 95, 1315–1325.
48 S. Jin, N. M. Stark and L. M. Matuana, Influence of astabilized cap layer on the photodegradation of co-extruded high density polyethylene/wood-flour com-posites, J. Vinyl Addit. Technol., 2013, 19, 239–249.
49 T. Qiang, D. Yu, A. Zhang, H. Gao, Z. Li, Z. Liu, W. Chenand Z. Han, Life cycle assessment on polylactide-basedwood plastic composites toughened with polyhydroxy-alkanoates, J. Clean. Prod., 2014, 66, 139–145.
50 R. Vidal, P. Martinez and D. Garrain, Life cycle assessmentof composite materials made of recycled thermoplastics
Perspective Photochemical & Photobiological Sciences
180 | Photochem. Photobiol. Sci., 2015, 14, 170–184 This journal is © The Royal Society of Chemistry and Owner Societies 2015

combined with rice husks and cotton linters, Int. J. LifeCycle Assess., 2009, 14, 73–82.
51 A. L. Andrady, Microplastics in the marine environment,Mar. Pollut. Bull., 2011, 62, 1596–1605.
52 S. L. Wright, R. C. Thompson and T. Galloway, Thephysical impacts of microplastics on marine organisms:A review, Environ. Pollut., 2013, 178, 483–492.
53 J. A. Ivar do Sul and M. F. Costa, The present and future ofmicroplastic pollution in the marine environment,Environ. Pollut., 2014, 185, 352–364.
54 A. Bakir, S. J. Rowland and R. C. Thompson, Enhanceddesorption of persistent organic pollutants from micro-plastics under simulated physiological conditions,Environ. Pollut., 2014, 185, 16–23.
55 J. P. G. L. Frias, V. Otero and P. Sobral, Evidence of micro-plastics in samples of zooplankton from Portuguesecoastal waters, Mar. Environ. Res., 2014, 95, 89–95.
56 O. Setälä, F. V. Lehtinen and M. Lehtiniemi, Ingestion andtransfer of microplastics in the planktonic food web,Environ. Pollut., 2014, 185, 77–83.
57 A. Geburtig and V. Wachtendorf, Determination of thespectral sensitivity and temperature dependence of poly-propylene crack formation caused by UV-irradiation,Polym. Degrad. Stab., 2010, 95, 2118–2123.
58 L. Douminge, X. Feaugas, J. Bernard and S. Mallarino,Extrinsic fluorescence as a sensitive method for studyingphoto-degradation of high density polyethylene correlatedwith mechanical stresses, Curr. Appl. Phys., 2013, 13,1751–1757.
59 L. Douminge, S. Mallarino, S. Cohendoz, X. Feaugas andJ. Bernard, Extrinsic fluorescence as a sensitive methodfor studying photo-degradation of high density polyethy-lene Part 1, Curr. Appl. Phys., 2010, 10, 1211–1215.
60 M. Ramanujam, V. Wachtendorf, P. J. Purohit, R. Mix,A. Schönhals and J. F. Friedrich, A detailed dielectricrelaxation spectroscopy study of artificial UV weatheredlow density polyethylene, Thermochim. Acta, 2012, 530, 73–78.
61 S. Kahlen, G. M. Wallner and R. W. Lang, Aging behaviorof polymeric solar absorber materials—Part 1 Engineeringplastics, Sol. Energy, 2010, 84, 1567–1576.
62 T. Imai, S. Takamatsu, K. Shiraishi, K. Marumoto andT. Itoh, Photovoltaic textiles manufactured with precisiondie coating, Procedia Eng., 2012, 47, 502–505.
63 A. J. Moulé, Power from plastic, Curr. Opin. Solid StateMater. Sci., 2010, 14, 123–130.
64 A. Bedeloglu, A. Demir, Y. Bozkurt and N. S. Sariciftci,Photovoltaic properties of polymer based organic solarcells adapted for non-transparent substrates, RenewableEnergy, 2010, 35, 2301–2306.
65 Z. Zhao, L. Rice, H. Efstathiadis and P. Haldar, Annealingand thickness related performance and degradation ofpolymer solar cells, Microelectron. Reliab., 2013, 53, 123–128.
66 M. D. Kempe, Ultraviolet light test and evaluationmethods for encapsulants of photovoltaic modules, Sol.Energy Mater. Sol. Cells, 2010, 94, 246–253.
67 O. Haillant, Accelerated weathering testing principles toestimate the service life of organic PV modules, Sol.Energy Mater. Sol. Cells, 2011, 95, 1284–1292.
68 M. Kempe and J. Wohlgemuth, Evaluation of Temperatureand Humidity on PV Module Component Degradation, inProceedings of the 39th IEEE Photovoltaic Specialists Confer-ence, June 16–21, 2013, Tampa Convention Center inTampa Bay, FL, 2013.
69 E. S. Kopp, V. P. Lonji, A. E. Brooks, P. L. Hidalgo-Gonzalez and A. D. Cronin, I–V curves and visual inspec-tion of 250 PV modules deployed over 2 years in Tucson,Paper presented at the 38th IEEE Photovoltaic SpecialistsConference, Austin Convention Center in Austin, TX, 2012.
70 NREL, Photovoltaic Module Qualification Plus TestingNational Renewable Energy Laboratory National Renew-able Energy. Laboratory Report No. NREL/TP-5200-60950Golden, CO, December 2013, p. 30.
71 F. J. M. Johannes, G. Reisinger, T. Hofmann, J. Hinken,H. Tantau, A. Ulbrich, S. Lambrecht, B. V. Elsner andU. Schurr, Glass–film-combination: Opto-physical pro-perties and energy saving potential of a novel greenhouseglazing systems, Energy Build., 2012, 50, 298–307.
72 C. Lamnatou and D. Chemisana, Solar radiation manipu-lations and their role in greenhouse claddings: Fluo-rescent solar concentrators, photoselective and othermaterials, Renewable Sustainable Energy Rev., 2013, 27,175–190.
73 A. M. Abdel-Ghany and I. M. Al-Helal, Solar energy utiliz-ation by a greenhouse: General relations, RenewableEnergy, 2011, 36, 189–196.
74 C. Espejo, A. Arribas, F. Monzó and P. P. Díez, Nanocom-posite films with enhanced radiometric properties forgreenhouse covering applications, J. Coat. Technol. Res.,2012, 5, 2017–2209.
75 J. F. Bornman, P. W. Barnes, S. A. Robinson, C. L. Ballaré,S. D. Flinte and M. M. Caldwell, Solar ultraviolet radiationand ozone depletion-driven climate change: effects on ter-restrial ecosystems, Photochem. Photobiol. Sci., 2015, 14,This issue.
76 Clariant, Clariant introduces stabilizers for UV protectionof greenhouse films, Addit. Polym., 2013, 2013, 3–4.
77 A. Dehbi, A. Hamid, I. Mourad and A. Bouaza, Degra-dation assessment of LDPE multilayer films used as agreen house cover: Natural and artificial aging Impacts,J. Appl. Polym. Sci., 2012, 124, 2702–2716.
78 A. Dehbi, A. Bouaza, A. Hamou, B. Youssef and J. M. Saiter,Artificial ageing of tri-layer polyethylene film used as green-house cover under the effect of the temperature and theUV-A simultaneously, Mater. Des., 2010, 31, 864–869.
79 G. Oreski, G. M. Wallner and R. W. Lang, Ageing charac-terization of commercial ethylene copolymer greenhousefilms by analytical and mechanical methods, Biosyst. Eng.,2009, 103, 489–496.
80 A. Al-Mahdouri, M. Baneshi, H. Gonome, J. Okajima andS. Maruyama, Sol. Energy Mater. Sol. Cells, 2013, 96,21–32.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 170–184 | 181

81 H. H. Redhwi, M. N. Siddiqui, A. L. Andrady andS. Hussain, Durability of LDPE nanocomposite with claysilica, and zinc oxide II. Weatherability of the nanocompo-sites, Polym. Compos., 2013, 34, 1878–1883.
82 H. H. Redhwi, M. N. Siddiqui, A. L. Andrady andS. Hussain, Durability of LDPE nanocomposites with claySilica, and zinc oxide-Part 1: Mechanical properties of thenanocomposite materials, J. Nanocompos. Mater., 2013,2013, 1–6.
83 R. Gaggino, Water-resistant panels made from recycledplastics and resin, Constr. Build. Mater., 2012, 35, 468–482.
84 C. Kingston, R. Zepp, A. L. Andrady, D. Boverfof, R. Fehir,D. Hawkins, J. Roberts, P. Sayre, B. Shelton, Y. Sultan,V. Vejins and W. Whlleben, Release characteristics ofselected carbon nanotube polymer composites, Carbon,2014, 68, 33–57.
85 P. Bijl, A. Heikkilä, S. Syrjälä, A. Aarva and A. Poikonen,Modelling of sample surface temperature in an outdoorweathering test, Polym. Test., 2011, 30.
86 O. Haillant, D. Dumbleton and A. Zielnik, An Arrheniusapproach to estimating organic photovoltaic moduleweathering acceleration factors, Sol. Energy Mater. Sol.Cells, 2011, 95, 1889–1895.
87 A. François-Heude, E. Richaud, E. Desnoux and X. Colin,Influence of temperature, UV-light wavelength and inten-sity on polypropylene photothermal oxidation, Polym.Degrad. Stab., 2014, 100, 10–20.
88 L. Tolvaj, L. Persze and L. Albert, Thermal degradation ofwood during photodegradation, J. Photochem. Photobiol.,B, 2011, 105, 90–93.
89 L. Persze and L. Tolvaj, Photodegradation of wood at elev-ated temperature: Colour change, J. Photochem. Photobiol.,B, 2012, 10844–10847.
90 L. Tolvaj and K. Mitsui, Correlation between hue angleand lightness of light irradiated wood, Polym. Degrad.Stab., 2010, 95, 638–642.
91 X. Wang, S. Liu, H. Chang and J. Liu, Sol-gel deposition ofTiO2 nanocoatings on wood surfaces with enhancedhydrophobicity and photostability, Wood Fiber Sci., 2014,46, 109–117.
92 J. Nässén, S. Hedenus, S. Karlsson and J. Holmberg, Con-crete vs. wood in buildings – An energy system approach,Build. Environ., 2012, 51, 361–369.
93 R. M. Rowell, Handbook of wood chemistry and wood com-posite, Taylor and Francis, CRC Press, 2012.
94 P. D. Evans, S. Kraushaar Gibson, I. Cullis, C. Liu andG. Sèbe, Photostabilization of wood using low molecularweight phenol formaldehyde resin and hindered aminelight stabilizer, Polym. Degrad. Stab., 2013, 98, 158–168.
95 G. Pajchrowski, A. Noskowiak, A. Lewandowska andW. Strykowski, Wood as a building material in the light ofenvironmental assessment of full life cycle of four build-ings, Constr. Build. Mater., 2014, 52, 428–436.
96 L. Gustavsson, A. Joelsson and R. Sathre, Life cycleprimary energy use and carbon emission of an eight-
storey wood-framed apartment building, Energy Build.,2010, 42, 230–242.
97 L. Tolvaj and D. Varga, Photodegradation of timber ofthree hardwood species caused by different light sources,Acta Silvatic. Lignaria Hung., 2012, 8, 145–156.
98 V. Živković, M. Arnold, K. Radmanović, K. Richter andH. Turkulin, Spectral sensitivity in the photodegradationof fir wood (Abies alba Mill) surfaces: Colour changesin natural weathering, Wood Sci. Technol., 2014, 48, 239–252.
99 K. K. Pandey and T. Vuorinen, Comparative study ofphotodegradation of wood by a UV laser and xenon lightsource, Polym. Degrad. Stab., 2008, 93, 2138–2146.
100 G. Agresti, G, L. Bonifazi, G. Calienno, A. Capobianco,C. Lo Monaco, R. Pelosi, R. Picchio and S. Serranti,Surface investigation of photo-degraded wood by colourmonitoring, infrared spectroscopy, and hyperspectralimaging, J. Spectrosc., 2013, 380536.
101 T. Volkmer, L. Arietano, C. Plummer, J. Strautmann andM. Noël, Loss of tensile strength in cellulose tissue on thesurface of spruce (Picea abies) caused by natural photo-degradation and delignification, Polym. Degrad. Stab.,2013, 98, 1118–1125.
102 C. M. Popescu, M. C. Popescu and C. Vasile, Structuralanalysis of photodegraded lime wood by means of FT-IRand 2D IR correlation spectroscopy, Int. J. Biol. Macromol.,2011, 48, 667–675.
103 A. L. Andrady, S. H. Hamid, X. Hu and A. Torikai, Effectsof increased solar ultraviolet radiation on materials,J. Photochem. Photobiol., B, 1998, 46, 96–103.
104 A. Heikkilä and P. Kärhä, Photoyellowing revisited: Deter-mination of an action spectrum of newspaper, Polym.Degrad. Stab., 2014, 99, 190–195.
105 V. Živković, M. Arnold, K. K. Pandey, K. Richter andH. Turkulin, Novel approach in analysis of light sourceeffects to spectral sensitivity of fir wood surfaces, in 6th
European Weathering Symposium: Natural and ArtificialAgeing of Polymers, ed. T. Reichert, Gesellschaft fuerUmweltsimulation, Bratislava, Slovačka, 2013, p. 643363.
106 T. C. Chang, H. Chang, C. L. Wu, H. Y. Lin andS. T. Chang, Stabilizing effect of extractives on the photo-oxidation of Acacia confusa wood, Polym. Degrad. Stab.,2010, 95, 1518–1522.
107 P. Nzokou and D. P. Kamdem, Influence of wood extrac-tives on the photo–discoloration of wood surfaces exposedto artificial weathering, Color Res. Appl., 2006, 31, 425–234.
108 D. Grifoni, L. Bacci, S. d. Lonardo, P. Pinelli, S. Scardigli,S. Camilli, F. Sabatini, G. Zipoli and A. Romani, UVprotective properties of cotton and flax fabrics dyedwith multifunctional plant extracts, Dyes Pigm., 2014, 105,89–96.
109 C. Hu, G. Jiang, M. Xiao, J. Zhou and Z. Yi, Effects ofheat treatment on water-soluble extractives and colorchanges of Merbau heartwood, J. Wood Sci., 2012, 58, 465–469.
Perspective Photochemical & Photobiological Sciences
182 | Photochem. Photobiol. Sci., 2015, 14, 170–184 This journal is © The Royal Society of Chemistry and Owner Societies 2015

110 J. Salla, K. K. Pandey, G. K. Prakash and K. M. Mahadevan,Photobleaching and dimensional stability of rubberwoodesterified by fatty acid chlorides, J. Wood Chem. Technol.,2012, 32, 121–136.
111 B. Forsthuber and G. Grüll, The effects of HALS in the pre-vention of photo-degradation of acrylic clear topcoatsand wooden surfaces, Polym. Degrad. Stab., 2010, 95, 746–755.
112 G. De Filpo, A. M. Palermo, F. Rachiele and F. P. Nicoletta,Preventing fungal growth in wood by titanium dioxidenanoparticles, Int. Biodeterior. Biodegrad., 2013, 85, 217–222.
113 P. D. Evans, Weathering of wood and wood composites,in Handbook of wood chemistry and wood composite,ed. R. M. Row, Taylor and Francis, CRC, 2012,pp. 151–213.
114 S. Vollmer and P. D. Evans, Performance of clear coatingson modified wood exposed to the weather for 2 years inAustralia, Int. Wood Prod. J., 2013, 4, 177–182.
115 B. S. W. Dawson, A. P. Singh, H. W. Kroese,M. A. Schwitzer, S. Gallagher, S. J. Riddiough and S. Wu,Enhancing exterior performance of clear coatings throughphotostabilisation of wooden surfaces Part 2: Coating andweathering performance, J. Coat. Technol. Res., 2008, 5,207–219.
116 P. A. van Meel, S. J. F. Erich, H. P. Huinink, K. Kopinga,J. Jong and O. C. G. Adan, Moisture transport in coatedwood, Prog. Org. Coat., 2011, 72, 686–694.
117 P. L. Chou, J. T. Chang, T. F. Yeh and S. Chang, Character-izing the conservation effect of clear coatings on photo-degradation of wood, Bioresour. Technol., 2008, 99, 1073–1079.
118 B. Lesar, M. Pavli, M. Petri, A. Seve, M. Skapin andM. Humar, Wax treatment of wood slows photodegrada-tion, Polym. Degrad. Stab., 2011, 96, 1271–1278.
119 K. K. Pandey and K. Srinivasa, Study of UV resistance andnatural weathering of coatings on chemically modifiedwood, International Research Group on Wood ProtectionReport No., Stockholm, Sweden, 2013.
120 S. Grelier, A. Castellan and L. Podgorski, Use of low mole-cular weight modified polystyrene to prevent photodegra-dation of clear softwoods for outdoor use, Polym. Degrad.Stab., 2007, 92, 1520–1527.
121 S. Olsson, J. M, M. Westin and E. Östmark, Grafting of2-hydroxy-4 (2, 3-epoxypropoxy)-benzophenone and epoxi-dized soybean oil to wood: Reaction conditions andeffects on the color stability of Scots pine, Polym. Degrad.Stab., 2012, 97, 1779–1786.
122 P. D. Evans, Review of the weathering and photostabilityof modified wood, Wood Mater. Sci. Eng., 2009, 4, 2–13.
123 I. Bhat, A. Khalil, K. B. Awang, I. O. Bakare andA. M. Issam, Effect of weathering on physical, mechanicaland morphological properties of chemically modifiedwood materials, Mater. Des., 2010, 31.
124 K. Mitsui, Acetylation of wood causes photobleaching,J. Photochem. Photobiol., B, 2010, 101, 210–214.
125 M. Jebrane, G. Sèbe, I. Cullis and P. D. Evans, Photostabi-lisation of wood using aromatic vinyl esters, Polym.Degrad. Stab., 2009, 94, 151–157.
126 T. Yuan, L. M. Zhang, F. Xu and R. C. Sun, Enhancedphotostability and thermal stability of wood by benzoyla-tion in an ionic liquid system, Ind. Crops Prod., 2013, 53,123–128.
127 S. I. Baur and A. J. Easteal, Improved photo-protection ofwood by chemical modification with silanes: NMR andESR studies, Polym. Adv. Technol., 2013, 24, 97–103.
128 D. Fauchadour, T. Jeanson, J. N. Bousseau andB. Echalier, Nanoparticles of cerium oxide, application tocoating technologies, Paint Coat. Ind., 2005, 21, 88–93.
129 A. Saadat-Monfared and M. Mohseni, Polyurethane nano-composite films containing nano-cerium oxide as UVabsorber; Part 2: Structural and mechanical studies uponUV exposure, Colloids Surf., A, 2014, 441, 752–757.
130 F. Bezati, V. Massardier, J. Balcaen and D. Froelich, Astudy on the dispersion, preparation, characterization andphoto-degradation of polypropylene traced with rare earthoxides, Polym. Degrad. Stab., 2011, 96, 51–59.
131 R. A. Mohammad, M. Mohseni, S. M. Mirabedini andM. H. Tabatabaei, Surface analysis and anti-graffiti behav-ior of a weathered polyurethane-based coating embeddedwith hydrophobic nano silica, Appl. Surf. Sci., 2012, 258,4391–4396.
132 M. Rashvand, Z. Ranjbar and S. Rastegar, Nano zinc oxideas a UV-stabilizer for aromatic polyurethane coatings,Prog. Org. Coat., 2011, 71, 362–368.
133 M. V. Cristea, B. Riedl and P. Blanchet, Effect of additionof nanosized UV absorbers on the physico-mechanicaland thermal properties of an exterior waterborne stain forwood, Prog. Org. Coat., 2011, 72, 755–762.
134 S. M. Fufa, B. P. Jelle and P. J. Hovde, Effects of TiO2
and clay nanoparticles loading on weathering perform-ance of coated wood, Prog. Org. Coat., 2013, 76, 1425–1429.
135 S. M. Fufa, B. P. Jelle and P. J. Hovde, Weathering per-formance of spruce coated with water based acrylic paintmodified with TiO2 and clay nanoparticles, Prog. Org.Coat., 2013, 76, 1543–1548.
136 B. Forsthuber, C. Schaller and G. Grull, Evaluation of thephoto stabilising efficiency of clear coatings comprisingorganic UV absorbers and mineral UV screeners on woodsurface, Wood Sci. Technol., 2013, 47, 281–297.
137 Q. Wang and P. J. Hauser, Developing a novel UV protec-tion process for cotton based on layer-by-layer self-assem-bly, Carbohydr. Polym., 2010, 81, 491–496.
138 P. Gies and A. McLennan, Everyday and high-UPF sun-protective clothing, Melanom Lett., 2012, 30, 7–8.
139 H. Z. Maged, B. E. H. Mamdouh, M. A. El-Naggar,A. Fathalla and M. A. Nisreen, Novel UV-protective formu-lations for cotton, PET fabrics and their blend utilizingirradiation technique, Eur. Polym. J., 2009, 45, 2926–2934.
140 B. Das, UV radiation protective clothing, Open Text. J.,2010, 3, 14–21.
Photochemical & Photobiological Sciences Perspective
This journal is © The Royal Society of Chemistry and Owner Societies 2015 Photochem. Photobiol. Sci., 2015, 14, 170–184 | 183

141 P. Kullavanijaya and H. W. Lim, Photoprotection, J. Am.Acad. Dermatol., 2005, 52, 937–958.
142 C. Kan and L. S. Yam, Effect of stretching onultraviolet protection of cotton and cotton/coolmax-blended weft knitted fabric in a wet state, Materials, 2014,7, 58–74.
143 C. A. Wilson, N. K. Bevin, R. M. Laing and B. E. Niven,Solar protection - effect of selected fabric and use charac-teristics on ultraviolet transmission, Text. Res. J., 2008, 78,95–104.
144 D. Grifoni, L. Bacci, G. Zipoli, L. Albanese and F. Sabatini,The role of natural dyes in the UV protection offabrics made of vegetable fibres, Dyes Pigm., 2011, 91,279–285.
145 D. Mihailović, Z. Šaponjić, M. Radoičić, T. Radetić,P. Jovančić, J. Nedeljković and M. Radetić, Functionali-zation of polyester fabrics with alginates and TiO2 nano-particles, Carbohydr. Polym., 2010, 79, 526–532.
146 B. A. Çakır, L. Budama, Ö. Topel and N. Hoda, Synthesisof ZnO nanoparticles using PS-b-PAA reverse micelle coresfor UV protective, self-cleaning and antibacterial textileapplications, Colloids Surf., A, 2012, 414, 132–139.
147 Z. Zhou, Y. Zhao and Z. Cai, Low-temperature growth ofZnO nanorods on PET fabrics with two-step hydrothermalmethod, Appl. Surf. Sci., 2010, 256, 4724–4728.
148 M. M. Abd El-Hady, A. Farouk and S. Sharaf, Flame retar-dancy and UV protection of cotton based fabrics usingnano-ZnO and polycarboxylic acids, Carbohydr. Polym.,2013, 92, 400–406.
149 D. Fakin, N. Veronovski, A. Ojstršek and M. Božič, Syn-thesis of TiO2–SiO2 colloid and its performance in reactivedyeing of cotton fabrics, Carbohydr. Polym., 2012, 88, 992–1001.
150 X. Liu, R. Song, W. Yu and N. Pan, Developing UV-protec-tive cotton fabrics based on SiOx nanoparticles, FibersPolym., 2012, 13, 489–494.
151 S. S. Uğur, M. Sarıışık and H. A. Aktaş, Nano-Al2O3 multi-layer film deposition on cotton fabrics by layer-by-layer deposition method, Mater. Res. Bull., 2011, 46, 1202–1206.
152 M. Gouda and S. M. Keshk, Evaluation of multifunctionalproperties of cotton fabric based on metal/chitosan film,Carbohydr. Polym., 2010, 80.
153 C. Tuchinda, S. Srivannaboon and H. W. Lim, Photopro-tection by window glass, automobile glass, and sun-glasses, J. Am. Acad. Dermatol., 2006, 54, 845–854.
154 J. T. Kim and M. S. Todorovic, Tuning control of buildingsglazing’s transmittance dependence on the solar radiationwavelength to optimize daylighting and building’s energyefficiency, Energy Build., 2013, 63, 108–118.
Perspective Photochemical & Photobiological Sciences
184 | Photochem. Photobiol. Sci., 2015, 14, 170–184 This journal is © The Royal Society of Chemistry and Owner Societies 2015