transcellular passage of neisseria meningitidis across a polarised
TRANSCRIPT

1
Transcellular passage of Neisseria meningitidis across a polarised respiratory
epithelium 2
Thomas C. Sutherland, Paola Quattroni, Rachel M. Exley, Christoph M Tang* 4
Centre for Molecular Microbiology and Infection, 6
Flowers Building,
Imperial College London, 8
London,
SW7 2AZ. 10
*For correspondence: Christoph Tang 12
Telephone: +44 (0)20 7594 3072 14
Fax: +44 (0)20 7594 3095
E-mail: [email protected] 16
18
Running Title: Epithelial traversal by N. meningitidis
20
Copyright © 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Infect. Immun. doi:10.1128/IAI.01377-09 IAI Accepts, published online ahead of print on 28 June 2010
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

2
Abstract
Neisseria meningitidis is a major cause of sepsis and meningitis but is also a 22
common commensal, present in the nasopharynx of between 8 to 20% of
healthy individuals. During carriage, the bacterium is found on the surface of 24
the nasopharyngeal epithelium and in deeper tissues, while to develop disease
the meningococcus must spread across the respiratory epithelium and enter 26
the systemic circulation. Therefore investigating the pathways by which N.
meningitidis crosses the epithelial barrier is relevant for understanding 28
carriage and disease, but has been hindered by the lack of appropriate models.
Here, we have established a physiologically relevant model of the upper 30
respiratory epithelial cell barrier to investigate the mechanisms responsible for
traversal of N. meningitidis. Calu-3 human respiratory epithelial cells were 32
grown on permeable cell culture membranes to form polarised monolayers of
cells joined by tight junctions. We show that the meningococcus crosses the 34
epithelial cell barrier by a transcellular route; traversal of the layer did not
disrupt its integrity and bacteria were detected within the cells of the 36
monolayer. We demonstrate that successful traversal of the epithelial cell
barrier by N. meningitidis requires expression of its Type four pili (Tfp) and 38
capsule, and is dependent on the host cell microtubule network. The Calu-3
model should be suitable for dissecting the pathogenesis of infections caused 40
by other respiratory pathogens as well as the meningococcus.
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

3
Introduction 42
Neisseria meningitidis is a leading cause of fatal sepsis and meningitis worldwide
(61). Despite being an important human pathogen, the bacterium is also a common 44
commensal, found in the nasopharynx of between 8 and 20% of healthy individuals
(25, 62). The human nasopharynx is lined by a columnar epithelium which forms the 46
first cellular barrier encountered by the meningococcus following its acquisition. This
cell layer consists of differentiated, polarised, respiratory epithelial cells joined by 48
tight junctions that form a barrier which excludes mucosal pathogens. The majority
of cells in the layer are ciliated, resulting in a brush border, although areas of un-50
ciliated cells are also present along with mucus secreting goblet cells (54). To cause
disease the meningococcus must spread from the nasopharynx, its only natural 52
reservoir, penetrate the upper respiratory epithelial cell barrier and enter the systemic
circulation. Furthermore, during asymptomatic colonisation bacteria are not confined 54
to the epithelial surface of the nasopharynx, but are also found in clusters beneath
the epithelial cell layer in tonsillar tissue (57). Therefore defining the route and 56
mechanisms of passage of the respiratory epithelial cell layer by N. meningitidis are
relevant for understanding both meningococcal carriage and disease. 58
Traversal of an epithelial barrier by a pathogen can be considered to consist of a 60
number of steps, which include i) adhesion to the apical surface, ii) invasion of
epithelial cells, iii) survival within cells, iv) movement to the basal side of cells, and iv) 62
escape from the basolateral aspect of the epithelium. The initial step in traversal by
N. meningitidis involves attachment of bacteria to epithelial cells. Work with 64
nasopharyngeal tissue in an organ culture model indicates that meningococcal
attachment is largely limited to non-ciliated cells of the respiratory epithelium (63). 66
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

4
For encapsulated meningococci, adhesion is mediated by bacterial surface-
associated filamentous structures called Type four pili (Tfp) (42, 46, 50, 64) which are 68
also required for colonisation of human nasopharyngeal tissue (16). Tfp perform
several important functions including DNA uptake (77), twitching motility (43) and 70
bacterial aggregation (28), and have been proposed to mediate meningococcal
attachment to host cells by binding to CD46 (33). After initial adhesion, N. 72
meningitidis forms microcolonies on the apical surface of epithelial cells and the
microvilli of infected cells becoming elongated and interweave through the 74
microcolonies (51, 63). Bacteria replicate within the microcolonies then disperse
across the cells in a Tfp-dependent manner (51). Following dispersal, the pili retract 76
and bacteria become tightly associated with the host plasma membrane; at this stage
bacteria may be internalised (50, 60, 63). 78
While the mechanisms that mediate attachment of N. meningitidis to epithelial cells 80
are well understood, there is a paucity of information about the subsequent steps
involved in traversal of the epithelial cell barrier. Indeed, there are even conflicting 82
data regarding the route of traversal with both paracellular and transcellular routes
identified in experiments with endometrial and gastrointestinal cells (3, 42, 50). 84
Although the related pathogen, Neisseria gonorrhoeae migrates through cells (76), it
is not possible to extrapolate findings from this unencapsulated bacterium to the 86
meningococcus, given the proposed role of capsule for survival within cells (60).
Therefore the aim of this study was to determine the route of traversal of N. 88
meningitidis across the respiratory epithelial cell barrier and define host and
pathogen factors involved in this key step in pathogenesis. We used Calu-3 cells to 90
establish a model for meningococcal traversal of the respiratory epithelial barrier.
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

5
Calu-3 human bronchial epithelial cells are one of the few respiratory cell lines that 92
differentiate into polarised monolayers when grown on porous membranes in vitro
(14, 20, 21, 56). Growing the cells in this way allows bacteria to escape from the 94
basolateral surface and provides them with the epithelial cell barrier encountered in
the nasopharynx without the need for organ culture models, which are not well suited 96
for defining mechanisms of cell entry and passage. Calu-3 monolayers display
features of differentiated human airway epithelium with junctional complexes (24, 56), 98
apical-basal differentiation, and possess transport and metabolism functions (20).
These characteristics have led to Calu-3 cells becoming widely used for in vitro 100
studies of drug delivery and toxicology of the human respiratory epithelium (5, 7, 14,
17, 38, 49). We show that N. meningitidis crosses the respiratory epithelial 102
monolayer by a transcellular route without disrupting the barrier function of the layer.
We also demonstrate that the bacterial capsule and Tfp are important for this 104
process. These structures are also required for passage when cells are grown with
an air interface. On the host side, an intact microtubule network is necessary for 106
efficient traversal while we observed no effect of the proposed Tfp receptor, CD46.
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

6
Materials and Methods 108
Bacterial strains and growth conditions
N. meningitidis was cultured in Brain Heart Infusion (BHI) broth (Oxoid) or on BHI 110
agar (1.5% wt/vol, Agar No.1, Oxoid) with 5% Levinthal’s supplement. Escherichia
coli was grown in Luria-Bertani (LB) broth (Gibco) at 37°C with shaking, or on LB 112
agar. Details of the strains used in this work are shown in Table 1. Antibiotics were
used at the following concentrations: kanamycin, 50 and 100 µg ml-1 and 114
erythromycin, 200 and 2 µg ml-1, for E. coli and N. meningitidis respectively, and
carbenicillin at 50 µg ml-1. 116
Cell culture 118
Calu-3 epithelial cells were cultured in DMEM:F12 (Invitrogen), while Chang epithelial
cells were propagated in RPMI 1640 (Invitrogen). Media were supplemented with 120
GlutamaxTM (Invitrogen) and 10% foetal calf serum (PAA Laboratories). All cells were
grown at 37°C in 5% CO2. Calu-3 monolayers were grown on approximately 1 cm2, 1 122
µm pore size, BD Falcon™ cell culture inserts (BD Bioscience) containing
polyethylene terephthalate membranes in 12 well tissue culture companion plates 124
(BD Bioscience). Cells (4 x 105) were seeded onto the apical side of membranes.
For assays performed with liquid covered culture (LCC), layers were maintained with 126
1 ml of culture medium in the apical chamber and 1.5 ml in the basolateral chamber.
Cells were allowed to grow and differentiate for five days, with the media changed 128
every second day. Air interface culture (AIC) monolayers were seeded and
maintained as above except that the apical media was removed after two days. The 130
trans-epithelial electrical resistance (TEER) across monolayers was measured with
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

7
an EVOM TEER meter attached to STX chopstick electrodes (World Precision 132
Instruments); only monolayers with TEER of over 1400 Ω were used in experiments.
134
Texas Red labelled 70,000 MW dextran (final concentration 0.1 µM, Invitrogen) was
added to the apical chamber of cell culture wells. Samples were taken from the 136
basolateral chamber at times afterwards, and fluorescence analysed with a
spectrophotometer (Varian Cary Eclipse). For microscopy, cover slips were seeded 138
with Calu-3 cells at a density of 4 x 105 cells/well and Chang cells at a density of 5 x
104 cells/well in 24 well plates. Cells were grown to semi-confluency overnight. 140
Challenge of cells with bacteria 142
All inocula consisted of 1 ml of a bacterial suspension per tissue culture well except
for infection of AIC layers where a volume of 200 µl per well was used. Bacteria 144
were harvested following overnight culture on solid media and re-suspended in 400
µl of PBS. The concentration of the bacterial suspension was adjusted in culture 146
media to give the desired multiplicity of infection (MOI). For traversal assays, media
in the basolateral chambers of monolayers was replaced immediately prior to 148
challenge. For competitive infections (2), inocula consisting of a 1:1 ratio of mutant
to wild-type bacteria with a final MOI of 40 were prepared and added to the apical 150
chamber. The ratio of bacterial strains in the inoculum was determined by serial
dilution and plating to media with and without antibiotics. After seven hours, cell 152
culture inserts containing infected monolayers were washed then transferred to new
wells containing 1.5 ml of culture media in the basolateral chamber and left for one 154
hour. The ratio of strains in the basolateral chamber was determined by plating to
solid media. This procedure was repeated at 23 hours post-challenge. The 156
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

8
competitive index was calculated by dividing the ratio of mutant to wild-type in the
output by the ratio in the input, and was expressed logarithmically as the logCI. 158
Adhesion assays were performed by inoculating polarised monolayers with bacteria 160
at an MOI of 40. After three hours monolayers were rinsed with Hank’s buffered
saline solution (HBSS) (Invitrogen) then treated with 1% saponin to lyse cells and 162
release all cell associated bacteria. Numbers of cell associated bacteria were
enumerated by plating to solid BHI media. The influence of purified CD46 on 164
adhesion was assessed by adding the purified protein to bacteria prior to infection of
cells. The inoculum was prepared at the required MOI, and 6.25 µg of purified CD46 166
(consisting of the extracellular domain alone) were added per 107 bacteria. Bacteria
and CD46 were incubated together at 37oC for 30 minutes, and then added to cells. 168
When required, different anti-CD46 mAbs or an isotype control at a concentration of
2 µg/ml were added to the infection medium for three hours. Details of antibodies 170
used in this study are given in Table 2.
172
For recovery of intracellular bacteria, Calu-3 monolayers were challenged with
bacteria at an MOI of 40. At eight or 24 hours after inoculation, monolayers were 174
washed three times with PBS on their apical and basolateral surfaces then incubated
with 200 µg/ml gentamicin for 30 minutes, again in both the apical and basolateral 176
chambers. Next, cell culture inserts were washed three times with PBS, and cells
were lysed by a 5 minute treatment with 1% saponin in PBS. The number of 178
intracellular bacteria was established by serial dilution of lysates and plating to solid
media. 180
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

9
Chemicals and Inhibitors 182
Taxol and nocodazole (Sigma) were dissolved in DMSO and used at a final
concentration of 2 µM. Nocodazole was added to media on the apical surface of 184
cells which were then incubated at 4oC for 45 minutes, then left at 37⁰C for 45
minutes prior to challenge. Taxol was added to the cells at the time of the challenge. 186
The number of bacteria traversing monolayers was expressed as the proportion of
the number of bacteria recovered from the basolateral chamber of wells containing 188
no cells. Latrunculin A (Sigma) was used at a final concentration of 1 µM. In all
cases the final concentration of DMSO in culture wells never exceeded 0.1%. 190
Fixation, labelling and staining 192
Antibodies were diluted in 10% horse serum in PBS. Samples for confocal
microscopy were washed twice with PBS, fixed for 15 minutes at room temperature in 194
3% paraformaldehyde (PFA) in PBS, then washed three times with PBS. For
labelling of intracellular and extracellular bacteria, cells were incubated for one hour 196
with an α lipopolysaccharide (LPS) antibody (αL3,7,9) after which cells were washed
twice in PBS then incubated for 30 minutes with an α-mouse RRX conjugated 198
secondary antibody to label extracellular bacteria. Cells were permeablised by
treatment with 0.2% Triton X-100 (T X-100) in PBS and the labelling process was 200
repeated except a Cy2 conjugated α-mouse secondary antibody was used to label
both intracellular and extracellular bacteria. Host cell structures were labelled after 202
permeabilisation; actin was stained with fluorescently labelled phalloidin. After
labelling, cover slips were washed twice in PBS, then in ddH2O, mounted in ProLong 204
Gold (Invitrogen) and analysed by fluorescence microscopy (BX40; Olympus) or a
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

10
confocal laser scanning microscopy (LSM510, Zeiss). Images were imported into 206
Adobe Photoshop CS2 for processing.
208
For TEM, samples were washed three times on ice with ice cold PBS then immersed
in EM grade glutaraldehyde (Agar Scientific) diluted to 0.5% in 200 mM sodium 210
cocodylate (Agar Scientific) at room temperature for 30 minutes. Next samples were
washed three times in 200 mM sodium cocodylate buffer and post-fixed in 1% 212
osmium tetroxide-1.5% potassium ferrocyanide at room temperature for one hour.
Cells were then washed in water, incubated in 0.5% magnesium-uranyl acetate 214
overnight at 4°C, washed again in water, dehydrated in ethanol, and embedded in
Epon. Cross sections were cut through fixed monolayers and placed on EM grids. 216
Lead citrate was added for contrast and grids were then examined using a Biacore
2000 transmission electron microscope. 218
Tfp and capsule expression 220
Monolayers were infected with MC58 at an MOI of 40 as described above. After 24
hours bacteria were recovered from the apical and basal chambers. A total of 10 222
colonies from the apical and 10 from the basal chambers were grown overnight on
solid media then harvested and resuspended in 0.5 ml PBS (Sigma, U.K.). For slot 224
blot analysis of Tfp expression, samples were combined with an equal volume of 2x
SDS-PAGE sample buffer and boiled for 10 minutes, then transferred to immubilon P 226
polyvinylidene fluoride (PVDF) membranes (Millipore) using a MINIFOLD slot blot
system (Whatman). The membranes were blocked in 5% milk in PBS for one hour at 228
room temperature with gentle shaking. Relevant antibodies were diluted in PBSTM
(PBS, 0.05% Tween 20, 0.5% skimmed milk), and incubated with the PVDF 230
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

11
membrane for 1-2 hours, followed by three washes with PBSTM for ten minutes
each. Following incubation with the primary mAb (SM1 which recognises class I pilin, 232
(70)), the PVDF membrane was washed with PBSTM three times for ten minutes
each. A goat anti-murine immunoglobulin antibody conjugated to horse radish 234
peroxidase (Dakocytomation) was used as the secondary antibody. Antibody binding
was detected using ECL Western Blotting detection reagents (GE Healthcare). 236
Capsule expression was analysed by ELISA. Suspensions of bacteria were 238
prepared in PBS as described above, and diluted to a concentration of 1x109.
Bacteria were heated at 56oC for 1 hour and then re-suspended in ELISA coating 240
buffer (Sigma). Samples (100 µl) were added to wells of 96 well plates and left
overnight at 4oC. Wells with coating buffer alone or those coated with MC58∆siaD 242
were included as controls. Next plates were washed with water, then incubated for
two hours with 2% bovine serum albumin (BSA) in PBS. Following blocking, plates 244
were washed three times with PBS/Tween 0.1% then incubated with α-serogroup B
capsule mAb for one hour at 37 oC. Plates were washed three times in PBS/Tween 246
0.1% and incubated with α murine Ig goat pAb conjugated to horse radish peroxidase
(αM HRP) for a further hour at 37 oC. After further washes, 100 µl of Sigmafast OPD 248
substrate (Sigma) was added to plates for six minutes, and the reaction stopped with
100 µl of 0.3 M HCl. Absorbance was read at 492 nm using Ascent software. 250
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

12
Results 252
Interactions between Calu-3 respiratory epithelial cells and N. meningitidis
Initially we compared the interactions of the meningococcus with Calu-3 and Chang 254
cells grown to semi-confluency then challenged with MC58, a clinical serogroup B
isolate of N. meningitidis, at a multiplicity of infection (MOI) of 100, which is in the 256
range of MOIs employed in other studies. This MOI represents a midpoint in a large
range of MOIs used to study the interaction of N. meningitidis with Chang cells (4, 258
73). MC58 adhered to and invaded into Calu-3 and Chang cells at similar levels
(average no. of adherent bacteria per cell 17.4 and 20 for Chang and Calu-3 cells, 260
respectively; average no. of internalised bacteria per cell 2.18 and 2.21 for Chang
and Calu-3 cells, respectively, Fig. S1). Furthermore adhesion of N. meningitidis to 262
Calu-3 cells was facilitated by Tfp (Fig. S1), in keeping with results using other
epithelial cell lines (47, 50, 69), and Calu-3 cells express CEACAMS (not shown), 264
which are receptors for Neisseria Opas (75).
266
As N. meningitidis adheres to and enters into Calu-3 cells, we next grew the cells on
permeable membranes in cell culture inserts so they formed a barrier between the 268
apical and basolateral chambers (Fig. S2). The development of tight junctions
between cells was monitored by measuring the TEER across layers. The level of 270
TEER across Calu-3 cells was influenced by a number of factors. Reducing the size
of pores in membranes led to higher levels of TEER but pores of less than 1 µm 272
impeded the passage of bacteria (not shown). Therefore we selected membranes
with 1 µm pores for all subsequent studies, which with seeding cells at a density of 4 274
x 105 per cm2, gave consistent levels of TEER of around 1850 Ω after 5 days (Fig.
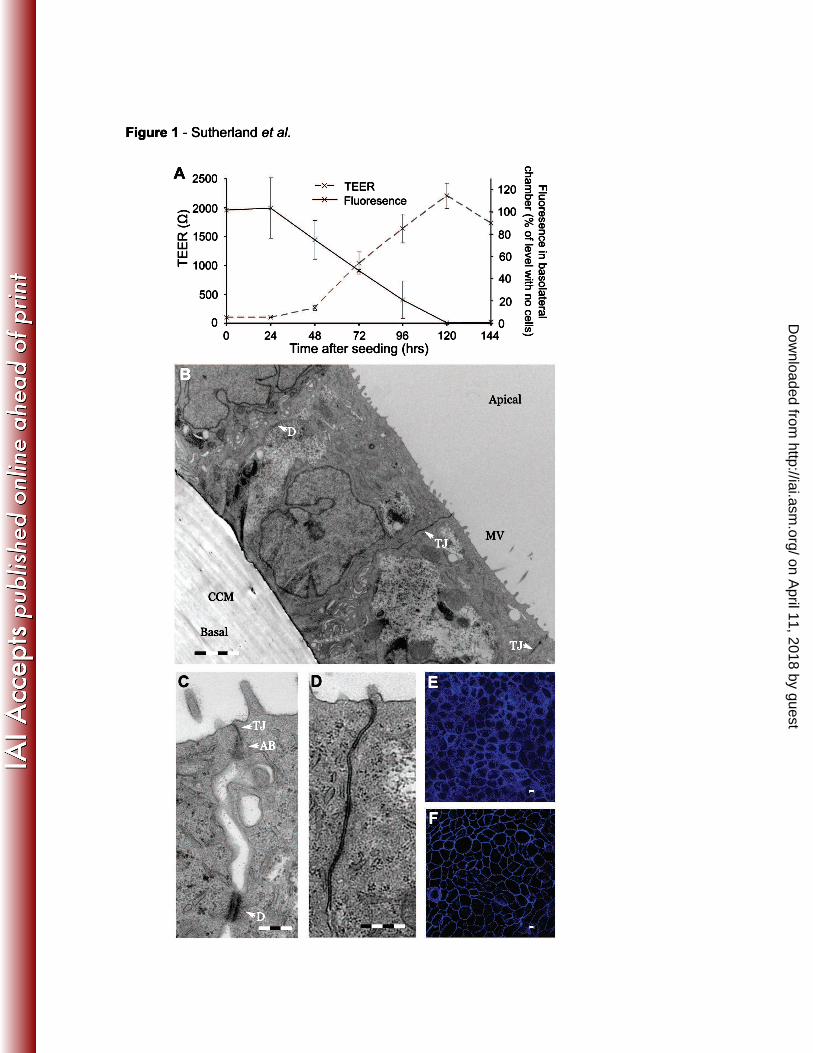
1A). The development of the epithelial cell barrier following seeding of wells was 276
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

13
also demonstrated by permeability of the monolayer to fluorescently labelled 70,000
MW dextran added to the apical chamber. Increasing TEER over time coincided with 278
decreasing permeability of the layer to the tracer (Fig. 1A), demonstrating that the
cells formed an effective barrier within five days of seeding. 280
Furthermore transmission electron microscopy (TEM) confirmed that Calu-3 cells 282
formed confluent monolayers with morphological features of a polarised respiratory
epithelium (Figs. 1B, C and D) consisting of a continuous monolayer of columnar 284
cells linked by junctional structures. There was apical-basal polarisation, with nuclei
located towards the basolateral side of cells, the development of tight junctions, and 286
microvilli on the apical surface (Fig. 1B). Full junctional complexes (with tight
junctions, adhesion belts and desmosomes) were present between cells (Fig. 1C), 288
and the membranes of adjacent cells were barely distinguishable (Fig. 1D). Immuno-
fluorescence microscopy of the tight junction proteins zonula occludens-1 (ZO-1, Fig. 290
1E) and occludin (Fig. 1F) showed the characteristic chicken-wire pattern formed by
their continuous presence at the margins of cells. At this time, while the tracer 292
diffused freely into the basolateral chamber in the absence of cells (Fig. S3A), no
tracer was detected in the basolateral chamber even after 24 hours (Fig. S3B). 294
Additionally, we challenged the apical surface of the monolayer with non-invasive E.
coli DH5α and monitored its appearance in the basolateral chamber over time. When 296
no cells were present on the cell culture membrane, E. coli was detected in the
basolateral chamber within two hours of challenge (Fig. S3C). In contrast, no 298
bacteria were detected in the presence of Calu-3 cell monolayers (Fig. S3D) even
after eight hours. 300
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

14
N. meningitidis traverses the epithelial cell barrier by a transcellular route 302
Next we investigated the traversal of N. meningitidis across Calu-3 monolayers five
days after seeding. The apical surfaces of layers were challenged with MC58 at an 304
MOI of 40, and eight and 24 hours later, the inserts containing infected membranes
were washed thoroughly in media then transferred into new wells. After one hour, 306
samples were taken from the basal chambers of the new wells and were plated to
BHI. N. meningitidis was detected traversing in small numbers (equivalent to 308
0.0035% of input) at eight hours post-challenge. By 24 hours, the number of bacteria
traversing over the course of one hour had increased by around four orders of 310
magnitude (Fig. 2A). Bacteria could occasionally be found in the basal chamber prior
to 8 hours but never earlier than six hours (not shown). At 16 hours intermediate 312
levels of traversing bacteria were found (not shown).
314
Traversal of the monolayer by N. meningitidis might result from loss of integrity of the
tight junctions or cell death. Therefore, the TEER across the monolayer was 316
monitored during the course of an infection. Although the TEER across the layer fell
following bacterial challenge, levels never dropped to below 40% of initial readings 318
and also fell in wells infected with E. coli and the uninfected control wells (Fig. 2B).
The integrity of the layer during infection was also monitored by adding the 320
fluorescent tracer to the apical surface of the monolayer at the time of challenge.
Although bacteria successfully traversed the monolayer, there was no increase in 322
fluorescence in the basolateral chamber beneath the infected cells even after 24
hours. Indeed, there was no significant difference in the level of fluorescence in the 324
basolateral chamber of infected and uninfected monolayers (Fig. 2C).
326
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

15
Infected monolayers were also fixed and examined for the presence of the tight
junction protein ZO-1. The distribution of ZO-1 in infected cells was indistinguishable 328
from uninfected monolayers (Fig. 3A). The protein was present at the margins of all
cells, with no gaps or disruption detected. Additionally, we examined infected 330
monolayers by TEM. At 8 hours following inoculation microcolonies of N.
meningitidis were present on the apical surface of the monolayer (Fig. 3B). These 332
bacteria were tightly associated with the apical surface of the cells of the monolayers,
and were often found on pedestal-like protrusions of the apical membrane (Fig. 3B 334
and C). After 24 hours, the number of adherent bacteria had increased and covered
a larger area of the apical surface of the monolayer (Fig. 3D). Meningococci were 336
still closely adherent to the apical surface (Fig. 3E) and examination of the margins of
cells revealed that the junctional structures were intact (Fig. 3F), consistent with the 338
results obtaining by ZO-1 labelling; even though gaps were evident between cells
below the tight junctions, these were present in infected and uninfected monolayers 340
(Fig. 1 and 3). Taken together these data demonstrate that there is no loss of
integrity of the Calu-3 monolayer coincident with traversal of N. meningitidis, 342
indicating that the meningococcus does not cross the epithelial barrier by a
paracellular route or following loss of cells from the layer. 344
These results suggest that N. meningitidis traverses the respiratory monolayer by a 346
transcellular route, which predicts that the cells of the monolayer should harbour a
population of intracellular bacteria during traversal. Therefore monolayers of cells 348
were challenged with MC58, then after eight or 24 hours were exposed to gentamicin
for one hour to kill extracellular bacteria. Cells were lysed by treatment with saponin 350
to release intracellular bacteria, which were recovered by plating lysates to solid
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

16
media. A population of gentamicin-protected bacteria (representing approximately 352
0.1% of the input) was consistently present in cells at eight hours post-challenge, and
this number increased by an order of magnitude by 24 hours post challenge (Fig. 354
4A). In contrast, no bacteria were recovered from gentamicin-treated membranes
which had been infected with N. meningitidis in the absence of cells (not shown). 356
It was possible that the recovered bacteria survived gentamicin treatment by 358
occupying extracellular sites between cells in the monolayer that were inaccessible to
the antibiotic. Therefore the infected monolayers were disrupted by treatment with 360
trypsin for 10 minutes or latrunculin A for 20 minutes prior to incubation with
gentamicin. Exposure of the monolayer to trypsin for 20 minutes is sufficient to 362
remove all cells from the membrane, while treatment with latrunculin A for 15 minutes
leads to a complete loss of TEER (not shown). Treatment with latrunculin or trypsin 364
did not alter the number of CFU recovered from the layer after exposure to
gentamicin (Fig. 4B), consistent with the bacteria being in an intracellular 366
compartment. In addition, we performed confocal microscopy of cross sections of
the monolayer at 24 hours following bacterial challenge. This revealed the presence 368
of N meningitidis within cells, including bacteria located in basal regions of cells (Fig.
4C). 370
Tfp and the polysaccharide capsule are required for traversal of the monolayer 372
For several pathogens, the mechanisms mediating adhesion to and entry into cells in
monolayers are different for those described with semi-confluent cells (22, 45). As 374
Tfp have been implicated in meningococcal traversal of gastrointestinal epithelial
cells (50), we examined the ability of a mutant lacking Tfp (MC58∆pilE) to cross the 376
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

17
respiratory epithelial cell barrier. The apical surface of Calu-3 monolayers were
challenged with a 1:1 ratio of the wild-type strain and MC58∆pilE, and the ratio of the 378
strains exiting the layer was determined at eight and 24 hours post-challenge. At
these times, inserts containing infected membranes were washed thoroughly in 380
media then transferred into new wells. Aliquots from the basolateral chambers of
new wells were taken one hour later, plated to media with and without antibiotics, and 382
the competitive index (CI) of the mutant calculated. Wells with inserts lacking cells
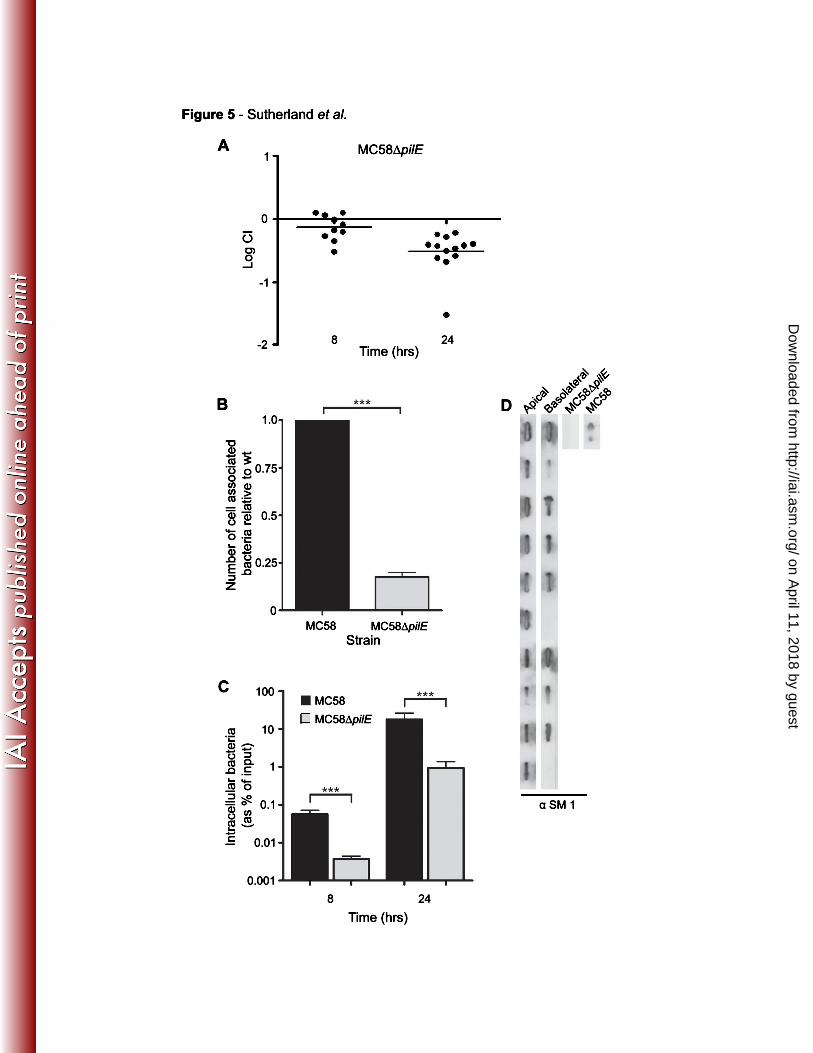
were challenged with the same inoculum as controls. We found that MC58∆pilE had 384
a significant defect for traversal across the Calu-3 monolayer after 24 hours with a
mean logCI of approximately -0.5 (Fig. 5A). To determine how Tfp contribute to 386
traversal, we analysed the adhesion of the pilus-deficient mutant and the wild-type
strain to the polarised respiratory monolayer in separate wells. Consistent with the 388
CI for MC58∆pilE for traversal, the Tfp mutant had a significant defect for adhesion to
the monolayer (p < 0.001, Fig. 5B). Furthermore the adhesion defect of MC58∆pilE 390
was reflected in reduced recovery of intracellular bacteria from MC58∆pilE infected
monolayers compared to MC58 infected cells (Fig. 5C). 392
It has been proposed that Tfp expression is downregulated after initial adhesion to 394
host cells (12). Therefore we examined pilus expression in bacteria following
passage through the monolayer. Slot blot analysis using the mAb SM1 was 396
performed on bacteria recovered from the apical and the basolateral chambers of
wells challenged with MC58. While all 10 colonies from the apical chamber 398
expressed Tfp, eight of 10 colonies were positive for Tfp in the basolateral chamber
(Fig. 5D). Identical findings were obtained when the experiment was performed on a 400
separate occasion.
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

18
402
Recent work has indicated that the sialic acid polysaccharide capsule of N.
meningitidis contributes to survival within epithelial cells (60). We hypothesised that 404
the capsule is also involved in traversal. Therefore we determined the CI for
traversal of MC58∆siaD, which is unable to express a capsule due to loss of SiaD, 406
the polysialic acid transferase (26). The unencapsulated strain had a marked defect
in traversal (Fig. 6A); after eight hours the mean logCI of the MC58∆siaD mutant was 408
-1.2, while by 24 hours the logCI had fallen to almost -4. To understand the basis of
this defect we examined the adhesion of MC58∆siaD to monolayers, consistent with 410
previous work (26), we found that the capsule deficient strain had a significant
increase in adhesion (approximately five fold) compared to MC58 (p < 0.001, Fig. 412
6B). We also recovered intracellular bacteria from MC58 and MC58∆siaD infected
monolayers 24 hours after challenge. In contrast to adhesion, there was a significant 414
reduction in the intracellular population of MC58∆siaD compared with MC58 at this
later time point (p < 0.001, Fig. 6C). Furthermore, as capsule expression is 416
downregulated during adhesion (12), we examined capsule expression in bacteria
from the apical and basolateral chambers at 24 hours post-challenge. All strains were 418
found to be encapsulated by ELISA (Fig. 6D). Both the pilus- and capsule-deficient
strains grew in media and crossed membranes without cells as MC58 (not shown), 420
so their attenuation cannot be ascribed to a growth defect or failure to pass through
the pores in the membranes. 422
An intact microtubule network is required for successful traversal 424
Transcellular traversal of cells by bacteria should be dependent on host as well as
bacterial factors. As microtubules are required for long range trafficking of vesicles, 426
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

19
including those containing pathogens, such as Salmonella enterica serovar
Typhimurium (S. typhimurium) (53) and N. gonorrhoeae (76), we next examined 428
whether disruption of the microtubule network using taxol or nocodazole affected
meningococcal traversal. Taxol inhibits the depolymerisation of tubulin subunits, 430
freezing intact microtubules (30). In contrast nocodazole de-polymerises
microtubules, leaving cells devoid of polymerised tubulin (11). Neither drug affected 432
bacterial replication in vitro (Fig. S4) or permeability of the layer to fluorescent tracer,
although some reduction in TEER was detected (Fig. S5). To confirm the effect of 434
these treatments on cells, monolayers were examined 24 hours after drug treatment
for the presence of microtubules and the tight junction protein occludin (Fig. S6). 436
Abnormalities in the microtubule networks of the cells of the monolayers, including
cells undergoing abnormal cell division, were seen in taxol treated cells (Fig. S6). In 438
contrast, β-tubulin labelling was diffuse throughout nocodazole treated cells,
consistent with microtubule depolymerisation (Fig. S5). The distribution of occludin 440
was unchanged following treatment with taxol or nocodazole, demonstrating (along
with the TEER and tracer data) that the layer remained intact despite drug treatment. 442
Treatment of cells with taxol or nocodazole led to a significant decrease in traversal
of N. meningitidis at eight and 24 hours post-challenge (p values for taxol and 444
nocodazole treated layers, < 0.05 and < 0.01 respectively, Fig. 7A). Interestingly,
treatment of the layers with taxol or nocodazole did not affect the number of 446
intracellular bacteria at eight or 24 hours (Fig. 7B). Taken together, these data
demonstrate that an intact microtubule network is necessary for successful traversal 448
of the layer by N. meningitidis.
450
Infection of Calu 3 monolayers grown under air interface culture (AIC)
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

20
Calu-3 monolayers can be grown under an air interface culture (24). Cells were 452
seeded as described above except the media was removed from the apical chamber
two days after seeding. Monolayers were then left for a further three days. 454
Consistent with previous work (24), the TEER across monolayers with AIC was
approximately half that seen with monlolayers grown under liquid covered culture 456
(LCC, Fig. 8A). TEM examination revealed that the AIC monolayers closely
resemble LCC monolayers (Fig. 8B). Competitive infections comparing traversal of 458
MC58 with pilus-deficient or capsule-deficient mutants with MC58 were performed
using the AIC monolayers. In contrast to LCC monolayers, no traversal had occurred 460
at eight hours post inoculation. However by 24 hours some bacteria had traversed
the monolayer. Both the pilus-deficient and capsule-deficient mutants had significant 462
defects for traversal (Fig. 8C and D). These results indicate that AIC monolayers
could be adapted to investigate N. meningitidis infection, and retain their selective 464
barrier function during meningococcal traversal.
466
The role of CD46 during adhesion of N. meningitidis to polarised respiratory
epithelial monolayers 468
CD46 is the proposed receptor for meningococcal Tfp (33). Therefore we examined
whether CD46 is expressed by Calu-3 cells. Western blot analysis was performed on 470
whole cell lysates cells from Calu-3 cells, as well as COS-7 and HeLa cells, which
are both known to express CD46 (39). We found Calu-3 cells expressed CD46 at 472
higher levels than HeLa or COS-7 cells (Fig. 9A). Next we next analysed the
distribution of CD46 in Calu-3 monolayers. CD46 was localised to the basolateral 474
aspect of cells in the monolayer (Fig. 9B), consistent with findings from other cell
types and tissues (36, 41, 66). Furthermore, challenge of monolayers with N. 476
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

21
meningitidis did not affect the basolateral distribution of CD46 (Fig. 9C), which was
confined to the lateral and basolateral margin of cells. 478
To test whether CD46 is involved in meningococcal adhesion to the monolayer, two
different approaches were used. First, MC58 was incubated with purified CD46 (6.25 480
µg CD46 per 107 bacteria) for 30 minutes prior to infection of Calu-3 monolayers, and
the adherent population compared with bacteria incubated in the absence of the 482
complement regulator. There was no significant difference in the level of adhesion of
CD46 bound bacteria compared to bacteria that were not pre-incubated with CD46 484
(Fig. 9D). Furthermore, the rate of bacterial adhesion was determined with anti-
CD46 mAbs in the infection medium (2 µg of anti- CD46 antibody per 1.6 x 107 CFU). 486
At three hours after challenge, the adherent population of bacteria was compared
with control wells without a mAb or with an isotype control. No significant difference 488
in the adhesion of meningococci to the monolayer was observed in the presence of
the mAbs (Fig.9E). These data are consistent with CD46 playing little role in the 490
adhesion of MC58 to the Calu-3 monolayers.
492
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

22
Discussion 494
The ability of bacteria to cross cellular barriers is critical during colonisation and
disease. These barriers can be formed in the host by a variety of cell types including 496
epithelial and endothelial cells, and at mucosal surfaces they form a key component
of innate immunity against infectious agents. The first cell barrier encountered by 498
bacteria in the nasopharynx is the pseudostratified columnar epithelium of the upper
respiratory tract. Here we characterised the interaction of N. meningitidis, an 500
important human pathogen with monolayers of polarised Calu-3 cells. Although this
cell line is derived from the bronchial epithelium, it is relevant for studying the biology 502
of the meningococcus as the respiratory tract is lined by a columnar epithelium from
the posterior nares down to the distal bronchioles. 504
Semi-confluent Calu-3 cells grown on impermeable substrates exhibited similar 506
patterns of adhesion and uptake of N. meningitidis as Chang cells, which have been
widely used for studying this bacterium. Adhesion of the meningococcus to Calu-3 508
cells was largely dependent on Tfp, consistent with previous work with other cell lines
(47, 50, 69), and invasion occured at low, but appreciable levels. When propagated 510
on semi-permeable membranes, Calu-3 cells polarised and formed monolayers (24,
56), which gave high levels of TEER and were impermeable to fluorescently labelled 512
tracers and non-pathogenic E. coli. Furthermore, the distribution of the tight junction
proteins ZO-1 and occludin, as well as electron microscopy, demonstrated that the 514
cells displayed all the characteristics of pseudostratified columnar epithelial cells
linked by a full range of junctional structures. 516
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

23
There is a distinction between pathogens which utilise a transcellular route to cross 518
epithelial cell monolayers (19), and those bacteria which cross by a paracellular route
following destruction of cells of the layer or disruption of tight junctions (1, 10, 18, 34, 520
35, 40, 55, 59, 68). There is conflicting evidence about the route that the
meningococcus takes to traverse the epithelial cell barrier in the nasopharynx. Work 522
with intestinal epithelial cells indicates that the bacterium migrates using a
transcellular route (50). In contrast, the meningococcus appeared to traverse an 524
endometrial epithelial:endothelial bilayer by both a para- and trans-cellular route (3).
Furthermore, the bacterium crosses the brain microvascular endothelial cells by a 526
paracellular route (8). We found that Calu-3 monolayers continued to display barrier
function during traversal of N. meningitidis. There was a minor reduction in TEER 528
across the monolayers during challenge but this was observed to some extent in both
infected and uninfected monolayers. However levels of TEER never fell to less than 530
40% of the initial readings, similar to levels following challenge with E. coli (which did
not cross the cell barrier), and monolayers remained impermeable to fluorescent 532
tracers with no change in the distribution of ZO-1 following infection. Additionally we
were unable to detect any cell destruction or alteration of junctional structures by 534
TEM. We also found that the monolayers allowed the selective passage of strains
during mixed infection experiments, confirming that traversal does not result from 536
loss of intergrity of the barrier. Finally, an intracellular population of bacteria was
detected in Calu-3 monolayers by recovery of viable organisms following treatment 538
with gentamicin, and by confocal microscopy. Attempts were made to obtain Z stack
images of infected monolayers but were unsuccessful because of auto-fluorescence 540
of membranes and bleaching of the signal when taking multiple images. To
circumvent this, membranes were folded back on themselves to allow acquisition of 542
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

24
high resolution, cross-sectional images of infected monolayers without including the
membrane itself (Fig. 5E); inside:out staining was used to confirm the intracellular 544
location of bacteria. Thus we have several independent lines of evidence that that N.
meningitidis crosses the respiratory epithelial barrier by a transcellular route. 546
Work with polarised epithelial cells in monolayers has provided novel insights into the 548
mechanisms and sites of adhesion of pathogens. For instance, the entry of Shigella
flexneri was shown to be restricted to the basolateral surface of epithelial cell 550
monolayers (45), while a novel adhesin was identified in Salmonella which is
specifically involved in adhesion to polarised cells (22). Here we found that Tfp 552
mediate attachment of the meningococcus to the monolayer, as well as to semi-
confluent cells, demonstrating that the mechanism of initial adhesion is similar to 554
non-polarised and polarised cells.
556
To identify bacterial factors involved in traversal, we performed competitive assays
between the wild-type and mutant strains. The advantage of this approach is that it 558
eliminates well to well variation in the overall level of traversal, and allows direct
comparison of the extent of traversal of strains. Consistent with previous results (50), 560
we found that the mutant lacking Tfp had a significant defect for traversal across the
monolayer compared with the wild-type strain. The defect for adhesion of the pilus 562
deficient strain to polarised monolayers was similar in magnitude to the reduction in
number of intracellular bacteria, and the overall defect of MC58∆pilE for traversal. 564
These data suggest that Tfp contribute to traversal during the attachment of the
bacterium to the apical surface of cells. However a significant proportion of bacteria 566
were pilus-negative following traversal of the monolayer, which is likely to reflect
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

25
antigenic variation from gene conversion with pilS resulting in a pilin-deficient 568
phenotype (9). It might be that Tfp are required for initial adhesion by mediating
bacterial aggregation, but subsequent cell entry requires individual (possibly Tfp-570
negative) bacteria to dissociate from microcolonies on the cells (32).
572
We found that Calu-3 monolayers express the proposed Tfp receptor CD46 (33), 574
which further widens the scope for experimentation using this cell culture model.
However other findings do not support a role of CD46 in meningococcal adhesion 576
(13, 23, 37, 52, 67). Therefore the Calu-3 monolayer offered another approach to
investigate this topic. Although there is evidence that CD46 is expressed on the 578
apical surface of the respiratory epithelium (58), CD46 was confined to the
basolateral surface of Calu-3 cells in the monolayer, similar to findings from 580
examination of the liver (36) and cell culture models (41, 66).. As expected from its
basolateral location, we did not find evidence that CD46 contributes to the adhesion 582
of N. meningitidis as addition of purified CD46 or anti-CD46 mAbs did not inhibit
bacterial attachment, in contrast to previous work (33). This discrepancy could be 584
because we used the four extracellular, complement regulatory, SCR domains of
CD46 rather than the full length molecule, while the mAbs may not be able to 586
compete effectively with interactions with the bacterium. Further studies are
necessary before it is possible to exclude a contribution of CD46 to initial binding of 588
N. meningitidis to Calu-3 cells.
590
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

26
There was a significant reduction in the recovery of capsule deficient (∆siaD) mutant
from within polarised Calu-3 monolayers despite a five fold increase in adhesion of 592
this strain, consistent with previous work indicating that the capsule is dispensable for
cell entry (27, 72). The polysaccharide capsule has been proposed to have an 594
important role in the survival of N. meningitidis in epithelial and endothelial cells (48,
60). We found that the ∆siaD exhibited a dramatically reduced rate of traversal of 596
monolayers, far in excess of its defect for intracellular survival, suggesting that the
capsule has further roles during exit of the bacteria from the basolateral pole of cells. 598
Thus the capsule is virtually indispensible for passage across the respiratory
epithelial barrier (during intracellular survival and escape from the basolateral 600
surface). This emphasises the need to consider the traversal of the epithelial barrier
by the gonococcus (which does not possess a capsule) and N. meningitidis 602
independently, and not assume findings from one pathogen can be extrapolated to
the other. 604
We found that traversal of Calu-3 monolayers by the meningococcus was impaired 606
by interfering with the microtubule network using either taxol or nocodazole. The
only example of apical to basal traversal of monolayers using a paracellular route 608
without disrupting tight junctions is the passage of leucocytes across epithelia.
However, this pathway is not affected by inhibiting the microtubule network with taxol 610
or nocodazole (29). Therefore, the impact of microtubule disruption on the passage
of bacteria provides further evidence for a transcellular route of meningococcal 612
traversal. These data also indicate that traversal by N. meningitidis does not occur
by the same mechanism as transcytosis of polymeric immunoglobulins, which is 614
initiated by receptor-mediated, clathrin-dependent endocytosis (44) and does not
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

27
require intact microtubules (31). We also found that the number of intracellular 616
bacteria at eight and 24 hours was not affected by microtubule disruption, suggesting
that microtubules are involved in basolateral escape of meningococci from cells 618
rather than entry.
620
To date, the intracellular niche occupied by N. meningitidis has not been defined. No
markers of intracellular compartments have been found to consistently co-localise 622
with intracellular meningococci (48, 50, 71), and the bacterium has even been
detected in the cytoplasm by some workers (65). Therefore Calu-3 monolayers could 624
be used to more precisely define the mechanisms and route of transcellular passage
of this important human pathogen. Calu-3 cells can also be grown with under AIC 626
conditions. This is a more lengthy process but it has been reported that the cells can
produce cilia and may produce an even more physiologically relevant model of the 628
nasopharyngeal epithelium (24). AIC monolayers still have barrier function and are
morphologically resemble monolayers grown under LCC. Furthermore, Calu-3 cells 630
could also be used to study the epithelial traversal of other pathogens present in the
respiratory tract including Streptococcus pneumoniae and Haemophilus influenzae. 632
The results should provide valuable information about the pathogenesis of these
pathogens, while defining the factors necessary for migration across the epithelium 634
could lead to the development of vaccines to prevent the diseases they cause.
636
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

28
Acknowledgements 638
We are grateful to Elisabeth Kugelberg for her support and advice, and to Mike
Hollinshead for his help with TEM. Purified CD46 protein was generously provided 640
by Prof. Susan Lea. Calu-3 cells were a kind gift from Dr Clive Robinson (St
George’s Hospital, London), and pEG2 was gratefully received from Dr Myron 642
Christodoulides (University of Southampton). T.C.S. is supported by a Wellcome
Trust PhD studentship, and R.M.E. is a Leverhulme Trust Career Development 644
Fellow. Work in C.M.T.’s laboratory is supported by the Medical Research Council
and the Wellcome Trust. 646
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

29
Figure legends
648
Fig. 1 Morphology of Calu-3 monolayers.
A. Development of the Calu-3 monolayer over time, showing increasing TEER and 650
decreasing permeability to Texas red labelled 70,000 MW dextran. Fluorescent
tracer was added to the apical chamber of monolayers at various times post seeding 652
and levels of fluorescence in the basolateral chamber were measured after six hours.
Error bars show the standard deviation from at least three separate experiments. B. 654
For TEM analysis, cells were grown for five days on cell culture inserts. Lower
magnification view (scale bar = 4 µm) shows cells organised into a monolayer of 656
columnar cells joined by tight junctions (TJ), and desmosomes (D). The cell culture
membrane (CCM) can be seen on the basolateral surface of the cells. Apical-basal 658
differentiation is demonstrated by the basal location of nuclei and the presence of
microvilli (MV) on the apical surface. C. Analysis of the cell boundaries shows the 660
formation of extensive junctional structures including full junctional complexes (JC)
composed of (from apical to basal surface) a tight junction (TJ), an adhesion belt 662
(AB), and a desmosome (D). Scale bar = 1 µm. D. Close apposition of cell
membranes within a tight junction. Scale bar = 1 µm. The integrity of the layer and 664
presence of tight junctions was further confirmed using confocal microscopy to
visualise the tight junction associated proteins, ZO-1 (E) and occludin (F). Scale bars 666
= 5 µm.
668
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

30
Fig. 2 N. meningitidis traverses the Calu-3 monolayer without affecting barrier
function. 670
A. Polarised monolayers were challenged with MC58 at an MOI of 40. The number
of bacteria traversing the layer was established by moving inserts to a fresh well at 672
eight and 24 hours and then sampling one hour later, from the basolateral chamber
of the new well. Results are shown as the percentage of bacteria in the input. Dots 674
represent individual wells from four separate experiments; horizontal bars represent
the mean. B. TEER was monitored in the presence and absence of bacteria for 24 676
hours following inoculation. Despite bacterial passage, TEER did not fall below 40%
of initial levels. C. Texas red labelled 70,000 MW dextran was added to the apical 678
side of cell monolayers which were then challenged with MC58 at an MOI of 40 and
fluorescence in the basolateral chamber was monitored over the next 24 hours. 680
Fluorescence did not increase despite bacterial passage. Diffusion of tracer across
inserts with no cells was included as a control. Error bars show the standard 682
deviation from at least three experiments.
684
Fig. 3 Microscopic analysis of infected monolayers.
A. Monolayers were inoculated with MC58 expressing GFP then fixed 24 hours later. 686
The tight junction protein ZO-1 was labelled and the layers examined by confocal
microscopy. Despite bacterial infection the pattern of ZO-1 labelling was unaffected. 688
Scale bars = 5 µm. B, C. For TEM, monolayers were challenged with MC58 and
fixed eight or 24 hours later. After eight hours the monolayer remained intact and 690
appeared undamaged despite the presence of bacterial colonies on the apical
surface. Bacteria adhered tightly to cells and were often on pedestal like protrusions 692
from the apical membrane. D. After 24 hours, there was no apparent disruption to
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

31
the integrity of the monolayer despite increased numbers of bacteria on the apical 694
surface. No gaps in the layer were observed (E). Adherent bacteria remained tightly
associated with the apical membrane. Enlargement of the region between cells (F) 696
revealed the presence of major junctional structures. Boxed regions in D indicate
sections enlarged in panels E and F. 698
Fig. 4 Identification of a population of intracellular bacteria in infected Calu-3 700
monolayers.
Monolayers of cells were challenged with N. meningitidis at an MOI of 40. A. After 702
eight or 24 hours, cells were treated with gentamicin to kill extracellular bacteria then
lysed with saponin to release intracellular bacteria which were enumerated by plating 704
of lysates. Each spot represents the result from an individual well from three
separate experiments. Horizontal bars represent the arithmetic mean. B. 706
Latrunculin A and trypsin were used to disrupt the monolayer before treatment with
gentamicin. Treatment with these chemicals caused no significant reduction in the 708
gentamicin-protected population of N meningitidis. No bacteria were recovered
following gentamicin treatment in the absence of Calu-3 cells. Error bars show the 710
standard deviation from at least three separate experiments. C. Representative
images showing intracellular bacteria (shown by pink arrowheads, present in the 712
monolayer at 24 hours post inoculation. Monolayers were infected with MC58
expressing GFP at an MOI of 40 and fixed after 24 hours. Extracellular bacteria were 714
labelled using an αL3,7,9 LPS mAb followed by a RRX conjugated secondary
antibody (red). Actin was stained with Alexa 647 phalloidin (blue). Scale bars, 5 µm. 716
Boxed areas are enlarged in the lower panels of i) and ii).
718
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

32
Fig. 5 Tfp are important for traversal of the epithelial cell barrier.
A. Monolayers of Calu-3 cells were challenged with a 1:1 mixture of MC58 and 720
MC58∆pilE on the apical surface and the ratio of the strains exiting the basolateral
surface over one hour was determined eight and 24 hours later. The mutant lacking 722
Tfp had a significant defect for traversal. Results are shown as the log competitive
index (logCI). Each spot represents the result from an individual well from three 724
separate experiments. Horizontal bars represent the arithmetic mean. B.
Monolayers of Calu-3 cells were challenged with either MC58 or MC58∆pilE. After 3 726
hours monolayers were washed and then lysed with 1% saponin to release all cell
associated bacteria. Enumeration of cell associated bacteria by plating revealed a 728
large reduction in the numbers of cell associated bacteria for the pilus deficient strain.
C. Intracellular bacteria were recovered by lysis of cells with saponin after 730
gentamicin treatment. MC58∆pilE is present in reduced numbers within the cells of
the monolayer after eight and 24 hours compared with MC58. No bacteria were 732
recovered following gentamicin treatment in the absence of Calu-3 cells (not shown).
Significant differences are indicated by *, < 0.05, **, < 0.01, ***, < 0.001 (Unpaired T-734
test). D. Slot blot analysis of pilin expression in 10 randomly selected colonies from
the apical and basolateral chambers of wells after a 24 hour infection with MC58. 736
Eight of 10 colonies expressed pilus in the basolateral chamber compared to 10 of 10
in the apical chamber. 738
Fig. 6 The polysaccharide capsule is crucial to the traversal of N. meningitidis. 740
A. The apical surfaces of Calu-3 cells were infected with a 1:1 mixture of MC58 and
MC58∆siaD, and the ratio of the strains exiting the basal surface over one hour at 742
eight and 24 hours was determined. The capsule deficient strain had a significant
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

33
defect for traversal. Data are shown as the log competitive index (logCI). Each spot 744
represents the result from an individual well from three separate experiments.
Horizontal bars represent the arithmetic mean. B. Monolayers of Calu-3 cells were 746
challenged with either MC58 or MC58∆siaD. After three hours monolayers were
washed and then lysed with 1% saponin to release cell associated bacteria. 748
Enumeration of cell associated bacteria by plating revealed a large increase in the
numbers of cell associated MC58∆siaD compared with the wild-type strain. C. 750
Intracellular bacteria were recovered by lysis of cells with saponin after gentamicin
treatment. Despite its increased adhesion, MC58∆siaD is present in reduced 752
numbers within the cells of the monolayer after 24 hours compared with MC58. No
bacteria were recovered following gentamicin treatment in the absence of Calu-3 754
cells (not shown). Significant differences are indicated by *, < 0.05, **, < 0.01, ***, <
0.001 (Unpaired T-test). D. Monolayers were infected with MC58 as described, and 756
bacteria were recovered from the apical and basolateral chambers 24 hours later.
Individual colonies (10 from each chamber) were grown overnight on solid media, 758
and analysed for capsule expression by ELISA using an anti-serogroup B mAb at a
variety of dilutions. The unencapsulated strain MC58∆siaD and the wild-type strain 760
were included as controls. All strains sampled from apical and basolateral chambers
were encapsulated. Spots represent the A492 of each strain. 762
Fig. 7 Disruption of microtubules within cells of the monolayer reduces 764
traversal of N. meningitidis.
A. Calu-3 monolayers were treated with nocodazole or taxol, then challenged with 766
MC58 at an MOI of 40. The number of bacteria in the basolateral chamber was
established by plating at eight and 24 hours post challenge. Both nocodazole and 768
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

34
taxol caused a significant reduction in the number of bacteria in the basolateral
chamber at both time points when compared to untreated layers. B. Monolayers of 770
Calu-3 cells were treated with nocodazole or taxol, then challenged. Intracellular
bacteria were recovered by lysis of cells with 1% saponin after exposure to 772
gentamicin for 30 minutes. There was no significant difference in the number of
intracellular bacteria at eight or 14 hours for either drug treatment compared to 774
untreated layers. No bacteria were recovered following gentamicin treatment in the
absence of Calu-3 cells (not shown). Error bars show the standard deviation from at 776
least three separate experiments. Significant differences are indicated by *, < 0.05,
**, < 0.01, ***, < 0.001 (Unpaired T-test). 778
Fig. 8 Challenge of monolayers grown with an air interface (AI) 780
A. Monolayers were seeded as for liquid covered monolayers but media was
removed from the apical chamber after 2 days. After 5 days levels of TEER in the AI 782
monolayers was approximately half that seen for layers grown in media. Significant
differences are indicated by *, < 0.05, **, < 0.01, ***, < 0.001 (Unpaired T-test). B. 784
TEM of AIC monolayers at 5 days. The gross morphology of the layer is not
significantly different from that seen in monolayers grown under standard conditions. 786
Lower magnification view (scale bar, 5 µm) shows cells organised into a monolayer
of columnar cells joined by tight junctions (TJ), and desmosomes (D). The cell 788
culture membrane (CCM) can be seen on the basolateral surface of the cells. Boxed
sections are enlarged in panels i) and ii) (scale bar, 1 µm). C. Competitive index of 790
MC58∆pilE across AI monolayers. In contrast to monolayers grown under media,
there was no traversal of N. meningitidis at 8 hours. However by 24 hours some 792
bacteria had traversed the layer. The pilus deficient mutant has a defect for traversal
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

35
compared to MC58. D. CI of MC58∆siaD for traversal across AI monolayers. The 794
capsule deficient mutant had a large defect for traversal. Results are shown as the
log competitive index (logCI). Each spot represents data from an individual well from 796
three separate experiments. Horizontal bars represent the arithmetic mean.
798
Fig. 9 The role of CD46 during binding of N. meningitidis to polarised
respiratory epithelial monolayers 800
A. CD46 expression detected by Western blot analysis of lysates of cell lines
(indicated). Full length CD46 migrates with a predicted molecular mass of between 802
50 and 79 kDa, while the purified SCR domains are detected as a band of 40 kDa.
B. Expression of CD46 in polarised monolayers examined by confocal microscopy. 804
Five day old monolayers immunolabelled for CD46 detected with a FITC-conjugated,
secondary antibody (green). Actin was counterstained with phalloidin conjugated to 806
an Alexa 595 fluorophore (red). C. Monolayers of Calu-3 cells were infected with
MC58 expressing eGFP (green) at an MOI of 40 for 8 hours, and immunolabelled 808
CD46 was detected with a RRX (red) conjugated secondary antibody; actin was
stained using phalloidin conjugated to Alexa 647 (blue). Representative images are 810
shown. Scale bars = 5 µm. D. Monolayers of Calu-3 cells were challenged with
MC58 at an MOI of 40 with or without 25 µg of purified CD46 per well (1 x 105 cells). 812
MC58 and purified CD46 were pre-incubated for 30 minutes prior to challenge.
Monolayers were also infected with MC58 pre-incubated with DMEM:F12 as a 814
control. E. Calu-3 cells were infected with MC58 at an MOI of 40 in presence or
absence of anti-human CD46 mAbs (indicated) at 2 µg/ml. Adhesion of N. 816
meningitidis to cells was calculated by recovering bacteria 3 hours later; error bars,
S.D. of experiments performed on three occasions. 818
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

36
Table 1. – Strains used in this study
Species Strain Description Antibiotic resistance
Origin/references
N. meningitidis
MC58 Serogroup B clinical isolate, ET-
5
- Isolated in the UK in 1989
MC58 eGFP
MC58 carrying pEG2
Ery (6)
MC58∆pilE Transposon inserted in pilE. Tfp
deficient
Kan (15)
MC58∆siaD Deletion mutagenesis of
sialyl transferase. capsule deficient
Ery (74)
E. coli DH5α Invitrogen
820
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

37
Table 2. – Antibodies used in this study.
Name Target Origin Working Dilution
Source/Reference
αL3,7,9 LPS 3,7,9 (4047)
Mouse monoclonal
4047
1/200 NIBSC
αZO-1 ZO-1 Rabbit polyclonal
1/30 Zymed
αOcc Occludin Rabbit polyclonal
1/30 Zymed
αβ-tub E7
β -Tubulin Mouse polyclonal
1/100 Developmental Studies Hybridoma
Bank, Iowa αSM1 PilE Mouse
monoclonal 1/1000 (70)
αmenB 95/750
N. meningitidis serogroup B
capsule
Mouse monoclonal
- NIBSC
αM HRP Mouse immunoglobulins
Goat polyclonal 1/1000 Dako
αβ-tub E7
β-Tubulin Mouse polyclonal
1/100 Developmental Studies Hybridoma
Bank, Iowa αM Cy2 Mouse IgG Donkey
polyclonal 1/200 Jackson
αM RRX Mouse IgG Donkey polyclonal
1/200 Jackson
αR Cy5 Rabbit IgG Donkey polyclonal
1/200 Jackson
122-2 CD46 Mouse
monoclonal 1/50 Santa Cruz
Biotechnology inc.
1.BB.442 CD46 Mouse
monoclonal 1/200 Santa Cruz
Biotechnology inc. E4.3
CD46
Mouse monoclonal
2µg/ml
Pharmingen
J4.48
CD46
Mouse monoclonal
2µg/ml
Santa Cruz Biotechnology inc.
Isotype control
IgG
Mouse monoclonal
2µg/ml
Dako
Phalloidin Alexa 595
Actin 1/100 Invitrogen
Phalloidin Alexa 647
Actin
1/100 Invitrogen
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

38
References 822
1. Attali, C., C. Durmort, T. Vernet, and A. M. Di Guilmi. 2008. The interaction 824
of Streptococcus pneumoniae with plasmin mediates transmigration across endothelial and epithelial monolayers by intercellular junction cleavage. Infect Immun 826
76:5350-6. 2. Beuzon, C. R., and D. W. Holden. 2001. Use of mixed infections with 828
Salmonella strains to study virulence genes and their interactions in vivo. Microbes Infect 3:1345-52. 830
3. Birkness, K. A., B. L. Swisher, E. H. White, E. G. Long, E. P. Ewing, Jr., and F. D. Quinn. 1995. A tissue culture bilayer model to study the passage of 832
Neisseria meningitidis. Infect Immun 63:402-9. 4. Capecchi, B., J. Adu-Bobie, F. Di Marcello, L. Ciucchi, V. Masignani, A. 834
Taddei, R. Rappuoli, M. Pizza, and B. Arico. 2005. Neisseria meningitidis NadA is a new invasin which promotes bacterial adhesion to and penetration into human 836
epithelial cells. Mol Microbiol 55:687-98. 5. Cavet, M. E., M. West, and N. L. Simmons. 1997. Transepithelial transport of 838
the fluoroquinolone ciprofloxacin by human airway epithelial Calu-3 cells. Antimicrob Agents Chemother 41:2693-8. 840
6. Christodoulides, M., J. S. Everson, B. L. Liu, P. R. Lambden, P. J. Watt, E. J. Thomas, and J. E. Heckels. 2000. Interaction of primary human endometrial cells 842
with Neisseria gonorrhoeae expressing green fluorescent protein. Mol Microbiol 35:32-43. 844
7. Cooney, D., M. Kazantseva, and A. J. Hickey. 2004. Development of a size-dependent aerosol deposition model utilising human airway epithelial cells for 846
evaluating aerosol drug delivery. Altern Lab Anim 32:581-90. 8. Coureuil, M., G. Mikaty, F. Miller, H. Lecuyer, C. Bernard, S. Bourdoulous, 848
G. Dumenil, R. M. Mege, B. B. Weksler, I. A. Romero, P. O. Couraud, and X. Nassif. 2009. Meningococcal type IV pili recruit the polarity complex to cross the 850
brain endothelium. Science 325:83-7. 9. Criss, A. K., K. A. Kline, and H. S. Seifert. 2005. The frequency and rate of 852
pilin antigenic variation in Neisseria gonorrhoeae. Mol Microbiol 58:510-9. 10. Cywes, C., and M. R. Wessels. 2001. Group A Streptococcus tissue invasion 854
by CD44-mediated cell signalling. Nature 414:648-52. 11. De Brabander, M. J., R. M. Van de Veire, F. E. Aerts, M. Borgers, and P. A. 856
Janssen. 1976. The effects of methyl (5-(2-thienylcarbonyl)-1H-benzimidazol-2-yl) carbamate, (R 17934; NSC 238159), a new synthetic antitumoral drug interfering with 858
microtubules, on mammalian cells cultured in vitro. Cancer Res 36:905-16. 12. Deghmane, A. E., D. Giorgini, M. Larribe, J. M. Alonso, and M. K. Taha. 860
2002. Down-regulation of pili and capsule of Neisseria meningitidis upon contact with epithelial cells is mediated by CrgA regulatory protein. Mol Microbiol 43:1555-64. 862
13. Edwards, J. L., E. J. Brown, S. Uk-Nham, J. G. Cannon, M. S. Blake, and M. A. Apicella. 2002. A co-operative interaction between Neisseria gonorrhoeae and 864
complement receptor 3 mediates infection of primary cervical epithelial cells. Cell Microbiol 4:571-84. 866
14. Ehrhardt, C., J. Fiegel, S. Fuchs, R. Abu-Dahab, U. F. Schaefer, J. Hanes, and C. M. Lehr. 2002. Drug absorption by the respiratory mucosa: cell culture 868
models and particulate drug carriers. J Aerosol Med 15:131-9.
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

39
15. Exley, R. M., L. Goodwin, E. Mowe, J. Shaw, H. Smith, R. C. Read, and C. 870
M. Tang. 2005. Neisseria meningitidis lactate permease is required for nasopharyngeal colonization. Infect Immun 73:5762-6. 872
16. Exley, R. M., R. Sim, L. Goodwin, M. Winterbotham, M. C. Schneider, R. C. Read, and C. M. Tang. 2009. Identification of meningococcal genes necessary for 874
colonization of human upper airway tissue. Infect Immun 77:45-51. 17. Fiegel, J., C. Ehrhardt, U. F. Schaefer, C. M. Lehr, and J. Hanes. 2003. 876
Large porous particle impingement on lung epithelial cell monolayers--toward improved particle characterization in the lung. Pharm Res 20:788-96. 878
18. Figueiredo, C. C., P. M. Deccache, L. M. Lopes-Bezerra, and V. Morandi. 2007. TGF-beta1 induces transendothelial migration of the pathogenic fungus 880
Sporothrix schenckii by a paracellular route involving extracellular matrix proteins. Microbiology 153:2910-21. 882
19. Finlay, B. B., B. Gumbiner, and S. Falkow. 1988. Penetration of Salmonella through a polarized Madin-Darby canine kidney epithelial cell monolayer. J Cell Biol 884
107:221-30. 20. Florea, B. I., M. L. Cassara, H. E. Junginger, and G. Borchard. 2003. Drug 886
transport and metabolism characteristics of the human airway epithelial cell line Calu-3. J Control Release 87:131-8. 888
21. Foster, K. A., M. L. Avery, M. Yazdanian, and K. L. Audus. 2000. Characterization of the Calu-3 cell line as a tool to screen pulmonary drug delivery. 890
Int J Pharm 208:1-11. 22. Gerlach, R. G., N. Claudio, M. Rohde, D. Jackel, C. Wagner, and M. 892
Hensel. 2008. Cooperation of Salmonella pathogenicity islands 1 and 4 is required to breach epithelial barriers. Cell Microbiol 10:2364-76. 894
23. Gill, D. B., M. Koomey, J. G. Cannon, and J. P. Atkinson. 2003. Down-regulation of CD46 by piliated Neisseria gonorrhoeae. J Exp Med 198:1313-22. 896
24. Grainger, C. I., L. L. Greenwell, D. J. Lockley, G. P. Martin, and B. Forbes. 2006. Culture of Calu-3 cells at the air interface provides a representative model of 898
the airway epithelial barrier. Pharm Res 23:1482-90. 25. Greenfield, S., P. R. Sheehe, and H. A. Feldman. 1971. Meningococcal 900
carriage in a population of "normal" families. J Infect Dis 123:67-73. 26. Hammerschmidt, S., R. Hilse, J. P. van Putten, R. Gerardy-Schahn, A. 902
Unkmeir, and M. Frosch. 1996. Modulation of cell surface sialic acid expression in Neisseria meningitidis via a transposable genetic element. Embo J 15:192-8. 904
27. Hammerschmidt, S., A. Muller, H. Sillmann, M. Muhlenhoff, R. Borrow, A. Fox, J. van Putten, W. D. Zollinger, R. Gerardy-Schahn, and M. Frosch. 1996. 906
Capsule phase variation in Neisseria meningitidis serogroup B by slipped-strand mispairing in the polysialyltransferase gene (siaD): correlation with bacterial invasion 908
and the outbreak of meningococcal disease. Mol Microbiol 20:1211-20. 28. Helaine, S., E. Carbonnelle, L. Prouvensier, J. L. Beretti, X. Nassif, and V. 910
Pelicic. 2005. PilX, a pilus-associated protein essential for bacterial aggregation, is a key to pilus-facilitated attachment of Neisseria meningitidis to human cells. Mol 912
Microbiol 55:65-77. 29. Hofman, P., L. D'Andrea, D. Carnes, S. P. Colgan, and J. L. Madara. 1996. 914
Intestinal epithelial cytoskeleton selectively constrains lumen-to-tissue migration of neutrophils. Am J Physiol 271:C312-20. 916
30. Horwitz, S. B. 1992. Mechanism of action of taxol. Trends Pharmacol Sci 13:134-6. 918
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

40
31. Hunziker, W., P. Male, and I. Mellman. 1990. Differential microtubule requirements for transcytosis in MDCK cells. Embo J 9:3515-25. 920
32. Ilver, D., H. Kallstrom, S. Normark, and A. B. Jonsson. 1998. Transcellular passage of Neisseria gonorrhoeae involves pilus phase variation. Infect Immun 922
66:469-73. 33. Kallstrom, H., M. K. Liszewski, J. P. Atkinson, and A. B. Jonsson. 1997. 924
Membrane cofactor protein (MCP or CD46) is a cellular pilus receptor for pathogenic Neisseria. Mol Microbiol 25:639-47. 926
34. Katz, J., V. Sambandam, J. H. Wu, S. M. Michalek, and D. F. Balkovetz. 2000. Characterization of Porphyromonas gingivalis-induced degradation of epithelial 928
cell junctional complexes. Infect Immun 68:1441-9. 35. Katz, J., Q. B. Yang, P. Zhang, J. Potempa, J. Travis, S. M. Michalek, and 930
D. F. Balkovetz. 2002. Hydrolysis of epithelial junctional proteins by Porphyromonas gingivalis gingipains. Infect Immun 70:2512-8. 932
36. Kinugasa, N., T. Higashi, K. Nouso, H. Nakatsukasa, Y. Kobayashi, M. Ishizaki, N. Toshikuni, K. Yoshida, S. Uematsu, and T. Tsuji. 1999. Expression of 934
membrane cofactor protein (MCP, CD46) in human liver diseases. Br J Cancer 80:1820-5. 936
37. Kirchner, M., D. Heuer, and T. F. Meyer. 2005. CD46-independent binding of neisserial type IV pili and the major pilus adhesin, PilC, to human epithelial cells. 938
Infect Immun 73:3072-82. 38. Li, Y., W. Wang, W. Parker, and J. P. Clancy. 2006. Adenosine regulation of 940
cystic fibrosis transmembrane conductance regulator through prostenoids in airway epithelia. Am J Respir Cell Mol Biol 34:600-8. 942
39. Liszewski, M. K., T. W. Post, and J. P. Atkinson. 1991. Membrane cofactor protein (MCP or CD46): newest member of the regulators of complement activation 944
gene cluster. Annu Rev Immunol 9:431-55. 40. Lytton, S. D., W. Fischer, W. Nagel, R. Haas, and F. X. Beck. 2005. 946
Production of ammonium by Helicobacter pylori mediates occludin processing and disruption of tight junctions in Caco-2 cells. Microbiology 151:3267-76. 948
41. Maisner, A., M. K. Liszewski, J. P. Atkinson, R. Schwartz-Albiez, and G. Herrler. 1996. Two different cytoplasmic tails direct isoforms of the membrane 950
cofactor protein (CD46) to the basolateral surface of Madin-Darby canine kidney cells. J Biol Chem 271:18853-8. 952
42. Merz, A. J., D. B. Rifenbery, C. G. Arvidson, and M. So. 1996. Traversal of a polarized epithelium by pathogenic Neisseriae: facilitation by type IV pili and 954
maintenance of epithelial barrier function. Mol Med 2:745-54. 43. Merz, A. J., M. So, and M. P. Sheetz. 2000. Pilus retraction powers bacterial 956
twitching motility. Nature 407:98-102. 44. Mostov, K. E., and D. L. Deitcher. 1986. Polymeric immunoglobulin receptor 958
expressed in MDCK cells transcytoses IgA. Cell 46:613-21. 45. Mounier, J., T. Vasselon, R. Hellio, M. Lesourd, and P. J. Sansonetti. 960
1992. Shigella flexneri enters human colonic Caco-2 epithelial cells through the basolateral pole. Infect Immun 60:237-48. 962
46. Nassif, X., J. L. Beretti, J. Lowy, P. Stenberg, P. O'Gaora, J. Pfeifer, S. Normark, and M. So. 1994. Roles of pilin and PilC in adhesion of Neisseria 964
meningitidis to human epithelial and endothelial cells. Proc Natl Acad Sci U S A 91:3769-73. 966
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

41
47. Nassif, X., C. Pujol, P. Morand, and E. Eugene. 1999. Interactions of pathogenic Neisseria with host cells. Is it possible to assemble the puzzle? Mol 968
Microbiol 32:1124-32. 48. Nikulin, J., U. Panzner, M. Frosch, and A. Schubert-Unkmeir. 2006. 970
Intracellular survival and replication of Neisseria meningitidis in human brain microvascular endothelial cells. Int J Med Microbiol 27:27. 972
49. Pezron, I., R. Mitra, D. Pal, and A. K. Mitra. 2002. Insulin aggregation and asymmetric transport across human bronchial epithelial cell monolayers (Calu-3). J 974
Pharm Sci 91:1135-46. 50. Pujol, C., E. Eugene, L. de Saint Martin, and X. Nassif. 1997. Interaction of 976
Neisseria meningitidis with a polarized monolayer of epithelial cells. Infect Immun 65:4836-42. 978
51. Pujol, C., E. Eugene, M. Marceau, and X. Nassif. 1999. The meningococcal PilT protein is required for induction of intimate attachment to epithelial cells following 980
pilus-mediated adhesion. Proc Natl Acad Sci U S A 96:4017-22. 52. Rahman, M., H. Kallstrom, S. Normark, and A. B. Jonsson. 1997. PilC of 982
pathogenic Neisseria is associated with the bacterial cell surface. Mol Microbiol 25:11-25. 984
53. Ramsden, A. E., D. W. Holden, and L. J. Mota. 2007. Membrane dynamics and spatial distribution of Salmonella-containing vacuoles. Trends Microbiol 15:516-986
24. 54. Reznik, G. K. 1990. Comparative anatomy, physiology, and function of the 988
upper respiratory tract. Environ Health Perspect 85:171-6. 55. Sajjan, U., Q. Wang, Y. Zhao, D. C. Gruenert, and M. B. Hershenson. 2008. 990
Rhinovirus Disrupts the Barrier Function of Polarized Airway Epithelial Cells. Am J Respir Crit Care Med 11:11. 992
56. Shen, B. Q., W. E. Finkbeiner, J. J. Wine, R. J. Mrsny, and J. H. Widdicombe. 1994. Calu-3: a human airway epithelial cell line that shows cAMP-994
dependent Cl- secretion. Am J Physiol 266:L493-501. 57. Sim, R. J., M. M. Harrison, E. R. Moxon, and C. M. Tang. 2000. 996
Underestimation of meningococci in tonsillar tissue by nasopharyngeal swabbing. Lancet 356:1653-4. 998
58. Sinn, P. L., G. Williams, S. Vongpunsawad, R. Cattaneo, and P. B. McCray, Jr. 2002. Measles virus preferentially transduces the basolateral surface of 1000
well-differentiated human airway epithelia. J Virol 76:2403-9. 59. Soriani, M., I. Santi, A. Taddei, R. Rappuoli, G. Grandi, and J. L. Telford. 1002
2006. Group B Streptococcus crosses human epithelial cells by a paracellular route. J Infect Dis 193:241-50. 1004
60. Spinosa, M. R., C. Progida, A. Tala, L. Cogli, P. Alifano, and C. Bucci. 2007. The Neisseria meningitidis capsule is important for intracellular survival in 1006
human cells. Infect Immun 75:3594-603. 61. Stephens, D. S. 2007. Conquering the meningococcus. FEMS Microbiol Rev 1008
31:3-14. 62. Stephens, D. S. 1999. Uncloaking the meningococcus: dynamics of carriage 1010
and disease. Lancet 353:941-2. 63. Stephens, D. S., L. H. Hoffman, and Z. A. McGee. 1983. Interaction of 1012
Neisseria meningitidis with human nasopharyngeal mucosa: attachment and entry into columnar epithelial cells. J Infect Dis 148:369-76. 1014
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

42
64. Stephens, D. S., and Z. A. McGee. 1981. Attachment of Neisseria meningitidis to human mucosal surfaces: influence of pili and type of receptor cell. J 1016
Infect Dis 143:525-32. 65. Tala, A., C. Progida, M. De Stefano, L. Cogli, M. R. Spinosa, C. Bucci, and 1018
P. Alifano. 2008. The HrpB-HrpA two-partner secretion system is essential for intracellular survival of Neisseria meningitidis. Cell Microbiol 25:25. 1020
66. Teuchert, M., A. Maisner, and G. Herrler. 1999. Importance of the carboxyl-terminal FTSL motif of membrane cofactor protein for basolateral sorting and 1022
endocytosis. Positive and negative modulation by signals inside and outside the cytoplasmic tail. J Biol Chem 274:19979-84. 1024
67. Tobiason, D. M., and H. S. Seifert. 2001. Inverse relationship between pilus-mediated gonococcal adherence and surface expression of the pilus receptor, CD46. 1026
Microbiology 147:2333-40. 68. Vermeer, P. D., J. McHugh, T. Rokhlina, D. W. Vermeer, J. Zabner, and M. 1028
J. Welsh. 2007. Vaccinia virus entry, exit, and interaction with differentiated human airway epithelia. J Virol 81:9891-9. 1030
69. Virji, M., C. Alexandrescu, D. J. Ferguson, J. R. Saunders, and E. R. Moxon. 1992. Variations in the expression of pili: the effect on adherence of 1032
Neisseria meningitidis to human epithelial and endothelial cells. Mol Microbiol 6:1271-9. 1034
70. Virji, M., J. E. Heckels, W. J. Potts, C. A. Hart, and J. R. Saunders. 1989. Identification of epitopes recognized by monoclonal antibodies SM1 and SM2 which 1036
react with all pili of Neisseria gonorrhoeae but which differentiate between two structural classes of pili expressed by Neisseria meningitidis and the distribution of 1038
their encoding sequences in the genomes of Neisseria spp. J Gen Microbiol 135:3239-51. 1040
71. Virji, M., H. Kayhty, D. J. Ferguson, C. Alexandrescu, J. E. Heckels, and E. R. Moxon. 1991. The role of pili in the interactions of pathogenic Neisseria with 1042
cultured human endothelial cells. Mol Microbiol 5:1831-41. 72. Virji, M., K. Makepeace, D. J. Ferguson, M. Achtman, and E. R. Moxon. 1044
1993. Meningococcal Opa and Opc proteins: their role in colonization and invasion of human epithelial and endothelial cells. Mol Microbiol 10:499-510. 1046
73. Virji, M., K. Makepeace, I. Peak, G. Payne, J. R. Saunders, D. J. Ferguson, and E. R. Moxon. 1995. Functional implications of the expression of PilC proteins in 1048
meningococci. Mol Microbiol 16:1087-97. 74. Virji, M., K. Makepeace, I. R. Peak, D. J. Ferguson, M. P. Jennings, and E. 1050
R. Moxon. 1995. Opc- and pilus-dependent interactions of meningococci with human endothelial cells: molecular mechanisms and modulation by surface polysaccharides. 1052
Mol Microbiol 18:741-54. 75. Virji, M., S. M. Watt, S. Barker, K. Makepeace, and R. Doyonnas. 1996. The 1054
N-domain of the human CD66a adhesion molecule is a target for Opa proteins of Neisseria meningitidis and Neisseria gonorrhoeae. Mol Microbiol 22:929-39. 1056
76. Wang, J. A., T. F. Meyer, and T. Rudel. 2007. Cytoskeleton and motor proteins are required for the transcytosis of Neisseria gonorrhoeae through polarized 1058
epithelial cells. Int J Med Microbiol 31:31. 77. Wolfgang, M., P. Lauer, H. S. Park, L. Brossay, J. Hebert, and M. Koomey. 1060
1998. PilT mutations lead to simultaneous defects in competence for natural transformation and twitching motility in piliated Neisseria gonorrhoeae. Mol Microbiol 1062
29:321-30.
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from