the infrastructure of the tegument o f moniliformis dubius...
TRANSCRIPT

137
The infrastructure of the tegument of Moniliformis dubius(Acanthocephala)
By W. L. NICHOLAS and E. H. MERCER
(From the Department of Zoology and the John Curtin School of Medical Research,Australian National University, Canberra, Australia)
With s plates
SummaryThe ultrastructure of the body wall of Moniliformis dubius has been studied in thelight and electron microscope. It consists of an apparently syncytial tegument, over-laid by a tenuous cuticle in the form of a finely fibrous extracellular fringe and is backedby a basement membrane and fibrous connective tissue. The tegument contains aframework of fibres, which, distally, is connected to a dense fibrous meshworkseparated from the cuticle by two membranes. Within the syncytial tegument arefound the usual cytoplasmic organelles: mitochondria (often degenerate in structure),Golgi clusters, small amounts of other smooth membranes, and numerous denseparticles (glycogen and perhaps ribosomes). Many mitochondria contain denseparticles. Evidence of vacuole formation at the surface of the tegument suggests thatpinocytosis plays a part in assimilation.
IntroductionT H E ultrastructure of the acanthocephalan body wall has not been describedin detail. Rothman (1961) published a brief note on the appearance of thebody wall of Macranthorhynchus hirudinacens under the electron microscopeand Nicholas and Hynes (1963) included an electron micrograph of a sectionthrough the body wall of Moniliformis dubius in a review of the group. Someobservations have also been made on the acanthor with the electron micro-scope by Burnham (1957).
Baer (1961) has described the acanthocephalan body wall as it appearsunder the light microscope. According to him it consists of a cuticle of twolayers, overlying a complex hypodermis, also divisible into two layers. Theoutermost layer of the cuticle is thin and apparently structureless. Beneaththis, the second layer, also thin, has fine striations perpendicular to the surface.The outer part of the hypodermis contains a complex system of fine, inter-woven fibres, which may appear felt-like, or as layers of fibres parallel to thesurface, alternating with felt-like layers. The inner part of the hypodermiscontains fibres orientated perpendicularly to the surface, its elements insertedinto the felt layer, distally, and into a basal membrane, proximally.
A system of fluid-filled canals lies within the deeper parts of the hypo-dermis. The whole of the hypodermis, like many of the other body tissues,appears syncytial. The body wall contains few large nuclei in relatively fixedpositions. In some acanthocephala, including M. dubius, these become very[Quart. J. micr. Sci., Vol. 106, pt. 2, pp. 137-146, 1965.]

138 Nicholas and Mercer—Tegument of Moniliformis
large and ramified; in others the nuclei of the body wall fragment duringlarval development.
Von Brand (1939) included some histochemical observations on the dis-tribution, of lipid and glycogei. in a study of the chemical composition ofMacranthorhynchus hirudinaceus, and Bullock (1949) made further observa-tions on both in a number of other species. Crompton (1963) made a com-prehensive histochemical study of Polymorphic minutus and, in addition,identified a thin layer of mucopolysaccharide outside the tegument, which hetermed the epicuticle.
The distinctions drawn in the past between cuticle and epicuticle on theone hand, with the implication that these are successive layers of a secretednon-living structure, and the hypodermis, on the other, implying a livingcellular structure, are misleading. As we shall show, both the cuticle and thehypodermis recognized by Baer (1961) are cytoplasmic. We shall use theterm tegument for both. Rothman (1959) and Threadgold (1963) in theirwork on platyhelminthes also adopted the term tegument, for analogousreasons. It would then seem logical to rename the epicuticle of Crompton(1963) as cuticle, although again, as we shall show, it hardly constitutes atough outer covering as is sometimes implied by the term.
M. dubius lives, as an adult, in the intestine of the rat, and in the larvalstages, in the haemocoel of cockroaches and other insects. Its life cycle hasbeen described by Burlingame and Chandler (1941) and Moore (1946).
Material and methodsMoniliformis dubius was maintained in the laboratory with rats as the de-
finitive host and cockroaches, Periplaneta americana, as the intermediate host,following the technique described by Burlingame and Chandler (1941). Weselected for our study immature worms, taken from rats which had beeninfected for two to three weeks, and sexually mature worms of both sexesfrom infections of more than six weeks standing. The worms were transferredfrom the intestine of the rat to Ringer's solution, rapidly rinsed, and cut intosmall pieces. Small segments of the body wall were then rapidly transferredto fixative.
Most of the specimens were fixed in 1% osmium tetroxide (Palade, 1952),dehydrated in a series of ethanol solutions of increasing strength and embeddedin Araldite. Sections, cut with diamond or glass knives, were stained witheither 3-5% aqueous uranyl acetate or lead citrate (Karnovsky, 1961) beforeexamination with a Siemens Elmiskop I electron microscope. Some specimenswere fixed in 1% potassium permanganate (Luft, 1956) followed by stainingwith uranyl acetate, and others, fixed in osmium tetroxide, were treated with1 % phosphotungstic acid in absolute alcohol before embedding.
Glutaraldehyde was used to fix tissues prior to treatment with enzymes(Sabatini, Bensch and Barrnett, 1963). A 3% solution in o-i M sodiumphosphate buffer of pH 7-3, containing a trace of CaCl2, was employed.
So that the structures seen under the electron microscope could be corre-

Nicholas and Mercer—Tegument of Moniliformis 139
lated with gross morphology, thicker sections (about 2-5 /x) were cut from thesame blocks and examined in the phase-contrast microscope. These in turnwere compared with conventional sections prepared from paraffin-embeddedtissue, stained in haematoxylin and eosin.
To identify glycogen deposits and ribosomes small pieces of the body wallfrom immature female worms were subjected to enzymic hydrolysis and thencompared with water-treated controls by light and electron microscopy. Forglycogen, tissues fixed in glutaraldehyde, washed in water, and digested inhuman saliva (7 h at room temperature) were transferred to the osmium fixa-tive and prepared for electron microscopy. For light microscopy, paraffinsections, prepared from Carnoy-fixed tissue, were treated with saliva andstained by the PAS reaction (Pearse, i960).
Ribonuclease (Sigma) digestion in water (nine hours) at 370 C was used forribosomes (Pearse, i960). Tissue fixed in glutaraldehyde before digestionand transferred to the osmium fixative afterwards was prepared for electronmicroscopy. For the light microscope, frozen sections of unembedded tissuewere examined in acridine orange by fluorescent microscopy (Bertalanffy,Masin, and Masin, 1958). Diminution in orange fluorescence followingdigestion of the sections was taken as an indication of RNA. For lipids, frozensections were cut from tissue fixed in formaldehyde-calcium (Baker, 1944),embedded in gelatine, and stained with Sudan black.
ResultsThe tegument of Moniliformis dubius, as seen in paraffin sections stained
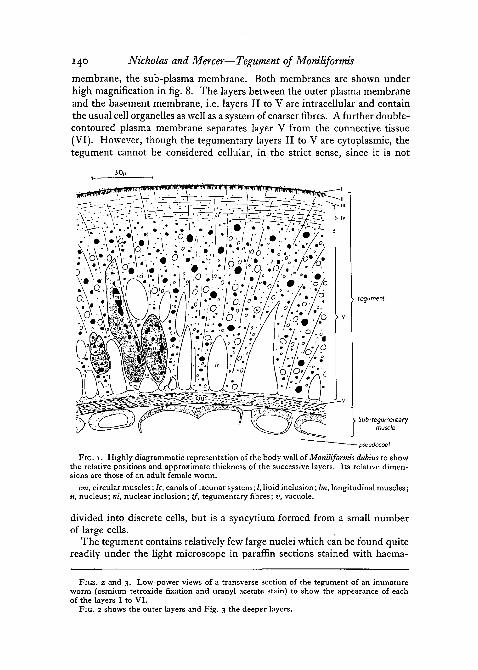
with haematoxylin and eosin, under the light microscope corresponds closelywith the tegument of other Acanthocephala described by Baer (1961) andCrompton (1963). Its structure as revealed by the light microscope is showndiagrammatically (fig. 1). Because of some confusion in nomenclature, whichhas arisen from the limitations of the light microscope, we have preferred toidentify the successive layers of the cuticle and tegument by the Romannumerals I to VI. The principle features observed under the electronmicroscope are shown in two low power electron micrographs (figs. 2 and 3).
The outermost layer I, which was named the epicuticle by Crompton(1963) in his study of Polymorphus minutus and which he concluded on histo-chemical grounds was formed from mucopolysaccharide, is clearly visibleunder the electron microscope (figs. 2 and 4). We have confirmed that in M.dubius layer I is PAS-positive and that the reaction is not diminished bysalivary digestion. It consists of a meshwork of fine fibres (less than 100 A indiameter), apparently randomly orientated. It shows no sharp outer boundaryand varies a good deal in thickness in different specimens, but is generallyabout 1 fx thick in adult worms and somewhat thinner in immature speci-mens.
Layer I is separated from the deeper layers of the tegument by a typicalplasma membrane ca 80 A thick and resolvable into 2 lines at high resolution(fig. 8). Beneath this and adjacent to it, there is a second more electron-dense
140 Nicholas and Mercer—Tegument of Moniliformis
membrane, the sub-plasma membrane. Both membranes are shown underhigh magnification in fig. 8. The layers between the outer plasma membraneand the basement membrane, i.e. layers II to V are intracellular and containthe usual cell organelles as well as a system of coarser fibres. A further double-contoured plasma membrane separates layer V from the connective tissue(VI). However, though the tegumentary layers II to V are cytoplasmic, thetegument cannot be considered cellular, in the strict sense, since it is not
o\-°. %\r>;i!/iH^:i--7^^r/vl [egument
Sub-tegumentarymuscle
- pseudocoel
FIG. I . Highly diagrammatic representation of the body wall of Moniliformis dubius to showthe relative positions and approximate thickness of the successive layers. Its relative dimen-sions are those of an adult female worm,
cm, circular muscles; Ic, canals of lacunar system; I, lipid inclusion; Im, longitudinal muscles;«, nucleus; ni, nuclear inclusion; tf, tegumentary fibres; v, vacuole.
divided into discrete cells, but is a syncytium formed from a small numberof large cells.
The tegument contains relatively few large nuclei which can be found quitereadily under the light microscope in paraffin sections stained with haema-
FIGS. 2 and 3. Low-power views of a transverse section of the tegument of an immatureworm (osmium tetroxide fixation and uranyl acetate stain) to show the appearance of eachof the layers I to VI.
FIG. 2 shows the outer layers and Fig. 3 the deeper layers.

FIGS. 2 and 3
W. L. NICHOLAS and E. H. MERCER
FIGS. 4 and 5
W. L. NICHOLAS and E. H. MERCER

Nicholas and Mercer—Tegument of Moniliformis 141
toxylin and eosin, but are difficult to find under the electron microscope inthe adult worm because they are widely separated in the tegument and the areasurveyed electron-microscopically is so small. They are easier to find in thesmaller immature worms. The nuclear membrane, the nucleolus and othernuclear contents resemble those described in higher animals.
Immediately beneath the sub-plasma membrane, in layer II there is a densemeshwork with long, tapering interstices running perpendicularly to thesurface (figs. 2, 4, and 8). The structure can only be recognized as a meshworkin partially (fig. 4) or wholly tangential sections. In strictly transverse orlongitudinal sections (i.e. sections at right angles to the surface) the structureappears to consist of branching septa perpendicular to the body surface,interspersed with less dense pores which taper as they approach the bodysurface. It is these septa which have been interpreted as fine striations byprevious workers using the light microscope. The septa in a large adult wormare about 3-3 \i deep. The pores between the septa taper from about o-6 ju,,proximally, to about 0-06 \L at the outer surface.
The interstices of the mesh contain membrane-bounded vesicles of varioussizes, many of them large enough to occlude the pores (fig. 8). Frequently,both the plasma membrane and the sub-plasma membrane appear to dipdown into the pore (fig. 8). The sub-plasma membrane cannot be followedvery far into the pores, but the plasma membrane often extends well down intothe pore (layer II), and it seems likely that the larger vesicles between thesepta and those in the deeper regions of the tegument are formed from aninvagination and pinching-off of the plasma membrane, their limiting mem-brane being derived from the plasma membrane (fig. 8). Rarely, two con-centric double-contoured membranes were seen to enclose a vesicle, andpossibly the outer of these two membranes is formed from the sub-plasmamembrane, though the latter has not been shown to be double-contoured atthe surface of the tegument (fig. 8).
The fibres which form a prominent part of the tegument under the lightmicroscope can be resolved in the electron microscope into bundles of finerfibres, which we shall term tegumentary fibres. The fibre bundles of thetegument follow an open spiral path and constitute a mechanical framework,distally continuous with the base of the meshwork (fig. 4) and proximallyjoined to the basement membrane (fig. 7). The individual fibres composingthe bundles are strongly stained by osmium tetroxide, potassium perman-ganate, uranyl acetate, and phosphotungstic acid, but poorly by lead citrate.They look tubular since only the periphery of the fibre shows affinity for
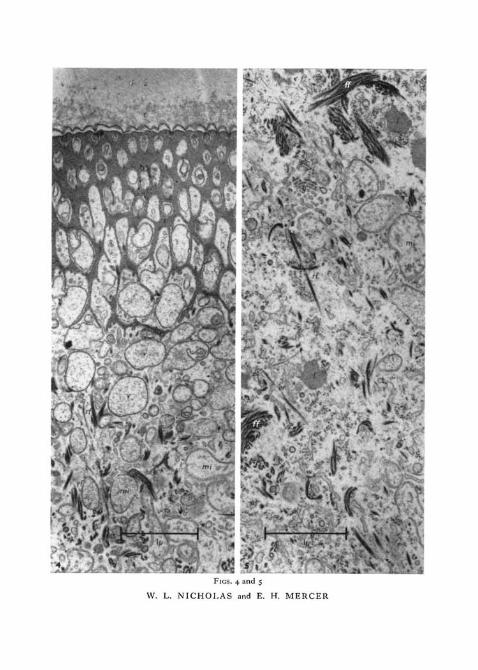
FIG. 4. This shows the peripheral region of the tegument (layers I to III) of an immatureworm in transverse section (osmium tetroxide fixation and uranyl acetate stain). The septahave been cut somewhat tangentially.
c, cuticle; mi, mitochondrion; rf, radial tegumentary fibres; s, septa; v, vesicles.FIG. 5. This shows layer IV in a transverse section from an immature worm (osmium
tetroxide fixation and uranyl acetate stain).er, endoplasmic reticulum; g, Golgi cluster; ff, feltwork of tegumentary fibres; /, lipid
inclusion; mi, mitochondrion; rf, radial tegumentary fibres.

142 Nicholas and Mercer—Tegument of Moniliformis
heavy metals and a 'core' does not (fig. 9). No banding has been observed inthese fibres.
No sharp distinction can be made between the meshwork and the fibrebundles, which resemble one another in their affinity for heavy metal stains,and, although at the periphery of the tegument the meshwork appears to beformed from closely packed and extremely fine fibrous material (fig. 8), at itsbase they aggregate into larger fibres continuous with those of the tegumentarybundles.
The fibre bundles containing relatively few fibres are predominantly radialin their orientation in layers III (fig. 2) and V (fig. 3). In layer IV they aremore numerous, contain more fibres, and form a feltwork (figs. 2 and 9).
Layer III, lying between the septa and the felt layer IV, from which it isnot sharply delimited, is filled with vesicles to the virtual exclusion of all otherstructures, apart from some radially orientated fibres (figs. 2 and 4). Many ofthese vesicles resemble those found in the pores and are probably derived fromthem. Similar vesicles are present in the deeper layers of the tegumentthough they are less numerous than at the base of the septa.
Mitochondria of somewhat atypical appearance are present throughout thetegumentary layers III to V (figs. 4, 5, 6, and n ) . In specimens taken fromimmature worms they were identified by the presence of two concentricdouble-contoured membranes, the inner membrane forming few poorlydeveloped cristae. In specimens taken from adult worms mitochondria weredifficult to identify with certainty, partly, we believe, because of difficulty inensuring rapid fixation of adult tissues, and partly because cristae were verypoorly developed. Vesicles containing myelinic forms were not infrequentand it seems possible that mitochondria show an increasing tendency todegenerate in the adult.
Lipid droplets are present throughout layers III, IV, and V ranging in sizefrom droplets which are clearly seen under the light microscope (in frozensections stained with Sudan black) to very small ultra-microscopic droplets(figs, 5 and 6). Their numbers varied greatly in different specimens and maybe very considerable (as observed in frozen sections under the light micro-scope and in Araldite sections under the electron microscope).
Other membranous organelles (elements of the reticulum) are sparselydistributed throughout layers III, IV, and V. Small stacks of parallel pairedmembranes (Golgi clusters) are common in immature specimens and less soin adults (figs. 5 and 6).
Small dense particles, 150 A to 400 A in diameter, are common throughoutthe cytoplasm. The larger particles are aggregates of smaller particles. From
Fie. 6. Layer V of an immature worm in transverse section (osmium tetroxide fixation anduranyl acetate stain), er, endoplasmic reticulum; g, Golgi cluster; /, lipid inclusion; mi, mito-chondrion; m, myelinic forms. An arrow points to intramitochondrial ribosome-like particles.
Fie. 7. This shows part of layer V, the basement membrane, and the connective tissueforming layer VI, in transverse section from an immature worm (osmium tetroxide fixationand uranyl acetate stain), bm, basement membrane; cf, connective tissue fibres; rl, radialtegumentary fibres; v, vesicles. The arrows point to the plasma membrane.

iI
FIGS. 6 and 7
W. L. NICHOLAS and E. H. MERCER
••^*-</.'7.z- •,wix::;*j.-*i-yis>w

W. L. NICHOLAS and E. H. MERCER

Nicholas and Mercer—Tegument of Moniliformis 143
their affinity for lead and their size the larger particles were thought to beglycogen. The diminution in their density when stained with lead followingsalivary digestion supports this identification. Some of the larger particles arepresent in vesicles and their density is not reduced following digestion. Thepresence of glycogen in tegumentary layers III to V was confirmed by adecrease in the PAS reaction following digestion.
The smaller particles are perhaps ribosomes. The presence of RNA in thetegument was established by fluorescence microscopy. An orange fluorescencecharacteristic of RNA in acridine orange was reduced following ribonucleasetreatment. The appearance of the tissue in the electron microscope was notsubstantially altered by ribonuclease digestion. Small groups of dense particlesare often found within structures which we have identified as mitochondria(fig. 6).
At the base of the tegument there is a double-contoured cell membrane,backed by a basement membrane (figs. 7 and 10.) Beyond this lies an extra-cellular space containing fine fibrils, groups of which show differing orienta-tions (figs. 3 and 7). These fine fibrils often show evidence of periodicity butit was not possible to identify them certainly with known fibrous structures.The coarser, radially-orientated tegumentary fibres appear attached to theplasma membrane, the point of insertion being marked by an electron-densethickening of the surface (fig. 7). Narrow projections of the plasma membrane,together with the fibre attachments, extend distally from the basement mem-brane. In some places, vesicles, resembling those found through the body ofthe tegument, line the distal surface of the plasma membrane (figs. 7 and 10).Sometimes the plasma membrane is drawn out to form an incompletelyenclosed vesicle (fig. 10). It seems logical to interpret the static picture seenhere as part of a dynamic process by which vesicles are either being pinchedoff from the plasma membrane, or, alternatively, are coalescing with it.Beneath the extracellular space lie circular and longitudinal muscle cells.
In their basic structure, the body walls of male, female, and immatureworms resemble one another. A very considerable increase in size accom-panies maturation, the female worms used in our study having about tentimes the diameter of the immature worms. Males are slightly smaller thanthe females. It is difficult to be precise, because the body is elastic andmoniliform, but all the layers (I to VI) increased in thickness, although wehave made no attempt to determine whether their relative proportions re-mained constant.
Other differences between the immature worm and the adult are that
FIG. 8. A high-power view of the outer surface of the tegument of an immature worm intransverse section (osmium tetroxide fixation and uranyl acetate stain). It shows structures,indicated by arrows, which have been tentatively identified as pinocytotic vacuoles in theprocess of formation. Also indicated is a vesicle («i) enclosed by two concentric plasmamembranes, c, cuticular fibres; pm, plasma membrane; sm, sub-plasma membrane; s, septain slightly tangential section; v, vesicle.
FIG. 9. A high-power view of the feltwork of fibres seen in layer IV of an adult worm intransverse section (osmium tetroxide fixation and uranyl acetate stain).

144 Nicholas and Mercer—Tegument of Moniliformis
membranous organelles (elements of the reticulum, Golgi clusters, mito-chondria) were less prominent in the latter. The tegumentary fibres on theother hand became more numerous.
DiscussionThe tegument of M. dubius shows many interesting features, some of which
cannot be satisfactorily interpreted by the purely descriptive approach adoptedin this paper. In this sense, our account must be considered as exploratoryand our interpretations as tentative. It confirms previous descriptions of thetegument (Baer, 1961) at the level resolvable by the light microscope.
The cytoplasmic fibres, which form so prominent a feature of the tegument,are unlike any previously described. From their architecture they clearlyserve a mechanical purpose. The dense meshwork found at the periphery ofthe tegument, layer II, is perhaps formed from the same material as the tegu-mentary fibres. The much finer fibres beneath the basement membrane aredifferent in appearance and superficially resemble fine collagen fibres. Thecuticle layer I (epicuticle of Crompton, 1963) is composed of still finerfilaments probably mucopolysaccharide in nature.
The invagination of the external surface membranes to form long tongueswithin the meshwork of fibres (layer II) of the tegument and the numerousvesicles seen immediately below this layer are strongly suggestive of pino-cytosis.
The formation of pinocytotic vacuoles over the whole surface of the bodycould provide a means of assimilating nutrients in an animal not providedwith an alimentary canal, but, in the absence of experimental evidence for thishypothesis, it must be considered tentative. A similar hypothesis has beenadvanced by Rothman (i960; 1963) for assimilation in cestodes which likeacanthocephala lack an alimentary canal, and by Threadgold (1963) to explainvacuolar structures seen in the tegument of another endoparasite, Fasciolahepatica, though this animal possesses a functional alimentary canal. Bjork-man and Thorsell (1964) studied the uptake of ferritin by F. hepatica with theelectron microscope, but, although they found ferritin had been taken up bythe tegument, they considered that the process was not pinocytosis, andpreferred the term 'transmembranosis'.
Crompton (1963) suggested that the epicuticular material in the acantho-cephalan P. minutus, which we have identified with layer I in M. dubius, wassecreted over the surface from the pores. If this view is correct, the structures
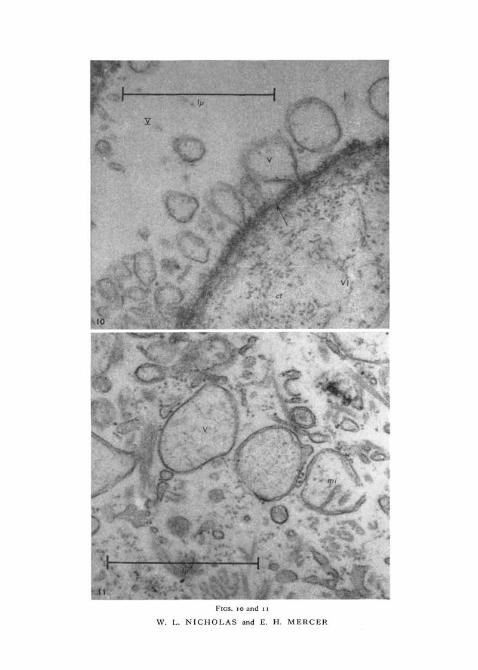
FlG. 10. This shows a high-power view of the basement membrane, which separates thelayers V and VI. Balloon-like extensions of the plasma membrane into layer V are shown.Transverse section of immature worm (osmium tetroxide fixation and uranyl acetate stain).Roman numerals identify the tegumentary layers and an arrow indicates the plasma mem-brane, bm, basement membrane; ct, connective tissue fibres; v, vesicle, apparently either inthe process of formation or of coalescing with the plasma membrane.
FIG. 11. This shows vesicles, with one double-contoured membrane, and mitochondriawith two concentric membranes. Transverse section of immature worm, layer III (osmiumtetroxide fixation and uranyl acetate stain), mi, mitochondrion; v, vesicle.
VI
f.uFIGS, IO and n
W. L. NICHOLAS and E. H. MERCER

Nicholas and Mercer—Tegument of Moniliformis 145
which we have suggested are pinocytotic vacuoles in the process of formationmight be secretory vacuoles in the act of discharging. These seem, however,rather too numerous to serve this purpose alone. Whatever the nature ofvacuolar structures the epicuticle must affect the passage of nutrients to thetegument.
Mitochondria show few poorly developed cristae, although these are betterdeveloped in the immature worms. There are signs (myelinic figures and lossof inner membrane) that mitochondria may degenerate in the adult. M. dubiusis a facultative anaerobe (Laurie, 1957,1959) and mitochondria may lose somedefinitive structural features in anaerobically respiring cells. Linnane, Vitols,and Nowland (1962) found that under anaerobic conditions, the yeast Toru-lopsis utilis lacked mitochondria. Instead a multi-membrane system waspresent, which when the cell was returned to aerobic conditions, was involvedin the formation of more typical mitochondria. Similarly Polakis, Bartley,and Meek (1964) found that the yeast Saccharomyces cerevisiae, when grownanaerobically, lacked mitochondria, though they developed under aerobicconditions in the presence of suitable substrates. Bjorkman and Thorsell(1962, 1964) found three types of mitochondria in the liver fluke, F. hepatica.In the tegument they were small and had very few cristae which they associ-ated with anaerobiosis and a low cytochrome content.
Cell boundaries were not apparent in the tegument under the light micro-scope, and, although it is not very practicable to search large cells under theelectron microscope for cell membranes, none was encountered, apart fromthe proximal and distal bounding membranes of the tegument. The absenceof recognizable cell membranes in the tegument would not be surprising inview of the ontogenetic development of the tegument. The tegument arisesin development by the considerable enlargement of a few cells, withoutconcomitant cell division. The nuclei of M. dubius become enlarged as theanimal develops in the definitive host but do not divide. Study of thedevelopment of the tegument from the embryonic stage seems necessary toestablish its true character (cellular or syncytial).
ReferencesBAER, J. G., 1961. In Traiti de Zoologie, edited by P. P. Grass6, vol. s, pp. 732-82. Paris
(Masson et Cie).BAKER, J. R., 1944. Quart. J. micr. Sci., 85, 1.BERTALANFFY, L. VON, MASIN, M., and MASIN, F., 1958. Cancer, n , 873.BJORKMAN, N., and THORSELL, W., 1962. Exp. Cell Res., 27, 342.BJORKMAN, N., and THORSELL, W., 1964. Exp. Cell Res., 33, 319.BRAND, T. VON, 1939. J. Parasit., 25, 329.BULLOCK, W. L., 1949. J. Morph., 84, 201.BURLINGAME, P. L., and CHANDLER, A. C, 1941. Amer. J. Hyg., 33D, 1.BURNHAM, K. D., 1957. Ph.D. Thesis. Iowa State University.CROMPTON, D. W. T., 1963. Parasitology, 53, 663.KARNOVSKY, M. J., 1961. J. biophys. biochem. Cytol., 11, 729.LAURIE, J. S., 1957. Exp. Parasit., 6, 245.LAURIE, J. S., 1959. Exp. Parasit., 8, 188.LINNANE, A. W., VITOLS, E., and NOWLAND, P. G., 1962. J. Cell Biol., 13, 345.

146 Nicholas and Mercer—Tegument of Moniliformis
Lwr, J. H., 1956. J. biophys. biochem. Cytol., 2, 799.MOORE, D. B., 1946. J. Parasit., 32, 257.NICHOLAS, W. L., and HYNES, H. B. N., 1963. In The Lower Metazoa, edited by E. C.
Dougherty. Berkeley (University of California Press).PALADE, G. E., 1952. J. exp. Med., 95, 285.PEARSE, A. G. E., i960. Histochemistry. London (Churchill).POLARIS, E. S., BARTLEY, W., and MEEK, G. A., 1964. Biochem. J., 90, 369.ROTHMAN, A. H., 1959. J. Parasit., 45 (4, suppl.), 28.ROTHMAN, A. H., i960. J. Parasit., 46 (5, suppl.), 10.ROTHMAN, A. H., 1961. J. Parasit., 47 (4, suppl.), 25.ROTHMAN, A. H., 1963. Trans. Amer. micr. Soc, 82, 22.SABATINI, D. D., BENSCH, K., and BARRNETT, R. J., 1963. J. Cell Biol., 17, 19.THREADGOLD, L. T., 1963. Exp. Cell Res., 30, 238.