the effects of temperature on growth rate and growth efficiency of juvenile common wolffish
TRANSCRIPT

The effects of temperature on growth rateand growth efficiency of juvenile commonwolffishIan McCarthy1*, Erlend Moksness2 and Dimitri A. Pavlov3
1 Division of Environmental and Evolutionary Biology, University Field Station,University of Glasgow, Rowardennan, Glasgow, G63 0AW, UK2 Institute of Marine Research, Flødevigen Marine Research Station, 4817 His,Norway3 Department of Biology, Moscow State University, 119899 Moscow, RussianFederation
The effects of temperature on the survival, growth rate and growth efficiency of larval andjuvenile common wolffish, Anarhichas lupus L. were studied at 0–31 days and 9–12 monthspost-hatching, respectively. The influence of temperature regime during egg incubation onsubsequent survival and growth was also examined. The fish were reared at constant watertemperatures of 5, 8, 11 and 14°C, and all groups were fed dry pellets. At age 1 month,maximum growth rates were observed at 11 and 14°C. Growth rates and survival of earlyjuveniles were dependent upon incubation history, high growth being obtained only if rearingtemperature exceeded the temperature of egg incubation. In juveniles at age 9–12 months, therelationships between temperature and growth, and temperature and growth efficiency wereparabolic: the optimum water temperatures for growth (Topt.G) and growth efficiency (Topt.GE)were 11°C and 9.7°C respectively. The growth rate and growth efficiency at these watertemperatures were 0.9% day–1 and 0.45 g weight gain per g food offered, respectively.
KEYWORDS: Growth, Growth efficiency, Temperature, Wolffish (Anarhichas lupus L.)
INTRODUCTION
The common wolffish, Anarhichas lupus L., is a marine fish that has been identifiedas a possible candidate species for cold-water aquaculture in northern Europe(Tilseth, 1990) and Atlantic Canada (Brown et al., 1995). A peculiarity of wolffishearly ontogeny, useful for artificial cultivation, is that the larvae (TL . 20 mm) hatchat an advanced stage, and complete their transition to the juvenile stage within afew days (Pavlov and Moksness, 1994b). In the past 5 y, laboratory studies havebeen conducted with the objective of controlling the life history of wolffish incaptivity, and investigating the culture conditions that promote optimal develop-ment and growth (Moksness and Pavlov, 1996).
Temperature is the major abiotic factor affecting growth in fish (Brett, 1979;Jobling, 1997). Therefore a knowledge of the temperature tolerance and optimum
* To whom correspondence should be addressed (e-mail: [email protected]).
0967–6120 © 1998 Chapman & Hall Ltd.
Aquaculture International 6, 207–218 (1998)

temperature for growth is important for an aquaculture species. Previous work hasexamined the temperature requirements of common wolffish at a number ofdifferent stages of the life cycle: egg maturation and incubation, and early larvalrearing (Pavlov and Moksness, 1994a,b; 1995; Moksness and Pavlov, 1996). Highquality eggs can be obtained when temperature is held below 10°C during finalmaturation (Pavlov and Moksness, 1996a), and a temperature regime where the eggsdevelop at 11°C during most of the embryonic period and are reared at 7°C duringthe sensitive stages of embryonic development has been proposed (Pavlov andMoksness, 1996b).
In captivity, wolffish larvae and juveniles have been reared from hatching tomaturation (Moksness, 1994; Pavlov and Moksness, 1994a). They can be fed on drypellets and attain a market size of 2 kg in 2 y (Moksness, 1994). The effect of feedcomposition and diet quality on the growth and feed conversion efficiency of larvaland juvenile common wolffish have been studied (Stefanussen et al., 1993; Moksnesset al., 1995), but the optimum water temperature for the growth and development oflarval and juvenile common wolffish remains unknown.
The aim of this study was to identify the optimum water temperature for thesurvival and growth of larval and juvenile common wolffish of different ages (, 1month old, 9–12 months old). Firstly, we examined whether there was anyconnection between the thermal history experienced during egg incubation on thesubsequent growth rates of larvae and juveniles , 1 month old. Secondly, in olderjuveniles (9–12 months post-hatching), the optimum water temperatures for growthand growth efficiency were determined.
MATERIALS AND METHODS
The experiments were carried out at the Institute of Marine Research, FlødevigenMarine Research Station, Norway, from February to July 1995. The larval andjuvenile common wolffish used in these experiments were reared from eggsobtained from adult broodstock held at the station. The origin of the broodstockand the methods of artificial fertilization and egg incubation were as described inearlier studies (Moksness, 1994; Pavlov, 1994; Pavlov and Moksness, 1994a, 1995,1996a). All groups of fish were kept in green-walled tanks (1 × 1 × 0.3 m; 260–280 l)and exposed to a 16 h light: 8 h dark photoperiod. Salinity varied between 33.6 and34.8‰, the average oxygen saturation was approximately 115%, and the dissolvedammonium concentration varied between 1.0 and 6.5 mmol NH4
1l–1.Desired experimental water temperatures were obtained using three sources: (1)
ambient water pumped directly from the sea channel which fluctuated between 5and 10°C during the study, and water from the same source that had either been (2)cooled to between 2 and 5°C or (3) heated to between 15 and 16°C. Water from twoof the three sources was mixed to provide the desired temperature of water flowinginto each tank. The water temperature in each tank was checked daily and carefullyregulated to minimize day-to-day fluctuations.
Experiment I
On 21 April (day 0), 60 newly hatched larvae were transferred into each of 12 smallfloating cages, 12 × 6 × 5 cm with a net bottom. The larvae originated from the eggs
208 I. McCarthy et al.

of a single female, and the eggs had been incubated at 7.0°C prior to hatching. Theaverage wet weight of another 60 larvae hatched from the eggs of the same femalewere measured to estimate the initial average wet weight of the experimentalgroups. The cages were placed in three tanks at temperatures of 5, 8 and 11°C. Thenumbers of cages at each temperature were three, three and six, respectively. At age18 days, three cages from the 11°C tank were transferred to a fourth tank with awater temperature of 14°C. Thus, four treatments (each containing three replicategroups) were obtained. These were designated treatments I.5, I.8, I.11 and I.11–14,respectively. The water temperatures during the experiment varied slightly fromthose stipulated (Table 1), but were not significantly different from 5, 8, 11 and 14°C(x2 = 0.49 – 2.32, 30 df, p # 1.00). Dead fish were removed from the cages daily andcounted and fish were transferred to a clean cage every 3 days to avoidaccumulation of uneaten food and faeces. The fish were fed dry pellets (SkrettingA/S ‘Tess Elite Plus 0.6’) six or seven times a day. The experiment was terminated on22 May, when the fish were aged 31 days. The wet weight of all surviving juvenileswas measured.
Experiment II
Experiment II was carried out on newly hatched larvae obtained from the eggs of asingle female. The eggs were incubated at a constant temperature of 11°C prior tohatching. Although this incubation temperature resulted in some larvae having alower number of fin rays in their caudal and anal fins, previous work has shown thatthis has no effect on their subsequent growth rates (Pavlov and Moksness, 1995). On20 June (day 0), 60 larvae were transferred to each of 12 small floating cagesidentical to those used in Experiment I. The average wet weight of another 60 larvaefrom the eggs of the same female were measured to estimate the initial average wetweight of the experimental groups. The cages were put into three tanks attemperatures of 8, 11 and 14°C. The numbers of cages at each water temperaturewere three, six and three respectively. At age 18 days, three cages from the 11°C tank
TABLE 1. The mean (SD) water temperatures (T°C)and specific growth rates (SGR, % day–1) for thefour treatments of juvenile common wolffish inExperiments I and II. The value presented for the11–14 °C group in Experiments I and II is the meanvalue over the whole experiment
Treatment (°C) T(°C) SGR
I. 58
1111–14
II. 811
11–1414
5.1 (0.5)7.7 (0.4)
10.8 (1.1)12.2 (1.8)
7.8 (0.5)11.0 (0.1)12.2 (1.6)14.0 (0.5)
1.86 (0.71)3.22 (0.40)4.14 (0.17)4.29 (0.24)1.71 (0.38)4.04 (0.34)4.81 (0.10)5.20 (0.20)
209Temperature effects on growth of common wolffish

were transferred to the 14°C tank. Thus, four treatments (each containing threereplicate groups) were obtained. These were designated treatments II.8, II.11, II.11–14, and II.14 respectively. The water temperatures during the experiment variedslightly from those stipulated (Table 1) but were not significantly different from 8, 11and 14°C (x2 = 0.33 – 0.67, 30 df, p # 1.00). All other experimental procedures(husbandry and feeding) were as described for Experiment I. The experiment wasterminated on 21 July when the fish were aged 31 days, and the wet weights ofsurviving fish were measured.
Experiment III
The fish used in Experiment III were juveniles that had hatched and start fed atFlødevigen during May and June 1994. The thermal history experienced by thesefish prior to the experiment was 6°C in May 1994, rising to approximately 12°C inSeptember–October, and then declining to 5°C by February 1995. Experiment IIIstarted on 20 February 1995 and lasted for 98 days. A total of 400 juvenile wolffishwere divided into four treatments, with two replicate tanks of 50 fish per treatment.Stipulated water temperatures were 5, 8, 11 and 14°C, respectively, and although thetemperatures during the experiment varied slightly from those stipulated (Table 2),they were not significantly different from 5, 8, 11 and 14°C (x2 = 1.32 – 3.51, 94–97 df,p # 1.00). The experimental groups were designated as III.5.1, III.5.2, III.8.1, III.8.2,III.11.1, III.11.2, III.14.1 and III.14.2, respectively.
The fish were fed a fish meal diet (NorSeaMink; Elite Plus, Skretting, Stavanger,Norway) by hand to apparent satiation two to six times per day. At each feeding, thefish were fed slowly and care was taken to ensure that food offered was eaten beforemore was presented. The fish usually fed quickly, and uneaten food on the bottomof the tank after 5 min was taken as indicating satiation. The amount of foodpresented to each group was calculated by weighing the amount of food in eachgroup’s feeding jar at the beginning and end of each week.
The average daily consumption rate (Con) of each group during the course of theexperiment were estimated using the following equation:
TABLE 2. The mean (SD) water temperatures (T°C) , initial and final numbers of fish (No, Nf), initialand final mean wet weight (Wo, Wf, g) survival rates expressed as a percentage of the initialnumber of fish in the group (S), total amount of feed offered (F, g) and total biomass gain (W, g)for the groups of common wolffish in experiment III. Mean values with the same superscript arenot significantly different from each other (Scheffe’s multiple comparison test, p < 0.05).
Group T(°C) No Wo (g) Nf Wf (g) S (%) F (g) W (g) SGR
5.15.28.18.2
11.111.214.114.2
4.9 (0.4)4.8 (0.4)7.7 (0.8)7.8 (0.5)
10.9 (0.8)11.0 (0.7)14.0 (0.7)13.9 (0.5)
5050505050505050
26.3 (2.5)a
26.0 (2.8)a
25.9 (2.4)a
25.6 (2.5)a
25.1 (2.7)a
25.7 (2.1)a
26.0 (2.8)a
25.4 (2.0)a
5049434847464746
42.0 (7.4)b
42.6 (7.3)b
52.8 (12.8)bcd
55.8 (12.5)cd
61.1 (20.5)cd
63.1 (15.5)c
55.4 (15.2)cd
51.2 (17.9)bd
10098869694929492
22312290233330223681378240524068
788792976
14001616161813001083
0.480.500.730.790.910.920.770.72
210 I. McCarthy et al.

Con (% day–1) = (FD*100)/(Nx*Wtmid*t) (1)
where FD is the amount of feed (g) offered to the group during the experiment, Nx isthe average number of fish in the group (initial 1 final/2) during the experiment,Wtmid is the average wet weight of the group (initial 1 final/2) and t is the length ofthe experiment in days.
Specific growth rate (SGR) was calculated according to the following equation:
SGR (% day–1) = [ln (Wt1) – ln (Wt0)]*100/t (2)
where Wt0 and Wt1 are the wet weights of the fish at start of the experiment and finalweight respectively, and t is the length of the experiment in days. The value of tvaried between experimental groups according to the days on which fish werekilled: 95 (groups III.5.1 and III.14.2), 96 (groups III.8.1 and III.11.1), 97 (groups III.5.2and III.14.1) and 98 days (groups III.8.2 and III.11.2), respectively.
Growth efficiency (GE, g weight gain per g food offered) was calculated from theformula:
GE = FD/W (3)
where FD is the amount of feed offered (g), and W is the biomass gain (g) by eachexperimental group during the course of the experiment. Correction has been madefor the mortalities in the groups.
Statistical analysisAll data are presented as means (SD). Statistical analysis was performed using oneway analysis of variance and Scheffe’s multiple comparison test as appropriate anddifferences present at the 5% level were considered significant. All statistical testsare described in Zar (1984).
RESULTS
Experiment I
In Experiment I, the highest mean survival of 56.0 (7.8)% was observed in the groupsof fish reared at 8°C (Fig. 1A), and the mean survival rate for fish reared at the otherthree temperatures was less than 31% [range 13.0 (8.2) to 30.3 (6.6)%]. Mortalitiesresulted from fish not weaning successfully on to the pelleted diet and wererecorded mainly at age 14–20 days in the 5 and 8°C treatments, and at age 11–20days in the 11 and 11–14°C treatments. The initial average wet weight was estimatedas 71.2 (3.7) mg. Final average wet weights increased significantly as the watertemperature increased from 5 to 11°C, but the average wet weights of larvae rearedat 11 and 14°C were similar (Fig. 1B). The specific growth rates of the larvae showeda similar temperature-dependent pattern and were highest in the 11 and 11–14°Ctreatments (Table 1). The mean SGR values were not significantly different betweenthe 11 and 11–14°C treatments (p , 0.44) but were significantly different between allother treatments (p , 0.05).
Experiment IIIn Experiment II, survival rates increased with increasing water temperature (Fig.1C), but were low and ranged between 16.5 (6.6) and 27.6 (4.9)%. Mortalities
211Temperature effects on growth of common wolffish
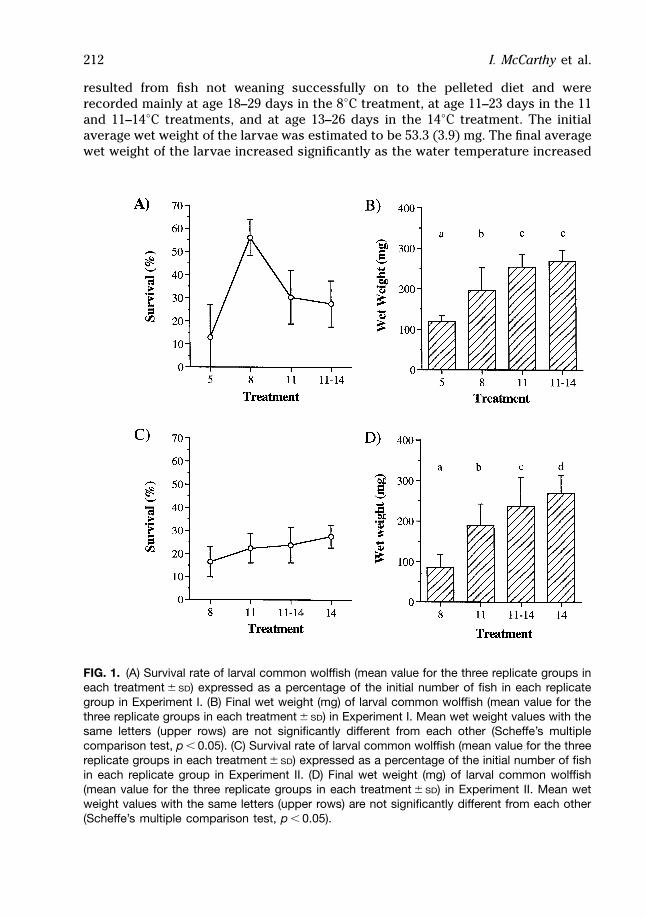
resulted from fish not weaning successfully on to the pelleted diet and wererecorded mainly at age 18–29 days in the 8°C treatment, at age 11–23 days in the 11and 11–14°C treatments, and at age 13–26 days in the 14°C treatment. The initialaverage wet weight of the larvae was estimated to be 53.3 (3.9) mg. The final averagewet weight of the larvae increased significantly as the water temperature increased
FIG. 1. (A) Survival rate of larval common wolffish (mean value for the three replicate groups ineach treatment 6 SD) expressed as a percentage of the initial number of fish in each replicategroup in Experiment I. (B) Final wet weight (mg) of larval common wolffish (mean value for thethree replicate groups in each treatment 6 SD) in Experiment I. Mean wet weight values with thesame letters (upper rows) are not significantly different from each other (Scheffe’s multiplecomparison test, p , 0.05). (C) Survival rate of larval common wolffish (mean value for the threereplicate groups in each treatment 6 SD) expressed as a percentage of the initial number of fishin each replicate group in Experiment II. (D) Final wet weight (mg) of larval common wolffish(mean value for the three replicate groups in each treatment 6 SD) in Experiment II. Mean wetweight values with the same letters (upper rows) are not significantly different from each other(Scheffe’s multiple comparison test, p , 0.05).
212 I. McCarthy et al.

from 5 to 14°C (Fig. 1D). The specific growth rates of the larvae showed a similartemperature-dependent pattern and were highest at 14°C (Table 1). The mean SGRvalues were significantly different among all four treatments (p , 0.05).
Experiment IIIIn Experiment III, the average initial wet weights of the experiment groups weresimilar, varying between 25.1 and 26.3 g (Table 2). The effects of water temperatureon survival, consumption and growth rates are shown in Table 2. Mortality ratesduring the experiment were lowest (# 2%) in the 5°C treatment and varied between4 and 8% in the other treatments, except for group III.8.1 where the mortality ratewas 14%. This group also exhibited poorer growth performance than group III.8.2(see Table 2). The fish in group III.8.1 exhibited no obvious differences inappearance or behaviour that might suggest any health problems and the reason forthe lower survival is not known.
The total amount of food eaten by the fish over the course of the experiment (F)increased with increasing water temperature; however, biomass gain was highest(. 1600 g) at 11°C. The average final wet weights of the experimental groupsincreased from 5 to 11°C, and declined at 14°C (Table 2), whereas estimatedconsumption rates increased with increasing water temperature from 5 to 14°C (Fig.2A). SGRs also increased with increasing water temperature from 5 to 11°C, werehighest at 11°C (0.91 and 0.92), and declined at 14°C to values comparable to thoserecorded at 8°C (Fig. 2B). The relationship between temperature and growth wasbest described by the following parabolic equation:
SGR = –0.012*T2 1 0.263*T – 0.529 (R2 = 0.959, n = 8, p , 0.001) (4)
where SGR is the specific growth rate (% day–1) and T is the water temperature (°C).Differentiation of Equation 4 indicated that the optimum water temperature forgrowth in juvenile common wolffish is 11°C (Table 2, Fig. 2B).
The relationship between water temperature and growth efficiency is shown inFig. 2C and was best described by the following parabolic equation:
GE = –0.006*T2 1 0.116*T – 0.073 (R2 = 0.916, n = 8, p , 0.001) (5)
where GE is the growth efficiency (g wet weight gain per g food offered) and T is thewater temperature (°C). Differentiation of Equation 5 indicated that the optimumwater temperature for growth efficiency in juvenile common wolffish is 9.7°C (Table2, Fig. 2C).
DISCUSSION
Larvae and 1 month old juveniles
A major constraint in the development of marine fish culture has been thesuccessful rearing of larvae beyond first-feeding. All marine fish species that arecurrently cultured have small pelagic eggs, and larvae which undergo met-amorphosis to the juvenile stage. In contrast to other marine fish, the commonwolffish has an early ontogeny similar to salmonids: large eggs (5.0–6.5 mm) andlarvae (20–25 mm) and the ability to start-feed on relatively large particulate fooditems at first-feeding. This should enable high larval survival rates and growth using
213Temperature effects on growth of common wolffish

formulated diets (Moksness et al., 1989; Tilseth, 1990; Pavlov and Moksness, 1994b)but the survival rates of larval and early juvenile common wolffish offered pelleteddiets have been low (Ringø et al., 1987; Moksness et al., 1989) and in this study,survival rates ranged from 1.8 to 63.6% in Experiment I and from 7.7 to 32.1% inExperiment II. In contrast, survival rates of 60–100% have been recorded forcommon wolffish fed live feed (Ringø et al.,1987; Moksness et al., 1989; Pavlov, 1995;Wiseman and Brown, 1996). Mortalities that occur during the larval phase in fish fedformulated diets may be due to the inability of the fish to wean on to the artificialfeed (Ringø et al., 1987; Strand et al., 1995; present study), but Strand et al., (1995)reported a survival rate of 82% for first-feeding common wolffish reared in a low-water-level raceway system and fed a specially formulated floating dry feed diet.The results of this study suggest that previous thermal history influences the
FIG. 2. The relationships between water temperature and (A) consumption (Con, % day–1), (B)specific growth rate (SGR, % day–1) and (C) growth efficiency (g weight gain per g food offered)for juvenile common wolffish in Experiment III.
214 I. McCarthy et al.

survival of early juvenile common wolffish (Fig. 1, Table 1). However, in bothExperiments I and II, the highest growth rates of the surviving fish were observed at14°C, which we conclude is the optimum water temperature for growth of earlyjuvenile common wolffish.
9–12 month old juveniles
In this study, the growth of 9–12 month old juvenile wolffish was examined over arange of water temperatures between 5 and 14°C with the aim of determining theoptimum temperature for growth and growth efficiency. The influence of watertemperature on consumption, growth and growth efficiency in fish has been wellstudied (see reviews by Brett, 1979; Elliott, 1982; Jobling, 1997). Within the thermalrange for a species, the relationships between water temperature and consumption,growth and growth efficiency are parabolic (Imsland et al., 1996; Bjornsson andTryggvadottir, 1996): each curve consisting of an ascending limb within the lowerpart of the thermal range, attaining a maximum value at its optimum watertemperature and then declining with any further increase in water temperature (seeFig. 2 in Jobling, 1997). Previous studies have shown that the optimum watertemperature for consumption (Topt.R) is higher than the optimum water temperaturefor growth (Topt.G), which in turn is higher than the optimum water temperature forgrowth efficiency (Topt.GE) (Woiwode and Adelman, 1991; Bjornsson and Tryggva-dottir, 1996; Jobling, 1997). The results of the present study (Fig. 2) are in agreementwith this rate–temperature model and suggest Topt.G and Topt.GE for juvenile commonwolffish are 11 and 9.7°C respectively. The water temperature at which consumptionrates are maximal cannot be ascertained from the results of this study as nomaximum value was observed between 5 and 14°C (Fig. 2A).
The results of this study predict a growth rate of 0.9% day–1 for juvenile commonwolffish at 11°C (Fig. 2B) and a maximum growth efficiency of 0.45 g wet weight gainper g food offered at 9.7°C (Fig. 2C). Previous work has shown that growth rates andgrowth efficiency in juvenile common wolffish are significantly influenced bynutritional factors such as feed composition and dietary protein source and by thephysical characteristics of the pellets (floating or sinking) (Stefanussen et al., 1993;Moksness et al., 1995; Strand et al., 1995). The results of these studies would suggestthat both increased growth and higher growth efficiency will be possible at 10–11°Cas the optimal dietary formulations for juvenile common wolffish become knownand as rearing conditions are optimized.
In this study, Topt.G and SGR recorded at that water temperature decreased withincreasing age. For early juvenile common wolffish (,1.0 g), a maximum growth rateof 5.4% day–1 was recorded at 14°C. In contrast, older juvenile wolffish (50–60 g, 9–12months old) exhibited a maximum growth rate of 0.9% day–1 at 11°C. Thisontogenetic variation has been reported for a number of fish species. Decreases intemperature optimum and maximum growth rate with increasing age have beenreported for cod (Gadus morhua) (Pedersen and Jobling, 1989), plaice (Pleuronectesplatessa) (Fonds et al., 1992), turbot (Scophthalmus maximus) (Imsland et al., 1996)and halibut (Hippoglossus hippoglossus) (Bjornsson and Tryggvadottir, 1996). How-ever, no such relationship has been reported for sockeye salmon (Brett, 1974) orbrown trout (Salmo trutta) (Elliott, 1982). From mathematical modelling of fishgrowth, Cuenco et al. (1985) predicted that Topt.G would decrease by 1–2°C for a 50
215Temperature effects on growth of common wolffish

fold increase in body weight. However, the available data indicate that a 3–4°Cdecrease in Topt.G is more likely (Imsland et al., 1996; Bjornsson and Tryggvadottir,1996; this study). Topt.G values for larger wolffish (. 60 g) have not been experimen-tally determined; however, assuming a 3°C decrease with a 50 fold increase in bodyweight, then Topt.G for 2.5 kg wolffish will be 8°C. The relationship betweentemperature and growth efficiency for juvenile common wolffish in this study wasparabolic (Fig. 2C). A similar relationship has been reported in salmonids (Brett etal., 1969; Elliott, 1982), cod (Jobling, 1988) and halibut (Bjornsson and Tryggvadottir,1996). Bjornsson and Tryggvadottir (1996) have reported a decrease in Topt.GE withincreasing body size (10 g to 5 kg), although this relationship has yet to be examinedfor common wolffish.
CONCLUSIONS AND RECOMMENDATIONS
1. In the experiments where larvae and early juveniles of common wolffish werereared from hatching for 1 month at temperatures between 5 and 14°C, thehighest growth rates were observed at 14°C.
2. The growth rates and survival of early juveniles appeared to be influenced bythe thermal incubation history experienced during the egg stage.
3. In fish of 9–12 months, the optimum water temperatures for growth rate andgrowth efficiencies were predicted to be 11 and 9.7°C, respectively. Therefore,water temperatures of 9–11°C should be regarded as optimal for juvenilecommon wolffish of this age.
ACKNOWLEDGEMENTS
We would like to thank the Norwegian Research Council (EM and DP) and theCarnegie Trust for the Universities of Scotland (IM) for financial support. Thetechnical assistance of Inger Henriksen, Vetle Madsen and many other members ofthe Flødevigen Research Station is highly appreciated. We are grateful to Dr. ChrisCarter, the editor of Aquaculture International and two anonymous referees for theircomments on an earlier version of this paper. This study was done according to theagreement of scientific co-operation between the Institute of Marine Research,Flødevigen Marine Research Station (Norway) and Moscow State University, Chair ofIchthyology (Russia).
REFERENCES
Bjornsson, B. and Tryggvadottir, S.V. (1996) Effects of size on optimal temperature for growthand growth efficiency of immature Atlantic halibut (Hippoglossus hippoglossus L.).Aquaculture 142, 33–42.
Brett, J.R. (1974) Tank experiments on the culture of pan-size sockeye (Oncorhynchus nerka)and pink salmon (O. gorbuscha) using environmental control. Aquaculture 4, 341–352.
Brett, J.R. (1979) Environmental factors and growth. In: Fish Physiology Vol. VIII Bioenergeticsand Growth (eds W.S. Hoar, D.J. Randall and J.R. Brett) Academic Press: New York, pp.599–675.
Brett, J.R., Shelbourn, J.E. and Shoop, C.T. (1969) Growth rate and body composition offingerling sockeye salmon, Oncorhynchus nerka, in relation to temperature and rationsize. Journal of the Fisheries Research Board of Canada 32, 2363–2394.
Brown, J., Helm, M. and Moir, J. (1995) New candidate species for aquaculture. In: Coldwater
216 I. McCarthy et al.

Aquaculture in Atlantic Canada 2nd edition (ed. A.D. Boghen) Tribune Press: Sackville NB,pp. 341–362.
Cuenco, M.L., Stickney, R.R. and Grant, W.E. (1985) Fish bioenergetics and growth inaquaculture ponds: II. Effects of interactions among size, temperature, dissolved oxygen,unionized ammonia and food on growth of individual fish. Ecological Modelling 27,191–206.
Elliott, J.M. (1982) The effects of temperature and ration size on the growth and energetics ofsalmonid fish in captivity. Comparative Biochemistry and Physiology 73B, 81–92.
Fonds, M., Cronie, R., Vethaak, A.D. and van der Puyl, P. (1992) Metabolism, food consumptionand growth of plaice (Pleuronectes platessa) and flounder (Platichthys flesus) in relation tofish size and temperature. Netherlands Journal of Sea Research 29, 127–143.
Imsland, A.K., Sunde, L.M., Folkvord, A. and Stefansson, S.O. (1996) The interaction oftemperature and fish size on growth of juvenile turbot. Journal of Fish Biology 49,926–940.
Jobling, M. (1988) A review of the physiological and nutritional energetics of cod, Gadusmorhua L., with particular reference to growth under farmed conditions. Aquaculture 70,1–19.
Jobling, M. (1997) Temperature and growth: modulation of growth rate via temperaturechange. In: Society for Experimental Biology Seminar Series 61: Global Warming:Implications for Freshwater and Marine Fish (eds C.M. Wood and D.G. McDonald)Cambridge University Press: Cambridge, pp. 225–253.
Moksness, E. (1994) Growth rates of the common wolffish, Anarhichas lupus L., and spottedwolffish, A. minor Olafsen, in captivity. Aquaculture and Fisheries Management 25,363–371.
Moksness, E. and Pavlov, D.A. (1996) Management by life cycle of wolffish, Anarhichas lupus L.,a new species for cold water aquaculture: a technical paper. Aquaculture Research 27,865–883.
Moksness, E., Gjøsæter, J., Reinert, A. and Fjallstein, I.S. (1989) Start-feeding and on-growing ofwolffish (Anarhichas lupus) in the laboratory. Aquaculture 77, 221–228.
Moksness, E., Rosenlund, G. and Lie, Ø. (1995) The effect of fish meal quality on growth ofjuvenile wolffish (Anarhichas lupus). Aquaculture Research 26, 109–115.
Pavlov, D.A. (1994) Maturation and artificial fertilisation of the eggs of captive commonwolffish (Anarhichas lupus L.) from the White Sea. Aquaculture and Fisheries Management25, 891–902.
Pavlov, D.A. (1995) Growth of juveniles of White Sea common wolffish, Anarhichas lupus L., incaptivity. Aquaculture Research 26, 195–203.
Pavlov, D.A. and Moksness, E. (1994a) Production and quality of eggs obtained from wolffish(Anarhichas lupus L.) reared in captivity. Aquaculture 122, 295–312.
Pavlov, D.A. and Moksness, E. (1994b) Reproductive biology, early ontogeny, and effect oftemperature on development in wolffish: comparison with salmon. Aquaculture Inter-national 2, 133–153.
Pavlov, D.A. and Moksness, E. (1995) Development of wolffish eggs at different temperatureregimes. Aquaculture International 3, 315–335.
Pavlov, D.A. and Moksness, E. (1996a) Repeat sexual maturation of wolffish (Anarhichas lupusL.) broodstock. Aquaculture 139, 249–263.
Pavlov, D.A. and Moksness, E. (1996b) Sensitive stages during embryonic development ofwolffish, Anarhichas lupus L. determining the final numbers of rays in unpaired fins andskeletal abnormalities. ICES Journal of Marine Science 53, 731–740.
Pedersen, T. and Jobling, M. (1989) Growth rates of large, sexually mature cod, Gadus morhua,in relation to condition and temperature during an annual cycle. Aquaculture 81,161–168.
217Temperature effects on growth of common wolffish

Ringø, E., Olsen, R.E. and Bøe, B. (1987) Initial feeding of wolffish (Anarhichas lupus L.) fry.Aquaculture 62, 33–43.
Stefanussen, D., Lie, Ø., Moksness, E. and Ugland, K.I. (1993) Growth of juvenile commonwolffish (Anarhichas lupus) fed practical fish feeds. Aquaculture 114, 103–111.
Strand, H.K., Hansen, T.K., Pedersen, A., Falk-Petersen, I.B. and Øiestad, V. (1995) First feedingof common wolffish on formulated dry diets in a low water-level raceway system.Aquaculture International 3, 1–10.
Tilseth, S. (1990) New species for cold-water farming. Aquaculture 85, 235–245.Wiseman, D.L. and Brown, J.A. (1996) Early growth and survival of larval striped wolffish
(Anarhichas lupus): a behavioural approach. Bulletin of the Aquaculture Association ofCanada 96–1,12–13.
Woiwode, J.G. and Adelman, I.R. (1991) Effects of temperature, photoperiod and ration size ongrowth of hybrid striped bass x white bass. Transactions of the American Fisheries Society120, 217–229.
Zar, J.H. (1984) Biostatistical analysis (2nd edition). Prentice Hall: London, 718 pp.
218 I. McCarthy et al.