the amino-acid sequence of porcine adenylate kinase from skeletal muscle
TRANSCRIPT

Eur. J. Biochem. 43,131-144 (1974)
The Amino-Acid Sequence of Porcine Adenylate Kinase from Skeletal Muscle
Albert HEIL, Gudrun M~LLER, Lafayette NODA, Thomas PINDER, Heiner SCHIRMER, Ilse SCHTRMER, and Inge VON ZABERN
Max-Plrtnck-Institut fur Medizinische Forschung, Heidelberg, and Department of Biochemistry, Dartmouth Medical School, Hanover, New Hampshire
(Received August 29/November 23, 1973)
1. Adenylate kinase (ATP :AMP phosphotransferase) has been purified 490-fold from porcine muscle with a final yield of 60 mg/kg muscle.
2. The amino-acid composition is Asp,,, Am,, ThrI4, Ser,,, Glu,,, Gln,, Pro,, Gly,,, Ala,, Cysz, Val,,, Met,, Ile,, Leu,,. Tyr,, Phe,, Lyszl, His,, Arg,,.
3. The protein molecule is a single polypeptide chain of 194 amino-acid residues with an acetyl- methionine a t the N-terminus and a lysine residue at the C-terminus.
4. Cyanogen bromide cleavage of carboxymethylated adenylate kinase yielded six fragments which were further degraded by using trypsin, chymotrypsin, thermolysin, subtilisin or 6-prote- ase. Sequence data on the resulting peptides are summarized in the present report, full details are given in a supplementary paper which has been deposited at CNRS from where copies can be obtained.
5. The primary structure of porcine adenylate kinase is : Ac-Met-Glu-Glu-Lys-Leu-Lys-Lys-
Ser-Lys-Ile-Ile-Phe-Val-Val-Gly-Gly-Pro-Gly - Ser - Gly- Lys - Gly -Thr- Gln- Cys- Glu- Lys- Ile -Val-
G~-Lys-Tyr-Gly-Tyr-Thr-His-Leu-Ser-Thr-Gly-~p-Leu-Leu-Arg-~a-Glu-Val-Ser-Ser-Gly-Ser-Ala-
Arg-Gly-Lys-Met-Leu-Ser-Glu-Ile-Met-Glu-Lys-Gly-Gln-Leu-Val-Pro-Leu-Glu-Thr-Val-Leu -Asp - Met-Leu-Arg-Asp-Ala-Met-Val-Ala-Lys-Val-Asp-Thr-Ser-Lys-Gly-Phe-Leu-Ile-Asp-Gly-Tyr-Pro-
Arg-Glu-Val-Lys-Gln-Gly-~lu-Glu-Phe-Glu-Arg-Lys-Ile-Gly-Gln-Pro-Thr-Leu-Leu-Leu-Tyr -Val -
Asp-Ala-Gly-Pro-Glu-Thr-Met-Thr-Lys-~g-Leu-Leu-Lys-Arg-Gly-Glu-Thr-Ser-Gly-Arg-Val-Asp - Asp-Asn-Glu-Glu-Thr-Ile-Lys-Lys-Arg-Leu-Glu-Thr-T~-T~-Lys-~a-Thr-Glu-Pro-Val-Ile-Ala-P~e-
Tyr-Glu-Lys-Arg-Gly-Ile-Val-Arg-Lys-Val-Asn-~a-Glu-Gly-Ser-Val-Asp-Asp-Val-Phe-~er-G~-Val-
Cys-Thr-His-Leu- Asp-Thr- Leu-Lys.
10 20
30 40 60
60 70
80 90
100 110
120 130 140
150 160
170 180
190
Adenylate kinases catalyze the reaction ATP + AMPS 2 ADP. These enzymes seem to be as ubiquitous in nature as the adenine nucleotides and have been isolated from Escherichia coli, baker’s yeast, from the muscles of cockroach, squid, carp, rabbit, swine and man, from liver mitochondria of rat, swine and cow, and from other sources [l]. As a fist
Abbreviations. CM, carboxymethyl; dansy1,b-dimethyl- aminonaphthalene- 1 -adphony1 .
Enzyntes (CBN recommendations 1972). Adenylate kinase (EC 2.7.4.3); hexokinase (EC 2.7.1.1); trypsin (EC 3.4.21.4); chymotrypsin (EC 3.4.21.1); subtilisin (EC 3.4.21.14); carboxypeptidase AM (EC 3.4.12.2) ; carboxypcptidase BM (EC 3.4.12.3); aminoacylase (EC 3.6.1.14); thermolysin (EC 3.4.24.4); a-proteaae (EC 3.4.21.-).
step in establishing the relationship between struc- ture and function of one adenylate kinase the crystal structure of the enzyme isolated from pig muscle is being studied [2]. Complementary to the X-ray diffraction investigation the amino-acid sequence of this protein, a single polypeptide chain of 194 residues, has been determined. The present paper describes the sequence analysis.
MATERIALS AND METHODS Chemicals were of reagent grade unless stated
otherwise in the following sections. N-Ethylmorpho- line was redistilled.
Em. J. Riochem. 43 (1974)

132 Amino-Acid Sequence of Porcine Adenyiate Kinase
Purijication of Adenylate Kinase
All procedures are carried out in a cold room (0-3 "C) or in an ice-bath. Porcine muscle (6 kg) trimmed reasonably free of fat is ground (motor- driven grinder with 118-inch holes) and is stirred gently for half an hour with 15 1 cold 0.01 M KCl. Particles are removed by draining and pressing in cheese-cloth bags (fraction I).
Using an ice-bath a t a pH meter and with efficient mechanical stirring, 2 M HCl is added rapidly to pH 3.5 (about 320 ml 2 M HC1) and the solution is gently stirred for 5 min. 2 M sodium hydroxide is quickly added to bring the solution to approximately pH5, then dropwise during about 20min to bring the pH to 7.0 (total about 375 ml 2 M NaOH). The suspension is allowed to stand for about 15 min. The denatured proteins are removed by centrifugation or by filtration with the aid of acid-washed Celite, (diatomaceous earth, about 0.15 x original weight of muscle) to give fraction 11.
The pH of the yellow solution is brought to 5.0 with 5 M acetic acid and the enzyme is adsorbed on a phosphocellulose column (8.73 x 25 cm, previously cycled and equilibrated with 0.1 M sodium acetate, pH 5.0; new lots of phosphocellulose should be tested before use for enzyme adsorbing property). A flow rate of about 60 ml per min is maintained by a peri- staltic pump. The position of adsorbed red proteins indicates roughly the location of the enzyme on the column. After adsorption (about 6 h) the column is washed with 2 column volumes of cold distilled water, then washed overnight with 0.05 M imidazole-HC1 buffer pH 7.0 (about 30 ml per min, total 16-18 1). The column is washed the next day with 0.07M imidazole-HC1 buffer pH 7.0, while collecting 20-ml aliquots and monitoring for enzymatic activity as well as absorbance at 280nm. Depending on the batch of phosphocellulose, the concentration of the imidazole buffers may need to be adjusted to remove as much inert protein as possible. The washing is continued until the effluent has an absorb- ance of 0.100 or less a t 280 nm and traces of enzyme (3-10 unitslml) begin to emerge from the column. Then the enzyme is eluted with one column volume of 5 mM AMP in 0.07 M imidazole adjusted to pH 7.0 followed by 21 0.015M ammonium sulfate in the same buffer. The 20-ml aliquots with more than 200 units/ml are collected. The use of AMP precludes measurements of absorbance at 280 nm for protein. Fraction I11 has a volume of approximately 1 1.
As soon as possible the enzyme is precipitated by adding solid ammonium sulfate to 9001, saturation a t pH 6.0 and is collected by centrifugation, preferably after flocculating overnight. The tan-colored paste may be stored in a deep-freeze to accumulate prep- arations for the next step or dissolved immediately in a minimal amount of buffer for gel separation on a
Sephadex G-75 column. For two combined prepara- tions ( 2 x 6 kg muscle) a column ( 5 . 2 0 ~ 160 cm) is used with 0.1 M imidazole-H,SO, buffer pH 7.0, with a flow rate of about 1.5 ml/min. Inert proteins start to come off at 0.25-0.35~column volume (900- 1300 ml) and the adenylate kinase activity comes off at 0.40-0.52xcolumn volume (1400 to 1900 ml). The 15-ml aliquots with enzymatic activity greater than 1500 unitslml are collected (fraction IV) and precipitated a t 9001, ammonium sulfate saturation and pH 6.0.
Crystallization is carried out a t a protein concen- tration of 30 mg/ml, 0.01 M phosphate pH 6.0 and about 62 O/, ammonium sulfate saturation. As crystal- lization proceeds the ammonium sulfate saturation may be slowly increased by a few percent to increase the yield.
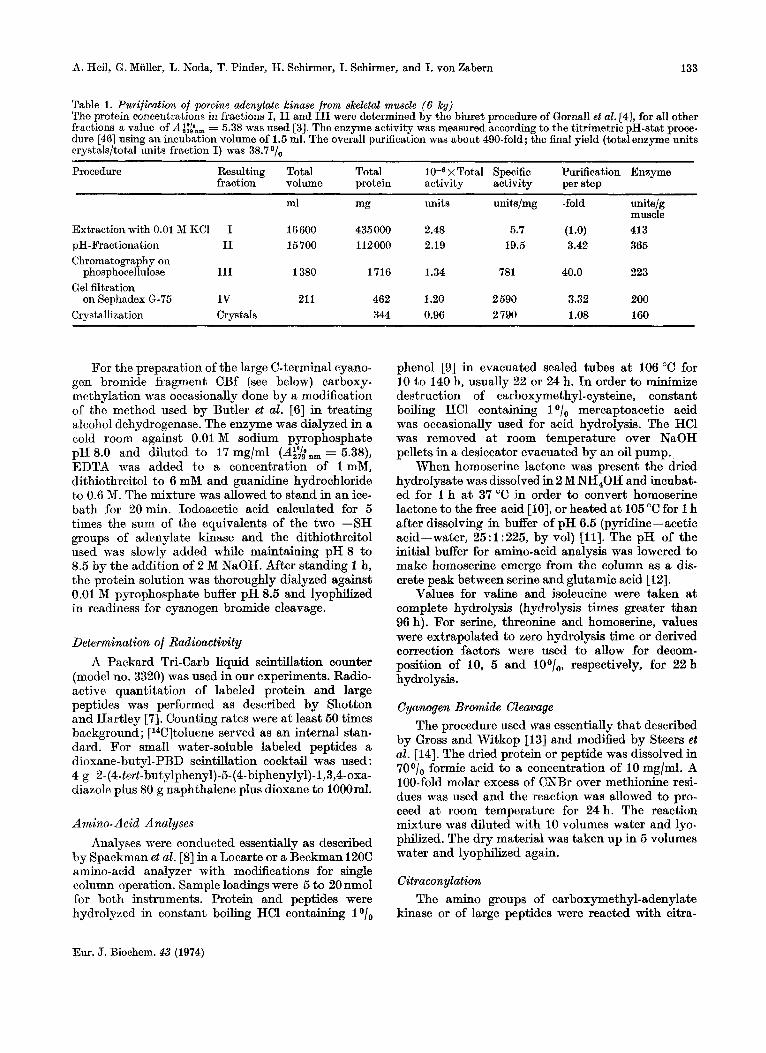
The above isolation procedure (Table 1) offers several advantages compared with that described by Schirmer et al. [3]. The yield of enzyme per kg muscle is much higher and fewer steps are involved in the preparation of the protein. The phosphocellulose step has been improved by eluting the enzyme with the substrate AMP.
Carboxymethylation Reaction of adenylate kinase with iodoacetate
was carried out essentially as described by Craven et al. [5]. Unlabeled iodoacetic acid was recrystallized twice from heptane ; iodo[2-14C]acetic acid (Radio- chemical Centre, Amersham) was used without further purification.
In a typical experiment 20 pmol (= 430 mg) freeze-dried adenylate kinase was suspended in 20 ml 0.5 M Tris-acetic acid buffer pH 8.7. Guanidine hydro- chloride (19.1 g, twice recrystallized from methanol) and 0.1 ml 200 mM dithiothreitol were added. The mixture was stirred for 15 rnin a t 0 "C and then the alkylation reaction was started in the dark by adding 1 ml 240 mM iodo[2-14C]acetic acid, the specific activity of which was 200 pCi/nmol. 0.5 M Tris- acetic acid buffer pH 8.7 was added to a final volume of 40 ml and the reaction was allowed to proceed for 1 h in the dark a t room temperature. Then a 5-fold molar excess of dithiothreitol over iodoacetate was added. The solution was stirred for 15 min to permit complete reaction of free iodoacetate and then dialyz- ed in the dark a t 4 "C against 10 mM acetic acid until the diffusate was free of radioactivity. The specific radioactivity of the protein was found to be 400pCi/ nmol.
All the incorporated radioactivity was associated with 8-carboxymethyl-cysteine residues. This was shown by purifying radioactive peptides and by electrophoretic studies on the amino-acids which were released by acid hydrolysis or complete enzymatic hydrolysis of the alkylated protein.
Eur. J. Biochem. 43 (1974)
A. Reil, G. Miiller, L. Noda, T. Pinder, H. Schirmer, I. Schirmer, and I. von Zabern 133
Table 1. Purification of porcine adenylate kinase from skeletal muscle ( 6 kg) The protein concentrations in fractions I, I1 and I11 were determined by the biuret procedure of Cornall et al. [4], for d l other fractions a value of A i:tn, = 5.38 was used [3]. The enzyme activity was measured according to the titrimetric pH-stat proce- dure [46] using an incubation volume of 1.5 ml. The overall purification was about 490-fold; the final yield (total enzyme units crystals/tot,al units fraction I) was 38.7 O/,,
Procedure ~ ~~~~~~ ~~
Resulting Total Total x Total Specific Purification Enzyme fraction volume protein activity activity per step
ml mg units units/mg -fold unitslg muscle
Extraction with 0.01 M KCI I 16 600 435 000 2.48 5.7 (1.0) 413 pH-Fractionation I1 15700 112000 2.19 19.5 3.42 365 Chromatography on
phosphocellulose I11 1380 1716 1.34 781 40.0 223 Gel filtration
Crystallization Crystals 344 0.96 2 790 1.08 160 on Sephadex 6-75 IV 21 1 462 1.20 2 590 3.32 200
For the preparation of the large C-terminal cyano- gcn bromide fragment CBf (see below) carboxy- methylation was occasionally done by a modification of the method used by Butler et al. [6] in treating alcohol dehydrogenase. The enzyme was dialyzed in a cold room against 0.01 M sodium pyrophosphate pH 8.0 and diluted to 17 mg/ml (A$bnm = 5.38), EDTA was added to a concentration of 1 mM, dithiothreitol to 6 mM and guanidine hydrochloride to 0.6 M. The mixture was allowed to stand in an ice- bath for 20 min. Iodoacetic acid calculated for 5 times the sum of the equivalents of the two -SH groups of adenylate kinase and the dithiothreitol used was slowly added while maintaining pH 8 to 8.5 by the addition of 2 M NaOH. After standing 1 h, the protein solution was thoroughly dialyzed against 0.01 M pyrophosphate buffer pH 8.5 and lyophilized in readiness for cyanogen bromide cleavage.
Determination of Radioactivity A Packard Tri-Carb liquid scintillation counter
(model no. 3320) was used in our experiments. Radio- active quantitation of labeled protein and large peptides was performed as described by Shotton and Hartley [7]. Counting rates were a t least 50 times background ; [14C]toluene served as an internal stan- dard. For small water-soluble labeled peptides a dioxane-butyl-PBD scintillation cocktail was used : 4 g 2- (4-tert-butylphenyl)-5- (4-biphenyly1)- 1,3,4-oxa- diazole plus 80 g naphthalene plus dioxane to IOOOml.
Amino-Acid Analyses Analyses were conducted essentially as described
by Spackman et al. [8] in a Locarte or a Beckman 120C a,mino-acid analyzer with modifications for single column operation. Sample loadings were 5 to 20 nmol for both instruments. Protein and peptides were hydrolyzed in constant boiling HC1 containing lo/,
phenol [9] in evacuated sealed tubes at 106 "C for 10 to 140 h, usually 22 or 24 h. In order to minimize destruction of carboxymethyl-cysteine, constant boiling HC1 containing I O/, mercaptoacetic acid was occasionally used for acid hydrolysis. The HC1 was removed a t room temperature over NaOH pellets in a desiccator evacuated by an oil pump.
When homoserine lactone was present the dried hydrolysate was dissolved in 2 M NH40H and incubat- ed for I h a t 37 "C in order to convert homoserine lactone to the free acid [lo], or heated a t 105 "C for 1 h after dissolving in buffer of pH 6.5 (pyridine-acetic acid-water, 25 : I : 225, by vol) [Ill. The pH of the initial buffer for amino-acid analysis was lowered to make homoserine emerge from the column as a dis- crete peak between serine and glutamic acid [12].
Values for valine and isoleucine were taken a t complete hydrolysis (hydrolysis times greater than 96 h). For serine, threonine and homoserine, values were extrapolated to zero hydrolysis time or derived correction factors were used to allow for decom- position of 10, 5 and 100/,, respectively, for 22 h hydrolysis.
Cyanogen Bromide Cleavage The procedure used was essentially that described
by Gross and Witkop [I31 and modified by Steers et al. [14]. The dried protein or peptide was dissolved in 70°/ , formic acid to a concentration of I0 mg/ml. A 100-fold molar excess of CNBr over methionine resi- dues was used and the reaction was allowed to pro- ceed a t room temperature for 24 h. The reaction mixture was diluted with 10 volumes water and lyo- philized. The dry material was taken up in 5 volumes water and lyophilized again.
Citraconybtion The amino groups of carboxymethyl-adenylate
kinase or of large peptides were reacted with citra-
Eur. J. Biochem. 43 (1974)

134 Amino- Acid Sequence of Porcine Adenylate Kinase
conic anhydride essentially as described by Gibbons and Perham [15]: (a) for confining the cleavage of the polypeptide chains by trypsin to arginine residues; (b) for unfolding polypeptide chains; (c) for avoiding the intermolecular aggregation of peptides in a mixture and (d) for identifying peptides in column eluates, the citraconyl group being a chromophore which absorbs a t 254nm. In a typical preparation 10 pmol (220 mg) freeze-dried carboxymethyl-aden- ylate kinase was suspended in 50mM N-ethyl- morpholine-acetic acid buffer pH 8.2 to a concentra- tion of 0.5 mM protein, i.e. 10.5 mM &-amino groups. A 30-fold excess (0.58 ml) citraconic anhydride (Eastman Chemical Co.) was added slowly while maintaining the pH 8.0-8.5 by addition of 2 M NaOH. The citraconylation was complete as evidenc- ed by failure to detect free amino groups by the 2,4,6-trinitrobenzene sulfonic acid assay [16,17]. The citraconylated protein was dialyzed against 50 mM N-ethylmorpholine-acetic acid buffer pH 8.2 at 2 "C and lyophilized. Significant loss of citraconyl groups occurred when the pH was lowered to 6 (about 200/, loss in 18 h at 0 "C). Care was taken to recitraconylate the protein or peptides immediately before chromatography or tryptic digestion. However, only 5- to 10-fold excess citraconic anhydride was used in recitraconylation.
Cit'raconylated peptides were unblocked ("de- citraconylated") by dissolving in water to a concen- tration of not over 10 mg/ml, adding formic acid to pH 3.0 and incubating at 60 "C for 2 h, or alternative- ly by dissolving the peptides to a concentration of 1 mM or less in a volatile buffer of pH 3.5, e.g. pyridine-acetic acid-water (1 : 10:89, by vol) and standing for 8 h at room temperature.
kinase obtained by the two different methods were virtually identical. Peptides were digested under similar conditions. 50-200 nmol peptide was dissolv- ed in 300 y10.5°/0 NH,HCO, and 10 y1 of a 1 mg/ml solution of trypsin was added.
Chymotryptic digestions were performed using the same concentrations and ratios as with trypsin. Worthington CDI (prepared from 3 x crystallized zymogen) was used except for peptide CBf for which Worthington CDS ("free of low-molecular-weight contaminants") was used. Digestion times varying from 1.3 to 16 h were used for peptide CBf with the resulting fingerprints being very different for samples from extremes of digestion times.
Recrystallized thermolysin was a gift of W. S&n- ger, Gottingen. Another batch, purchased from Merck (Darmstadt), used without recrystallization to prepare certain peptides from CBf in larger amounts showed a higher enzymatic activity. The digestion conditions were essentially the same as for trypsin (method b), except that the incubation temperature was 55 "C and the incubation period varied. Peptides CRb and CBd were digested for 4 h. For CBf, however, after judging the degree of digestion by the result of hgerprints, an incubation period of 1 h with an enzyme to substrate ratio of I : 50 was chosen in order to limit the number of fragments.
&-Protease from C. atrox venom (No. 20030 from Pierce Chemical Co., Rockford, Illinois, U.S.A.), was used to digest peptide CBf following the method of Pfleiderer [19]. The enzyme to substrate ratio was 1:20.
Subtilisin (Serva, Heidelberg) digestions were performed according to the method b of tryptic digestions.
Enzymatic Hydrolysis Methods of Isolation of Peptides Tryptic digestions were performed in one of two
ways: (a) the protein or peptide was dissolved in water t o a concentration of about 10 mgfml, then placed in a 30 "C water bath with a pH-stat set a t 8.0, The reaction was started by adding lo/, of the weight of protein in trypsin dissolved in 1 mM HCI. Samples of trypsin used were Worthington TRL 100371s treated with L - 1 - tosylamido - 2 - phenylethyl chloromethyl ketone [18] or Worthington TRTPCK. After the NaOR uptake rate had slowed down, usually 10 min, a second aliquot of 1 trypsin was added and then a third. The reaction was allowed to proceed for a total of two hours although it was 95O/, complete within IOmin after the addition of the first aliquot of trypsin. (b) The protein was dissolved in 0.501, NH,HCO, pH8.0 to a concentration of about 1Omglml and the reaction was started by adding trypsin equal to 2-501, of the weight of substrate. The reaction was allowed to proceed for four hours a t 37 "C. Fingerprints of tryptic digests of adenylate
With relatively large amounts of peptides (I0 to 20 pmol) gel filtration on Sephadex G-50 or G-25 was used as the first purification step. Subsequent purifi- cation steps were carried out using high-voltage paper electrophoresis or descending paper chromatog- raphy singly or in combinations. Peptide mixtures were streaked on Whatman no. 3MM paper to a concentration of about I mglcm or one-fifth that amount on Whatman no. 1 paper. Internal fluorescent markers [20] were spotted every 2 or 3 cm along the origin line. Electrophoresis was performed a t 3000 volts (about 60 volts/cm) in a modified Michl appa- ratus 1211. One of the following three systems were used: pH6.5 (pyridine-acetic acid-water, 25: 1~225, by vol, cooled with pyridine- toluene, 2 : 23) ; pH 3.5 (pyridine-acetic acid-water, 1 : 10: 89, cooled with Varsol) or pH 1.9 (formic acid-acetic acid-water, 1 : 4 : 45 ; Varsol as coolant). Visible dyes, xylene cyanol FF and e-dinitrophenyl-lysine or crystal violet, along With a mixture of 17 amino acids,
Eur. J. Biochem. 43 (1974)

A. Heil, G. Miiller, L. Noda, T. Pinder, H. Schirmer, I. Schirmer, and I. von Zabern 135
were spotted on the edges of the sheet and used as reference markers [22J Details of the manipulations were as given by Ryle et al. [23], and by Weeds and Hartley [24].
Descending chromatography was carried out as described by Waley and Watson [25] by use of the following single phase solvent system : butan-1-01- acetic acid-water-pyridine (15:3: 10: 12, by vol). Peptides were located by staining guide strips with the cadmium-ninhydrin reagent of Heilmann et al. 1261 or the hypochlorite-starch-iodide reagent of Rydon and Smith [27]. In appropriate cases the following staining reactions were used: Pauly test for tyrosine and histidine [28], Sakaguchi test €or arginine [29] (excluding the h a 1 spray with iodine) and iodoplatinate test for methionine [30]. Peptides were eluted with 0.02 to 0.1 M NH,OH or 30’1, pyridine or 50°/, acetic acid.
Fingerprints, two-dimensional peptide maps, were carried out as described by Svasti and Milstein [31]. Electrophoresis a t pH 6.5 was performed on the pep- tide mixture using Whatman no. 3MM paper, follow- ed by descending chromatography a t a right angle to the first direction. I n some cases the neutral band was cut out after descending chromatography and rerun in pH 1.9 electrophoresis.
Combined Dansyl-Edmn Xequencing Method Peptides were sequenced by the dansyl-Edman
technique as described by Gray [32,33]. Reagents used for this procedure were phenylisothiocyanate and trifiuoroacetic acid from Merck which were re- distilled before use and dansyl chloride purchased from Sigma Chemical Co. (St. Louis, Mo., U.S.A.). Sequanal grade reagents purchased from Pierce Chemical Co. (Rockford, Illinois, U.S.A.) were used directly for work on peptide CBf. Dansyl-amino acids were identified after chromatography on poly- amide sheets (Schleicher & Schiill, Dassel, Germany or Gallard-Schlesinger Chemical Co., Carle Place, New York, U.S.A.) according to the method of Woods and Wang [34]. The following additional solvents were used in the same direction as solvent 11: ethyl acetate-methanol-acetic acid (20: 1 : 1, by vol) [35] to separate dansyl-threonine from dansyl-serine and dansyl-glutamic acid from dansyl-aspartic acid ; 1.20/, acetic acid-0.64O/, pyridine-25°/, ethanol in water folIowed by 0.05 M Na,PO,-ethanol (3: 1, by vol) [36] to separate the monodansyl derivatives of lysine, histidine and arginine.
Amide Assignments Amide assignments [37] were made by comparing
the electrophoretic mobility of the peptide relative to aspartic acid a t pH 6.5 ( , u A ~ ~ ) before and after removal of the residue in question by the Edman reagents. h i d e assignments could frequently be
made simply on the basis of the mobility of the intact peptide.
Determination of C-Terminal Residues C-Terminal residues were identified by digesting
the peptide with carboxypeptidase A and/or carb- oxypeptidase B. Both enzymes (purchased from Sigma Chemical Co.) had been treated with diiso- propylfluorophosphate and were used according to the procedures of Ambler [38]. Samples of digests in 0.2 M N-ethylmorpholine pH 8.0-8.5 were taken in a time sequence, enzyme reaction was stopped by an appropriate mixture of citrate-HC1 pH 2.2, and the samples were stored in a deep-freeze until analyzed in an amino-acid analyzer.
Detection of the N-Terminal Acetyl Group The N-terminal blocking group was investigated
by hydrazinolysis and subsequent dansylation a t low pH as described by Schmer and Kreil[39]. The method described was scaled down to a sample size of 250 nmol. The reference substances 1-acetyl-2-dansylhydrazine and the corresponding formyl derivative were prepar- ed by dansylation of hydrazineacetate (Pierce) and formylhydrazide (Eastman Chemical Co.) under the same conditions as used for peptide samples. The acyldansylhydrazides were identified by thin-layer chromatography on silica gel 60 sheets (Merck, Darmstadt).
Nomenclature of Peptides Cyanogen bromide cleavage of porcine adenylate
kinase provided the primary fragments for further investigation. Hence, the system of nomenclature is based on these CB-fragments. Specifically they are designated consecutively, starting from the N- terminus of the protein, as fragments CBa to CBf or simply as “a” to “f”. By this convention f indicates cyanogen bromide fragment f. An exception is made for two subfragments of CBf found in low yield; namely, CBfN covering the N-terminal portion and CBfc covering the C-terminal portion of CBf.
Peptides obtained by enzymatic digestions are designated as follows: T, tryptic peptides; T(c), peptides obtained by tryptic digestion after blocking lysine residues by citraconylation ; C, chymotryptic peptides ; Th, thermolytic peptides ; S, subtilisin peptides ; aP, ci-protease peptides.
Secondary digestion products were named accord- ing to their position in the primary fragment, e.g. fT(c)3 indicates the 3rd tryptic peptide from the amino end of citraconylated cyanogen bromide fragment f. When subsequent digestions were per- formed additional designations were added using the same convention, e.g. fT(c)3C1 indicates the N-terminal peptide obtained from a chymotryptic digestion of the above fragment, fT(c)3.
Em. J. Biochem. 43 (1974)

136 Amino-Acid Sequence of Porcine Adenylate Kinase
Table 2. Amino-acid composition of carboxymethylated ade- nylate kinase The values listed in the first column are averages of six analy- ses. Tryptophan was determined after Ba(OH), hydrolysis [40]. Other details of the analyses are presented under Meth- ods. Molecular weight of protein was taken as 21500 [3]
Amino acid Residues Composition inferred from sequence results
Carboxymethyl-cysteine Aspartic acid Asparagine Threonine Serine Glutamic acid Glutamine 1 Proline Glycine Alanine Valine Methionine Isoleucine Leucine Tyrosine Phen ylalanine Lysine Histidine Arginine Tryptophan
num ber/mol protcin
1.7 2 11 2 13.0
13.6 14 10.7 11
19 6 25.2
6.2 6 19.2 19 8.0 8
17.2 17 6.0 6 8.5 9
18.6 18 6.8 7 4.9 5
20.9 21 1.8 2
11.0 11 <0.1 0
No. of residues 194
The designations used for methionine-containing tryptic peptides which were used for overlapping cyanogen bromide fragments contain the letters used to identify the CB peptides which are overlapped. For example, Tab is the overlapping peptide for CBa and CBb.
RESULTS Amino- Acid Composition
The results obtained from analyses of six different preparations of carboxymethylated adenylate kinase are given in Table 2 together with the amino-acid composition of the protein as sequenced. There are no discrepancies between the two sets of data as can be seen in Table 2. Adenylate kinase is composed of 194 amino-acid residues and has a calculated molecu- lar weight of 21700. For this calculation it was taken into account that in the native enzyme the cysteine residues are not carboxymethylated and that the N-terminal amino acid is acetylated. Adenyl- ate kinase consists of a single polypeptide chain since no cystine is present and since there is only one (blocked) amino-terminal residue and only one carboxyl-terminal residue.
Procedure of Sequence Analysis As mentioned in the section on Nomenclature
cleavage of carboxymethylated adenylate b a s e
Iodo[Z-%] acetate
Treatment with CNBr, Tiypsin, then isolation of then citraconic anhydride;
then isolation of t
[CE-fragments with citraconylated lysine residues I 1
Larger citraconylateda I
less th, I 15 resid
Sequen
Trypsin; then isolation of I Removal of the
trypsin; then isolation of
the citraconyl citraconyl groups; Removal of
groups
? analysis Sequence analysis
Removal of the citracon yl groups; chymotrypsin or thermolysin; then isolation I Of
tryptic peptides for overlapping the
Methionine-containing peptides for overlapping the CB-fragments
Fig. 1. Outline of the sequence analysis of adenylate kinase
Eur. J. Biochem. 43 (1974)
A. Heil, G. Muller, L. Noda, T. Pinder, H. Schirmer, I. Schirmer, and I. von Zabern 137
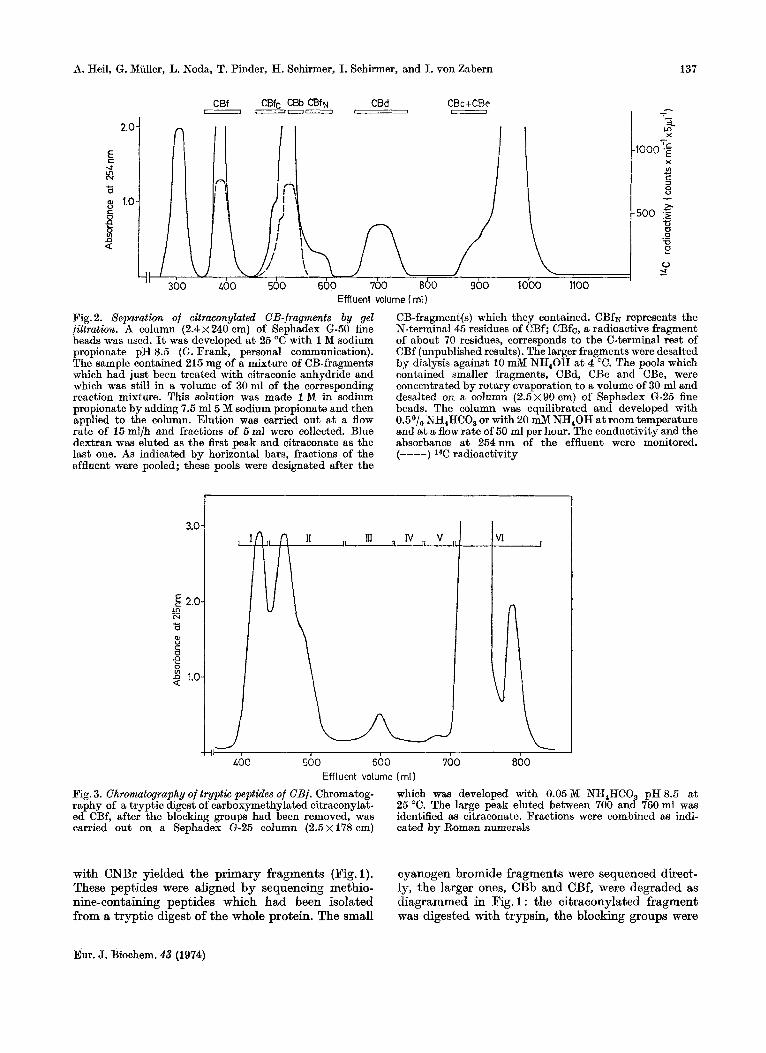
Effluent volume ( ml) Fig. 2. Separation of citraconylated CB-fragments by gel filtration. A column (2 .4~240 om) of Sephadex G-50 fine beads was used. It was developed a t 25 "C with 1 M sodium propionate pH 8.5 (GI. Frank, personal communication). The sample contained 215 mg of a mixture of CB-fragments which had just been treated with citraconic anhydride and which was still in a volume of 30 ml of the corresponding reaction mixture. This solution was made 1 M in sodium propionate by adding 7.5 m15 M sodium propionate and then applied to the column. Elution was carried out a t a flow rate of 15ml/h and fractions of 5ml were collected. Blue dextran was eluted as the first peak and citraconate as the last one. As indicated bv horizontal bars, fractions of the
CB-fragment(s) which they contained. CBfN represents the N-terminal45 residues of CBf; CBfc, a radioactive fragment of about 70 residues, corresponds to the C-terminal rest of CBf (unpublished results). The larger fragments were desalted by dialysis against 10 mM NH,OH a t 4 "C. The pools which contained smaller fragments, CBd, CBc and CBe, were concentrated by rotary evaporation to a volume of 30 ml and desalted on a column (2 .5~90cm) of Sephadex G-25 fine beads. The column was equilibrated and developed with 0.5O/, NH,HCO, or with 20 mM NH,OH a t room temperature and at a flow rate of 50 ml per hour. The conductivity and the absorbance a t 254nm of the effluent were monitored. (----) 14C radioactivitv
effluent were pooled; these pools were designated after the
3.0
g 2.0 Ln
3 8
-0
N
c 0
2 1.0
JI I
:I L
500 600 700 800 11- I
GOO Effluent volume (ml )
Fig. 3. Chromatography of tryptic peptides of CBf. Chromatog- raphy of a tryptic digest of carboxymethylated citraconylat- ed CBf, after the blocking groups had been removed, was carried out on a Sephadex G-25 column (2.5 x 178 cm) cated by Roman numerals
which was developed with 0.05 M NH,HCO, pH 8.5 a t 25 "C. The large peak eluted between 700 and 760 ml was identified as citraconate. Fractions were combined as indi-
with CNBr yielded the primary fragments (Fig. 1). These peptides were aligned by sequencing methio- nine-containing peptides which had been isolated from a tryptic digest of the whole protein. The small
cyanogen bromide fragments were sequenced direct- ly, the larger ones, CBb and CBf, were degraded as diagrammed in Fig. 1 : the citraconylated fragment was digested with trypsin, the blocking groups were
Eur. J. Biochem. 43 (1974)
138 Amino-Acid Sequence of Porcine Adenylate Kinase
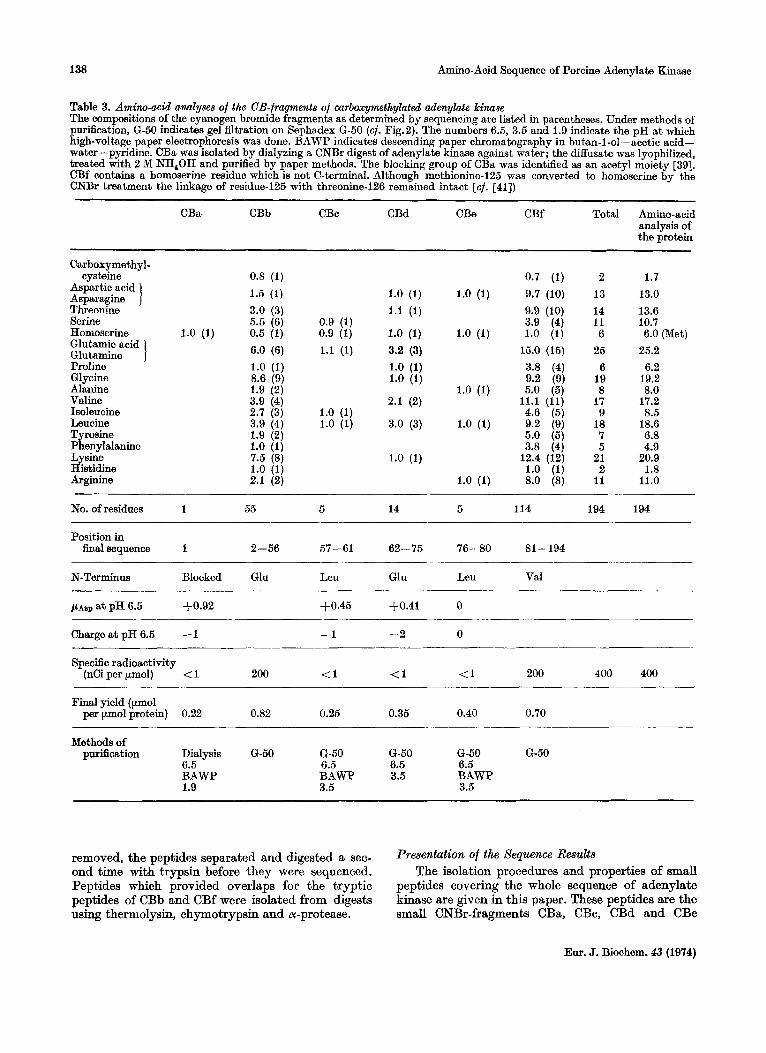
Table 3. Amim-mid analyses of the CB-fragments of carboxymthylated adenylate kilulae The compositions of the cyanogen bromide fragments as determined by sequencing are listed in parentheses. Under methods of purification, G-50 indicates gel filtration on Sephadex G-50 (cf. Fig.2). The numbers 6.5, 3.5 and 1.9 indicate the pH a t which high-voltage paper electrophoresis was done. BAWP indicates descending paper chromatography in butan-1-01-acetic acid- water-pyridine. CBa wm isolated by dialyzing a CNBr digest of adenylate kinase against water; the diffusate was lyophilized, treated with 2 M NH,OH and purified by paper methods. The blocking group of CBa was identified as an acetyl moiety [39]. CBf contains a homoserine residue which is not C-terminal. Although methionine-125 was converted to homoserine by the CNBr treatment the linkage of residue-125 with threonine-126 remained intact [cf. [all)
____ ~~
CBa CBb CBc CBd CBe CBf Total Amino-acid analysis of the protein
Carboxymethyl- cysteine
Aspartic acid Asparagine 1 Threonine Serine Homoserine Glutamic acid Glutamine 1 Proline 6 1 y c i n e Alanine Valine Isoleucine Leucine Tyrosine Phenylalanine Lysine Histidine Arginine
0.8 (1) 1.5 (1) 3.0 (3) 5.5 (6)
1.0 (1) 0.5 (1) 6.0 (6) 1.0 (1) 8.6 (9) 1.9 (2) 3.9 (4)
3.9 (4) 1.9 (2) 1.0 (1) 7.5 (8) 1.0 (1) 2.1 (2)
2.7 (3)
0.7 (1) 1.0 (1) 1.0 (1) 9.7 (10) 1.1 (1) 9.9 (10)
0.9 (1) 3.9 (4) 0.9 (1) 1.0 (1) 1.0 (1) 1.0 (1) 1.1 (1) 3.2 (3) 15.0 (15)
1.0 (1) 3.8 (4) 1.0 (1) 9.2 (9) . ,
1.0 (1) 5.0 (5j 2.1 (2) 11.1 (11) , .
1.0 (1) 4.6 ' (5 j 1.0 (1) 3.0 (3) 1.0 (1) 9.2 (9)
5.0 (51
2 13 14 11 6
25 6
19 8
17 9
18 7 5
21 2
11
1.7 13.0 13.6 10.7 6.0 (Met)
25.2 6.2
19.2 8.0
17.2 8.5
18.6 6.8 4.9
20.9 1.8
11.0
No. of residues 1 55 5 14 5 114 194 194
Position in finalsequence 1 2-56 57-61 62-75 76-80 81 - 194
N-Terminus Blocked Glu Leu Gln Leu Val
, U A S ~ at pH 6.5 3-0.92 +0.45 $0.41 0
Charge a t pH 6.5 - 1 -1 -2 0
Specific radioactivity (nCi per pmol) < 1 200 (1 <1 (1 200 400 400
Final yield (ymol per pmol protein) 0.22 0.82 0.25 0.35 0.40 0.70
Methods of purification Dialysis G-50 G-50 6-50 6-50 6-50
6.5 6.5 6.5 6.5 BAWP BAWP 3.5 BAWP 1.9 3.5 3.5
removed, the peptides separated and digested a sec- ond time with trypsin before they were sequenced. Peptides which provided overlaps for the tryptic peptides of CBb and CBf were isolated from digests using thermolysin, chymotrypsin and or-protease.
I k ~ n t a t i o n Of the Sequence L k d t s The isolation procedures and properties of small
peptides covering the whole sequence of adenylate kinase are given in this paper. These peptides are the small CNBr-fragments CBa, CBc, CBd and CBe
Eur. J. Biochem. 43 (1974)

A. Heil, G. Miiller, L. Noda, T. Pinder, H. Schirmer, I. Schirmer, and I. von Zabern 139
Table 4. Amino-acid analyses of tqptk p p t k l e s of CBb Under methobs of purification, G-50 indicates gel filtration of the tryptic peptides of citraconylated CBb on a column (2.4 x 240 cm) of Sephadex G-50 fine beads which was developed with 20 mM NH,OH at 4 "C. The effluent was pooled in two frac- tions: the first one contained the radioactive peptide bT(c)l, the second one the much smaller peptides bT(c)2 and bT(c)3
Peptides derived from bT(c)l bT(c)2 bT(c)3 Total Amino-
T i T2 T3 T4 T5 T6 T7 T8 analy- acid
sis of CBb
Carboxymethyl- c ysteine
Aspartic acid Asparagine ] Threonine Serine
0.6 1 0.8 1.1 1 1.5
0.9 1.8 3 3.0 1.0 1.1 1.1 3.0 6 5.5
Homoeerine 1.0 1 0.5 2.1 1.0 1.1 6 6.0 Glutamic acid
Glutamine ] 2*2 Proline 1.0 1 1.0 G1 ycine 4.0 1.0 2.4 1.0 1.1 9 8.6 Alanine 2.0 2 1.9 Valine 1.8 0.9 1.1 4 3.9 Methionine 0 0 Isoleucine 1.7 0.9 3 2.7 Leucine 0.9 3.0 4 3.9 Tyrosine 2.0 2 1.9 Phenylalanine 1.0 1 1.0 Lysine 1.0 1.0 1.0 1.0 1.0 1.0 1.0 1.0 8 7.5 Histidine 1 .o 1 1.0 Arginine 1.0 0.9 2 2.1
No. of residues 3 2 1 2 12 6 4 13 9 3 55 55
N-Terminus Glu Leu Lys Ser Ile Gly Ile Tyr Ala Gly
p ~ s p at pH 6.5 +0.50 -0.64 -0.91 -0.64 -0.27 +0.32 -0.40 -0.10 0 -0.62
Methods of 6.5 6.5 6.5 6.5 6.5 6.5 6.5 6.6 G-50 6-50 purification 3.5 BAWP 3.5 BAWP 3.5 3.5 3.5 3.5 6.6 6.5
BAWP BAWP 1.9 BAWP BAWP BAWP BAWP
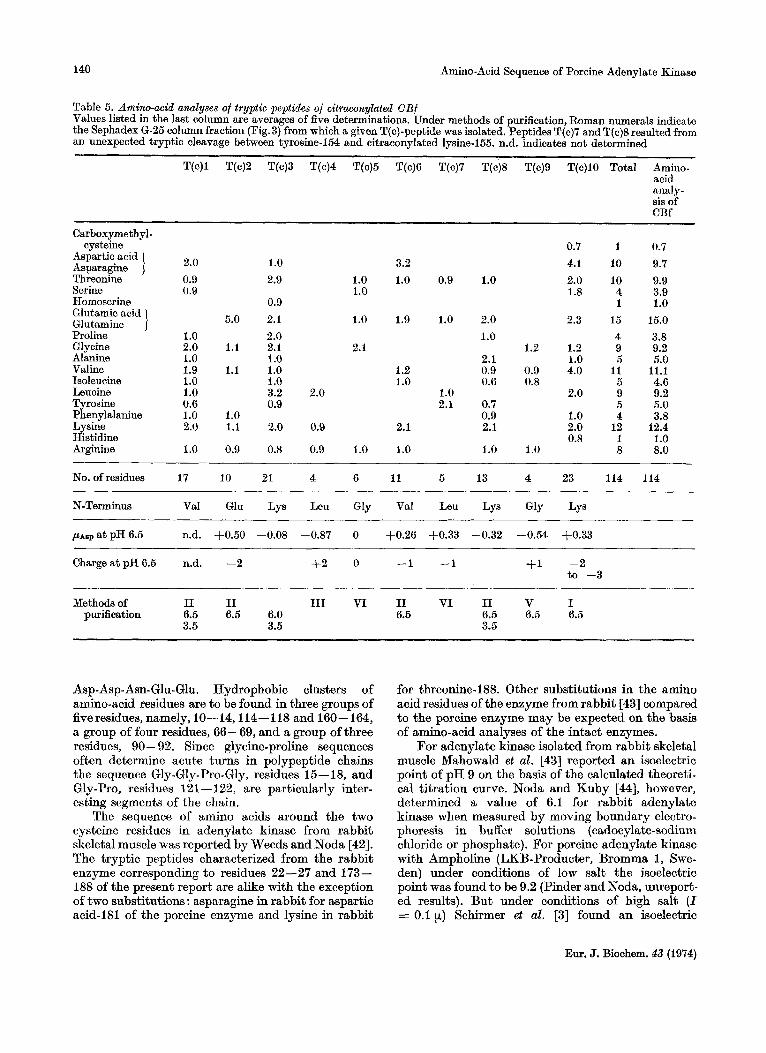
(Fig.2 and Table 3). the tryptic peptides of CBb (Table 4) and the principal tryptic peptides of CBf (Fig.3 and Table 5).
Data on the isolation, amino-acid compositions and sequences of overlapping peptides and furfher details are published in a supplementary paper.
All results from which the sequence of adenylate kinase was deduced are delineated in Fig.4 and 5. The complete primary structure of the protein is given in Fig. 6.
DISCUSSION The primary structure of adenylate b a s e from
porcine skeletal muscle is presented in Fig.6. The sequence of amino-acid residues in this single poly- peptide chain can be confirmed by, and at the same
time aid in, the interpretation of X-ray diffraction studies currently being conducted. The molecule is characterized by the absence of tryptophan, the presence of two cysteines and the absence of cystine, the presence of two histidines, five phenylalanines, seven tyrosines and six each of proline and methio- nine.
Clusters of basic and acidic amino-acid residues may be of sigdicance in maintaining the confor- mation of the molecule or in facilitating interactions with other cellular components. Four lysines inter- spersed with a leucine and a serine are to be found together as residues 4-9. Two lysines and an arginine forming a triplet of basic residues occupy positions 147- 149, and interspersed throughout the sequence are five lysine-arginine pairs. A striking cluster of acidic amino acids is found in the region 140-144,
Eur. J. Biochem. 43 (1974)
140 Amino-Acid Sequence of Porcine Adenylate Kinase
Table 5. Amino-acid analyses of tryptic peptides of citraconylated CBf Values listed in the last column are averages of five determinations. Under methods of purification, Roman numerals indicate the Sephadex 6-25 column fraction (Fig.3) from which a given T(c)-peptide was isolated. Peptides T(c)7 and T(c)8 resulted from an unexpected tryptic cleavage between tyrosine-154 and citraconylated lysine-155. n.d. indicates not determined
T(c)l T(c)2 T(c)3 T(c)4 T(c)5 T(c)6 T(c)7 T(c)8 T(c)9 T(c)lO Total Amino- acid analy- sis of CBf
Carboxyme thy1 - cysteine 0.7 1 0.7
2.0 1.0 3.2 4.1 10 9.7 Aspartic acid Asparagine Threonine 0.9 2.9 1.0 1.0 0.9 1.0 2.0 10 9.9 Swine 0.9 1.0 1.8 4 3.9 Homoserine 0.9 1 1.0
5.0 2.1 1.0 1.9 1.0 2.0 2.3 15 15.0 Glutamic acid Glutamine ] Proline 1 .o 2.0 1.0 4 3.8 Glycine 2.0 1.1 2.1 2.1 1.2 1.2 9 9.2 Alanine 1 .o 1 .o 2.1 1.0 5 5.0 Valine 1.9 1.1 1.0 1.2 0.9 0.9 4.0 11 11.1 Isoleucine 1 .o 1 .o 1.0 0.6 0.8 5 4.6 Leucine 1.0 3.2 2.0 1.0 2.0 9 9.2 Tyrosine 0.6 0.9 2.1 0.7 5 5.0 Phenylalanine 1.0 1.0 0.9 1.0 4 3.8 Lysine 2.0 1.1 2.0 0.9 2.1 2.1 2.0 12 12.4 Histidine 0.8 1 1.0 Arginine 1.0 0.9 0.8 0.9 1.0 1.0 1.0 1.0 8 8.0
No. of residues 17 10 21 4 6 11
N-Terminus Val Glu Lys Leu Gly Val Leu Lys Gly Lys
5 13 4 23 114 114
ASP a t PH 6.5 n.d. +0.50 -0.08 -0.87 0 +0.26 +0.33 -0.32 -0.54 +0.33
Charge a t pH 6.5 n.d. -2 t 2 0 -1 -1 +I -2
Methods of I1 I1 I11 VI I1 VI I1 V I
to -3
purification 6.5 6.5 6.0 6.5 6.5 6.5 6.5 3.5 3.5 3.5
Asp-Asp-Asn-Glu-Glu. Hydrophobic clusters of amino-acid residues are to be found in three groups of fiveresidues, namely, 10--14,114-118 and 160-164, a group of four residues, 66-69, and a group of three residues, 90 -92. Since glycine-proline sequences often determine acute turns in polypeptide chains the sequence Gly-Gly-Pro-Gly, residues 15-18, and Gly-Pro, residues 121 - 122, are particularly inter- esting segments of the chain.
The sequence of amino acids around the two cysteine residues in adenylate kinase from rabbit skeletal muscle was reported by Weeds and Noda [42]. The tryptic peptides characterized from the rabbit enzyme corresponding to residues 22-27 and 173- 188 of the present report are alike with the exception of two substitutions : asparagine in rabbit for aspartic acid-I81 of the porcine enzyme and lysine in rabbit
for threonine-188. Other substitutions in the amino acid residues of the enzyme from rabbit [43] compared to the porcine enzyme may be expected on the basis of amino-acid analyses of the intact enzymes.
For adenylate kinase isolated from rabbit skeletal muscle Mahowald et al. [43] reported an isoelectric point of pH 9 on the basis of the calculated theoreti- cal titration curve. Noda and Kuby [44], however, determined a value of 6.1 for rabbit adenylate kinase when measured by moving boundary electro- phoresis in buffer solutions (cadocylate-sodium chloride or phosphate). For porcine adenylate kinase with Ampholine (LKB-Producter, Bromma 1, Swe- den) under conditions of low salt the isoelectric point was found to be 9.2 (Pinder and Noda, unreport- ed results). But under conditions of high salt ( I = 0.1 p) Schirmer et al. [3] found an isoelectric
Em. J. Biochem. 43 (1974)

A. Heil, G. Muller, L. Noda, T. Pinder, H. Schirmer, I. Schirmer, and I. von Zabern 141
10 2 0 A c - Hse Glu-Glu-Lys- Leu- Lys-Lys- S e r - L y s - I l e - I le -Phe-Va l - Val -Gly -Gly-Pro-Gly-Ser- Gly-Lys-
CBa CBb Hb-, --------- ---------
---Ah+Htl--ib777--------+ ++ --------
T2 T 3 T 4 T5
bC 1 bC2 & Tab I----+ cl-7 1-7 -7 -7 17 1
30 40 - Gly -Thr -Gln- CMCys - G l u - Lys - 1 l e - Val- Gln-Lys - Tyr -G ly -Tyr -Thr-His- Leu- Ser-Thr- Gly-Asp- Leu-
CBb
b T ( c ) l
T6 A F T 7 T 8 l b c 2 / 7 - 7 - 7 7 + 7 7 7 7 + 7 7 7 7 7 - - - - - T -
I
bThl bTh2 bTh4 bTh5 7----+--177-7/t777-7+&77777
+---- ---I bTh3 +--+
50 60 -Leu-Arg -A la -Glu -Val - Ser-Ser -Gly- Ser -A la -Arg- Gly -Lys- lise Leu- Ser- G lu- I le - Hse Glu- Lys-
CBb CBc CBd
70 80 - GI y - Gln- Leu - Val - Pro - Leu- Glu - Thr-Val -Leu - Asp-Hse Leu - Arg- Asp- Ala - Hse Va I - Ala- L ys
CBd CBf +
+ 7 7 7 - - 7 7 7 - - 7 7 - - 7 7 T - -
7 7 4 l - - 7 7 - - - + k - - - dTh l dTh2 dTh3
,Tde Tef
Fig.4. Summary of the sequence data on cyanogen bromide fragments CBa- CBe. The supporting data are given for the sequence determination of residues 1-83. Residues 2 -36, the N-terminal portion of CBb, were confirmed by automatic sequence analysis (Wittmann-Liebold, personal communica- tion). In peptides which overlap CB-fragments methionine is present instead of homoserine. A y indicates that the amide assignment was made by comparing the electrophoretic
point of about 6.0 for porcine adenylate kinase. On the basis of excess basic groups (21 Lys + 11 Arg + 2 His - 11 Asp - 19 Glu - 1 terminal carboxyl group) the isoelectric point would be expected to be at an alkaline pH if one assumes that all basic and acidic side chains are in an aqueous environment.
mobilities a t pH 6.5 before and after the marked residue was removed. (-) The residue was present in the amino-acid analysis of the peptide delineated by vertical lines. Vertical lines connecting peptides indicate that a further digestion was performed giving rise to the lower set of peptides; (-) the residue was determined by the dansyl-Edman technique and (-) the residue was determined by carboxy- peptidase digestion
The low isoelectric points observed might be explain- ed by binding of buffer anions such as phosphate which would be favored at higher ionic strength.
The goal of elucidating the catalytic mechanism of a so-called “high-energy’’ phosphate-transferring enzyme seems closer to attainment as data on adenyl-
Eur. J. Biochem. 43 (1974)

I r r r
m r $%
f -c c Lc
I $1 I?, T-El I I T T
.-r I
I t r I I ~ I i:r I r:r i r i r
Amino-Acid Sequence of Poroine Adenylate Kinase
Eur. J. Biochem. 43 (1974)

A. Heil, G. Muller, L. Noda, T. Pinder, H. Schirmer, I. Schirmer, and I. von Zabern 143
1 2 3 4 5 6 7 8 9 10 11 12, 13 14 15 16 17 16 19 20 Ac - Met-Glu - Glu - Lys-Leu - Lys-Lys -Ser-Lys-lle -l ie - Phe -Val - Val- Gly-Gly-Pro -Gly-Ser- Gly-
2l 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 Lys-Gly -Thr-Gln - Cys - Glu - Lys-Ile-Val- Gln - Lys -Tyr -Gly-Tyr - Thr - His - Leu-Ser - Thr - Gly-
41 12 43 44 45 16 47 L8 49 50 51 52 53 54 55 56 57 58 59 60 Asp-Leu - Leu-Arg- Ala-Glu - Val-Ser-Ser- Gly -Ser-A[a-Arg- Gly-Lys- Met- Leu-Ser-Glu-Ile -
61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 Met-Glu-Lys-Gly-Gin- Leu- C/a(-pro-Leu-G/u-Tbr-l/o/-Leu-Asp-Met-L~u-A~-As~-Al~ - Mef-
81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 Val -Ah- Lys-Val -Asp-Thr-Ser - Lys-Gly +he-Leu-lle -Asp-Gly -Tyr- Pro - Arg - Glu -Val - Lys - 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 Gln - Gly -Glu - Glu - Phe -Glu -A rg - Lys-Ile-Gly-Gln -Pro - Thr - Leu - Leu -Leu - Tyr -Val -Asp -A la -
121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 G(y - Pro - Glu -Thr-Met- Thr- Lys -Arg-Leu-Leu-Lys - Arg -G/y-Glu- Thr-Ser- Gly-Atg -Val-Asp-
141 11.2 143 144 145 146 147 148 149 150 151 152 153 154 155 156 157 158 159 160 Asp -Am- Glu - Glu - Thr -Ile - Lys-lys-Arg- Leu -Glu -Thr-Tyr - Tyr - Lys-Ala - Thr-Glu - Pro -Val-
161 162 163 164 165 166 167 168 169 170 171 172 173 171 175 176 177 178 179 180 I le -Ala - Phe-vr- Glu - Lys-Arg-Gly-/le -Val -Arg- Lys - Val-Asn -Ala -Glu - Gly -Ser-Vul- Asp - 181 182 183 184 185 186 187 188 189 190 191 192 193 194
Asp -Val - Phe -Ser-Gln - Val - Cys -Thr -His- Leu -Asp - Thr- Leu - Lys
Fig. 6. The amino-acid sequence of porcine adenylate kinase from skeletal muscle
ate b a s e accumulate. X-Ray diffraction studies being carried out in Heidelberg by G. E. Schulz and his colleagues [2], nuclear magnetic resonance studies by M. Cohn and her colleagues [45] and chemical studies will lead to a better understanding of the dynamic nature of phosphoryl transfer. Reactive groups identified include: thiols which facilitate but do not appear to be essential for enzymatic activity [46] ; imidazoles at least one of which is essential [45, 471; thioethers of methionine one or two of which may be important for catalysis or for maintaining the enzyme conformation [48] and an &-amino group (N. C. Price, unreported results) which is at least near the active site.
We are indebted to Dr B. Wittmann-Liebold (Berlin) who confirmed the N-terminal portions of the large cyanogen bromide fragments by automatic sequence analpis.
Annex The following document has been deposited at the
Archives originales du Centre de Documentation du C.N.R.S., F-75971 Paris-Cedex-20, France where it may be ordered as microfiche or photocopies. Reference No.: A. 0. - 630.
Supplemental Material to the Amino-Acid Sequence of Porcine Adenylate Kinase from Skeletal Muscle (with six tables and one figure).
REFERENCES 1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11. 12.
13.
14.
15.
16. 17.
Noda,L. (1973) in The Enzymes (Boyer,P.D., ed.) vol. 8 Part A, pp. 279-305, Academic Press, New York and London.
Schulz, G. E., Biedermann, K., Kabsch, W. & Schirmer, R. H. (1973) J . MoZ. Bi01. 80. 857-864.
Schirmer; I., Schirmer, R. H., Schulz, G. E. & Thuma, E.
Gornall. A. G.. Bardawill. C. J. & David. M. M. f1949b (1970) FEBB Lett. 10, 333-338.
- , J . Biol. Chem. 177, 751-766.
Biol. Chem. 240, 2468-2477.
R. (1969) Biochem. J. 112, 679-689.
Craven, G. R., Steers, E. & Anfinsen, C. B. (1965) J.
Butler, P. J. G., Harris, J. I., Hartley, B. S. & Leberman,
Shotton, D. M. & Hartley, B. S. (1973) Biochem. J .
Spackman, D. H., Stein, W. H. & Moore, S. (1958) Anal. Chem. 30, 1190-1206.
Sanger, F. & Thompson, E. 0. P. (1963) Biochim. Bio- phys. Acta, 71, 468-471.
Tang, J. t Hartley, B. 5. (1967) Biochem. J. 102, 593-599.
Ambler, R. P. (1965) Biochem. J . 96, 32P. Schroeder, W. A., Shelton, J. R. & Robberson, B. (1967)
Biochim. Biophys. Acta, 147, 590-592. Gross, E. & Witkop, B. (1962) J . Biol. Chem. 237,
Steers, E., Craven, G. R., Anfinsen, C. B. & Bethune,
Gibbons,I. & Perham,R.N. (1970) Bwchem. J . 116,
Habeeb, A. F. S. A. (1966) Anal. Biochem. 14, 328-336. Fields, R. (1971) Biochem. J . 124, 581-590.
131, 643-675.
1856 - 1860.
J. L. (1965) J . Biol. Chem. 240, 2478-2484.
843-849.
Eur. J. Biochem. 43 (1974)

144 A. Heil et al. : Amino-Acid Sequence of Porcine Adenylate Kinase
18. Wang, S. & Carpenter, F. H. (1965) J. Biol. Chem. 240,
19. Mella, K., Volz, M. & Pfleiderer, G. (1967) Anal. Bio-
20. Brown, J. R. & Hartley, B. S. (1966) Biochem. J. 101,
21. Michl, H. (1951) Monatsh. Chem. verw. Teile anderer
22. Stevenson, K. J. (1971) Anal. Biochem. 40, 29-34. 23. Ryle, A. P., Sanger, F., Smith, L. F. & Kitai, R. (i955)
24. Weeds,A. G. & Hartley, B. S. (1968) Biochem. J. 107,
25. Waley, S. G. & Watson, J. (1953) Biochem. J. 55,
26. Heilmann, J., Barollier, J. & Watzke, E. (1957) Hoppe-
27. Rydon, H. N. & Smith, P. W. G. (1952) Nature (Lond.)
28. Offord, R. E. (1969) in Data for Biochemical Research, 2nd Edition (Dawson, R. M. C . , Elliott, D. C. , Elliott, W. H. & Jones, K. M., eds) p. 530, Oxford University Press, London.
29. Irreverre, F. (1965) Biochim. Biophys. Acta, 111, 551 - 552.
30. Njaa, L. R. (1963) Acta Chem. S c a d . 17, 1169-1170. 31. Svasti, J. & Milstein, C. (1972) Biochem. J . 128, 427-
32. Gray, W. R. (1972a) Methods Enzymol. 25, 121-138.
1619-1625.
chem. 21, 219-226.
214-228.
W ~ S S . 82,489-493.
Biochem. J. 60, 541 -556.
531 -548.
328-337.
Seyler’s 2. Physiol. Chem. 309, 219-220.
169, 922-923.
444.
33. Gray, W. R. (1972b) Methods Enzymol. 25, 333-344. 34. Woods, K. R. & Wang, K. T. (1967) Biochim. Biophys.
35. Croshaw, K., Kessup, S. J. & Ramwell, P. W. (1967)
36. Hartley, B. S. (1970) Biochem. J. 119, 805-822. 37. Offord, R. E. (1966) Nature (Lond.) 211, 591-593, 38. Ambler, R. P. (1972) Methods Enzymol. 25, 143-154. 39. Schmer, G. & Kreil, G. (1969) Anal. Biochem. 29, 186-
40. Noltmann, E.A., Mahowald, T. A. & Kuby, S. A. (1962)
41. Schroeder, W. A., Shelton, J. B. & Shelton, J. R. (1969)
42. Weeds, A. G. & Noda, L. (1968) Biochem. J. 107, 311-
43. Mahowald, T. A., Noltmann, E. A. & Kuby, S. A. (1962)
44. Noda, L. & Kuby, S. A. (1957) J. Biol. Chem. 226,
45. Cohn,M., Leigh, J. S. t Reed, G. H. (1972) Cold 8pring Harbor Symp. Quant. Biol. 36,533-540.
46. Kress, L. F., Bono, V. H. & Noda, L. (1966) J. Bio2. Chem. 241, 2293-2300.
47. Schirmer, R. H., Schirmer, I. & Noda, L. (1970) Bio- chim. Biophys. Acta, 207, 165-177.
48. Kress, L. F. & Noda, L. (1967) J . Biol. Chem. 242,
A d a , 133, 369-370.
Biochem. J . 103, 79-85.
192.
J. Biol. Chem. 237, 1146-1154.
Arch. Biochem. Biophys. 130, 551 -555.
312.
J. Biol. Chem. 237, 1138-1145.
551-558.
558-564.
A. Heil, G. Miiller, H. Schirmer, I. Schirmer, and I. von Zabern, Max-Planck-Institut fiir Medizinische Forschung, D-6900 Heidelberg 1, JahnstraBe 29, Federal Republic of Germany
L. Noda and T. Pinder, Department of Biochemistry, Dartmouth Medical School, Hanover, New Hampshire, U.S.A. 03755
Eur. J. Biochem. 43 (1974)