research article leaf anatomy and photochemical behaviour
TRANSCRIPT

Research ArticleLeaf Anatomy and Photochemical Behaviour of Solanumlycopersicum L Plants from Seeds Irradiated with Low-LETIonising Radiation
V De Micco1 R Paradiso1 G Aronne1 S De Pascale1 M Quarto2 and C Arena3
1 Department of Agricultural and Food Sciences University of Naples Federico II via Universita 100 80055 Portici Italy2 Department of Physics University of Naples Federico II Via Cinthia 4-80126 Naples Italy3 Department of Biology University of Naples Federico II Via Cinthia 4-80126 Naples Italy
Correspondence should be addressed to V De Micco demiccouninait
Received 5 February 2014 Accepted 17 March 2014 Published 23 April 2014
Academic Editor Charles Hocart
Copyright copy 2014 V De Micco et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Plants can be exposed to ionising radiation not only in Space but also on Earth due to specific technological applications or afternuclear disasters The response of plants to ionising radiation depends on radiation qualityquantity andor plant characteristicsIn this paper we analyse some growth traits leaf anatomy and ecophysiological features of plants of Solanum lycopersicum LldquoMicrotomrdquo grown from seeds irradiated with increasing doses of X-rays (03 10 20 50 and 100 Gy) Both juvenile and compoundleaves fromplants developed from irradiated and control seeds were analysed through light and epifluorescencemicroscopy Digitalimage analysis allowed quantifying anatomical parameters to detect the occurrence of signs of structural damage Fluorescenceparameters and total photosynthetic pigment content were analysed to evaluate the functioning of the photosynthetic machineryRadiation did not affect percentage and rate of seed germination Plants from irradiated seeds accomplished the crop cycleand showed a more compact habitus Dose-depended tendencies of variations occurred in phenolic content while other leafanatomical parameters did not show distinct trends after irradiationThe sporadic perturbations of leaf structure observed duringthe vegetative phase after high levels of radiation were not so severe as to induce any significant alterations in photosyntheticefficiency
1 Introduction
The effect of ionising radiation on plants is studied withindifferent frameworks On one hand high levels of radiationare an undesirable factor affecting plant growth in Spaceor on Earth in case of nuclear disasters such as those ofChernobyl and Fukushima [1 2] On the other hand ionisingradiation is applied as a tool to induce a wide spectrum ofmutations in breeding programs or as means for microbialdecontamination [3 4] Although focusing on different typesof radiation and on different biological endpoints mostexperiments aiming to highlight the response of plants toionising radiation have a common aspect they analyse theeffect of a single type of radiation with wide range of dosesin order to build dose-response curves and to test plant
radioresistance [2] Generally the response to low doses (ieup to 10Gy) is interesting for Space-related issues the use ofincreasingly higher doses is important not only for their roleas positive controls (plant reaction beingmore likely expectedat high doses) but also to generate information useful forterrestrial applications of radioecology [2 5] Focusing onSpace exploration there is common agreement that long-term human permanence in Space relies on the possibility toregenerate resources in Bioregenerative Life Support Systems(BLSSs) where plants can play a key role [6 7] Plant growthis not prevented in Space however although plants canbe more resistant than other organisms to specific Spacefactors (including reduced gravity and cosmic radiation)there is also evidence of altered growth and reduced fitness[2 8 9] Many studies have indicated the occurrence of
Hindawi Publishing Corporatione Scientific World JournalVolume 2014 Article ID 428141 13 pageshttpdxdoiorg1011552014428141
2 The Scientific World Journal
either positive or negative phenomena in plants exposedto low-LET (linear energy transfer) (eg X- and gamma-rays) and high-LET (eg protons and heavy ions) ionisingradiation [2] Definitely in many experiments carried out inSpace plants were exposed to various combinations of bothaltered gravity and radiation Ground-based research helpsdistinguish between plant reactions triggered by each oneof the two factors On Earth the simulation of the complexSpace radiation spectrum including the radiation emittedfrom the Sun and galactic cosmic rays is impossible eventhough single particles can be produced at large particleaccelerators [10] The exposure likely to be encountered byorganisms in Space depends on the shielding features of thespace vehicles andor planetary platforms as well as on theevolution of the solar cycle during the permanence in Spacehowever it can be estimated that during a relatively quietsolar cycle a permanence of one year in Space would exposeorganisms to a dose of less than 10Gy [10ndash12] In Space a largefraction of the dose is delivered by protons (approximately87) which are low-LET particles with similar effectivenesscompared to X-rays Therefore initial studies can exploitconventional X-rays as the reference radiation to obtaindata which are the necessary basis for future comparisonusing accelerated ion beams [13 14] The severity of theeffects of low- and high-LET ionising radiation dependson several factors related to the radiation itself (eg typetotal dose and dose rate) and to plant features (eg speciescultivar plant age complexity of the target organ or tissueand level of ploidy) [2 15] An interesting phenomenonin plants known as hormesis is the occurrence of positiveresponses to low doses of ionising radiation which wouldstimulate processes such as germination and growth [16]A recent paper by Marcu et al [17] showed enhancedgermination plant growth and synthesis of photosyntheticpigments when seeds were irradiated with low doses ofgamma rays but opposite responses when doses were highSparsely and densely ionising radiation can also determinemutations inducing favourable agronomical traits such asearlier maturity higher yields and resistance to diseases [1819] On the other hand there is evidence of altered geneexpression due to plantsrsquo exposure to ionising radiation lead-ing to detrimental consequences culminating in plant death[20]
The very prolific literature on radiation-induced geneticaberrations andmetabolic alterations (especially linked to theROSmdashreactive oxygen speciesmdashproduction) is accompaniedby scarce information at the structural level [2] Giventhat the physics of plant structure ultimately regulate majormetabolic and physiological processes [21] the developmentof well-structured above-ground organs is needed to obtainefficient photosynthesis The photosynthetic process can beconsidered one of themain functions accomplished by higherplants in plant-based modules of BLSSs for air regenerationThus knowing how ionising radiation affectsmorphogenesisand even more leaf development is interesting to evaluatepossible impairment in photosynthesis which would con-strain the maximisation of resource efficiency in the BLSSsIndeed the optimisation of resource use in the BLSSs isconsidered a challenge in Space exploration [6 22ndash24]
Space factors can directly alter the inner structure ofplants or impair photosynthesis due to the alteration of gasexchange or of functioning of the photosystems Indeed thelack of convective forces due to microgravity can alter theavailability of gases at the carboxylation sites [25] On theother hand high levels of ionising radiation can negativelyaffect pigment-protein complexes and enzymes responsiblefor light absorption electron transport and carbon reductioncycle [2] Moreover high radiation levels may also inducephotoprotection mechanisms and affect the behaviour of theD1 protein in the PSII repair cycle [26 27] Photoprotectionmechanisms can rely on physiological adjustments [14 28]andor on the intensification of structural and biochemicalbarriers such as trichomes and phenolic compounds acting asnatural screens against radiation [29] Indeed the increasedcontent of phenolic compounds linked to chloroplast mem-branes has been shown in bean leaves irradiated with highdoses of X-rays [30]
The main aim of this paper is to analyse whether X-ray irradiation of Solanum lycopersicum L ldquoMicrotomrdquo atthe seed stage affects leaf development and anatomy fluo-rescence parameters and pigment content This study hasbeen conducted within a wider experimentation aiming toanalyse the effect of irradiation performed at seed stage onvarious growth processes of dwarf tomato In this paperwe focus on the analysis of leaf morphofunctional traits intwo leaf types In particular we analyse leaves maintainingjuvenile traits (ie the first two true leaves characterisedby simpler morphology than successive leaves) and adultcompound leaves in order to evaluate whether similarstructures characterised by different complexity and ageshow differential responses Indeed it is recognised that theincrease in morphological complexity confers more efficientbuffering capacity to biological systems for dealing withradiation-induced damage [31] An X-ray source at 250 kVwas used to irradiate the seeds because X-rays constitutethe reference radiation to assess the damage caused by anyother radiation source at the same dose The dwarf cultivarMicrotomwas selected as plant material because it is a modelsystem widely used for research in molecular biology [32ndash34] Moreover its compact habit short life cycle and fruitdevelopment not requiring hand pollination are desirablefeatures in BLSSs and would make this cultivar a modelsystem also for research in plant Space biology
2 Materials and Methods
21 Plant Material and Irradiation Procedure The experi-ment was conducted using seeds of Solanum lycopersicum LldquoMicrotomrdquo provided by Holland Online Vof (AmsterdamThe Netherlands) Seed irradiation was performed on April2013 Dry seeds were placed into Petri dishes in one layerand irradiated with five doses of X-rays (03 Gy 10Gy 20Gy50Gy and 100Gy) 250 kVp at dose rate of 1 Gymin Aset of three Petri dishes with 15 seeds each was used foreach irradiation treatment and for a nonirradiated controlDoses up to 20Gy were chosen in order to build a reference-response range to explore plant sensitivity as generally set in
The Scientific World Journal 3
experiments to evaluate the effect of radiation on biologicalsystems [13 14 28 30] The highest doses (50Gy and 100Gy)are generally applied since they can be considered as positivecontrols as there ismore likely a plant reaction at these dosesX-rays were delivered as one dose per each Petri dish [35] inorder to avoid cumulative effects on the same sample X-rayswere produced by a Thomson tube (TR 300 F 250 kVp Sta-bilipan Siemens Forchheim Germany) with tungsten cath-ode filtered by 1mm thick copper foil and with 15mA anodiccurrent Before irradiation a physical dosimetry was per-formed by using an ionization chamber (VictoreenModlingAustria) The X-ray intensity was measured at the estab-lished distance where seeds were positioned under the X-raytube
After the exposure to X-rays (on 24 April) the irradiatedseeds and controls were placed into Petri dishes on three lay-ers of filter paper moistened with distilled water Petri disheswere incubated in the dark at temperature of 20∘C andmoni-tored daily to analyse germination percentage and rate (con-sidered as the number of days taken to reach the maximumgermination percentage) Seeds were classified as germinatedwhen the emerging root grew as long as the seed maximumdiameter
22 Growth Conditions The growth experiment was carriedout in a greenhouse with a polyethylene film cover Thegreenhouse was equipped with a black plastic shading net(70 shading) in order to keep the temperature and lightintensity close to the levels feasible in test bed evaluationof BLSSs technologies [22 23] Wavelength composition ofthe sunlight including the whole PAR range spectrum forphotosynthetic activity was not affected
Seedlings were transplanted into pots (4 cm diameter) onpeat-based compost (peat soil 1 1 in volume) and trans-ferred to the greenhouse 5 days after sowing (DAS) Thenthey were repotted in 10 cm pots using the same substrateat the stadium of fourth true leaf (ie two juvenile and twocompound leaves) at 22DAS In the cultivar Microtom thefirst two true leaves maintain juvenile traits and are charac-terised by simple morphology they are trifoliate with littleor not evident lobes Successive leaves show more complexmorphology having pennate-compound lamina with highlyindented lobes
During the whole growth period plants were irrigatedwith tapwater at 2-day interval in order to reach the containercapacity (till the beginning of drainage)
The temperature inside the greenhouse was recordedevery 10 minutes by means of a data logger (Tynitag Ultra2Gemini Data Loggers Chichester UK) Photosynthetic pho-ton flux density (PPFD) at the canopy level was recordeddaily at 1200 pm with a Delta OHMDO9847multifunctionmeter
The whole crop cycle (up to fruit ripening) lasted from 24April until 6 August 2013
During the whole growth cycle the mean values of tem-perature inside the greenhouse were 284216∘C (daynight)and the light intensity recorded at noon at the canopy levelwas 446 120583mol mminus2 sminus1 on average
23 Biometric and Microscopy Analyses Biometric analyseswere performed once a week on 10 plants per treatmentthroughout the crop cycle Plant height and number of leaveswere recorded Moreover single leaves of the main stem werephotographed with a digital camera (Nikon D3100 NikonEurope BV) in order to examine leaf area increments withthe software program AnalySIS 32 (Olympus HamburgGermany)
From 5 plants per treatment the second juvenile leaf(JL) and the first compound leaf (CL) were collected whenthey were fully developed One median leaflet (one of thetwo opposite leaflets below the apex) per leaf was used formicroscopy analyses while the others were used for pigmentextraction
Leaflets destined to microscopy analyses were immedi-ately fixed in FAA (40 formaldehydeglacial acetic acid50ethanol-5590 by volume) for several days Leaflets werecut under a dissection microscope (ZSX9 Olympus) toobtain subsamples of 5 times 5mm in the middle of laminaincluding the main vein Subsamples were dehydrated inan ethanol series and embedded in the acrylic resin JB4(Polysciences Warrington PA USA) Semithin cross sec-tions (5 120583m thick) were cut through a rotative microtomeSections were stained with 0025 Toluidine blue in 01Mcitrate buffer at pH 4 [36] mounted with Canadian Balsamand observed under a light microscope (BX60 Olympus)Unstained sections were mounted with mineral oil for fluo-rescence and observed under an epifluorescence microscope(BX60 Olympus) equipped with a mercury lamp band-passfilter 330ndash385 nm dichromatic mirror 400 nm and aboveand barrier filter 420 nm and above With these filters it waspossible to detect the presence of simple phenolics that areautofluorescent at such wavelengths [37 38] Three sectionsper leaflet were analysed and images were collected by meansof a digital camera (CAMEDIA C4040 Olympus) at variousmagnifications
24 Digital Image Analysis All digital images were analysedwith the AnalySIS 32 (Olympus) software for image analysisThe thickness of the palisade and the spongy parenchymaswere measured in five regions along the leaf lamina Thecell area and shape of upper and lower epidermis palisadeand spongy parenchyma were quantified in 15 cells pereach tissue per section More specifically cell shape wascharacterised as (a) aspect ratio (maximum widthheightratio of a bounding rectangle for the cell defining how itis elongated) (b) sphericity (roundness of a particle withspherical particles having a maximum value of 1) and (c)convexity (the fraction of the cellrsquos surface area and thearea of its convex a turgid cell has a maximum value of1) [39 40] The cell area occupied by phenolic compoundswas measured in 3 regions (150 times 200 120583m2 each) of themesophyll per section selected avoiding veins Thus thearea occupied by phenolic compounds was calculated asthe percentage of tissuepicture occupied by compoundsappearing autofluorescent at the above-reported filter settings[41]
4 The Scientific World Journal
C 100 Gy50 Gy20 Gy10 Gy03 Gy
Figure 1 Plants of S lycopersicum L ldquoMicrotomrdquo developed fromcontrol and irradiated seeds at 40 DAS (Days After Sowing) Plantsfrom seeds exposed to increasing doses of X-rays (from 0 to 100Gy)are shown from left to right
25 Fluorescence Measurements and Pigment Content Flu-orescence measurements and pigment extraction were con-ducted on 5 pennate-compound leaves of 5 plants subjectedto each irradiation treatment and in controls at 40 DAS
Chlorophyll a fluorescence measurements were carriedout by means of a pulse amplitude modulated fluorometer(Junior-PAM Walz Germany) equipped with a monitoringLeaf-Clip JUNIOR-B (Walz Germany) On 30min dark-adapted leaves the background fluorescence signal Fo wasinduced by internal light provided by a blue LED of about2-3 120583mol photonssdotmminus2sdotsminus1 at a frequency of 05 kHz Themaximal fluorescence level in the dark-adapted state (Fm)was measured by 1 s saturating light pulse (10 000120583molphotonssdotmminus2sdotsminus1) at a frequency of 10 kHz the maximal PSIIphotochemical efficiency (FvFm) was calculated as FvFm =(FmminusFo)FmThemeasurements in the light were carried outby exposing each leaf to a PPFD of 420120583mol photonssdotmminus2sdotsminus1for 5min This level of PPFD was chosen because it fallsin the range of maximal quantum yield for the Microtomcultivar as indicated by fluorescence fast-response curves tolight (data not shown) The steady-state fluorescence signal(Ft) and the maximal fluorescence (Fm1015840) under illuminationwere measured setting the light measure at a frequency of10 kHz Fm1015840 was determined by a 1 s saturating light pulse(10 000120583mol photonssdotmminus2sdotsminus1)
The quantum yield of the PSII electron transport (ΦPSII)and nonphotochemical quenching (NPQ) were expressedaccording to Genty et al [42] and Bilger and Bjorkman [43]
After fluorescence determinations leaves were collectedfrom plants for the photosynthetic pigment content determi-nation namely chlorophylls and carotenoids Pigments wereextracted with a mortar and pestle in ice-cold 100 acetoneand quantified by a spectrophotometer (UV-VIS Cary 100Agilent Technologies USA) according to Lichtenthaler [44]
26 Statistical Analysis All results were subjected to statis-tical analysis using the SPSS statistical package (SPSS IncChicago IL USA) For the time course of plant height num-ber of leaves and leaf area interpolation equations (second-order polynomial curve) were calculated For anatomy andecophysiology data were subjected to ANOVA multiplecomparison tests were performed with Student-Newman-Keuls and Duncan coefficients using 119875 lt 005 as the level of
probability Data on sphericity convexity and percentage ofphenolics were transformed through arcsine function beforestatistical analysis
3 Results
31 Effect of Ionizing Radiation on Seed Germination Expo-sure of dry seeds to X-rays did not affect their percentageof germination which ranged between 978 and 100 withno significant differences among treatments Similarly X-raysdid not determine significant alterations in germination rateswhich ranged between 50 and 67 days
32 Effect of Ionizing Radiation on Plant Growth Plants fromboth control and irradiated seeds completed their crop cycleup to fruit ripening in about 100 DAS plants from irradiatedseeds did not show any apparent alteration in above-groundgrowth (Figure 1)
Figure 2 shows the effects of the seed irradiation treat-ments on plant height leaf number and plant leaf areaof the main stem The irradiation dose differently affectedthese growth parameters In plants of the nonirradiatedcontrol plant height reached the maximum value of 602 cmat approximately 71 DAS Irradiation of seeds with 03Gydid not significantly influence plant height Starting from71 DAS until the end of the cycle height of plants fromseeds irradiated with 10Gy was significantly higher than thatreached after irradiation with 20 50 and 100Gy Final heightof control plants and of those from seeds irradiated withX-ray doses up to 10Gy was significantly higher than thatreached after irradiation with 20Gy and above (Figure 2(a))
Theprogression of the number of leaves and leaf areawerenot consistent with the time course of plant height (Figure 2)Indeed in the control at the end of the crop cycle plantswere characterized by 97 leaves corresponding to 625 cm2leaf area Plants from irradiated seeds formed a number ofleaves significantly higher than control at the end of the cyclewith maximum value in plants from seeds irradiated with10Gy (Figure 2(b)) Similarly under all the radiation dosesbut 100Gy plants showed significantly larger leaf area thanthe control with maximum value at 03 Gy (Figure 2(c))
33 Effect of Ionizing Radiation on LeafAnatomy Microscopyanalysis of the leaf sections showed a regular anatomicalorganisation in both juvenile and compound leaves formedafter seed irradiation at different levels of X-rays Sincethe effect of the treatments was similar in both types ofleaves microscopy views are shown only for compoundleaves (Figure 3) Both leaf types were characterised by atypical dorsiventral structure which did not face disruptivestructural alterations after irradiation of seeds with X-raysMicroscopy observations of leaves from seeds irradiatedwith up to 20Gy did not reveal evident differences com-pared with control leaves (Figures 3(a)ndash3(d)) At 50 and100Gy sporadic perturbations in the mesophyll structurewere detectable (Figures 3(e)ndash3(h)) More specifically thepresence of more shrunk mesophyll cells (Figures 3(g) and3(h)) due to palisade cells collapsed in the lower portion
The Scientific World Journal 5
0
10
20
30
40
50
60
70
80
0 20 40 60 80 100
Plan
t hei
ght (
mm
)
DAS
(a)
0
5
10
15
20
Num
ber o
f lea
ves (
Np
lant
)
0 20 40 60 80 100DAS
(b)
0 20 40 60 80 100DAS
0
20
40
60
80
100
120
140
Plan
t lea
f are
a (cm
2pl
antminus1)
C03 Gy10 Gy
20 Gy50 Gy100 Gy
(c)
Figure 2 Time course of the plant height (a) number of leaves (b) and plant leaf area (c) in S lycopersicum L ldquoMicrotomrdquo as a function ofthe X-ray doses applied at the seed stage Mean values and standard errors are shown (119899 = 5) All second order polynomial curves had 1199032values above 097
(Figure 3(g)) and spongy cells characterised by more angularshape (Figure 3(h)) was found The occurrence of irregular-shaped cells of leaves from seeds irradiated with 50 and100Gy was confirmed by significant lower convexity valuesthan control and low-dose treatments (Tables 1 and 2) Theepidermis of compound leaves from irradiated seeds tendedto have less elongated cells (ie lower aspect ratio and highersphericity) while in the palisade parenchyma an oppositetrend was observed (Table 2) No specific tendency of cellshape variation was found in the juvenile leaves (Table 1)Themain veins did not show any irregularities in vessel walls afterany irradiation treatments (Figures 3(i) and 3(j)) Epifluores-cence microscopy showed the autofluorescence of phenoliccompounds which were mainly localised along chloroplast
membranes in both control leaves and those from irradiatedplants (Figures 4(a) and 4(b)) No differences in autofluores-cence distribution were observed in mesophyll cells of juve-nile and compound leaves from control and irradiated seedsThe percentage of phenolic compounds showed a nonsignifi-cant tendency to increase in juvenile leaves at low irradiationdoses followed by a significant decrease at 50 and 100Gy(Figure 4(c)) In compound leaves phenolic compoundstended to decrease after all irradiation treatments with thelowest values in leaves from seeds irradiated at 50 and 100Gy(Figure 4(d))
The other analysed anatomical parameters did not show aclear-cut dose-dependent response (Figure 5 Tables 1 and 2)In juvenile leaves the size of cells significantly decreased after
6 The Scientific World Journal
(b) (c)
(d) (e)
(f)
(g) (h)
(i)
(j)
(a)
Figure 3 Light microscopy views of cross sections of leaf lamina of S lycopersicum L ldquoMicrotomrdquo compound leaves from plants developedfrom control (a) and irradiated seeds 03 Gy (b) 10Gy (c) 20Gy (d) 50Gy (e g) and 100Gy (f h) Detail of xylem in the main vein is shownin leaves from control (i) and 100Gy irradiated seeds (j) Images (andashh) are at the same magnification Bar = 50 120583m
the irradiation of seeds only in palisade parenchyma withthe lowest values at 20Gy (Figures 5(c) 5(e) 5(g) and 5(i))By contrast in compound leaves the application of 100Gyresulted in increased cell size compared with control andleaves from seeds irradiated up to 50Gy (Figures 5(d) 5(f)5(h) and 5(j))
34 Effect of Ionizing Radiation on Fluorescence Measure-ments and Pigment Content No significant differences were
detectable in fluorescence parameters and total photosyn-thetic pigment content in plants grown from irradiated seedsat different X-rays doses (Figure 6) A tendency to NPQincrease was found at 03 and 10Gy followed by a decreaseat doses higher than 20Gy However the lack of significantdifferences in ΦPSII and NPQ among treatments indicatedthat X-rays delivered at the seed stage on tomato plantsdid not alter the capability of photosynthetic apparatus oftomato plants either to convert light energy in photochem-istry or to perform thermal dissipation mechanisms The
The Scientific World Journal 7
(a) (b)
Phen
olic
com
poun
ds (
)
aa a
aab
b
0
10
20
30
40
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf JL
(c)
Phen
olic
com
poun
ds (
) aab
abc ab
c bc
0
10
20
30
40
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf CL
(d)
Figure 4 Epifluorescence microscopy views of lamina cross sections of S lycopersicum L ldquoMicrotomrdquo compound leaves from plantsdeveloped from control (a) and 100Gy irradiated seeds (b) Phenolic compounds along chloroplast membranes are autofluorescent Percentof mesophyll occupied by phenolic compounds () is shown in (c) and (d) for juvenile (JL) and compound (CL) leaves respectively Meanvalues and standard errors are reported (119899 = 15) different letters correspond to significantly different values after multiple comparison tests(119875 lt 005)
maximum photochemical efficiency of PSII (FvFm) washigher in leaves from irradiated seeds than control anddid not show variations among different X-rays treatmentsthis indicated the maintenance of higher level of photo-synthetic activity after irradiation irrespectively from dose(Figure 6(c))
The total chlorophyll as well as the total carotenoidcontent did not show significant changes due to irradiationof seeds (Figure 7)
4 Discussion
Ionising radiation can have positive null or negative effectson plant development depending on the properties of theradiation the features of the plant material and possibleinteractions with other environmental factors [2] There iscommon agreement that plants are characterised by highradioresistance moreover low-LET ionising radiation hasbeen reported to increase germination percentage rootlength plant height and other growth parameters especiallyat low doses which induce hormesis [16 17 45ndash47] Ourresults suggest that S lycopersicum ldquoMicrotomrdquo is charac-terised by high radioresistance at the seed stage indeed
irradiation of dry seeds with a wide range of X-ray dosesdid not hamper germination and plant growth up to thecompletion of the crop cycle An hormesis effect was evidentsince growth either remained unaffected or was positivelyaffected especially by low doses of X-rays Apart from plantsgrown from seeds irradiated with 03 and 10Gy all plantsfrom irradiated seeds were characterised by a more compacthabitus higher number of larger leaves was not accompaniedby increased height The radiation-induced more compactgrowth pattern is positive in BLSSs in Spacewhere availabilityof volume for growth is a major technical constraint [7 4849] In the cultivar Microtom irradiation with doses equalto or higher than 20Gy caused a decrease in plant heightin agreement with the induction of dwarf or semidwarfgrowth reported as a general effect of exposure to ionisingradiation [18] Hence irradiation on the ldquoMicrotomrdquo seedswould intensify the trait of dwarf growth which alreadycharacterise this cultivar and should be taken into account forcultivar selection for the BLSSs [50] Any variations in growthparameters in Space-oriented ecologically closed systemscan determine valuable changes in terms of optimisationof volume and resource recycling These changes can bestraightaway converted into economic saving or waste [9]For example the increase in the number and size of leaves
8 The Scientific World Journal
Palisade parenchyma Spongy parenchyma
ab b ab a ab ab
a a a a a a
0010203040506
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf JL
Lam
ina t
hick
ness
(mm
)
(a)
Palisade parenchyma Spongy parenchyma
b b b b b a
a ab ab b abab
Leaf CL
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
010203040506
Lam
ina t
hick
ness
(mm
)
(b)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a a aa a
a
0200400600800
1000Upper epidermis
Cel
l are
a (120583
m2)
(c)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
1000
a a a a aa
Upper epidermis
Cel
l are
a (120583
m2)
(d)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
a bc bc c bc b
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(e)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
ab ab b ab ab a
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(f)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a aa
a a a
0200400600800
100012001400
Cel
l are
a (120583
m2)
(g)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
100012001400
b b b b ba
Cel
l are
a (120583
m2)
(h)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
a aa
a a a
0
200
400
600
800
Cel
l are
a (120583
m2)
(i)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
b b b b ba
0
200
400
600
800
Cel
l are
a (120583
m2)
(j)
Figure 5 Lamina thickness (a b) and cell area of upper epidermis (c d) palisade (e f) spongy (g h) parenchymas and lower epidermis (i j)of S lycopersicum L ldquoMicrotomrdquo leaves of control and irradiated plants at different doses of X-rays in both juvenile (JL) and compound (CL)leaves Mean values and standard errors are shown (119899 = 15 in (a b) 119899 = 45 in (cndashj)) Different letters correspond to significantly differentvalues according to multiple comparison tests (119875 lt 005)
The Scientific World Journal 9
04
05
06
07
08
a a a a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
ΦPS
II
(a)
00
02
04
06
08
NPQ a
a aa a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
(b)
070
075
080
085
090
a
b b b b b
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
FF
m
(c)
Figure 6 Quantum yield of PSII electron transport (ΦPSII a) nonphotochemical quenching (NPQ b) and maximal PSII photochemicalefficiency (FvFm c) in compound (CL) S lycopersicum L ldquoMicrotomrdquo leaves of control and plants from seeds irradiated at different dosesof X-rays Mean values and standard deviations are shown (119899 = 5) Different letters correspond to significantly different values according tomultiple comparison tests (119875 lt 005)
0
1
2
3
4
a
aa
a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Chl a
+ b
(mg g
minus1)
(a)
0
02
04
06
a
aa a
a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Car x
+ c
(mg g
minus1)
(b)
Figure 7 Total chlorophyll content (Chl a+b) (a) and total carotenoid content (Car x+c) (b) in compound (CL) S lycopersicum L ldquoMicrotomrdquoleaves of control and plants from seeds irradiated at different doses of X-rays Mean values and standard errors are shown (119899 = 5) Differentletters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
induced in ldquoMicrotomrdquo especially by low doses of irradiationcan be beneficial to atmosphere regeneration of pressurisedmodules provided that formed leaves do not show structuralaberrations which reduce their function
General anatomical structure of both juvenile and com-pound ldquoMicrotomrdquo leaves was only slightly perturbed inplants from seeds subjected to the highest levels of radiation
Clear-cut dose-depended tendencies of variations were notfound in lamina thickness and cell size and shape of thevarious tissues The formation of smaller palisade cells wasfound only in juvenile leaves of irradiated seeds while anincrease in cell size was found only in spongy parenchymaand lower epidermis of compound leaves from seeds irra-diated at 100Gy The diverse growth behaviour found in
10 The Scientific World Journal
Table 1 Cell shape in different tissues of juvenile leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown (119899 = 75)Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf JL Aspect ratio Sphericity Convexity
Upper epidermis
C 197 plusmn 016 a 0308 plusmn 0050 a 0926 plusmn 0021 ab03Gy 200 plusmn 013 a 0290 plusmn 0038 a 0941 plusmn 0013 a10Gy 210 plusmn 012 a 0239 plusmn 0037 a 0898 plusmn 0016 ab20Gy 217 plusmn 014 a 0258 plusmn 0045 a 0909 plusmn 0019 abc50Gy 170 plusmn 008 a 0376 plusmn 0037 a 0927 plusmn 0016 ab100Gy 197 plusmn 014 a 0286 plusmn 0042 a 0876 plusmn 0028 c
Palisade parenchyma
C 290 plusmn 010 a 0109 plusmn 0015 a 0879 plusmn 0010 a03 Gy 304 plusmn 012 a 0099 plusmn 0016 a 0805 plusmn 0016 b10Gy 309 plusmn 013 a 0093 plusmn 0019 a 0792 plusmn 0015 b20Gy 283 plusmn 011 a 0101 plusmn 0017 a 0784 plusmn 0017 b50Gy 314 plusmn 014 a 0082 plusmn 0016 a 0772 plusmn 0016 b100Gy 275 plusmn 011 a 0119 plusmn 0020 a 0805 plusmn 0017 b
Spongy parenchyma
C 190 plusmn 008 a 0307 plusmn 0033 a 0893 plusmn 0012 a03 Gy 180 plusmn 007 a 0341 plusmn 0029 a 0876 plusmn 0013 a10Gy 203 plusmn 010 a 0290 plusmn 0031 a 0901 plusmn 0012 a20Gy 194 plusmn 012 a 0344 plusmn 0040 a 0881 plusmn 0015 a50Gy 174 plusmn 009 a 0393 plusmn 0035 a 0883 plusmn 0014 a100Gy 198 plusmn 012 a 0322 plusmn 0037 a 0878 plusmn 0015 a
Lower epidermis
C 178 plusmn 009 bc 0343 plusmn 0045 ab 0919 plusmn 0019 a03 Gy 225 plusmn 017 ab 0263 plusmn 0044 b 0933 plusmn 0024 a10Gy 219 plusmn 011 abc 0218 plusmn 0034 b 0888 plusmn 0020 ab20Gy 180 plusmn 013 bc 0378 plusmn 0047 ab 0926 plusmn 0017 a50Gy 167 plusmn 001 c 0461 plusmn 0051 a 0924 plusmn 0021 a100Gy 238 plusmn 021 a 0257 plusmn 0057 b 0857 plusmn 0030 b
the various tissues of structures of different age is in agree-ment with the different light-mediated control of cell devel-opment in the various leaf tissues [51] Altered mesophyllcharacterized by partly collapsed cells with irregular cellshape was found only sporadically after the exposure of seedsto 50 and 100Gy in theMicrotomcultivar Suchmodificationswere less marked than those found in leaves of bean wherethe target organs for irradiation were the leaves themselves[30] leaves directly exposed to X-rays in bean showed theformation of more loose parenchyma with larger cells likelydue to a radiation-induced damage to cell walls that reducedmechanical constraints to cell enlargement [30 52] Furtherinvestigations on cell wall composition would be useful toelucidate whether a possible alteration of cell walls wasresponsible for cell enlargement in the spongy parenchymaand epidermis of leaves from seeds of ldquoMicrotomrdquo irradiatedwith the highest dose of X-rays However in the cultivarMicrotom the slight alterations of mesophyll structure didnot impair photosynthetic efficiency as demonstrated by thelack of significant dose-dependent variation trends in thequantum yield of PSII electron transport (ΦPSII) nonphoto-chemical quenching (NPQ) and maximal PSII photochemi-cal efficiency (FvFm) The increased values of FvFm closeto 08 which is considered an optimal value for healthy plants[53] suggested that the photosynthetic apparatus of the leavesfrom irradiated seeds has higher levels of photochemical
efficiency irrespective of dose The unchanged or increasedcontent of both chlorophyll and carotenoid pigments con-firmed that the photosynthetic apparatus remained stablealso in terms of light harvesting capacity and indicatedno deleterious effect of radiation on plant photosyntheticmachinery RecentlyMarcu et al [17] showed that irradiationof lettuce seeds with gamma rays up to 30Gy enhancedthe content of chlorophyll a chlorophyll b and carotenoidswhile higher doses (up to 70Gy) resulted in their decreaseHowever the effect of ionising radiation on photosyntheticpigment content is quite controversial Increased pigmentcontent is ascribed to the phenomenon of hormesis whilea lower content is generally ascribed to the inhibition ofchlorophyll and carotenoid synthesis or to oxidation due tothe high levels of radiation-induced water radiolysis [2 1454ndash56] Some plants possess natural screens against radiationwhen their tissues contain phenolic compounds with antioxi-dant functions and can counteract photoinhibitory processes[29 57] An increase of phenolic compounds linked to thechloroplast membranes has been recently found in Phaseolusvulgaris irradiated leaves exposed to high levels of X-rays[28 30] Changes in phenolic content after the exposure toSpace can be attributed either to direct effects on the synthesisor mobilization of phenolic compounds or to indirect effectsdue to changes in ultrastructural organization such as theincrease in the number of chloroplasts per cell [30 58]
The Scientific World Journal 11
Table 2 Cell shape in different tissues of compound leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown(119899 = 75) Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf CL Aspect ratio Sphericity Convexity
Upper epidermis
C 166 plusmn 007 c 0422 plusmn 0042 a 0946 plusmn 0010 a03 Gy 216 plusmn 017 ab 0244 plusmn 0033 c 0925 plusmn 0016 ab10Gy 171 plusmn 011 bc 0401 plusmn 0042 a 0914 plusmn 0013 b20Gy 186 plusmn 010 abc 0328 plusmn 0041 ab 0919 plusmn 0020 ab50Gy 225 plusmn 018 a 0247 plusmn 0041 c 0897 plusmn 0019 b100Gy 188 plusmn 009 abc 0319 plusmn 0045 ab 0902 plusmn 0016 b
Palisade parenchyma
C 355 plusmn 012 a 0073 plusmn 0014 b 0890 plusmn 0012 a03 Gy 289 plusmn 010 b 0102 plusmn 0013 ab 0847 plusmn 0018 b10Gy 300 plusmn 014 b 0104 plusmn 0021 ab 0826 plusmn 0017 b20Gy 302 plusmn 011 b 0094 plusmn 0013 ab 0810 plusmn 0014 b50Gy 283 plusmn 012 b 0107 plusmn 0021 ab 0750 plusmn 0021 c100Gy 275 plusmn 012 b 0122 plusmn 0023 a 0809 plusmn 0015 b
Spongy parenchyma
C 193 plusmn 010 a 0323 plusmn 0035 a 0904 plusmn 0011 a03 Gy 183 plusmn 010 a 0375 plusmn 0042 a 0879 plusmn 0013 a10Gy 176 plusmn 007 a 0357 plusmn 0034 a 0869 plusmn 0011 a20Gy 180 plusmn 009 a 0366 plusmn 0038 a 0885 plusmn 0016 a50Gy 171 plusmn 006 a 0372 plusmn 0034 a 0879 plusmn 0015 a100Gy 188 plusmn 010 a 0339 plusmn 0037 a 0883 plusmn 0015 a
Lower epidermis
C 148 plusmn 008 b 0545 plusmn 0045 a 0957 plusmn 0015 a03 Gy 210 plusmn 017 a 0295 plusmn 0046 b 0929 plusmn 0019 ab10Gy 190 plusmn 012 ab 0334 plusmn 0043 b 0893 plusmn 0019 b20Gy 178 plusmn 011 ab 0388 plusmn 0047 b 0929 plusmn 0017 ab50Gy 172 plusmn 009 ab 0367 plusmn 0040 b 0914 plusmn 0016 b100Gy 191 plusmn 014 ab 0362 plusmn 0054 b 0888 plusmn 0025 b
The lowering of phenolic content at high levels of X-rays inldquoMicrotomrdquo leaves is not in agreement with general trendsindicating enhanced phenolics production in plants grownin Space [58] or in various plant tissues directly irradiatedwith gamma rays [59] However the low content of phenolicscan be an indirect effect of increasing volume occupiedby intercellular spaces due to the loosening of parenchymacells
In conclusion our results show that irradiation of the dryseeds of the Microtom cultivar did not hamper germinationand the development of functional leaves The formationof more compact plants characterised by a higher numberof larger leaves can be valuable in terms of increasingresource regeneration in the BLSSs High levels of radiationinduced only slight structural perturbations which weresimilar in juvenile and compound leaves Such perturbationsdid not affect photosynthetic efficiency since the maximalPSII photochemical efficiency was even increased after irra-diation at all doses However the tendency to accumulatelower amounts of phenolic compounds along chloroplastmembranes after irradiation of seeds with high doses ofradiation should be further investigated because it coulddetermine the reduction of natural photoprotection [57]Finally the overall results indicate that theMicrotom cultivarirradiated at the seed stage has high radioresistance duringthe growth which is a desirable feature for plant cultivation
in Space Radioresistance together with dwarf growth andshort life cycle makes this cultivar a valuable candidate forthe cultivation in BLSSs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are grateful to the ldquoCompagnia di San Paolordquo(Turin) and the ldquoPolo delle Scienze e delle Tecnologie perla Vitardquo of the University of Naples Federico II for thefinancial support The authors also thank Maria Giordanoand Pasquale Scognamiglio for their support in plant culti-vation and microscopy analyses The authors acknowledgefour anonymous editors for helpful comments on a previousversion of the paper
References
[1] A Real S Sundell-Bergman J F Knowles D S Woodheadand I Zinger ldquoEffects of ionising radiation exposure on plantsfish and mammals relevant data for environmental radiation
12 The Scientific World Journal
protectionrdquo Journal of Radiological Protection vol 24 no 4 ppA123ndashA137 2004
[2] V De Micco C Arena D Pignalosa and M Durante ldquoEffectsof sparsely and densely ionizing radiation on plantsrdquo Radiationand Environmental Biophysics vol 50 no 1 pp 1ndash19 2011
[3] N Kumari P Kumar D Mitra B Prasad B N Tiwary andL Varshney ldquoEffects of ionizing radiation on microbial decon-tamination phenolic contents and antioxidant properties oftriphalardquo Journal of Food Science vol 74 no 3 pp M109ndashM1132009
[4] A Tanaka N Shikazono and Y Hase ldquoStudies on biologicaleffects of ion beams on lethality molecular nature of mutationmutation rate and spectrum of mutation phenotype for muta-tion breeding in higher plantsrdquo Journal of Radiation Researchvol 51 no 3 pp 223ndash233 2010
[5] M Durante and A Kronenberg ldquoGround-based research withheavy ions for space radiation protectionrdquo Advances in SpaceResearch vol 35 no 2 pp 180ndash184 2005
[6] E Hu S I Bartsev and H Liu ldquoConceptual design of a biore-generative life support system containing crops and silkwormsrdquoAdvances in Space Research vol 45 no 7 pp 929ndash939 2010
[7] V deMicco G Aronne G Colla R Fortezza and S de PascaleldquoAgro-biology for bioregenerative life support systems in long-term space missions general constraints and the Italian effortsrdquoJournal of Plant Interactions vol 4 no 4 pp 241ndash252 2009
[8] T W Halstead and F R Dutcher ldquoPlants in spacerdquo AnnualReview of Plant Physiology vol 38 pp 317ndash345 1987
[9] V De Micco S De Pascale R Paradiso and G AronneldquoMicrogravity effects on different stages of higher plant life cycleand completion of the seed-to-seed cyclerdquo Plant Biology vol 16pp 31ndash38 2014
[10] C J Bennett C Pirim and T M Orlando ldquoSpace-weatheringof solar system bodies a laboratory perspectiverdquo ChemicalReviews vol 113 pp 9086ndash9150 2013
[11] R A Kerr ldquoRadiation will make astronautsrsquo trip to Mars evenriskierrdquo Science vol 340 article 1031 2014
[12] D M Hassler C Zeitlin R F Wimmer-Schweingruber et alldquoMarsrsquo surface radiation environment measured with the MarsScience Laboratoryrsquos Curiosity Roverrdquo Science vol 343 no6169 Article ID 1244797 2014
[13] M Durante and F A Cucinotta ldquoHeavy ion carcinogenesis andhuman space explorationrdquo Nature Reviews Cancer vol 8 no 6pp 465ndash472 2008
[14] C Arena V De Micco and A De Maio ldquoGrowth alterationand leaf biochemical responses in P vulgaris plants exposed todifferent doses of ionizing radiationrdquo Plant Biology vol 16 pp194ndash202 2014
[15] R W Holst and D J Nagel ldquoRadiation effects on plantsrdquo inPlants For Environmental Studies W Wang J W Gorsuch andJ S Hughes Eds pp 37ndash81 Lewis Publishers Boca Raton FlaUSA 1997
[16] T D Luckey Hormesis With Ionizing Radiation CRC PressBoca Raton Fla USA 1980
[17] D Marcu V Cristea and L Daraban ldquoDose-dependent effectsof gamma radiation on lettuce (Lactuca sativa var capitata)seedlingsrdquo International Journal of Radiation Biology vol 89 pp219ndash223 2013
[18] M Mei H Deng Y Lu et al ldquoMutagenic effects of heavy ionradiation in plantsrdquo Advances in Space Research vol 14 no 10pp 363ndash372 1994
[19] Y LiM Liu Z Cheng andY Sun ldquoSpace environment inducedmutations prefer to occur at polymorphic sites of rice genomesrdquoAdvances in Space Research vol 40 no 4 pp 523ndash527 2007
[20] D S Kim J-B Kim E J Goh et al ldquoAntioxidant response ofArabidopsis plants to gamma irradiation genome-wide expres-sion profiling of the ROS scavenging and signal transductionpathwaysrdquo Journal of Plant Physiology vol 168 no 16 pp 1960ndash1971 2011
[21] T J Brodribb ldquoXylem hydraulic physiology the functionalbackbone of terrestrial plant productivityrdquo Plant Science vol177 no 4 pp 245ndash251 2009
[22] R M Wheeler J C Sager R P Prince andW M Knott ldquoCropproduction for advanced life support systemsmdashobservationfrom the Kennedy Space Center Breadboard Projectrdquo NASATM-2003-211184 NASA Kennedy Space Center Florida 2003
[23] R M Wheeler C L Mackowiak G W Stutte et al ldquoCropproductivities and radiation use efficiencies for bioregenerativelife supportrdquo Advances in Space Research vol 41 no 5 pp 706ndash713 2008
[24] R Paradiso V De Micco R Buonomo G Aronne G Bar-bieri and S De Pascale ldquoSoilless cultivation of soybean forBioregenerative Life-Support Systems a literature review andthe experience of the MELiSSA ProjectmdashFood characterisationPhase Irdquo Plant Biology vol 16 pp 69ndash78 2014
[25] D M Porterfield ldquoThe biophysical limitations in physiologicaltransport and exchange in plants grown in microgravityrdquoJournal of Plant Growth Regulation vol 21 no 2 pp 177ndash1902002
[26] K Yokthongwattana and A Melis ldquoPhotoinhibition and recov-ery in oxygenic photosynthesis mechanism of a photosystem IIdamage and repair cyclerdquo in Photoprotection PhotoinhibitionGene Regulation and Environment B Demmig-Adams WW Adams III and A K Mattoo Eds pp 175ndash191 SpringerDordecht The Netherlands 2006
[27] M T Giardi G Rea M D Lambreva et al ldquoMutations ofphotosystem II D1 protein that empower efficient phenotypesof Chlamydomonas reinhardtii under extreme environment inSpacerdquo PLoS ONE vol 8 Article ID e64352 2013
[28] C Arena V De Micco G Aronne M Pugliese A Virzo DeSanto andADeMaio ldquoResponse ofPhaseolus vulgarisL plantsto low-let ionizing radiation growth and oxidative stressrdquo ActaAstronautica vol 91 pp 107ndash114 2013
[29] V Lattanzio P A Kroon and S Quideau ldquoPlant phenolicsmdashsecondary metabolites with diverse functionsrdquo in RecentAdvances in Polyphenol Research F Daay and V Lattanzio Edspp 1ndash35 Wiley-Blackwell Oxford UK 2008
[30] V De Micco C Arena and G Aronne ldquoAnatomical alterationsof Phaseolus vulgaris L mature leaves irradiated with X-raysrdquoPlant Biology vol 16 pp 187ndash193 2014
[31] A D Krikorian ldquoPlants and somatic embryos in space whathave we learnedrdquo Gravitational and Space Biology Bulletin vol11 no 2 pp 5ndash14 1998
[32] R Meissner Y Jacobson S Melamed et al ldquoA new modelsystem for tomato geneticsrdquoThe Plant Journal vol 12 no 6 pp1465ndash1472 1997
[33] E Emmanuel and A A Levy ldquoTomato mutants as tools forfunctional genomicsrdquo Current Opinion in Plant Biology vol 5no 2 pp 112ndash117 2002
[34] E Martı C Gisbert G J Bishop M S Dixon and J LGarcıa-Martınez ldquoGenetic and physiological characterizationof tomato cv Micro-Tomrdquo Journal of Experimental Botany vol57 pp 2037ndash2047 2006
The Scientific World Journal 13
[35] T Kurimoto J V H Constable and A Huda ldquoEffects ofionizing radiation exposure on arabidopsis thalianardquo HealthPhysics vol 99 no 1 pp 49ndash57 2010
[36] L Reale D Gigante F Landucci F Ferranti and R VenanzonildquoMorphological and histo-anatomical traits reflect die-back inPhragmites australis (Cav) Steudrdquo Aquatic Botany vol 103 pp122ndash128 2012
[37] K Fukuzawa ldquoUltraviolet microscopyrdquo in Methods in LigninChemistry S Y Lin andCWDence Eds pp 110ndash131 SpringerBerlin Germany 1992
[38] S E Ruzin Plant Microtechnique and Microscopy OxfordUniversity Press New York NY USA 1999
[39] V De Micco G Aronne J-P Joseleau and K Ruel ldquoXylemdevelopment and cell wall changes of soybean seedlings grownin spacerdquo Annals of Botany vol 101 no 5 pp 661ndash669 2008
[40] S Van Buggenhout T Grauwet A Van Loey and M Hen-drickx ldquoStructureprocessing relation of vacuum infused straw-berry tissue frozen under different conditionsrdquo European FoodResearch and Technology vol 226 no 3 pp 437ndash448 2008
[41] V De Micco and G Aronne ldquoBiometric anatomy of seedlingsdeveloped onboard of Foton-M2 in an automatic system sup-porting growthrdquoActa Astronautica vol 62 no 8-9 pp 505ndash5132008
[42] B Genty J M Briantais and N R Baker ldquoThe relationshipbetween quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescencerdquo Biochimica andBiophysica Acta vol 990 pp 87ndash92 1989
[43] W Bilger and O Bjorkman ldquoRole of the xanthophyll cycle inphotoprotection elucidated by measurements of light-inducedabsorbance changes fluorescence and photosynthesis in leavesof Hedera canariensisrdquo Photosynthesis Research vol 25 no 3pp 173ndash185 1990
[44] H K Lichtenthaler ldquoChlorophylls and carotenoids pigmentsof photosynthetic biomembranesrdquoMethods in Enzymology vol148 pp 350ndash382 1987
[45] J P Maity DMishra A Chakraborty A Saha S C Santra andS Chanda ldquoModulation of some quantitative and qualitativecharacteristics in rice (Oryza sativa L) and mung (Phaseolusmungo L) by ionizing radiationrdquo Radiation Physics and Chem-istry vol 74 no 5 pp 391ndash394 2005
[46] L Cheng H Yang B Lin et al ldquoEffect of gamma-ray radiationon physiological morphological characters and chromosomeaberrations of minitubers in Solanum tuberosum Lrdquo Interna-tional Journal of Radiation Biology vol 86 no 9 pp 791ndash7992010
[47] B Al-Safadi and R Elias ldquoImprovement of caper (Capparisspinosa L) propagation using in vitro culture and gammairradiationrdquo Scientia Horticulturae vol 127 no 3 pp 290ndash2972011
[48] O Monje G W Stutte G D Goins D M Porterfield and GE Bingham ldquoFarming in space environmental and biophysicalconcernsrdquo Advances in Space Research vol 31 no 1 pp 151ndash1672003
[49] V De Micco M Scala and G Aronne ldquoEffects of simulatedmicrogravity on male gametophyte of Prunus Pyrus andBrassica speciesrdquo Protoplasma vol 228 no 1ndash3 pp 121ndash1262006
[50] V De Micco R Buonomo R Paradiso S De Pascale and GAronne ldquoSoybean cultivar selection for Bioregenerative LifeSupport Systems (BLSS)mdashtheoretical selectionrdquo Advances inSpace Research vol 49 no 10 pp 1415ndash1421 2012
[51] T Kozuka S-G Kong M Doi K-I Shimazaki and ANagatani ldquoTissue-autonomous promotion of palisade celldevelopment by phototropin 2 in Arabidopsisrdquo Plant Cell vol23 no 10 pp 3684ndash3695 2011
[52] E Kovacs and A Keresztes ldquoEffect of gamma and UV-BCradiation on plant cellsrdquoMicron vol 33 no 2 pp 199ndash210 2002
[53] R Ogaya J Penuelas D Asensio and J Llusia ldquoChlorophyllfluorescence responses to temperature and water availabilityin two co-dominant Mediterranean shrub and tree speciesin a long-term field experiment simulating climate changerdquoEnvironmental and Experimental Botany vol 73 no 1 pp 89ndash93 2011
[54] R Zaka C M Vandecasteele and M T Misset ldquoEffects oflow chronic doses of ionizing radiation on antioxidant enzymesand G6PDH activities in Stipa capillata (Poaceae)rdquo Journal ofExperimental Botany vol 53 no 376 pp 1979ndash1987 2002
[55] J-H KimM-H Baek Y C Byung GW Seung and J-S KimldquoAlterations in the photosynthetic pigments and antioxidantmachineries of red pepper (Capsicum annuum L) seedlingsfrom gamma-irradiated seedsrdquo Journal of Plant Biology vol 47no 4 pp 314ndash321 2004
[56] N A Al-Enezi and J M Al-Khayri ldquoAlterations of DNA ionsand photosynthetic pigments content in date palm seedlingsinduced by X-irradiationrdquo International Journal of Agricultureand Biology vol 14 pp 329ndash336 2012
[57] V de Micco C Arena L Vitale G Aronne and A V de SantoldquoAnatomy and photochemical behaviour of mediterranean Cis-tus incanus winter leaves under natural outdoor and warmerindoor conditionsrdquo Botany vol 89 no 10 pp 677ndash688 2011
[58] L H Levine H G Levine E C Stryjewski V Prima and WC Piastuch ldquoEffect of spaceflight on isoflavonoid accumulationin etiolated soybean seedlingsrdquo Journal of Gravitational Physiol-ogy vol 8 no 2 pp 21ndash27 2001
[59] X Fan ldquoAntioxidant capacity of fresh-cut vegetables exposedto ionizing radiationrdquo Journal of the Science of Food andAgriculture vol 85 no 6 pp 995ndash1000 2005
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 The Scientific World Journal
either positive or negative phenomena in plants exposedto low-LET (linear energy transfer) (eg X- and gamma-rays) and high-LET (eg protons and heavy ions) ionisingradiation [2] Definitely in many experiments carried out inSpace plants were exposed to various combinations of bothaltered gravity and radiation Ground-based research helpsdistinguish between plant reactions triggered by each oneof the two factors On Earth the simulation of the complexSpace radiation spectrum including the radiation emittedfrom the Sun and galactic cosmic rays is impossible eventhough single particles can be produced at large particleaccelerators [10] The exposure likely to be encountered byorganisms in Space depends on the shielding features of thespace vehicles andor planetary platforms as well as on theevolution of the solar cycle during the permanence in Spacehowever it can be estimated that during a relatively quietsolar cycle a permanence of one year in Space would exposeorganisms to a dose of less than 10Gy [10ndash12] In Space a largefraction of the dose is delivered by protons (approximately87) which are low-LET particles with similar effectivenesscompared to X-rays Therefore initial studies can exploitconventional X-rays as the reference radiation to obtaindata which are the necessary basis for future comparisonusing accelerated ion beams [13 14] The severity of theeffects of low- and high-LET ionising radiation dependson several factors related to the radiation itself (eg typetotal dose and dose rate) and to plant features (eg speciescultivar plant age complexity of the target organ or tissueand level of ploidy) [2 15] An interesting phenomenonin plants known as hormesis is the occurrence of positiveresponses to low doses of ionising radiation which wouldstimulate processes such as germination and growth [16]A recent paper by Marcu et al [17] showed enhancedgermination plant growth and synthesis of photosyntheticpigments when seeds were irradiated with low doses ofgamma rays but opposite responses when doses were highSparsely and densely ionising radiation can also determinemutations inducing favourable agronomical traits such asearlier maturity higher yields and resistance to diseases [1819] On the other hand there is evidence of altered geneexpression due to plantsrsquo exposure to ionising radiation lead-ing to detrimental consequences culminating in plant death[20]
The very prolific literature on radiation-induced geneticaberrations andmetabolic alterations (especially linked to theROSmdashreactive oxygen speciesmdashproduction) is accompaniedby scarce information at the structural level [2] Giventhat the physics of plant structure ultimately regulate majormetabolic and physiological processes [21] the developmentof well-structured above-ground organs is needed to obtainefficient photosynthesis The photosynthetic process can beconsidered one of themain functions accomplished by higherplants in plant-based modules of BLSSs for air regenerationThus knowing how ionising radiation affectsmorphogenesisand even more leaf development is interesting to evaluatepossible impairment in photosynthesis which would con-strain the maximisation of resource efficiency in the BLSSsIndeed the optimisation of resource use in the BLSSs isconsidered a challenge in Space exploration [6 22ndash24]
Space factors can directly alter the inner structure ofplants or impair photosynthesis due to the alteration of gasexchange or of functioning of the photosystems Indeed thelack of convective forces due to microgravity can alter theavailability of gases at the carboxylation sites [25] On theother hand high levels of ionising radiation can negativelyaffect pigment-protein complexes and enzymes responsiblefor light absorption electron transport and carbon reductioncycle [2] Moreover high radiation levels may also inducephotoprotection mechanisms and affect the behaviour of theD1 protein in the PSII repair cycle [26 27] Photoprotectionmechanisms can rely on physiological adjustments [14 28]andor on the intensification of structural and biochemicalbarriers such as trichomes and phenolic compounds acting asnatural screens against radiation [29] Indeed the increasedcontent of phenolic compounds linked to chloroplast mem-branes has been shown in bean leaves irradiated with highdoses of X-rays [30]
The main aim of this paper is to analyse whether X-ray irradiation of Solanum lycopersicum L ldquoMicrotomrdquo atthe seed stage affects leaf development and anatomy fluo-rescence parameters and pigment content This study hasbeen conducted within a wider experimentation aiming toanalyse the effect of irradiation performed at seed stage onvarious growth processes of dwarf tomato In this paperwe focus on the analysis of leaf morphofunctional traits intwo leaf types In particular we analyse leaves maintainingjuvenile traits (ie the first two true leaves characterisedby simpler morphology than successive leaves) and adultcompound leaves in order to evaluate whether similarstructures characterised by different complexity and ageshow differential responses Indeed it is recognised that theincrease in morphological complexity confers more efficientbuffering capacity to biological systems for dealing withradiation-induced damage [31] An X-ray source at 250 kVwas used to irradiate the seeds because X-rays constitutethe reference radiation to assess the damage caused by anyother radiation source at the same dose The dwarf cultivarMicrotomwas selected as plant material because it is a modelsystem widely used for research in molecular biology [32ndash34] Moreover its compact habit short life cycle and fruitdevelopment not requiring hand pollination are desirablefeatures in BLSSs and would make this cultivar a modelsystem also for research in plant Space biology
2 Materials and Methods
21 Plant Material and Irradiation Procedure The experi-ment was conducted using seeds of Solanum lycopersicum LldquoMicrotomrdquo provided by Holland Online Vof (AmsterdamThe Netherlands) Seed irradiation was performed on April2013 Dry seeds were placed into Petri dishes in one layerand irradiated with five doses of X-rays (03 Gy 10Gy 20Gy50Gy and 100Gy) 250 kVp at dose rate of 1 Gymin Aset of three Petri dishes with 15 seeds each was used foreach irradiation treatment and for a nonirradiated controlDoses up to 20Gy were chosen in order to build a reference-response range to explore plant sensitivity as generally set in
The Scientific World Journal 3
experiments to evaluate the effect of radiation on biologicalsystems [13 14 28 30] The highest doses (50Gy and 100Gy)are generally applied since they can be considered as positivecontrols as there ismore likely a plant reaction at these dosesX-rays were delivered as one dose per each Petri dish [35] inorder to avoid cumulative effects on the same sample X-rayswere produced by a Thomson tube (TR 300 F 250 kVp Sta-bilipan Siemens Forchheim Germany) with tungsten cath-ode filtered by 1mm thick copper foil and with 15mA anodiccurrent Before irradiation a physical dosimetry was per-formed by using an ionization chamber (VictoreenModlingAustria) The X-ray intensity was measured at the estab-lished distance where seeds were positioned under the X-raytube
After the exposure to X-rays (on 24 April) the irradiatedseeds and controls were placed into Petri dishes on three lay-ers of filter paper moistened with distilled water Petri disheswere incubated in the dark at temperature of 20∘C andmoni-tored daily to analyse germination percentage and rate (con-sidered as the number of days taken to reach the maximumgermination percentage) Seeds were classified as germinatedwhen the emerging root grew as long as the seed maximumdiameter
22 Growth Conditions The growth experiment was carriedout in a greenhouse with a polyethylene film cover Thegreenhouse was equipped with a black plastic shading net(70 shading) in order to keep the temperature and lightintensity close to the levels feasible in test bed evaluationof BLSSs technologies [22 23] Wavelength composition ofthe sunlight including the whole PAR range spectrum forphotosynthetic activity was not affected
Seedlings were transplanted into pots (4 cm diameter) onpeat-based compost (peat soil 1 1 in volume) and trans-ferred to the greenhouse 5 days after sowing (DAS) Thenthey were repotted in 10 cm pots using the same substrateat the stadium of fourth true leaf (ie two juvenile and twocompound leaves) at 22DAS In the cultivar Microtom thefirst two true leaves maintain juvenile traits and are charac-terised by simple morphology they are trifoliate with littleor not evident lobes Successive leaves show more complexmorphology having pennate-compound lamina with highlyindented lobes
During the whole growth period plants were irrigatedwith tapwater at 2-day interval in order to reach the containercapacity (till the beginning of drainage)
The temperature inside the greenhouse was recordedevery 10 minutes by means of a data logger (Tynitag Ultra2Gemini Data Loggers Chichester UK) Photosynthetic pho-ton flux density (PPFD) at the canopy level was recordeddaily at 1200 pm with a Delta OHMDO9847multifunctionmeter
The whole crop cycle (up to fruit ripening) lasted from 24April until 6 August 2013
During the whole growth cycle the mean values of tem-perature inside the greenhouse were 284216∘C (daynight)and the light intensity recorded at noon at the canopy levelwas 446 120583mol mminus2 sminus1 on average
23 Biometric and Microscopy Analyses Biometric analyseswere performed once a week on 10 plants per treatmentthroughout the crop cycle Plant height and number of leaveswere recorded Moreover single leaves of the main stem werephotographed with a digital camera (Nikon D3100 NikonEurope BV) in order to examine leaf area increments withthe software program AnalySIS 32 (Olympus HamburgGermany)
From 5 plants per treatment the second juvenile leaf(JL) and the first compound leaf (CL) were collected whenthey were fully developed One median leaflet (one of thetwo opposite leaflets below the apex) per leaf was used formicroscopy analyses while the others were used for pigmentextraction
Leaflets destined to microscopy analyses were immedi-ately fixed in FAA (40 formaldehydeglacial acetic acid50ethanol-5590 by volume) for several days Leaflets werecut under a dissection microscope (ZSX9 Olympus) toobtain subsamples of 5 times 5mm in the middle of laminaincluding the main vein Subsamples were dehydrated inan ethanol series and embedded in the acrylic resin JB4(Polysciences Warrington PA USA) Semithin cross sec-tions (5 120583m thick) were cut through a rotative microtomeSections were stained with 0025 Toluidine blue in 01Mcitrate buffer at pH 4 [36] mounted with Canadian Balsamand observed under a light microscope (BX60 Olympus)Unstained sections were mounted with mineral oil for fluo-rescence and observed under an epifluorescence microscope(BX60 Olympus) equipped with a mercury lamp band-passfilter 330ndash385 nm dichromatic mirror 400 nm and aboveand barrier filter 420 nm and above With these filters it waspossible to detect the presence of simple phenolics that areautofluorescent at such wavelengths [37 38] Three sectionsper leaflet were analysed and images were collected by meansof a digital camera (CAMEDIA C4040 Olympus) at variousmagnifications
24 Digital Image Analysis All digital images were analysedwith the AnalySIS 32 (Olympus) software for image analysisThe thickness of the palisade and the spongy parenchymaswere measured in five regions along the leaf lamina Thecell area and shape of upper and lower epidermis palisadeand spongy parenchyma were quantified in 15 cells pereach tissue per section More specifically cell shape wascharacterised as (a) aspect ratio (maximum widthheightratio of a bounding rectangle for the cell defining how itis elongated) (b) sphericity (roundness of a particle withspherical particles having a maximum value of 1) and (c)convexity (the fraction of the cellrsquos surface area and thearea of its convex a turgid cell has a maximum value of1) [39 40] The cell area occupied by phenolic compoundswas measured in 3 regions (150 times 200 120583m2 each) of themesophyll per section selected avoiding veins Thus thearea occupied by phenolic compounds was calculated asthe percentage of tissuepicture occupied by compoundsappearing autofluorescent at the above-reported filter settings[41]
4 The Scientific World Journal
C 100 Gy50 Gy20 Gy10 Gy03 Gy
Figure 1 Plants of S lycopersicum L ldquoMicrotomrdquo developed fromcontrol and irradiated seeds at 40 DAS (Days After Sowing) Plantsfrom seeds exposed to increasing doses of X-rays (from 0 to 100Gy)are shown from left to right
25 Fluorescence Measurements and Pigment Content Flu-orescence measurements and pigment extraction were con-ducted on 5 pennate-compound leaves of 5 plants subjectedto each irradiation treatment and in controls at 40 DAS
Chlorophyll a fluorescence measurements were carriedout by means of a pulse amplitude modulated fluorometer(Junior-PAM Walz Germany) equipped with a monitoringLeaf-Clip JUNIOR-B (Walz Germany) On 30min dark-adapted leaves the background fluorescence signal Fo wasinduced by internal light provided by a blue LED of about2-3 120583mol photonssdotmminus2sdotsminus1 at a frequency of 05 kHz Themaximal fluorescence level in the dark-adapted state (Fm)was measured by 1 s saturating light pulse (10 000120583molphotonssdotmminus2sdotsminus1) at a frequency of 10 kHz the maximal PSIIphotochemical efficiency (FvFm) was calculated as FvFm =(FmminusFo)FmThemeasurements in the light were carried outby exposing each leaf to a PPFD of 420120583mol photonssdotmminus2sdotsminus1for 5min This level of PPFD was chosen because it fallsin the range of maximal quantum yield for the Microtomcultivar as indicated by fluorescence fast-response curves tolight (data not shown) The steady-state fluorescence signal(Ft) and the maximal fluorescence (Fm1015840) under illuminationwere measured setting the light measure at a frequency of10 kHz Fm1015840 was determined by a 1 s saturating light pulse(10 000120583mol photonssdotmminus2sdotsminus1)
The quantum yield of the PSII electron transport (ΦPSII)and nonphotochemical quenching (NPQ) were expressedaccording to Genty et al [42] and Bilger and Bjorkman [43]
After fluorescence determinations leaves were collectedfrom plants for the photosynthetic pigment content determi-nation namely chlorophylls and carotenoids Pigments wereextracted with a mortar and pestle in ice-cold 100 acetoneand quantified by a spectrophotometer (UV-VIS Cary 100Agilent Technologies USA) according to Lichtenthaler [44]
26 Statistical Analysis All results were subjected to statis-tical analysis using the SPSS statistical package (SPSS IncChicago IL USA) For the time course of plant height num-ber of leaves and leaf area interpolation equations (second-order polynomial curve) were calculated For anatomy andecophysiology data were subjected to ANOVA multiplecomparison tests were performed with Student-Newman-Keuls and Duncan coefficients using 119875 lt 005 as the level of
probability Data on sphericity convexity and percentage ofphenolics were transformed through arcsine function beforestatistical analysis
3 Results
31 Effect of Ionizing Radiation on Seed Germination Expo-sure of dry seeds to X-rays did not affect their percentageof germination which ranged between 978 and 100 withno significant differences among treatments Similarly X-raysdid not determine significant alterations in germination rateswhich ranged between 50 and 67 days
32 Effect of Ionizing Radiation on Plant Growth Plants fromboth control and irradiated seeds completed their crop cycleup to fruit ripening in about 100 DAS plants from irradiatedseeds did not show any apparent alteration in above-groundgrowth (Figure 1)
Figure 2 shows the effects of the seed irradiation treat-ments on plant height leaf number and plant leaf areaof the main stem The irradiation dose differently affectedthese growth parameters In plants of the nonirradiatedcontrol plant height reached the maximum value of 602 cmat approximately 71 DAS Irradiation of seeds with 03Gydid not significantly influence plant height Starting from71 DAS until the end of the cycle height of plants fromseeds irradiated with 10Gy was significantly higher than thatreached after irradiation with 20 50 and 100Gy Final heightof control plants and of those from seeds irradiated withX-ray doses up to 10Gy was significantly higher than thatreached after irradiation with 20Gy and above (Figure 2(a))
Theprogression of the number of leaves and leaf areawerenot consistent with the time course of plant height (Figure 2)Indeed in the control at the end of the crop cycle plantswere characterized by 97 leaves corresponding to 625 cm2leaf area Plants from irradiated seeds formed a number ofleaves significantly higher than control at the end of the cyclewith maximum value in plants from seeds irradiated with10Gy (Figure 2(b)) Similarly under all the radiation dosesbut 100Gy plants showed significantly larger leaf area thanthe control with maximum value at 03 Gy (Figure 2(c))
33 Effect of Ionizing Radiation on LeafAnatomy Microscopyanalysis of the leaf sections showed a regular anatomicalorganisation in both juvenile and compound leaves formedafter seed irradiation at different levels of X-rays Sincethe effect of the treatments was similar in both types ofleaves microscopy views are shown only for compoundleaves (Figure 3) Both leaf types were characterised by atypical dorsiventral structure which did not face disruptivestructural alterations after irradiation of seeds with X-raysMicroscopy observations of leaves from seeds irradiatedwith up to 20Gy did not reveal evident differences com-pared with control leaves (Figures 3(a)ndash3(d)) At 50 and100Gy sporadic perturbations in the mesophyll structurewere detectable (Figures 3(e)ndash3(h)) More specifically thepresence of more shrunk mesophyll cells (Figures 3(g) and3(h)) due to palisade cells collapsed in the lower portion
The Scientific World Journal 5
0
10
20
30
40
50
60
70
80
0 20 40 60 80 100
Plan
t hei
ght (
mm
)
DAS
(a)
0
5
10
15
20
Num
ber o
f lea
ves (
Np
lant
)
0 20 40 60 80 100DAS
(b)
0 20 40 60 80 100DAS
0
20
40
60
80
100
120
140
Plan
t lea
f are
a (cm
2pl
antminus1)
C03 Gy10 Gy
20 Gy50 Gy100 Gy
(c)
Figure 2 Time course of the plant height (a) number of leaves (b) and plant leaf area (c) in S lycopersicum L ldquoMicrotomrdquo as a function ofthe X-ray doses applied at the seed stage Mean values and standard errors are shown (119899 = 5) All second order polynomial curves had 1199032values above 097
(Figure 3(g)) and spongy cells characterised by more angularshape (Figure 3(h)) was found The occurrence of irregular-shaped cells of leaves from seeds irradiated with 50 and100Gy was confirmed by significant lower convexity valuesthan control and low-dose treatments (Tables 1 and 2) Theepidermis of compound leaves from irradiated seeds tendedto have less elongated cells (ie lower aspect ratio and highersphericity) while in the palisade parenchyma an oppositetrend was observed (Table 2) No specific tendency of cellshape variation was found in the juvenile leaves (Table 1)Themain veins did not show any irregularities in vessel walls afterany irradiation treatments (Figures 3(i) and 3(j)) Epifluores-cence microscopy showed the autofluorescence of phenoliccompounds which were mainly localised along chloroplast
membranes in both control leaves and those from irradiatedplants (Figures 4(a) and 4(b)) No differences in autofluores-cence distribution were observed in mesophyll cells of juve-nile and compound leaves from control and irradiated seedsThe percentage of phenolic compounds showed a nonsignifi-cant tendency to increase in juvenile leaves at low irradiationdoses followed by a significant decrease at 50 and 100Gy(Figure 4(c)) In compound leaves phenolic compoundstended to decrease after all irradiation treatments with thelowest values in leaves from seeds irradiated at 50 and 100Gy(Figure 4(d))
The other analysed anatomical parameters did not show aclear-cut dose-dependent response (Figure 5 Tables 1 and 2)In juvenile leaves the size of cells significantly decreased after
6 The Scientific World Journal
(b) (c)
(d) (e)
(f)
(g) (h)
(i)
(j)
(a)
Figure 3 Light microscopy views of cross sections of leaf lamina of S lycopersicum L ldquoMicrotomrdquo compound leaves from plants developedfrom control (a) and irradiated seeds 03 Gy (b) 10Gy (c) 20Gy (d) 50Gy (e g) and 100Gy (f h) Detail of xylem in the main vein is shownin leaves from control (i) and 100Gy irradiated seeds (j) Images (andashh) are at the same magnification Bar = 50 120583m
the irradiation of seeds only in palisade parenchyma withthe lowest values at 20Gy (Figures 5(c) 5(e) 5(g) and 5(i))By contrast in compound leaves the application of 100Gyresulted in increased cell size compared with control andleaves from seeds irradiated up to 50Gy (Figures 5(d) 5(f)5(h) and 5(j))
34 Effect of Ionizing Radiation on Fluorescence Measure-ments and Pigment Content No significant differences were
detectable in fluorescence parameters and total photosyn-thetic pigment content in plants grown from irradiated seedsat different X-rays doses (Figure 6) A tendency to NPQincrease was found at 03 and 10Gy followed by a decreaseat doses higher than 20Gy However the lack of significantdifferences in ΦPSII and NPQ among treatments indicatedthat X-rays delivered at the seed stage on tomato plantsdid not alter the capability of photosynthetic apparatus oftomato plants either to convert light energy in photochem-istry or to perform thermal dissipation mechanisms The
The Scientific World Journal 7
(a) (b)
Phen
olic
com
poun
ds (
)
aa a
aab
b
0
10
20
30
40
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf JL
(c)
Phen
olic
com
poun
ds (
) aab
abc ab
c bc
0
10
20
30
40
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf CL
(d)
Figure 4 Epifluorescence microscopy views of lamina cross sections of S lycopersicum L ldquoMicrotomrdquo compound leaves from plantsdeveloped from control (a) and 100Gy irradiated seeds (b) Phenolic compounds along chloroplast membranes are autofluorescent Percentof mesophyll occupied by phenolic compounds () is shown in (c) and (d) for juvenile (JL) and compound (CL) leaves respectively Meanvalues and standard errors are reported (119899 = 15) different letters correspond to significantly different values after multiple comparison tests(119875 lt 005)
maximum photochemical efficiency of PSII (FvFm) washigher in leaves from irradiated seeds than control anddid not show variations among different X-rays treatmentsthis indicated the maintenance of higher level of photo-synthetic activity after irradiation irrespectively from dose(Figure 6(c))
The total chlorophyll as well as the total carotenoidcontent did not show significant changes due to irradiationof seeds (Figure 7)
4 Discussion
Ionising radiation can have positive null or negative effectson plant development depending on the properties of theradiation the features of the plant material and possibleinteractions with other environmental factors [2] There iscommon agreement that plants are characterised by highradioresistance moreover low-LET ionising radiation hasbeen reported to increase germination percentage rootlength plant height and other growth parameters especiallyat low doses which induce hormesis [16 17 45ndash47] Ourresults suggest that S lycopersicum ldquoMicrotomrdquo is charac-terised by high radioresistance at the seed stage indeed
irradiation of dry seeds with a wide range of X-ray dosesdid not hamper germination and plant growth up to thecompletion of the crop cycle An hormesis effect was evidentsince growth either remained unaffected or was positivelyaffected especially by low doses of X-rays Apart from plantsgrown from seeds irradiated with 03 and 10Gy all plantsfrom irradiated seeds were characterised by a more compacthabitus higher number of larger leaves was not accompaniedby increased height The radiation-induced more compactgrowth pattern is positive in BLSSs in Spacewhere availabilityof volume for growth is a major technical constraint [7 4849] In the cultivar Microtom irradiation with doses equalto or higher than 20Gy caused a decrease in plant heightin agreement with the induction of dwarf or semidwarfgrowth reported as a general effect of exposure to ionisingradiation [18] Hence irradiation on the ldquoMicrotomrdquo seedswould intensify the trait of dwarf growth which alreadycharacterise this cultivar and should be taken into account forcultivar selection for the BLSSs [50] Any variations in growthparameters in Space-oriented ecologically closed systemscan determine valuable changes in terms of optimisationof volume and resource recycling These changes can bestraightaway converted into economic saving or waste [9]For example the increase in the number and size of leaves
8 The Scientific World Journal
Palisade parenchyma Spongy parenchyma
ab b ab a ab ab
a a a a a a
0010203040506
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf JL
Lam
ina t
hick
ness
(mm
)
(a)
Palisade parenchyma Spongy parenchyma
b b b b b a
a ab ab b abab
Leaf CL
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
010203040506
Lam
ina t
hick
ness
(mm
)
(b)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a a aa a
a
0200400600800
1000Upper epidermis
Cel
l are
a (120583
m2)
(c)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
1000
a a a a aa
Upper epidermis
Cel
l are
a (120583
m2)
(d)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
a bc bc c bc b
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(e)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
ab ab b ab ab a
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(f)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a aa
a a a
0200400600800
100012001400
Cel
l are
a (120583
m2)
(g)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
100012001400
b b b b ba
Cel
l are
a (120583
m2)
(h)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
a aa
a a a
0
200
400
600
800
Cel
l are
a (120583
m2)
(i)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
b b b b ba
0
200
400
600
800
Cel
l are
a (120583
m2)
(j)
Figure 5 Lamina thickness (a b) and cell area of upper epidermis (c d) palisade (e f) spongy (g h) parenchymas and lower epidermis (i j)of S lycopersicum L ldquoMicrotomrdquo leaves of control and irradiated plants at different doses of X-rays in both juvenile (JL) and compound (CL)leaves Mean values and standard errors are shown (119899 = 15 in (a b) 119899 = 45 in (cndashj)) Different letters correspond to significantly differentvalues according to multiple comparison tests (119875 lt 005)
The Scientific World Journal 9
04
05
06
07
08
a a a a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
ΦPS
II
(a)
00
02
04
06
08
NPQ a
a aa a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
(b)
070
075
080
085
090
a
b b b b b
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
FF
m
(c)
Figure 6 Quantum yield of PSII electron transport (ΦPSII a) nonphotochemical quenching (NPQ b) and maximal PSII photochemicalefficiency (FvFm c) in compound (CL) S lycopersicum L ldquoMicrotomrdquo leaves of control and plants from seeds irradiated at different dosesof X-rays Mean values and standard deviations are shown (119899 = 5) Different letters correspond to significantly different values according tomultiple comparison tests (119875 lt 005)
0
1
2
3
4
a
aa
a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Chl a
+ b
(mg g
minus1)
(a)
0
02
04
06
a
aa a
a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Car x
+ c
(mg g
minus1)
(b)
Figure 7 Total chlorophyll content (Chl a+b) (a) and total carotenoid content (Car x+c) (b) in compound (CL) S lycopersicum L ldquoMicrotomrdquoleaves of control and plants from seeds irradiated at different doses of X-rays Mean values and standard errors are shown (119899 = 5) Differentletters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
induced in ldquoMicrotomrdquo especially by low doses of irradiationcan be beneficial to atmosphere regeneration of pressurisedmodules provided that formed leaves do not show structuralaberrations which reduce their function
General anatomical structure of both juvenile and com-pound ldquoMicrotomrdquo leaves was only slightly perturbed inplants from seeds subjected to the highest levels of radiation
Clear-cut dose-depended tendencies of variations were notfound in lamina thickness and cell size and shape of thevarious tissues The formation of smaller palisade cells wasfound only in juvenile leaves of irradiated seeds while anincrease in cell size was found only in spongy parenchymaand lower epidermis of compound leaves from seeds irra-diated at 100Gy The diverse growth behaviour found in
10 The Scientific World Journal
Table 1 Cell shape in different tissues of juvenile leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown (119899 = 75)Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf JL Aspect ratio Sphericity Convexity
Upper epidermis
C 197 plusmn 016 a 0308 plusmn 0050 a 0926 plusmn 0021 ab03Gy 200 plusmn 013 a 0290 plusmn 0038 a 0941 plusmn 0013 a10Gy 210 plusmn 012 a 0239 plusmn 0037 a 0898 plusmn 0016 ab20Gy 217 plusmn 014 a 0258 plusmn 0045 a 0909 plusmn 0019 abc50Gy 170 plusmn 008 a 0376 plusmn 0037 a 0927 plusmn 0016 ab100Gy 197 plusmn 014 a 0286 plusmn 0042 a 0876 plusmn 0028 c
Palisade parenchyma
C 290 plusmn 010 a 0109 plusmn 0015 a 0879 plusmn 0010 a03 Gy 304 plusmn 012 a 0099 plusmn 0016 a 0805 plusmn 0016 b10Gy 309 plusmn 013 a 0093 plusmn 0019 a 0792 plusmn 0015 b20Gy 283 plusmn 011 a 0101 plusmn 0017 a 0784 plusmn 0017 b50Gy 314 plusmn 014 a 0082 plusmn 0016 a 0772 plusmn 0016 b100Gy 275 plusmn 011 a 0119 plusmn 0020 a 0805 plusmn 0017 b
Spongy parenchyma
C 190 plusmn 008 a 0307 plusmn 0033 a 0893 plusmn 0012 a03 Gy 180 plusmn 007 a 0341 plusmn 0029 a 0876 plusmn 0013 a10Gy 203 plusmn 010 a 0290 plusmn 0031 a 0901 plusmn 0012 a20Gy 194 plusmn 012 a 0344 plusmn 0040 a 0881 plusmn 0015 a50Gy 174 plusmn 009 a 0393 plusmn 0035 a 0883 plusmn 0014 a100Gy 198 plusmn 012 a 0322 plusmn 0037 a 0878 plusmn 0015 a
Lower epidermis
C 178 plusmn 009 bc 0343 plusmn 0045 ab 0919 plusmn 0019 a03 Gy 225 plusmn 017 ab 0263 plusmn 0044 b 0933 plusmn 0024 a10Gy 219 plusmn 011 abc 0218 plusmn 0034 b 0888 plusmn 0020 ab20Gy 180 plusmn 013 bc 0378 plusmn 0047 ab 0926 plusmn 0017 a50Gy 167 plusmn 001 c 0461 plusmn 0051 a 0924 plusmn 0021 a100Gy 238 plusmn 021 a 0257 plusmn 0057 b 0857 plusmn 0030 b
the various tissues of structures of different age is in agree-ment with the different light-mediated control of cell devel-opment in the various leaf tissues [51] Altered mesophyllcharacterized by partly collapsed cells with irregular cellshape was found only sporadically after the exposure of seedsto 50 and 100Gy in theMicrotomcultivar Suchmodificationswere less marked than those found in leaves of bean wherethe target organs for irradiation were the leaves themselves[30] leaves directly exposed to X-rays in bean showed theformation of more loose parenchyma with larger cells likelydue to a radiation-induced damage to cell walls that reducedmechanical constraints to cell enlargement [30 52] Furtherinvestigations on cell wall composition would be useful toelucidate whether a possible alteration of cell walls wasresponsible for cell enlargement in the spongy parenchymaand epidermis of leaves from seeds of ldquoMicrotomrdquo irradiatedwith the highest dose of X-rays However in the cultivarMicrotom the slight alterations of mesophyll structure didnot impair photosynthetic efficiency as demonstrated by thelack of significant dose-dependent variation trends in thequantum yield of PSII electron transport (ΦPSII) nonphoto-chemical quenching (NPQ) and maximal PSII photochemi-cal efficiency (FvFm) The increased values of FvFm closeto 08 which is considered an optimal value for healthy plants[53] suggested that the photosynthetic apparatus of the leavesfrom irradiated seeds has higher levels of photochemical
efficiency irrespective of dose The unchanged or increasedcontent of both chlorophyll and carotenoid pigments con-firmed that the photosynthetic apparatus remained stablealso in terms of light harvesting capacity and indicatedno deleterious effect of radiation on plant photosyntheticmachinery RecentlyMarcu et al [17] showed that irradiationof lettuce seeds with gamma rays up to 30Gy enhancedthe content of chlorophyll a chlorophyll b and carotenoidswhile higher doses (up to 70Gy) resulted in their decreaseHowever the effect of ionising radiation on photosyntheticpigment content is quite controversial Increased pigmentcontent is ascribed to the phenomenon of hormesis whilea lower content is generally ascribed to the inhibition ofchlorophyll and carotenoid synthesis or to oxidation due tothe high levels of radiation-induced water radiolysis [2 1454ndash56] Some plants possess natural screens against radiationwhen their tissues contain phenolic compounds with antioxi-dant functions and can counteract photoinhibitory processes[29 57] An increase of phenolic compounds linked to thechloroplast membranes has been recently found in Phaseolusvulgaris irradiated leaves exposed to high levels of X-rays[28 30] Changes in phenolic content after the exposure toSpace can be attributed either to direct effects on the synthesisor mobilization of phenolic compounds or to indirect effectsdue to changes in ultrastructural organization such as theincrease in the number of chloroplasts per cell [30 58]
The Scientific World Journal 11
Table 2 Cell shape in different tissues of compound leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown(119899 = 75) Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf CL Aspect ratio Sphericity Convexity
Upper epidermis
C 166 plusmn 007 c 0422 plusmn 0042 a 0946 plusmn 0010 a03 Gy 216 plusmn 017 ab 0244 plusmn 0033 c 0925 plusmn 0016 ab10Gy 171 plusmn 011 bc 0401 plusmn 0042 a 0914 plusmn 0013 b20Gy 186 plusmn 010 abc 0328 plusmn 0041 ab 0919 plusmn 0020 ab50Gy 225 plusmn 018 a 0247 plusmn 0041 c 0897 plusmn 0019 b100Gy 188 plusmn 009 abc 0319 plusmn 0045 ab 0902 plusmn 0016 b
Palisade parenchyma
C 355 plusmn 012 a 0073 plusmn 0014 b 0890 plusmn 0012 a03 Gy 289 plusmn 010 b 0102 plusmn 0013 ab 0847 plusmn 0018 b10Gy 300 plusmn 014 b 0104 plusmn 0021 ab 0826 plusmn 0017 b20Gy 302 plusmn 011 b 0094 plusmn 0013 ab 0810 plusmn 0014 b50Gy 283 plusmn 012 b 0107 plusmn 0021 ab 0750 plusmn 0021 c100Gy 275 plusmn 012 b 0122 plusmn 0023 a 0809 plusmn 0015 b
Spongy parenchyma
C 193 plusmn 010 a 0323 plusmn 0035 a 0904 plusmn 0011 a03 Gy 183 plusmn 010 a 0375 plusmn 0042 a 0879 plusmn 0013 a10Gy 176 plusmn 007 a 0357 plusmn 0034 a 0869 plusmn 0011 a20Gy 180 plusmn 009 a 0366 plusmn 0038 a 0885 plusmn 0016 a50Gy 171 plusmn 006 a 0372 plusmn 0034 a 0879 plusmn 0015 a100Gy 188 plusmn 010 a 0339 plusmn 0037 a 0883 plusmn 0015 a
Lower epidermis
C 148 plusmn 008 b 0545 plusmn 0045 a 0957 plusmn 0015 a03 Gy 210 plusmn 017 a 0295 plusmn 0046 b 0929 plusmn 0019 ab10Gy 190 plusmn 012 ab 0334 plusmn 0043 b 0893 plusmn 0019 b20Gy 178 plusmn 011 ab 0388 plusmn 0047 b 0929 plusmn 0017 ab50Gy 172 plusmn 009 ab 0367 plusmn 0040 b 0914 plusmn 0016 b100Gy 191 plusmn 014 ab 0362 plusmn 0054 b 0888 plusmn 0025 b
The lowering of phenolic content at high levels of X-rays inldquoMicrotomrdquo leaves is not in agreement with general trendsindicating enhanced phenolics production in plants grownin Space [58] or in various plant tissues directly irradiatedwith gamma rays [59] However the low content of phenolicscan be an indirect effect of increasing volume occupiedby intercellular spaces due to the loosening of parenchymacells
In conclusion our results show that irradiation of the dryseeds of the Microtom cultivar did not hamper germinationand the development of functional leaves The formationof more compact plants characterised by a higher numberof larger leaves can be valuable in terms of increasingresource regeneration in the BLSSs High levels of radiationinduced only slight structural perturbations which weresimilar in juvenile and compound leaves Such perturbationsdid not affect photosynthetic efficiency since the maximalPSII photochemical efficiency was even increased after irra-diation at all doses However the tendency to accumulatelower amounts of phenolic compounds along chloroplastmembranes after irradiation of seeds with high doses ofradiation should be further investigated because it coulddetermine the reduction of natural photoprotection [57]Finally the overall results indicate that theMicrotom cultivarirradiated at the seed stage has high radioresistance duringthe growth which is a desirable feature for plant cultivation
in Space Radioresistance together with dwarf growth andshort life cycle makes this cultivar a valuable candidate forthe cultivation in BLSSs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are grateful to the ldquoCompagnia di San Paolordquo(Turin) and the ldquoPolo delle Scienze e delle Tecnologie perla Vitardquo of the University of Naples Federico II for thefinancial support The authors also thank Maria Giordanoand Pasquale Scognamiglio for their support in plant culti-vation and microscopy analyses The authors acknowledgefour anonymous editors for helpful comments on a previousversion of the paper
References
[1] A Real S Sundell-Bergman J F Knowles D S Woodheadand I Zinger ldquoEffects of ionising radiation exposure on plantsfish and mammals relevant data for environmental radiation
12 The Scientific World Journal
protectionrdquo Journal of Radiological Protection vol 24 no 4 ppA123ndashA137 2004
[2] V De Micco C Arena D Pignalosa and M Durante ldquoEffectsof sparsely and densely ionizing radiation on plantsrdquo Radiationand Environmental Biophysics vol 50 no 1 pp 1ndash19 2011
[3] N Kumari P Kumar D Mitra B Prasad B N Tiwary andL Varshney ldquoEffects of ionizing radiation on microbial decon-tamination phenolic contents and antioxidant properties oftriphalardquo Journal of Food Science vol 74 no 3 pp M109ndashM1132009
[4] A Tanaka N Shikazono and Y Hase ldquoStudies on biologicaleffects of ion beams on lethality molecular nature of mutationmutation rate and spectrum of mutation phenotype for muta-tion breeding in higher plantsrdquo Journal of Radiation Researchvol 51 no 3 pp 223ndash233 2010
[5] M Durante and A Kronenberg ldquoGround-based research withheavy ions for space radiation protectionrdquo Advances in SpaceResearch vol 35 no 2 pp 180ndash184 2005
[6] E Hu S I Bartsev and H Liu ldquoConceptual design of a biore-generative life support system containing crops and silkwormsrdquoAdvances in Space Research vol 45 no 7 pp 929ndash939 2010
[7] V deMicco G Aronne G Colla R Fortezza and S de PascaleldquoAgro-biology for bioregenerative life support systems in long-term space missions general constraints and the Italian effortsrdquoJournal of Plant Interactions vol 4 no 4 pp 241ndash252 2009
[8] T W Halstead and F R Dutcher ldquoPlants in spacerdquo AnnualReview of Plant Physiology vol 38 pp 317ndash345 1987
[9] V De Micco S De Pascale R Paradiso and G AronneldquoMicrogravity effects on different stages of higher plant life cycleand completion of the seed-to-seed cyclerdquo Plant Biology vol 16pp 31ndash38 2014
[10] C J Bennett C Pirim and T M Orlando ldquoSpace-weatheringof solar system bodies a laboratory perspectiverdquo ChemicalReviews vol 113 pp 9086ndash9150 2013
[11] R A Kerr ldquoRadiation will make astronautsrsquo trip to Mars evenriskierrdquo Science vol 340 article 1031 2014
[12] D M Hassler C Zeitlin R F Wimmer-Schweingruber et alldquoMarsrsquo surface radiation environment measured with the MarsScience Laboratoryrsquos Curiosity Roverrdquo Science vol 343 no6169 Article ID 1244797 2014
[13] M Durante and F A Cucinotta ldquoHeavy ion carcinogenesis andhuman space explorationrdquo Nature Reviews Cancer vol 8 no 6pp 465ndash472 2008
[14] C Arena V De Micco and A De Maio ldquoGrowth alterationand leaf biochemical responses in P vulgaris plants exposed todifferent doses of ionizing radiationrdquo Plant Biology vol 16 pp194ndash202 2014
[15] R W Holst and D J Nagel ldquoRadiation effects on plantsrdquo inPlants For Environmental Studies W Wang J W Gorsuch andJ S Hughes Eds pp 37ndash81 Lewis Publishers Boca Raton FlaUSA 1997
[16] T D Luckey Hormesis With Ionizing Radiation CRC PressBoca Raton Fla USA 1980
[17] D Marcu V Cristea and L Daraban ldquoDose-dependent effectsof gamma radiation on lettuce (Lactuca sativa var capitata)seedlingsrdquo International Journal of Radiation Biology vol 89 pp219ndash223 2013
[18] M Mei H Deng Y Lu et al ldquoMutagenic effects of heavy ionradiation in plantsrdquo Advances in Space Research vol 14 no 10pp 363ndash372 1994
[19] Y LiM Liu Z Cheng andY Sun ldquoSpace environment inducedmutations prefer to occur at polymorphic sites of rice genomesrdquoAdvances in Space Research vol 40 no 4 pp 523ndash527 2007
[20] D S Kim J-B Kim E J Goh et al ldquoAntioxidant response ofArabidopsis plants to gamma irradiation genome-wide expres-sion profiling of the ROS scavenging and signal transductionpathwaysrdquo Journal of Plant Physiology vol 168 no 16 pp 1960ndash1971 2011
[21] T J Brodribb ldquoXylem hydraulic physiology the functionalbackbone of terrestrial plant productivityrdquo Plant Science vol177 no 4 pp 245ndash251 2009
[22] R M Wheeler J C Sager R P Prince andW M Knott ldquoCropproduction for advanced life support systemsmdashobservationfrom the Kennedy Space Center Breadboard Projectrdquo NASATM-2003-211184 NASA Kennedy Space Center Florida 2003
[23] R M Wheeler C L Mackowiak G W Stutte et al ldquoCropproductivities and radiation use efficiencies for bioregenerativelife supportrdquo Advances in Space Research vol 41 no 5 pp 706ndash713 2008
[24] R Paradiso V De Micco R Buonomo G Aronne G Bar-bieri and S De Pascale ldquoSoilless cultivation of soybean forBioregenerative Life-Support Systems a literature review andthe experience of the MELiSSA ProjectmdashFood characterisationPhase Irdquo Plant Biology vol 16 pp 69ndash78 2014
[25] D M Porterfield ldquoThe biophysical limitations in physiologicaltransport and exchange in plants grown in microgravityrdquoJournal of Plant Growth Regulation vol 21 no 2 pp 177ndash1902002
[26] K Yokthongwattana and A Melis ldquoPhotoinhibition and recov-ery in oxygenic photosynthesis mechanism of a photosystem IIdamage and repair cyclerdquo in Photoprotection PhotoinhibitionGene Regulation and Environment B Demmig-Adams WW Adams III and A K Mattoo Eds pp 175ndash191 SpringerDordecht The Netherlands 2006
[27] M T Giardi G Rea M D Lambreva et al ldquoMutations ofphotosystem II D1 protein that empower efficient phenotypesof Chlamydomonas reinhardtii under extreme environment inSpacerdquo PLoS ONE vol 8 Article ID e64352 2013
[28] C Arena V De Micco G Aronne M Pugliese A Virzo DeSanto andADeMaio ldquoResponse ofPhaseolus vulgarisL plantsto low-let ionizing radiation growth and oxidative stressrdquo ActaAstronautica vol 91 pp 107ndash114 2013
[29] V Lattanzio P A Kroon and S Quideau ldquoPlant phenolicsmdashsecondary metabolites with diverse functionsrdquo in RecentAdvances in Polyphenol Research F Daay and V Lattanzio Edspp 1ndash35 Wiley-Blackwell Oxford UK 2008
[30] V De Micco C Arena and G Aronne ldquoAnatomical alterationsof Phaseolus vulgaris L mature leaves irradiated with X-raysrdquoPlant Biology vol 16 pp 187ndash193 2014
[31] A D Krikorian ldquoPlants and somatic embryos in space whathave we learnedrdquo Gravitational and Space Biology Bulletin vol11 no 2 pp 5ndash14 1998
[32] R Meissner Y Jacobson S Melamed et al ldquoA new modelsystem for tomato geneticsrdquoThe Plant Journal vol 12 no 6 pp1465ndash1472 1997
[33] E Emmanuel and A A Levy ldquoTomato mutants as tools forfunctional genomicsrdquo Current Opinion in Plant Biology vol 5no 2 pp 112ndash117 2002
[34] E Martı C Gisbert G J Bishop M S Dixon and J LGarcıa-Martınez ldquoGenetic and physiological characterizationof tomato cv Micro-Tomrdquo Journal of Experimental Botany vol57 pp 2037ndash2047 2006
The Scientific World Journal 13
[35] T Kurimoto J V H Constable and A Huda ldquoEffects ofionizing radiation exposure on arabidopsis thalianardquo HealthPhysics vol 99 no 1 pp 49ndash57 2010
[36] L Reale D Gigante F Landucci F Ferranti and R VenanzonildquoMorphological and histo-anatomical traits reflect die-back inPhragmites australis (Cav) Steudrdquo Aquatic Botany vol 103 pp122ndash128 2012
[37] K Fukuzawa ldquoUltraviolet microscopyrdquo in Methods in LigninChemistry S Y Lin andCWDence Eds pp 110ndash131 SpringerBerlin Germany 1992
[38] S E Ruzin Plant Microtechnique and Microscopy OxfordUniversity Press New York NY USA 1999
[39] V De Micco G Aronne J-P Joseleau and K Ruel ldquoXylemdevelopment and cell wall changes of soybean seedlings grownin spacerdquo Annals of Botany vol 101 no 5 pp 661ndash669 2008
[40] S Van Buggenhout T Grauwet A Van Loey and M Hen-drickx ldquoStructureprocessing relation of vacuum infused straw-berry tissue frozen under different conditionsrdquo European FoodResearch and Technology vol 226 no 3 pp 437ndash448 2008
[41] V De Micco and G Aronne ldquoBiometric anatomy of seedlingsdeveloped onboard of Foton-M2 in an automatic system sup-porting growthrdquoActa Astronautica vol 62 no 8-9 pp 505ndash5132008
[42] B Genty J M Briantais and N R Baker ldquoThe relationshipbetween quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescencerdquo Biochimica andBiophysica Acta vol 990 pp 87ndash92 1989
[43] W Bilger and O Bjorkman ldquoRole of the xanthophyll cycle inphotoprotection elucidated by measurements of light-inducedabsorbance changes fluorescence and photosynthesis in leavesof Hedera canariensisrdquo Photosynthesis Research vol 25 no 3pp 173ndash185 1990
[44] H K Lichtenthaler ldquoChlorophylls and carotenoids pigmentsof photosynthetic biomembranesrdquoMethods in Enzymology vol148 pp 350ndash382 1987
[45] J P Maity DMishra A Chakraborty A Saha S C Santra andS Chanda ldquoModulation of some quantitative and qualitativecharacteristics in rice (Oryza sativa L) and mung (Phaseolusmungo L) by ionizing radiationrdquo Radiation Physics and Chem-istry vol 74 no 5 pp 391ndash394 2005
[46] L Cheng H Yang B Lin et al ldquoEffect of gamma-ray radiationon physiological morphological characters and chromosomeaberrations of minitubers in Solanum tuberosum Lrdquo Interna-tional Journal of Radiation Biology vol 86 no 9 pp 791ndash7992010
[47] B Al-Safadi and R Elias ldquoImprovement of caper (Capparisspinosa L) propagation using in vitro culture and gammairradiationrdquo Scientia Horticulturae vol 127 no 3 pp 290ndash2972011
[48] O Monje G W Stutte G D Goins D M Porterfield and GE Bingham ldquoFarming in space environmental and biophysicalconcernsrdquo Advances in Space Research vol 31 no 1 pp 151ndash1672003
[49] V De Micco M Scala and G Aronne ldquoEffects of simulatedmicrogravity on male gametophyte of Prunus Pyrus andBrassica speciesrdquo Protoplasma vol 228 no 1ndash3 pp 121ndash1262006
[50] V De Micco R Buonomo R Paradiso S De Pascale and GAronne ldquoSoybean cultivar selection for Bioregenerative LifeSupport Systems (BLSS)mdashtheoretical selectionrdquo Advances inSpace Research vol 49 no 10 pp 1415ndash1421 2012
[51] T Kozuka S-G Kong M Doi K-I Shimazaki and ANagatani ldquoTissue-autonomous promotion of palisade celldevelopment by phototropin 2 in Arabidopsisrdquo Plant Cell vol23 no 10 pp 3684ndash3695 2011
[52] E Kovacs and A Keresztes ldquoEffect of gamma and UV-BCradiation on plant cellsrdquoMicron vol 33 no 2 pp 199ndash210 2002
[53] R Ogaya J Penuelas D Asensio and J Llusia ldquoChlorophyllfluorescence responses to temperature and water availabilityin two co-dominant Mediterranean shrub and tree speciesin a long-term field experiment simulating climate changerdquoEnvironmental and Experimental Botany vol 73 no 1 pp 89ndash93 2011
[54] R Zaka C M Vandecasteele and M T Misset ldquoEffects oflow chronic doses of ionizing radiation on antioxidant enzymesand G6PDH activities in Stipa capillata (Poaceae)rdquo Journal ofExperimental Botany vol 53 no 376 pp 1979ndash1987 2002
[55] J-H KimM-H Baek Y C Byung GW Seung and J-S KimldquoAlterations in the photosynthetic pigments and antioxidantmachineries of red pepper (Capsicum annuum L) seedlingsfrom gamma-irradiated seedsrdquo Journal of Plant Biology vol 47no 4 pp 314ndash321 2004
[56] N A Al-Enezi and J M Al-Khayri ldquoAlterations of DNA ionsand photosynthetic pigments content in date palm seedlingsinduced by X-irradiationrdquo International Journal of Agricultureand Biology vol 14 pp 329ndash336 2012
[57] V de Micco C Arena L Vitale G Aronne and A V de SantoldquoAnatomy and photochemical behaviour of mediterranean Cis-tus incanus winter leaves under natural outdoor and warmerindoor conditionsrdquo Botany vol 89 no 10 pp 677ndash688 2011
[58] L H Levine H G Levine E C Stryjewski V Prima and WC Piastuch ldquoEffect of spaceflight on isoflavonoid accumulationin etiolated soybean seedlingsrdquo Journal of Gravitational Physiol-ogy vol 8 no 2 pp 21ndash27 2001
[59] X Fan ldquoAntioxidant capacity of fresh-cut vegetables exposedto ionizing radiationrdquo Journal of the Science of Food andAgriculture vol 85 no 6 pp 995ndash1000 2005
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

The Scientific World Journal 3
experiments to evaluate the effect of radiation on biologicalsystems [13 14 28 30] The highest doses (50Gy and 100Gy)are generally applied since they can be considered as positivecontrols as there ismore likely a plant reaction at these dosesX-rays were delivered as one dose per each Petri dish [35] inorder to avoid cumulative effects on the same sample X-rayswere produced by a Thomson tube (TR 300 F 250 kVp Sta-bilipan Siemens Forchheim Germany) with tungsten cath-ode filtered by 1mm thick copper foil and with 15mA anodiccurrent Before irradiation a physical dosimetry was per-formed by using an ionization chamber (VictoreenModlingAustria) The X-ray intensity was measured at the estab-lished distance where seeds were positioned under the X-raytube
After the exposure to X-rays (on 24 April) the irradiatedseeds and controls were placed into Petri dishes on three lay-ers of filter paper moistened with distilled water Petri disheswere incubated in the dark at temperature of 20∘C andmoni-tored daily to analyse germination percentage and rate (con-sidered as the number of days taken to reach the maximumgermination percentage) Seeds were classified as germinatedwhen the emerging root grew as long as the seed maximumdiameter
22 Growth Conditions The growth experiment was carriedout in a greenhouse with a polyethylene film cover Thegreenhouse was equipped with a black plastic shading net(70 shading) in order to keep the temperature and lightintensity close to the levels feasible in test bed evaluationof BLSSs technologies [22 23] Wavelength composition ofthe sunlight including the whole PAR range spectrum forphotosynthetic activity was not affected
Seedlings were transplanted into pots (4 cm diameter) onpeat-based compost (peat soil 1 1 in volume) and trans-ferred to the greenhouse 5 days after sowing (DAS) Thenthey were repotted in 10 cm pots using the same substrateat the stadium of fourth true leaf (ie two juvenile and twocompound leaves) at 22DAS In the cultivar Microtom thefirst two true leaves maintain juvenile traits and are charac-terised by simple morphology they are trifoliate with littleor not evident lobes Successive leaves show more complexmorphology having pennate-compound lamina with highlyindented lobes
During the whole growth period plants were irrigatedwith tapwater at 2-day interval in order to reach the containercapacity (till the beginning of drainage)
The temperature inside the greenhouse was recordedevery 10 minutes by means of a data logger (Tynitag Ultra2Gemini Data Loggers Chichester UK) Photosynthetic pho-ton flux density (PPFD) at the canopy level was recordeddaily at 1200 pm with a Delta OHMDO9847multifunctionmeter
The whole crop cycle (up to fruit ripening) lasted from 24April until 6 August 2013
During the whole growth cycle the mean values of tem-perature inside the greenhouse were 284216∘C (daynight)and the light intensity recorded at noon at the canopy levelwas 446 120583mol mminus2 sminus1 on average
23 Biometric and Microscopy Analyses Biometric analyseswere performed once a week on 10 plants per treatmentthroughout the crop cycle Plant height and number of leaveswere recorded Moreover single leaves of the main stem werephotographed with a digital camera (Nikon D3100 NikonEurope BV) in order to examine leaf area increments withthe software program AnalySIS 32 (Olympus HamburgGermany)
From 5 plants per treatment the second juvenile leaf(JL) and the first compound leaf (CL) were collected whenthey were fully developed One median leaflet (one of thetwo opposite leaflets below the apex) per leaf was used formicroscopy analyses while the others were used for pigmentextraction
Leaflets destined to microscopy analyses were immedi-ately fixed in FAA (40 formaldehydeglacial acetic acid50ethanol-5590 by volume) for several days Leaflets werecut under a dissection microscope (ZSX9 Olympus) toobtain subsamples of 5 times 5mm in the middle of laminaincluding the main vein Subsamples were dehydrated inan ethanol series and embedded in the acrylic resin JB4(Polysciences Warrington PA USA) Semithin cross sec-tions (5 120583m thick) were cut through a rotative microtomeSections were stained with 0025 Toluidine blue in 01Mcitrate buffer at pH 4 [36] mounted with Canadian Balsamand observed under a light microscope (BX60 Olympus)Unstained sections were mounted with mineral oil for fluo-rescence and observed under an epifluorescence microscope(BX60 Olympus) equipped with a mercury lamp band-passfilter 330ndash385 nm dichromatic mirror 400 nm and aboveand barrier filter 420 nm and above With these filters it waspossible to detect the presence of simple phenolics that areautofluorescent at such wavelengths [37 38] Three sectionsper leaflet were analysed and images were collected by meansof a digital camera (CAMEDIA C4040 Olympus) at variousmagnifications
24 Digital Image Analysis All digital images were analysedwith the AnalySIS 32 (Olympus) software for image analysisThe thickness of the palisade and the spongy parenchymaswere measured in five regions along the leaf lamina Thecell area and shape of upper and lower epidermis palisadeand spongy parenchyma were quantified in 15 cells pereach tissue per section More specifically cell shape wascharacterised as (a) aspect ratio (maximum widthheightratio of a bounding rectangle for the cell defining how itis elongated) (b) sphericity (roundness of a particle withspherical particles having a maximum value of 1) and (c)convexity (the fraction of the cellrsquos surface area and thearea of its convex a turgid cell has a maximum value of1) [39 40] The cell area occupied by phenolic compoundswas measured in 3 regions (150 times 200 120583m2 each) of themesophyll per section selected avoiding veins Thus thearea occupied by phenolic compounds was calculated asthe percentage of tissuepicture occupied by compoundsappearing autofluorescent at the above-reported filter settings[41]
4 The Scientific World Journal
C 100 Gy50 Gy20 Gy10 Gy03 Gy
Figure 1 Plants of S lycopersicum L ldquoMicrotomrdquo developed fromcontrol and irradiated seeds at 40 DAS (Days After Sowing) Plantsfrom seeds exposed to increasing doses of X-rays (from 0 to 100Gy)are shown from left to right
25 Fluorescence Measurements and Pigment Content Flu-orescence measurements and pigment extraction were con-ducted on 5 pennate-compound leaves of 5 plants subjectedto each irradiation treatment and in controls at 40 DAS
Chlorophyll a fluorescence measurements were carriedout by means of a pulse amplitude modulated fluorometer(Junior-PAM Walz Germany) equipped with a monitoringLeaf-Clip JUNIOR-B (Walz Germany) On 30min dark-adapted leaves the background fluorescence signal Fo wasinduced by internal light provided by a blue LED of about2-3 120583mol photonssdotmminus2sdotsminus1 at a frequency of 05 kHz Themaximal fluorescence level in the dark-adapted state (Fm)was measured by 1 s saturating light pulse (10 000120583molphotonssdotmminus2sdotsminus1) at a frequency of 10 kHz the maximal PSIIphotochemical efficiency (FvFm) was calculated as FvFm =(FmminusFo)FmThemeasurements in the light were carried outby exposing each leaf to a PPFD of 420120583mol photonssdotmminus2sdotsminus1for 5min This level of PPFD was chosen because it fallsin the range of maximal quantum yield for the Microtomcultivar as indicated by fluorescence fast-response curves tolight (data not shown) The steady-state fluorescence signal(Ft) and the maximal fluorescence (Fm1015840) under illuminationwere measured setting the light measure at a frequency of10 kHz Fm1015840 was determined by a 1 s saturating light pulse(10 000120583mol photonssdotmminus2sdotsminus1)
The quantum yield of the PSII electron transport (ΦPSII)and nonphotochemical quenching (NPQ) were expressedaccording to Genty et al [42] and Bilger and Bjorkman [43]
After fluorescence determinations leaves were collectedfrom plants for the photosynthetic pigment content determi-nation namely chlorophylls and carotenoids Pigments wereextracted with a mortar and pestle in ice-cold 100 acetoneand quantified by a spectrophotometer (UV-VIS Cary 100Agilent Technologies USA) according to Lichtenthaler [44]
26 Statistical Analysis All results were subjected to statis-tical analysis using the SPSS statistical package (SPSS IncChicago IL USA) For the time course of plant height num-ber of leaves and leaf area interpolation equations (second-order polynomial curve) were calculated For anatomy andecophysiology data were subjected to ANOVA multiplecomparison tests were performed with Student-Newman-Keuls and Duncan coefficients using 119875 lt 005 as the level of
probability Data on sphericity convexity and percentage ofphenolics were transformed through arcsine function beforestatistical analysis
3 Results
31 Effect of Ionizing Radiation on Seed Germination Expo-sure of dry seeds to X-rays did not affect their percentageof germination which ranged between 978 and 100 withno significant differences among treatments Similarly X-raysdid not determine significant alterations in germination rateswhich ranged between 50 and 67 days
32 Effect of Ionizing Radiation on Plant Growth Plants fromboth control and irradiated seeds completed their crop cycleup to fruit ripening in about 100 DAS plants from irradiatedseeds did not show any apparent alteration in above-groundgrowth (Figure 1)
Figure 2 shows the effects of the seed irradiation treat-ments on plant height leaf number and plant leaf areaof the main stem The irradiation dose differently affectedthese growth parameters In plants of the nonirradiatedcontrol plant height reached the maximum value of 602 cmat approximately 71 DAS Irradiation of seeds with 03Gydid not significantly influence plant height Starting from71 DAS until the end of the cycle height of plants fromseeds irradiated with 10Gy was significantly higher than thatreached after irradiation with 20 50 and 100Gy Final heightof control plants and of those from seeds irradiated withX-ray doses up to 10Gy was significantly higher than thatreached after irradiation with 20Gy and above (Figure 2(a))
Theprogression of the number of leaves and leaf areawerenot consistent with the time course of plant height (Figure 2)Indeed in the control at the end of the crop cycle plantswere characterized by 97 leaves corresponding to 625 cm2leaf area Plants from irradiated seeds formed a number ofleaves significantly higher than control at the end of the cyclewith maximum value in plants from seeds irradiated with10Gy (Figure 2(b)) Similarly under all the radiation dosesbut 100Gy plants showed significantly larger leaf area thanthe control with maximum value at 03 Gy (Figure 2(c))
33 Effect of Ionizing Radiation on LeafAnatomy Microscopyanalysis of the leaf sections showed a regular anatomicalorganisation in both juvenile and compound leaves formedafter seed irradiation at different levels of X-rays Sincethe effect of the treatments was similar in both types ofleaves microscopy views are shown only for compoundleaves (Figure 3) Both leaf types were characterised by atypical dorsiventral structure which did not face disruptivestructural alterations after irradiation of seeds with X-raysMicroscopy observations of leaves from seeds irradiatedwith up to 20Gy did not reveal evident differences com-pared with control leaves (Figures 3(a)ndash3(d)) At 50 and100Gy sporadic perturbations in the mesophyll structurewere detectable (Figures 3(e)ndash3(h)) More specifically thepresence of more shrunk mesophyll cells (Figures 3(g) and3(h)) due to palisade cells collapsed in the lower portion
The Scientific World Journal 5
0
10
20
30
40
50
60
70
80
0 20 40 60 80 100
Plan
t hei
ght (
mm
)
DAS
(a)
0
5
10
15
20
Num
ber o
f lea
ves (
Np
lant
)
0 20 40 60 80 100DAS
(b)
0 20 40 60 80 100DAS
0
20
40
60
80
100
120
140
Plan
t lea
f are
a (cm
2pl
antminus1)
C03 Gy10 Gy
20 Gy50 Gy100 Gy
(c)
Figure 2 Time course of the plant height (a) number of leaves (b) and plant leaf area (c) in S lycopersicum L ldquoMicrotomrdquo as a function ofthe X-ray doses applied at the seed stage Mean values and standard errors are shown (119899 = 5) All second order polynomial curves had 1199032values above 097
(Figure 3(g)) and spongy cells characterised by more angularshape (Figure 3(h)) was found The occurrence of irregular-shaped cells of leaves from seeds irradiated with 50 and100Gy was confirmed by significant lower convexity valuesthan control and low-dose treatments (Tables 1 and 2) Theepidermis of compound leaves from irradiated seeds tendedto have less elongated cells (ie lower aspect ratio and highersphericity) while in the palisade parenchyma an oppositetrend was observed (Table 2) No specific tendency of cellshape variation was found in the juvenile leaves (Table 1)Themain veins did not show any irregularities in vessel walls afterany irradiation treatments (Figures 3(i) and 3(j)) Epifluores-cence microscopy showed the autofluorescence of phenoliccompounds which were mainly localised along chloroplast
membranes in both control leaves and those from irradiatedplants (Figures 4(a) and 4(b)) No differences in autofluores-cence distribution were observed in mesophyll cells of juve-nile and compound leaves from control and irradiated seedsThe percentage of phenolic compounds showed a nonsignifi-cant tendency to increase in juvenile leaves at low irradiationdoses followed by a significant decrease at 50 and 100Gy(Figure 4(c)) In compound leaves phenolic compoundstended to decrease after all irradiation treatments with thelowest values in leaves from seeds irradiated at 50 and 100Gy(Figure 4(d))
The other analysed anatomical parameters did not show aclear-cut dose-dependent response (Figure 5 Tables 1 and 2)In juvenile leaves the size of cells significantly decreased after
6 The Scientific World Journal
(b) (c)
(d) (e)
(f)
(g) (h)
(i)
(j)
(a)
Figure 3 Light microscopy views of cross sections of leaf lamina of S lycopersicum L ldquoMicrotomrdquo compound leaves from plants developedfrom control (a) and irradiated seeds 03 Gy (b) 10Gy (c) 20Gy (d) 50Gy (e g) and 100Gy (f h) Detail of xylem in the main vein is shownin leaves from control (i) and 100Gy irradiated seeds (j) Images (andashh) are at the same magnification Bar = 50 120583m
the irradiation of seeds only in palisade parenchyma withthe lowest values at 20Gy (Figures 5(c) 5(e) 5(g) and 5(i))By contrast in compound leaves the application of 100Gyresulted in increased cell size compared with control andleaves from seeds irradiated up to 50Gy (Figures 5(d) 5(f)5(h) and 5(j))
34 Effect of Ionizing Radiation on Fluorescence Measure-ments and Pigment Content No significant differences were
detectable in fluorescence parameters and total photosyn-thetic pigment content in plants grown from irradiated seedsat different X-rays doses (Figure 6) A tendency to NPQincrease was found at 03 and 10Gy followed by a decreaseat doses higher than 20Gy However the lack of significantdifferences in ΦPSII and NPQ among treatments indicatedthat X-rays delivered at the seed stage on tomato plantsdid not alter the capability of photosynthetic apparatus oftomato plants either to convert light energy in photochem-istry or to perform thermal dissipation mechanisms The
The Scientific World Journal 7
(a) (b)
Phen
olic
com
poun
ds (
)
aa a
aab
b
0
10
20
30
40
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf JL
(c)
Phen
olic
com
poun
ds (
) aab
abc ab
c bc
0
10
20
30
40
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf CL
(d)
Figure 4 Epifluorescence microscopy views of lamina cross sections of S lycopersicum L ldquoMicrotomrdquo compound leaves from plantsdeveloped from control (a) and 100Gy irradiated seeds (b) Phenolic compounds along chloroplast membranes are autofluorescent Percentof mesophyll occupied by phenolic compounds () is shown in (c) and (d) for juvenile (JL) and compound (CL) leaves respectively Meanvalues and standard errors are reported (119899 = 15) different letters correspond to significantly different values after multiple comparison tests(119875 lt 005)
maximum photochemical efficiency of PSII (FvFm) washigher in leaves from irradiated seeds than control anddid not show variations among different X-rays treatmentsthis indicated the maintenance of higher level of photo-synthetic activity after irradiation irrespectively from dose(Figure 6(c))
The total chlorophyll as well as the total carotenoidcontent did not show significant changes due to irradiationof seeds (Figure 7)
4 Discussion
Ionising radiation can have positive null or negative effectson plant development depending on the properties of theradiation the features of the plant material and possibleinteractions with other environmental factors [2] There iscommon agreement that plants are characterised by highradioresistance moreover low-LET ionising radiation hasbeen reported to increase germination percentage rootlength plant height and other growth parameters especiallyat low doses which induce hormesis [16 17 45ndash47] Ourresults suggest that S lycopersicum ldquoMicrotomrdquo is charac-terised by high radioresistance at the seed stage indeed
irradiation of dry seeds with a wide range of X-ray dosesdid not hamper germination and plant growth up to thecompletion of the crop cycle An hormesis effect was evidentsince growth either remained unaffected or was positivelyaffected especially by low doses of X-rays Apart from plantsgrown from seeds irradiated with 03 and 10Gy all plantsfrom irradiated seeds were characterised by a more compacthabitus higher number of larger leaves was not accompaniedby increased height The radiation-induced more compactgrowth pattern is positive in BLSSs in Spacewhere availabilityof volume for growth is a major technical constraint [7 4849] In the cultivar Microtom irradiation with doses equalto or higher than 20Gy caused a decrease in plant heightin agreement with the induction of dwarf or semidwarfgrowth reported as a general effect of exposure to ionisingradiation [18] Hence irradiation on the ldquoMicrotomrdquo seedswould intensify the trait of dwarf growth which alreadycharacterise this cultivar and should be taken into account forcultivar selection for the BLSSs [50] Any variations in growthparameters in Space-oriented ecologically closed systemscan determine valuable changes in terms of optimisationof volume and resource recycling These changes can bestraightaway converted into economic saving or waste [9]For example the increase in the number and size of leaves
8 The Scientific World Journal
Palisade parenchyma Spongy parenchyma
ab b ab a ab ab
a a a a a a
0010203040506
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf JL
Lam
ina t
hick
ness
(mm
)
(a)
Palisade parenchyma Spongy parenchyma
b b b b b a
a ab ab b abab
Leaf CL
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
010203040506
Lam
ina t
hick
ness
(mm
)
(b)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a a aa a
a
0200400600800
1000Upper epidermis
Cel
l are
a (120583
m2)
(c)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
1000
a a a a aa
Upper epidermis
Cel
l are
a (120583
m2)
(d)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
a bc bc c bc b
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(e)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
ab ab b ab ab a
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(f)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a aa
a a a
0200400600800
100012001400
Cel
l are
a (120583
m2)
(g)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
100012001400
b b b b ba
Cel
l are
a (120583
m2)
(h)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
a aa
a a a
0
200
400
600
800
Cel
l are
a (120583
m2)
(i)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
b b b b ba
0
200
400
600
800
Cel
l are
a (120583
m2)
(j)
Figure 5 Lamina thickness (a b) and cell area of upper epidermis (c d) palisade (e f) spongy (g h) parenchymas and lower epidermis (i j)of S lycopersicum L ldquoMicrotomrdquo leaves of control and irradiated plants at different doses of X-rays in both juvenile (JL) and compound (CL)leaves Mean values and standard errors are shown (119899 = 15 in (a b) 119899 = 45 in (cndashj)) Different letters correspond to significantly differentvalues according to multiple comparison tests (119875 lt 005)
The Scientific World Journal 9
04
05
06
07
08
a a a a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
ΦPS
II
(a)
00
02
04
06
08
NPQ a
a aa a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
(b)
070
075
080
085
090
a
b b b b b
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
FF
m
(c)
Figure 6 Quantum yield of PSII electron transport (ΦPSII a) nonphotochemical quenching (NPQ b) and maximal PSII photochemicalefficiency (FvFm c) in compound (CL) S lycopersicum L ldquoMicrotomrdquo leaves of control and plants from seeds irradiated at different dosesof X-rays Mean values and standard deviations are shown (119899 = 5) Different letters correspond to significantly different values according tomultiple comparison tests (119875 lt 005)
0
1
2
3
4
a
aa
a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Chl a
+ b
(mg g
minus1)
(a)
0
02
04
06
a
aa a
a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Car x
+ c
(mg g
minus1)
(b)
Figure 7 Total chlorophyll content (Chl a+b) (a) and total carotenoid content (Car x+c) (b) in compound (CL) S lycopersicum L ldquoMicrotomrdquoleaves of control and plants from seeds irradiated at different doses of X-rays Mean values and standard errors are shown (119899 = 5) Differentletters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
induced in ldquoMicrotomrdquo especially by low doses of irradiationcan be beneficial to atmosphere regeneration of pressurisedmodules provided that formed leaves do not show structuralaberrations which reduce their function
General anatomical structure of both juvenile and com-pound ldquoMicrotomrdquo leaves was only slightly perturbed inplants from seeds subjected to the highest levels of radiation
Clear-cut dose-depended tendencies of variations were notfound in lamina thickness and cell size and shape of thevarious tissues The formation of smaller palisade cells wasfound only in juvenile leaves of irradiated seeds while anincrease in cell size was found only in spongy parenchymaand lower epidermis of compound leaves from seeds irra-diated at 100Gy The diverse growth behaviour found in
10 The Scientific World Journal
Table 1 Cell shape in different tissues of juvenile leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown (119899 = 75)Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf JL Aspect ratio Sphericity Convexity
Upper epidermis
C 197 plusmn 016 a 0308 plusmn 0050 a 0926 plusmn 0021 ab03Gy 200 plusmn 013 a 0290 plusmn 0038 a 0941 plusmn 0013 a10Gy 210 plusmn 012 a 0239 plusmn 0037 a 0898 plusmn 0016 ab20Gy 217 plusmn 014 a 0258 plusmn 0045 a 0909 plusmn 0019 abc50Gy 170 plusmn 008 a 0376 plusmn 0037 a 0927 plusmn 0016 ab100Gy 197 plusmn 014 a 0286 plusmn 0042 a 0876 plusmn 0028 c
Palisade parenchyma
C 290 plusmn 010 a 0109 plusmn 0015 a 0879 plusmn 0010 a03 Gy 304 plusmn 012 a 0099 plusmn 0016 a 0805 plusmn 0016 b10Gy 309 plusmn 013 a 0093 plusmn 0019 a 0792 plusmn 0015 b20Gy 283 plusmn 011 a 0101 plusmn 0017 a 0784 plusmn 0017 b50Gy 314 plusmn 014 a 0082 plusmn 0016 a 0772 plusmn 0016 b100Gy 275 plusmn 011 a 0119 plusmn 0020 a 0805 plusmn 0017 b
Spongy parenchyma
C 190 plusmn 008 a 0307 plusmn 0033 a 0893 plusmn 0012 a03 Gy 180 plusmn 007 a 0341 plusmn 0029 a 0876 plusmn 0013 a10Gy 203 plusmn 010 a 0290 plusmn 0031 a 0901 plusmn 0012 a20Gy 194 plusmn 012 a 0344 plusmn 0040 a 0881 plusmn 0015 a50Gy 174 plusmn 009 a 0393 plusmn 0035 a 0883 plusmn 0014 a100Gy 198 plusmn 012 a 0322 plusmn 0037 a 0878 plusmn 0015 a
Lower epidermis
C 178 plusmn 009 bc 0343 plusmn 0045 ab 0919 plusmn 0019 a03 Gy 225 plusmn 017 ab 0263 plusmn 0044 b 0933 plusmn 0024 a10Gy 219 plusmn 011 abc 0218 plusmn 0034 b 0888 plusmn 0020 ab20Gy 180 plusmn 013 bc 0378 plusmn 0047 ab 0926 plusmn 0017 a50Gy 167 plusmn 001 c 0461 plusmn 0051 a 0924 plusmn 0021 a100Gy 238 plusmn 021 a 0257 plusmn 0057 b 0857 plusmn 0030 b
the various tissues of structures of different age is in agree-ment with the different light-mediated control of cell devel-opment in the various leaf tissues [51] Altered mesophyllcharacterized by partly collapsed cells with irregular cellshape was found only sporadically after the exposure of seedsto 50 and 100Gy in theMicrotomcultivar Suchmodificationswere less marked than those found in leaves of bean wherethe target organs for irradiation were the leaves themselves[30] leaves directly exposed to X-rays in bean showed theformation of more loose parenchyma with larger cells likelydue to a radiation-induced damage to cell walls that reducedmechanical constraints to cell enlargement [30 52] Furtherinvestigations on cell wall composition would be useful toelucidate whether a possible alteration of cell walls wasresponsible for cell enlargement in the spongy parenchymaand epidermis of leaves from seeds of ldquoMicrotomrdquo irradiatedwith the highest dose of X-rays However in the cultivarMicrotom the slight alterations of mesophyll structure didnot impair photosynthetic efficiency as demonstrated by thelack of significant dose-dependent variation trends in thequantum yield of PSII electron transport (ΦPSII) nonphoto-chemical quenching (NPQ) and maximal PSII photochemi-cal efficiency (FvFm) The increased values of FvFm closeto 08 which is considered an optimal value for healthy plants[53] suggested that the photosynthetic apparatus of the leavesfrom irradiated seeds has higher levels of photochemical
efficiency irrespective of dose The unchanged or increasedcontent of both chlorophyll and carotenoid pigments con-firmed that the photosynthetic apparatus remained stablealso in terms of light harvesting capacity and indicatedno deleterious effect of radiation on plant photosyntheticmachinery RecentlyMarcu et al [17] showed that irradiationof lettuce seeds with gamma rays up to 30Gy enhancedthe content of chlorophyll a chlorophyll b and carotenoidswhile higher doses (up to 70Gy) resulted in their decreaseHowever the effect of ionising radiation on photosyntheticpigment content is quite controversial Increased pigmentcontent is ascribed to the phenomenon of hormesis whilea lower content is generally ascribed to the inhibition ofchlorophyll and carotenoid synthesis or to oxidation due tothe high levels of radiation-induced water radiolysis [2 1454ndash56] Some plants possess natural screens against radiationwhen their tissues contain phenolic compounds with antioxi-dant functions and can counteract photoinhibitory processes[29 57] An increase of phenolic compounds linked to thechloroplast membranes has been recently found in Phaseolusvulgaris irradiated leaves exposed to high levels of X-rays[28 30] Changes in phenolic content after the exposure toSpace can be attributed either to direct effects on the synthesisor mobilization of phenolic compounds or to indirect effectsdue to changes in ultrastructural organization such as theincrease in the number of chloroplasts per cell [30 58]
The Scientific World Journal 11
Table 2 Cell shape in different tissues of compound leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown(119899 = 75) Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf CL Aspect ratio Sphericity Convexity
Upper epidermis
C 166 plusmn 007 c 0422 plusmn 0042 a 0946 plusmn 0010 a03 Gy 216 plusmn 017 ab 0244 plusmn 0033 c 0925 plusmn 0016 ab10Gy 171 plusmn 011 bc 0401 plusmn 0042 a 0914 plusmn 0013 b20Gy 186 plusmn 010 abc 0328 plusmn 0041 ab 0919 plusmn 0020 ab50Gy 225 plusmn 018 a 0247 plusmn 0041 c 0897 plusmn 0019 b100Gy 188 plusmn 009 abc 0319 plusmn 0045 ab 0902 plusmn 0016 b
Palisade parenchyma
C 355 plusmn 012 a 0073 plusmn 0014 b 0890 plusmn 0012 a03 Gy 289 plusmn 010 b 0102 plusmn 0013 ab 0847 plusmn 0018 b10Gy 300 plusmn 014 b 0104 plusmn 0021 ab 0826 plusmn 0017 b20Gy 302 plusmn 011 b 0094 plusmn 0013 ab 0810 plusmn 0014 b50Gy 283 plusmn 012 b 0107 plusmn 0021 ab 0750 plusmn 0021 c100Gy 275 plusmn 012 b 0122 plusmn 0023 a 0809 plusmn 0015 b
Spongy parenchyma
C 193 plusmn 010 a 0323 plusmn 0035 a 0904 plusmn 0011 a03 Gy 183 plusmn 010 a 0375 plusmn 0042 a 0879 plusmn 0013 a10Gy 176 plusmn 007 a 0357 plusmn 0034 a 0869 plusmn 0011 a20Gy 180 plusmn 009 a 0366 plusmn 0038 a 0885 plusmn 0016 a50Gy 171 plusmn 006 a 0372 plusmn 0034 a 0879 plusmn 0015 a100Gy 188 plusmn 010 a 0339 plusmn 0037 a 0883 plusmn 0015 a
Lower epidermis
C 148 plusmn 008 b 0545 plusmn 0045 a 0957 plusmn 0015 a03 Gy 210 plusmn 017 a 0295 plusmn 0046 b 0929 plusmn 0019 ab10Gy 190 plusmn 012 ab 0334 plusmn 0043 b 0893 plusmn 0019 b20Gy 178 plusmn 011 ab 0388 plusmn 0047 b 0929 plusmn 0017 ab50Gy 172 plusmn 009 ab 0367 plusmn 0040 b 0914 plusmn 0016 b100Gy 191 plusmn 014 ab 0362 plusmn 0054 b 0888 plusmn 0025 b
The lowering of phenolic content at high levels of X-rays inldquoMicrotomrdquo leaves is not in agreement with general trendsindicating enhanced phenolics production in plants grownin Space [58] or in various plant tissues directly irradiatedwith gamma rays [59] However the low content of phenolicscan be an indirect effect of increasing volume occupiedby intercellular spaces due to the loosening of parenchymacells
In conclusion our results show that irradiation of the dryseeds of the Microtom cultivar did not hamper germinationand the development of functional leaves The formationof more compact plants characterised by a higher numberof larger leaves can be valuable in terms of increasingresource regeneration in the BLSSs High levels of radiationinduced only slight structural perturbations which weresimilar in juvenile and compound leaves Such perturbationsdid not affect photosynthetic efficiency since the maximalPSII photochemical efficiency was even increased after irra-diation at all doses However the tendency to accumulatelower amounts of phenolic compounds along chloroplastmembranes after irradiation of seeds with high doses ofradiation should be further investigated because it coulddetermine the reduction of natural photoprotection [57]Finally the overall results indicate that theMicrotom cultivarirradiated at the seed stage has high radioresistance duringthe growth which is a desirable feature for plant cultivation
in Space Radioresistance together with dwarf growth andshort life cycle makes this cultivar a valuable candidate forthe cultivation in BLSSs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are grateful to the ldquoCompagnia di San Paolordquo(Turin) and the ldquoPolo delle Scienze e delle Tecnologie perla Vitardquo of the University of Naples Federico II for thefinancial support The authors also thank Maria Giordanoand Pasquale Scognamiglio for their support in plant culti-vation and microscopy analyses The authors acknowledgefour anonymous editors for helpful comments on a previousversion of the paper
References
[1] A Real S Sundell-Bergman J F Knowles D S Woodheadand I Zinger ldquoEffects of ionising radiation exposure on plantsfish and mammals relevant data for environmental radiation
12 The Scientific World Journal
protectionrdquo Journal of Radiological Protection vol 24 no 4 ppA123ndashA137 2004
[2] V De Micco C Arena D Pignalosa and M Durante ldquoEffectsof sparsely and densely ionizing radiation on plantsrdquo Radiationand Environmental Biophysics vol 50 no 1 pp 1ndash19 2011
[3] N Kumari P Kumar D Mitra B Prasad B N Tiwary andL Varshney ldquoEffects of ionizing radiation on microbial decon-tamination phenolic contents and antioxidant properties oftriphalardquo Journal of Food Science vol 74 no 3 pp M109ndashM1132009
[4] A Tanaka N Shikazono and Y Hase ldquoStudies on biologicaleffects of ion beams on lethality molecular nature of mutationmutation rate and spectrum of mutation phenotype for muta-tion breeding in higher plantsrdquo Journal of Radiation Researchvol 51 no 3 pp 223ndash233 2010
[5] M Durante and A Kronenberg ldquoGround-based research withheavy ions for space radiation protectionrdquo Advances in SpaceResearch vol 35 no 2 pp 180ndash184 2005
[6] E Hu S I Bartsev and H Liu ldquoConceptual design of a biore-generative life support system containing crops and silkwormsrdquoAdvances in Space Research vol 45 no 7 pp 929ndash939 2010
[7] V deMicco G Aronne G Colla R Fortezza and S de PascaleldquoAgro-biology for bioregenerative life support systems in long-term space missions general constraints and the Italian effortsrdquoJournal of Plant Interactions vol 4 no 4 pp 241ndash252 2009
[8] T W Halstead and F R Dutcher ldquoPlants in spacerdquo AnnualReview of Plant Physiology vol 38 pp 317ndash345 1987
[9] V De Micco S De Pascale R Paradiso and G AronneldquoMicrogravity effects on different stages of higher plant life cycleand completion of the seed-to-seed cyclerdquo Plant Biology vol 16pp 31ndash38 2014
[10] C J Bennett C Pirim and T M Orlando ldquoSpace-weatheringof solar system bodies a laboratory perspectiverdquo ChemicalReviews vol 113 pp 9086ndash9150 2013
[11] R A Kerr ldquoRadiation will make astronautsrsquo trip to Mars evenriskierrdquo Science vol 340 article 1031 2014
[12] D M Hassler C Zeitlin R F Wimmer-Schweingruber et alldquoMarsrsquo surface radiation environment measured with the MarsScience Laboratoryrsquos Curiosity Roverrdquo Science vol 343 no6169 Article ID 1244797 2014
[13] M Durante and F A Cucinotta ldquoHeavy ion carcinogenesis andhuman space explorationrdquo Nature Reviews Cancer vol 8 no 6pp 465ndash472 2008
[14] C Arena V De Micco and A De Maio ldquoGrowth alterationand leaf biochemical responses in P vulgaris plants exposed todifferent doses of ionizing radiationrdquo Plant Biology vol 16 pp194ndash202 2014
[15] R W Holst and D J Nagel ldquoRadiation effects on plantsrdquo inPlants For Environmental Studies W Wang J W Gorsuch andJ S Hughes Eds pp 37ndash81 Lewis Publishers Boca Raton FlaUSA 1997
[16] T D Luckey Hormesis With Ionizing Radiation CRC PressBoca Raton Fla USA 1980
[17] D Marcu V Cristea and L Daraban ldquoDose-dependent effectsof gamma radiation on lettuce (Lactuca sativa var capitata)seedlingsrdquo International Journal of Radiation Biology vol 89 pp219ndash223 2013
[18] M Mei H Deng Y Lu et al ldquoMutagenic effects of heavy ionradiation in plantsrdquo Advances in Space Research vol 14 no 10pp 363ndash372 1994
[19] Y LiM Liu Z Cheng andY Sun ldquoSpace environment inducedmutations prefer to occur at polymorphic sites of rice genomesrdquoAdvances in Space Research vol 40 no 4 pp 523ndash527 2007
[20] D S Kim J-B Kim E J Goh et al ldquoAntioxidant response ofArabidopsis plants to gamma irradiation genome-wide expres-sion profiling of the ROS scavenging and signal transductionpathwaysrdquo Journal of Plant Physiology vol 168 no 16 pp 1960ndash1971 2011
[21] T J Brodribb ldquoXylem hydraulic physiology the functionalbackbone of terrestrial plant productivityrdquo Plant Science vol177 no 4 pp 245ndash251 2009
[22] R M Wheeler J C Sager R P Prince andW M Knott ldquoCropproduction for advanced life support systemsmdashobservationfrom the Kennedy Space Center Breadboard Projectrdquo NASATM-2003-211184 NASA Kennedy Space Center Florida 2003
[23] R M Wheeler C L Mackowiak G W Stutte et al ldquoCropproductivities and radiation use efficiencies for bioregenerativelife supportrdquo Advances in Space Research vol 41 no 5 pp 706ndash713 2008
[24] R Paradiso V De Micco R Buonomo G Aronne G Bar-bieri and S De Pascale ldquoSoilless cultivation of soybean forBioregenerative Life-Support Systems a literature review andthe experience of the MELiSSA ProjectmdashFood characterisationPhase Irdquo Plant Biology vol 16 pp 69ndash78 2014
[25] D M Porterfield ldquoThe biophysical limitations in physiologicaltransport and exchange in plants grown in microgravityrdquoJournal of Plant Growth Regulation vol 21 no 2 pp 177ndash1902002
[26] K Yokthongwattana and A Melis ldquoPhotoinhibition and recov-ery in oxygenic photosynthesis mechanism of a photosystem IIdamage and repair cyclerdquo in Photoprotection PhotoinhibitionGene Regulation and Environment B Demmig-Adams WW Adams III and A K Mattoo Eds pp 175ndash191 SpringerDordecht The Netherlands 2006
[27] M T Giardi G Rea M D Lambreva et al ldquoMutations ofphotosystem II D1 protein that empower efficient phenotypesof Chlamydomonas reinhardtii under extreme environment inSpacerdquo PLoS ONE vol 8 Article ID e64352 2013
[28] C Arena V De Micco G Aronne M Pugliese A Virzo DeSanto andADeMaio ldquoResponse ofPhaseolus vulgarisL plantsto low-let ionizing radiation growth and oxidative stressrdquo ActaAstronautica vol 91 pp 107ndash114 2013
[29] V Lattanzio P A Kroon and S Quideau ldquoPlant phenolicsmdashsecondary metabolites with diverse functionsrdquo in RecentAdvances in Polyphenol Research F Daay and V Lattanzio Edspp 1ndash35 Wiley-Blackwell Oxford UK 2008
[30] V De Micco C Arena and G Aronne ldquoAnatomical alterationsof Phaseolus vulgaris L mature leaves irradiated with X-raysrdquoPlant Biology vol 16 pp 187ndash193 2014
[31] A D Krikorian ldquoPlants and somatic embryos in space whathave we learnedrdquo Gravitational and Space Biology Bulletin vol11 no 2 pp 5ndash14 1998
[32] R Meissner Y Jacobson S Melamed et al ldquoA new modelsystem for tomato geneticsrdquoThe Plant Journal vol 12 no 6 pp1465ndash1472 1997
[33] E Emmanuel and A A Levy ldquoTomato mutants as tools forfunctional genomicsrdquo Current Opinion in Plant Biology vol 5no 2 pp 112ndash117 2002
[34] E Martı C Gisbert G J Bishop M S Dixon and J LGarcıa-Martınez ldquoGenetic and physiological characterizationof tomato cv Micro-Tomrdquo Journal of Experimental Botany vol57 pp 2037ndash2047 2006
The Scientific World Journal 13
[35] T Kurimoto J V H Constable and A Huda ldquoEffects ofionizing radiation exposure on arabidopsis thalianardquo HealthPhysics vol 99 no 1 pp 49ndash57 2010
[36] L Reale D Gigante F Landucci F Ferranti and R VenanzonildquoMorphological and histo-anatomical traits reflect die-back inPhragmites australis (Cav) Steudrdquo Aquatic Botany vol 103 pp122ndash128 2012
[37] K Fukuzawa ldquoUltraviolet microscopyrdquo in Methods in LigninChemistry S Y Lin andCWDence Eds pp 110ndash131 SpringerBerlin Germany 1992
[38] S E Ruzin Plant Microtechnique and Microscopy OxfordUniversity Press New York NY USA 1999
[39] V De Micco G Aronne J-P Joseleau and K Ruel ldquoXylemdevelopment and cell wall changes of soybean seedlings grownin spacerdquo Annals of Botany vol 101 no 5 pp 661ndash669 2008
[40] S Van Buggenhout T Grauwet A Van Loey and M Hen-drickx ldquoStructureprocessing relation of vacuum infused straw-berry tissue frozen under different conditionsrdquo European FoodResearch and Technology vol 226 no 3 pp 437ndash448 2008
[41] V De Micco and G Aronne ldquoBiometric anatomy of seedlingsdeveloped onboard of Foton-M2 in an automatic system sup-porting growthrdquoActa Astronautica vol 62 no 8-9 pp 505ndash5132008
[42] B Genty J M Briantais and N R Baker ldquoThe relationshipbetween quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescencerdquo Biochimica andBiophysica Acta vol 990 pp 87ndash92 1989
[43] W Bilger and O Bjorkman ldquoRole of the xanthophyll cycle inphotoprotection elucidated by measurements of light-inducedabsorbance changes fluorescence and photosynthesis in leavesof Hedera canariensisrdquo Photosynthesis Research vol 25 no 3pp 173ndash185 1990
[44] H K Lichtenthaler ldquoChlorophylls and carotenoids pigmentsof photosynthetic biomembranesrdquoMethods in Enzymology vol148 pp 350ndash382 1987
[45] J P Maity DMishra A Chakraborty A Saha S C Santra andS Chanda ldquoModulation of some quantitative and qualitativecharacteristics in rice (Oryza sativa L) and mung (Phaseolusmungo L) by ionizing radiationrdquo Radiation Physics and Chem-istry vol 74 no 5 pp 391ndash394 2005
[46] L Cheng H Yang B Lin et al ldquoEffect of gamma-ray radiationon physiological morphological characters and chromosomeaberrations of minitubers in Solanum tuberosum Lrdquo Interna-tional Journal of Radiation Biology vol 86 no 9 pp 791ndash7992010
[47] B Al-Safadi and R Elias ldquoImprovement of caper (Capparisspinosa L) propagation using in vitro culture and gammairradiationrdquo Scientia Horticulturae vol 127 no 3 pp 290ndash2972011
[48] O Monje G W Stutte G D Goins D M Porterfield and GE Bingham ldquoFarming in space environmental and biophysicalconcernsrdquo Advances in Space Research vol 31 no 1 pp 151ndash1672003
[49] V De Micco M Scala and G Aronne ldquoEffects of simulatedmicrogravity on male gametophyte of Prunus Pyrus andBrassica speciesrdquo Protoplasma vol 228 no 1ndash3 pp 121ndash1262006
[50] V De Micco R Buonomo R Paradiso S De Pascale and GAronne ldquoSoybean cultivar selection for Bioregenerative LifeSupport Systems (BLSS)mdashtheoretical selectionrdquo Advances inSpace Research vol 49 no 10 pp 1415ndash1421 2012
[51] T Kozuka S-G Kong M Doi K-I Shimazaki and ANagatani ldquoTissue-autonomous promotion of palisade celldevelopment by phototropin 2 in Arabidopsisrdquo Plant Cell vol23 no 10 pp 3684ndash3695 2011
[52] E Kovacs and A Keresztes ldquoEffect of gamma and UV-BCradiation on plant cellsrdquoMicron vol 33 no 2 pp 199ndash210 2002
[53] R Ogaya J Penuelas D Asensio and J Llusia ldquoChlorophyllfluorescence responses to temperature and water availabilityin two co-dominant Mediterranean shrub and tree speciesin a long-term field experiment simulating climate changerdquoEnvironmental and Experimental Botany vol 73 no 1 pp 89ndash93 2011
[54] R Zaka C M Vandecasteele and M T Misset ldquoEffects oflow chronic doses of ionizing radiation on antioxidant enzymesand G6PDH activities in Stipa capillata (Poaceae)rdquo Journal ofExperimental Botany vol 53 no 376 pp 1979ndash1987 2002
[55] J-H KimM-H Baek Y C Byung GW Seung and J-S KimldquoAlterations in the photosynthetic pigments and antioxidantmachineries of red pepper (Capsicum annuum L) seedlingsfrom gamma-irradiated seedsrdquo Journal of Plant Biology vol 47no 4 pp 314ndash321 2004
[56] N A Al-Enezi and J M Al-Khayri ldquoAlterations of DNA ionsand photosynthetic pigments content in date palm seedlingsinduced by X-irradiationrdquo International Journal of Agricultureand Biology vol 14 pp 329ndash336 2012
[57] V de Micco C Arena L Vitale G Aronne and A V de SantoldquoAnatomy and photochemical behaviour of mediterranean Cis-tus incanus winter leaves under natural outdoor and warmerindoor conditionsrdquo Botany vol 89 no 10 pp 677ndash688 2011
[58] L H Levine H G Levine E C Stryjewski V Prima and WC Piastuch ldquoEffect of spaceflight on isoflavonoid accumulationin etiolated soybean seedlingsrdquo Journal of Gravitational Physiol-ogy vol 8 no 2 pp 21ndash27 2001
[59] X Fan ldquoAntioxidant capacity of fresh-cut vegetables exposedto ionizing radiationrdquo Journal of the Science of Food andAgriculture vol 85 no 6 pp 995ndash1000 2005
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 The Scientific World Journal
C 100 Gy50 Gy20 Gy10 Gy03 Gy
Figure 1 Plants of S lycopersicum L ldquoMicrotomrdquo developed fromcontrol and irradiated seeds at 40 DAS (Days After Sowing) Plantsfrom seeds exposed to increasing doses of X-rays (from 0 to 100Gy)are shown from left to right
25 Fluorescence Measurements and Pigment Content Flu-orescence measurements and pigment extraction were con-ducted on 5 pennate-compound leaves of 5 plants subjectedto each irradiation treatment and in controls at 40 DAS
Chlorophyll a fluorescence measurements were carriedout by means of a pulse amplitude modulated fluorometer(Junior-PAM Walz Germany) equipped with a monitoringLeaf-Clip JUNIOR-B (Walz Germany) On 30min dark-adapted leaves the background fluorescence signal Fo wasinduced by internal light provided by a blue LED of about2-3 120583mol photonssdotmminus2sdotsminus1 at a frequency of 05 kHz Themaximal fluorescence level in the dark-adapted state (Fm)was measured by 1 s saturating light pulse (10 000120583molphotonssdotmminus2sdotsminus1) at a frequency of 10 kHz the maximal PSIIphotochemical efficiency (FvFm) was calculated as FvFm =(FmminusFo)FmThemeasurements in the light were carried outby exposing each leaf to a PPFD of 420120583mol photonssdotmminus2sdotsminus1for 5min This level of PPFD was chosen because it fallsin the range of maximal quantum yield for the Microtomcultivar as indicated by fluorescence fast-response curves tolight (data not shown) The steady-state fluorescence signal(Ft) and the maximal fluorescence (Fm1015840) under illuminationwere measured setting the light measure at a frequency of10 kHz Fm1015840 was determined by a 1 s saturating light pulse(10 000120583mol photonssdotmminus2sdotsminus1)
The quantum yield of the PSII electron transport (ΦPSII)and nonphotochemical quenching (NPQ) were expressedaccording to Genty et al [42] and Bilger and Bjorkman [43]
After fluorescence determinations leaves were collectedfrom plants for the photosynthetic pigment content determi-nation namely chlorophylls and carotenoids Pigments wereextracted with a mortar and pestle in ice-cold 100 acetoneand quantified by a spectrophotometer (UV-VIS Cary 100Agilent Technologies USA) according to Lichtenthaler [44]
26 Statistical Analysis All results were subjected to statis-tical analysis using the SPSS statistical package (SPSS IncChicago IL USA) For the time course of plant height num-ber of leaves and leaf area interpolation equations (second-order polynomial curve) were calculated For anatomy andecophysiology data were subjected to ANOVA multiplecomparison tests were performed with Student-Newman-Keuls and Duncan coefficients using 119875 lt 005 as the level of
probability Data on sphericity convexity and percentage ofphenolics were transformed through arcsine function beforestatistical analysis
3 Results
31 Effect of Ionizing Radiation on Seed Germination Expo-sure of dry seeds to X-rays did not affect their percentageof germination which ranged between 978 and 100 withno significant differences among treatments Similarly X-raysdid not determine significant alterations in germination rateswhich ranged between 50 and 67 days
32 Effect of Ionizing Radiation on Plant Growth Plants fromboth control and irradiated seeds completed their crop cycleup to fruit ripening in about 100 DAS plants from irradiatedseeds did not show any apparent alteration in above-groundgrowth (Figure 1)
Figure 2 shows the effects of the seed irradiation treat-ments on plant height leaf number and plant leaf areaof the main stem The irradiation dose differently affectedthese growth parameters In plants of the nonirradiatedcontrol plant height reached the maximum value of 602 cmat approximately 71 DAS Irradiation of seeds with 03Gydid not significantly influence plant height Starting from71 DAS until the end of the cycle height of plants fromseeds irradiated with 10Gy was significantly higher than thatreached after irradiation with 20 50 and 100Gy Final heightof control plants and of those from seeds irradiated withX-ray doses up to 10Gy was significantly higher than thatreached after irradiation with 20Gy and above (Figure 2(a))
Theprogression of the number of leaves and leaf areawerenot consistent with the time course of plant height (Figure 2)Indeed in the control at the end of the crop cycle plantswere characterized by 97 leaves corresponding to 625 cm2leaf area Plants from irradiated seeds formed a number ofleaves significantly higher than control at the end of the cyclewith maximum value in plants from seeds irradiated with10Gy (Figure 2(b)) Similarly under all the radiation dosesbut 100Gy plants showed significantly larger leaf area thanthe control with maximum value at 03 Gy (Figure 2(c))
33 Effect of Ionizing Radiation on LeafAnatomy Microscopyanalysis of the leaf sections showed a regular anatomicalorganisation in both juvenile and compound leaves formedafter seed irradiation at different levels of X-rays Sincethe effect of the treatments was similar in both types ofleaves microscopy views are shown only for compoundleaves (Figure 3) Both leaf types were characterised by atypical dorsiventral structure which did not face disruptivestructural alterations after irradiation of seeds with X-raysMicroscopy observations of leaves from seeds irradiatedwith up to 20Gy did not reveal evident differences com-pared with control leaves (Figures 3(a)ndash3(d)) At 50 and100Gy sporadic perturbations in the mesophyll structurewere detectable (Figures 3(e)ndash3(h)) More specifically thepresence of more shrunk mesophyll cells (Figures 3(g) and3(h)) due to palisade cells collapsed in the lower portion
The Scientific World Journal 5
0
10
20
30
40
50
60
70
80
0 20 40 60 80 100
Plan
t hei
ght (
mm
)
DAS
(a)
0
5
10
15
20
Num
ber o
f lea
ves (
Np
lant
)
0 20 40 60 80 100DAS
(b)
0 20 40 60 80 100DAS
0
20
40
60
80
100
120
140
Plan
t lea
f are
a (cm
2pl
antminus1)
C03 Gy10 Gy
20 Gy50 Gy100 Gy
(c)
Figure 2 Time course of the plant height (a) number of leaves (b) and plant leaf area (c) in S lycopersicum L ldquoMicrotomrdquo as a function ofthe X-ray doses applied at the seed stage Mean values and standard errors are shown (119899 = 5) All second order polynomial curves had 1199032values above 097
(Figure 3(g)) and spongy cells characterised by more angularshape (Figure 3(h)) was found The occurrence of irregular-shaped cells of leaves from seeds irradiated with 50 and100Gy was confirmed by significant lower convexity valuesthan control and low-dose treatments (Tables 1 and 2) Theepidermis of compound leaves from irradiated seeds tendedto have less elongated cells (ie lower aspect ratio and highersphericity) while in the palisade parenchyma an oppositetrend was observed (Table 2) No specific tendency of cellshape variation was found in the juvenile leaves (Table 1)Themain veins did not show any irregularities in vessel walls afterany irradiation treatments (Figures 3(i) and 3(j)) Epifluores-cence microscopy showed the autofluorescence of phenoliccompounds which were mainly localised along chloroplast
membranes in both control leaves and those from irradiatedplants (Figures 4(a) and 4(b)) No differences in autofluores-cence distribution were observed in mesophyll cells of juve-nile and compound leaves from control and irradiated seedsThe percentage of phenolic compounds showed a nonsignifi-cant tendency to increase in juvenile leaves at low irradiationdoses followed by a significant decrease at 50 and 100Gy(Figure 4(c)) In compound leaves phenolic compoundstended to decrease after all irradiation treatments with thelowest values in leaves from seeds irradiated at 50 and 100Gy(Figure 4(d))
The other analysed anatomical parameters did not show aclear-cut dose-dependent response (Figure 5 Tables 1 and 2)In juvenile leaves the size of cells significantly decreased after
6 The Scientific World Journal
(b) (c)
(d) (e)
(f)
(g) (h)
(i)
(j)
(a)
Figure 3 Light microscopy views of cross sections of leaf lamina of S lycopersicum L ldquoMicrotomrdquo compound leaves from plants developedfrom control (a) and irradiated seeds 03 Gy (b) 10Gy (c) 20Gy (d) 50Gy (e g) and 100Gy (f h) Detail of xylem in the main vein is shownin leaves from control (i) and 100Gy irradiated seeds (j) Images (andashh) are at the same magnification Bar = 50 120583m
the irradiation of seeds only in palisade parenchyma withthe lowest values at 20Gy (Figures 5(c) 5(e) 5(g) and 5(i))By contrast in compound leaves the application of 100Gyresulted in increased cell size compared with control andleaves from seeds irradiated up to 50Gy (Figures 5(d) 5(f)5(h) and 5(j))
34 Effect of Ionizing Radiation on Fluorescence Measure-ments and Pigment Content No significant differences were
detectable in fluorescence parameters and total photosyn-thetic pigment content in plants grown from irradiated seedsat different X-rays doses (Figure 6) A tendency to NPQincrease was found at 03 and 10Gy followed by a decreaseat doses higher than 20Gy However the lack of significantdifferences in ΦPSII and NPQ among treatments indicatedthat X-rays delivered at the seed stage on tomato plantsdid not alter the capability of photosynthetic apparatus oftomato plants either to convert light energy in photochem-istry or to perform thermal dissipation mechanisms The
The Scientific World Journal 7
(a) (b)
Phen
olic
com
poun
ds (
)
aa a
aab
b
0
10
20
30
40
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf JL
(c)
Phen
olic
com
poun
ds (
) aab
abc ab
c bc
0
10
20
30
40
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf CL
(d)
Figure 4 Epifluorescence microscopy views of lamina cross sections of S lycopersicum L ldquoMicrotomrdquo compound leaves from plantsdeveloped from control (a) and 100Gy irradiated seeds (b) Phenolic compounds along chloroplast membranes are autofluorescent Percentof mesophyll occupied by phenolic compounds () is shown in (c) and (d) for juvenile (JL) and compound (CL) leaves respectively Meanvalues and standard errors are reported (119899 = 15) different letters correspond to significantly different values after multiple comparison tests(119875 lt 005)
maximum photochemical efficiency of PSII (FvFm) washigher in leaves from irradiated seeds than control anddid not show variations among different X-rays treatmentsthis indicated the maintenance of higher level of photo-synthetic activity after irradiation irrespectively from dose(Figure 6(c))
The total chlorophyll as well as the total carotenoidcontent did not show significant changes due to irradiationof seeds (Figure 7)
4 Discussion
Ionising radiation can have positive null or negative effectson plant development depending on the properties of theradiation the features of the plant material and possibleinteractions with other environmental factors [2] There iscommon agreement that plants are characterised by highradioresistance moreover low-LET ionising radiation hasbeen reported to increase germination percentage rootlength plant height and other growth parameters especiallyat low doses which induce hormesis [16 17 45ndash47] Ourresults suggest that S lycopersicum ldquoMicrotomrdquo is charac-terised by high radioresistance at the seed stage indeed
irradiation of dry seeds with a wide range of X-ray dosesdid not hamper germination and plant growth up to thecompletion of the crop cycle An hormesis effect was evidentsince growth either remained unaffected or was positivelyaffected especially by low doses of X-rays Apart from plantsgrown from seeds irradiated with 03 and 10Gy all plantsfrom irradiated seeds were characterised by a more compacthabitus higher number of larger leaves was not accompaniedby increased height The radiation-induced more compactgrowth pattern is positive in BLSSs in Spacewhere availabilityof volume for growth is a major technical constraint [7 4849] In the cultivar Microtom irradiation with doses equalto or higher than 20Gy caused a decrease in plant heightin agreement with the induction of dwarf or semidwarfgrowth reported as a general effect of exposure to ionisingradiation [18] Hence irradiation on the ldquoMicrotomrdquo seedswould intensify the trait of dwarf growth which alreadycharacterise this cultivar and should be taken into account forcultivar selection for the BLSSs [50] Any variations in growthparameters in Space-oriented ecologically closed systemscan determine valuable changes in terms of optimisationof volume and resource recycling These changes can bestraightaway converted into economic saving or waste [9]For example the increase in the number and size of leaves
8 The Scientific World Journal
Palisade parenchyma Spongy parenchyma
ab b ab a ab ab
a a a a a a
0010203040506
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf JL
Lam
ina t
hick
ness
(mm
)
(a)
Palisade parenchyma Spongy parenchyma
b b b b b a
a ab ab b abab
Leaf CL
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
010203040506
Lam
ina t
hick
ness
(mm
)
(b)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a a aa a
a
0200400600800
1000Upper epidermis
Cel
l are
a (120583
m2)
(c)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
1000
a a a a aa
Upper epidermis
Cel
l are
a (120583
m2)
(d)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
a bc bc c bc b
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(e)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
ab ab b ab ab a
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(f)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a aa
a a a
0200400600800
100012001400
Cel
l are
a (120583
m2)
(g)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
100012001400
b b b b ba
Cel
l are
a (120583
m2)
(h)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
a aa
a a a
0
200
400
600
800
Cel
l are
a (120583
m2)
(i)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
b b b b ba
0
200
400
600
800
Cel
l are
a (120583
m2)
(j)
Figure 5 Lamina thickness (a b) and cell area of upper epidermis (c d) palisade (e f) spongy (g h) parenchymas and lower epidermis (i j)of S lycopersicum L ldquoMicrotomrdquo leaves of control and irradiated plants at different doses of X-rays in both juvenile (JL) and compound (CL)leaves Mean values and standard errors are shown (119899 = 15 in (a b) 119899 = 45 in (cndashj)) Different letters correspond to significantly differentvalues according to multiple comparison tests (119875 lt 005)
The Scientific World Journal 9
04
05
06
07
08
a a a a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
ΦPS
II
(a)
00
02
04
06
08
NPQ a
a aa a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
(b)
070
075
080
085
090
a
b b b b b
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
FF
m
(c)
Figure 6 Quantum yield of PSII electron transport (ΦPSII a) nonphotochemical quenching (NPQ b) and maximal PSII photochemicalefficiency (FvFm c) in compound (CL) S lycopersicum L ldquoMicrotomrdquo leaves of control and plants from seeds irradiated at different dosesof X-rays Mean values and standard deviations are shown (119899 = 5) Different letters correspond to significantly different values according tomultiple comparison tests (119875 lt 005)
0
1
2
3
4
a
aa
a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Chl a
+ b
(mg g
minus1)
(a)
0
02
04
06
a
aa a
a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Car x
+ c
(mg g
minus1)
(b)
Figure 7 Total chlorophyll content (Chl a+b) (a) and total carotenoid content (Car x+c) (b) in compound (CL) S lycopersicum L ldquoMicrotomrdquoleaves of control and plants from seeds irradiated at different doses of X-rays Mean values and standard errors are shown (119899 = 5) Differentletters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
induced in ldquoMicrotomrdquo especially by low doses of irradiationcan be beneficial to atmosphere regeneration of pressurisedmodules provided that formed leaves do not show structuralaberrations which reduce their function
General anatomical structure of both juvenile and com-pound ldquoMicrotomrdquo leaves was only slightly perturbed inplants from seeds subjected to the highest levels of radiation
Clear-cut dose-depended tendencies of variations were notfound in lamina thickness and cell size and shape of thevarious tissues The formation of smaller palisade cells wasfound only in juvenile leaves of irradiated seeds while anincrease in cell size was found only in spongy parenchymaand lower epidermis of compound leaves from seeds irra-diated at 100Gy The diverse growth behaviour found in
10 The Scientific World Journal
Table 1 Cell shape in different tissues of juvenile leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown (119899 = 75)Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf JL Aspect ratio Sphericity Convexity
Upper epidermis
C 197 plusmn 016 a 0308 plusmn 0050 a 0926 plusmn 0021 ab03Gy 200 plusmn 013 a 0290 plusmn 0038 a 0941 plusmn 0013 a10Gy 210 plusmn 012 a 0239 plusmn 0037 a 0898 plusmn 0016 ab20Gy 217 plusmn 014 a 0258 plusmn 0045 a 0909 plusmn 0019 abc50Gy 170 plusmn 008 a 0376 plusmn 0037 a 0927 plusmn 0016 ab100Gy 197 plusmn 014 a 0286 plusmn 0042 a 0876 plusmn 0028 c
Palisade parenchyma
C 290 plusmn 010 a 0109 plusmn 0015 a 0879 plusmn 0010 a03 Gy 304 plusmn 012 a 0099 plusmn 0016 a 0805 plusmn 0016 b10Gy 309 plusmn 013 a 0093 plusmn 0019 a 0792 plusmn 0015 b20Gy 283 plusmn 011 a 0101 plusmn 0017 a 0784 plusmn 0017 b50Gy 314 plusmn 014 a 0082 plusmn 0016 a 0772 plusmn 0016 b100Gy 275 plusmn 011 a 0119 plusmn 0020 a 0805 plusmn 0017 b
Spongy parenchyma
C 190 plusmn 008 a 0307 plusmn 0033 a 0893 plusmn 0012 a03 Gy 180 plusmn 007 a 0341 plusmn 0029 a 0876 plusmn 0013 a10Gy 203 plusmn 010 a 0290 plusmn 0031 a 0901 plusmn 0012 a20Gy 194 plusmn 012 a 0344 plusmn 0040 a 0881 plusmn 0015 a50Gy 174 plusmn 009 a 0393 plusmn 0035 a 0883 plusmn 0014 a100Gy 198 plusmn 012 a 0322 plusmn 0037 a 0878 plusmn 0015 a
Lower epidermis
C 178 plusmn 009 bc 0343 plusmn 0045 ab 0919 plusmn 0019 a03 Gy 225 plusmn 017 ab 0263 plusmn 0044 b 0933 plusmn 0024 a10Gy 219 plusmn 011 abc 0218 plusmn 0034 b 0888 plusmn 0020 ab20Gy 180 plusmn 013 bc 0378 plusmn 0047 ab 0926 plusmn 0017 a50Gy 167 plusmn 001 c 0461 plusmn 0051 a 0924 plusmn 0021 a100Gy 238 plusmn 021 a 0257 plusmn 0057 b 0857 plusmn 0030 b
the various tissues of structures of different age is in agree-ment with the different light-mediated control of cell devel-opment in the various leaf tissues [51] Altered mesophyllcharacterized by partly collapsed cells with irregular cellshape was found only sporadically after the exposure of seedsto 50 and 100Gy in theMicrotomcultivar Suchmodificationswere less marked than those found in leaves of bean wherethe target organs for irradiation were the leaves themselves[30] leaves directly exposed to X-rays in bean showed theformation of more loose parenchyma with larger cells likelydue to a radiation-induced damage to cell walls that reducedmechanical constraints to cell enlargement [30 52] Furtherinvestigations on cell wall composition would be useful toelucidate whether a possible alteration of cell walls wasresponsible for cell enlargement in the spongy parenchymaand epidermis of leaves from seeds of ldquoMicrotomrdquo irradiatedwith the highest dose of X-rays However in the cultivarMicrotom the slight alterations of mesophyll structure didnot impair photosynthetic efficiency as demonstrated by thelack of significant dose-dependent variation trends in thequantum yield of PSII electron transport (ΦPSII) nonphoto-chemical quenching (NPQ) and maximal PSII photochemi-cal efficiency (FvFm) The increased values of FvFm closeto 08 which is considered an optimal value for healthy plants[53] suggested that the photosynthetic apparatus of the leavesfrom irradiated seeds has higher levels of photochemical
efficiency irrespective of dose The unchanged or increasedcontent of both chlorophyll and carotenoid pigments con-firmed that the photosynthetic apparatus remained stablealso in terms of light harvesting capacity and indicatedno deleterious effect of radiation on plant photosyntheticmachinery RecentlyMarcu et al [17] showed that irradiationof lettuce seeds with gamma rays up to 30Gy enhancedthe content of chlorophyll a chlorophyll b and carotenoidswhile higher doses (up to 70Gy) resulted in their decreaseHowever the effect of ionising radiation on photosyntheticpigment content is quite controversial Increased pigmentcontent is ascribed to the phenomenon of hormesis whilea lower content is generally ascribed to the inhibition ofchlorophyll and carotenoid synthesis or to oxidation due tothe high levels of radiation-induced water radiolysis [2 1454ndash56] Some plants possess natural screens against radiationwhen their tissues contain phenolic compounds with antioxi-dant functions and can counteract photoinhibitory processes[29 57] An increase of phenolic compounds linked to thechloroplast membranes has been recently found in Phaseolusvulgaris irradiated leaves exposed to high levels of X-rays[28 30] Changes in phenolic content after the exposure toSpace can be attributed either to direct effects on the synthesisor mobilization of phenolic compounds or to indirect effectsdue to changes in ultrastructural organization such as theincrease in the number of chloroplasts per cell [30 58]
The Scientific World Journal 11
Table 2 Cell shape in different tissues of compound leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown(119899 = 75) Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf CL Aspect ratio Sphericity Convexity
Upper epidermis
C 166 plusmn 007 c 0422 plusmn 0042 a 0946 plusmn 0010 a03 Gy 216 plusmn 017 ab 0244 plusmn 0033 c 0925 plusmn 0016 ab10Gy 171 plusmn 011 bc 0401 plusmn 0042 a 0914 plusmn 0013 b20Gy 186 plusmn 010 abc 0328 plusmn 0041 ab 0919 plusmn 0020 ab50Gy 225 plusmn 018 a 0247 plusmn 0041 c 0897 plusmn 0019 b100Gy 188 plusmn 009 abc 0319 plusmn 0045 ab 0902 plusmn 0016 b
Palisade parenchyma
C 355 plusmn 012 a 0073 plusmn 0014 b 0890 plusmn 0012 a03 Gy 289 plusmn 010 b 0102 plusmn 0013 ab 0847 plusmn 0018 b10Gy 300 plusmn 014 b 0104 plusmn 0021 ab 0826 plusmn 0017 b20Gy 302 plusmn 011 b 0094 plusmn 0013 ab 0810 plusmn 0014 b50Gy 283 plusmn 012 b 0107 plusmn 0021 ab 0750 plusmn 0021 c100Gy 275 plusmn 012 b 0122 plusmn 0023 a 0809 plusmn 0015 b
Spongy parenchyma
C 193 plusmn 010 a 0323 plusmn 0035 a 0904 plusmn 0011 a03 Gy 183 plusmn 010 a 0375 plusmn 0042 a 0879 plusmn 0013 a10Gy 176 plusmn 007 a 0357 plusmn 0034 a 0869 plusmn 0011 a20Gy 180 plusmn 009 a 0366 plusmn 0038 a 0885 plusmn 0016 a50Gy 171 plusmn 006 a 0372 plusmn 0034 a 0879 plusmn 0015 a100Gy 188 plusmn 010 a 0339 plusmn 0037 a 0883 plusmn 0015 a
Lower epidermis
C 148 plusmn 008 b 0545 plusmn 0045 a 0957 plusmn 0015 a03 Gy 210 plusmn 017 a 0295 plusmn 0046 b 0929 plusmn 0019 ab10Gy 190 plusmn 012 ab 0334 plusmn 0043 b 0893 plusmn 0019 b20Gy 178 plusmn 011 ab 0388 plusmn 0047 b 0929 plusmn 0017 ab50Gy 172 plusmn 009 ab 0367 plusmn 0040 b 0914 plusmn 0016 b100Gy 191 plusmn 014 ab 0362 plusmn 0054 b 0888 plusmn 0025 b
The lowering of phenolic content at high levels of X-rays inldquoMicrotomrdquo leaves is not in agreement with general trendsindicating enhanced phenolics production in plants grownin Space [58] or in various plant tissues directly irradiatedwith gamma rays [59] However the low content of phenolicscan be an indirect effect of increasing volume occupiedby intercellular spaces due to the loosening of parenchymacells
In conclusion our results show that irradiation of the dryseeds of the Microtom cultivar did not hamper germinationand the development of functional leaves The formationof more compact plants characterised by a higher numberof larger leaves can be valuable in terms of increasingresource regeneration in the BLSSs High levels of radiationinduced only slight structural perturbations which weresimilar in juvenile and compound leaves Such perturbationsdid not affect photosynthetic efficiency since the maximalPSII photochemical efficiency was even increased after irra-diation at all doses However the tendency to accumulatelower amounts of phenolic compounds along chloroplastmembranes after irradiation of seeds with high doses ofradiation should be further investigated because it coulddetermine the reduction of natural photoprotection [57]Finally the overall results indicate that theMicrotom cultivarirradiated at the seed stage has high radioresistance duringthe growth which is a desirable feature for plant cultivation
in Space Radioresistance together with dwarf growth andshort life cycle makes this cultivar a valuable candidate forthe cultivation in BLSSs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are grateful to the ldquoCompagnia di San Paolordquo(Turin) and the ldquoPolo delle Scienze e delle Tecnologie perla Vitardquo of the University of Naples Federico II for thefinancial support The authors also thank Maria Giordanoand Pasquale Scognamiglio for their support in plant culti-vation and microscopy analyses The authors acknowledgefour anonymous editors for helpful comments on a previousversion of the paper
References
[1] A Real S Sundell-Bergman J F Knowles D S Woodheadand I Zinger ldquoEffects of ionising radiation exposure on plantsfish and mammals relevant data for environmental radiation
12 The Scientific World Journal
protectionrdquo Journal of Radiological Protection vol 24 no 4 ppA123ndashA137 2004
[2] V De Micco C Arena D Pignalosa and M Durante ldquoEffectsof sparsely and densely ionizing radiation on plantsrdquo Radiationand Environmental Biophysics vol 50 no 1 pp 1ndash19 2011
[3] N Kumari P Kumar D Mitra B Prasad B N Tiwary andL Varshney ldquoEffects of ionizing radiation on microbial decon-tamination phenolic contents and antioxidant properties oftriphalardquo Journal of Food Science vol 74 no 3 pp M109ndashM1132009
[4] A Tanaka N Shikazono and Y Hase ldquoStudies on biologicaleffects of ion beams on lethality molecular nature of mutationmutation rate and spectrum of mutation phenotype for muta-tion breeding in higher plantsrdquo Journal of Radiation Researchvol 51 no 3 pp 223ndash233 2010
[5] M Durante and A Kronenberg ldquoGround-based research withheavy ions for space radiation protectionrdquo Advances in SpaceResearch vol 35 no 2 pp 180ndash184 2005
[6] E Hu S I Bartsev and H Liu ldquoConceptual design of a biore-generative life support system containing crops and silkwormsrdquoAdvances in Space Research vol 45 no 7 pp 929ndash939 2010
[7] V deMicco G Aronne G Colla R Fortezza and S de PascaleldquoAgro-biology for bioregenerative life support systems in long-term space missions general constraints and the Italian effortsrdquoJournal of Plant Interactions vol 4 no 4 pp 241ndash252 2009
[8] T W Halstead and F R Dutcher ldquoPlants in spacerdquo AnnualReview of Plant Physiology vol 38 pp 317ndash345 1987
[9] V De Micco S De Pascale R Paradiso and G AronneldquoMicrogravity effects on different stages of higher plant life cycleand completion of the seed-to-seed cyclerdquo Plant Biology vol 16pp 31ndash38 2014
[10] C J Bennett C Pirim and T M Orlando ldquoSpace-weatheringof solar system bodies a laboratory perspectiverdquo ChemicalReviews vol 113 pp 9086ndash9150 2013
[11] R A Kerr ldquoRadiation will make astronautsrsquo trip to Mars evenriskierrdquo Science vol 340 article 1031 2014
[12] D M Hassler C Zeitlin R F Wimmer-Schweingruber et alldquoMarsrsquo surface radiation environment measured with the MarsScience Laboratoryrsquos Curiosity Roverrdquo Science vol 343 no6169 Article ID 1244797 2014
[13] M Durante and F A Cucinotta ldquoHeavy ion carcinogenesis andhuman space explorationrdquo Nature Reviews Cancer vol 8 no 6pp 465ndash472 2008
[14] C Arena V De Micco and A De Maio ldquoGrowth alterationand leaf biochemical responses in P vulgaris plants exposed todifferent doses of ionizing radiationrdquo Plant Biology vol 16 pp194ndash202 2014
[15] R W Holst and D J Nagel ldquoRadiation effects on plantsrdquo inPlants For Environmental Studies W Wang J W Gorsuch andJ S Hughes Eds pp 37ndash81 Lewis Publishers Boca Raton FlaUSA 1997
[16] T D Luckey Hormesis With Ionizing Radiation CRC PressBoca Raton Fla USA 1980
[17] D Marcu V Cristea and L Daraban ldquoDose-dependent effectsof gamma radiation on lettuce (Lactuca sativa var capitata)seedlingsrdquo International Journal of Radiation Biology vol 89 pp219ndash223 2013
[18] M Mei H Deng Y Lu et al ldquoMutagenic effects of heavy ionradiation in plantsrdquo Advances in Space Research vol 14 no 10pp 363ndash372 1994
[19] Y LiM Liu Z Cheng andY Sun ldquoSpace environment inducedmutations prefer to occur at polymorphic sites of rice genomesrdquoAdvances in Space Research vol 40 no 4 pp 523ndash527 2007
[20] D S Kim J-B Kim E J Goh et al ldquoAntioxidant response ofArabidopsis plants to gamma irradiation genome-wide expres-sion profiling of the ROS scavenging and signal transductionpathwaysrdquo Journal of Plant Physiology vol 168 no 16 pp 1960ndash1971 2011
[21] T J Brodribb ldquoXylem hydraulic physiology the functionalbackbone of terrestrial plant productivityrdquo Plant Science vol177 no 4 pp 245ndash251 2009
[22] R M Wheeler J C Sager R P Prince andW M Knott ldquoCropproduction for advanced life support systemsmdashobservationfrom the Kennedy Space Center Breadboard Projectrdquo NASATM-2003-211184 NASA Kennedy Space Center Florida 2003
[23] R M Wheeler C L Mackowiak G W Stutte et al ldquoCropproductivities and radiation use efficiencies for bioregenerativelife supportrdquo Advances in Space Research vol 41 no 5 pp 706ndash713 2008
[24] R Paradiso V De Micco R Buonomo G Aronne G Bar-bieri and S De Pascale ldquoSoilless cultivation of soybean forBioregenerative Life-Support Systems a literature review andthe experience of the MELiSSA ProjectmdashFood characterisationPhase Irdquo Plant Biology vol 16 pp 69ndash78 2014
[25] D M Porterfield ldquoThe biophysical limitations in physiologicaltransport and exchange in plants grown in microgravityrdquoJournal of Plant Growth Regulation vol 21 no 2 pp 177ndash1902002
[26] K Yokthongwattana and A Melis ldquoPhotoinhibition and recov-ery in oxygenic photosynthesis mechanism of a photosystem IIdamage and repair cyclerdquo in Photoprotection PhotoinhibitionGene Regulation and Environment B Demmig-Adams WW Adams III and A K Mattoo Eds pp 175ndash191 SpringerDordecht The Netherlands 2006
[27] M T Giardi G Rea M D Lambreva et al ldquoMutations ofphotosystem II D1 protein that empower efficient phenotypesof Chlamydomonas reinhardtii under extreme environment inSpacerdquo PLoS ONE vol 8 Article ID e64352 2013
[28] C Arena V De Micco G Aronne M Pugliese A Virzo DeSanto andADeMaio ldquoResponse ofPhaseolus vulgarisL plantsto low-let ionizing radiation growth and oxidative stressrdquo ActaAstronautica vol 91 pp 107ndash114 2013
[29] V Lattanzio P A Kroon and S Quideau ldquoPlant phenolicsmdashsecondary metabolites with diverse functionsrdquo in RecentAdvances in Polyphenol Research F Daay and V Lattanzio Edspp 1ndash35 Wiley-Blackwell Oxford UK 2008
[30] V De Micco C Arena and G Aronne ldquoAnatomical alterationsof Phaseolus vulgaris L mature leaves irradiated with X-raysrdquoPlant Biology vol 16 pp 187ndash193 2014
[31] A D Krikorian ldquoPlants and somatic embryos in space whathave we learnedrdquo Gravitational and Space Biology Bulletin vol11 no 2 pp 5ndash14 1998
[32] R Meissner Y Jacobson S Melamed et al ldquoA new modelsystem for tomato geneticsrdquoThe Plant Journal vol 12 no 6 pp1465ndash1472 1997
[33] E Emmanuel and A A Levy ldquoTomato mutants as tools forfunctional genomicsrdquo Current Opinion in Plant Biology vol 5no 2 pp 112ndash117 2002
[34] E Martı C Gisbert G J Bishop M S Dixon and J LGarcıa-Martınez ldquoGenetic and physiological characterizationof tomato cv Micro-Tomrdquo Journal of Experimental Botany vol57 pp 2037ndash2047 2006
The Scientific World Journal 13
[35] T Kurimoto J V H Constable and A Huda ldquoEffects ofionizing radiation exposure on arabidopsis thalianardquo HealthPhysics vol 99 no 1 pp 49ndash57 2010
[36] L Reale D Gigante F Landucci F Ferranti and R VenanzonildquoMorphological and histo-anatomical traits reflect die-back inPhragmites australis (Cav) Steudrdquo Aquatic Botany vol 103 pp122ndash128 2012
[37] K Fukuzawa ldquoUltraviolet microscopyrdquo in Methods in LigninChemistry S Y Lin andCWDence Eds pp 110ndash131 SpringerBerlin Germany 1992
[38] S E Ruzin Plant Microtechnique and Microscopy OxfordUniversity Press New York NY USA 1999
[39] V De Micco G Aronne J-P Joseleau and K Ruel ldquoXylemdevelopment and cell wall changes of soybean seedlings grownin spacerdquo Annals of Botany vol 101 no 5 pp 661ndash669 2008
[40] S Van Buggenhout T Grauwet A Van Loey and M Hen-drickx ldquoStructureprocessing relation of vacuum infused straw-berry tissue frozen under different conditionsrdquo European FoodResearch and Technology vol 226 no 3 pp 437ndash448 2008
[41] V De Micco and G Aronne ldquoBiometric anatomy of seedlingsdeveloped onboard of Foton-M2 in an automatic system sup-porting growthrdquoActa Astronautica vol 62 no 8-9 pp 505ndash5132008
[42] B Genty J M Briantais and N R Baker ldquoThe relationshipbetween quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescencerdquo Biochimica andBiophysica Acta vol 990 pp 87ndash92 1989
[43] W Bilger and O Bjorkman ldquoRole of the xanthophyll cycle inphotoprotection elucidated by measurements of light-inducedabsorbance changes fluorescence and photosynthesis in leavesof Hedera canariensisrdquo Photosynthesis Research vol 25 no 3pp 173ndash185 1990
[44] H K Lichtenthaler ldquoChlorophylls and carotenoids pigmentsof photosynthetic biomembranesrdquoMethods in Enzymology vol148 pp 350ndash382 1987
[45] J P Maity DMishra A Chakraborty A Saha S C Santra andS Chanda ldquoModulation of some quantitative and qualitativecharacteristics in rice (Oryza sativa L) and mung (Phaseolusmungo L) by ionizing radiationrdquo Radiation Physics and Chem-istry vol 74 no 5 pp 391ndash394 2005
[46] L Cheng H Yang B Lin et al ldquoEffect of gamma-ray radiationon physiological morphological characters and chromosomeaberrations of minitubers in Solanum tuberosum Lrdquo Interna-tional Journal of Radiation Biology vol 86 no 9 pp 791ndash7992010
[47] B Al-Safadi and R Elias ldquoImprovement of caper (Capparisspinosa L) propagation using in vitro culture and gammairradiationrdquo Scientia Horticulturae vol 127 no 3 pp 290ndash2972011
[48] O Monje G W Stutte G D Goins D M Porterfield and GE Bingham ldquoFarming in space environmental and biophysicalconcernsrdquo Advances in Space Research vol 31 no 1 pp 151ndash1672003
[49] V De Micco M Scala and G Aronne ldquoEffects of simulatedmicrogravity on male gametophyte of Prunus Pyrus andBrassica speciesrdquo Protoplasma vol 228 no 1ndash3 pp 121ndash1262006
[50] V De Micco R Buonomo R Paradiso S De Pascale and GAronne ldquoSoybean cultivar selection for Bioregenerative LifeSupport Systems (BLSS)mdashtheoretical selectionrdquo Advances inSpace Research vol 49 no 10 pp 1415ndash1421 2012
[51] T Kozuka S-G Kong M Doi K-I Shimazaki and ANagatani ldquoTissue-autonomous promotion of palisade celldevelopment by phototropin 2 in Arabidopsisrdquo Plant Cell vol23 no 10 pp 3684ndash3695 2011
[52] E Kovacs and A Keresztes ldquoEffect of gamma and UV-BCradiation on plant cellsrdquoMicron vol 33 no 2 pp 199ndash210 2002
[53] R Ogaya J Penuelas D Asensio and J Llusia ldquoChlorophyllfluorescence responses to temperature and water availabilityin two co-dominant Mediterranean shrub and tree speciesin a long-term field experiment simulating climate changerdquoEnvironmental and Experimental Botany vol 73 no 1 pp 89ndash93 2011
[54] R Zaka C M Vandecasteele and M T Misset ldquoEffects oflow chronic doses of ionizing radiation on antioxidant enzymesand G6PDH activities in Stipa capillata (Poaceae)rdquo Journal ofExperimental Botany vol 53 no 376 pp 1979ndash1987 2002
[55] J-H KimM-H Baek Y C Byung GW Seung and J-S KimldquoAlterations in the photosynthetic pigments and antioxidantmachineries of red pepper (Capsicum annuum L) seedlingsfrom gamma-irradiated seedsrdquo Journal of Plant Biology vol 47no 4 pp 314ndash321 2004
[56] N A Al-Enezi and J M Al-Khayri ldquoAlterations of DNA ionsand photosynthetic pigments content in date palm seedlingsinduced by X-irradiationrdquo International Journal of Agricultureand Biology vol 14 pp 329ndash336 2012
[57] V de Micco C Arena L Vitale G Aronne and A V de SantoldquoAnatomy and photochemical behaviour of mediterranean Cis-tus incanus winter leaves under natural outdoor and warmerindoor conditionsrdquo Botany vol 89 no 10 pp 677ndash688 2011
[58] L H Levine H G Levine E C Stryjewski V Prima and WC Piastuch ldquoEffect of spaceflight on isoflavonoid accumulationin etiolated soybean seedlingsrdquo Journal of Gravitational Physiol-ogy vol 8 no 2 pp 21ndash27 2001
[59] X Fan ldquoAntioxidant capacity of fresh-cut vegetables exposedto ionizing radiationrdquo Journal of the Science of Food andAgriculture vol 85 no 6 pp 995ndash1000 2005
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
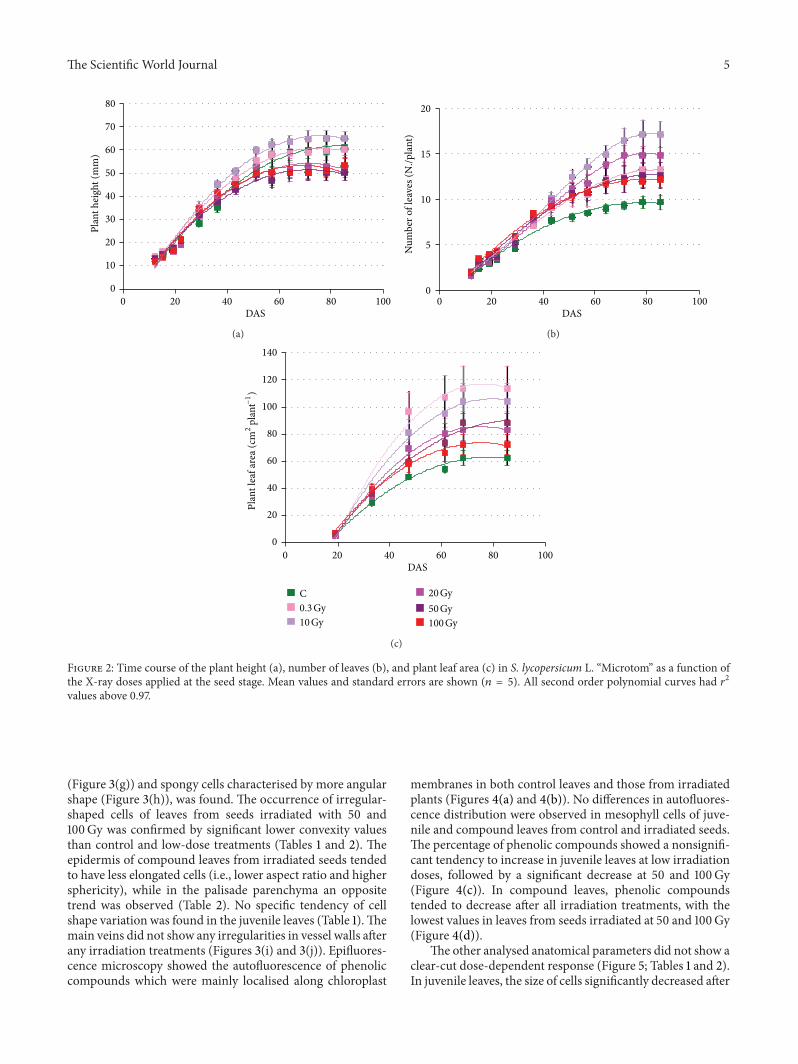
The Scientific World Journal 5
0
10
20
30
40
50
60
70
80
0 20 40 60 80 100
Plan
t hei
ght (
mm
)
DAS
(a)
0
5
10
15
20
Num
ber o
f lea
ves (
Np
lant
)
0 20 40 60 80 100DAS
(b)
0 20 40 60 80 100DAS
0
20
40
60
80
100
120
140
Plan
t lea
f are
a (cm
2pl
antminus1)
C03 Gy10 Gy
20 Gy50 Gy100 Gy
(c)
Figure 2 Time course of the plant height (a) number of leaves (b) and plant leaf area (c) in S lycopersicum L ldquoMicrotomrdquo as a function ofthe X-ray doses applied at the seed stage Mean values and standard errors are shown (119899 = 5) All second order polynomial curves had 1199032values above 097
(Figure 3(g)) and spongy cells characterised by more angularshape (Figure 3(h)) was found The occurrence of irregular-shaped cells of leaves from seeds irradiated with 50 and100Gy was confirmed by significant lower convexity valuesthan control and low-dose treatments (Tables 1 and 2) Theepidermis of compound leaves from irradiated seeds tendedto have less elongated cells (ie lower aspect ratio and highersphericity) while in the palisade parenchyma an oppositetrend was observed (Table 2) No specific tendency of cellshape variation was found in the juvenile leaves (Table 1)Themain veins did not show any irregularities in vessel walls afterany irradiation treatments (Figures 3(i) and 3(j)) Epifluores-cence microscopy showed the autofluorescence of phenoliccompounds which were mainly localised along chloroplast
membranes in both control leaves and those from irradiatedplants (Figures 4(a) and 4(b)) No differences in autofluores-cence distribution were observed in mesophyll cells of juve-nile and compound leaves from control and irradiated seedsThe percentage of phenolic compounds showed a nonsignifi-cant tendency to increase in juvenile leaves at low irradiationdoses followed by a significant decrease at 50 and 100Gy(Figure 4(c)) In compound leaves phenolic compoundstended to decrease after all irradiation treatments with thelowest values in leaves from seeds irradiated at 50 and 100Gy(Figure 4(d))
The other analysed anatomical parameters did not show aclear-cut dose-dependent response (Figure 5 Tables 1 and 2)In juvenile leaves the size of cells significantly decreased after
6 The Scientific World Journal
(b) (c)
(d) (e)
(f)
(g) (h)
(i)
(j)
(a)
Figure 3 Light microscopy views of cross sections of leaf lamina of S lycopersicum L ldquoMicrotomrdquo compound leaves from plants developedfrom control (a) and irradiated seeds 03 Gy (b) 10Gy (c) 20Gy (d) 50Gy (e g) and 100Gy (f h) Detail of xylem in the main vein is shownin leaves from control (i) and 100Gy irradiated seeds (j) Images (andashh) are at the same magnification Bar = 50 120583m
the irradiation of seeds only in palisade parenchyma withthe lowest values at 20Gy (Figures 5(c) 5(e) 5(g) and 5(i))By contrast in compound leaves the application of 100Gyresulted in increased cell size compared with control andleaves from seeds irradiated up to 50Gy (Figures 5(d) 5(f)5(h) and 5(j))
34 Effect of Ionizing Radiation on Fluorescence Measure-ments and Pigment Content No significant differences were
detectable in fluorescence parameters and total photosyn-thetic pigment content in plants grown from irradiated seedsat different X-rays doses (Figure 6) A tendency to NPQincrease was found at 03 and 10Gy followed by a decreaseat doses higher than 20Gy However the lack of significantdifferences in ΦPSII and NPQ among treatments indicatedthat X-rays delivered at the seed stage on tomato plantsdid not alter the capability of photosynthetic apparatus oftomato plants either to convert light energy in photochem-istry or to perform thermal dissipation mechanisms The
The Scientific World Journal 7
(a) (b)
Phen
olic
com
poun
ds (
)
aa a
aab
b
0
10
20
30
40
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf JL
(c)
Phen
olic
com
poun
ds (
) aab
abc ab
c bc
0
10
20
30
40
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf CL
(d)
Figure 4 Epifluorescence microscopy views of lamina cross sections of S lycopersicum L ldquoMicrotomrdquo compound leaves from plantsdeveloped from control (a) and 100Gy irradiated seeds (b) Phenolic compounds along chloroplast membranes are autofluorescent Percentof mesophyll occupied by phenolic compounds () is shown in (c) and (d) for juvenile (JL) and compound (CL) leaves respectively Meanvalues and standard errors are reported (119899 = 15) different letters correspond to significantly different values after multiple comparison tests(119875 lt 005)
maximum photochemical efficiency of PSII (FvFm) washigher in leaves from irradiated seeds than control anddid not show variations among different X-rays treatmentsthis indicated the maintenance of higher level of photo-synthetic activity after irradiation irrespectively from dose(Figure 6(c))
The total chlorophyll as well as the total carotenoidcontent did not show significant changes due to irradiationof seeds (Figure 7)
4 Discussion
Ionising radiation can have positive null or negative effectson plant development depending on the properties of theradiation the features of the plant material and possibleinteractions with other environmental factors [2] There iscommon agreement that plants are characterised by highradioresistance moreover low-LET ionising radiation hasbeen reported to increase germination percentage rootlength plant height and other growth parameters especiallyat low doses which induce hormesis [16 17 45ndash47] Ourresults suggest that S lycopersicum ldquoMicrotomrdquo is charac-terised by high radioresistance at the seed stage indeed
irradiation of dry seeds with a wide range of X-ray dosesdid not hamper germination and plant growth up to thecompletion of the crop cycle An hormesis effect was evidentsince growth either remained unaffected or was positivelyaffected especially by low doses of X-rays Apart from plantsgrown from seeds irradiated with 03 and 10Gy all plantsfrom irradiated seeds were characterised by a more compacthabitus higher number of larger leaves was not accompaniedby increased height The radiation-induced more compactgrowth pattern is positive in BLSSs in Spacewhere availabilityof volume for growth is a major technical constraint [7 4849] In the cultivar Microtom irradiation with doses equalto or higher than 20Gy caused a decrease in plant heightin agreement with the induction of dwarf or semidwarfgrowth reported as a general effect of exposure to ionisingradiation [18] Hence irradiation on the ldquoMicrotomrdquo seedswould intensify the trait of dwarf growth which alreadycharacterise this cultivar and should be taken into account forcultivar selection for the BLSSs [50] Any variations in growthparameters in Space-oriented ecologically closed systemscan determine valuable changes in terms of optimisationof volume and resource recycling These changes can bestraightaway converted into economic saving or waste [9]For example the increase in the number and size of leaves
8 The Scientific World Journal
Palisade parenchyma Spongy parenchyma
ab b ab a ab ab
a a a a a a
0010203040506
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf JL
Lam
ina t
hick
ness
(mm
)
(a)
Palisade parenchyma Spongy parenchyma
b b b b b a
a ab ab b abab
Leaf CL
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
010203040506
Lam
ina t
hick
ness
(mm
)
(b)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a a aa a
a
0200400600800
1000Upper epidermis
Cel
l are
a (120583
m2)
(c)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
1000
a a a a aa
Upper epidermis
Cel
l are
a (120583
m2)
(d)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
a bc bc c bc b
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(e)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
ab ab b ab ab a
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(f)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a aa
a a a
0200400600800
100012001400
Cel
l are
a (120583
m2)
(g)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
100012001400
b b b b ba
Cel
l are
a (120583
m2)
(h)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
a aa
a a a
0
200
400
600
800
Cel
l are
a (120583
m2)
(i)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
b b b b ba
0
200
400
600
800
Cel
l are
a (120583
m2)
(j)
Figure 5 Lamina thickness (a b) and cell area of upper epidermis (c d) palisade (e f) spongy (g h) parenchymas and lower epidermis (i j)of S lycopersicum L ldquoMicrotomrdquo leaves of control and irradiated plants at different doses of X-rays in both juvenile (JL) and compound (CL)leaves Mean values and standard errors are shown (119899 = 15 in (a b) 119899 = 45 in (cndashj)) Different letters correspond to significantly differentvalues according to multiple comparison tests (119875 lt 005)
The Scientific World Journal 9
04
05
06
07
08
a a a a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
ΦPS
II
(a)
00
02
04
06
08
NPQ a
a aa a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
(b)
070
075
080
085
090
a
b b b b b
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
FF
m
(c)
Figure 6 Quantum yield of PSII electron transport (ΦPSII a) nonphotochemical quenching (NPQ b) and maximal PSII photochemicalefficiency (FvFm c) in compound (CL) S lycopersicum L ldquoMicrotomrdquo leaves of control and plants from seeds irradiated at different dosesof X-rays Mean values and standard deviations are shown (119899 = 5) Different letters correspond to significantly different values according tomultiple comparison tests (119875 lt 005)
0
1
2
3
4
a
aa
a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Chl a
+ b
(mg g
minus1)
(a)
0
02
04
06
a
aa a
a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Car x
+ c
(mg g
minus1)
(b)
Figure 7 Total chlorophyll content (Chl a+b) (a) and total carotenoid content (Car x+c) (b) in compound (CL) S lycopersicum L ldquoMicrotomrdquoleaves of control and plants from seeds irradiated at different doses of X-rays Mean values and standard errors are shown (119899 = 5) Differentletters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
induced in ldquoMicrotomrdquo especially by low doses of irradiationcan be beneficial to atmosphere regeneration of pressurisedmodules provided that formed leaves do not show structuralaberrations which reduce their function
General anatomical structure of both juvenile and com-pound ldquoMicrotomrdquo leaves was only slightly perturbed inplants from seeds subjected to the highest levels of radiation
Clear-cut dose-depended tendencies of variations were notfound in lamina thickness and cell size and shape of thevarious tissues The formation of smaller palisade cells wasfound only in juvenile leaves of irradiated seeds while anincrease in cell size was found only in spongy parenchymaand lower epidermis of compound leaves from seeds irra-diated at 100Gy The diverse growth behaviour found in
10 The Scientific World Journal
Table 1 Cell shape in different tissues of juvenile leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown (119899 = 75)Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf JL Aspect ratio Sphericity Convexity
Upper epidermis
C 197 plusmn 016 a 0308 plusmn 0050 a 0926 plusmn 0021 ab03Gy 200 plusmn 013 a 0290 plusmn 0038 a 0941 plusmn 0013 a10Gy 210 plusmn 012 a 0239 plusmn 0037 a 0898 plusmn 0016 ab20Gy 217 plusmn 014 a 0258 plusmn 0045 a 0909 plusmn 0019 abc50Gy 170 plusmn 008 a 0376 plusmn 0037 a 0927 plusmn 0016 ab100Gy 197 plusmn 014 a 0286 plusmn 0042 a 0876 plusmn 0028 c
Palisade parenchyma
C 290 plusmn 010 a 0109 plusmn 0015 a 0879 plusmn 0010 a03 Gy 304 plusmn 012 a 0099 plusmn 0016 a 0805 plusmn 0016 b10Gy 309 plusmn 013 a 0093 plusmn 0019 a 0792 plusmn 0015 b20Gy 283 plusmn 011 a 0101 plusmn 0017 a 0784 plusmn 0017 b50Gy 314 plusmn 014 a 0082 plusmn 0016 a 0772 plusmn 0016 b100Gy 275 plusmn 011 a 0119 plusmn 0020 a 0805 plusmn 0017 b
Spongy parenchyma
C 190 plusmn 008 a 0307 plusmn 0033 a 0893 plusmn 0012 a03 Gy 180 plusmn 007 a 0341 plusmn 0029 a 0876 plusmn 0013 a10Gy 203 plusmn 010 a 0290 plusmn 0031 a 0901 plusmn 0012 a20Gy 194 plusmn 012 a 0344 plusmn 0040 a 0881 plusmn 0015 a50Gy 174 plusmn 009 a 0393 plusmn 0035 a 0883 plusmn 0014 a100Gy 198 plusmn 012 a 0322 plusmn 0037 a 0878 plusmn 0015 a
Lower epidermis
C 178 plusmn 009 bc 0343 plusmn 0045 ab 0919 plusmn 0019 a03 Gy 225 plusmn 017 ab 0263 plusmn 0044 b 0933 plusmn 0024 a10Gy 219 plusmn 011 abc 0218 plusmn 0034 b 0888 plusmn 0020 ab20Gy 180 plusmn 013 bc 0378 plusmn 0047 ab 0926 plusmn 0017 a50Gy 167 plusmn 001 c 0461 plusmn 0051 a 0924 plusmn 0021 a100Gy 238 plusmn 021 a 0257 plusmn 0057 b 0857 plusmn 0030 b
the various tissues of structures of different age is in agree-ment with the different light-mediated control of cell devel-opment in the various leaf tissues [51] Altered mesophyllcharacterized by partly collapsed cells with irregular cellshape was found only sporadically after the exposure of seedsto 50 and 100Gy in theMicrotomcultivar Suchmodificationswere less marked than those found in leaves of bean wherethe target organs for irradiation were the leaves themselves[30] leaves directly exposed to X-rays in bean showed theformation of more loose parenchyma with larger cells likelydue to a radiation-induced damage to cell walls that reducedmechanical constraints to cell enlargement [30 52] Furtherinvestigations on cell wall composition would be useful toelucidate whether a possible alteration of cell walls wasresponsible for cell enlargement in the spongy parenchymaand epidermis of leaves from seeds of ldquoMicrotomrdquo irradiatedwith the highest dose of X-rays However in the cultivarMicrotom the slight alterations of mesophyll structure didnot impair photosynthetic efficiency as demonstrated by thelack of significant dose-dependent variation trends in thequantum yield of PSII electron transport (ΦPSII) nonphoto-chemical quenching (NPQ) and maximal PSII photochemi-cal efficiency (FvFm) The increased values of FvFm closeto 08 which is considered an optimal value for healthy plants[53] suggested that the photosynthetic apparatus of the leavesfrom irradiated seeds has higher levels of photochemical
efficiency irrespective of dose The unchanged or increasedcontent of both chlorophyll and carotenoid pigments con-firmed that the photosynthetic apparatus remained stablealso in terms of light harvesting capacity and indicatedno deleterious effect of radiation on plant photosyntheticmachinery RecentlyMarcu et al [17] showed that irradiationof lettuce seeds with gamma rays up to 30Gy enhancedthe content of chlorophyll a chlorophyll b and carotenoidswhile higher doses (up to 70Gy) resulted in their decreaseHowever the effect of ionising radiation on photosyntheticpigment content is quite controversial Increased pigmentcontent is ascribed to the phenomenon of hormesis whilea lower content is generally ascribed to the inhibition ofchlorophyll and carotenoid synthesis or to oxidation due tothe high levels of radiation-induced water radiolysis [2 1454ndash56] Some plants possess natural screens against radiationwhen their tissues contain phenolic compounds with antioxi-dant functions and can counteract photoinhibitory processes[29 57] An increase of phenolic compounds linked to thechloroplast membranes has been recently found in Phaseolusvulgaris irradiated leaves exposed to high levels of X-rays[28 30] Changes in phenolic content after the exposure toSpace can be attributed either to direct effects on the synthesisor mobilization of phenolic compounds or to indirect effectsdue to changes in ultrastructural organization such as theincrease in the number of chloroplasts per cell [30 58]
The Scientific World Journal 11
Table 2 Cell shape in different tissues of compound leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown(119899 = 75) Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf CL Aspect ratio Sphericity Convexity
Upper epidermis
C 166 plusmn 007 c 0422 plusmn 0042 a 0946 plusmn 0010 a03 Gy 216 plusmn 017 ab 0244 plusmn 0033 c 0925 plusmn 0016 ab10Gy 171 plusmn 011 bc 0401 plusmn 0042 a 0914 plusmn 0013 b20Gy 186 plusmn 010 abc 0328 plusmn 0041 ab 0919 plusmn 0020 ab50Gy 225 plusmn 018 a 0247 plusmn 0041 c 0897 plusmn 0019 b100Gy 188 plusmn 009 abc 0319 plusmn 0045 ab 0902 plusmn 0016 b
Palisade parenchyma
C 355 plusmn 012 a 0073 plusmn 0014 b 0890 plusmn 0012 a03 Gy 289 plusmn 010 b 0102 plusmn 0013 ab 0847 plusmn 0018 b10Gy 300 plusmn 014 b 0104 plusmn 0021 ab 0826 plusmn 0017 b20Gy 302 plusmn 011 b 0094 plusmn 0013 ab 0810 plusmn 0014 b50Gy 283 plusmn 012 b 0107 plusmn 0021 ab 0750 plusmn 0021 c100Gy 275 plusmn 012 b 0122 plusmn 0023 a 0809 plusmn 0015 b
Spongy parenchyma
C 193 plusmn 010 a 0323 plusmn 0035 a 0904 plusmn 0011 a03 Gy 183 plusmn 010 a 0375 plusmn 0042 a 0879 plusmn 0013 a10Gy 176 plusmn 007 a 0357 plusmn 0034 a 0869 plusmn 0011 a20Gy 180 plusmn 009 a 0366 plusmn 0038 a 0885 plusmn 0016 a50Gy 171 plusmn 006 a 0372 plusmn 0034 a 0879 plusmn 0015 a100Gy 188 plusmn 010 a 0339 plusmn 0037 a 0883 plusmn 0015 a
Lower epidermis
C 148 plusmn 008 b 0545 plusmn 0045 a 0957 plusmn 0015 a03 Gy 210 plusmn 017 a 0295 plusmn 0046 b 0929 plusmn 0019 ab10Gy 190 plusmn 012 ab 0334 plusmn 0043 b 0893 plusmn 0019 b20Gy 178 plusmn 011 ab 0388 plusmn 0047 b 0929 plusmn 0017 ab50Gy 172 plusmn 009 ab 0367 plusmn 0040 b 0914 plusmn 0016 b100Gy 191 plusmn 014 ab 0362 plusmn 0054 b 0888 plusmn 0025 b
The lowering of phenolic content at high levels of X-rays inldquoMicrotomrdquo leaves is not in agreement with general trendsindicating enhanced phenolics production in plants grownin Space [58] or in various plant tissues directly irradiatedwith gamma rays [59] However the low content of phenolicscan be an indirect effect of increasing volume occupiedby intercellular spaces due to the loosening of parenchymacells
In conclusion our results show that irradiation of the dryseeds of the Microtom cultivar did not hamper germinationand the development of functional leaves The formationof more compact plants characterised by a higher numberof larger leaves can be valuable in terms of increasingresource regeneration in the BLSSs High levels of radiationinduced only slight structural perturbations which weresimilar in juvenile and compound leaves Such perturbationsdid not affect photosynthetic efficiency since the maximalPSII photochemical efficiency was even increased after irra-diation at all doses However the tendency to accumulatelower amounts of phenolic compounds along chloroplastmembranes after irradiation of seeds with high doses ofradiation should be further investigated because it coulddetermine the reduction of natural photoprotection [57]Finally the overall results indicate that theMicrotom cultivarirradiated at the seed stage has high radioresistance duringthe growth which is a desirable feature for plant cultivation
in Space Radioresistance together with dwarf growth andshort life cycle makes this cultivar a valuable candidate forthe cultivation in BLSSs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are grateful to the ldquoCompagnia di San Paolordquo(Turin) and the ldquoPolo delle Scienze e delle Tecnologie perla Vitardquo of the University of Naples Federico II for thefinancial support The authors also thank Maria Giordanoand Pasquale Scognamiglio for their support in plant culti-vation and microscopy analyses The authors acknowledgefour anonymous editors for helpful comments on a previousversion of the paper
References
[1] A Real S Sundell-Bergman J F Knowles D S Woodheadand I Zinger ldquoEffects of ionising radiation exposure on plantsfish and mammals relevant data for environmental radiation
12 The Scientific World Journal
protectionrdquo Journal of Radiological Protection vol 24 no 4 ppA123ndashA137 2004
[2] V De Micco C Arena D Pignalosa and M Durante ldquoEffectsof sparsely and densely ionizing radiation on plantsrdquo Radiationand Environmental Biophysics vol 50 no 1 pp 1ndash19 2011
[3] N Kumari P Kumar D Mitra B Prasad B N Tiwary andL Varshney ldquoEffects of ionizing radiation on microbial decon-tamination phenolic contents and antioxidant properties oftriphalardquo Journal of Food Science vol 74 no 3 pp M109ndashM1132009
[4] A Tanaka N Shikazono and Y Hase ldquoStudies on biologicaleffects of ion beams on lethality molecular nature of mutationmutation rate and spectrum of mutation phenotype for muta-tion breeding in higher plantsrdquo Journal of Radiation Researchvol 51 no 3 pp 223ndash233 2010
[5] M Durante and A Kronenberg ldquoGround-based research withheavy ions for space radiation protectionrdquo Advances in SpaceResearch vol 35 no 2 pp 180ndash184 2005
[6] E Hu S I Bartsev and H Liu ldquoConceptual design of a biore-generative life support system containing crops and silkwormsrdquoAdvances in Space Research vol 45 no 7 pp 929ndash939 2010
[7] V deMicco G Aronne G Colla R Fortezza and S de PascaleldquoAgro-biology for bioregenerative life support systems in long-term space missions general constraints and the Italian effortsrdquoJournal of Plant Interactions vol 4 no 4 pp 241ndash252 2009
[8] T W Halstead and F R Dutcher ldquoPlants in spacerdquo AnnualReview of Plant Physiology vol 38 pp 317ndash345 1987
[9] V De Micco S De Pascale R Paradiso and G AronneldquoMicrogravity effects on different stages of higher plant life cycleand completion of the seed-to-seed cyclerdquo Plant Biology vol 16pp 31ndash38 2014
[10] C J Bennett C Pirim and T M Orlando ldquoSpace-weatheringof solar system bodies a laboratory perspectiverdquo ChemicalReviews vol 113 pp 9086ndash9150 2013
[11] R A Kerr ldquoRadiation will make astronautsrsquo trip to Mars evenriskierrdquo Science vol 340 article 1031 2014
[12] D M Hassler C Zeitlin R F Wimmer-Schweingruber et alldquoMarsrsquo surface radiation environment measured with the MarsScience Laboratoryrsquos Curiosity Roverrdquo Science vol 343 no6169 Article ID 1244797 2014
[13] M Durante and F A Cucinotta ldquoHeavy ion carcinogenesis andhuman space explorationrdquo Nature Reviews Cancer vol 8 no 6pp 465ndash472 2008
[14] C Arena V De Micco and A De Maio ldquoGrowth alterationand leaf biochemical responses in P vulgaris plants exposed todifferent doses of ionizing radiationrdquo Plant Biology vol 16 pp194ndash202 2014
[15] R W Holst and D J Nagel ldquoRadiation effects on plantsrdquo inPlants For Environmental Studies W Wang J W Gorsuch andJ S Hughes Eds pp 37ndash81 Lewis Publishers Boca Raton FlaUSA 1997
[16] T D Luckey Hormesis With Ionizing Radiation CRC PressBoca Raton Fla USA 1980
[17] D Marcu V Cristea and L Daraban ldquoDose-dependent effectsof gamma radiation on lettuce (Lactuca sativa var capitata)seedlingsrdquo International Journal of Radiation Biology vol 89 pp219ndash223 2013
[18] M Mei H Deng Y Lu et al ldquoMutagenic effects of heavy ionradiation in plantsrdquo Advances in Space Research vol 14 no 10pp 363ndash372 1994
[19] Y LiM Liu Z Cheng andY Sun ldquoSpace environment inducedmutations prefer to occur at polymorphic sites of rice genomesrdquoAdvances in Space Research vol 40 no 4 pp 523ndash527 2007
[20] D S Kim J-B Kim E J Goh et al ldquoAntioxidant response ofArabidopsis plants to gamma irradiation genome-wide expres-sion profiling of the ROS scavenging and signal transductionpathwaysrdquo Journal of Plant Physiology vol 168 no 16 pp 1960ndash1971 2011
[21] T J Brodribb ldquoXylem hydraulic physiology the functionalbackbone of terrestrial plant productivityrdquo Plant Science vol177 no 4 pp 245ndash251 2009
[22] R M Wheeler J C Sager R P Prince andW M Knott ldquoCropproduction for advanced life support systemsmdashobservationfrom the Kennedy Space Center Breadboard Projectrdquo NASATM-2003-211184 NASA Kennedy Space Center Florida 2003
[23] R M Wheeler C L Mackowiak G W Stutte et al ldquoCropproductivities and radiation use efficiencies for bioregenerativelife supportrdquo Advances in Space Research vol 41 no 5 pp 706ndash713 2008
[24] R Paradiso V De Micco R Buonomo G Aronne G Bar-bieri and S De Pascale ldquoSoilless cultivation of soybean forBioregenerative Life-Support Systems a literature review andthe experience of the MELiSSA ProjectmdashFood characterisationPhase Irdquo Plant Biology vol 16 pp 69ndash78 2014
[25] D M Porterfield ldquoThe biophysical limitations in physiologicaltransport and exchange in plants grown in microgravityrdquoJournal of Plant Growth Regulation vol 21 no 2 pp 177ndash1902002
[26] K Yokthongwattana and A Melis ldquoPhotoinhibition and recov-ery in oxygenic photosynthesis mechanism of a photosystem IIdamage and repair cyclerdquo in Photoprotection PhotoinhibitionGene Regulation and Environment B Demmig-Adams WW Adams III and A K Mattoo Eds pp 175ndash191 SpringerDordecht The Netherlands 2006
[27] M T Giardi G Rea M D Lambreva et al ldquoMutations ofphotosystem II D1 protein that empower efficient phenotypesof Chlamydomonas reinhardtii under extreme environment inSpacerdquo PLoS ONE vol 8 Article ID e64352 2013
[28] C Arena V De Micco G Aronne M Pugliese A Virzo DeSanto andADeMaio ldquoResponse ofPhaseolus vulgarisL plantsto low-let ionizing radiation growth and oxidative stressrdquo ActaAstronautica vol 91 pp 107ndash114 2013
[29] V Lattanzio P A Kroon and S Quideau ldquoPlant phenolicsmdashsecondary metabolites with diverse functionsrdquo in RecentAdvances in Polyphenol Research F Daay and V Lattanzio Edspp 1ndash35 Wiley-Blackwell Oxford UK 2008
[30] V De Micco C Arena and G Aronne ldquoAnatomical alterationsof Phaseolus vulgaris L mature leaves irradiated with X-raysrdquoPlant Biology vol 16 pp 187ndash193 2014
[31] A D Krikorian ldquoPlants and somatic embryos in space whathave we learnedrdquo Gravitational and Space Biology Bulletin vol11 no 2 pp 5ndash14 1998
[32] R Meissner Y Jacobson S Melamed et al ldquoA new modelsystem for tomato geneticsrdquoThe Plant Journal vol 12 no 6 pp1465ndash1472 1997
[33] E Emmanuel and A A Levy ldquoTomato mutants as tools forfunctional genomicsrdquo Current Opinion in Plant Biology vol 5no 2 pp 112ndash117 2002
[34] E Martı C Gisbert G J Bishop M S Dixon and J LGarcıa-Martınez ldquoGenetic and physiological characterizationof tomato cv Micro-Tomrdquo Journal of Experimental Botany vol57 pp 2037ndash2047 2006
The Scientific World Journal 13
[35] T Kurimoto J V H Constable and A Huda ldquoEffects ofionizing radiation exposure on arabidopsis thalianardquo HealthPhysics vol 99 no 1 pp 49ndash57 2010
[36] L Reale D Gigante F Landucci F Ferranti and R VenanzonildquoMorphological and histo-anatomical traits reflect die-back inPhragmites australis (Cav) Steudrdquo Aquatic Botany vol 103 pp122ndash128 2012
[37] K Fukuzawa ldquoUltraviolet microscopyrdquo in Methods in LigninChemistry S Y Lin andCWDence Eds pp 110ndash131 SpringerBerlin Germany 1992
[38] S E Ruzin Plant Microtechnique and Microscopy OxfordUniversity Press New York NY USA 1999
[39] V De Micco G Aronne J-P Joseleau and K Ruel ldquoXylemdevelopment and cell wall changes of soybean seedlings grownin spacerdquo Annals of Botany vol 101 no 5 pp 661ndash669 2008
[40] S Van Buggenhout T Grauwet A Van Loey and M Hen-drickx ldquoStructureprocessing relation of vacuum infused straw-berry tissue frozen under different conditionsrdquo European FoodResearch and Technology vol 226 no 3 pp 437ndash448 2008
[41] V De Micco and G Aronne ldquoBiometric anatomy of seedlingsdeveloped onboard of Foton-M2 in an automatic system sup-porting growthrdquoActa Astronautica vol 62 no 8-9 pp 505ndash5132008
[42] B Genty J M Briantais and N R Baker ldquoThe relationshipbetween quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescencerdquo Biochimica andBiophysica Acta vol 990 pp 87ndash92 1989
[43] W Bilger and O Bjorkman ldquoRole of the xanthophyll cycle inphotoprotection elucidated by measurements of light-inducedabsorbance changes fluorescence and photosynthesis in leavesof Hedera canariensisrdquo Photosynthesis Research vol 25 no 3pp 173ndash185 1990
[44] H K Lichtenthaler ldquoChlorophylls and carotenoids pigmentsof photosynthetic biomembranesrdquoMethods in Enzymology vol148 pp 350ndash382 1987
[45] J P Maity DMishra A Chakraborty A Saha S C Santra andS Chanda ldquoModulation of some quantitative and qualitativecharacteristics in rice (Oryza sativa L) and mung (Phaseolusmungo L) by ionizing radiationrdquo Radiation Physics and Chem-istry vol 74 no 5 pp 391ndash394 2005
[46] L Cheng H Yang B Lin et al ldquoEffect of gamma-ray radiationon physiological morphological characters and chromosomeaberrations of minitubers in Solanum tuberosum Lrdquo Interna-tional Journal of Radiation Biology vol 86 no 9 pp 791ndash7992010
[47] B Al-Safadi and R Elias ldquoImprovement of caper (Capparisspinosa L) propagation using in vitro culture and gammairradiationrdquo Scientia Horticulturae vol 127 no 3 pp 290ndash2972011
[48] O Monje G W Stutte G D Goins D M Porterfield and GE Bingham ldquoFarming in space environmental and biophysicalconcernsrdquo Advances in Space Research vol 31 no 1 pp 151ndash1672003
[49] V De Micco M Scala and G Aronne ldquoEffects of simulatedmicrogravity on male gametophyte of Prunus Pyrus andBrassica speciesrdquo Protoplasma vol 228 no 1ndash3 pp 121ndash1262006
[50] V De Micco R Buonomo R Paradiso S De Pascale and GAronne ldquoSoybean cultivar selection for Bioregenerative LifeSupport Systems (BLSS)mdashtheoretical selectionrdquo Advances inSpace Research vol 49 no 10 pp 1415ndash1421 2012
[51] T Kozuka S-G Kong M Doi K-I Shimazaki and ANagatani ldquoTissue-autonomous promotion of palisade celldevelopment by phototropin 2 in Arabidopsisrdquo Plant Cell vol23 no 10 pp 3684ndash3695 2011
[52] E Kovacs and A Keresztes ldquoEffect of gamma and UV-BCradiation on plant cellsrdquoMicron vol 33 no 2 pp 199ndash210 2002
[53] R Ogaya J Penuelas D Asensio and J Llusia ldquoChlorophyllfluorescence responses to temperature and water availabilityin two co-dominant Mediterranean shrub and tree speciesin a long-term field experiment simulating climate changerdquoEnvironmental and Experimental Botany vol 73 no 1 pp 89ndash93 2011
[54] R Zaka C M Vandecasteele and M T Misset ldquoEffects oflow chronic doses of ionizing radiation on antioxidant enzymesand G6PDH activities in Stipa capillata (Poaceae)rdquo Journal ofExperimental Botany vol 53 no 376 pp 1979ndash1987 2002
[55] J-H KimM-H Baek Y C Byung GW Seung and J-S KimldquoAlterations in the photosynthetic pigments and antioxidantmachineries of red pepper (Capsicum annuum L) seedlingsfrom gamma-irradiated seedsrdquo Journal of Plant Biology vol 47no 4 pp 314ndash321 2004
[56] N A Al-Enezi and J M Al-Khayri ldquoAlterations of DNA ionsand photosynthetic pigments content in date palm seedlingsinduced by X-irradiationrdquo International Journal of Agricultureand Biology vol 14 pp 329ndash336 2012
[57] V de Micco C Arena L Vitale G Aronne and A V de SantoldquoAnatomy and photochemical behaviour of mediterranean Cis-tus incanus winter leaves under natural outdoor and warmerindoor conditionsrdquo Botany vol 89 no 10 pp 677ndash688 2011
[58] L H Levine H G Levine E C Stryjewski V Prima and WC Piastuch ldquoEffect of spaceflight on isoflavonoid accumulationin etiolated soybean seedlingsrdquo Journal of Gravitational Physiol-ogy vol 8 no 2 pp 21ndash27 2001
[59] X Fan ldquoAntioxidant capacity of fresh-cut vegetables exposedto ionizing radiationrdquo Journal of the Science of Food andAgriculture vol 85 no 6 pp 995ndash1000 2005
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 The Scientific World Journal
(b) (c)
(d) (e)
(f)
(g) (h)
(i)
(j)
(a)
Figure 3 Light microscopy views of cross sections of leaf lamina of S lycopersicum L ldquoMicrotomrdquo compound leaves from plants developedfrom control (a) and irradiated seeds 03 Gy (b) 10Gy (c) 20Gy (d) 50Gy (e g) and 100Gy (f h) Detail of xylem in the main vein is shownin leaves from control (i) and 100Gy irradiated seeds (j) Images (andashh) are at the same magnification Bar = 50 120583m
the irradiation of seeds only in palisade parenchyma withthe lowest values at 20Gy (Figures 5(c) 5(e) 5(g) and 5(i))By contrast in compound leaves the application of 100Gyresulted in increased cell size compared with control andleaves from seeds irradiated up to 50Gy (Figures 5(d) 5(f)5(h) and 5(j))
34 Effect of Ionizing Radiation on Fluorescence Measure-ments and Pigment Content No significant differences were
detectable in fluorescence parameters and total photosyn-thetic pigment content in plants grown from irradiated seedsat different X-rays doses (Figure 6) A tendency to NPQincrease was found at 03 and 10Gy followed by a decreaseat doses higher than 20Gy However the lack of significantdifferences in ΦPSII and NPQ among treatments indicatedthat X-rays delivered at the seed stage on tomato plantsdid not alter the capability of photosynthetic apparatus oftomato plants either to convert light energy in photochem-istry or to perform thermal dissipation mechanisms The
The Scientific World Journal 7
(a) (b)
Phen
olic
com
poun
ds (
)
aa a
aab
b
0
10
20
30
40
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf JL
(c)
Phen
olic
com
poun
ds (
) aab
abc ab
c bc
0
10
20
30
40
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf CL
(d)
Figure 4 Epifluorescence microscopy views of lamina cross sections of S lycopersicum L ldquoMicrotomrdquo compound leaves from plantsdeveloped from control (a) and 100Gy irradiated seeds (b) Phenolic compounds along chloroplast membranes are autofluorescent Percentof mesophyll occupied by phenolic compounds () is shown in (c) and (d) for juvenile (JL) and compound (CL) leaves respectively Meanvalues and standard errors are reported (119899 = 15) different letters correspond to significantly different values after multiple comparison tests(119875 lt 005)
maximum photochemical efficiency of PSII (FvFm) washigher in leaves from irradiated seeds than control anddid not show variations among different X-rays treatmentsthis indicated the maintenance of higher level of photo-synthetic activity after irradiation irrespectively from dose(Figure 6(c))
The total chlorophyll as well as the total carotenoidcontent did not show significant changes due to irradiationof seeds (Figure 7)
4 Discussion
Ionising radiation can have positive null or negative effectson plant development depending on the properties of theradiation the features of the plant material and possibleinteractions with other environmental factors [2] There iscommon agreement that plants are characterised by highradioresistance moreover low-LET ionising radiation hasbeen reported to increase germination percentage rootlength plant height and other growth parameters especiallyat low doses which induce hormesis [16 17 45ndash47] Ourresults suggest that S lycopersicum ldquoMicrotomrdquo is charac-terised by high radioresistance at the seed stage indeed
irradiation of dry seeds with a wide range of X-ray dosesdid not hamper germination and plant growth up to thecompletion of the crop cycle An hormesis effect was evidentsince growth either remained unaffected or was positivelyaffected especially by low doses of X-rays Apart from plantsgrown from seeds irradiated with 03 and 10Gy all plantsfrom irradiated seeds were characterised by a more compacthabitus higher number of larger leaves was not accompaniedby increased height The radiation-induced more compactgrowth pattern is positive in BLSSs in Spacewhere availabilityof volume for growth is a major technical constraint [7 4849] In the cultivar Microtom irradiation with doses equalto or higher than 20Gy caused a decrease in plant heightin agreement with the induction of dwarf or semidwarfgrowth reported as a general effect of exposure to ionisingradiation [18] Hence irradiation on the ldquoMicrotomrdquo seedswould intensify the trait of dwarf growth which alreadycharacterise this cultivar and should be taken into account forcultivar selection for the BLSSs [50] Any variations in growthparameters in Space-oriented ecologically closed systemscan determine valuable changes in terms of optimisationof volume and resource recycling These changes can bestraightaway converted into economic saving or waste [9]For example the increase in the number and size of leaves
8 The Scientific World Journal
Palisade parenchyma Spongy parenchyma
ab b ab a ab ab
a a a a a a
0010203040506
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf JL
Lam
ina t
hick
ness
(mm
)
(a)
Palisade parenchyma Spongy parenchyma
b b b b b a
a ab ab b abab
Leaf CL
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
010203040506
Lam
ina t
hick
ness
(mm
)
(b)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a a aa a
a
0200400600800
1000Upper epidermis
Cel
l are
a (120583
m2)
(c)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
1000
a a a a aa
Upper epidermis
Cel
l are
a (120583
m2)
(d)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
a bc bc c bc b
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(e)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
ab ab b ab ab a
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(f)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a aa
a a a
0200400600800
100012001400
Cel
l are
a (120583
m2)
(g)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
100012001400
b b b b ba
Cel
l are
a (120583
m2)
(h)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
a aa
a a a
0
200
400
600
800
Cel
l are
a (120583
m2)
(i)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
b b b b ba
0
200
400
600
800
Cel
l are
a (120583
m2)
(j)
Figure 5 Lamina thickness (a b) and cell area of upper epidermis (c d) palisade (e f) spongy (g h) parenchymas and lower epidermis (i j)of S lycopersicum L ldquoMicrotomrdquo leaves of control and irradiated plants at different doses of X-rays in both juvenile (JL) and compound (CL)leaves Mean values and standard errors are shown (119899 = 15 in (a b) 119899 = 45 in (cndashj)) Different letters correspond to significantly differentvalues according to multiple comparison tests (119875 lt 005)
The Scientific World Journal 9
04
05
06
07
08
a a a a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
ΦPS
II
(a)
00
02
04
06
08
NPQ a
a aa a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
(b)
070
075
080
085
090
a
b b b b b
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
FF
m
(c)
Figure 6 Quantum yield of PSII electron transport (ΦPSII a) nonphotochemical quenching (NPQ b) and maximal PSII photochemicalefficiency (FvFm c) in compound (CL) S lycopersicum L ldquoMicrotomrdquo leaves of control and plants from seeds irradiated at different dosesof X-rays Mean values and standard deviations are shown (119899 = 5) Different letters correspond to significantly different values according tomultiple comparison tests (119875 lt 005)
0
1
2
3
4
a
aa
a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Chl a
+ b
(mg g
minus1)
(a)
0
02
04
06
a
aa a
a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Car x
+ c
(mg g
minus1)
(b)
Figure 7 Total chlorophyll content (Chl a+b) (a) and total carotenoid content (Car x+c) (b) in compound (CL) S lycopersicum L ldquoMicrotomrdquoleaves of control and plants from seeds irradiated at different doses of X-rays Mean values and standard errors are shown (119899 = 5) Differentletters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
induced in ldquoMicrotomrdquo especially by low doses of irradiationcan be beneficial to atmosphere regeneration of pressurisedmodules provided that formed leaves do not show structuralaberrations which reduce their function
General anatomical structure of both juvenile and com-pound ldquoMicrotomrdquo leaves was only slightly perturbed inplants from seeds subjected to the highest levels of radiation
Clear-cut dose-depended tendencies of variations were notfound in lamina thickness and cell size and shape of thevarious tissues The formation of smaller palisade cells wasfound only in juvenile leaves of irradiated seeds while anincrease in cell size was found only in spongy parenchymaand lower epidermis of compound leaves from seeds irra-diated at 100Gy The diverse growth behaviour found in
10 The Scientific World Journal
Table 1 Cell shape in different tissues of juvenile leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown (119899 = 75)Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf JL Aspect ratio Sphericity Convexity
Upper epidermis
C 197 plusmn 016 a 0308 plusmn 0050 a 0926 plusmn 0021 ab03Gy 200 plusmn 013 a 0290 plusmn 0038 a 0941 plusmn 0013 a10Gy 210 plusmn 012 a 0239 plusmn 0037 a 0898 plusmn 0016 ab20Gy 217 plusmn 014 a 0258 plusmn 0045 a 0909 plusmn 0019 abc50Gy 170 plusmn 008 a 0376 plusmn 0037 a 0927 plusmn 0016 ab100Gy 197 plusmn 014 a 0286 plusmn 0042 a 0876 plusmn 0028 c
Palisade parenchyma
C 290 plusmn 010 a 0109 plusmn 0015 a 0879 plusmn 0010 a03 Gy 304 plusmn 012 a 0099 plusmn 0016 a 0805 plusmn 0016 b10Gy 309 plusmn 013 a 0093 plusmn 0019 a 0792 plusmn 0015 b20Gy 283 plusmn 011 a 0101 plusmn 0017 a 0784 plusmn 0017 b50Gy 314 plusmn 014 a 0082 plusmn 0016 a 0772 plusmn 0016 b100Gy 275 plusmn 011 a 0119 plusmn 0020 a 0805 plusmn 0017 b
Spongy parenchyma
C 190 plusmn 008 a 0307 plusmn 0033 a 0893 plusmn 0012 a03 Gy 180 plusmn 007 a 0341 plusmn 0029 a 0876 plusmn 0013 a10Gy 203 plusmn 010 a 0290 plusmn 0031 a 0901 plusmn 0012 a20Gy 194 plusmn 012 a 0344 plusmn 0040 a 0881 plusmn 0015 a50Gy 174 plusmn 009 a 0393 plusmn 0035 a 0883 plusmn 0014 a100Gy 198 plusmn 012 a 0322 plusmn 0037 a 0878 plusmn 0015 a
Lower epidermis
C 178 plusmn 009 bc 0343 plusmn 0045 ab 0919 plusmn 0019 a03 Gy 225 plusmn 017 ab 0263 plusmn 0044 b 0933 plusmn 0024 a10Gy 219 plusmn 011 abc 0218 plusmn 0034 b 0888 plusmn 0020 ab20Gy 180 plusmn 013 bc 0378 plusmn 0047 ab 0926 plusmn 0017 a50Gy 167 plusmn 001 c 0461 plusmn 0051 a 0924 plusmn 0021 a100Gy 238 plusmn 021 a 0257 plusmn 0057 b 0857 plusmn 0030 b
the various tissues of structures of different age is in agree-ment with the different light-mediated control of cell devel-opment in the various leaf tissues [51] Altered mesophyllcharacterized by partly collapsed cells with irregular cellshape was found only sporadically after the exposure of seedsto 50 and 100Gy in theMicrotomcultivar Suchmodificationswere less marked than those found in leaves of bean wherethe target organs for irradiation were the leaves themselves[30] leaves directly exposed to X-rays in bean showed theformation of more loose parenchyma with larger cells likelydue to a radiation-induced damage to cell walls that reducedmechanical constraints to cell enlargement [30 52] Furtherinvestigations on cell wall composition would be useful toelucidate whether a possible alteration of cell walls wasresponsible for cell enlargement in the spongy parenchymaand epidermis of leaves from seeds of ldquoMicrotomrdquo irradiatedwith the highest dose of X-rays However in the cultivarMicrotom the slight alterations of mesophyll structure didnot impair photosynthetic efficiency as demonstrated by thelack of significant dose-dependent variation trends in thequantum yield of PSII electron transport (ΦPSII) nonphoto-chemical quenching (NPQ) and maximal PSII photochemi-cal efficiency (FvFm) The increased values of FvFm closeto 08 which is considered an optimal value for healthy plants[53] suggested that the photosynthetic apparatus of the leavesfrom irradiated seeds has higher levels of photochemical
efficiency irrespective of dose The unchanged or increasedcontent of both chlorophyll and carotenoid pigments con-firmed that the photosynthetic apparatus remained stablealso in terms of light harvesting capacity and indicatedno deleterious effect of radiation on plant photosyntheticmachinery RecentlyMarcu et al [17] showed that irradiationof lettuce seeds with gamma rays up to 30Gy enhancedthe content of chlorophyll a chlorophyll b and carotenoidswhile higher doses (up to 70Gy) resulted in their decreaseHowever the effect of ionising radiation on photosyntheticpigment content is quite controversial Increased pigmentcontent is ascribed to the phenomenon of hormesis whilea lower content is generally ascribed to the inhibition ofchlorophyll and carotenoid synthesis or to oxidation due tothe high levels of radiation-induced water radiolysis [2 1454ndash56] Some plants possess natural screens against radiationwhen their tissues contain phenolic compounds with antioxi-dant functions and can counteract photoinhibitory processes[29 57] An increase of phenolic compounds linked to thechloroplast membranes has been recently found in Phaseolusvulgaris irradiated leaves exposed to high levels of X-rays[28 30] Changes in phenolic content after the exposure toSpace can be attributed either to direct effects on the synthesisor mobilization of phenolic compounds or to indirect effectsdue to changes in ultrastructural organization such as theincrease in the number of chloroplasts per cell [30 58]
The Scientific World Journal 11
Table 2 Cell shape in different tissues of compound leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown(119899 = 75) Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf CL Aspect ratio Sphericity Convexity
Upper epidermis
C 166 plusmn 007 c 0422 plusmn 0042 a 0946 plusmn 0010 a03 Gy 216 plusmn 017 ab 0244 plusmn 0033 c 0925 plusmn 0016 ab10Gy 171 plusmn 011 bc 0401 plusmn 0042 a 0914 plusmn 0013 b20Gy 186 plusmn 010 abc 0328 plusmn 0041 ab 0919 plusmn 0020 ab50Gy 225 plusmn 018 a 0247 plusmn 0041 c 0897 plusmn 0019 b100Gy 188 plusmn 009 abc 0319 plusmn 0045 ab 0902 plusmn 0016 b
Palisade parenchyma
C 355 plusmn 012 a 0073 plusmn 0014 b 0890 plusmn 0012 a03 Gy 289 plusmn 010 b 0102 plusmn 0013 ab 0847 plusmn 0018 b10Gy 300 plusmn 014 b 0104 plusmn 0021 ab 0826 plusmn 0017 b20Gy 302 plusmn 011 b 0094 plusmn 0013 ab 0810 plusmn 0014 b50Gy 283 plusmn 012 b 0107 plusmn 0021 ab 0750 plusmn 0021 c100Gy 275 plusmn 012 b 0122 plusmn 0023 a 0809 plusmn 0015 b
Spongy parenchyma
C 193 plusmn 010 a 0323 plusmn 0035 a 0904 plusmn 0011 a03 Gy 183 plusmn 010 a 0375 plusmn 0042 a 0879 plusmn 0013 a10Gy 176 plusmn 007 a 0357 plusmn 0034 a 0869 plusmn 0011 a20Gy 180 plusmn 009 a 0366 plusmn 0038 a 0885 plusmn 0016 a50Gy 171 plusmn 006 a 0372 plusmn 0034 a 0879 plusmn 0015 a100Gy 188 plusmn 010 a 0339 plusmn 0037 a 0883 plusmn 0015 a
Lower epidermis
C 148 plusmn 008 b 0545 plusmn 0045 a 0957 plusmn 0015 a03 Gy 210 plusmn 017 a 0295 plusmn 0046 b 0929 plusmn 0019 ab10Gy 190 plusmn 012 ab 0334 plusmn 0043 b 0893 plusmn 0019 b20Gy 178 plusmn 011 ab 0388 plusmn 0047 b 0929 plusmn 0017 ab50Gy 172 plusmn 009 ab 0367 plusmn 0040 b 0914 plusmn 0016 b100Gy 191 plusmn 014 ab 0362 plusmn 0054 b 0888 plusmn 0025 b
The lowering of phenolic content at high levels of X-rays inldquoMicrotomrdquo leaves is not in agreement with general trendsindicating enhanced phenolics production in plants grownin Space [58] or in various plant tissues directly irradiatedwith gamma rays [59] However the low content of phenolicscan be an indirect effect of increasing volume occupiedby intercellular spaces due to the loosening of parenchymacells
In conclusion our results show that irradiation of the dryseeds of the Microtom cultivar did not hamper germinationand the development of functional leaves The formationof more compact plants characterised by a higher numberof larger leaves can be valuable in terms of increasingresource regeneration in the BLSSs High levels of radiationinduced only slight structural perturbations which weresimilar in juvenile and compound leaves Such perturbationsdid not affect photosynthetic efficiency since the maximalPSII photochemical efficiency was even increased after irra-diation at all doses However the tendency to accumulatelower amounts of phenolic compounds along chloroplastmembranes after irradiation of seeds with high doses ofradiation should be further investigated because it coulddetermine the reduction of natural photoprotection [57]Finally the overall results indicate that theMicrotom cultivarirradiated at the seed stage has high radioresistance duringthe growth which is a desirable feature for plant cultivation
in Space Radioresistance together with dwarf growth andshort life cycle makes this cultivar a valuable candidate forthe cultivation in BLSSs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are grateful to the ldquoCompagnia di San Paolordquo(Turin) and the ldquoPolo delle Scienze e delle Tecnologie perla Vitardquo of the University of Naples Federico II for thefinancial support The authors also thank Maria Giordanoand Pasquale Scognamiglio for their support in plant culti-vation and microscopy analyses The authors acknowledgefour anonymous editors for helpful comments on a previousversion of the paper
References
[1] A Real S Sundell-Bergman J F Knowles D S Woodheadand I Zinger ldquoEffects of ionising radiation exposure on plantsfish and mammals relevant data for environmental radiation
12 The Scientific World Journal
protectionrdquo Journal of Radiological Protection vol 24 no 4 ppA123ndashA137 2004
[2] V De Micco C Arena D Pignalosa and M Durante ldquoEffectsof sparsely and densely ionizing radiation on plantsrdquo Radiationand Environmental Biophysics vol 50 no 1 pp 1ndash19 2011
[3] N Kumari P Kumar D Mitra B Prasad B N Tiwary andL Varshney ldquoEffects of ionizing radiation on microbial decon-tamination phenolic contents and antioxidant properties oftriphalardquo Journal of Food Science vol 74 no 3 pp M109ndashM1132009
[4] A Tanaka N Shikazono and Y Hase ldquoStudies on biologicaleffects of ion beams on lethality molecular nature of mutationmutation rate and spectrum of mutation phenotype for muta-tion breeding in higher plantsrdquo Journal of Radiation Researchvol 51 no 3 pp 223ndash233 2010
[5] M Durante and A Kronenberg ldquoGround-based research withheavy ions for space radiation protectionrdquo Advances in SpaceResearch vol 35 no 2 pp 180ndash184 2005
[6] E Hu S I Bartsev and H Liu ldquoConceptual design of a biore-generative life support system containing crops and silkwormsrdquoAdvances in Space Research vol 45 no 7 pp 929ndash939 2010
[7] V deMicco G Aronne G Colla R Fortezza and S de PascaleldquoAgro-biology for bioregenerative life support systems in long-term space missions general constraints and the Italian effortsrdquoJournal of Plant Interactions vol 4 no 4 pp 241ndash252 2009
[8] T W Halstead and F R Dutcher ldquoPlants in spacerdquo AnnualReview of Plant Physiology vol 38 pp 317ndash345 1987
[9] V De Micco S De Pascale R Paradiso and G AronneldquoMicrogravity effects on different stages of higher plant life cycleand completion of the seed-to-seed cyclerdquo Plant Biology vol 16pp 31ndash38 2014
[10] C J Bennett C Pirim and T M Orlando ldquoSpace-weatheringof solar system bodies a laboratory perspectiverdquo ChemicalReviews vol 113 pp 9086ndash9150 2013
[11] R A Kerr ldquoRadiation will make astronautsrsquo trip to Mars evenriskierrdquo Science vol 340 article 1031 2014
[12] D M Hassler C Zeitlin R F Wimmer-Schweingruber et alldquoMarsrsquo surface radiation environment measured with the MarsScience Laboratoryrsquos Curiosity Roverrdquo Science vol 343 no6169 Article ID 1244797 2014
[13] M Durante and F A Cucinotta ldquoHeavy ion carcinogenesis andhuman space explorationrdquo Nature Reviews Cancer vol 8 no 6pp 465ndash472 2008
[14] C Arena V De Micco and A De Maio ldquoGrowth alterationand leaf biochemical responses in P vulgaris plants exposed todifferent doses of ionizing radiationrdquo Plant Biology vol 16 pp194ndash202 2014
[15] R W Holst and D J Nagel ldquoRadiation effects on plantsrdquo inPlants For Environmental Studies W Wang J W Gorsuch andJ S Hughes Eds pp 37ndash81 Lewis Publishers Boca Raton FlaUSA 1997
[16] T D Luckey Hormesis With Ionizing Radiation CRC PressBoca Raton Fla USA 1980
[17] D Marcu V Cristea and L Daraban ldquoDose-dependent effectsof gamma radiation on lettuce (Lactuca sativa var capitata)seedlingsrdquo International Journal of Radiation Biology vol 89 pp219ndash223 2013
[18] M Mei H Deng Y Lu et al ldquoMutagenic effects of heavy ionradiation in plantsrdquo Advances in Space Research vol 14 no 10pp 363ndash372 1994
[19] Y LiM Liu Z Cheng andY Sun ldquoSpace environment inducedmutations prefer to occur at polymorphic sites of rice genomesrdquoAdvances in Space Research vol 40 no 4 pp 523ndash527 2007
[20] D S Kim J-B Kim E J Goh et al ldquoAntioxidant response ofArabidopsis plants to gamma irradiation genome-wide expres-sion profiling of the ROS scavenging and signal transductionpathwaysrdquo Journal of Plant Physiology vol 168 no 16 pp 1960ndash1971 2011
[21] T J Brodribb ldquoXylem hydraulic physiology the functionalbackbone of terrestrial plant productivityrdquo Plant Science vol177 no 4 pp 245ndash251 2009
[22] R M Wheeler J C Sager R P Prince andW M Knott ldquoCropproduction for advanced life support systemsmdashobservationfrom the Kennedy Space Center Breadboard Projectrdquo NASATM-2003-211184 NASA Kennedy Space Center Florida 2003
[23] R M Wheeler C L Mackowiak G W Stutte et al ldquoCropproductivities and radiation use efficiencies for bioregenerativelife supportrdquo Advances in Space Research vol 41 no 5 pp 706ndash713 2008
[24] R Paradiso V De Micco R Buonomo G Aronne G Bar-bieri and S De Pascale ldquoSoilless cultivation of soybean forBioregenerative Life-Support Systems a literature review andthe experience of the MELiSSA ProjectmdashFood characterisationPhase Irdquo Plant Biology vol 16 pp 69ndash78 2014
[25] D M Porterfield ldquoThe biophysical limitations in physiologicaltransport and exchange in plants grown in microgravityrdquoJournal of Plant Growth Regulation vol 21 no 2 pp 177ndash1902002
[26] K Yokthongwattana and A Melis ldquoPhotoinhibition and recov-ery in oxygenic photosynthesis mechanism of a photosystem IIdamage and repair cyclerdquo in Photoprotection PhotoinhibitionGene Regulation and Environment B Demmig-Adams WW Adams III and A K Mattoo Eds pp 175ndash191 SpringerDordecht The Netherlands 2006
[27] M T Giardi G Rea M D Lambreva et al ldquoMutations ofphotosystem II D1 protein that empower efficient phenotypesof Chlamydomonas reinhardtii under extreme environment inSpacerdquo PLoS ONE vol 8 Article ID e64352 2013
[28] C Arena V De Micco G Aronne M Pugliese A Virzo DeSanto andADeMaio ldquoResponse ofPhaseolus vulgarisL plantsto low-let ionizing radiation growth and oxidative stressrdquo ActaAstronautica vol 91 pp 107ndash114 2013
[29] V Lattanzio P A Kroon and S Quideau ldquoPlant phenolicsmdashsecondary metabolites with diverse functionsrdquo in RecentAdvances in Polyphenol Research F Daay and V Lattanzio Edspp 1ndash35 Wiley-Blackwell Oxford UK 2008
[30] V De Micco C Arena and G Aronne ldquoAnatomical alterationsof Phaseolus vulgaris L mature leaves irradiated with X-raysrdquoPlant Biology vol 16 pp 187ndash193 2014
[31] A D Krikorian ldquoPlants and somatic embryos in space whathave we learnedrdquo Gravitational and Space Biology Bulletin vol11 no 2 pp 5ndash14 1998
[32] R Meissner Y Jacobson S Melamed et al ldquoA new modelsystem for tomato geneticsrdquoThe Plant Journal vol 12 no 6 pp1465ndash1472 1997
[33] E Emmanuel and A A Levy ldquoTomato mutants as tools forfunctional genomicsrdquo Current Opinion in Plant Biology vol 5no 2 pp 112ndash117 2002
[34] E Martı C Gisbert G J Bishop M S Dixon and J LGarcıa-Martınez ldquoGenetic and physiological characterizationof tomato cv Micro-Tomrdquo Journal of Experimental Botany vol57 pp 2037ndash2047 2006
The Scientific World Journal 13
[35] T Kurimoto J V H Constable and A Huda ldquoEffects ofionizing radiation exposure on arabidopsis thalianardquo HealthPhysics vol 99 no 1 pp 49ndash57 2010
[36] L Reale D Gigante F Landucci F Ferranti and R VenanzonildquoMorphological and histo-anatomical traits reflect die-back inPhragmites australis (Cav) Steudrdquo Aquatic Botany vol 103 pp122ndash128 2012
[37] K Fukuzawa ldquoUltraviolet microscopyrdquo in Methods in LigninChemistry S Y Lin andCWDence Eds pp 110ndash131 SpringerBerlin Germany 1992
[38] S E Ruzin Plant Microtechnique and Microscopy OxfordUniversity Press New York NY USA 1999
[39] V De Micco G Aronne J-P Joseleau and K Ruel ldquoXylemdevelopment and cell wall changes of soybean seedlings grownin spacerdquo Annals of Botany vol 101 no 5 pp 661ndash669 2008
[40] S Van Buggenhout T Grauwet A Van Loey and M Hen-drickx ldquoStructureprocessing relation of vacuum infused straw-berry tissue frozen under different conditionsrdquo European FoodResearch and Technology vol 226 no 3 pp 437ndash448 2008
[41] V De Micco and G Aronne ldquoBiometric anatomy of seedlingsdeveloped onboard of Foton-M2 in an automatic system sup-porting growthrdquoActa Astronautica vol 62 no 8-9 pp 505ndash5132008
[42] B Genty J M Briantais and N R Baker ldquoThe relationshipbetween quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescencerdquo Biochimica andBiophysica Acta vol 990 pp 87ndash92 1989
[43] W Bilger and O Bjorkman ldquoRole of the xanthophyll cycle inphotoprotection elucidated by measurements of light-inducedabsorbance changes fluorescence and photosynthesis in leavesof Hedera canariensisrdquo Photosynthesis Research vol 25 no 3pp 173ndash185 1990
[44] H K Lichtenthaler ldquoChlorophylls and carotenoids pigmentsof photosynthetic biomembranesrdquoMethods in Enzymology vol148 pp 350ndash382 1987
[45] J P Maity DMishra A Chakraborty A Saha S C Santra andS Chanda ldquoModulation of some quantitative and qualitativecharacteristics in rice (Oryza sativa L) and mung (Phaseolusmungo L) by ionizing radiationrdquo Radiation Physics and Chem-istry vol 74 no 5 pp 391ndash394 2005
[46] L Cheng H Yang B Lin et al ldquoEffect of gamma-ray radiationon physiological morphological characters and chromosomeaberrations of minitubers in Solanum tuberosum Lrdquo Interna-tional Journal of Radiation Biology vol 86 no 9 pp 791ndash7992010
[47] B Al-Safadi and R Elias ldquoImprovement of caper (Capparisspinosa L) propagation using in vitro culture and gammairradiationrdquo Scientia Horticulturae vol 127 no 3 pp 290ndash2972011
[48] O Monje G W Stutte G D Goins D M Porterfield and GE Bingham ldquoFarming in space environmental and biophysicalconcernsrdquo Advances in Space Research vol 31 no 1 pp 151ndash1672003
[49] V De Micco M Scala and G Aronne ldquoEffects of simulatedmicrogravity on male gametophyte of Prunus Pyrus andBrassica speciesrdquo Protoplasma vol 228 no 1ndash3 pp 121ndash1262006
[50] V De Micco R Buonomo R Paradiso S De Pascale and GAronne ldquoSoybean cultivar selection for Bioregenerative LifeSupport Systems (BLSS)mdashtheoretical selectionrdquo Advances inSpace Research vol 49 no 10 pp 1415ndash1421 2012
[51] T Kozuka S-G Kong M Doi K-I Shimazaki and ANagatani ldquoTissue-autonomous promotion of palisade celldevelopment by phototropin 2 in Arabidopsisrdquo Plant Cell vol23 no 10 pp 3684ndash3695 2011
[52] E Kovacs and A Keresztes ldquoEffect of gamma and UV-BCradiation on plant cellsrdquoMicron vol 33 no 2 pp 199ndash210 2002
[53] R Ogaya J Penuelas D Asensio and J Llusia ldquoChlorophyllfluorescence responses to temperature and water availabilityin two co-dominant Mediterranean shrub and tree speciesin a long-term field experiment simulating climate changerdquoEnvironmental and Experimental Botany vol 73 no 1 pp 89ndash93 2011
[54] R Zaka C M Vandecasteele and M T Misset ldquoEffects oflow chronic doses of ionizing radiation on antioxidant enzymesand G6PDH activities in Stipa capillata (Poaceae)rdquo Journal ofExperimental Botany vol 53 no 376 pp 1979ndash1987 2002
[55] J-H KimM-H Baek Y C Byung GW Seung and J-S KimldquoAlterations in the photosynthetic pigments and antioxidantmachineries of red pepper (Capsicum annuum L) seedlingsfrom gamma-irradiated seedsrdquo Journal of Plant Biology vol 47no 4 pp 314ndash321 2004
[56] N A Al-Enezi and J M Al-Khayri ldquoAlterations of DNA ionsand photosynthetic pigments content in date palm seedlingsinduced by X-irradiationrdquo International Journal of Agricultureand Biology vol 14 pp 329ndash336 2012
[57] V de Micco C Arena L Vitale G Aronne and A V de SantoldquoAnatomy and photochemical behaviour of mediterranean Cis-tus incanus winter leaves under natural outdoor and warmerindoor conditionsrdquo Botany vol 89 no 10 pp 677ndash688 2011
[58] L H Levine H G Levine E C Stryjewski V Prima and WC Piastuch ldquoEffect of spaceflight on isoflavonoid accumulationin etiolated soybean seedlingsrdquo Journal of Gravitational Physiol-ogy vol 8 no 2 pp 21ndash27 2001
[59] X Fan ldquoAntioxidant capacity of fresh-cut vegetables exposedto ionizing radiationrdquo Journal of the Science of Food andAgriculture vol 85 no 6 pp 995ndash1000 2005
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

The Scientific World Journal 7
(a) (b)
Phen
olic
com
poun
ds (
)
aa a
aab
b
0
10
20
30
40
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf JL
(c)
Phen
olic
com
poun
ds (
) aab
abc ab
c bc
0
10
20
30
40
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf CL
(d)
Figure 4 Epifluorescence microscopy views of lamina cross sections of S lycopersicum L ldquoMicrotomrdquo compound leaves from plantsdeveloped from control (a) and 100Gy irradiated seeds (b) Phenolic compounds along chloroplast membranes are autofluorescent Percentof mesophyll occupied by phenolic compounds () is shown in (c) and (d) for juvenile (JL) and compound (CL) leaves respectively Meanvalues and standard errors are reported (119899 = 15) different letters correspond to significantly different values after multiple comparison tests(119875 lt 005)
maximum photochemical efficiency of PSII (FvFm) washigher in leaves from irradiated seeds than control anddid not show variations among different X-rays treatmentsthis indicated the maintenance of higher level of photo-synthetic activity after irradiation irrespectively from dose(Figure 6(c))
The total chlorophyll as well as the total carotenoidcontent did not show significant changes due to irradiationof seeds (Figure 7)
4 Discussion
Ionising radiation can have positive null or negative effectson plant development depending on the properties of theradiation the features of the plant material and possibleinteractions with other environmental factors [2] There iscommon agreement that plants are characterised by highradioresistance moreover low-LET ionising radiation hasbeen reported to increase germination percentage rootlength plant height and other growth parameters especiallyat low doses which induce hormesis [16 17 45ndash47] Ourresults suggest that S lycopersicum ldquoMicrotomrdquo is charac-terised by high radioresistance at the seed stage indeed
irradiation of dry seeds with a wide range of X-ray dosesdid not hamper germination and plant growth up to thecompletion of the crop cycle An hormesis effect was evidentsince growth either remained unaffected or was positivelyaffected especially by low doses of X-rays Apart from plantsgrown from seeds irradiated with 03 and 10Gy all plantsfrom irradiated seeds were characterised by a more compacthabitus higher number of larger leaves was not accompaniedby increased height The radiation-induced more compactgrowth pattern is positive in BLSSs in Spacewhere availabilityof volume for growth is a major technical constraint [7 4849] In the cultivar Microtom irradiation with doses equalto or higher than 20Gy caused a decrease in plant heightin agreement with the induction of dwarf or semidwarfgrowth reported as a general effect of exposure to ionisingradiation [18] Hence irradiation on the ldquoMicrotomrdquo seedswould intensify the trait of dwarf growth which alreadycharacterise this cultivar and should be taken into account forcultivar selection for the BLSSs [50] Any variations in growthparameters in Space-oriented ecologically closed systemscan determine valuable changes in terms of optimisationof volume and resource recycling These changes can bestraightaway converted into economic saving or waste [9]For example the increase in the number and size of leaves
8 The Scientific World Journal
Palisade parenchyma Spongy parenchyma
ab b ab a ab ab
a a a a a a
0010203040506
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf JL
Lam
ina t
hick
ness
(mm
)
(a)
Palisade parenchyma Spongy parenchyma
b b b b b a
a ab ab b abab
Leaf CL
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
010203040506
Lam
ina t
hick
ness
(mm
)
(b)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a a aa a
a
0200400600800
1000Upper epidermis
Cel
l are
a (120583
m2)
(c)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
1000
a a a a aa
Upper epidermis
Cel
l are
a (120583
m2)
(d)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
a bc bc c bc b
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(e)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
ab ab b ab ab a
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(f)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a aa
a a a
0200400600800
100012001400
Cel
l are
a (120583
m2)
(g)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
100012001400
b b b b ba
Cel
l are
a (120583
m2)
(h)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
a aa
a a a
0
200
400
600
800
Cel
l are
a (120583
m2)
(i)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
b b b b ba
0
200
400
600
800
Cel
l are
a (120583
m2)
(j)
Figure 5 Lamina thickness (a b) and cell area of upper epidermis (c d) palisade (e f) spongy (g h) parenchymas and lower epidermis (i j)of S lycopersicum L ldquoMicrotomrdquo leaves of control and irradiated plants at different doses of X-rays in both juvenile (JL) and compound (CL)leaves Mean values and standard errors are shown (119899 = 15 in (a b) 119899 = 45 in (cndashj)) Different letters correspond to significantly differentvalues according to multiple comparison tests (119875 lt 005)
The Scientific World Journal 9
04
05
06
07
08
a a a a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
ΦPS
II
(a)
00
02
04
06
08
NPQ a
a aa a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
(b)
070
075
080
085
090
a
b b b b b
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
FF
m
(c)
Figure 6 Quantum yield of PSII electron transport (ΦPSII a) nonphotochemical quenching (NPQ b) and maximal PSII photochemicalefficiency (FvFm c) in compound (CL) S lycopersicum L ldquoMicrotomrdquo leaves of control and plants from seeds irradiated at different dosesof X-rays Mean values and standard deviations are shown (119899 = 5) Different letters correspond to significantly different values according tomultiple comparison tests (119875 lt 005)
0
1
2
3
4
a
aa
a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Chl a
+ b
(mg g
minus1)
(a)
0
02
04
06
a
aa a
a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Car x
+ c
(mg g
minus1)
(b)
Figure 7 Total chlorophyll content (Chl a+b) (a) and total carotenoid content (Car x+c) (b) in compound (CL) S lycopersicum L ldquoMicrotomrdquoleaves of control and plants from seeds irradiated at different doses of X-rays Mean values and standard errors are shown (119899 = 5) Differentletters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
induced in ldquoMicrotomrdquo especially by low doses of irradiationcan be beneficial to atmosphere regeneration of pressurisedmodules provided that formed leaves do not show structuralaberrations which reduce their function
General anatomical structure of both juvenile and com-pound ldquoMicrotomrdquo leaves was only slightly perturbed inplants from seeds subjected to the highest levels of radiation
Clear-cut dose-depended tendencies of variations were notfound in lamina thickness and cell size and shape of thevarious tissues The formation of smaller palisade cells wasfound only in juvenile leaves of irradiated seeds while anincrease in cell size was found only in spongy parenchymaand lower epidermis of compound leaves from seeds irra-diated at 100Gy The diverse growth behaviour found in
10 The Scientific World Journal
Table 1 Cell shape in different tissues of juvenile leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown (119899 = 75)Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf JL Aspect ratio Sphericity Convexity
Upper epidermis
C 197 plusmn 016 a 0308 plusmn 0050 a 0926 plusmn 0021 ab03Gy 200 plusmn 013 a 0290 plusmn 0038 a 0941 plusmn 0013 a10Gy 210 plusmn 012 a 0239 plusmn 0037 a 0898 plusmn 0016 ab20Gy 217 plusmn 014 a 0258 plusmn 0045 a 0909 plusmn 0019 abc50Gy 170 plusmn 008 a 0376 plusmn 0037 a 0927 plusmn 0016 ab100Gy 197 plusmn 014 a 0286 plusmn 0042 a 0876 plusmn 0028 c
Palisade parenchyma
C 290 plusmn 010 a 0109 plusmn 0015 a 0879 plusmn 0010 a03 Gy 304 plusmn 012 a 0099 plusmn 0016 a 0805 plusmn 0016 b10Gy 309 plusmn 013 a 0093 plusmn 0019 a 0792 plusmn 0015 b20Gy 283 plusmn 011 a 0101 plusmn 0017 a 0784 plusmn 0017 b50Gy 314 plusmn 014 a 0082 plusmn 0016 a 0772 plusmn 0016 b100Gy 275 plusmn 011 a 0119 plusmn 0020 a 0805 plusmn 0017 b
Spongy parenchyma
C 190 plusmn 008 a 0307 plusmn 0033 a 0893 plusmn 0012 a03 Gy 180 plusmn 007 a 0341 plusmn 0029 a 0876 plusmn 0013 a10Gy 203 plusmn 010 a 0290 plusmn 0031 a 0901 plusmn 0012 a20Gy 194 plusmn 012 a 0344 plusmn 0040 a 0881 plusmn 0015 a50Gy 174 plusmn 009 a 0393 plusmn 0035 a 0883 plusmn 0014 a100Gy 198 plusmn 012 a 0322 plusmn 0037 a 0878 plusmn 0015 a
Lower epidermis
C 178 plusmn 009 bc 0343 plusmn 0045 ab 0919 plusmn 0019 a03 Gy 225 plusmn 017 ab 0263 plusmn 0044 b 0933 plusmn 0024 a10Gy 219 plusmn 011 abc 0218 plusmn 0034 b 0888 plusmn 0020 ab20Gy 180 plusmn 013 bc 0378 plusmn 0047 ab 0926 plusmn 0017 a50Gy 167 plusmn 001 c 0461 plusmn 0051 a 0924 plusmn 0021 a100Gy 238 plusmn 021 a 0257 plusmn 0057 b 0857 plusmn 0030 b
the various tissues of structures of different age is in agree-ment with the different light-mediated control of cell devel-opment in the various leaf tissues [51] Altered mesophyllcharacterized by partly collapsed cells with irregular cellshape was found only sporadically after the exposure of seedsto 50 and 100Gy in theMicrotomcultivar Suchmodificationswere less marked than those found in leaves of bean wherethe target organs for irradiation were the leaves themselves[30] leaves directly exposed to X-rays in bean showed theformation of more loose parenchyma with larger cells likelydue to a radiation-induced damage to cell walls that reducedmechanical constraints to cell enlargement [30 52] Furtherinvestigations on cell wall composition would be useful toelucidate whether a possible alteration of cell walls wasresponsible for cell enlargement in the spongy parenchymaand epidermis of leaves from seeds of ldquoMicrotomrdquo irradiatedwith the highest dose of X-rays However in the cultivarMicrotom the slight alterations of mesophyll structure didnot impair photosynthetic efficiency as demonstrated by thelack of significant dose-dependent variation trends in thequantum yield of PSII electron transport (ΦPSII) nonphoto-chemical quenching (NPQ) and maximal PSII photochemi-cal efficiency (FvFm) The increased values of FvFm closeto 08 which is considered an optimal value for healthy plants[53] suggested that the photosynthetic apparatus of the leavesfrom irradiated seeds has higher levels of photochemical
efficiency irrespective of dose The unchanged or increasedcontent of both chlorophyll and carotenoid pigments con-firmed that the photosynthetic apparatus remained stablealso in terms of light harvesting capacity and indicatedno deleterious effect of radiation on plant photosyntheticmachinery RecentlyMarcu et al [17] showed that irradiationof lettuce seeds with gamma rays up to 30Gy enhancedthe content of chlorophyll a chlorophyll b and carotenoidswhile higher doses (up to 70Gy) resulted in their decreaseHowever the effect of ionising radiation on photosyntheticpigment content is quite controversial Increased pigmentcontent is ascribed to the phenomenon of hormesis whilea lower content is generally ascribed to the inhibition ofchlorophyll and carotenoid synthesis or to oxidation due tothe high levels of radiation-induced water radiolysis [2 1454ndash56] Some plants possess natural screens against radiationwhen their tissues contain phenolic compounds with antioxi-dant functions and can counteract photoinhibitory processes[29 57] An increase of phenolic compounds linked to thechloroplast membranes has been recently found in Phaseolusvulgaris irradiated leaves exposed to high levels of X-rays[28 30] Changes in phenolic content after the exposure toSpace can be attributed either to direct effects on the synthesisor mobilization of phenolic compounds or to indirect effectsdue to changes in ultrastructural organization such as theincrease in the number of chloroplasts per cell [30 58]
The Scientific World Journal 11
Table 2 Cell shape in different tissues of compound leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown(119899 = 75) Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf CL Aspect ratio Sphericity Convexity
Upper epidermis
C 166 plusmn 007 c 0422 plusmn 0042 a 0946 plusmn 0010 a03 Gy 216 plusmn 017 ab 0244 plusmn 0033 c 0925 plusmn 0016 ab10Gy 171 plusmn 011 bc 0401 plusmn 0042 a 0914 plusmn 0013 b20Gy 186 plusmn 010 abc 0328 plusmn 0041 ab 0919 plusmn 0020 ab50Gy 225 plusmn 018 a 0247 plusmn 0041 c 0897 plusmn 0019 b100Gy 188 plusmn 009 abc 0319 plusmn 0045 ab 0902 plusmn 0016 b
Palisade parenchyma
C 355 plusmn 012 a 0073 plusmn 0014 b 0890 plusmn 0012 a03 Gy 289 plusmn 010 b 0102 plusmn 0013 ab 0847 plusmn 0018 b10Gy 300 plusmn 014 b 0104 plusmn 0021 ab 0826 plusmn 0017 b20Gy 302 plusmn 011 b 0094 plusmn 0013 ab 0810 plusmn 0014 b50Gy 283 plusmn 012 b 0107 plusmn 0021 ab 0750 plusmn 0021 c100Gy 275 plusmn 012 b 0122 plusmn 0023 a 0809 plusmn 0015 b
Spongy parenchyma
C 193 plusmn 010 a 0323 plusmn 0035 a 0904 plusmn 0011 a03 Gy 183 plusmn 010 a 0375 plusmn 0042 a 0879 plusmn 0013 a10Gy 176 plusmn 007 a 0357 plusmn 0034 a 0869 plusmn 0011 a20Gy 180 plusmn 009 a 0366 plusmn 0038 a 0885 plusmn 0016 a50Gy 171 plusmn 006 a 0372 plusmn 0034 a 0879 plusmn 0015 a100Gy 188 plusmn 010 a 0339 plusmn 0037 a 0883 plusmn 0015 a
Lower epidermis
C 148 plusmn 008 b 0545 plusmn 0045 a 0957 plusmn 0015 a03 Gy 210 plusmn 017 a 0295 plusmn 0046 b 0929 plusmn 0019 ab10Gy 190 plusmn 012 ab 0334 plusmn 0043 b 0893 plusmn 0019 b20Gy 178 plusmn 011 ab 0388 plusmn 0047 b 0929 plusmn 0017 ab50Gy 172 plusmn 009 ab 0367 plusmn 0040 b 0914 plusmn 0016 b100Gy 191 plusmn 014 ab 0362 plusmn 0054 b 0888 plusmn 0025 b
The lowering of phenolic content at high levels of X-rays inldquoMicrotomrdquo leaves is not in agreement with general trendsindicating enhanced phenolics production in plants grownin Space [58] or in various plant tissues directly irradiatedwith gamma rays [59] However the low content of phenolicscan be an indirect effect of increasing volume occupiedby intercellular spaces due to the loosening of parenchymacells
In conclusion our results show that irradiation of the dryseeds of the Microtom cultivar did not hamper germinationand the development of functional leaves The formationof more compact plants characterised by a higher numberof larger leaves can be valuable in terms of increasingresource regeneration in the BLSSs High levels of radiationinduced only slight structural perturbations which weresimilar in juvenile and compound leaves Such perturbationsdid not affect photosynthetic efficiency since the maximalPSII photochemical efficiency was even increased after irra-diation at all doses However the tendency to accumulatelower amounts of phenolic compounds along chloroplastmembranes after irradiation of seeds with high doses ofradiation should be further investigated because it coulddetermine the reduction of natural photoprotection [57]Finally the overall results indicate that theMicrotom cultivarirradiated at the seed stage has high radioresistance duringthe growth which is a desirable feature for plant cultivation
in Space Radioresistance together with dwarf growth andshort life cycle makes this cultivar a valuable candidate forthe cultivation in BLSSs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are grateful to the ldquoCompagnia di San Paolordquo(Turin) and the ldquoPolo delle Scienze e delle Tecnologie perla Vitardquo of the University of Naples Federico II for thefinancial support The authors also thank Maria Giordanoand Pasquale Scognamiglio for their support in plant culti-vation and microscopy analyses The authors acknowledgefour anonymous editors for helpful comments on a previousversion of the paper
References
[1] A Real S Sundell-Bergman J F Knowles D S Woodheadand I Zinger ldquoEffects of ionising radiation exposure on plantsfish and mammals relevant data for environmental radiation
12 The Scientific World Journal
protectionrdquo Journal of Radiological Protection vol 24 no 4 ppA123ndashA137 2004
[2] V De Micco C Arena D Pignalosa and M Durante ldquoEffectsof sparsely and densely ionizing radiation on plantsrdquo Radiationand Environmental Biophysics vol 50 no 1 pp 1ndash19 2011
[3] N Kumari P Kumar D Mitra B Prasad B N Tiwary andL Varshney ldquoEffects of ionizing radiation on microbial decon-tamination phenolic contents and antioxidant properties oftriphalardquo Journal of Food Science vol 74 no 3 pp M109ndashM1132009
[4] A Tanaka N Shikazono and Y Hase ldquoStudies on biologicaleffects of ion beams on lethality molecular nature of mutationmutation rate and spectrum of mutation phenotype for muta-tion breeding in higher plantsrdquo Journal of Radiation Researchvol 51 no 3 pp 223ndash233 2010
[5] M Durante and A Kronenberg ldquoGround-based research withheavy ions for space radiation protectionrdquo Advances in SpaceResearch vol 35 no 2 pp 180ndash184 2005
[6] E Hu S I Bartsev and H Liu ldquoConceptual design of a biore-generative life support system containing crops and silkwormsrdquoAdvances in Space Research vol 45 no 7 pp 929ndash939 2010
[7] V deMicco G Aronne G Colla R Fortezza and S de PascaleldquoAgro-biology for bioregenerative life support systems in long-term space missions general constraints and the Italian effortsrdquoJournal of Plant Interactions vol 4 no 4 pp 241ndash252 2009
[8] T W Halstead and F R Dutcher ldquoPlants in spacerdquo AnnualReview of Plant Physiology vol 38 pp 317ndash345 1987
[9] V De Micco S De Pascale R Paradiso and G AronneldquoMicrogravity effects on different stages of higher plant life cycleand completion of the seed-to-seed cyclerdquo Plant Biology vol 16pp 31ndash38 2014
[10] C J Bennett C Pirim and T M Orlando ldquoSpace-weatheringof solar system bodies a laboratory perspectiverdquo ChemicalReviews vol 113 pp 9086ndash9150 2013
[11] R A Kerr ldquoRadiation will make astronautsrsquo trip to Mars evenriskierrdquo Science vol 340 article 1031 2014
[12] D M Hassler C Zeitlin R F Wimmer-Schweingruber et alldquoMarsrsquo surface radiation environment measured with the MarsScience Laboratoryrsquos Curiosity Roverrdquo Science vol 343 no6169 Article ID 1244797 2014
[13] M Durante and F A Cucinotta ldquoHeavy ion carcinogenesis andhuman space explorationrdquo Nature Reviews Cancer vol 8 no 6pp 465ndash472 2008
[14] C Arena V De Micco and A De Maio ldquoGrowth alterationand leaf biochemical responses in P vulgaris plants exposed todifferent doses of ionizing radiationrdquo Plant Biology vol 16 pp194ndash202 2014
[15] R W Holst and D J Nagel ldquoRadiation effects on plantsrdquo inPlants For Environmental Studies W Wang J W Gorsuch andJ S Hughes Eds pp 37ndash81 Lewis Publishers Boca Raton FlaUSA 1997
[16] T D Luckey Hormesis With Ionizing Radiation CRC PressBoca Raton Fla USA 1980
[17] D Marcu V Cristea and L Daraban ldquoDose-dependent effectsof gamma radiation on lettuce (Lactuca sativa var capitata)seedlingsrdquo International Journal of Radiation Biology vol 89 pp219ndash223 2013
[18] M Mei H Deng Y Lu et al ldquoMutagenic effects of heavy ionradiation in plantsrdquo Advances in Space Research vol 14 no 10pp 363ndash372 1994
[19] Y LiM Liu Z Cheng andY Sun ldquoSpace environment inducedmutations prefer to occur at polymorphic sites of rice genomesrdquoAdvances in Space Research vol 40 no 4 pp 523ndash527 2007
[20] D S Kim J-B Kim E J Goh et al ldquoAntioxidant response ofArabidopsis plants to gamma irradiation genome-wide expres-sion profiling of the ROS scavenging and signal transductionpathwaysrdquo Journal of Plant Physiology vol 168 no 16 pp 1960ndash1971 2011
[21] T J Brodribb ldquoXylem hydraulic physiology the functionalbackbone of terrestrial plant productivityrdquo Plant Science vol177 no 4 pp 245ndash251 2009
[22] R M Wheeler J C Sager R P Prince andW M Knott ldquoCropproduction for advanced life support systemsmdashobservationfrom the Kennedy Space Center Breadboard Projectrdquo NASATM-2003-211184 NASA Kennedy Space Center Florida 2003
[23] R M Wheeler C L Mackowiak G W Stutte et al ldquoCropproductivities and radiation use efficiencies for bioregenerativelife supportrdquo Advances in Space Research vol 41 no 5 pp 706ndash713 2008
[24] R Paradiso V De Micco R Buonomo G Aronne G Bar-bieri and S De Pascale ldquoSoilless cultivation of soybean forBioregenerative Life-Support Systems a literature review andthe experience of the MELiSSA ProjectmdashFood characterisationPhase Irdquo Plant Biology vol 16 pp 69ndash78 2014
[25] D M Porterfield ldquoThe biophysical limitations in physiologicaltransport and exchange in plants grown in microgravityrdquoJournal of Plant Growth Regulation vol 21 no 2 pp 177ndash1902002
[26] K Yokthongwattana and A Melis ldquoPhotoinhibition and recov-ery in oxygenic photosynthesis mechanism of a photosystem IIdamage and repair cyclerdquo in Photoprotection PhotoinhibitionGene Regulation and Environment B Demmig-Adams WW Adams III and A K Mattoo Eds pp 175ndash191 SpringerDordecht The Netherlands 2006
[27] M T Giardi G Rea M D Lambreva et al ldquoMutations ofphotosystem II D1 protein that empower efficient phenotypesof Chlamydomonas reinhardtii under extreme environment inSpacerdquo PLoS ONE vol 8 Article ID e64352 2013
[28] C Arena V De Micco G Aronne M Pugliese A Virzo DeSanto andADeMaio ldquoResponse ofPhaseolus vulgarisL plantsto low-let ionizing radiation growth and oxidative stressrdquo ActaAstronautica vol 91 pp 107ndash114 2013
[29] V Lattanzio P A Kroon and S Quideau ldquoPlant phenolicsmdashsecondary metabolites with diverse functionsrdquo in RecentAdvances in Polyphenol Research F Daay and V Lattanzio Edspp 1ndash35 Wiley-Blackwell Oxford UK 2008
[30] V De Micco C Arena and G Aronne ldquoAnatomical alterationsof Phaseolus vulgaris L mature leaves irradiated with X-raysrdquoPlant Biology vol 16 pp 187ndash193 2014
[31] A D Krikorian ldquoPlants and somatic embryos in space whathave we learnedrdquo Gravitational and Space Biology Bulletin vol11 no 2 pp 5ndash14 1998
[32] R Meissner Y Jacobson S Melamed et al ldquoA new modelsystem for tomato geneticsrdquoThe Plant Journal vol 12 no 6 pp1465ndash1472 1997
[33] E Emmanuel and A A Levy ldquoTomato mutants as tools forfunctional genomicsrdquo Current Opinion in Plant Biology vol 5no 2 pp 112ndash117 2002
[34] E Martı C Gisbert G J Bishop M S Dixon and J LGarcıa-Martınez ldquoGenetic and physiological characterizationof tomato cv Micro-Tomrdquo Journal of Experimental Botany vol57 pp 2037ndash2047 2006
The Scientific World Journal 13
[35] T Kurimoto J V H Constable and A Huda ldquoEffects ofionizing radiation exposure on arabidopsis thalianardquo HealthPhysics vol 99 no 1 pp 49ndash57 2010
[36] L Reale D Gigante F Landucci F Ferranti and R VenanzonildquoMorphological and histo-anatomical traits reflect die-back inPhragmites australis (Cav) Steudrdquo Aquatic Botany vol 103 pp122ndash128 2012
[37] K Fukuzawa ldquoUltraviolet microscopyrdquo in Methods in LigninChemistry S Y Lin andCWDence Eds pp 110ndash131 SpringerBerlin Germany 1992
[38] S E Ruzin Plant Microtechnique and Microscopy OxfordUniversity Press New York NY USA 1999
[39] V De Micco G Aronne J-P Joseleau and K Ruel ldquoXylemdevelopment and cell wall changes of soybean seedlings grownin spacerdquo Annals of Botany vol 101 no 5 pp 661ndash669 2008
[40] S Van Buggenhout T Grauwet A Van Loey and M Hen-drickx ldquoStructureprocessing relation of vacuum infused straw-berry tissue frozen under different conditionsrdquo European FoodResearch and Technology vol 226 no 3 pp 437ndash448 2008
[41] V De Micco and G Aronne ldquoBiometric anatomy of seedlingsdeveloped onboard of Foton-M2 in an automatic system sup-porting growthrdquoActa Astronautica vol 62 no 8-9 pp 505ndash5132008
[42] B Genty J M Briantais and N R Baker ldquoThe relationshipbetween quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescencerdquo Biochimica andBiophysica Acta vol 990 pp 87ndash92 1989
[43] W Bilger and O Bjorkman ldquoRole of the xanthophyll cycle inphotoprotection elucidated by measurements of light-inducedabsorbance changes fluorescence and photosynthesis in leavesof Hedera canariensisrdquo Photosynthesis Research vol 25 no 3pp 173ndash185 1990
[44] H K Lichtenthaler ldquoChlorophylls and carotenoids pigmentsof photosynthetic biomembranesrdquoMethods in Enzymology vol148 pp 350ndash382 1987
[45] J P Maity DMishra A Chakraborty A Saha S C Santra andS Chanda ldquoModulation of some quantitative and qualitativecharacteristics in rice (Oryza sativa L) and mung (Phaseolusmungo L) by ionizing radiationrdquo Radiation Physics and Chem-istry vol 74 no 5 pp 391ndash394 2005
[46] L Cheng H Yang B Lin et al ldquoEffect of gamma-ray radiationon physiological morphological characters and chromosomeaberrations of minitubers in Solanum tuberosum Lrdquo Interna-tional Journal of Radiation Biology vol 86 no 9 pp 791ndash7992010
[47] B Al-Safadi and R Elias ldquoImprovement of caper (Capparisspinosa L) propagation using in vitro culture and gammairradiationrdquo Scientia Horticulturae vol 127 no 3 pp 290ndash2972011
[48] O Monje G W Stutte G D Goins D M Porterfield and GE Bingham ldquoFarming in space environmental and biophysicalconcernsrdquo Advances in Space Research vol 31 no 1 pp 151ndash1672003
[49] V De Micco M Scala and G Aronne ldquoEffects of simulatedmicrogravity on male gametophyte of Prunus Pyrus andBrassica speciesrdquo Protoplasma vol 228 no 1ndash3 pp 121ndash1262006
[50] V De Micco R Buonomo R Paradiso S De Pascale and GAronne ldquoSoybean cultivar selection for Bioregenerative LifeSupport Systems (BLSS)mdashtheoretical selectionrdquo Advances inSpace Research vol 49 no 10 pp 1415ndash1421 2012
[51] T Kozuka S-G Kong M Doi K-I Shimazaki and ANagatani ldquoTissue-autonomous promotion of palisade celldevelopment by phototropin 2 in Arabidopsisrdquo Plant Cell vol23 no 10 pp 3684ndash3695 2011
[52] E Kovacs and A Keresztes ldquoEffect of gamma and UV-BCradiation on plant cellsrdquoMicron vol 33 no 2 pp 199ndash210 2002
[53] R Ogaya J Penuelas D Asensio and J Llusia ldquoChlorophyllfluorescence responses to temperature and water availabilityin two co-dominant Mediterranean shrub and tree speciesin a long-term field experiment simulating climate changerdquoEnvironmental and Experimental Botany vol 73 no 1 pp 89ndash93 2011
[54] R Zaka C M Vandecasteele and M T Misset ldquoEffects oflow chronic doses of ionizing radiation on antioxidant enzymesand G6PDH activities in Stipa capillata (Poaceae)rdquo Journal ofExperimental Botany vol 53 no 376 pp 1979ndash1987 2002
[55] J-H KimM-H Baek Y C Byung GW Seung and J-S KimldquoAlterations in the photosynthetic pigments and antioxidantmachineries of red pepper (Capsicum annuum L) seedlingsfrom gamma-irradiated seedsrdquo Journal of Plant Biology vol 47no 4 pp 314ndash321 2004
[56] N A Al-Enezi and J M Al-Khayri ldquoAlterations of DNA ionsand photosynthetic pigments content in date palm seedlingsinduced by X-irradiationrdquo International Journal of Agricultureand Biology vol 14 pp 329ndash336 2012
[57] V de Micco C Arena L Vitale G Aronne and A V de SantoldquoAnatomy and photochemical behaviour of mediterranean Cis-tus incanus winter leaves under natural outdoor and warmerindoor conditionsrdquo Botany vol 89 no 10 pp 677ndash688 2011
[58] L H Levine H G Levine E C Stryjewski V Prima and WC Piastuch ldquoEffect of spaceflight on isoflavonoid accumulationin etiolated soybean seedlingsrdquo Journal of Gravitational Physiol-ogy vol 8 no 2 pp 21ndash27 2001
[59] X Fan ldquoAntioxidant capacity of fresh-cut vegetables exposedto ionizing radiationrdquo Journal of the Science of Food andAgriculture vol 85 no 6 pp 995ndash1000 2005
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
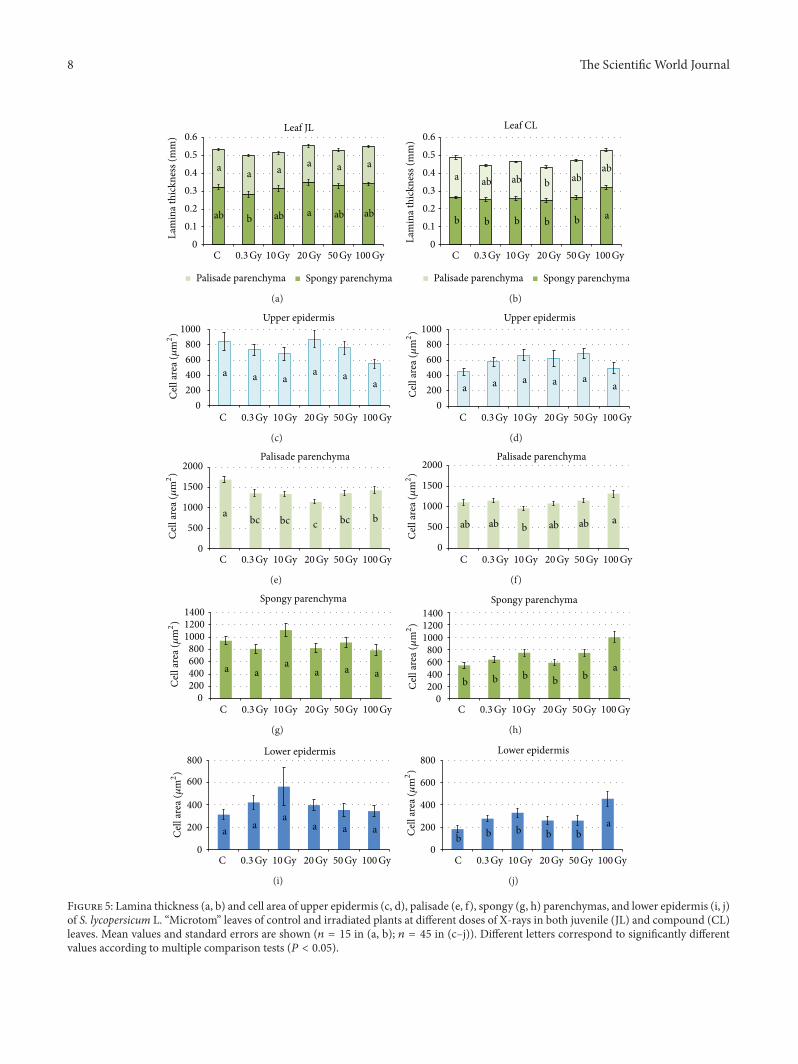
8 The Scientific World Journal
Palisade parenchyma Spongy parenchyma
ab b ab a ab ab
a a a a a a
0010203040506
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Leaf JL
Lam
ina t
hick
ness
(mm
)
(a)
Palisade parenchyma Spongy parenchyma
b b b b b a
a ab ab b abab
Leaf CL
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
010203040506
Lam
ina t
hick
ness
(mm
)
(b)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a a aa a
a
0200400600800
1000Upper epidermis
Cel
l are
a (120583
m2)
(c)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
1000
a a a a aa
Upper epidermis
Cel
l are
a (120583
m2)
(d)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
a bc bc c bc b
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(e)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Palisade parenchyma
ab ab b ab ab a
0
500
1000
1500
2000
Cel
l are
a (120583
m2)
(f)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
a aa
a a a
0200400600800
100012001400
Cel
l are
a (120583
m2)
(g)
Spongy parenchyma
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy0
200400600800
100012001400
b b b b ba
Cel
l are
a (120583
m2)
(h)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
a aa
a a a
0
200
400
600
800
Cel
l are
a (120583
m2)
(i)
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Lower epidermis
b b b b ba
0
200
400
600
800
Cel
l are
a (120583
m2)
(j)
Figure 5 Lamina thickness (a b) and cell area of upper epidermis (c d) palisade (e f) spongy (g h) parenchymas and lower epidermis (i j)of S lycopersicum L ldquoMicrotomrdquo leaves of control and irradiated plants at different doses of X-rays in both juvenile (JL) and compound (CL)leaves Mean values and standard errors are shown (119899 = 15 in (a b) 119899 = 45 in (cndashj)) Different letters correspond to significantly differentvalues according to multiple comparison tests (119875 lt 005)
The Scientific World Journal 9
04
05
06
07
08
a a a a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
ΦPS
II
(a)
00
02
04
06
08
NPQ a
a aa a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
(b)
070
075
080
085
090
a
b b b b b
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
FF
m
(c)
Figure 6 Quantum yield of PSII electron transport (ΦPSII a) nonphotochemical quenching (NPQ b) and maximal PSII photochemicalefficiency (FvFm c) in compound (CL) S lycopersicum L ldquoMicrotomrdquo leaves of control and plants from seeds irradiated at different dosesof X-rays Mean values and standard deviations are shown (119899 = 5) Different letters correspond to significantly different values according tomultiple comparison tests (119875 lt 005)
0
1
2
3
4
a
aa
a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Chl a
+ b
(mg g
minus1)
(a)
0
02
04
06
a
aa a
a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Car x
+ c
(mg g
minus1)
(b)
Figure 7 Total chlorophyll content (Chl a+b) (a) and total carotenoid content (Car x+c) (b) in compound (CL) S lycopersicum L ldquoMicrotomrdquoleaves of control and plants from seeds irradiated at different doses of X-rays Mean values and standard errors are shown (119899 = 5) Differentletters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
induced in ldquoMicrotomrdquo especially by low doses of irradiationcan be beneficial to atmosphere regeneration of pressurisedmodules provided that formed leaves do not show structuralaberrations which reduce their function
General anatomical structure of both juvenile and com-pound ldquoMicrotomrdquo leaves was only slightly perturbed inplants from seeds subjected to the highest levels of radiation
Clear-cut dose-depended tendencies of variations were notfound in lamina thickness and cell size and shape of thevarious tissues The formation of smaller palisade cells wasfound only in juvenile leaves of irradiated seeds while anincrease in cell size was found only in spongy parenchymaand lower epidermis of compound leaves from seeds irra-diated at 100Gy The diverse growth behaviour found in
10 The Scientific World Journal
Table 1 Cell shape in different tissues of juvenile leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown (119899 = 75)Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf JL Aspect ratio Sphericity Convexity
Upper epidermis
C 197 plusmn 016 a 0308 plusmn 0050 a 0926 plusmn 0021 ab03Gy 200 plusmn 013 a 0290 plusmn 0038 a 0941 plusmn 0013 a10Gy 210 plusmn 012 a 0239 plusmn 0037 a 0898 plusmn 0016 ab20Gy 217 plusmn 014 a 0258 plusmn 0045 a 0909 plusmn 0019 abc50Gy 170 plusmn 008 a 0376 plusmn 0037 a 0927 plusmn 0016 ab100Gy 197 plusmn 014 a 0286 plusmn 0042 a 0876 plusmn 0028 c
Palisade parenchyma
C 290 plusmn 010 a 0109 plusmn 0015 a 0879 plusmn 0010 a03 Gy 304 plusmn 012 a 0099 plusmn 0016 a 0805 plusmn 0016 b10Gy 309 plusmn 013 a 0093 plusmn 0019 a 0792 plusmn 0015 b20Gy 283 plusmn 011 a 0101 plusmn 0017 a 0784 plusmn 0017 b50Gy 314 plusmn 014 a 0082 plusmn 0016 a 0772 plusmn 0016 b100Gy 275 plusmn 011 a 0119 plusmn 0020 a 0805 plusmn 0017 b
Spongy parenchyma
C 190 plusmn 008 a 0307 plusmn 0033 a 0893 plusmn 0012 a03 Gy 180 plusmn 007 a 0341 plusmn 0029 a 0876 plusmn 0013 a10Gy 203 plusmn 010 a 0290 plusmn 0031 a 0901 plusmn 0012 a20Gy 194 plusmn 012 a 0344 plusmn 0040 a 0881 plusmn 0015 a50Gy 174 plusmn 009 a 0393 plusmn 0035 a 0883 plusmn 0014 a100Gy 198 plusmn 012 a 0322 plusmn 0037 a 0878 plusmn 0015 a
Lower epidermis
C 178 plusmn 009 bc 0343 plusmn 0045 ab 0919 plusmn 0019 a03 Gy 225 plusmn 017 ab 0263 plusmn 0044 b 0933 plusmn 0024 a10Gy 219 plusmn 011 abc 0218 plusmn 0034 b 0888 plusmn 0020 ab20Gy 180 plusmn 013 bc 0378 plusmn 0047 ab 0926 plusmn 0017 a50Gy 167 plusmn 001 c 0461 plusmn 0051 a 0924 plusmn 0021 a100Gy 238 plusmn 021 a 0257 plusmn 0057 b 0857 plusmn 0030 b
the various tissues of structures of different age is in agree-ment with the different light-mediated control of cell devel-opment in the various leaf tissues [51] Altered mesophyllcharacterized by partly collapsed cells with irregular cellshape was found only sporadically after the exposure of seedsto 50 and 100Gy in theMicrotomcultivar Suchmodificationswere less marked than those found in leaves of bean wherethe target organs for irradiation were the leaves themselves[30] leaves directly exposed to X-rays in bean showed theformation of more loose parenchyma with larger cells likelydue to a radiation-induced damage to cell walls that reducedmechanical constraints to cell enlargement [30 52] Furtherinvestigations on cell wall composition would be useful toelucidate whether a possible alteration of cell walls wasresponsible for cell enlargement in the spongy parenchymaand epidermis of leaves from seeds of ldquoMicrotomrdquo irradiatedwith the highest dose of X-rays However in the cultivarMicrotom the slight alterations of mesophyll structure didnot impair photosynthetic efficiency as demonstrated by thelack of significant dose-dependent variation trends in thequantum yield of PSII electron transport (ΦPSII) nonphoto-chemical quenching (NPQ) and maximal PSII photochemi-cal efficiency (FvFm) The increased values of FvFm closeto 08 which is considered an optimal value for healthy plants[53] suggested that the photosynthetic apparatus of the leavesfrom irradiated seeds has higher levels of photochemical
efficiency irrespective of dose The unchanged or increasedcontent of both chlorophyll and carotenoid pigments con-firmed that the photosynthetic apparatus remained stablealso in terms of light harvesting capacity and indicatedno deleterious effect of radiation on plant photosyntheticmachinery RecentlyMarcu et al [17] showed that irradiationof lettuce seeds with gamma rays up to 30Gy enhancedthe content of chlorophyll a chlorophyll b and carotenoidswhile higher doses (up to 70Gy) resulted in their decreaseHowever the effect of ionising radiation on photosyntheticpigment content is quite controversial Increased pigmentcontent is ascribed to the phenomenon of hormesis whilea lower content is generally ascribed to the inhibition ofchlorophyll and carotenoid synthesis or to oxidation due tothe high levels of radiation-induced water radiolysis [2 1454ndash56] Some plants possess natural screens against radiationwhen their tissues contain phenolic compounds with antioxi-dant functions and can counteract photoinhibitory processes[29 57] An increase of phenolic compounds linked to thechloroplast membranes has been recently found in Phaseolusvulgaris irradiated leaves exposed to high levels of X-rays[28 30] Changes in phenolic content after the exposure toSpace can be attributed either to direct effects on the synthesisor mobilization of phenolic compounds or to indirect effectsdue to changes in ultrastructural organization such as theincrease in the number of chloroplasts per cell [30 58]
The Scientific World Journal 11
Table 2 Cell shape in different tissues of compound leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown(119899 = 75) Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf CL Aspect ratio Sphericity Convexity
Upper epidermis
C 166 plusmn 007 c 0422 plusmn 0042 a 0946 plusmn 0010 a03 Gy 216 plusmn 017 ab 0244 plusmn 0033 c 0925 plusmn 0016 ab10Gy 171 plusmn 011 bc 0401 plusmn 0042 a 0914 plusmn 0013 b20Gy 186 plusmn 010 abc 0328 plusmn 0041 ab 0919 plusmn 0020 ab50Gy 225 plusmn 018 a 0247 plusmn 0041 c 0897 plusmn 0019 b100Gy 188 plusmn 009 abc 0319 plusmn 0045 ab 0902 plusmn 0016 b
Palisade parenchyma
C 355 plusmn 012 a 0073 plusmn 0014 b 0890 plusmn 0012 a03 Gy 289 plusmn 010 b 0102 plusmn 0013 ab 0847 plusmn 0018 b10Gy 300 plusmn 014 b 0104 plusmn 0021 ab 0826 plusmn 0017 b20Gy 302 plusmn 011 b 0094 plusmn 0013 ab 0810 plusmn 0014 b50Gy 283 plusmn 012 b 0107 plusmn 0021 ab 0750 plusmn 0021 c100Gy 275 plusmn 012 b 0122 plusmn 0023 a 0809 plusmn 0015 b
Spongy parenchyma
C 193 plusmn 010 a 0323 plusmn 0035 a 0904 plusmn 0011 a03 Gy 183 plusmn 010 a 0375 plusmn 0042 a 0879 plusmn 0013 a10Gy 176 plusmn 007 a 0357 plusmn 0034 a 0869 plusmn 0011 a20Gy 180 plusmn 009 a 0366 plusmn 0038 a 0885 plusmn 0016 a50Gy 171 plusmn 006 a 0372 plusmn 0034 a 0879 plusmn 0015 a100Gy 188 plusmn 010 a 0339 plusmn 0037 a 0883 plusmn 0015 a
Lower epidermis
C 148 plusmn 008 b 0545 plusmn 0045 a 0957 plusmn 0015 a03 Gy 210 plusmn 017 a 0295 plusmn 0046 b 0929 plusmn 0019 ab10Gy 190 plusmn 012 ab 0334 plusmn 0043 b 0893 plusmn 0019 b20Gy 178 plusmn 011 ab 0388 plusmn 0047 b 0929 plusmn 0017 ab50Gy 172 plusmn 009 ab 0367 plusmn 0040 b 0914 plusmn 0016 b100Gy 191 plusmn 014 ab 0362 plusmn 0054 b 0888 plusmn 0025 b
The lowering of phenolic content at high levels of X-rays inldquoMicrotomrdquo leaves is not in agreement with general trendsindicating enhanced phenolics production in plants grownin Space [58] or in various plant tissues directly irradiatedwith gamma rays [59] However the low content of phenolicscan be an indirect effect of increasing volume occupiedby intercellular spaces due to the loosening of parenchymacells
In conclusion our results show that irradiation of the dryseeds of the Microtom cultivar did not hamper germinationand the development of functional leaves The formationof more compact plants characterised by a higher numberof larger leaves can be valuable in terms of increasingresource regeneration in the BLSSs High levels of radiationinduced only slight structural perturbations which weresimilar in juvenile and compound leaves Such perturbationsdid not affect photosynthetic efficiency since the maximalPSII photochemical efficiency was even increased after irra-diation at all doses However the tendency to accumulatelower amounts of phenolic compounds along chloroplastmembranes after irradiation of seeds with high doses ofradiation should be further investigated because it coulddetermine the reduction of natural photoprotection [57]Finally the overall results indicate that theMicrotom cultivarirradiated at the seed stage has high radioresistance duringthe growth which is a desirable feature for plant cultivation
in Space Radioresistance together with dwarf growth andshort life cycle makes this cultivar a valuable candidate forthe cultivation in BLSSs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are grateful to the ldquoCompagnia di San Paolordquo(Turin) and the ldquoPolo delle Scienze e delle Tecnologie perla Vitardquo of the University of Naples Federico II for thefinancial support The authors also thank Maria Giordanoand Pasquale Scognamiglio for their support in plant culti-vation and microscopy analyses The authors acknowledgefour anonymous editors for helpful comments on a previousversion of the paper
References
[1] A Real S Sundell-Bergman J F Knowles D S Woodheadand I Zinger ldquoEffects of ionising radiation exposure on plantsfish and mammals relevant data for environmental radiation
12 The Scientific World Journal
protectionrdquo Journal of Radiological Protection vol 24 no 4 ppA123ndashA137 2004
[2] V De Micco C Arena D Pignalosa and M Durante ldquoEffectsof sparsely and densely ionizing radiation on plantsrdquo Radiationand Environmental Biophysics vol 50 no 1 pp 1ndash19 2011
[3] N Kumari P Kumar D Mitra B Prasad B N Tiwary andL Varshney ldquoEffects of ionizing radiation on microbial decon-tamination phenolic contents and antioxidant properties oftriphalardquo Journal of Food Science vol 74 no 3 pp M109ndashM1132009
[4] A Tanaka N Shikazono and Y Hase ldquoStudies on biologicaleffects of ion beams on lethality molecular nature of mutationmutation rate and spectrum of mutation phenotype for muta-tion breeding in higher plantsrdquo Journal of Radiation Researchvol 51 no 3 pp 223ndash233 2010
[5] M Durante and A Kronenberg ldquoGround-based research withheavy ions for space radiation protectionrdquo Advances in SpaceResearch vol 35 no 2 pp 180ndash184 2005
[6] E Hu S I Bartsev and H Liu ldquoConceptual design of a biore-generative life support system containing crops and silkwormsrdquoAdvances in Space Research vol 45 no 7 pp 929ndash939 2010
[7] V deMicco G Aronne G Colla R Fortezza and S de PascaleldquoAgro-biology for bioregenerative life support systems in long-term space missions general constraints and the Italian effortsrdquoJournal of Plant Interactions vol 4 no 4 pp 241ndash252 2009
[8] T W Halstead and F R Dutcher ldquoPlants in spacerdquo AnnualReview of Plant Physiology vol 38 pp 317ndash345 1987
[9] V De Micco S De Pascale R Paradiso and G AronneldquoMicrogravity effects on different stages of higher plant life cycleand completion of the seed-to-seed cyclerdquo Plant Biology vol 16pp 31ndash38 2014
[10] C J Bennett C Pirim and T M Orlando ldquoSpace-weatheringof solar system bodies a laboratory perspectiverdquo ChemicalReviews vol 113 pp 9086ndash9150 2013
[11] R A Kerr ldquoRadiation will make astronautsrsquo trip to Mars evenriskierrdquo Science vol 340 article 1031 2014
[12] D M Hassler C Zeitlin R F Wimmer-Schweingruber et alldquoMarsrsquo surface radiation environment measured with the MarsScience Laboratoryrsquos Curiosity Roverrdquo Science vol 343 no6169 Article ID 1244797 2014
[13] M Durante and F A Cucinotta ldquoHeavy ion carcinogenesis andhuman space explorationrdquo Nature Reviews Cancer vol 8 no 6pp 465ndash472 2008
[14] C Arena V De Micco and A De Maio ldquoGrowth alterationand leaf biochemical responses in P vulgaris plants exposed todifferent doses of ionizing radiationrdquo Plant Biology vol 16 pp194ndash202 2014
[15] R W Holst and D J Nagel ldquoRadiation effects on plantsrdquo inPlants For Environmental Studies W Wang J W Gorsuch andJ S Hughes Eds pp 37ndash81 Lewis Publishers Boca Raton FlaUSA 1997
[16] T D Luckey Hormesis With Ionizing Radiation CRC PressBoca Raton Fla USA 1980
[17] D Marcu V Cristea and L Daraban ldquoDose-dependent effectsof gamma radiation on lettuce (Lactuca sativa var capitata)seedlingsrdquo International Journal of Radiation Biology vol 89 pp219ndash223 2013
[18] M Mei H Deng Y Lu et al ldquoMutagenic effects of heavy ionradiation in plantsrdquo Advances in Space Research vol 14 no 10pp 363ndash372 1994
[19] Y LiM Liu Z Cheng andY Sun ldquoSpace environment inducedmutations prefer to occur at polymorphic sites of rice genomesrdquoAdvances in Space Research vol 40 no 4 pp 523ndash527 2007
[20] D S Kim J-B Kim E J Goh et al ldquoAntioxidant response ofArabidopsis plants to gamma irradiation genome-wide expres-sion profiling of the ROS scavenging and signal transductionpathwaysrdquo Journal of Plant Physiology vol 168 no 16 pp 1960ndash1971 2011
[21] T J Brodribb ldquoXylem hydraulic physiology the functionalbackbone of terrestrial plant productivityrdquo Plant Science vol177 no 4 pp 245ndash251 2009
[22] R M Wheeler J C Sager R P Prince andW M Knott ldquoCropproduction for advanced life support systemsmdashobservationfrom the Kennedy Space Center Breadboard Projectrdquo NASATM-2003-211184 NASA Kennedy Space Center Florida 2003
[23] R M Wheeler C L Mackowiak G W Stutte et al ldquoCropproductivities and radiation use efficiencies for bioregenerativelife supportrdquo Advances in Space Research vol 41 no 5 pp 706ndash713 2008
[24] R Paradiso V De Micco R Buonomo G Aronne G Bar-bieri and S De Pascale ldquoSoilless cultivation of soybean forBioregenerative Life-Support Systems a literature review andthe experience of the MELiSSA ProjectmdashFood characterisationPhase Irdquo Plant Biology vol 16 pp 69ndash78 2014
[25] D M Porterfield ldquoThe biophysical limitations in physiologicaltransport and exchange in plants grown in microgravityrdquoJournal of Plant Growth Regulation vol 21 no 2 pp 177ndash1902002
[26] K Yokthongwattana and A Melis ldquoPhotoinhibition and recov-ery in oxygenic photosynthesis mechanism of a photosystem IIdamage and repair cyclerdquo in Photoprotection PhotoinhibitionGene Regulation and Environment B Demmig-Adams WW Adams III and A K Mattoo Eds pp 175ndash191 SpringerDordecht The Netherlands 2006
[27] M T Giardi G Rea M D Lambreva et al ldquoMutations ofphotosystem II D1 protein that empower efficient phenotypesof Chlamydomonas reinhardtii under extreme environment inSpacerdquo PLoS ONE vol 8 Article ID e64352 2013
[28] C Arena V De Micco G Aronne M Pugliese A Virzo DeSanto andADeMaio ldquoResponse ofPhaseolus vulgarisL plantsto low-let ionizing radiation growth and oxidative stressrdquo ActaAstronautica vol 91 pp 107ndash114 2013
[29] V Lattanzio P A Kroon and S Quideau ldquoPlant phenolicsmdashsecondary metabolites with diverse functionsrdquo in RecentAdvances in Polyphenol Research F Daay and V Lattanzio Edspp 1ndash35 Wiley-Blackwell Oxford UK 2008
[30] V De Micco C Arena and G Aronne ldquoAnatomical alterationsof Phaseolus vulgaris L mature leaves irradiated with X-raysrdquoPlant Biology vol 16 pp 187ndash193 2014
[31] A D Krikorian ldquoPlants and somatic embryos in space whathave we learnedrdquo Gravitational and Space Biology Bulletin vol11 no 2 pp 5ndash14 1998
[32] R Meissner Y Jacobson S Melamed et al ldquoA new modelsystem for tomato geneticsrdquoThe Plant Journal vol 12 no 6 pp1465ndash1472 1997
[33] E Emmanuel and A A Levy ldquoTomato mutants as tools forfunctional genomicsrdquo Current Opinion in Plant Biology vol 5no 2 pp 112ndash117 2002
[34] E Martı C Gisbert G J Bishop M S Dixon and J LGarcıa-Martınez ldquoGenetic and physiological characterizationof tomato cv Micro-Tomrdquo Journal of Experimental Botany vol57 pp 2037ndash2047 2006
The Scientific World Journal 13
[35] T Kurimoto J V H Constable and A Huda ldquoEffects ofionizing radiation exposure on arabidopsis thalianardquo HealthPhysics vol 99 no 1 pp 49ndash57 2010
[36] L Reale D Gigante F Landucci F Ferranti and R VenanzonildquoMorphological and histo-anatomical traits reflect die-back inPhragmites australis (Cav) Steudrdquo Aquatic Botany vol 103 pp122ndash128 2012
[37] K Fukuzawa ldquoUltraviolet microscopyrdquo in Methods in LigninChemistry S Y Lin andCWDence Eds pp 110ndash131 SpringerBerlin Germany 1992
[38] S E Ruzin Plant Microtechnique and Microscopy OxfordUniversity Press New York NY USA 1999
[39] V De Micco G Aronne J-P Joseleau and K Ruel ldquoXylemdevelopment and cell wall changes of soybean seedlings grownin spacerdquo Annals of Botany vol 101 no 5 pp 661ndash669 2008
[40] S Van Buggenhout T Grauwet A Van Loey and M Hen-drickx ldquoStructureprocessing relation of vacuum infused straw-berry tissue frozen under different conditionsrdquo European FoodResearch and Technology vol 226 no 3 pp 437ndash448 2008
[41] V De Micco and G Aronne ldquoBiometric anatomy of seedlingsdeveloped onboard of Foton-M2 in an automatic system sup-porting growthrdquoActa Astronautica vol 62 no 8-9 pp 505ndash5132008
[42] B Genty J M Briantais and N R Baker ldquoThe relationshipbetween quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescencerdquo Biochimica andBiophysica Acta vol 990 pp 87ndash92 1989
[43] W Bilger and O Bjorkman ldquoRole of the xanthophyll cycle inphotoprotection elucidated by measurements of light-inducedabsorbance changes fluorescence and photosynthesis in leavesof Hedera canariensisrdquo Photosynthesis Research vol 25 no 3pp 173ndash185 1990
[44] H K Lichtenthaler ldquoChlorophylls and carotenoids pigmentsof photosynthetic biomembranesrdquoMethods in Enzymology vol148 pp 350ndash382 1987
[45] J P Maity DMishra A Chakraborty A Saha S C Santra andS Chanda ldquoModulation of some quantitative and qualitativecharacteristics in rice (Oryza sativa L) and mung (Phaseolusmungo L) by ionizing radiationrdquo Radiation Physics and Chem-istry vol 74 no 5 pp 391ndash394 2005
[46] L Cheng H Yang B Lin et al ldquoEffect of gamma-ray radiationon physiological morphological characters and chromosomeaberrations of minitubers in Solanum tuberosum Lrdquo Interna-tional Journal of Radiation Biology vol 86 no 9 pp 791ndash7992010
[47] B Al-Safadi and R Elias ldquoImprovement of caper (Capparisspinosa L) propagation using in vitro culture and gammairradiationrdquo Scientia Horticulturae vol 127 no 3 pp 290ndash2972011
[48] O Monje G W Stutte G D Goins D M Porterfield and GE Bingham ldquoFarming in space environmental and biophysicalconcernsrdquo Advances in Space Research vol 31 no 1 pp 151ndash1672003
[49] V De Micco M Scala and G Aronne ldquoEffects of simulatedmicrogravity on male gametophyte of Prunus Pyrus andBrassica speciesrdquo Protoplasma vol 228 no 1ndash3 pp 121ndash1262006
[50] V De Micco R Buonomo R Paradiso S De Pascale and GAronne ldquoSoybean cultivar selection for Bioregenerative LifeSupport Systems (BLSS)mdashtheoretical selectionrdquo Advances inSpace Research vol 49 no 10 pp 1415ndash1421 2012
[51] T Kozuka S-G Kong M Doi K-I Shimazaki and ANagatani ldquoTissue-autonomous promotion of palisade celldevelopment by phototropin 2 in Arabidopsisrdquo Plant Cell vol23 no 10 pp 3684ndash3695 2011
[52] E Kovacs and A Keresztes ldquoEffect of gamma and UV-BCradiation on plant cellsrdquoMicron vol 33 no 2 pp 199ndash210 2002
[53] R Ogaya J Penuelas D Asensio and J Llusia ldquoChlorophyllfluorescence responses to temperature and water availabilityin two co-dominant Mediterranean shrub and tree speciesin a long-term field experiment simulating climate changerdquoEnvironmental and Experimental Botany vol 73 no 1 pp 89ndash93 2011
[54] R Zaka C M Vandecasteele and M T Misset ldquoEffects oflow chronic doses of ionizing radiation on antioxidant enzymesand G6PDH activities in Stipa capillata (Poaceae)rdquo Journal ofExperimental Botany vol 53 no 376 pp 1979ndash1987 2002
[55] J-H KimM-H Baek Y C Byung GW Seung and J-S KimldquoAlterations in the photosynthetic pigments and antioxidantmachineries of red pepper (Capsicum annuum L) seedlingsfrom gamma-irradiated seedsrdquo Journal of Plant Biology vol 47no 4 pp 314ndash321 2004
[56] N A Al-Enezi and J M Al-Khayri ldquoAlterations of DNA ionsand photosynthetic pigments content in date palm seedlingsinduced by X-irradiationrdquo International Journal of Agricultureand Biology vol 14 pp 329ndash336 2012
[57] V de Micco C Arena L Vitale G Aronne and A V de SantoldquoAnatomy and photochemical behaviour of mediterranean Cis-tus incanus winter leaves under natural outdoor and warmerindoor conditionsrdquo Botany vol 89 no 10 pp 677ndash688 2011
[58] L H Levine H G Levine E C Stryjewski V Prima and WC Piastuch ldquoEffect of spaceflight on isoflavonoid accumulationin etiolated soybean seedlingsrdquo Journal of Gravitational Physiol-ogy vol 8 no 2 pp 21ndash27 2001
[59] X Fan ldquoAntioxidant capacity of fresh-cut vegetables exposedto ionizing radiationrdquo Journal of the Science of Food andAgriculture vol 85 no 6 pp 995ndash1000 2005
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

The Scientific World Journal 9
04
05
06
07
08
a a a a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
ΦPS
II
(a)
00
02
04
06
08
NPQ a
a aa a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
(b)
070
075
080
085
090
a
b b b b b
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
FF
m
(c)
Figure 6 Quantum yield of PSII electron transport (ΦPSII a) nonphotochemical quenching (NPQ b) and maximal PSII photochemicalefficiency (FvFm c) in compound (CL) S lycopersicum L ldquoMicrotomrdquo leaves of control and plants from seeds irradiated at different dosesof X-rays Mean values and standard deviations are shown (119899 = 5) Different letters correspond to significantly different values according tomultiple comparison tests (119875 lt 005)
0
1
2
3
4
a
aa
a a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Chl a
+ b
(mg g
minus1)
(a)
0
02
04
06
a
aa a
a a
C 03 Gy 10 Gy 20 Gy 50 Gy 100 Gy
Car x
+ c
(mg g
minus1)
(b)
Figure 7 Total chlorophyll content (Chl a+b) (a) and total carotenoid content (Car x+c) (b) in compound (CL) S lycopersicum L ldquoMicrotomrdquoleaves of control and plants from seeds irradiated at different doses of X-rays Mean values and standard errors are shown (119899 = 5) Differentletters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
induced in ldquoMicrotomrdquo especially by low doses of irradiationcan be beneficial to atmosphere regeneration of pressurisedmodules provided that formed leaves do not show structuralaberrations which reduce their function
General anatomical structure of both juvenile and com-pound ldquoMicrotomrdquo leaves was only slightly perturbed inplants from seeds subjected to the highest levels of radiation
Clear-cut dose-depended tendencies of variations were notfound in lamina thickness and cell size and shape of thevarious tissues The formation of smaller palisade cells wasfound only in juvenile leaves of irradiated seeds while anincrease in cell size was found only in spongy parenchymaand lower epidermis of compound leaves from seeds irra-diated at 100Gy The diverse growth behaviour found in
10 The Scientific World Journal
Table 1 Cell shape in different tissues of juvenile leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown (119899 = 75)Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf JL Aspect ratio Sphericity Convexity
Upper epidermis
C 197 plusmn 016 a 0308 plusmn 0050 a 0926 plusmn 0021 ab03Gy 200 plusmn 013 a 0290 plusmn 0038 a 0941 plusmn 0013 a10Gy 210 plusmn 012 a 0239 plusmn 0037 a 0898 plusmn 0016 ab20Gy 217 plusmn 014 a 0258 plusmn 0045 a 0909 plusmn 0019 abc50Gy 170 plusmn 008 a 0376 plusmn 0037 a 0927 plusmn 0016 ab100Gy 197 plusmn 014 a 0286 plusmn 0042 a 0876 plusmn 0028 c
Palisade parenchyma
C 290 plusmn 010 a 0109 plusmn 0015 a 0879 plusmn 0010 a03 Gy 304 plusmn 012 a 0099 plusmn 0016 a 0805 plusmn 0016 b10Gy 309 plusmn 013 a 0093 plusmn 0019 a 0792 plusmn 0015 b20Gy 283 plusmn 011 a 0101 plusmn 0017 a 0784 plusmn 0017 b50Gy 314 plusmn 014 a 0082 plusmn 0016 a 0772 plusmn 0016 b100Gy 275 plusmn 011 a 0119 plusmn 0020 a 0805 plusmn 0017 b
Spongy parenchyma
C 190 plusmn 008 a 0307 plusmn 0033 a 0893 plusmn 0012 a03 Gy 180 plusmn 007 a 0341 plusmn 0029 a 0876 plusmn 0013 a10Gy 203 plusmn 010 a 0290 plusmn 0031 a 0901 plusmn 0012 a20Gy 194 plusmn 012 a 0344 plusmn 0040 a 0881 plusmn 0015 a50Gy 174 plusmn 009 a 0393 plusmn 0035 a 0883 plusmn 0014 a100Gy 198 plusmn 012 a 0322 plusmn 0037 a 0878 plusmn 0015 a
Lower epidermis
C 178 plusmn 009 bc 0343 plusmn 0045 ab 0919 plusmn 0019 a03 Gy 225 plusmn 017 ab 0263 plusmn 0044 b 0933 plusmn 0024 a10Gy 219 plusmn 011 abc 0218 plusmn 0034 b 0888 plusmn 0020 ab20Gy 180 plusmn 013 bc 0378 plusmn 0047 ab 0926 plusmn 0017 a50Gy 167 plusmn 001 c 0461 plusmn 0051 a 0924 plusmn 0021 a100Gy 238 plusmn 021 a 0257 plusmn 0057 b 0857 plusmn 0030 b
the various tissues of structures of different age is in agree-ment with the different light-mediated control of cell devel-opment in the various leaf tissues [51] Altered mesophyllcharacterized by partly collapsed cells with irregular cellshape was found only sporadically after the exposure of seedsto 50 and 100Gy in theMicrotomcultivar Suchmodificationswere less marked than those found in leaves of bean wherethe target organs for irradiation were the leaves themselves[30] leaves directly exposed to X-rays in bean showed theformation of more loose parenchyma with larger cells likelydue to a radiation-induced damage to cell walls that reducedmechanical constraints to cell enlargement [30 52] Furtherinvestigations on cell wall composition would be useful toelucidate whether a possible alteration of cell walls wasresponsible for cell enlargement in the spongy parenchymaand epidermis of leaves from seeds of ldquoMicrotomrdquo irradiatedwith the highest dose of X-rays However in the cultivarMicrotom the slight alterations of mesophyll structure didnot impair photosynthetic efficiency as demonstrated by thelack of significant dose-dependent variation trends in thequantum yield of PSII electron transport (ΦPSII) nonphoto-chemical quenching (NPQ) and maximal PSII photochemi-cal efficiency (FvFm) The increased values of FvFm closeto 08 which is considered an optimal value for healthy plants[53] suggested that the photosynthetic apparatus of the leavesfrom irradiated seeds has higher levels of photochemical
efficiency irrespective of dose The unchanged or increasedcontent of both chlorophyll and carotenoid pigments con-firmed that the photosynthetic apparatus remained stablealso in terms of light harvesting capacity and indicatedno deleterious effect of radiation on plant photosyntheticmachinery RecentlyMarcu et al [17] showed that irradiationof lettuce seeds with gamma rays up to 30Gy enhancedthe content of chlorophyll a chlorophyll b and carotenoidswhile higher doses (up to 70Gy) resulted in their decreaseHowever the effect of ionising radiation on photosyntheticpigment content is quite controversial Increased pigmentcontent is ascribed to the phenomenon of hormesis whilea lower content is generally ascribed to the inhibition ofchlorophyll and carotenoid synthesis or to oxidation due tothe high levels of radiation-induced water radiolysis [2 1454ndash56] Some plants possess natural screens against radiationwhen their tissues contain phenolic compounds with antioxi-dant functions and can counteract photoinhibitory processes[29 57] An increase of phenolic compounds linked to thechloroplast membranes has been recently found in Phaseolusvulgaris irradiated leaves exposed to high levels of X-rays[28 30] Changes in phenolic content after the exposure toSpace can be attributed either to direct effects on the synthesisor mobilization of phenolic compounds or to indirect effectsdue to changes in ultrastructural organization such as theincrease in the number of chloroplasts per cell [30 58]
The Scientific World Journal 11
Table 2 Cell shape in different tissues of compound leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown(119899 = 75) Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf CL Aspect ratio Sphericity Convexity
Upper epidermis
C 166 plusmn 007 c 0422 plusmn 0042 a 0946 plusmn 0010 a03 Gy 216 plusmn 017 ab 0244 plusmn 0033 c 0925 plusmn 0016 ab10Gy 171 plusmn 011 bc 0401 plusmn 0042 a 0914 plusmn 0013 b20Gy 186 plusmn 010 abc 0328 plusmn 0041 ab 0919 plusmn 0020 ab50Gy 225 plusmn 018 a 0247 plusmn 0041 c 0897 plusmn 0019 b100Gy 188 plusmn 009 abc 0319 plusmn 0045 ab 0902 plusmn 0016 b
Palisade parenchyma
C 355 plusmn 012 a 0073 plusmn 0014 b 0890 plusmn 0012 a03 Gy 289 plusmn 010 b 0102 plusmn 0013 ab 0847 plusmn 0018 b10Gy 300 plusmn 014 b 0104 plusmn 0021 ab 0826 plusmn 0017 b20Gy 302 plusmn 011 b 0094 plusmn 0013 ab 0810 plusmn 0014 b50Gy 283 plusmn 012 b 0107 plusmn 0021 ab 0750 plusmn 0021 c100Gy 275 plusmn 012 b 0122 plusmn 0023 a 0809 plusmn 0015 b
Spongy parenchyma
C 193 plusmn 010 a 0323 plusmn 0035 a 0904 plusmn 0011 a03 Gy 183 plusmn 010 a 0375 plusmn 0042 a 0879 plusmn 0013 a10Gy 176 plusmn 007 a 0357 plusmn 0034 a 0869 plusmn 0011 a20Gy 180 plusmn 009 a 0366 plusmn 0038 a 0885 plusmn 0016 a50Gy 171 plusmn 006 a 0372 plusmn 0034 a 0879 plusmn 0015 a100Gy 188 plusmn 010 a 0339 plusmn 0037 a 0883 plusmn 0015 a
Lower epidermis
C 148 plusmn 008 b 0545 plusmn 0045 a 0957 plusmn 0015 a03 Gy 210 plusmn 017 a 0295 plusmn 0046 b 0929 plusmn 0019 ab10Gy 190 plusmn 012 ab 0334 plusmn 0043 b 0893 plusmn 0019 b20Gy 178 plusmn 011 ab 0388 plusmn 0047 b 0929 plusmn 0017 ab50Gy 172 plusmn 009 ab 0367 plusmn 0040 b 0914 plusmn 0016 b100Gy 191 plusmn 014 ab 0362 plusmn 0054 b 0888 plusmn 0025 b
The lowering of phenolic content at high levels of X-rays inldquoMicrotomrdquo leaves is not in agreement with general trendsindicating enhanced phenolics production in plants grownin Space [58] or in various plant tissues directly irradiatedwith gamma rays [59] However the low content of phenolicscan be an indirect effect of increasing volume occupiedby intercellular spaces due to the loosening of parenchymacells
In conclusion our results show that irradiation of the dryseeds of the Microtom cultivar did not hamper germinationand the development of functional leaves The formationof more compact plants characterised by a higher numberof larger leaves can be valuable in terms of increasingresource regeneration in the BLSSs High levels of radiationinduced only slight structural perturbations which weresimilar in juvenile and compound leaves Such perturbationsdid not affect photosynthetic efficiency since the maximalPSII photochemical efficiency was even increased after irra-diation at all doses However the tendency to accumulatelower amounts of phenolic compounds along chloroplastmembranes after irradiation of seeds with high doses ofradiation should be further investigated because it coulddetermine the reduction of natural photoprotection [57]Finally the overall results indicate that theMicrotom cultivarirradiated at the seed stage has high radioresistance duringthe growth which is a desirable feature for plant cultivation
in Space Radioresistance together with dwarf growth andshort life cycle makes this cultivar a valuable candidate forthe cultivation in BLSSs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are grateful to the ldquoCompagnia di San Paolordquo(Turin) and the ldquoPolo delle Scienze e delle Tecnologie perla Vitardquo of the University of Naples Federico II for thefinancial support The authors also thank Maria Giordanoand Pasquale Scognamiglio for their support in plant culti-vation and microscopy analyses The authors acknowledgefour anonymous editors for helpful comments on a previousversion of the paper
References
[1] A Real S Sundell-Bergman J F Knowles D S Woodheadand I Zinger ldquoEffects of ionising radiation exposure on plantsfish and mammals relevant data for environmental radiation
12 The Scientific World Journal
protectionrdquo Journal of Radiological Protection vol 24 no 4 ppA123ndashA137 2004
[2] V De Micco C Arena D Pignalosa and M Durante ldquoEffectsof sparsely and densely ionizing radiation on plantsrdquo Radiationand Environmental Biophysics vol 50 no 1 pp 1ndash19 2011
[3] N Kumari P Kumar D Mitra B Prasad B N Tiwary andL Varshney ldquoEffects of ionizing radiation on microbial decon-tamination phenolic contents and antioxidant properties oftriphalardquo Journal of Food Science vol 74 no 3 pp M109ndashM1132009
[4] A Tanaka N Shikazono and Y Hase ldquoStudies on biologicaleffects of ion beams on lethality molecular nature of mutationmutation rate and spectrum of mutation phenotype for muta-tion breeding in higher plantsrdquo Journal of Radiation Researchvol 51 no 3 pp 223ndash233 2010
[5] M Durante and A Kronenberg ldquoGround-based research withheavy ions for space radiation protectionrdquo Advances in SpaceResearch vol 35 no 2 pp 180ndash184 2005
[6] E Hu S I Bartsev and H Liu ldquoConceptual design of a biore-generative life support system containing crops and silkwormsrdquoAdvances in Space Research vol 45 no 7 pp 929ndash939 2010
[7] V deMicco G Aronne G Colla R Fortezza and S de PascaleldquoAgro-biology for bioregenerative life support systems in long-term space missions general constraints and the Italian effortsrdquoJournal of Plant Interactions vol 4 no 4 pp 241ndash252 2009
[8] T W Halstead and F R Dutcher ldquoPlants in spacerdquo AnnualReview of Plant Physiology vol 38 pp 317ndash345 1987
[9] V De Micco S De Pascale R Paradiso and G AronneldquoMicrogravity effects on different stages of higher plant life cycleand completion of the seed-to-seed cyclerdquo Plant Biology vol 16pp 31ndash38 2014
[10] C J Bennett C Pirim and T M Orlando ldquoSpace-weatheringof solar system bodies a laboratory perspectiverdquo ChemicalReviews vol 113 pp 9086ndash9150 2013
[11] R A Kerr ldquoRadiation will make astronautsrsquo trip to Mars evenriskierrdquo Science vol 340 article 1031 2014
[12] D M Hassler C Zeitlin R F Wimmer-Schweingruber et alldquoMarsrsquo surface radiation environment measured with the MarsScience Laboratoryrsquos Curiosity Roverrdquo Science vol 343 no6169 Article ID 1244797 2014
[13] M Durante and F A Cucinotta ldquoHeavy ion carcinogenesis andhuman space explorationrdquo Nature Reviews Cancer vol 8 no 6pp 465ndash472 2008
[14] C Arena V De Micco and A De Maio ldquoGrowth alterationand leaf biochemical responses in P vulgaris plants exposed todifferent doses of ionizing radiationrdquo Plant Biology vol 16 pp194ndash202 2014
[15] R W Holst and D J Nagel ldquoRadiation effects on plantsrdquo inPlants For Environmental Studies W Wang J W Gorsuch andJ S Hughes Eds pp 37ndash81 Lewis Publishers Boca Raton FlaUSA 1997
[16] T D Luckey Hormesis With Ionizing Radiation CRC PressBoca Raton Fla USA 1980
[17] D Marcu V Cristea and L Daraban ldquoDose-dependent effectsof gamma radiation on lettuce (Lactuca sativa var capitata)seedlingsrdquo International Journal of Radiation Biology vol 89 pp219ndash223 2013
[18] M Mei H Deng Y Lu et al ldquoMutagenic effects of heavy ionradiation in plantsrdquo Advances in Space Research vol 14 no 10pp 363ndash372 1994
[19] Y LiM Liu Z Cheng andY Sun ldquoSpace environment inducedmutations prefer to occur at polymorphic sites of rice genomesrdquoAdvances in Space Research vol 40 no 4 pp 523ndash527 2007
[20] D S Kim J-B Kim E J Goh et al ldquoAntioxidant response ofArabidopsis plants to gamma irradiation genome-wide expres-sion profiling of the ROS scavenging and signal transductionpathwaysrdquo Journal of Plant Physiology vol 168 no 16 pp 1960ndash1971 2011
[21] T J Brodribb ldquoXylem hydraulic physiology the functionalbackbone of terrestrial plant productivityrdquo Plant Science vol177 no 4 pp 245ndash251 2009
[22] R M Wheeler J C Sager R P Prince andW M Knott ldquoCropproduction for advanced life support systemsmdashobservationfrom the Kennedy Space Center Breadboard Projectrdquo NASATM-2003-211184 NASA Kennedy Space Center Florida 2003
[23] R M Wheeler C L Mackowiak G W Stutte et al ldquoCropproductivities and radiation use efficiencies for bioregenerativelife supportrdquo Advances in Space Research vol 41 no 5 pp 706ndash713 2008
[24] R Paradiso V De Micco R Buonomo G Aronne G Bar-bieri and S De Pascale ldquoSoilless cultivation of soybean forBioregenerative Life-Support Systems a literature review andthe experience of the MELiSSA ProjectmdashFood characterisationPhase Irdquo Plant Biology vol 16 pp 69ndash78 2014
[25] D M Porterfield ldquoThe biophysical limitations in physiologicaltransport and exchange in plants grown in microgravityrdquoJournal of Plant Growth Regulation vol 21 no 2 pp 177ndash1902002
[26] K Yokthongwattana and A Melis ldquoPhotoinhibition and recov-ery in oxygenic photosynthesis mechanism of a photosystem IIdamage and repair cyclerdquo in Photoprotection PhotoinhibitionGene Regulation and Environment B Demmig-Adams WW Adams III and A K Mattoo Eds pp 175ndash191 SpringerDordecht The Netherlands 2006
[27] M T Giardi G Rea M D Lambreva et al ldquoMutations ofphotosystem II D1 protein that empower efficient phenotypesof Chlamydomonas reinhardtii under extreme environment inSpacerdquo PLoS ONE vol 8 Article ID e64352 2013
[28] C Arena V De Micco G Aronne M Pugliese A Virzo DeSanto andADeMaio ldquoResponse ofPhaseolus vulgarisL plantsto low-let ionizing radiation growth and oxidative stressrdquo ActaAstronautica vol 91 pp 107ndash114 2013
[29] V Lattanzio P A Kroon and S Quideau ldquoPlant phenolicsmdashsecondary metabolites with diverse functionsrdquo in RecentAdvances in Polyphenol Research F Daay and V Lattanzio Edspp 1ndash35 Wiley-Blackwell Oxford UK 2008
[30] V De Micco C Arena and G Aronne ldquoAnatomical alterationsof Phaseolus vulgaris L mature leaves irradiated with X-raysrdquoPlant Biology vol 16 pp 187ndash193 2014
[31] A D Krikorian ldquoPlants and somatic embryos in space whathave we learnedrdquo Gravitational and Space Biology Bulletin vol11 no 2 pp 5ndash14 1998
[32] R Meissner Y Jacobson S Melamed et al ldquoA new modelsystem for tomato geneticsrdquoThe Plant Journal vol 12 no 6 pp1465ndash1472 1997
[33] E Emmanuel and A A Levy ldquoTomato mutants as tools forfunctional genomicsrdquo Current Opinion in Plant Biology vol 5no 2 pp 112ndash117 2002
[34] E Martı C Gisbert G J Bishop M S Dixon and J LGarcıa-Martınez ldquoGenetic and physiological characterizationof tomato cv Micro-Tomrdquo Journal of Experimental Botany vol57 pp 2037ndash2047 2006
The Scientific World Journal 13
[35] T Kurimoto J V H Constable and A Huda ldquoEffects ofionizing radiation exposure on arabidopsis thalianardquo HealthPhysics vol 99 no 1 pp 49ndash57 2010
[36] L Reale D Gigante F Landucci F Ferranti and R VenanzonildquoMorphological and histo-anatomical traits reflect die-back inPhragmites australis (Cav) Steudrdquo Aquatic Botany vol 103 pp122ndash128 2012
[37] K Fukuzawa ldquoUltraviolet microscopyrdquo in Methods in LigninChemistry S Y Lin andCWDence Eds pp 110ndash131 SpringerBerlin Germany 1992
[38] S E Ruzin Plant Microtechnique and Microscopy OxfordUniversity Press New York NY USA 1999
[39] V De Micco G Aronne J-P Joseleau and K Ruel ldquoXylemdevelopment and cell wall changes of soybean seedlings grownin spacerdquo Annals of Botany vol 101 no 5 pp 661ndash669 2008
[40] S Van Buggenhout T Grauwet A Van Loey and M Hen-drickx ldquoStructureprocessing relation of vacuum infused straw-berry tissue frozen under different conditionsrdquo European FoodResearch and Technology vol 226 no 3 pp 437ndash448 2008
[41] V De Micco and G Aronne ldquoBiometric anatomy of seedlingsdeveloped onboard of Foton-M2 in an automatic system sup-porting growthrdquoActa Astronautica vol 62 no 8-9 pp 505ndash5132008
[42] B Genty J M Briantais and N R Baker ldquoThe relationshipbetween quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescencerdquo Biochimica andBiophysica Acta vol 990 pp 87ndash92 1989
[43] W Bilger and O Bjorkman ldquoRole of the xanthophyll cycle inphotoprotection elucidated by measurements of light-inducedabsorbance changes fluorescence and photosynthesis in leavesof Hedera canariensisrdquo Photosynthesis Research vol 25 no 3pp 173ndash185 1990
[44] H K Lichtenthaler ldquoChlorophylls and carotenoids pigmentsof photosynthetic biomembranesrdquoMethods in Enzymology vol148 pp 350ndash382 1987
[45] J P Maity DMishra A Chakraborty A Saha S C Santra andS Chanda ldquoModulation of some quantitative and qualitativecharacteristics in rice (Oryza sativa L) and mung (Phaseolusmungo L) by ionizing radiationrdquo Radiation Physics and Chem-istry vol 74 no 5 pp 391ndash394 2005
[46] L Cheng H Yang B Lin et al ldquoEffect of gamma-ray radiationon physiological morphological characters and chromosomeaberrations of minitubers in Solanum tuberosum Lrdquo Interna-tional Journal of Radiation Biology vol 86 no 9 pp 791ndash7992010
[47] B Al-Safadi and R Elias ldquoImprovement of caper (Capparisspinosa L) propagation using in vitro culture and gammairradiationrdquo Scientia Horticulturae vol 127 no 3 pp 290ndash2972011
[48] O Monje G W Stutte G D Goins D M Porterfield and GE Bingham ldquoFarming in space environmental and biophysicalconcernsrdquo Advances in Space Research vol 31 no 1 pp 151ndash1672003
[49] V De Micco M Scala and G Aronne ldquoEffects of simulatedmicrogravity on male gametophyte of Prunus Pyrus andBrassica speciesrdquo Protoplasma vol 228 no 1ndash3 pp 121ndash1262006
[50] V De Micco R Buonomo R Paradiso S De Pascale and GAronne ldquoSoybean cultivar selection for Bioregenerative LifeSupport Systems (BLSS)mdashtheoretical selectionrdquo Advances inSpace Research vol 49 no 10 pp 1415ndash1421 2012
[51] T Kozuka S-G Kong M Doi K-I Shimazaki and ANagatani ldquoTissue-autonomous promotion of palisade celldevelopment by phototropin 2 in Arabidopsisrdquo Plant Cell vol23 no 10 pp 3684ndash3695 2011
[52] E Kovacs and A Keresztes ldquoEffect of gamma and UV-BCradiation on plant cellsrdquoMicron vol 33 no 2 pp 199ndash210 2002
[53] R Ogaya J Penuelas D Asensio and J Llusia ldquoChlorophyllfluorescence responses to temperature and water availabilityin two co-dominant Mediterranean shrub and tree speciesin a long-term field experiment simulating climate changerdquoEnvironmental and Experimental Botany vol 73 no 1 pp 89ndash93 2011
[54] R Zaka C M Vandecasteele and M T Misset ldquoEffects oflow chronic doses of ionizing radiation on antioxidant enzymesand G6PDH activities in Stipa capillata (Poaceae)rdquo Journal ofExperimental Botany vol 53 no 376 pp 1979ndash1987 2002
[55] J-H KimM-H Baek Y C Byung GW Seung and J-S KimldquoAlterations in the photosynthetic pigments and antioxidantmachineries of red pepper (Capsicum annuum L) seedlingsfrom gamma-irradiated seedsrdquo Journal of Plant Biology vol 47no 4 pp 314ndash321 2004
[56] N A Al-Enezi and J M Al-Khayri ldquoAlterations of DNA ionsand photosynthetic pigments content in date palm seedlingsinduced by X-irradiationrdquo International Journal of Agricultureand Biology vol 14 pp 329ndash336 2012
[57] V de Micco C Arena L Vitale G Aronne and A V de SantoldquoAnatomy and photochemical behaviour of mediterranean Cis-tus incanus winter leaves under natural outdoor and warmerindoor conditionsrdquo Botany vol 89 no 10 pp 677ndash688 2011
[58] L H Levine H G Levine E C Stryjewski V Prima and WC Piastuch ldquoEffect of spaceflight on isoflavonoid accumulationin etiolated soybean seedlingsrdquo Journal of Gravitational Physiol-ogy vol 8 no 2 pp 21ndash27 2001
[59] X Fan ldquoAntioxidant capacity of fresh-cut vegetables exposedto ionizing radiationrdquo Journal of the Science of Food andAgriculture vol 85 no 6 pp 995ndash1000 2005
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

10 The Scientific World Journal
Table 1 Cell shape in different tissues of juvenile leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown (119899 = 75)Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf JL Aspect ratio Sphericity Convexity
Upper epidermis
C 197 plusmn 016 a 0308 plusmn 0050 a 0926 plusmn 0021 ab03Gy 200 plusmn 013 a 0290 plusmn 0038 a 0941 plusmn 0013 a10Gy 210 plusmn 012 a 0239 plusmn 0037 a 0898 plusmn 0016 ab20Gy 217 plusmn 014 a 0258 plusmn 0045 a 0909 plusmn 0019 abc50Gy 170 plusmn 008 a 0376 plusmn 0037 a 0927 plusmn 0016 ab100Gy 197 plusmn 014 a 0286 plusmn 0042 a 0876 plusmn 0028 c
Palisade parenchyma
C 290 plusmn 010 a 0109 plusmn 0015 a 0879 plusmn 0010 a03 Gy 304 plusmn 012 a 0099 plusmn 0016 a 0805 plusmn 0016 b10Gy 309 plusmn 013 a 0093 plusmn 0019 a 0792 plusmn 0015 b20Gy 283 plusmn 011 a 0101 plusmn 0017 a 0784 plusmn 0017 b50Gy 314 plusmn 014 a 0082 plusmn 0016 a 0772 plusmn 0016 b100Gy 275 plusmn 011 a 0119 plusmn 0020 a 0805 plusmn 0017 b
Spongy parenchyma
C 190 plusmn 008 a 0307 plusmn 0033 a 0893 plusmn 0012 a03 Gy 180 plusmn 007 a 0341 plusmn 0029 a 0876 plusmn 0013 a10Gy 203 plusmn 010 a 0290 plusmn 0031 a 0901 plusmn 0012 a20Gy 194 plusmn 012 a 0344 plusmn 0040 a 0881 plusmn 0015 a50Gy 174 plusmn 009 a 0393 plusmn 0035 a 0883 plusmn 0014 a100Gy 198 plusmn 012 a 0322 plusmn 0037 a 0878 plusmn 0015 a
Lower epidermis
C 178 plusmn 009 bc 0343 plusmn 0045 ab 0919 plusmn 0019 a03 Gy 225 plusmn 017 ab 0263 plusmn 0044 b 0933 plusmn 0024 a10Gy 219 plusmn 011 abc 0218 plusmn 0034 b 0888 plusmn 0020 ab20Gy 180 plusmn 013 bc 0378 plusmn 0047 ab 0926 plusmn 0017 a50Gy 167 plusmn 001 c 0461 plusmn 0051 a 0924 plusmn 0021 a100Gy 238 plusmn 021 a 0257 plusmn 0057 b 0857 plusmn 0030 b
the various tissues of structures of different age is in agree-ment with the different light-mediated control of cell devel-opment in the various leaf tissues [51] Altered mesophyllcharacterized by partly collapsed cells with irregular cellshape was found only sporadically after the exposure of seedsto 50 and 100Gy in theMicrotomcultivar Suchmodificationswere less marked than those found in leaves of bean wherethe target organs for irradiation were the leaves themselves[30] leaves directly exposed to X-rays in bean showed theformation of more loose parenchyma with larger cells likelydue to a radiation-induced damage to cell walls that reducedmechanical constraints to cell enlargement [30 52] Furtherinvestigations on cell wall composition would be useful toelucidate whether a possible alteration of cell walls wasresponsible for cell enlargement in the spongy parenchymaand epidermis of leaves from seeds of ldquoMicrotomrdquo irradiatedwith the highest dose of X-rays However in the cultivarMicrotom the slight alterations of mesophyll structure didnot impair photosynthetic efficiency as demonstrated by thelack of significant dose-dependent variation trends in thequantum yield of PSII electron transport (ΦPSII) nonphoto-chemical quenching (NPQ) and maximal PSII photochemi-cal efficiency (FvFm) The increased values of FvFm closeto 08 which is considered an optimal value for healthy plants[53] suggested that the photosynthetic apparatus of the leavesfrom irradiated seeds has higher levels of photochemical
efficiency irrespective of dose The unchanged or increasedcontent of both chlorophyll and carotenoid pigments con-firmed that the photosynthetic apparatus remained stablealso in terms of light harvesting capacity and indicatedno deleterious effect of radiation on plant photosyntheticmachinery RecentlyMarcu et al [17] showed that irradiationof lettuce seeds with gamma rays up to 30Gy enhancedthe content of chlorophyll a chlorophyll b and carotenoidswhile higher doses (up to 70Gy) resulted in their decreaseHowever the effect of ionising radiation on photosyntheticpigment content is quite controversial Increased pigmentcontent is ascribed to the phenomenon of hormesis whilea lower content is generally ascribed to the inhibition ofchlorophyll and carotenoid synthesis or to oxidation due tothe high levels of radiation-induced water radiolysis [2 1454ndash56] Some plants possess natural screens against radiationwhen their tissues contain phenolic compounds with antioxi-dant functions and can counteract photoinhibitory processes[29 57] An increase of phenolic compounds linked to thechloroplast membranes has been recently found in Phaseolusvulgaris irradiated leaves exposed to high levels of X-rays[28 30] Changes in phenolic content after the exposure toSpace can be attributed either to direct effects on the synthesisor mobilization of phenolic compounds or to indirect effectsdue to changes in ultrastructural organization such as theincrease in the number of chloroplasts per cell [30 58]
The Scientific World Journal 11
Table 2 Cell shape in different tissues of compound leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown(119899 = 75) Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf CL Aspect ratio Sphericity Convexity
Upper epidermis
C 166 plusmn 007 c 0422 plusmn 0042 a 0946 plusmn 0010 a03 Gy 216 plusmn 017 ab 0244 plusmn 0033 c 0925 plusmn 0016 ab10Gy 171 plusmn 011 bc 0401 plusmn 0042 a 0914 plusmn 0013 b20Gy 186 plusmn 010 abc 0328 plusmn 0041 ab 0919 plusmn 0020 ab50Gy 225 plusmn 018 a 0247 plusmn 0041 c 0897 plusmn 0019 b100Gy 188 plusmn 009 abc 0319 plusmn 0045 ab 0902 plusmn 0016 b
Palisade parenchyma
C 355 plusmn 012 a 0073 plusmn 0014 b 0890 plusmn 0012 a03 Gy 289 plusmn 010 b 0102 plusmn 0013 ab 0847 plusmn 0018 b10Gy 300 plusmn 014 b 0104 plusmn 0021 ab 0826 plusmn 0017 b20Gy 302 plusmn 011 b 0094 plusmn 0013 ab 0810 plusmn 0014 b50Gy 283 plusmn 012 b 0107 plusmn 0021 ab 0750 plusmn 0021 c100Gy 275 plusmn 012 b 0122 plusmn 0023 a 0809 plusmn 0015 b
Spongy parenchyma
C 193 plusmn 010 a 0323 plusmn 0035 a 0904 plusmn 0011 a03 Gy 183 plusmn 010 a 0375 plusmn 0042 a 0879 plusmn 0013 a10Gy 176 plusmn 007 a 0357 plusmn 0034 a 0869 plusmn 0011 a20Gy 180 plusmn 009 a 0366 plusmn 0038 a 0885 plusmn 0016 a50Gy 171 plusmn 006 a 0372 plusmn 0034 a 0879 plusmn 0015 a100Gy 188 plusmn 010 a 0339 plusmn 0037 a 0883 plusmn 0015 a
Lower epidermis
C 148 plusmn 008 b 0545 plusmn 0045 a 0957 plusmn 0015 a03 Gy 210 plusmn 017 a 0295 plusmn 0046 b 0929 plusmn 0019 ab10Gy 190 plusmn 012 ab 0334 plusmn 0043 b 0893 plusmn 0019 b20Gy 178 plusmn 011 ab 0388 plusmn 0047 b 0929 plusmn 0017 ab50Gy 172 plusmn 009 ab 0367 plusmn 0040 b 0914 plusmn 0016 b100Gy 191 plusmn 014 ab 0362 plusmn 0054 b 0888 plusmn 0025 b
The lowering of phenolic content at high levels of X-rays inldquoMicrotomrdquo leaves is not in agreement with general trendsindicating enhanced phenolics production in plants grownin Space [58] or in various plant tissues directly irradiatedwith gamma rays [59] However the low content of phenolicscan be an indirect effect of increasing volume occupiedby intercellular spaces due to the loosening of parenchymacells
In conclusion our results show that irradiation of the dryseeds of the Microtom cultivar did not hamper germinationand the development of functional leaves The formationof more compact plants characterised by a higher numberof larger leaves can be valuable in terms of increasingresource regeneration in the BLSSs High levels of radiationinduced only slight structural perturbations which weresimilar in juvenile and compound leaves Such perturbationsdid not affect photosynthetic efficiency since the maximalPSII photochemical efficiency was even increased after irra-diation at all doses However the tendency to accumulatelower amounts of phenolic compounds along chloroplastmembranes after irradiation of seeds with high doses ofradiation should be further investigated because it coulddetermine the reduction of natural photoprotection [57]Finally the overall results indicate that theMicrotom cultivarirradiated at the seed stage has high radioresistance duringthe growth which is a desirable feature for plant cultivation
in Space Radioresistance together with dwarf growth andshort life cycle makes this cultivar a valuable candidate forthe cultivation in BLSSs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are grateful to the ldquoCompagnia di San Paolordquo(Turin) and the ldquoPolo delle Scienze e delle Tecnologie perla Vitardquo of the University of Naples Federico II for thefinancial support The authors also thank Maria Giordanoand Pasquale Scognamiglio for their support in plant culti-vation and microscopy analyses The authors acknowledgefour anonymous editors for helpful comments on a previousversion of the paper
References
[1] A Real S Sundell-Bergman J F Knowles D S Woodheadand I Zinger ldquoEffects of ionising radiation exposure on plantsfish and mammals relevant data for environmental radiation
12 The Scientific World Journal
protectionrdquo Journal of Radiological Protection vol 24 no 4 ppA123ndashA137 2004
[2] V De Micco C Arena D Pignalosa and M Durante ldquoEffectsof sparsely and densely ionizing radiation on plantsrdquo Radiationand Environmental Biophysics vol 50 no 1 pp 1ndash19 2011
[3] N Kumari P Kumar D Mitra B Prasad B N Tiwary andL Varshney ldquoEffects of ionizing radiation on microbial decon-tamination phenolic contents and antioxidant properties oftriphalardquo Journal of Food Science vol 74 no 3 pp M109ndashM1132009
[4] A Tanaka N Shikazono and Y Hase ldquoStudies on biologicaleffects of ion beams on lethality molecular nature of mutationmutation rate and spectrum of mutation phenotype for muta-tion breeding in higher plantsrdquo Journal of Radiation Researchvol 51 no 3 pp 223ndash233 2010
[5] M Durante and A Kronenberg ldquoGround-based research withheavy ions for space radiation protectionrdquo Advances in SpaceResearch vol 35 no 2 pp 180ndash184 2005
[6] E Hu S I Bartsev and H Liu ldquoConceptual design of a biore-generative life support system containing crops and silkwormsrdquoAdvances in Space Research vol 45 no 7 pp 929ndash939 2010
[7] V deMicco G Aronne G Colla R Fortezza and S de PascaleldquoAgro-biology for bioregenerative life support systems in long-term space missions general constraints and the Italian effortsrdquoJournal of Plant Interactions vol 4 no 4 pp 241ndash252 2009
[8] T W Halstead and F R Dutcher ldquoPlants in spacerdquo AnnualReview of Plant Physiology vol 38 pp 317ndash345 1987
[9] V De Micco S De Pascale R Paradiso and G AronneldquoMicrogravity effects on different stages of higher plant life cycleand completion of the seed-to-seed cyclerdquo Plant Biology vol 16pp 31ndash38 2014
[10] C J Bennett C Pirim and T M Orlando ldquoSpace-weatheringof solar system bodies a laboratory perspectiverdquo ChemicalReviews vol 113 pp 9086ndash9150 2013
[11] R A Kerr ldquoRadiation will make astronautsrsquo trip to Mars evenriskierrdquo Science vol 340 article 1031 2014
[12] D M Hassler C Zeitlin R F Wimmer-Schweingruber et alldquoMarsrsquo surface radiation environment measured with the MarsScience Laboratoryrsquos Curiosity Roverrdquo Science vol 343 no6169 Article ID 1244797 2014
[13] M Durante and F A Cucinotta ldquoHeavy ion carcinogenesis andhuman space explorationrdquo Nature Reviews Cancer vol 8 no 6pp 465ndash472 2008
[14] C Arena V De Micco and A De Maio ldquoGrowth alterationand leaf biochemical responses in P vulgaris plants exposed todifferent doses of ionizing radiationrdquo Plant Biology vol 16 pp194ndash202 2014
[15] R W Holst and D J Nagel ldquoRadiation effects on plantsrdquo inPlants For Environmental Studies W Wang J W Gorsuch andJ S Hughes Eds pp 37ndash81 Lewis Publishers Boca Raton FlaUSA 1997
[16] T D Luckey Hormesis With Ionizing Radiation CRC PressBoca Raton Fla USA 1980
[17] D Marcu V Cristea and L Daraban ldquoDose-dependent effectsof gamma radiation on lettuce (Lactuca sativa var capitata)seedlingsrdquo International Journal of Radiation Biology vol 89 pp219ndash223 2013
[18] M Mei H Deng Y Lu et al ldquoMutagenic effects of heavy ionradiation in plantsrdquo Advances in Space Research vol 14 no 10pp 363ndash372 1994
[19] Y LiM Liu Z Cheng andY Sun ldquoSpace environment inducedmutations prefer to occur at polymorphic sites of rice genomesrdquoAdvances in Space Research vol 40 no 4 pp 523ndash527 2007
[20] D S Kim J-B Kim E J Goh et al ldquoAntioxidant response ofArabidopsis plants to gamma irradiation genome-wide expres-sion profiling of the ROS scavenging and signal transductionpathwaysrdquo Journal of Plant Physiology vol 168 no 16 pp 1960ndash1971 2011
[21] T J Brodribb ldquoXylem hydraulic physiology the functionalbackbone of terrestrial plant productivityrdquo Plant Science vol177 no 4 pp 245ndash251 2009
[22] R M Wheeler J C Sager R P Prince andW M Knott ldquoCropproduction for advanced life support systemsmdashobservationfrom the Kennedy Space Center Breadboard Projectrdquo NASATM-2003-211184 NASA Kennedy Space Center Florida 2003
[23] R M Wheeler C L Mackowiak G W Stutte et al ldquoCropproductivities and radiation use efficiencies for bioregenerativelife supportrdquo Advances in Space Research vol 41 no 5 pp 706ndash713 2008
[24] R Paradiso V De Micco R Buonomo G Aronne G Bar-bieri and S De Pascale ldquoSoilless cultivation of soybean forBioregenerative Life-Support Systems a literature review andthe experience of the MELiSSA ProjectmdashFood characterisationPhase Irdquo Plant Biology vol 16 pp 69ndash78 2014
[25] D M Porterfield ldquoThe biophysical limitations in physiologicaltransport and exchange in plants grown in microgravityrdquoJournal of Plant Growth Regulation vol 21 no 2 pp 177ndash1902002
[26] K Yokthongwattana and A Melis ldquoPhotoinhibition and recov-ery in oxygenic photosynthesis mechanism of a photosystem IIdamage and repair cyclerdquo in Photoprotection PhotoinhibitionGene Regulation and Environment B Demmig-Adams WW Adams III and A K Mattoo Eds pp 175ndash191 SpringerDordecht The Netherlands 2006
[27] M T Giardi G Rea M D Lambreva et al ldquoMutations ofphotosystem II D1 protein that empower efficient phenotypesof Chlamydomonas reinhardtii under extreme environment inSpacerdquo PLoS ONE vol 8 Article ID e64352 2013
[28] C Arena V De Micco G Aronne M Pugliese A Virzo DeSanto andADeMaio ldquoResponse ofPhaseolus vulgarisL plantsto low-let ionizing radiation growth and oxidative stressrdquo ActaAstronautica vol 91 pp 107ndash114 2013
[29] V Lattanzio P A Kroon and S Quideau ldquoPlant phenolicsmdashsecondary metabolites with diverse functionsrdquo in RecentAdvances in Polyphenol Research F Daay and V Lattanzio Edspp 1ndash35 Wiley-Blackwell Oxford UK 2008
[30] V De Micco C Arena and G Aronne ldquoAnatomical alterationsof Phaseolus vulgaris L mature leaves irradiated with X-raysrdquoPlant Biology vol 16 pp 187ndash193 2014
[31] A D Krikorian ldquoPlants and somatic embryos in space whathave we learnedrdquo Gravitational and Space Biology Bulletin vol11 no 2 pp 5ndash14 1998
[32] R Meissner Y Jacobson S Melamed et al ldquoA new modelsystem for tomato geneticsrdquoThe Plant Journal vol 12 no 6 pp1465ndash1472 1997
[33] E Emmanuel and A A Levy ldquoTomato mutants as tools forfunctional genomicsrdquo Current Opinion in Plant Biology vol 5no 2 pp 112ndash117 2002
[34] E Martı C Gisbert G J Bishop M S Dixon and J LGarcıa-Martınez ldquoGenetic and physiological characterizationof tomato cv Micro-Tomrdquo Journal of Experimental Botany vol57 pp 2037ndash2047 2006
The Scientific World Journal 13
[35] T Kurimoto J V H Constable and A Huda ldquoEffects ofionizing radiation exposure on arabidopsis thalianardquo HealthPhysics vol 99 no 1 pp 49ndash57 2010
[36] L Reale D Gigante F Landucci F Ferranti and R VenanzonildquoMorphological and histo-anatomical traits reflect die-back inPhragmites australis (Cav) Steudrdquo Aquatic Botany vol 103 pp122ndash128 2012
[37] K Fukuzawa ldquoUltraviolet microscopyrdquo in Methods in LigninChemistry S Y Lin andCWDence Eds pp 110ndash131 SpringerBerlin Germany 1992
[38] S E Ruzin Plant Microtechnique and Microscopy OxfordUniversity Press New York NY USA 1999
[39] V De Micco G Aronne J-P Joseleau and K Ruel ldquoXylemdevelopment and cell wall changes of soybean seedlings grownin spacerdquo Annals of Botany vol 101 no 5 pp 661ndash669 2008
[40] S Van Buggenhout T Grauwet A Van Loey and M Hen-drickx ldquoStructureprocessing relation of vacuum infused straw-berry tissue frozen under different conditionsrdquo European FoodResearch and Technology vol 226 no 3 pp 437ndash448 2008
[41] V De Micco and G Aronne ldquoBiometric anatomy of seedlingsdeveloped onboard of Foton-M2 in an automatic system sup-porting growthrdquoActa Astronautica vol 62 no 8-9 pp 505ndash5132008
[42] B Genty J M Briantais and N R Baker ldquoThe relationshipbetween quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescencerdquo Biochimica andBiophysica Acta vol 990 pp 87ndash92 1989
[43] W Bilger and O Bjorkman ldquoRole of the xanthophyll cycle inphotoprotection elucidated by measurements of light-inducedabsorbance changes fluorescence and photosynthesis in leavesof Hedera canariensisrdquo Photosynthesis Research vol 25 no 3pp 173ndash185 1990
[44] H K Lichtenthaler ldquoChlorophylls and carotenoids pigmentsof photosynthetic biomembranesrdquoMethods in Enzymology vol148 pp 350ndash382 1987
[45] J P Maity DMishra A Chakraborty A Saha S C Santra andS Chanda ldquoModulation of some quantitative and qualitativecharacteristics in rice (Oryza sativa L) and mung (Phaseolusmungo L) by ionizing radiationrdquo Radiation Physics and Chem-istry vol 74 no 5 pp 391ndash394 2005
[46] L Cheng H Yang B Lin et al ldquoEffect of gamma-ray radiationon physiological morphological characters and chromosomeaberrations of minitubers in Solanum tuberosum Lrdquo Interna-tional Journal of Radiation Biology vol 86 no 9 pp 791ndash7992010
[47] B Al-Safadi and R Elias ldquoImprovement of caper (Capparisspinosa L) propagation using in vitro culture and gammairradiationrdquo Scientia Horticulturae vol 127 no 3 pp 290ndash2972011
[48] O Monje G W Stutte G D Goins D M Porterfield and GE Bingham ldquoFarming in space environmental and biophysicalconcernsrdquo Advances in Space Research vol 31 no 1 pp 151ndash1672003
[49] V De Micco M Scala and G Aronne ldquoEffects of simulatedmicrogravity on male gametophyte of Prunus Pyrus andBrassica speciesrdquo Protoplasma vol 228 no 1ndash3 pp 121ndash1262006
[50] V De Micco R Buonomo R Paradiso S De Pascale and GAronne ldquoSoybean cultivar selection for Bioregenerative LifeSupport Systems (BLSS)mdashtheoretical selectionrdquo Advances inSpace Research vol 49 no 10 pp 1415ndash1421 2012
[51] T Kozuka S-G Kong M Doi K-I Shimazaki and ANagatani ldquoTissue-autonomous promotion of palisade celldevelopment by phototropin 2 in Arabidopsisrdquo Plant Cell vol23 no 10 pp 3684ndash3695 2011
[52] E Kovacs and A Keresztes ldquoEffect of gamma and UV-BCradiation on plant cellsrdquoMicron vol 33 no 2 pp 199ndash210 2002
[53] R Ogaya J Penuelas D Asensio and J Llusia ldquoChlorophyllfluorescence responses to temperature and water availabilityin two co-dominant Mediterranean shrub and tree speciesin a long-term field experiment simulating climate changerdquoEnvironmental and Experimental Botany vol 73 no 1 pp 89ndash93 2011
[54] R Zaka C M Vandecasteele and M T Misset ldquoEffects oflow chronic doses of ionizing radiation on antioxidant enzymesand G6PDH activities in Stipa capillata (Poaceae)rdquo Journal ofExperimental Botany vol 53 no 376 pp 1979ndash1987 2002
[55] J-H KimM-H Baek Y C Byung GW Seung and J-S KimldquoAlterations in the photosynthetic pigments and antioxidantmachineries of red pepper (Capsicum annuum L) seedlingsfrom gamma-irradiated seedsrdquo Journal of Plant Biology vol 47no 4 pp 314ndash321 2004
[56] N A Al-Enezi and J M Al-Khayri ldquoAlterations of DNA ionsand photosynthetic pigments content in date palm seedlingsinduced by X-irradiationrdquo International Journal of Agricultureand Biology vol 14 pp 329ndash336 2012
[57] V de Micco C Arena L Vitale G Aronne and A V de SantoldquoAnatomy and photochemical behaviour of mediterranean Cis-tus incanus winter leaves under natural outdoor and warmerindoor conditionsrdquo Botany vol 89 no 10 pp 677ndash688 2011
[58] L H Levine H G Levine E C Stryjewski V Prima and WC Piastuch ldquoEffect of spaceflight on isoflavonoid accumulationin etiolated soybean seedlingsrdquo Journal of Gravitational Physiol-ogy vol 8 no 2 pp 21ndash27 2001
[59] X Fan ldquoAntioxidant capacity of fresh-cut vegetables exposedto ionizing radiationrdquo Journal of the Science of Food andAgriculture vol 85 no 6 pp 995ndash1000 2005
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

The Scientific World Journal 11
Table 2 Cell shape in different tissues of compound leaves of S lycopersicum L ldquoMicrotomrdquo Mean values plusmn standard errors are shown(119899 = 75) Different letters correspond to significantly different values according to multiple comparison tests (119875 lt 005)
Leaf CL Aspect ratio Sphericity Convexity
Upper epidermis
C 166 plusmn 007 c 0422 plusmn 0042 a 0946 plusmn 0010 a03 Gy 216 plusmn 017 ab 0244 plusmn 0033 c 0925 plusmn 0016 ab10Gy 171 plusmn 011 bc 0401 plusmn 0042 a 0914 plusmn 0013 b20Gy 186 plusmn 010 abc 0328 plusmn 0041 ab 0919 plusmn 0020 ab50Gy 225 plusmn 018 a 0247 plusmn 0041 c 0897 plusmn 0019 b100Gy 188 plusmn 009 abc 0319 plusmn 0045 ab 0902 plusmn 0016 b
Palisade parenchyma
C 355 plusmn 012 a 0073 plusmn 0014 b 0890 plusmn 0012 a03 Gy 289 plusmn 010 b 0102 plusmn 0013 ab 0847 plusmn 0018 b10Gy 300 plusmn 014 b 0104 plusmn 0021 ab 0826 plusmn 0017 b20Gy 302 plusmn 011 b 0094 plusmn 0013 ab 0810 plusmn 0014 b50Gy 283 plusmn 012 b 0107 plusmn 0021 ab 0750 plusmn 0021 c100Gy 275 plusmn 012 b 0122 plusmn 0023 a 0809 plusmn 0015 b
Spongy parenchyma
C 193 plusmn 010 a 0323 plusmn 0035 a 0904 plusmn 0011 a03 Gy 183 plusmn 010 a 0375 plusmn 0042 a 0879 plusmn 0013 a10Gy 176 plusmn 007 a 0357 plusmn 0034 a 0869 plusmn 0011 a20Gy 180 plusmn 009 a 0366 plusmn 0038 a 0885 plusmn 0016 a50Gy 171 plusmn 006 a 0372 plusmn 0034 a 0879 plusmn 0015 a100Gy 188 plusmn 010 a 0339 plusmn 0037 a 0883 plusmn 0015 a
Lower epidermis
C 148 plusmn 008 b 0545 plusmn 0045 a 0957 plusmn 0015 a03 Gy 210 plusmn 017 a 0295 plusmn 0046 b 0929 plusmn 0019 ab10Gy 190 plusmn 012 ab 0334 plusmn 0043 b 0893 plusmn 0019 b20Gy 178 plusmn 011 ab 0388 plusmn 0047 b 0929 plusmn 0017 ab50Gy 172 plusmn 009 ab 0367 plusmn 0040 b 0914 plusmn 0016 b100Gy 191 plusmn 014 ab 0362 plusmn 0054 b 0888 plusmn 0025 b
The lowering of phenolic content at high levels of X-rays inldquoMicrotomrdquo leaves is not in agreement with general trendsindicating enhanced phenolics production in plants grownin Space [58] or in various plant tissues directly irradiatedwith gamma rays [59] However the low content of phenolicscan be an indirect effect of increasing volume occupiedby intercellular spaces due to the loosening of parenchymacells
In conclusion our results show that irradiation of the dryseeds of the Microtom cultivar did not hamper germinationand the development of functional leaves The formationof more compact plants characterised by a higher numberof larger leaves can be valuable in terms of increasingresource regeneration in the BLSSs High levels of radiationinduced only slight structural perturbations which weresimilar in juvenile and compound leaves Such perturbationsdid not affect photosynthetic efficiency since the maximalPSII photochemical efficiency was even increased after irra-diation at all doses However the tendency to accumulatelower amounts of phenolic compounds along chloroplastmembranes after irradiation of seeds with high doses ofradiation should be further investigated because it coulddetermine the reduction of natural photoprotection [57]Finally the overall results indicate that theMicrotom cultivarirradiated at the seed stage has high radioresistance duringthe growth which is a desirable feature for plant cultivation
in Space Radioresistance together with dwarf growth andshort life cycle makes this cultivar a valuable candidate forthe cultivation in BLSSs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors are grateful to the ldquoCompagnia di San Paolordquo(Turin) and the ldquoPolo delle Scienze e delle Tecnologie perla Vitardquo of the University of Naples Federico II for thefinancial support The authors also thank Maria Giordanoand Pasquale Scognamiglio for their support in plant culti-vation and microscopy analyses The authors acknowledgefour anonymous editors for helpful comments on a previousversion of the paper
References
[1] A Real S Sundell-Bergman J F Knowles D S Woodheadand I Zinger ldquoEffects of ionising radiation exposure on plantsfish and mammals relevant data for environmental radiation
12 The Scientific World Journal
protectionrdquo Journal of Radiological Protection vol 24 no 4 ppA123ndashA137 2004
[2] V De Micco C Arena D Pignalosa and M Durante ldquoEffectsof sparsely and densely ionizing radiation on plantsrdquo Radiationand Environmental Biophysics vol 50 no 1 pp 1ndash19 2011
[3] N Kumari P Kumar D Mitra B Prasad B N Tiwary andL Varshney ldquoEffects of ionizing radiation on microbial decon-tamination phenolic contents and antioxidant properties oftriphalardquo Journal of Food Science vol 74 no 3 pp M109ndashM1132009
[4] A Tanaka N Shikazono and Y Hase ldquoStudies on biologicaleffects of ion beams on lethality molecular nature of mutationmutation rate and spectrum of mutation phenotype for muta-tion breeding in higher plantsrdquo Journal of Radiation Researchvol 51 no 3 pp 223ndash233 2010
[5] M Durante and A Kronenberg ldquoGround-based research withheavy ions for space radiation protectionrdquo Advances in SpaceResearch vol 35 no 2 pp 180ndash184 2005
[6] E Hu S I Bartsev and H Liu ldquoConceptual design of a biore-generative life support system containing crops and silkwormsrdquoAdvances in Space Research vol 45 no 7 pp 929ndash939 2010
[7] V deMicco G Aronne G Colla R Fortezza and S de PascaleldquoAgro-biology for bioregenerative life support systems in long-term space missions general constraints and the Italian effortsrdquoJournal of Plant Interactions vol 4 no 4 pp 241ndash252 2009
[8] T W Halstead and F R Dutcher ldquoPlants in spacerdquo AnnualReview of Plant Physiology vol 38 pp 317ndash345 1987
[9] V De Micco S De Pascale R Paradiso and G AronneldquoMicrogravity effects on different stages of higher plant life cycleand completion of the seed-to-seed cyclerdquo Plant Biology vol 16pp 31ndash38 2014
[10] C J Bennett C Pirim and T M Orlando ldquoSpace-weatheringof solar system bodies a laboratory perspectiverdquo ChemicalReviews vol 113 pp 9086ndash9150 2013
[11] R A Kerr ldquoRadiation will make astronautsrsquo trip to Mars evenriskierrdquo Science vol 340 article 1031 2014
[12] D M Hassler C Zeitlin R F Wimmer-Schweingruber et alldquoMarsrsquo surface radiation environment measured with the MarsScience Laboratoryrsquos Curiosity Roverrdquo Science vol 343 no6169 Article ID 1244797 2014
[13] M Durante and F A Cucinotta ldquoHeavy ion carcinogenesis andhuman space explorationrdquo Nature Reviews Cancer vol 8 no 6pp 465ndash472 2008
[14] C Arena V De Micco and A De Maio ldquoGrowth alterationand leaf biochemical responses in P vulgaris plants exposed todifferent doses of ionizing radiationrdquo Plant Biology vol 16 pp194ndash202 2014
[15] R W Holst and D J Nagel ldquoRadiation effects on plantsrdquo inPlants For Environmental Studies W Wang J W Gorsuch andJ S Hughes Eds pp 37ndash81 Lewis Publishers Boca Raton FlaUSA 1997
[16] T D Luckey Hormesis With Ionizing Radiation CRC PressBoca Raton Fla USA 1980
[17] D Marcu V Cristea and L Daraban ldquoDose-dependent effectsof gamma radiation on lettuce (Lactuca sativa var capitata)seedlingsrdquo International Journal of Radiation Biology vol 89 pp219ndash223 2013
[18] M Mei H Deng Y Lu et al ldquoMutagenic effects of heavy ionradiation in plantsrdquo Advances in Space Research vol 14 no 10pp 363ndash372 1994
[19] Y LiM Liu Z Cheng andY Sun ldquoSpace environment inducedmutations prefer to occur at polymorphic sites of rice genomesrdquoAdvances in Space Research vol 40 no 4 pp 523ndash527 2007
[20] D S Kim J-B Kim E J Goh et al ldquoAntioxidant response ofArabidopsis plants to gamma irradiation genome-wide expres-sion profiling of the ROS scavenging and signal transductionpathwaysrdquo Journal of Plant Physiology vol 168 no 16 pp 1960ndash1971 2011
[21] T J Brodribb ldquoXylem hydraulic physiology the functionalbackbone of terrestrial plant productivityrdquo Plant Science vol177 no 4 pp 245ndash251 2009
[22] R M Wheeler J C Sager R P Prince andW M Knott ldquoCropproduction for advanced life support systemsmdashobservationfrom the Kennedy Space Center Breadboard Projectrdquo NASATM-2003-211184 NASA Kennedy Space Center Florida 2003
[23] R M Wheeler C L Mackowiak G W Stutte et al ldquoCropproductivities and radiation use efficiencies for bioregenerativelife supportrdquo Advances in Space Research vol 41 no 5 pp 706ndash713 2008
[24] R Paradiso V De Micco R Buonomo G Aronne G Bar-bieri and S De Pascale ldquoSoilless cultivation of soybean forBioregenerative Life-Support Systems a literature review andthe experience of the MELiSSA ProjectmdashFood characterisationPhase Irdquo Plant Biology vol 16 pp 69ndash78 2014
[25] D M Porterfield ldquoThe biophysical limitations in physiologicaltransport and exchange in plants grown in microgravityrdquoJournal of Plant Growth Regulation vol 21 no 2 pp 177ndash1902002
[26] K Yokthongwattana and A Melis ldquoPhotoinhibition and recov-ery in oxygenic photosynthesis mechanism of a photosystem IIdamage and repair cyclerdquo in Photoprotection PhotoinhibitionGene Regulation and Environment B Demmig-Adams WW Adams III and A K Mattoo Eds pp 175ndash191 SpringerDordecht The Netherlands 2006
[27] M T Giardi G Rea M D Lambreva et al ldquoMutations ofphotosystem II D1 protein that empower efficient phenotypesof Chlamydomonas reinhardtii under extreme environment inSpacerdquo PLoS ONE vol 8 Article ID e64352 2013
[28] C Arena V De Micco G Aronne M Pugliese A Virzo DeSanto andADeMaio ldquoResponse ofPhaseolus vulgarisL plantsto low-let ionizing radiation growth and oxidative stressrdquo ActaAstronautica vol 91 pp 107ndash114 2013
[29] V Lattanzio P A Kroon and S Quideau ldquoPlant phenolicsmdashsecondary metabolites with diverse functionsrdquo in RecentAdvances in Polyphenol Research F Daay and V Lattanzio Edspp 1ndash35 Wiley-Blackwell Oxford UK 2008
[30] V De Micco C Arena and G Aronne ldquoAnatomical alterationsof Phaseolus vulgaris L mature leaves irradiated with X-raysrdquoPlant Biology vol 16 pp 187ndash193 2014
[31] A D Krikorian ldquoPlants and somatic embryos in space whathave we learnedrdquo Gravitational and Space Biology Bulletin vol11 no 2 pp 5ndash14 1998
[32] R Meissner Y Jacobson S Melamed et al ldquoA new modelsystem for tomato geneticsrdquoThe Plant Journal vol 12 no 6 pp1465ndash1472 1997
[33] E Emmanuel and A A Levy ldquoTomato mutants as tools forfunctional genomicsrdquo Current Opinion in Plant Biology vol 5no 2 pp 112ndash117 2002
[34] E Martı C Gisbert G J Bishop M S Dixon and J LGarcıa-Martınez ldquoGenetic and physiological characterizationof tomato cv Micro-Tomrdquo Journal of Experimental Botany vol57 pp 2037ndash2047 2006
The Scientific World Journal 13
[35] T Kurimoto J V H Constable and A Huda ldquoEffects ofionizing radiation exposure on arabidopsis thalianardquo HealthPhysics vol 99 no 1 pp 49ndash57 2010
[36] L Reale D Gigante F Landucci F Ferranti and R VenanzonildquoMorphological and histo-anatomical traits reflect die-back inPhragmites australis (Cav) Steudrdquo Aquatic Botany vol 103 pp122ndash128 2012
[37] K Fukuzawa ldquoUltraviolet microscopyrdquo in Methods in LigninChemistry S Y Lin andCWDence Eds pp 110ndash131 SpringerBerlin Germany 1992
[38] S E Ruzin Plant Microtechnique and Microscopy OxfordUniversity Press New York NY USA 1999
[39] V De Micco G Aronne J-P Joseleau and K Ruel ldquoXylemdevelopment and cell wall changes of soybean seedlings grownin spacerdquo Annals of Botany vol 101 no 5 pp 661ndash669 2008
[40] S Van Buggenhout T Grauwet A Van Loey and M Hen-drickx ldquoStructureprocessing relation of vacuum infused straw-berry tissue frozen under different conditionsrdquo European FoodResearch and Technology vol 226 no 3 pp 437ndash448 2008
[41] V De Micco and G Aronne ldquoBiometric anatomy of seedlingsdeveloped onboard of Foton-M2 in an automatic system sup-porting growthrdquoActa Astronautica vol 62 no 8-9 pp 505ndash5132008
[42] B Genty J M Briantais and N R Baker ldquoThe relationshipbetween quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescencerdquo Biochimica andBiophysica Acta vol 990 pp 87ndash92 1989
[43] W Bilger and O Bjorkman ldquoRole of the xanthophyll cycle inphotoprotection elucidated by measurements of light-inducedabsorbance changes fluorescence and photosynthesis in leavesof Hedera canariensisrdquo Photosynthesis Research vol 25 no 3pp 173ndash185 1990
[44] H K Lichtenthaler ldquoChlorophylls and carotenoids pigmentsof photosynthetic biomembranesrdquoMethods in Enzymology vol148 pp 350ndash382 1987
[45] J P Maity DMishra A Chakraborty A Saha S C Santra andS Chanda ldquoModulation of some quantitative and qualitativecharacteristics in rice (Oryza sativa L) and mung (Phaseolusmungo L) by ionizing radiationrdquo Radiation Physics and Chem-istry vol 74 no 5 pp 391ndash394 2005
[46] L Cheng H Yang B Lin et al ldquoEffect of gamma-ray radiationon physiological morphological characters and chromosomeaberrations of minitubers in Solanum tuberosum Lrdquo Interna-tional Journal of Radiation Biology vol 86 no 9 pp 791ndash7992010
[47] B Al-Safadi and R Elias ldquoImprovement of caper (Capparisspinosa L) propagation using in vitro culture and gammairradiationrdquo Scientia Horticulturae vol 127 no 3 pp 290ndash2972011
[48] O Monje G W Stutte G D Goins D M Porterfield and GE Bingham ldquoFarming in space environmental and biophysicalconcernsrdquo Advances in Space Research vol 31 no 1 pp 151ndash1672003
[49] V De Micco M Scala and G Aronne ldquoEffects of simulatedmicrogravity on male gametophyte of Prunus Pyrus andBrassica speciesrdquo Protoplasma vol 228 no 1ndash3 pp 121ndash1262006
[50] V De Micco R Buonomo R Paradiso S De Pascale and GAronne ldquoSoybean cultivar selection for Bioregenerative LifeSupport Systems (BLSS)mdashtheoretical selectionrdquo Advances inSpace Research vol 49 no 10 pp 1415ndash1421 2012
[51] T Kozuka S-G Kong M Doi K-I Shimazaki and ANagatani ldquoTissue-autonomous promotion of palisade celldevelopment by phototropin 2 in Arabidopsisrdquo Plant Cell vol23 no 10 pp 3684ndash3695 2011
[52] E Kovacs and A Keresztes ldquoEffect of gamma and UV-BCradiation on plant cellsrdquoMicron vol 33 no 2 pp 199ndash210 2002
[53] R Ogaya J Penuelas D Asensio and J Llusia ldquoChlorophyllfluorescence responses to temperature and water availabilityin two co-dominant Mediterranean shrub and tree speciesin a long-term field experiment simulating climate changerdquoEnvironmental and Experimental Botany vol 73 no 1 pp 89ndash93 2011
[54] R Zaka C M Vandecasteele and M T Misset ldquoEffects oflow chronic doses of ionizing radiation on antioxidant enzymesand G6PDH activities in Stipa capillata (Poaceae)rdquo Journal ofExperimental Botany vol 53 no 376 pp 1979ndash1987 2002
[55] J-H KimM-H Baek Y C Byung GW Seung and J-S KimldquoAlterations in the photosynthetic pigments and antioxidantmachineries of red pepper (Capsicum annuum L) seedlingsfrom gamma-irradiated seedsrdquo Journal of Plant Biology vol 47no 4 pp 314ndash321 2004
[56] N A Al-Enezi and J M Al-Khayri ldquoAlterations of DNA ionsand photosynthetic pigments content in date palm seedlingsinduced by X-irradiationrdquo International Journal of Agricultureand Biology vol 14 pp 329ndash336 2012
[57] V de Micco C Arena L Vitale G Aronne and A V de SantoldquoAnatomy and photochemical behaviour of mediterranean Cis-tus incanus winter leaves under natural outdoor and warmerindoor conditionsrdquo Botany vol 89 no 10 pp 677ndash688 2011
[58] L H Levine H G Levine E C Stryjewski V Prima and WC Piastuch ldquoEffect of spaceflight on isoflavonoid accumulationin etiolated soybean seedlingsrdquo Journal of Gravitational Physiol-ogy vol 8 no 2 pp 21ndash27 2001
[59] X Fan ldquoAntioxidant capacity of fresh-cut vegetables exposedto ionizing radiationrdquo Journal of the Science of Food andAgriculture vol 85 no 6 pp 995ndash1000 2005
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

12 The Scientific World Journal
protectionrdquo Journal of Radiological Protection vol 24 no 4 ppA123ndashA137 2004
[2] V De Micco C Arena D Pignalosa and M Durante ldquoEffectsof sparsely and densely ionizing radiation on plantsrdquo Radiationand Environmental Biophysics vol 50 no 1 pp 1ndash19 2011
[3] N Kumari P Kumar D Mitra B Prasad B N Tiwary andL Varshney ldquoEffects of ionizing radiation on microbial decon-tamination phenolic contents and antioxidant properties oftriphalardquo Journal of Food Science vol 74 no 3 pp M109ndashM1132009
[4] A Tanaka N Shikazono and Y Hase ldquoStudies on biologicaleffects of ion beams on lethality molecular nature of mutationmutation rate and spectrum of mutation phenotype for muta-tion breeding in higher plantsrdquo Journal of Radiation Researchvol 51 no 3 pp 223ndash233 2010
[5] M Durante and A Kronenberg ldquoGround-based research withheavy ions for space radiation protectionrdquo Advances in SpaceResearch vol 35 no 2 pp 180ndash184 2005
[6] E Hu S I Bartsev and H Liu ldquoConceptual design of a biore-generative life support system containing crops and silkwormsrdquoAdvances in Space Research vol 45 no 7 pp 929ndash939 2010
[7] V deMicco G Aronne G Colla R Fortezza and S de PascaleldquoAgro-biology for bioregenerative life support systems in long-term space missions general constraints and the Italian effortsrdquoJournal of Plant Interactions vol 4 no 4 pp 241ndash252 2009
[8] T W Halstead and F R Dutcher ldquoPlants in spacerdquo AnnualReview of Plant Physiology vol 38 pp 317ndash345 1987
[9] V De Micco S De Pascale R Paradiso and G AronneldquoMicrogravity effects on different stages of higher plant life cycleand completion of the seed-to-seed cyclerdquo Plant Biology vol 16pp 31ndash38 2014
[10] C J Bennett C Pirim and T M Orlando ldquoSpace-weatheringof solar system bodies a laboratory perspectiverdquo ChemicalReviews vol 113 pp 9086ndash9150 2013
[11] R A Kerr ldquoRadiation will make astronautsrsquo trip to Mars evenriskierrdquo Science vol 340 article 1031 2014
[12] D M Hassler C Zeitlin R F Wimmer-Schweingruber et alldquoMarsrsquo surface radiation environment measured with the MarsScience Laboratoryrsquos Curiosity Roverrdquo Science vol 343 no6169 Article ID 1244797 2014
[13] M Durante and F A Cucinotta ldquoHeavy ion carcinogenesis andhuman space explorationrdquo Nature Reviews Cancer vol 8 no 6pp 465ndash472 2008
[14] C Arena V De Micco and A De Maio ldquoGrowth alterationand leaf biochemical responses in P vulgaris plants exposed todifferent doses of ionizing radiationrdquo Plant Biology vol 16 pp194ndash202 2014
[15] R W Holst and D J Nagel ldquoRadiation effects on plantsrdquo inPlants For Environmental Studies W Wang J W Gorsuch andJ S Hughes Eds pp 37ndash81 Lewis Publishers Boca Raton FlaUSA 1997
[16] T D Luckey Hormesis With Ionizing Radiation CRC PressBoca Raton Fla USA 1980
[17] D Marcu V Cristea and L Daraban ldquoDose-dependent effectsof gamma radiation on lettuce (Lactuca sativa var capitata)seedlingsrdquo International Journal of Radiation Biology vol 89 pp219ndash223 2013
[18] M Mei H Deng Y Lu et al ldquoMutagenic effects of heavy ionradiation in plantsrdquo Advances in Space Research vol 14 no 10pp 363ndash372 1994
[19] Y LiM Liu Z Cheng andY Sun ldquoSpace environment inducedmutations prefer to occur at polymorphic sites of rice genomesrdquoAdvances in Space Research vol 40 no 4 pp 523ndash527 2007
[20] D S Kim J-B Kim E J Goh et al ldquoAntioxidant response ofArabidopsis plants to gamma irradiation genome-wide expres-sion profiling of the ROS scavenging and signal transductionpathwaysrdquo Journal of Plant Physiology vol 168 no 16 pp 1960ndash1971 2011
[21] T J Brodribb ldquoXylem hydraulic physiology the functionalbackbone of terrestrial plant productivityrdquo Plant Science vol177 no 4 pp 245ndash251 2009
[22] R M Wheeler J C Sager R P Prince andW M Knott ldquoCropproduction for advanced life support systemsmdashobservationfrom the Kennedy Space Center Breadboard Projectrdquo NASATM-2003-211184 NASA Kennedy Space Center Florida 2003
[23] R M Wheeler C L Mackowiak G W Stutte et al ldquoCropproductivities and radiation use efficiencies for bioregenerativelife supportrdquo Advances in Space Research vol 41 no 5 pp 706ndash713 2008
[24] R Paradiso V De Micco R Buonomo G Aronne G Bar-bieri and S De Pascale ldquoSoilless cultivation of soybean forBioregenerative Life-Support Systems a literature review andthe experience of the MELiSSA ProjectmdashFood characterisationPhase Irdquo Plant Biology vol 16 pp 69ndash78 2014
[25] D M Porterfield ldquoThe biophysical limitations in physiologicaltransport and exchange in plants grown in microgravityrdquoJournal of Plant Growth Regulation vol 21 no 2 pp 177ndash1902002
[26] K Yokthongwattana and A Melis ldquoPhotoinhibition and recov-ery in oxygenic photosynthesis mechanism of a photosystem IIdamage and repair cyclerdquo in Photoprotection PhotoinhibitionGene Regulation and Environment B Demmig-Adams WW Adams III and A K Mattoo Eds pp 175ndash191 SpringerDordecht The Netherlands 2006
[27] M T Giardi G Rea M D Lambreva et al ldquoMutations ofphotosystem II D1 protein that empower efficient phenotypesof Chlamydomonas reinhardtii under extreme environment inSpacerdquo PLoS ONE vol 8 Article ID e64352 2013
[28] C Arena V De Micco G Aronne M Pugliese A Virzo DeSanto andADeMaio ldquoResponse ofPhaseolus vulgarisL plantsto low-let ionizing radiation growth and oxidative stressrdquo ActaAstronautica vol 91 pp 107ndash114 2013
[29] V Lattanzio P A Kroon and S Quideau ldquoPlant phenolicsmdashsecondary metabolites with diverse functionsrdquo in RecentAdvances in Polyphenol Research F Daay and V Lattanzio Edspp 1ndash35 Wiley-Blackwell Oxford UK 2008
[30] V De Micco C Arena and G Aronne ldquoAnatomical alterationsof Phaseolus vulgaris L mature leaves irradiated with X-raysrdquoPlant Biology vol 16 pp 187ndash193 2014
[31] A D Krikorian ldquoPlants and somatic embryos in space whathave we learnedrdquo Gravitational and Space Biology Bulletin vol11 no 2 pp 5ndash14 1998
[32] R Meissner Y Jacobson S Melamed et al ldquoA new modelsystem for tomato geneticsrdquoThe Plant Journal vol 12 no 6 pp1465ndash1472 1997
[33] E Emmanuel and A A Levy ldquoTomato mutants as tools forfunctional genomicsrdquo Current Opinion in Plant Biology vol 5no 2 pp 112ndash117 2002
[34] E Martı C Gisbert G J Bishop M S Dixon and J LGarcıa-Martınez ldquoGenetic and physiological characterizationof tomato cv Micro-Tomrdquo Journal of Experimental Botany vol57 pp 2037ndash2047 2006
The Scientific World Journal 13
[35] T Kurimoto J V H Constable and A Huda ldquoEffects ofionizing radiation exposure on arabidopsis thalianardquo HealthPhysics vol 99 no 1 pp 49ndash57 2010
[36] L Reale D Gigante F Landucci F Ferranti and R VenanzonildquoMorphological and histo-anatomical traits reflect die-back inPhragmites australis (Cav) Steudrdquo Aquatic Botany vol 103 pp122ndash128 2012
[37] K Fukuzawa ldquoUltraviolet microscopyrdquo in Methods in LigninChemistry S Y Lin andCWDence Eds pp 110ndash131 SpringerBerlin Germany 1992
[38] S E Ruzin Plant Microtechnique and Microscopy OxfordUniversity Press New York NY USA 1999
[39] V De Micco G Aronne J-P Joseleau and K Ruel ldquoXylemdevelopment and cell wall changes of soybean seedlings grownin spacerdquo Annals of Botany vol 101 no 5 pp 661ndash669 2008
[40] S Van Buggenhout T Grauwet A Van Loey and M Hen-drickx ldquoStructureprocessing relation of vacuum infused straw-berry tissue frozen under different conditionsrdquo European FoodResearch and Technology vol 226 no 3 pp 437ndash448 2008
[41] V De Micco and G Aronne ldquoBiometric anatomy of seedlingsdeveloped onboard of Foton-M2 in an automatic system sup-porting growthrdquoActa Astronautica vol 62 no 8-9 pp 505ndash5132008
[42] B Genty J M Briantais and N R Baker ldquoThe relationshipbetween quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescencerdquo Biochimica andBiophysica Acta vol 990 pp 87ndash92 1989
[43] W Bilger and O Bjorkman ldquoRole of the xanthophyll cycle inphotoprotection elucidated by measurements of light-inducedabsorbance changes fluorescence and photosynthesis in leavesof Hedera canariensisrdquo Photosynthesis Research vol 25 no 3pp 173ndash185 1990
[44] H K Lichtenthaler ldquoChlorophylls and carotenoids pigmentsof photosynthetic biomembranesrdquoMethods in Enzymology vol148 pp 350ndash382 1987
[45] J P Maity DMishra A Chakraborty A Saha S C Santra andS Chanda ldquoModulation of some quantitative and qualitativecharacteristics in rice (Oryza sativa L) and mung (Phaseolusmungo L) by ionizing radiationrdquo Radiation Physics and Chem-istry vol 74 no 5 pp 391ndash394 2005
[46] L Cheng H Yang B Lin et al ldquoEffect of gamma-ray radiationon physiological morphological characters and chromosomeaberrations of minitubers in Solanum tuberosum Lrdquo Interna-tional Journal of Radiation Biology vol 86 no 9 pp 791ndash7992010
[47] B Al-Safadi and R Elias ldquoImprovement of caper (Capparisspinosa L) propagation using in vitro culture and gammairradiationrdquo Scientia Horticulturae vol 127 no 3 pp 290ndash2972011
[48] O Monje G W Stutte G D Goins D M Porterfield and GE Bingham ldquoFarming in space environmental and biophysicalconcernsrdquo Advances in Space Research vol 31 no 1 pp 151ndash1672003
[49] V De Micco M Scala and G Aronne ldquoEffects of simulatedmicrogravity on male gametophyte of Prunus Pyrus andBrassica speciesrdquo Protoplasma vol 228 no 1ndash3 pp 121ndash1262006
[50] V De Micco R Buonomo R Paradiso S De Pascale and GAronne ldquoSoybean cultivar selection for Bioregenerative LifeSupport Systems (BLSS)mdashtheoretical selectionrdquo Advances inSpace Research vol 49 no 10 pp 1415ndash1421 2012
[51] T Kozuka S-G Kong M Doi K-I Shimazaki and ANagatani ldquoTissue-autonomous promotion of palisade celldevelopment by phototropin 2 in Arabidopsisrdquo Plant Cell vol23 no 10 pp 3684ndash3695 2011
[52] E Kovacs and A Keresztes ldquoEffect of gamma and UV-BCradiation on plant cellsrdquoMicron vol 33 no 2 pp 199ndash210 2002
[53] R Ogaya J Penuelas D Asensio and J Llusia ldquoChlorophyllfluorescence responses to temperature and water availabilityin two co-dominant Mediterranean shrub and tree speciesin a long-term field experiment simulating climate changerdquoEnvironmental and Experimental Botany vol 73 no 1 pp 89ndash93 2011
[54] R Zaka C M Vandecasteele and M T Misset ldquoEffects oflow chronic doses of ionizing radiation on antioxidant enzymesand G6PDH activities in Stipa capillata (Poaceae)rdquo Journal ofExperimental Botany vol 53 no 376 pp 1979ndash1987 2002
[55] J-H KimM-H Baek Y C Byung GW Seung and J-S KimldquoAlterations in the photosynthetic pigments and antioxidantmachineries of red pepper (Capsicum annuum L) seedlingsfrom gamma-irradiated seedsrdquo Journal of Plant Biology vol 47no 4 pp 314ndash321 2004
[56] N A Al-Enezi and J M Al-Khayri ldquoAlterations of DNA ionsand photosynthetic pigments content in date palm seedlingsinduced by X-irradiationrdquo International Journal of Agricultureand Biology vol 14 pp 329ndash336 2012
[57] V de Micco C Arena L Vitale G Aronne and A V de SantoldquoAnatomy and photochemical behaviour of mediterranean Cis-tus incanus winter leaves under natural outdoor and warmerindoor conditionsrdquo Botany vol 89 no 10 pp 677ndash688 2011
[58] L H Levine H G Levine E C Stryjewski V Prima and WC Piastuch ldquoEffect of spaceflight on isoflavonoid accumulationin etiolated soybean seedlingsrdquo Journal of Gravitational Physiol-ogy vol 8 no 2 pp 21ndash27 2001
[59] X Fan ldquoAntioxidant capacity of fresh-cut vegetables exposedto ionizing radiationrdquo Journal of the Science of Food andAgriculture vol 85 no 6 pp 995ndash1000 2005
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

The Scientific World Journal 13
[35] T Kurimoto J V H Constable and A Huda ldquoEffects ofionizing radiation exposure on arabidopsis thalianardquo HealthPhysics vol 99 no 1 pp 49ndash57 2010
[36] L Reale D Gigante F Landucci F Ferranti and R VenanzonildquoMorphological and histo-anatomical traits reflect die-back inPhragmites australis (Cav) Steudrdquo Aquatic Botany vol 103 pp122ndash128 2012
[37] K Fukuzawa ldquoUltraviolet microscopyrdquo in Methods in LigninChemistry S Y Lin andCWDence Eds pp 110ndash131 SpringerBerlin Germany 1992
[38] S E Ruzin Plant Microtechnique and Microscopy OxfordUniversity Press New York NY USA 1999
[39] V De Micco G Aronne J-P Joseleau and K Ruel ldquoXylemdevelopment and cell wall changes of soybean seedlings grownin spacerdquo Annals of Botany vol 101 no 5 pp 661ndash669 2008
[40] S Van Buggenhout T Grauwet A Van Loey and M Hen-drickx ldquoStructureprocessing relation of vacuum infused straw-berry tissue frozen under different conditionsrdquo European FoodResearch and Technology vol 226 no 3 pp 437ndash448 2008
[41] V De Micco and G Aronne ldquoBiometric anatomy of seedlingsdeveloped onboard of Foton-M2 in an automatic system sup-porting growthrdquoActa Astronautica vol 62 no 8-9 pp 505ndash5132008
[42] B Genty J M Briantais and N R Baker ldquoThe relationshipbetween quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescencerdquo Biochimica andBiophysica Acta vol 990 pp 87ndash92 1989
[43] W Bilger and O Bjorkman ldquoRole of the xanthophyll cycle inphotoprotection elucidated by measurements of light-inducedabsorbance changes fluorescence and photosynthesis in leavesof Hedera canariensisrdquo Photosynthesis Research vol 25 no 3pp 173ndash185 1990
[44] H K Lichtenthaler ldquoChlorophylls and carotenoids pigmentsof photosynthetic biomembranesrdquoMethods in Enzymology vol148 pp 350ndash382 1987
[45] J P Maity DMishra A Chakraborty A Saha S C Santra andS Chanda ldquoModulation of some quantitative and qualitativecharacteristics in rice (Oryza sativa L) and mung (Phaseolusmungo L) by ionizing radiationrdquo Radiation Physics and Chem-istry vol 74 no 5 pp 391ndash394 2005
[46] L Cheng H Yang B Lin et al ldquoEffect of gamma-ray radiationon physiological morphological characters and chromosomeaberrations of minitubers in Solanum tuberosum Lrdquo Interna-tional Journal of Radiation Biology vol 86 no 9 pp 791ndash7992010
[47] B Al-Safadi and R Elias ldquoImprovement of caper (Capparisspinosa L) propagation using in vitro culture and gammairradiationrdquo Scientia Horticulturae vol 127 no 3 pp 290ndash2972011
[48] O Monje G W Stutte G D Goins D M Porterfield and GE Bingham ldquoFarming in space environmental and biophysicalconcernsrdquo Advances in Space Research vol 31 no 1 pp 151ndash1672003
[49] V De Micco M Scala and G Aronne ldquoEffects of simulatedmicrogravity on male gametophyte of Prunus Pyrus andBrassica speciesrdquo Protoplasma vol 228 no 1ndash3 pp 121ndash1262006
[50] V De Micco R Buonomo R Paradiso S De Pascale and GAronne ldquoSoybean cultivar selection for Bioregenerative LifeSupport Systems (BLSS)mdashtheoretical selectionrdquo Advances inSpace Research vol 49 no 10 pp 1415ndash1421 2012
[51] T Kozuka S-G Kong M Doi K-I Shimazaki and ANagatani ldquoTissue-autonomous promotion of palisade celldevelopment by phototropin 2 in Arabidopsisrdquo Plant Cell vol23 no 10 pp 3684ndash3695 2011
[52] E Kovacs and A Keresztes ldquoEffect of gamma and UV-BCradiation on plant cellsrdquoMicron vol 33 no 2 pp 199ndash210 2002
[53] R Ogaya J Penuelas D Asensio and J Llusia ldquoChlorophyllfluorescence responses to temperature and water availabilityin two co-dominant Mediterranean shrub and tree speciesin a long-term field experiment simulating climate changerdquoEnvironmental and Experimental Botany vol 73 no 1 pp 89ndash93 2011
[54] R Zaka C M Vandecasteele and M T Misset ldquoEffects oflow chronic doses of ionizing radiation on antioxidant enzymesand G6PDH activities in Stipa capillata (Poaceae)rdquo Journal ofExperimental Botany vol 53 no 376 pp 1979ndash1987 2002
[55] J-H KimM-H Baek Y C Byung GW Seung and J-S KimldquoAlterations in the photosynthetic pigments and antioxidantmachineries of red pepper (Capsicum annuum L) seedlingsfrom gamma-irradiated seedsrdquo Journal of Plant Biology vol 47no 4 pp 314ndash321 2004
[56] N A Al-Enezi and J M Al-Khayri ldquoAlterations of DNA ionsand photosynthetic pigments content in date palm seedlingsinduced by X-irradiationrdquo International Journal of Agricultureand Biology vol 14 pp 329ndash336 2012
[57] V de Micco C Arena L Vitale G Aronne and A V de SantoldquoAnatomy and photochemical behaviour of mediterranean Cis-tus incanus winter leaves under natural outdoor and warmerindoor conditionsrdquo Botany vol 89 no 10 pp 677ndash688 2011
[58] L H Levine H G Levine E C Stryjewski V Prima and WC Piastuch ldquoEffect of spaceflight on isoflavonoid accumulationin etiolated soybean seedlingsrdquo Journal of Gravitational Physiol-ogy vol 8 no 2 pp 21ndash27 2001
[59] X Fan ldquoAntioxidant capacity of fresh-cut vegetables exposedto ionizing radiationrdquo Journal of the Science of Food andAgriculture vol 85 no 6 pp 995ndash1000 2005
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology