leaf morphology and anatomy in two contrasting...
TRANSCRIPT

Acta Botanica Hungarica 50(1–2), pp. 97–113, 2008DOI: 10.1556/ABot.50.2008.1–2.7
LEAF MORPHOLOGY AND ANATOMY IN TWOCONTRASTING ENVIRONMENTS FOR C3 AND C4GRASSES OF DIFFERENT INVASION POTENTIAL
L. J. HAN1, A. MOJZES2,3 and T. KALAPOS2
1School of Life Sciences, Changchun Normal University, 3 North Line, Jichang HighwayChangchun 130032, Jilin, China
2Department of Plant Taxonomy and Ecology, Eötvös Loránd UniversityH-1117 Budapest, Pázmány P. s. 1/C, Hungary; E-mail: [email protected]
3Institute of Ecology and Botany, Hungarian Academy of SciencesH-2163 Vácrátót, Alkotmány u. 2–4, Hungary
(Received 28 March, 2007; Accepted 30 July, 2007)
Leaf morphology, coarse structure and anatomy were compared for two invasive C4, twonon-invasive C4, and two expanding native C3 grass species grown in their original,high-light semiarid temperate habitat, and in a growth room under variable moderate lightand favourable supply of water and nutrients. It was hypothesised that (H1) among C4grasses leaf structural response will be greater for invasive than for non-invasive species,and (H2) for plants of high spreading capacity C4 species will be less responsive than C3 spe-cies. Leaf mass per area was lower in the growth room than in the field by 43.4–54% and5.7–21.2% for grasses of high spreading capacity and for non-invasive C4 species, respec-tively. Little or no response was observed in the proportion of epidermis and mesophyll,but the proportional area of veins plus sclerenchyma was greater in the field than in thegrowth room for the invasive C4 Sorghum halepense, and the spreading C3 Bromus inermisand Calamagrostis epigeios, while it did not differ for the two non-invasive C4 grasses and theinvasive C4 Cynodon dactylon. Leaf intervenial distance was invariant for C4 grasses (exceptfor the non-invasive Chrysopogon gryllus) and the C3 C. epigeios, but changed by 25.1% for theC3 B. inermis. These results suggest that among C4 grasses invasive species exceed non-inva-sive ones in the plasticity of leaf coarse structure, but not that of leaf morphology and anat-omy. However, leaf structure was not less plastic in invasive C4 than in expanding C3grasses except for intervenial distance.
Key words: Bothriochloa ischaemum, Bromus inermis, Calamagrostis epigeios, Chrysopogon gryl-lus, Cynodon dactylon, leaf intervenial distance, leaf mass per area, leaf structure, plasticity,Sorghum halepense
0236–6495/$ 20.00 © 2008 Akadémiai Kiadó, Budapest

INTRODUCTION
Grasses (Poaceae) usually inhabit high-light environments and theirabundance declines in biotopes with moderate to low light intensity (e.g. inforest understorey, Chapman (1996), Bredenkamp et al. (2002)). In addition totheir characteristic shoot morphology with erect leaves, morphology and ana-tomical structure of individual leaves must play an important role in this habi-tat preference. There is considerable variation among grasses in the capacity topersist in shaded environments. Grasses with the C4 photosynthetic pathwayare considered to have lower tolerance of low irradiance than grasses with theC3 photosynthetic metabolism, possibly due to the extra energy requirementof the C4 dicarboxylic pathway, to insufficient time for evolutionary adapta-tion to shaded environments (Pearcy and Ehleringer 1984) or to a limited plas-ticity or acclimation potential to low light intensity at leaf structural and bio-chemical levels (Sage and McKown 2006). A possible additional source of vari-ation among grasses in the tolerance of low light is the invasive potential of thespecies. We hypothesise that invasive grasses (particularly those with the C4photosynthetic pathway) capable of rapid spreading across a wide spectrumof environments should better tolerate heterogeneous light environments – in-cluding low-light habitats – than related species with low capacity for colonis-ation.
Numerous plant traits have been identified so far to be associated withthe invasiveness of plants, including life history traits (Burke and Grime 1996,Kolar and Lodge 2001, Radford and Cousens 2000, Rejmánek and Richardson1996, White et al. 2001), present geographical distribution, reflecting the toler-ance of the range of climates (Burke and Grime 1996, Dukes and Mooney1999), as well as attributes associated with carbon gain and resource-use effi-ciency (McDowell 2002, Pattison et al. 1998, Williams et al. 1995, Yamashita etal. 2000, 2002). The role of leaf morphology and anatomy in a species’ invasionpotential, however, is not sufficiently investigated yet, particularly for thegroup of grasses (Molnár et al. 2000, Yamashita et al. 2000, 2002). In addition tochanges in leaf biochemistry, ultrastructure and plant biomass allocation, leafmorphological and anatomical modifications are frequent components ofplant adjustment to contrasting habitat light conditions (Boardman 1977,Fekete and Szujkó-Lacza 1973, Givnish 1988, Lambers et al. 1998, Mendes et al.2001, Mojzes et al. 2005, Oguchi et al. 2003, Sims and Pearcy 1992).
In this study, we aimed at comparing leaf morphological and anatomicalstructure of six grass species grown in two contrasting environments: underfull sun in their natural semiarid temperate habitats and in a growth room un-der variable moderate light and good supply of water and nutrients. Condi-tions with a lower radiation load in the growth room can be considered as
98 HAN, L. J., MOJZES, A. and KALAPOS, T.
Acta Bot. Hung. 50, 2008

analogous to (but do not mimicking) those in shade compared to full sunpatches of forest steppe vegetation mosaic. In addition to lower light intensity– as its main component – it also includes lower frequency or shorter periodsof high temperature and temporal water shortage, and is associated withlower mechanical stress caused by wind. Thus, rather than focusing on the ef-fect of light intensity exclusively, we explored phenotypical responses tochanges in several associated environmental factors. Two hypotheses weretested:
H1: invasive C4 grasses have a higher capacity to adjust leaf morphologyand anatomy to contrasting environments than non-invasive C4 grasses do.This is presumed because spreading species probably encounter heteroge-neous environments more often than their stationary relatives.
H2: Among grasses of high colonisation capacity (either invasive aliens orexpanding natives) C4 species possess a lesser degree of plasticity in leaf mor-phology and anatomy than C3 species do. In a recent paper Sage and McKown(2006) argues on such difference among photosynthetic pathways.
MATERIALS AND METHODS
Species studied
Six perennial grass species were included in this study: four C4 and twoC3 species. Among the C4 grasses bermudagrass (Cynodon dactylon (L.) Pers.)and Johnsongrass (Sorghum halepense (L.) Pers.) are invasive (Grace et al. 2001,Holm et al. 1977), while gold beard grass (Chrysopogon gryllus (Torn.) Trin.) andyellow bluestem (Bothriochloa ischaemum (L.) Keng) are non-invasive natives.The latter species may reach local dominance as the canopy of sympatricgrasses opens up on disturbance (Zólyomi and Fekete 1994). Each C4 speciesstudied belongs to the NADP-ME biochemical subtype except C. dactylon,which is a NAD-ME C4 plant. Both C3 species studied here are native and pos-sess high capacity for spatial expansion. Smooth brome (Bromus inermisLeyss.) is a mid-successional species native in Europe, frequent in semiariddisturbed areas, and is an invasive alien throughout North America (e.g.Grace et al. 2001, Grilz and Romo 1994, Willson and Stubbendieck 2000). Thenoxious weed chee reedgrass (Calamagrostis epigeios (L.) Roth) is able to colo-nise rapidly a wide range of disturbed habitats, particularly forest clearings,abandoned fields and wastelands (Rebele and Lehmann 2001).
LEAF STRUCTURAL RESPONSE OF INVASIVE GRASSES 99
Acta Bot. Hung. 50, 2008

Field sampling and growth conditions
Our aim was to compare the morphology and anatomy of leaves devel-oped under two contrasting conditions: in the field with semiarid climate andhigh-light conditions, and in a growth room of low-light environment with ad-equate water and nutrient supply. Whole plants in soil monoliths were col-lected in the field and transferred to laboratory for growing plants undersemicontrolled conditions. Except for B. inermis, species were sampled in for-est steppe vegetation and adjacent arable land (cornfield) near the villageIsaszeg, 25 km East of Budapest in the summers of 2002–2004. Annual meantemperature is 9 °C, yearly precipitation is about 600 mm. Chernozem soil cov-ers the loess bedrock typically (Fekete et al. 2000). Bromus inermis was trans-planted from forest steppe vegetation on calcareous sandy soil near the villageFülöpháza, on the Great Hungarian Plain. Annual mean temperature is 10.4°C, yearly precipitation is 505 mm (Kovács-Láng et al. 2000). Plants with theiroriginal soil were placed in 4-litre pots, shoots were cut back to 1 cm above soilsurface and newly emerged shoots were grown in the growth room of theEötvös Loránd University. In the growth room, plants received natural sun-light supplemented with a 1,000 W halogen lamp over a photoperiod of 12 h insummer and 9 h in winter. Pots were rotated every 3 weeks so that we canminimise spatial heterogeneity of light environment in the growth room. Themean maximum photosynthetic photon flux density (PPFD) at the height ofshoots on clear days was 810 µmol m–2 s–1 in summer and 180 µmol m–2 s–1 inwinter (i.e. 40% and 9% of full sunlight, respectively). Air temperature and rel-ative humidity were automatically recorded hourly by using an HOBO ProRH/Temp sensor (Onset Computers Inc., Bourne, MA, USA), which was puton the table where pots were placed. Daily mean air temperature was 24.0 ± 4.4°C in summer and 18.5 ± 2.3 °C in winter. Relative air humidity ranged fromabout 20% to 80% during the day. Plants were watered adequately and sup-plied with 0.5 ml nutrient solution per pot (containing 13% N, 4.5% P2O5, 6.5%K2O and micronutrients) in three-week intervals. The second fully developedleaf from the top of 10 shoots per species was sampled in October 2004. Fiveleaves per species were used for morphological and another five for anatomi-cal measurements (n = 5). For comparison with field-grown plants, leaveswere sampled in the same manner from plants in their original habitats (or forS. halepense from a similar degraded biotope in Budapest) in June 2005. Aftersampling, leaves were transported to the laboratory immediately in closedcontainers with the cut leaf base immersed in water. Morphological measure-ments were completed on the same or following day.
100 HAN, L. J., MOJZES, A. and KALAPOS, T.
Acta Bot. Hung. 50, 2008

Data collection
Leaf morphology and coarse structure – Leaf length was measured by using aruler read to the nearest mm, while leaf width was determined in the middle ofthe leaf blade under a binocular microscope equipped with a measuring lensto the accuracy of 0.1 mm. One-sided leaf surface area was determined by aLeaf Area Meter (LI-COR 3000A, LI-COR Inc., Lincoln, Nebraska). Leaf thick-ness (T*, mm) was measured in the middle portion of the leaf lamina, halfwaybetween the leaf edge and the central vein by using a thickness meter (Dial In-dicator and Magnetic Base, a division of Siechert and Wood, Inc. Pasadena,USA, accuracy 0.01 mm). Leaf samples were dried to constant weight at 90 °Cand than dry mass was measured. Leaf mass per area (dry mass per unit leafarea, LMA, g m–2) and leaf bulk tissue density (dry mass per unit leaf volume,D = LMA/T, g cm–3) were calculated from these data.
Leaf anatomy – The middle portion of the leaf blade was severed and fixedin a 1:1:1 mixture of 96% alcohol, glycerine and distilled water until process-ing. Leaf blade cross sections were obtained by hand cutting without embed-ding by using elderpith and razor blades. Leaf cross-sections were perma-nently mounted in the same solute used for sample storage, observed withoutstaining and photographed under a light microscope (Nikon Eclipse E400,Nikon Inc., Yokohama, Japan) using a digital camera (Nikon CoolPix 4500).Quantitative leaf anatomical measurements were made on A4 sized printoutsof digital photographs. Distance data were measured by using a ruler, the var-ious tissue components were cut out, their mass was measured on an analyti-cal balance and their area was calculated by using the mass per area ratio ofthat sort of paper determined beforehand. For the sake of accuracy, scalingwas also photographed and printed out in the same way as leaf micrographs.Distance between vein centres (intervenial distance) were measured, and theproportional area of three component tissues (epidermis, mesophyll and thesum of vascular tissue and sclerenchyma) were calculated on a leaf blade crosssection halfway between the central vein and the leaf edge by using a magnifi-cation of 180 and 435 for C3 species and C4 species, respectively. The outer andinner bundle sheaths were included in the mesophyll and the vascular tissues,respectively. The thickness of mesophyll (for C3 species only) and epidermisthickness were determined at two points of an intervenial region. The thicknessof the outer, parenchymatous bundle sheath was measured on 3–4 randomlyselected bundle sheath cells per primary vein. These measurements were per-formed at a magnification of 850. Five replicates per environment were usedfor each species. For thickness parameters and for intervenial distance, aver-age values measured on the same cross section was used as replicates.
LEAF STRUCTURAL RESPONSE OF INVASIVE GRASSES 101
Acta Bot. Hung. 50, 2008
* Abbreviations: LMA = leaf mass per area; T = leaf thickness; D = leaf bulk tissue density

Statistical analyses
To test our hypotheses, we analysed intraspecific differences for eachvariable between means under the two growth conditions. Multiple compari-sons among species in the same environment were performed as well, but dueto the low number of species, these results were discussed only for robust dif-ferences among photosynthesis types or C4 subtypes regardless of the growthenvironment. Two-way ANOVA with growth conditions and species asgrouping variables was used with subsequent least significant difference(LSD) test to analyse significant differences among means. For variables wherethe homoscedasticity assumption of ANOVA was fulfilled within the samespecies or within the same light conditions only, unpaired t-test (with separatevariance estimates if necessary) was applied for comparisons of the means ofgrowth conditions, and one-way ANOVA with LSD test was used to test sig-nificant differences among the means of species, respectively. When data didnot meet the assumptions of ANOVA, nonparametric Kruskal-Wallis test withsubsequent post hoc test was used instead for multiple comparisons. Each sta-tistics was performed by using the Statistica version 7.0 software (StatSoft Inc.2004), and differences were considered significant at p < 0.05 level.
RESULTS
Leaf coarse structure
Leaf mass per area (LMA) was significantly greater in the field than in thegrowth room for each species except for C. gryllus (Fig. 1A). Variation in thisparameter was greater for S. halepense and C. dactylon than for B. ischaemum,while both C3 grasses showed about twofold differences (Table 1). For C.epigeios, S. halepense and C. dactylon, these differences in LMA resulted fromhigher leaf thickness (T) of plants grown in full sun compared with those de-veloped under moderate light conditions (Fig. 1B), while leaf bulk tissue den-sity (D) did not differ significantly between the two light environments forthese species (Fig. 1C). For B. inermis, both components of LMA (i.e. T and D)were significantly lower for leaves developed in the growth room than forthose grown in the field. Greater leaf bulk tissue density was the main deter-minant of higher LMA of field-grown leaves for B. ischaemum only, but the dif-ference was only marginally significant. Chrysopogon gryllus showed remark-able invariance in each leaf coarse structural parameter (Fig. 1, Table 1). Undermoderate irradiance, leaf thickness of this grass was the greatest among thespecies studied, and under field conditions it was similar to that of B. inermis.
102 HAN, L. J., MOJZES, A. and KALAPOS, T.
Acta Bot. Hung. 50, 2008

LEAF STRUCTURAL RESPONSE OF INVASIVE GRASSES 103
Acta Bot. Hung. 50, 2008
0
20
40
60
80
Leaf
mas
spe
rar
ea(L
MA,g
m–2
)
Field Growth roomBC
ab
*
C
ab
*
B bA
*
A
a
*
A
a
*
ab
0
0.07
0.14
0.21
0.28
Leaf
thic
knes
s(T
,mm
)
C
b
*
D
b
*
C c
A aB
a
*
B
a
*
0
0.2
0.4
0.6
0.8
Leafb
ulk
tissu
ede
nsity
(D,g
cm-3)
ABa
*
A abAB b
C
c
m
B b B b
B. inermis C.epigeios C.gryllus B.ischaemumS.halepense C.dactylon
C3 expanding C4 non-invasive C4 invasive
A
B
C
Fig. 1. Leaf coarse structure for six grasses of different photosynthetic pathway type andcapacity for area expansion developed in the field in full sun semiarid habitat or undervariable moderate light and good supply of water and nutrients in a growth room. A) Leafmass per area (LMA, g m–2); B) leaf thickness (T, mm); C) leaf bulk tissue density(D = LMA/T, g cm–3). Asterisks indicate significant differences (p < 0.05) between growthconditions, “m” denotes marginal significance (p = 0.0538). Within a given growthcondition, the same letters indicate statistically insignificant differences among means ofspecies (lower case letters for growth room, upper case characters for field conditions). For
each variable mean ± 1 SE of 5 replicates are shown
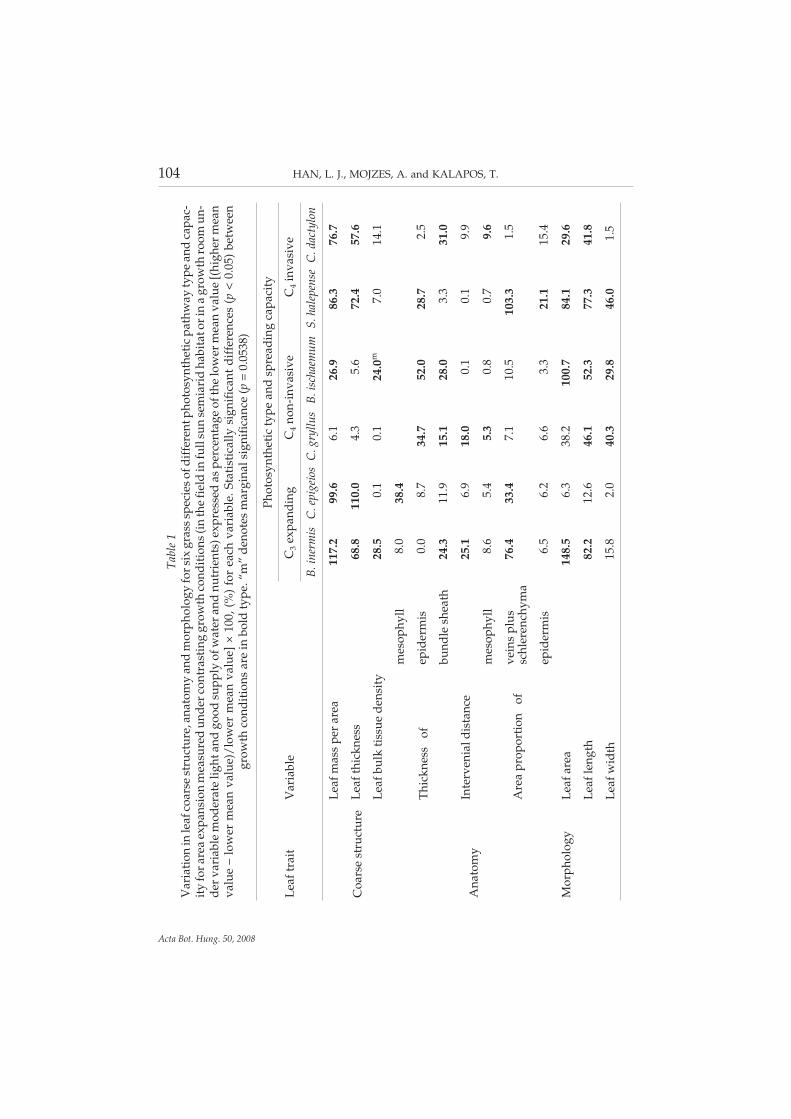
104 HAN, L. J., MOJZES, A. and KALAPOS, T.
Acta Bot. Hung. 50, 2008
Tabl
e1
Var
iati
onin
leaf
coar
sest
ruct
ure,
anat
omy
and
mor
phol
ogy
fors
ixgr
ass
spec
ies
ofdi
ffer
entp
hoto
synt
heti
cpa
thw
ayty
pean
dca
pac-
ity
fora
rea
expa
nsio
nm
easu
red
unde
rcon
tras
ting
grow
thco
ndit
ions
(in
the
fiel
din
full
sun
sem
iari
dha
bita
tori
na
grow
thro
omun
-de
rvar
iabl
em
oder
ate
light
and
good
supp
lyof
wat
eran
dnu
trie
nts)
expr
esse
das
perc
enta
geof
the
low
erm
ean
valu
e[(
high
erm
ean
valu
e–
low
erm
ean
valu
e)/l
ower
mea
nva
lue]
×10
0,(%
)fo
rea
chva
riab
le.S
tati
stic
ally
sign
ific
ant
diff
eren
ces
(p<
0.05
)be
twee
ngr
owth
cond
itio
nsar
ein
bold
type
.“m
”de
note
sm
argi
nals
igni
fica
nce
(p=
0.05
38)
Phot
osyn
thet
icty
pean
dsp
read
ing
capa
city
Leaf
trai
tV
aria
ble
C3
expa
ndin
gC
4no
n-in
vasi
veC
4in
vasi
ve
B.in
erm
isC
.epi
geio
sC
.gry
llus
B.is
chae
mum
S.ha
lepe
nse
C.d
acty
lon
Coa
rse
stru
ctur
e
Leaf
mas
spe
rar
ea11
7.2
99.6
6.1
26.9
86.3
76.7
Leaf
thic
knes
s68
.811
0.0
4.3
5.6
72.4
57.6
Leaf
bulk
tiss
uede
nsit
y28
.50.
10.
124
.0m
7.0
14.1
Ana
tom
y
Thic
knes
sof
mes
ophy
ll8.
038
.4
epid
erm
is0.
08.
734
.752
.028
.72.
5
bund
lesh
eath
24.3
11.9
15.1
28.0
3.3
31.0
Inte
rven
iald
ista
nce
25.1
6.9
18.0
0.1
0.1
9.9
Are
apr
opor
tion
of
mes
ophy
ll8.
65.
45.
30.
80.
79.
6
vein
spl
ussc
hler
ench
yma
76.4
33.4
7.1
10.5
103.
31.
5
epid
erm
is6.
56.
26.
63.
321
.115
.4
Mor
phol
ogy
Leaf
area
148.
56.
338
.210
0.7
84.1
29.6
Leaf
leng
th82
.212
.646
.152
.377
.341
.8
Leaf
wid
th15
.82.
040
.329
.846
.01.
5

In contrast, leaves of the other three C4 grasses were significantly thinner thanthose of the two C3 species under both light regimes.
Leaf anatomy
The mesophyll was significantly thinner in the growth room than in fullsun for C. epigeios, but remained similar for B. inermis (Table 2, this variablewas obtained for C3 species only). Epidermis thickness was significantly lowerin the growth room than in the field for C. gryllus, B. ischaemum and S.halepense, but did not vary significantly between light conditions for C. epigeios,B. inermis and C. dactylon. Field-grown leaves of B. inermis, B. ischaemum and C.dactylon possessed significantly thinner parenchymatous bundle sheath thanthose developed under moderate light conditions; an opposite pattern wasfound for C. gryllus, whereas C. epigeios and S. halepense did not show signifi-cant differences in this trait between the two environments. Among species, C.dactylon had the thickest parenchymatous bundle sheath under both irradian-ce levels. Average distance between veins was remarkably invariant for the C4grasses in the two contrasting environments. This trait differed significantlyonly for C. gryllus and B. inermis, but in an opposite way: it was greater undergrowth room conditions for B. inermis, but was lower for C. gryllus. When thesix species are compared concerning their photosynthesis types, mean inter-venial distance was significantly shorter for the C4 grasses than for the two C3grasses, and among the C4 types, this was greater for the NAD-ME C. dactylonthan for the other three NADP-ME type C4 species, regardless of the growthenvironment. Area proportion of mesophyll decreased slightly, but signifi-cantly under lower light conditions for C. gryllus, while increased for C.dactylon, and did not change significantly for the other species. Proportionalarea of veins plus sclerenchyma was significantly greater in field-grownleaves of B. inermis, C. epigeios and S. halepense than in those developed in thegrowth room, whereas did not vary significantly between light conditions forthe other three species. For S. halepense, this was at the expense of the area ofepidermis, which was proportionally lower in leaves exposed to full sun thanin leaves grown under moderate light level, while in the two C3 grasses bothmesophyll and epidermis areas tended to be lower under field conditions thanin the growth room.
Leaf morphology
Except for C. gryllus and C. epigeios, leaf area was significantly greater inthe growth room than in natural habitats (Fig. 2A). This was because each ofthe four species possessed significantly longer leaves in moderate than under
LEAF STRUCTURAL RESPONSE OF INVASIVE GRASSES 105
Acta Bot. Hung. 50, 2008

106 HAN, L. J., MOJZES, A. and KALAPOS, T.
Acta Bot. Hung. 50, 2008
Tabl
e2
Qua
ntita
tive
light
mic
rosc
opic
leaf
anat
omic
alch
arac
teri
stic
s(m
ean
±1
SE,n
=5)
for
six
gras
ses
ofdi
ffer
entp
hoto
synt
hetic
path
way
type
and
capa
city
fora
rea
expa
nsio
nde
velo
ped
inth
efie
ldin
full
sun
sem
iari
dha
bita
toru
nder
vari
able
mod
erat
elig
htan
dgo
odsu
p-pl
yof
wat
eran
dnu
trie
nts
ina
grow
thro
om.F
orea
chsp
ecie
s,st
atis
tical
lysi
gnifi
cant
diff
eren
ces
(p<
0.05
)bet
wee
ngr
owth
cond
ition
sar
ein
bold
type
.For
each
vari
able
,sig
nific
anti
nter
spec
ific
diff
eren
ces
with
ina
give
ngr
owth
cond
ition
are
indi
cate
das
inFi
gure
1
Phot
osyn
thet
icty
pean
dsp
read
ing
capa
city
Var
iabl
eG
row
thco
ndit
ions
C3
expa
ndin
gC
4no
n-in
vasi
veC
4in
vasi
veB.
iner
mis
C.e
pige
ios
C.g
ryllu
sB.
isch
aem
umS.
hale
pens
eC
.dac
tylo
n
Thic
knes
sof
mes
ophy
ll(µ
m)
Gro
wth
room
117.
8±2.
8a
108.
6±9.
2a
Fiel
d10
9.1±
4.5
A15
0.2±
9.8
B
epid
erm
is(µ
m)
Gro
wth
room
17.8
±0.5
c13
.5±0
.4b
13.1
±0.5
b15
.0±0
.7b
17.8
±0.8
c10
.1±0
.4a
Fiel
d17
.8±1
.9C
14.7
±0.6
B17
.7±0
.9C
22.8
±0.8
D22
.9±1
.3D
10.4
±0.5
A
bund
lesh
eath
(µm
)G
row
thro
om17
.3±1
.0d
12.5
±0.6
bc10
.2±0
.6a
12.1
±0.3
b14
.0±0
.7c
28.5
±1.5
eFi
eld
13.9
±0.8
C14
.0±0
.6C
11.7
±0.6
B9.
5±0.
3A
14.5
±0.8
C21
.7±1
.2D
Inte
rven
iald
ista
nce
(µm
)G
row
thro
om27
5.8±
10.7
d28
3.1±
19.2
d67
.8±2
.7a
65.4
±1.7
a10
8.1±
5.2
b13
0.3±
6.9
cFi
eld
220.
5±10
.5E
265.
0±8.
6F
80.0
±3.7
B65
.4±4
.1A
108.
0±7.
0C
143.
2±7.
4D
Are
apr
opor
-ti
onof
mes
ophy
ll(%
)G
row
thro
om59
.2±2
.6b
56.0
±1.3
ab54
.4±0
.8ab
50.7
±0.8
a58
.7±0
.8b
56.3
±1.7
abFi
eld
54.5
±1.1
B53
.1±0
.9A
B57
.3±0
.6C
51.1
±1.4
A58
.3±0
.6C
51.4
±0.9
Ave
ins
plus
schl
eren
chym
a(%
)
Gro
wth
room
8.7±
0.5
a13
.8±0
.5b
17.8
±1.5
c8.
6±1.
0a
6.3±
1.1
a13
.0±1
.1b
Fiel
d15
.4±0
.8BC
18.5
±1.5
D16
.6±1
.4C
D9.
5±0.
9A
12.8
±0.7
B13
.1±0
.8B
epid
erm
is(%
)G
row
thro
om32
.1±2
.7ab
30.2
±1.1
ab27
.8±1
.3a
40.7
±1.6
c35
.0±1
.6b
30.7
±2.0
abFi
eld
30.1
±1.6
B28
.4±1
.0A
B26
.1±1
.3A
39.4
±1.0
D28
.9±0
.9A
B35
.4±0
.5C

LEAF STRUCTURAL RESPONSE OF INVASIVE GRASSES 107
Acta Bot. Hung. 50, 2008
0
18
36
54
72
Leaf
are
a(c
m2)
Field Growth room
B
bc
*
Cabc
Babc
Aab
C
c
*A
**
a
0
18
36
54
72
Le
af
length
(cm
)
C
c
*
E
d
*
DE
d
B
b
*
D
d
*
A
a
*
0
5
10
15
20
Le
af
wid
th(m
m)
B. inermis C.epigeios C. gryllus B.ischaemum S.halepense C.dactylon
DE c
D b
C
a
*
A a
*
E
d
*
B a
C3expanding C4non-invasive C4invasive
A
B
C
Fig. 2. Leaf morphology for six grasses of different photosynthetic pathway type and capac-ity for area expansion developed in the field in full sun semiarid habitat or under variablemoderate light and good supply of water and nutrients in a growth room. A) Leaf area(cm2); B) leaf length (cm); C) leaf width (mm). For each variable mean ± 1 SE of 5 replicates
are shown. Significant differences are indicated as in Figure 1

high-light conditions (Fig. 2B), but for S. halepense and B. ischaemum, signifi-cantly greater leaf width also contributed to the variation in total leaf size (Fig.2C). Among species, B. inermis exhibited the greatest differences in leaf areaand length between the two light regimes (Table 1). Chrysopogon gryllus alsoproduced significantly longer, but narrower leaves in the growth room, result-ing in leaf area similar to that developed in the field. Neither the length nor thewidth of leaves showed significant differences between light conditions for C.epigeios. Under moderate light level, S. halepense tended to have larger leavesthan the other species; however, differences were significant in comparisonwith B. ischaemum and C. dactylon only.
DISCUSSION
Leaf responses to marked change in growth environment were heteroge-neous in morphology, coarse structure and anatomy, and within groups spe-cies often responded differently, that for certain variables makes difficult todraw unambiguous conclusions to the group as a whole. However, this alsoimplies an existing diversity in the mechanism of leaf structural responses toaltered growth environment even within a plant functional group (i.e. peren-nial grasses). In line with our hypothesis (H1), among C4 grasses the invasive S.halepense and C. dactylon exhibited a higher degree of plasticity in leaf coarsestructure between contrasting environments than the non-invasive C. gryllusand B. ischaemum. Both C4 invaders altered substantially leaf coarse structure(LMA and thickness) in response to moderate light environment, whereasboth traits remained invariable for the non-invasive C. gryllus, and LMAchanged much less for B. ischaemum. This is in line with prior evidence on theimportant role of leaf mass per area and its plasticity in the performance andsuccess of certain invasive exotic or expanding native species (Gloser andGloser 1996, Juraimi et al. 2004, McDowell 2002, Smith and Knapp 2001, Wil-liams et al. 1995). Leaf bulk tissue density appears to be the least responsive tosuch environmental change irrespective of the species’ invasion capacity. Forthe invasive S. halepense, thicker leaves in the field than in the growth roommust have resulted from a greater proportion of veins plus sclerenchyma, thatindicates larger investment into vascular and support tissues under temporalwater shortage and mechanical stress caused by wind in the natural habitat.Knapp and Gilliam (1985) also reported a greater area of the bundle sheath-primary vascular bundle complex associated with greater leaf thickness andleaf mass per area for Andropogon gerardii in the high-light (burned) environ-ment compared to the light-limited, unburned site in a tallgrass prairie. How-
108 HAN, L. J., MOJZES, A. and KALAPOS, T.
Acta Bot. Hung. 50, 2008

ever, no such investment was observed for the other invasive C4 grass, C. dac-tylon and for the two non-invasive C4 grasses in our study.
Leaf morphology responded markedly to the change in growth environ-ment, although the response was not consistently different between invasiveand non-invasive C4 species. The greatest change in leaf length and width oc-curred for the invasive S. halepense closely followed by the two non-invasivespecies, while the invasive C. dactylon produced the smallest difference. Grassleaves usually grow larger, longer but narrower under moderate light inten-sity than in unshaded conditions (Langer 1979). Among C4 grasses in ourstudy, both invasive species and the non-invasive B. ischaemum developedlarger leaves in the growth room than in the field, that were longer, but notnarrower (wider instead for S. halepense and B. ischaemum). Only C. gryllus pro-duced narrower leaves in the growth room than in the original grassland, butsimultaneous increase in leaf length compensated for it, thus leaf area did notchange for this species.
Leaf anatomy was less responsive than morphology or coarse structurefor the grasses studied, and no clear difference appeared between invasiveand non-invasive C4 species. Most leaf anatomical traits varied for the non-in-vasive C. gryllus, but the magnitude of response (5.3–34.7%) remained belowthat of the fewer anatomical variables changing significantly for the two inva-sive C4 grasses (9.6–103.3%). This suggests some anatomical adjustment abilityfor the non-invasive C. gryllus as well. Considering that anatomical changes ofthe other non-invasive grass, B. ischaemum was comparable with that of the in-vasive C. dactylon and S. halepense, invasives do not appear to exceed non-invasives in leaf anatomical response to the environment, at least for the C4grasses studied here. Thinner parenchymatous bundle sheath cell layer inleaves of C. gryllus in the growth room than in the field may partly be responsi-ble for shorter distances between veins in the growth room. In contrast, veindensity remained unchanged for the other C4 grasses in this study under bothlight conditions. For three of the four C4 species studied, thicker epidermis inthe field than in the growth room may provide greater protection againsthigher transpirational water loss, wind damage and wind-carried dust parti-cles in natural environment (Mauseth 1988). Plasticity of leaf traits for the na-tive B. ischaemum appeared to be intermediate between that of the non-inva-sive C. gryllus and the two invasive C4 species. This may be associated with thespecies’ ability to reach local dominance mainly in disturbed grassland vege-tation (Zólyomi and Fekete 1994). Among the two C3 species of high spreadingcapacity in this study, B. inermis showed particularly high responsiveness thatappears to be consistent with the species’ forest edge phytocoenological affin-ity (Szujkó-Lacza and Rajczy 1986, Zólyomi and Fekete 1994). Phenotypicplasticity in photosynthesis and leaf coarse structure has been documented to
LEAF STRUCTURAL RESPONSE OF INVASIVE GRASSES 109
Acta Bot. Hung. 50, 2008

be greater for species adapted to spatially or temporally heterogeneous or un-predictable environments (e.g. for early or middle successional species) thanfor those from more homogeneous and stable environments (e.g. for late suc-cessionals, Abrams and Mostoller 1995, Strauss-Debenedetti and Bazzaz 1991,Yamashita et al. 2000). Furthermore, early or mid-successional status of a spe-cies often couples with high potential for invasiveness (Grace et al. 2001,Yamashita et al. 2000, 2002). Thus, the flexibility of leaf morphology and struc-ture may also help B. inermis to successfully invade temperate grasslands inNorth America (Grace et al. 2001, Grilz and Romo 1994, Willson and Stubben-dieck 2000).
Our results do not support the hypothesis of greater ability for structuraladjustment to the growth environment for C3 than for C4 grasses (H2), a com-parison we could make for species of high capacity to spread. Overall, therewere similar changes in leaf morphology, coarse structure and anatomy in re-sponse to the growth environment, although certain variables behaved differ-ently in the two groups. Leaf intervenial distance was the most prominentvariable of this sort, which showed less variation among C4 than C3 species,most probably due to the strong need of close proximity of mesophyll andbundle sheath cells in C4 photosynthesis that may restrict the ability of shadedC4 plants to increase vein spacing (Sage and McKown 2006). Albeit leaf coarsestructure changed at a similar or greater extent for the two C3 species than forC4 invasives, in leaf morphology and anatomy the responsiveness of the C3 B.inermis and the C4 S. halepense was similarly high, while that of the C3 C. epigeiosand the C4 C. dactylon was comparably moderate. Thus, based on these dataneither C3 nor C4 grasses can be qualified as more plastic in leaf anatomy andmorphology. Similar to these results, no consistent differences were found inother comparisons of C3 and C4 species in the ability to reduce leaf thickness inresponse to shading (Sage and McKown 2006). Shorter intervenial distanceswe observed here for C4 grasses than for C3 grasses, and for C4 NADP-ME spe-cies than for the C4 NAD-ME C. dactylon are consistent with previous results(Dengler et al. 1994, Kawamitsu et al. 1985, Ogle 2003).
In conclusion, among C4 grasses studied here invasive species appear topossess greater leaf coarse structural, but not consistently higher morphologi-cal and anatomical plasticity between contrasting light environments if com-pared with non-invasive ones. A higher plasticity may contribute to the suc-cessful establishment and persistence of invasive C4 grasses in heterogeneouslight environments. Ecophysiological plasticity also proved to play an impor-tant role in the invasion success of some tropical and subtropical species(Pattison et al. 1998, Williams et al. 1995, Yamashita et al. 2000). However, ourresults do not support the hypothesis that among grasses of high capacity forcolonisation C4 species are inferior to C3 species in the capacity of adjusting
110 HAN, L. J., MOJZES, A. and KALAPOS, T.
Acta Bot. Hung. 50, 2008

leaf morphology and structure to the environment. Nevertheless, further stud-ies involving a greater number of species are necessary to reach more generalconclusions.
*
Acknowledgements – Support from the Hungarian Scientific Research Fund (OTKAT038028, W15301) is acknowledged. This work is a result of the first author’s visit to Hun-gary supported by the intergovernmental scientific exchange program between China andHungary (administered by the Hungarian Scholarship Board).
REFERENCES
Abrams, M. D. and Mostoller, S. A. (1995): Gas exchange, leaf structure and nitrogen in con-trasting successional tree species growing in open and understory sites during adrought. – Tree Physiol. 15: 361–370.
Boardman, N. K. (1977): Comparative photosynthesis of sun and shade plants. – Ann. Rev.Plant Physiol. 28: 355–377.
Bredenkamp, G. J., Spada, F. and Kazmierczak, E. (2002): On the origin of northern andsouthern hemisphere grasslands. – Plant Ecol. 163: 209–229.
Burke, M. J. W. and Grime, J. P. (1996): An experimental study of plant community in-vasibility. – Ecology 77(3): 776–790.
Chapman, G. P. (1996): The biology of grasses. – CAB International, Wallingford, 273 pp.Dengler, N. G., Dengler, R. E., Donnelly, P. M. and Hattersley, P. W. (1994): Quantitative
leaf anatomy of C3 and C4 grasses (Poaceae): bundle sheath and mesophyll surfacearea relationships. – Ann. Bot. 73: 241–255.
Dukes, J. S. and Mooney, H. A. (1999): Does global change increase the success of biologicalinvaders? – Trends Ecol. Evol. 14(4): 135–139.
Fekete, G. and Szujkó-Lacza, J. (1973): Leaf anatomical and photosynthetical reactions ofQuercus pubescens Willd. to environmental factors in various ecosystems. I. Leaf ana-tomical reactions. – Acta Bot. Acad. Sci. Hung. 18(1–2): 59–89.
Fekete, G., Virágh, K., Aszalós, R. and Précsényi, I. (2000): Static and dynamic approaches tolandscape heterogeneity in the Hungarian forest-steppe zone. – J. Veg. Sci. 11: 375–382.
Givnish, J. T. (1988): Adaptation to sun and shade: a whole-plant perspective. – Austral. J.Plant Physiol. 15: 63–92.
Gloser, V. and Gloser, J. (1996): Acclimation capability of Calamagrostis epigejos and C.arundinacea to changes in radiation environment. – Photosynthetica 32(2): 203–212.
Grace, J. B., Smith, M. D., Grace, S. L., Collins, S. L. and Stohlgren, T. J. (2001): Interactions be-tween fire and invasive plants in temperate grasslands of North America. – In: Galley, K. E.M. and Wilson, T. P. (eds): Proceedings of the Invasive Species Workshop: the Role ofFire in the Control and Spread of Invasive Species. Fire Conference 2000: the First Na-tional Congress on Fire Ecology, Prevention and Management. Miscellaneous Publi-cation, No. 11. Tall Timbers Research Station, Tallahassee, pp. 40–65.
Grilz, P. L. and Romo, J. T. (1994): Water relations and growth of Bromus inermis Leyss(smooth brome) following spring or autumn burning in a fescue prairie. – Amer. Midl.Nat. 132: 340–348.
LEAF STRUCTURAL RESPONSE OF INVASIVE GRASSES 111
Acta Bot. Hung. 50, 2008

Holm, L. G., Plucknett, D. L., Pancho, J. V. and Herberger, J. P. (1977): The world’s worstweeds. Distribution and biology. – The University Press of Hawaii, Honolulu, 609 pp.
Juraimi, A. S., Drennan, D. S. H. and Anuar, N. (2004): The effects of shading on the growth,development and partitioning of biomass in Bermudagrass (Cynodon dactylon (L.)Pers.). – J. Biol. Sci. 4(6): 756–762.
Kawamitsu, Y., Hakoyama, S., Agata, W. and Takeda, T. (1985): Leaf interveinal distancescorresponding to anatomical types in grasses. – Plant Cell Physiol. 26(3): 589–593.
Knapp, A. K. and Gilliam, F. S. (1985): Response of Andropogon gerardii (Poaceae) to fire-induced high vs. low irradiance environments in tallgrass prairie: leaf structure andphotosynthetic pigments. – Amer. J. Bot. 72(11): 1668–1671.
Kolar, C. S. and Lodge, D. M. (2001): Progress in invasion biology: predicting invaders. –Trends Ecol. Evol. 16(4): 199–204.
Kovács-Láng, E., Kröel-Dulay, Gy., Kertész, M., Fekete, G., Bartha, S., Mika, J.,Dobi-Wantuch, I., Rédei, T., Rajkai, K. and Hahn, I. (2000): Changes in the composi-tion of sand grassland along a climatic gradient in Hungary and implication for cli-mate change. – Phytocoenologia 30(3–4): 385–407.
Lambers, H., Chapin III, F. S. and Pons, T. L. (1998): Plant physiological ecology. – SpringerVerlag, New York, 540 pp.
Langer, R. H. M. (1979): How grasses grow. 2nd ed. – Edward Arnold (Publishers) Limited,London, 66 pp.
Mauseth, J. D. (1988): Plant anatomy. – The Benjamin/Cummings Publishing Company, Inc.,California, 560 pp.
McDowell, S. C. L. (2002): Photosynthetic characteristics of invasive and noninvasive spe-cies of Rubus (Rosaceae). – Amer. J. Bot. 89(9): 1431–1438.
Mendes, M. M., Gazarini, L. C. and Rodrigues, M. L. (2001): Acclimation of Myrtus com-munis to contrasting Mediterranean light environments – effects on structure andchemical composition of foliage and plant water relations. – Environ. Exp. Bot. 45:165–178.
Mojzes, A., Kalapos, T. and Virágh, K. (2005): Leaf anatomical plasticity of Brachypodiumpinnatum (L.) Beauv. growing in contrasting microenvironments in a semiarid loessforest-steppe vegetation mosaic. – Community Ecol. 6(1): 49–56.
Molnár, E., Bagi, I., Csintalan, Zs. and Nyakas, A. (2000): The invasion success of a native grass-land species in the Great Hungarian Plain. – In: Ferenciková, D., Gáborcik, N., Ondrásek,L., Uhlrarová, E. and Zimková, M. (eds): Grassland ecology V. Proceedings of the 5thEcological Conference, 23–25 Nov 1999, Grassland and Mountain Agriculture Re-search Institute, Banská Bystrica, pp. 423–433.
Ogle, K. (2003): Implications of intervenial distance for quantum yield in C4 grasses: a mod-elling and meta-analysis. – Oecologia 136: 532–542.
Oguchi, R., Hikosaka, K. and Hirose, T. (2003): Does the photosynthetic light-acclimationneed change in leaf anatomy? – Plant Cell Environ. 26: 505–512.
Pattison, R. R., Goldstein, G. and Ares, A. (1998): Growth, biomass allocation and photosyn-thesis of invasive and native Hawaiian rainforest species. – Oecologia 117: 449–459.
Pearcy, R. W. and Ehleringer, J. (1984): Comparative ecophysiology of C3 and C4 plants. –Plant Cell Environ. 7: 1–13.
Radford, I. J. and Cousens, R. D. (2000): Invasiveness and comparative life-history traits ofexotic and indigenous Senecio species in Australia. – Oecologia 125: 531–542.
Rebele, F. and Lehmann, C. (2001): Biological flora of Central Europe: Calamagrostisepigejos (L.) Roth. – Flora 196: 325–344.
112 HAN, L. J., MOJZES, A. and KALAPOS, T.
Acta Bot. Hung. 50, 2008

Rejmánek, M. and Richardson, D. M. (1996): What attributes make some plant species moreinvasive? – Ecology 77(6): 1655–1661.
Sage, R. F. and McKown, A. D. (2006): Is C4 photosynthesis less phenotypically plastic thanC3 photosynthesis? – J. Exp. Bot. 57(2): 303–317.
Sims, D. A. and Pearcy, R. W. (1992): Response of leaf anatomy and photosynthetic capacityin Alocasia macrorrhiza (Araceae) to a transfer from low to high light. – Amer. J. Bot.79(4): 449–455.
Smith, M. D. and Knapp, A. K. (2001): Physiological and morphological traits of exotic, in-vasive exotic, and native plant species in tallgrass prairie. – Int. J. Plant Sci. 162(4):785–792.
StatSoft Inc. (2004): Statistica (data analysis software system), version 7.0. – StatSoft Inc., Tulsa.Strauss-Debenedetti, S. and Bazzaz, F. A. (1991): Plasticity and acclimation to light in tropi-
cal Moraceae of different successional positions. – Oecologia 87: 377–387.Szujkó-Lacza, J. and Rajczy, M. (1986): Similarity investigations on a loess steppe fragment
in a forest-steppe in Hungary. – Annls hist.-nat. Mus. natn. Hung. 78: 49–62.White, T. A., Campbell, B. D. and Kemp, P. D. (2001): Laboratory screening of the juvenile
responses of grassland species to warm temperature pulses and water deficits to pre-dict invasiveness. – Funct. Ecol. 15: 103–112.
Williams, D. G., Mack, R. N. and Black, R. A. (1995): Ecophysiology of introduced Pennise-tum setaceum on Hawaii: the role of phenotypic plasticity. – Ecology 76(5): 1569–1580.
Willson, G. D. and Stubbendieck, J. (2000): A provisional model for smooth brome manage-ment in degraded tallgrass prairie. – Ecol. Rest. 18(1): 34–38.
Yamashita, N., Koike, N. and Ishida, A. (2002): Leaf ontogenetic dependence of light accli-mation in invasive and native subtropical trees of different successional status. – PlantCell Environ. 25: 1341–1356.
Yamashita, N., Ishida, A., Kushima, H. and Tanaka, N. (2000): Acclimation to sudden in-crease in light favoring an invasive over native trees in subtropical islands, Japan. –Oecologia 125: 412–419.
Zólyomi, B. and Fekete, G. (1994): The Pannonian loess steppe: differentiation in space andtime. – Abstracta Botanica 18(1): 29–41.
LEAF STRUCTURAL RESPONSE OF INVASIVE GRASSES 113
Acta Bot. Hung. 50, 2008