research article an oral vaccine against candidiasis
TRANSCRIPT

RESEARCH ARTICLE
An oral vaccine against candidiasis generated by a yeast moleculardisplay systemSeiji Shibasaki1, Wataru Aoki2, Takashi Nomura3, Ayuko Miyoshi1, Senji Tafuku3, Tomomitsu Sewaki3 &Mitsuyoshi Ueda2
1 General Education Center and Gradudate School of Pharmacy, Hyogo University of Health Sciences, Kobe, Japan
2 Division of Applied Life Sciences, Graduate School of Agriculture, Kyoto University, Kyoto, Japan
3 Genolac BL Corporation, Okinawa, Japan
This paper deals with the development of an oral vaccine against Enolase 1 (Eno1p) of Candida albicans. The approachtaken by using cell surface engineering of Saccharomyces cerevisiae is original and a proper example of how modern oralvaccines can be produced.
Keywords
Eno1p; Candida albicans; Saccharomyces
cerevisiae; cell surface engineering.
Correspondence
Seiji Shibasaki, General Education Center
and Gradudate School of Pharmacy,
Hyogo University of Health Sciences,
1-3-6 Minatojima, Chuo-ku, Kobe 650-8530,
Japan.
Tel.: +81 78 304 3159
fax: +81 78 304 2859
e-mail: [email protected]
Received 4 May 2013; revised 4 June 2013;
accepted 14 July 2013. Final version
published online 14 August 2013.
doi:10.1111/2049-632X.12068
Editor: Willem van Eden
Abstract
Enolase 1 (Eno1p) of Candida albicans is an immunodominant antigen. However,conventional technologies for preparing an injectable vaccine require purificationof the antigenic protein and preparation of an adjuvant. To develop a novel type oforal vaccine against candidiasis, we generated Saccharomyces cerevisiae cellsthat display the Eno1p antigen on their surfaces. Oral delivery of the engineeredS. cerevisiae cells prolonged survival rate of mice that were subsequentlychallenged with C. albicans. Given that a vaccine produced using moleculardisplay technology avoids the need for protein purification, this oral vaccine offersa promising alternative to the use of conventional and injectable vaccines forpreventing a range of infectious diseases.
Introduction
Candidiasis is a serious infectious disease caused by thefungus Candida albicans and other Candida species (Nucci& Marr, 2005; Pfaller & Diekema, 2007). Superficial orsystemic candidiasis is observed when the host immunityis compromised by AIDS, chemotherapies for cancertreatment or the administration of immunosuppressants.Together, C. albicans and Candida glabrata are responsiblefor 70–80% of Candida infections in patients with candidiasisor candidemia. In general, options for treating candidiasisremain unsatisfactory because of delayed diagnosis and alack of reliable tools to detect Candida species inside thebody of a patient (Noble & Johnson, 2007).Pharmacotherapy of candidiasis often involves the admin-
istration of caspofungin, micafungin, anidulafungin and
amphotericin B. However, mutants of Candida with reducedsusceptibility to these drugs have emerged (Rodloff et al.,2011). Although genetic and molecular mechanisms thatconfer resistance of C. albicans to antifungal drugs havebeen studied for a long time (Morschh€auser, 2002), atpresent, there is no general strategy to prevent the emer-gence of resistance to antifungal drugs. In addition,unwanted side effects of chemical drugs against Candidaspecies as well as other fungi also pose a serious problem(Benko et al., 1999). Therefore, prevention of infection byvaccination against Candida species is thought to be animportant complementary strategy to pharmacotherapy inefforts to control candidiasis.Vaccination with Eno1 protein (Eno1p) of C. albicans,
which encodes the glycolytic enzyme enolase 1 (2-phospho-D-glycerate hydrolyase), can protect against
Pathogens and Disease (2013), 69, 262–268, © 2013 Federation of European Microbiological Societies. Published by John Wiley & Sons Ltd. All rights reserved262
Pathogens and Disease ISSN 2049-632X

infection with C. albicans (Montagnoli et al., 2004; Li et al.,2011). Besides Eno1p, the C. albicans proteins, e.g. hyphalwall protein, glyceraldehyde-3-phosphate dehydrogenaseand phosphoglycerate kinase can also induce protectiveimmune functions against candidiasis when administeredwith appropriate adjuvant compounds to enhance theimmune response to the antigen (Xin & Cutler, 2011).However, the technologies available to purify antigenicproteins, prepare adjuvant, and deliver both in a convenientand noninvasive manner pose a challenge to vaccinedevelopment.In recent years, we have developed the use of molecular
display technology, which uses genetically engineeredmicroorganisms to produce foreign proteins (Shibasakiet al., 2007). In general, the approach involves fusion of aheterologous protein, such as an antigen, to a cell-wallprotein, thereby anchoring the hybrid protein to the cellsurface and exposing the heterologous protein to theexterior of the cell. The use of molecular display to preparetarget proteins is especially well established for use with theyeast Saccharomyces cerevisiae, and it is also termed cellsurface engineering (Shibasaki et al., 2009). For instance,an antigen from red sea bream iridovirus (RSIV) wasdisplayed on the surface of yeast cells to develop an oralvaccine for use by fisheries (Tamaru et al., 2006). A majorcomponent of the yeast cell wall, b-glucan, is expected toserve as an adjuvant (Rodr�ıguez et al., 2009), thus obviat-ing the need to prepare an adjuvant for codelivery. Yeastsare generally recognized as safe (GRAS) organisms, andthus appropriate for the preparation of oral vaccines withoutthe need for antigen purification, as is required forrecombinant proteins produced by Escherichia coli. Anadvantage of systems that use molecular display is that theyprovide antigenic proteins more rapidly and convenientlythan conventional approaches to produce vaccines.Here, we show that the antigens displayed on yeast
generated using molecular display technology offer promiseas a novel type of oral vaccine against candidiasis, whichserves as a model of infectious disease. The Eno1p proteinfrom C. albicans was selected as the model antigenicprotein to be displayed on the surface of S. cerevisiae cells.Delivery of the cells to mice by oral or intranasal adminis-tration, or by subcutaneous injection, increased the titers ofanti-Eno1p antibodies and prolonged the lives of miceinfected with lethal levels of C. albicans. These resultssuggest that the yeast molecular display system mightprovide a convenient tool to protect against various infec-tious diseases.
Materials and methods
Strains and media
The E. coli strain DH5a [F�endA1 hsdR17 (rK�, mK
+)supE44 thi1 recA1 gyrA96 DlacU169 ðφ80 lacZDM15](Hanahan, 1983) was used as a host for manipulation ofrecombinant DNA. The E. coli strain BL21 [F� ompT hsdSB(rB
�, mB�) gal dcm (DE3)] was used to produce antigenic
proteins. Both E. coli strains were grown in Luria–Bertani
(LB) medium [1% (w/v) tryptone, 0.5% (w/v) yeast extract,0.5% (w/v) NaCl and 0.1% (w/v) glucose]. The S. cerevisiaestrain BY4741 (MATa his3-1 leu2 met15 ura3) was used forcell surface display of antigenic proteins. YPD medium [1%(w/v) yeast extract, 2% (w/v) polypeptone and 2% (w/v)glucose] was used for the transformation of yeast cells.Yeast cells that carried a plasmid were grown in SDCmedium [2% (w/v) glucose, 0.67% (w/v) yeast nitrogen basewithout amino acids, 1% (w/v) casamino acids and supple-mented with appropriate amino acids].
Plasmid construction and transformation of microbialcells
The plasmid pQE-eno1, which was used to produceEno1p in E. coli, was constructed by first using PCR togenerate the Eno1p-encoding sequence using primers5′-ATGGATCCTCTTACGCCACTAAAATCCACGCC-3′ and5′-TAAAGCTTTTACAATTGAGAAGCCTTTTGGAAATCTT-TACC-3′ and genomic DNA of C. albicans strain SC5314(American Type Culture Collection) (Hube et al., 1997),which was purified as described previously (Aoki et al.,2011). The fragment of Eno1p-encoding sequence wasinserted into the pQE30 plasmid (Qiagen, Hilden, Germany)that had been digested with BamHI and HindIII to constructthe recombinant plasmid pQE-eno1.The plasmid pULD-eno1, which was used to display
Eno1p from C. albicans on the surface of S. cerevisiaecells, was constructed by amplifying the Eno1p-encodingsequence by PCR using primers 5′-ACGCGGCCGCTCTT-ACGCCACTAAAATCCACGCC-3′ and 5′- TGCTCGAGCA-ATTGAGAAGCCTTTTGGAAATCTTTACC-3′ and the samegenomic DNA of C. albicans described above. The fragmentof the gene encoding ENO1 was inserted into plasmidpULD1 (Kuroda et al., 2009) that had been digested withNotI and XhoI. Thus, the Eno1p-encoding sequence wasfused to the 5′ end of the cell-wall anchoring protein,a-agglutinin-encoding sequence in the plasmid. The con-structed plasmid pQE-eno1 was introduced into E. coli BL21as described previously (Hanahan, 1983) for propagation.The constructed plasmid pULD1-eno1 and parent plasmidpULD1 were introduced into S. cerevisiae BY4741 using thelithium acetate method (Ito et al., 1983) for surface displayof the protein. The nucleotide sequence of all constructedplasmids was confirmed by using an ABI PRISM 3130Genetic Analyzer (Applied Biosystems, Foster City, CA).
Protein production and purification
Expression of the pQE30-eno1 plasmid in E. coli BL21(DE3) produced Eno1p as a fusion protein with anN-terminal His6 tag. The cells were inoculated in 14 mLLB medium, containing 100 lg L�1 ampicillin, and grownin shaken flasks overnight at 37 °C. Fresh LB mediumcontaining 100 lg L�1 ampicillin (120 mL) was inoculatedwith 6 mL of the overnight cultures and the cells weregrown at 37 °C to an OD600 of 0.6–1.0. Gene expressionwas then induced by the addition of isopropyl b-D-thio-galactoside (IPTG; Wako Pure Chemical, Osaka, Japan)
Pathogens and Disease (2013), 69, 262–268, © 2013 Federation of European Microbiological Societies. Published by John Wiley & Sons Ltd. All rights reserved 263
S. Shibasaki et al. Yeast-based oral vaccine against candidiasis

to a final concentration of 1 mM. After 3–4 h of cultivationat 37 °C, the cell cultures were harvested by centrifugation(4000 g, 20 min, 4 °C). The cell pellets were subsequentlyresuspended in 10 mL of B-PER bacterial protein extrac-tion reagent (Thermo Fisher Scientific, IL), and shakengently for 10 min at room temperature (25 °C). Solubleproteins were separated from insoluble proteins by centri-fugation at 27 000 g for 20 min. Supernatant containingsoluble proteins were purified by recovering the His6-Eno1fusion protein after passage through nickel-chelated aga-rose (Thermo Fisher Scientific) columns. The column wasequilibrated with 10 mL of B-PER Bacterial Protein Extrac-tion Reagent before application of the supernatant. Afterwashing the column with wash buffer of the B-PERreagent, the bound proteins were released with elutionbuffer [50 mM Tris, 300 mM NaCl, 200 mM imidazole,10% (v/v) glycerol]. Endotoxin was removed from theeluate by passage through Detoxi-Gel endotoxin-removingcolumns (Pierce, Rockford, IL), resulting in levels of < 0.1endotoxin units mL�1, as indicated using Limulus Amebo-cyte Lysate (LAL) PYROGENT single-test vials (Lonza,Walkersville, MD) as described in the manufacturer’sprotocol.
Immunofluorescence staining
Yeast cells were collected by centrifugation at 6000 g for5 min, washed with phosphate-buffered saline (PBS; 50 mMphosphate, 150 mMsodium chloride, pH 7.4) and adjusted to3.2 9 108 cells mL�1 with PBS. Thereafter, 200 lL of thiscell suspension was centrifuged at 6000 g for 5 min. Col-lected cells were incubatedwith 200 lL of a 1 : 250 dilution ofrabbit IgG in PBS containing 1% (w/v) BSA at room temper-ature for 1 h (Shibasaki et al., 2007).Surface-blocked cells were incubated with 3 lg mL�1 of
mouse monoclonal antibody against the FLAG tag (Sig-ma-Aldrich, St Louis, MO) in PBS for 1.5 h at roomtemperature. After washing with PBS, the cells wereincubated with 3 lg mL�1 of AlexaFluor488-conjugatedgoat antimouse IgG antibody (Invitrogen, CA) in PBS for1.5 h at room temperature, and then washed again. Fluo-rescence of the yeast cell surface was observed using anOlympus BX51 microscope (Olympus, Tokyo, Japan).Fluorescence units were measured using the SpectraMaxM2 Microplate Reader (Molecular Devices, CA) with
107
108
106
Seru
m Ig
G ti
ter
(log 10
)
105
104
103
102
80
100
Surv
ival
rat
io (%
)
20
40
60
Time (day)
00 10 20 30
040
(a)
(b)
Fig. 1 Subcutaneous administration of the recombinant antigens with
IFA (incomplete Freund’s adjuvant). (a) Antibody response (P < 0.01).
(b) Survival test. Closed circles, administration of Eno1p; closed
triangles, administration of PBS.
107
106
Seru
m Ig
G ti
ter
(log 10
)
105
104
103
102
80
100
Surv
ival
rat
io (%
)
20
40
60
Time (day)
00 10 20 30 40
(a)
(b)
Fig. 2 Intranasal administration of the recombinant antigens with CT
(cholera toxin). (a) Antibody response (P < 0.05). (b) Survival test.
Closed circles, administration of Eno1p; closed triangles, administration
of PBS.
Pathogens and Disease (2013), 69, 262–268, © 2013 Federation of European Microbiological Societies. Published by John Wiley & Sons Ltd. All rights reserved264
Yeast-based oral vaccine against candidiasis S. Shibasaki et al.

excitation and emission wavelengths of 495 and 519 nm,respectively.
Animals
Female C57BL/6 mice were obtained from Japan SLC, Inc.(Shizuoka, Japan). Mice were maintained in a specific-path-ogen-free manner and allowed to drink and eat ad libitum.
Immunization by intranasal or subcutaneousadministration using recombinant proteins
Seven-week-old female C57BL/6mice (eight per group) wereimmunized with 30 lg of E. coli-expressed recombinantEno1 protein by intranasal delivery (i.n.) or subcutaneousinjection (s.c.), using it in 20-lL volumes containing 1 lg ofcholera toxin (Sigma-Aldrich) (i.n.) or 100-lL volumes con-taining Freund’s incomplete adjuvant (DIFCO Laboratories,Detroit, MI) (s.c.). Eight female C57BL/6 mice of the sameage, which received adjuvant alone mixed with PBS, servedas a control group. Mice were immunized at weeks 0, 2 and 4.Blood samples were collected at week 6 from the tail vein fordetermination of the titer of serum IgG that binds to Eno1p.
Immunization by oral administration using yeast cellsthat display Eno1p on their surfaces
Seven-week-old female C57BL/6 mice were used forimmunization experiments. A solution of S. cerevisiae cellsthat display Eno1p on their surfaces (1.6 9 109 cells/400 lL) were administered to 10 mice per dose at weeks
0, 1 and 3 (priming) and at week 7 (booster). Wild-typeS. cerevisiae was used as a control. All inoculums weresuspended in PBS (400 lL per animal) and administered viaan intragastric tube after 2 h of fasting, once per day for5 days per week. Blood samples were collected at week 9from the tail vein to determine the titer of serum IgG.
Challenge with C. albicans
For survival studies, mice were infected with 1.1 9 105 cellsof C. albicans resuspended in 100 lL PBS by tail-veininjection 2 weeks after the last immunization. Mice wereobserved daily for 4 weeks after challenge. All animalexperimental protocols were approved by the InstitutionalAnimal Care and Use Committee, and animal experimentswere conducted according to the institutional ethical guide-lines for animal experiments. The outcomes of C. albicanschallenge were shown using the Kaplan-Meier method.Significant differences between groups were tested usingthe log-rank method.
Determination of endpoint titer
Indirect enzyme-linked immunosorbent assay (ELISA) wasconducted for antibody analysis for antisera collected at week6 (i.n. or s.c.) or week 9 (p.o.). Briefly, 96-well microtiter plates(Nalge Nunc International, Rochester, NY) were coated with50 lL per well of E. coli-expressed recombinant Eno1(0.01 lg lL�1). The plates were blocked with 1% bovineserum albumin (BSA) dissolved in PBS containing 0.05%Tween-20. Serially diluted antisera and horseradish peroxi-
(a)
(c) (d)
(b)
Fig. 3 Immunofluorescence microscopic observation of displayed Eno1 on Saccharomyces cerevisiae. (a,b) BY4741 (control cell). (c,d) BY4741
harboring pULD1-eno1. (a,c) Bright-field images. (b,d) fluorescence microscopic images after staining using the anti-FLAG antibody. Scale
bars = 10 lm.
Pathogens and Disease (2013), 69, 262–268, © 2013 Federation of European Microbiological Societies. Published by John Wiley & Sons Ltd. All rights reserved 265
S. Shibasaki et al. Yeast-based oral vaccine against candidiasis
dase (HRP)-conjugated goat antimouse IgG (1 : 4000, Pro-mega, Madison, WI), followed by its substrate, were added towells. After 20 min incubation at room temperature, thereaction was stopped by adding 1 M sulfuric acid, and theabsorbance was measured at 450 nm (A450 ) using a micro-plate reader (Bio-Rad Laboratories Inc., Redmond, WA). Theserum IgG antibody titer was defined either as the serumdilution that gave an A450 value equal to 0.1.
Results and discussion
Extraction of Eno1p from E. coli for s.c. injection andi.n. administration
The use of PCR to amplify the Eno1p-encoding sequencefrom genomic DNA extracted from C. albicans as a templateyielded a fragment of the expected size, which was cloned
into the pQE30 vector that was designed to produceHis-tagged recombinant proteins. After confirmation thatthe nucleotide sequence of the insert within pQE-Eno1 wasconfirmed as correct by comparison with the Candidagenome database (http://www.candidagenome.org/), theplasmid was introduced into E. coli BL21, and the recombi-nant protein was produced in the cytosol, and then recov-ered, and purified by passage of the extract through anickel-chelated column. The recombinant protein was fur-ther purified by passage through an endotoxin-removingcolumn. The concentration of endotoxin in eluate from thenickel-chelated column ranged from 0.06 to 0.125 EU(endotoxin units) mL�1. Analysis of Eno1 proteins usingsodium dodecyl sulfate polyacrylamide gel electrophoresis(SDS-PAGE) and Western blotting indicated that anti-Hisantibody recognized the purified protein (data not shown).
Vaccination by s.c. and i.n. administration of Eno1p
We used both s.c. injection and i.n. administration toexamine the efficacy with which delivery of recombinantEno1 could induce immunity. The ability of vaccination withEno1p to protect against candidiasis was determined bytesting survival after administration of a lethal dose ofC. albicans.Administration of Eno1p with adjuvant by s.c. administra-
tion elevated titers of antibody that bound to Eno1p(Fig. 1a). At 35 days after challenge with a lethal dose ofC. albicans, 12.5% of mice vaccinated with Eno1p werealive, whereas none of the control animals infected with thesame dose of C. albicans SC5314 (Hube et al., 1997)survived longer than 23 days after infection (Fig. 1b).As was observed for s.c. administration, i.n. administra-
tion of Eno1p with adjuvant increased titers of Eno1p-spe-cific antibody (Fig. 2a). However, approximately twice asmany animals (25% compared with 12.5%) survived chal-lenge with C. albicans after i.n. administration of the vaccinecompared with s.c. injection (Fig. 2b). Although our findingthat vaccination with Eno1p can protect against subsequentexposure to C. albicans is consistent with the previousreport that showed the ability of Eno1p to induce immunity toC. albicans in mice (Montagnoli et al., 2004), our findingsreveal for the first time that i.n. administration of Eno1pappears to induce protective immunity against C. albicansmore effectively than s.c. delivery of the protein.
Preparation of Eno1p-displaying yeast for oraladministration
A S. cerevisiae strain that displays Eno1p on its surface wasgenerated by transforming yeast strain BY4741 with thepULD1-eno1 plasmid and confirmation of successful trans-formation with this plasmid by auxotrophic selection andcolony direct PCR. The strain that harbored pULD1-eno1was named BY4741/eno1. Display of Eno1p on the cellsurface of BY4741/eno1 was observed by immunofluores-cence microscopy. Whereas no fluorescence was observedon the surfaces of cells of the parent strain BY4741(Fig. 3a and b), clear fluorescence of AlexaFluor488 was
Seru
m Ig
G ti
ter
(log 10
)
105
104
103
102
80
100
Surv
ival
rat
io (%
)
20
40
60
Time (day)
00 10 20 30 40
(a)
(b)
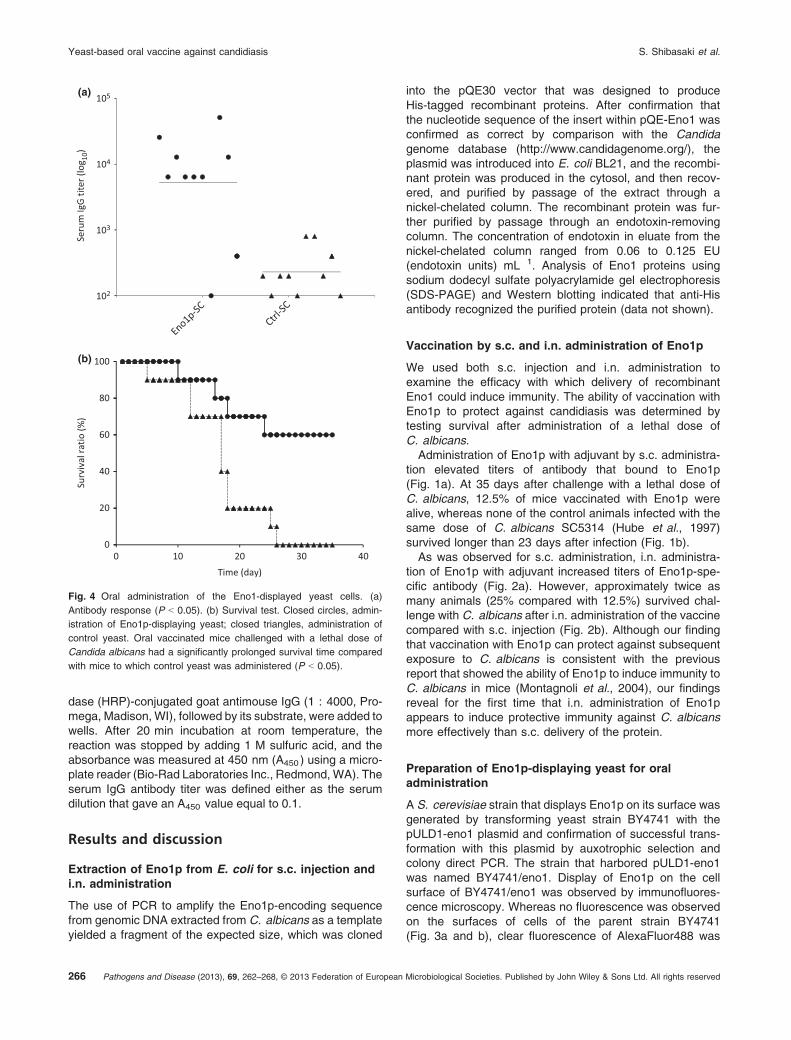
Fig. 4 Oral administration of the Eno1-displayed yeast cells. (a)
Antibody response (P < 0.05). (b) Survival test. Closed circles, admin-
istration of Eno1p-displaying yeast; closed triangles, administration of
control yeast. Oral vaccinated mice challenged with a lethal dose of
Candida albicans had a significantly prolonged survival time compared
with mice to which control yeast was administered (P < 0.05).
Pathogens and Disease (2013), 69, 262–268, © 2013 Federation of European Microbiological Societies. Published by John Wiley & Sons Ltd. All rights reserved266
Yeast-based oral vaccine against candidiasis S. Shibasaki et al.

observed around the surfaces of BY4741/eno1 cells (Fig. 3cand d).To quantify changes in the relative amounts of Eno1p
displayed on the surfaces of yeast cells during cultivation,the fluorescence of yeast cells stained with AlexaFluor488was analysed using a multiwell plate reader. The amounts ofdisplayed Eno1p increased for the first 24 h of the growthcycle (data not shown).
Vaccination by oral administration of yeast cells thatdisplay Eno1p on their surfaces
Cultures used for oral administration of Eno1p-displayingyeast cells to mice were grown for 24 h, given that this wasthe length of culture when cells displayed the largest numberof Eno1p-fusion proteins on their surfaces. Mice werevaccinated by oral administration of the cells four timesover the course of a 7-week period before challenge with alethal dose of C. albicans. The average titer of antibodyagainst Eno1p generated after oral administration of yeastcells that display Eno1p was 5.2 9 103, although the valuevaried substantially between animals, 1 9 102 to 5.2 9 104
(Fig. 4a). Although this dispersion was different from resultsof conventional administration of the antigenic protein(Figs 1a and 2a), the yeast positively gave enough immu-nological stimuli to almost all mice. Examination of survivalrate after challenge with C. albicans for 35 days indicatedthat 60% of mice that received oral administration ofEno1p-displaying cells survived longer than mice thatreceived oral administration of control cells (Fig. 4b). Thissurvival rate is better than that associated with the moreconventional modes of immunogen administration, i.e. s.c.
injection and i.n. administration. These results suggest thatdisplay of a suitable antigen on the surfaces of orallydelivered yeast cells generated by molecular display mightprovide a convenient and effective type of oral vaccineagainst various infectious diseases. Moreover, this yeastoral vaccine can be prepared rapidly because unlikeproteins produced in E. coli, it does not require a compli-cated purification step (Fig. 5). These advantages are alsorelevant for emerging pandemics.The immunological responses shown in Fig. 4a show that
administration of control S. cerevisiae gave an immuneresponse IgG titer of 2 9 102 as well as of IncompleteFreund’s adjuvant and cholera toxin (Figs 1a and 2a).Saccharomyces cerevisiae is thought to have a betteradjuvant function as a result of b-glucan in its cell wall(Berner et al., 2008; Rodr�ıguez et al., 2009),
Conclusions
After Eno1p was successfully produced by E. coli andpurified by passage through endotoxin-removing columns,the protein was given to mice before challenge with a lethaldose of C. albicans cells. Survival tests confirmed theprotective efficacies of this antigen: after more than1 month, survival rates associated with s.c. injection andi.n. infection were 12.5% and 25%, respectively. Next,Eno1p was produced on the surface of S. cerevisiae cellsand was administered orally to mice. Oral delivery of yeastcells that displayed Eno1p on their surfaces protected 60%of mice against candidiasis. The present study demon-strates the considerable potential of molecular display ofimmunogens on microbial cells to generate oral vaccines for
Yeast Antigenic protein displaying yeast
DNA E. coli Antigenicprotein
Removingof ET
Purification +adjuvant
(a)
(b)
Fig. 5 Comparison between conventional and novel oral vaccines. (a) Conventional vaccination using recombinant protein requires a complicated
procedure for purification of the antigen, removal of endotoxin (ET), and preparation and delivery of an adjuvant. (b) Oral vaccine avoids the
complicated steps to produce recombinant antigenic protein in Escherichia coli. In addition, the yeast cell surface acts as an adjuvant to promote an
immune response.
Pathogens and Disease (2013), 69, 262–268, © 2013 Federation of European Microbiological Societies. Published by John Wiley & Sons Ltd. All rights reserved 267
S. Shibasaki et al. Yeast-based oral vaccine against candidiasis

convenient protection against infectious diseases. The onlyrequirement is the availability of a DNA sequence thatencodes the antigenic protein; no purification step is neededto produce this oral vaccine. However, whether it retainsprotective effect after attenuation of whole cells as inthe previous study using another type of antigen-displayingcell (Adachi et al., 2010) needs to be examined, becauseS. cerevisiae sometimes causes invasive infection(Enache-Angoulvant & Hennequin, 2005).For further development of a promising vaccine that
protects against candidiasis, we have used proteomicanalyses to investigate other candidate antigenic proteins(Aoki et al., 2013). This should lead to novel antigens thathave a stronger effect as a vaccine.
Acknowledgements
This work was supported by the regional innovation creationR&D programs of the Ministry of Economy, Trade andIndustry, Japan. The authors declare no conflict of interest.
References
Adachi K, Kawana K, Yokoyama T et al. (2010) Oral immunization
with a Lactobacillus casei vaccine expressing human papilloma-
virus (HPV) type 16 E7 is an effective strategy to induce mucosal
cytotoxic lymphocytes against HPV16 E7. Vaccine 28: 2810–2817.
Aoki W, Kitahara N, Miura N, Morisaka H, Yamamoto Y, Kuroda K &
Ueda M (2011) Comprehensive characterization of secreted
aspartic proteases encoded by a virulence gene family in Candida
albicans. J Biochem 150: 431–438.Aoki W, Ueda T, Tatsukami Y, Kitahara N, Morisaka H, Kuroda K &
Ueda M (2013) Time-course proteomic profile of Candida
albicans during adaptation to a fetal serum. Pathog Dis 67: 67–75.Benko I, Hern�adi F, Megyeri A, Kiss A, Somogyi G, Tegyey Z,
Kraicsovits F & Kov�acs P (1999) Comparison of the toxicity of
fluconazole and other azole antifungal drugs to murine and
human granulocyte-macrophage progenitor cells in vitro. J Anti-
microb Chemother 43: 675–681.Berner VK, Sura ME & Hunter KW Jr (2008) Conjugation of protein
antigen to microparticulate beta-glucan from Saccharomyces ce-
revisiae: a new adjuvant for intradermal and oral immunizations.
Appl Microbiol Biotechnol 80: 1053–1061.Enache-Angoulvant A & Hennequin C (2005) Invasive Saccharo-
myces infection: a comprehensive review. Clin Infect Dis 41:
1559–1568.Hanahan D (1983) Studies on transformation of Escherichia coli
with plasmids. J Mol Biol 166: 557–580.
Hube B, Sanglard D, Odds FC, Hess D, Monod M, Sch€afer W,
Brown AJ & Gow NA (1997) Disruption of each of the secreted
aspartyl proteinase genes SAP1, SAP2, and SAP3 of Candida
albicans attenuates virulence. Infect Immun 65: 3529–3538.Ito H, Fukuda Y, Murata K & Kimura A (1983) Transformation of
intact yeast cells treated with alkali cations. J Bacteriol 153: 163–168.
Kuroda K, Matsui K, Higuchi S, Kotaka A, Sahara H, Hata Y & Ueda
M (2009) Enhancement of display efficiency in yeast display
system by vector engineering and gene disruption. Appl Microbiol
Biotechnol 82: 713–719.Li W, Hu X, Zhang X, Ge Y, Zhao S, Hu Y & Ashman RB (2011)
Immunisation with the glycolytic enzyme enolase confers effec-
tive protection against Candida albicans infection in mice.
Vaccine 29: 5526–5533.Montagnoli C, Sandini S, Bacci A, Romani L & La Valle R (2004)
Immunogenicity and protective effect of recombinant enolase of
Candida albicans in a murine model of systemic candidiasis. Med
Mycol 42: 319–324.Morschh€auser J (2002) The genetic basis of fluconazole resis-
tance development in Candida albicans. Biochim Biophys Acta
1587: 240–248.Noble SM & Johnson AD (2007) Genetics of Candida albicans, a
diploid human fungal pathogen. Annu Rev Genet 41: 193–211.Nucci M & Marr KA (2005) Emerging fungal diseases. Clin Infect Dis
41: 521–526.Pfaller MA & Diekema DJ (2007) Epidemiology of invasive candi-
diasis: a persistent public health problem. Clin Microbiol Rev 20:
133–163.Rodloff C, Koch D & Schaumann R (2011) Epidemiology and
antifungal resistance in invasive candidiasis. Eur J Med Res 16:
187–195.Rodr�ıguez I, Chamorro R, Novoa B & Figueras A (2009) b-Glucan
administration enhances disease resistance and some innate
immune responses in zebrafish (Danio rerio). Fish Shellfish
Immunol 27: 369–373.Shibasaki S, Kawabata A, Ishii J, Yagi S, Kadonosono T, Kato M,
Fukuda N, Kondo A & Ueda M (2007) Construction of a novel
synergistic system for production and recovery of secreted
recombinant proteins by the cell surface engineering. Appl
Microbiol Biotechnol 75: 821–828.Shibasaki S, Maeda H & Ueda M (2009) Molecular display
technology using yeast – arming technology. Anal Sci 25: 41–49.Tamaru Y, Ohtsuka M, Kato K, Manabe S, Kuroda K, Sanada M &
Ueda M (2006) Application of the arming system for the
expression of the 380R antigen from red sea bream iridovirus
(RSIV) on the surface of yeast cells: a first step for the
development of an oral vaccine. Biotechnol Prog 22: 949–953.Xin H & Cutler JE (2011) Vaccine and monoclonal antibody that
enhance mouse resistance to candidiasis. Clin Vaccine Immunol
18: 1656–1667.
Pathogens and Disease (2013), 69, 262–268, © 2013 Federation of European Microbiological Societies. Published by John Wiley & Sons Ltd. All rights reserved268
Yeast-based oral vaccine against candidiasis S. Shibasaki et al.