physiological characteristics underlyingthedistribution ... · 578 shilo andyetinson the...
TRANSCRIPT

Vol 38, No. 4APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Oct. 1979, p. 577-5840099-2240/79/10-0577/08$02.00/0
Physiological Characteristics Underlying the DistributionPatterns of Luminous Bacteria in the Mediterranean Sea and
the Gulf of ElatM. SHILO* AND T. YETINSON
Department ofMicrobiological Chemistry, The Hebrew University-Hadassah Medical School, Jerusalem,Israel
Received for publication 18 June 1979
Physiological characteristics of luminous bacteria isolated from the Mediter-ranean and Gulf of Elat were compared to determine their relationship to thespecific seasonal and geographic distribution patterns of these bacteria. Theeffects of temperature on growth rate and yield, relative sensitivity to photooxi-dation, resistance to high salt concentration (8%), and ability to grow in nutrient-poor conditions appear to control these patterns. The winter appearance ofPhotobacterium fischeri and the succession of winter and summer types ofBeneckea harveyi in the eastem Mediterranean are explained by different tem-perature requirements for growth. Sensitivity to photooxidation explains thedisappearance of P. leiognathi, present in the main body of the Gulf of Elatthroughout the year, from the shallow coastal strip. B. harveyi is present in thiscoastal strip which is higher in nutrients and in productivity than the open waters.Competition experiments between B. harveyi and P. leiognathi in batch andcontinuous culture indicate that the oligotrophic P. leiognathi is outcompeted byB. harveyi in rich and even in relatively poor media. The distribution pattemfound in the Bardawil hypersaline lagoon is explained by selection of salinity-resistant mutants of B. harveyi from the Mediterranean Sea.
Luminous marine bacteria show a character-istic geographic and seasonal distribution pat-tern. This was recognized as early as 1889 byBeijerinck (4, 5), who described the seasonalfluctuation of luminous bacteria of the NorthSea where Bacillus phosphoreum (syn., Photo-bacterium phosphoreum), dominant throughoutthe entire year, was partially replaced in thesummer months of August and September by P.splendidum (syn., Beneckea harveyi or B. splen-dida). A different seasonal distribution patternhas been described recently from the southernCalifornia coast of the Pacific Ocean (20) whereP. fischeri, present throughout the year, waspartially replaced by B. harveyi during the sum-mer. In autumn and in winter, P. phosphoreumwas an additional minor component of this lu-minous bacterial community. Another fluctua-tion pattern was found in the eastern Mediter-ranean (M. Shelubsky-Shilo, M.S. thesis, He-brew University of Jerusalem, 1943), where sev-eral types of B. harveyi occur in sequence duringthe year and where an intrusion of a differenttype of luminous bacterium, now recognized asP. fischeri, takes place in the winter. In a pre-vious paper (23) we described the distributionpattern of different luminous bacterial types in
the eastern Mediterranean and the Gulf of Elat:P. fischeri, a typical Mediterranean winter type,is absent from the Gulf of Elat, whereas the Gulfof Elat species, P. leiognathi, is never found ineastern Mediterranean waters. Seasonal differ-ences in B. harveyi strains were marked in thecoastal region of the Mediterranean Sea. B.harveyi was the dominant species in the shallowcoastal strip of the Gulf of Elat; however, it wasnot isolated from deeper open waters of the gulf.In the present paper, we consider whether thephysiological characteristics of the different lu-minous bacterial types can explain their geo-graphic and seasonal patterns, and we evaluatethe role of specific environmental factors in gov-erning their distribution.
Ecological conditions for single environmentalfactors such as temperature, light, and nutrientand salt concentrations were simulated experi-mentally to determine their effect on pure cul-tures of luminous bacteria. The findings wererelated to the observed population dynamics ofluminous bacteria (23).
MATERIALS AND METHODSBacterial strains and cultivation conditions. B.
harveyi, P. leiognathi, and P. fischeri isolates from
577
on Septem
ber 9, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

578 SHILO AND YETINSON
the Mediterranean Sea, the hypersaline Bardawil la-goon, and the Gulf of Elat were compared (23). Forgrowth of the bacteria, yeast extract-peptone medium(YP) and basal medium containing 3 or 8% NaCl wasused (23). The same media with 1.5% Difco agar servedfor plating.
Photooxidation experiments. Luminous bacteriawere grown in a shaker on YP medium (3% NaCl) at26°C, harvested by centrifugation (15 min at 10,000x g), and suspended in sterile seawater. Laboratoryexperiments on photooxidation were carried out byusing a Widioscope (AB Wiktoks Mekaniska, Jarfalla,Sweden), with a halogen lamp (250 W, 24 V) as thelight source. These photooxidation experiments werecarried out in sealed, flat-bottom tubes 1 cm in diam-eter, at a light intensity of 106 ergs.cm-2s-'. Thesuspensions were kept in an atmosphere of pure 02 at25°C, with continuous stirring by a magnetic stirrer.In the experiments where a photosensitizer, toluidineblue (1 ,uM), was added, the light source was an illu-minated Warburg monometer unit with tungstenlamps (40 W, 220 V) (B. Braun Melsungen AG, W.Germany).
In situ experiments to detect photodynamic damagewere carried out at station 3 (23) in the Gulf of Elat.Different strains of luminous bacteria were placed incellophane dialysis tubes, 1 cm in diameter, and sus-pended in different depths of the water column aspreviously described (1, 9). A parallel series of cello-phane tubes covered by black plastic sheets served ascontrols.SOD activity determination. Cells were har-
vested, suspended in 0.01 M potassium phosphate atpH 7.8, and disrupted for 30 s with glass beads (0.10 to0.11 mm in diameter) in a Nossal Homogenizer (CaseWestern Reserve University, Cleveland, Ohio). Celldebris was removed by centrifugation at 30,000 x g for15 min, and the cell-free extracts used for the enzymeassay were obtained by the procedure of McCordand Fridovich (15). Protein was determined by themethod of Davis (8); at 4°C at a current of 3 mA pergel cylinder, superoxide dismutase (SOD) activity waslocalized by the negative staining procedure describedby Beauchamp and Fridovich (3). The gels werescanned at 560 mm.Transfer of bacteria from low to high salt con-
centration. Transfer of bacteria from 3 to 8% NaClmedia was carried out both by abrupt change (shock)and by gradual change. For abrupt change, cells in the3% NaCl medium were harvested by centrifuging at10,000 x g and suspended in 8% NaCl in either artificialseawater or 8% NaCl-YP medium. Gradual change insalinity (3 to 8% NaCl) was brought about by dialyzingthe cell suspension against a continuously increasinggradient of NaCl over 8 h. Survival of the cells afterdifferent periods of exposure in the high NaCl concen-tration was determined by plating on 3% NaCl YPagar plates and counting the colonies. In parallel, theportion of the population capable of forming colonieson 8% NaCl YP agar was estimated and expressed aspercent of total on 3% NaCl.
Competition experiments between differentluminous bacterial species. Experiments were car-ried out both in batch and in continuous culture con-ditions with an inoculum mixture of B. harveyi and P.
leiognathi strains of equal density. The ratio betweenthese strains was followed throughout growth. Themedia used were 3% NaCl-YP medium and basalmedium with 0.3% glycerol (23). For the competitionexperiments in continuous culture conditions, a chem-ostat (Bioflow model C30; New Brunswick ScientificCo., Inc., New Brunswick, N.J.) with a flow rate of 0.3ml.min-', agitated at 400 rpm, and an aeration of 0.2(vol/vol) min-' was used. The total volume in theculture vessel was 300 ml. Plating was carried out onYP agar containing 3% NaCl and 0.2% soluble starchto differentiate between B. harveyi strains which areamylase positive and P. leiognathi strains which areamylase negative. Extracellular amylase productionwas detected as described in a previous paper (23).
RESULTSGrowth temperatures. Luminous bacteria
strains isolated from different environments orduring different seasons differed markedly ingrowth temperature minima and maxima andalso in growth rates and cell yields for differenttemperatures (Fig. 1 and Table 1). All Mediter-ranean winter strains, B. harveyi as well as P.fischeri, grew at 10°C, but many did not grow at40°C. On the other hand, summer B. harveyiisolates grew well at 40°C but did not grow at10°C. At 15°C, growth of summer isolates wassignificantly lower than that of the winterstrains, whereas at 34°C, the summer strainsoutgrew the winter ones, although the differencein rate was minimal (Table 1). Thus, the sea-sonal selection of thermal types of luminousbacteria in the coastal waters of the easternMediterranean basin seems to be due to differentgrowth temperature spectra of the principaltypes of luminous bacteria.
Sensitivity to photooxidative damage.Luminous bacteria differ in sensitivity to pho-tooxidation, as demonstrated in the laboratory
E 500-B.h
400 Oh 66)
o0 /rj 3)300-0
-, 200 (E 5)- 0
0 10 20 30 40 50TEMPERATURE (OC)
FIG. 1. Growth yield of different strains of lumi-nous bacteria at different temperatures in YP me-dium. Yield was measured after 24 h in Klett units atthe stationary phase ofgrowth; 100 Klett units (filter420) = 0.7 x 108 cells per ml.
APPL. ENVIRON. MICROBIOL.
on Septem
ber 9, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from
LUMINOUS BACTERIA DISTRIBUTION PATTERNS 579
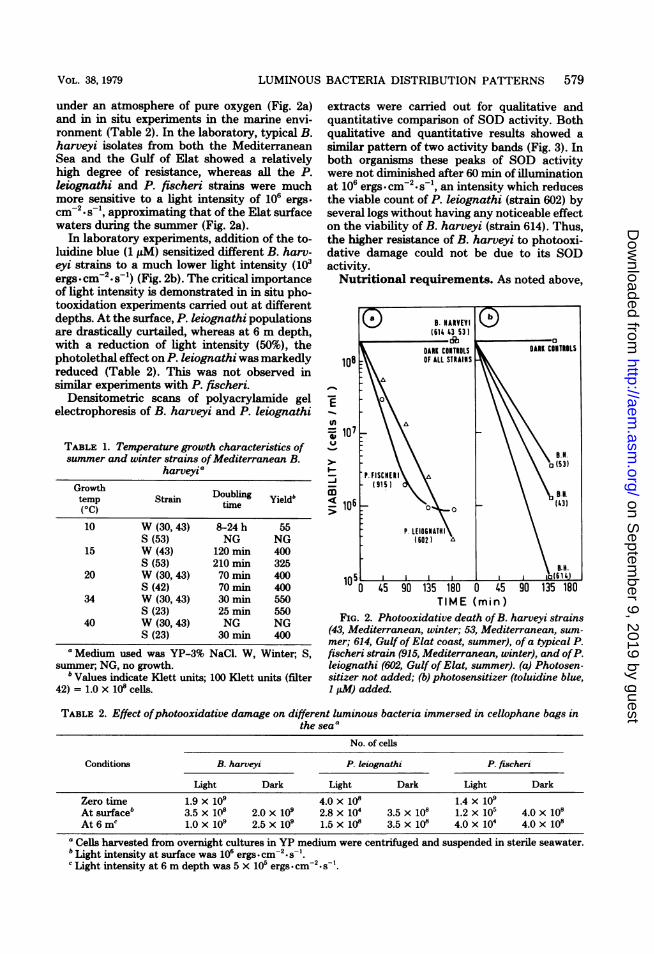
under an atmosphere of pure oxygen (Fig. 2a)and in in situ experiments in the marine envi-ronment (Table 2). In the laboratory, typical B.harveyi isolates from both the MediterraneanSea and the Gulf of Elat showed a relativelyhigh degree of resistance, whereas all the P.leiognathi and P. fischeri strains were muchmore sensitive to a light intensity of 106 ergs.
-2 -1cm *s , approximating that of the Elat surfacewaters during the summer (Fig. 2a).
In laboratory experiments, addition of the to-luidine blue (1 ,uM) sensitized different B. harv-eyi strains to a much lower light intensity (103ergs.cm-2.s-) (Fig. 2b). The critical importanceof light intensity is demonstrated in in situ pho-tooxidation experiments carried out at differentdepths. At the surface, P. leiognathi populationsare drastically curtailed, whereas at 6 m depth,with a reduction of light intensity (50%), thephotolethal effect on P. leiognathi was markedlyreduced (Table 2). This was not observed insimilar experiments with P. fischeri.
Densitometric scans of polyacrylamide gelelectrophoresis of B. harveyi and P. leiognathi
TABLE 1. Temperature growth characteristics ofsummer and winter strains ofMediterranean B.
harveyiaGrowthtemp Strain Doubling Yield'(OC) time10 W (30,43) 8-24h 55
S (53) NG NG15 W (43) 120 min 400
S (53) 210min 32520 W (30, 43) 70 min 400
S (42) 70 min 40034 W (30, 43) 30 mi 550
S (23) 25 min 55040 W (30, 43) NG NG
S (23) 30 min 400a Medium used was YP-3% NaCl. W, Winter, S,
summer; NG, no growth.b Values indicate Klett units; 100 Klett units (filter
42) = 1.0 x 108 cells.
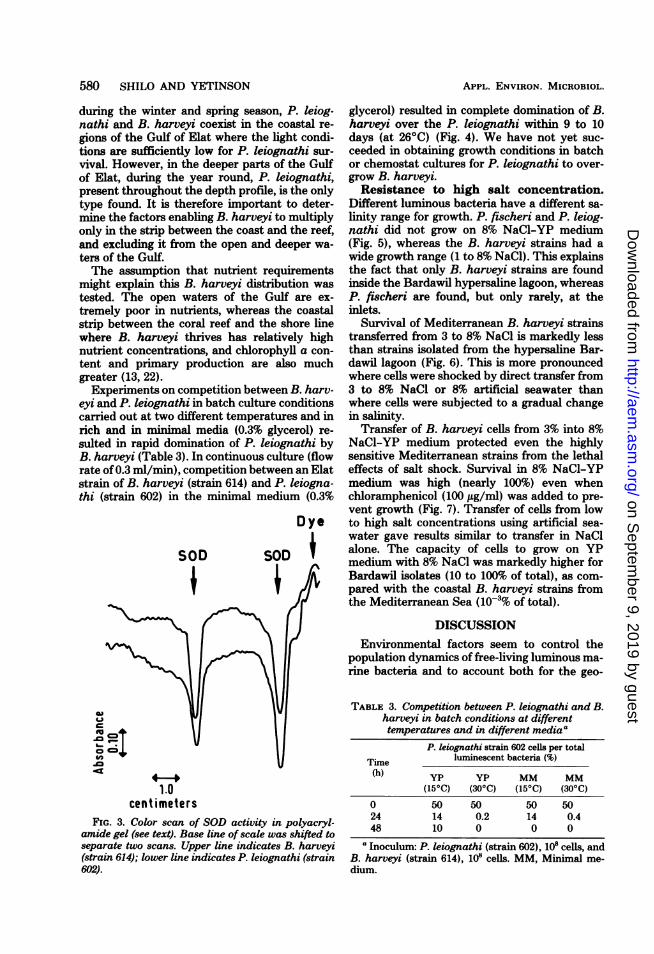
extracts were carried out for qualitative andquantitative comparison of SOD activity. Bothqualitative and quantitative results showed asimilar pattern of two activity bands (Fig. 3). Inboth organisms these peaks of SOD activitywere not diminished after 60 min of illuminationat 106 ergs-cm-2-s-', an intensity which reducesthe viable count of P. leiognathi (strain 602) byseveral logs without having any noticeable effecton the viability of B. harveyi (strain 614). Thus,the higher resistance of B. harveyi to photooxi-dative damage could not be due to its SODactivity.Nutritional requirements. As noted above,
-BHARVEYI(61X 43 53)dbo
DARK CONTROLS DARK CONTROLS
10E OF ALL STRAINS
107 . ItE10
>- ~~~~~~~~~~~~~(53)P. FISCNCR
(915)
BSN.
106 ~~~~~(43)
TIME (min)FIG. 2. Photooxidative death of B. harveyi strains
(43, Mediterranean, winter; 53, Mediterranean, sum-
mer; 614, Gulf of Elat coast, summer), of a typical P.fischeri strain (915, Mediterranean, winter), and ofP.leiognathi (602, Gulf of Elat, summer). (a) Photosen-sitizer not added; (b) photosensitizer (toluidine blue,1 pAM) added.
TABLE 2. Effect ofphotooxidative damage on different luminous bacteria immersed in cellophane bags inthe seaa
No. of cells
Conditions B. harveyi P. leiognathi P. fischeri
Light Dark Light Dark Light Dark
Zero time 1.9 x 109 4.0 x 108 1.4 x 109At surfaceb 3.5 x 108 2.0 x 109 2.8 x 104 3.5 x 108 1.2 x 105 4.0 x 108At 6 mc 1.0 x 109 2.5 x 109 1.5 x 108 3.5 x 108 4.0 x 104 4.0 x 108
a Cells harvested from overnight cultures in YP medium were centrifuged and suspended in sterile seawater.b Light intensity at surface was 106 ergs cm-2 s-'.c Light intensity at 6 m depth was 5 x 105 ergs.cm2.s-'.
VOL. 38, 1979
on Septem
ber 9, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from
580 SHILO AND YETINSON
during the winter and spring season, P. leiog-nathi and B. harveyi coexist in the coastal re-
gions of the Gulf of Elat where the light condi-tions are sufficiently low for P. leiognathi sur-vival. However, in the deeper parts of the Gulfof Elat, during the year round, P. leiognathi,present throughout the depth profile, is the onlytype found. It is therefore important to deter-mine the factors enabling B. harveyi to multiplyonly in the strip between the coast and the reef,and excluding it from the open and deeper wa-ters of the Gulf.The assumption that nutrient requirements
might explain this B. harveyi distribution was
tested. The open waters of the Gulf are ex-
tremely poor in nutrients, whereas the coastalstrip between the coral reef and the shore linewhere B. harveyi thrives has relatively highnutrient concentrations, and chlorophyll a con-
tent and primary production are also muchgreater (13, 22).Experiments on competition between B. harv-
eyi and P. leiognathi in batch culture conditionscarried out at two different temperatures and inrich and in minimal media (0.3% glycerol) re-
sulted in rapid domination of P. leiognathi byB. harveyi (Table 3). In continuous culture (flowrate of0.3 ml/min), competition between an Elatstrain of B. harveyi (strain 614) and P. leiogna-thi (strain 602) in the minimal medium (0.3%
SOD
Dye
SOD
c
L)
-to= o;
1.0
centimetersFIG. 3. Color scan of SOD activity in polyacryl-
amide gel (see text). Base line of scale was shifted toseparate two scans. Upper line indicates B. harveyi(strain 614); lower line indicates P. leiognathi (strain602).
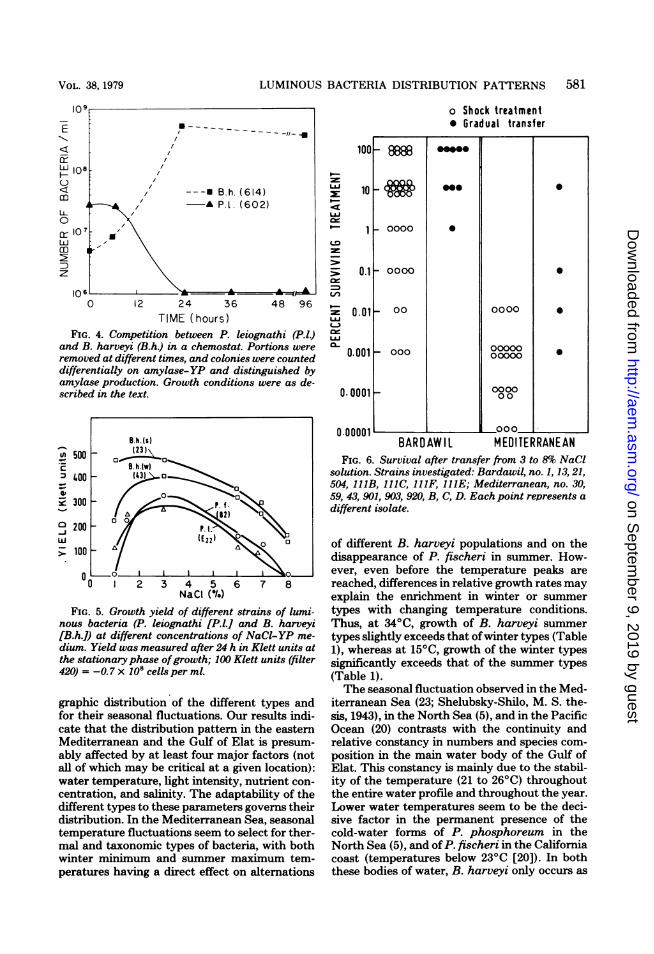
glycerol) resulted in complete domination of B.harveyi over the P. leiognathi within 9 to 10days (at 2600) (Fig. 4). We have not yet suc-ceeded in obtaining growth conditions in batchor chemostat cultures for P. leiognathi to over-grow B. harveyi.Resistance to high salt concentration.
Different luminous bacteria have a different sa-
linity range for growth. P. fischeri and P. leiog-nathi did not grow on 8% NaCl-YP medium(Fig. 5), whereas the B. harveyi strains had a
wide growth range (1 to 8% NaCl). This explainsthe fact that only B. harveyi strains are foundinside the Bardawil hypersaline lagoon, whereasP. fischeri are found, but only rarely, at theinlets.
Survival of Mediterranean B. harveyi strainstransferred from 3 to 8% NaCl is markedly lessthan strains isolated from the hypersaline Bar-dawil lagoon (Fig. 6). This is more pronouncedwhere cells were shocked by direct transfer from3 to 8% NaCl or 8% artificial seawater thanwhere cells were subjected to a gradual changein salinity.
Transfer of B. harveyi cells from 3% into 8%NaCl-YP medium protected even the highlysensitive Mediterranean strains from the lethaleffects of salt shock. Survival in 8% NaCl-YPmedium was high (nearly 100%) even whenchloramphenicol (100 ,ug/ml) was added to pre-vent growth (Fig. 7). Transfer of cells from lowto high salt concentrations using artificial sea-water gave results similar to transfer in NaClalone. The capacity of cells to grow on YPmedium with 8% NaCl was markedly higher forBardawil isolates (10 to 100% of total), as com-
pared with the coastal B. harveyi strains fromthe Mediterranean Sea (10-3% of total).
DISCUSSIONEnvironmental factors seem to control the
population dynamics of free-living luminous ma-rine bacteria and to account both for the geo-
TABLE 3. Competition between P. leiognathi and B.harveyi in batch conditions at differenttemperatures and in different mediaa
P. leiognathi strain 602 cells per totalTime luminescent bacteria (%)(h) yp yp MM MM
(150C) (300C) (150C) (300C)
0 50 50 50 5024 14 0.2 14 0.448 10 0 0 0
aInoculum: P. leiognathi (strain 602), 108 cells, andB. harveyi (strain 614), 10' cells. MM, Minimal me-dium.
APPL. ENVIRON. MICROBIOL.
,--a
on Septem
ber 9, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from
LUMINOUS BACTERIA DISTRIBUTION PATTERNS 581
_~ -----/-
z ~ ~~~~~~~~~~~~~~/
WI108 //< F , ---U B.h. (614)A0-A122(602)
LL /0
gQlo7
TIME (hOUrS )FIG. 4. Competition between P. leiognathi (P.IJ)
and B. harveyi (B.h.) in a chemostat. Portions wereremoved at different times, and colonies were counteddifferentially on amylase-YP and distinguished byamylase production. Growth conditions were as de-scribed in the text.
B.h.(s)
v}500 _o-
_23._ B. h.(w)
:140 23) 4 6 7
I0~~~~~
TIE (hou )
FIG. 5. Growth yield of different strains of lumi-nous bacteria (P. leiognathi [P.l.] and B. harveyi[B.h.0 at different concentrations of NaCI-YP me-dium. Yield was measured after 24 h in Klett units atthe stationary phase ofgrowth; 100 Klett units (filter420) = -0.7 x1OScells per ml.
graphic distribution of the different types andfor their seasonal fluctuations. Our results indi-cate that the distribution pattern in the easternMediterranean and the Gulf of Elat is presum-ably affected by at least four major factors (notall of which may be critical at a given location):water temperature, light intensity, nutrient con-centration, and salinity. The adaptability of thedifferent types to these parameters governs theirdistribution. In the Mediterranean Sea, seasonaltemperature fluctuations seem to select for ther-mal and taxonomic types of bacteria, with bothwinter miIiimum and sunumer maximnum tem-peratures having a direct effect on alternations
0000
o Shock treatment* Gradual transfer
100
LLI
LAJ
1-
cx
L-)
U.J0l-
10 _
- 888
0
0
0
0.1 - 0000
0-01_
0.001 - 000
O.0001 _
n nnnni
00 0000
00000
00000
000000
u .u u uu I,BARDAWIIl MEDITERRANEAN
FIG. 6. Survival after transfer from 3 to 8% NaClsolution. Strains investigated: Bardawil, no. 1, 13,21,504, 111B, 111C, 111F, 111E; Mediterranean, no. 30,59, 43, 901, 903, 920, B, C, D. Each point represents a
different isolate.
of different B. harveyi populations and on thedisappearance of P. fischeri in summer. How-ever, even before the temperature peaks are
reached, differences in relative growth rates mayexplain the enrichment in winter or summer
types with changing temperature conditions.Thus, at 340C, growth of B. harveyi summer
types slightly exceeds that ofwinter types (Table1), whereas at 150C, growth of the winter typessignificantly exceeds that of the summer types(Table 1).The seasonal fluctuation observed in the Med-
iterranean Sea (23; Shelubsky-Shilo, M. S. the-sis, 1943), in the North Sea (5), and in the PacificOcean (20) contrasts with the continuity andrelative constancy in numbers and species com-
position in the main water body of the Gulf ofElat. This constancy is mainly due to the stabil-ity of the temperature (21 to 260C) throughoutthe entire water profile and throughout the year.Lower water temperatures seem to be the deci-sive factor in the permanent presence of thecold-water forms of P. phosphoreum in theNorth Sea (5), and of P. fischeri in the Californiacoast (temperatures below 230C [20]). In boththese bodies of water, B. harveyi only occurs as
000
0
VOL. 38, 1979
I
on Septem
ber 9, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

582 SHILO AND YETINSON
__)-J-jw
(-0
wm
z
TIM E (hours)FIG. 7. Survival and capability ofgrowth oftypical B. harveyi (B.h) Mediterranean (4) and Bardawil (714)
strains in 8% NaCI-YP medium after sudden transfer from 3 to 8% NaCI in different media. Solid line andopen symbols indicate viability tested by capability offorming colonies on 3% YP medium. Broken line andclosed symbols indicate capability for growth on 8% NaCl-YP medium. Treatment media: A, YP-8% NaCI;E, 8% NaCI; 0, 8% artificial seawater (ASW).
a summer intrusion. Already in 1916, Beijerinckobserved that the growth temperature optimumof B. harveyi in laboratory media was muchhigher than that of P. phosphoreum and thatthis could explain the distribution of these spe-cies. In the eastern Mediterranean, with an an-nual oscillation between 15 and 300C, P. fischeriis found only during the winter and the spring.In the Gulf of Elat, where temperatures never
fall below 200C, P. fischeri is never found.The selection of different symbiotic luminous
bacteria inhabiting the light organ of differentfishes also appears to be governed by the watertemperatures. Thus, some of the cold-water
deepsea fishes, including representatives belong-ing to several families of different orders, areinhabited by P. phosphoreum (20). This groupof luminous bacteria is characterized by its ca-pacity to grow at 40C and also seems to survivehigh pressure better than either P. fischeri or B.harveyi (6). Leiognathidae, which are typicaltropical shallow-water Pacific fishes living intemperatures from 25 to 300C, have light organsinhabited by the warm-water bacterium, P.leiognathi (17). Monocentridae, subtropical andtemperate relatively shallow-water fishes, arefound in Australian, Japanese, South African,and South American coastal waters, where tem-
APPL. ENVIRON. MICROBIOL.
on Septem
ber 9, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

LUMINOUS BACTERIA DISTRIBUTION PATTERNS 583
peratures range from 15 to 20°C. The symbioticluminous bacterium inhabiting their light organsis the mesotropic species, P. fischeri (10, 19).
In the Gulf of Elat, P. leiognathi, the solefree-living luminous bacterium of the open wa-ter, is present in the entire column below 2 mdepth; its absence from shallow coastal areasand the surface layer of open water in summercan now be explained as due to solar radiation-induced decay. Only the more photoresistant B.harveyi can survive under the strong light con-ditions prevailing in the shallow coastal regionduring the summer. In in situ experiments, asingle day of exposure to sunlight of a mixed P.leiognathi and B. harveyi culture submerged incellophane tubes reduced the viable count of P.leiognathi by several orders of magnitude,whereas the B. harveyi population was hardlyaffected.
Differences in sensitivity to photodynamic ef-fects are widely known from different organisms;however, this may be the first description oflethal photooxidation governing composition ofindigenous bacterial communities in oceans. Dif-ferences in resistance to photooxidation mayexplain the distribution of luminous bacteria onthe Hawaiian coast, where P. leiognathi, P.phosphoreum, and B. harveyi were present inapproximately equal numbers in the open watersoff Oahu, whereas B. harveyi was the sole orpredominant species in the coastal waters (16).A photooxidative decay model has likewise
been presented for enteric bacteria in seawaterby Chamberlin and Mitchell (7) in their analysisof the field data of Gameson and Gould (11)where indications were found that there is alinear relationship between decay rates and lightintensity. Thus, light was proposed as the majorfactor controlling mortality of enteric bacteria inmarine environments. This does not exclude thepossibility that additional factors can accentuateor may even be essential to photooxidative dam-age.The intensity of light was found to be critical;
the slight decrease in light intensity at severalmeters depth below the surface was sufficient tomarkedly reduce the photolethal effect on P.leiognathi (Table 2). The addition of toluidineblue photosensitizer destroyed the natural re-sistance of B. harveyi to photooxidation, and itbecame as sensitive to light and oxygen as P.leiognathi and P. fischeri (Fig. 5). This suggeststhat the high sensitivity of P. leiognathi and P.fischeri to light may be due to the presence of anatural photosensitizer such as the phorphyrinpigments.
Nutrient concentration may also control thedistribution of some species. B. harveyi thrives
in such eutrophic marine enviroments as theMediterranean shore, enriched by local sewageoutfalls, by flood waters from agricultural land,by winter mixing, and by upwelling during sum-mer (O. Oren, Ph.D. thesis, Hebrew Universityof Jerusalem, 1970). In the Gulf of Elat, B.harveyi was present only in the coastal stripbetween the coral reef and the shore, a regionmarkedly higher in nutrient concentration, pri-mary production and chlorophyll a content thanthe open water region (13, 22). The Gulf of Elat,which is surrounded by desert, is characterizedby low nutrient concentrations throughout theentire water column. We suggest that the oligo-trophic conditions prevailing in the Gulf favorthe selective enrichment of P. leiognathi overB. harveyi. Competition experiments betweenGulf of Elat B. harveyi and P. leiognathi strainsin batch culture as well as in the chemostatconfirmed the superiority of B. harveyi over P.leiognathi when rich or less rich minimal mediawere used (Fig. 4, Table 3). We suggest that ourinability to obtain conditions for selective en-richment of P. leiognathi is due to the fact thateven the minimal medium is "rich" when com-pared with the nutrient concentrations of Gulfof Elat water. We predict that this oligotrophicspecies has a lower threshold level for nutrientconcentrations than B. harveyi does.The effect of salinity has been investigated in
populations from the Bardawil lagoon, a hyper-saline body of water surrounded by the Medi-terranean Sea and separated from it by a narrowbar but with a constant inflow of Mediterraneanwater taking place. For survival and growth inthe lagoon, tolerance to high salt concentrations(8%) is required. Most B. harveyi types and allP. fischeri types appear to be eliminated in thelagoon owing to their sensitivity to salt shock.Only halo-tolerant types or mutants of B. harv-eyi are capable of surviving and multiplying inthis environment. This inference is supported bythe distribution of luminous bacteria along atransect extending from the opening of the la-goon toward its inner, most saline part, sampledin August 1978. The percentage of cells capableof colony formation on 8% YP agar showed amarked increase from the Mediterranean inletstoward the hypersaline part of the lagoon, indi-cating that selection or enrichment for saline-tolerant types had taken place. An interestingcase of differences in sensitivity to cation shockamong yeast strains and their mutants has beendescribed by Bard et al. (2) where differences inthe membrane sterol composition between thewild type and its mutants seemed to underliethe sensitivity to lethal cation pulses in theyeast.
VOL. 38, 1979
on Septem
ber 9, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

584 SHILO AND YETINSON
In conclusion, the notion that luminous bac-teria can serve as valuable tools in indicating thepresence of different water masses is supported.Moreover, information has been obtained on thephysiological factors which govern and controldistribution ofluminous bacteria, and these find-ings may now be extended to other bacterialgroups.
ACKNOWLEDGMENTS
We thank Miriam Fine for her part in carrying out theactivity tests and gel electrophoresis of SOD; Julie Seijffersfor her help in the photooxidation experiments; W. J. Hastingsand E. G. Ruby for critical reading of the manuscript; andNechama Ben-Eliahu for editorial assistance.
This work was supported by a grant from the DeutscheForschungsmeinschaft.
LITERATURE CITED
1. Abeliovich, A., and M. Shilo. 1972. Photooxidativedeath in blue-green algae. J. Bacteriol. 111:682-689.
2. Bard, M., N. D. Lees, K. S. Burrows, and F. W.Kleinhans. 1978. Difference in crystal violet uptakeand cation-induced death among yeast sterol mutants.J. Bacteriol. 135:1146-1148.
3. Beauchamp, C. O., and I. Fridovich. 1971. Superoxidedismutase: improved assays and an assay applicable topolyacrylamide gels. Anal. Biochem. 44:276-287.
4. Beijerinck, M. W. 1889. Le Photobacterium luminosum,bacterie luminesce de la Mer du Nord. Arch. Neerl. Sci.Exacta Nat. Haarlem. 23:401-405.
5. Beijerinck, M. W. 1916. Leuchtbakterien der Nordsee inAugust und September. Folia Microbiol. (Delft) 4:15-40.
6. Brown, D. E., F. H. Johnson, and D. A. Marsland.1942. The pressure temperature relations of bacterialluminescence. J. Cell. Comp. Physiol. 20:151-168.
7. Chamberlin, C. E., and R. M. Mitchell. 1978. A decaymodel for enteric bacteria in natural waters, p. 325. In
R. Mitchell (ed.), Water pollution microbiology. Wiley-Interscience, New York.
8. Davis, B. J. 1964. Disc electrophoresis. 2. Method andapplication to human serum proteins. Ann. N.Y. Acad.Sci. 121:404-427.
9. Eloff, J. N., Y. Steinitz, and M. Shilo. 1976. Photooxi-dation of cyanobacteria in natural conditions. Appl.Environ. Microbiol. 31:119-126.
10. Fitzgerald, J. M. 1977. Classification ofluminous bacteriafrom the light organ of the Australian pinecone fish,Cleidopus gloriamaris. Arch. Microbiol. 112:153-156.
11. Gameeson, A. L. H., and D. J. Gould. 1975. Proceedingsof the International Symposium on Discharge of Sew-age from Sea Outfalls. Pergamon Press, Inc., Elmsford,N.Y.
12. Krinsky, N. L. 1968. The protective functions of carote-noid pigments. Photophysics 3:123-195.
13. Levanon-Spanier, I., E. Padan, and Z. Reiss. 1979.Primary production in a desert-enclosed sea-the Gulfof Elat (Aqaba), Red Sea. Deep Sea Res., in press.
14. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folinphenol reagent. J. Biol. Chem. 193:265-275.
15. McCord, J. J., and L. Fridovich. 1969. Superoxide dis-mutase-an enzymic function for erythrocuperin (Hem-ocuperin). J. Biol. Chem. 244:6049-6055.
16. Reichelt, J. L., and P. Baumann. 1973. Taxonomy ofthe marine, luminous bacteria. Arch. Mikrobiol. 94:283-330.
17. Reichelt, J. L., K. Nealson, and J. W. Hastings. 1977.The specificity of symbiosis: pony fish and luminescentbacteria. Arch. Microbiol. 112:157-161.
18. Ruby, E. G., and K. H. Nealson. 1976. Symbiotic asso-ciation of Photobacterium fischeri with the marineluminous fish Monocentris japonica: a model of sym-biosis based on bacterial studies. Biol. Bull. (WoodsHole, Mass.) 151:574-586.
19. Ruby, E. G., and J. G. Morin. 1978. Specificity of sym-biosis between deep-sea fishes and psychrotrophic lu-minous bacteria. Deep Sea Res. 25:161-167.
20. Ruby, E. G., and K. Nealson. 1978. Seasonal changes inthe species composition of luminous bacteria in near-shore seawater. Limnol. Oceanogr. 23:530-533.
21. Spikes, J. D., and R. Livingston. 1969. The molecularbiology of photodynamic action sensitized oxidations inbiological systems. Adv. Rad. Biol. 3:29-121.
22. Sournia, A. 1977. Notes on primary productivity ofcoastal waters in the Gulf of Elt (Red Sea). Int. Rev.Ges. Hydrobiol. 62:813-819.
23. Yetinson, T., and M. Shilo. 1979. Seasonal and geo-graphic distribution of luminous bacteria in the EasternMediterranean Sea and the Gulf of Elat. Appl. Environ.Microbiol. 37:1230-1238.
APPL. ENVIRON. MICROBIOL.
on Septem
ber 9, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from