peninsula’)oa.upm.es/39649/1/hugo_mas_gisbert.pdf · 2016-03-09 · los ojos de mis hijos. y no...
TRANSCRIPT

UNIVERSIDAD POLITÉCNICA DE MADRID
ESCUELA TÉCNICA SUPERIOR DE INGENIEROS DE MONTES
Fenología y capacidad de dispersión de Monochamus galloprovincialis (Olivier 1795)
en la Península Ibérica
(‘Phenology and dispersal ability of M. galloprovincialis (Olivier 1795) at Iberian
Peninsula’)
TESIS DOCTORAL
Hugo Mas i Gisbert
Ingeniero de Montes
2015


DEPARTAMENTO DE SILVOPASCICULTURA
ESCUELA TÉCNICA SUPERIOR DE INGENIEROS DE MONTES
UNIVERSIDAD POLITÉCNICA DE MADRID
Fenología y capacidad de dispersión de Monochamus galloprovincialis (Olivier 1795) en la Península Ibérica
(‘Phenology and dispersal ability of Monochamus galloprovincialis (Olivier 1795) at Iberian Peninsula’)
TESIS DOCTORAL
Autor:
Hugo Mas i Gisbert. Ingeniero de Montes
Dirigida por:
Dr. Iñaki Etxebeste Larrañaga. Doctor en Biología
Dr. Diego Gallego Cambronero. Doctor en Biología
Tutor:
Jose Antonio Omeñaca González. Doctor Ingeniero de Montes
Madrid 2015


Tribunal nombrado por el Mgfco. y Excmo. Sr. Rector de la Universidad
Politécnica de Madrid, el día de de 2015.
Presidente D. Vocal D. Vocal D. Vocal D. Secretario D.
Realizado del acto de defensa y lectura de la Tesis el día de de 2015 en Madrid.
Calificación ……………………………………………
EL PRESIDENTE LOS VOCALES
EL SECRETARIO


Una nota del autor
‘But I don't want to go among mad people’, Alice remarked. ‘Oh, you can't help that', said the Cat: ‘we're all mad here. I'm mad. You're mad.'
‘How do you know I'm mad?' said Alice. ‘You must be,' said the Cat, ‘or you wouldn't have come here.'
Lewis Carroll, Alice’s Adventure in Wonderland
Hace un cuarto de siglo, arañada en un muro frente al cual se encontraba la última parada del autobús que, desde el primer día y durante cuatro años, tuve que coger con tanta frecuencia para remontar la interminable cuesta que separaba mi casa del instituto donde cursé el bachillerato unificado polivalente, una frase que pudo haber resultado extraña para un púber de catorce años supuso, de alguna forma, el umbral conceptual que toda persona cruza quién sabe en qué momento para descubrir la llanura de la vida y el conocimiento, de la sociedad y la política, arrasada tantas veces por la belleza de nuestros propios enemigos: «Tierno habría votado NO».
Aunque en cierto modo las enseñanzas del Viejo Profesor marcaran entones mi educación humanista, sólo pretendo traer aquí un aspecto muy concreto de su filosofía, la defensa férrea del agnosticismo que Enrique Tierno Galván abanderó durante toda su vida, y que aun siendo un concepto algo distante de mi ateísmo consustancial, describe bien la sensación íntima de un comportamiento transfronterizo en el que me he visto agradablemente embarrado tantas veces en mi vida y, especialmente, en el ejercicio de mi profesión como Ingeniero de Montes a cargo del Laboratori de Sanitat Forestal de la Generalitat Valenciana durante la última década: vivir en el ecotono, a caballo entre dos determinaciones: la investigación científica y la gestión forestal.
No es fácil saber en qué momento lo decidí. Podría parecer que fue la inercia laboral o la extinción de una etapa burocrática en mi universidad lo que me empujó a tomar la determinación de meterme dentro de un estrecho túnel para escribir una disertación de centenares de páginas sobre los hábitos de un pequeño insecto, y del que sólo salgo ahora, después de tres largos y extraños meses, con una tesis casi inacabada y consciente de nuevo de con qué aridez y esfuerzo se teje el proceso creativo. Sin embargo, con todo, creo sinceramente que ese frágil resorte intelectual que precipita las voluntades quizás ya se disparó hace ahora mucho más tiempo, cuando con escasos once años construí un rudimentario laboratorio de entomología en el desván de una masía en la montaña mediterránea de Alcoi donde transcurrió mi infancia. En aquel momento empecé a ser consciente de mi profunda admiración por esa locura que es el intento de objetivar la realidad (o al menos intersubjetivarla), una locura de la que ya nunca conseguiría desprenderme.
Hoy esa disertación está a punto de ser juzgada por cinco locos. Y no es un asunto trivial. Ha llovido desde entonces todo el peso de mi historia, y sin saber muy bien cómo, en estos años, decidí de alguna manera llevarme la vida por delante y intentar discurrir con mayor o menor ingenio las trazas y los modos de ejecutar mis ideas, apurar rápidamente la paciencia y resolver problemas, aproximar el volumen de una vaca al de una esfera, inventar en lo posible y utilizar las técnicas que me dio mi tiempo para aprovechar la naturaleza y vivir en ella; ser, en definitiva, forestal en la medida en que se es Hombre, arrinconando eventualmente la detersiva teoría y mundo de las ideas para meter las manos en el barro y tomar las decisiones como buenamente pude. Y aún así ahora veo que nunca olvidé del todo lo que un día quise ser dentro de aquel desván. Hay un hilo, pues, que recorre entero el laberinto de Dédalo, y que ha mantenido

unidos estos dos mundos, y otros muchos, en mí y a lo largo de mi vida: la aventura del conocimiento teórico y la escuela de la praxis.
En definitiva, el tiempo no es más que espejos que giran reflejando nuestra tierra inalcanzable. De niño tuve la sensación de que el Hombre había nacido en una era dominada por los artrópodos. Ahora, más viejo, más cansado, soy consciente de que en ella también va a extinguirse, y ésta es una de las razones que renuevan mi ilusión y mi interés por el apasionante insectorum sive minimorum animalium Theatrum.
Esta es mi tesis. Nada más. Llevo escribiéndola desde los once años, y la seguiré escribiendo a través de los ojos de mis hijos. Y no sé cómo, pero quiero darles gracias así, defectuosamente, a ellos y a mis padres, a mis hermanos, mis amigos y a mi compañera, por este otro hilo que ha hilvanado toda mi vida, y a mí a ellos.
Al final del camino el Viejo Profesor estaba seguro de que Dios tendría a su lado un sitio reservado para un buen agnóstico. Espero yo que la Ciencia lo tenga para un modesto gestor.

CAPÍTULO 1 INTRODUCIÓN 1 EVOLUCIÓN DE LA DISTRIBUCIÓN GEOGRÁFICA DE NEMATODO DEL PINO (NMP) EN EUROPA 1 EL GÉNERO MONOCHAMUS DEJEAN (COLEOPTERA: CERAMBYCIDAE) 4 MONOCHAMUS COMO VECTOR DE BURSAPHELENCHUS XYLOPHILUS (STEINER & BUHRER) NICKLE 5 BIOLOGÍA DE MONOCHAMUS GALLOPROVINCIALIS 7 SINTOMATOLOGÍA Y DESARROLLO DE LA ENFERMEDAD DEL MARCHITAMIENTO DEL PINO 8 CONTROL DEL NEMATODO DE LA MADERA DEL PINO 9 ESTRUCTURA DE LA TESIS 11 OBJETIVOS DE LA TESIS 13 BIBLIOGRAFÍA 15
CAPÍTULO 2 FENOLOGÍA DE LA EMERGENCIA DE MONOCHAMUS GALLOPROVINCIALIS (COL.: CERAMBYCIDAE) EN LA PENÍNSULA IBÉRICA 21
INTRODUCCIÓN 21 MATERIAL Y MÉTODOS 24
Recolección de material colonizado 24 Cría de los insectos 25 Datos climáticos 25 Análisis estadístico 26
RESULTADOS 29 Periodo de emergencia 29 Proporción de sexos en la emergencia 30 Acumulación de grados día. 33 Re-evaluación de la metodología de cálculo de los umbrales de desarrollo utilizada por NAVES y DE SOUSA (2009) 34
DISCUSIÓN 38 Patrón de emergencia de adultos 38 Proporción de sexos en la emergencia 40 Acumulación de grados día 41
BIBLIOGRAFÍA 45 CAPÍTULO 3 FENOLOGÍA DEL VUELO DE MONOCHAMUS GALLOPROVINCIALIS (COL.: CERAMBYCIDAE) EN LA PENÍNSULA IBÉRICA 49
INTRODUCCIÓN 49 MATERIAL Y MÉTODOS 52
Análisis estadísticos 58 RESULTADOS 59 DISCUSIÓN 70 BIBLIOGRAFÍA 76
CAPÍTULO 4 COMUNIDAD DE COLEÓPTEROS SAPROXÍLICOS ASOCIADA A MONOCHAMUS
GALLOPROVINCIALIS (COL.: CERAMBYCIDAE) EN EL LEVANTE DE LA PENÍNSULA IBÉRICA79 ANTECEDENTES 79 INTRODUCCIÓN 82
Escarabajos saproxílicos, especies invasoras y plagas 82 El binomio Monochamus spp. y Bursaphelenchus xylophilus 82 Redes de muestreo sistemático, redes vigía 83 Estudios ecosistémicos forestales 84
SECCIÓN 4.1. ANÁLISIS DE LA ESTRUCTURA DE LAS COMUNIDADES DE COLEÓPTEROS SAPROXÍLICOS EN LOS PINARES DEL LEVANTE DE LA PÉNÍNSULA IBÉRICA MEDIANTES REDES DE INTERACCIÓN. 86
Introducción 86 Material y métodos 87 Resultados 94
SECCIÓN 4.2 COMUNIDADES DE COLEÓPTEROS SAPROXÍLICOS ASOCIADAS A MONOCHAMUS GALLOPROVINCIALIS (OLIVIER 1795) EN EL LEVANTE DE LA PENÍNSULA IBÉRICA. 99
Introducción 99 Material y Métodos 101

Resultados 104 DISCUSIÓN GENERAL 116 BIBLIOGRAFÍA 123
CAPÍTULO 5 DISPERSIÓN Y POBLACIÓN DE MONOCHAMUS GALLOPROVINCIALIS (COL.: CERAMBYCIDAE) EN LA PENÍNSULA IBÉRICA 129
INTRODUCIÓN 129 SECCIÓN 5.1. RESPUESTA TEMPORAL DE INDIVIDUOS, NATIVOS Y LIBERADOS, DE MONOCHAMUS GALLOPROVINCIALIS A ATRAYENTES CAIROMONALES Y FEROMONALES EN UNA MASA AISLADA. 132
Introducción 132 Material y métodos 134 Resultados 136 Discusión 138
SECCIÓN 5.2 INFLUENCIA DE LA ESTRUCTURA DE LA MASA EN LA DENSIDAD POBLACIONAL DE MONOCHAMUS GALLOPROVINCIALIS 141
Introducción 141 Material y métodos 142 Resultados 144 Discusión 146
SECCIÓN 5.3 INFLUENCIA DEL VIENTO Y LAS VARIABLES DE MASA FORESTAL EN LAS RECAPTURAS DE MONOCHAMUS GALLOPROVINCIALIS 148
Introducción 148 Material y Métodos 149 Resultados 157 Discusión 167
SECCIÓN 5.4 CAPACIDAD DE VUELO A LARGA DISTANCIA EN CONDICIONES NATURALES MONOCHAMUS GALLOPROVINCIALIS 173
Introducción 173 Material y métodos 174 Resultados 180 Discusión 183
BIBLIOGRAFÍA 187 CAPÍTULO 6 SÍNTESIS 193
SECCIÓN 6.1 FENOLOGÍA Y DISPERSIÓN DE MONOCHAMUS GALLOPROVINCIALIS EN LA PENÍNSULA IBÉRICA 193 SECCIÓN 6.2 RECOMENDACIONES PARA LA GESTIÓN DEL NMP EN FUNCIÓN DE SU FENOLOGÍA Y DISPERSIÓN. UN MODELO DE GESTIÓN FORESTAL APLICADO AL CONTROL. 202
Antecedentes 202 Recomendaciones 203
BIBLIOGRAFÍA 211 CAPÍTULO 7 CONCLUSIONES 215 ANEJOS 218

Resumen La aparición y avance de la enfermedad del marchitamiento del pino (Pine Wilt Desease, PWD), causada por Bursaphelenchus xylophilus (Nematoda; Aphelenchoididae), el nematodo de la madera del pino (NMP), en el suroeste de Europa, ha puesto de manifiesto la necesidad de estudiar la fenología y la dispersión de su único vector conocido en Europa, Monochamus galloprovincialis (Col., Cerambycidae).
El análisis de 12 series de emergencias entre 2010 y 2014, registradas en Palencia, València y Teruel, con material procedente de diversos puntos de la península ibérica, demostró una alta variabilidad en la fenología de M. galloprovincialis y la divergencia térmica respecto de las poblaciones portuguesas. Para éstas, el establecimiento de los umbrales térmicos de desarrollo de las larvas post-dormantes del vector (12,2 y 33,5ºC) permitió la predicción de la emergencia mediana para la fecha en la que se acumulaban de 822 grados-día. Ninguna de las series analizadas en este trabajo necesitó de dichos grados-día estimados para la emergencia mediana. Asimismo, la emergencia se adelantó en las regiones más calurosas, mientras que se retrasó en las zonas más templadas. Más allá de la posible variabilidad entre poblaciones locales peninsulares, se detectaron indicios de que la diferencia en la acumulación de calor durante el otoño puede afectar el grado de maduración de las larvas invernantes, y su posterior patrón temporal de emergencia. Por último, también fueron observados comportamientos de protandria en las emergencias.
Respecto a la fenología de su vuelo, entre los años 2010 y 2015, fueron ejecutados un total de 8 experimentos de captura de M. galloprovincialis mediante trampas cebadas con atrayentes en diferentes regiones (Castellón, Teruel, Segovia y Alicante) permitiendo el seguimiento del periodo de vuelo. Su análisis permitió constatar la disminución de las capturas y el acortamiento del periodo de vuelo con la altitud, el inicio del vuelo en el mes de mayo/junio a partir de los 14ºC de temperatura media diaria, la influencia de las altas temperaturas en la disminución de las capturas estivales (potencial causante de perfiles bimodales en las curvas de vuelo en las zonas menos frías), la evolución de la proporción de sexos a lo largo del periodo de vuelo (que muestra una mayor captura de hembras al inicio y de machos al final) y el comportamiento diurno y ligado a las altas temperaturas del vuelo circadiano del insecto.
Dos redes de muestreo sistemático de insectos saproxílicos instaladas en la Comunitat Valencia (Red MUFFET, 15 parcelas, año 2013) y en Murcia (Red ESFP, 20 parcelas, años 2008-2010) permitieron el estudio de la comunidad de insectos relacionada con M. galloprovincialis. Cada una de las parcelas contaba con una trampa cebada con atrayentes y una estación meteorológica. El registro de más de 250 especies de coleópteros saproxílicos demostró el potencial que tiene el empleo de redes de trampas vigía para la detección temprana de organismos exóticos, además de permitir la caracterización y evaluación de las comunidades de entomofauna útil, representando una de las mejores herramientas de la gestión integrada de plagas. En este caso, la comunidad de saproxílicos estudiada mostró ser muy homogénea respecto a la variación ambiental de las zonas de muestreo, y que pese a las pequeñas variaciones entre las comunidades de los diferentes ecosistemas, el rol que M. galloprovincialis desempeña en ellas a lo largo de todo el gradiente estudiado es el mismo. Con todo, el análisis mediante redes de interacción mostró su relevancia ecológica al actuar de conector entre los diferentes niveles tróficos.
Por último, un total de 12 experimentos de marcaje-liberación-recaptura desarrollados entre 2009 y 2012 en Castellón, Teruel, Valencia y Murcia permitieron evaluar el comportamiento dispersivo de M. galloprovincialis. Las detecciones mediante trampas cebadas de los insectos liberados se dieron por lo

menos 8 días después de la emergencia. La abundancia de población pareció relacionada con la continuidad, la naturalización de la masa, y con la afección previa de incendios. La dispersión no estuvo influida por la dirección ni la intensidad de los vientos dominantes. La abundancia de material hospedante (en lo referente a las variables de masa y a los índices de competencia) influyó en la captura del insecto en paisajes fragmentados, aunque la ubicación de las trampas optimizó el número de capturas cuando se ubicaron en el límite de la masa y en zonas visibles. Por último también se constató que M. galloprovincialis posee suficiente capacidad de dispersión como para recorrer hasta 1500 m/día, llegando a alcanzar distancias máximas de 13600m o de 22100 m.

Abstract The detection and expansion of the Pine Wilt Desease (PWD), caused by Bursaphelenchus xylophilus (Nematoda; Aphelenchoididae), Pine Wood Nematode (PWN), in southwestern Europe since 1999, has triggered off the study of the phenology and the dispersion of its unique vector in the continent, Monochamus galloprovincialis (Coleoptera, Cerambycidae).
The analysis of 12 emergence series between 2010 and 2014 registered in Palencia, Teruel and Valencia (Spain), registered from field colonized material collected at several locations of the Iberian Peninsula, showed a high variability in the emergence phenology of M. galloprovincialis. In addition, these patterns showed a very acute thermal divergence regarding a development model fitted earlier in Portugal. Such model forecasted the emergence of 50% of M. galloprovincialis individuals in the Setúbal Peninsula (Portugal) when an average of 822 degree-days (DD) were reached, based on the accumulation of heat from the 1st of March until emergence and lower and upper thresholds of 12.2 ºC and 33,5 °C respectively. In our results, all analyzed series needed less than 822 DD to complete the 50% of the emergence. Also, emergency occurred earlier in the hottest regions, while it was delayed in more temperate areas. Beyond the possible variability between local populations, the difference in the heat accumulation during the fall season may have affected the degree of maturation of overwintering larvae, and subsequently, the temporal pattern of M. galloprovincialis emergences. Therefore these results suggest the need to differentiate local management strategies for the PWN vector, depending on the location, and the climatic variables of each region. Finally, protandrous emergence patterns were observed for M. galloprovincialis in most of the studied data-sets.
Regarding the flight phenology of M. galloprovincialis, a total of 8 trapping experiments were carried out in different regions of the Iberian Peninsula (Castellón, Teruel, Segovia and Alicante) between 2010 and 2015. The use of commercial lures and traps allowed monitoring of the flight period of M. galloprovincialis. The analyses of such curves, helped confirming different aspects. First, a decline in the number of catches and a shortening of the flight period was observed as the altitude increased. Flight period was recorded to start in May / June when the daily average temperature went over 14 ° C. A significant influence of high temperatures on the decrease of catches in the summer was found in many occasions, which frequently lead to a bimodal profile of the flight curves in warm areas. The evolution of sex ratio along the flight period shows a greater capture of females at the beginning of the period, and of males at the end. In addition, the circadian response of M. galloprovincialis to lured traps was described for the first time, concluding that the insect is diurnal and that such response is linked to high temperatures.
Two networks of systematic sampling of saproxylic insects were installed in the Region of Valencia (Red MUFFET, 15 plots, 2013) and Murcia (Red ICPF, 20 plots, 2008-2010). These networks, intended to serve the double purpose of early-detection and long term monitoring of the saproxylic beetle assemblies, allowed the study of insect communities related to M. galloprovincialis. Each of the plots had a trap baited with attractants and a weather station. The registration of almost 300 species of saproxylic beetles demonstrated the potential use of such trapping networks for the early detection of exotic organisms, while at the same time allows the characterization and evaluation of useful entomological fauna communities, representing one of the best tools for the integrated pest management. In this particular case, the studied community of saproxylic beetles was very homogeneous with respect to environmental variation of the

sampling areas, and despite small variations between communities of different ecosystems, the role that M. galloprovincialis apparently plays in them across the studied gradient seems to be the same. However, the analysis through food-webs showed the ecological significance of M. galloprovincialis as a connector between different trophic levels.
Finally, 12 mark-release-recapture experiments were carried out between 2009 and 2012 in Castellón, Teruel, Valencia and Murcia (Spain) with the aim to describe the dispersive behavior of M. galloprovincialis as well as the stand and landscape characteristics that could influence its abundance and dispersal. No insects younger than 8 days were caught in lured traps. Population abundance estimates from mark-release-recapture data, seemed related to forest continuity, naturalization, and to prior presence of forest fires. On the other hand, M. galloprovincialis dispersal was not found to be significantly influenced by the direction and intensity of prevailing winds. The abundance of host material, very related to stand characteristics and spacing indexes, influenced the insect abundance in fragmented landscapes. In addition, the location of the traps optimized the number of catches when they were placed in the edge of the forest stands and in visible positions. Finally it was also found that M. galloprovincialis is able to fly up to 1500 m / day, reaching maximum distances of up to 13600 m or 22100 m.

1
Capítulo 1 Introdución
El siglo XXI parece haber diluido las fronteras físicas del mundo. El espacio que distancia los continentes es cada vez más reducido y apenas unas horas separan las antípodas del planeta. La globalización de la economía y el aumento del comercio internacional han roto todas las barreras que, hasta ahora, habían separado la biota del mundo, durante millones de años de evolución, en diferentes e independientes comunidades, y ha brindado la oportunidad de explorar La Tierra a muchos organismos sedentarios. Materiales inertes y organismos vivos transitan por la superficie del planeta a una velocidad difícil de controlar. Cientos, miles de artrópodos, nematodos, bacterias, virus, hongos van ampliando las fronteras de su hábitat original gracias al Hombre (LEVINE Y D'ANTONIO 2003, HOLMES et al. 2009, HULME 2009, KENIS et al. 2009, MARINI et al. 2011). Aunque una buena parte de ellos, lo hacen de manera imperceptible, inofensiva e invisible a los intereses del ser humano, hay un grupo que constituye un elemento fundamental del cambio global y que contribuye definitivamente a la degradación de los ecosistemas y a la pérdida de nuestro acervo más primario, la biodiversidad: son las especies invasoras (PYŠEK Y RICHARDSON 2010). En los últimos años, y de forma continuada, nuevas plagas han apareciendo en regiones donde no se las esperaba. La plasticidad ecológica con la que proceden, la carencia eventual de enemigos naturales, y la repentina susceptibilidad de hospedantes que no han coevolucionado con ellas abren nuevos horizontes a su persistente expansión (WORK et al. 2005, KIRKENDALL Y FACCOLI 2010). Uno de las invasiones que con mayor impacto ha irrumpido en los bosques europeos durante las últimas décadas, la protagoniza el nematodo de la madera del pino (NMP), Bursaphelenchus xylophilus (Steiner et Buhrer) Nickle (DWINELL 1997, FUTAI 2013), causante de la enfermedad del marchitamiento del pino (Pine Wilt Desease, PWD). Este organismo es transmitido globalmente por algunas especies de escarabajos longicornios del género Monochamus (Col.: Cerambycidae; AKBULUT Y STAMPS 2012). Su detección en Portugal en el año 1999 (MOTA et al. 1999) y la confirmación de la especie M. galloprovincialis (Olivier) como su vector de facto en el continente europeo (SOUSA et al. 2001), supusieron una gran preocupación y la toma de drásticas medidas para la erradicación y contención de la enfermedad, las cuales, más de tres lustros después, no han conseguido arrancarla de raíz ni detener su perseverante propagación por la región occidental de la península ibérica (MAGRAMA 2015). Evolución de la distribución geográfica de nematodo del pino (NMP) en Europa El origen de la enfermedad data de principios del siglo XX. Después de su introducción en Japón en dicha época, el nematodo causó importantes pérdidas de madera, que alcanzaron su punto máximo en 2,4 millones de metros cúbicos en 1979, a la vez que se invertían cerca de 30 millones de dólares americanos anualmente en las medidas de control durante los años siguientes (YOSHIMURA et al. 1999). A partir de entonces, el nematodo de la madera del pino se fue extendiendo hacia China y Corea, en 1982 y 1988, donde se han observado mortandades muy severas (ZHAO et al. 2008). Tras su detección por primera vez en Europa en 1999 (MOTA et al. 1999), y a pesar de los esfuerzos de gestión intensiva realizada por el estado portugués

2
(RODRIGUES 2008), la enfermedad se propagó a través de todo el Portugal continental llegando a penetrar algunas regiones vecinas de España (ABELLEIRA et al. 2011, ROBERTSON et al. 2011, VICENTE et al. 2012, INÁCIO et al. 2015, ZAMORA et al. 2015). En 1999, la prospección realizada inicialmente demostró que la distribución del nematodo no se limitaba a árboles aislados, sino a un área en la región continental de la península de Setúbal, de cerca de 309000 ha que fue delimitada como Zona Restringida (ZR) y que, de acuerdo a las normas establecidas por la Comisión Europea, agrupaba la Zona Afectada (ZA) y una zona de seguridad, al menos 20 km de ancho alrededor de la ZA, llamado Zona de Amortiguamiento o Zona Tampón (ZT). La parte restante del territorio en el que no se detectó NMP fue clasificada como Zona Libre (ZL). En ella fueron realizadas inspecciones periódicas y prospección de los bosques de coníferas de manera intensiva, esencialmente en torno a "zonas de riesgo", que fueron definidas como puntos de entrada, almacenamiento y/o procesamiento de la madera. Con los años, los límites de ZA y ZT se fue modificando, y hubo ampliaciones, especialmente hacia el límite sur y a lo largo de la costa, de manera que la zona geográfica de la ZR evolucionó desde aproximadamente las 309000 hectáreas en 1999/2000 hasta cerca de 1,01 millones de hectáreas en 2006/2007. Esta propagación de la enfermedad llevó a la Comisión Europea a imponer, en 2006/2007, la implementación de un Faja de Contención Fitosanitaria (FCF) en las afueras de la ZR, de forma que quedase libre de coníferas hospedantes de NMP y de su insecto vector, con el fin de reducir al mínimo el riesgo de su propagación natural. No obstante, en 2008, se detectaron nuevos brotes de NMP en varios municipios de la región central en Portugal, anteriormente considerada Zona Libre, y el estado portugués decidió declarar toda la parte continental como Zona Restringida, dejando una Zona Tampón de 20 km de ancho a lo largo de toda la frontera con España. Esto se plasmó en la Ordenanza 553-B / 2008 de 27 de junio, derogada por el Decreto-Ley N ° 95/2011, de 8 de agosto y este último modificado y vuelto a publicar a través del Decreto-Ley N ° 123/2015 de 3 de julio. En este caso, el conjunto de medidas y requisitos para la explotación y la transformación industrial de coníferas fue ampliado al haberse adoptado nuevas directrices, incorporadas en un Programa de Acción renovado, que tuvo en cuenta el conocimiento científico adquirido durante ese periodo y la nueva realidad de la enfermedad en Portugal (Figura 1.1). Por otro lado, en 2009, el NMP fue detectado en Madeira (FONSECA et al. 2012), donde, siguiendo las directrices generales establecidas para el control de este agente nocivo en el territorio continental portugués, también se estableció un Plan de Acción para el Control de NMP en esta Región Autónoma.

3
Figura 1.1 Evolución de las zonas afectadas por el nematodo de la madera del pino (NMP) desde 1999 y la
ubicación del cinturón profiláctico de corta a hecho (FCF) Asimismo, desde el año 2008 han sido detectados cuatro procesos infectivos dentro del territorio español y cerca de la frontera con Portugal (ABELLEIRA et al. 2011, ROBERTSON et al. 2011, ZAMORA et al. 2015). La inmediatez en la detección ha permitido acometer las labores de erradicación y control de un modo efectivo. El primer foco fue detectado en Villanueva de la Sierra, Cáceres, a finales de 2008 y ha sido erradicado completamente tras cuatro años sin nuevos positivos. Mientras que en proceso de erradicación se encuentran los focos de As Neves, en Pontevedra, declarado en 2010; Valverde del Fresno, en Cáceres, declarado en 2012; y Sancti-Spíritus, también en Cáceres, declarado a finales de 2013 (Figura 1.2). Las zonas demarcadas respectivas incluyen el foco y una circunferencia de 14 km. de radio a su alrededor (MAGRAMA 2015).

4
Figura 1.2 Distribución de la enfermedad del machitamiento del pino en la península ibérica.
Por último, los modelos de expansión de la enfermedad publicados hasta la fecha (ROBINET et al. 2011, PUKKALA et al. 2014) prevén un escenario graves daños en para las coníferas de la península ibérica. Y una reciente evaluación del impacto económico cuantitativo de la enfermedad del marchitamiento de pino en Europa, ha pronosticado una pérdida acumulada de 11,9 mil millones de euros entre hasta 2030, sólo en España, con el 67,5% de sus bosques de coníferas severamente afectadas (SOLIMAN et al. 2012). El género Monochamus Dejean (Coleoptera: Cerambycidae) Los vectores de esta enfermedad pertenecen son especies del género Monochamus, perteneciente al orden Coleoptera, familia Cerambycidae, subfamilia Laminae y tribu Monochamini; una de las mayores tribus de Cerambycidae, con más de 600 especies agrupadas dentro de cerca de 100 géneros (DILLON Y DILLON 1941). Tradicionalmente se ha aceptado la inclusión de esta familia en la Superfamilia Chrysomeloidea, formada además por otras dos familias: Chrysomelidae y Bruchidae (CROWSON 1955). La familia Cerambycidae es de hábitos estrictamente fitófagos y muchas de sus especies se alimentan de tejidos vegetales de plantas leñosas. Los cerambícidos asociados a ecosistemas forestales suelen ocupar hábitats saproxílicos, y cumplen funciones ecosistémicas muy importantes. De hecho, su actividad coadyuva al reciclaje de los minerales y su retorno al suelo, ya que sus galerías ayudan a la invasión posterior de otros insectos y hongos de pudrición de la madera, así como a su fragmentación (COULSON Y WITTER 1984, GROVE 2002). El género Monochamus no es una excepción, y tanto la fase adulta como la larvaria se alimentan fundamentalmente de tejidos vegetales. Ninguna de las especies del género Monochamus se reproduce en árboles sanos, sino que sólo se sienten atraídas por árboles muy estresados, moribundos o recién muertos, y por la madera recién talada. Los árboles que llevan muertos desde hace algún tiempo se consideran, por lo general, hospedadores inadecuados (LINSLEY 1959, HELLRIGL 1971, LINIT 1988, AKBULUT Y STAMPS 2012). Las especies de Monochamus ovopositan en hendiduras

5
excavadas en la corteza de los huéspedes adecuados, que las hembras roen con sus mandíbulas. Las larvas recién eclosionadas se alimentan inicialmente de floema y cambium para posteriormente entrar en el xilema y construir una galería que termina en una cámara de pupación, de donde emergen al exterior, después de completada la metamorfosis, a través de un agujero circular (PERSHING Y LINIT 1986). Tras la emergencia, el adulto de Monochamus requiere un período para la maduración sexual. En este periodo se alimenta de braquiblastos, floema y corteza tierna de ramillos y brotes (LINSLEY 1959, HANKS 1999). El género Monochamus se compone de unas 150 especies distribuidas en todo el mundo, aunque con mayor abundancia en África ecuatorial, donde numerosas especies atacan el café y el cacao (HELLRIGL 1971). En términos generales la biología de las especies de Asia, América del Norte y la región eurosiberiana son similares, aunque existen diferencias importantes en la distribución geográfica, la preferencia de hospedadores y lugar de ovoposición, así como en número de estadios larvarios o la longitud del ciclo biológico (HELLRIGL 1971, LINIT 1988, KISHI 1995). En lo que respecta a la península ibérica, de acuerdo con GONZÁLEZ et al. (2007), el género Monochamus se compone exclusivamente de dos especies: Monochamus sutor Linneo 1758, y M. galloprovincialis (Olivier 1795). Monochamus sutor es una especie de dominio paleártico, llegando hasta Japón por Siberia. En la península ibérica es una especie rara que se distribuye únicamente por Pirineos y sus estribaciones. La segunda especie, M. galloprovincialis, es mucho más común y se encuentra distribuida en el área circunmediterránea, Europa central, Cáucaso, Siberia, Mongolia y China. Ambas comparten el mismo grupo filogenético entre las especies descritas para el continente europeo (CESARI et al. 2005). Si bien prácticamente todas las especies de Pinus, e incluso Abies y Picea, han sido descritos como hospedantes, ensayos realizados en masas puras de Pinus pinea L. no han podido detectar ningún M. galloprovincialis (Sánchez-Husillos com. personal).
Monochamus como vector de Bursaphelenchus xylophilus (Steiner & Buhrer) Nickle Hasta la fecha, por tanto, los únicos vectores que se han demostrado efectivos en la trasmisión del NMP, son especies del género Monochamus (KOBAYASHI et al. 1984, LINIT 1988, 1990; KISHI 1995; ZHANG y LINIT 1998)(KOBAYASHI et al. 1984, LINIT 1988, 1990, SOUSA et al. 2001, AKBULUT Y STAMPS 2012). Esta asociación entre cerambícidos y una especie de Bursaphelenchus es rara, ya que la mayoría de las especies de Bursaphelenchus son transmitidos por escolítidos (Scolytinae) y, con menor frecuencia, por otras especies del grupo Curculionidae (BRAASCH 2001, RYSS et al. 2005, PENAS et al. 2006). Las siete de especies que han sido identificados como vectores son: M. carolinensis (Olivier, 1792), M. mutador (LeConte, 1850), M. scutellatus (Say, 1824) y M. titillator (Fabricius, 1775) en América del Norte; M. alternatus (Hope, 1842)y M. saltuarius (Gebler, 1830) en el Noreste de Asia (LINIT 1988, KISHI 1995); y M. galloprovincialis en Europa (SOUSA et al. 2001, AKBULUT Y STAMPS 2012). En América del Norte, M. carolinensis se considera el vector local más importante (LINIT 1987, 1988) mientras que en Japón es M. alternatus (MORIMOTO Y IWASAKI 1972, KOBAYASHI et al. 1984, MAMIYA 1988, KISHI 1995). De acuerdo con Evans et al. (1996), además del ya mencionado M. galloprovincialis, M. sutor, M. sartor (Fabricius 1787), M. urussovi (Fischer 1806) y M . saltuarius son potenciales vectores del NMP. La identificación de M. galloprovincialis como vector europeo de la NMP tuvo lugar tras su detección en Portugal (SOUSA et al. 2001) y actualmente es, de hecho, el vector fundamental de la enfermedad en la península ibérica. La interacción entre ambos organismos es una relación mutualista, obligatoria para el nematodo y facultativa para el escarabajo. Esto es así porque el nematodo necesita del escarabajo para su transporte y dispersión, mientras que el escarabajo se

6
ve beneficiado de la muerte del arbolado provocado del el nematodo ya que supone un incremento en material hospedador donde realizar la ovoposición. El potencial reproductivo de M. galloprovincialis es menor que el de los principales vectores americano y asiático (M. carolinensis y M. alternatus). También la media de nematodos transportados por cada adulto del vector europeo (2000-2300 nematodos por escarabajo) difiere considerablemente de la media transportada por M. carolinensis en EEUU, con 19000 nematodos por escarabajo (NAVES et al. 2006a). Consecuentemente, la tasa de expansión natural de la enfermedad en la península ibérica, mediante este vector, podría ser ligeramente inferior comparada con la dispersión llevada a cabo por sus homólogos en Asia (ROBINET et al. 2009, ROBINET et al. 2011). Las observaciones de NAVES et al. (2006a) al respecto de la distribución de B. xylophilus en el cuerpo de M. galloprovincialis mostraron patrones muy similares a los estudiados para las especies de América del Norte y Asia del Este (LINIT 1988): el 91% de los adultos inmaduros emergidos de trozas infectadas transportaban nematodos y el mayor número de éstos se ubicaban en la región torácica de los escarabajos. En términos generales el nematodo es transportado en las tráqueas y en las invaginaciones del exoesqueleto de los insectos (TOGASHI 1985, 1990). Los periodos de transmisión de nematodos por parte de M. galloprovincialis son similares a los de M. carolinensis (LINIT 1989, 1990), siendo más frecuente durante las primeras 6 semanas después de la emergencia. En concreto la tasa de transmisión del NMP es baja durante los primeros días después de la emergencia del vector y el 50% de las transmisiones exitosas se producen entre dos y cuatro semanas después, disminuyendo drásticamente más adelante, aunque aún se mantenga la posibilidad de encontrar nematodos en el vector (NAVES et al. 2007b). El éxito de esta inoculación de los nematodos en la planta hospedadora es un punto clave en la agresividad de la enfermedad, y básicamente puede darse de dos formas: a través de la alimentación para la maduración sexual (transmisión primaria) o a través de la ovoposición (transmisión secundaria) (LINIT 1988). La transmisión primaria ocurre durante el periodo en que los adultos recién emergidos necesitan unos días de alimentación para alcanzar la madurez sexual. Durante este periodo los escarabajos roen principalmente corteza tierna de brotes y ramillos en árboles sanos. Esta alimentación provoca heridas, que son aprovechadas por las larvas de cuarto estadio del NMP para entrar en el hospedador (MAMIYA Y ENDA 1972, WINGFIELD Y BLANCHETTE 1983, LUZZI et al. 1984, MALEK Y APPLEBY 1984, MAMIYA 1988, LINIT 1990), dejándose caer desde las tráqueas o desde las invaginaciones donde son transportadas. Si el hospedador es susceptible a B. xylophilus, el nematodo se desarrolla a través de su fase fitófaga, alimentándose de los tejidos del xilema y desembocando en un rápido aumento de su población que, eventualmente, provoca una interrupción del flujo de agua y mata al árbol (LINIT 1988, FUTAI 2013). La transmisión secundaria, por ovoposición, es menos importante ya que Monochamus sólo es capaz de hacer las puestas en árboles moribundos o recién muertos, por lo que esta infección por nematodos no puede ser causa de muerte de arbolado, aunque si de reservorio de la enfermedad (LINIT 1988, FUTAI 2013). En términos de peligrosidad de la enfermedad, la transmisión primaria a árboles sanos a través de las heridas provocadas por la alimentación de maduración sexual de Monochamus es verdaderamente la más efectiva, y es especialmente importante en las regiones donde el NMP ha sido introducido, siendo una componente esencial de la epidemiología de la enfermedad del marchitamiento del pino (MAMIYA Y NICKLE 1984, KISHI 1995, YOSHIMURA et al. 1999). En cambio, la transmisión por ovoposición es la forma más frecuente de transmisión en

7
América del Norte, donde es nativo B. xylophilus (WINGFIELD Y BLANCHETTE 1983, LUZZI et al. 1984, DWINELL Y NICKLE 1989, LINIT 1989, EDWARDS Y LINIT 1992) y parece ser el mecanismo original de transmisión en el que se fundamentó la evolución de la asociación entre el agente patógeno, su vector y los hospedadores (DWINELL Y NICKLE 1989). Esto explicaría por qué en estas regiones la enfermedad es mucho menos agresiva y no causa la muerte de los pinos autóctonos (WINGFIELD Y BLANCHETTE 1983, RUTHERFORD Y WEBSTER 1987, DWINELL Y NICKLE 1989, BERGDAHL et al. 1991). No obstante, aunque la transmisión secundaria no dé como resultado la muerte de nuevos hospedadores, no deja de ser un mecanismo de propagación del NMP de cierta relevancia. Esta transmisión secundaria afecta al comercio de la madera desde el área de distribución natural del nematodo (DWINELL Y NICKLE 1989), incluso a áreas potencialmente amenazadas como Escandinavia, donde no se espera que la enfermedad se desarrolle debido a las bajas temperaturas de verano (ØKLAND et al. 2010, SOLIMAN et al. 2012). La propagación natural del NMP entre sus hospedadores está por tanto directamente vinculada a la dispersión de sus vectores. Biología de Monochamus galloprovincialis Monochamus galloprovincialis, como el resto de las especies paleárticas de su género, vive en coníferas moribundas, en declive o recién cortadas, y por lo general es considerada como una plaga secundaria en los bosques. Debido a que presenta principalmente este comportamiento xilófago, M. galloprovincialis, ha sido considerado siempre un parásito secundario, por lo que no ha sido una especie especialmente estudiada hasta la confirmación de su rol como vector del NMP (LINSLEY 1959, HELLRIGL 1971, LINIT 1987, HANKS 1999). En ambientes mediterráneos, M. galloprovincialis es capaz de desarrollar una única generación anual (FRANCARDI Y PENNACCHIO 1996, KOUTROUMPA et al. 2008, NAVES et al. 2008) mientras que TOMMINEN (1993) observó que la mayoría de las larvas completaron su desarrollo en 2 años en condiciones de campo en Finlandia. De acuerdo con estas observaciones, estudios realizados en latitudes intermedias, señalan desarrollos intermedios (KOUTROUMPA et al. 2008). El ciclo biológico de M. galloprovincialis, y en concreto sus patrones de emergencia, han sido estudiados en Setúbal (Portugal) y Orléans (Francia) (NAVES et al. 2007a, KOUTROUMPA et al. 2008, NAVES Y DE SOUSA 2009), mostrando el comienzo de las emergencias en mayo, con un máximo en julio y su finalización en septiembre. Nunca han sido estudiados en España, aunque, desde el punto de vista de la gestión, en varios lugares de este país han sido analizadas sus curvas de vuelo mediante el uso de trampas cebadas con atrayentes cairomonales y feromonales (MAGRAMA 2015). No obstante, aunque la consistencia entre curvas de vuelo y curvas de emergencia es esperable (NAVES et al. 2008), ambas curvas pueden mostrar variaciones muy significativas en la duración (KOUTROUMPA et al. 2008) además de un desfase en el periodo inicial de las capturas de las trampas como consecuencia de la ineficacia en la atracción de inmaduros por parte de los atrayentes comúnmente utilizados, generalmente Galloprotect ® (SEDQ S.L., Barcelona). Por ello el conocimiento de los patrones de emergencia de esta especie a lo largo de la península ibérica continúa siendo necesario para acometer una gestión eficaz. La longevidad de M. galloprovincialis es similar entre machos y hembras, 61 y 64 días de media respectivamente (NAVES et al. 2006b). La tasa de ovoposición de las hembras oscila entre los 37 y los 87 huevos durante su vida fértil (HELLRIGL 1971, FRANCARDI Y PENNACCHIO 1996, NAVES et al. 2006b). Y la duración de su desarrollo larvario parece estar

8
influida por la temperatura, siendo el umbral inferior de desarrollo los 12,2ºC (KOUTROUMPA et al. 2008, NAVES Y DE SOUSA 2009). Los agentes de mortalidad más importantes de M. galloprovincialis, evaluados para la zona afectada por el PWD de Portugal, son el hongo Beauveria bassiana (Bals.-Criv.) Vuill., y la avispa parásita, Cynopterus flavator (Fabricius 1793) (NAVES et al. 2008). No se han encontrado parasitoides relevantes de huevos, aunque las larvas pueden ser parasitadas (en porcentajes bajos) por diferentes bracónidos (Hymenoptera) (NAVES et al. 2005). Diversos trabajos han citado a M. galloprovincialis en sus listados de especies saproxílicas (e.g. LOMBARDERO et al. 2012, ETXEBESTE et al. 2013), sin embargo las relaciones con su entomofauna asociada son esencialmente desconocidas, y es probable que del mismo modo que otras especies de cerambícidos (e. g. DODDS et al. 2001) M. galloprovincialis desempeñe funciones tróficas relevantes en el hábitat saproxílico. Un mayor conocimiento de las interacciones tróficas de las larvas de M. galloprovincialis, así como de su entomofauna asociada, podría ayudar en la mejora de los planes de gestión (KENIS Y HILSZCZANSKI 2004). Aunque M. galloprovincialis puede ser encontrado en todas las especies de pino presentes en la península ibérica, con la mencionada excepción de P. pinea, diferentes estudios en condiciones de laboratorio indican que las especie de pino preferidas tanto para su alimentación como para su ovoposición serían tanto P. pinaster como P. sylvestris (NAVES et al. 2006b, SANCHEZ-HUSILLOS et al. 2013). SANCHEZ-HUSILLOS et al. (2013), mostrando además que M. galloprovincialis es capaz de alimentarse y reproducirse con éxito sobre P. pinea, si bien el vector no ha sido capturado nunca en masas puras de la mencionada especie. Sintomatología y Desarrollo de La Enfermedad del Marchitamiento del Pino Una vez que el NMP ha conseguido salir del vector y acceder a los tejidos vegetales del árbol hospedador, comienza a alimentarse de parénquima y células epiteliales en el tejido leñoso de los canales resiníferos causando una reducción progresiva, y finalmente un cese completo, del flujo de resina. Como consecuencia de esta actividad, a la vez que de la multiplicación y migración del NMP a lo largo del árbol, se da una acumulación progresiva de gran cantidad de sustancias anormales cerca de las membranas de las punteaduras, lo que finalmente provoca el bloqueo del flujo de agua, resultando en la cavitación gradual de las traqueidas y la posterior destrucción de los vasos conductores de agua (FUTAI 2013). El cese de la transpiración hace que las acículas de la copa del árbol viren desde el verde-amarillento hasta el marrón en pocas semanas. En la fase final de la enfermedad, el árbol infectado muere debido a un estrés hídrico severo (MALEK Y APPLEBY 1984, FUTAI 2013). Consecuentemente, la sintomatología de dicha enfermedad es una marchitez caracterizada por los siguientes rasgos:
1. Reducción y cese de la producción de exudados de resina. A mitad de julio los árboles afectados no presentan exudación de resina. En condiciones experimentales los árboles inoculados presentan reducción de resina y dos semanas después de la inoculación cesa completamente su producción.
2. Reducción y cese de la transpiración. La transpiración cesa de 20-30 días después de la inoculación. Durante este periodo no se observa síntomas de la enfermedad.
3. Marchitamiento y amarilleado de las acículas por la disminución de la transpiración. La desecación de la madera y disminución de la humedad en los vasos conductores de savia son las características de la aparición de la enfermedad.

9
4. Muerte del árbol. El árbol enfermo muere de finales de agosto a octubre, alrededor de 30-40 días después de la aparición de los primeros síntomas. No obstante, las acículas marrón-pardas se mantienen en el árbol hasta el próximo verano.
Control del nematodo de la madera del pino Existen diversas razones dificultan el control de la enfermedad ocasionada por el nematodo de la madera del pino: la gran cantidad de hospedadores del organismo patógeno, su extremadamente corto ciclo biológico, su alta fecundidad y fertilidad, la alternancia entre fases fitófaga y micófaga y la resiliencia de sus estadios larvarios. Por otro lado su vector, de hábitos xilófagos, permanece la mayor parte del tiempo oculto en el interior de la madera o debajo de la corteza, refugiado de posibles tratamientos insecticidas ycon una muy baja presión de depredación de sus enemigos naturales conocidos. Por tanto, debido a que el nematodo de la madera de pino permanece constantemente oculto, ya sea en el árbol o el cuerpo de su vector, las estrategias de gestión para reducir el daño de NMP son enfocados necesariamente al control de sus vectores. Varios métodos de control están disponibles actualmente, como las aplicaciones aéreas de plaguicidas (KAMATA 2008), el control biológico utilizando parasitoides, depredadores y hongos y nematodos entomopatógenos (SHIMAZU 2008) y la atracción mediante feromonas (PAJARES et al. 2010, TEALE et al. 2011, MACIAS-SAMANO et al. 2012) o atrayentes (MILLER et al. 2013); pero para optimizar el uso de cualesquiera de estos métodos contra los vectores, resulta esencial el conocimiento de la fenología de M. galloprovincialis en condiciones de campo. De hecho, hasta la fecha, ninguno de los métodos tradicionales de control de plagas y enfermedades ha sido aplicado con éxito en grandes áreas en ninguna de las regiones donde ha sido detectada la enfermedad (ZHAO et al. 2008, VICENTE et al. 2012). El uso de insecticidas en el tratamiento de grandes superficies arboladas ha sido y es aún utilizado ampliamente para el control de los vectores asiáticos en países como Japón, China o Corea. Sustancias químicas como fenitrotión, fentión, diazinone y carbaril se han aplicado por aspersión aérea a finales de verano y otoño (KOBAYASHI et al. 1984, MAMIYA Y NICKLE 1984). En Corea del Sur la pulverización aérea y terrestre de insecticidas para prevenir la alimentación maduración y oviposición de M. alternatus adultos ha sido utilizada para su control en el momento de la emergencia de los adultos (LA et al. 1999, PARK et al. 2014). En consonancia, diversas investigaciones han sido enfocadas a la determinación de modelos fenológicos para optimizar el momento adecuado del tratamiento al objeto de aumentar la eficiencia en el control del vector, minimizando los daños medioambientales (PARK et al. 2014, JUNG et al. 2015). No obstante, esta contaminación medioambiental, sumada a la destrucción de los enemigos naturales o a los cambios provocados en los ecosistemas, fueron la piedra angular de las quejas manifestadas contra las aplicaciones aéreas de insecticidas en Japón (TOGASHI 1990). Hoy en día los insecticidas se utilizan principalmente como un método preventivo para reducir la actividad de alimentación de Monochamus adultos en árboles aislados de alto valor o sobre bosques pequeños y aislados con baja incidencia del marchitamiento (WEBSTER 1999). En el caso europeo, la Directiva 2009/128/CEE, de 21 de octubre de 2009, por la que se establece el marco de la actuación comunitaria para conseguir un uso sostenible de los plaguicidas, indica que la pulverización aérea debe prohibirse en términos generales, aunque permite algunas excepciones en los casos en que ésta presente claras ventajas en términos de menor impacto en la salud humana y el medio ambiente en comparación con otros métodos de pulverización, o cuando no haya ninguna alternativa viable, siempre que se empleen las mejores técnicas disponibles para reducir la deriva durante la aplicación del producto.

10
Ante esta perspectiva, las herramientas de control del NMP aplicada en la actualidad en Europa y España quedan reducidas básicamente a los tratamientos selvícolas y a la inspección. Por un lado, en concreto, la tala y destrucción completa de los pies sintomáticos detectados antes del inicio del periodo de emergencia del insecto para evitar la dispersión de los vectores infectados. Y por otro lado la detección temprana de nuevos focos de la enfermedad o la prospección de sus más probables vías de entrada en los distintos territorios. En este sentido, tanto las redes de alerta temprana (CHADÈS et al. 2011) como los modelos fenológicos son extremadamente interesantes para el aumento de la eficiencia y la eficacia de la gestión de este agente patógeno (NAVES et al. 2007a, KOUTROUMPA et al. 2008, NAVES Y DE SOUSA 2009, PARK et al. 2014, JUNG et al. 2015). No obstante, además de la eliminación de los árboles infectados, el uso de trampas cebadas con atrayentes para la captura de adultos de M. galloprovincialis durante la temporada de vuelo ha sido propuesto para el control de la enfermedad en estudios recientes (SANCHEZ-HUSILLOS et al. 2015, TORRES-VILA et al. 2015). En este sentido, el complejo de sustancias con mayor actividad atractiva incluye un compuestos cairomonales emitidos por el hospedador ( pineno) y dos cairomonas emitidas por escolítidos del género Ips y Orthotomicus (ipsenol y 2-metil-3-buten-2-ol) (IBEAS et al. 2007, PAJARES et al. 2010, ETXEBESTE et al. 2012). Estos trabajos han permitido desarrollar un atrayente cairomonal muy eficaz en la captura de M. galloprovincialis, mejorado posteriormente con el descubrimiento de una feromona de agregación emitida por los machos de esta especie (PAJARES et al. 2010). Con ello ha podido desarrollarse y comercializarse un cebo cairo-feromonal (SEDQ, Barcelona) que permite una alta tasa de capturas de ambos sexos de M. galloprovincialis durante prácticamente todo su periodo de vuelo. Su utilización puede ser enfocada, por tanto, a la monitorización de sus periodos de vuelo, a la reducción de la población de M. galloprovincialis en determinadas zonas sensibles (SANCHEZ-HUSILLOS et al. 2015) y al desarrollo de nuevas líneas de investigación que aporten conocimientos sobre aspectos tan trascendentes como su dispersión, entre otros. Como se ha comentado con anterioridad, la propagación del NMP entre sus hospedadores queda limitada a la dispersión de sus vectores. Si bien la dispersión natural de Monochamus no deja de ser un proceso esencial en la biología de la especie, no sólo para evitar la endogamia poblacional, sino también para localizar árboles hospedadores susceptibles de ataque (CLOBERT et al. 2012, DAVID et al. 2014, ETXEBESTE et al. 2015), su rol como vector de B. xylophilus le confiere un interés especial desde el punto de vista de la sanidad forestal. Además, en este caso, a la capacidad natural de dispersión del vector hay que añadirle también la dispersión antropocórica sensu lato. Y en este sentido conviene destacar que el actual aumento del flujo de materiales y personas a lo ancho del planeta, y la globalización, en definitiva, del comercio internacional, ha impulsado la aparición de especies invasoras en todos los continentes (LEVINE Y D'ANTONIO 2003, ROBINET et al. 2009, ROBINET et al. 2011), siendo éstas invasiones consideradas una de las principales causas de pérdida de la biodiversidad, además de originar grandes daños económicos y ambientales. Si bien la expansión local de la enfermedad se ve afectada por la capacidad dispersiva a corta distancia (short distance dispersal, SDD) de Monochamus (TAKASU et al. 2000, ROBINET et al. 2011), los eventos de dispersión a larga distancia (long distance dispersal, LDD) tienen una importancia desmesurada cuando se trata de especies invasoras o de cuarentena, y más aún cuando se trata de vectores de organismos patógenos exóticos (NATHAN et al. 2003, CLOBERT et al. 2012, NATHAN et al. 2012). Estos eventos LDD se caracterizan por un alto grado de aleatoriedad, y los registros a menudo proceden de conjuntos de datos anecdóticos, incompletos, y potencialmente sesgados, lo cual, pese a representar ejemplos manifiestos de la

11
capacidad dispersiva de estos organismos, hace muy difícil su caracterización (TURCHIN 1998, NATHAN et al. 2003). En referencia a esto, la capacidad dispersiva natural de M. galloprovincialis es potencialmente muy alta, de acuerdo con David et al (2014), demostrando mediante experimentos de laboratorio que estos insectos son capaces de acumular distancias de vuelo promedio de aproximadamente 16 km a lo largo de su vida, habiéndose observado incluso distancias de vuelo puntuales mucho mayores, como el caso de un individuo que mostró una capacidad de volar hasta 62,72 kilometros, siempre bajo condiciones de laboratorio (DAVID et al. 2014).
Estructura de la tesis
Este trabajo de investigación consta de dos partes fundamentales que abordan el estudio de la biología M. galloprovincialis desde dos perspectivas diferentes: su fenología y su dispersión. La importancia de ambas en lo que respecta al control de la enfermedad que vectorizan es de primera magnitud y su impacto en la gestión real de este organismo patógeno de cuarentena es insoslayable. Todos los trabajos que componen esta investigación fueron realizados en un contexto temporal muy concreto, que se inicia con el desarrollo de un atrayente cairomonal y feromonal eficaz para la captura de M. galloprovincialis (PAJARES et al. 2010), y con la mejora de la metodología de trampeo para la captura de este insecto (ÁLVAREZ et al. 2015). Estos dos hitos suponen en sí la puesta a punto de una herramienta eficaz que permite, además de su aplicación directa en la gestión, la posibilidad de realizar estudios científicos que aporten información sobre la fenología de los vuelos, la densidad de población o la dispersión de esta especie, hasta entonces muy difíciles de acometer. Además, como consecuencia del diseño de estos experimentos, una gran cantidad de insectos tuvieron que ser criados en condiciones de laboratorio durante esos años, lo cual permitió investigar también la fenología de su emergencia. Asimismo estos trabajos fueron realizados en un contexto espacial también muy concreto: las reuniones de los extintos talleres de feromonas organizados por el MAGRAMA (2015) para la puesta en común de resultados y el diseño consensuado de experimentos en diferentes regiones de la península ibérica y llevados a cabo por diferentes administraciones autonómicas para dar mayor representatividad a los mismos. En definitiva, durante esos años, diversas líneas de investigación fueron abiertas en España y Portugal como consecuencia de la aparición de una tecnología que las posibilitaba, y en respuesta a una perentoria necesidad de dar herramientas para control de la enfermedad en la península ibérica. La primera parte de esta memoria, dedicada a la fenología del insecto, consta de tres trabajos diferentes. El primero de ellos trata sobre la fenología de su emergencia. En él se analizaron doce curvas de emergencias distribuidas a lo largo de la península con la intención de determinar si la fenología de la emergencia de M. galloprovincialis era homogénea en todo el territorio peninsular y si, por tanto, los modelos predictivos publicados por (NAVES Y DE SOUSA 2009) para la Península de Setúbal (punto de entrada de la enfermedad en Europa) eran válidos para predecirlas en otras regiones.

12
El segundo trabajo está dedicado al estudio de la fenología de vuelo en la península ibérica. Una amplia serie de experimentos de trampeo de M. galloprovincialis fueron instalados a lo largo de la península con el objetivo de estudiar las poblaciones del insecto en esas zonas, su capacidad dispersiva y su fenología de vuelo. La contabilización de las capturas de las trampas permitió tasar las fechas en las que se inicia y se acaba el periodo de vuelo del insecto y evaluar su relación con el clima o con las características del medio. La obviedad de que mucha fauna beneficiosa era capturada en toda Europa como efecto colateral de que el atrayente de M. galloprovincialis tuviese un componente cairomonal, así como la importancia que en los últimos tiempos empezaron a adquirir las redes de alerta temprana como herramienta fundamental para la gestión de la fauna invasora, propiciaron la instalación de diferentes redes de muestreo permanente en la región levantina de la península ibérica. Algunas de estas redes (Red EFSP y Red MUFFET) aportarán una importante y cuantiosa información sobre la biodiversidad de coleópteros saproxílicos de los pinares mediterráneos de esta región cuando puedan ser analizadas series temporales más largas. Hasta la fecha, pese a los pocos años de muestreo, se ha podido realizar una primera aproximación a los datos con el objetivo de determinar el papel de M. galloprovincialis en la comunidad de insectos a la que está asociado, los cuales quedan incluidos en el tercer trabajo que cierra la primera parte de esta tesis. El segundo bloque fundamental de esta memoria engloba toda una serie de experimentos realizados con el objetivo de conocer aspectos sobre la densidad de población o la dispersión de M. galloprovincialis en el levante de la península ibérica. Formado por un único capítulo dividido en cuatro secciones, en la primera de ellas se analiza la respuesta temporal del insecto a los atrayentes cairomonales y feomonales utilizados, toda vez que esta respuesta no es inmediata tras la emergencia sino que parece condicionada a la madurez sexual de los individuos. La segunda sección analiza la influencia de la estructura de la masa en la densidad de población de M. galloprovincialis. Una tercera sección está dedicada al estudio de la influencia del viento y de las diferentes variables que caracterizan las masas forestales en las capturas y en la dispersión de M. galloprovincialis, mientras que la cuarta y última sección se centra en el análisis de su dispersión a larga distancia. En resumen, la normativa europea anterior al año 2012 (Decisión de la Comisión 133/2006/CE) obligaba a la creación de una faja profiláctica de 3 km de radio alrededor de cualquier detección de la enfermedad del marchitamiento del pino en España dentro la cual todas las coníferas debían ser cortadas y destruidas in situ. La distancia del radio de corta se basaba en la única y antigua publicación científica existente hasta ese momento sobre la capacidad de dispersión a larga distancia de los insectos vectores de la enfermedad (SHIBATA 1986). La necesidad de aportar conocimiento sobre este asunto concreto llevó al diseño e instalación de una larga serie de más de 10 experimentos, entre los años 2010 y 2012 en el levante peninsular, enfocados a resolver preguntas sobre su densidad de población, capacidad de vuelo y su comportamiento dispersivo. Su análisis compone el cuerpo fundamental de esta tesis, y la secuencia temporal de los ensayos es fundamental para comprender la investigación en su conjunto, ya que los resultados de los experimentos de un año fueron convocando el diseño experimental de los de los años posteriores, por lo que su lectura en este momento concreto debería ser realizada desde una perspectiva diacrónica.

13
La metodología fue común a todos los experimentos. Se trató esencialmente de experimentos de marcaje-liberación- recaptura de M. galloprovincialis, diferenciándose entre ellos exclusivamente en la cantidad y distribución espacial de las trampas, las características (paisajísticas y estructurales) de las áreas de muestreo, el origen de los insectos liberados (nativos o criado en laboratorio) o la ubicación relativa del punto de liberación. Los resultados más relevantes fueron urgentemente comunicados en congresos nacionales o internacionales sobre la materia. Por último, incluido en un capítulo de síntesis de todos los resultados aportados por este trabajo de investigación, se añade un apartado con recomendaciones para la gestión apoyadas en los resultados obtenidos, con la diáfana intención de darle un contenido práctico a un lustro de trabajo de investigación en el Laboratori de Sanitat Forestal de la Generalitat Valenciana que ha desembocado en la redacción de esta tesis doctoral.
Objetivos de la tesis
Desde la detección del primer proceso infectivo dentro del territorio europeo, las medidas, tanto para la erradicación como para la prevención de la enfermedad del marchitamiento del pino en Europa, han ido adaptándose y acoplándose al nuevo conocimiento que los esfuerzos de investigación iban aportando. Las diferentes velocidades de la investigación científica y la gestión a menudo desembocan en desfases y falta de acoples entre el conocimiento objetivo y la aplicación de medidas de gestión actualizadas. En el caso concreto del NMP, diferentes normativas europeas y nacionales han ido sucediéndose desde la primera detección en 1999. En los últimos quince años una gran cantidad de información ha sido publicada en el foro de discusión científica y, como consecuencia, muchas de las medidas originales han sido ajustadas o desestimadas, así como nuevas propuestas han aparecido en la gestión de esta enfermedad. Actualmente toda la normativa, y en definitiva la mayoría de los esfuerzos de contención práctica de la enfermedad, han ido centrándose principalmente en la gestión de su vector, M. galloprovincialis. Esta especie ha sido considerablemente bien estudiada en muchísimos aspectos y ha sido mucha la información que han aportado los científicos portugueses sobre todo lo relacionado con su biología y su rol como vector de B. xylophilus. No obstante aún queda mucho trabajo por hacer hasta la consecución de un conocimiento razonablemente exhaustivo y útil sobre su comportamiento, que desemboque en una transferencia tecnológica y de conocimiento que mejore el control de la dispersión de la enfermedad. Algunas de las lagunas de conocimiento que aún existen, y en especial en el territorio con mayor potencialidad de dispersión de la enfermedad en Europa, la península ibérica, están relacionadas con el conocimiento de su fenología, tanto en lo referido a su patrón de emergencias como en lo referido a sus curvas de vuelo. También son desconocidas sus relaciones con otros organismos con los que convive, y se conoce poco de su capacidad de dispersión natural. Esto cobra especial relevancia cuando, como se ha comentado en apartados anteriores, las principales medidas de erradicación y contención tienen mucho que ver con el ajuste de un calendario de actuaciones silvícolas a la fenología del insecto, y con el ajuste de la planificación territorial de los trabajos a su capacidad de dispersión natural. El objetivo general de esta tesis es aportar conocimiento sobre estos aspectos de tantísima relevancia práctica que permita mejorar las herramientas de gestión de una de las enfermedades

14
de cuarentena más peligrosas para los ecosistemas forestales europeos. De forma detallada los objetivos particulares se resumirían de la siguiente forma: Bloque A. Fenología de M. galloprovincialis A. 1 Fenología de la emergencia de M. galloprovincialis
- Describir la variabilidad temporal de las emergencias de M. galloprovincialis a lo largo de la península ibérica
- Evaluar los modelos de desarrollo basados en la acumulación de grados día existentes para M. galloprovincialis
- Describir la evolución de la sex-ratio en las emergencias de M. galloprovincialis A. 2 Fenología de vuelo de M. galloprovincialis
- Describir la incidencia de la altura y la temperatura en el vuelo de M. galloprovincialis
- Describir las temperaturas mínimas requeridas para el inicio del vuelo de M. galloprovincialis
- Describir el ritmo circadiano en las capturas de M. galloprovincialis - Describir la evolución de la sex-ratio en las capturas de M. galloprovincialis ¨
A. 3 Comunidad de coleópteros saproxílicos asociada a M. galloprovincialis - Evaluar el potencial de las redes de trampas vigía en la detección temprana de
organismos exóticos - Describir la comunidad asociada y la distribución de M. galloprovincialis a lo largo
de los pinares del este y sudeste ibérico - Estudiar el papel desempeñado por M. galloprovincialis en las redes tróficas de
insectos saproxílicos
Bloque B. Población y despersión de M. galloprovincialis B. 1 Niveles poblacionales de M. galloprovincialis
- Estudiar el tamaño de las poblaciones de M. galloprovincialis en relación a la fragmentación y tamaño de las masas forestales
- Estudiar las características de las masas forestales que afectan a la abundancia de M. galloprovincialis
B. 2 Comportamiento dispersivo en inmaduros de M. galloprovincialis - Estudiar la respuesta de M. galloprovincialis recientemente emergidos a los cebos
comerciales para la especie - Estudiar la capacidad de dispersión en inmaduros de M. galloprovincialis en
condiciones de ausencia de hospedante B. 3 Comportamiento dispersivo en maduros de M. galloprovincialis
- Estudiar la influencia del viento en la dispersión de M. galloprovincialis - Estudiar la influencia de la configuración paisajística en la dispersión de M.
galloprovincialis - Describir los patrones de dispersión a corta y larga distancia en M. galloprovincialis - Describir las características de las masas forestales que afectan la recaptura de M.
galloprovincialis liberados
Bloque C. Síntesis y recomendaciones para la gestión - Describir recomendaciones para la mejora de la gestión del NMP

15
Bibliografía
ABELLEIRA, A., A. PICOAGA, J. P. MANSILLA y O. AGUIN (2011). Detection of
Bursaphelenchus xylophilus, Causal Agent of Pine Wilt Disease on Pinus pinaster in Northwestern Spain. Plant Disease 95(6): 776-776.
AKBULUT, S. y W. T. STAMPS (2012). Insect vectors of the pinewood nematode: a review of the biology and ecology of Monochamus species. Forest Pathology 42(2): 89-99.
ÁLVAREZ, G., I. ETXEBESTE, D. GALLEGO, G. DAVID, L. BONIFACIO, H. JACTEL, E. SOUSA y J. A. PAJARES (2015). Optimization of traps for live trapping of Pine Wood Nematode vector Monochamus galloprovincialis. Journal of Applied Entomology 139(8): 618-626.
BERGDAHL, D., S. HALIK, J. TOMMINEN y H. AKAR (1991). Frequency of infestation of Monochamus notatus and M. scutellatus by Bursaphelenchus xylophilus in Vermont. Phytopathology 81: 120.
BRAASCH, H. (2001). Bursaphelenchus species in conifers in Europe: distribution and morphological relationships. EPPO bulletin 31(2): 127-142.
CESARI, M., O. MARESCALCHI, V. FRANCARDI y B. MANTOVANI (2005). Taxonomy and phylogeny of European Monochamus species: first molecular and karyological data. Journal of Zoological Systematics and Evolutionary Research 43(1): 1-7.
CHADÈS, I., T. G. MARTIN, S. NICOL, M. A. BURGMAN, H. P. POSSINGHAM y Y. M. BUCKLEY (2011). General rules for managing and surveying networks of pests, diseases, and endangered species. Proceedings of the National Academy of Sciences 108(20): 8323-8328.
CLOBERT, J., M. BAGUETTE, T. G. BENTON, J. M. BULLOCK y S. DUCATEZ (2012). Dispersal ecology and evolution, Oxford University Press.
COULSON, R. N. y J. A. WITTER (1984). Forest entomology: ecology and management, John Wiley & Sons.
CROWSON, R. (1955). The Natural Classification of the Families of Coleoptera. . London, Nathaniel Lloyd.
DAVID, G., B. GIFFARD, D. PIOU y H. JACTEL (2014). Dispersal capacity of Monochamus galloprovincialis, the European vector of the pine wood nematode, on flight mills. Journal of Applied Entomology 138(8): 566-576.
DILLON, L. S. y E. S. DILLON (1941). The tribe Monochamini in the western hemisphere (Coleoptera: Cerambycidae). Reading Public Museum and Art Gallery Scientific Publications(1).
DODDS, K. J., C. GRABER y F. M. STEPHEN (2001). Facultative intraguild predation by larval Cerambycidae (Coleoptera) on bark beetle larvae (Coleoptera : Scolytidae). Environmental Entomology 30(1): 17-22.
DWINELL, L. D. (1997). THE PINEWOOD NEMATODE:Regulation and Mitigation. Annual Review of Phytopathology 35(1): 153-166.
DWINELL, L. D. y W. R. NICKLE (1989). An overview of the pine wood nematode ban in North America. General Technical Report-Southeastern Forest Experiment Station, USDA Forest Service(SE-55).
EDWARDS, O. y M. LINIT (1992). Transmission of Bursaphelenchus xylophilus through oviposition wounds of Monochamus carolinensis (Coleoptera: Cerambycidae). Journal of Nematology 24(1): 133.
ETXEBESTE, I., G. ÁLVAREZ, G. PÉREZ y J. A. PAJARES (2012). Field response of the six-toothed pine bark beetle, Ips sexdentatus (Col.: Curculionidae, Scolytinae), to pheromonal blend candidates. Journal of Applied Entomology 136: 431-444.
ETXEBESTE, I., J. L. LENCINA y J. PAJARES (2013). Saproxylic community, guild and species responses to varying pheromone components of a pine bark beetle. Bulletin of Entomological Research 103: 497-510.

16
ETXEBESTE, I., E. SANCHEZ-HUSILLOS, G. ÁLVAREZ, H. MAS I GISBERT y J. PAJARES (2015). Dispersal of Monochamus galloprovincialis (Col.: Cerambycidae) as recorded by mark-release recapture using pheromone traps. Journal of Applied Entomology: In press.
FONSECA, L., J. M. S. CARDOSO, A. LOPES, M. PESTANA, F. ABREU, N. NUNES, M. MOTA y I. ABRANTES (2012). The pinewood nematode, Bursaphelenchus xylophilus, in Madeira Island. Helminthologia 49(2): 96-103.
FRANCARDI, V. y F. PENNACCHIO (1996). Note sulla bioecologia di Monochamus galloprovincialis (Olivier) in Toscana e in Liguria (Coleoptera Cerambycidae). Redia 79: 153-169.
FUTAI, K. (2013). Pine Wood Nematode, Bursaphelenchus xylophilus. Annual Review of Phytopathology 51(1): 61-83.
GONZÁLEZ, C., E. VIVES y A. ZUZARTE (2007). Nuevo catálogo de los Cerambycidae (Coleoptera) de la Península Ibérica, islas Baleares e islas atlánticas: Canarias, Açores y Madeira. Monografías SEA 12: 1-211.
GROVE, S. J. (2002). Saproxylic insect ecology and the sustainable management of forests. Annual Review of Ecology and Systematics 33: 1-23.
HANKS, L. (1999). Influence of the larval host plant on reproductive strategies of cerambycid beetles. Annual review of entomology 44(1): 483-505.
HELLRIGL, K. G. (1971). Die Bionomie der europaischen Monochamus-Arten (Coleopt., Cerambycid.) und ihre Bedeutung fur die Forstund Holzwirtschaft. Redia 52: 367-509.
HOLMES, T. P., J. E. AUKEMA, B. VON HOLLE, A. LIEBHOLD y E. SILLS (2009). Economic impacts of invasive species in forests. Annals of the New York Academy of Sciences 1162(1): 18-38.
HULME, P. E. (2009). Trade, transport and trouble: managing invasive species pathways in an era of globalization. Journal of Applied Ecology 46(1): 10-18.
IBEAS, F., D. GALLEGO, J. DIEZ y J. PAJARES (2007). An operative kairomonal lure for managing pine sawyer beetle Monochamus galloprovincialis (Coleoptera: Cerymbycidae). Journal of Applied Entomology 131(1): 13-20.
INÁCIO, M. L., F. NÓBREGA, P. VIEIRA, L. BONIFÁCIO, P. NAVES, E. SOUSA y M. MOTA (2015). First detection of Bursaphelenchus xylophilus associated with Pinus nigra in Portugal and in Europe. Forest Pathology 45(3): 235-238.
JUNG, C. S., S.-H. KOH, Y. NAM, J. J. AHN, C. Y. LEE y W. I. CHOI (2015). A Model for Predicting Spring Emergence of Monochamus saltuarius (Coleoptera: Cerambycidae) from Korean white pine, Pinus koraiensis.
KAMATA, N. (2008). Integrated pest management of pine wilt disease in Japan: Tactics and strategies. Pine wilt disease, Springer: 304-322.
KENIS, M., M.-A. AUGER-ROZENBERG, A. ROQUES, L. TIMMS, C. PÉRÉ, M. J. COCK, J. SETTELE, S. AUGUSTIN y C. LOPEZ-VAAMONDE (2009). Ecological effects of invasive alien insects. Ecological Impacts of Non-Native Invertebrates and Fungi on Terrestrial Ecosystems, Springer: 21-45.
KENIS, M. y J. HILSZCZANSKI (2004). Natural Enemies of Cerambycidae and Buprestidae Infesting Living Trees. Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis. F. Lieutier, K. R. Day, A. Battisti, J.-C. Grégoire y H. F. Evans. Dordrecht, Kluwer Academic Publishers: 475-498.
KIRKENDALL, L. R. y M. FACCOLI (2010). Bark beetles and pinhole borers (Curculionidae, Scolytinae, Platypodinae) alien to Europe. ZooKeys(56): 227.
KISHI, Y. (1995). Pine wood nematode and the Japanese pine sawyer, Thomas Company Limited.
KOBAYASHI, F., A. YAMANE y T. IKEDA (1984). The japanese pine sawyer beetle as the vector of pine wilt disease. Annual review of entomology 29(1): 115-135.

17
KOUTROUMPA, F. A., B. VINCENT, G. ROUX-MORABITO, C. MARTIN y F. LIEUTIER (2008). Fecundity and larval development of Monochamus galloprovincialis (Coleoptera Cerambycidae) in experimental breeding. Annals of Forest Science 65(7).
LA, Y., Y. MOON, W. YEO, S. SHIN y W. BAK (1999). Recent status of pine wilt disease in Korea. Sustainability of pine forests in relation to pine wilt and decline. Proceedings of International Symposium, Tokyo, Japan, 27-28 October, 1998., Shokado Shoten.
LEVINE, J. M. y C. M. D'ANTONIO (2003). Forecasting biological invasions with increasing international trade. Conservation Biology 17(1): 322-326.
LINIT, M. (1987). The insect component of pine wilt disease in the United States. Pathogenicity of the pine wood nematode: 66-73.
LINIT, M. (1988). Nematode Vector Relationships in the Pine Wilt Disease System. Journal of Nematology 20(2): 227-235.
LINIT, M. (1989). Temporal pattern of pinewood nematode exit from the insect vector Monochamus carolinensis. Journal of Nematology 21(1): 105.
LINIT, M. (1990). Transmission of pinewood nematode through feeding wounds of Monochamus carolinensis (Coleoptera: Cerambycidae). Journal of Nematology 22(2): 231.
LINSLEY, E. G. (1959). Ecology of cerambycidae. Annual review of entomology 4(1): 99-138. LOMBARDERO, M. J., M. ALONSO-RODRÍGUEZ y E. P. ROCA-POSADA (2012). Tree
insects and pathogens display opposite tendencies to attack native vs. non-native pines. Forest Ecology and Management 281: 121-129.
LUZZI, M., R. WILKINSON y A. TARJAN (1984). Transmission of the pinewood nematode, Bursaphelenchus xylophilus, to slash pine trees and log bolts by a cerambycid beetle, Monochamus titillator, in Florida. Journal of Nematology 16(1): 37.
MACIAS-SAMANO, J. E., D. WAKARCHUK, J. G. MILLAR y L. M. HANKS (2012). 2-Undecyloxy-1-ethanol in combination with other semiochemicals attracts three Monochamus species (Coleoptera: Cerambycidae) in British Columbia, Canada. The Canadian Entomologist 144(06): 764-768.
MAGRAMA. (2015). "Nematodo de la madera del pino." 2015, de http://www.magrama.gob.es/es/agricultura/temas/sanidad-vegetal/nematodo-de-la-madera-del-pino/#.
MALEK, R. B. y J. E. APPLEBY (1984). Epidemiology of pine wilt in Illinois. Disease distribution. Plant Disease 68(3): 180-186.
MAMIYA, Y. (1988). History of Pine Wilt Disease in Japan. Journal of Nematology 20(2): 219-226.
MAMIYA, Y. y N. ENDA (1972). Transmission of Bursaphelenchus lignicolus (Nematoda: Aphelenchoididae) by Monochamus alternatus (Coleoptera: Cerambycidae). Nematologica 18(2): 159-162.
MAMIYA, Y. y W. NICKLE (1984). The pine wood nematode. Plant and insect nematodes.: 589-626.
MARINI, L., R. A. HAACK, R. J. RABAGLIA, E. P. TOFFOLO, A. BATTISTI y M. FACCOLI (2011). Exploring associations between international trade and environmental factors with establishment patterns of exotic Scolytinae. Biological invasions 13(10): 2275-2288.
MILLER, D. R., C. M. CROWE, B. F. BARNES, K. J. GANDHI y D. A. DUERR (2013). Attaching lures to multiple-funnel traps targeting saproxylic beetles (Coleoptera) in pine stands: inside or outside funnels? Journal of economic entomology 106(1): 206-214.
MORIMOTO, K. y A. IWASAKI (1972). Role of Monochamus alternatus (Coleoptera: Cerambycidae) as a vector of Bursaphelenchus lignicolus (Nematoda: Aphelenchoididae). Journal of the Japanese Forestry Society 54(6): 177-183.
MOTA, M. M., H. BRAASCH, M. A. BRAVO, A. C. PENAS, W. BURGERMEISTER, K. METGE y E. SOUSA (1999). First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1: 727-734.

18
NATHAN, R., E. KLEIN, J. ROBLEDO-ARNUNCIO y E. REVILLA (2012). Dispersal kernels: review. Dispersal Ecology and Evolution. Oxford University Press. Oxford. pp: 187-210.
NATHAN, R., G. PERRY, J. T. CRONIN, A. E. STRAND y M. L. CAIN (2003). Methods for estimating long‐distance dispersal. Oikos 103(2): 261-273.
NAVES, P., S. CAMACHO, E. DE SOUSA y J. QUARTAU (2007a). Transmission of the pine wood nematode Bursaphelenchus xylophilus through feeding activity of Monochamus galloprovincialis (Col., Cerambycidae). Journal of Applied Entomology 131(1): 21-25.
NAVES, P., M. KENIS y E. SOUSA (2005). Parasitoids associated with Monochamus galloprovincialis (Oliv.)(Coleoptera: Cerambycidae) within the pine wilt nematode-affected zone in Portugal. Journal of Pest Science 78(2): 57-62.
NAVES, P. M., S. CAMACHO, E. M. DE SOUSA y J. A. QUARTAU (2006a). Entrance and distribution of the pinewood nematode Bursaphelenchus xylophilus on the body of its vector Monochamus galloprovincialis (Coleoptera: Cerambycidae). Entomologia generalis 29(1): 71-80.
NAVES, P. M., S. CAMACHO, E. M. DE SOUSA y J. A. QUARTAU (2007b). Transmission of the pine wood nematode Bursaphelenchus xylophilus through feeding activity of Monochamus galloprovincialis (Col., Cerambycidae). Journal of Applied Entomology 131(1): 21-25.
NAVES, P. M. y E. M. DE SOUSA (2009). Threshold temperatures and degree-day estimates for development of post-dormancy larvae of Monochamus galloprovincialis (Coleoptera: Cerambycidae). Journal of Pest Science 82(1): 1-6.
NAVES, P. M., E. M. DE SOUSA y J. A. QUARTAU (2006b). Feeding and oviposition preferences of Monochamus galloprovincialis for certain conifers under laboratory conditions. Entomologia experimentalis et applicata 120(2): 99-104.
NAVES, P. M., E. SOUSA y J. M. RODRIGUES (2008). Biology of Monochamus galloprovincialis (Coleoptera: Cerambycidae) in the pine wilt disease affected zone, Southern Portugal. Silva Lusitana 16(2): 133-148.
ØKLAND, B., O. SKARPAAS, M. SCHROEDER, C. MAGNUSSON, Å. LINDELÖW y K. THUNES (2010). Is eradication of the pinewood nematode (Bursaphelenchus xylophilus) likely? An evaluation of current contingency plans. Risk analysis 30(9): 1424-1439.
PAJARES, J. A., G. ALVAREZ, F. IBEAS, D. GALLEGO, D. R. HALL y D. I. FARMAN (2010). Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. Journal of Chemical Ecology 36(6): 570-583.
PARK, C., D. KIM, S. LEE, Y. MOON, Y. CHUNG y D. S. KIM (2014). A forecasting model for the adult emergence of overwintered Monochamus alternatus (Coleoptera: Cerambycidae) larvae based on degree-days in Korea. Applied Entomology and Zoology 49(1): 35-42.
PENAS, A. C., M. A. BRAVO, P. NAVES, L. BONIFÁCIO, E. SOUSA y M. MOTA (2006). Species of Bursaphelenchus Fuchs, 1937 (Nematoda: Parasitaphelenchidae) and other nematode genera associated with insects from Pinus pinaster in Portugal. Annals of Applied Biology 148(2): 121-131.
PERSHING, J. y M. LINIT (1986). Biology of Monochamus carolinensis (Coleoptera: Cerambycidae) on scotch pine in Missouri. Journal of the Kansas Entomological Society: 706-711.
PUKKALA, T., T. MOYKKYNEN y C. ROBINET (2014). Comparison of the potential spread of pinewood nematode (Bursaphelenchus xylophilus) in Finland and Iberia simulated with a cellular automaton model. Forest Pathology 44(5): 341-352.
PYŠEK, P. y D. M. RICHARDSON (2010). Invasive species, environmental change and management, and health. Annual Review of Environment and Resources 35: 25-55.

19
ROBERTSON, L., S. C. ARCOS, M. ESCUER, R. S. MERINO, G. ESPARRAGO, A. ABELLERA y A. NAVAS (2011). Incidence of the pinewood nematode Bursaphelenchus xylophlius Steiner & Buhrer, 1934 (Nickle, 1970) in Spain. Nematology 13: 755-757.
ROBINET, C., A. ROQUES, H. PAN, G. FANG, J. YE, Y. ZHANG y J. SUN (2009). Role of Human-Mediated Dispersal in the Spread of the Pinewood Nematode in China. PLoS ONE 4(2): e4646.
ROBINET, C., N. VAN OPSTAL, R. BAKER y A. ROQUES (2011). Applying a spread model to identify the entry points from which the pine wood nematode, the vector of pine wilt disease, would spread most rapidly across Europe. Biological invasions 13(12): 2981-2995.
RODRIGUES, J. M. (2008). National Eradication Programme for the Pinewood Nematode. Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems. M. Mota y P. Vieira, Springer Netherlands: 5-14.
RUTHERFORD, T. y J. WEBSTER (1987). Distribution of pine wilt disease with respect to temperature in North America, Japan, and Europe. Canadian Journal of Forest Research 17(9): 1050-1059.
RYSS, A., P. VIEIRA, M. MOTA y O. KULINICH (2005). A synopsis of the genus Bursaphelenchus Fuchs, 1937 (Aphelenchida: Parasitaphelenchidae) with keys to species. Nematology 7(3): 393-458.
SANCHEZ-HUSILLOS, E., G. ÁLVAREZ-BAZ, I. ETXEBESTE y J. A. PAJARES (2013). Shoot feeding, oviposition, and development of Monochamus galloprovincialis on Pinus pinea relative to other pine species. Entomologia experimentalis et applicata: 1-10.
SANCHEZ-HUSILLOS, E., I. ETXEBESTE y J. PAJARES (2015). Effectiveness of mass trapping in the reduction of Monochamus galloprovincialis Olivier (Col.: Cerambycidae) populations. Journal of Applied Entomology.
SHIBATA, E. (1986). Dispersal movement of the adult Japanese Pine Sawyer, Monochamus alternatus Hope (Coleoptera, Cerambycidae) in a young pine forest. Applied Entomology and Zoology 21(1): 184-186.
SHIMAZU, M. (2008). Biological control of the Japanese pine sawyer beetle, Monochamus alternatus. Pine Wilt Disease, Springer: 351-370.
SOLIMAN, T., M. C. MOURITS, W. VAN DER WERF, G. M. HENGEVELD, C. ROBINET y A. G. O. LANSINK (2012). Framework for modelling economic impacts of invasive species, applied to pine wood nematode in Europe.
SOUSA, E., M. A. BRAVO, J. PIRES, P. NAVES, A. C. PENAS, L. BONIFACIO y M. M. MOTA (2001). Bursaphelenchus xylophilus (Nematoda; Aphelenchoididae) associated with Monochamus galloprovincialis (Coleoptera; Cerambycidae) in Portugal. Nematology 3: 89-91.
TAKASU, F., N. YAMAMOTO, K. KAWASAKI, K. TOGASHI, Y. KISHI y N. SHIGESADA (2000). Modeling the expansion of an introduced tree disease. Biological Invasions 2(2): 141-150.
TEALE, S. A., J. D. WICKHAM, F. ZHANG, J. SU, Y. CHEN, W. XIAO, L. M. HANKS y J. G. MILLAR (2011). A male-produced aggregation pheromone of Monochamus alternatus (Coleoptera: Cerambycidae), a major vector of pine wood nematode. Journal of economic entomology 104(5): 1592-1598.
TOGASHI, K. (1985). Transmission curves of Bursaphelenchus xylophilus (Nematoda, Aphelenchoididae) from its vector, Monochamus alternatus (Coleoptera, Cerambycidae), to pine trees with reference to population performance. Applied Entomology and Zoology 20(3): 246-251.
TOGASHI, K. (1990). A field experiment on dispersal of newly emerged adults of Monochamus alternatus (Coleoptera, Cerambycidae). Researches on Population Ecology 32(1): 1-13.

20
TOMMINEN, J. (1993). Development of Monochamus galloprovincialis Olivier (Coleoptera, Cerambycidae) in Cut Trees of Young Pines (Pinus sylvestris L.) and Log Bolts in Southern Finland. Entomologica Fennica 4(3): 137-142.
TORRES-VILA, L. M., C. ZUGASTI, J. M. DE-JUAN, M. J. OLIVA, C. MONTERO, F. J. MENDIOLA, Y. CONEJO, Á. SÁNCHEZ, F. FERNÁNDEZ, F. PONCE y G. ESPÁRRAGO (2015). Mark-recapture of Monochamus galloprovincialis with semiochemical-baited traps: population density, attraction distance, flight behaviour and mass trapping efficiency. Forestry 88(2): 224-236.
TURCHIN, P. (1998). Quantitative analysis of movement: measuring and modeling population redistribution in animals and plants, Sinauer Associates Sunderland.
VICENTE, C., M. ESPADA, P. VIEIRA y M. MOTA (2012). Pine Wilt Disease: a threat to European forestry. European Journal of Plant Pathology 133(1): 89-99.
VIVES (2000). Insecta, Coleoptera, Cerambycidae. Serie Fauna Ibérica. Museo Nacional de Ciencias Naturales, CSIC, Madrid. 12.
WEBSTER, J. (1999). Pine wilt disease: a world wide survey. Sustainability of pine forests in relation to pine wilt and decline. Proceedings of International Symposium, Tokyo, Japan, 27-28 October, 1998., Shokado Shoten.
WINGFIELD, M. y R. BLANCHETTE (1983). The pine-wood nematode, Bursaphelenchus xylophilus, in Minnesota and Wisconsin: insect associates and transmission studies. Canadian Journal of Forest Research 13(6): 1068-1076.
WORK, T. T., D. G. MCCULLOUGH, J. F. CAVEY y R. KOMSA (2005). Arrival rate of nonindigenous insect species into the United States through foreign trade. Biological invasions 7(2): 323-332.
YOSHIMURA, A., K. KAWASAKI, F. TAKASU, K. TOGASHI, K. FUTAI y N. SHIGESADA (1999). Modeling the spread of pine wilt disease caused by nematodes with pine sawyers as vector. Ecology 80(5): 1691-1702.
ZAMORA, P., V. RODRÍGUEZ, F. RENEDO, A. V. SANZ, J. C. DOMÍNGUEZ, G. PÉREZ-ESCOLAR, J. MIRANDA, B. ÁLVAREZ, A. GONZÁLEZ-CASAS, E. MAYOR, M. DUEÑAS, A. MIRAVALLES, A. NAVAS, L. ROBERTSON y A. B. MARTÍN (2015). First Report of Bursaphelenchus xylophilus Causing Pine Wilt Disease on Pinus radiata in Spain. Plant Disease: PDIS-03-15-0252-PDN.
ZHAO, B. G., K. FUTAI, J. R. SUTHERLAND y Y. TAKEUCHI (2008). Pine wilt disease, Springer.

21
Capítulo 2
Fenología de la emergencia de Monochamus
galloprovincialis (Col.: Cerambycidae) en la península ibérica
Introducción La enfermedad del marchitamiento del pino (Pine Wilt Disease, PWD) es una de las más importantes amenazas para los bosques de coníferas del mundo (VICENTE et al. 2012, FUTAI 2013). El causante de esta enfermedad es el Nematodo de la Madera del Pino (NMP) Bursaphelenchus xylophilus (Steiner et Bührer), el cual es considerado Organismo de Cuarentena por la Unión Europea desde 1993 (Directiva 77/93, EEC). En 1999 fue comunicado por primera vez en Europa después de su identificación como responsable de la muerte de una gran cantidad de pinos de la especie Pinus pinaster Aiton cerca de Setúbal (40 km al sudeste de Lisboa) en Portugal (MOTA et al. 1999). Actualmente la enfermedad se ha expandido por casi todo el territorio portugués y la parte continental del país ha sido declarada Zona Demarcada (ABELLEIRA et al. 2011, VICENTE et al. 2012, ZAMORA et al. 2015). Desde la introducción del NMP en Portugal hasta hoy, España ha informado de cuatro detecciones de PWD cerca de la frontera con Portugal (MAGRAMA 2015, ZAMORA et al. 2015). Aunque estos brotes están siendo controlados o han sido ya erradicados, la mera aparición de 4 focos en pocos años significa que este país podría fácilmente ejercer el papel de puente en la dispersión de esta enfermedad desde Portugal hacia el resto de Europa (ROBINET et al. 2011, PUKKALA et al. 2014). Por lo tanto, la inspección y el control del NMP se han convertido en objetivo prioritario en las políticas forestales españolas y europeas actuales, y la acción principal para ello está siendo enfocada al control de sus vectores (Directiva UE 2014/5353/CE). La propagación de esta enfermedad está fundamentalmente vinculada a la dispersión de sus vectores (TOGASHI Y SHIGESADA 2006, ROBINET et al. 2009, ROBINET et al. 2011). Varias especies de escarabajos barrenadores de la madera, especialmente Cerambycidae, Buprestidae y Curculionidae, se han descrito en asociación con el NMP en todo el mundo, aunque no ha sido demostrada capacidad de transmisión efectiva a los pinos para la mayor parte de ellos (LINIT 1988, SOUSA et al. 2001, SOUSA et al. 2002, ROBERTSON et al. 2008, ROBERTSON et al. 2011). Sólo algunas especies de escarabajos longicornios del género Monochamus (Cerambycidae) han sido reconocidas, hasta la fecha, como principales vectores de NMP, y por tanto, capaces de transmitirlo a sus hospedadores (KOBAYASHI et al. 1984, LINIT 1988, SOUSA et al. 2001). Poco después de la primera detección del NMP en Portugal, se demostró que M. galloprovincialis podía comportarse de facto como un vector eficaz de la enfermedad en la península ibérica (SOUSA et al. 2001). Además, existen cuatro especies endémicas de Monochamus en el continente europeo que son generalmente consideradas como vectores potenciales de B. xylophilus: M. sutor (Linnaeus), M. sartor (Fabricius), M. urussovi

22
(Fischer) y M. saltuarius (Gebler) (EPPO 2009, EVANS et al. 2009, AKBULUT Y STAMPS 2012). Este hecho ha generado una gran preocupación en Europa, puesto que la introducción del NMP supone un enorme riesgo para muchos bosques europeos debido a diferentes factores: la amplia distribución de especies de árboles hospedantes susceptibles de contraer la enfermedad (TRÖLTZSCH et al. 2009), la existencia de diferentes especies de Monochamus (EVANS et al. 2009, AKBULUT Y STAMPS 2012), y la idoneidad de las condiciones climáticas para el desarrollo de la enfermedad, especialmente en los países mediterráneos (RUTHERFORD et al. 1992, PÉREZ et al. 2008). Hasta la fecha, el control del NMP ha estado ceñido a la detección temprana de los árboles infectados y su destrucción completa antes de la emergencia de los vectores, de forma que éstos no puedan generar nuevos focos de la enfermedad. El desarrollo de trampas y atrayentes que permiten el seguimiento de M. galloprovincialis (PAJARES et al. 2004, PAJARES et al. 2010, ÁLVAREZ et al. 2015) ha permitido el desarrollo de alternativas de gestión centradas en el control poblacional del vector (SANCHEZ-HUSILLOS et al. 2015, TORRES-VILA et al. 2015). Sin embargo estas estrategias no pueden, por sí mismas, sustituir completamente los métodos utilizados hasta hoy. Uno de los principales factores que afectan la erradicación y control de especies invasoras es la correcta sincronización entre las medidas adoptadas y la fenología de las especies a gestionar (BOGICH et al. 2008). Desde esta perspectiva, la utilización de modelos fenológicos en la gestión de las plagas ha demostrado ser una herramienta fundamental para la creación de programas de vigilancia y control de plagas forestales, muy útil para la calificación del riesgo de infestación (LOGAN 1988, BENTZ et al. 1991, WERMELINGER Y SEIFERT 1998, BENTZ 2006, BAIER et al. 2007, REGNIERE et al. 2012) y completamente necesaria para la organización temporal de las medidas de control dirigidas al control del vector (PARK et al. 2014). La temperatura ambiental es el factor abiótico principal que modula la biología, ecología y dinámica poblacional de gran parte de los insectos. El carácter ectotermo del desarrollo fisiológico permite la modelización de la tasa de desarrollo en función de la temperatura externa (KONTODIMAS et al. 2004). En este marco, diversos modelos basados en el uso de unidades de calor (grados-día, °D) han sido utilizados para el estudio de la fenología y la predicción del desarrollo de la población en diversas especies de insectos (por ejemplo QUESADA-MORAGA Y SANTIAGO-ALVAREZ 2000, MYERS et al. 2014). En concreto, la relación de la temperatura con los patrones emergencia de imagos de Monochamus ha sido ampliamente estudiada en M. alternatus en Corea, Japón y China (TOGASHI 1985, 1990, TOGASHI et al. 2008, PARK et al. 2014) y en M. Saltuarius en Japón y Corea (JIKUMARU et al. 1994, JIKUMARU Y TOGASHI 1996, JUNG et al. 2015). En el caso de M. galloprovincialis en Europa, han sido publicados algunos estudios locales en Italia, Francia y Portugal que han mostrado patrones de emergencia con perfiles unimodales, que se extienden desde finales de primavera a mediados de verano(FRANCARDI Y PENNACCHIO 1996, KOUTROUMPA et al. 2008, FRANCARDI et al. 2009, NAVES Y DE SOUSA 2009). No obstante no existen representaciones ni estudios generales de la emergencia para toda la península ibérica. En este contexto, NAVES y DE SOUSA (2009) propusieron un modelo de predicción de la emergencia de M. galloprovincialis en la península de Setúbal basado en la acumulación de grados-día a partir del 1 de marzo, estableciendo el umbral mínimo de desarrollo larvario en 12.2 ± 0.8ºC. La media de acumulación de calor para la emergencia del 50% de la población en condiciones de laboratorio fue de 822 °D. Este modelo funcionó con precisión en la predicción de las emergencias en años anteriores en la misma región de estudio (Setúbal, Portugal), pero se

23
desconoce si es igualmente preciso en otras regiones de la península Ibérica y Europa y puede, por tanto, ser considerada una herramienta de aplicación más general. Los objetivos iniciales de este trabajo, por tanto, son (i) ampliar el conocimiento sobre la fenología de la emergencia de M. galloprovincialis en la península Ibérica, (ii) evaluar la utilidad del mencionado modelo de desarrollo larvario portuqués en la predicción de la emergencia de M. galloprovincialis en varias localidades de la península Ibérica y (iii) situar en el contexto de la dispersión de la NMP las diferencias o similitudes entre los diferentes patrones de emergencia que pudieren observarse en dicho territorio. Alternativamente, se han evaluado otros modelos de desarrollo no-lineares con el objetivo de mejorar la precisión con la que predecir la emergencia de M. galloprovincialis.
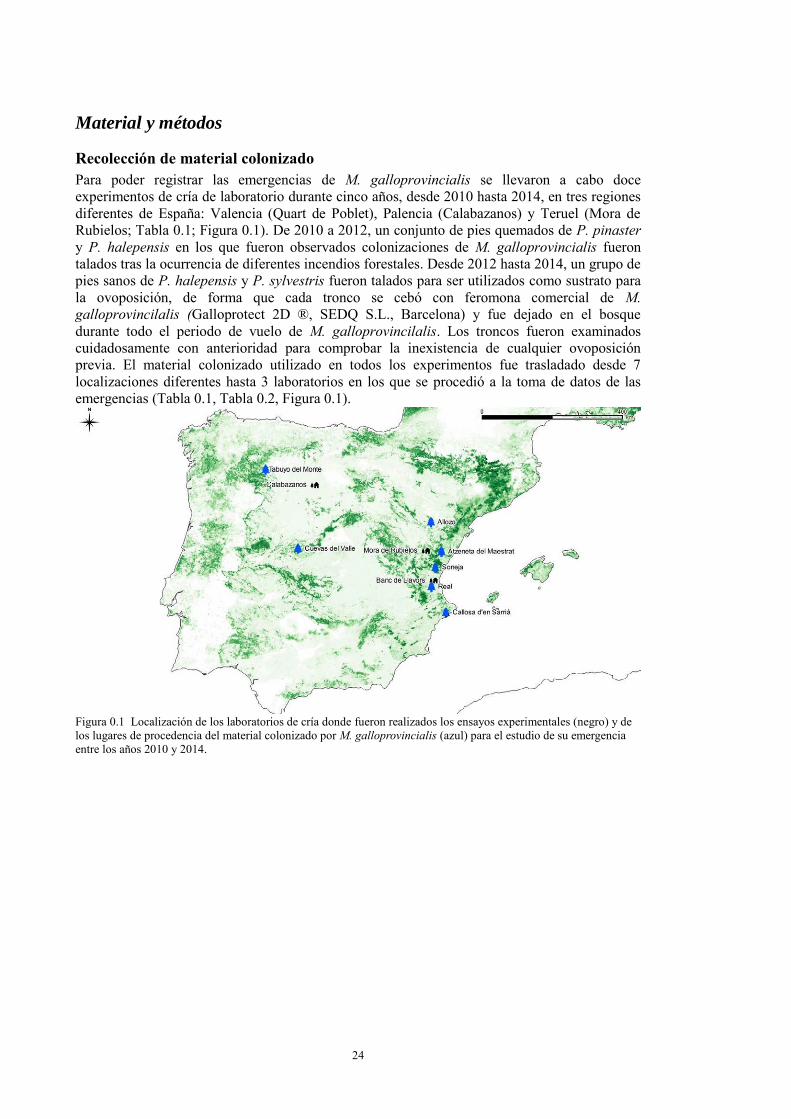
24
Material y métodos
Recolección de material colonizado Para poder registrar las emergencias de M. galloprovincialis se llevaron a cabo doce experimentos de cría de laboratorio durante cinco años, desde 2010 hasta 2014, en tres regiones diferentes de España: Valencia (Quart de Poblet), Palencia (Calabazanos) y Teruel (Mora de Rubielos; Tabla 0.1; Figura 0.1). De 2010 a 2012, un conjunto de pies quemados de P. pinaster y P. halepensis en los que fueron observados colonizaciones de M. galloprovincialis fueron talados tras la ocurrencia de diferentes incendios forestales. Desde 2012 hasta 2014, un grupo de pies sanos de P. halepensis y P. sylvestris fueron talados para ser utilizados como sustrato para la ovoposición, de forma que cada tronco se cebó con feromona comercial de M. galloprovincilalis (Galloprotect 2D ®, SEDQ S.L., Barcelona) y fue dejado en el bosque durante todo el periodo de vuelo de M. galloprovincilalis. Los troncos fueron examinados cuidadosamente con anterioridad para comprobar la inexistencia de cualquier ovoposición previa. El material colonizado utilizado en todos los experimentos fue trasladado desde 7 localizaciones diferentes hasta 3 laboratorios en los que se procedió a la toma de datos de las emergencias (Tabla 0.1, Tabla 0.2, Figura 0.1).
Figura 0.1 Localización de los laboratorios de cría donde fueron realizados los ensayos experimentales (negro) y de los lugares de procedencia del material colonizado por M. galloprovincialis (azul) para el estudio de su emergencia entre los años 2010 y 2014.

25
Tabla 0.1 Características del material colonizado por M. galloprovincialis y localización de los ensayos Exp Año Localización Procedencia Tipo Traslado Hopedante Clase Fecha 1 2010 Calabazanos Cuevas del Valle Jaula 2010-02-15 P. pinaster Incendio 2009-07-28 2 2010 Mora de Rubielos Alloza Jaula 2010-05-19 P. halepensis Incendio 2009-07-24 3 2010 Valencia Soneja Jaula 2010-05-14 P. halepensis Incendio 2009-06-20 4 2011 Calabazanos Cuevas del Valle Jaula 2011-04-12 P. pinaster Incendio 2009-07-28 5 2011 Mora de Rubielos Callosa d’En Sarrià Jaula 2011-05-25 P. halepensis Incendio 2010-05-20 6 2011 Valencia Callosa d’En Sarrià Jaula 2011-05-16 P. halepensis Incendio 2010-05-20 7 2012 Calabazanos Real de Montroi Jaula 2012-05-22 P. halepensis Incendio 2011-07-02 8 2012 Valencia Atzaneta del Maestrat Bidón 2012-02-02 P. halepensis Corta 2011-08-31 9 2012 Valencia Real de Montroi Jaula 2012-05-18 P. halepensis Incendio 2011-07-02 10 2013 Calabazanos Tabuyo del Monte Bidón 2012-09-30 P. sylvestris Corta 2012-06-27 11 2013 Calabazanos Tabuyo del Monte Jaula 2012-09-30 P. sylvestris Corta 2012-06-27 12 2014 Calabazanos Tabuyo del Monte Jaula 2013-10-17 P. sylvestris Corta 2013-06-19 Tabla 0.2 Características de las regiones de procedencia d material colonizado Procedencia Material Clima* Altitud Especia principal Masa Tm Tmin Tmax Cuevas del Valle Csbo 850 P. pinaster Natural 13 7 20 Alloza BSk 670 P. halepensis Repoblación 13 7 19 Soneja Csa 300 P. halepensis Natural 16 10 21 Callosa d’En Sarrià Csa 250 P. halepensis Natural 17 9,5 24 Real de Montroi Csao 200 P. halepensis Natural 17 11 23 Atzaneta del Maestrat Cfa 450 P. halepensis Repoblación 14 8 20 Tabuyo del Monte Csb 1050-1130 P. sylvestris Repoblación 10 4 16 * BSk, estepa fría; Cfa, templado sin estación seca con verano caluroso; CSa, templado con verano seco y caluroso; Csb, templado con verano seco y templado;
Cría de los insectos Se dispuso de dos conjuntos de árboles: por un lado árboles quemados en incendios forestales y colonizados por M. galloprovincialis de forma natural y, por otro, árboles talados y cebados con atrayente cairo-feromonal (Galloprotect 2D, SEDQ, Barcelona) para forzar la colonización. Ambos conjuntos fueron cortados en trozas de 0,5-1 m de longitud, transportadas a laboratorio y almacenadas en contenedores de plástico (90 cm de altura, 60 cm de diámetro) o en jaulas (3 m de altura, 4 de longitud y 4 de anchura) cubiertos con una tela de tul para su aireación y protección, hasta la emergencia de los adultos de M. galloprovincialis (Tabla 0.1). El almacenamiento se realizó en el exterior, pero a cubierto de la lluvia para evitar la proliferación de hongos. En el caso concreto del material hospedante del Experimento nº 9, llevado a cabo en Valencia en el año 2012 (Tabla 0.1), parte de las trozas fueron trasladadas a una cámara de frío (4ºC; aprox. 15 días) con el objetivo de ralentizar la emergencia, y evaluar el efecto del frío en el patrón general de las emergencias. Todos los contenedores y jaulas fueron revisados diariamente. Se recolectaron todos los escarabajos emergidos, los cuales fueron sexados, pesados y medidos (longitud del élitro derecho o longitud total).
Datos climáticos Las acumulaciones de grados días fueron calculadas usando la metodología descrita por ROLTSCH et al. (1999) y utilizada en los cálculos realizados por NAVES y DE SOUSA (2009). Para tal computación se construyó un algoritmo en el entorno de programación estadístico R(R CORE TEAM 2015), el método de onda sinoidal (BASKERVILLE Y EMIN 1969, ALLEN 1976, ROLTSCH et al. 1999). El intervalo de cálculo fue el comprendido desde

26
el primer día de marzo hasta la emergencia del 50% de los adultos, contemplando como umbral mínimo de desarrollo los 12.2 ± 0.8ºC, mientras que el máximo se estableció entre los 32 y 35ºC (NAVES Y DE SOUSA 2009). Este modelo requiere únicamente de las temperaturas máximas y mínimas diarias del aire. Para la limitación de las temperaturas por encima del umbral superior en el modelo, fue utilizado una corrección de corte intermedio que penaliza el desarrollo para los días en los que la temperatura máxima supera el umbral máximo(ROLTSCH et al. 1999). La acumulación de grados-día tomando como intervalo de cálculo el momento de la ovoposición fue también calculada, siguiendo el mismo procedimiento. Para ello, se emplearon las fechas de los incendios y las emergencias medias de los años anteriores como referencia. En estos cálculos fueron utilizadas las medidas de un registrador de datos de temperatura instalado en cada localización experimental (HOBO pendant UA-001-64, Onset, USA), contrastadas a su vez con los datos de estaciones meteorológicas oficiales cercanas (a una distancia menor de 5 km de cada localización experimental) pertenecientes a la Agencia Estatal de Meteorología (AEMET) y a la Fundación CEAM (CEAMET) de la Generalitat Valenciana. Si las trozas fueron trasladadas entre dos localizaciones (del monte al laboratorio), los datos climáticos de ambos lugares fueron consecuentemente concatenados en los análisis. En dos de los experimentos, los desarrollados en Quart de Poblet (Valencia) con material colonizado de Atzeneta del Maestrat en 2012, y en Calabazanos (Palencia) con material colonizado de Tabuyo del Monte en 2013 (en los cuales las trozas fueron almacenadas en contenedores cerrados de 200 l de capacidad) se midió la temperatura del interior del contenedor y la temperatura del interior de la madera con sondas adecuadas (HOBO U23-004,Onset, USA).
Análisis estadístico Una primera aproximación consistió en la graficación de las curvas de densidad de emergencias por localidad y año, de tal forma que tales curvas estuviesen estandarizadas a modo de densidades estadísticas, y por tanto comparables entre sí. Para poder analizar la tendencia en la razón de sexo a lo largo del periodo de la emergencia, se calcularon las razones de sexo en los percentiles 20, 40, 60, 80 y 100 de las emergencias, dividiendo el número emergido de M. galloprovincialis machos por las hembras para cada uno de los cinco periodos delimitados por los percentiles. Para ilustrar las tendencias en la razón de sexo, ésta se modelizó frente a la fecha en la que se registrarón resultando en un modelo linear donde la pendiente señala la tendencia. Tras el computo inicial de los grados día acumulados para las series de emergencias siguiendo el método descrito anteriormente, se observaron desviaciones significativas de los percentiles descritos para los experimentos en NAVES y DE SOUSA (2009; ver resultados). Aunque los modelos de desarrollo lineares han sido empleados en gran número de ocasiones, los umbrales térmicos estimados por este método han sido frecuentemente mejorados en precisión por modelos no lineares (KONTODIMAS et al. 2004). Teniendo en cuenta que los umbrales de desarrollo utilizados para el computo de los grados-día estaba basado en un modelo linear de desarrollo, y tomando, por tanto, como referencia los datos publicados por (NAVES Y DE SOUSA 2009), se procedió a reajustar el modelo de desarrollo con el objetivo de comprobar si era posible conseguir un ajuste mejor para los datos registrados en este trabajo. En un primer paso, se procedió a reajustar el modelo de acuerdo con la aproximación lineal utilizada en los análisis de NAVES y DE SOUSA (2009; Figura 0.2), para luego hacer lo mismo utilizando otras aproximaciones no lineales, citadas por PARK et al. (2014) y JUNG et al. (2015) y aparentemente válidas para otras especies de Monochamus. Existen numerosas aproximaciones no lineares desarrolladas por diferentes autores; dos de los modelos más éxitosos son los denominados Briere y Lactin, y dos de sus variantes (KONTODIMAS et al. 2004 y las referencias citadas en él; Tabla 0.3). La parametrización de estas funciones fue realizada por

27
mínimos cuadrados. Los puntos de inicio de las iteraciones para ajustar los parámetros de cada modelo fueron elegidos con base en los parámetros obtenidos por (JUNG et al. 2015). Finalmente, también se re-analizaron los mismos datos siguiendo un modelo mecánico conocido como Sharpe-Schoolfield-Ikemoto (SSI). Para ajuste de éste último modelo, la optimización de los parámetros del modelo se realizó siguiendo el algoritmo de Nelder-Mead (IKEMOTO et al. 2013). Este modelo se define como:
Los diferentes parámetros se hallan descritos en la tabla 2.4 en base a lo descrito por IKEMOTO et al. (2013). Una vez obtenidos los nuevos umbrales térmicos de desarrollo, éstos fueron utilizados para recalcular la acumulación de grados día de cada emergente bajo los umbrales marcados por cada modelo. Finalmente se calcularon los grados-día para los percentiles 5% , 25%, 50%, 75%, 95% y 99%, para así poder comparar los ajustes derivados de los modelos no-linears con los demás resultados. Tabla 0.3 Modelos no lineales de tasas de desarrollo larvario, obtenidos de la revisión metodológica de Kontodimas et al (2004), utilizados para la reinterpretación de los datos de Naves & Sousa (2009). En los dos modelos de Lactin (Lactin 1 y Lactin 2), T es la temperatura de cría (ºC), ρ es una constante que define la tasa de la temperatura óptima, Tmax es el umbral superior de temperatura, y ΔT es el rango de temperatura sobre los cuales la ralentización del desarrollo fisiológico se convierte en predominante. En ellos, λ permite la estimación de un umbral inferior de temperatura. En ambos modelos Briére (1 y 2), T es la temperatura de cría (ºC) y Tmin es la temperatura del umbral inferior de desarrollo, y a y m son constantes empíricas. Ecuación no-linear Modelo Referencia
Lactin 1 Lactin et al. (1995)
Lactin 2 Lactin et al. (1995) Briere 1 Briere et al. (1999) Briere 2 Briere et al. (1999)
Tabla 0.4 Parámetros del model SSI Símbolo Descripción
Tasa media de desarrollo (1/día) Temperatura absoluta (K) (273.15 K = 0 C) Gas constante (1.987 cal/deg/mol)
Entalpía de activación de la reacción catalizada po la enzima (cal/mol) Cambio en la entalpía asociada con la inactivación de la enzima por bajas temperaturas (cal/mol) Cambio en la entalpía asociada con la inactivación de la enzima por altas temperaturas (cal/mol)
Temperatura a la cual la enzima es la mitad active y la mitad inactive por bajas temperaturas (K) Temperatura a la cual la enzima es la mitad active y la mitad inactive por altas temperaturas (K) Temperatura optima intrínseca a la cual la probabilidad de que la enzima esté en estado activo es máxima
(K) Tasa de desarrollo a la temperature optima intrínseca (1/day) asumiendo que no hay inactivación
enzimática

28
Figura 0.2 Relación lineal entre la tasa de desarrollo larvario de M. galloprovincialis y la temperatura obtenida para los datos publicados por Naves & Sousa (2009) en su Tabla 1. .

29
Resultados
Periodo de emergencia Las distribuciones de la densidad de las emergencias de los adultos de M. galloprovincialis se presentan en Figura 0.3. La actividad de la emergencia de los adultos se produjo durante aproximadamente 2 meses casi todos los años, y de una forma unimodal (aunque cierta bimodalidad es observable en algunos casos). No obstante, el periodo de emergencia mostró una gran variación entre los diferentes años y lugares.
Figura 0.3 Curvas de emergencia de M. galloprovincialis en los diferentes ensayos analizados. En el eje de las abcisas representa la densidad estadística de insectos emergidos por día de tal forma que el área bajo la curva representa el 100% de las emergencias. El eje de las ordenadas muestra los días ordinales. Las abreviaturas utilizadas conrresponden las diferentes localizaciones y años (Cal, Calabazabos; Val, Valencia; Mor, Mora de Rubielos). La diferencia máxima entre las fechas a las que se produjo el 50% de las emergencias acumuladas fue de 35 días, y se dio entre la fecha más temprana, el 11 de junio en 2010 en Valencia, y la fecha más tardía, el 16 de julio de 2010 en Calabazanos (Palencia). La curva de emergencia más temprana se observó en Quart de Poblet (Valencia) en 2010. Tanto su inicio, como su mediana, como el final de su periodo de emergencia fueron anteriores a todos los demás períodos estudiados. Por otra parte, la curva de emergencia más tardía se observó en Calabazanos (Palencia) en 2013. En términos generales, las curvas de emergencia fueron anteriores en Valencia que en Palencia (Tabla 0.5)

30
Tabla 0.5 Fechas de emergencia de adultos de M. galloprovincialis separada por diferentes percentiles de emergencia acumulada. N= número de adultos emergidos totales. SEM= error estándar de la media. Experimento Fecha de emergencia 5% 25% 50% 75% 95% 99% Media N SEM Exp 1 Calabazanos.2010.Jaula 29-Jun 09-Jul 16-Jul 24-Jul 06-Ago 18-Ago 16-Jul 993 0,39 Exp 4 Calabazanos.2011.Jaula 10-Jun 18-Jun 28-Jun 21-Jul 04-Ago 17-Ago 03-Jul 652 0,74 Exp 7 Calabazanos.2012.Jaula 29-May 05-Jun 22-Jun 30-Jun 12-Jul 17-Jul 19-Jun 400 0,76 Exp 10 Calabazanos.2013.Bidón.ext 02-Jul 15-Jul 21-Jul 28-Jul 09-Ago 18-Ago 20-Jul 335 0,6 Exp 10 Calabazanos.2013.Bidón.int 02-Jul 15-Jul 21-Jul 28-Jul 09-Ago 18-Ago 20-Jul 335 0,6 Exp 11 Calabazanos.2013.Jaula 01-Jul 15-Jul 22-Jul 27-Jul 05-Ago 16-Ago 20-Jul 1044 0,32 Exp 12 Calabazanos.2014.Jaula 02-Jun 23-Jun 04-Jul 16-Jul 25-Jul 14-Ago 02-Jul 938 0,56 Exp 2 Mora.2010.Jaula 12-Jun 24-Jun 02-Jul 10-Jul 23-Jul 26-Jul 01-Jul 186 0,89 Exp 5 Mora.2011.Jaula 01-Jun 16-Jun 30-Jun 06-Jul 24-Jul 04-Ago 27-Jun 79 1,77 Exp 3 Valencia.2010.Jaula 24-May 02-Jun 11-Jun 19-Jun 04-Jul 08-Jul 11-Jun 556 0,53 Exp 6 Valencia.2011.Jaula 01-Jun 10-Jun 21-Jun 30-Jun 16-Jul 25-Jul 21-Jun 502 0,64 Exp 8 Valencia.2012.Bidón 10-Jun 20-Jun 25-Jun 04-Jul 14-Jul 14-Jul 26-Jun 73 1,11 Exp 9 Valencia.2012.Jaula 26-May 08-Jun 26-Jun 05-Jul 17-Jul 27-Jul 23-Jun 1635 0,42
Proporción de sexos en la emergencia La proporción de sexos en las emergencias totales muestra patrones equilibrados entre ambos sexos, con valores en la razón de sexo oscilando entre 0.83 y 1.19 machos por hembra (Tabla 0.6). Tabla 0.6 Frecuencias de emergentes de M. galloprovincialis por sexo, año y localidad experimental. N= número de adultos emergidos totales.
Experimento Machos Hembras Razón de sexo N
Calabazanos 2010 519 474 1.09 993
Mora de Rubielos 2010 99 87 1.14 186
Valencia 2010 268 288 0.93 556
Calabazanos 2011 313 316 0.99 629
Mora de Rubielos 2011 43 36 1.19 79
Valencia 2011 245 257 0.95 502
Calabazanos 2013 746 633 1.18 1379
Calabazanos 2014 502 453 1.11 955
Valencia 2012 775 935 0.83 1710 La evolución temporal de la proporción de sexos a lo largo el periodo de emergencia puede observarse en la Figura 0.4. Tal y como se puede observar, en la mayor parte de los registros la razón de sexo evolucionó negativamente, y en los casos en los que tal tendencia no se registró, la razón se mantuvo en beneficio de los machos (Calabazanos y Mora de Rubielos 2010). La evolución más dramática se observó los años 2010 y 2011 en Valencia, donde al inicio del periodo de emergencia se observaron cerca de tres machos cada dos hembras emergidas, mientras que al final del periodo, emergieron dos hembras por cada macho.

31
Figura 0.4 Evolución temporal de la razón de sexos (machos a hembras) a lo largo el periodo de emergencia correspondiente. La razón de sexos ha sido calculada para las emergencias acumuladas hasta la fecha en la que se alcanzaba cada percentil en intervalos del 20%. La masa de los insectos varió de forma significativa entre los años analizados al respecto (F4, 3115 883.7 P<0.001), y entre los sexos (F4, 3115 883.7 P= 0.034; Figura 0.5), si bien la diferencia en masa entre sexos no se manifestó significativamente entre los individuos del mismo año. En el caso de la longitud elitral, tal y como se puede observar en el diagrama de cajas de la Figura 0.6, se mantuvo la misma tendencia. En este caso, la diferencia en la forma de medir la longitud (total o elitral), no permite comparar los experimentos entre sí. A modo de referencia, las hembras fueron 0.5-1 mm más largas, y 0.25 mg más pesadas.

32
Figura 0.5 Diagramas de cajas de los pesos de M. galloprovincialis al emerger represeantados por sexos y experimentos.

33
Figura 0.6 Diagramas de cajas de las longitudes elitrales de M. galloprovincialis al emerger represeantados por sexos y experimentos.
Acumulación de grados día. Las acumulaciones de calor (grados-día) para las diferentes tasas de emergencia de adultos de M. galloprovincialis basadas en la propuesta de NAVES y DE SOUSA (2009), en la que se tiene en cuenta la acumulación de grados-día desde el 1 de marzo con un umbral mínimo de desarrollo 12,2 ± 0,8 ° C, se presenta en la Tabla 0.7. En ella puede ser observada una gran variabilidad entre las diferentes áreas de estudio en los resultados. Las máximas acumulaciones de calor hasta la aparición de 50% de los adultos en los experimentos llevados a cabo en jaulas fueron observadas en Valencia (2012) y en Calabazanos (2011), con 699,15 °D y 643,54 °D, respectivamente, todas ellas menores que los 822 °D mostrados por NAVES y DE SOUSA (2009). Por otra parte, la mínima acumulación de calor se produjo en Mora de Rubielos (2010 y 2011) y Valencia (2011), con 220,87; 309,54 y 304,08 °D respectivamente.

34
Tabla 0.7 Acumulación de calor (en grados-día) a partir del 1 de marzo del año de emergencia para todos los ensayos separada por percentiles. N= número de adultos emergidos totales. SEM= error estándar de la media. Experimento Fecha de emergencia 5% 25% 50% 75% 95% 99% Media N SEM Exp 1 Calabazanos.2010.Jaula 322,87 432,92 507,06 582,04 701,73 804,46 509,05 993 3,79 Exp 4 Calabazanos.2011.Jaula 428,81 512,52 643,54 899,31 1067,21 1245,03 703 652 8,63 Exp 7 Calabazanos.2012.Jaula 390,85 448,59 557,97 642,16 721,93 756,87 556,23 400 5,71 Exp 10 Calabazanos.2013.Barril.ext 648,29 824,08 909,87 1008,7 1170,21 1288,85 905,9 335 8,14 Exp 10 Calabazanos.2013.Barril.int 300,32 458,46 534,75 623,96 768,94 866,41 531,21 335 7,2 Exp 11 Calabazanos.2013.Jaula 311,56 476,41 568,82 635,37 743,42 876,14 555,04 1044 3,93 Exp 12 Calabazanos.2014.Jaula 204,12 315,65 394,45 463,07 547,03 704,75 386,85 938 3,6 Exp 2 Mora.2010.Jaula 154,48 201,28 220,87 278,18 412,32 448,78 244,31 186 5,4 Exp 5 Mora.2011.Jaula 163,76 203,95 309,54 365,25 524,14 609,76 311,95 79 12,81 Exp 3 Valencia.2010.Jaula 187,77 250,7 341,81 409,13 545,63 598,27 341,37 556 4,67 Exp 6 Valencia.2011.Jaula 170,05 216,19 304,08 406,05 608,07 740,41 330,17 502 6,4 Exp 8 Valencia.2012.Barril 647,12 782,12 859,01 987,88 1142,2 1146,29 885,23 73 16,34 Exp 9 Valencia.2012.Jaula 364,63 489,88 699,15 817,79 970,67 1108,37 686,2 1635 4,95
La acumulación de calor utilizando los umbrales propuestos por NAVES y DE SOUSA (2009) pero teniendo como fecha de inicio de la contabilización la fecha de la ovoposición, mostraron resultados no coherentes ya que la fecha exacta de la puesta de huevos era desconocida, por lo que no fue tenida en cuenta en los análisis.
Re-evaluación de la metodología de cálculo de los umbrales de desarrollo utilizada por NAVES y DE SOUSA (2009) Como consecuencia, por tanto, de la acusada variabilidad entre las acumulaciones de calor de los diferentes ensayos (Tabla 0.7), y también con respecto a los resultados de NAVES y DE SOUSA (2009), se procedió a reajustar el modelo desde diferentes enfoques con el objeto de comprobar si era posible conseguir un ajuste mejor para todos los datos: la aproximación lineal ya utilizada por NAVES y DE SOUSA (2009; Figura 0.2), así como otros ajustes no lineales desarrollados por diferentes autores (KONTODIMAS et al. 2004) y también citados para la modelización de las tasas de desarrollo larvario de M. alternatus por (PARK et al. 2014) y (JUNG et al. 2015). Las temperaturas de los umbrales de desarrollo inferiores (Tmin) y superiores (Tmax) de los modelos analizados pueden ser observadas en la siguiente Figura 0.7 y en la Tabla 0.8. El modelo Lactin 2 no se presenta ya que el parámetro lambda la hace incomparable con el resto de los modelos ajustados.
Figura 0.7 Temperaturas máximas y mínimas de desarrollo según modelos lineales y no lineales de la tasa de desarrollo larvario para los datos aportados por Naves & Sousa (2009).

36
Tabla 0.8 Temperaturas umbrales de desarrollo para los diferentes modelos analizados sobre los datos proporcionados por Naves & Sousa (2009). DTmin es la temperatura mínima de desarrollo (umbral inferior) y DTmax es la temperatura máxima de desarrollo (umbral superior)
Model DTmin DTmax Naves et al. (2009) 12.2 ºC Entre 32-35 ºC Linear 11.8 ºC No disponible Lactin 1 12.23 ºC 33.96 ºC Briere 1 12.02 ºC 35.77 ºC Briere 2 10.15 ºC 32.55 ºC
Para el caso del modelo Sharpe-Schoolfield-Ikemoto (SSI), el ajuste según dicho modelo puede ser observado en la Figura 0.8 y en la Tabla 0.9.
Figura 0.8 Ajuste según el modelo Sharpe-Schoolfield-Ikemoto (SSI) para los datos proporcionados por Naves & Sousa (2009). Tabla 0.9 Temperaturas de desarrollo para el modelos SSI sobre los datos proporcionados por NAVES y DE SOUSA (2009).TL es la temperatura a la cual la enzima es la mitad activa y la mitad inactiva por bajas temperaturas, TH es la temperatura a la cual la enzima es la mitad activa y la mitad inactiva por altas temperaturas y TΦ es la temperatura optima intrínseca a la cual la probabilidad de que la enzima esté en estado activo es máxima. Los temperaturas umbrales tenidas en cuenta son las medias de TL y TH.
Parametro Estimate Inferior Superior Media Mediana 1 T-Phi 21.85 15.65 26.24 22.24 22.08 6 TL 12.52 -38.12 18.24 11.28 13.15 7 TH 31.87 28.33 34.08 32.14 31.87
Una vez establecidos los diferentes umbrales térmicos, se procedió a simular los percentiles de emergentes en base a los datos publicados por NAVES y DE SOUSA (2009) en la Tabla 1 de su trabajo. Para ello, la fecha de emergencia de cada individuo se extrajo de una distribución normal con la media y desviación descritas en la mencionada tabla. Posteriormente, se calculó la acumulación de calor de cada individuo simulado utilizando los umbrales de la Tabla 0.8. Los percentiles de la acumulación de calor fueron entonces comparados con los registros obtenidos en los diferentes experimentos (Tabla 0.10), donde puede observarse que la acumulación de calor para que se dé la emergencia del 50% de la población, contabilizada según los diferentes modelos, no muestra una variación de gran magnitud de los 822 °D indicados por NAVES y DE SOUSA (2009).

37
Tabla 0.10 Acumulación de grados-día según los diferentes modelos analizados. NAVES y DE SOUSA (2009) mostraron un umbral superior de desarrollo a una temperatura de 32-35ºC. Por tanto, para el cálculo exacto respecto de los modelos lineales, han sido tomadas las temperaturas de 33,5ºC y 35ºC. Ambas temperaturas también han sido utilizadas para el cálculo del calor acumulado según modelo lineal teniendo en cuenta la temperatura mínima de 11,8ºC calculada en este capítulo sobre los datos de Naves & Souza (2009). Lineal (35;12.2) Lineal (33.5;12.2) Lineal (35;11.8) Lineal (33.5;11.8) Lactin 1 Briere 1 Briere 2 SSI 1% 422.1 422.1 433.50 433.50 421.24 427.23 480.56 412.97 10% 640.8 640.8 664.32 664.32 639.72 656.50 740.01 605.12 50% 891.0 891.0 924.00 924.00 889.65 900.36 1066.55 864.90 90% 1188.0 1188.0 1212.32 1212.32 1186.20 1198.80 1369.65 1153.20 99% 1593.5 1593.5 1625.70 1625.70 1591.09 1607.99 1758.49 1546.83

38
Discusión
En el caso de M. galloprovincialis, hasta ahora, los patrones de emergencia sólo habían sido analizados localmente en Francia, Italia y en Portugal (FRANCARDI Y PENNACCHIO 1996, KOUTROUMPA et al. 2008, NAVES Y DE SOUSA 2009). Este trabajo permite la comparación de los resultados de estos estudios previos, con los que aquí se presentan, realizados en 12 localidades diferentes de la península ibérica. En resumen, nuestros resultados evidencian variaciones significativas, tanto temporales como geográficas, entre las diferentes doce curvas de emergencia analizadas (Figura 0.3). De forma general, los insectos criados en material de reproducción procedente de zonas cálidas muestran curvas de emergencia adelantadas con respecto a las curvas de emergencia de aquellos que vienen de los lugares más fríos y acumulaciones de calor menores. Estos patrones son también diferentes de los registrados y previstos en la Península de Setúbal en Portugal (NAVES Y DE SOUSA 2009). Por lo tanto, esto sugiere la necesidad de diferenciar modelos de gestión locales del vector del NMP dependiendo de la ubicación geográfica y teniendo en cuenta las variables climáticas de cada región.
Patrón de emergencia de adultos Las doce curvas de emergencia analizadas en este estudio muestran que existen variaciones geográficas significativas entre los patrones de emergencia de M. galloprovincialis en la península ibérica. Aparentemente esta variación parece estar influida por las condiciones climáticas de la región en la que se desarrolla la larva, de forma que los insectos desarrollados y emergidos de trozas provenientes de zonas cálidas muestran curvas de emergencia adelantadas con respecto a los emergidos de trozas provenientes de zonas frías. En este mismo sentido también se observan variaciones interanuales en las curvas de emergencia. Éstas también parecen estar influidas por la temperatura, en concreto por las condiciones climáticas del otoño-invierno anterior a la emergencia, por la traslación del material colonizado desde zonas cálidas a zonas más frías, o por cualquier tratamiento de frío dado al material antes de la emergencia. En todos estos casos las condiciones frías producen un retraso relativo en la emergencia con respecto a las condiciones cálidas. De acuerdo con KOUTROUMPA et al. (2008), la duración del desarrollo larvario de M. galloprovincialis varía ampliamente entre larvas, incluso del mismo estadio, y la temperatura ciertamente juega un rol importante en ello. Y, de hecho, un porcentaje de estas larvas puede llegar emerger el año siguiente (HELLRIGL 1971, TOMMINEN 1993, NAVES et al. 2007, KOUTROUMPA et al. 2008). Esto coincide con los estudios sobre M. alternatus que muestran un ciclo complejo en el que las larvas que evolucionan hasta el estado adulto en el mismo año o en el siguiente en función de los estadios larvarios entrantes en el periodo invernal (TOGASHI 1989, 1990). De acuerdo con esto, el patrón de emergencia que muestran los resultados, tan extendido a lo largo de varios meses, podría sugerir que el ciclo de vida de M. galloprovincialis es, en efecto, tan complicado como el mostrado por PARK et al. (2014) para M. alternatus. Este autor señala dos condiciones que pueden modificar el patrón de emergencia: (i) la estructura de edad de las larvas entrantes en el invierno, que puede influir en el desplazamiento horizontal de la curva de

39
emergencia, y (ii) la heterogeneidad medioambiental posterior al paso del invierno, que puede causar variación del desarrollo larvario. Los resultados de este trabajo indican, en términos generales, que las curvas de emergencia en zonas cálidas se producen antes que en zonas frías, por lo que la temperatura parece jugar un papel fundamental estas diferemcias. En efecto, PARK et al. (2014) consideran que, aparentemente, un mayor número de larvas de M. alternatus en tercer estadio (en vez de en cuarto) en el periodo invernal supone un retraso en la curva de emergencia por el estadio adicional que deben alcanzar las larvas antes de la emergencia. Además, el hecho de que todos los estadios larvarios de M. galloprovincialis pueden ser encontrados antes y después del invierno (KOUTROUMPA et al. 2008) posibilita que las condiciones apuntadas por PARK et al. (2014) puedan ser la explicación de las variaciones en las emergencias observadas en nuestros resultados. En efecto, siguiendo un gradiente térmico, la proporción de larvas en uno u otro estadio debería variar de forma que, en las zonas más cálidas, la mayoría de las larvas antes del invierno estuviesen en L4, mientras que en las más frías, como en el caso de KOUTROUMPA et al. (2008), se presentasen en más diversidad de estadios. Respecto a los ensayos realizados en Calabazanos (Palencia) entre 2010 y 2014 conviene discutir lo siguiente: En 2010 las trozas fueron traslocadas al laboratorio para el estudio de las emergencias antes de terminado el periodo invernal (15/02/2010), mientras que en el año 2011 se produjo un retraso en la traslocación (realizada el 12/04/2011). En ambos casos el material hospedante fue P. pinaster proveniente de un incendio. Los resultados de 2011 muestran un adelanto en la curva de emergencia respecto a la de 2010, y con un perfil bimodal. La traslocación tardía que sufrieron las trozas desde una zona cálida (Cuevas del Valle) a una zona más fría (Calabazanos) puede explicar este adelanto de la curva de vuelo así como la distribución bimodal de las emergencias. Las altas temperaturas posteriores al paso del invierno y anteriores a la traslocación son las responsables de que se produzca, en este caso, una aceleración en el desarrollo larvario posterior al invierno. En los años 2013 y 2014, el material infestado fue P. sylvestris proveniente de corta. En este caso también se observa un adelanto de la curva de emergencia en el experimento realizado en Calabazanos el año 2014 respecto a la observada en 2013. El año 2014 se caracterizó en esa zona por haber tenido un otoño-invierno muy cálido con respecto al año anterior. Las temperaturas anormalmente altas del periodo previo al invierno pudieron suponer una aceleración del desarrollo larvario y, por tanto, permitir que un mayor número de larvas alcanzasen estadios larvarios superiores (L4 principalmente) antes del invierno, originándose un adelanto de la curva, en coherencia nuevamente con lo apuntado por (PARK et al. 2014). En el experimento realizado en 2012, el material hospedadorfue P. halepensis proveniente de incendio, originario de Valencia, una región completamente diferente, con un clima más cálido que el de Calabazanos. La curva de emergencia, por tanto, debe ser preferiblemente comparada con los ensayos realizados con el mismo material hospedante de la región de Valencia. De dichos ensayos, realizados en Quart de Poblet (Valencia), entre 2010 y 2012, conviene destacar: El material hospedador utilizado en todos los ensayos fue P. halepensis proveniente de incendio. Los años 2011 y 2012 parte de este material fue traslocado de zonas cálidas a zonas frías (en concreto a los laboratorios de Mora de Rubielos en 2011 y a Calabazanos en 2012) en la primavera tardía, pasado el periodo invernal y casi iniciado el periodo de vuelo. Por otro lado, en 2012, al material colonizado de Quart de Poblet se le aplicó un tratamiento de frío a un

40
conjunto aleatorio de trozas con el objetivo de retrasar la emergencia de individuos empleados en otros experimentos. En los casos concretos de la traslocación de trozas desde la zona cálida de Callosa d’En Sarrià (Valencia) a la fría de Mora de Rubielos (Teruel) en 2011, y desde la cálida de Real de Montroi (Valencia) a la fría de Calabazanos (Palencia) en 2012, se pueden observar nuevamente curvas de emergencia bimodales con un segundo pico retrasado respecto a las curvas de emergencia de trozas no traslocadas (que componen los ensayos realizados en Valencia los años 2010 y 2011), lo que en apariencia podría significar que que se hubiese producido una ralentización en el desarrollo larvario posterior al periodo invernal como consecuencia del frío ambiental de las nuevas ubicaciones tras la traslocación. De acuerdo con KOUTROUMPA et al. (2008), en términos de desarrollo larvario, las larvas provenientes de huevos tardíos alcanzan a aquellas que provienen de huevos tempranos, incluso considerando sólo el primer año de desarrollo, de forma que la emergencia se dé en un periodo breve de tiempo. Este grado de sincronización de las emergencias ha sido apuntada ya por otros autores como una estrategia de otros insectos perforadores para vencer la resistencia de los hospedantes y, de esta manera, asegurar el éxito en la colonización de nuevos árboles (RAFFA Y BERRYMAN 1983, BENTZ 2006). Si bien en el caso de Monochamus spp. la necesidad de superar las defensas de los árboles no existe, la emergencia concentrada en un breve periodo de tiempo puede conferirles otro tipo de ventajas evolutivas como pueden ser la accesibilidad temporal al material de cría y las condiciones climáticas favorables. La existencia de una dormancia/diapausa en el 4º estadio larvario sugerida por NAVES et al. (2007) y KOUTROUMPA et al. (2008) y que puede suceder más hallá de la hibernación parece completar además, el ciclo de aquellas larvas que no pudieran llegar al cuarto estadío antes del periodo invernal. Por tanto, las curvas de emergencia de M. galloprovincialis en la península Ibérica deberían ser, en términos generales, unimodales, tal y como ha sido constatado en los estudios previos a éste (FRANCARDI Y PENNACCHIO 1996, KOUTROUMPA et al. 2008, NAVES Y DE SOUSA 2009). La observación de curvas de emergencia bimodales en nuestros resultados supone la existencia de un segundo periodo de ralentización o parada del desarrollo larvario provocado por la aplicación artificial de temperaturas frías después de la diapausa/dormancia invernal. Este mismo efecto puede observarse en el ensayo realizado en Valencia el año 2012. Ese año se realizó una aplicación de frío a una parte aleatoria de las trozas colonizadas con posterioridad al inicio del periodo de emergencia. En este caso también se observó una curva bimodal con un segundo pico desplazado hacia la derecha. Las bajas temperaturas posteriores al paso del invierno y anteriores a la finalización del periodo de emergencia nuevamente produjeron, en este caso, una deceleración o parada en el desarrollo larvario posterior al invierno y, por tanto, un retraso en la emergencia de una parte de los adultos. Por último, en el ensayo realizado en Mora de Rubielos en 2010, con material hospedante de P. halepensis proveniente de incendio mostró una curva de emergencia más retrasada que las curvas observadas en Valencia, de acuerdo con un régimen climático más frío, característico de la zona.
Proporción de sexos en la emergencia En referencia a las proporciones de sexos observadas en las emergencias, nuestros resultados muestran que éstas son favorables a los machos al inicio del periodo de emergencia, y que esta

41
proporción va invirtiéndose a medida que avanza el dicho periodo. Y aunque generalmente no es observable una desviación muy evidente de la proporción 1:1, puede concluirse que la emergencia de los machos es ligeramente anterior a la de las hembras (Figura 0.4) y que, por tanto, M. galloprovincialis muestra comportamientos de protandria en sus emergencias, es decir, aparición de los machos anterior a la de las hembras. Entre las series analizadas, aquellas que no han mostrado este patrón, han mantenido una mayor proporción de machos emergentes. Son numerosos los ejemplos de protandria en el reino animal, aunque estos patrones conductuales y sus ventajas no son completamente comprendidas actualmente, y hasta siete diferentes hipótesis han sido formuladas para explicar la protandria desde los años 70 (MORBEY Y YDENBERG 2001). Una de ellas, la protandría como estrategia reproductiva, según la cual la aparición de los machos es anterior a la de las hembras para así maximizar el éxito reproductivo (es decir, aumentar el número de hembras fecundadas), es la más postulada en el caso de los artrópodos, ya que enfatiza los beneficios del apareamiento múltiple de los machos (BLANCKENHORN et al. 2007), aunque en puridad también deberían ser tenidos en cuenta otros muchos factores que podrían afectar al apareamiento (patrón de mortalidad, longevidad, grados de poliandria o poliginia, territorialidad, variación temporal en la fecundidad de las hembras, etc.) y que son realtivamente desconocidos en el caso de M. galloprovincialis. Por otro lado, la protandria observada podría también deberse a diferencias en la velocidad de desarrollo larvario entre sexos (BLANCKENHORN et al. 2007), pero, por lo que conocemos, los trabajos que han estudiado el desarrollo larvario en M. galloprovincialis nada han investigado sobre esta cuestión (KOUTROUMPA et al. 2008, NAVES Y DE SOUSA 2009). La diferencie entre el tamaño de ambos sexos podría, asimismo, estar relacionada con un desarrollo larvario más prolongado y, consecuentemente, con una emergencia más tardía de las hembras, como ha sido puesto de manifiesto en otras especies (WIKLUND Y SOLBRECK 1982). En la mayoría de los artrópodos la hembra es de mayor tamaño que el macho, por lo que su velocidad de desarrollo es supuestamente más lenta (BLANCKENHORN et al. 2007). No obstante, la diferencia de tamaños en M. galloprovincialis no es muy acusada, las hembras son generalmente algo mayores que los machos, tanto en peso, por lo que, en definitiva, muestra que son necesarios nuevos estudios sobre la velocidad de desarrollo larvario de M. galloprovincialias, discriminada por sexos, que permitan conocer tanto las causas como las posibles ventajas de la protandria observada en las emergencias analizadas en este trabajo. Tal y como se ha observado en otros sistemas (BENTZ et al. 1991, BAIER et al. 2007), el detallar los requerimientos térmicos de cada una de las fases larvarias pude mejorar mucho el potencial predictor de los modelos de desarrollo.
Acumulación de grados día El cálculo de la acumulación de grados-día para que se produzca la emergencia de M. galloprovincialis, utilizando la propuesta realizada por NAVES y DE SOUSA (2009) de utilización de un umbral mínimo de desarrollo de 12.2 ± 0.8ºC y de contabilización de grados-día desde el 1 de marzo hasta la emergencia del 50% de la población, mostró una gran variabilidad entre las diferentes zonas de estudio, si bien es verdad que pudo ser observada cierta convergencia dentro de cada una de ellas. Sin embargo, en ningún caso, la emergencia del 50% de los individuos se produjo a los 822 propuestos por NAVES y DE SOUSA (2009) en su modelo de predicción, sino a cantidades de grados-día muy inferiores. Además, tras el re-análisis de los datos de NAVES y DE SOUSA (2009) y el re-ajuste a diferentes modelos no lineares que pudieran dar mejores resultados, tampoco se observó una importante variabilidad en la acumulación de calor entre estos nuevos resultados y los publicados por estos autores para Portugal, independientemente del modelo utilizado (Tabla 0.10).

42
No se observó, en resumen, un ajuste mejor para todos los datos, por lo que la poca concordancia entre los resultados de acumulación de calor en las diferentes localizaciones de la península ibérica, incluyendo tanto los experimentos de este trabajo como los trabajos de NAVES y DE SOUSA (2009), no puede deberse a la precisión del modelo propuesto por estos autores. En este sentido, algunas variaciones en los patrones de emergencia a diferentes temperaturas fueron observados por (JUNG et al. 2015) para M. alternatus en Corea. Y PRITCHARD et al. (1996) apuntaron que la acumulación de calor requerida por un insecto podría ser también variable en relación a la temperatura de exposición. Mas recientemente, se ha demostrado que la que el efecto del calor en la acumulación de la tasa de desarrollo no es siempre aditiva (WU et al. 2014). Los resultados de KOUTROUMPA et al. (2008) ya resaltaron que existía una muy alta variación en la edad observada entre cada estadio larvario, y que ésta no podía ser explicada solamente a través de una combinación de temperatura, calidad del material hospedador y día de ovoposición. TOGASHI (1985, 1989, 1990) también sugirió para M. alternatus, al igual que KOUTROUMPA et al. (2008) para M. gallopovincialis, que la calidad del material hospedante podría influir en el desarrollo larvario de forma que la buena calidad del alimento ralentizaría el desarrollo mientras que la baja calidad lo aceleraría. La calidad de las trozas no ha sido tenida en cuenta en este estudio, aunque es cierto que las características del material hospedante fueron muy homogéneas y que las trozas utilizadas fueron seleccionadas por su máxima colonización en caso provenir de incendios, y por la máxima idoneidad para la colonización en el caso de provenir de cortas (además de utilizarse el mismo día de corta), por lo que no se estima que éste sea un factor influyente en las curvas de emergencia estudiadas de este trabajo. En el mismo sentido, KOUTROUMPA et al. (2008) indicaron que, pese a que esto puediese influir en el desarrollo, en sus ensayos se observó mucha menos variación de edad en momento de la emergencia que entre ninguno de los estadios larvarios previos, por lo que se deduce que su efecto sobre el patrón de emergencia no debería ser muy acusado. Ciertamente algunas condiciones experimentales pueden haber interferido, pero las observaciones realizadas en esta memoria sugieren, al menos, que el rol de la temperatura es el fundamental, aunque insuficiente, para explicar de manera óptima la amplia variabilidad en la distribución de las emergencias. Por otro lado, se han observado diferencias en la adaptación térmica entre las poblaciones locales de muchas especies de insectos (SINCLAIR et al. 2012). Y debido a estas variaciones, las respuestas térmicas de las poblaciones locales podrían diferir entre ellas. En la práctica, éstas son tratados como fuentes de error en la predicción de los períodos de emergencia (JUNG et al. 2015). Como especies nativas en la península Ibérica, es posible que algunas características del ciclo biológico de M. galloprovincialis hayan divergido entre las poblaciones locales en respuesta a diferencias en el clima local. Por lo tanto, esta variabilidad aparentemente podría explicarse por la existencia, dentro de la península Ibérica, de adaptaciones locales en las diferentes regiones de la misma, quizás consecuencia de la existencia de poblaciones genéticamente diferenciadas (HARAN et al. 2015). Ningún estudio en este sentido ha sido publicado hasta la fecha por lo que nuestros resultados justifican la necesidad de abrir nuevas líneas de investigación que abarquen estos aspectos. En cualquier caso, el modelo propuesto por NAVES y DE SOUSA (2009) no puede considerarse como un modelo general para predecir la aparición de M. galloprovincialis en toda la Península Ibérica. En términos globales las emergencias de M. galloprovincialis en la península Ibérica están relacionadas con una acumulación de calor menor en las zonas cálidas que en las zonas frías, lo cual está en consonancia con el adelanto de las curvas en zonas cálidas y el retraso en zonas frías. Las altas temperaturas del otoño-invierno anterior a la dormancia podrían acelerar el desarrollo larvario y cambiar la composición de la estructura de edad de las larvas (mayor

43
cantidad de larvas de 4º estadio) antes de la llegada del invierno provocando de esta forma curvas de emergencia más tempranas (PARK et al. 2014) y, como consecuencia, acumulaciones de calor menores. Eso sería así porque el modelo de NAVES y DE SOUSA (2009) contabiliza la acumulación de calor a partir del 1 de marzo hasta la mediana de las emergencias. El análisis de la acumulación de calor a partir del mismo momento de la ovoposición, en vez de la propuesta de NAVES y DE SOUSA (2009), podría ajustar mejor el modelo a escala de la península ibérica. No obstante el desconocimiento de la función de distribución de las puestas de M. galloprovincialis en nuestros ensayos hace imposible realizar un cálculo adecuado del mismo. De todos modos, debe ser tenido en cuenta que el efecto del patrón temporal de ovoposición sobre el patrón de emergencia no debería ser muy acusado, según KOUTROUMPA et al. (2008). Estos resultados ponen definitivamente en evidencia la necesidad de un estudio detallado de los requerimientos térmicos de cada una de las fases larvarias de M. galloprovincialis, al objeto de comprender mejor su desarrollo, incluyendo las posibles diferencias genéticas entre diferentes poblaciones. Si bien no se han podido encontrar patrones generales par la península, la comparación detallada entre series de características comunes permite discutir diferentes aspectos de la fenología de la emergencia de M. galloprovincialis. En concreto, en los ensayos realizados en Calabazanos (Palencia), se produce significativamente una menor acumulación de calor para que se dé el 50% de las emergencias en el año 2014 que en 2013. En este caso, unas anormalmente elevadas temperaturas durante el periodo pre-dormancia y el otoño aparentemente pudieron haber permitido un mayor desarrollo de las larvas anteriores al periodo invernal dando lugar a una composición de edad de dicho conjunto de larvas con un mayor número de ellas en cuarto estadio. Esto está en desacuerdo con el modelo propuesto por NAVES y DE SOUSA (2009), que toma como primer día de suma de los grados-día acumulados el 1 de marzo. Según nuestros datos, dicha asunción aparentemente no sería adecuada en el caso de analizar años con temperaturas otoñales e invernales con grandes diferencias. Estos resultados ponen de manifiesto las consecuencias que un aumento de las temperaturas otoñales e invernales debidos al cambio climático pueden tener en la epidemiología del PWD. Si bien estos efectos se han analizado desde una perspectiva de la distribución de M. galloprovincialis (PÉREZ et al. 2008), tal efecto no ha sido estudiado en el contexto del desarrollo larvario, si bien, resultados como los de 2014, ponen de maniefiesto que tal cambio puede resultar en una extensión del periodo de vuelo del vector, acarreando una mayor probabilidad de expandir la enfermedad. En el caso de los ensayos realizados en Quart de Poblet (Valencia) los experimentos realizados en 2010 con trozas provenientes de Soneja, y en 2011 con trozas provenientes de Callosa d’En Sarrià (ambas de zonas con regímenes térmicos similares), los resultados muestran una acumulación de calor coincidente. El año 2012 la acumulación de calor es mucho mayor ya que debido al tratamiento de frío, la emergencia se prolongó el tiempo. Por otro lado, los ensayos realizados en bidón (Calabazanos 2013 y Quart de Poblet 2012), un ambiente cerrado de mayor inercia térmica, muestran acumulaciones de temperatura mucho más elevadas que las de cualquier otro ensayo, no obstante la toma de datos de temperatura muestra cifras aberrantes por la poca circulación de aire y deben ser tomadas en cuenta con mucha cautela. Aún así, los grados-día observados en estos experimentos son los únicos que coinciden con los observados por NAVES y DE SOUSA (2009), aunque en su caso la temperatura considerada fue la temperatura ambiente, tomada en el exterior de los contenedores de las trozas. El caso de Calabazanos 2013 es el único de todos les ensayos en el que un mismo material hospedante ha sido dividido en dos bloques y cada uno recibido diferente tratamiento en cuanto a su ubicación hasta la emergencia (bidón y jaula). Los resultados muestran que, pese al considerable aumento de temperatura en el interior del barril por la falta de circulación del aire y

44
por confinamiento, se produce una sincronización de las emergencias de todo el material, tanto el ubicado en bidón como el ubicado en jaula. Estos resultados sugieren nuevamente la existencia de adaptaciones locales que determinen el patrón de emergencia del insecto, de forma que la acumulación de calor podría ser simplemente una condición necesaria pero no suficiente para el inicio de la emergencia, mientras que podrían existir otro tipo de señales que determinarían el inicio de la emergencia una vez alcanzada una concreta acumulación de calor. En este sentido TOGASHI et al. (2010) mostró la existencia de efectos del fotoperiodo en el desarrollo larvario de M. urussovi en trozas de árboles. En el caso estudiado, una vez alcanzados los °D necesarios para completar el desarrollo larvario, un exceso de calor podría no afectar a la curva de emergencia, siempre que no se superase el umbral superior de 35,5 ºC propuesto por NAVES y DE SOUSA (2009). Esta temperatura sólo se da en los bidones de cría una vez que la mayor parte de los individuos ha emergido, lo que podría indicar que el efecto de que se supere el umbral superior no es muy acentuado. Por otro lado, en los casos en los que se produce una traslocación del material hospedante entre zonas con distintos regímenes climáticos (Calabazanos2011, Calabazanos2012, Mora2011) se observan resultados erráticos en los análisis de la acumulación de calor, lo que sugiere que, en cierta medida, la manipulación del material produce alguna interferencia en el desarrollo larvario que se ve reflejada en su patrón de emergencia. Esta sincronización, en resumen, sugiere que el mecanismo que determina el patrón de emergencia de M. galloprovincialis en una zona concreta podría ser muy complejo, influido por diferentes variables como la temperatura, el fotoperiodo, la calidad del material hospedador, la distribución de la ovoposición (KOUTROUMPA et al. 2008), y la variabilidad genética o incluso la existencia de algún otro tipo de señales de inicio. Tal y como se ha mencionado anteriormente, estudios futuros deberían centrarse en el efecto detallado de esas variables en el desarrollo larvario de M. galloprovincialis, de manera que las medidas de control destinadas a la gestión sostenible del NMP puedan aplicarse en el momento adecuado.

45
Bibliografía
ABELLEIRA, A., A. PICOAGA, J. P. MANSILLA y O. AGUIN (2011). Detection of Bursaphelenchus xylophilus, Causal Agent of Pine Wilt Disease on Pinus pinaster in Northwestern Spain. Plant Disease 95(6): 776-776.
AKBULUT, S. y W. T. STAMPS (2012). Insect vectors of the pinewood nematode: a review of the biology and ecology of Monochamus species. Forest Pathology 42(2): 89-99.
ALLEN, J. C. (1976). Modified Sine Wave Method for Calculating Degree Days. Environmental Entomology 5(3): 388-396.
ÁLVAREZ, G., I. ETXEBESTE, D. GALLEGO, G. DAVID, L. BONIFACIO, H. JACTEL, E. SOUSA y J. A. PAJARES (2015). Optimization of traps for live trapping of Pine Wood Nematode vector Monochamus galloprovincialis. Journal of Applied Entomology 139(8): 618-626.
BAIER, P., J. PENNERSTORFER y A. SCHOPF (2007). PHENIPS - A comprehensive phenology model of Ips typographus (L.) (Col., Scolytinae) as a tool for hazard rating of bark beetle infestation. Forest Ecology and Management 249(3): 171-186.
BASKERVILLE, G. L. y P. EMIN (1969). Rapid Estimation of Heat Accumulation from Maximum and Minimum Temperatures. Ecology 50(3): 514-517.
BENTZ, B. J. (2006). Mountain pine beetle population sampling: inferences from Lindgren pheromone traps and tree emergence cages. Canadian Journal of Forest Research-Revue Canadienne De Recherche Forestiere 36(2): 351-360.
BENTZ, B. J., J. A. LOGAN y G. D. AMMAN (1991). Temperature-Dependent Development of the Mountain Pine-Beetle (Coleoptera, Scolytidae) and Simulation of Its Phenology. Canadian Entomologist 123(5): 1083-1094.
BLANCKENHORN, W. U., A. F. G. DIXON, D. J. FAIRBAIRN, M. W. FOELLMER, P. GIBERT, K. VAN DER LINDE, R. MEIER, S. NYLIN, S. PITNICK, C. SCHOFF, M. SIGNORELLI, T. TEDER y C. WIKLUND (2007). Proximate causes of Rensch's rule: Does sexual size dimorphism in arthropods result from sex differences in development time? American Naturalist 169(2): 245-257.
BOGICH, T. L., A. M. LIEBHOLD y K. SHEA (2008). To sample or eradicate? A cost minimization model for monitoring and managing an invasive species. Journal of Applied Ecology 45(4): 1134-1142.
EPPO (2009). PM 7/4(2): Bursaphelenchus xylophilus. EPPO Bulletin 39(3): 344-353. EVANS, H. F., O. KULINICH, C. MAGNUSSON, C. ROBINET y T. SCHROEDER (2009).
Report of a Pest Risk Analysis for Bursaphelenchus xylophilus, European and Mediterranean Plant Protection Organization (EPPO): 17.
FRANCARDI, V., J. DE SILVA, F. PENNACCHIO y P. F. ROVERSI (2009). Pine volatiles and terpenoid compounds attractive to European xylophagous species, vectors of Bursaphelenchus spp. nematodes. Phytoparasitica 37(4): 295-302.
FRANCARDI, V. y F. PENNACCHIO (1996). Note sulla bioecologia di Monochamus galloprovincialis (Olivier) in Toscana e in Liguria (Coleoptera Cerambycidae). Redia 79: 153-169.
FUTAI, K. (2013). Pine Wood Nematode, Bursaphelenchus xylophilus. Annual Review of Phytopathology 51(1): 61-83.
HARAN, J., A. ROQUES, A. BERNARD, C. ROBINET y G. ROUX (2015). Altitudinal Barrier to the Spread of an Invasive Species: Could the Pyrenean Chain Slow the Natural Spread of the Pinewood Nematode? PLoS ONE 10(7): e0134126.
HELLRIGL, K. G. (1971). Die Bionomie der europaischen Monochamus-Arten (Coleopt., Cerambycid.) und ihre Bedeutung fur die Forstund Holzwirtschaft. Redia 52: 367-509.
IKEMOTO, T., I. KURAHASHI y P. J. SHI (2013). Confidence interval of intrinsic optimum temperature estimated using thermodynamic SSI model. Insect Science 20(3): 420-428.

46
JIKUMARU, S. y K. TOGASHI (1996). Effect of temperature on the post-diapause development of Monochamus saltuarius (Gebler) (Coleoptera: Cerambycidae). Applied Entomology and Zoology 31(1): 145-148.
JIKUMARU, S., K. TOGASHI, A. TAKETSUNE y F. TAKAHASHI (1994). Oviposition Biology of Monochamus saltuarius (Coleopter, Cerambycidae) at a constant-temperature. Applied Entomology and Zoology 29(4): 555-561.
JUNG, C. S., S.-H. KOH, Y. NAM, J. J. AHN, C. Y. LEE y W. I. CHOI (2015). A Model for Predicting Spring Emergence of Monochamus saltuarius (Coleoptera: Cerambycidae) from Korean white pine, Pinus koraiensis.
KOBAYASHI, F., A. YAMANE y T. IKEDA (1984). The japanese pine sawyer beetle as the vector of pine wilt disease. Annual Review of Entomology 29: 115-135.
KONTODIMAS, D. C., P. A. ELIOPOULOS, G. J. STATHAS y L. P. ECONOMOU (2004). Comparative temperature-dependent development of Nephus includens (Kirsch) and Nephus bisignatus (Boheman) (Coleoptera : Coccinellidae) preying on Planococcus citri (Risso) (Homoptera : Pseudococcidae): Evaluation of a linear and various nonlinear models using specific criteria. Environmental Entomology 33(1): 1-11.
KOUTROUMPA, F. A., B. VINCENT, G. ROUX-MORABITO, C. MARTIN y F. LIEUTIER (2008). Fecundity and larval development of Monochamus galloprovincialis (Coleoptera Cerambycidae) in experimental breeding. Annals of Forest Science 65(7).
LINIT, M. J. (1988). Nematode Vector Relationships in the Pine Wilt Disease System. Journal of Nematology 20(2): 227-235.
LOGAN, J. A. (1988). Toward an Expert System for Development of Pest Simulation-Models. Environmental Entomology 17(2): 359-376.
MAGRAMA. (2015). "Nematodo de la madera del pino." 2015, de http://www.magrama.gob.es/es/agricultura/temas/sanidad-vegetal/nematodo-de-la-madera-del-pino/#.
MORBEY, Y. E. y R. C. YDENBERG (2001). Protandrous arrival timing to breeding areas: a review. Ecology Letters 4(6): 663-673.
MOTA, M. M., H. BRAASCH, M. A. BRAVO, A. C. PENAS, W. BURGERMEISTER, K. METGE y E. SOUSA (1999). First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1: 727-734.
MYERS, S. W., K. E. ZYLSTRA, J. A. FRANCESE, D. M. BORCHERT y S. M. BAILEY (2014). Phenology and flight periodicity of Sirex noctilio (Hymenoptera: Siricidae) in central New York, U.S.A. Agricultural and Forest Entomology 16(2): 129-135.
NAVES, P. y E. DE SOUSA (2009). Threshold temperatures and degree-day estimates for development of post-dormancy larvae of Monochamus galloprovincialis (Coleoptera: Cerambycidae). Journal of Pest Science 82(1): 1-6.
NAVES, P. M., E. M. DE SOUSA y J. A. QUARTAU (2007). Winter dormancy of the pine sawyer Monochamus galloprovincialis (Col., Cerambycidae) in Portugal. Journal of Applied Entomology 131(9-10): 669-673.
PAJARES, J. A., G. ALVAREZ, F. IBEAS, D. GALLEGO, D. R. HALL y D. I. FARMAN (2010). Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. Journal of Chemical Ecology 36(6): 570-583.
PAJARES, J. A., F. IBEAS, J. J. DIEZ y D. GALLEGO (2004). Attractive responses by Monochamus galloprovincialis (Col., Cerambycidae) to host and bark beetle semiochemicals. Journal of Applied Entomology 128(9-10): 633-638.
PARK, C., D. KIM, S. LEE, Y. MOON, Y. CHUNG y D. S. KIM (2014). A forecasting model for the adult emergence of overwintered Monochamus alternatus (Coleoptera: Cerambycidae) larvae based on degree-days in Korea. Applied Entomology and Zoology 49(1): 35-42.
PÉREZ, G., J. DÍEZ, F. IBEAS y J. PAJARES (2008). Assessing pine wilt disease risk under a climate change scenario in Northwestern Spain. Managing Forest Ecosystems: The

47
Challenge of Climate Change. F. Bravo, R. Jandl, V. LeMay y K. von Gadow, Springer: 269-282.
PRITCHARD, G., L. D. HARDER y R. A. MUTCH (1996). Development of aquatic insect eggs in relation to temperature and strategies for dealing with different thermal environments. Biological Journal of the Linnean Society 58(2): 221-244.
PUKKALA, T., T. MOYKKYNEN y C. ROBINET (2014). Comparison of the potential spread of pinewood nematode (Bursaphelenchus xylophilus) in Finland and Iberia simulated with a cellular automaton model. Forest Pathology 44(5): 341-352.
QUESADA-MORAGA, E. y C. SANTIAGO-ALVAREZ (2000). Temperature related effects on embryonic development of the Mediterranean locust, Dociostaurus maroccanus. Physiological Entomology 25(2): 191-195.
R CORE TEAM (2015). R: A language and environment for statistical computing. Vienna, Austria, R Foundation for Statistical Computing.
RAFFA, K. F. y A. A. BERRYMAN (1983). The Role of Host Plant-Resistance in the Colonization Behavior and Ecology of Bark Beetles (Coleoptera, Scolytidae). Ecological Monographs 53(1): 27-49.
REGNIERE, J., J. POWELL, B. BENTZ y V. NEALIS (2012). Effects of temperature on development, survival and reproduction of insects: Experimental design, data analysis and modeling. Journal of Insect Physiology 58(5): 634-647.
ROBERTSON, L., S. C. ARCOS, M. ESCUER, R. S. MERINO, G. ESPARRAGO, A. ABELLERA y A. NAVAS (2011). Incidence of the pinewood nematode Bursaphelenchus xylophlius Steiner & Buhrer, 1934 (Nickle, 1970) in Spain. Nematology 13: 755-757.
ROBERTSON, L., A. GARCIA-ALVAREZ, S. C. ARCOS, M. A. DIEZ-ROJO, J. PEDRO MANSILLA, R. SANZ, C. MARTINEZ, M. ESCUER, L. CASTRESANA, A. NOTARIO, A. BELLO y M. ARIAS (2008). Potential Insect Vectors of Bursaphelenchus spp. (Nematoda: Parasitaphelenchidae) in Spanish Pine Forests. Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems. M. M. Mota y P. Vieira: 221-233.
ROBINET, C., A. ROQUES, H. PAN, G. FANG, J. YE, Y. ZHANG y J. SUN (2009). Role of Human-Mediated Dispersal in the Spread of the Pinewood Nematode in China. PLoS ONE 4(2): e4646.
ROBINET, C., N. VAN OPSTAL, R. BAKER y A. ROQUES (2011). Applying a spread model to identify the entry points from which the pine wood nematode, the vector of pine wilt disease, would spread most rapidly across Europe. Biological Invasions 13(12): 2981-2995.
ROLTSCH, W. J., F. G. ZALOM, A. J. STRAWN, J. F. STRAND y M. J. PITCAIRN (1999). Evaluation of several degree-day estimation methods in California climates. International Journal of Biometeorology 42(4): 169-176.
RUTHERFORD, T. A., E. RIGA y J. M. WEBSTER (1992). Temperature-mediated behavioral relationships in Bursaphelenchus xylophilus, B. mucronatus, and their hybrids. Journal of Nematology 24(1): 40-44.
SANCHEZ-HUSILLOS, E., I. ETXEBESTE y J. PAJARES (2015). Effectiveness of mass trapping in the reduction of Monochamus galloprovincialis Olivier (Col.: Cerambycidae) populations. Journal of Applied Entomology.
SINCLAIR, B. J., C. M. WILLIAMS y J. S. TERBLANCHE (2012). Variation in Thermal Performance among Insect Populations*. Physiological and Biochemical Zoology 85(6): 594-606.
SOUSA, E., M. A. BRAVO, J. PIRES, P. NAVES, A. C. PENAS, L. BONIFACIO y M. M. MOTA (2001). Bursaphelenchus xylophilus (Nematoda; Aphelenchoididae) associated with Monochamus galloprovincialis (Coleoptera; Cerambycidae) in Portugal. Nematology 3: 89-91.

48
SOUSA, E., P. NAVES, L. BONIFACIO, M. A. BRAVO, A. C. PENAS, J. PIRES y M. SERRAO (2002). Preliminary survey for insects associated with Bursaphelenchus xylophilus in Portugal. Bulletin OEPP 32(3): 499-502.
TOGASHI, K. (1985). Transmission curves of Bursaphelenchus xylophilus (Nematoda, Aphelenchoididae) from its vector, Monochamus alternatus (Coleoptera, Cerambycidae), to pine trees with reference to population performance. Applied Entomology and Zoology 20(3): 246-251.
TOGASHI, K. (1989). Development of Monochamus alternatus Hope (Coleoptera, Cerambycidae) in relation to oviposition time. Japanese Journal of Applied Entomology and Zoology 33(1): 1-8.
TOGASHI, K. (1990). A field experiment on dispersal of newly emerged adults of Monochamus alternatus (Coleoptera, Cerambycidae). Researches on Population Ecology 32(1): 1-13.
TOGASHI, K., H. KASUGA, H. YAMASHITA y K. IGUCHI (2008). Effect of host tree species on larval body size and pupal-chamber tunnel of Monochamus alternatus (Coleoptera : Cerambycidae). Applied Entomology and Zoology 43(2): 235-240.
TOGASHI, K., H. KASUGA, H. YAMASHITA y K. IGUCHI (2010). Larval diapause of Monochamus urussovi and photoperiodic effects on larval development in tree bolts. Journal of Applied Entomology 134(8): 672-674.
TOGASHI, K. y N. SHIGESADA (2006). Spread of the pinewood nematode vectored by the Japanese pine sawyer: modeling and analytical approaches. Population Ecology 48(4): 271-283.
TOMMINEN, J. (1993). Development of Monochamus galloprovincialis Olivier (Coleoptera, Cerambycidae) in Cut Trees of Young Pines (Pinus sylvestris L.) and Log Bolts in Southern Finland. Entomologica Fennica 4(3): 137-142.
TORRES-VILA, L. M., C. ZUGASTI, J. M. DE-JUAN, M. J. OLIVA, C. MONTERO, F. J. MENDIOLA, Y. CONEJO, Á. SÁNCHEZ, F. FERNÁNDEZ, F. PONCE y G. ESPÁRRAGO (2015). Mark-recapture of Monochamus galloprovincialis with semiochemical-baited traps: population density, attraction distance, flight behaviour and mass trapping efficiency. Forestry 88(2): 224-236.
TRÖLTZSCH, K., J. VAN BRUSSELEN y A. SCHUCK (2009). Spatial occurrence of major tree species groups in Europe derived from multiple data sources. Forest Ecology and Management 257(1): 294-302.
VICENTE, C., M. ESPADA, P. VIEIRA y M. MOTA (2012). Pine Wilt Disease: a threat to European forestry. European Journal of Plant Pathology 133(1): 89-99.
WERMELINGER, B. y M. SEIFERT (1998). Analysis of the temperature dependent development of the spruce bark beetle Ips typographus (L) (Col., Scolytidae). Journal of Applied Entomology 122(1-5): 185-191.
WIKLUND, C. y C. SOLBRECK (1982). Adaptive Versus Incidental Explanations for the Occurrence of Protandry in a Butterfly, Leptidea-Sinapis L. Evolution 36(1): 56-62.
WU, T. H., S. F. SHIAO y T. OKUYAMA (2014). Development of insects under fluctuating temperature: a review and case study. Journal of Applied Entomology.
ZAMORA, P., V. RODRÍGUEZ, F. RENEDO, A. V. SANZ, J. C. DOMÍNGUEZ, G. ESCOLAR, J. MIRANDA, B. ÁLVAREZ, A. GONZÁLEZ-CASAS, E. MAYOR, M. DUEÑAS, A. MIRAVALLES, A. NAVAS, L. ROBERTSON y A. B. MARTÍN (2015). First report of Bursaphelenchus xylophilus causing Pine Wilt Disease on Pinus radiata in Spain. Plant Disease.

49
Capítulo 3
Fenología del vuelo de Monochamus
galloprovincialis (Col.: Cerambycidae) en la península ibérica
Introducción
Monochamus galloprovincialis Olivier (Coleoptera: Cerambycidae) es el vector europeo del Nematodo de la Madera del Pino (NMP), Bursaphelenchus xylophilus (Steiner & Bührer), causante de la Enfermedad del Marchitamiento del Pino (PWD es su acrónimo en inglés). Desde su detección en Portugal y en España, planes de erradicación y contingencia han sido establecidos en diferentes países con el objetivo de establecer medidas para contener la dispersión de la enfermedad por el continente europeo (VICENTE et al. 2012, MAGRAMA 2015b). Actualmente las herramientas aplicadas para el control del NMP en Europa y en la península Ibérica se reducen a la selvicultura y a la inspección fitosanitaria, esto es: tala y destrucción completa de los pies sintomáticos antes del inicio del periodo de emergencia de M. galloprovincialis (para evitar de esta forma la dispersión de los vectores portadores de NMP), y prospección de las zonas en función del riesgo de entrada de la enfermedad. No obstante, diversos trabajos durante la última década (PAJARES et al. 2004, IBEAS et al. 2007, PAJARES et al. 2010), han dado lugar al desarrollo y comercialización de un cebo cairomonal y feromonal (Galloprotect 2D. SEDQ, Barcelona) que permite obtener una alta tasa de capturas de ambos sexos de M. galloprovincialis, lo que facilita una buena monitorización de su periodo de vuelo (SANCHEZ-HUSILLOS et al. 2015, TORRES-VILA et al. 2015). A su vez, diversos avances han posibilitado la mejora de las trampas empleadas (ÁLVAREZ et al. 2015). La temperatura es la variable ambiental que más condiciona la ontogenia y fenología de los seres vivos, y particularmente la de los insectos, que al ser organismos poiquilotermos padecen directamente de las fluctuaciones de la temperatura ambiental (GILBERT Y RAWORTH 1996, GILLOOLY et al. 2002). En este sentido, aunque hay diversos estudios que prueban que la temperatura afecta al desarrollo larvario de M. galloprovincialis (KOUTROUMPA et al. 2008, NAVES Y DE SOUSA 2009), no se conoce con suficiente profundidad la incidencia de ésta sobre el periodo de vuelo del insecto adulto. En consecuencia, tampoco se conoce detalladamente la influencia de variables ambientales complejas como la altitud en la supervivencia o en el establecimiento de sus poblaciones en zonas de alta montaña. El trabajo publicado recientemente por HARAN et al. (2015) muestra que, en Pirineos, a altidudes superiores a los 1590 mnsm y con mínimas medias en invierno por debajo de los -7ºC, las poblaciones de M. galloprovincialis son tan bajas que se proponen esos valores ambientales como límite de distribución de la especie.

50
En efecto, algunos autores han estudiado la influencia de la temperatura en el desarrollo larvario de M. galloprovincialis (KOUTROUMPA et al. 2008, NAVES Y DE SOUSA 2009), basándose en modelos de acumulación de calor que vinculan la contabilización de los requerimientos térmicos a la finalización de la dormancia invernal. Sin embargo, otros estudios (ver Capítulo 2) sugieren que la temperatura del otoño e invierno posteriores a la ovoposición también podrían influir en este desarrollo provocando adelantos o retrasos en la fenología de las emergencias de los insectos (GALLINAT et al. 2015). En consonancia con esta sugerencia, HARAN et al. (2015) propusieron la media de las temperaturas mínimas de los meses de invierno como variable relacionada con la supervivencia y la capacidad de dispersión de M. galloprovincialis en zonas frías. Hay que tener en cuenta que este parámetro puede afectar a las larvas de un insecto cuyo desarrollo se expande desde el verano hasta la primavera del año siguiente, incluso llegando a tener ciclos semivoltinos en las zonas más frías de su amplia distribución geográfica (TOMMINEN 1993). Las curvas de vuelo de M. galloprovincialis descritas por NAVES et al. (2008) mostraron periodos que se extendían desde mayo hasta finales de octubre en Portugal, teniendo un máximo de capturas entre los meses de junio y julio, si bien es cierto que en sus resultados fueron detectadas diferencias entre los patrones de vuelo de los diferentes años. A su vez, diferentes estudios y documentos técnicos vinculados a la gestión del NMP han mostrado curvas de vuelo de M. galloprovincialis con perfiles bimodales a lo ancho de toda la península Ibérica, en las que un primer pico se da en el mes de junio, para posteriormente registrar un descenso en las capturas en el mes de julio y volver a mostrar un segundo pico, que suele ser el máximo, a final de verano (MAGRAMA 2015a, RODRIGUES Com. Pers.). La falta de comprensión de este fenómeno en la fenología de M. galloprovincialis ha determinado que uno de los objetivos de este trabajo sea estudiar la incidencia de la temperatura en el inicio del vuelo y en el descenso de las capturas estivales producidas en la parte intermedia del periodo de vuelo en alguna de las localidades. Por otro lado, en la zona afectada por el B. xylophilus en Portugal, existen masas importantes de Pinus pinaster Aiton, Pinus pinea L., y algunos pies dispersos de P. halepensis Miller. Sin embargo, P. pinaster ha sido el principal afectado por la enfermedad, y consecuentemente es considerado el hospedador local más importante para M. galloprovincialis. Esta especie de pino ocupa casi un millón de hectáreas en Portugal, alrededor del 30% de la superficie forestal del país (DAS FLORESTAS 2001). Por el contrario, en Europa del Norte y Central, Pinus sylvestris L. es el hospedador más importante de M. galloprovincialis (HELLRIGL 1971, FRANCARDI Y PENNACCHIO 1996). Además, al ser P. sylvestris la especie de Pinus más extendida del mundo, y tener su límite de distribución suroccidental en la península Ibérica (RICHARDSON et al. 1998), el estudio del comportamiento de M. galloprovincialis sobre esta especie cobra especial relevancia. Por lo tanto, si bien la preocupación sobre la potencial dispersión de la enfermedad a lo ancho de Europa se centra en el tándem M. galloprovincialis/P. pinaster (FVO-CE, Com. Pers.), los estudios sobre la idoneidad de otras especies hospedadoras para el desarrollo de M. galloprovincialis sugieren que tanto P. sylvestris como P. halepensis podrían ser adecuados para la alimentación y la oviposición del insecto en Portugal, por lo que también podrían serlo en el resto de Europa (NAVES et al. 2006, SANCHEZ-HUSILLOS et al. 2013). Además, la reciente detección del NMP en pies de P. nigra Arnold y P. radiata Don amplia el espectro de hospedantes a considerar (INÁCIO et al. 2015, ZAMORA et al. 2015). El caso de P. sylvestris es especialmente delicado por diversas razones: en primer lugar está ampliamente extendido por todo el continente europeo y se trata de una especie de alto valor ecológico y comercial. En segundo lugar es una especie altamente susceptible a la enfermedad causada por el NMP (NUNES DA SILVA et al. 2015); y por último, como se ha comentado, M.

51
galloprovincialis muestra una alta atracción hacia esta especie en sus preferencias tanto para la alimentación como para la ovoposición (NAVES et al. 2006, SANCHEZ-HUSILLOS et al. 2013). En la península Ibérica, P. sylvestris ocupa los ecosistemas de los pisos oromediterráneo y subalpino, generalmente a altitudes comprendidas entre 800 y los 2040 msnm, en las zonas más elevadas de las cadenas montañosas peninsulares. Desde esta perspectiva, uno de los pinares de P. sylvestris más extensos, valiosos e importantes de la península ibérica es el Pinar de Valsaín, ubicado en la Sierra de Guadarrama, dentro del Sistema Central, y que ha sido declarado recientemente Parque Nacional. Este pinar se encuentra, aunque ocupando las cotas más altas, en contacto con formaciones vegetales dominadas por P. pinaster formando un corredor forestal casi continuo que cruza de Este a Oeste el Sistema Central, desde Guadarrama, pasando por la Sierra de Gredos, hasta la Sierra de Gata (donde han sido detectados los últimos dos focos de la enfermedad en España) y la portuguesa Serra da Malcata. Este corredor forestal supone una de las zonas con mayor riesgo para dispersión natural de la enfermedad en la península y a su vez una de las mejores zonas donde estudiar la influencia de la altura en el vuelo de M. galloprovincialis (MAGRAMA 2015b). Por ello, un segundo objetivo de este trabajo es estudiar la incidencia del gradiente altitudinal de alta montaña sobre la curva de vuelo y en las poblaciones de M. galloprovincialis en dicha región. En resumen, los diferentes experimentos fueron diseñados con el objetivo de averiguar:
Influencia de la altitud en las capturas de M. galloprovincialis. Influencia de las temperaturas en el inicio del periodo de vuelo de M. galloprovincialis. Influencia de las altas temperaturas en el descenso de capturas estivales de M.
galloprovincialis. Estudio de los vuelos circadianos de M. galloprovincialis.

52
Material y métodos
Un total de 8 experimentos de captura de M. galloprovincialis mediante trampas cebadas con atrayentes cairomonales y feromonales fueron llevados a cabo en diferentes regiones de la península Ibérica entre los años 2010 y 2015 (Tabla 3.1, Figura 3.1). La metodología fue esencialmente común a todos ellos en lo referente al dispositivo de captura. Se utilizaron trampas de bandas cruzadas modelo Crosstrap® (ECONEX, Murcia), trampas multiembudo o trampas experimentales ‘TORRE LSF’ de equivalente efectividad (HERNÁNDEZ et al. 2011, ÁLVAREZ et al. 2015). Se realizó captura con muerte (en húmedo con propilenglicol, o en seco recurriendo al uso de insecticidas en el interior del bote colector) o en vivo, utilizando trampas modificadas para evitar la huida de los insectos (ÁLVAREZ et al. 2015). Como atrayente se utilizó el cebo comercial para M. galloprovincialis (Galloprotect Plus®, SEDQ S.L., Barcelona) que libera dos feromonas de escolítidos (ipsenol y 2-metil-3buten-1-ol, a 2.5 y 12 mg/día respectivamente), una cairomona del hospedador (α-pineno, a 500 mg/día) y una feromona específica del género Monochamus (2-undecyloxy-1-ethanol a 2 mg/día (PAJARES et al. 2010, MACIAS-SAMANO et al. 2012, PAJARES et al. 2013). La renovación de los cebos se realizó según las indicaciones del fabricante (cada 6 semanas) y la revisión de las trampas fue realizada periódicamente (Tabla 3.1) hasta la finalización del periodo de vuelo del insecto. Se realizó un registro de temperatura en los diferentes periodos de muestreo, que fueron recogidos en estaciones cercanas al área de estudio o mediante sensores de temperatura HOBO pendant UA-001-64 (Onset, USA) instalados en las diferentes zonas de estudio. La siguiente tabla resume las localizaciones y los periodos de muestreo correspondientes a los diferentes trampeos.
Tabla 3.1 Localización y características de los diferentes ensayos de captura de M. galloprovincialis
Exp Año Localidad Provincia Fecha Inicio Fecha Fin Nº trampas Frecuencia Muestreo 1 2010 Jerica Castelló 29/05/2010 25/10/2010 16 3 muestreos/semana 2 2011 Altura Castelló 15/06/2011 20/11/2011 19 2 muestreos/10 días 3 2010 Mora de Rubielos Teruel 17/05//2010 15/11/2010 18 3 muestreos/semana 4 2011 Mora de Rubielos Teruel 10/05/2011 04/11/2011 38 3 muestreos/semana 5 2011 Valsaín/Cercedilla Segovia/Madrid 27/05/2011 28/10/2011 17 Semanal 6 2012 Valsaín/Cercedilla Segovia/Madrid 01/06/2012 29/10/2012 27 Quincenal 7 2013 Valsaín/Cercedilla Segovia/Madrid 07/07/2013 14/10/2013 27 Quincenal 8 2015 Alcoi Alacant 28/08/2015 06/09/2015 2 Horaria
Figura 3.1 Localización de los ensayos de captura de M. galloprovincialis

53
Figura 3.1 Localización de los ensayos de captura de M. galloprovincialis
Todos los experimentos fueron diseñados con el objetivo de dar respuesta a los objetivos enunciados:
Influencia de la altitud en las capturas de M. galloprovincialis
Para la evaluación de la incidencia de la altitud en la captura de M. galloprovincialis fueron realizados 3 ensayos (Exp5, Exp6 y Exp7 en Tabla 3.1). En todos ellos un conjunto de trampas fue instalado consecutivamente durante los años 2011, 2012 y 2013 en la Sierra de Guadarrama. Se siguió un itinerario que ascendía en cota de manera en tramos de 100 m, y de orientación (vertientes Norte y Sur) del Sistema Central peninsular (Figura 3.2 y Figura 3.3). En la vertiente Norte de la Sierra, un transecto de trampeo discurrió por el monte Pinar de Valsaín (San Ildefonso-La Granja, Segovia), desde la Cruz de la Gallega hasta el Puerto de la Fuenfría, y desde allí, por la vertiente Sur, un segundo transecto de trampas descendió hasta la Colonia de las Dehesas (Cercedilla, Madrid) (Figura 3.3). La zona muestrada cubrío una masa natural adulta de P. sylvestris.

54
Figura 3.2 Mapa de la Península Ibérica con la cobertura estimada de coníferas según EFI y la localización de los experimentos desarrollados en la Sierra de Guadarrama (Segovia-Madrid)
Figura 3.3 Localización de las trampas de los experimentos desarrollados en la Sierra de
Guadarrama (Segovia-Madrid) En 2011 se instalaron 17 trampas, 9 de ellas en Valsaín (exposición Norte), en un rango aproximado entre los 1350 y los 1850 metros de altitud, y las 8 restantes en Cercedilla (exposición Sur), en el mismo rango altitudinal. El periodo de trampeo se extendió desde el 27 de mayo al 28 de octubre. Se instaló un sensor de temperatura modelo HOBO Pendant UA-001-64 (Onset, USA) a 1700 m para obtener los datos de temperatura. La frecuencia de muestreo fue semanal. Durante el año 2011 también fueron incorporadas al estudio un grupo de 5 trampas colocadas para el muestreo de M. galloprovincialis en el entorno de un aserradero en la parte más baja del Pinar de Valsaín entre los 1150 y los 1450 m de altitud (Tabla 3.2). Durante 2012 se amplió el dispositivo de trampas hasta las 27. El itinerario en vertiente Norte se continuó en descenso hasta los 1150 metros y se incrementó el número de trampas hasta las altitudes superiores. En el itinerario en vertiente Sur, el trampeo descendió hasta los 1250 m. El dispositivo fue revisado desde el 1 de junio hasta el 29 de octubre. Se instalaron un total de cinco sensores meteorológicos HOBO Pendant UA-001-64 (Onset, USA) siguiendo un gradiente altitudinal (Tabla 3.2). En 2013 se mantuvo el dispositivo de 27 trampas del año anterior. El periodo de revisiones comprendió desde el 7 de julio al 14 de octubre. Se mantuvieron únicamente los sensores meteorológicos a 1600 metros de altitud en ambas vertientes, así como el sensor ubicado a 1800 m. La frecuencia de muestreo fue quincenal (Tabla 3.2).
Tabla 3.2 Distribución del número de trampas por años y altitud, ubicadas en la Sierra de Guadarrama Año Altitud 2011 2012 2013 1150 4 1 1 1250 0 3 3 1350 3 4 4 1450 4 3 3 1550 3 3 3 1650 5 5 5

55
Año Altitud 2011 2012 2013 1750 3 5 5 1850 1 3 3
Influencia de las temperaturas en el inicio del periodo de vuelo de M. galloprovincialis
Para el estudio de la influencia de las temperaturas en el inicio del periodo de vuelo de M. galloprovincialis fue utilizado el ensayo realizado en la Sierra de Guadarrama en el año 2012, descrito en el apartado anterior (Exp6, Tabla 3.1). En este caso, del total de las 27 trampas utilizadas en este ensayo y colocadas entre los 1150 y los 1850 msnm, fue registrada la temperatura en 5 de ellas (ubicadas a 1363 m, 1431 m, 1644 m, 1685 m, y 1693 m sobre el nivel del mar, respectivamente) mediante la colocación de cinco sensores de temperatura HOBO pendant UA-001-64 (Onset, USA). La revisión se llevó a cabo entre el 1 de junio y el 29 de octubre de 2012. La frecuencia de muestreo fue quincenal (Figura 3.2 y Figura 3.3)
Influencia de las altas temperaturas en el descenso de capturas estivales de M.
galloprovincialis
Para la evaluación de la incidencia de la temperatura estival en la captura de M. galloprovincialis fueron realizados 4 ensayos durante los años 2010 y 2011, dos de ellos en la provincia de Teruel y dos de ellos en la de Castellón, respectivamente (Ex1, Exp2, Exp3 y Exp4 en la Tabla 3.1). El dispositivo de captura utilizado en la provincia de Teruel en el año 2010 constó de 18 trampas “TorreLSF” metálicas (PÉREZ et al. 2004, HERNÁNDEZ ALONSO et al. 2007). El ensayo se realizó en una masa adulta de P, pinaster y P. nigra salzmannii ubicada en el paraje Camarracho, en los términos municipales de Cabra de Mora, Mora de Rubielos y Valbona, de la provincia de Teruel. Las trampas se ubicaron en un gradiente altitudinal que osciló entre los 930 m y los 1365 msnm (Figura 3.1 y Figura 3.4). La frecuencia de muestreo fue de 3 veces por semana entre el 17 de mayo y el 15 de noviembre de 2010 (HERNÁNDEZ et al. 2011).

56
Figura 3.4 Área de estudio y localización de las trampas utilizadas en el experimentos Exp3 y Exp 4 ubicados en la provincia de
Teruel durante los años 2010 y 2011. Un segundo ensayo fue realizado en esa misma zona de estudio en 2011. En él se incrementó el número de trampas hasta un total de 38, de las cuales 22 fueron de tipo “TorreLSF” metálicas y 16 de tipo Lindgren modificadas(ÁLVAREZ et al. 2015). Se mantuvo la frecuencia de muestreo de 3 veces por semana entre el 10 de mayo y el 4 de noviembre de 2011. Otros dos ensayos fueron realizados en la provincia de Castellón: En el año 2010, un dispositivo de 16 trampas Lindgren modificadas fue instalado en el MUP La Muela, Roquetillo y Feliciano (Jérica, Castellón). El área de muestreo fue una repoblación naturalizada de P. halepensis a una altitud entre 520 y 850 m. (Figura 3.1 y Figura 3.5) La frecuencia de muestreo fue de 3 veces semanales entre el 29 de mayo y el 25 de octubre de 2010. En el año 2011, un conjunto de 19 trampas Lindgren modificadas (ÁLVAREZ et al. 2015) fueron instaladas en el Monte de Utilidad Pública (MUP) Las Boqueras (Altura, Castellón) entre el 15.06.2011 y el 20.11.2011 (Figura 3.1 y Figura 3.5). El área de estudio estuvo compuesta por una masa natural de P. halepensis a una altitud entre los 800m y 900 m, con presencia reducida de otras especies acompañantes provenientes de repoblación (P. nigra austriaca, P. pinaster y P. sylvestris). La frecuencia de muestreo fue de 2 veces cada 10 días entre el 15 de junio de 2011 y el 20 de noviembre de 2011.

57
Figura 3.5 Áreas de muestreo de los ensayos realizados en Castelló en el año 2010 (Exp1) y 2011 (Exp2), respectivamente
Evolución de la sex-ratio durante el vuelo de M. galloprovincialis
Para la evaluación de las diferencias entre las curvas de vuelo de machos y hembras de M. galloprovincialis fueron utilizados los mismos 4 experimentos descritos en el apartado anterior, realizados en Castellón y Teruel en los años 2010 y 2011.
Vuelos circadianos de M. galloprovincialis
Un dispositivo de 2 trampas Lindgren modificadas (ÁLVAREZ et al. 2015) fue instalado en el Parque Natural La Font Roja de Alcoi (Alicante) en el año 2015. La zona de muestreo fue una masa natural de P. halepensis a una altitud de 800 msnm (Figura 3.1 y Figura 3.6). La frecuencia de muestreo fue horaria entre el 28.08.2015 y el 06.09.2015, con el objetivo de registrar las capturas de M. galloprovincialis por horas y poder evaluar la frecuencia circadiana de sus vuelos. La temperatura fue registrada mediante un HOBO pendant UA-001-64 (Onset, USA) durante todo el periodo de muestreo.
Figura 3.6 Área de muestreo y localización de las trampas utilizadas en el estudio de los vuelos circadianos de M. galloprovincialis
en Alcoi (Alicante), Exp2015/a

58
Análisis estadísticos De la misma manera, los análisis estadísticos atendieron a los diferentes objetivos propuestos en este capítulo. Todos los cálculos y representaciones han sido realizados bajo el entorno de programación y lenguaje estadístico R (R CORE TEAM 2015).
Influencia de la altitud en las capturas de M. galloprovincialis
Se analizó, además de la altitud, la posible influencia de otras variables, como la exposición de la ladera o la cobertura de coníferas, en las capturas de M. galloprovincialis durante los años 2011, 2012 y 2013. Para ello fue realizada una modelización linear generalizada mixta, tomando el año de estudio como factor aleatorio, y la altitud, exposición y cobertura de coníferas como variables fijas. La transformación de la exposición para el análisis fue realizada según ROBERTS y COOPER (1989). En el análisis de la influencia de la altitud en las capturas, fue utilizado un modelo de regresión logístico binomial relacionando la altura con las capturas relativas acumuladas para cada año de estudio. Este análisis fue realizado siguiendo dos aproximaciones distintas: capturas acumuladas por trampas individuales y por capturas de trampas acumuladas por diferentes rangos altitudinales. Asimismo se analizó la influencia de la altitud en la duración del periodo de vuelo, mediante una regresión polinomial.
Influencia de las temperaturas en el inicio del periodo de vuelo de M. galloprovincialis
Para llevar a cabo el análisis, en primer lugar, se modelizó la variación local de la temperatura con la altitud (Anejo 3.1). Posteriormente, se realizó una regresión lineal, para estudiar la influencia de las temperaturas (máximas, mínimas y medias) en el periodo de vuelo. Complementariamente, este estudio también se abordó calculando la acumulación de calor como la suma de temperaturas durante cada periodo (grados-hora) y llevando a una gráfica las capturas proporcionales.
Influencia de las altas temperaturas en el descenso de capturas estivales de M.
galloprovincialis
Para este análisis se procedió a detectar anomalías en las series temporales de temperaturas y capturas. Ambas series se ajustaron a un modelo aditivo generalizado (GAM) con el objetivo de suavizar la curva. La primera derivada de los modelos ajustados se utilizó para evaluar la significancia de las tendencias. Para el análisis, se utilizaron las temperaturas medias diarias durante periodo de muestreo, y se estandarizaron las capturas a medias de las capturas totales por día.
Evolución de la sex-ratio durante el vuelo de M. galloprovincialis
Para el análisis de las posibles diferencias existentes en la sucesión temporal de capturas de M. galloprovincialis entre machos y hembras, la razón sexual (número de machos dividido por el número de hembras, sex ratio) de las capturas fue representada frente a la fecha de muestreo y se ajustó un modelo de regresión lineal. Para el ajuste del modelo fueron descartadas las fechas con capturas insuficientes.

59
Vuelos circadianos de M. galloprovincialis
Las capturas horarias de M. galloprovincialis se representaron gráficamente mediante histogramas y diagramas de cajas. En el caso del histograma, se computó también la densidad estadística de las capturas. Para evaluar gráficamente la distribución de las capturas en función de la hora del día y de a temperatura, se generó un ráster utilizando las capturas como variable z y las mencionadas variables como las coordenadas x e y.
Resultados
Un total de 7373 M. galloprovincialis fueron capturados en el total de los ocho ensayos realizados (Tabla 3.3).
Tabla 3.3 Capturas totales de M. galloprovincialis entre los años 2010 y 2015 en el total de los ensayos Nº Exp Localización Año Total Machos Hembras
Exp1 Jérica 2010 1147 516 631
Exp2 Altura 2011 1995 808 1187
Exp3 Mora 2010 1106 684 622
Exp4 Mora 2011 1671 773 898
Exp5 Valsaín 2011 402 - -
Exp6 Valsaín 2012 635 - -
Exp7 Valsaín 2013 357 - -
Exp8 Alcoi 2015 60 26 34
Influencia de la altitud en las capturas de M. galloprovincialis
No se observó ningún patrón claro en la distribución de las capturas en función ni de la orientación ni tampoco de la cobertura de coníferas en el área de muestreo de cada trampa. En las figuras del Anejo 3.2 puede observarse la frecuencia de capturas en función de las variables evaluadas (altitud, orientación y cobertura vegetal), así como el perfil de las curvas de vuelo para cada trampa (Anejo 3.3). No obstante, sí que pudo observarse cierto patrón en la distribución de las frecuencias de captura en función de la altitud (Figura 3.7). En el Anejo 3.4 puede observarse en concreto la distribución de las capturas en función de la altitud, representación a la cual se le ha añadido una línea de tendencia basada en un GLM de tipo logístico. En la tabla del Anejo 3.5 aparecen los efectos fijos del modelo mínimo ajustado.

60
Figura 3.7 Capturas de M. galloprovincialis por trampa en función de la altitud de muestreo para los experimentos desarrollados en la Sierra de Guardarrama (Exp5, Exp 6 y Exp7) durante el periodo de
2011 a 2013.
Las curvas de vuelo acumuladas para el año 2011, organizadas según el rango de altitud en el que se ubicaron, y ajustadas a un modelo de regresión logística binomial puede observarse en la Figura 3.8. En ella no son observables diferencias en las formas de las curvas a los diferentes rangos altitudinales. Los análisis realizados para las capturas de 2012 mostraron patrones muy semejantes pese a que la frecuencia de muestreo fue menor (Anejos 3.6, 3.7 y 3.8). La inclusión de la variable altitud en el modelo (Tabla 3.4) no mejora el ajuste (p=0.203), por lo que no hay influencia de la altitud en la forma de la curva de vuelo.
Figura 3.8 Capturas acumuladas de M. galloprovincialis organizadas según el rango de altitud de muestreo (considerando intervalos de 100 m) para el año 2011. Se representa también el ajuste de un modelo de regresión logística binomial a la
distribución de las capturas.
Tabla 3.4 Parámetros ajustados por una modelización lineal generalizada y estadísticos asociados.
Estimador SE z Pr(>|z|)
Intercepto -14.57 1.94 -7.5 <0.001

61
Fecha ordinal 0.067535 0.009 7.6 <0.001
El mismo análisis fue realizado teniendo en cuenta las capturas por trampa, sin estar éstas agrupadas por rangos altitudinales. Este análisis mostró patrones muy similares a los expuestos anteriormente, aunque más confusos en su interpretación (Anejo 3.9). Igualmente fueron analizadas las diferencias en las curvas de capturas acumuladas para las dos laderas del área de estudio (ladera sur de Cercedilla y ladera norte del Pinar de Valsaín) en el año 2011. De nuevo no pudo observarse diferencia alguna entre las curvas en función de la altitud, ni en función de la orientación de la ladera (Anejo 3.10). Los resultados concluyen que el vuelo de M. galloprovincialis no se da de forma diferente en función de la altitud, aunque el porcentaje de capturas acumuladas por fecha y altitud muestra que el periodo de tiempo en la que se éstas se producen se hace más corto según se asciende en altitud (Tabla 3.5).
Tabla 3.5 Porcentaje de capturas de M. galloprovincialis acumuladas para los diferentes rangos altitudinales
Altura (msnm) Fecha 1350 1450 1550 1650 1750 1850 2011-05-27 0.0 0.0 0.0 0.0 0.0 NA 2011-06-08 0.0 0.0 0.0 0.0 0.0 0.0 2011-06-16 0.0 0.0 0.0 0.0 0.0 0.0 2011-06-27 5.1 13.5 6.8 3.6 5.8 0.0 2011-07-04 5.1 17.3 10.3 8.0 13.4 12.5 2011-07-18 16.5 26.9 22.8 28.9 39.8 25.0 2011-07-28 35.7 46.2 32.1 48.8 61.7 37.5 2011-08-06 47.5 51.9 60.4 67.0 68.5 43.8 2011-08-17 58.9 65.4 66.0 82.0 79.8 68.8 2011-08-24 69.2 71.2 72.7 83.0 86.4 81.2 2011-09-01 81.0 73.1 75.0 87.8 90.2 93.8 2011-09-08 84.2 76.9 80.7 89.2 93.7 93.8 2011-09-20 92.9 88.5 85.2 95.0 98.8 100.0 2011-10-08 100.0 98.1 95.5 99.5 100.0 100.0 2011-10-21 100.0 100.0 100.0 100.0 100.0 100.0 2011-10-28 100.0 100.0 100.0 100.0 100.0 100.0
La representación gráfica de la duración del periodo de vuelo en función de los rangos altitudinales muestra una variación de forma análoga a la variación de la abundancia de M. galloprovincialis a lo largo del gradiente altitudinal estudiado en 2011 (Figura 3.9). De nuevo, el análisis de los datos de 2012 muestra patrones muy similares a los aquí comentados (Anejos 3.7 y 3.8)

62
Figura 3.9 Duración del periodo de vuelo de M. galloprovincialis en función de la altitud de muestreo para el año 2012 en la Sierra
de Guadarrama (Exp 6)
Influencia de las temperaturas en el inicio del periodo de vuelo de M. galloprovincialis
En primer lugar se procedió a la modelización de la variación de la temperatura con la altitud. Los modelos ajustados relacionaron un descenso de 0,7 grados cada 100 de ascensión (Anejo 3.1). En la Figura 3.10 puede observarse la evolución de las temperaturas con la altitud y las capturas obtenidas en las trampas anejas a los registradores de temperatura, en el año 2012.
Figura 3.10 Evolución de las temperaturas (máximas, medias y mínimas) de los meses de verano y otoño de 2012 para las
diferentes altitudes muestreadas. Las líneas discontinuas verticales marcan los días en los que se registraron las capturas, y las capturas correspondientes al tramo anterior están a la izquierda de tal barra
En la Figura 3.11 se muestran las capturas proporcionales en función de la temperatura media en la parte inicial del vuelo de M. galloprovincialis para el año 2012. Asimismo se observó un efecto significativo de la temperatura media en las capturas (F1 13 8,48 p=0.012), que no fue observado en el caso de contabilizar temperaturas máximas ni temperaturas mínimas.

63
Figura 3.11 Distribución de las capturas proporcionales de M. galloprovincialis en el inicio del periodo de vuelo en función de la
temperatura media en el año 2012 para el Exp6 de Sierra de Guadarrama (Segovia-Madrid). De ambas figuras puede deducirse que el vuelo de M. galloprovincialis se inicia en el mes de mayo-junio, cuando las temperaturas medias son superiores a los 14-15ºC.
Influencia de las altas temperaturas en el descenso de capturas estivales de M.
galloprovincialis
La Figura 3.12 muestra las capturas acumuladas en las diferentes zonas de estudio (Jérica 2010, Altura 2100, y Teruel 2010-2011) y en función de la temperatura. En todas ellas es posible observar una desaceleración en la acumulación de las capturas coincidiendo con las temperaturas medias más altas (generalmente a finales del mes de julio y agosto).
a b
c d

64
Figura 3.12 Capturas acumuladas de M. galloprovincialis para las diferentes zonas de estudio: a) Exp1. Jérica. 2010, b) Exp2. Altura. 2011, c) Exp 3. Mora. 2010, d) Exp 4. Mora. 2011. Las series temporales de capturas se han ajustado a un modelo aditivo
generalizado con el objetivo de suavizar la curva
La representación de las capturas diarias de M. galloprovincialis junto a la evolución temporal de las temperaturas medias permite observar esta tendencia de una forma más clara (Figura 3.13Figura 3.14, Figura 3.15 y Figura 3.16).
En el caso concreto de Jérica (Castellón), en el año 2010 (Figura 3.13) se observa una coincidencia entre la subida de las temperaturas a finales de julio (hasta la temperatura media máxima del periodo de vuelo) y un descenso en el número de capturas relativo, observable en la Figura 3.12 anterior como una desaceleración en la acumulación de capturas en ese periodo.
Figura 3.13 Distribución de las capturas diarias de M. galloprovincialis (arriba) y de las temperaturas medias del periodo de vuelo
(abajo) para el ensayo realizado en Jérica (Castellón) en el año 2010. Los tramos marcados en azul y en rojo representan incrementos y descensos significativamente diferentes de cero en los modelos ajustados
En el caso de Altura (Castellón), en 2011 (Figura 3.14), aunque de forma menos acentuada, la subida de las temperaturas medias de julio se corresponde con una ligera atenuación en las capturas.
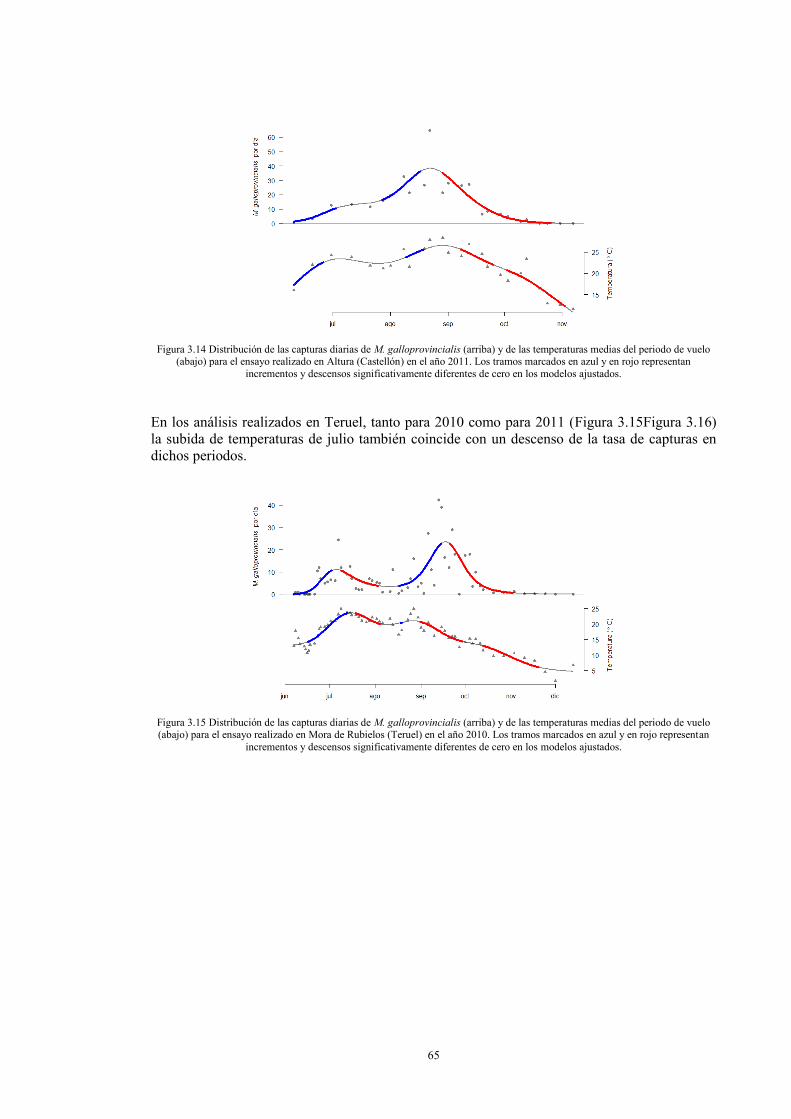
65
Figura 3.14 Distribución de las capturas diarias de M. galloprovincialis (arriba) y de las temperaturas medias del periodo de vuelo
(abajo) para el ensayo realizado en Altura (Castellón) en el año 2011. Los tramos marcados en azul y en rojo representan incrementos y descensos significativamente diferentes de cero en los modelos ajustados.
En los análisis realizados en Teruel, tanto para 2010 como para 2011 (Figura 3.15Figura 3.16) la subida de temperaturas de julio también coincide con un descenso de la tasa de capturas en dichos periodos.
Figura 3.15 Distribución de las capturas diarias de M. galloprovincialis (arriba) y de las temperaturas medias del periodo de vuelo (abajo) para el ensayo realizado en Mora de Rubielos (Teruel) en el año 2010. Los tramos marcados en azul y en rojo representan
incrementos y descensos significativamente diferentes de cero en los modelos ajustados.

66
Figura 3.16 Distribución de las capturas diarias de M. galloprovincialis (arriba) y de las temperaturas medias del periodo de vuelo (abajo) para el ensayo realizado en Mora de Rubielos (Teruel) en el año 2011. Los tramos marcados en azul y en rojo representan
incrementos y descensos significativamente diferentes de cero en los modelos ajustados.
Evolución de la sex-ratio durante el vuelo de M. galloprovincialis
En la Tabla 3.2 pueden observarse las capturas generales de M. galloprovincialis discriminadas por sexos. En la mayoría de los experimentos la cantidad total de hembras capturadas es mayor a la cantidad de machos. Sólo en el caso del ensayo realizado en Mora en 2010, la captura de machos es superior, aunque muy ligeramente. La proporción sexual entre machos y hembras (sex ratio) de M. galloprovincialis capturados, analizada a lo largo del periodo de muestreo, puede observarse en las Figura 3.17, Figura 3.18 y Figura 3.19. En tres de los experimentos pudo ajustarse significativamente un modelo lineal (Jérica 2010, Mora 2010 y Mora 2011) mientras que el experimento realizado en Altura en 2011 no muestra ningún patrón significativo. En general puede observarse un aumento significativo de la proporción de machos con respecto a la de hembras con el paso del tiempo; esto es, la proporción entre sexos es favorable a las hembras al inicio del periodo de vuelo, mientras que al final, se capturan más machos que hembras.
Figura 3.17 Evolución de la proporción de sexos de las capturas durante el periodo de vuelo de M. galloprovincialis en el
experimento realizado en Jérica (Castellón) ene l año 2010. La proporción de sexos ha sido calculada como el cociente del número de machos entre el número de hembras que se han capturado

67
Figura 3.18 Evolución de la proporción de sexos de las capturas durante el periodo de vuelo de M. galloprovincialis en el
experimento realizado en Mora de Rubielos (Teruel) en el año 2010. La proporción de sexos ha sido calculada como el cociente del número de machos entre el número de hembras que se han capturado. Los datos correspondientes a bajas capturas no han sido
tomados en cuenta durante la modelización.
Figura 3.19 Evolución de la proporción de sexos de las capturas durante el periodo de vuelo de M. galloprovincialis en el
experimento realizado en Mora de Rubielos (Teruel) en el año 2011. La proporción de sexos ha sido calculada como el cociente del número de machos entre el número de hembras que se han capturado.
Vuelos circadianos de M. galloprovincialis
La Figura 3.20 muestra la distribución de las capturas a lo largo de ciclos circadianos de 24 h. Puede observarse que todas las capturas se obtuvieron en horario diurno y que el máximo se dio en las horas centrales del día, entre las 13 h y las 17 h.

68
Figura 3.20 Histograma de la media de las capturas horarias de M. galloprovincialis en
2015
En la Figura 3 21 puede observarse la evolución de las temperaturas a lo largo del día durante el periodo de muestreo, mostrando máximos de temperaturas medias entre las 12h y las 18h.
Figura 3 21 Evolución de la media horaria de las temperaturas tomadas con intervalos de
5 minutos en el periodo de muestreo
Por último, en la Figura 3.22 se representa la frecuencia de capturas en función de la hora y de la temperatura. La distribución de las capturas se analizó en función de la hora de captura, y la temperatura media registrada durante la hora anterior.

69
Figura 3.22 Frecuencia de capturas en función de la hora del día y de la temperatura
media horaria
En resumen, los resultados muestran que los insectos se capturaron predominantemente durante las horas centrales del día, momento en el que se dan las temperaturas máximas diarias (Figura 3.22).

70
Discusión
La incidencia de la temperatura en la biología de M. galloprovincialis ha sido poco estudiada hasta la fecha. Algunos estudios evalúan su efecto sobre el desarrollo larvario (KOUTROUMPA et al. 2008, NAVES Y DE SOUSA 2009) y recientemente también se ha estudiado su influencia sobre la capacidad de dispersión del insecto, aunque sólo en el caso concreto de los Pirineos como barrera a la propagación de la enfermedad (HARAN et al. 2015). De igual manera, aunque es conocida la amplia distribución global de M. galloprovincialis, su distribución altitudinal tampoco ha sido estudiada en profundidad (HARAN et al. 2015). Nuestros resultados muestran, en primer lugar, que efectivamente existe una influencia de una variable sintética como la altitud en la población de M. galloprovincialis, registrada ésta como frecuencia de capturas en trampas cebadas con atrayentes. Sin embargo la altitud se halla correlacionada con otras variables de alta significación biológica como la temperatura, humedad, o la irradiación. El modelo representado en la Figura 3.7 sugiere que a partir de 1350 msnm la captura de estos insectos decrece, y que en este decrecimiento no hay efecto ni de la orientación de la ladera ni de la cobertura de pinar en esa zona, sino esencialmente los parámetros correlacionados con la altitud (Anejo 3.2). Por otro lado, si bien el transecto estudiado recoge una variabilidad suficiente para el estudio de la influencia de la altitud en la abundancia de M. galloprovincialis, es posible que otras variables locales así como las condiciones microclimáticas de cada punto de muestro hayan incidido en el nivel de capturas. Sin embargo la comparación de las capturas a lo largo de los tres años de estudio muestra un patrón similar para los niveles de capturas en las diferentes alturas estudiadas (Figura 3.7). Estos resultados son coherentes, en términos generales, con el reciente trabajo de (HARAN et al. 2015) en la cordillera de los Pirineos. En él, no obstante, se plantean los 1590 msnm como umbral de máxima altitud para la supervivencia de M. galloprovincialis, puesto que, a esta altitud, se dan medias de temperaturas mínimas inferiores a -7ºC y observan capturas muy bajas (10 individuos en más de 3 meses de muestreo). Estas frecuencias de capturas son coherentes con los modelos obtenidos en nuestros ensayos a una mayor altitud, superior a los 1700 m (Figura 3.7). Pero no lo son tanto a los 1600 m, donde las capturas obtenidas en Valsaín son algo mayores que las citadas por HARAN et al. (2015) para Pirineos. Además, en este mismo sentido, nuestros resultados apuntan a que existe una alta probabilidad de encontrar M. galloprovincialis a más de 1900 msnm (presumiblemente hasta el límite altitudinal al cual es posible encontrar P. sylvestris), si bien es cierto que esta extrapolación debe ser tomada con cautela ya que las altitudes superiores a los 1900 msnm quedan fuera del rango de la modelización (Figura 3.10). De cualquier forma la influencia de la latitud puede permitir esos desfases altitudinales, recordando siempre que la altitud en sí misma no es un parámetro de significación biológica, sino sus parámetros correlacionados. En cualquier caso, estas diferencias entre las capturas observadas en Pirineos y en el Sistema Central podrían tener que ver con la propia geomorfología de ambas cadenas, y en concreto con la altitud y con el porcentaje de superficie existente por encima del límite de 1590 msnm citado por HARAN et al. (2015) en ambas cordilleras, ya que el Sistema Central es una cadena montañosa con una altitud general considerablemente menor que la de los Pirineos (2592 m en Sistema Central frente a 3404 m en Pirineos). Además, según HARAN et al. (2015) , si bien la parte central de Los Pirineos (zona donde se acumulan las mayores altitudes) podría constituir cierta barrera natural a la dispersión del insecto, la presencia de corredores biológicos a ambos flacos de la cadena montañosa hace permeable dicha barrera y que en consecuencia podría no afectar a la expansión del NMP hacia el norte de Europa. Consecuentemente, en el caso del Sistema Central, la presencia de corredores biológicos con material hospedante es mucho mayor debido a que es muy poca, y muy inconexa, la superficie que está por encima de los 1590

71
msnm en dicho sistema montañoso. Por ello parece razonable que el campeo de M. galloprovincialis por las zonas más elevadas del Sistema central sea mayor que el de Pirineos. Por otro lado, este efecto de la altitud sobre la frecuencia de capturas, no se manifiesta en el perfil temporal de las mismas, es decir, en la forma de las curvas de vuelo. Nuestros resultados, de hecho, muestran patrones o formas de curva de vuelo muy similares a las diferentes altitudes estudiadas (Figura 3.8, Anejo 3.3). Y no obstante sí que es posible detectar una influencia de la altitud en la duración de dichos periodos de vuelo, de forma que a mayores altitudes la duración del periodo de vuelo tiende a ser más corta, y este acortamiento se da por ambos flancos de la curva: un inicio de vuelo más tardío y una finalización del mismo más temprana (Tabla 3.5, Figura 3.9). El inicio del periodo de vuelo, tratándose de una especie que atraviesa el periodo invernal en estado larvario, está influido necesariamente por la ocurrencia de la emergencia y por las variables ambientales que la determinan. En el Capítulo 2 de esta memoria se muestra que, en términos generales, las curvas de emergencia de las zonas más frías de la península ibérica son más tardías que las de las zonas más cálidas, lo cual es coherente con un inicio del periodo de vuelo también más tardío en las zonas más frías (y en consecuencia en las zonas más altas). No obstante, en este aspecto, debe tenerse en consideración que la técnica utilizada para la monitorización de las curvas de vuelo (trampas cebadas con atrayentes feromonales y cairomonales) no parece ser eficaz hasta que el adulto de M. galloprovincialis ha completado su maduración sexual (ver también el Capítulo 5; PAJARES et al. 2010, SANCHEZ-HUSILLOS et al. 2015). Para alcanzarla, los adultos recién emergidos necesitan alimentarse, fundamentalmente de corteza tierna de ramillos de pino, durante un periodo de tiempo determinado, que diversos autores citan siempre menor a los 14 días (HELLRIGL 1971, FRANCARDI Y PENNACCHIO 1996; SÁNCHEZ-HUSILLOS et al. Comm. pers., NAVES et al. 2008, DAVID et al. 2014). Estas observaciones son consistentes con las descritas en el apartado 1 del Capítulo 5 de este trabajo, en el que, durante un ensayo realizado en una masa muy pequeña y aislada (<10 ha), se obtuvo una muy alta tasa de recapturas (aproximadamente el 36,7% de los insectos liberados), las más tempranas tan sólo 8 días posteriores a la liberación de los insectos inmaduros. Por lo tanto, para una correcta monitorización del periodo de vuelo mediante trampas cebadas debe ser tenido en cuenta este lapso de tiempo en el cual los adultos de M. galloprovincialis ya han emergido pero son sexualmente inmaduros y, por tanto, no susceptibles de ser capturados por las trampas. En esta cuestión, y en definitiva, nuestros resultados, muestran que el inicio del periodo de vuelo está correlacionado con los primeros periodos en los meses de mayo o junio durante los cuales la media de las temperaturas supera los 14-15ºC (Figura 3.10Figura 3.11). Estos resultados son consistentes con los mostrados por HOCH et al. (2015) para la región de Baden en Austria, con inicios del periodo de vuelo exactamente a las mismas temperaturas. Dicho estudio es el único en el que se ha realizado, hasta la fecha, un análisis de la influencia de la temperatura en la curva de vuelo de M. galloprovincialis. El resto de publicaciones al respecto, se limitan a análisis descriptivos de los periodos en Portugal (NAVES et al. 2008) y España (MAGRAMA 2015b). Todos estos estudios coinciden en que el periodo de vuelo se extiende aproximadamente, y de manera continua, desde mediados de mayo hasta finales de octubre o principios de noviembre, siendo el inicio del vuelo más temprano en el Sur de Europa (España, Portugal continental y en Madeira) que en Europa Central (Austria). No obstante, es posible detectar algunas discordancias entre las características de las diferentes curvas publicadas:

72
NAVES y DE SOUSA (2009) indicaron que el máximo de las capturas se daba entre los meses de junio y julio. Sin embargo, el patrón de las capturas difirió significativamente entre los años estudiados para una misma localización, lo cual hace pensar en que pueda existir algún tipo de incidencia de alguna variable medioambiental en la forma de dichas curvas de vuelo (en este caso, especialmente, de la temperatura). Además, este estudio se realizó con anterioridad al descubrimiento y comercialización del atrayente para M. galloprovincialis por lo que el cebo utilizado para su captura es diferente al utilizado en el resto de estudios mencionados, y no pueden ser comparados con el rigor necesario. Aun así, estos resultados de máximos de capturas en junio-julio son coincidentes con los mostrados por (HOCH et al. 2015) para las regiones de Austria. En el caso centroeuropeo las curvas de vuelo mostraron perfiles unimodales. Las curvas de vuelo analizadas en los experimentos realizados en la Sierra de Guadarrama para la evaluación de la influencia de la altitud en las capturas (en el primer apartado de este mismo capítulo) también muestran perfiles netamente unimodales (Anejo 3.3). En este caso conviene tener en cuenta que el ensayo es realizado en ecosistemas de montaña, por encima de los 1350 msnm y hasta los 1950 msnm en el Sistema Central, en los que el régimen de temperaturas es notablemente más frío que en el resto de zonas estudiadas en la península Ibérica. Por contraposición, en los últimos años, algunos documentos técnicos provenientes de los trabajos de monitorización del M. galloprovincialis para la gestión del NMP en España y Portugal, han mostrado perfiles bimodales en las curvas de vuelo de la mayor parte de la península Ibérica. La forma de estas curvas es muy similar a la que muestran los resultados de esta memoria: con un máximo relativo de capturas a finales del mes de junio y principios de julio, un máximo absoluto de capturas a finales del mes de agosto y septiembre (Figura 3.13, Figura 3.14, Figura 3.15 y Figura 3.16). Respecto al análisis de la influencia de la temperatura en estos descensos observados en las capturas estivales, conviene tener en cuenta que, en términos generales, la temperatura del aire afecta tanto a la densidad de insectos como a su actividad (en este caso, a su actividad de vuelo). Una asunción básica en este tipo de análisis suele radicar en que, una vez alcanzado el umbral inferior de temperaturas a partir del cual el insecto puede generar suficiente energía para mantener el vuelo, la ocurrencia de éste es relativamente independiente de la temperatura. Sin embargo, esto parece estar más bien relacionado con la capacidad de vuelo que no con la ‘apetencia’ por el mismo, cuando ambas son necesarias y no suficientes para que tenga lugar el vuelo del insecto (TAYLOR 1963). Definitivamente, los umbrales inferiores de temperatura son, en climas templados, un factor climático determinante en el vuelo de los insectos, que además puede ser medido mediante capturas en trampas. No obstante, tampoco existe duda de los que límites superiores de temperatura para el vuelo de los insectos existan, aunque su determinación reviste mayor dificultad y éstos actúan con menor frecuencia que los primeros (TAYLOR 1963, LOGAN et al. 2006). En este sentido, nuestros análisis mostraron que las altas temperaturas podrían estar relacionadas con el descenso en las capturas estivales, lo cual se fundamente precisamente en la hipótesis clásica de que la actividad (el vuelo) del insectos (M. galloprovincialis en este caso) se incrementaría gradualmente con el aumento de la temperatura hasta alcanzar un óptimo, a partir del cual, la subida de las temperaturas causaría un descenso relativamente abrupto en la actividad de los insectos. Esta hipótesis que ha sido demostrada tanto en campo como en laboratorio, mostrando que el número de vuelos por minuto sigue el mismo patrón descrito para una gran cantidad de grupos taxonómicos (TAYLOR 1963, GILBERT Y RAWORTH 1996, LOGAN Y POWELL 2003).

73
Y en efecto, en nuestros resultados este rasgo se observa de manera reiteradamente en los diferentes años y áreas de estudio. En todos los casos analizados es posible observar una desaceleración en la acumulación de las capturas coincidiendo con los valores máximos de las temperaturas medias estivales, las cuales podrían funcionar como límite superior de temperatura para la ‘apetencia’ por el vuelo (Figura 3.12). Quedaría por identificar los valores de temperatura de ese umbral en futuras investigaciones. Desde este enfoque, como ya ha sido comentado, puede explicarse la bimodalidad observada en los resultados. Puesto que los periodos de vuelo suelen extenderse a lo largo de varios meses, generalmente entre mayo y agosto, en el momento de las temperaturas máximas estivales, aún quedarían insectos no emergidos e inmaduros que no se capturarían en las trampas. Este reclutamiento de individuos, susceptibles de ser capturados una vez restablecidas las condiciones climáticas óptimas para el vuelo tras el periodo de temperaturas máximas, podría ser responsable de que el segundo pico de capturas a finales de agosto y septiembre fuese especialmente acusado. En otras palabras, la posible inhibición del vuelo de los adultos en un momento en el que no todos ellos son susceptibles de ser capturados (sino que están en proceso de serlo), podría explicar que el especial aumento de las capturas posterior se debiese, según TAYLOR (1963), tanto al aumento de la actividad de vuelo (propia del aumento de la temperatura), como al aumento puntual de los individuos reclutados susceptibles de ser atraídos por las trampas. La unimodalidad en las curvas observada en los trabajos centroeuropeos (HOCH et al. 2015) y en las cotas más elevadas del Sistema Central de la península ibérica, se explicaría fundamentalmente porque las temperaturas estivales en esas regiones no alcanzan las temperaturas umbrales que podrían inhibir el vuelo de los adultos, por lo que no se produciría el descenso estival en la tasa de capturas observable en el resto de la península. Por tanto, dichas curvas serían unimodales y adaptadas simplemente a un desfase con respecto a las curvas de emergencia de duración aproximadamente igual a lapso de tiempo medio necesario para que se produzca la maduración sexual de los adultos de M. galloprovincialis que posibilite su captura en las trampas. Por último, el análisis de la evolución de la proporción sexual de las capturas a lo largo del periodo de vuelo, también parece ser coherente con la hipótesis mencionada. La feromona específica del género Monochamus, 2-undecyloxy-1-ethanol, es emitida por los machos de M. galloprovincialis y funciona como una feromona sexual y agregativa ya que es capaz de atraer a ambos sexos (PAJARES et al. 2010). Aun así, la gran mayoría de muestreos realizados con este atrayente constatan una proporción sexual de las capturas sesgada a favor de las hembras, es decir, éstas que son capturadas generalmente en mayor cantidad que los machos (PAJARES et al. 2010, HERNÁNDEZ et al. 2011, PAJARES et al. 2013, ETXEBESTE et al. 2015, SANCHEZ-HUSILLOS et al. 2015). También nuestros resultados son coincidentes en este sentido (Tabla 3.2). Pero a pesar de ello, el análisis de la evolución de la proporción de sexos de las capturas a lo largo del periodo de vuelo muestra un aumento significativo de la proporción de machos con respecto a la de hembras con el paso del tiempo, es más, la proporción entre sexos es favorable a las hembras al inicio del periodo de vuelo (se capturan más hembras que machos) mientras que es favorable a los machos al final del mismo (Figura 3.17, Figura 3.18 y Figura 3.19). Puesto que la proporción sexual de las emergencias no muestra este sesgo, sino que se mantiene en una proporción 1:1 incluso ligeramente favorable a los machos al inicio de la misma (Capítulo 2), y puesto que la longevidad no es mayor en machos que en hembras (Etxebeste Com. Pers.), parece razonable pensar que la respuesta de los machos a la emisión de feromonas

74
por parte de otros machos aumenta con el paso del tiempo, o por el contrario, la capacidad de respuesta de las hembras a dicha feromona la que disminuye. El primer caso podría estar relacionado con un mecanismo de compensación (trade-off) entre la necesidad reproductiva y el final de la vida del individuo, habitual en otras especies (LATTY Y REID 2009, DE LOOF 2011), y según el cual, conforme se aproxima el final del periodo de vida de algunos especímenes, éstos optimizan sus estrategias para conseguir el máximo número de hembras fecundadas. Es decir, a una mayor edad existiría un mayor estrés del individuo, lo cual derivaría en una apuesta por la progenie por encima de otros factores, en este caso, optimizando la detección de feromonas emitidas por otros machos, las cuales actuarían como señal de una mayor probabilidad de presencia de hembras en ese punto (PURESWARAN et al. 2006, LATTY Y REID 2009). Y en el marco de la bimodalidad del periodo de vuelo, este aumento de la respuesta de los machos longevos a las feromonas de otros machos podría suponer, en tal caso, un nuevo aporte de individuos extra en la parte final del periodo de vuelo, lo cual también estaría en consonancia con la aparición del acusado segundo pico que puede ser observado en nuestros resultados. En el segundo caso, y de forma armónica con lo expuesto, también sería razonable pensar que, con el paso de la estación, y una vez que las hembras fuesen fecundadas, ovopositasen y fuesen cumpliendo su ciclo, su urgencia por la fecundación disminuyese y, consecuentemente lo hiciese también su atracción por la feromona emitida por los machos. En cualquier caso, nuevos estudios deberían ser encaminados en este sentido con el objetivo de comprender con mayor profundidad la fenología del vuelo, y tomando en consideración los aspectos recién mencionados en nuestras hipótesis. Por último, también debe ser tenido en cuenta que, para el análisis del efecto de la temperatura en el vuelo de los insectos, en general, el estudio del tiempo que éstos pasan a temperaturas comprendidas entre determinados umbrales debería tener más efecto sobre las capturas diarias que la media de las temperaturas (TAYLOR 1963) , para lo cual, resulta determinante conocer los periodos circadianos del vuelo de M. galloprovincialis. Aunque sus hábitos de vuelo han sido considerados crepusculares en algunos estudios (BONIFÁCIO 2009) o diurnos en otros (BERGER 2012), hasta donde conocemos es la primera vez que se ha realizado un estudio con la profundidad y la frecuencia de muestreo (horaria) del que se presenta en este trabajo. Nuestros resultados muestran que el periodo circadiano del vuelo de M. galloprovincialis en el momento del muestreo (finales de agosto y principios de septiembre) es diurno y que las capturas aumentan con el aumento de la temperatura, produciéndose máximos de capturas en las horas más calurosas del día (Figura 3.20). El periodo de tiempo estudiado se ha ceñido exclusivamente al principio del mes de septiembre, momento en el que era esperable el máximo de capturas. En este periodo, aunque puede detectarse la existencia de un umbral inferior de temperatura que aparentemente inhibe el vuelo del insecto (Figura 3 21 y Figura 3.22), no parece observarse el efecto de un umbral superior en las capturas, como sería esperable en virtud de lo argumentado sobre la bimodalidad de las curvas de vuelo líneas arriba. El régimen de temperaturas del periodo de muestreo (finales de agosto y principios de septiembre) no es el más caluroso en el área de estudio. Y este ensayo, por lo tanto, debería completarse con el análisis de los vuelos circadianos en los periodos de máximas temperaturas, así como en el momento del inicio del periodo de vuelo, con el objeto de poder completar la evaluación de la influencia de las tres variables que fundamentalmente podrían influir en el vuelo circadiano del insecto: irradiancia (o periodo horario), temperatura y época del año. Las capturas obtenidas al inicio y al final del periodo diurno, 9h y 22h (Figura 3.22), aunque escasas, aparecen vinculadas a temperaturas suficientemente altas (superiores a los 18ºC), por lo

75
que en periodos de temperaturas medias más elevadas a lo largo de todo el día, sería esperable que las capturas se diesen en un rango horario más ancho, y no especialmente ceñidas a las horas centrales del día. En estos periodos calurosos del verano, incluso podría darse, en estas horas centrales, temperaturas máximas que sobrepasasen un umbral por encima del cual se inhibiese el vuelo del insecto, provocando una bimodalidad circadiana o que su vuelo quedase fundamentalmente retrasado a la parte final del día (momento de descenso de la temperatura), lo cual explicaría las observaciones crepusculares de BONIFÁCIO (2009). En cualquier caso y en resumen, nuestros resultados indican que, durante el periodo de máximas capturas de su curva de vuelo, M. galloprovincialis tiene un vuelo diurno ajustado a las horas más calurosas del día, entre las 13h y las 17h.

76
Bibliografía
ÁLVAREZ, G., I. ETXEBESTE, D. GALLEGO, G. DAVID, L. BONIFACIO, H. JACTEL, E.
SOUSA y J. A. PAJARES (2015). Optimization of traps for live trapping of Pine Wood Nematode vector Monochamus galloprovincialis. Journal of Applied Entomology 139(8): 618-626.
BERGER, P. (2012). Coléoptères Cerambycidae de la faune de France continentale et de Corse: actualisation de l'ouvrage d'André Villiers, 1978, ARE.
BONIFÁCIO, L. F. P. (2009). Impacte e evolução da doença da murchidão dos pinheiros (pine wilt disease) na zona afectada a sul do Rio Tejo.
DAS FLORESTAS, D. D. G. (2001). Inventário Florestal Nacional. 3ª Revisão. Direcção Geral de Florestas, Lisboa.
DAVID, G., B. GIFFARD, D. PIOU y H. JACTEL (2014). Dispersal capacity of Monochamus galloprovincialis, the European vector of the pine wood nematode, on flight mills. Journal of Applied Entomology 138(8): 566-576.
DE LOOF, A. (2011). Longevity and aging in insects: Is reproduction costly; cheap; beneficial or irrelevant? A critical evaluation of the “trade-off” concept. Journal of Insect Physiology 57(1): 1-11.
ETXEBESTE, I., E. SANCHEZ-HUSILLOS, G. ÁLVAREZ, H. MAS I GISBERT y J. PAJARES (2015). Dispersal of Monochamus galloprovincialis (Col.: Cerambycidae) as recorded by mark-release recapture using pheromone traps. Journal of Applied Entomology: In press.
FRANCARDI, V. y F. PENNACCHIO (1996). Note sulla bioecologia di Monochamus galloprovincialis galloprovincialis (Olivier) in Toscana e in Liguria (Coleoptera Cerambycidae). Redia 79: 153-169.
GALLINAT, A. S., R. B. PRIMACK y D. L. WAGNER (2015). Autumn, the neglected season in climate change research. Trends in ecology & evolution 30(3): 169-176.
GILBERT, N. y D. A. RAWORTH (1996). Insects and temperature - A general theory. Canadian Entomologist 128(1): 1-13.
GILLOOLY, J. F., E. L. CHARNOV, G. B. WEST, V. M. SAVAGE y J. H. BROWN (2002). Effects of size and temperature on developmental time. Nature 417(6884): 70-73.
HARAN, J., A. ROQUES, A. BERNARD, C. ROBINET y G. ROUX (2015). Altitudinal Barrier to the Spread of an Invasive Species: Could the Pyrenean Chain Slow the Natural Spread of the Pinewood Nematode? PLoS ONE 10(7): e0134126.
HELLRIGL, K. G. (1971). Die Bionomie der europaischen Monochamus-Arten (Coleopt., Cerambycid.) und ihre Bedeutung fur die Forstund Holzwirtschaft. Redia 52: 367-509.
HERNÁNDEZ ALONSO, R., V. PÉREZ FORTEA, G. SÁNCHEZ PEÑA, J. CASTELLÁ SOLÁ, J. PALENCIA ADRUBAU, J. M. GIL BONO y A. ORTIZ SÁNCHEZ (2007). Ensayos de trampeo de escolítidos perforadores subcorticales en pinares mediante el uso de feromonas 2002-2005. Ecología(21): 43-55.
HERNÁNDEZ, R., A. ORTIZ, V. PÉREZ, J. GIL y G. SÁNCHEZ (2011). " Monochamus galloprovinciales"(Olivier, 1795)(Coleoptera: Cerambycidae), comportamiento y distancias de vuelo. Boletín de sanidad vegetal. Plagas 37(1): 79-96.
HOCH, G., D. MITTERMAYR y H. KREHAN (2015). Monochamus-Arten als potenzielle Vektoren des Kiefernholznematoden in Österreich: Lockstofffallen zum Monitoring des Fluges. Forstschutz Aktuell 60(61): 61.
IBEAS, F., D. GALLEGO, J. DIEZ y J. PAJARES (2007). An operative kairomonal lure for managing pine sawyer beetle Monochamus galloprovincialis (Coleoptera: Cerymbycidae). Journal of Applied Entomology 131(1): 13-20.
INÁCIO, M. L., F. NÓBREGA, P. VIEIRA, L. BONIFÁCIO, P. NAVES, E. SOUSA y M. MOTA (2015). First detection of Bursaphelenchus xylophilus associated with Pinus nigra in Portugal and in Europe. Forest Pathology 45(3): 235-238.

77
KOUTROUMPA, F. A., B. VINCENT, G. ROUX-MORABITO, C. MARTIN y F. LIEUTIER (2008). Fecundity and larval development of Monochamus galloprovincialis (Coleoptera Cerambycidae) in experimental breeding. Annals of Forest Science 65(7).
LATTY, T. M. y M. L. REID (2009). First in line or first in time? Effects of settlement order and arrival date on reproduction in a group-living beetle Dendroctonus ponderosae. Journal of Animal Ecology 78(3): 549-555.
LOGAN, J. A. y J. A. POWELL (2003). Modelling Mountain Pine Beetle Phenological Response to Temperature. Mountain Pine Beetle Symposium: Challenges and Solutions. October 30-31, 2003, .
T.L. Shore, J.E. Brooks, and J.E. Stone (editors). e,, Victoria, BC. 298 p. T. L. Shore, J. E. Brooks y J. E. Stone. Kelowna, British Columbia, Natural Resources Canada, Canadian Forest Service, Pacific Forestry Centre, Victoria, BC: 210-222.
LOGAN, J. D., W. WOLESENSKY y A. JOERN (2006). Temperature-dependent phenology and predation in arthropod systems. Ecological Modelling 196(3-4): 471-482.
MACIAS-SAMANO, J. E., D. WAKARCHUK, J. G. MILLAR y L. M. HANKS (2012). 2-Undecyloxy-1-ethanol in combination with other semiochemicals attracts three Monochamus species (Coleoptera: Cerambycidae) in British Columbia, Canada. The Canadian Entomologist 144(06): 764-768.
MAGRAMA. (2015a). "Nematodo de la madera del pino." 2015, de http://www.magrama.gob.es/es/agricultura/temas/sanidad-vegetal/nematodo-de-la-madera-del-pino/#.
MAGRAMA (2015b). Trabajos de seguimiento y control del vector del nematodo de la madera del pino. Ministerio de Agricultura, Pesca y Alimentación. Internal report: 32 pages.
NAVES, P. y E. DE SOUSA (2009). Threshold temperatures and degree-day estimates for development of post-dormancy larvae of Monochamus galloprovincialis (Coleoptera: Cerambycidae). Journal of Pest Science 82(1): 1-6.
NAVES, P. M., E. M. DE SOUSA y J. A. QUARTAU (2006). Feeding and oviposition preferences of Monochamus galloprovincialis for certain conifers under laboratory conditions. Entomologia Experimentalis Et Applicata 120(2): 99-104.
NAVES, P. M., E. SOUSA y J. M. RODRIGUES (2008). Biology of Monochamus galloprovincialis (Coleoptera: Cerambycidae) in the pine wilt disease affected zone, Southern Portugal. Silva Lusitana 16(2): 133-148.
NUNES DA SILVA, M., A. SOLLA, L. SAMPEDRO, R. ZAS y M. W. VASCONCELOS (2015). Susceptibility to the pinewood nematode (PWN) of four pine species involved in potential range expansion across Europe. Tree Physiology.
PAJARES, J. A., G. ÁLVAREZ, D. R. HALL, P. DOUGLAS, F. CENTENO, N. IBARRA, M. SCHROEDER, S. A. TEALE, Z. WANG y S. YAN (2013). 2‐(Undecyloxy)‐ethanol is a major component of the male‐produced aggregation pheromone of Monochamus sutor. Entomologia experimentalis et applicata 149(2): 118-127.
PAJARES, J. A., G. ALVAREZ, F. IBEAS, D. GALLEGO, D. R. HALL y D. I. FARMAN (2010). Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. Journal of Chemical Ecology 36(6): 570-583.
PAJARES, J. A., F. IBEAS, J. J. DIEZ y D. GALLEGO (2004). Attractive responses by Monochamus galloprovincialis (Col., Cerambycidae) to host and bark beetle semiochemicals. Journal of Applied Entomology 128(9-10): 633-638.
PÉREZ, V., R. HERNÁNDEZ, J. CASTELLÁ, G. SÁNCHEZ y J. PALENCIA (2004). Ensayos de atracción y captura de Ips acuminatus (Coleoptera: Scolytidae). Ecología(18): 35-46.
PURESWARAN, D., B. SULLIVAN y M. AYRES (2006). Fitness consequences of pheromone production and host selection strategies in a tree-killing bark beetle (Coleoptera: Curculionidae: Scolytinae). Oecologia 148(4): 720-728.

78
R CORE TEAM (2015). R: A language and environment for statistical computing. Vienna, Austria, R Foundation for Statistical Computing.
RICHARDSON, D. M., P. W. RUNDEL y D. RICHARDSON (1998). Ecology and biogeography of Pinus: an introduction. Ecology and biogeography of Pinus: 3-46.
ROBERTS, D. W. y S. V. COOPER (1989). Concepts and techniques of vegetation mapping. General technical report INT-US Department of Agriculture, Forest Service, Intermountain Research Station (USA).
RODRIGUES, J. M. (Com. Pers.). SANCHEZ-HUSILLOS, E., G. ÁLVAREZ-BAZ, I. ETXEBESTE y J. A. PAJARES (2013).
Shoot feeding, oviposition, and development of Monochamus galloprovincialis on Pinus pinea relative to other pine species. Entomologia Experimentalis Et Applicata: 1-10.
SANCHEZ-HUSILLOS, E., I. ETXEBESTE y J. PAJARES (2015). Effectiveness of mass trapping in the reduction of Monochamus galloprovincialis Olivier (Col.: Cerambycidae) populations. Journal of Applied Entomology.
TAYLOR, L. (1963). Analysis of the effect of temperature on insects in flight. The Journal of Animal Ecology: 99-117.
TOMMINEN, J. (1993). Development of Monochamus galloprovincialis Olivier (Coleoptera, Cerambycidae) in Cut Trees of Young Pines (Pinus sylvestris L.) and Log Bolts in Southern Finland. Entomologica Fennica 4(3): 137-142.
TORRES-VILA, L. M., C. ZUGASTI, J. M. DE-JUAN, M. J. OLIVA, C. MONTERO, F. J. MENDIOLA, Y. CONEJO, Á. SÁNCHEZ, F. FERNÁNDEZ, F. PONCE y G. ESPÁRRAGO (2015). Mark-recapture of Monochamus galloprovincialis with semiochemical-baited traps: population density, attraction distance, flight behaviour and mass trapping efficiency. Forestry 88(2): 224-236.
VICENTE, C., M. ESPADA, P. VIEIRA y M. MOTA (2012). Pine Wilt Disease: a threat to European forestry. European Journal of Plant Pathology 133(1): 89-99.
ZAMORA, P., V. RODRÍGUEZ, F. RENEDO, A. V. SANZ, J. C. DOMÍNGUEZ, G. PÉREZ-ESCOLAR, J. MIRANDA, B. ÁLVAREZ, A. GONZÁLEZ-CASAS, E. MAYOR, M. DUEÑAS, A. MIRAVALLES, A. NAVAS, L. ROBERTSON y A. B. MARTÍN (2015). First Report of Bursaphelenchus xylophilus Causing Pine Wilt Disease on Pinus radiata in Spain. Plant Disease: PDIS-03-15-0252-PDN.

79
Capítulo 4
Comunidad de coleópteros saproxílicos asociada a Monochamus galloprovincialis (Col.: Cerambycidae) en el levante de la
Península Ibérica
Antecedentes
A principios de la década de 1980 se despertó una preocupación general ante el continuo y progresivo deterioro del estado de salud de los bosques que, de manera generalizada, se registraba en el viejo continente. Esta preocupación comenzó en Europa Central ante los episodios de lluvia ácida producida por contaminación transnacional a larga distancia, originada por la combustión masiva de carbón con alto nivel de azufre. Como respuesta a la creciente inquietud en 1985 se establece el Programa de Cooperación Internacional para la Evaluación y Seguimiento de los Efectos de la Contaminación Atmosférica en los Bosques (ICP-FOREST 2011). En este marco, en 1986 se publica un reglamento comunitario sobre protección de los bosques contra los efectos de la contaminación atmosférica que pone en marcha, de forma coordinada, las acciones de seguimiento en todos los países comunitarios. Un año más tarde, queda constituida la Red Europea de Seguimiento de Daños en los Bosques (Red CE de Nivel I), cuyo objetivo es estudiar a gran escala, una serie de parámetros de sencilla medición, sobre las condiciones ecológicas de la estación y la vitalidad del arbolado. Desde 1987 se realizan muestreos sistemáticos con periodicidad anual para la evaluación del estado de salud de los bosques, y que abarcan de manera representativa, el total de la superficie forestal de la UE. No obstante, pese a la alta utilidad de esta red en la detección de las zonas decadentes o dañadas y el seguimiento de su evolución, se constata que no es posible establecer, a partir de la información que proporciona, el papel del clima y la contaminación, en el declive de los bosques europeos (ICP-FOREST 2011). Para dar respuesta a esta carencia, en 1994 se publica en España el Reglamento CEE/1091, en el que se detalla la metodología adecuada para una nueva red de daños (Red CE de Nivel II), que queda definida como “Red de Parcelas para el Seguimiento Continuo e Intensivo de los Ecosistemas Forestales”; El objetivo de esta otra red es adquirir información sobre la evolución actual y sobre la relación entre las características de la estación, los factores de estrés y el estado fitosanitario de los bosques a escala nacional y europea (Reglamento CE 1091/94). Para ello se protocolizan mediciones más intensivas, precisas y descriptivas que en la Red de Nivel I, sobre por ejemplo suelos, producción foliar, crecimiento arbóreo, deposiciones atmosféricas o condiciones meteorológicas de las parcelas. En 2003 entra en vigor el Reglamento Europeo Forest Focus, bajo cuya estructura se amparan las redes y las actividades en ellas realizadas. Con este reglamento se incrementa el número de

80
objetivos de las redes, incluyendo el seguimiento de los efectos del cambio climático, de la biodiversidad forestal y de la gestión sostenible de los sistemas forestales. El seguimiento del estado y de la evolución de los bosques que se ha realizado desde entonces, incluye evaluaciones con arreglo a métodos normalizados, y ha creado una importante plataforma para el intercambio de conocimientos sobre el estado de salud del arbolado en los sistemas forestales europeos. Los resultados y las recomendaciones derivados de estos trabajos constituyen la base científica para las decisiones políticas sobre el control de la contaminación atmosférica y otras políticas medioambientales de protección del entorno natural. En España, los resultados obtenidos por la Red CE de Nivel I ofrecen un balance periódico sobre la variación espacial y temporal del estado de los bosques y su relación con los factores de estrés, tanto biótico como abiótico, y entre ellos la Contaminación Atmosférica. Para ello, se recurre a una red sistemática internacional a gran escala consistente en más de 5700 puntos de seguimiento dispuestos en una cuadrícula de 16 x 16 km que cubre toda Europa de un modo aleatorio y sistemático. El número de puntos de la Red CE de Nivel I en España es actualmente 620 (con tan sólo 19 puntos en la Comunitat Valenciana y 12 en la Región de Murcia). Esta densidad de puntos se traslada a unos resultados muy fidedignos a nivel europeo, buenos a nivel regional continental, aceptables a nivel estatal, pero poco representativos a nivel autonómico en el caso español (MAGRAMA 2015). Debido a esto, se consideró necesario disponer en la Comunitat Valenciana de una red propia que cumpliera con los requisitos establecidos por la Unión Europea. Es por ello, en el año 2008 se puso en marcha de la red autonómica de Evaluación Fitosanitaria en las masas forestales de la Comunitat Valenciana (Red EFIVA) cuyo principal objetivo es la evaluación del estado de salud, observación de posibles daños y toma de muestras de determinados organismos patógenos de los bosques en la Comunitat Valenciana. Se pasó de una malla sistemática de 16x16 km a una de 8x8 km, lo que ha incrementado en 129 parcelas las 19 ya existentes, de forma que los resultados ofrecidos por la red EFIVA fuesen fidedignos y representativos de la realidad de los bosques valencianos. La metodología y los procedimientos empleados en la toma de datos de esta red permite comparar los resultados que de ella se obtienen con los de otras comunidades autónomas y otros países o regiones europeas (CITMA 2015 -b). Finalmente, de acuerdo con los tres pilares antes citados (Redes de Nivel I y II, y Forest Focus), la Direcció General de Gestió del Medi Natural de la Generalitat Valenciana dio, en 2013, un paso adelante en la investigación con la nueva Red de Muestreo Forestal y Fitosanitario de Ecosistemas Tipo (Red MUFFET 1 ). Tomando como modelo la Red de Estaciones de Seguimiento Forestal Permanenete (Red ESFP) establecida por la Dirección General de Patrimonio Natural y Biodiversidad de la Región de Murcia en 2008, los objetivos acometidos por esta red fueron más ambiciosos que los de las redes de evaluación fitosanitaria anteriormente comentadas, teniendo aplicación en diferentes aspectos medioambientales: desde la salud de los bosques hasta el estudio de su biodiversidad, pasando por el establecimiento de especies bioindicadoras de cambios climáticos y medioambientales, el descubrimiento de nuevas especies foráneas introducidas en el territorio gracias a su funcionamiento como red de alerta temprana, o el estudio detallado de las interrelaciones en el ecosistema forestal y sus dinámicas. Este tipo de redes tienen además el añadido de establecer series temporales de datos de gran relevancia científica (e. g. LINDENMAYER Y LIKENS 2009). 1 La Red MUFFET toma su nombre en honor al físico y naturalista inglés Thomas Muffet (1553-1604) coautor y promotor fundamental de la póstuma edición y de la difusión del primer tratado moderno sobre entomología, la guía ilustrada de insectos Insectorum sive minimorum animalium Theatrum (1634), obra que marca, definitivamente, el inicio de la Entomología Moderna.

81
En este capítulo se analizan preliminarmente los datos aportados por los primeros años de muestreo de ambas redes con el objetivo general de mostrar una herramienta potencial de análisis ecológico y de gestión relacionada, no sólo con M. galloprovincialis, sino con toda la comunidad de insectos saproxílicos asociada a esta especie. Pese alenorme interés de los resultados potenciales, deben ser asumidas, no obstante, las inevitables limitaciones que impone la actual carencia de series temporales de datos en los análisis y resultados presentados, y debe ser remarcada que la permanencia y, por tanto, el análisis a lo largo del tiempo, es una de las vocaciones fundamentales de esta herramienta.

82
Introducción
Escarabajos saproxílicos, especies invasoras y plagas Las especies invasoras son un elemento fundamental del cambio global y están contribuyendo definitivamente a la pérdida de biodiversidad y a la degradación de los ecosistemas en todo el mundo (PIMENTEL et al. 2005, PYŠEK Y RICHARDSON 2010). Entre estas especies destacan por su importancia los escarabajos perforadores de la madera, que agrupan a las principales plagas forestales, nativas y exóticas, y que son causantes de grandes pérdidas económicas y medioambientales en los bosques de todo el mundo (BROCKERHOFF et al. 2006). La facilidad para ser transportados en embalajes de madera y la dificultad de su detección en tales situaciones los convierten en un grupo alta capacidad invasora (HAACK 2001, MCCULLOUGH et al. 2006). La gravedad de algunas invasiones biológicas reside en las implicaciones de la llegada de una nueva especie a un escosistema con el que no ha coevolucionado. En términos generales, los efectos sobre el ecosistema suelen ser desfavorables y frecuentemente conlleva la extinción local de especies. Las especies invasoras lo son por la elevada adaptabilidad ecológica que poseen, la carencia de enemigos naturales, y la susceptibilidad de hospedantes que no han coevolucionado con ellas. Esto permite su expansión y su establecimiento en el territorio (WORK et al. 2005, KIRKENDALL Y FACCOLI 2010). La estimación actual es que la tendencia a la aparición de nuevas especies invasoras sea creciente en los próximos años debido, fundamentalmente, a dos causas: i), la continua globalización de los mercados y el aumento del movimiento de personas y materiales, que facilitan el desplazamiento de esas especies a otros territorios (LEVINE Y D'ANTONIO 2003, HULME 2009, KENIS et al. 2009, MARINI et al. 2011); y ii) la facilitación del establecimiento de especies tropicales en zonas templadas debido al calentamiento global (ROQUES et al. 2010).
El binomio Monochamus spp. y Bursaphelenchus xylophilus Uno de los casos más relevantes de las últimas décadas en Asia y Europa, es la invasión del nematodo de la madera del pino (NMP) Bursaphelenchus xylophilus, vectorizado forzosamente por las especies del género Monochamus spp. Bursaphelenchus xylophilus es una especie de cuarentena fitosanitaria incluida en la lista A2 de EPPO, causante de la enfermedad del marchitamiento del pino (pine wilt disease, PWD). Ésta es una de las más importantes enfermedades forestales a nivel global, llegando a causar graves daños en Asia, y considerada una grave amenaza para los pinares europeos (EVANS et al. 1996, EVANS et al. 2009, VICENTE et al. 2012). Poco después de la detección del NMP en Portugal en 1999 (MOTA et al. 1999), se demostró que M. galloprovincialis (Olivier 1795), especie muy habitual en la región mediterránea, era un vector eficaz de dicha enfermedad en la Península Ibérica (SOUSA et al. 2001). Muy posiblemente la entrada del NMP en Europa se produjo a través de vectores infectados alojados en el interior de material de embalaje de madera (palets, madera de estiba, etc.), que

83
fueron descargados en Setubal (Portugal), muy cerca de Lisboa (EVANS et al. 2009). De hecho, aunque los coleópteros perforadores de la madera podrían colonizar nuevas regiones a través del comercio y transporte de madera en rollo o de plantas para plantación (SIITONEN 2000, PIEL et al. 2008, LIEBHOLD et al. 2012), es el material de embalaje de madera el que tiene asociado un mayor riesgo respecto al transporte de este grupo de especies invasoras y es, por lo tanto, la vía de entrada mas común de estos organismos en nuevos territorios (HAACK 2001, STANAWAY et al. 2001, BROCKERHOFF et al. 2006).
Redes de muestreo sistemático, redes vigía En este contexto, la rápida detección de especies exóticas es de fundamental importancia para implementar a tiempo las medidas fitosanitarias de los programas de erradicación pertinentes, así como para poder estimar las tasas de entrada de especies invasoras (BROCKERHOFF et al. 2006, RASSATI et al. 2014). Es por ello que en muchos países, como Canadá, Australia, Nueva Zelanda o EEUU están combinando la inspección directa tradicional de los bienes importados en los puntos de entrada fronterizos con programas de trampeo de insectos saproxílicos (HAACK 2001, BROCKERHOFF et al. 2006, RABAGLIA et al. 2008, WYLIE et al. 2008, RASSATI et al. 2014). Estos programas tienen como objetivo la identificación de los lugares donde es más probable tanto la llegada como el establecimiento de las especies invasoras. Con ello se pretende aumentar la eficiencia en la detección temprana y optimizar el uso de recursos (BOGICH et al. 2008). La instalación de trampas cebadas con atrayentes debería ser realizada en los puntos de entrada fronterizos, así como en los bosques circundantes donde las especies recién llegadas podrían establecerse (RASSATI et al. 2014, RASSATI et al. 2015b). En términos generales, el empleo de redes de seguimiento permanente y de trampas vigía para la detección temprana de organismos exóticos y especies de cuarentena, representa una de las mejores herramientas actuales de la gestión integrada de plagas (RASSATI et al. 2014, RASSATI et al. 2015a). Y desde esta perspectiva, asumiendo la alta diversidad de organismos que deben ser detectados, tanto las trampas como los atrayentes deben ser tan generalistas como sea posible (RASSATI et al. 2014, RASSATI et al. 2015a). Los requerimientos metodológicos para la detección temprana de especies invasoras, convergen con los requerimientos de los estudios ecológicos de las especies que pueden llegar a amenazar. La información reacabada sobre especies locales en tales redes de seguimiento, permite la caracterización y evaluación de la biodiversidad y conformadción de las comunidades presentes en el hábitat de estudio así como su evolución temporal frente a cambios en el ecosistema. En este sentido, si bien los insectos perforadores de la madera no nativos son considerados como unas de las mayores amenazas ambientales en todo el mundo (HAACK 2001, WORK et al. 2005, KIRKENDALL Y FACCOLI 2010), los insectos perforadores de la madera nativos, y los insectos saproxílicos en general, juegan un papel importante en los procesos de descomposición y recirculación de nutrientes retenidos en la biomasa de madera en los ecosistemas forestales. En este sentido, la desaparición de los bosques maduros y de la madera muerta es la causa de que esta comunidad de insectos sea una de las más amenazadas en el ecositema forestal (GROVE 2002). Muchas especies de escarabajos saproxílicos de amplia distribución en Europa han sufrido un deterioro significativo de sus poblaciones (NIETO Y ALEXANDER 2010). De acuerdo con estos autores, las principales amenazas sobre estas las especies de coleópteros saproxílicos europeos son la pérdida de hábitat por tala y extracción de madera, la expansión de los terrenos agrícolas y la expansión urbana. En menor medida, la gestión de madera muerta para la prevención y extinción de incendios forestales también representa una amenaza sobre estas comunidades. NIETO y ALEXANDER (2010) exponen

84
asimismo que se dedican pocos esfuerzos en Europa para la vigilancia sistemática de especies de escarabajos saproxílicos, por lo que apuntan a la clara necesidad de elaborar conjuntamente la información obtenida, así como, un plan europeo más amplio de acciones de conservación.
Estudios ecosistémicos forestales Los arboles castigados por agentes abióticos (como la sequía), son más susceptibles de ser infectados que los sanos, ya que estos últimos tienen defensas más sólidas contra el ataque (ALLEN 1976, MCDOWELL et al. 2008). Comunidades enteras de insectos saproxílicos forestales dependen de este principio. Para encontrar árboles adecuados para la reproducción, muchas especies de insectos buscan señales químicas y visuales emitidas por árboles bajo algún tipo de estrés, un recurso muy intermitente espacial y temporalmente (GROVE 2002). A su vez, la comunidad de insectos depredadores y parasitoides depende de la comunidad de insectos fitófagos y micófagos. En general, las poblaciones los diferentes gremios tróficos de sarpoxílicos, no sólo están moduladas por la ocasional disponibilidad de su hábitat, y también están moduladas directamente por factores abióticos como el clima. Ciertas poblaciones de insectos pueden incrementar notablemente su tamaño en los años con inviernos suaves y cortos, ya que les permite alagar su periodo reproductivo además de la reducción de la mortandad invernal. De este modo la temperatura y la precipitación además modular el crecimiento de los árboles, no solo influye en la reproducción de insectos, sino también en las interacciones insecto/árbol e insecto/insecto. Ligeros cambios en las temperaturas y precipitaciones pueden convertir a especies vegetales, antes resistentes, en especies vulnerables, aumentando la intensidad de los ataques de insectos que tiempo atrás se comportaban con un carácter secundario, o permitiendo que nuevas especies agresivas aparezcan en zonas en las que antiguamente estaban ausentes (LINDNER et al. 2010). En efecto, el equilibrio dinámico de los ecosistemas forestales, puede ser desestabilizado por diferentes factores, entre los que destacan los incrementos poblacionales de insectos que se alimentan de arboles, que pueden derivar en brotes epidémicos bajo condiciones ambientales favorables. De este modo, una sequía puede predisponer a los árboles al ataque de insectos, haciéndolos más vulnerables por debilitamiento. El resultado neto puede ser la pérdida de superficie boscosa, ejecutada por los insectos y modulada por el clima (DUNN Y CRUTCHFIELD 2006). Pueden definirse entonces, de manera simplificada, tres tipos de actores: los árboles, los insectos y el clima. Su coincidencia espacio-temporal define un sistema altamente complejo, compuesto de muchos actores en interacción continua. El clima describe el contexto de la dinámica a pequeña escala: la temperatura modula la reproducción de insectos, y en interacción con la precipitación, también el crecimiento de los árboles. Es importante destacar que existen bucles de retroalimentación que puede mantener la estabilidad del ecosistema o llevar a la inestabilidad así como amplificar pequeños efectos a una escala mayor. DUNN y CRUTCHFIELD (2006) utilizan en su trabajo el papel que los escolítidos (Curculionidae, Scolytinae) en la deforestación a gran escala y en el cambio climático. Ciertas especies de estos escolítidos y en ciertas condiciones ambientales, pueden ejecutar procesos de deforestación muy importantes. Aunque el ejemplo es necesariamente incompleto, ya que solo se refiere a un tipo de ecosistema forestal, sirve para ilustrar la complejidad de las interacciones que están implicadas en la dinámica forestal. Los escolítidos representan un agente biótico que actúa a escala temporal mínima, es decir, de años. Sin embargo algunas especies de escolítidos de los bosques boreales norteamericanos, en muy poco tiempo, pueden causar la deforestación de grandes áreas, generando perturbaciones ecológicas sin precedentes (KURZ et al. 2008).

85
Al analizar la relación entre los componentes de este circuito retroalimentado, la primera conclusión es que se conoce muy poco sobre las interacciones entre insectos y árboles, y entre éstos y el clima (DUNN Y CRUTCHFIELD 2006). Se hace necesario por tanto poner mayor atención en algunos aspectos, como son las interacciones entre especies (depredación, simbiosis, competencia o mutualismo), así como las relaciones intraespecíficas (dinámica poblacionales, depredación, competencia o cooperación intraespecíficas). El presente capítulo tiene como objetivo remarcar la doble función de las redes de seguimiento permanente y de trampas vigía, las cuales, además de constituir una de las mejores herramientas para la detección temprana de organismos exóticos y especies de cuarentena, permiten la captura de una gran variedad de especies nativas gracias al carácter genérico de las trampas y cebos utilizados, lo cual hace posible la caracterización y evaluación de las comunidades presentes en los hábitats de estudio. Estos estudios ecológicos, afrontados también desde la perspectiva del análisis de redes de interacciones, permiten realizar análisis de evolución temporal o de estructura de las comunidades de insectos saproxílicos, en este caso, asociadas a la especie objeto de estudio: M. galloprovincialis, cuya relación con la comunidad de insectos con los que se interrelaciona se encuentra verdaderamente poco estudiada (SLAMA 1998, KENIS Y HILSZCZANSKI 2004). Los primeros datos aportados por ambas redes son analizados parcialmente en el presente de capítulo. El verdadero valor de estas redes de muestreo sistemático se basa en su vocación de permanencia en el tiempo y en la consecuente capacidad de aportar información de series temporales de muestreos. Los resultados y los análisis que aquí se presentan son un estudio preliminar de los aspectos más fundamentales relacionados con el protagonista principal de esta memoria, M. galloprovincialis, y a su vez suponen una primera aproximación al potencial de estudio y conocimiento ecológico que este tipo de redes puede generar. Este Capítulo 4 ha sido estructurado en dos secciones que son discutidas de manera conjunta al final del capítulo. La primera está dedicada al análisis de los datos proporcionados por la Red EFSP, con objetivo de caracterizar la estructura de las comunidades de coleópteros saproxílicos y el papel particular que cumple M. galloprovincialis en los pinares del sudeste de la península Ibérica a través de su análisis mediante redes de interacción. La segunda sección del capítulo está enfocada al análisis de los datos aportados por la Red MUFFET con el objetivo de estudiar las comunidades de insectos saproxílicos asociadas a M. galloprovincialis en el área de estudio.

86
Sección 4.1. Análisis de la estructura de las comunidades de coleópteros
saproxílicos en los pinares del levante de la Pénínsula Ibérica mediantes
redes de interacción.
Introducción De acuerdo con CARNICER et al. (2009) las comunidades ecológicas se definen como entidades complejas y dinámicas que están integradas por poblaciones que, a su vez, son temporalmente variables e interactúan de maneras muy diversas. Ejemplos de estas interacciones son la competencia, depredación, herbivoría, dispersión, polinización o parasitismo, entre otros. Esto causa continuos cambios en la arquitectura de los flujos de energía y de materia en el espacio y el tiempo. Tal abrumadora complejidad puede ser analizada en representaciones gráficas mediante redes, en las que, básicamente, las especies se representan como nodos y las interacciones entre las especies se representan como vínculos. El análisis de redes es una valiosa herramienta para el estudio de la diversidad de especies y las interacciones en las grandes redes tróficas (BASCOMPTE et al. 2003). Un alto número de comunidades ecológicas se han estudiado bajo este punto de vista, descubriéndose patrones de interacción especializados como el anidamiento de redes mutualistas (BASCOMPTE et al. 2003) o la modularidad en redes antagónicas (THÉBAULT Y FONTAINE 2010), proporcionando así información sobre la función y la evolución de los componentes del sistema. Los patrones de interacción actúan como una variable modelando la estructura de la red de interacciones, la reducción de la competencia interespecífica efectiva y mejorando el número de especies que coexisten (BASTOLLA et al. 2009). Además, estas interacciones pueden determinar la respuesta de las comunidades a las perturbaciones medioambientales (GUIMERA Y AMARAL 2005, IVES Y CARPENTER 2007, LAWRENCE et al. 2012, QUINTO et al. 2012). Los enfoques acerca de qué son y cómo tratar estas redes han proporcionado una manera muy visual de sintetizar la estructura de los sistemas ecológicos y sus interacciones. De este modo se pueden estudiar sus propiedades emergentes y analizar la estabilidad y robustez de las comunidades. Sin embargo, los enfoques de las redes se han visto obstaculizados por la falta de información explícita tanto espacial como temporal. Afortunadamente, un creciente número de estudios están llenando progresivamente estas lagunas, proporcionando tanto la mejora de enfoques espacialmente explícitos como descripciones exhaustivas de la dinámica temporal de las redes (CARNICER et al. 2009, THOMPSON et al. 2012). Según CARNICER et al. (2009), otro desafío aún sin resolver consiste en identificar y evaluar cuantitativamente la importancia relativa de los diversos procesos ecológicos y evolutivos que, en última instancia, conforman la estructuras de redes. De hecho, la mayoría de las redes ecológicas comparten algunas propiedades, pero la importancia relativa de la evolución y los procesos ecológicos que moldean estos patrones emergentes, aún no se entienden en su plenitud. En términos más generales, las redes ecológicas se caracterizan por tener la mayoría de las interacciones concentradas en pocas especies, que luego ejercen una fuerte influencia en las demás. Sin embargo, a corto plazo, las comunidades pueden varíar en composición de especies, debido variaciones en las fenologías, o en los comporamientos migratorios y dispersivos, entre otros. Por otro lado, en escalas de tiempo mucho más largas, las especies evolucionan y coevolucionan en un conjunto de rasgos morfológicos y etológicos que hacen variar las especies con las que interactúan. Las redes tróficas son un ejemplo de redes complejas. Como su propio nombre indica, el análisis de este tipo de redes y de su topología conlleva complicadas computaciones. Además

87
las redes tróficas están caracterizadas por una alta complejidad estructural, por una dinámica evolutiva o por una alta diversidad de conexiones, entre otros (STROGATZ 2001, THOMPSON et al. 2012). Sin embargo, el mero hecho de caracterizar la anatomía de una red ayuda en la comprensión de la función de los diferentes nodos de esa red, y de la red en sí; en otras palabras, la estructura siempre afecta a la función (ver por ejemplo STROGATZ 2001).
Material y métodos
Diseño experimental
Una red para el muestreo sistemático de insectos saproxílicos (sensu lato) vinculados a los pinares mediterráneos fue instalada en la Región de Murcia, Sudeste de la Península Ibérica, entre los años 2008 y 2010. Esta red fue denominada Red de Estaciones de Seguimiento Forestal Permanante (Red ESFP). La red consistió en un conjunto de 20 parcelas (o estaciones ESFP) distribuidas a lo largo de toda la Región de Murcia (Tabla 4.1, Figura 4.1). El área de estudio comprendió las masas forestales más representativas de las diferentes formaciones vegetales de pinar del sudeste de la Península Ibérica. La ubicación de las estaciones EFSP se realizó de manera aleatoria y estratificada, utilizándose la variable altitud como modulador de la mayor parte de la variabilidad ambiental a media escala en la Región de Murcia.
Tabla 4.1 Características de las estaciones forestales de seguimiento permanenete (ESFP) de la región de Murcia. Parcela Localización Especie principal Especie acompañante Tipo1 Altura Dominante Altitud ESFP1 Jumilla 1 P. halepensis BM 10 823 ESFP2 Mazarron 2 P. halepensis BI 5 459 ESFP3 Murcia 3 P. halepensis R 4 407 ESFP4 Espuña 4 P. halepensis BM 6 518 ESFP5 Espuña 5 P. nigra BM 10 1436 ESFP6 Espuña 6 P. halepensis P. pinaster BM 10 1288 ESFP7 Lorca 1 P. halepensis R 5 831 ESFP8 Zarcilla 8 P. halepensis R 4 903 ESFP9 Murcia 9 P. halepensis BM 10 544 ESFP10 Zarcilla 10 P. halepensis BM 4 789 ESFP11 Caravaca 11 P. halepensis BM 8 883 ESFP12 Caravaca 12 P. halepensis BM 5 864 ESFP13 Caravaca 13 P. halepensis P. pinaster BM 10 1154 ESFP14 Caravaca 14 P. halepensis BM 4 1044 ESFP15 Moratalla 15 P. nigra BM 6 1226 ESFP16 Abarán 16 P. halepensis R 4 349 ESFP17 Abarán 17 P. halepensis BM 5 509 ESFP18 Yecla 18 P. halepensis BM 8 896 ESFP19 Jumilla 19 P. halepensis BM 5 927 ESFP20 Jumilla 20 P. halepensis BM 3 852 1BM, bosque maduro; BI, Bosque en islas; R, repoblación.

88
Figura 4.1 Localización de las parcelas de muestreo de la Red ESFP (Murcia)
Para la captura de los insectos fueron utilizadas trampas de bandas cruzadas, modelo “crosstrap®” (Sanidad Agrícola ECONEX S.L., Siscar, Murcia, España). Éstas fueron cebadas con un compuesto atractivo para una amplia variedad de escarabajos perforadores de la madera (RASSATI et al. 2014, RASSATI et al. 2015b). Este atrayente está compuesto por un dispensador de (-)α-pineno (velocidad de emisión de 300 mg/día), un dispensador de etanol (velocidad de emisión de 2000 mg/día) y un dispensador de ipsdienol (95.28%), ipsenol (4.7%) y (s)-(+)-cis-verbenol (0.02%). Todos los atrayentes fueron fueron provistos por la empresa Sanidad Agrícola Econex S.L. (Siscar-Santomera, Murcia, España) y SEDQ S.L. (Barcelona, España). La presión de muestreo fue mensual durante todo el periodo de estudio, y la captura de los insectos se realizó en húmedo, utilizando etilenglicol como conservante en el bote recolector. La diana de este método de trampeo es la comunidad de saproxílicos sensu lato, de acuerdo con la definición propuesta en NIETO y ALEXANDER (2010). Los individuos capturados fueron cribados e identificados en laboratorio. La temperatura fue controlada mediante el uso de un registrador continuo HOBO pendant UA-001-64 (Onset, USA). Cada registrador se programó para realizar una medida de temperatura del aire cada dos horas. La pluviometría fue medida mediante el uso de un pluviómetro de cuchara modelo Rain-o-m100 (Onset, USA), conectado con un registrador HOBO pendant event UA-003-64 (Onset, USA).
Análisis de datos
Determinación de las interacciones entre especies
A partir de los valores de abundancia de cada una de las especies de insectos saproxílicos capturadas en la red ESPF durante el periodo de 2008 a 2010, se elaboraron dos filtros para determinar y cuantificar las interacciones entre las especies (Tabla 4.2). Un primer filtro identificó interacciones por coincidencia espacio temporal, y un segundo filtro identificó interacciones a partir de información biológica.

89
Tabla 4.2 Tipos de relaciones consideradas para establecer la red de interacciones.
Interacción Características Competencia Bidireccional, desfavorable para ambas especies Depredación Unidireccional, favorable para la especie receptora (depredador) Comensalismo Bidireccional, efectos indeterminados.
Determinación de las interacciones por coincidencia espacio temporal
Se consideró que para que existiese interacción entre dos especies debía existir coincidencia espacio temporal, es decir coincidencia corológica y fenológica de dichas especies en el momento de colonizar un árbol en el adecuado estado de vigor. La coincidencia temporal se determinó directamente a partir del valor de la correlación de las fenologías de cada par de especies: en los casos de valores de correlación negativos se consideró que no existía interacción posible entre ambas especies. Con los valores de correlación positivos se consideró que sí podía existir interacción, siendo ésta además cuantificada con el valor de la correlación. De este modo, a mayor grado de solapamiento fenológico, mayor correlación y, por lo tanto, mayor posibilidad de interacción.
Determinación de las interacciones a partir de información biológica.
Un segundo filtro fue establecido a partir de la información biológica existente para cada especie. Para ello se utilizaró la información hallada en la literatura, así como la experiencia del equipo de investigación y consultas a especialistas. Se consideraron sólo especies de Coleoptera y una especie de Hymenoptera. Diferentes especies de los órdenes Diptera e Hymenoptera (salvo la especie incluida) no fueron incluidas en el estudio. Se elaboró una tabla de doble entrada colocando, en las filas, la clase a la que se adscribió cada especie (Tabla 4.3), y en las columnas, los aspectos de la biología de cada especie que pudieran limitar las interacciones entre cada par de especies (Tabla 4.4).

90
Tabla 4.3 Categorías a las que se adscribe cada especie capturada en la red ESFP.
Clase Descripción
Escolítido Especies corticícolas exclusivamente floemofagas, de tamaño entre 2 y 6 mm. Colonizan árboles en estados iniciales de decaimiento.
Depredador Especies de hábitos exclusivamente depredadores, principalmente de escolítidos. De tamaños entre 3 y 20 mm, pertenecen a las familias Carabidae, Histeridae, Trogossitidae, Tenebrionidae, Colidiidae, Rhyzophagidae y Cleridae, entre otras. Todas las especies son depredadores tanto en estado adulto como en el larvario. Las especies de mayor tamaño cazan en estado adulto sobre la superficie del árbol. Las especies de menor tamaño cazan en el interior de las galerías de sus presas. Todos los estados larvarios depredan, principalmente larvas de escolítidos, subcorticalmente. La familia Carabidae depreda sobre la corteza en todos sus estadios.
Depredador facultativo Especies de hábitos floemo-xilófagos, pero que en la fase floemófaga consumen larvas subcorticales, principalmente escolítidos. Su tamaño oscila entre los 4 y 30 mm, perteneciendo a las familias Cerambycidae, Buprestidae, Elateridae y Curculionidae, entre otras. Llegan a árboles en un estado de decaimiento avanzado y frecuentemente atacados por escolítidos.
Ambrosía Especies pertenecientes a la subfamilia Scolitinae, de hábitos xilomicetófagos. Son especies de tamaños entre 2 y 4 mm. Viven en galerías en la madera profunda de árboles moribundos o recientemente muertos, donde se alimentan de hongos que germinan a partir de esporas transportadas por los propios escolítidos. Los machos son ápteros y las hembras pueden realizar partenogénesis.
Xilófago Especies estrictamente xilófagas, de entre 2 y 30 mm. En estado larvario viven en la madera profunda de árboles en estados avanzados de colonización por otras especies. Pertenecen a las familias Bostrichidae, Anobiidae, Priocenidae y Oedemeridae, así como Braconidae (Hymenoptera), entre otras.
Rizofágido Especies que se alimentan de micelio de hongos que crecen en el espacio subcortical de árboles en avanzado estado de degradación, con tamaño de unos 3 mm. Pertenecen a la familia Eucmonidae.
Comensal Especies no extrictamente saproxilófagas, generalmente saprófitas, que son atraídas por restos vegetales en descomposición. De tamaños entre los 3 y 30 mm, pertenecen a las familias Cerambycidae, Cucujidae, Melodonthidae, Nitidulidae, Aphodidae, Alleculidae, Tenebrionidae, Mycetophagidae, Mycteridae, Cetonidae y Dermestidae, entre otras.
Defoliador Especies no saproxilófagas, sino que se alimentan de acículas de pino en todos sus estadios vitales. De tamaños entre 4 y 6 mm, pertenecen a la familia Curculionidae.

91
Tabla 4.4 Aspectos de la biología de las especies que pueden limitar las interacciones entre ellas.
Hábito Descripción
Especie hospedadora Se tiene en cuenta la especie o especies vegetales susceptibles de ataque y/o colonización por parte de la especie de insecto forestal. Se consideran las especies P. halepensis, P. nigra, P. pinaster, Quercus spp., Juniperus spp., Cystus spp., Pistacea spp., Thapsia spp., Ficus carica, Thymelaea spp., Retama spp., Gramineas y Fabaceas.
Zona de alimentación Área de la planta en la que se desarrolla total o parcialmente la vida de la especie. Se consideran como distintas el área subcortical incluyendo el floema y el área del xilema.
Área de caza En el caso de los depredadores, se anota como distinto si la especie caza en el exterior de la corteza o en el área subcortical, en cualquier fase de su ciclo vital.
Parte del árbol colonizada Se considera como hábitos diferentes si las especies viven estrictamente en el fuste o ramas gruesas de los árboles o si son capaces de colonizar ramas de menor diámetro.
Hábitos detritívoros Se establece una categoría especial para especies comensales de hábitos detritívoros estrictos.
Hábitos defoliadores Se establece una categoría especial para especies defoliadoras.
A partir de estas categorías fueron inferidas sus posibles relaciones mediante un sistema de filtrado jerárquico (Figura 4.2). Para cada parcela se obtuvo una matriz cuadrada de dimensión igual al número de especies, donde se representaron, de forma binaria, la posibilidad de interacción entre pares de especies. El 0 indicó imposibilidad de interacción y el 1 indicó posibilidad de interacción. Esta matriz incorporó información tanto de relaciones bidireccionales (o aristas) por competencia, como unidireccionales (o arcos) por depredación.
Figura 4.2 Diagrama de decisiones para establecer posibilidad de interacción a partir de aspectos de la biología de pares de especies. El conector con línea continua indica opción positiva, mientras que con
línea discontinua indica opción negativa.
Posteriormente, para cada una de las 20 parcelas de la red ESFP se elaboró una matriz sintética de las interacciones posibles entre pares de especies mediante álgebra de matrices de modo que la matriz de coincidencias feno-corológicas, con los valores de correlación fenológica positiva, se multiplicó por la matriz binaria obtenida del análisis de los aspectos biológicos. Como resultado se obtuvo una matriz cuadrada de dimensión igual al número de especies encontradas en la estación, con información cuantitativa de la posible interacción entre cada par de especies.

92
Representación gráfica y análisis de las redes de interacciones.
Para llevar a cabo el estudio de la estructura de su red de interacciones, en que las especies son los nodos y las interacciones son trazos, se obtuvieron los grafos correspondientes a las 20 estaciones de seguimiento forestal permanente (ESFP). Para ello se utilizó el software Pakej (BATAGELJ Y MRVAR 2006). Para la representación gráfica se utilizó el algoritmo de Kamada-Kawai para una representación plana, en dos dimensiones y en la modalidad libre, donde no se fuerza la topología. Las interacciones unidireccionales de depredación, arcs en terminología de Pajek (BATAGELJ Y MRVAR 2006), se representan como un trazo con una única punta de flecha. La punta señala al depredador, de acuerdo con el flujo de materia en una red trófica. Las interacciones de competencia, bidireccionales se representan mediante un trazo con dos puntas de flecha, denominadas edges en Pajek (BATAGELJ Y MRVAR 2006). La cuantificación de cada interacción se representa con el grosor del trazo. De forma general se utiliza en la presente memoria la doneminación “conexión” par referirse a ellas. Una vez construida cada red, se calcularon los parámetros fundamentales (Tabla 4.5). Tabla 4.5 Parámetros medidos en cada una de las redes de interacciones obtenidas Parámetro Definición Interpetación Nodos Numero de nodos en la red. Medida de complejidad de la red. Grado Numero de interacciones que recibe y emite cada nodo. Medida de complejidad de la red. Grado máximo Número de conexiones en el nodo más conectado Medida de complejidad de la red. Grado medio Valor medio del grado de todos los nodos de la red Medida de conectividad global. Conexiones totales
Suma de todas las conexiones de la red Medida de complejidad de la red.
Densidad Número de conexiones en relación al número total conexiones posibles
Medida de las saturación de la conectividad
Conectores principales
Determinación de los nodos que conectan diferentes subredes.
Determinación de las especies clave en el sistema.
Selección de indicadores
Con la información extraída de la redes de interacciones se realizó una selección de parámetros indicadores de calidad forestal. Para ello se utilizó análisis de redundancias RDA (Redundance Detendred Analysis) de la extensión Vegan (OKSANEN et al. 2008) y modelos lineares generalizados (GLM) para estudiar tendencias entre los parámetros calculados de la red y ciertas variables ambientales extraídas de cada parcela (Tabla 4.6).
Tabla 4.6 Variables utilizadas en la selección de indicadores de calidad forestal.
Variable Código Definición Rango
Altitud Altitud Altitud de la ESFP, medida con GPS y altímetro digital.
349 – 1436 m
Temperatura media tm Temperatura media calculada a partir de las medidas del registrador en cada ESFP.
13.29 – 21.14 ºC
Temperatura máxima del mes más cálido
tmax_mc Temperatura máxima absoluta en el mes más cálido registrado durante el periodo de muestreo.
38.60 – 51.72 ºC
Temperatura máxima del mes más frío
tmax_mf Temperatura máxima absoluta en el mes más frío registrado durante el periodo de muestreo.
4.21 – 26.39 ºC
Temperatura máxima media
tmax_med Temperatura máxima media mensual durante el periodo de muestreo.
26.02 – 38.35 ºC

93
Variable Código Definición Rango
Temperatura mínima del mes más cálido
tmin_mc Temperatura mínima absoluta en el mes más cálido registrado durante el periodo de muestreo.
9.67– 21.76 ºC
Temperatura mínima del mes más frío
tmin_mf Temperatura mínima absoluta en el mes más frío registrado durante el periodo de muestreo.
-12.36 – 1.33 ºC
Temperatura mínima media
tmin_med Temperatura mínima media mensual durante el periodo de muestreo.
0.58 – 11.73 ºC
Precipitación acumulada pt Precipitación total acumulada mensualmente durante el periodo de muestreo.
20 – 425 l/m2
Especie de árbol presente
Sp_arbol Variable categórica, Especies de árboles forestales dominantes en la ESFP.
Phale Pnigr Phale-Ppina
Tipo de formación forestal
Tipología Tipo de formación boscosa, según una categoría sintética del IFN3.
Bosque_isla Bosque_maduro Repoblación
Diámetro medio de los árboles
DBH_media Valor medio del diámetro a la altura normal (1,5 m) de todos los árboles de la parcela.
5 – 25.16 cm
Crecimiento diametral medio
Crec_med Valor del crecimiento medio mensual de los árboles marcados con dendrómetros en cada parcela.
-0,07 – 0,35 cm

94
Resultados
Resultados generales
Se capturaron un total de 69234 ejemplares pertenecientes a 141 especies repartidas en 30 familias y dos órdenes de la comunidad de insectos saproxílicos sensu lato (Anejo 4.1). De ese número total de ejemplares capturados, el 77,6 % pertenece a la subfamilia Scolytinae (18 especies), mientras que el 20,1 % se reparte entre las familias Colydiidae (1 especie), Tenebrionidae (5 especies), Cerambycidae (22 especies), Cleridae (6 especies) y Trogossitidae (2 especies). El 0,97 % restante se reparte entre 87 especies pertenecientes a 24 familias (Figura 4.3).
Figura 4.3 Abundancia de individuos por familias. (A) Abundancia absoluta. (B) Representación a escala
logarítmica.

95
Redes de Interacciones
Las figuras del Anejo 4.2 ilustran los grafos que representan la red de interacciones para cada parcela de la Red ESFP. A continuación, en el texto, se han incluido los grafos de las estaciones ESFP2 y ESFP15 como ejemplo (Figura 4.4 y Figura 4.5). Los colores representan los tipos funcionales definidos en el Anejo 4.1. Para este caso, el color azul representa a depredadores y el color rosa a escolítidos. El tamaño del nodo es directamente proporcional a la abundancia de la especie en cada parcela de muestreo. El tamaño de la punta de la flecha en los trazos es directamente proporcional a la intensidad de la interacción. En la Tabla 4.7se muestra un resumen de las características principales de las redes de interacciones construidas.
Figura 4.4 Grafo que representa la red de intereacciones de la estación ESFP 2
Figura 4.5 Grafo que representa la red de intereacciones de la estación ESFP 15

96
Tabla 4.7 Resumen de características principales de las redes de interacciones construidas.
ESP Nº sp Grado Grado medio Nº conexiones Densidad Elementos más conectados
ESFP1 43 0 - 40 13.54 582 0,16 Plegaderus otti (depredador) Hylurgus micklizti (escolitido)
ESFP2 23 0 - 18 6,17 142 0,14 Aulonium ruficorne (depredador) Orthotomicus erosus (escolitido)
ESFP3 29 0 - 24 7,24 210 0,13 Orthotomicus erosus (escolitido)
ESFP4 39 0 - 34 12,20 554 0,19 Tenmoscheisla coerulea (depredador) Scobicia chevrieri (xilofago)
ESFP5 38 0 - 35 11,05 420 0,15 Allonix quadrimaculata (depredador)
ESFP6 48 0 - 40 16,96 814 0,18 Thanasimus formicarius (depredador) Acmaoederella bipunctata (facultativo)
ESFP7 43 0 - 33 13,35 574 0,16 Acanthocinus griseus (facultativo)
ESFP8 46 0 - 42 15,56 716 0,17 Acmaoederella bipunctata (facultativo)
ESFP9 36 0 - 38 14,39 518 0,21 Corticeus pini (depredador) Calcophora mariana (facultativo)
ESFP10 39 0 - 48 15,08 890 0,13 Tenmoscheisla coerulea (depredador) Acmaoederella bipunctata (facultativo)
ESFP11 40 0 - 34 10,5 420 0,14 Monochamus galloprovincialis (facultativo)
ESFP12 54 0 - 40 13,26 716 0,13 Monochamus galloprovincialis (facultativo)
ESFP13 35 0 - 34 9,6 336 0,14 Corticeus pini (depredador) Monochamus galloprovincialis (facultativo)
ESFP14 57 0 - 57 15,4 878 0,14 Acanthocinus griseus (facultativo)
ESFP15 54 0 - 60 26,29 1420 0,25 Antaxia lencinae (facultativo) Cylister filiformis (depredador)
ESFP16 36 0 - 34 7,56 272 0,11 Aulonium ruficorne (depredador) Orthotomicus erosus (escolitido)
ESFP17 37 0 - 42 13,84 512 0,19 Monochamus galloprovincialis (facultativo)
ESFP18 34 0 - 26 11,18 380 0,17 Acmaoederella bipunctata (facultativo) Acanthocinus griseus (facultativo)
ESFP19 35 0 - 38 13,6 476 0,2 Monochamus galloprovincialis (facultativo)
ESFP20 36 0 - 33 12,78 460 0,19 Aulonium ruficorne (facultativo) Monochamus galloprovincialis (facultativo)
El número de conexiones varía entre 142 y 1420, estando el 75 % de los valores por debajo de 716, con media en 564 y mediana en 515. De hecho, tan solo un valor, el de la parcela 15, se encuentra en el 4º cuartil. Variando entre 23 y 59 especies, la mitad las parcelas acumularon entre 30 y 40 especies. Respecto al número máximo de conexiones por nodo o grado máximo, sus valores varían entre 18 y 60, con una distribución similar a la del número de conexiones, con una discontinuidad en los rangos de secuencias observadas, entre los valores de 50 y 55. Lo mismo se observa en la distribución de frecuencias del grado medio, que varía entre 6,17 y 27,24, con una discontinuidad entre los valores medios de 20 y 25 conexiones. Respecto a una primera aproximación a la selección de indicadores, el análisis de la RDA mostró que los dos primeros ejes del RDA explicaron el 72,13 % de la variabilidad de los datos, aunque el eje 2 apenas aportó explicación. En la Figura 4.6 se representan gráficamente

97
los dos primeros ejes del RDA. El eje 1 está relacionado con el gradiente de temperaturas mínimas, de modo que las estaciones con menor temperatura mínima se colocan en la parte positive del eje y las de mayor temperatura mínima en la parte negativa. En esta ordenación se segrega claramente la variable número de conexiones, que se relaciona positivamente con estaciones con temperaturas mínimas más frías, como son las ESFP15, 6, 10, 14, 12 y 8. Las demás variables aparecen agrupadas muy cerca del centro del los ejes. El eje 2 apenas parece tener relación con las variables estudiadas.
Figura 4.6 Representación gráfica de los dos primeros ejes del RDA.
Se encontró una relación entre la temperatura mínima del mes más frío y la variable “número de conexiones” (Figura 4.7). Se observa una relación lineal explicativa que se ajusta a la fórmula: Nº Conexiones= 4,022-0,229xTmin_mf (1) R2adj= 0,34; F= 10,75; P= 0,004 Y por último, en la representación gráfica se observa que un grupo de ESFPs (13, 11, 5, 19, 20 y 18) se alejan de la tendencia lineal.
-10 0 10 20
-15
-10
-50
510
15
RDA1
RD
A2
VerticesGrado_maxGrado_med Conexionesdensidad
ESFP1
ESFP2
ESFP3 ESFP4
ESFP5
ESFP6
ESFP7
ESFP8
ESFP9
ESFP10
ESFP11
ESFP12
ESFP13
ESFP14
ESFP15
ESFP16
ESFP17
ESFP18
ESFP19ESFP20tmin_mf
pt
tmax_mf
tmin_mc
tmax_med
tmin_med
DBH_media
Cre_med
-10
1

98
Figura 4.7 Representación gráfica de la relación entre la variable temperatura mínima del mes más
frío y el número de conexiones.
-12 -10 -8 -6 -4 -2 0
200
400
600
800
1000
1200
1400
Temperatura mínima del mes más frío
Nº d
e co
nexi
ones
1
23
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19 20

99
Sección 4.2 Comunidades de coleópteros saproxílicos asociadas a
Monochamus galloprovincialis (Olivier 1795) en el levante de la
Península Ibérica.
Introducción Un objetivo fundamental de la investigación ecológica es entender los mecanismos que influyen en la estabilidad, la complejidad y la dinámica temporal de los ecosistemas. Esto es esencial para ser capaces de predecir, y, finalmente, mitigar, las consecuencias del aumento de las perturbaciones ambientales, tales como la pérdida de hábitat, el cambio climático o las invasiones de especies exóticas (MCCANN 2000, MONTOYA et al. 2006). El estudio de las interacciones entre los organismos que componen las redes ecológicas ha jugado un papel central en el desarrollo de la ecología como disciplina científica (HARDY 1924, MCCANN 2000). Las especies del género Monochamus, y en concreto M. galloprovincialis, son insectos saproxílicos de hábitos secundarios que buscan pinos muy debilitados para completar su ciclo biológico (VIVES 2000). El estudio de este género de coleópteros ha despertado un gran interés en los últimos años ya que sus especies son los únicos vectores conocidos del nematodo de la madera del pino Bursaphelenchus xylophilus (Steiner et Buhrer) Nickle (NMP), agente causal de la enfermedad del marchitamiento del pino. La introducción del NMP en Portugal causó una gran mortandad de P.pinaster en la península de Setúbal entre los años 2001 y 2007. Diversos estudios relacionaron, en ese periodo, la presencia de M. galloprovincialis con las de otros insectos saproxílicos (pertenecientes las familias Cerambicidae, Curculionidae (Scolytinae) y Buprestidae, en el interior de los árboles muertos (BONIFÁCIO 2009). No obstante solamente M. galloprovincialis fue identificado como vector efectivo de la enfermedad (SOUSA et al. 2001). Las larvas de los cerambícidos son consideradas por norma general fitófagas (CRAIGHEAD 1923, LINSLEY 1959), aunque el canibalismo durante dicha fase larvaria (VICTORSSON Y WIKARS 1996) y la depredación de escarabajos cortícolas han sido constatados en algunos estudios (HELLRIGL 1971). Asimismo, la colonización por parte de cerambícidos de pies moribundos o recién muertos por ataque de escolítidos es sumamente frecuente, y de hecho muchas asociaciones entre cerambícidos y escolítidos han sido identificadas hasta la fecha (CHANSLER 1967, MOSER et al. 1971, DAHLSTEN Y STEPHEN 1974, DIXON Y PAYNE 1979, DODDS et al. 2001). En este mismo sentido, algunas especies de escolítidos producen feromonas de agregación que permiten realizar ataques coordinados contra sus hospedadores. Asociado a ello, un conjunto de escarabajos saproxílicos pueden responder cairomonalmente a los infoquímicos de los escolítidos (ETXEBESTE et al. 2013b). Y son muchas las evidencias de la exitencia de estas respuestas semioquímicas de cerambícidos a los volátiles de pinos hospedantes así como a los atrayentes kairomonales producidos por escolítidos (BILLINGS Y CAMERON 1984, FATZINGER et al. 1987, SCHROEDER Y WESLIEN 1994, PAJARES et al. 2004, IBEAS et al. 2007, PAJARES et al. 2010). En concreto, el complejo de sustancias con mayor actividad atractiva para M. galloprovincialis incluye un compuesto cairomonal emitido por el hospedador (α-pineno) y dos compuestos de la feromona emitida por escolítidos del género Ips y Orthotomicus (ipsenol y metil-butenol) de acuerdo con PAJARES et al. (2004) e IBEAS et al. (2007), aunque existe controversia en el origen del metil-butenol, ya que se ha demostrado que

100
algunas especies de pino emiten dicho compuesto (SEYBOLD et al. 2006). Todo esto parece evidenciar una relación entre ambos grupos que va a tender a la coincidencia espacio-temporal en el floema y la región subcortical de los árboles muy decadentes o recién muertos. Con todo, estas interrelaciones entre las especies saproxílicas que colonizan las coníferas no se ajustan simplemente a una rudimentaria cadena trófica, sino que están revestidas de una mayor complejidad, formado redes reticulares que afectan a un mayor número de especies conectadas, donde los casos de omnivoría, predación-competencia intragremio, o compartimentalización de los recursos no son, en absoluto, aislados (AMEZAGA Y RODR GUEZ 1998, DODDS et al. 2001, AUKEMA Y RAFFA 2004, HUNTER 2009, ETXEBESTE et al. 2013b). Son, por tanto, muchas y muy complejas las interrelaciones que suceden en la región subcortical de las coníferas mediterráneas y su conocimiento sería de gran ayuda en la gestión de las plagas forestales desde la perspectiva del control integrado de plagas. El carácter genérico de las trampas y los atrayentes utilizados en las redes de trampas vigía, en general, puede hacer posible afrontar estudios para la caracterización y evaluación de estas comunidades (RASSATI et al. 2014, RASSATI et al. 2015a, RASSATI et al. 2015b). No obstante, la identificación y determinación de la diversidad entomológica es indiscutiblemente complicada, y paralelamente a la inventariación, se necesitan mecanismos ágiles y baratos para estimar y cuantificar la biodiversidad, la ecología o el medio ambiente. Un enfoque hacia la monitorización restringida de especies indicadoras, representantes o sustitutos de sus valores, puede ser uno de esos métodos (SAMWAYS 1994, NEW et al. 1995, MARTÍN-PIERA 2001). The Oxford Dictionary of Zoology define “especie indicadora” como aquella especie que posee una estrecha relación con respecto a uno o varios factores ambientales y, por tanto, cuando está presente, indica una condición medioambiental o un grupo de condiciones. Seleccionar correctamente aquellas especies se vuelve muy importante con la pretensión de que su estudio suponga una buena, barata y rápida aproximación de la realidad y ayude a conseguir una buena gestión del espacio natural. Ello dependerá de que satisfaga diversas condiciones: fácil muestreo y clasificación, distribución cosmopolita, adecuada representación en las muestras, abundancia significativa, facilidad y seguridad de almacenamiento, pertenencia a grupos taxonómicos bien conocidos, identificación rápida, distribución temporal y espacial predictible (para asegurar una continuidad a largo plazo), entre otras (MCGEOCH 1998). En referencia al muestreo de comunidades, de acuerdo con DE CÁCERES (2015), la determinación de la ocurrencia o la abundancia de un pequeño conjunto de especies indicadoras, como alternativa a la toma de muestras de toda la comunidad, ha sido particularmente útil en el monitoreo ambiental a largo plazo para la conservación o gestión ecológica. Las especies, por lo tanto, deberían ser elegidas como indicadores sólo si i) reflejan el estado biótico o abiótico del medio ambiente, ii) presentan pruebas de los efectos del cambio ambiental, o iii) predicen la diversidad de otras especies, taxa o comunidades dentro de un área. En definitiva, el objetivo de este trabajo es realizar una primera aproximación al estudio de la comunidad de insectos saproxílicos asociada a M. galloprovincialis mediante el análisis preliminar de los primeros datos aportados por la Red MUFFET.

101
Material y Métodos
Diseño experimental
Una red de muestreo sistemático de insectos saproxílicos (sensu lato) vinculados a los pinares mediterráneos fueron instalados en la Comunitat Valenciana, Este de la Península Ibérica, durante el año 2013. Esta red fue denominada Red de Muestreo Fitosanitario y Forestal de Ecosistemas Tipo (Red Muffet) de la Comunitat Valenciana. La red Muffet consistió en un conjunto de 15 parcelas (o estaciones Muffet) distribuidas a lo largo de toda el territorio de la Comunitat Valenciana. El área de estudio comprendió las masas forestales más representativas de las diferentes formaciones vegetales de pinar del este de la Península Ibérica. La distribución de las estaciones Muffet se realizó de manera aleatoria y estratificada, utilizándose la especie dominante y la altitud como variables moduladoras de la mayor parte de la variabilidad ambiental de los pinares a media escala en la Comunitat Valenciana. De esta forma, la variabilidad de las cinco especies de pinares del territorio valenciano (P. halepensis, P.pinea, P.pinaster, P.nigra y P.sylvestris) a sus diferentes altitudes fue cubierta por la red Muffet (Tabla 4.8, Figura 4.8 y Anejo 4.4)
Tabla 4.8 Características generales de las parcelas de muestre de la Red Muffet (otras características pueden ser cotejadas en el Anejo 4.4)
Parcela Localización Sp. principal Sp. acompañante Tipo1 Altura dom. Altitud Muffet 1 Guardamar del Segura P. pinea P. halepensis BM 4 11 Muffet 2 Crevillent P. halepensis BI 1 296 Muffet 3 Biar P. pinea P. halepensis BM 5 850 Muffet 4 Agres P. halepensis R 7 791 Muffet 5 El Saler P. halepensis P. pinea BM 10 2 Muffet 6 Puebla de San Miguel P. sylvestris P. nigra BM 10 1733 Muffet 7 Puebla de San Miguel P. nigra BM 10 1297 Muffet 8 Puebla de San Miguel P. pinaster BM 10 877 Muffet 9 Pobla Tornesa P. pinaster P. halepensis BM 8 360 Muffet 10 Penyagolosa P. sylvestris P. nigra BM 14 1524 Muffet 11 Penyagolosa P. nigra BM 10 1131 Muffet 12 Penyagolosa P. pinaster P. sylvestris BM 20 1471 Muffet 13 Aitana P. nigra P. pinaster R 6 1343 Muffet 14 Jérica P. halepensis RN 8 498 Muffet 16 Morella P. nigra BM 12 1005 1BM, bosque maduro; BI, Bosque en islas; R, repoblación.

102
Figura 4.8 Localización de las parcelas de la Red Muffet.
Para la captura de los insectos fueron utilizadas trampas de bandas cruzadas, modelo “crosstrap®” (Sanidad Agrícola ECONEX S.L., Siscar, Murcia, España). Éstas fueron cebadas con un compuesto atractivo para una amplia variedad de escarabajos perforadores de la madera (RASSATI et al. 2014). Este atrayente está compuesto por un dispensador de (-)α-pineno (velocidad de emisión de 300 mg/día), un dispensador de etanol (velocidad de emisión de 2000 mg/día) y un dispensador de ipsdienol (95.28%), ipsenol (4.7%) y (s)-(+)-cis-verbenol (0.02%). Todos los atrayentes fueron fueron provistos por la empresa Sanidad Agrícola Econex S.L. (Siscar-Santomera, Murcia, España) y SEDQ S.L. (Barcelona, España). La presión de muestreo fue mensual durante todo el periodo de estudio, y la captura de los insectos se realizó en húmedo, utilizando etilenglicol como conservante en el bote recolector. La diana de esta método de trampeo es la comunidad de saproxílicos sensu lato, de acuerdo con la definición propuesta en NIETO y ALEXANDER (2010), ampliada a los escolítidos y otros insectos floemofagos. Los individuos capturados fueron cribados e identificados en laboratorio. La temperatura fue controlada mediante el uso de un registrador continuo HOBO pendant UA-001-64 (Onset, USA). Cada registrador se programó para realizar una medida de temperatura del aire cada dos horas. El registrador se ubicó colgado de una rama, generalmente de matorrales, aproximadamente a 1 m de altura, intentando reducir al máximo la incidencia directa del sol. La pluviometría fue medida mediante el uso de un pluviómetro de cuchara modelo Rain-o-m100 (Onset, USA), conectado con un registrador HOBO pendant event UA-003-64 (Onset, USA).
Análisis estadísticos
El registro de temperaturas se utilizó para el cálculo de la acumulación de calor, tomando en cuenta un umbral de desarrollo mínimo de 12,2 ± 0.8ºC como aproximación general, basado en el modelo de desarrollo descrito para M. galloprovincialis en Portugal (NAVES Y DE SOUSA 2009). Las acumulaciones (grado-día) se calcularon utilizando el método de onda sinusoidal con una corrección intermedia para altas temperaturas, utilizando un umbral superior de 32 ºC (BASKERVILLE Y EMIN 1969, ALLEN 1976, ROLTSCH et al. 1999).

103
Para el estudio de comunidades, se utilizó la matriz de disimilitudes en base al cálculo de distancias de Bray-Curtis, al optimizar éste la variabilidad en relación a otros índices de disimilitud. Tal matriz se empleó para una ordenación de las parcelas, definidas como únicas para cada mes y localización, siguiendo el método de ordenación NMDS (Non-metric Multidimensional Scaling) y como resultado de 1000 permutaciones. Las variables ambientales de cada parcela se ajustaron a la ordenación resultante maximizando las correlaciones a los factores definidios en la ordenación. Se tuvieron en cuenta las siguientes variables: especie hospedadora principal, altitud dobre el nivel del mar, altura dominante de los pies, cobertura arbórea y total (medida como fracción de cabida cubierta, FCC), presencia relativa de las especies acompañantes (P. halepensis, P. nigra, P. sylvestris, P. pinaster, P. pinea, Q. Ilex, Q. coccifera, Eucalyptus sp, J. oxycedrus, J. thurifera, J. communis) (Anejo 4.4). Por otro lado, para seleccionar las especies que acompañan a M. galloprovincialis, e identificar las que son indicadoras de dicha comunidad acompañante fue realizado un análisis de especies indicadoras mediante la librería indicspecies del entorno R (DE CÁCERES 2015). Para la realización este análisis es necesario un vector que contenga una clasificación en grupos, que fue obtenido utilizando un análisis clúster no jerárquico (Non-hierárchichal clustering) con método de agrupamiento de k-medias y la optimización SSI (WEINGESSEL et al. 1999), utilizando la librería VEGAN del entorno R. La mejor partición, en este caso, se indica por el valor más alto SSI. Todos los cálculos y representaciones han sido realizados bajo el entorno de programación y lenguaje estadístico R (R CORE TEAM 2015).

104
Resultados
Resultados generales
Se capturaron 47696 ejemplares pertenecientes a 272 especies y a 61 familias distintas (Anejo 4.5). De las familias capturadas, el 57% corresponde a la familia Curculionidae (donde está incluido la subfamilia Scolitynae), el 8 % a Monotomidae, el 7% a Tenebrionidae, el 6% a Cerambycidae y Cleridae respectivamente y el 16% restante se corresponde a otras familias (Tabla 4.9, Figura 4.9 y Figura 4.10).
Tabla 4.9 Clasificación de familias de insectos capturados.
Familia Tot.individuos Nº especies
Familia Tot. Individuos Nº especies
Anthicidae 5 1 Laemophloeidae 104 2 Anthocoridae 227 1 Lampyridae 5 1 Anthribidae 12 1 Latridiidae 147 3 Aphodidae 1 1 Leiodidae 7 4 Apionidae 6 - Malachiidae 10 4 Blattidae 1 - Melandrydae 3 1 Bostrichidae 35 7 Meloidae 1 1 Bothrideridae 2 1 Melolonthidae 4 4 Buprestidae 78 6 Monotomidae 4015 3 Cantharidae 3 1 Mycetophagidae 211 6 Carabidae 22 1 Mycteridae 3 1 Cerambycidae 2819 6 Nitidulidae 405 6 Cetoniidae 4 3 Oedemeridae 11 1 Chrysomelidae 22 10 Pentatomidae 2 1 Cicadidae 77 1 Phalacridae 1 1 Ciidae 3 - Prionoceridae 32 1 Cleridae 2873 7 Ptinidae 324 19 Coccinellidae 24 8 Raphididae 9 1 Coreidae 4 1 Reduviidae 2 2 Cryptophagidae 135 3 Ripiphoridae 1 1 Curculionidae 27389 41 Salpingidae 24 3 Cydnidae 11 2 Scirtidae 1 1 Dasytidae 69 7 Scraptidae 17 2 Dermestidae 105 16 Silphidae 1 1 Elateridae 553 9 Siricidae 43 2 Endomychidae 1 1 Staphylinidae 1235 - Erotylidae 1 1 Tenebrionidae 3306 8 Eucinetidae 4 2 Throscidae 2 1 Eucnemidae 1 1 Trogossitidae 975 2 Histeridae 165 6 Zopheridae 2029 1 No colópteros 114 - Total 47696

105
Figura 4.9 Frecuencia de capturas por especie en la Red Muffet durante el año 2013. Sólo se representan las especies con capturas mayores de 2 individuos.

106
Figura 4.10 Frecuencia de capturas de especies por parcela de la Red Muffet durante el año 2013. Sólo se representan las especies con capturas mayores de 2 individuos.

107
Fenología del vuelo
En la Figura 4.11 pueden observarse los periodos de vuelo de todas las especies capturadas en la red Muffet el año 2013, a excepción de aquellas con un número de individuos capturados menor de dos. La acumulación de calor (grados–día) utilizando el modelo de onda sinusoidal basado en los umbrales de desarrollo descritos, para Monochamus galloprovincialis, por NAVES y DE SOUSA (2009) puede observarse en la Figura 4.12. En términos generales, la ¡Error! No se encuentra el origen de la referencia. muestra una evidente variación estacional del vuelo de varios conjuntos de especies. Un primer grupo, con vuelos claramente estivales o de primavera tardía, ocupa las posiciones centrales del eje de abcisas de la tabla. Un segundo grupo de especies, aquellas que se encuentran en las posiciones altas de la gráfica, presumiblemente van a tener un periodo de vuelo más tardío (más cercano al otoño). Por último, un tercer grupo de especies que se encuentran en la parte inferior del eje de abcisas de la tabla, y que serán aquellas que tengan periodos de vuelo más primaverales. Pese a que los análisis han sido realizados a partir de los primeros datos tomados por la Red MUFFET (instalada a finales de 2012), y no se dispone de una serie temporal de más de 2 años, que permitiría un análisis más fiable de los periodos de vuelo de los insectos diana, algunos rasgos generales son observables y, aunque exceden los objetivos del presente capítulo y deberán ser pospuestos para el futuro próximo análisis más profundos, merece la pena que sean comentados al objeto de describir someramente la comunidad de saproxílicos asociada a la especie M. galloprovincialis. Se puede observar como Oxypleurus nodieri (Mulsant, 1893) muestra un vuelo con el 50% de las capturas acumuladas a mediados del mes de abril. En este sentido, existen otras especies como Litargus connexus (Geoffroy, 1785), Pityophagus laevior (Abeille, 1872), y Acanthocinus aedilis (Linnaeus, 1758), que muestran también un vuelo temprano, con medianas a finales de mayo, o principios de junio. Otros insectos como Rhagium inquisitor (Linnaeus, 1758) y Hylastes attenuatus (Erichson, 1836) tienen las medianas de sus vuelos a mediados de junio, mientras que especies como Hylastes linearis (Erichson, 1836), y Xyleborus eurygraphus (Ratzeburg, 1837) las obtienen a finales del mes de junio. En el mes de julio, se obtienen las medianas de las capturas de Hylurgus ligniperda (Fabricius, 1787) y de Xyleborus eurygraphus (Ratzeburg, 1837), Crypturgus mediterraneus (Eichhoff, 1871), Hylastes ater (Paykull, 1800) o Hylurgus ligniperda (Fabricius, 1787). Los depredadores Aulonium ruficorne (Olivier, 1790), Cryptolestes ferrugineus (Stephens, 1831), y Temnoschelia coerulea (Olivier, 1790) tienen el 50% de sus capturas también a finales del mes de julio. Orthotomicus erosus (Wollaston, 1857), Acanthocinus griseus (Fabricius, 1792), y Monochamus galloprovincialis (Olivier, 1795) presentan el 50% de las capturas en el mes de agosto, al igual que otras muchas especies como Ips sexdentatus (Börner, 1776), Spondylis buprestoides (Linnaeus, 1758), Thanasimus femoralis (Zetterstedt, 1828), Rhizophagus ferrugineus (Paykull, 1800), Arhopalus rusticus (Linnaeus, 1758) y Arhopalus syriacus (Reitter, 1895).

108
En el mes de septiembre es donde se contabilizan mayor número de insectos con periodos del 50% de las capturas acumuladas: Acanthocinus hispanicus (Sama & Schurmann, 1980), Hylurgus miklitzi (Wachtl, 1881), Ips mannsfeldi (Wachtl, 1879), Pityogenes calcaratus (Eichhoff, 1878), Pogonocherus perroudi (Mulsant, 1839), Paromalus parallelepipedus (Herbst, 1792), Rhizophagus depressus (Fabricius, 1792), Corticeus pini (Panzer, 1799) o Arhopalus ferus (Mulsant, 1839). En referencia a los casos concretos de especies de un mismo género, los casos más destacables son los siguientes. Las tres especies del género Arhopalus registradas en este estudio tienen vuelos similares, en los meses de verano: Arhopalus ferus (Mulsant, 1839), Arhopalus rusticus (Linnaeus, 1758) y Arhopalus syriacus (Reitter, 1895) registran capturas prácticamente simultaneas. En cambio, las especies el género Thanasimus muestran diferencias temporales en el periodo de vuelo: el periodo de vuelo de T. formicarius es propio de primavera-verano, mientras que el de T. femoralis se da algo más retrasado. Y las especies el género Rhizophagus vuelan prácticamente sin solapadamiento, con un pico invernal en el caso de R. depressus y con un pico estival en el caso de R. ferrugineus.

109
Figura 4.11 Periodos de vuelo de las especies capturadas en la Red Muffet durante el año 2013. Sólo se representan las especies con capturas mayores de 2 individuos (Anejo 4.6 y Anejo 4.7). La línea interior del diagrama de
caajas representa la mediana de las capturas. La fecha está indicada en días julianos.

110
Figura 4.12 Acumulación de calor para el periodo de vuelo de las especies capturadas tomando los umbrales de desarrollo propuestos por Naves et al (2009) para M. galloprovincialis. Sólo se representan las especies con capturas mayores de 2 individuos (Anejo 4.6 y Anejo 4.7). La fecha está indicada en días julianos

111
Análisis de comunidades y especies indicadoras
El análisis de comunidades ha sido realizado exclusivamente para las especies coincidentes temporalmente con M. galloprovincialis, por lo que sólo han sido tenidas en cuenta las especies capturadas entre los meses de junio a octubre, ambos incluidos. El Escalamiento Multidimensional (NMDS, Non-metric Multidimensional Scaling) realizado con el objetivo de ver qué variables medioambientales (Anejo 4.4) podrían estar relacionadas con las diferentes especies y abundancias de coleópteros saproxílicos capturadas en cada una de las parcelas Muffet en el año 2013 mostró un valor del stress fue de 0,193, por lo que la información aportada por el análisis es relativamente baja, es decir, se observó una baja disimilaridad (FAITH et al. 1987) (Figura 4.13).
Figura 4.13 Ordenación NMDS (1000 permutaciones, ajuste a la 66esima iteración) de las variables relacionadas
con la abundancia de las diferentes especies capturadas en la Red MUFFET en el año 2013 La Tabla 4.10 muestra el peso medio por especie en la ordenación NMDS de las muestras, junto con los vectores de máxima correlación de las variables ambientales de las parcelas tras 1000 permutaciones:

112
Tabla 4.10 Ordenación NMDS con 1000 permutaciones
MDS1 MDS2 r2 Pr(>r) Grados día acumulados -0.99648 -0.08388 0.5407 0.000999 *** Altura 0.92594 -0.37767 0.5592 0.000999 *** FFC 0.99943 0.03372 0.1874 0.000999 *** P. halepensis -0.87797 0.47871 0.6676 0.000999 *** P. nigra 0.83068 -0.55674 0.3399 0.000999 *** P. pinea -0.70747 0.70674 0.2235 0.000999 *** P. sylvestris 0.74270 -0.66962 0.1960 0.000999 *** Q. ilex 0.84415 -0.53611 0.0994 0.024975 * Eucalipto -0.97335 -0.22934 0.2061 0.000999 ***
La Figura 4.13 muestra que la comunidad de saproxílicos estudiada parece muy homogénea, y aunque existan factores ambientales que generen gradientes, el peso medio específico de ellos en la composición faunísitca es muy bajo. La centralidad de M. galloprovincialis en el gráfico, y el rango de los ejes, es señal concreta de ello. Aún así, uno de los mayores gradientes observables es el generado por el trinomio, con alta colinealidad, formado por la altitud sobre el nivel del mar (msnm), los grados día acumulados a la fecha de muestreo, y la proporción de P. halepensis en el punto de muestreo. La colinealidad viene dada porque los pinares de P. halepensis ocupan las regiones más cálidas y, a su vez, más degradadas del territorio valenciano, mientras que los ecosistemas alpinos del interior de la región, a mayor altura y ocupados principalmente por pino laricio y silvestre se caracterizan por tener coberturas mayores. Utilizando el gradiente de P. halepensis como referencia se puede obtener la siguiente ordenación (Figura 4.14), en la que se muestra la posición central de M. galloprovincialis (y no sólo de esta especie, sino de la gran mayoría) y la mayor variabilidad de las parcelas. Es decir, se muestra que aunque la variabilidad de las parcelas de estudio es alta, la de las comunidades de colópteros saproxílcos capturados es muy baja.
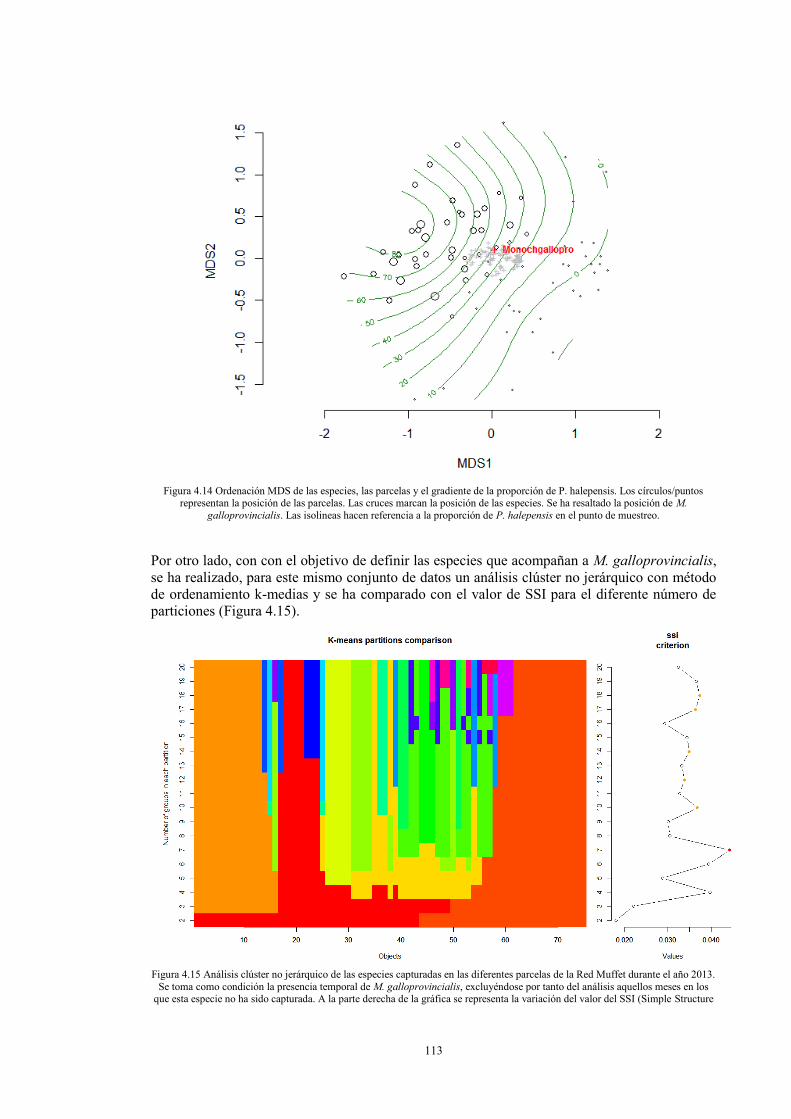
113
Figura 4.14 Ordenación MDS de las especies, las parcelas y el gradiente de la proporción de P. halepensis. Los círculos/puntos
representan la posición de las parcelas. Las cruces marcan la posición de las especies. Se ha resaltado la posición de M. galloprovincialis. Las isolineas hacen referencia a la proporción de P. halepensis en el punto de muestreo.
Por otro lado, con con el objetivo de definir las especies que acompañan a M. galloprovincialis, se ha realizado, para este mismo conjunto de datos un análisis clúster no jerárquico con método de ordenamiento k-medias y se ha comparado con el valor de SSI para el diferente número de particiones (Figura 4.15).
Figura 4.15 Análisis clúster no jerárquico de las especies capturadas en las diferentes parcelas de la Red Muffet durante el año 2013.
Se toma como condición la presencia temporal de M. galloprovincialis, excluyéndose por tanto del análisis aquellos meses en los que esta especie no ha sido capturada. A la parte derecha de la gráfica se representa la variación del valor del SSI (Simple Structure

114
Index), cuto valor máximo marca el número de particiones óptima del análisis de conglomerados. Puede observarse que el número de bloques o grupos óptimos, marcado por el valor máximo de SSI, es de siete. A partir del SSI óptimo, el análisis de especies indicadoras con 7 clusters realizado mediante la librería indicspecies package (DE CÁCERES 2015) mostró las especies que podrían funcionar como indicadoras de algunos de los grupos (Tabla 4.11) o combinaciones de grupos (Tabla 4.12). De todas maneras, M. galloprovincialis no se asoció a ningún grupo en particular (y en cualquier caso nunca significativamente), al igual que A. griseus (Anejos 4.8 y 4.9) Tabla 4.11 Composición y especies indicadoras (significativamente) de los grupos derivados del análisis clúster realizado sobre las
especies capturadas en las parcelas de la Red MUFFET en el año 2013, excluyendo del análisis los meses en los que M. galloprovincialis no ha sido capturado.
Grupo Parcelas/Mes Especies Indicadoras 1 Muf13-Sep, Muf16-Sep, Muf9-Oct, Muf13-Oct, Muf16-Oct
-
2 Muf3-Jun, Muf4-Jun, Muf8-Jun, Muf14-Jun, Muf3-Jul, Muf8-Jul, Muf3-Aug, Muf7-Aug, Muf8-Aug,Muf6-Sep, Muf7-Sep, Muf8-Sep, Muf3-Oct,Muf6-Oct, Muf7-Oct,Muf8-Oct
Hylurgus ligniperda, Hymenorus doublieri
3 Muf6-Jun, Muf9-Jun, Muf10-Jun, Muf11-Jun, Muf12-Jun, Muf13-Jun, Muf16-Jun, Muf13-Jul
Hylaster attenuates, Hylobius abietis, Pityophagus laevior, Acanthocinus aedilis
4 Muf5-Jun, Muf2-Jul, Muf5-Jul Pityogenes calcaratus, Brachytemnus porcatus, Chrysobothris solieri
5 Muf6-Jul, Muf7-Jul, Muf10-Jul, Muf11-Jul, Muf12-Jul, Muf6-Aug, Muf10-Aug, Muf11-Aug, Muf12-Aug, Muf10-Sep, Muf11-Sep, Muf12-Sep, Muf10-Oct, Muf11-Oct, Muf12-Oct
Ips sexdentatus, Ips mannsfeldi
6 Muf7-Jun, Muf9-Jul, Muf16-Jul, Muf9-Aug, Muf13-Aug, Muf16-Aug, Muf3-Sep
Arhopalus syriacus, Lacon punctatus
7 Muf1-Jun, Muf2-Jun, Muf1-Jul, Muf4-Jul, Muf14-Jul, Muf1-Aug, Muf2-Aug, Muf4-Aug, Muf5-Aug, Muf14-Aug, Muf1-Sep, Muf2-Sep, Muf4-Sep, Muf5-Sep, Muf9-Sep, Muf14-Sep, Muf1-Oct, Muf2-Oct, Muf4-Oct, Muf5-Oct, Muf14-Oct
-

115
Tabla 4.12 Especies indicadoras (significativamente asociadas) de las combinaciones de grupos derivados del análisis clúster realizado sobre las especies capturadas en las parcelas de la Red MUFFET en el año 2013, excluyendo del análisis los meses en los que M. galloprovincialis no ha sido capturado.
Grupo Especies Indicadoras 3+6 Rhagium inquisitor, Oxypleurus nodieri 5+6 Thanasimus femoralis 6+7 Arhopalus ferus 1+3+5 Rhizophagus depressus 2+4+7 Hylurgus miklitzi, Xyleborinus saxesenii 2+5+6 Spondylis buprestoides 2+3+5+6 Temnoscheila coerulea 2+3+4+5+7 Orthotomicus erosus, Aulonium ruficorne

116
Discusión General
Los resultados presentados suponen el aporte de una lista de 298 especies de coleópteros saproxílicos para las regiones orientales de la Península Ibérica. De estas especies, 95 y 156 fueron exclusivas de Murcia y Valencia respectivamente, y únicamente 46 especies fueron descritas en ambas regiones. Estos resultados, en sí mismo, constituyen una contribución muy valiosa en términos de registro de la diversidad biológica en los pinares ibéricos (NIETO Y ALEXANDER 2010). En consonancia con los resultados de RASSATI et al. (2014), RASSATI et al. (2015a), RASSATI et al. (2015b) la metodología de muestreo, compuesta por una trampa de interceptación de vuelo cebada con atrayentes formados por α-pineno, etanol, ipsdienol y cis-verbenol, muestra ser un sistema eficaz de captura y un cebo de muy amplio espectro para las especies de insectos saproxílicos forestales mediterráneas, de manera que las especies capturadas representan una parte muy importante de la diversidad de éstos en los pinares del este y sureste ibérico. En lo que se refiere al estudio de la comunidad asociada a de M. galloprovincialis, los primeros análisis de nuestros resultados muestran que se trata de una especie ubicua, presente en toda la variabilidad medioambiental de los pinares del este y sudeste ibérico, sin que sea posible asociarla significativamente a alguna comunidad de coleópteros saproxílicos (Anejo 4.9), mientras que otras especies, sin embargo, sí parecen estar asociadas a diferentes ecosistemas (Tabla 4.11). Es decir, el rol que M. galloprovincialis juega en las comunidades de entomofauna de los ecosistemas en los que está presente es el mismo a lo largo de todo el gradiente estudiado. Aún así los resultados apuntan a la existencia de una posible relación de especial importancia entre el escolítido O. erosus y M. galloprovincialis similar a las establecidas entre otras especies del género Monochamus y otras especeies de escolítidos, como los binomios M. carolinensis - Ips calligraphus o M. titillator - Dendroctonus frontalis (COULSON Y WITTER 1984, MILLER 1984, 1985, FLAMM et al. 1989, DODDS et al. 2001). La relación entre ambas especies, no obstante, deberá ser objeto de estudios más concretos y de mayor profundidad en el futuro. Asimismo, además de ser una especie de muy amplia distribución en la zona de estudio, los análisis realizados desde la perspectiva del estudio de las redes complejas de interacciones muestran que M. galloprovincilis se trata de una especie clave en muchos de los ecosistemas analizados ya que actúa como conector fundamental entre las distintas subredes que forman las especies de insectos saproxílicos presentes en cada uno de los ecosistemas, facilitando las interacciones entre los diferentes componentes de las redes. La utilización de las redes de interacciones como método de análisis de la biodiversidad y de la estructura de las comunidades muestra ser una herramienta interesante, y potencialmente muy útil para el desarrollo de un indicador de cambios en la calidad forestal a corto plazo. En el caso de los datos analizados en la región de Murcia se aprecia que hay ciertas redes, como las correspondientes a las ESFP 15, 9 y 19, que presentan densidades de conectividad altas, con un máximo de 0,25 (Tabla 4.7). El menor valor de conectividad es 0,11, estando la mayor parte de valores de densidad entre este último y 0,2. Esto indica que se trata de redes con poca saturación de interacciones y que probablemente respondan bien a la realidad, ya que las especies han evolucionado separando sus nichos ecológicos, y desarrolando mecanismos eficaces para evitar la competencia (DODDS et al. 2001, AUKEMA Y RAFFA 2004, HUNTER 2009). En el número de conexiones también se aprecia un patrón parecido, ya que las ESFPs 15, 10, 14 y 6 poseen más de 400 interacciones, destacando la ESFP15 con 710 interacciones. La ESFP

117
con menor número es la 2, con tan solo 71 conexiones. El resto se sitúan entre las 105 y 358 interacciones. Esto vuelve a incidir en que se trata de redes poco saturadas (Tabla 4.7). Respecto a las especies conectoras, es decir, especies que hacen de puente entre distintas subredes o partes de la red, destaca por su redundancia M. galloprovincialis, conector en 6 estaciones de la Red ESFP (11, 12, 13, 17, 19, 20), la mitad de ellas en exclusividad. La siguiente especie en importancia como conector es el escolítido O. erosus, conector en 3 de las redes (2, 3 y 16) (Tabla 4.7). Estas dos especies podrían tener el valor de ser especies clave en casi la mitad de las estaciones forestales, ya que facilitarían mucho las interacciones entre los diferentes componentes de la red. La desaparición de estas especies podría desestructurar el sistema con efectos poco predecibles (BELLINGERI Y BODINI 2013). De los resultados obtenidos en la Red MUFFET, se han obtenido 115 curvas de vuelo, que muestran la variación de la fenología del vuelo de las diferentes especies capturados en la dicha red de muestreo a lo largo del año 2013 (Figura 4.11). Los vuelos de las distintas especies se van solapando en una continua sucesión de periodos de vuelo, sin vacíos entre ellos. En este punto debe destacarse el inconveniente real de que el periodo de muestreo sea inferior a los dos años. Los días ordinales se empiezan a contar el 1 de enero y en ese momento el vuelo del grupo de especies de vuelo otoñal-invernal ya ha empezado, por tanto, la contabilización de los grados-día acumulados para que se produzca el vuelo se produce de manera desfasada. Es decir, en estas situaciones, el gráfico induce claramente a un error en su interpretación, como en el caso de Tomicus destruens o Rhyzophagus depressus. En análisis futuros, la fecha deberá ser tratada como una variable circular que permita una mejor interpretación de todos los datos. En el caso concreto de M. galloprovincialis, la observación de su periodo de vuelo y su acumulación de grados-día lo sitúan como una especie de vuelo estival, de acuerdo con lo expuesto en la bibliografía (NAVES et al. 2007, NAVES et al. 2008) y, por tanto, coincidente fenológicamente con la mayoría de las especies de la comunidad de insectos saproxílicos de la zona de estudio, por lo que, al menos, es posible que exista interacción entre ellas (Figura 4.11 y Figura 4.14). Estos resultados son coincidentes con los periodos de vuelo mostrados en los resultados del Capítulo 3 de este mismo trabajo. Esta amplia comunidad de insectos saproxílicos con la que coincide fenológicamente el vuelo de M. galloprovincialis ha sido analizada desde una perspectiva de análisis de comunidades teniendo en cuenta las diferentes variables de los ecosistemas del área de estudio, que representan toda la variabilidad medioambiental de los pinares de la Comunitat Valenciana. El análisis NMDS, ceñido a las especies fenológicamente coincidentes con M. galloprovincialis, mostró que la comunidad de saproxílicos estudiada es muy homogénea respecto a la variación ambiental de las zonas de muestreo. La centralidad de M. galloprovincialis en el gráfico de la Figura 4.13 es señal de que, aunque existen factores ambientales que generen gradientes, el peso medio específico de ellos es muy bajo. El posterior análisis de la posición de M. galloprovincialis con respecto a los gradientes ambientales más relevantes (en concreto el conjunto formado por la altitud sobre el nivel del mar, los grados día acumulados a la fecha de muestreo, y la proporción de P. halepensis en el punto de muestreo) mostró que M. galloprovincialis es, es una especie ubicua en el área de estudio, con presencia en toda la variabilidad medioambiental de los pinares de la región, desde el nivel del mar hasta las altitudes superiores a los 1500 msnm (Figura 4.14), Esto está en consonancia con la amplia distribución geográfica y ambiental observada a nivel global para M. galloprovincialis (VIVES 2000). En esta misma figura, de nuevo, se muestra que, en términos generales, que la variabilidad de las comunidades de colópteros saproxílcos capturados es muy baja, a pesar de la mayor variabilidad de las zonas de estudio.

118
Las relaciones de M. galloprovincialis con el resto de insectos con los que interacciona en el ecosistema apenas son conocidas (KENIS Y HILSZCZANSKI 2004). Del análisis de conglomerados realizado para el estudio de la comunidad de coleópteros saproxílicos posiblemente asociada a M. galloprovincialis (Figura 4.15, Tabla 4.10 y Tabla 4.11) merece la pena apuntar algunas de las observaciones más interesantes. Este análisis muestra varios grupos de interés que aparecen determinados significativamente por la captura de algunas especies en periodos concretos del año. Uno de los grupos más interesantes es el grupo 6, que engloba los muestreos de verano y otoño en los ecosistemas de montaña, que en la región oriental de la península ibérica están dominados por pinares de P.sylvestris y P. nigra. Una de las especies indicadora de este grupo, junto a I. sexdentatus, es I. mannsfeldi, que es una especie rara asociada al hábitat natural del P. nigra. Este grupo está relativamente asociado al grupo 3, otro grupo que engloba el muestreo primaveral de zonas de ecosistemas de montaña, en concreto, de una de las zonas con mayor interés en términos de biodiversidad de todo el levante peninsular, que es el macizo de Penyagolosa (Muf10, Muf11 y Muf12) (CITMA 2015 -a, b) Por último, el grupo 7 engloba la mayoría de las parcelas más cálidas, ocupadas en su totalidad por P. halepensis, aun presentando especies acompañantes como el P.pinea o el P.pinaster (Tabla 4.14). En resumen, este análisis apunta a la posibilidad de que existan hábitats relativamente diferenciados por la estructura de sus comunidades de insectos saproxílicos; y aunque el corto periodo de muestreo analizados no permite mayor claridad en el análisis, es esperable que análisis futuros de series temporales lo suficientemente largas permitan sacar conclusiones más sólidas. No obstante esto, los análisis de conglomerados realizados sobre los datos de capturas sí que mostraron, nuevamente, que las comunidades de saproxílicos del Este peninsular son muy homogéneas y que M. galloprovincialis no pudo ser asociado a ningún grupo o bloque en particular. En un primer análisis teniendo en cuenta exclusivamente las especies significativamente asociadas a los clusters, M. galloprovincialis no aparece como tal vinculada a ningún bloque ni a ningún conjunto de bloques (Tabla 4.10 y Tabla 4.11). Sólo después de un análisis teniendo en cuenta todas las especies (aunque su relación con los diferentes grupos no sea significativa) es posible asignarla a un conjunto de bloques que agrupa a la gran mayoría de ellos (Anejo 4.9). De esta forma, los resultados muestran que M. galloprovincialis sólo aparece ligado a una comunidad de saproxílicos cuando ésta reúne a todas las especies contenidas en 6 de los 7 grupos o bloques iniciales del clúster (Stat 0,838, p=0,17), es decir, todos menos el grupo 4, lo que equivale a decir que se trata de una especie de amplia distribución, presente en la gran mayoría de las parcelas de muestreo. Y además, debe tenerse en cuenta que el grupo 4 lo forman muy pocos muestreos y ceñidos a los meses de junio y julio, en las parcelas 2 y 5, parcelas muy cálidas en las que M. galloprovincialis ha sido capturado al menos en la parela Muffet5 en muestreos posteriores (resultados de 2014 y 2015 no mostrados), por lo que es esperable que, teniendo en cuenta los datos aportados por más años de muestreo, esta especie sea también capturada en dichas parcelas y en dichos periodos demostrando su total ubicuidad en el área de estudio. Asimismo, esta mima situación se da con otra especie de cerambícido, A. griseus (Anejo 4.9). De acuerdo con BONIFÁCIO (2009), estas dos especies, junto al también cerambícido A. syriacus, fueron los cerambícidos más frecuentes en la colonización de los árboles muertos por como consecuencia de la Enfermedad del Marchitamiento del Pino en la Península de Setúbal (Portugal) tras la detección de B. xylophilus en 1999, mostrando patrones de distribución en el tronco que revelaron comportamientos de exclusión interespecífica, a fin de reducir la competencia de sus larvas por los recursos disponibles (YOSHIKAWA 1987). De esta manera, A. syriacus tuvo tendencia a ocupar las partes basales de los troncos, A. griseus las partes intermedias y M. galloprovincialis las partes superiores de los fustes (BONIFÁCIO 2009). Este comportamiento de exclusión interespecífica está en consonancia con la coincidencia de sus

119
curvas de vuelo observada en nuestros resultados (¡Error! No se encuentra el origen de la referencia.). Un desfase entre sus fenologías de vuelo podría derivar en que la distribución de la colonización del tronco del árbol estuviese correlacionada en mayor medida con la secuencia temporal de llegada al fuste (es decir, con el periodo de vuelo reproductivo) más que con las preferencias concretas del material de cría para cada especie, tal y como sucede en la competencia interespecífica de otras especies de perforadores (AMEZAGA Y RODR GUEZ 1998). Por otro lado, la especie de escolítidos más coincidente con M. galloprovincialis en la región subcortical de los árboles muertos entre 2000 y 2007 como consecuencia de la PWD en la Península de Setúbal (Portugal) fue O. erosus (BONIFÁCIO 2009). Y esta situación no es, en absoluto, exclusiva de ese episodio. En efecto, la gestión de los focos de perforadores surgidos en la Comunitat Valenciana como consecuencia de la intensa sequía sufrida en el levante de la Península Ibérica desde la primavera de 2013, que provocó la muerte de cientos de miles de pies de P. halepensis y P. pinaster, supuso la colocación de centenares de trampas cebadas con atrayentes cairomonales y feromonales específicos para la gestión de O. erosus durante los meses de verano de 2014 y 2015, por parte del gobierno de la Generalitat Valenciana. El muestreo de estas trampas, mostró abundantes capturas de M. galloprovincialis (Pérez-Laorga, Com. Pers.). La asociación entre especies del género Monochamus y escolítidos ha sido estudiada en otras regiones y para otras especies: en el sur de EEUU, las altas poblaciones de M. titillator suelen encontrarse asociadas a poblaciones de D. frontalis en árboles muertos como consecuencia de ataques de los escolítidos. Entre ambas especies, estas interacciones han sido descritas como de “competencia” (COULSON et al. 1976), aunque otros estudios posteriores han sugerido que dicha interacción podría ser definida como comensalismo (FLAMM et al. 1989). De igual forma MILLER (1984) sugirió que la competencia de M. titilator e I. calligraphus por los recursos del floema podía tener un efecto negativo sobre algunos depredadores subcorticales de éste último. Posteriormente DODDS et al. (2001) mostraron que también existía interacción entre M. carolinensis e I. calligraphus, una relación en la que el primero de ellos actuaba como depredador facultativo de las larvas del segundo y, por tanto, podía ser considerado como un depredador intragremio durante los periodos de desarrollo subcortical, antes de su etapa xilofágica. La depredación intragremio podría reportar numerosos beneficios al depredador facultativo en el sentido de que se trata de una interacción compleja que contempla tanto la competencia como la depredación (POLIS et al. 1989). En primer lugar, la depredación supondría un aporte nutritivo importante en caso de recursos limitados o de bajos contenidos nutritivos en el floema (SLANSKY JR 1992). Esta mejora en la calidad nutricional puede tener una ventaja adaptativa para los cerambícidos por la aceleración del desarrollo larvario y el consecuente acortamiento de dicha fase (HELLRIGL 1971). Pero además de ello, la depredación intragremio supone también la reducción de competencia por el recurso limitado del floema, aunque también del número de depredadores presentes en el interior del floema (MILLER 1985). No obstante, esta capacidad de actuar como depredador facultativo, muy probablemente, sea dependiente del tamaño de las larvas y, por tanto, la propia condición cambie con el desarrollo y el crecimiento de las mismas. DODDS et al. (2001) mostró que los primeros estadios larvarios de M. carolinensis, los de menor tamaño, no depredaban con tanta frecuencia larvas de I. calligraphus como sí lo hacían las de mayor tamaño, de estadios larvario más avanzados. POLIS et al. (1989) sugirieron que la depredación intragremio podría reducir o eliminar drásticamente las poblaciones locales de presas y que podría ser importante en el declive de las poblaciones de D. frontalis conforme a una depredación intragremio dependiente de la densidad y a lo largo de varias generaciones de M. titilator. Sea como fuere, las interacciones que se

120
producen subcorticalmente entre cerambícidos y escolítidos son poco conocidas en términos generales, y casi completamente desconocidas para el caso concreto de M. galloprovincialis. No obstante, AKBULUT y STAMPS (2012) encontraron similitudes notables entre M. carolinensis y M. galloprovincialis en muchas de sus características biológicas, como la aparente supervivencia desde el huevo hasta la larva, la supervivencia generacional y el número de adultos emergidos. Y otros estudios, además, revelaron resultados similares también entre M. carolinensis y el vector del NMP de Asia Oriental, M. alternatus, lo que sugiere que todas estas especies podrían tener patrones de ciclos biológicos similares (NAVES et al. 2006, NAVES et al. 2008, AKBULUT Y STAMPS 2012) y que la investigación sobre una especie podría ser ampliamente aplicable a todas las demás especies del género. Según esto, efectivamente, nuestros resultados, aunque enmarcados en un análisis de comunidades mucho más generalizado, parecen mostrar una evidente relación entre ambas especies. La ordenación por bloques realizada sobre los datos obtenidos por la red MUFFET en 2013 mostraron que O. erosus es también una especie ubicua, vinculada finalmente a otras especies de amplia distribución y muy comunes en la zona de estudio, como Aulonium ruficorne, Cryptolestes ferrugineus, M. galloprovincialis o A. griseus (Anejos 4.8 y 4.9). A su vez, los resultados de la Red de EFSP de Murcia destacaron a ambas especies, O. erosus y M. galloprovincialis, como las especies clave en la conexión interna de las redes (conector) en muchas de las estaciones de estudio y, en algunas ocasiones, conjuntamente (Tabla 4.7). En el análisis de las curvas de vuelos realizado con los datos recogidos por la Red MUFFET puede observarse que la curva de vuelo de O.erosus aparece adelantada con respecto a la de M. galloprovincialis (Figura 4.13). La estrategia para sobrevivir al invierno por ambas especies es diferente, ya que M. galloprovincialis pasa este periodo en estado larvario, mientras que O. erosus lo hace en el estado adulto, refugiado en las resquebrajaduras de la corteza e incluso dentro de antiguas galerías (MENDEL Y HALPERIN 1982). La presencia de adultos de O. erosus en el periodo invernal conlleva la posible extensión de su periodo de vuelo a lo largo de todo el año si se dan temperaturas benignas en inviernos muy suaves ya que el rango de temperaturas dentro del cual puede producirse el vuelo de esta especie es muy amplio, entre los 12ºC y los 38ºC (MENDEL Y HALPERIN 1982). No obstante, la situación normal en la región oriental de la península ibérica es que exista una parada invernal del vuelo y que éste tenga un pico adelantado unos meses con respecto al de M. galloprovincialis. Esta situación de desfase en el vuelo y, por tanto, en la llegada de las poblaciones de ambas especies a la colonización de los árboles debilitados, podría suponer cierta sincronización temporal de manera que la emergencia y el vuelo de M. galloprovincialis tenga lugar en un periodo del año en el que O. erosus ya haya accedido a los árboles más debilitados, acelerando su proceso de decadencia y generando, de esta manera, una mayor disponibilidad de material hospedante adecuado para la reproducción de M. galloprovincialis, teniendo en cuenta, además, los beneficios reportados por la predación intragremio ya comentados líneas arriba.
(1998) demostraron la existencia de compartimentalización de los recursos alimenticios, floema en este caso, entre diferentes especies de escolítidos ibéricos, entre ellos, O. erosus, debida entre otros factores al espesor de la corteza. La compartimentalización de los nichos y los recursos ha sido frecuentemente considerada como un mecanismo para minimizar la competición interespecífica (SCHOENER 1974). Los resultados de estos autores mostraron además que la secuencia de llegada al hospedante es significativa a la hora de seleccionar de los nichos más adecuados por parte de las especies pioneras.

121
En este mismo sentido, también los compuestos semioquímicos de agregación pueden jugar un papel importante en la compartimentalización del hábitat entre los diferentes perforadores de la madera (BOUHOT et al. 1988), y la respuesta positiva por parte de M. galloprovincialis a componentes fundamentales de la feromona de O. erosus, como 2-metil-3-buten-2-ol o ipsdienol es una nueva prueba de la interacción entre ambas especies (GIESEN et al. 1984, HUERTA et al. 2004, SEYBOLD et al. 2006, IBEAS et al. 2007, PAJARES et al. 2010, ETXEBESTE et al. 2013b) aunque existe controversia en el origen del 2-metil-3-buten-2-ol ya que se ha demostrado que algunas especies de pino emiten dicho compuesto (SEYBOLD et al. 2006). En este caso, ambas especies tienen preferencia por el mismo espesor de corteza y por partes del árbol similares a la hora de colonizar los árboles debilitados (AMEZAGA Y RODR GUEZ 1998, BONIFÁCIO 2009), por lo que parece razonable pensar que pueda existir una adaptación o sincronización del ciclo de M. galloprovincialis al de O. erosus al objeto de que el cerambícido, tanto en su condición de xilófago como en su condición de posible depredador facultativo, pudiera obtener beneficios de las altas poblaciones previas de O. erosus ya instaladas en los árboles hospedadores. En resumen, hay indicios que hacen pensar en la existencia una fuerte interacción entre estas dos especies, que debería ser estudiada en futuras investigaciones con el objetivo de afinar con mayor precisión la gestión de sus poblaciones. Y no obstante, O. erosus aparece frecuentemente asociado a otros escolítidos (MENDEL Y HALPERIN 1982, AMEZAGA Y RODR GUEZ 1998, BONIFÁCIO 2009, ETXEBESTE et al. 2013b). Por lo que las relaciones entre M. galloprovincialis con estos otros escolítidos podría ser también razonable. (1998) mostraron el solapamiento espacial y temporal existente los nichos de O. erosus e Ips sexdentatus, y ETXEBESTE et al. (2013b) sugirieron la existencia de interacción entre ambas especies, apuntando a la posibilidad de que se diesen entre ambas ciertos comportamientos de exclusión interespecífica basándose en la mayor tolerancia de O. erosus a la verbenona, que posee un demostrado poder inhibitorio para I. sexdentatus(PAIVA et al. 1988, KOHNLE et al. 1992, ROMÓN et al. 2007, ETXEBESTE Y PAJARES 2011, ETXEBESTE et al. 2013a);. O. erosus es una especie menos agresiva que I. sexdentatus en la península ibérica (GIL Y PAJARES 1986) siendo la primera considerada en la mayoría de los foros de gestión como una especie poco importante, con un carácter fundamentalmente secundario, y cuyas poblaciones tienen grandes incrementos tras periodos de sequía. En cambio, I. sexdentatus es considerada una especie primaria capaz de provocar grandes mortandades en arbolado sano cuando sus poblaciones aumentan de forma epidémica (ETXEBESTE Y PAJARES 2011, ETXEBESTE et al. 2013a). La mayor parte de la zona de estudio está ocupada por formaciones vegetales de P. halepensis, especie que no es hospedador de I. sexdentatus, por lo que las capturas de esta especie quedan relegadas a los ecosistemas de altitud del interior de la Comunitat Valenciana y Murcia (con P.nigra y/o P.sylvestris), de las cuales parece ser una especie indicadora (Tabla 4.11), y a las fragmentadas formaciones de P. pinaster del levante de la península ibérica. Por ello, las posibles relaciones entre la fenología o la pertenencia a similares comunidades de saproxílcos de I. sexdentatus y M. galloprovincialis no pueden ser inferidas en este estudio. Sí resultaría interesante ahondar en el estudio de las posibles interacciones entre M. galloprovincialis y O. erosus. Y sería de similar interés conocer la existencia de equivalentes interacciones entre M. galloprovincialis e I. sexdentatus en los ecosistemas en los que éste compite con ventaja por su carácter primario frente a O. erosus y, por tanto, la posibilidad de interacción con el cerambícido es mayor.

123
Bibliografía
AKBULUT, S. y W. T. STAMPS (2012). Insect vectors of the pinewood nematode: a review of
the biology and ecology of Monochamus species. Forest Pathology 42(2): 89-99. ALLEN, J. C. (1976). Modified Sine Wave Method for Calculating Degree Days.
Environmental Entomology 5(3): 388-396. (1998). Resource partitioning of four sympatric bark
beetles depending on swarming dates and tree species. Forest Ecology and Management 109(1): 127-135.
AUKEMA, B. H. y K. F. RAFFA (2004). Does aggregation benefit bark beetles by diluting predation? Links between a group‐colonisation strategy and the absence of emergent multiple predator effects. Ecological Entomology 29(2): 129-138.
BASCOMPTE, J., P. JORDANO, C. J. MELIÁN y J. M. OLESEN (2003). The nested assembly of plant–animal mutualistic networks. Proceedings of the National Academy of Sciences 100(16): 9383-9387.
BASKERVILLE, G. L. y P. EMIN (1969). Rapid Estimation of Heat Accumulation from Maximum and Minimum Temperatures. Ecology 50(3): 514-517.
BASTOLLA, U., M. A. FORTUNA, A. PASCUAL-GARCÍA, A. FERRERA, B. LUQUE y J. BASCOMPTE (2009). The architecture of mutualistic networks minimizes competition and increases biodiversity. Nature 458(7241): 1018-1020.
BATAGELJ, V. y A. MRVAR (2006). Pajek datasets h ttp. vlado. fmf. uni-lj. si/pub/networks/data.
BELLINGERI, M. y A. BODINI (2013). Threshold extinction in food webs. Theoretical Ecology 6(2): 143-152.
BILLINGS, R. F. y R. S. CAMERON (1984). Kairomonal responses of Coleoptera, Monochamus titillator (Cerambycidae), Thanasimus dubius (Cleridae), and Temnochila virescens (Trogositidae), to behavioral chemicals of southern pine bark beetles (Coleoptera: Scolytidae). Environmental Entomology 13(6): 1542-1548.
BOGICH, T. L., A. M. LIEBHOLD y K. SHEA (2008). To sample or eradicate? A cost minimization model for monitoring and managing an invasive species. Journal of Applied Ecology 45(4): 1134-1142.
BONIFÁCIO, L. F. P. (2009). Impacte e evolução da doença da murchidão dos pinheiros (pine wilt disease) na zona afectada a sul do Rio Tejo.
BOUHOT, L., F. LIEUTIER y D. DEBOUZIE (1988). Spatial and temporal distribution of attacks by Tomicus piniperda L. and Ips sexdentatus Boern.(Col., Scolytidae) on Pinus sylvestris. Journal of Applied Entomology 106(1‐5): 356-371.
BROCKERHOFF, E. G., A. M. LIEBHOLD y H. JACTEL (2006). The ecology of forest insect invasions and advances in their management. Canadian Journal of Forest Research 36(2): 263-268.
CARNICER, J., P. JORDANO y C. J. MELIÁN (2009). The temporal dynamics of resource use by frugivorous birds: a network approach. Ecology 90(7): 1958-1970.
CHANSLER, J. F. (1967). Biology and life history of Dendroctonus adjunctus (Coleoptera: Scolytidae). Annals of the Entomological Society of America 60(4): 760-767.
CITMA (2015 -a). Banc de Dades de la Biodiversitat. www.bdb.cma.gva.es. Consulta realizada el 08/08/2015. Conselleria d’Infraestructures, Territori i Medi Ambient (Generalitat Valenciana).
CITMA (2015 -b). Revisión y evaluación de la red autonómica de valuación itosanitaria en las masas forestales de la Comunidad Valenciana. Red EFIVA. 2015. . Consellería de Infraestructuras Territorio y Medio Ambiente. Generalitat Valenciana Documento Interno.

124
COULSON, R. N., A. M. MAYYASI, J. L. FOLTZ y F. P. HAIN (1976). Interspecific competition between Monochamus titillator and Dendroctonus frontalis. Environmental Entomology 5(2): 235-247.
COULSON, R. N. y J. A. WITTER (1984). Forest entomology: ecology and management, John Wiley & Sons.
CRAIGHEAD, F. C. (1923). North American cerambycid larvae. DAHLSTEN, D. y F. STEPHEN (1974). Natural enemies and insect associates of the mountain
pine beetle, Dendroctonus ponderosae (Coleoptera: Scolytidae), in sugar pine. The Canadian Entomologist 106(11): 1211-1217.
DE CÁCERES, M. (2015). https://cran.r-project.org/web/packages/indicspecies/vignettes/indicspeciesTutorial.pdf.
DIXON, W. N. y T. PAYNE (1979). Sequence of arrival and spatial distribution of entomophagous and associate insects on southern pine beetle-infested trees. MP.
DODDS, K. J., C. GRABER y F. M. STEPHEN (2001). Facultative intraguild predation by larval Cerambycidae (Coleoptera) on bark beetle larvae (Coleoptera: Scolytidae). Environmental Entomology 30(1): 17-22.
DUNN, D. y J. P. CRUTCHFIELD (2006). Insects, Trees, and Climate: The bioacoustic ecology of deforestation and entomogenic climate change. arXiv preprint q-bio/0612019.
ETXEBESTE, I., G. ALVAREZ y J. PAJARES (2013a). Log colonization by Ips sexdentatus prevented by increasing host unsuitability signaled by verbenone. Entomologia Experimentalis Et Applicata 147(3): 231-240.
ETXEBESTE, I., J. L. LENCINA y J. PAJARES (2013b). Saproxylic community, guild and species responses to varying pheromone components of a pine bark beetle. Bulletin of entomological research 103(05): 497-510.
ETXEBESTE, I. y J. A. PAJARES (2011). Verbenone protects pine trees from colonization by the six-toothed pine bark beetle, Ips sexdentatus Boern. (Col.: Scolytinae). Journal of Applied Entomology 135(4): 258-268.
EVANS, H., D. MCNAMARA, H. BRAASCH, J. CHADOEUF y C. MAGNUSSON (1996). Pest risk analysis (PRA) for the territories of the European Union (as PRA area) on Bursaphelenchus xylophilus and its vectors in the genus Monochamus. EPPO Bulletin 26(2): 199-249.
EVANS, H. F., O. KULINICH, C. MAGNUSSON, C. ROBINET y T. SCHROEDER (2009). Report of a Pest Risk Analysis for Bursaphelenchus xylophilus, European and Mediterranean Plant Protection Organization (EPPO): 17.
FAITH, D. P., P. R. MINCHIN y L. BELBIN (1987). Compositional dissimilarity as a robust measure of ecological distance. Vegetatio 69(1-3): 57-68.
FATZINGER, C., B. SIEGFRIED, R. WILKINSON y J. NATION (1987). Trans-verbenol, turpentine, and ethanol as trap baits for the black turpentine beetle, Dendroctonus terebrans, and other forest Coleoptera in north Florida. Journal of Entomological Science 22(3): 201-209.
FLAMM, R., R. COULSON, P. BECKLEY, P. PULLEY y T. WAGNER (1989). Maintenance of a phloem-inhabiting guild. Environmental Entomology 18(3): 381-387.
GIESEN, H., U. KOHNLE, J. VITÉ, M. L. PAN y W. FRANCKE (1984). Das aggregationspheromon des mediterranen Kiefernborkenkäfers Ips (Orthotomicus) erosus. Zeitschrift für Angewandte Entomologie 98(1‐5): 95-97.
GIL, L. A. y J. A. PAJARES (1986). Los Escolítidos de las Coníferas en la Península Ibérica. Madrid, Instituto Nacional de Investigaciones Agrarias.
GROVE, S. J. (2002). Saproxylic insect ecology and the sustainable management of forests. Annual Review of Ecology and Systematics 33: 1-23.
GUIMERA, R. y L. A. N. AMARAL (2005). Functional cartography of complex metabolic networks. Nature 433(7028): 895-900.

125
HAACK, R. A. (2001). Intercepted Scolytidae (Coleoptera) at US ports of entry: 1985–2000. Integrated Pest Management Reviews 6(3-4): 253-282.
HARDY, S. A. C. (1924). The herring in relation to its animate environment, HM Stationery Office.
HELLRIGL, K. G. (1971). Die Bionomie der europaischen Monochamus-Arten (Coleopt., Cerambycid.) und ihre Bedeutung fur die Forstund Holzwirtschaft. Redia 52: 367-509.
HUERTA, A., F. ROBREDO, J. DIEZ y J. A. PAJARES (2004). Life-cycle characteristics of Tetrastichus turionum, a parasitoid of the European pine shoot moth, Rhyacionia buoliana. Journal of Applied Entomology 128(9-10): 628-632.
HULME, P. E. (2009). Trade, transport and trouble: managing invasive species pathways in an era of globalization. Journal of Applied Ecology 46(1): 10-18.
HUNTER, M. D. (2009). Trophic promiscuity, intraguild predation and the problem of omnivores. Agricultural and Forest Entomology 11(2): 125-131.
IBEAS, F., D. GALLEGO, J. DIEZ y J. PAJARES (2007). An operative kairomonal lure for managing pine sawyer beetle Monochamus galloprovincialis (Coleoptera: Cerymbycidae). Journal of Applied Entomology 131(1): 13-20.
ICP-FOREST (2011). The Condition of Forests in Europe. Executive Report. Institute for World Forestry y European commission. Brussels.
IVES, A. R. y S. R. CARPENTER (2007). Stability and diversity of ecosystems. Science 317(5834): 58-62.
KENIS, M., M.-A. AUGER-ROZENBERG, A. ROQUES, L. TIMMS, C. PÉRÉ, M. J. COCK, J. SETTELE, S. AUGUSTIN y C. LOPEZ-VAAMONDE (2009). Ecological effects of invasive alien insects. Ecological Impacts of Non-Native Invertebrates and Fungi on Terrestrial Ecosystems, Springer: 21-45.
KENIS, M. y J. HILSZCZANSKI (2004). Natural enemies of Cerambycidae and Buprestidae infesting living trees. Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis, Springer: 475-498.
KIRKENDALL, L. R. y M. FACCOLI (2010). Bark beetles and pinhole borers (Curculionidae, Scolytinae, Platypodinae) alien to Europe. ZooKeys(56): 227.
KOHNLE, U., S. DENSBORN, D. DUHME y J. VITÉ (1992). Bark beetle attack on host logs reduced by spraying with repellents. Journal of Applied Entomology 114(1‐5): 83-90.
KURZ, W. A., C. C. DYMOND, G. STINSON, G. J. RAMPLEY, E. T. NEILSON, A. L. CARROLL, T. EBATA y L. SAFRANYIK (2008). Mountain pine beetle and forest carbon feedback to climate change. Nature 452(7190): 987-990.
LAWRENCE, D., F. FIEGNA, V. BEHRENDS, J. G. BUNDY, A. B. PHILLIMORE, T. BELL y T. G. BARRACLOUGH (2012). Species interactions alter evolutionary responses to a novel environment. PLoS-Biology 10(5): 941.
LEVINE, J. M. y C. M. D'ANTONIO (2003). Forecasting biological invasions with increasing international trade. Conservation Biology 17(1): 322-326.
LIEBHOLD, A. M., E. G. BROCKERHOFF, L. J. GARRETT, J. L. PARKE y K. O. BRITTON (2012). Live plant imports: the major pathway for forest insect and pathogen invasions of the US. Frontiers in Ecology and the Environment 10(3): 135-143.
LINDENMAYER, D. B. y G. E. LIKENS (2009). Adaptive monitoring: a new paradigm for long-term research and monitoring. Trends in Ecology & Evolution 24(9): 482-486.
LINDNER, M., M. MAROSCHEK, S. NETHERER, A. KREMER, A. BARBATI, J. GARCIA-GONZALO, R. SEIDL, S. DELZON, P. CORONA y M. KOLSTRÖM (2010). Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. Forest Ecology and Management 259(4): 698-709.
LINSLEY, E. G. (1959). Ecology of cerambycidae. Annual Review of Entomology 4(1): 99-138. MAGRAMA. (2015). "Nematodo de la madera del pino." 2015, de
http://www.magrama.gob.es/es/agricultura/temas/sanidad-vegetal/nematodo-de-la-madera-del-pino/#.

126
MARINI, L., R. A. HAACK, R. J. RABAGLIA, E. P. TOFFOLO, A. BATTISTI y M. FACCOLI (2011). Exploring associations between international trade and environmental factors with establishment patterns of exotic Scolytinae. Biological Invasions 13(10): 2275-2288.
MARTÍN-PIERA, F. (2001). Area networks for conserving Iberian insects: A case study of dung beetles (col., Scarabaeoidea). Journal of Insect Conservation 5(4): 233-252.
MCCANN, K. S. (2000). The diversity–stability debate. Nature 405(6783): 228-233. MCCULLOUGH, D. G., T. T. WORK, J. F. CAVEY, A. M. LIEBHOLD y D. MARSHALL
(2006). Interceptions of nonindigenous plant pests at US ports of entry and border crossings over a 17-year period. Biological Invasions 8(4): 611-630.
MCDOWELL, N., W. T. POCKMAN, C. D. ALLEN, D. D. BRESHEARS, N. COBB, T. KOLB, J. PLAUT, J. SPERRY, A. WEST y D. G. WILLIAMS (2008). Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New phytologist 178(4): 719-739.
MCGEOCH, M. A. (1998). The selection, testing and application of terrestrial insects as bioindicators. Biological Reviews of the Cambridge Philosophical Society 73(02): 181-201.
MENDEL, Z. y J. HALPERIN (1982). The biology and behavior ofOrthotomicus erosus in Israel. Phytoparasitica 10(3): 169-181.
MILLER, M. C. (1984). Mortality contribution of insect natural enemies to successive generations of Ips calligraphus (Germar)(Coleoptera, Scolytidae) in loblolly pine. Zeitschrift für Angewandte Entomologie 98(1‐5): 495-500.
MILLER, M. C. (1985). The effect of Monochamus titillator (F.)(Col., Cerambycidae) foraging on the emergence of Ips calligraphus (Germ.)(Col., Scolytidae) insect associates. Zeitschrift für Angewandte Entomologie 100(1‐5): 189-197.
MONTOYA, J. M., S. L. PIMM y R. V. SOLÉ (2006). Ecological networks and their fragility. Nature 442(7100): 259-264.
MOSER, J. C., R. THATCHER y L. PICKARD (1971). Relative abundance of southern pine beetle associates in East Texas. Annals of the Entomological Society of America 64(1): 72-77.
MOTA, M. M., H. BRAASCH, M. A. BRAVO, A. C. PENAS, W. BURGERMEISTER, K. METGE y E. SOUSA (1999). First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1: 727-734.
NAVES, P., S. CAMACHO, E. DE SOUSA y J. QUARTAU (2007). Transmission of the pine wood nematode Bursaphelenchus xylophilus through feeding activity of Monochamus galloprovincialis (Col., Cerambycidae). Journal of Applied Entomology 131(1): 21-25.
NAVES, P. y E. DE SOUSA (2009). Threshold temperatures and degree-day estimates for development of post-dormancy larvae of Monochamus galloprovincialis (Coleoptera: Cerambycidae). Journal of Pest Science 82(1): 1-6.
NAVES, P. M., E. M. DE SOUSA y J. A. QUARTAU (2006). Feeding and oviposition preferences of Monochamus galloprovincialis for certain conifers under laboratory conditions. Entomologia experimentalis et applicata 120(2): 99-104.
NAVES, P. M., E. SOUSA y J. M. RODRIGUES (2008). Biology of Monochamus galloprovincialis (Coleoptera: Cerambycidae) in the pine wilt disease affected zone, Southern Portugal. Silva Lusitana 16(2): 133-148.
NEW, T., R. PYLE, J. THOMAS, C. THOMAS y P. HAMMOND (1995). Butterfly conservation management. Annual Review of Entomology 40(1): 57-83.
NIETO, A. y K. ALEXANDER (2010). European Red List of Saproxylic Beetles. Luxemburg, Publications Office of the European Union: 45 p, ISBN 978-92-79-14152-2.
OKSANEN, J., K. ROELAND, P. LEGENDRE, B. O'HARA, G. L. SIMPSON, P. SOLYMOS, H. H. STEVENS y H. WAGNER (2008). vegan: Community Ecology Package. R package version 1.15-1.

127
PAIVA, M. R., M. F. PESSOA y J. VITÉ (1988). Reduction in the pheromone attractant response of Orthotomicus erosus (Woll.) and Ips sexdentatus Boern.(Col., Scolytidae) 1. Journal of Applied Entomology 106(1‐5): 198-200.
PAJARES, J. A., G. ALVAREZ, F. IBEAS, D. GALLEGO, D. R. HALL y D. I. FARMAN (2010). Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. Journal of Chemical Ecology 36(6): 570-583.
PAJARES, J. A., F. IBEAS, J. J. DIEZ y D. GALLEGO (2004). Attractive responses by Monochamus galloprovincialis (Col., Cerambycidae) to host and bark beetle semiochemicals. Journal of Applied Entomology 128(9-10): 633-638.
PIEL, F., M. GILBERT, C. DE CANNIÈRE y J. C. GRÉGOIRE (2008). Coniferous round wood imports from Russia and Baltic countries to Belgium. A pathway analysis for assessing risks of exotic pest insect introductions. Diversity and distributions 14(2): 318-328.
PIMENTEL, D., R. ZUNIGA y D. MORRISON (2005). Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecological Economics 52(3): 273-288.
POLIS, G. A., C. A. MYERS y R. D. HOLT (1989). The ecology and evolution of intraguild predation: potential competitors that eat each other. Annual review of ecology and systematics: 297-330.
PYŠEK, P. y D. M. RICHARDSON (2010). Invasive species, environmental change and management, and health. Annual Review of Environment and Resources 35: 25-55.
QUINTO, J., M. Á. MARCOS-GARCÍA, C. DÍAZ-CASTELAZO, V. RICO-GRAY, H. BRUSTEL, E. GALANTE y E. MICÓ (2012). Breaking down complex saproxylic communities: understanding sub-networks structure and implications to network robustness.
R CORE TEAM (2015). R: A language and environment for statistical computing. Vienna, Austria, R Foundation for Statistical Computing.
RABAGLIA, R., D. DUERR, R. ACCIAVATTI y I. RAGENOVICH (2008). Early detection and rapid response for non-native bark and ambrosia beetles. US Department of Agriculture Forest Service, Forest Health Protection.
RASSATI, D., M. FACCOLI, L. MARINI, R. A. HAACK, A. BATTISTI y E. P. TOFFOLO (2015a). Exploring the role of wood waste landfills in early detection of non-native wood-boring beetles. Journal of Pest Science: 1-10.
RASSATI, D., M. FACCOLI, E. PETRUCCO TOFFOLO, A. BATTISTI y L. MARINI (2015b). Improving the early detection of alien wood‐boring beetles in ports and surrounding forests. Journal of Applied Ecology 52(1): 50-58.
RASSATI, D., E. P. TOFFOLO, A. ROQUES, A. BATTISTI y M. FACCOLI (2014). Trapping wood boring beetles in Italian ports: a pilot study. Journal of Pest Science 87(1): 61-69.
ROLTSCH, W. J., F. G. ZALOM, A. J. STRAWN, J. F. STRAND y M. J. PITCAIRN (1999). Evaluation of several degree-day estimation methods in California climates. International Journal of Biometeorology 42(4): 169-176.
ROMÓN, P., J. C. ITURRONDOBEITIA, K. GIBSON, B. S. LINDGREN y A. GOLDARAZENA (2007). Quantitative association of bark beetles with pitch canker fungus and effects of verbenone on their semiochemical communication in Monterey pine forests in northern Spain. Environmental Entomology 36(4): 743-750.
ROQUES, A., B. RICHARDSON y I. HOOD (2010). Alien forest insects in a warmer world and a globalised economy: impacts of changes in trade, tourism and climate on forest biosecurity. New Zealand Journal of Forestry Science, New Zealand Forest Research Institute.
SAMWAYS, M. J. (1994). Insect Conservation Biology (Conservation Biology, No 2), Springer Science & Business Media.

128
SCHOENER, T. W. (1974). Resource partitioning in ecological communities. Science 185(4145): 27-39.
SCHROEDER, L. y J. WESLIEN (1994). Reduced offspring production in bark beetleTomicus piniperda in pine bolts baited with ethanol and α-pinene, which attract antagonistic insects. Journal of Chemical Ecology 20(7): 1429-1444.
SEYBOLD, S. J., J. C. LEE, A. LUXOVA, S. M. HAMUD, P. JIROŠ y R. L. PENROSE (2006). Chemical ecology of bark beetles in California’s urban forests. I. CITRUS AND BIOLOGICAL CONTROL. YESTERDAY, TODAY, TOMORROW: 87.
SIITONEN, J. (2000). Beetles (Coleoptera) imported to Finland on Russian roundwood. Aktuelt fra skogforskningen(4): 11-17.
SLAMA, M. (1998). Longhorn beetles Cerambycidae of Czech and Slovak Republics. Czech Republic: Tercie (publ.) (in Czech).
SLANSKY JR, F. (1992). Allelochemical-nutrient interactions in herbivore nutritional ecology. Herbivores: their interactions with secondary plant metabolites 2: 135-174.
SOUSA, E., M. A. BRAVO, J. PIRES, P. NAVES, A. C. PENAS, L. BONIFACIO y M. M. MOTA (2001). Bursaphelenchus xylophilus (Nematoda; Aphelenchoididae) associated with Monochamus galloprovincialis (Coleoptera; Cerambycidae) in Portugal. Nematology 3: 89-91.
STANAWAY, M. A., M. P. ZALUCKI, P. S. GILLESPIE, C. M. RODRIGUEZ y G. V. MAYNARD (2001). Pest risk assessment of insects in sea cargo containers. Australian journal of Entomology 40(2): 180-192.
STROGATZ, S. H. (2001). Exploring complex networks. Nature 410(6825): 268-276. THÉBAULT, E. y C. FONTAINE (2010). Stability of ecological communities and the
architecture of mutualistic and trophic networks. Science 329(5993): 853-856. THOMPSON, R. M., U. BROSE, J. A. DUNNE, R. O. HALL JR, S. HLADYZ, R. L.
KITCHING, N. D. MARTINEZ, H. RANTALA, T. N. ROMANUK, D. B. STOUFFER y J. M. TYLIANAKIS (2012). Food webs: reconciling the structure and function of biodiversity. Trends in Ecology & Evolution 27(12): 689-697.
VICENTE, C., M. ESPADA, P. VIEIRA y M. MOTA (2012). Pine Wilt Disease: a threat to European forestry. European Journal of Plant Pathology 133(1): 89-99.
VICTORSSON, J. y L.-O. WIKARS (1996). Sound production and cannibalism in larvae of the pine-sawyer beetle Monochamus sutor L.(Coleoptera: Cerambycidae). Entomologisk Tidskrift 117(1/2): 29-33.
VIVES (2000). Insecta, Coleoptera, Cerambycidae. Serie Fauna Ibérica. Museo Nacional de Ciencias Naturales, CSIC, Madrid. 12.
WEINGESSEL, A., E. DIMITRIADOU y S. DOLNICAR (1999). An examination of indexes for determining the number of clusters in binary data sets.
WORK, T. T., D. G. MCCULLOUGH, J. F. CAVEY y R. KOMSA (2005). Arrival rate of nonindigenous insect species into the United States through foreign trade. Biological Invasions 7(2): 323-332.
WYLIE, F., M. GRIFFITHS y J. KING (2008). Development of hazard site surveillance programs for forest invasive species: a case study from Brisbane, Australia. Australian forestry 71(3): 229-235.
YOSHIKAWA, K. (1987). A study of the subcortical insect community in pine trees. II Vertical distribution. Applied Entomology and Zoology 22(2): 195-206.

129
Capítulo 5
Dispersión y población de Monochamus
galloprovincialis (Col.: Cerambycidae) en la península ibérica
Introdución
Dos únicas especies componen la representación del género Monochamus (Coleoptera, Cerambycidae) en la península Ibérica: Monochamus sutor Linneo, y Monochamus galloprovincialis (Olivier). Monochamus sutor es una especie de dominio paleártico, cuya distribución llega hasta Japón atravesando Siberia. No obstante, en la península Ibérica es una especie rara que se distribuye únicamente por Pirineos y sus estribaciones. La segunda especie, M. galloprovincialis es mucho más común y se encuentra distribuida en el área circunmediterránea, Europa central, Cáucaso, Siberia, Mongolia y China. En la península Ibérica vive sobre prácticamente todas las especies de Pinus, e incluso Abies y Picea (GONZÁLEZ et al. 2007). Monochamus galloprovincialis, y en general todas las especies del género, son consideradas de hábitos secundarios, colonizadoras de pinos muy debilitados y preferentemente con presencia de escolítidos (VIVES 2000). Prueba de ello es la respuesta de M. galloprovincialis a un compuesto cairomonal emitido por el hospedador ( -pineno) y dos compuestos de la feromona emitida por escolítidos del género Ips y Orthotomicus (ipsenol y 2-metil-3-buten-2-ol), de acuerdo con PAJARES et al. (2004) e IBEAS et al. (2007), señales características de coníferas estresadas y que están siendo colonizadas por estos insectos. El interés por el estudio de este género de coleópteros Monochamus se ha incrementado significativamente en los últimos años ya que sus especies son los únicos vectores conocidos del nematodo de la madera del pino Bursaphelenchus xylophilus (Steiner et Buhrer). La primera detección de NMP en Europa se produjo en la península de Setúbal en 1999 (MOTA et al. 1999). A partir de ese momento, se demarcó la zona afectada y aplicaron medidas de control. Desde 2008, todo el conjunto de Portugal continental fue designado zona demarcada y se delimitó una zona de amortiguamiento que ocupó 20 km de ancho lo largo de la frontera con España. La zona infestada se dividió en dos zonas: la zona de intervención (ZI), donde realmente se identificaron los árboles infestados y el resto. Después de varios años de aplicación de medidas de control contra el NMP, las autoridades portuguesas consideraron que la erradicación no era viable y por lo tanto decidieron contener el PWN en la zona infestada y concentrar así sus acciones de control en la zona de amortiguamiento de la zona demarcada de Portugal continental, al tiempo que se intentaba retrasar y controlar la propagación real de la plaga mediante la aplicación de determinadas medidas en la zona infestada (VICENTE et al. 2012).

130
En España la primera detección de B. xylophilus fue octubre de 2008, en la Sierra de Dios Padre, en Extremadura. Tras aplicar las medidas de erradicación contenidas en la Decisión de la Comisión 2006/133/CE, el foco fue declarado erradicado por las autoridades españolas el 16 de enero de 2013. Después de este primer foco, hubo otras tres nuevas detecciones en España: As Neves, Galicia (2010); Valverde del Fresno, Extremadura (2012); Sancti-Spiritus, Castilla y León (2013). Actualmente todas ellas se encuentran en curso de erradicación. En todos estos casos fueron establecidas zonas demarcadas y medidas de erradicación conforme a la legislación vigente (ABELLEIRA et al. 2011, MAGRAMA 2015a, ZAMORA et al. 2015). La dispersión de esta enfermedad está mediada por la dispersión de sus vectores, sea ésta de tipo natural o sea facilitada por los grandes movimientos de personas y materiales que acompañan a la globalización de la economía y al comercio internacional. Fundamentalmente es este último tipo de dispersión el responsable de la aparición repentina de todo tipo de especies invasoras a lo ancho del mundo, capaces de generar grandes daños a la biodiversidad, al medio ambiente o a la economía (LEVINE Y D'ANTONIO 2003, ROBINET et al. 2009, ROBINET et al. 2011). No obstante, una vez alcanzados nuevos territorios, la dispersión natural adquiere un especial protagonismo por ser el mecanismo esencial de las especies para alcanzar nuevos entornos y para facilitar el intercambio genético entre diferentes poblaciones o dentro de las mismas (CLOBERT et al. 2012). La propagación del PWD a larga distancia está relacionada esencialmente con el transporte humano (TAKASU et al. 2000, TOGASHI Y SHIGESADA 2006, ROBINET et al. 2011), mientras que la dispersión local está vinculada al comportamiento natural de las diferentes especies de Monochamus (YOSHIMURA et al. 1999, TAKASU et al. 2000, ROBINET et al. 2011). La dispersión de los organismos vivos, se representa frecuentemente mediante una curva de probabilidad del alcance de diferentes distancias y ha sido estudiada para muchos organismos (TURCHIN 1998, OKUBO Y LEVIN 2001, NATHAN et al. 2012). En términos generales, tal curva se caracteriza por tener a una gran parte de los eventos de dispersión a corta distancia, es decir, una mayoría permanece relativamente cerca de su lugar de nacimiento (o emergencia en el caso de Monochamus), mientras que unos pocos pueden realizan grandes desplazamientos a larga distancia, caracterizando una curva leptocúrvica con una cola muy difícilmente estimable (NATHAN et al. 2003b). Uno de los mecanismos más utilizados para el estudio de la capacidad potencial de vuelo de los insectos son los molinos de vuelo, pero estos ensayos de laboratorio son difícilmente ajustables a las condiciones naturales habida cuenta que ninguna otra variable medioambiental es evaluada en ellos(RANIUS 2006). Además, la tendencia común es a que sobredimensionen las capacidades de vuelo reales (RANIUS 2006, TAYLOR et al. 2010). Por lo tanto, para la evaluación de la capacidad dispersiva de los insectos en condiciones naturales deben ser utilizados, complementariamente, otros métodos entre los que destacan los ensayos de marcaje-liberación-recaptura. El éxito relativo de estos experimentos radica en la puesta a punto de una técnica eficaz de captura. El uso de atrayentes cairomonales y feromonales y la existencia de dispositivos de trampeo adecuados han permitido que este tipo de experimentos hayan sido ampliamente utilizados para evaluar la dispersión de escarabajos perforadores de la madera (TURCHIN Y THOENY 1993, TURCHIN Y ODENDAAL 1996, FRANKLIN et al. 2000, BANCROFT Y SMITH 2005). El desarrollo en los últimos tiempos de un atrayente cairo-feromonal muy eficaz en la captura de M. galloprovincialis como fruto de los trabajos de PAJARES et al. (2004), IBEAS et al. (2007), PAJARES et al. (2010), y el incremento de la eficacia del dispositivo de capturas tras las mejoras implementadas en las trampas (ÁLVAREZ et al. 2015) han permitido la realización

131
de estudios de marcaje-liberación-recaptura de M. galloprovincialis que tiempo atrás habrían sido inabordables e infructuosos. Estas técnicas han sido utilizadas desde hace décadas para estudiar procesos de dispersión de especies (TURCHIN Y THOENY 1993, NATHAN et al. 2003b), así como para estudiar movimientos individuales y capacidades de dispersión a larga distancia (KOENIG et al. 1996). Ambos métodos han sido utilizados en los últimos años para estudiar la capacidad dispersiva de M. galloprovincialis. En concreto, el uso de molinos de ha mostrado promedios de distancias de vuelo próximas a los 16 km (DAVID et al. 2014) mientras que la dispersión en masas continuas ha sido recientemente estudiada mostrando distancias de dispersión considerablemente menores, con medianas de insectos dispersores entre 233 y 532 m, y 99% a distancias entre 2344 m y 3496 m (ETXEBESTE et al. 2015). El presente capítulo tiene el objetivo general de aportar datos sobre el comportamiento, la capacidad y las características del vuelo de M. galloprovincialis en condiciones naturales, y el papel que ciertos factores ambientales como la topografía, el paisaje o el viento pueden tener en la modulación de dicho vuelo utilizando técnicas de marcaje-liberación-recaptura. Los objetivos concretos han sido estructurados en diferentes secciones, con la intención de facilitar la comprensión del trabajo. En concreto, se recogen los resultados de una larga serie de experimentos de marcaje-liberación-recaptura realizados en la región oriental de la península ibérica entre los años 2010 y 2015 (Tabla 5.1). La metodología es común a todos ellos, diferenciándose exclusivamente en la cantidad de trampas y su distribución espacial, en las características (paisajísticas y estructurales) de las áreas de muestreo, el origen de los insectos liberados (nativos o criados en laboratorio) o la ubicación relativa del punto de liberación.
Tabla 5.1 Experimentos analizados en el capítulo 5 de dispersión y población de M. galloprovincialis Experimento
Localización
T.M. Provincia Año Sección Exp2009/a Bovalar Atzeneta del Mestrat Castelló 2009 5.4 Exp2009/b Camarracho Mora de Ruebilos Teruel 2009 5.4 Exp2010/a Bovalar-Mas de Timor Atzeneta del Mestrat Castelló 2010 5.2 Exp2010/b La Muela Jérica Castelló 2010 5.3/5.4 Exp2010/c Camarracho Mora de Ruebilos Teruel 2010 5.4 Exp2011/a Boqueras Altura Castelló 2011 5.3/5.4 Exp2011/b Coll de la Bassa Atzeneta del Mestrat Castelló 2011 5.3/5.4 Exp2011/c Camarracho Mora de Ruebilos Teruel 2011 5.4 Exp2011/d Mosaico Jumilla Jumilla Murcia 2011 5.4 Exp2012/a Masa aislada Caudete Caudete València 2012 5.1 Exp2012/b Mosaico Atzeneta del Maestrat Atzeneta del Mestrat Castelló 2012 5.3 Exp2012/c Mosaico Camporrobles Camporrobles València 2012 5.4

132
Sección 5.1. Respuesta temporal de individuos, nativos y liberados, de
Monochamus galloprovincialis a atrayentes cairomonales y feromonales
en una masa aislada.
Introducción El desarrollo de un atrayente cairomonal y feromonal eficaz (PAJARES et al. 2004, IBEAS et al. 2007, PAJARES et al. 2010) y la mejora de las trampas utilizadas para su captura (ÁLVAREZ et al. 2015) ha permitido llevar a cabo monitorizaciones estandarizadas de su población (MAGRAMA 2015b) y diferentes investigaciones sobre importantes parámetros ecológicos como su fenología, densidad de población, dispersión o capacidad de vuelo a larga distancia, entre otros (HERNÁNDEZ et al. 2011, GALLEGO et al. 2012, SANCHEZ-HUSILLOS et al. 2015a, TORRES-VILA et al. 2015). El atrayente utilizado en ensayos científicos y muestreos, y comercializado con el nombre Galloprotect Plus ®, (SEDQ, Barcelona) está compuesto por una mezcla cairomonal formada por dos componentes feromonales de escolítidos (ipsenol y 2-metil-3-buten-2-ol ), con posibilidad de añadir un volátil de hospedante, α-pineno como sinergizante (PAJARES et al. 2004, IBEAS et al. 2007, PAJARES et al. 2010), y por una feromona de Monochamus, de tipo sexual y agregativa, emitida por los machos: 2-undeciloxi-1-etanol. La atracción hacia este compuesto parece vinculada a la madurez de los adultos, tanto en el caso de los machos como de las hembras, como prueban los hechos de que la feromona sea producida por machos maduros alimentándose de ramillos de pino (PAJARES et al. 2010) y que la atracción de hembras hacia las plumas con olores de machos en bioensayos con olfatómetros, solo se dé cuando ambos sexos son maduros (IBEAS et al. 2008). Los comportamientos de madurez sexual de M. galloprovincialis han sido observados, siempre, después de un periodo de días tras la emergencia y tras de un periodo de alimentación en los ramillos de pinos sanos (HELLRIGL 1971, FRANCARDI Y PENNACCHIO 1996, DAVID et al. 2014, ETXEBESTE et al. 2015, SANCHEZ-HUSILLOS et al. 2015a). En estudios de campo con insectos criados en laboratorio marcados y liberados, sólo han capturado insectos sexualmente maduros, y generalmente a partir de la segunda semana tras la liberación (ETXEBESTE et al. 2015, SANCHEZ-HUSILLOS et al. 2015a). Según BONIFACIO (2009), el comportamiento de algunos insectos tras la emergencia incluye un vuelo fisiológico para fortalecer los músculos de vuelo instantes después de la emergencia. La incapacidad de las trampas para muestrear este periodo podría suponer un importante problema en su utilización como método de control del NMP, por lo que es necesario conocer de forma detallada el funcionamiento y la efectividad de las trampas en los primeros días tras la emergencia del insecto, y ahondar en el conocimiento del comportamiento de M. galloprovincialis en estado inmaduro. El objetivo, por tanto, de este trabajo es evaluar la efectividad de las trampas cebadas con atrayentes cairomonales y feromonales para capturas individuos marcados y liberados y la evolución diaria de las capturas a lo largo de su periodo de maduración sexual (durante los primeros días tras la emergencia de los insectos) mediante el uso de insectos inmaduros criados en laboratorio y liberados en una masa de reducidas dimensiones y muy aislada que pueda actuar, en la medida de lo posible, como medida de confinamiento de la población liberada.

134
Material y métodos Un experimento de marcaje-liberación se llevó a cabo en una masa forestal aislada y de reducida superficie (10 ha) en el término municipal de Caudete de las Fuentes (Valencia, España; 39 ° 49'18 "N, 0 ° 38'31" W) durante el año 2012, entre el 20.06.2012 y el 6.11.2012 (el estudio es codificado a partir de ahora como Exp2012a). La zona de estudio general es una matriz agrícola de secano (fundamentalmente cereal, almendro y viña) con muy poco arbolado disperso. En un radio de 3 km alrededor de la masa estudiada sólo es posible observar algún bosquete de P. halepensis de superficies muy pequeñas (<5ha) o árboles aislados (Figura 5.1). La primera masa de pinar continua y extensa (>10 ha) cercana a la masa forestal de estudio se encuentra a más de 3 km hacia el Sur. En términos generales, la región está formada por una amplia llanura con alguna ondulación, rodeada por regiones montañosas al Norte y Este, y la cuenca del río Cabriel en su vertiente occidental y meridional. Está situada a una altitud de 770 msnm, aunque tiene elevaciones que sobrepasan los 800 m y destacan los vértices geodésicos de tercer orden de la Ermita (850 m) y Atalaya de Caudete (860 m). El río Madre, afluente del Magro, atraviesa el término de oeste a este.
Figura 5.1 Localización de la masa aislada en la que fue realizado el Exp2012/a. En verde, la presencia de masa forestal de P.
halepensis según el IFN3 (2008) Diez trampas modificadas (ÁLVAREZ et al. 2015) y cebadas con atrayente cairomonal y feromonal Galloprotect 2D ® (PAJARES et al. 2010) fueron colocadas a lo largo de una gradilla de 90 m de lado en la masa forestal de la zona de estudio, una repoblación regular de P. halepensis de 9 ha cercana al núcleo urbano de Caudete de las Fuentes (Figura 5.1) Un total de 215 M. galloprovincialis (100 machos y 115 hembras criados en laboratorio y marcados) fueron liberados en un punto central del interior de la masa de estudio en una sola

135
tanda de liberación. El punto de liberación se estableció en el centroide de la masa. Las trampas fueron revisadas diariamente durante los primeros 21 días y semanalmente, a partir de entonces, hasta finalizado el periodo de vuelo del insecto. La captura se realizó con muerte para reducir al máximo la probabilidad de escape de los insectos. Las representaciones gráficas de las capturas fueron acompañadas por las tendencias ajustadas por modelos aditivos generalizados (GAM) en el caso de las curvas de vuelo, y por un ajuste binomial mediante modelos lineares generalizados (GLM), en el caso de las capturas acumuladas. Todos los cálculos y representaciones han sido realizados bajo el entorno de programación y lenguaje estadístico R (R CORE TEAM 2015).

136
Resultados Un 35,3 % de los insectos liberados fue recapturado (Tabla 5.2, Figura 5.2, Figura 5.3). El primer M. galloprovincialis recapturado fue macho y tuvo lugar el octavo día después de la liberación de los insectos (Figura 5.4). El noveno día fueron capturados 2 machos y 2 hembras, iniciándose, a partir de este momento, la captura de los insectos criados en laboratorio y liberados en la masa de estudio de forma relativamente constate (Figura 5.4).
Tabla 5.2 Capturas de M. galloprovincialis nativos y liberados en el Exp2012/a
T01 T02 T03 T04 T05 T06 T07 T08 T09 T10 Nativos 31 26 27 34 17 13 23 22 19 26
Liberados 2 6 4 18 19 3 1 29 1 1
Figura 5.2 Capturas totales de M. galloprovincialis nativos en Exp2012/a separadas por sexo (hembras en rojo, machos en
azul)
Figura 5.3 Capturas totales de M. galloprovincialis liberados en
Exp2012/a separadas por sexo (hembras en rojo, machos en azul)
La curva de vuelo registrada por las capturas tanto de insectos nativos como de insectos liberados y recapturados sigue un perfil similar durante la primera mitad del periodo de vuelo. A partir de la primera semana de agosto, las capturas de insectos liberados descienden hasta dejar de capturarse mientras que un segundo pico se observa en los insectos nativos capturados hasta finales del mes de octubre (Figura 5.4 y Figura 5.5).

137
Figura 5.4 Perfil temporal de las capturas diarias por trampa de M.
galloprovincialis nativos y liberados, y separadas por sexo (hembras en rojo, machos en azul) en Exp2112/a
Figura 5.5 Capturas diarias por trampa acumuladas de M. galloprovincialis
nativos y liberados, y separadas por sexo (hembras en rojo, machos en azul) en Exp2112/a

138
Discusión El muestreo intensivo (diario) desde el día posterior a la liberación de M. galloprovincialias inmaduros y criados en laboratorio supone, hasta donde conocemos, la primera evaluación detallada del periodo de respuesta en campo de M. galloprovincialis a los atrayentes cairomonales y feromonales que componen el cebo estándar utilizado en la mayoría de los ensayos recientes que implican captura de individuos (Capítulo 3; SANCHEZ-HUSILLOS et al. 2013, ETXEBESTE et al. 2015, SANCHEZ-HUSILLOS et al. 2015a, b). Así, los resultados muestran que la primera captura tuvo lugar el octavo día (un insecto macho) mientras que el noveno día se obtuvieron un total de 4 nuevas capturas (dos machos y dos hembras) (Figura 5.5). La eficacia del atrayente Galloprotect 2D parece definitivamente ligado a la madurez sexual de ambos sexos, pese a que una parte de la mezcla atrayente son cairomonas del hospedante. En este sentido, estas cairomonas emulan la señal emitida por árboles debilitados y/o colonizados por escolítidos, que son escogidos por M. galloprovincialis como material de reproducción, por lo que parece razonable que la respuesta a este tipo de señales se dé también una vez alcanzada dicha madurez sexual. Los trabajos de IBEAS et al (2008) mostraron la existencia de una atracción de hembras exclusivamente maduras hacia machos exclusivamente maduros en olfactómetro, sin que se observase atracción alguna en el caso de que uno de los sexos no hubiese alcanzado la madurez sexual. Para alcanzar la madurez sexual, los adultos de M. galloprovincialis recién emergidos necesitan un periodo de intensa alimentación, fundamentalmente de corteza tierna de ramillos de pino (HELLRIGL 1971, FRANCARDI Y PENNACCHIO 1996). En concreto, diversos estudios abordan este aspecto desde diferentes perspectivas: HELLRIGL (1971) indica que el periodo de maduración de las gónadas puede durar hasta 14 días, mientras que FRANCARDI y PENNACCHIO (1996) observan periodos de inmadurez de entre 4 y 6 días (constatando cópulas de adultos jóvenes sólo 2-3 días después de la emergencia. NAVES et al. (2008) observan que la aparición de mordeduras de puesta se produce a partir del día 11 posterior a la emergencia; y DAVID et al. (2014) observan que no es posible encontrar huevos hasta 10 días después de la emergencia. Recientemente, los estudios fisiológicos de SANCHEZ-HUSILLOS et al. (2015b) indican que la maduración sexual de M. galloprovincialis se alcanza después de 12 días. Todos estos datos, pese a su variabilidad, son consistentes con las observaciones en campo realizadas en este ensayo, que marcan los 8-9 días como el periodo de ineficacia de las trampas con respecto a la emergencia de los adultos. Este periodo, en el marco del control de la enfermedad del marchitamiento del pino, puede suponer un problema para la monitorización de la dispersión de este insecto en caso de que exista un vuelo inicial dispersivo en M. galloprovincialis, tal y como indica BONIFÁCIO (2009). Sin embargo, este vuelo fisiológico no reproductivo, aunque habitual en algunos insectos (ZHERA et l. 1997, LÓPEZ GUILLEN et al. 201), no se encuentra suficientemente estudiado ni comprobado en el caso de M. galloprovincialis. Respecto a la evolución temporal de las capturas (curva de vuelo), tanto la referente a los insectos liberados como la de los insectos nativos puede ser observada en la Figura 5.4 y Figura 5.5. Aunque la instalación del experimento fue colocada con unos días de retraso con respecto al inicio del periodo de vuelo de M. galloprovincialis en la zona de estudio, puede observarse una alta similitud entre las capturas de insectos liberados y nativos (siempre tomando en consideración los desajustes provocados por los primeros 8 días tras la liberación y por el retraso en la instalación del conjunto de trampas).

139
No obstante, a partir del mes de agosto, se produce una fuerte divergencia entre ambas curvas de vuelo. Por un lado la captura por día de insectos criados en laboratorio y liberados decrece hasta cero, mientras que, tras un descenso de las capturas a inicios del mes de agosto, éstas vuelven a incrementarse mostrando unas tasas constantes de capturas de entorno a dos individuos por día a finales de verano e inicios de otoño, (Figura 5.4), coincidente con el segundo pico de capturas obtenidas en una gran cantidad de ensayos de trampeo en la península ibérica desde 2010 (Capítulo 3).
Estos resultados concuerdan con la hipótesis planteada en el Capítulo 3 de esta tesis, en la que se sugiere que puede existir cierta inhibición del vuelo de los insectos en los periodos más calurosos del verano, como puede ser el caso del descenso de la tasa de capturas de M. galloprovincialis nativos a mediados de julio (Figura 5.4). En ese momento de inhibición del vuelo, una parte de la población aún se encuentra en las cámaras de pupación sin haber emergido, y otra parte, aun habiendo emergido, no ha completado su maduración sexual por lo que no es susceptible de ser atraída por los atrayentes utilizados. El restablecimiento de las temperaturas adecuadas para que se produzca el vuelo con normalidad provocará, no sólo el aumento de una actividad de vuelo temporalmente inhibida, sino el reclutamiento de una parte de la población que, hasta esa fecha, no había sido susceptible de ser atraídas por las trampas por encontrarse no emergida o sexualmente inmadura. La mejora en la detección por parte de los machos más viejos de la feromona producida por otros machos podría suponer, además, un nuevo aporte de población capturada a este segundo pico. En el caso de los insectos criados en laboratorio, la llegada de las altas temperaturas estivales podría inhibir el vuelo de estos insectos pero, puesto que se realizó una única liberación de individuos, no habría un reclutamiento extra de individuos supuestamente no emergidos o inmaduros (Figura 5.4).
Por último, la alta tasa de captura registrada en este experimento (35,3 %) es considerablemente mayor que las del resto de experimentos de captura-recaptura analizados en este trabajo (Capítulo 5.3) así como en otras publicaciones (HERNÁNDEZ et al. 2011, TORRES-VILA et al. 2015), mientras que es del mismo orden de magnitud que los resultados de los trabajos realizados en 2009 y 2010 por ETXEBESTE et al. (2015). Aparentemente, existen varios parámetros que podrían haber influido en la tasa de recapturas de M. galloprovincialis: el tamaño de la masa forestal, su aislamiento y la densidad de trampas utilizada. La densidad de trampas utilizada en este ensayo (1,25 trampas/ha, en una superficie de 8 ha) es mucho mayor al resto de los analizados en el Capítulo 5.3 de este trabajo (por ejemplo, 0.2 trampas/ha en Exp2010b y 0.36 trampas/ha en Exp2011/a), lo cual es coherente con una mayor tasa de recapturas observada aquí. No obstante, el diseño experimental de TORRES-VILA et al. (2015) cuenta con una densidad mucho mayor (6,25 trampas/ha) y una superficie muestreada menor (4 ha), y sin embargo la tasa de recapturas máxima obtenida en dicho experimento es del 19%. Esta diferencia podría estar relacionada con el nivel de aislamiento del área de muestreo de este estudio. La capacidad de dispersión de M. galloprovincialis en una masa continua ha sido estudiada por ETXEBESTE et al. (2015), fijando en 532 m la distancia a la que se dispersa el 50% de la población según el mejor modelo mecánico estudiado. Parece razonable pensar que el área muestreada en los trabajos de TORRES-VILA et al. (2015) , una parcela de 200 m x 200 m en una masa continua, es demasiado reducida, y que un porcentaje muy elevado de los insectos han abandonado el área de estudio y no han podido ser capturados, lo cual justificaría la diferencia entre su tasa de recaptura máxima (19%) y la obtenida en los ensayos de ETXEBESTE et al. (2015), con medias superiores al 30%. Según esta lógica, el aislamiento de la masa estudiada en este trabajo supondría un freno para la dispersión de M. galloprovincialis,

140
ya que la tasa de recapturas, aún con una densidad de trampas menor a la utilizada por TORRES-VILA et al. (2015) es significativamente superior. No obstante, la densidad de trampas utilizada en este ensayo es muy superior a la utilizada en los diseños experimentales de ETXEBESTE et al. (2015) y, sin embargo, las tasas de recapturas son muy similares. Esto parece indicar que, aunque es esperable que el aislamiento suponga cierto freno a la dispersión, esto no significa, ni mucho menos, que la dispersión quede anulada y que, en condiciones de aislamiento como las estudiadas en este trabajo, M. galloprovincialis se comporte como una población cerrada con cierto grado de confinamiento. Por contra, estas diferencias de tasa de recapturas parecen indicar que M. galloprovincialis es capaz de abandonar la masa en búsqueda de nuevos entornos. Esto está en consonancia con la distribución de las recapturas en las trampas. Como puede observarse, aunque la distribución de los insectos nativos en la masa es relativamente homogénea (Figura 5.2), las recapturas se dan mayoritariamente en solo 3 de las 10 trampas, y todas ellas están ubicadas en la parte noroccidental de la masa (Figura 5.3). Puesto que no es esperable una influencia clara de la dirección y densidad de los vientos dominantes en su dispersión dentro de esta masa, esta acumulación de recapturas parecen mostrar la existencia de un efecto borde en la masa, posiblemente relacionado con el regreso de los M. galloprovincialis a la masa después de no encontrar masas cercanas a las que trasladarse. Efectivamente, como puede observarse en la distribución de masas con material hospedante en la Figura 5.1, en la zona norte y noroeste alrededor de la masa estudiada no es posible encontrar manchas de pinar o pinos aislados, mientras que algunas masas dispersas y pies aislados sí son observables en sentido sur y sureste alrededor de la masa estudiada. Estas manchas podrían actuar como islas en la dispersión hacia masas más extensas del sur del área de estudio, mientras que la ausencia de éstas hacia el norte, podrían suponer el regreso de los insectos dispersantes a la masa de partida, razón por la cual las capturas se acumularían, precisamente, en las trampas más cercanas al borde noroccidental de la masa.

141
Sección 5.2 Influencia de la estructura de la masa en la densidad
poblacional de Monochamus galloprovincialis
Introducción Dado el importante papel que está llamado a desempeñar la especie M. galloprovincialis en la dispersión en Europa de la Enfermedad del Marchitamiento del Pino, causada por el Nematodo de la Manera del Pino, B. xylophilus, el conocimiento de sus procesos de selección de hábitat a gran y pequeña escala así como, las variables que afectan a su densidad de población, podría ser de gran utilidad en el proceso de toma de decisiones para la erradicación o contingencia de la enfermedad. Como corresponde a especies forestales de carácter secundario, M. galloprovincialis oviposita exclusivamente en árboles muy estresados o recién muertos, aprovechándose de unos mecanismos de defensa debilitados y de una calidad nutricional de los tejidos leñosos aún no significativamente mermada (HANKS 1999, NAVES et al. 2006a, NAVES et al. 2006c). No obstante, mientras la presencia de arbolado sano o estresado está asegurada en todo momento en cualquier rodal, esto puede no ser así en el caso de arbolado muy decadente o recién muerto, lo que provoca que la ventana de actuación para la ovoposición de estas especies sea especialmente corta y frágil. El caso de los escasos 40 días en los que un árbol muerto por daños mecánicos se mantiene atractivo como material de cría adecuado para M. carolinensis es una clara prueba de ello (ALYA Y HAIN 1985). Tales recursos son efímeros y su disponibilidad es impredecible, muchas veces consecuencia de eventos extremos (incendios, sequía, tormentas, nevadas…) y otras influida por una serie de variables (densidad de la masa, competencia, edad, naturalización, condiciones edáficas, ubicación fitoclimática, falta de gestión,…) por lo que este tipo de insectos han desarrollado mecanismos muy eficientes en la detección de los hospedadores adecuados (SAINT-GERMAIN et al. 2004b, SAINT‐GERMAIN et al. 2007). La existencia de estos mecanismos es la base de la ecología química aplicada a la gestión de plagas y, consecuentemente, del desarrollo de los dispositivos de captura basados en atrayentes cairomonales o feromonales para la captura de insectos perforadores de la madera. Como se ha citado anteriormente, los estudios realizados en este marco conceptual por (PAJARES et al. 2004, IBEAS et al. 2007, IBEAS et al. 2008, PAJARES et al. 2010) desembocaron en el desarrollo de un cebo comercial útil para la monitorización y la captura masiva de M. galloprovincialis (SANCHEZ-HUSILLOS et al. 2015a, TORRES-VILA et al. 2015) . Y aunque recientemente se han llevado realizado estudios demográficos de cerambícidos a través de ensayos de captura-recaptura (DRAG et al. 2011, TIKKAMÄKI Y KOMONEN 2011, TORRES-VILA et al. 2012, TORRES‐VILA et al. 2013, SANCHEZ-HUSILLOS et al. 2015a) ningún estudio ha sido enfocado fundamentalmente a evaluar el efecto de la estructura de las masas en la población de M. galloprovincialis presente en ellas. El objetivo, por tanto, de este estudio es comparar las poblaciones de M. galloprovincialis en dos zonas de diferente edad, naturalización, aislamiento e historial de gestión forestal; y evaluar si los trampeos pueden servir como aproximación a la abundancia de la especie.

142
Material y métodos
Localización y diseño experimental
Dos experimentos de captura-marcado-recaptura de M. galloprovincialis fueron realizados de manera simultánea en dos masas forestales, de características diferentes y distantes entre sí, en los términos municipales de Llucena del Cid y Atzeneta del Maestrat (Castellón) entre los meses de junio y noviembre de 2010. En cada área de estudio fueron instaladas un total de 6 trampas multiembudo cebadas con cebo comercial Galloprotect 2D, agrupadas en 3 bloques. Cada bloque consistió en 2 trampas separadas entre sí de 10 a15 m, con el objetivo de crear una única área de captura (MILLER 2006, GRAHAM et al. 2010, MCMAHON et al. 2010, MILLER et al. 2011) Asimismo, cada bloque fue separado entre sí 500 m siguiendo un transecto a lo largo de cada una de las masas forestales (Figura 5.6). La separación entre las dos masas estudiadas fue de 7,2 km, y entre ambas no hubo continuidad forestal, aunque sí presencia fragmentada de manchas de pinar de pequeña extensión y pinos individuales. Las trampas fueron modificadas para la captura en vivo. La primera área de estudio estuvo localizada en Atzeneta del Maestrat (Castellón), en el monte Bovalar, CS3008M1. Se trató de un pinar adulto de P. halepensis de 88 ha, a una altitud sobre el nivel del mar de entre 375-640 m, con distribución uniforme y alta cobertura (FCC=80%), 8 m de altura dominante de los pies y continuidad forestal en dirección noreste hasta los términos municipales de Culla y Torre d’En Besora. Su grado de naturalización fue alto, con presencia de Q. ilex como especie acompañante con individuos dispersos (FCC=5%). Un gran incendio forestal, en agosto de 2007, alcanzó el monte hasta el límite exacto de la zona de muestreo de este ensayo (Figura 5.6) La segunda área de estudio estuvo localizada en Llucena del Cid (Castelló), el monte Mas de Timor, CS3064M1. Se trató de una repoblación joven de P. halepensis no naturalizada, con una altura dominante de 4 m, con sotobosque pobre o ausente y una cobertura alta (FCC 80%). La masa está situada a una altitud entre 500-660 msnm, con presencia de monte bravo de P. nigra y P. pinaster provenientes de repoblación en zonas anejas al área muestreada y sin que exista continuidad forestal a su alrededor, salvo pies aislados y manchas fragmentadas de regeneración natural de P. halepensis (Figura 5.6). En la localización 1 (Bovalar) las trampas fueron colocadas el 11/06/2010 y fueron muestreadas cada 4-5 días. En la localización 2 (Mas de Timor) las trampas fueron colocadas el día 14/06/2010 y fueron muestreadas, también cada 4-5 días, durante el mismo periodo. En ambas masas todos los insectos capturados fueron marcados individualmente y liberados, en cada revisión de trampas, en el mismo punto de captura. Con el objetivo de evitar el posible efecto sumidero que pudiese existir sobre los insectos liberados en el mismo punto de captura, las trampas fueron retiradas durante 2-3 días antes de la liberación de los insectos. Pasado ese periodo, éstas fueron recolocadas en los mismos puntos. Se sustituyó el cebo comercial cada 45 días.

143
Figura 5.6 Áreas de estudio. En color, superpuesto sobre la fotografía aérea de la zona, las formaciones de pinar
en la región según el IFN3 (¿?’)
Análisis estadístico
Todos los análisis y cálculos fueron realizados con el entorno y lenguaje estadístico R (R CORE TEAM 2015). Los análisis demográficos se realizaron utilizando la librería RMark como interfaz para el paquete MARK (IVAN 2008). Se consideró que las poblaciones de M. galloprovincialis durante los periodos de muestreo fueron de tipo abierto, teniendo en cuenta nacimientos, muertes, emigración e inmigración ocurridos durante esos períodos. Se siguió, por lo tanto, la metodología de Jolly-Seber, bajo la parametrización POPAN, para estimar tres grupos de parámetros primarios, que conducen a tres parámetros obtenidos: nacimientos (Bi), tamaño de la población en el intervalo de muestreo (Ni) y el tamaño total de la población (superpoblación, Ntot). Los parámetros primarios representan: pi, la probabilidad de captura en ocasión i; ø'i, la probabilidad de que un insecto sobreviva entre las ocasiones i e i + 1; y bi, la probabilidad de que un insecto de la superpoblación entre en la población entre los momento i e i + 1 (también conocido como penti, probabilidad de entrada). Estos parámetros primarios se modelan como constantes, (∙) en notación MARK, linealmente dependientes del tiempo (T), dependientes del sexo (Sex), o por tener interacciones aditivas o multiplicativas entre el momento de muestreo y el sexo (T × Sexo). Una gama de modelos para cada parámetro primario fueron definidos y posteriormente ejecutados. El tratamiento factorial de tiempo no fue considerado por el hecho de que en más de una ocasión del muestreo no se obtuvo ninguna recaptura en todos los conjuntos de datos. Los mejores modelos fueron seleccionados utilizando el criterio de información de cuasi-Akaike (AICC). Las estimaciones de Ntot se derivaron de los parámetros medios ponderados de los mejores modelos (ΔAICc <10). Las pérdidas durante la captura, es decir, escarabajos encontrados muertos o en mal estado en las trampas, fueron tenidas en cuenta en el cálculo.

144
Resultados Un total de 575 M. galloprovincialis fueron capturados en el monte Bovalar (240 ♂, 335 ♀) de los cuales 23 (16 ♂, 7 ♀) fueron recapturados en más de una ocasión. De forma paralela, un total de 278 M. galloprovincialis fueron capturados en el monte Mas de Timor (138 ♂, 140 ♀) de los cuales 22 (15 ♂, 7 ♀) fueron recapturados en más de una ocasión. Las medias de las capturas por trampas mostraron diferencias significativas (F1 21 40,97 p<0.001) entre ambas zonas de estudio (Figura 5.7)
Figura 5.7 Capturas medias por trampa de M. galloprovincialis en las
diferentes zonas de estudio (Bovalar y Mas de Timor) Una de las recapturas correspondió a un insecto liberado en el punto 2 de Mas de Timor y recapturada en el punto 1 de Bovalar, 35 días después de su liberación, habiendo recorrido una distancia de 7,2 km. Esta captura no es tenida en cuenta en la estimación de las poblaciones, salvo para asumir el supuesto de que las poblaciones de M. galloprovincialis durante los periodos de muestreo fueron de tipo abierto. Los modelos Jolly-Seber con mejor ajuste (Tabla 5.3) mostraron probabilidades de supervivencia dependientes del tiempo (φ), mientras que la probabilidad de captura (p) o bien se mantuvo constante o bien fue dependiente del sexo, o del sexo y el tiempo para el modelo con mayor (0,99) pero en el monte Bovalar. La probabilidad de entrada (penti) fue dependiente del tiempo y el tamaño de la población en el periodo de muestreo se mantuvo fijo (N). En definitiva, la estructura del modelo de ambos ensayos es, en términos generales, igual, exceptuando la probabilidad de recaptura, fundamentalmente constante en Mas de Timor, y dependiente del sexo y el tiempo en Bovalar.
Tabla 5.3 Resumen del mejor ajuste de los modelos Jolly-Seber (parametrización Popan) y la estimación de la abundancia de la población derivada (Ntot±SE) en los montes Mas de Timor (Llucena del Cid, Castellón) y Bovalar (Atzeneta del Maestrat,
Castellón) en el año 2010. Los parámetros del modelo se mantienen constantes (.), fijos (por ejemplo, 1), con diferencias entre sexos (Sex), con respuesta lineal al tiempo (Time) o presentando interacciones entre parámetros (por ejemplo, Time*Sex). No. P, significa número de parámetros en el modelo. La estimación Ntot derivada de la ponderación del peso de los mejores modelos (ΔAICc <10).
Localiz. Sex Modelo No. P AICc Peso Población bruta (Ntot±SE)
Hembras Machos Total
Mas de Timor
1 φ (Time)p(1)pent(Time)N(1) 6 595.1 0.6226 1326±317.8 1338±318 2664
2 φ (Time)p(Sex)pent(Time)N(1) 7 596.1 0.3774
Bovalar
2 φ (Time)p(Time * Sex)pent(Time)N(1) 9 722.8 0.9994
4165±772.7 4078±770.3 8243 4 φ (~Time * Sex)p(Sex)pent(Time)N(~1) 9 738.6 0.00037 3 φ (~Time * Sex)p(Sex)pent(1)N(~1) 8 739.3 0.00026 1 φ (Time)p(Time)pent(Time)N(1) 7 745.7 1.102e-05

145
La estimación del tamaño de la población para el monte Mas de Timor fue de 2664 M. galloprovincialis mientras que para Bovalar asciendió a 8243 M. galloprovincialis (Tabla 5.3), aunque en la Figura 5.8 puede observarse la fluctuación concreta de la población en Mas de Timor y Bovalar, respectivamente, a lo largo del periodo de vuelo del insecto.
Figura 5.8 Evolución temporal de las estimaciones de abundancia de la población M. galloprovincialis por fecha en el experimento
realizado en 2010 en Mas de Timó (Llucena del Cid, Castelló) y Bovalar (Atzeneta del Maestrat, Castelló), respectivamente. El tamaño de la población ha sido derivado de los datos de marcado y recaptura, modelados bajo método POPAN y a partir de la media
ponderada de los mejores parámetros del modelo

146
Discusión Estando la dispersión del NMP indisolublemente ligada a la ecología de M. galloprovincialis, el conocimiento de las densidades poblacionales de esta especie aparece como uno de los factores fundamentales a tener en cuenta tanto en los análisis de riesgo como en los planes de erradicación y contención, es decir, en el control de la enfermedad, en sentido amplio. Aunque diferentes estudios demográficos de M. galloprovincialis han sido realizados recientemente en la península ibérica (SANCHEZ-HUSILLOS et al. 2015a, TORRES-VILA et al. 2015), ninguno de ellos ha estado enfocado a la evaluación de las características ambientales que puedan influir en estas densidades. Y hasta donde conocemos, éste es el primer estudio demográfico de M. galloprovincialis realizado simultáneamente en dos masas de diferentes características con el objetivo de evaluar sus respectivas densidades poblacionales y las variables que las determinan. Los resultados que aportan los modelos ajustados muestran una población de M. galloprovincialis en la masa forestal ubicada en el monte Bovalar cuatro veces mayor que la del monte Mas de Timor (Tabla 5.3, Figura 5.8). Bovalar forma parte de una masa forestal madura y relativamente continua de P. halepensis ubicada en las cotas más altas de una elevación de terreno que se extiende de Sur a Norte a lo largo de más de 10 km, y en la que este monte ocupa la zona más meridional, en contacto directo con los límites del incendio que asoló la región en 2007. Por el contrario la masa forestal de Mas de Timor es una repoblación joven, en absoluto naturalizada y con un alto grado de aislamiento, por lo que quedó relativamente alejada de los límites del mencionado incendio (Figura 5.6). Tanto la continuidad (y, por tanto, el tamaño) y naturalización de las masas, como la afección del incendio tres años antes de la fecha de estudio, son variables que pueden justificar, por sí mismas, las diferencias poblacionales. En efecto, M. galloprovincialis es considerado una plaga forestal de hábitos secundarios, es decir, que su colonización se da sobre pies muy debilitados, moribundos o recién muertos (HELLRIGL 1971, FRANCARDI Y PENNACCHIO 1996, NAVES et al. 2006a, AKBULUT Y STAMPS 2012, RASSATI et al. 2012), material que no siempre está disponible de forma constante y duradera en la naturaleza (GROVE 2002). La existencia de este efímero material hospedante está ligada a la aparición súbita de grandes cantidades, como consecuencia de incendios o de acontecimientos meteorológicos extremos que provocan muertes por daños fisiológicos o mecánicos en el arbolado, o a la mortalidad natural en las masas forestales (vinculada, generalmente, a las variables de masa y a la gestión forestal). En el caso de estudio, el incendio que tuvo lugar en el año 2007 puso a disposición de M. galloprovincialis una gran cantidad de material hospedante en el que realizar la ovoposición inmediatamente después del incendio. Éste afectó de lleno, como se ha indicado, al monte Bovalar, mientras que sus límites quedaron alejados de Mas de Timor, como consecuencia del aislamiento forestal de esta masa. La relación de los incendios forestales con los aumentos poblacionales de escolítidos y, por lo tanto, con la aparición de focos de perforadores ha sido frecuentemente estudiada (TRÄGÅRDH 1940, HANULA et al. 2002, MCHUGH et al. 2003, BILLINGS et al. 2004, SAINT-GERMAIN et al. 2004b, a, DEROSE Y LONG 2009) y supone que la puesta de material hospedante adecuado a disposición de insectos saproxílicos de hábitos secundarios como M. galloprovincialis se puede prolongar en el tiempo tras la afección por el incendio, en este caso, como consecuencia del ataque de escolítidos. Efectivamente, los trabajos de limpieza de árboles afectados por Tomicus destruens ejecutados por el Servicio de Gestión

147
Forestal de la Generalitat Valenciana (Pérez-Laorga, Com. Pers.), se prolongaron durante varios años en toda la zona del incedio, incluido el monte Bovalar. Y en este mismo sentido, la diferencia de poblaciones locales observada también parece ser coherente con la diferencia de aislamiento, edad y de naturalización entre ambas masas, y es esperable que en una masa continua, adulta, con mayor diversidad de fustes y con mayor biomasa, la probabilidad de encontrar material hospedante adecuado (bien por muerte de arbolado, bien por muerte de ramas) sea más alta que en una repoblación joven y apenas naturalizada. No obstante, en futuros estudios, la presencia de material vegetal susceptible de colonización debería ser cuantificada en la medida de lo posible a través de la realización de transectos ad hoc en las diferentes zonas. Por otro lado, en el ámbito de la gestión, la monitorización de las poblaciones de plagas forestales (incluido M. galloprovincialis) mediante el uso de trampas cebadas con atrayentes cairomonales y feromonales es utilizada de manera general como una aproximación a la cuantificación relativa de las poblaciones de las plagas, entre diferentes zonas o en diferentes momentos, con el objetivo de ajustar la gestión o de prevenir irrupciones epidémicas (ver por ejemplo SCHROEDER 2012, MARINI et al. 2013). Nuestros resultados muestran que esta aproximación tan generalizada debe ser tomada con cautela, o matizada consecuentemente, ya que, si bien es posible observar una relación directa entre el número de capturas de las trampas y el tamaño de la población estimada, esta relación no es en absoluto proporcional. En concreto, mientras que las capturas analizadas para ambas zonas muestran diferencias entre sus medias a razón de 2:1, es decir, el doble de capturas en Bovalar que en Mas de Timor (Figura 5.7); las estimaciones del tamaño de población muestran proporciones algo mayores (3:1) (Tabla 5.3). Esta situación debe ser tenida en cuenta, a futuro, en los análisis de riesgo de plagas, así como en la evaluación de la eficacia relativa del empleo de trampas cebadas con atrayentes cairomonales y feromonales en la disminución de la población en casos de brotes epidémicos. Por último, la captura de uno de los individuos en una masa diferente a la que fue liberado, y que, por tanto, atravesó los más de 7 km que separan ambas zonas de estudio, más allá de mostrar la alta capacidad de dispersión de esta especie (ya constatada en estudios paralelos, ver Capítulo 5.4, demuestra la conveniencia de asumir, en futuros estudios demográficos, que las poblaciones de M. galloprovincialis durante periodos de muestreo largos son de tipo abierto (SANCHEZ-HUSILLOS et al. 2015a), y que la consideración de éstas como tipo cerrado (TORRES-VILA et al. 2015) podría provocar desajustes en las estimaciones.

148
Sección 5.3 Influencia del viento y las variables de masa forestal en las
recapturas de Monochamus galloprovincialis
Introducción La detección de B. Xylophilus en Portugal en el año 1999 (MOTA et al. 1999) generó una gran preocupación en Europa por los potencialmente enormes efectos negativos de esta enfermedad sobre los pinares europeos, y fueron tomadas una batería de medidas para frenar la expansión de la enfermedad por el interior de la Unión Europea (Decisiones de la Comisión Europea 2000/58/CE y 2012/535/UE, primera y última en relación al NMP). La mayoría de las medidas están enfocadas al control de su vector, M. galloprovincialis, dado que la dispersión de la enfermedad está directamente vinculada a la capacidad dispersiva de éste, tanto a su dispersión natural como a su dispersión antropocórica (ROBINET et al. 2009). La dispersión es un mecanismo esencial en la dinámica de poblaciones de todas las especies, y, aunque no en todos, sí en la mayor parte de los insectos pterigota, está ligada al vuelo, ya que ofrece evidentes ventajas frente a otros tipos de desplazamiento. La capacidad de vuelo y dispersión de M. galloprovincialis ha sido estudiada recientemente, y aunque los bioensayos con molinos de vuelo han mostrado capacidades potenciales de vuelo cercanas a los 16 km de promedio (DAVID et al. 2014), estudios de marcaje-liberación-recaptura han establecido curvas de dispersión en masas arboladas con porcentajes muy bajos de insectos que alcanzan los 5 km (ETXEBESTE et al. 2015). Ambos métodos son a la vez incompletos y complementarios, puesto que ambos poseen defectos diferentes y difícilmente subsanables. Por un lado los bioensayos con molinos de vuelo no toman en cuenta ninguna de las variables ambientales que pueden interferir en el vuelo de los insectos, a la vez que existe una constante estimulación al vuelo provocada por los mecanismos de sujeción de los insectos al molino (RANIUS 2006). Y por otro lado, la utilización de trampas cebadas con atrayentes es defectuosa en la medida en que el muestreo riguroso de áreas amplias de territorio supone una la utilización de una cantidad inabarcable de trampas, especialmente en insectos con grandes capacidades de vuelo, como M. galloprovincialis (NATHAN et al. 2003a, YAMAMURA et al. 2003). En este contexto, el inicio, la selección de la dirección o la finalización del vuelo en condiciones naturales se da como resultado de una serie de cambios endógenos en el comportamiento del insecto, ligado tanto al tiempo de vuelo como a la percepción de nutrido grupo de señales de todo tipo: térmicas, olfactorias, visuales, etc. (COMPTON 2002). En concreto, los compuestos volátiles emitidos por plantas u otros insectos poseen la capacidad potencial de proveer de información altamente específica a los insectos receptores (no sólo sobre la presencia de hospedantes, sino también sobre sus estados fisiológicos o sobre la presencia de otros herbívoros o depredadores). Pero la probabilidad de su detección disminuye con el aumento de la distancia al centro de emisión, y los gradientes de olor están sujetos a las turbulencias del aire y no son fácilmente modelizables (MURLIS et al. 1992, CARDÉ 1996). Además, éstos son necesariamente detectados a barlovento y frecuentemente de manera intermitente, por lo que los insectos deben volar contra la dirección dominante del viento para alcanzar la fuente de emisión (COMPTON 2002). Aun así, en el proceso de detección de la fuente de emisión de los atrayentes, la disponibilidad de señales visuales de la propia fuente de olor, la distribución aérea de olor afectada por turbulencias, el viento o los niveles de luz, entre otras variables, influyen en el éxito de estas maniobras (CARDÉ 1996). En definitiva, el vuelo dispersivo de M. galloprovincialis parece ser un evento muy complejo apenas entendido y cuya comprensión facilitaría la toma de decisiones en el manejo de la

149
expansión del NMP. El objetivo de este trabajo es proporcionar información sobre la influencia de determinadas variables medioambientales como el viento o las variables de masa forestal en los vuelos y la dispersión del M. galloprovincialis mediante experimentos de marcaje-liberación-recaptura realizados en campo.
Material y Métodos
Localización y diseño experimental
Un conjunto de 4 experimentos de marcaje-liberación-recaptura de M. galloprovincialis fueron instalados en la provincia de Castelló entre los años 2010 y 2012 (Figura 5.9). Los experimentos se desarrollaron secuencialmente y de manera coordinada a lo largo de tres años de forma que cada experimento fue diseñado teniendo en cuenta los resultados obtenidos en los experimentos anteriores, por lo cual, algunos aspectos de su delineación experimental fueron consecuentemente modificados año tras año para la consecución de nuevos objetivos. Los dispositivos de capturas estuvieron formados por trampas cross-vane (Crosstrap®) o multi-embudo modificadas (ÁLVAREZ et al. 2015) cebadas con atrayente cairomonal-feromonal Galloprotect Plus ®, compuesto por dos difusores: uno con feromona de agregación (2-undeciloxi-1-etanol) y otro con sustancias cairomonales (Ipsenol y 2-metil-3buten-1-ol, completado con dos difusores de (α)-pineno (PAJARES et al. 2004, IBEAS et al. 2007, PAJARES et al. 2010) . La renovación de los cebos se realizó según las indicaciones fijadas del fabricante (cada 6 semanas). El marcado de los insectos fue individualizado. Para la fijación de las etiquetas (mayoritariamente sobre el pronoto y en pocos casos la parte proximal del élitro izquierdo) se utilizó un pegamento de cianoacrilato con consistencia de gel.
Figura 5.9 Localización de las diferentes áreas de estudio donde han sido realizados los experimentos: Jérica, (Exp2010/b), Altura (Exp2011/a) y Atzeneta del Maestrat
(Exp2012/b y Exp2012/b), todos ellos en la provincia de Castellón. El diseño experimental común a todos los experimentos fue la colocación de un conjunto de trampas, cebadas con atrayentes, a distancias concretas alrededor de un punto fijo desde el cual fueron liberados los ejemplares de M. galloprovincialis previamente marcados. Las trampas se colgaron de las ramas inferiores de los pinos o de una cuerda entre dos árboles, quedando la parte superior de la trampa a aproximadamente 1,8 m del suelo.

150
La liberación de insectos no nativos supuso su cría en laboratorio. Ésta se realizó a partir de trozas provenientes de árboles colonizados por M. galloprovincialis en condiciones naturales. Las trozas fueron trasportadas a laboratorio e introducidas en jaulas y bidones de cría a temperatura ambiente a la espera de la emergencia de los adultos. Los adultos, una vez emergidos, fueron aislados individualmente en botes de plástico con un papel humedecido y reservados a 4ºC hasta el momento de su marcado y liberación. Se intentó no mantener a los insectos en estas condiciones más de 3 días.
Experimento 2010/b (Jérica, Castellón)
Se realizó en el MUP La Muela, Roquetillo y Feliciano (Jérica, Castelló) entre en 29/05/2010 y el 01/11/2010. El área de muestreo es una repoblación naturalizada de P. halepensis a una altitud entre 520 y 850 m. Se distribuyeron 16 trampas en tres círculos concéntricos alrededor de un punto de liberación de los insectos elegido al azar dentro del monte, y a diferentes distancias de éste (4 trampas a 100m, 4 trampas a 250m y 8 trampas a 500m). Las trampas estuvieron enfrentadas dos a dos y se mantuvo una estructura de roseta entre círculos concéntricos (Figura 5.10). Otras 6 trampas multi-embudo modificadas fueron colocadas fuera del área de muestreo principal para que actuasen como trampas vigía de posibles vuelos de larga distancia: a 2500 m de distancia del punto de liberación (39º 52’ 39.72’’ N; 0º 32’ 14.03’’ O) y 3 más a aproximadamente 5000 m. La frecuencia de muestreo fue semanal. Un total de 487 individuos de M. galloprovincialis fueron liberados en la zona de estudio, habiendo sido criados en laboratorio. Asimismo un total de 670 insectos (430♀, 240♂) capturados fueron marcados y liberados de nuevo desde el punto de liberación.
Figura 5.10 Localización de las 16 trampas del Exp2010/b en el área de muestreo principal. Fuera de esta área fueron colocadas
otras 6 trampas vigía de vuelos de larga distancia. En color verde las masas forestales de P. halepensis según el IFN3 (2008).

151
Experimento 2011/a (Altura, Castelló)
Un segundo experimento en esta sección se desarrolló en el MUP Las Boqueras (Altura, Castelló) entre el 15/06/2011 y el 20/11/2011. Las altitudes en la zona oscilaron entre los 400-1200 msnm. La amplia zona de estudio (> 48.000 ha) estuvo dominada por rodales irregulares de pino P. halepensis, aunque pudieron encontrarse otras especies de pino (P. nigra, P. pinaster, o P. sylvestris), nunca en proporciones superiores al 5% del rodal. Treinta y seis trampas modificadas fueron utilizadas para monitorizar la dispersión de M. galloprovincialis bajo dos regímenes de captura. Por un lado, un conjunto de 19 trampas se colocaron con una distribución regular al tresbolillo (disposición 3-4-5-4-3) en un rodal de pinos continuo aunque heterogéneo en su estructura de masa de aproximadamente 400 ha. Este conjunto de trampas se mantuvo en el rodal durante intervalos de 6 días de promedio, y fueron retiradas del monte durante intervalos complementarios de 4 días de promedio, de manera que durante este periodo no existiese ninguna fuente de feromona artificial en el rodal (momento aprovechado para la liberación de los insectos con el objetivo de poder evitar posibles interferencias de las trampas en la dispersión de los insectos). Por otra parte, 17 trampas adicionales fueron instaladas en manchas de bosque con un predominio de P. halepensis en dos anillos concéntricos a distancias de 4 a 6 km y 10.8 km adaptándose la distribución de las manchas de vegetación a esas distancias (Figura 5.11). Un total de 1568 ejemplares de M. galloprovincialis fueron liberados en la zona de estudio. De éstos, un grupo de 473 individuos habían sido criados en laboratorio y fueron liberados desde el punto de liberación, coincidente con la trampa central (Figura 5.11), en el momento en que las trampas se habían retirado del monte, es decir, en ausencia de fuentes artificiales de feromona y cairomona. Otro grupo de 463 individuos de origen nativo (es decir, capturados y marcados) fue liberado en dos puntos de la red central, a 250 m de la trampa central (equidistante entre la trampa 15 y la 8, y la trampa 15 y la 14, respectivamente). Un último grupo de 632 M. galloprovincialis nativos fueron capturados, marcados y liberados en el mismo punto de su captura en el momento de retirada de las trampas (es decir, de nuevo sin presencia de fuentes artificiales de atrayente en campo) (Figura 5.11). Para más detalles, los procedimientos y resultados de esta experiencia se hallan redactados en ETXEBESTE et al. (2015).
Figura 5.11 Localización general de las trampas del Exp2011/a, ubicado en Altura (Castellón) en el área de estudio total y en el área
central, respectivamente.
Experimento 2011/b (Atzeneta del Maestrat, Castelló)

152
Se desarrolló en los Términos Municipales de Atzaneta del Maestrat, Torre d’En Besora, Culla, Benafigos y Vistabella del Maestrat (Castelló) entre el 29/06/2011 y el 22/11/2011. El área de muestreo es un mosaico agrícola-forestal compuesto, en términos generales, por una masa arbolada continua de P. halepensis en una elevación del terreno que se extiende de sur a norte a lo largo de más de 10 km, flanqueada por dos valles a este y oeste ocupados principalmente por terrenos de cultivo (olivos y almendros), y flanqueados éstos, de nuevo a este y a oeste respectivamente, por masas forestales continuas que ocupan las cotas más altas de las elevaciones de terreno. Se colocaron un total de 19 trampas tipo multi-embudo y Crosstrap® modificadas. conforme a la siguiente distribución: 10 trampas en la misma masa forestal donde se liberaron los insectos, a aproximadamente 3, 4 y 5 km del punto de liberación (40º 14’ 14.73’’N; 0º 7’ 31.57’’ O) en dirección norte y sur; 5 trampas en masas arboladas separadas por terrenos de cultivo y situadas a aproximadamente 3, 4, 5, 6 y 7 km al Oeste del punto de liberación; y 3 trampas en masas arboladas separadas por terrenos de cultivo a aproximadamente 3, 4 y 5 km al Este del punto de liberación (Figura 5.12). La frecuencia de muestreo fue semanal. Los insectos capturados fueron marcados y liberados desde el punto de liberación tras cada muestreo semanal. No se liberaron insectos criados en laboratorio.
Figura 5.12 Localización general de las trampas en el área de estudio del Exp2011/b, ubicado en Atxeneta del Maestrat (Castellón).
Experimento 2012/b (Atzeneta del Maestrat, Castelló)
Un experimento de marcaje-liberación-recaptura de M. galloprovincialis fue realizado en una parcela de 1,5 km por 1,5 km en los términos municipales de Atzeneta del Maestrat (Castelló) y Torre d’En Besora (Castelló) en el año 2012. El paisaje del área de estudio es un fondo de valle

153
de matriz agrícola de secano (fundamentalmente cereales, almendros y olivos) con un mosaico forestal en el cual predominan masas adultas de P. halepensis, formando manchas de extensión variable. El valle se encuentra flanqueado por dos elevaciones de terreno que se extienden en dirección noreste- suroeste a lo largo de más de 10 km y ocupadas por masas forestales adultas de P. halepensis relativamente continuas (Figura 5.13). En la parcela de estudio fueron colocadas un total de 16 trampas tipo Crosstrap® modificada (GALLEGO et al. 2012), dispuestas en gradilla con una separación entre trampas de 500 m. La gradilla fue colocada en el área de estudio de manera aleatoria de modo que las trampas ocuparon puntos en el interior de las masas forestales o fuera de ellas. El experimento se mantuvo entre el 26/06/2012 y 02/11/2012. Un total de 476 individuos de M. galloprovincialis fueron liberados en la zona de estudio, habiendo sido criados en laboratorio provenientes de árboles quemados en el incendio forestal ocurrido el 02/07/2011 en Real de Montroi (Valencia) (Capítulo 2). Tras la emergencia, los insectos fueron conservados de forma individualizada a una temperatura de 4-8ºC, con hidratación pero sin alimentación. El tiempo entre la emergencia de los adultos y su liberación fue menor de una semana, y sólo fueron liberados los insectos que se encontraban en condiciones óptimas para la evaluación de su dispersión. La liberación de los insectos inmaduros se realizó desde el punto central de la gradilla (Figura 5.13). Se realizó un muestreo semanal de todas las trampas.
Figura 5.13 Localización general de las trampas en el área de estudio del Exp2012/b, ubicado en Atzeneta del Maestrat (Castellón)
Análisis estadístico
Todos los cálculos y representaciones han sido realizados bajo el entorno de programación y lenguaje estadístico R (R CORE TEAM 2015).

154
Influencia del viento
El estudio del viento fue abordado mediante el cómputo de una regresión entre las variables ‘viento’ y ‘desplazamiento’. Para ello se procedió de la siguiente manera:
1. Tomando las fechas de suelta y recaptura como referencia, se filtraron los datos meteorológicos comprendidos entre ambas fechas, de forma que pudiese evaluarse exclusivamente las variables de viento que afectasen al periodo de dispersión de cada insecto.
2. Los datos de viento disponibles estuvieron compuestos por tres observaciones diarias de dirección e intensidad tomadas a las 9, 14 y 18 horas. Estos datos se convirtieron a variables circulares utilizando la librería circular de R (AGOSTINELLI Y LUND 2013).
3. Para computar la dirección dominante se siguió el siguiente algoritmo a. Si el número de días con registros válidos de vientos entre la suelta y la
recaptura es 1, no se realizó ningún cálculo, y se seleccionó la dirección correspondiente al día de suelta.
b. Si el número de días con registros válidos de vientos entre la suelta y la recaptura es 2, se tomó como dominante el primero cronológicamente, asumiendo que en el caso de que el desplazamiento de M. galloprovincialis se viese afectado por el viento, lo sería inmediatamente después de la suelta, ya que los insectos se lanzan al vuelo al liberarlos.
c. En el resto de los casos (la gran mayor parte), se calculó la densidad estadística de observaciones de vientos. Una vez computada la densidad, se halló el pico de tal curva, que se estableció como la dirección dominante. En el Anejo 6.1 pueden observarse algunos ejemplos de la estimación del viento dominante para el periodo de vuelo de cada insecto particular.
4. La velocidad asociada a la dirección dominante, se calculó teniendo en cuenta el registro asociado a la dirección del viento cuando la lectura es única (3.a y 3.b), y para el resto los registros en un intervalo de +/- 50 grados de la dirección dominante. La velocidad asociada resultante, fue la velocidad media de los registros. El empleo de abanicos más agudos dejaron registros fuera con mucha frecuencia, resultando en datos no disponibles, de ahí que se emplease ese intervalo.
Influencia de las características de la masa
La influencia de las características de masa así como de otras variables sintéticas que pudieran relacionadas con la tasa de capturas de las trampas fue evaluada, de manera secuencial, para los experimentos: Exp2010/b, Exp2011/a y Exp2012/b. En todos ellos, diferentes características del rodal donde fue ubicada cada una de las trampas fueron determinadas en un inventario silvícola cualitativo para cada experimento (Tabla 5.4). Se aplicaron modelos lineales generalizados y se realizaron análisis de varianza para cada variable y el sexo, y para insectos nativos y capturados-liberados-recapturados. Sólo se tuvieron en cuenta los niveles de los factores que tiene tres o más trampas asociadas. Tabla 5.4 Características de masa y variables sintéticas utilizadas en la valoración de la colocación óptima de las trampas. Variables Clasificación Espesura Trabada, Completa, Incompleta clara, Incompleta hueca, Completa en bosquetes, Incompleta en bosquetes FCC >70% (Alta), 40%-70% (Media), 10%-40% (Baja), <10% (Muy Baja) Edad Maduro, Joven Estado Diseminado, Repoblado, Monte bravo, Latizal, Fustal

155
Visibilidad Buena, Regular, Mala Topografía Loma, Ladera, Barranco, Llano Ubicación trampa Dentro de masa, Límite de masa, Fuera de masa Exposición Solana, Umbría Orientación N, S, E, O Naturalización Naturalizado, En proceso, No naturalizado
Debido al tamaño y a la fragmentación del paisaje en el área de estudio del Exp2011/b, se analizó la influencia de variables de masa continuas sobre el número de capturas. Para el cálculo de las variables, se realizó un inventario LiDAR con un pixel de 25 metros a partir de los modelos de inventario LiDAR ajustados para el Monte de Utilidad Pública (V42) Dehesa del Rebollo. Para ello se utilizó la información LiDAR proveniente del Plan Nacional de Ortofotografía Aérea (PNOA-LiDAR) con una densidad de 0,5 puntos/m2 (1 pulso LIDAR cada 2 m2) y que fue capturada en el año 2009.
Para cada uno de los pixeles de 25 metros del área de estudio, a partir de la altura sobre el suelo de cada retorno de la nube de puntos LiDAR, se estimó una batería de estadísticos de estructura de la vegetación (altura mínima, máxima, mediana, media, desviaciones típicas, percentiles de alturas, estadísticos de dispersión, etc.). Se definió una altura sobre el suelo de 2 metros para diferenciar los retornos LiDAR correspondientes a vegetación arbórea de los estratos inferiores (matorral o estrato de regeneración).
Los estadísticos estimados permitieron describir la estructura de la vegetación de forma continua en la superficie del territorio y se mostró su relación con las principales variables de masa forestal. La fracción de cabida cubierta del LiDAR (LFCC) se definió para cada pixel como el porcentaje de primeros retornos por encima de 2 m.
Los modelos LiDAR utilizados permitieron estimar, a partir de los estadísticos de estructura de la vegetación del LiDAR, las siguientes variables de masa: i) número de pies (N), área basimétrica (G), diámetro cuadrático medio (Dg), altura dominante (Ho), volumen con Corteza (VCC), incremento en Volumen (IAVC) y Biomasa aérea (W). Los modelos utilizados fueron potenciales y multivariables, ajustados con el método de mínimos cuadrados ordinarios a partir de la linealización de los modelos aplicando logaritmos a ambos lados (Tabla 5.5).
Tabla 5.5 Valores del sesgo, error absoluto medio (eam) y error cuadrático medio (ecm) para los valores absolutos y relativos obtenidos en el ajuste de los modelos aplicados al área de estudio.
Estrato Variable promedio sesgo Sesgo relativo (%) eam eam relativo (%) ecm ecm relativo (%)
Pinus halepensis
Ho 12,90 0,90 7,00 1,70 13,50 2,20 17,10 N 237,60 18,20 7,70 73,40 30,90 107,50 45,30 G 10,30 -0,20 2,20 1,80 17,90 2,60 25,00 V 45,40 -0,20 0,50 10,00 22,00 13,20 29,00
IAVC 1,30 0,00 0,30 0,30 26,80 0,40 35,30 W 23,00 -0,10 0,40 5,00 21,80 7,10 31,00
Los promedios de cada variable de masa fueron obtenidos de la superficie contenida en un radio de 500 m alrededor de la trampa, como aproximación al área de muestreo efectivo (sampling efective area) establecido para estos dispositivos de trampeo por (ETXEBESTE et al. 2015). Asimismo, para incluir también la competencia entre los pies de la masa como variable analizada, fue utilizado el índice de Hart estimado como:

156
Esta expresión es la más adecuada para el cálculo de la competencia en masas irregulares de P. halepensis según (RUIZ-PEINADO et al. 2001). El procesado espacial general de todos datos fue realizado utilizando el programa Quantum - GIS 2.10.1-Pisa.

157
Resultados
Influencia del viento
La influencia del viento fue analizada para cada experimento teniendo en cuenta el diferente momento horario de toma de datos del viento (9h, 14h y 18h). Por economía de espacio, en la Figura 5.14 se representa un ejemplo la regresión entre el viento y el desplazamiento para cada experimento, puesto que el resto de análisis (tomando en cuenta el viento a diferentes horas) mostraron la misma de falta de correlación entre ambas variables y, por tanto, resultados equivalentes (Anejo 5.2). El análisis de la influencia del sexo y de la procedencia de los insectos (nativos o criados en laboratorio) en sus desplazamientos en función del viento dominante a las tres horas diferentes del día tampoco mostró correlación alguna.
a) Exp2010/b (Jérica, Castellón)
b) Exp2011/a (Altura, Castelló)
c) Exp2011/b (Atzeneta del Maestrat, Castelló) d) Expe2012/b (Atzeneta del Maestrat, Castelló)
Figura 5.14 Representación gráfica de la regresión entre el viento (tomado a las 18 h) y el desplazamiento de M. galloprovincialis
durante su periodo en libertad para todos los experimentos realizados. En la Figura 5.15 puede observarse la distribución de las capturas en función de la distancia al punto de liberación y el rumbo del desplazamiento. No se observa ningún sesgo direccional significativo en el desplazamiento realizado por los individuos recapturados.

158
a) Exp2010/b (Jérica, Castellón) b) Exp2011/a (Altura, Castelló)
c) Exp2011/b (Atzeneta del Maestrat, Castelló) d) Expe2012/b (Atzeneta del Maestrat, Castelló)
Figura 5.15 Figura 5.15. Distribución de las capturas de M. galloprovincialis en función de la distancia al punto de liberación y el
rumbo del desplazamiento para todos los experimentos realizados

159
Influencia de las variables de masa forestal
Año 2010. Exp2010/b
En el experimento Exp2010/b (Jérica, Castellón) se capturaron un total de 1228 M. galloprovincialis, de los cuales 1141 lo fueron en una única ocasión. El sex ratio total en las trampas fue 0,81 machos por hembra (Figura 5.16)
Figura 5.16 Capturas totales de M. galloprovincialis por trampa discriminadas por sexo (en rojo las hembras, en azul los machos) en
el área de estudio del Exp2010/b Los resultados de los análisis realizados en el año 2010 (Exp2010/b) no mostraron correlación entre las capturas de insectos nativos y las recapturas de insectos capturados-marcados-liberados totales por trampa (ρ= 0,25, p=0,342).
Respecto a la captura de insectos nativos se observó efecto de las siguientes variables cualitativas sobre el número de capturas:
Espesura (F2 12 5.64, p= 0.019), mostrando más capturas la espesura incompleta en bosquetes.
Visibilidad de la trampa (F2 28 4.04, p= 0.029), Se obtuvieron más capturas en las trampas más visibles.
Orientación (F1 22 5.95, p= 0.023), se capturó un mayor número de insectos a orientación norte que a orientación “todas direcciones”.
Ubicación relativa de la trampa con respecto a la masa forestal (F1,27 23.37, p<0.05), las trampas colocadas en el límite de la masa mostraron mayor número de capturas que las colocadas en el interior.
No se observó efecto significativo en las variables Cobertura y Topografía. El estado de la masa no pudo ser evaluado por poca representatividad en el muestreo. Respecto a las recapturas, no se observó efecto de ninguna variable sobre la captura de las trampas.

160
Año 2011. Exp2011/b
En el Experimento EXP2011/b (Altura, Castellón) se capturaron1835 M. galloprovincialis, de los cuales 135 lo fueron en una única ocasión. El sex ratio total en las trampas fue 0,66 machos por hembra (Figura 5.17).
Figura 5.17 Capturas de M. galloprovincialis nativos por trampa en Altura (Castellón) en el año 2011 (Exp2011/b). En rojo la proporción de hembras y en azul la proporción de machos. El tamaño del círculo es mayor a mayor número de
capturas Los resultados del experimento realizado en 2011 (Exp2011/b) mostraron una correlación ρ= 0,62, con probabilidad 0,004, entre las capturas y recapturas totales por trampa.
En dicho experimento, respecto a la captura de insectos nativos, se observó efecto significativo de las siguientes variables cualitativas sobre el número de capturas:
Cobertura: la cobertura alta (FCC>70%) mostró mayores capturas que la cobertura entre 40-70% de FCC (F1 27 4.9, p= 0.035).
Espesura: el efecto fue equivalente en el caso de la espesura completa, con mayores capturas (F3 33 4.36, p= 0.011).
Visibilidad de la trampa: las trampas colocadas bajo dosel arbóreo mostraron significativamente mayores capturas (F1,31 12.98, p=0.001).
Ubicación relativa de las trampas: las trampas colocadas en el interior de las masas capturaron significativamente un mayor número de insectos (F1,31 7.0, p=0.013)
Estado de la masa: los fustales mostraron un mayor número de capturas que los latizales (F1 31 6,05 p=0,019).

161
Edad: Los rodales maduros mostraron mayores capturas que los jóvenes (F1,35 5,02 p=0,032)
El resto de variables analizadas (orientación y topografía) no mostraron efectos significativos sobre el número de capturas.
En los análisis realizados para los insectos recapturados, aunque se detectaron patrones muy similares a los descritos para los insectos nativos, no se observó efecto significativo de ninguna de las variables analizadas, probablemente como consecuencia del menor tamaño de la muestra.
Los análisis de las variables de masa continuas obtenidas del inventario LIDAR mostraron altas correlaciones entre el número de capturas de insectos nativos y dichas variables (Tabla 5.6)
Tabla 5.6 Matriz de correlaciones de las variables de masa obtenidas mediante inventario LIDAR (LFCC, fracción de cabida cubierta; Ho, altura dominante; G, área basimétrica y V, volumen) y el número de capturas (Mongal).
Mongal MEANLFCC SUMLFCC MEANHo SUMHo MEANG SUMG MEANV SUMV Mongal 1 0.662 0.6611 0.692 0.6915 0.692 0.6915 0.6559 0.6556 MEANLFCC 0.662 1 1 0.9727 0.9735 0.9821 0.9824 0.9743 0.9745 SUMLFCC 0.6611 1 1 0.972 0.9728 0.9817 0.982 0.9739 0.9741 MEANHo 0.692 0.9727 0.972 1 0.9999 0.9942 0.9939 0.9893 0.9891 SUMHo 0.6915 0.9735 0.9728 0.9999 1 0.9946 0.9943 0.9896 0.9895 MEANG 0.692 0.9821 0.9817 0.9942 0.9946 1 1 0.9948 0.9948 SUMG 0.6915 0.9824 0.982 0.9939 0.9943 1 1 0.9947 0.9948 MEANV 0.6559 0.9743 0.9739 0.9893 0.9896 0.9948 0.9947 1 1 SUMV 0.6556 0.9745 0.9741 0.9891 0.9895 0.9948 0.9948 1 1
Como ejemplo, en la Figura 5.18 se muestra la relación lineal existente entre número de capturas y la fracción de cabida cubierta (FCC) así como la altura dominante (Ho).
Figura 5.18 Representación gráfica de la relación entre la captura de M. galloprovincialis y las variables fracción de cabida cubierta (FCC) y altura dominante (Ho) obtenidas de inventario
LIDAR en el ensayo realizado en Altura (Castellón) en 2011 (Exp2011/a).

162
En las Figura 5.19 y Figura 5.20, pueden observarse las variables tomadas de referencia para el ajuste de un modelo (Ho y FCC) representadas en el área de estudio junto a la localización de las trampas.
Figura 5.19 Representación gráfica de las alturas dominantes (Ho)
en el área de estudio (mayor altura representada con mayor intensidad de color)
Figura 5.20 Representación gráfica de la fracción de cabida
cubierta (FCC) en el área de estudio (mayor cobertura representada con mayor intensidad de color)
Asimismo también fue posible observar altas correlaciones entre dichas variables y el número de recapturas (Tabla 5.7).
Tabla 5.7 Matriz de correlaciones de las variables de masa obtenidas mediante inventario LIDAR (LFCC, fracción de cabida cubierta; Ho, altura dominante; G, área basimétrica y V, volumen) y el número de recapturas de M. galloprovincialis (Recapts).
Recapts MEANLFCC SUMLFCC MEANHo SUMHo MEANG SUMG MEANV Recapts 1 0.623 0.6198 0.6527 0.6485 0.6508 0.6476 0.6158 MEANLFCC 0.623 1 1 0.9727 0.9735 0.9821 0.9824 0.9743 SUMLFCC 0.6198 1 1 0.972 0.9728 0.9817 0.982 0.9739 MEANHo 0.6527 0.9727 0.972 1 0.9999 0.9942 0.9939 0.9893 SUMHo 0.6485 0.9735 0.9728 0.9999 1 0.9946 0.9943 0.9896 MEANG 0.6508 0.9821 0.9817 0.9942 0.9946 1 1 0.9948 SUMG 0.6476 0.9824 0.982 0.9939 0.9943 1 1 0.9947 MEANV 0.6158 0.9743 0.9739 0.9893 0.9896 0.9948 0.9947 1 SUMV 0.6135 0.9745 0.9741 0.9891 0.9895 0.9948 0.9948 1
Los modelos ajustados (Figura 5.21) también mostraron una relación lineal entre el aumento número de recapturas y el aumento de la fracción de cabida cubierta (FCC) así como el de la altura dominante (Ho), como ya sucedió con respecto al número de capturas.

163
Figura 5.21 Representación gráfica de la relación entre las recapturas de M. galloprovincialis y las variables fracción de cabida cubierta (FCC) y altura dominante (Ho) obtenidas de inventario
LIDAR en el ensayo realizado en Altura (Castellón) en 2011 (Exp2011/a).
También fue estudiada la relación entre el número de capturas y la competencia, expresada a través del índice de competencia de Hart para P. halepensis en la península Ibérica (MONTERO et al. 2000)
El modelo ajustado mostró una relación lineal entre el aumento de capturas y la disminución del índice de competencia de Hart (t -4.635, p<0.05) (Figura 5.22)
Figura 5.22 Representación gráfica de la relación entre las capturas de M. galloprovincialis y el Índice de Hart, calculado sobre variables obtenidas de inventario LIDAR en el ensayo realizado
en Altura (Castellón) en 2011 (Exp2011/a).

164
Año 2012. Exp2012/b
En el experimento EXP2012/b (Atzeneta del Maestrat, Castellón) se capturaron un total de 1547 M. galloprovincialis nativos. Un total de 85 insectos fueron recapturados de los 476 inmaduros liberados. El sex ratio total en las trampas fue 1,17 machos por hembra.
Respecto a la captura de insectos nativos (Figura 5.23), se repiten los mismos patrones observados en 2011 de efecto de las variables sobre las capturas. En resumen puede constatarse efecto significativo, fundamentalmente, de la ubicación de la trampa dentro o fuera de los parches de vegetación (F2 28 28,23 p<0,05) y, consecuentemente, de todas las variables vinculadas a la presencia de pinar en el área de estudio, es decir, máximas capturas en: coberturas altas, mayores al 70% (F1 27 19,4 p<0,05); espesuras completas (F1 24 27,57 p<0,05), orientación Oeste (F1 29 38,9 p<0,05), ubicación topográfica en ladera frente a llano (F1 29 38,9 p<0,05), ubicación de la trampa interior a la masa (F1 28 21.31 p<0,05). Sin embargo, el efecto observado no fue significativo en el caso de la visibilidad de la trampa (sin tener en cuenta las trampas ubicadas fuera de la masa forestal que, naturalmente, siempre son visible). A modo de resumen, la relación del porcentaje de superficie ocupada por masa forestal en los 500 m alrededor de cada trampa con el número de capturas obtenido también mostró una correlación significativa (R2 = 0.94, P<0.001).
Figura 5.23 Capturas de M. galloprovincialis nativos por trampa en Atzeneta del Maestrat (Castellón) en el año 2012 (Exp2012/b). En rojo la proporción de hembras y en azul la proporción de machos. El tamaño del círculo es mayor a mayor número de capturas.

165
Los resultados de los análisis no mostraron correlación significativa entre las capturas de insectos nativos y las recapturas de insectos criados-marcados-liberados totales por trampa (ρ= 0,09, p=0,747).
En los análisis realizados para los insectos criados en laboratorio, liberados y recapturados, no se observó efecto significativo de ninguna de las variables analizadas, la presencia de masa forestal alrededor de la trampa no influyó significativamente en el número de recapturas (R2=0,08, P=0,768). No obstante el 79% de las capturas se dio exclusivamente en dos trampas, ubicadas ambas en la masa forestal más cercana al punto de liberación, aproximadamente 250 m de distancia (Figura 5.24).
Figura 5.24 Capturas por trampa de M. galloprovincialis inmaduros y liberados en la zona de estudio (Altura, Castellón) en el año 2011 (Exp2012/b). El tamaño del círculo es mayor a mayor número de capturas.

167
Discusión El conocimiento de las variables que influyen en la dispersión de M. galloprovincialis es fundamental para afrontar el control del PWD, ya que su difusión a lo largo de Europa, depende, en último término, de la dispersión del vector. No obstante, pese a su gran importancia, esta cuestión ha sido poco abordada en los estudios desarrollados hasta ahora. El efecto de las plumas de semioquímicos en el vuelo de los insectos, aunque en absoluto suficientemente comprendido, ha sido estudiado en laboratorio para algunas especies de cerambícidos, mostrándose que la difusión de volátiles afecta a la dirección del vuelo del insecto y a su comportamiento en el proceso de búsqueda de alimentación o apareamiento (BARATA Y ARAÚJO 2001). No obstante son esperables grandes diferencias entre los ensayos de laboratorio y los experimentos de campo, en la medida en que el movimiento del aire, en condiciones naturales, está sujeto a múltiples y muy dinámicas turbulencias atmosféricas (MURLIS et al. 1992, CARDÉ 1996). Nuestros resultados muestran que la recapturas de M. galloprovincialis liberados (tanto nativos como criados en laboratorio) no está influida significativamente por la dirección ni la intensidad del viento dominante, al menos a la escala estudiada y dentro de los rangos de intensidad registrados en los ensayos. Esto hace suponer que existen otras variables que condicionan de manera más determinante el vuelo de esta especie, o que el viento no afecta a la dispersión de este insecto, no durante todo el tiempo transcurrido entre la suelta y la recaptura, sino sólo puntualmente, por lo que el conocimiento de su efecto es imposible dentro de los márgenes de este estudio. El análisis ha sido realizado en cuatro experimentos ubicados en áreas de estudio muy diferentes en tamaño y en estructura: desde masas relativamente fragmentadas pero con presencia continua de pinos (Exp2010/b y Exp2011/a), hasta masas verdaderamente fragmentadas, representantes de mosaicos agroforestales, con amplias zonas totalmente desprovistas de pinos (Exp2011/b y Exp2012/b). En ninguno de los experimentos se ha podido comprobar que existiese una orientación principal en los rumbos de los trayectos realizados por los insectos liberados-recapturados (Figura 5.15) y, en cualquier caso, estos rumbos no han mostrado correlación alguna con los de los vientos dominantes de los periodos en los cada uno de uno de los individuos ha estado libre (es decir, desde su fecha de liberación hasta su fecha de recaptura) (Figura 5.14). Asimismo, la influencia del viento tampoco se ha observado por razón de género o de procedencia de los insectos (nativos o criados en laboratorio) Estos resultados no son coincidentes con los escasos estudios realizados hasta la fecha sobre este aspecto. BONIFÁCIO (2009)observó que el vuelo de M. galloprovincialis se orientaba mayoritariamente a favor de viento, concluyendo que éste condicionaba la dispersión de los volátiles y, por tanto, la orientación de los insectos hacia las fuentes emisoras. Los estudios de BONIFÁCIO (2009) se basaron en muestreos de visu y en el vareo de árboles para la recaptura de los insectos marcados y liberados, con una frecuencia de muestreo diaria. En su caso, además, la parcela de estudio tuvo dimensiones muy reducidas (0.3 ha) en relación con las estudiadas en este trabajo, y las distancias recorridas por los insectos recapturados fueron, consecuentemente, mucho menores a las aportadas aquí. Por otro lado, los estudios de BONIFÁCIO (2009), si bien incluyeron trampeos con atrayentes, éstos fueron diferentes a los utilizados en este trabajo, ya que fueron realizados con anterioridad a la publicación de la feromona emitida por los machos de M. galloprovincialis (PAJARES et al. 2010), por lo que las metodologías de captura son diferentes y ambos resultados no pueden ser rigurosamente comparados.

168
Las emisiones de compuestos semioquímicos, en condiciones naturales, consisten en breves ráfagas de olor de muy diversa intensidad y que a mayor distancia de la fuente emisora son más débiles y más intermitentes (MURLIS et al. 1992). La alta frecuencia de muestreo (diaria) y la reducida escala (0,3 ha) utilizadas en los trabajos de BONIFÁCIO (2009) permiten que sus resultados puedan ser comparados relativamente con los de ensayos de laboratorio con túneles de viento (BARATA Y ARAÚJO 2001), pero no deberían ser considerados representativos del comportamiento dispersivo de M. galloprovincialis en condiciones naturales, toda vez que los estudios más recientes (DAVID et al. 2014, ETXEBESTE et al. 2015, SANCHEZ-HUSILLOS et al. 2015b, TORRES-VILA et al. 2015) han mostrado una capacidad de vuelo de M. galloprovincialis mucho mayor a la estudiada por BONIFÁCIO (2009). Por todo ello, parece más interesante relacionar la dispersión de M. galloprovincialis con el comportamiento de las señales de olor a una mayor escala espacial y temporal, aun teniendo en cuenta el incremento de complejidad ya que, a estas escalas, la intensidad de la señal disminuye, aparecen fuertes intermitencias y aumenta la probabilidad de que las turbulencias atmosféricas modifiquen las pautas observadas en laboratorio (MURLIS et al. 1992). En este sentido, y de nuevo con resultados divergentes con los nuestros, GALLEGO et al. (2012) mostraron también la existencia de una correlación entre la orientación de los desplazamientos de los insectos y la dirección de los vientos dominantes en un estudio realizado sobre un paisaje de mosaico agroforestal y en una superficie mucho mayor. En él se forzaba a insectos maduros a dispersarse en busca de alimentación o material de reproducción desde una zona desprovista de masa forestal. No obstante, estos resultados deben ser tomados con cautela, ya que una fuerte co-linealidad es observable entre la orientación de la distribución de las manchas de pinar y la dirección de los vientos dominantes, por lo que parece razonable pensar que podría ser la presencia de estas masas forestales en un paisaje fragmentado la variable determinante en la dirección principal de la dispersión, con independencia del efecto que el viento produzca en ella. El contraejemplo a esta situación concreta puede ser observado en el Exp2012/b de este trabajo (Figura 5.13), que también fue realizado sobre un paisaje de mosaico agroforestal, y en condiciones muy similares a las estudiadas por GALLEGO et al. (2012), pero con la utilización de insectos inmaduros y con una presencia homogénea de trampas en el área de estudio. En este experimento nuestros resultados tampoco muestran una correlación entre la dirección de los vientos dominantes y la dirección de los vuelos, sino que el máximo de capturas se da en las trampas ubicadas en los parches de vegetación de pinar más cercanos al punto de liberación, independientemente de la dirección que tomen los vientos dominantes, lo cual también es coincidente con los resultados de GALLEGO et al. (2012). Y si bien es cierto que la madurez sexual de los insectos podría marcar alguna diferencia, no es esperable que esto suceda ya que los estudios de SANCHEZ-HUSILLOS et al. (2015b) han mostrado patrones de dispersión equivalentes para ambos grupos, aunque con un lapso temporal entre ellos determinado por maduración sexual de los insectos inmaduros. Según HARDIE y MINKS (1999), ZHANG y SCHLYTER (2004), la amplia diversidad de señales infoquímicas presentes en el ambiente determina una secuencia en la elección del material hospedante por parte de insectos perforadores, de forma que, en su proceso dispersivo, en primer lugar se produce una selección del hábitat (detección de la masa forestal), en segundo lugar una selección de la especie hospedante más adecuada dentro de la masa y, por último, la elección del árbol hospedador concreto más adecuado para la colonización. Y son diferentes señales infoquímicas (cualitativa y cuantitativamente) las que determinan cada paso de este proceso. En coherencia con ello, CARDÉ (1996) indicaron que las masas vegetales tienden a formar grandes y difusas plumas de olor, lo cual se considera que mejora la viabilidad de las

169
estrategias anemotácticas, puesto que no requieren necesariamente el encuentro cara a cara típico de la búsqueda de plumas discretas de feromonas con una fuente de emisión puntual. Por todo ello, parece razonable pensar que, en zonas de mosaico agroforestal, el viento pueda tener una mayor influencia en la detección de los parches de vegetación, aunque otras muchas variables puedan intervenir en los rumbos de dichos vuelos, como por ejemplo la disponibilidad de señales visuales de la propia fuente de olor o, fundamentalmente, la cercanía de las mismas. Y sin embargo, en virtud de lo expuesto, también es razonable pensar que, una vez alcanzadas las zonas forestales, cubiertas fundamentalmente por material hospedante, la presencia y distribución de los paquetes de olor, de manera continua y masiva, en el ambiente marque un comportamiento diferente y tomen más importancia las emisiones puntuales de feromonas y otros atrayentes de cara a la elección de las especies e individuos hospedantes concretos (HARDIE Y MINKS 1999, ZHANG Y SCHLYTER 2004). En resumen, los resultados de este trabajo deben ser contextualizados convenientemente dada la complejidad del comportamiento de las plumas de olor a largas distancias. Y, aunque no está en discusión que el viento condiciona la dispersión de los volátiles y que la detección de éstos condiciona a su vez la orientación de los vuelos puntuales de M. galloprovincialis, nuestros estudios han evaluado la influencia del viento en la dispersión del insecto en superficies muy extensas, con registros de vuelos muy largos, y con promedios de periodos en libertad muy amplios (promedio de 14 días entre la liberación y la recaptura), y en estas condiciones naturales, no puede asumirse que exista una influencia significativa del viento en la dispersión de M. galloprovincialis, y, en caso de existir, no puede ser conocida con base en los datos de los que actualmente se dispone. La poca coincidencia con los resultados de estudios previos se debe a que éstos se desarrollan sobre una escala espacial y temporal muy pequeña y con diferente metodología (BONIFÁCIO 2009) o a la existencia de fuertes co-linealidades entre la distribución de las masas y los vientos dominantes (GALLEGO et al. 2012). Con respecto a la influencia de las variables de masa en la captura de M. galloprovincialis, para una adecuada interpretación de los resultados, éstos deben ser analizados de forma secuencial, según se han ido desarrollando los experimentos. Los análisis de las variables discretas (cobertura, espesura, estado, edad, visibilidad de la trampa, ubicación, topografía u orientación) en el año 2010 (Exp2010/b) muestran, fundamentalmente, correlaciones de las capturas máximas con la espesura incompleta por bosquetes, con la alta visibilidad de las trampas y con la ubicación de éstas en el límite de las masas forestales. En este ensayo, el área de estudio abarcó unas 80 ha y estuvo compuesta en esencia por una masa boscosa relativamente continua. El hecho de que los promedios de las capturas de insectos nativos fueran similares en todas las trampas (Figura 5.16), permite interpretar que las variables correlacionadas con los máximos de capturas representan razonablemente bien los criterios básicos para la optimización de las capturas de M. gallprovincialis mediante el trampeo en una misma zona de muestreo, y que, en definitiva, la colocación óptima de las trampas, para maximizar el número de capturas de un rodal, debería adecuarse a una alta visibilidad y una ubicación en zonas límite de la masa. Las máximas capturas en espesuras incompletas en bosquetes parecen reforzar esta misma idea. En 2011 (Exp2011/a), sin embargo, el área muestreada es considerablemente más extensa (aproximadamente 400 ha) y más fragmentada que en 2010, y en ella es posible observar rodales bien diferenciados con estructura de masa muy diferente: desde zonas absolutamente despobladas hasta fustales maduros con espesura completa (Figura 5.17). En este caso, además, la densidad de trampas es menor a la del ensayo desarrollado en 2010 (500 m de distancia entre trampas y distribución al tresbolillo). El análisis de la relación entre dichas variables de masa y el número de capturas de insectos nativos por trampa muestra correlaciones de capturas

170
máximas con: coberturas altas (FCC>70%), espesuras completas, baja visibilidad de las trampas, ubicación de éstas en interior de la masa, rodales maduros y fustales (frente a latizales). Estos resultados, aunque en aparente contradicción con los obtenidos en 2010, representan un enfoque diferente a mayor escala y, por tanto, complementan los análisis previos para explicar con mayor detalle la presencia y las capturas de M. galloprovincialis en campo. Al tratarse de una superficie mayor y con alta diversidad en su estructura forestal, los resultados obtenidos indican preliminarmente que los lugares, dentro de un paisaje claramente fragmentado, en los que es más probable encontrar altas poblaciones de M. galloprovincialis son las masas forestales maduras y densas, lo cual parece estar relacionado, obviamente, con la presencia de material hospedante para la alimentación y la reproducción de M. galloprovincialis. En efecto, en consonancia con lo expuesto, un análisis más detallado de la correlación de las capturas con las diferentes variables de masa, pero tomando esta vez variables continuas obtenidas mediante un inventario LIDAR, y asumiendo un radio de acción de las trampas de 535 m de promedio (ETXEBESTE et al. 2015), mostró relaciones lineales y altas correlaciones (Tabla 5.6) entre las capturas y las diferentes variables: densidad de pies, altura dominante, área basimétrica, fracción de cabida cubierta y volumen (Figura 5.18). Esto parece corroborar que, efectivamente, la presencia de M. galloprovincialis en el territorio está vinculada a la presencia de material hospedante y que la relación entre ambos es lineal y directa, de forma que a medida que aumentan las variables de masa, aumentan también las capturas de M. galloprovincialis. En el año 2012, se avanzó un paso más allá en el estudio de la fragmentación del paisaje y el ensayo es realizado en un mosaico agroforestal, absolutamente fragmentado, con zonas completamente desprovistas de pino y manchas de pinar de dimensiones variables esparcidas por el área de muestreo. En este caso, la variable fundamentalmente correlacionada con la captura de M. galloprovincialis es la presencia de material hospedante en el lugar de colocación de la trampa, es decir, si ésta ha sido ubicada en una masa forestal o fuera de ella, y cuál es el porcentaje de masa forestal en el entorno de las mismas. Las mismas correlaciones observadas en el ensayo anterior (Exp2011/b) son corroboradas aquí de nuevo aunque aparecen algunos matices que pueden ser explicados: la influencia significativa de la orientación o la topografía se debe a que, como puede observarse en las Figura 5.19 y Figura 5.20, la única zona de pinar relativamente continuo y extenso está ubicada en la elevación de terreno que existe al este de la parcela de estudio, y las trampas están ubicadas en una ladera con orientación oeste, mientras que el resto de ellas se ubican en el llano contiguo, por lo que estas dos variables están correlacionadas con la presencia de masa forestal. Por otro lado, la alta correlación entre el porcentaje de masa forestal de la superficie englobada en un radio de 500 m alrededor de la trampa y el número de capturas (R2 = 0.94, P<0.001) avala la necesidad de acometer estudios dirigidos a analizar el efecto de la fragmentación del paisaje en la densidad de M. galloprovincialis, pues es fácilmente observable en la Figura 5.17 que las máximas capturas se dan en las trampas ubicadas en los rodales menos fragmentados, es decir, los ubicados en la elevación del terreno ocupada por una masa de pinar continua al este del área de estudio. En definitiva, aunque todo esto parece evidente, su análisis a una escala mayor no lo es tanto, y nuestros resultados parecen mostrar que, no sólo la cobertura de pinar de las diferentes regiones, sino también la fragmentación del paisaje, pueden tener grandes implicaciones en las densidades de población locales de las diferentes especies de Monochamus y, consecuentemente, en las estrategias de control de la enfermedad en Europa, a través, principalmente, de unos análisis de riesgo ajustados. De hecho, es paradigmático el caso de Bélgica, donde el muestreo de 100

171
trampas sobre 30500 km² sólo ha capturado 7 Monochamus adultos lo largo de 3 años (J.C. Grégoire, Com. Pers.), lo cual parece estar relacionado, también, con la cobertura local de coníferas. Nuestros resultados, en este sentido, avalan la necesidad de acometer estudios a escala europea con el objetivo de tratar de relacionar la abundancia de Monochamus spp. con las características del paisaje, tales como la cobertura local de pino o de otras coníferas, la edad de los rodales, la densidad, el estado sanitario, la fragmentación, etc. y, definitivamente, la resistencia del paisaje o la conectividad. La propia biología del insecto está, naturalmente, relacionada con estos resultados en la medida en que M. galloprovincialis es considerada una plaga secundaria que coloniza árboles recién muertos o moribundos (VIVES 2000). En condiciones naturales, este material adecuado para su reproducción es puesto a su disposición, de manera puntal, como consecuencia de incendios o de grandes daños provocados por eventos meteorológicos extremos (nevadas, vientos). No obstante, de manera continua y extendida a todo el territorio aunque no te forma tan intensa, es la mortalidad natural del arbolado en las masas forestales la fuente basal de material de cría para esta especie, por lo que parece razonable pensar que su abundancia estará vinculada a la cantidad de árboles extremadamente debilitados o moribundos en las masas, es decir, a su estado fitosanitario. La alta competencia entre los pies de una masa forestal está directamente relacionada con la falta de vigor, la debilidad del arbolado y, finalmente, la muerte de ejemplares. Diversos índices de competencia, utilizados en gestión forestal para la programación de las claras (Índice de Har-Becking, Reineke), pueden servir de indicadores de la aparición de pies debilitados o muertos en las masas forestales por excesiva competencia, y suponer una aproximación más completa y directa de la presencia de material hospedante para M. galloprovincialis que las variables de masa, en la medida en que estos índices integran varias de ellas en sus expresiones (RUIZ-PEINADO et al. 2001) Los análisis de la relación entre el número de capturas y el índice de Hart (S) muestran, efectivamente, una relación lineal inversa entre ambas variables, de forma que a menores valores del índice (mayor competencia) las capturas de M. galloprovincialis son mayores. No obstante, el índice de Hart adecuado para el mantenimiento de una masa forestal en el estado adecuado de competencia de forma que se minimice la debilidad provocada por ésta en el arbolado depende de la calidad de estación y de la edad (RUIZ-PEINADO et al. 2001) por lo que no es posible dar un valor genérico que minimice la abundancia de M. galloprovincialis. Las tablas de producción proporcionadas por RUIZ-PEINADO et al. (2001) para P. halepensis en la península Ibérica muestran un rango de valores del índice de Hart entre 30 y 51, siendo mayores los valores en el caso de calidades de estación bajas. Dentro de ese intervalo se dan, precisamente, los máximos de capturas en nuestro ensayo (Figura 5.22), nuestros resultados parecen indicar que las calidades de estación más altas de los pinares de P. halepensis de la península ibérica van a estar correlacionadas con abundancias elevadas de M. galloprovincialis mientras que son esperables índices de Hart mayores en las calidades de estación inferiores (no productivas) y, por tanto, menores abundancias del insecto. Estos resultados son coherentes con los mostrados en el Capítulo 4, que muestran que en la única parcela de la Red MUFFET en la que no fue capturado M. galloprovincialis en 2013 y 2014 fue la Muffet2, que representa un pinar de una especialmente baja calidad de estación ubicado en el piso bioclimático semiárido del sur de la provincia de Alicante. En términos generales, nuestros resultados parecen abrir la puerta a que, desde la perspectiva del control de la enfermedad, puedan ser planteados modelos de gestión enfocados a llevar a las masas forestales hacia estructuras de baja densidad y baja competencia con el objetivo de minimizar la abundancia del vector allá donde fuere necesario.

172
Por último, aparte de lo discutido hasta aquí, merece la pena mencionar que los análisis del Exp2012/b ofrecen un sorprendente resultado: la proporción de sexos total en los insectos nativos capturados está sesgada a favor de los machos, y este sesgo es contrario a todas las observaciones realizadas en los estudios acometidos hasta la fecha (HERNÁNDEZ et al. 2011, GALLEGO et al. 2012, ETXEBESTE et al. 2015, SANCHEZ-HUSILLOS et al. 2015a). Puesto que la feromona utilizada en los dispositivos de captura es segregada por los machos (PAJARES et al. 2010), es esperable que, aunque captura individuos de ambos sexos, la proporción de los mismos muestre un sesgo favorablea las hembra, como ha sido constatado hasta la fecha en un gran número de experimentos. La explicación a esta observación podría estar en el carácter pionero de los machos en la búsqueda de nuevos entornos (MORBEY Y YDENBERG 2001), lo cual sería coherente con una mayor presencia de este sexo en las zonas muy fragmentadas. En resumen, el análisis de la influencia de la estructura de la masa en la captura de M. galloprovincialis a lo largo de estos tres ensayos secuenciales muestra que, en primer lugar, la abundancia de M. galloprovincialis está relacionada significativamente con la abundancia de material hospedante, de manera que el insecto está presente fundamentalmente dentro de las masas forestales, y que esta abundancia es mayor a medida que aumentan las diferentes variables de la masa (densidad de pies, área basimétrica, volumen y altura dominante). Los resultados muestran también que existe una relación lineal inversa entre el índice de competencia de Hart y la abundancia de M. galloprovincialis, por lo que el aumento de competencia dentro de las masas también está relacionado con el aumento de la abundancia del insecto. También la fragmentación del paisaje puede influir en la abundancia de M. galloprovincialis así como en la proporción de sexos presentes en las manchas de vegetación. Y en segundo lugar, los resultados muestran que, a igualdad de condiciones de abundancia de M. galloprovincialis, las trampas serán más eficientes en capturas si son colocadas en zonas visibles y en el límite de las masas objeto de muestreo.

173
Sección 5.4 Capacidad de vuelo a larga distancia en condiciones
naturales Monochamus galloprovincialis
Introducción
El Nematodo de la Madera del Pino (PWN), Bursaphelenchus xylophilus (Steiner and Buhrer 1934), es el agente causante de la Enfermedad del Marchitamiento del Pino (EMP). Aunque es originario de Norte América, actualmente también está establecido en los continentes asiático y europeo. La dispersión de la enfermedad comienza a principios del siglo XX en Japón (MAMIYA 1988) y se extiende posteriormente a China (BAOJUN Y QOULI 1989), Taiwán (RUAN 1996)y Corea del Sur (LA et al. 1999). En 1999 se introduce en la Unión Europea a través de Portugal (MOTA et al. 1999, MOTA Y VIEIRA 2008). La irrupción de la enfermedad en los diferentes países ha supuesto la toma de medidas de contención y erradicación muy intensas. Entre ellas el establecimiento de cinturones de aislamiento para las zonas libres de plaga, en los que se ha cortado y destruido in situ las coníferas, al objeto de impedir la dispersión natural de B. xylophilus y de sus vectores. La anchura de los cinturones de aislamiento ha alcanzado los 3 km en España (JUNTA DE EXTREMADURA, 2008) y los 4 km en China (ZHAO et al. 2008).
La dispersión de esta enfermedad está fundamentalmente ligada a la dispersión de sus vectores (YOSHIMURA et al. 1999, TOGASHI Y SHIGESADA 2006, ROBINET et al. 2009, ROBINET et al. 2011, PUKKALA et al. 2014). El PWN es transportado de árbol a árbol, principalmente, por especies del género Monochamus. Sus especies más destacables como vectores del PWN en las diferentes regiones del planeta son M. alternatus (Asia), M. carolinensis (América) y M. galloprovincialis (Europa). Estas especies presentan una destacable similitud entre sí, en lo referente a su biología y ecología (AKBULUT Y STAMPS 2012). A pesar de ser el principal vector de B. xylophilus en Europa, la capacidad de dispersión natural de M. galloprovincialis ha sido poco estudiada hasta la fecha. Esto se debe a que las especies que componen el género han sido consideradas plagas secundarias inocuas para el arbolado sano; y por ello, no han sido estudiadas en profundidad con anterioridad a la presencia del PWN en sus regiones (AKBULUT Y STAMPS 2012). En este ámbito, diversas técnicas de marcado-liberación-recaptura se han probado extensamente desde hace décadas para estudiar procesos de dispersión de especies, así como para estudiar movimientos individuales y capacidades de dispersión a larga distancia (KOENIG et al. 1996, NATHAN et al. 2003b). Los estudios de PAJARES et al. (2004), IBEAS et al. (2007), PAJARES et al. (2010) han permitido el desarrollo de un atrayente cairo-feromonal muy eficaz en la captura de ambos sexos de M. galloprovincialis. Este atrayente ha sido recientemente distribuido comercialmente por la empresa SEDQ (Barcelona). Disponer de este cebo ha permitido realizar estudios sobre la biología y ecología de M. galloprovincialis en los últimos años. HERNÁNDEZ et al. (2011) y GALLEGO et al. (2012), de acuerdo con las propuestas metodológicas expuestas en Taller sobre Complejos Feromonales de Insectos Forestales, promovido por el Servicio de Sanidad Forestal y Equilibrios Biológicos del Ministerio de Agricultura, Alimentación y Medio Ambiente, realizaron experiencias en los que se pusieron a prueba las técnicas de marcado-recaptura de M. galloprovincialis. Sus trabajos, desarrollados en Teruel y Murcia respectivamente, son revisados aquí y sus resultados son incorporados a los obtenidos mediante estudios similares que, paralela y coordinadamente, fueron realizados en la

174
Comunitat Valenciana, en la misma época, en el marco general de la gestión de plagas y enfermedades forestales. El objetivo de este trabajo es la evaluación de la capacidad de dispersión natural a larga distancia de M. galloprovincialis y, subsidiariamente, la evaluación de la eficacia de las medidas de profilaxis contra la Enfermedad del Marchitamiento del Pino basadas en el establecimiento de cinturones de aislamiento (eliminación total de especies hospedantes susceptibles) en zonas libres de plaga para evitar la dispersión natural de M. galloprovincialis y B. xylophilus. Los resultados de esta sección fueron presentados en un congreso específico de la enfermedad de marchitamiento del pino acontecido en Braunschweig (Alemania) en 2013 y organizado por la International Union of Forest Research Organization (MAS et al. 2013).
Material y métodos
Nueve experimentos tuvieron lugar en tres diferentes regiones del levante de la Península Ibérica: Teruel, Comunitat Valenciana y Murcia, entre los años 2009 y 2012 (Figura 5.25). Todos ellos se basaron en el marcaje-liberación-recaptura de individuos de M. galloprovincialis. Las capturas se consiguieron mediante la utilización trampas cebadas con atrayentes cairomonales-feromonales. El atrayente utilizado fue Galloprotect Plus® La renovación de los cebos se realizó según las indicaciones del fabricante (cada 6 semanas). El marcado de los insectos fue individualizado y para la fijación de las etiquetas (mayoritariamente sobre el pronoto y en pocos casos la parte proximal del élitro izquierdo) se utilizó un pegamento de cianoacrilato con consistencia de gel.
Figura 5.25 Localización de las diferentes áreas de estudio donde han sido realizados los experimentos: Mora de Rubielos (Exp2009/b, Exp2010/c,
Exp2011/c), Jérica (Exp2010/b), Altura (Exp2011/a), Atzeneta del Maestrat (Exp2009/a, Exp2011/b), Jumilla (Exp2011/d) y Camporrobles (Exp2012/c)
El diseño experimental común a todos los experimentos fue la colocación de un conjunto de trampas, cebadas con atrayentes, a distancias concretas alrededor de un punto fijo desde el cual fueron liberados los ejemplares de M. galloprovincialis previamente marcados. Las trampas se colgaron de las ramas inferiores de los pinos o de una cuerda entre dos árboles, quedando la parte superior de la trampa a aproximadamente 1,8 m del suelo. Los ocho experimentos se desarrollaron secuencialmente y de manera coordinada a lo largo de tres años de forma que el

175
diseño de cada bloque de experimentos anuales contempló el objetivo de superar los resultados obtenidos en los años anteriores. En 2009 se desarrolló el Experimento 1 (Teruel) y el Experimento 2 (Castellón). En 2010, el Experimento 3 (Teruel) y el Experimento 4 (Castellón). En 2011 se llevaron a cabo los experimentos 5 (Teruel), 6 y 7 (Castellón) y 8 (Murcia).
Año 2009. En ambos experimentos (Exp2009/a y Exp2009/b) los individuos de M. galloprovincialis liberados fueron insectos nativos capturados en esas zonas.
1. Experimento 2009/a. Se realizó en el MUP Boalar d’Atzeneta (Atzeneta del Maestrat,
Castelló), desde el 26/06/2009 hasta el 20/11/2009. El área de estudio es una masa natural de P. halepensis, situada entre 375 y 640 m de altitud, en la que se colocaron 4 trampas multi-embudo de 12 unidades (Phero Tech Inc. Delta, BC, Canadá) a 900 m del punto de liberación (40º 12’ 54.85’’ N; 0º 8’ 33.83’’ O) y fueron alejándose 100 m, todas ellas, a medida que se produjeron las capturas a cada distancia. La frecuencia de muestreo fue semanal. El marcaje de los M. galloprovincialis no fue individualizado, pero sí diferenciado entre fechas de liberación (semanal).
2. Experimento 2009/b. Se realizó en el paraje denominado Carramacho (Mora de
Rubielos y Cabra de Mora, Teruel) desde el 24/07/2009 hasta finalizado el mes de noviembre. La formación vegetal es una masa natural de P. pinaster y P. nigra salzmannii, entre 1000 y 1300 m de altitud, en la que se colocaron, a 500 m del punto de liberación (40º 18’ 16.08’’ N; 0º 48’ 0.91’’ O) en diversas orientaciones, 6 trampas tipo “TorreLSF” metálica (PÉREZ et al. 2004, HERNÁNDEZ ALONSO et al. 2007). Cada trampa se alejó una vez que se produjo una recaptura en ella. La frecuencia de muestreo fue diaria. Los M. galloprovincialis liberados fueron marcados masivamente (no hubo marcado individualizado) (HERNÁNDEZ et al. 2011).
Año 2010. Como respuesta a los resultados de distancias de vuelo máximas obtenidos en 2009, se diseñaron los experimentos 3 y 4 al objeto de ampliar el área de muestreo y obtener distancias de vuelo mayores. En los experimentos realizados en 2010 se combinó la utilización de M. galloprovincialis criados en laboratorio y la de insectos nativos capturados por las propias trampas. El marcado de los insectos fue individualizado. La cría en laboratorio se realizó a partir de trozas provenientes de árboles colonizados por M. galloprovincialis en condiciones naturales. Las trozas fueron trasportadas a laboratorio e introducidas en jaulas y bidones de cría a temperatura ambiente a la espera de la emergencia de los adultos. Los adultos, una vez emergidos, fueron aislados individualmente en botes de plástico con un papel humedecido y reservados a 4ºC hasta el momento de su marcado y liberación. Se intentó no mantener a los insectos en estas condiciones más de 3 días. Las trampas multi-embudo utilizadas en 2010 incluyeron una modificación que permite mayor eficacia en la captura en vivo de los M. galloprovincialis al impedir su huída. Esta modificación consiste en la utilización de botes recolectores del doble de profundidad así como la extensión del cuello del embudo hacia dentro del bote colector.
3. Experimento 2010/b. Conforme a la metodología ya descrita en el apartado 5.3 del
capítulo, se realizó en el MUP La Muela, Roquetillo y Feliciano (Jérica, Castelló). El área de muestreo es una repoblación naturalizada de P. halepensis a una altitud entre 520 y 850 m. Se distribuyeron 6 trampas multi-embudo modificadas a 2500 m de distancia del punto de liberación (39º 52’ 39.72’’ N; 0º 32’ 14.03’’ O) y 3 más a aproximadamente 5000 m. Las trampas permanecieron fijas en sus posiciones durante todo el periodo, y la frecuencia de muestreo fue semanal.

176
4. Experimento 2010/c. Se realizó en la misma zona del Experimento 1 entre el
07/06/2010 y el 12/12/2010. Se aumentó la cantidad de trampas colocadas hasta un total de 18 trampas tipo “TorreLSF”, que cubrieron un área mayor que en 2009. Quedaron distribuidas inicialmente de la siguiente forma: 6 trampas entre 500 y 1000m, 4 entre 100 y 1500 y 8 entre 1500 y 2000 (Figura 5.26). Posteriormente, las trampas con menos capturas fueron recolocadas a distancias mayores a lo largo del periodo de muestreo llegando a alcanzar posiciones superiores a los 7000 m. La revisión se hizo 3 veces por semana (lunes, miércoles y viernes) (HERNÁNDEZ et al. 2011).
Figura 5.26 Localización de las trampas utilizadas en el Experimento 2010/c en la zona de
Mora de Rubielos, Cabra de Mora y Teruel (Teruel)
Año 2011. El análisis de los resultados de los experimentos realizados en 2010, que mostraron evidencias de la capacidad de M. galloprovincialis de recorrer largas distancias a través de áreas no forestales, permitió diseñar de forma más adecuada los experimentos 5, 6, 7 y 8 (llevados a cabo en 2011 en Teruel, Castellón y Murcia) con el objetivo de constatar vuelos aún mayores que los obtenidos en los años anteriores. En todos los experimentos de 2011 las trampas permanecieron fijas en sus posiciones durante todo el periodo de muestreo.
5. Experimento 2011/a. Conforme a la metodología ya descrita en el apartado 5.3 de este capítulo, se desarrolló en el MUP Las Boqueras (Altura, Castelló) entre el 15/06/2011 y el 20/11/2011. Se trata de una masa natural de P. halepensis a una altitud entre los 800m y 900 m, con presencia de otras especies acompañantes provenientes de repoblación (P. nigra austriaca, P. pinaster y P. sylvestris). Se colocaron 12 trampas Multi-embudo modificadas en las zonas arboladas existentes a distancias de entre 5000 m y 10000 m del punto de liberación (39º 49’ 16.15’’ N; 0º 38’ 31.48’’ O) y en todas las orientaciones, priorizando la fácil accesibilidad conforme a la red de pistas y carreteras de la zona. La frecuencia de muestreo fue semanal.
6. Experimento 2011/b. Conforme a la metodología ya descrita en el apartado 5.3 de este
capítulo, se desarrolló en los Términos Municipales de Atzaneta del Maestrat, Torre d’En Besora, Culla, Benafigos y Vistabella del Maestrat (Castelló) entre el 29/06/2011 y el 22/11/2011. El área de muestreo se trata de un mosaico agrícola-forestal

177
compuesto, en términos generales, por una masa arbolada continua de P. halepensis en una elevación del terreno que se extiende de sur a norte a lo largo de más de 10 km, flanqueada por dos valles a este y oeste ocupados principalmente por terrenos de cultivo (olivos y almendros), y flanqueados éstos, de nuevo a este y oeste, por masas forestales continuas que ocupan las elevaciones de terreno. Se colocaron un total de 19 trampas, Tipo multi-embudo modificada y Crosstrap® modificada (GALLEGO et al. 2012) conforme a la siguiente distribución: 10 trampas en la misma masa forestal donde se liberaron los insectos, a aproximadamente 3, 4 y 5 km del punto de liberación (40º 14’ 14.73’’N; 0º 7’ 31.57’’ O) en dirección norte y sur; 5 trampas en masas arboladas separadas por terrenos de cultivo y situadas a aproximadamente 3, 4, 5, 6 y 7 km al Oeste del punto de liberación; y 3 trampas en masas arboladas separadas por terrenos de cultivo a aproximadamente 3, 4 y 5 km al Este del punto de liberación. La frecuencia de muestreo fue semanal.
7. Experimento 2011/c. Se desarrolló en la misma localidad que los experimentos
Exp2009/b y Exp2010/c, utilizándose el mismo punto de liberación del año anterior (Figura 5.26). Se instalaron 6 trampas a una distancia de 3 km de radio alrededor del punto de suelta. Asimismo se instalaron otras 32 trampas a distintas distancias del punto (entre 5000 m y 15500 m) en zonas arboladas y priorizando la fácil accesibilidad conforme a la red de pistas y carreteras de la zona, aumentando considerablemente el área de muestreo. Se utilizaron dos modelos de trampas: TorreLSF y multi-embudo modificada. La frecuencia de muestreo fue de dos veces por semana (HERNÁNDEZ et al. 2011)
8. Experimento 2011/d. Se desarrolló en el Norte del T.M de Jumilla (Murcia) en un
mosaico agrícola-forestal en el que predomina el pinar de P. halepensis en estado de latizal formando manchas de extensión variable (Figura 5.27). El experimento constó de 17 trampas Crosstrap® (GALLEGO et al. 2012) ubicadas tanto en zonas arboladas como en pinos dispersos en zonas agrícolas y se mantuvo entre el 01/07/2011 y el 30/10/2011. El punto de liberación (38º 37’ 10.28’’ N; 1º 22’ 27.62’’ O) se estableció en una zona agrícola, aproximadamente en el centroide del polígono formado por las trampas. La distancia mínima entre trampas fue de 632 m y la media de 4853 m. La trampa más próxima al punto de liberación se encontró a 2110 m y la más alejada a 8339 m. El 44 % de las trampas quedó ubicado a más de 3 km del punto de liberación y el 56% restante entre los 2 km y los 3 km. Las trampas se revisaron 2 veces por semana.

178
Figura 5.27 Localización de las trampas utilizadas en el Experimento 2011/d en Jumilla
(Murcia)
9. Experimento 2012/c. Se desarrolló en un paisaje de mosaico agroforestal en Camporrobles (Valencia, 39 ° 49'18 "N, 0 ° 38'31" W) durante el año 2012, entre el 06/07/2012 y el 01/11/2012 (Figura 5.28). La altitud en la zona es de 700 msnm aproximadamente. La zona de estudio es una matriz agrícola de secano (fundamentalmente cereal) en la que, en un radio de 3 km, sólo es posible observar un bosquete de pinar de P. halepensis de 5 ha de superficie (Figura 5.28). A distancias mayores de 2 km, algún otro bosquete de pinar de P. halepensis es posible de encontrar, siempre en golpes de superficies muy reducidas, como parte de pinares anejos a casas de campo. La primera gran masa de pinar cercana al área de estudio se encuentra a más de 3 km al oeste, en la provincia de Cuenca, en los alrededores del Embalse de Contreras. Nueve trampas modificadas y cebadas con atrayente cairomonal-feromonal fueron colocadas con una disposición en gradilla en la única masa forestal de la zona de estudio, un bosquete de 5 ha de superficie en el Cerro de la Ermita de San Cristóbal (Camporrobles), (UTM 636425/4389126/30S). Asimismo, otras 4 trampas fueron colocadas en los límites de la masa forestal circundante al Embalse de Contreras, a más de 3 km del punto de liberación, con la intención de que actuasen como vigía de posibles dispersiones a larga distancia de los insectos liberados (Figura 5.28). Un total 387 adultos inmaduros de M. galloprovincialis, criados en laboratorio y marcados individualmente, fueron liberados fuera de la masa muestreada a 3 diferentes distancias de la misma (500, 1000 y 1500 metros) en 3 tandas distintas. Las trampas fueron revisadas semanalmente. Para la liberación de insectos fueron utilizados 4 puntos de liberación, tres al Este de la masa muestreada y uno de ellos al Oeste de la misma (Figura 5.28).

179
Figura 5.28 Área de muestro y ubicación de trampas y puntos de liberación del Experimento 2012/c

180
Resultados Los resultados obtenidos cada año condicionaron el diseño experimental y los objetivos de los experimentos desarrollados en años posteriores.
En 2009, en el Experimento 2009/b se constató la capacidad de M. galloprovincialis de alcanzar distancias de 1800 m. De los 243 insectos liberados fueron capturados 12 ejemplares en 6 trampas, lo que supuso un 4,96% del total. Cinco de ellos fueron capturados a distancias superiores a los 1000 m. La ausencia de marcaje individualizado impidió la constatación de la velocidad de dispersión. El Experimento 2009/a, realizado de forma paralela en el tiempo al Experimento 1, mostró distancias de vuelo cercanas a los 1100 m. El porcentaje de capturas, con un número menor de trampas que el Experimento 1, fue muy bajo (1,42% de los 282 insectos liberados). La máxima velocidad de dispersión fue de 143 m/día.
En 2010, en el Experimento 2010/c, situado en la misma ubicación que el Experimento 2009/b se mostró que M. galloprovincialis es capaz de llegar a recorrer 7100 m en 18 días. El porcentaje de recapturas fue de un 7,32%, habiéndose liberado un total de 1106 insectos. De los 81 que fueron capturados, 18 individuos sobrepasaron la distancia máxima de dispersión encontrada en 2009, y 11 de ellos (14% de las recapturas) superaron los 3000 m, distancia de referencia en la delimitación de áreas de erradicación (EPPO, 2009). La máxima velocidad de dispersión se dio en un ejemplar que fue capturado a 1900 m del punto de suelta, en la primera revisión, dos días después de su liberación (mínimo 950 m/día).
En el mismo año, el Experimento 2010/b mostró también vuelos superiores a los 3000 m. Aunque el porcentaje de recapturas fue muy bajo con respecto a otros experimentos debido a la baja presión de muestreo (de los 487 insectos liberados sólo se recuperaron 3) se obtuvieron resultados positivos en las dos distancias muestreadas (3000 m y 5000 m). Asimismo se produjo el avistamiento, fotografiado y recuperación, por parte de personas ajenas al experimento, de un ejemplar macho de M. galloprovincialis liberado por los autores a 22100 m de distancia 74 días antes de su captura. La mayor velocidad de dispersión se constata en este mismo insecto, 299 m/día.
Los diseños experimentales de 2011 pretendieron, consecuentemente, ampliar la distancia de dispersión de M. galloprovincialis constatada en el año 2010. Por el propio carácter secuencial del diseño los experimentos, los realizados este último año recogen, en términos generales, los resultados más interesantes.
En el Experimento 2011/c se amplió considerablemente el área de muestreo realizada en la misma zona en 2010 (Experimento 3) y se aumentó el número de trampas. La mayor distancia detectada ascendió a 13600 m, recorridos en 10 días. En este ensayo, además, la totalidad de las recapturas obtenidas (23 insectos, un 2.31% del total liberado) alcanzaron los 3000 m, que eran las trampas más próximas, y 8 de ellas sobrepasaron los 5000 m. La máxima velocidad de dispersión alcanzó los 1500 m/día, aunque también hubo capturas obtenidas a 3000 m después de más de 60 días (Figura 5.29-a).
Los experimentos 2011/a y 2011/b proporcionaron datos en el mismo sentido de modo que en el Experimento 6, de los 1440 insectos liberados, 4 de ellos se recapturaron por encima de los 5000 m de distancia del punto de liberación, obteniéndose una distancia máxima a los 7800 m. La máxima velocidad de dispersión fue 403 m/día. En el Experimento 7 se mostró que, de los 17 insectos recapturados (2.57% del total liberado), 9 se recuperaron a distancias mayores de 3000 m, 6 a distancias superiores a 4000 m y 2 de ellos a más de 5000 m. Se obtuvo una

181
velocidad máxima de dispersión de 841 m/día, aunque también se dieron capturas en las distancias mínimas (3000 m) después de más de 40 días (Figura 5.29-b). En el Experimento 2011/d las recapturas supusieron un 11.24% del total: 47 recapturas de los 418 inicialmente marcados y liberados. La mayor distancia registrada fue a 8317 m, no obstante se trató de un insecto que había perdido la etiqueta (sólo se observaron los restos del pegamento), por lo que se desconoce el tiempo que tardó en recorrer la distancia y, como consecuencia, no ha podido ser contabilizado en el cálculo de velocidades de dispersión, ni en el histograma bivariante de la Figura 5.29-c. La velocidad máxima de dispersión fue de 801 m/día, aunque, como ya sucedió en los experimentos 5 y 7, también se constataron periodos de tiempo largos (48 días) en capturas a distancias cercanas a los 3 km. Asimismo, el porcentaje de capturas por encima de los 3000 m fue del 1.7% (7 individuos) del total de insectos liberados.
De forma global y a modo de resumen, a lo largo de los 3 años, se ha obtenido una distancia máxima de dispersión natural de 13600 m recorridos en 10 días (Mora de Rubielos, Teruel, 2011). No obstante, se debe tener en cuenta que uno de los ejemplares liberados en Jérica (Castelló) en 2010 fue fotografiado y capturado por personas ajenas a la investigación, fuera del área de muestreo, a 22100 m del punto de liberación. Se trató de un ejemplar macho de 82 días de vida, que estuvo 8 días a 4 ºC sin alimentación después de la emergencia antes de ser liberado. La media total de las velocidades de dispersión es de 230 m/día, alcanzando un valor máximo de 1500 m/día. En el conjunto de los experimentos no se observaron diferencias entre las recapturas de machos y hembras, ni entre las distancias de vuelo alcanzadas por ambos sexos. De los 21 insectos que recorrieron más de 5 km, 11 son hembras y 10 machos. El noveno experimento (Exp2012/c) queda fuera del análisis porque su objetivo fue constatar máximos alcances en el vuelo de M. galloprovincialis sexualmente inmaduros liberados en áreas desprovistas de material hospedante, y las distancias constatados en él son menores que las observadas en experimentos anteriores. No obstante, por la especificidad de su diseño experimental, los resultados deben ser analizados aparte. En él la tasa de recapturas fue muy baja, 4 capturas de los 387 insectos liberados. Tres de estas capturas se dieron a los 500 m de distancia, la cuarta se obtuvo a 3000m (Tabla 5.8)
Tabla 5.8 Recapturas de M. galloprovincialis inmaduros en Exp2012/c
Fecha liberación Fecha captura Tiempo transcurrido Pto liberación Pto Recaptura Dist 11/07/2012 25/07/2012 14 PL1 T6 500 06/07/2012 25/07/2012 19 PL1 TC1 3000 13/07/2012 01/08/2012 19 PL4 T1 500 18/07/2012 07/08/2012 21 PL4 T6 500
1
Figura 5.29 Histogramas bivariantes de las recapturas con respecto al tiempo en libertad y distancia del punto de liberación (Fig5.29-a: Exp 2011/c. Fig 5.29-b: Exp 2011/a y Exp2011/b. Fig 5.29-c: Exp2011/d)

183
Discusión En los organismos patógenos que se dispersan con autonomía, es decir, sin necesidad de vector, la capacidad de realizar vuelos de largo alcance no significa necesariamente la capacidad de desarrollar nuevas poblaciones (e infecciones) a largas distancias. Si el porcentaje de vuelos de largo alcance es muy bajo, pequeñas poblaciones podrían no sobrevivir debido a, por ejemplo, la incapacidad de encontrar parejas adecuadas en baja abundancia, o debido a las perturbaciones (ambientales o antropogénicas) de un hábitat inadecuado (SMITH et al. 2004). No obstante, como vector de B. xylophilus, M. galloprovincialis puede transportar varios centenares de nematodos en su cuerpo (NAVES et al. 2006b), por lo que la presencia de pocos individuos portantes del nematodo es suficiente para provocar la aparición de un nuevo foco de infección de la enfermedad (TOGASHI Y SHIGESADA 2006, NAVES et al. 2007), sin embargo este proceso puede estar regulado por efectos Allee, por los que un número mínimo de invasores y hospedantes deben de coincidir espacialmente para poder iniciar un nuevo foco de PWD (YOSHIMURA et al. 1999). Como consecuencia, en este caso, la colección de datos puntuales de vuelos de largo recorrido es importante en sí misma ya que define las distancias dentro de las cuales puede darse una dispersión natural de la enfermedad. AKBULUT y STAMPS (2012) consideran que la capacidad de vuelo de las especies del género Monochamus en condiciones naturales es pobre y que sus individuos no son buenos voladores. Fundamentan su afirmación en los resultados de estudios japoneses (KOBAYASHI et al. 1973, IDO et al. 1975, KAWABATA et al. 1975, KAWABATA 1979, OGAWA Y HAGIWARA 1980), que indican que la dispersión de la mayoría de la población se reduce a unos pocos centenares de metros, con máximos puntuales constatados entre los 2400m y los 3300 m. Nuestros resultados, al contrario, muestran que la capacidad de dispersión natural a larga distancia de M. galloprovincialis es muy superior. La recaptura de 31 individuos (18.5% de los 168 recapturados) a más de 4000 m del punto de liberación o el dato de un ejemplar a 13600 m supera ampliamente los límites constatados en los experimentos de KAWABATA (1979) y de OGAWA y HAGIWARA (1980) para M. alternatus .(KOBAYASHI et al. 1984). Estos resultados están en mayor consonancia con las recientes publicaciones sobre la capacidad de vuelo o dispersiva de esta especie, que muestran promedios de vuelos potenciales de 16 km en bioensayos con molinos de vuelo (DAVID et al. 2014) o modelos de dispersión en los que el 99% de los M. galloprovincialis abastan los 3500 m, ajustando la posibilidad a que se produzcan vuelos mayores en una pequeña parte de la población (ETXEBESTE et al. 2015).
Los primeros estudios, además, fundamentaron la determinación de la anchura de los cinturones de aislamiento planteados en los programas de erradicación de B. xylophilus en España como respuesta a la normativa europea (JUNTA DE EXTREMADURA, 2008; XUNTA DE GALICIA, 2010) y en China (ZHAO et al. 2008). En estos programas se estimó que una buena profilaxis contra el PWD sería la corta y destrucción de todas las coníferas en cinturones de anchura determinada (3 km alrededor de la zona de detección de la enfermedad para el caso de Europa y 4 km en China). En este sentido, nuestros resultados contradicen la utilidad de tales medidas. Los experimentos desarrollados en la última fase del estudio (año 2011) fueron diseñados con la experiencia previa de que M. galloprovincialis era capaz de superar los 3000 m de recorrido, observado en 2010. Los experimentos 5, 7 y 8, que contaron con una presión de muestreo mayor a partir de dicha distancia, arrojan resultados importantes que muestran porcentajes de recapturas a más de 3000 m del 2.3%, 2.6% y 1.7% respectivamente. Asimismo, además de la ya mencionada recaptura puntual a 13600m, el dato más sorprendente e inesperado se produce con la recaptura de un ejemplar a 22100 m del punto de liberación. No tenemos posibilidad de afirmar si se ha producido una dispersión natural o si se ha producido un transporte como insecto-polizón. No obstante, el hecho de que haya alcanzado tal distancia es un dato relevante desde el punto de vista de los objetivos de este estudio. Nuestros resultados

184
apoyan la tesis de que las medidas profilácticas establecidas en China y España para evitar la dispersión natural de Monochamus spp y B. xylophilus consistentes en la eliminación del material hospedante a lo largo bandas de varios kilómetros de anchura resultan ineficaces en el control de la dispersión de los vectores y, por tanto, de la enfermedad; aunque conviene tener en cuenta que todos nuestros estudios han sido realizados con insectos no infectados por nematodos y que desconocemos cómo podría afectar la infección a su capacidad de vuelo.
Generalmente, en los estudios de dispersión son habituales los desajustes y la baja coincidencia de resultados entre experimentos realizados con métodos diferentes (marcado-liberación-recaptura, molinos de vuelo, telemetría o estudios genéticos) aun tratándose de una misma especie (DRAG et al. 2011, DAVID et al. 2014). En este contexto, los experimentos de marcado-liberación-recaptura pueden adolecer de subestimar la proporción de dispersiones naturales a larga distancia en la medida en que estos individuos pueden abandonar completamente el área de muestreo (TURCHIN Y THOENY 1993, KOENIG et al. 1996, TURCHIN 1998, JONSELL et al. 2003, FRANZEN Y NILSSON 2007) y, de esta forma, ofrecer valores de distancias de dispersión erróneamente bajos con respecto a otras metodologías. Nuestros experimentos, enfocados primordialmente a la obtención de datos de vuelos de largo alcance, y con superficies de muestreo más grandes de lo habitual en este tipo de ensayos (SMITH et al. 2001, SMITH et al. 2004, BANCROFT Y SMITH 2005, DRAG et al. 2011, ETXEBESTE et al. 2015, SANCHEZ-HUSILLOS et al. 2015a, TORRES-VILA et al. 2015) asumen la baja probabilidad de recapturas de M. galloprovincialis a tan larga distancia, debido a la baja presión de muestreo para áreas tan amplias, a cambio de la fuerte potencia de los pocos datos que puedan obtenerse. Ninguna de las distribuciones de las recapturas de los experimentos realizados en 2011 (Figuras 5.29-a, 5.29-b, 5.29-c) se ajusta bien a una función de dispersión típica, exponencial negativa (KAREIVA 1983, TURCHIN Y THOENY 1993), lo que puede indicar que se trata de eventos de dispersión a larga distancia (ETXEBESTE et al. 2015). De esta forma, nuestros resultados pueden compararse con los resultados obtenidos por (AKBULUT Y LINIT 1999), en experimentos desarrollados con molinos de vuelo (que obtuvieron distancias de vuelo de 10 km para M. carolinensis) o por (DAVID et al. 2014) ,que obtuvieron promedios de 16 km para M. galloprovincialis, más que con otros experimentos de marcado-liberación-recaptura realizados en Japón (KOBAYASHI et al. 1984). Numerosos autores han criticado el empleo de los molinos de vuelo, ya que las condiciones experimentales, sobre todo la manipulación de los individuos estudiados y el ambiente de laboratorio, suponen un gran sesgo en las lecturas (TAYLOR et al. 2010 y las citas en este trabajo). Sin embargo otra posible interpretación de las lecturas registradas en los molinos de vuelo se produce al interpretar estos registros como las distancias acumuladas resultantes de la suma de cada uno de los segmentos volados por el insecto bajo un entorno de caminos aleatorios o teorías de difusión (TURCHIN 1998, OKUBO Y LEVIN 2001, TAYLOR et al. 2010). De hecho, una parte de los resultados del experimento Exp2011/a, no mostrados en este trabajo, fue incluida en el trabajo de ETXEBESTE et al. (2015) del que el autor de esta memoria es coautor. En él, los resultados de este experimento fueron utilizados para comprobar la predicción de las capturas a larga distancia utilizando los diferentes modelos de dispersión a corta distancia obtenidos por sus autores en experimentos similares, todos ellos realizados en años anteriores (2009 y 2010) sobre en áreas de estudio de menor superficie. En las conclusiones de dicho trabajo se indica que los modelos ajustados por los autores predicen los eventos de dispersión a larga distancia registrados en 2011 en la Comunitat Valenciana (dos recapturas a 5000 m), aunque sugieren que son necesarios más estudios que determinen el grado en el que las características del paisaje y de la masa pueden influir en la dispersión de M. galloprovincialis.

185
Por último, nuestros resultados destacan también la importancia de profundizar en el estudio de la dispersión de esta especie centrándose en dos aspectos diferentes: analizar la dispersión del porcentaje de la población que permanece en áreas próximas a su lugar de emergencia (SMITH et al. 2001, SMITH et al. 2004, BANCROFT Y SMITH 2005, DRAG et al. 2011), y analizar la proporción de la población que se dispersa a largas distancias, acometiendo, de esta forma, una de las tareas pendientes en los estudios de marcado-liberación-recaptura indicada por TURCHIN y ODENDAAL (1996) y KOENIG et al. (1996): la estimación de las colas de las funciones de dispersión habituales en este tipo de experimentos. Nuestros estudios permiten pensar que la colocación de trampas a larga distancia, aún en densidad más baja que la utilizada en los experimentos habituales, puede ofrecer resultados reveladores en este sentido. El último ensayo, realizado en 2012, merece discusión aparte por su especial diseño experimental. Una de las grandes preocupaciones en torno a la gestión de M. galloprovincialis como dispersor del PWD, radica en el conocimiento de su comportamiento dispersivo durante el periodo de tiempo en el que no es susceptible de ser capturado por las trampas utilizadas (Capítulo 5.1), presumiblemente, periodo en el que aún no ha alcanzado la madurez sexual (HELLRIGL 1971, FRANCARDI Y PENNACCHIO 1996, NAVES et al. 2008, DAVID et al. 2014, SANCHEZ-HUSILLOS et al. 2015b). Asimismo, la dispersión a larga distancia de la enfermedad del marchitamiento del pino está más correlacionada con la dispersión artificial provocada por el movimiento de materiales y personas propio de la economía globalizada y el comercio internacional que con la dispersión natural del insecto (ROBINET et al. 2009, ROBINET et al. 2011). El estudio del comportamiento y capacidades de los imagos recién emergidos en zonas alejadas de sus propias áreas de alimentación y reproducción (zonas alejadas de masas forestales) es de gran ayuda para la comprensión de la posible aparición de nuevos focos en zonas alejadas de las áreas infectadas. En este sentido, pese a la baja tasa recaptura obtenida en dicho ensayo (lo cual impide un adecuado tratamiento estadístico y el ajuste a un modelo de dispersión razonable), de nuevo un dato resulta revelador por la gran distancia registrada, 3 km. Si bien es cierto que M. galloprovincialis no parece ser una especie que, a lo largo de su historia evolutiva, haya podido adaptarse a situaciones de emergencia de los imagos en punto situados a largas distancias del material hospedante (antes al contrario, la situación habitual en condiciones naturales respondería a emergencias dentro de la propia masa forestal), los 3 km de distancia de vuelo registrada en una zona sin posibilidad de alimentarse parecen indicar que M. galloprovincialis tiene la capacidad de alcanzar zonas alejadas sin necesidad de alimentarse previamente. Estos resultados coinciden con los mostrados por SANCHEZ-HUSILLOS et al. (2015b), que indican que fisiológicamente estos eventos son posibles con base en las reservas de lípidos en el momento de la emergencia, aunque también indican que la alimentación inmediatamente posterior a la emergencia debería mejorar la capacidad fisiológica de vuelo, también de acuerdo con los resultados de DAVID et al. (2014) para bioensayos con molinos de vuelo. Y en cualquier caso, esta distancia de 3 km es coherente también con los resultados de dispersión de insectos inmaduros mostrados por SANCHEZ-HUSILLOS et al. (2015b) en ensayos similares al presentado aquí. Monochamus galloprovincialis posee suficiente capacidad de dispersión como para recorrer distancias de varios kilómetros en pocos días. Se ha registrado una velocidad de dispersión de al menos, (ya que el insecto es muy difícil que llegue al lugar de captura en línea recta), 1500 m/día y longevidades de individuos cercanas a los 3 meses. La máxima distancia de dispersión observada ha sido de 13600 m. También ha sido constatada una recaptura a 22100 m del punto de liberación, distancia de la que no puede asegurarse que se haya producido de forma natural o con el concurso humano (insecto-polizón). El hecho de que algunos escarabajos puedan dispersarse a estas distancias tan grandes plantea un problema muy importante en la erradicación de la enfermedad y en la contención de su expansión. Estos datos, junto a los altos

186
porcentajes de recapturas a más de 3000 m (cercanas al 2% de la población liberada en todas las regiones estudiadas) hacen patente que las medidas profilácticas establecidas en España y China para evitar la dispersión natural de Monochamus spp. y B. xylophilus, consistentes en la eliminación de las especies hospedantes en bandas de varios kilómetros de anchura, podrían resultar ineficaces en el control de la dispersión de los vectores y, por tanto, de la enfermedad.

187
Bibliografía
ABELLEIRA, A., A. PICOAGA, J. P. MANSILLA y O. AGUIN (2011). Detection of Bursaphelenchus xylophilus, Causal Agent of Pine Wilt Disease on Pinus pinaster in Northwestern Spain. Plant Disease 95(6): 776-776.
AGOSTINELLI, C. y U. LUND (2013). R package 'circular': Circular Statistics (version 0.4-7). URL https://r-forge.r-project.org/projects/circular/.
AKBULUT, S. y M. J. LINIT (1999). Flight Performance of Monochamus carolinensis (Coleoptera: Cerambycidae) with Respect to Nematode Phoresis and Beetle Characteristics. Environmental Entomology 28(6): 1014-1020.
AKBULUT, S. y W. T. STAMPS (2012). Insect vectors of the pinewood nematode: a review of the biology and ecology of Monochamus species. Forest Pathology 42(2): 89-99.
ÁLVAREZ, G., I. ETXEBESTE, D. GALLEGO, G. DAVID, L. BONIFACIO, H. JACTEL, E. SOUSA y J. A. PAJARES (2015). Optimization of traps for live trapping of Pine Wood Nematode vector Monochamus galloprovincialis. Journal of Applied Entomology 139(8): 618-626.
ALYA, A. y F. HAIN (1985). Life histories of Monochamus carolinensis and M. titillator (Coleoptera: Cerambycidae) in the Piedmont of North Carolina. Journal of Entomological Science 20(4): 390-397.
BANCROFT, J. y M. SMITH (2005). Dispersal and influences on movement for Anoplophora glabripennis calculated from individual mark‐recapture. Entomologia experimentalis et applicata 116(2): 83-92.
BAOJUN, Y. y W. QOULI (1989). Distribution of the pinewood nematode in China and susceptibility of some Chinese and exotic pines to the nematode. Canadian Journal of Forest Research 19(12): 1527-1530.
BARATA, E. N. y J. ARAÚJO (2001). Olfactory orientation responses of the eucalyptus woodborer, Phoracantha semipunctata, to host plant in a wind tunnel. Physiological Entomology 26(1): 26-37.
BILLINGS, R., S. CLARKE, V. ESPINO-MENDOZA, P. CORDÓN CABRERA, B. MELÉNDEZ FIGUEROA, J. RAMÓN CAMPOS y G. BAEZA (2004). Bark beetle outbreaks and fire: a devastating combination for Central America’s pine forests. Unasylva 55(217): 7.
BONIFACIO, L. (2009). Impacte e Evolução da Doença da Muchidão dos Pinheiros (Pine Wilt Disease) na Zona Afectada a Sul do Rio Tejo. Doutoramento em Biologia PhD, Universidade de Lisboa, Lisboa, 222
BONIFÁCIO, L. F. P. (2009). Impacte e evolução da doença da murchidão dos pinheiros (pine wilt disease) na zona afectada a sul do Rio Tejo.
CARDÉ, R. T. (1996). Odour plumes and odour-mediated flight in insects. Olfaction in mosquito-host interactions 200: 54-70.
CLOBERT, J., M. BAGUETTE, T. G. BENTON, J. M. BULLOCK y S. DUCATEZ (2012). Dispersal ecology and evolution, Oxford University Press.
COMPTON, S. G. (2002). Sailing with the wind: dispersal by small flying insects. Dispersal ecology: 113-133.
DAVID, G., B. GIFFARD, D. PIOU y H. JACTEL (2014). Dispersal capacity of Monochamus galloprovincialis, the European vector of the pine wood nematode, on flight mills. Journal of Applied Entomology 138(8): 566-576.
DEROSE, R. J. y J. N. LONG (2009). Wildfire and spruce beetle outbreak: Simulation of interacting disturbances in the central Rocky Mountains. Ecoscience 16(1): 28-38.
DRAG, L., D. HAUCK, P. POKLUDA, K. ZIMMERMANN y L. CIZEK (2011). Demography and dispersal ability of a threatened saproxylic beetle: a mark-recapture study of the Rosalia Longicorn (Rosalia alpina). PLoS ONE 6(6): e21345.
ETXEBESTE, I., E. SANCHEZ-HUSILLOS, G. ÁLVAREZ, H. MAS I GISBERT y J. PAJARES (2015). Dispersal of Monochamus galloprovincialis (Col.: Cerambycidae) as

188
recorded by mark-release recapture using pheromone traps. Journal of Applied Entomology: In press.
FRANCARDI, V. y F. PENNACCHIO (1996). Note sulla bioecologia di Monochamus galloprovincialis Olivier) in Toscana e in Liguria (Coleoptera Cerambycidae). Redia 79: 153-169.
FRANKLIN, A. J., C. DEBRUYNE y J. C. GRÉGOIRE (2000). Recapture of Ips typographus L.(Col., Scolytidae) with attractants of low release rates: localized dispersion and environmental influences. Agricultural and Forest Entomology 2(4): 259-270.
FRANZEN, M. y S. G. NILSSON (2007). What is the required minimum landscape size for dispersal studies? Journal of Animal Ecology 76(6): 1224-1230.
GALLEGO, D., F. SANCHEZ-GARCÍA, H. MAS, M. CAMPO y J. L. L. GUTIÉRREZ (2012). Estudio de la capacidad de vuelo a larga distancia de" Monochamus galloprovincialis"(Oliver 1795).(Coleoptera: Cerambycidae) en un mosaico agro-forestal. Boletín de sanidad vegetal. Plagas 38(1): 109-124.
GONZÁLEZ, C., E. VIVES y A. ZUZARTE (2007). Nuevo catálogo de los Cerambycidae (Coleoptera) de la Península Ibérica, islas Baleares e islas atlánticas: Canarias, Açores y Madeira. Monografías SEA 12: 1-211.
GRAHAM, E. E., R. F. MITCHELL, P. F. REAGEL, J. D. BARBOUR, J. G. MILLAR y L. M. HANKS (2010). Treating panel traps with a fluoropolymer enhances their efficiency in capturing cerambycid beetles. Journal of economic entomology 103(3): 641-647.
GROVE, S. J. (2002). Saproxylic insect ecology and the sustainable management of forests. Annual Review of Ecology and Systematics 33: 1-23.
HANKS, L. (1999). Influence of the larval host plant on reproductive strategies of cerambycid beetles. Annual Review of Entomology 44(1): 483-505.
HANULA, J. L., J. R. MEEKER, D. R. MILLER y E. L. BARNARD (2002). Association of wildfire with tree health and numbers of pine bark beetles, reproduction weevils and their associates in Florida. Forest Ecology and Management 170(1): 233-247.
HARDIE, J. y A. K. MINKS (1999). Pheromones of non-lepidopteran insects associated with agricultural plants, CABI publishing.
HELLRIGL, K. G. (1971). Die Bionomie der europaischen Monochamus-Arten (Coleopt., Cerambycid.) und ihre Bedeutung fur die Forstund Holzwirtschaft. Redia 52: 367-509.
HERNÁNDEZ ALONSO, R., V. PÉREZ FORTEA, G. SÁNCHEZ PEÑA, J. CASTELLÁ SOLÁ, J. PALENCIA ADRUBAU, J. M. GIL BONO y A. ORTIZ SÁNCHEZ (2007). Ensayos de trampeo de escolítidos perforadores subcorticales en pinares mediante el uso de feromonas 2002-2005. Ecología(21): 43-55.
HERNÁNDEZ, R., A. ORTIZ, V. PÉREZ, J. GIL y G. SÁNCHEZ (2011). " Monochamus galloprovinciales"(Olivier, 1795)(Coleoptera: Cerambycidae), comportamiento y distancias de vuelo. Boletín de sanidad vegetal. Plagas 37(1): 79-96.
IBEAS, F., J. J. DÍEZ y J. A. PAJARES (2008). Olfactory sex attraction and mating behaviour in the pine sawyer Monochamus galloprovincialis (Coleoptera: Cerambycidae). Journal of Insect Behavior 21(3): 101-110.
IBEAS, F., D. GALLEGO, J. DIEZ y J. PAJARES (2007). An operative kairomonal lure for managing pine sawyer beetle Monochamus galloprovincialis (Coleoptera: Cerymbycidae). Journal of Applied Entomology 131(1): 13-20.
IDO, N., J. TAKEDA, K. KOBAYASHI, A. TAKETANI, M. OKUDA y R. HOSODA (1975). Investigation o f the dispersal of pine sawyer adults. Trans. 86th Ann. Meet. Jpn. For. Soc.: 335-336.
IVAN, J. (2008). Density estimation. In: Program MARK: a gentle introduction. Ed. by Cooch E, White G, 20–21, 20–19. http://www.phidot.org/software/mark/docs/ book/ [accessed on 15 October 2014].
JONSELL, M., M. SCHROEDER y T. LARSSON (2003). The saproxylic beetle Bolitophagus reticulatus: its frequency in managed forests, attraction to volatiles and flight period. Ecography 26(4): 421-428.

189
KAREIVA, P. (1983). Local movement in herbivorous insects: applying a passive diffusion model to mark-recapture field experiments. Oecologia 57(3): 322-327.
KAWABATA, K. (1979). Migration of the pine sawyer among small islands. Trans. 32nd Ann. Meet. Kyushu Branch Jpn. For. Soc.: 281-282.
KAWABATA, K., M. KOJO y Y. KATSU (1975). Examination of attractants. Trans. 28th Ann. Meet. Kyushu Branch Jpn. For. Soc.: 161-162.
KOBAYASHI, F., M. OKUDA, R. HOSODA, A. TAKETANI y N. IDO (1973). Mark recapture tests on the behaviors of pine sawyer adults. Trans . . 84th Ann. Meet. Jpn. For. Soc.: 327-328.
KOBAYASHI, F., A. YAMANE y T. IKEDA (1984). The Japanese Pine Sawyer Beetle As The Vector Of Pine Wilt Disease. Annual Review of Entomology 29: 115-135.
KOENIG, W. D., D. VAN VUREN y P. N. HOOGE (1996). Detectability, philopatry, and the distribution of dispersal distances in vertebrates. Trends in ecology & evolution 11(12): 514-517.
LA, Y., Y. MOON, W. YEO, S. SHIN y W. BAK (1999). Recent status of pine wilt disease in Korea. Sustainability of pine forests in relation to pine wilt and decline. Proceedings of International Symposium, Tokyo, Japan, 27-28 October, 1998., Shokado Shoten. 239-241.
LEVINE, J. M. y C. M. D'ANTONIO (2003). Forecasting biological invasions with increasing international trade. Conservation Biology 17(1): 322-326.
MAGRAMA. (2015a). "Nematodo de la madera del pino." 2015, de http://www.magrama.gob.es/es/agricultura/temas/sanidad-vegetal/nematodo-de-la-madera-del-pino/#.
MAGRAMA (2015b). Trabajos de seguimiento y control del vector del nematodo de la madera del pino. Ministerio de Agricultura, Pesca y Alimentación. Internal report: 32 pages.
MAMIYA, Y. (1988). History of Pine Wilt Disease in Japan. Journal of Nematology 20(2): 219-226.
MARINI, L., A. LINDELOW, A. M. JONSSON, S. WULFF y L. M. SCHROEDER (2013). Population dynamics of the spruce bark beetle: a long-term study. Oikos 122(12): 1768-1776.
MAS, H., HERNANDEZ, R., VILLAROYA, G., SANCHEZ, G., PÉREZ-LAORGA, E., GONZALEZ, E., et al. (2013). Dispersal behavior and long distance flight capacity of Monochamus galloprovincialis (Olivier 1795). In: Schröder T (ed.), Pine Wilt Disease Conference. pp. 22, Braunschweig,ISSN: 1866-590X. 2013.
MCHUGH, C. W., T. E. KOLB y J. L. WILSON (2003). Bark beetle attacks on ponderosa pine following fire in northern Arizona. Environmental Entomology 32(3): 510-522.
MCMAHON, M. D., K. F. RAFFA, E. V. NORDHEIM y B. H. AUKEMA (2010). Too close for comfort: effect of trap spacing distance and pattern on statistical inference of behavioral choice tests in the field. Entomologia experimentalis et applicata 136(1): 66-71.
MILLER, D. R. (2006). Ethanol and (−)-α-pinene: attractant kairomones for some large wood-boring beetles in southeastern USA. Journal of Chemical Ecology 32(4): 779-794.
MILLER, D. R., C. ASARO, C. M. CROWE y D. A. DUERR (2011). Bark beetle pheromones and pine volatiles: attractant kairomone lure blend for longhorn beetles (Cerambycidae) in pine stands of the southeastern United States. Journal of economic entomology 104(4): 1245-1257.
MORBEY, Y. E. y R. C. YDENBERG (2001). Protandrous arrival timing to breeding areas: a review. Ecology Letters 4(6): 663-673.
MOTA, M. M., H. BRAASCH, M. A. BRAVO, A. C. PENAS, W. BURGERMEISTER, K. METGE y E. SOUSA (1999). First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1: 727-734.
MOTA, M. M. y P. VIEIRA (2008). Pine wilt disease: a worldwide threat to forest ecosystems, Springer.

190
MURLIS, J., J. S. ELKINTON y R. T. CARDE (1992). Odor plumes and how insects use them. Annual Review of Entomology 37(1): 505-532.
NATHAN, R., E. KLEIN, J. J. ROBLEDO-ARNUNCIO y E. REVILLA (2012). Dispersal kernels: review. Dispersal ecology and evolution. J. Clobert, M. Baguette, T. G. Benton y J. M. Bullock. Oxford, Oxford University Press: 187-210.
NATHAN, R., G. PERRY, J. T. CRONIN, A. E. STRAND y M. L. CAIN (2003a). Methods for estimating long-distance dispersal. Oikos 103(2): 261-273.
NATHAN, R., G. PERRY, J. T. CRONIN, A. E. STRAND y M. L. CAIN (2003b). Methods for estimating long‐distance dispersal. Oikos 103(2): 261-273.
NAVES, P., S. CAMACHO, E. DE SOUSA y J. QUARTAU (2007). Transmission of the pine wood nematode Bursaphelenchus xylophilus through feeding activity of Monochamus galloprovincialis (Col., Cerambycidae). Journal of Applied Entomology 131(1): 21-25.
NAVES, P., E. DE SOUSA y J. A. QUARTAU (2006a). Reproductive traits of Monochamus galloprovincialis (Coleoptera : Cerambycidae) under laboratory conditions. Bulletin of Entomological Research 96(3): 289-294.
NAVES, P. M., S. CAMACHO, E. M. DE SOUSA y J. A. QUARTAU (2006b). Entrance and distribution of the pinewood nematode Bursaphelenchus xylophilus on the body of its vector Monochamus galloprovincialis (Coleoptera: Cerambycidae). Entomologia generalis 29(1): 71-80.
NAVES, P. M., E. M. DE SOUSA y J. A. QUARTAU (2006c). Feeding and oviposition preferences of Monochamus galloprovincialis for certain conifers under laboratory conditions. Entomologia Experimentalis Et Applicata 120(2): 99-104.
NAVES, P. M., E. SOUSA y J. M. RODRIGUES (2008). Biology of Monochamus galloprovincialis (Coleoptera: Cerambycidae) in the pine wilt disease affected zone, Southern Portugal. Silva Lusitana 16(2): 133-148.
OGAWA, S. y Y. HAGIWARA (1980). Expansion of the pine infestation caused by pine wood nematodes. Shinrin Boeki (Forest Pests) 29: 115-117.
OKUBO, A. y S. A. LEVIN (2001). Diffusion and ecological problems: modern perspectives, Springer Science & Business Media.
PAJARES, J. A., G. ALVAREZ, F. IBEAS, D. GALLEGO, D. R. HALL y D. I. FARMAN (2010). Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. Journal of Chemical Ecology 36(6): 570-583.
PAJARES, J. A., F. IBEAS, J. J. DIEZ y D. GALLEGO (2004). Attractive responses by Monochamus galloprovincialis (Col., Cerambycidae) to host and bark beetle semiochemicals. Journal of Applied Entomology 128(9-10): 633-638.
PÉREZ, V., R. HERNÁNDEZ, J. CASTELLÁ, G. SÁNCHEZ y J. PALENCIA (2004). Ensayos de atracción y captura de Ips acuminatus (Coleoptera: Scolytidae). Ecología(18): 35-46.
PUKKALA, T., T. MOYKKYNEN y C. ROBINET (2014). Comparison of the potential spread of pinewood nematode (Bursaphelenchus xylophilus) in Finland and Iberia simulated with a cellular automaton model. Forest Pathology 44(5): 341-352.
R CORE TEAM (2015). R: A language and environment for statistical computing. Vienna, Austria, R Foundation for Statistical Computing.
RANIUS, T. (2006). Measuring the dispersal of saproxylic insects: a key characteristic for their conservation. Population Ecology 48(3): 177-188.
RASSATI, D., E. P. TOFFOLO, A. BATTISTI y M. FACCOLI (2012). Monitoring of the pine sawyer beetle Monochamus galloprovincialis by pheromone traps in Italy. Phytoparasitica 40(4): 329-336.
ROBINET, C., A. ROQUES, H. PAN, G. FANG, J. YE, Y. ZHANG y J. SUN (2009). Role of Human-Mediated Dispersal in the Spread of the Pinewood Nematode in China. PLoS ONE 4(2): e4646.

191
ROBINET, C., N. VAN OPSTAL, R. BAKER y A. ROQUES (2011). Applying a spread model to identify the entry points from which the pine wood nematode, the vector of pine wilt disease, would spread most rapidly across Europe. Biological Invasions 13(12): 2981-2995.
RUAN, X. S. (1996). The occurrence and control of Bursaphelenchus xylophilus in Taiwan. Plant Quarantine (Shanghai) 10(2): 86-87.
RUIZ-PEINADO, R., G. M. GONZÁLEZ y I. C. R. DE VIÑAS (2001). Growth and yield models for Pinus halepensis Mill. Investigación agraria. Sistemas y recursos forestales 10(1): 179-201.
SAINT-GERMAIN, M., P. DRAPEAU y C. HÉBERT (2004a). Landscape-scale habitat selection patterns of Monochamus scutellatus (Coleoptera: Cerambycidae) in a recently burned black spruce forest. Environmental Entomology 33(6): 1703-1710.
SAINT-GERMAIN, M., P. DRAPEAU y C. HÉBERT (2004b). Xylophagous insect species composition and patterns of substratum use on fire-killed black spruce in central Quebec. Canadian Journal of Forest Research 34(3): 677-685.
SAINT‐GERMAIN, M., C. M. BUDDLE y P. DRAPEAU (2007). Primary attraction and random landing in host‐selection by wood‐feeding insects: a matter of scale? Agricultural and Forest Entomology 9(3): 227-235.
SANCHEZ-HUSILLOS, E., I. ETXEBESTE, G. ÁLVAREZ-BAZ y J. PAJARES (2013). Physiological development of Monochamus galloprovincialis immature adults through shoot feeding. Pine Wilt Disease Conference, Braunschweig, Berichte aus dem Julius Kühn-Institut. 26-27.
SANCHEZ-HUSILLOS, E., I. ETXEBESTE y J. PAJARES (2015a). Effectiveness of mass trapping in the reduction of Monochamus galloprovincialis Olivier (Col.: Cerambycidae) populations. Journal of Applied Entomology.
SANCHEZ-HUSILLOS, E., I. ETXEBESTE y J. PAJARES (2015b). Physiological development and dispersal ability of newly emerged Monochamus galloprovincialis (Coleoptera: Cerambycidae). Entomologia Experimentalis Et Applicata: Enviado.
SCHROEDER, L. M. (2012). Monitoring of Ips typographus and Pityogenes chalcographus: influence of trapping site and surrounding landscape on catches. Agricultural and Forest Entomology: no-no.
SMITH, M. T., J. BANCROFT, G. LI, R. GAO y S. TEALE (2001). Dispersal of Anoplophora glabripennis (Cerambycidae). Environmental Entomology 30(6): 1036-1040.
SMITH, M. T., P. C. TOBIN, J. BANCROFT, G. LI y R. GAO (2004). Dispersal and spatiotemporal dynamics of Asian longhorned beetle (Coleoptera: Cerambycidae) in China. Environmental Entomology 33(2): 435-442.
TAKASU, F., N. YAMAMOTO, K. KAWASAKI, K. TOGASHI, Y. KISHI y N. SHIGESADA (2000). Modeling the expansion of an introduced tree disease. Biological Invasions 2(2): 141-150.
TAYLOR, R., L. S. BAUER, T. M. POLAND y K. N. WINDELL (2010). Flight performance of Agrilus planipennis (Coleoptera: Buprestidae) on a flight mill and in free flight. Journal of Insect Behavior 23(2): 128-148.
TIKKAMÄKI, T. y A. KOMONEN (2011). Estimating population characteristics of two saproxylic beetles: a mark-recapture approach. Journal of Insect Conservation 15(3): 401-408.
TOGASHI, K. y N. SHIGESADA (2006). Spread of the pinewood nematode vectored by the Japanese pine sawyer: modeling and analytical approaches. Population Ecology 48(4): 271-283.
TORRES-VILA, L. M., Á. SANCHEZ-GONZÁLEZ, F. PONCE-ESCUDERO, D. MARTÍN-VERTEDOR y J. J. FERRERO-GARCÍA (2012). Assessing mass trapping efficiency and population density of Cerambyx welensii Küster by mark-recapture in dehesa open woodlands. European Journal of Forest Research 131(4): 1103-1116.

192
TORRES-VILA, L. M., C. ZUGASTI, J. M. DE-JUAN, M. J. OLIVA, C. MONTERO, F. J. MENDIOLA, Y. CONEJO, Á. SÁNCHEZ, F. FERNÁNDEZ, F. PONCE y G. ESPÁRRAGO (2015). Mark-recapture of Monochamus galloprovincialis with semiochemical-baited traps: population density, attraction distance, flight behaviour and mass trapping efficiency. Forestry 88(2): 224-236.
TORRES‐VILA, L. M., Á. SÁNCHEZ‐GONZÁLEZ, J. MERINO‐MARTÍNEZ, F. PONCE‐ESCUDERO, Y. CONEJO‐RODRÍGUEZ, D. MARTÍN‐VERTEDOR y J. J. FERRERO‐GARCÍA (2013). Mark–recapture of Cerambyx welensii in dehesa woodlands: dispersal behaviour, population density, and mass trapping efficiency with low trap densities. Entomologia experimentalis et applicata 149(3): 273-281.
TRÄGÅRDH, I. (1940). Neue Beobachtungen über den Schusterbock (Monochamus sutor L.) in Schweden. Zeitschrift für Angewandte Entomologie 27(1): 142-149.
TURCHIN, P. (1998). Quantitative analysis of movement: measuring and modeling population redistribution in animals and plants. Sunderland, Mass., Sinauer.
TURCHIN, P. y F. J. ODENDAAL (1996). Measuring the effective sampling area of a pheromone trap for monitoring population density of southern pine beetle (Coleoptera: Scolytidae). Environmental Entomology 25(3): 582-588.
TURCHIN, P. y W. T. THOENY (1993). Quantifying dispersal of southern pine beetles with mark recapture experiments and a diffusion-model. Ecological Applications 3(1): 187-198.
VICENTE, C., M. ESPADA, P. VIEIRA y M. MOTA (2012). Pine Wilt Disease: a threat to European forestry. European Journal of Plant Pathology 133(1): 89-99.
VIVES (2000). Insecta, Coleoptera, Cerambycidae. Serie Fauna Ibérica. Museo Nacional de Ciencias Naturales, CSIC, Madrid. 12.
YAMAMURA, K., M. KISHITA, N. ARAKAKI, F. KAWAMURA y Y. SADOYAMA (2003). Estimation of dispersal distance by mark-recapture experiments using traps: correction of bias caused by the artificial removal by traps. Population Ecology 45(2): 149-155.
YOSHIMURA, A., K. KAWASAKI, F. TAKASU, K. TOGASHI, K. FUTAI y N. SHIGESADA (1999). Modeling the spread of pine wilt disease caused by nematodes with pine sawyers as vector. Ecology 80(5): 1691-1702.
ZAMORA, P., V. RODRÍGUEZ, F. RENEDO, A. V. SANZ, J. C. DOMÍNGUEZ, G. PÉREZ-ESCOLAR, J. MIRANDA, B. ÁLVAREZ, A. GONZÁLEZ-CASAS, E. MAYOR, M. DUEÑAS, A. MIRAVALLES, A. NAVAS, L. ROBERTSON y A. B. MARTÍN (2015). First Report of Bursaphelenchus xylophilus Causing Pine Wilt Disease on Pinus radiata in Spain. Plant Disease: PDIS-03-15-0252-PDN.
ZHANG, Q. H. y F. SCHLYTER (2004). Olfactory recognition and behavioural avoidance of angiosperm nonhost volatiles by conifer‐inhabiting bark beetles. Agricultural and Forest Entomology 6(1): 1-20.
ZHAO, B. G., K. FUTAI, J. R. SUTHERLAND y Y. TAKEUCHI (2008). Pine wilt disease, Springer.

193
Capítulo 6 Síntesis
Sección 6.1 Fenología y dispersión de Monochamus galloprovincialis en
la península ibérica
En el ámbito forestal europeo, una de las especies invasoras con mayor impacto potencial negativo sobre las masas de pinares del continente es el nematodo de la madera del pino (NMP) B. xylophilus, detectado en Portugal en el año 1999 (MOTA et al. 1999) y cuyo vector en la región europea es un cerambícido muy común en las masas forestales de la orla mediterránea y de muy amplia distribución global, M. galloprovincialis (SOUSA et al. 2001). Esta situación ha generado una gran preocupación y la consecuente toma de drásticas medidas de erradicación y de contención de la enfermedad. Actualmente el NMP se ha expandido por todo Portugal continental y ha sido detectado en cuatro focos en la parte fronteriza de España desde 2010 (ABELLEIRA et al. 2011, ROBERTSON et al. 2011, INÁCIO et al. 2015, MAGRAMA 2015, ZAMORA et al. 2015). En la medida en que B. xylophilus necesita forzosamente del concurso de un vector para su dispersión (YOSHIMURA et al. 1999, TOGASHI Y SHIGESADA 2006, FUTAI 2013), tanto en el ámbito de la gestión, como en el de la investigación, han fijado su atención sobre M. galloprovincialis. Efectivamente, desde la primera detección de NMP en Europa, diferentes y abundantes estudios han sido llevados a cabo con el objetivo de ampliar el conocimiento sobre la biología y la ecología de este coleóptero vector. El actual control del NMP en Europa se basa fundamentalmente en la detección temprana y destrucción de los árboles infectados dentro de unos plazos de tiempo marcados por la emergencia de los vectores. No obstante, el reciente desarrollo de un dispositivo para la captura y de un atrayente adecuados para el seguimiento de M. galloprovincialis (PAJARES et al. 2004, PAJARES et al. 2010, ÁLVAREZ et al. 2015) han supuesto la aparición de herramientas alternativas de gestión, enfocadas a su control poblacional (SANCHEZ-HUSILLOS et al. 2015a, TORRES-VILA et al. 2015) y han permitido abrir líneas de investigación sobre su fenología de vuelo y dispersión antes inabordables. En este sentido, uno de los principales factores que afectan la erradicación y control de especies invasoras es la correcta sincronización entre las medidas adoptadas y la fenología de las especies a gestionar (BOGICH et al. 2008). Desde esta perspectiva, y siempre bajo la óptica del vínculo con la gestión, en esta memoria se ha pretendido discurrir con la mayor atención y examinar la biología y la ecología de este insecto, en especial, en los aspectos relacionados con su fenología y su dispersión en el territorio de la península ibérica. En el primer bloque de esta tesis, los estudios realizados sobre la fenología de la emergencia de los imagos de M. galloprovincialis (Capítulo 2) muestran que existe una gran variabilidad en los patrones observados en la península Ibérica. Hasta ahora, éstos habían sido analizados

194
localmente en Francia, Italia y en Portugal (FRANCARDI Y PENNACCHIO 1996, KOUTROUMPA et al. 2008, NAVES Y DE SOUSA 2009). Para poder registrar las emergencias de M. galloprovincialis se llevaron a cabo doce experimentos de cría durante cinco años, desde 2010 hasta 2014, en tres regiones diferentes de España: Valencia (Quart de Poblet), Palencia (Calabazanos) y Teruel (Mora de Rubielos). Todos ellos fueron realizados a partir de árboles colonizados en campo, que posteriormente fueron trasladados a laboratorio, donde se ubicaron en bidones o jaulas de cría. Según nuestros resultados, de forma general, los insectos desarrollados en zonas cálidas muestran curvas de emergencia adelantadas y acumulaciones de calor menores con respecto a los desarrollados en lugares más fríos (Capítulo 2). Estos patrones son también diferentes de los registrados y previstos en la península de Setúbal en Portugal (NAVES Y DE SOUSA 2009), por lo que el modelo predictivo planteado por estos autores no puede ser extrapolado al resto de la península. Esta propuesta para la predicción de la emergencia del 50% de la población se basa en la utilización de un umbral mínimo de desarrollo de 12,2 ± 0,8ºC y de la contabilización de grados-día desde el 1 de marzo, fecha posterior a la dormancia invernal mostrada por esta especie. No obstante, nuestros resultados concuerdan mejor con la hipótesis de PARK et al. (2014) que plantea que dos condiciones podrían modificar el patrón de la emergencia de otro vector del NMP: (i) la estructura de edad de las larvas entrantes en el invierno, que puede influir en el desplazamiento horizontal de la curva de emergencia, y (ii) la heterogeneidad medioambiental posterior al paso del invierno, que podría causar variación del desarrollo larvario. Por lo cual, las temperaturas del otoño e invierno anteriores al 1 de marzo podrían tener responsabilidad añadida en la variabilidad observada. En términos globales, las emergencias de M. galloprovincialis en la península Ibérica no se ajustan al modelo predictivo de NAVES y DE SOUSA (2009), sino que están relacionadas con una acumulación de calor menor en las zonas cálidas que en las zonas frías, lo cual está en consonancia con el adelanto temporal de las curvas en zonas cálidas y el retraso en zonas frías. Las altas temperaturas del otoño-invierno anterior a la dormancia podrían acelerar el desarrollo larvario y cambiar la composición de la estructura de edad de las larvas (mayor cantidad de larvas de 4º estadio) antes de la llegada del invierno provocando de esta forma curvas de emergencia más tempranas (PARK et al. 2014) y, como consecuencia, acumulaciones de calor menores (Capítulo 2). En cualquier caso, nuestros resultados ponen en evidencia la necesidad de un estudio detallado de los requerimientos térmicos de cada una de las fases larvarias de M. galloprovincialis. No obstante, aunque parece indiscutible el rol fundamental que juega la temperatura en el desarrollo larvario, también debe ser tenido en cuenta también que, como especies nativas en la península ibérica, es posible que algunas características del ciclo biológico de M. galloprovincialis hayan divergido entre las poblaciones locales en respuesta a diferencias en el clima local (SINCLAIR et al. 2012). Por lo cual, la variabilidad observada aparentemente podría explicarse por la existencia, dentro de la península Ibérica, de adaptaciones locales en las diferentes regiones de la misma, quizás consecuencia de la existencia de poblaciones genéticamente diferenciadas (HARAN et al. 2015). Nuestros resultados sugieren, en definitiva, la necesidad de diferenciar modelos de gestión locales del vector del NMP dependiendo de la ubicación geográfica y teniendo en cuenta las variables climáticas de cada región ya que, en resumen, el mecanismo que determina el patrón de emergencia de M. galloprovincialis en una zona concreta podría ser muy complejo, influido por diferentes variables como la temperatura, el fotoperiodo, la calidad del material hospedador, la distribución de la ovoposición (KOUTROUMPA et al. 2008), y la variabilidad genética o incluso la existencia de algún otro tipo de señales de inicio. Estudios futuros deberían centrarse en el efecto detallado de esas variables en el desarrollo larvario de M. galloprovincialis, de

195
manera que las medidas de control destinadas a la gestión sostenible del NMP puedan aplicarse en el momento adecuado. Por último, también han sido observados fenómenos de protandria en las emergencias de M. galloprovincialis (Capítulo 2), probablemente como estrategia reproductiva, según la cual la aparición de los machos anterior a la de las hembras maximiza el éxito reproductivo al aumentar el número de hembras fecundadas (BLANCKENHORN et al. 2007), es decir, se trataría de una estrategia que favorecería el apareamiento múltiple de los machos. Si bien es verdad que algunos estudios han analizado la influencia de la temperatura en el desarrollo larvario de M. galloprovincialis (KOUTROUMPA et al. 2008, NAVES Y DE SOUSA 2009), hasta la fecha muy pocos estudios han estudiado su influencia en el vuelo del insecto tras la emergencia (HARAN et al. 2015, HOCH et al. 2015). De hecho, sólo el reciente desarrollo de trampas y atrayentes eficaces para su captura (PAJARES et al. 2004, PAJARES et al. 2010, ÁLVAREZ et al. 2015) ha permitido la monitorización de su curva de vuelo y, por tanto, el estudio de las variables que pudieren influir en él. El estudio de la influencia de la temperatura en el vuelo de M. galloprovincialis se incluye en el Capítulo 3 de este trabajo. Para llevarlo a cabo, un total de 8 experimentos de captura de M. galloprovincialis mediante trampas cebadas con atrayentes cairomonales y feromonales fueron realizados en diferentes regiones de la península Ibérica (Castellón, Teruel, Segovia y Alicante) entre los años 2010 y 2015, de manera que se registró el vuelo del insecto mediante capturas en trampas durante todo su periodo de vuelo y con frecuencia periódica (Capítulo 3). Su análisis ha permitido observar algunos resultados interesantes con respecto a la influencia de la altitud en las capturas del insecto, a la influencia de las temperaturas en el inicio de su periodo de vuelo y en sus comportamientos circadianos, y la de las altas temperaturas en un descenso de capturas estivales observado con mucha frecuencia. En primer lugar, los resultados muestran que efectivamente existe influencia de una variable sintética como la altitud, correlacionada con otras con significación biológica, como la temperatura, en la captura del insecto en el Sistema Central. Y aunque es posible observar una disminución de las capturas a partir de los 1350 msnm, el modelo ajustado parece indicar que existe una alta probabilidad de encontrar M. galloprovincialis a más de 1900 msnm, y presumiblemente hasta el límite altitudinal donde es posible encontrar formaciones de P. sylvestris, si bien es cierto que esta extrapolación debe ser tomada con cautela ya que las altitudes superiores a los 1900 msnm quedan fuera del rango de la modelización. Esto, en cualquier caso, no parece ser totalmente coherente con los resultados aportados por HARAN et al. (2015) para Pirineos, que establece los 1590 mnsm y las temperaturas mínimas medias por debajo de -7ºC como valores ambientales límite de la distribución de la especie, por lo que nuevos estudios deberían ser orientados en este sentido (Capítulo 3). Por otro lado, respecto a la variación temporal de las capturas a lo largo del periodo de vuelo del insecto, sí que es posible detectar una influencia de la altitud en la duración de dichos periodos de vuelo, de forma que a mayores altitudes (a partir de los 1600 msnm) la duración del periodo de vuelo tiende a ser más corta, mostrando tanto un inicio de vuelo más tardío como una finalización del mismo más temprana. En el Capítulo 2 se muestra que, en términos generales, las curvas de emergencia de las zonas más frías de la península ibérica son más tardías que las de las zonas más cálidas, lo cual es completamente coherente con un inicio del periodo de vuelo también más tardío en las zonas más frías (y en consecuencia en las zonas a mayor altitud). En concreto, nuestros resultados muestran que el inicio del periodo de vuelo está correlacionado con las temperaturas medias diarias, de manera que se inicia cuando éstas superan los 14-15ºC en los meses de mayo o junio, siendo estos resultados consistentes con los mostrados por

196
(HOCH et al. 2015) para la región de Baden en Austria, con inicios del periodo de vuelo exactamente a las mismas temperaturas (Capítulo 3). Asimismo, tanto las curvas de vuelo austríacas (HOCH et al. 2015) como las mostradas para la Sierra de Guadarrama (Sistema Central), a altitudes superiores a los 1150 msnm, presentan perfiles unimodales, mientras que una clara bimodalidad ha sido observada en el resto de experimentos realizados en diferentes regiones de la península ibérica (Capítulo 3). En todos estos casos analizados es posible observar una desaceleración en la acumulación de las capturas que coincide con los valores máximos de las temperaturas medias estivales, las cuales podrían funcionar como límite superior de temperatura para la «apetencia» por el vuelo. De este mismo modo, la unimodalidad en las curvas observada en los trabajos centroeuropeos (HOCH et al. 2015) y en las cotas más elevadas del Sistema Central de la península ibérica se explicaría porque las temperaturas estivales en esas regiones no alcanzan las temperaturas umbrales que podrían inhibir el vuelo de los adultos. No obstante, quedaría por identificar los valores de temperatura de ese umbral en futuras investigaciones. Además, en el perfil de las curvas bimodales, el segundo pico de capturas representa frecuentemente el máximo absoluto de la curva. Esto podría ser consecuencia de la suma de varios factores, como la larga duración de la curva de emergencia (Capítulo 2) y el periodo de inmadurez sexual de los imagos recién emergidos, durante el cual no se sentirían atraídos por el atrayente utilizado (Capítulo 5.1). Desde este enfoque, en efecto, podría explicarse la bimodalidad, frecuentemente asimétrica, observada en los resultados. Puesto que los periodos de emergencia suelen extenderse a lo largo de varios meses, generalmente entre mayo y agosto (Capítulo 2), en el momento de darse las temperaturas máximas estivales, aún quedarían insectos no emergidos e insectos inmaduros que no se capturarían en las trampas. Este reclutamiento de individuos, susceptibles de ser capturados una vez restablecidas las condiciones climáticas óptimas para el vuelo tras el periodo de temperaturas máximas estivales, podría ser en parte, responsable de que el segundo pico de capturas a finales de agosto y septiembre fuese especialmente acusado. Por otro lado, de forma general, la proporción de sexos de las capturas con el atrayente utilizado, es siempre favorable a las hembras (Capítulo 3). Esto es coherente con la formulación del atrayente, que incluye una feromona emitida por machos maduros de M. galloprovincialis, que con el resto de componentes sinergiza la atracción de ambos sexos, aunque segada hacia las hembras (PAJARES et al. 2010). No obstante, el análisis de la evolución de la proporción de sexos en las capturas a lo largo de todo el periodo de vuelo muestra un aumento significativo de la proporción de machos con respecto a la de hembras con el paso del tiempo, es decir, la proporción de sexos en las capturas empieza siendo favorable a las hembras para acabar siendo favorable a los machos al final del periodo de vuelo. Esto podría tener que ver con una disminución de la capacidad de respuesta de las hembras a las feromonas emitidas por los machos una vez cumplido su ciclo reproductivo, o con un aumento en la capacidad de respuesta de los machos a las feromonas emitidas por otros machos al final del periodo de vuelo como un mecanismo de compensación entre la necesidad reproductiva y el final de la vida del insecto (Capítulo 3). En este último caso, este nuevo reclutamiento de individuos podría suponer también parte de la explicación del segundo pico de capturas en las curvas bimodales. No obstante, tras estas observaciones, futuras investigaciones deberían ir encaminadas al análisis de la variación a lo largo del tiempo del comportamiento sexual de machos y hembras y de su capacidad de respuesta a la feromona (Capítulo 3). Por último, para llevar a cabo un estudio sobre los hábitos de vuelo circadianos de M. galloprovincialis, un conjunto de trampas cebadas con atrayentes fue muestreado con frecuencia horaria durante 2 semanas, a la vez que se registró la temperatura en el área de estudio (Capítulo

197
3). Monochamus galloprovincialis ha sido descrito como un insecto de hábitos crepusculares en algunos estudios (BONIFÁCIO 2009) o diurnos en otros (BERGER 2012). Pero hasta donde conocemos es la primera vez que se ha realizado un estudio de frecuencia de muestreo horaria. Los resultados muestran que el periodo circadiano del vuelo de M. galloprovincialis, en el momento del muestreo (finales de agosto y principios de septiembre), es diurno y que las capturas aumentan con el aumento de la temperatura, produciéndose máximos de capturas en las horas más calurosas del día. El periodo de tiempo estudiado se ciñó exclusivamente al momento en el que era esperable el máximo de capturas, es decir, principios de septiembre (Capítulo 3). En este periodo, aunque parece detectarse la existencia de un umbral inferior de temperatura que aparentemente inhibe el vuelo del insecto, no parece observarse el efecto de un umbral superior en las capturas, como sería esperable en virtud de lo argumentado sobre la bimodalidad de las curvas de vuelo líneas arriba (Capítulo 3). Si bien es cierto que las temperaturas alcanzadas en estas fechas no fueron especialmente altas. En cualquier caso, sería interesante extender el estudio de los vuelos circadianos de M. galloprovincialis a los periodos de máximas temperaturas estivales, así como al momento inicial del periodo de vuelo, con el objeto de poder completar la evaluación de la influencia de las tres variables que más probablemente podrían influir en el vuelo circadiano del insecto: irradiancia (o periodo horario), temperatura y fotoperiodo (época del año). La monitorización de M. galloprovincialis mediante el uso de trampas cebadas con atrayentes cairomonales y feromonales permite observar que otros muchos insectos saproxílicos son capturados junto a la especie diana. Desde esta perspectiva, y asumiendo que de entre todas las especies invasoras forestales destacan por su importancia los escarabajos perforadores de la madera (BROCKERHOFF et al. 2006), la rápida detección de estas especies exóticas es de fundamental importancia para implementar a tiempo las medidas fitosanitarias de los programas de erradicación pertinentes, así como para poder estimar las tasas de entrada de las mismas (BROCKERHOFF et al. 2006, RASSATI et al. 2015a). Por lo tanto, empleo de redes de seguimiento permanente y de trampas vigía para la detección temprana de organismos exóticos y especies de cuarentena, representa una de las mejores herramientas actuales de la gestión integrada de plagas (RASSATI et al. 2014, RASSATI et al. 2015b). Así mismo, la alta diversidad de organismos que son capturados con atrayentes generalistas (RASSATI et al. 2014, RASSATI et al. 2015b) abre una nueva e interesante línea de conocimiento permitiendo que, además de su funcionamiento como trampas vigía, dichas redes de muestreo permitan la caracterización y evaluación de las comunidades de insectos presentes en el hábitat de estudio (RASSATI et al. 2014). En este contexto, dos redes de muestreo sistemático de insectos saproxílicos instaladas en la Comunitat Valenciana (Red MUFFET, 15 parcelas, año 2013) y en Murcia (Red ESFP, 20 parcelas, años 2008-2010) permitieron el estudio de la comunidad de insectos relacionada con M. galloprovincialis. Cada una de las parcelas contaba con una trampa cebada con atrayentes genéricos (volátiles de hospedante y feromonas de escolítidos) y una estación meteorológica. De esta manera fue muestreada toda la variabilidad medioambiental de las formaciones de pinar ubicadas en el este y sudeste de la península ibérica, desde el nivel del mar hasta los 1700 m de altitud (Capítulo 4). Este análisis supone, en sí mismo, la presentación de una herramienta con un gran potencial en el ámbito de la investigación y como herramienta de gestión. Las redes ESFP y MUFFET analizadas han mostrado ser sistemas muy útiles de muestreo de las comunidades de insectos saproxílicos propias de los pinares sudorientales peninsulares tanto en el registro de la biodiversidad (con el aporte de una primera lista de 298 especies de coleópteros saproxílicos que abarca todos los ecosistemas dominados por pinares en el levante peninsular), como para la realización de estudios ecosistémicos forestales. Además, queda reforzada la importancia de las mismas como herramientas para alerta temprana o y para la vigilancia (redes vigía) (Capítulo 4).

198
En lo que se refiere al análisis específico de la comunidad asociada a de M. galloprovincialis, los resultados muestran que se trata de una especie ubicua, presente en toda la variabilidad medioambiental de los pinares del este y sudeste ibéricos, sin que sea posible asociarla significativamente a ninguna comunidad concreta de coleópteros saproxílicos. En otras palabras, la comunidad general de saproxílicos estudiada es muy homogénea respecto a la variación ambiental de las zonas de muestreo, y aunque pueden existir ciertas variaciones entre las comunidades de los diferentes ecosistemas, el rol que M. galloprovincialis desempeña en ellas, a lo largo de todo el gradiente estudiado, debe ser siempre el mismo. Con todo, los resultados apuntan a la existencia de posibles interacciones ecológicas claves entre el escolítido Orthotomicus erosus y M. galloprovincialis similar a las descritas entre otras especies, como los binomios M. carolinensis - Ips calligraphus o M. titillator - Dendroctonus frontalis (COULSON et al. 1976, MILLER 1984, 1985, FLAMM et al. 1989, DODDS et al. 2001b). Esta relación está fundamentada en la coincidencia fenológica y corológica de las dos especies mostrada en los resultados y en los roles que ambas desempeñan en los ecosistemas en los que están presentes (Capítulo 4.2). No obstante, abundante bibliografía muestra también la similitud en sus respuestas a infoquímicos (GIESEN et al. 1984, HUERTA et al. 2004, SEYBOLD et al. 2006, IBEAS et al. 2007, PAJARES et al. 2010, ETXEBESTE et al. 2012, ETXEBESTE et al. 2013a, ETXEBESTE et al. 2013b). Ambas especies muestran una preferencia compartida por las mismas características del material de cría (como el espesor de corteza, el diámetro o ubicación relativa en el árbol), así como la posible existencia de relaciones intragremio de ambas especies (AMEZAGA Y RODR GUEZ 1998, DODDS et al. 2001a, BONIFÁCIO 2009, HUNTER 2009), en la que habría que probar la depredación facultativa de las larvas del cerambícido sobre las del escolítido en árboles atacados De la misma manera, desde la perspectiva de análisis de redes, ambas se comportan como especies conectoras (de forma individual o conjuntamente) de una buena parte de los ecosistemas estudiados en la Red EFSP. Es decir, son especies que hacen de puente y facilitan las interacciones entre los distintos niveles tróficos (representados como distintas subredes o partes de la red), por lo cual podrían tener el valor añadido de ser especies clave en estos ecosistemas (Capítulo 4.1). En resumen, hay indicios que hacen pensar en la existencia una interacción clave entre estas dos especies, que debería ser estudiada en futuras investigaciones con el objetivo de explorar sus posibilidades en la gestión de sus poblaciones. Los resultados de la última parte de esta memoria, englobados en el Capítulo 5, dan respuesta a una serie de cuestiones vinculadas a la dispersión y a la población de M. galloprovincialis. Para la consecución de dicho objetivo, un total de 12 experimentos de marcaje-liberación-recaptura de adultos nativos o criados en laboratorio fueron desarrollados entre 2009 y 2012 en diferentes regiones de la península ibérica (Castellón, Teruel, Valencia y Murcia). Como primera conclusión, el análisis de la respuesta de los adultos recién emergidos al atrayente feromonal y cairomonal utilizado, fija en 8-9 días el periodo en el que, tanto machos como hembras, empiezan a responder al atrayente en campo (Capítulo 5.1). Estos resultados son coherentes con la mayoría de estudios que, basándose en uno u otro aspecto, evalúan la maduración sexual de esta especie (HELLRIGL 1971, FRANCARDI Y PENNACCHIO 1996, DAVID et al. 2014, SANCHEZ-HUSILLOS et al. 2015a, b), aunque hasta este momento ningún estudio había evaluado con la suficiente presión de muestreo (diaria en este caso) dicha respuesta en campo y en condiciones naturales.

199
Esta cuestión está íntimamente relacionada con la correspondencia entre las curvas de emergencia (Capítulo 2) y las curvas de vuelo (Capítulo 3), ya que éstas no pueden ser monitorizadas desde el mismo día de la emergencia, sino una exclusivamente vez pasado este periodo en el que supuestamente se produce la maduración sexual del insecto y, por tanto, se inicia la capacidad de respuesta a los atrayentes cairomonales y feromonales. Lo cual debe ser tenido en cuenta en la organización de las medidas de control de la enfermedad, especialmente en el calendario de actuaciones para la retirada de pies afectados por la enfermedad. Es decir, el seguimiento de las curvas de vuelo de M. galloprovincialis tendría un periodo de desajuste con respecto al inicio de las emergencias de los insectos, que sería la fecha adecuada a tener en cuenta. En cualquier caso, la incapacidad de capturar insectos en los primeros días tras la emergencia supone un problema en el control de la enfermedad, toda vez que se presupone la existencia de un vuelo fisiológico, o no reproductivo, durante estos primeros días, posiblemente para el fortalecimiento de los músculos alares, o de acumulación de reservas de grasa de cara a los esfuerzos de reproducción, en cualquier caso implica una maduración del insecto. La dispersión anterior a la posibilidad de captura de este insecto, asumiendo que el 95% de las infecciones de NMP se producen en las primeras 6 semanas (NAVES et al. 2007), supondría un obstáculo importante en la contención de la dispersión natural de la enfermedad mediante el uso de trampas. No obstante este vuelo fisiológico no se encuentra suficientemente estudiado ni comprobado en el caso de M. galloprovincialis. Y los únicos estudios que abordan, aunque indirectamente, esta cuestión (BONIFACIO 2009) parecen indicar que los desplazamientos del insecto durante los primeros días tras su emergencia son extremadamente reducidos (más de un 80% de M. galloprovincialis avistados tras su liberación en una parcela de 0.3 ha). Además el pico de infecciones registrado en el mencionado trabajo, se produce entre las semanas segunda y sexta tras la emergencia de M. galloprovincialis, lo que en la práctica se traduce a que los insectos ya responden al cebo semioquímico (NAVES et al. 2007). En este mismo sentido, los resultados de (SANCHEZ-HUSILLOS et al. 2015a) también muestran curvas muy similares de dispersión de insectos maduros y de inmaduros, simplemente mostrando un desfase de dos semanas entre ambas. Debe tenerse en cuenta que en estos estudios el muestreo tuvo una periodicidad semanal, por lo que es previsible que el desfase real, en caso de haber sido muestreado con más frecuencia, fuese más parecido al que muestran nuestros resultados. En esta misma sección (Capítulo 5.1) también se muestra la evidencia del “efecto borde” en la dispersión de este insecto en paisajes con mosaico agroforestal. Los análisis de la dispersión a larga distancia muestran la clara capacidad de M. galloprovincialis de dispersarse más allá de los límites de las masas forestales (Capítulo 5.4), siendo capaz de atravesar zonas sin arbolado en los paisajes en mosaico. No obstante, nuestros resultados muestran que se registran más capturas en el borde de la mancha forestal de origen. Esto podría explicarse por la retención de individuos que se resisten a salir de la mancha o bien por el retorno de algunos individuos dispersantes tras no encontrar zonas favorables más allá de los límites de la masa forestal Por último, también el diferente patrón de recaptura observado entre insectos nativos e insectos criados en laboratorio en un experimento de marcaje-liberación-recaptura realizado una en masa aislada (Capítulo 5.1) es coherente con la explicación apuntada en el Capítulo 3. En este caso, la llegada de las altas temperaturas estivales podría inhibir el vuelo de estos insectos pero, puesto que se realizó una única liberación de individuos criados en laboratorio, no habría un reclutamiento extra de individuos supuestamente no emergidos o inmaduros, por lo que el aumento de capturas al final del verano no sería evidente en el caso de los insectos criados en cautividad mientras que sí debería serlo en el de insectos nativos, como han demostrado las capturas. (Capítulo 5.1).

200
En la segunda sección de este último capítulo (Capítulo 5.2) se intenta estimar el nivel de población a partir de las capturas en trampas, una cuestión de máxima relevancia en el uso de trampas cebadas con atrayentes para el seguimiento de las poblaciones de M. galloprovincialis. Además también analiza la relación entre las características de la masa forestal y el tamaño poblaciones de esta especie. Aunque diferentes estudios demográficos de M. galloprovincialis han sido realizados recientemente en la península ibérica (SANCHEZ-HUSILLOS et al. 2015a, TORRES-VILA et al. 2015), ninguno de ellos ha estado enfocado a la evaluación de las características ambientales que puedan influir en la densidad poblacional. Este trabajo es, por tanto, el primer estudio demográfico de M. galloprovincialis realizado simultáneamente en dos masas de diferentes características con el objetivo de evaluar sus respectivas densidades poblacionales y las variables que las determinan. Los resultados muestran que tanto variables como la continuidad forestal, (por tanto, el mayor tamaño del parche de la mancha forestal), como una mayor naturalización de las masas, o la afección de un incendio pocos años antes de la fecha de estudio, están relacionadas con el mayor de las poblaciones de M. galloprovincialis. Por otro lado, en el ámbito de la gestión, la monitorización de las poblaciones de insectos de interés forestal (incluido M. galloprovincialis) mediante el uso de trampas cebadas con atrayentes cairomonales y feromonales es utilizada de manera general como una aproximación a la cuantificación relativa de las poblaciones, entre diferentes zonas o en diferentes momentos, con el objetivo de ajustar la gestión o de prevenir irrupciones epidémicas (BROCKERHOFF et al. 2006). Nuestros resultados muestran que esta aproximación tan generalizada debe ser tomada con cautela, o matizada consecuentemente, ya que, si bien es posible observar una relación directa entre el número de capturas de las trampas y el tamaño de la población estimada, esta relación no es aparentemente lineal. De este modo, mientras que las capturas medias para ambas zonas se diferencian a razón de 2:1, las estimaciones del tamaño de población muestran una relación de 3:1. Esta consideración debe ser tenida en cuenta, en los análisis de riesgo de expansión del NMP, así como en la evaluación de la eficacia del empleo de trampas cebadas con atrayentes cairomonales y feromonales en la disminución de la población en casos de brotes epidémicos. Los análisis contemplados en la tercera sección de este capítulo (Capítulo 5.3) se centran en la evaluación, mediante procedimientos de marcaje-liberación-recaptura, de la influencia del viento y las variables de masas en la dispersión de M. galloprovincialis. Se ha evaluado también la relación con las capturas de una serie de variables sintéticas, con el objetivo práctico de obtener las condiciones que debe cumplir una colocación óptima de las trampas en futuros estudios, en el sentido de maximizar el número de capturas. En resumen, aunque no está en discusión que el viento condiciona la dispersión de los volátiles y que la detección de éstos condiciona a su vez la orientación de los vuelos puntuales de M. galloprovincialis, nuestros estudios han evaluado la influencia del viento en la dispersión del insecto en superficies muy extensas, con registros de vuelos muy largos, y con periodos en libertad muy amplios (promedios de14 días entre la liberación y la recaptura). Los resultados indican que no puede asumirse que exista una influencia significativa ni de la intensidad ni de la dirección de los vientos dominantes en la dispersión de M. galloprovincialis a la escala del trabajo. Los resultados contradicen en parte a los de estudios previos, debido diferencias de escala (BONIFÁCIO 2009) o a la existencia de fuertes colinealidades entre la distribución de las masas y los vientos dominantes (GALLEGO et al. 2012).

201
En la segunda parte esta misma sección (Capítulo 5.3), los resultados muestran la correlación de la abundancia de material hospedante con la presencia del insecto en paisajes agroforestales con fragmentación forestal y el mayor número de capturas de M. galloprovincialis. En este sentido se observa una fuerte correlación de las capturas con todas las variables de masa habitualmente utilizadas en la caracterización forestal de las mismas, como densidad de pies, altura dominante, área basimétrica, fracción de cabida cubierta o volumen (Capítulo 5.3). De la aplicación de esta metodología sobre masas forestales continuas de menor superficie (próxima a las 100 ha), utilizando variables sintéticas y cualitativas (como la visibilidad de las trampas o la ubicación relativa de éstas con respecto a la masa, la espesura, edad, topografía, exposición, orientación o naturalización de las mismas) puede concluirse que la colocación óptima de las trampas, para maximizar el número de capturas en un rodal, debería adecuarse a una alta visibilidad y una ubicación próxima a los límites de la masa. (Capítulo 5.3). Nuestros resultados, en este sentido, avalan la necesidad de acometer estudios a escala europea con el objetivo de tratar de relacionar la abundancia de Monochamus spp. con variables de resistencia o conectividad del paisaje, tales como la cobertura local de pino o de otras coníferas, la edad de los rodales, la densidad, el estado sanitario o la fragmentación (Capítulo 5.3). Por otro lado también se ha puesto en evidencia la correlación del máximo de capturas con mínimos del índice de competencia de Hart, que describe la alta competencia entre pies. En la medida en que estos índices informan sobre la necesidad de aclarar la masa para que ésta crezca con el vigor adecuado, nuestros resultados parecen abrir la puerta a que, desde la perspectiva del control de la enfermedad, puedan ser planteados modelos de gestión enfocados a llevar a las masas forestales hacia estructuras de baja densidad y baja competencia con el objetivo de minimizar la abundancia del vector allá donde fuere necesario (Capítulo 5.3). Por último, parece detectarse un sesgo en las capturas favorables a los machos los paisajes muy fragmentados (cuando lo habitual es que exista un sesgo hacia las hembras). Esto podría deberse al carácter pionero de los machos como colonizadores de nuevos espacios. Si esta relación se probara, indicaría que el tamaño de las manchas forestales y la fragmentación del paisaje,) podría, no solo influir en la abundancia de M. galloprovincialis (Capítulo 5.2) sino también en la proporción de sexos presentes en ellos. Por último, la última sección de este capítulo (Capítulo 5.4) representa una colección de datos de distancias de vuelos máximas tomadas durante 3 años en diferentes zonas del levante de la península ibérica (Murcia, Teruel y Castellón). Con ellos puede concluirse que M. galloprovincialis posee suficiente capacidad de dispersión como para recorrer distancias de varios kilómetros en pocos días, a velocidades de dispersión de al menos 1500 m/día (ya que es razonable pensar que el insecto no llega al lugar de captura en línea recta) y longevidades de individuos cercanas a los 3 meses. La máxima distancia de dispersión observada fue de 13600 m, aunque también fue constatada una recaptura a 22100 m del punto de liberación, aunque en este último caso no puede asegurarse que produjese de forma natural o con el concurso humano, posiblemente como polizón en un vehículo. El hecho de que algunos individuos puedan dispersarse a estas distancias tan grandes plantea un problema muy importante en la erradicación de la enfermedad y en la contención de su expansión. Estos datos, junto a los altos porcentajes de recapturas a más de 3000 m, sobre el 2%, hacen patente que las medidas profilácticas establecidas en España y China para evitar la dispersión natural de Monochamus sp y B. xylophilus, consistentes en la eliminación de las especies hospedantes en bandas de varios kilómetros de anchura, podrían resultar ineficaces en el control de la dispersión de los vectores y, por tanto, de la enfermedad.

202
Sección 6.2 Recomendaciones para la gestión del NMP en función de su
fenología y dispersión. Un modelo de gestión forestal aplicado al control.
Antecedentes La primera detección de Bursaphelenchus xylophilus en Europa se produce en la Península de Setúbal en 1999 (MOTA et al. 1999). A partir de ese momento, se demarca la zona afectada y aplican medidas de control. Desde 2008, todo el conjunto de Portugal continental es designado zona demarcada y se delimita una zona de amortiguamiento que ocupa 20 kilómetros de ancho lo largo de la frontera con España. La zona infestada se divide en dos zonas: la zona de intervención (ZI), donde realmente se identificaron los árboles infestados y el resto. Después de varios años de aplicación de medidas de control contra el NMP, las autoridades portuguesas consideran que la erradicación no es viable y por lo tanto deciden contener PWN en la zona infestada y concentrar sus acciones de control en la zona de amortiguamiento de la zona demarcada del Portugal continental. Al mismo tiempo, se intenta retrasar y controlar la propagación real de la plaga mediante la aplicación de determinadas medidas en la zona infestada (Rodríguez, J. M., com. pers. ;VICENTE et al. 2012). Bursaphelenchus xylophilus se detecta en España por primera vez en octubre de 2008, en la Sierra de Dios Padre, en la Comunidad Autónoma (CA) de Extremadura (ROBERTSON et al. 2011). La detección se produce en un único árbol infectado, fuera de la masa forestal, y situado a 12,5 km de una industria de procesamiento de la madera y a 40 km de la frontera con Portugal. La detección del árbol sintomático se produce durante la prospección sistemática de una cuadrícula de 2 x 2 km, en el trayecto de uno de los puntos de muestreo a otro. Después de aplicar las medidas de erradicación contenidas en la Decisión de la Comisión 2006/133/CE el foco es declarado erradicado por las autoridades españolas el 16 de enero de 2013. Tras la detección de este primer foco, otros tres han sido detectados en España, actualmente en curso de erradicación. En noviembre de 2010, se declara un foco en As Neves (Galicia) (ABELLEIRA et al. 2011). En febrero de 2012, se encuentra de nuevo un único árbol infectado en Valverde del Fresno (Extremadura) a 595 metros de la frontera con Portugal. En diciembre de 2013, en Sancti-Spíritus (Castilla y León), se encuentran 135 árboles infestados en un rodal de bosque mixto de 150 hectáreas en una zona de alto riesgo debido a la proximidad de una ruta de transporte principal de madera de Portugal (ZAMORA et al. 2015). La zona había sido objeto de reconocimiento en el año 2012 como parte de la cuadrícula para la prospección de PWN, y las muestras tomadas de los árboles sintomáticos y asintomáticos dieron resultados negativos. En todos estos brotes, son establecidas zonas demarcadas y medidas de erradicación conforme a la legislación vigente (MAGRAMA 2015). Una reciente evaluación del impacto económico cuantitativo de la enfermedad del marchitamiento de pino en Europa ha pronosticado una pérdida acumulada para España de 11,9 mil millones € desde 2008 a 2030. En el mejor de los casos España quedaría con el 67,5% de sus bosques de coníferas severamente afectadas (SOLIMAN et al. 2012). Los puntos débiles en el control de la enfermedad son dos. El primero está relacionado con la dificultad intrínseca del control de la enfermedad por razones ya mencionadas (Capítulo 1): la gran cantidad de hospedantes susceptibles al NMP, su ciclo biológico extremadamente corto en condiciones favorables, su alta fecundidad y fertilidad, la alternancia entre fases fitófaga y micófaga, la resiliencia de sus estadios larvarios, o el hecho de que su vector, de hábitos xilófagos, permanezca la mayor parte del tiempo oculto en el interior de la madera o debajo de la corteza, fuera de alcance de posibles tratamientos insecticidas. El segundo punto del problema tiene que ver con las dificultades administrativas, en especial, con el régimen de propiedad del suelo en las zonas afectadas del centro de Portugal y del N. de España (Galicia y

203
zonas de Castilla-León), con altos porcentajes de propiedad privada distribuida en parcelas muy pequeñas y cuyos propietarios son de difícil localización (Rodríguez Com. Pers.; Sánchez, Com. Pers.).
Recomendaciones Las estrategias de gestión para reducir el daño de NMP en la península ibérica deben ser enfocados necesariamente al control de su vector, M. galloprovincialis. La dispersión de este vector puede ser de dos tipos: natural y facilitada por el hombre. Ambos tipos de dispersión son incluidas en los planes de contingencia nacionales y autonómicos, así como en la normativa europea al respecto. Actualmente las herramientas de control del NMP aplicada en Europa y en la península ibérica se reducen a la selvicultura y a la inspección fitosanitaria, en concreto, a la tala y destrucción completa de los pies sintomáticos detectados antes del inicio del periodo de emergencia de M. galloprovincialis para evitar la dispersión de los vectores infectados, y a la prospección de las zonas con mayor riesgo de entrada de la enfermedad, respectivamente. Las recomendaciones propuestas a continuación suponen mejoras para estas dos herramientas de control del NMP.
Inspección fitosanitaria: monitorización
Los trabajos de PAJARES et al. (2010) e IBEAS et al. (2007) han permitido el desarrollo y comercialización de un cebo cairo-feromonal que permite una alta tasa de capturas de ambos sexos de M. galloprovincialis durante prácticamente todo su periodo de vuelo. Esto hace posible trampeos, no sólo para la monitorización de los periodos de vuelo, sino también para la reducción de la población de M. galloprovincialis en determinadas zonas sensibles (SANCHEZ-HUSILLOS et al. 2015a, TORRES-VILA et al. 2015). El diseño de la trampa también ha sido optimizado, aumentando la tasa de capturas del insecto en vivo (ÁLVAREZ et al. 2015), condición indispensable para la detección de nematodos en vector. La adecuada instalación de las trampas, en tiempo y ubicación servirá para maximizar las capturas. Desde esta perspectiva el objetivo de los trampeos puede ser diferente:
1. Monitorización 2. Extracción de población
a. Maximización del nivel de capturas (minimización de la abundancia de población)
b. Maximización del control de la enfermedad (minimización de la dispersión de la enfermedad)
De acuerdo con las curvas de emergencia mostradas en los resultados del Capítulo 2 de este trabajo, el percentil 1 de las emergencias más tempranas se da el 26 de mayo mientras que el percentil 50 se da el 11 de junio. Según NAVES y DE SOUSA (2009), estos percentiles son algo más tardíos en la península de Setúbal, Portugal, ocurriendo el percentil 1 el 31 de mayo y el percentil 50 el 10 de Julio. Conforme a los resultados de NAVES et al. (2007), más del 95% de las transmisiones se dan en las primeras 8-9 semanas. Esto marca una posible ventana de actuación para la captura de los vectores con el objetivo de contener o monitorizar la presencia de la enfermedad. En este caso debe ser tenida en cuenta que existen diferencias fenológicas en la emergencia en península ibérica, posiblemente por adaptaciones locales de las poblaciones

204
de M. galloprovincialis(Capítulo 2). Por lo tanto la gestión debe adaptarse al conocimiento de la fenología local de la emergencia del insecto, en la medida de lo posible. Por otro lado, de acuerdo con la fenología del vuelo presentada en los resultados del Capítulo 3, los máximos del vuelo del vector se producen en el mes de septiembre. Esto marca una nueva y diferente ventana de actuación con el fin de maximizar el número de capturas del vector. La emisión correcta y constante de los volátiles que conforman el atrayente Galloprotect 2D garantizado por el fabricante (SEDQ S.L., Barcelona) entre 45-50 días y el periodo de vuelo completo de M. galloprovincialis, según los resultados mostrados en el Capítulo 3, tiene una duración aproximada de 6 meses (de finales de mayo a principios de noviembre). Por lo tanto, son necesarias 1-2 reposiciones de los atrayentes para cubrir el muestreo del periodo de vuelo completo, siempre que no aparezcan en el mercado nuevos productos con otra duración. Más allá de las directrices para la captura masiva, discutidas por SANCHEZ-HUSILLOS et al. (2015a) y TORRES-VILA et al. (2015), y que exigirían el trampeo de M. galloprovincialis durante todo su periodo de vuelo, el uso de las trampas como herramienta para el control del NMP puede ser optimizado para la consecución de otros objetivos diferentes a la captura masiva. A continuación se incluyen recomendaciones para su optimización:
1. Si no hubiere posibilidad de acometer la colocación de las 3 tandas de atrayente, en función de las diferentes ventanas de actuación comentadas, la segunda (y última) reposición de atrayente debería ser descartada, de manera que quedasen cubiertos los meses de junio-julio (en una primera colocación) y agosto-septiembre (en la reposición). En la primera parte del trampeo, el objetivo sería minimizar la propagación del nematodo, y en la segunda minimizar la población de vectores.
2. Si no hubiese posibilidad de sustituir los atrayentes, y sólo fuese posible muestrear
durante un periodo de 50 días (duración estándar de la emisión correcta de los atrayentes), los objetivos del trampeo deberían ser concretados:
a. Monitorización vs. extracción de población. b. Minimización de la propagación del NMP vs. minimización de la población de
vectores. Esta doble dicotomía está fuertemente relacionada con la zona de colocación de las trampas y con la gestión diseñada para esa zona:
1. Zonas con presencia de la enfermedad (zonas demarcadas). La gestión está orientada a la monitorización o a la minimización de la propagación de la plaga. En todos estos casos el periodo de muestreo adecuado sería junio-julio: La monitorización puede ser, bien de la intensidad de presencia de la enfermedad, o
bien de la velocidad de dispersión de la misma, muestreando los bordes de las zonas afectadas, lo que ayudará en el proceso de toma de decisiones sobre la optimización de los recursos disponibles.
La minimización de la propagación del NMP tendrá lugar en las zonas limítrofes entre áreas infectadas y no infectadas.
2. Zonas no infectadas:
Riesgo alto de infección. La utilización del trampeo puede ir encaminado a: o Detección temprana de la enfermedad a través de la detección de vectores
infectados (monitorización). Ventana de actuación: junio-julio.

205
o Extracción de la posible población infectada que aparezca (si se sospecha que la entrada de la enfermedad es inminente). Ventana de actuación: junio-julio.
o Extracción del máximo posible de población de M. galloprovincialis con el objetivo de generar una zona de menor incidencia potencial de la enfermedad, es decir, con baja abundancia de vectores. Ventana de actuación: agosto-septiembre.
Riesgo bajo de infección. La decisión de gestión más probable será de carácter,
preventivo, mediante la monitorización de las poblaciones de M. galloprovincialis con el objetivo de conocer sus dinámicas poblacionales. Es decir, las trapas deberían ser colocadas en agosto-septiembre, cuando las capturas por trampa serán mayores.
Respecto a los diferentes parámetros que pueden regir la ubicación relativa de las trampas, algunos aspectos han sido analizados con anterioridad a esta memoria y se encuentran disponibles en la literatura. La colocación en altura de las trampas fue estudiada y discutida por BONIFÁCIO (2009) recomendándose su colocación a 5 m de altura, en la copa de los árboles (donde es más probable encontrar a los insectos), si la operatividad lo permite. Los trabajos de ETXEBESTE et al. (2015) y SANCHEZ-HUSILLOS et al. (2015a) aportaron información práctica al respecto de la distancia de separación entre trampas o de la densidad de éstas para cumplir con los distintos objetivos: monitorización o extracción de población del vector. ETXEBESTE et al. (2015) analizaron la dispersión de M. galloprovincialis dentro de un bosque continuo, ajustada a dos tipos de modelos: mecánico y empírico. El modelo mecánico mostró que el 50% de los individuos se dispersaban entre 480-532 m, mientras que el 99% de la población lo hacían a aproximadamente unos 2500 m. Sin embargo, la utilización de un modelo empírico ajustó que el 50% de la dispersión escarabajos alcanzaban los 230 m en su dispersión y que el 99% se dispersaba hasta los 3500 m. En este sentido, el modelo empírico proporcionó resultados más conservadores, por lo que se recomienda que sea preferiblemente éste el tenido en cuenta en decisiones de gestión. No obstante, conviene recordar que los resultados de ETXEBESTE et al. (2015) son referidos a masas continuas y que los resultados analizados en los Capítulos 6 y 7 de este trabajo parecen indicar que estos rangos de dispersión son mayores en casos de masas muy abiertas, mosaicos agroforestales o ausencia de árboles hospedantes en las inmediaciones. Por otro lado, ETXEBESTE et al. (2015) estimaron el radio de acción estacional de estas trampas (seasonal sampling range), situándolo en un promedio de 535 m. Ésta es, por tanto, la distancia óptima que debe regir la colocación de trampas para monitorización de M. galloprovincialis (aproximadamente 500 m de radio o 1 km entre trampas). Complementariamente, en el mismo trabajo también se indicó que es necesaria una mayor densidad de trampas (o una menor distancia entre ellas) para llevar a cabo una captura masiva del escarabajo de manera eficaz. Aunque el estudio está realizado sobre densidades moderadas de población del insecto, puede tomarse, como aproximación, una densidad de 1 trampa/ha, 60 m de radio o 120 m entre trampas para el trampeo masivo óptimo (ETXEBESTE et al. 2015, SANCHEZ-HUSILLOS et al. 2015a). Si bien otros estudios han criticado la aplicación de este método de reducción de población (ver TORRES-VILA et al. 2015), tales conclusiones pueden ser deberse al sesgo por la realización del estudio en un área muy inferior (200x200 m) a la capacidad dispersiva media de M. galloprovincialis.

206
Un nuevo aporte, en este sentido, suponen los resultados mostrados en el Capítulo 6. Según éstos el lugar de colocación de las trampas está correlacionado con el número de capturas, de manera que, en caso de poder elegir, las trampas deberán estar colocadas en lugares visibles y en zonas cercanas a los límites de las masas del rodal que se quiera muestrear.
Tratamientos silvícolas. Un modelo de gestión enfocado al control del NMP
Como ya se ha indicado, la segunda herramienta de control del NMP aplicada en Europa y en la península Ibérica es la silvicultura aplicada al control del NMP, en concreto, la tala y destrucción completa de los pies sintomáticos detectados antes del inicio del periodo de emergencia de M. galloprovincialis para evitar la dispersión de los vectores infectados. Esta herramienta es un instrumento activo, aplicable a partir del momento en que la infección es detectada, y está concebida para frenar la dispersión de los vectores infectados. Su eficacia depende de la capacidad del gestor de detectar el mayor número posible de árboles sintomáticos y de su capacidad de destruirlos conforme a un calendario fijado en respuesta a la fenología de la emergencia de M. galloprovincialis. Lo que, a continuación, se propone es un modelo de gestión silvícola diseñado específicamente para la gestión del NMP, tanto en referente a su prevención, contención o erradicación. La situación derivada de la detección y expansión del NMP en Portugal ha provocado la necesidad de diseñar un modelo de gestión específico para la prevención y contención de la dispersión de esta enfermedad y aplicable a regiones con diferentes características en relación a la presencia o riesgo de presencia del organismo patógeno. Se necesita por tanto un modelo de aplicación inmediata, que dé resultados rápidamente y que pueda mantenerse en el futuro. Existen una serie de factores clave a tener en cuenta a la hora de diseñar el modelo:
Zona de implementación: Zona Tampón, Zona Demarcada, Zona libre, etc. Objetivos finales:
o Reducir la probabilidad de dispersión de la enfermedad (reducir la probabilidad de infección).
Reducción del material hospedante Reducción de la abundancia de población del vector de la enfermedad
(M. galloprovincialis) o Incrementar la probabilidad de detección de la enfermedad y destrucción de los
focos Mejora de los sistemas de detección de los focos de infección
(muestreos sistemáticos) El objetivo de los modelos de gestión teóricos de P. pinaster es la consecución de una estructura de masa regular, con rotaciones de corta cada 15-20 años, recomendando dejar a los pies alcanzar una edad cercana a los 80-100 años para realizar una corta a hecho y una siembra posterior. No obstante, es frecuente en montes de P. pinaster puro que se corten a hecho a mayor edad. En casos de montes ordenados de pinares resinados y en pinares del Sistema Ibérico, los turnos de corta se han establecido frecuentemente en los 100 años (JCYL 1901). Este modelo genera una fase, que se sitúa a una edad del arbolado comprendida entre los 20 y los 40 años, donde hay hiperdensidad, mucha competencia y, por tanto, aparición de

207
decaimiento y muerte del arbolado (WYCKOFF Y CLARK 2002, BRAVO-OVIEDO et al. 2006, LINARES et al. 2010). El nuevo modelo, por tanto, tiene como objetivo evitar esta fase de hiperdensidad en la que se producen decaimiento y muerte de pies de manera significativa. El principal factor de predisposición al decaimiento del pino rodeno relacionado con la estructura de las masas se debe a causas fisiológicas, en concreto, a una pérdida de funcionalidad en la estructura de movimiento de la savia. A altas densidades, y alta competencia entre pies, el crecimiento primario en busca de la luz provoca copas muy mermadas y fustes muy altos y muy delgados, por lo que la traslocación de nutrientes se complica al no ser suficientemente grande la copa para mover la savia. Esto provoca la pérdida de vigor, la aparición de síntomas de decaimiento y, por tanto, el aumento de la susceptibilidad a plagas y enfermedades (WYCKOFF Y CLARK 2002, BRAVO-OVIEDO et al. 2006, LINARES et al. 2010). El modelo planteado se basa en los criterios para la ordenación forestal recogidos por PROSILVA para la aplicación de selvicultura próxima a la naturaleza (LANDA 2009, GARCÍA GÜEMES Y CALAMA 2013, PASCUAL et al. 2013, SABÍN GALÁN 2013, SABÍN GALÁN et al. 2013). Este tipo de ordenación establece criterios que se ajustan a los requerimientos del problema concreto provocado por la dispersión de la PWD a la vez que responde a los principios fundamentales que debe garantizar la gestión forestal sostenible: persistencia, conservación, mejora de la capacidad productiva de suelo y vuelo, y máximo rendimiento de bienes y servicios para la sociedad (SABÍN GALÁN et al. 2013). SABÍN GALÁN et al. (2013) definen los cambios fundamentales que deben realizarse en la redacción de planes de gestión bajo criterios de selvicultura próxima a la naturaleza, muchos de los cuales son perfectamente aplicables al modelo propuesto. La ordenación según los criterios establecidos supone redefinir algunos de los conceptos clásicos utilizados y afrontar algunas de las partes desde una perspectiva diferente, no obstante, la mayor parte del contenido es coincidente con los Planes de Gestión Forestal Sostenible (PGFS) tradicionales (SABÍN GALÁN 2013, SABÍN GALÁN et al. 2013) Dentro de este marco conceptual, el modelo fundamental que mejor se adapta a la consecución de los objetivos planteados es el de entresaca. El objetivo de la aplicación de los criterios silvícolas que se exponen a continuación será la consecución de una estructura de masa irregular, con espesura incompleta o no trabada, mucho vigor, sin solapamiento de copas ni existencia de masa debajo de las copas. La aplicación del modelo estará supeditada a la accesibilidad y a la posibilidad física de practicar la selvicultura. Por su parte, las diferentes características del modelo deberán adaptarse a la situación de las masas a gestionar, principalmente existencias y crecimiento. En caso de pinares abiertos no es de aplicación el modelo silvícola. Estos pinares ya estarán suficientemente abiertos como para permitir una inspección eficiente. La retirada de pies decadentes o recién muertos, inevitablemente, deberá responder a cortas específicas, no siempre rentables. En cualquier caso, la superficie mínima de actuación para mejora de la rentabilidad económica será consecuentemente mayor. Para el caso de pinares suficientemente capitalizados, es decir, con suficientes pies por hectárea, se describen los aspectos fundamentales del modelo de gestión propuesto: Objetivos
1. Llevar la masa a una estructura irregular con volúmenes objetivo de 150-200 m3/ha

208
2. Que la gestión silvícola sea rentable o autofinanciable. La inversión inicial (y la rentabilidad) dependerá en gran medida de la dotación de infraestructura (básicamente la red de pistas) de la zona.
Características del modelo silvícola a aplicar a masas con el volumen objetivo: Rotaciones de corta cada 5 y 7 años (mucha frecuencia de corta) (SABÍN
GALÁN 2013), lo que permitirán visitar (y gestionar) toda la superficie cada poco tiempo, para evitar grandes periodos sin revisión que puedan dar lugar a mortalidades/decaimientos del arbolado. De esta forma se facilitará la inspección de la enfermedad.
En cada corta se sacará más de 25-30 m3/corta, que es el Límite Inferior Comercial, para que sea rentable y ejecutable. Este volumen deberá ser similar al crecimiento de la masa en ese periodo. Se consideran, como cifras de referencia, crecimientos de 4 m3/ha×año para las calidades medias de estación y 7 m3/ha×año para las calidades altas (ABEJÓN Y LORANCA 1989). Para estos crecimientos se plantearán dos periodos de rotación. La superficie mínima para su aplicación será de 10-20 ha.
Se plantean dos modelos silvícolas diferentes en función de la calidad de estación, con dos periodos de rotación y masas objetivo diferentes:
o Buena calidad de estación: se llevará a unas existencias de 200 m3/ha, con periodo de rotación de 5 años.
o Calidad media de estación: se llevará a unas existencias de 150 m3/ha, con periodo de rotación de 7 años.
Diseño de actuaciones Primera intervención Para todas las masas:
Apertura y establecimiento (si no existe) de una red de pistas adecuada.
Apertura y establecimiento de una Red de Saca:
Criterios de selección de árboles de porvenir:
o Selección en función del vigor (tamaño de copa).
o Extracción de pies dominados y poco vigorosos.
o Selección enfocada a la individualización de copas.
o Como orientación para la selección pueden contemplarse las siguientes características en los árboles de porvenir: clase sociológica dominante o codominante, copa equilibrada o con posibilidad de reequilibrarse, ramas delgadas, distancia entre pies mayor de 6 m salvo cuando dos pies cercanos presenten un modelo de crecimiento unitario, buena calidad tecnológica del fuste, recto y sin defectos (a igualdad de condiciones de vigor).
Para las masas jóvenes regulares: Claras moderadas. Se abrirán las calles y se cortarán los pies de árboles enfermos, los
dominados y todos aquellos que tengan mayor posibilidad de morir en los próximos

209
años. Por último, se eliminará algo de competencia a los mejores pies seleccionados. De este modo la masa continuará capitalizándose y se podrá volver a intervenir en la misma en un periodo corto de rotación.
Objetivo: evitar la fase no deseable (edades 20-40 años) de hiperdensidad, en la que hay mucha competencia y mucho decaimiento/mortalidad.
Para las masas adultas capitalizadas: Claras fuertes: Extracción que puede ser mayor del 30% de la masa. Aproximadamente
el 20% puede abrirse para la realización de vías de saca. En las masas en las que sea posible, se intentará dejar la masa en los volúmenes objetivo: 150-200 m3/ha.
Intervenciones posteriores Para las masas adultas capitalizadas:
Rotación de corta cada 5-7 años. Corta del crecimiento. La masa crecerá a un ritmo de 4-7 m3/ha×año. Se cortará de 25 a 30 m3/corta (Límite Inferior Comercial)
El criterio de selección de los árboles de porvenir seguirá siendo en función del vigor (tamaño de la copa). No se utilizarán criterios basados en las calidades (calidad de la madera, del fuste, etc.) ni por diámetros, salvo en igualdad de condiciones. Los árboles adultos que no tengan un especial valor ecológico o que no aumenten su valor económico por no tener buena calidad tecnológica o buen crecimiento se podrán eliminar para dar paso al arbolado joven o bien para abrir huecos con el fin de promover la regeneración.
El objetivo será tener una Fracción de Cabida Cubierta (FCC) del 60-70% con espesura incompleta y pies muy vigorosos.
Señalamiento de la Corta:
o Pies con decaimiento y poco vigorosos
o Corta de cosecha de arbolado adulto con dos objetivos, individualizar las copas y abrir huecos. Estos huecos serán colonizados rápidamente por regenerado, que será clareado en la siguiente intervención que se realice en la masa.
o La corta será diferente en:
Regenerado: cortas de mejora (dejando 2 pies/hueco)
Masa joven: cortas para la individualización de copas, sin abrir huecos.
Masa adulta: cortas para la individualización de copas y apertura de huecos.
La madera se clasificará en las siguientes clases:
o Madera aserrada
o Embalaje
o Trituración

210
Mejoras del modelo
o Se distribuye la inversión del clareo en el tiempo. o No se fuerza la regeneración de una sola especie: Silvicultura Adaptativa. o Permite versatilidad y capacidad de adaptación a un mercado de la madera cambiante.
Se obtiene un monte similar siempre a lo largo del tiempo (irregular) o La productividad del monte no cambia con el modelo de gestión. En este caso será 4-5
m3/ha×año aproximadamente. o Se corta cada árbol en su momento. Un árbol vigoroso que tenga buena calidad y esté
aumentando su valor económico cada año que pase, puede ser que un árbol se corte con 80cm de diámetro y 140 años.
o La FCC del estrato dominante será aproximadamente del 60%. El resto (40%) serán huecos en regeneración.
o Las calles de desembosque y las pistas harán el monte transitable. o Se evitan los costes de la regeneración artificial y los posibles problemas para la
regeneración derivados de la apertura de superficies demasiado grandes, como la desecación producida por el exceso de insolación.
o Se evitan las cortas a hecho, que pueden provocar dispersiones más largas de M. galloprovincialis a la vez que generan impacto visual y rechazo social.
o Permite mantener el uso recreativo del monte.

211
Bibliografía
ABEJÓN, J. G. y J. G. LORANCA (1989). Tablas de producción de densidad variable para"
Pinus pinaster Ait." en el Sistema Central. ABELLEIRA, A., A. PICOAGA, J. P. MANSILLA y O. AGUIN (2011). Detection of
Bursaphelenchus xylophilus, Causal Agent of Pine Wilt Disease on Pinus pinaster in Northwestern Spain. Plant Disease 95(6): 776-776.
ÁLVAREZ, G., I. ETXEBESTE, D. GALLEGO, G. DAVID, L. BONIFACIO, H. JACTEL, E. SOUSA y J. A. PAJARES (2015). Optimization of traps for live trapping of Pine Wood Nematode vector Monochamus galloprovincialis. Journal of Applied Entomology 139(8): 618-626.
(1998). Resource partitioning of four sympatric bark beetles depending on swarming dates and tree species. Forest Ecology and Management 109(1): 127-135.
BERGER, P. (2012). Coléoptères Cerambycidae de la faune de France continentale et de Corse: actualisation de l'ouvrage d'André Villiers, 1978, ARE.
BLANCKENHORN, W. U., A. F. G. DIXON, D. J. FAIRBAIRN, M. W. FOELLMER, P. GIBERT, K. VAN DER LINDE, R. MEIER, S. NYLIN, S. PITNICK, C. SCHOFF, M. SIGNORELLI, T. TEDER y C. WIKLUND (2007). Proximate causes of Rensch's rule: Does sexual size dimorphism in arthropods result from sex differences in development time? American Naturalist 169(2): 245-257.
BOGICH, T. L., A. M. LIEBHOLD y K. SHEA (2008). To sample or eradicate? A cost minimization model for monitoring and managing an invasive species. Journal of Applied Ecology 45(4): 1134-1142.
BONIFÁCIO, L. F. P. (2009). Impacte e evolução da doença da murchidão dos pinheiros (pine wilt disease) na zona afectada a sul do Rio Tejo.
BRAVO-OVIEDO, A., H. STERBA, M. DEL RÍO y F. BRAVO (2006). Competition-induced mortality for Mediterranean Pinus pinaster Ait. and P. sylvestris L. Forest Ecology and Management 222(1): 88-98.
BROCKERHOFF, E. G., A. M. LIEBHOLD y H. JACTEL (2006). The ecology of forest insect invasions and advances in their management. Canadian Journal of Forest Research 36(2): 263-268.
COULSON, R. N., A. M. MAYYASI, J. L. FOLTZ y F. P. HAIN (1976). Interspecific competition between Monochamus titillator and Dendroctonus frontalis. Environmental Entomology 5(2): 235-247.
DAVID, G., B. GIFFARD, D. PIOU y H. JACTEL (2014). Dispersal capacity of Monochamus galloprovincialis, the European vector of the pine wood nematode, on flight mills. Journal of Applied Entomology 138(8): 566-576.
DODDS, K. J., C. GRABER y F. M. STEPHEN (2001a). Facultative intraguild predation by larval Cerambycidae (Coleoptera) on bark beetle larvae (Coleoptera : Scolytidae). Environmental Entomology 30(1): 17-22.
DODDS, K. J., C. GRABER y F. M. STEPHEN (2001b). Facultative intraguild predation by larval Cerambycidae (Coleoptera) on bark beetle larvae (Coleoptera: Scolytidae). Environmental Entomology 30(1): 17-22.
ETXEBESTE, I., G. ALVAREZ y J. PAJARES (2013a). Log colonization by Ips sexdentatus prevented by increasing host unsuitability signaled by verbenone. Entomologia Experimentalis Et Applicata 147(3): 231-240.
ETXEBESTE, I., G. ÁLVAREZ, G. PÉREZ y J. A. PAJARES (2012). Field response of the six-toothed pine bark beetle, Ips sexdentatus (Col.: Curculionidae, Scolytinae), to pheromonal blend candidates. Journal of Applied Entomology 136: 431-444.

212
ETXEBESTE, I., J. L. LENCINA y J. PAJARES (2013b). Saproxylic community, guild and species responses to varying pheromone components of a pine bark beetle. Bulletin of Entomological Research 103: 497-510.
ETXEBESTE, I., E. SANCHEZ-HUSILLOS, G. ÁLVAREZ, H. MAS I GISBERT y J. PAJARES (2015). Dispersal of Monochamus galloprovincialis (Col.: Cerambycidae) as recorded by mark-release recapture using pheromone traps. Journal of Applied Entomology: In press.
FLAMM, R., R. COULSON, P. BECKLEY, P. PULLEY y T. WAGNER (1989). Maintenance of a phloem-inhabiting guild. Environmental Entomology 18(3): 381-387.
FRANCARDI, V. y F. PENNACCHIO (1996). Note sulla bioecologia di Monochamus galloprovincialis (Olivier) in Toscana e in Liguria (Coleoptera Cerambycidae). Redia 79: 153-169.
FUTAI, K. (2013). Pine Wood Nematode, Bursaphelenchus xylophilus. Annual Review of Phytopathology 51(1): 61-83.
GALLEGO, D., F. SANCHEZ-GARCÍA, H. MAS, M. CAMPO y J. L. L. GUTIÉRREZ (2012). Estudio de la capacidad de vuelo a larga distancia de" Monochamus galloprovincialis"(Oliver 1795).(Coleoptera: Cerambycidae) en un mosaico agro-forestal. Boletín de sanidad vegetal. Plagas 38(1): 109-124.
GARCÍA GÜEMES, C. y R. CALAMA (2013). La Selvicultura próxima a la naturaleza como estrategia adaptativa al cambio climático. Congresos Forestales.
GIESEN, H., U. KOHNLE, J. VITÉ, M. L. PAN y W. FRANCKE (1984). Das aggregationspheromon des mediterranen Kiefernborkenkäfers Ips (Orthotomicus) erosus. Zeitschrift für Angewandte Entomologie 98(1‐5): 95-97.
HARAN, J., A. ROQUES, A. BERNARD, C. ROBINET y G. ROUX (2015). Altitudinal Barrier to the Spread of an Invasive Species: Could the Pyrenean Chain Slow the Natural Spread of the Pinewood Nematode? PLoS ONE 10(7): e0134126.
HELLRIGL, K. G. (1971). Die Bionomie der europaischen Monochamus-Arten (Coleopt., Cerambycid.) und ihre Bedeutung fur die Forstund Holzwirtschaft. Redia 52: 367-509.
HOCH, G., D. MITTERMAYR y H. KREHAN (2015). Monochamus-Arten als potenzielle Vektoren des Kiefernholznematoden in Österreich: Lockstofffallen zum Monitoring des Fluges. Forstschutz Aktuell 60(61): 61.
HUERTA, A., F. ROBREDO, J. DIEZ y J. A. PAJARES (2004). Life-cycle characteristics of Tetrastichus turionum, a parasitoid of the European pine shoot moth, Rhyacionia buoliana. Journal of Applied Entomology 128(9-10): 628-632.
HUNTER, M. D. (2009). Trophic promiscuity, intraguild predation and the problem of omnivores. Agricultural and Forest Entomology 11(2): 125-131.
IBEAS, F., D. GALLEGO, J. DIEZ y J. PAJARES (2007). An operative kairomonal lure for managing pine sawyer beetle Monochamus galloprovincialis (Coleoptera: Cerymbycidae). Journal of Applied Entomology 131(1): 13-20.
INÁCIO, M. L., F. NÓBREGA, P. VIEIRA, L. BONIFÁCIO, P. NAVES, E. SOUSA y M. MOTA (2015). First detection of Bursaphelenchus xylophilus associated with Pinus nigra in Portugal and in Europe. Forest Pathology 45(3): 235-238.
JCYL (1901). Proyecto de Ordenación del Monte de Utilidad Pública núm. 105, ‘Pinar Viejo’. Coca (Segovia). Junta de Castilla y León.
KOUTROUMPA, F. A., B. VINCENT, G. ROUX-MORABITO, C. MARTIN y F. LIEUTIER (2008). Fecundity and larval development of Monochamus galloprovincialis (Coleoptera Cerambycidae) in experimental breeding. Annals of Forest Science 65(7).
LANDA, F. (2009). Aplicación de Selvicultura Prosilva en montes privados, aplicación práctica en un monte de pino silvestre. Congresos Forestales.
LINARES, J. C., J. J. CAMARERO y J. A. CARREIRA (2010). Competition modulates the adaptation capacity of forests to climatic stress: insights from recent growth decline and death in relict stands of the Mediterranean fir Abies pinsapo. Journal of Ecology 98(3): 592-603.

213
MAGRAMA. (2015). "Nematodo de la madera del pino." 2015, de http://www.magrama.gob.es/es/agricultura/temas/sanidad-vegetal/nematodo-de-la-madera-del-pino/#.
MILLER, M. C. (1984). Mortality contribution of insect natural enemies to successive generations of Ips calligraphus (Germar)(Coleoptera, Scolytidae) in loblolly pine. Zeitschrift für Angewandte Entomologie 98(1‐5): 495-500.
MILLER, M. C. (1985). The effect of Monochamus titillator (F.)(Col., Cerambycidae) foraging on the emergence of Ips calligraphus (Germ.)(Col., Scolytidae) insect associates. Zeitschrift für Angewandte Entomologie 100(1‐5): 189-197.
MOTA, M. M., H. BRAASCH, M. A. BRAVO, A. C. PENAS, W. BURGERMEISTER, K. METGE y E. SOUSA (1999). First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1: 727-734.
NAVES, P., S. CAMACHO, E. DE SOUSA y J. QUARTAU (2007). Transmission of the pine wood nematode Bursaphelenchus xylophilus through feeding activity of Monochamus galloprovincialis (Col., Cerambycidae). Journal of Applied Entomology 131(1): 21-25.
NAVES, P. M. y E. M. DE SOUSA (2009). Threshold temperatures and degree-day estimates for development of post-dormancy larvae of Monochamus galloprovincialis (Coleoptera: Cerambycidae). Journal of Pest Science 82(1): 1-6.
PAJARES, J. A., G. ALVAREZ, F. IBEAS, D. GALLEGO, D. R. HALL y D. I. FARMAN (2010). Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. Journal of Chemical Ecology 36(6): 570-583.
PAJARES, J. A., F. IBEAS, J. J. DIEZ y D. GALLEGO (2004). Attractive responses by Monochamus galloprovincialis (Col., Cerambycidae) to host and bark beetle semiochemicals. Journal of Applied Entomology 128(9-10): 633-638.
PARK, C., D. KIM, S. LEE, Y. MOON, Y. CHUNG y D. S. KIM (2014). A forecasting model for the adult emergence of overwintered Monochamus alternatus (Coleoptera: Cerambycidae) larvae based on degree-days in Korea. Applied Entomology and Zoology 49(1): 35-42.
PASCUAL, A., P. SABÍN, A. FERNÁNDEZ, F. BRAVO, C. ORDÓÑEZ, F. RODRÍGUEZ y F. SOSTENIBLE (2013). los señalamientos forestales: una herramienta imprescindible para mejorar nuestros bosques. Foresta(57): 54-57.
RASSATI, D., M. FACCOLI, L. MARINI, R. A. HAACK, A. BATTISTI y E. P. TOFFOLO (2015a). Exploring the role of wood waste landfills in early detection of non-native wood-boring beetles. Journal of Pest Science: 1-10.
RASSATI, D., M. FACCOLI, E. PETRUCCO TOFFOLO, A. BATTISTI y L. MARINI (2015b). Improving the early detection of alien wood‐boring beetles in ports and surrounding forests. Journal of Applied Ecology 52(1): 50-58.
RASSATI, D., E. P. TOFFOLO, A. ROQUES, A. BATTISTI y M. FACCOLI (2014). Trapping wood boring beetles in Italian ports: a pilot study. Journal of Pest Science 87(1): 61-69.
ROBERTSON, L., S. C. ARCOS, M. ESCUER, R. S. MERINO, G. ESPARRAGO, A. ABELLERA y A. NAVAS (2011). Incidence of the pinewood nematode Bursaphelenchus xylophlius Steiner & Buhrer, 1934 (Nickle, 1970) in Spain. Nematology 13: 755-757.
SABÍN GALÁN, P. (2013). Retos y oportunidades de la gestión bajo criterios de Selvicultura Próxima a la Naturaleza en el norte peninsular. Congresos-CARGA FINAL.
SABÍN GALÁN, P., M. ZURITA LAINA, T. VILLA y P. RODRIGUEZ-NORIEGA (2013). Redacción de proyectos de ordenación bajo criterios de selvicultura próxima a la naturaleza. Congresos Forestales.
SANCHEZ-HUSILLOS, E., I. ETXEBESTE y J. PAJARES (2015a). Effectiveness of mass trapping in the reduction of Monochamus galloprovincialis Olivier (Col.: Cerambycidae) populations. Journal of Applied Entomology.

214
SANCHEZ-HUSILLOS, E., I. ETXEBESTE y J. PAJARES (2015b). Physiological development and dispersal ability of newly emerged Monochamus galloprovincialis (Coleoptera: Cerambycidae). Entomologia Experimentalis Et Applicata: Enviado.
SEYBOLD, S. J., J. C. LEE, A. LUXOVA, S. M. HAMUD, P. JIROŠ y R. L. PENROSE (2006). Chemical ecology of bark beetles in California’s urban forests. I. CITRUS AND BIOLOGICAL CONTROL. YESTERDAY, TODAY, TOMORROW: 87.
SINCLAIR, B. J., C. M. WILLIAMS y J. S. TERBLANCHE (2012). Variation in Thermal Performance among Insect Populations*. Physiological and Biochemical Zoology 85(6): 594-606.
SOLIMAN, T., M. C. MOURITS, W. VAN DER WERF, G. M. HENGEVELD, C. ROBINET y A. G. O. LANSINK (2012). Framework for modelling economic impacts of invasive species, applied to pine wood nematode in Europe.
SOUSA, E., M. A. BRAVO, J. PIRES, P. NAVES, A. C. PENAS, L. BONIFACIO y M. M. MOTA (2001). Bursaphelenchus xylophilus (Nematoda; Aphelenchoididae) associated with Monochamus galloprovincialis (Coleoptera; Cerambycidae) in Portugal. Nematology 3: 89-91.
TOGASHI, K. y N. SHIGESADA (2006). Spread of the pinewood nematode vectored by the Japanese pine sawyer: modeling and analytical approaches. Population Ecology 48(4): 271-283.
TORRES-VILA, L. M., C. ZUGASTI, J. M. DE-JUAN, M. J. OLIVA, C. MONTERO, F. J. MENDIOLA, Y. CONEJO, Á. SÁNCHEZ, F. FERNÁNDEZ, F. PONCE y G. ESPÁRRAGO (2015). Mark-recapture of Monochamus galloprovincialis with semiochemical-baited traps: population density, attraction distance, flight behaviour and mass trapping efficiency. Forestry 88(2): 224-236.
VICENTE, C., M. ESPADA, P. VIEIRA y M. MOTA (2012). Pine Wilt Disease: a threat to European forestry. European Journal of Plant Pathology 133(1): 89-99.
WYCKOFF, P. H. y J. S. CLARK (2002). The relationship between growth and mortality for seven co‐occurring tree species in the southern Appalachian Mountains. Journal of Ecology 90(4): 604-615.
YOSHIMURA, A., K. KAWASAKI, F. TAKASU, K. TOGASHI, K. FUTAI y N. SHIGESADA (1999). Modeling the spread of pine wilt disease caused by nematodes with pine sawyers as vector. Ecology 80(5): 1691-1702.
ZAMORA, P., V. RODRÍGUEZ, F. RENEDO, A. V. SANZ, J. C. DOMÍNGUEZ, G. PÉREZ-ESCOLAR, J. MIRANDA, B. ÁLVAREZ, A. GONZÁLEZ-CASAS, E. MAYOR, M. DUEÑAS, A. MIRAVALLES, A. NAVAS, L. ROBERTSON y A. B. MARTÍN (2015). First Report of Bursaphelenchus xylophilus Causing Pine Wilt Disease on Pinus radiata in Spain. Plant Disease: PDIS-03-15-0252-PDN.

215
Capítulo 7 Conclusiones
Bloque A. Fenología de M. galloprovincialis.
o A.1 Fenología de la emergencia de M. galloprovincialis. La emergencia de M. galloprovincialis mostró una alta variabilidad tanto en la
ocurrencia temporal como en los requerimientos térmicos a lo largo de la península ibérica.
Los periodos de emergencia están desfasados en la península ibérica, de modo que en las regiones más cálidas son un mes más tempranas que en las zonas más templadas.
Los requerimientos térmicos para alcanzar la emergencia fueron menores que los registrados y previstos en la península de Setúbal en Portugal, por lo que el modelo predictivo planteado para esta región no puede ser extrapolado al resto de la península ibérica.
Se detectaron indicios de que la diferencia en la acumulación de calor durante el otoño puede afectar el grado de maduración de las larvas invernantes, y a su posterior patrón temporal de emergencia que a su vez también puede estar influido por la heterogeneidad medioambiental posterior al paso del invierno.
Monochamus galloprovincialis mostró comportamientos de protandria en sus emergencias.
A.2 Fenología del vuelo de M. galloprovincialis Monochamus galloprovincialis aparece hasta alturas próximas a los 1900
msnm en el Sistema Central. Altitud y la variable correlacionada temperatura modulan las capturas y el
periodo de vuelo de M. galloprovincialis, de modo que el insecto se hace más raro y su periodo de vuelo es más corto a partir de ciertas altitudes.
El inicio del periodo de vuelo está correlacionado con las temperaturas medias diarias, de manera que se inicia cuando se superan los 14ºC en los meses de mayo o junio.
El perfil de la curva de vuelo de M. galloprovincialis es unimodal en las zonas frías del Sistema Central, mientras que es bimodal en zonas más cálidas del resto de la península.
Se propone la existencia de un límite superior de temperatura que limita el vuelo del insecto en fechas centrales del verano, causante de la forma bimodal de las curvas de vuelo en zonas cálidas.
Existen cambios en la proporción de sexos de las capturas a lo largo del periodo de vuelo. El sesgo inicial favorable a las hembras se convierte en

216
favorable a los machos al final del periodo, lo cual podría explicarse por el progresivo agotamiento del vigor reproductivo de las hembras o por el aumento de la capacidad de respuesta a la feromona por parte de los machos al final del periodo como estrategia reproductiva.
El máximo vuelo de M. galloprovincialis ocurre en las horas más cálidas del día, lo que contradice los hábitos crepusculares citados anteriormente.
o A.3 Comunidad de coleópteros saproxílicos asociada a M. galloprovincialis Las redes de trampas vigía tienen buen potencial para la detección temprana de
organismos exóticos, además de permitir la caracterización y evaluación de las comunidades de entomofauna útil, representando una importante herramienta en la gestión integrada de plagas.
Monochamus galloprovincialis es una especie ubicua, presente en toda la variabilidad medioambiental de los pinares del este y sudeste ibérico, sin que sea posible asociarla significativamente a ninguna comunidad de coleópteros saproxílicos concreta.
La comunidad de coleópteros saproxílicos de los pinares orientales de la península ibérica mostró ser muy homogénea respecto a la variación ambiental de las zonas de muestreo, y M. galloprovincialis desempeña el mismo rol en ellas.
El análisis de la estructura de las comunidades de coleópteros saproxílicos mediante el uso de redes de interacciones demuestra ser una herramienta útil para los estudios ecosistémicos. Dichos análisis mostraron también la relevancia ecológica de M. galloprovincialis al actuar de conector entre los diferentes niveles tróficos.
Se detectaron indicios que apoyan la existencia de una interacción entre el escolítido O. erosus y M. galloprovincialis basada en la coincidencia fenológica y corológica de las dos especies y los roles que ambas desempeñan en los ecosistemas en los que están presentes.
Bloque B. Población y dispersión de M. galloprovincialis
o Se confirma que M. galloprovincialis no responde en campo a los atrayentes cairomonales y feromonales utilizados hasta 8 días después de la emergencia.
o El tamaño de las poblaciones de M. galloprovincialis parece estar relacionado con la fragmentación y tamaño de las manchas forestales, con su naturalización, y con la afección previa de incendios forestales pocos años antes.
o Los niveles de captura de M. galloprovincialis en trampas cebadas está positivamente relacionado, aunque de forma no lineal, con el tamaño estimado de la población.
o No se ha podido demostrar la influencia clara de la dirección e intensidad de los vientos dominantes en la dispersión de M. galloprovincialis a escala de paisaje, con registros de vuelo a larga distancia y de larga duración.
o Existe una correlación directa entre la estructura de la masa forestal de material hospedante y la captura de M. galloprovincialis.
o Las capturas de M. galloprovincialis está también relacionada con la ubicación de las trampas, optimizándose en posiciones de borde de la masa y en zonas visibles.

217
o Se detecta una resistencia por parte de adultos en vuelo de M. galloprovincialis a no abandonar las manchas de bosque en paisajes de mosaico agroforestal, de modo que aumentan las capturas en el borde de las masas, pese a la capacidad de vuelo entre manchas forestales
o Monochamus galloprovincialis posee una alta capacidad de dispersión en estado inmaduro y es capaz de recorrer al menos 3 km sin alimentarse, en busca del alimento necesario para su maduración, si no lo encuentra en el entorno cercano.
o Monochamus galloprovincialis posee suficiente capacidad de dispersión a larga distancia como para recorrer una media de al menos 1500 m/día , llegando a alcanzar excepcionalmente distancias máximas de 13600 m o de 22100 m.

218
Anejos

Anejos Anejo 3.1 Modelización de la variación de la temperatura en función de la altitud en Valsaín (Segovia) y Cercedilla (Madrid)
(Intercept) msnm sin(((2 * pi)/24) * id) cos(((2 * pi)/24) * id) 12.489.414 -0.007521 -0.283884 -0.202698

Anejo 3.2 Frecuencia de capturas en función de las variables altura sobre el nivel del mar, orientación y cobertura vegetal.

Anejo 3.3 Distribución temporal de las capturas de M. galloprovincialis en Guadarrama en los años 2011 y 2012 (curvas de vuelo) entre los 1363 m y los 1835 m dobre el nivel del mar.
Año 2011
Año 2012

Anexo 3.4 Distribución de capturas de M. galloprovincialis en Valsaín y Cercedilla (años 2011, 2012, 2013) en función de la altitud. A la representación se le ha añadido una línea de tendencia basada en un MLG de tipo logístico.
Anejo 3.5 Efectos fijos del modelo mínimo ajustado de la distribución de capturas de M. galloprovincialis en Valsaín y Cercedilla (años 2011, 2012, 2013) en función de la altitud
Fixed effects Estimate Std. Error z value Pr(>|z|)
(Intercept) 309.744 0.14266 21.712 < 2e-16
poly(Lmsnm, 2)1 -413.421 0.28104 -14.710 < 2e-16
poly(Lmsnm, 2)2 -254.984 0.26814 -9.509 < 2e-16
Asp -0.67174 0.08276 -8.117 4.78e-16

Anexo 3.6 Curvas de vuelo acumuladas para el año 2012, utilizando las trampas organizadas según el rango de altitud al que se ubicaron, y ajustadas a un modelo de regresión logística binomial.
Coefficients: Estimate Std. Error z value Pr(>|z|)
(Intercept) -1.763.106 265.146 -6.650 2.94e-11
JDate 0.08568 0.01321 6.487 8.78e-11 Anejo 3.7 Porcentaje de capturas acumuladas por fecha y altura para el año 2012 en Valsaín y Cercedilla
1250 1350 1450 1550 1650 1750 1850 2012-06-10 0.0 0.0 0.5 0.0 0.0 0.0 0.0 2012-06-18 5.9 0.4 1.5 1.2 2.5 0.0 0.0 2012-07-02 14.7 20.7 35.7 12.9 21.4 4.1 16.7 2012-07-12 20.6 44.7 44.6 18.7 32.6 16.8 20.8 2012-08-06 41.2 88.1 84.0 66.1 80.5 85.2 77.1 2012-09-03 73.5 98.8 94.5 88.2 93.0 94.6 100.0 2012-10-05 97.1 100.0 99.5 98.9 100.0 100.0 100.0 2012-10-29 100.0 100.0 100.0 100.0 100.0 100.0 100.0

Anejo 3.8 Duración del periodo de vuelo en función de los rangos altitudinales. Año 2012
Anejo 3.9 Curvas de vuelo acumuladas para el año 2011 por trampa en función de la altitud de colocación de las mismas.

Anexo 3.10 Curvas de capturas acumuladas para las dos laderas del área de estudio (ladera sur de Cercedilla y ladera norte de Valsaín) en el año 2011

Anejo 4.1. Listado de órdenes, familias y especies de la comunidad de insectos saproxílicos considerada. Se indica también la tipología funcional asignada, así como la abundancia total de ejemplares capturados.
Orden Familia Especie Código Tipología Abundancia Coleoptera Alleculidae Heliotaurus ruficollis Helruf Comensal 62 Isomira antennata Isoant Comensal 2 Isomira hispanica Isohis Comensal 1 Proctenius luteus Prolut Comensal 1 Anobiidae Ernobius lucidus Ernluc Xilofago 1 Ernobius rufus Ernruf Xilofago 2 Aphodidae Cheirodes submetalica Chesub Comensal 1 Pleurophorus caeseus Plecae Comensal 3 Attelabidae Lasiorhynchites oeruleocephalus Lasoer Defoliador 1 Bostrichidae Bosthychus capuccinus Boscap Xilofago 2 Scobicia chevrieri Scoche Xilofago 14 Xyloperta picea Xylpic Xilofago 35 Xyloperta praetusa Xylpra Xilofago 8 Buprestidae Acmaeodera rubromaculata segurensis Acmrub Facultativo 10 Acmaeoderella cyanipennis Acacya Facultativo 1 Acmaeoderella moroderi Acamor Facultativo 10 Acmaodera degener Acmdeg Facultativo 2 Acmaoderea bipunctata Acmbip Facultativo 136 Acmaoderella adpsersula Acmadp Facultativo 1 Anthaxia carmen Antcar Facultativo 30 Anthaxia funerula Antfun Facultativo 36 Anthaxia lencinai Antlen Facultativo 11 Anthaxia millefiori Antmil Facultativo 9 Anthaxia nigritula Antnig Facultativo 20 Anthaxia parallela Antpar Facultativo 1 Anthaxia rugicollis Antrug Facultativo 38 Anthaxia scutellaris Antscu Facultativo 1 Buprestis nonenmaculata Bupnon Facultativo 23 Buprestis octoguttata Bupoct Facultativo 3 Chalcophora mariana Calmar Facultativo 2 Phaenops cyaena Phacya Facultativo 153 Carabidae Amara similata Amasim Depredador 2 Aphaenops cuaenops Aphcua Depredador 4 Cryptophonus tenebrosus Cryten Depredador 1 Dromius agilis Droagi Depredador 2 Dromius chovauti Drocho Depredador 21 Lebia cyanocephalus Lebcya Depredador 3 Mesolestes scapularis Messca Depredador 1 Microlestes abeillei Micneg Depredador 1 Microlestes corticalis Miccor Depredador 2 Microlestes reitteri Micrei Depredador 4 Paradromius linearis Parlin Depredador 1 Singilis alternans Sinalt Depredador 3 Syntomus fuscomaculatus Synfus Depredador 9 Cerambycidae Acanthocinus aedilis Acaaed Facultativo 3 Acanthocinus griseus Acagri Facultativo 88 Acanthocinus hispanicus Acahis Facultativo 23 Arrophalus syriacus Arrsyr Facultativo 484 Chlorophorus trifasciatus Chltri Facultativo 25 Clhorophorus varius Chlvar Facultativo 2 Cribroleptura otini Crioti Facultativo 9 Deilus fugax Deifug Facultativo 10 Monochamus galloprovincialis Mongal Facultativo 765 Natrius brevipennis Natbre Xilofago 1 Oxipleurus nodieri Oxinor Facultativo 531 Penichroa timida Pentim Facultativo 2 Phymatodes testaceus Phytes Facultativo 2 Pogonocherus perroudi Pogper Facultativo 85 Purpuricenus ferrugineus Purfer Facultativo 38 Ragium inquisitor Ragium Facultativo 39 Semanotus laurasi Semlar Facultativo 1 Spondylis buprestoides Spobup Facultativo 55 Stromatium unicolor Struni Facultativo 1 Trichoferus fasciculatus Trifas Facultativo 1 Trichoferus magnani Trimag Facultativo 1 Vesperus xatarti Vesxat Comensal 29 Cetonidae Netocia cuprea Netcup Comensal 6 Netocia morio Netmor Comensal 12 Netocia oblonga Netobl Comensal 4 Protaetia morio Promor Comensal 3 Protaetia oblonga Proobl Comensal 6 Tropinota squalida Trosqu Comensal 10 Cleridae Allonyx quadrimaculata Aloqua Depredador 48 Clerus mutiliarius Clemut Depredador 2 Opilio domesticus Opidom Depredador 40 Thanasimus femoralis Tamfem Depredador 160 Thanasimus formicarius Tanfor Depredador 1394 Tricodes leucopsideus Trcleu Depredador 4 Coccinelidae Coccinella septempunctata Cocsep Depredador 1 Thea turiphera Thetur Depredador 1 Colydiidae Aulonium ruficorne Aloruf Depredador 6062 Cucujidae Ahasverus advena Ahaadv Comensal 2 Curculionidae Magdalis rufa Magruf Facultativo 20 Pachyrhinus squamosus Pacsqu Defoliador 16 Ryncholus sp Rynsp Escolitido 189 Dermestidae Orphilus beali Orpbea Comensal 16 Orphilus niger Orpnig Comensal 1 Elateridae Cardiophorus signatus Carsig Facultativo 22 Lacon punctatus Lacon Facultativo 45 Eucmenidae Anelastidius feisthameli Anefei Rizofago 2 Histeridae Cylister elongatus Cylelo Depredador 10

Orden Familia Especie Código Tipología Abundancia Cylister filiformis Cylfil Depredador 59 Onthophilus beali Ontbea Depredador 1 Onthophilus niger Ontnig Depredador 2 Plegaderus otti Pleott Depredador 2 Sternocoelis acutangulus Steacu Depredador 1 Teretrius parasitica Terpar Depredador 16 Malachiidae Attalus pictus Attpic Depredador 1 Melodonthidae Amphimallon korbi Ampkor Comensal 1 Euserica segurana Eusseg Comensal 2 Rhizotrogus cicatricosus Rhicic Comensal 3 Rhizotrogus coiffaiti Rhicoi Comensal 2 Rhizotrogus patruellis Rhipat Comensal 8 Mycetophagidae Litargus connexus Litcon Comensal 3 Thyphaea stercorea Thyste Comensal 2 Mycteridae Mycterus curculionides Myccur Comensal 36 Nitidulidae Carpophilus hemiptera Carhem Comensal 1 Pityophagus laevior Pytio Depredador 178 Oedemeridae Chrisantia amata Chrham Xilofago 6 Chrisantia solieri Chrsol Xilofago 9 Nacerdes raymondi Nacray Xilofago 43 Oedemera basalis Oedbas Xilofago 1 Spharedrus lencinae Spalen Xilofago 2 Priocenidae Lobonix aeneus Lobaen Xilofago 93 Salpingidae Sphaeriestes castaneus Sphcas Depredador 2 Scolytidae Carpoborus mínimus Carmin Escolitido 3 Carpoborus pini Carpin Escolitido 122 Hylastes ater Hyaate Escolitido 92 Hylastes attenuatus Hyaatt Escolitido 288 Hylastes linearis Hyalin Escolitido 907 Hylurgus ligniperda Hyllig Escolitido 1285 Hylurgus micklizti Hylmic Escolitido 27780 Ips acuminatus Ipsacu Escolitido 1 Ips mannsfeldi Ipsman Escolitido 29 Ips sexdentatus Ipssex Escolitido 1032 Orthotomicus erosus Ipsero Escolitido 21628 Orthotomicus longicollis Ortlon Escolitido 5 Phloeosinus tujae Photuj Escolitido 1 Pytiogenes calcaratus Pytcal Escolitido 92 Pytioptorus pubescens Pytpub Escolitido 1 Tomicus destruens Tomdes Escolitido 254 Xileborinus xaxeseni Xilxas Ambrosia 163 Xyleborus eurigraphus Xyleur Ambrosia 57 Tenebrionidae Corticeus pini Corpin Depredador 2637 Corticeus rufitorax Corruf Depredador 1 Lagria grenieri Laggre Comensal 2 Nalassus calpensis Nalcal Comensal 21 Nesotes carbo Nescar Comensal 2 Trogossitidae Nemozoma elongatum Nemelo Depredador 1 Tennoscheila coerulea Tencoe Depredador 1322 Hymenoptera Braconidae Sirex juvencus Sirjuv Xilofago 89

Anejo 4.2. Grafos que representan la red de interacciones para cada parcela de la Red ESFP de la región de Murcia, utilizando el algoritmo de Kamada-Kawai en modo libre. Los grafos de las ESFP números 2 y 15 han sido incluidas como ejemplos en el tecto de la memoria .Los colores representan los tipos funcionales definidos en el Anejo 4.1. El tamaño del nodo es directamente proporcional a la abundancia de la especie en cada parcela de muestreo. En los casos de extrema desproporción en las abundancias de algunas especies, no se representó dicho vector de abundancias, teniendo entonces todos los nodos el mismo tamaño. El tamaño de la punta de la flecha en los trazos es directamente proporcional a la intensidad de la interacción. Relación de colores con los que se representa el vector de tipos funcionales en los grafos: Excolítido (rosa), Depredador (Azul), Depredador facultativo (naranja), Ambrosía (amarillo), Xilófago (Morado), Comensal (verde), Defoliador (rojo).

Anejo 4.2.1. Grafo de la red de interacciones de la ESFP 1
Anejo 4.2.2. Grafo de la red de interacciones de la ESFP 2
Anejo 4.2.3. Grafo de la red de interacciones de la ESFP 3 Anejo 4.2.4. Grafo de la red de interacciones de la ESFP 4

Anejo 4.2.5. Grafo de la red de interacciones de la ESFP 5
Anejo 4.2.6. Grafo de la red de interacciones de la ESFP 6
Anejo 4.2.7. Grafo de la red de interacciones de la ESFP 7 Anejo 4.2.8. Grafo de la red de interacciones de la ESFP 8

Anejo 4.2.9. Grafo de la red de interacciones de la ESFP 9
Anejo 4.2.10. Grafo de la red de interacciones de la ESFP 10
Anejo 4.2.11. Grafo de la red de interacciones de la ESFP 11 Anejo 4.2.12. Grafo de la red de interacciones de la ESFP 12

Anejo 4.2.13. Grafo de la red de interacciones de la ESFP 13
Anejo 4.2.14. Grafo de la red de interacciones de la ESFP 14
Anejo 4.2.15. Grafo de la red de interacciones de la ESFP 15 Anejo 4.2.16. Grafo de la red de interacciones de la ESFP 16

Anejo 4.2.17. Grafo de la red de interacciones de la ESFP 17
Anejo 4.2.18. Grafo de la red de interacciones de la ESFP 18
Anejo 4.2.19. Grafo de la red de interacciones de la ESFP 19 Anejo 4.2.20. Grafo de la red de interacciones de la ESFP 20

Anejo 4.3. Características de las parcelas de la Red ESFP de la Región de Murcia.
Estación
X Y Alt Cod. Denominacion monte T.M. Comarca Alt (m) Alt (M) max (m)
Superf. (ha)
Exposición
Especie princpal
Distribución
Origen Ho FCC tot FCC arb Desarrollo Sp. arbórea 2
ESFP1 611686 4257801 823 MU95 Sierra del Carche Jumilla Altiplano 467 1362 3754 Umbria P. halepensis Uniforme Natural 10 100 100 Latizal Pistacia lentiscus ESFP2 655038 4167637 459 MU128 Sierra del Algarrobo Mazarr
ón Litoral 104 699 2086 Umbria P. halepensis Uniforme Repob 5 50 50 Latizal Pistacia lentiscus
ESFP3 661507 4196569 407 MU174 El Valle y Carrasco Murcia Centro 51 607 5228 Mixta P. halepensis Uniforme Repob 4 90 90 Latizal - ESFP4 636520 4193585 518 MU28 Sierra Espuña de Alhama Alhama
de Murcia
Espuña 256 1440 3943 Mixta P. halepensis Uniforme Repob 6 100 69 Latizal - ESFP5 626321 4192278 1436 MU29 Sierra Espuña de Totana Totana Espuña 354 1580 6070 Umbria P.nigra Uniforme Repob 10 90 70 Fustal Quercus ilex ESFP6 625727 4191067 1288 MU29 Sierra Espuña de Totana Totana Espuña 354 1580 6070 Solana P. pinaster Uniforme Repob 10 90 90 Fustal - ESFP7 618199 4189861 831 MU85 Sierra y Llano de las
Cabras Lorca Lorca 607 881 558 Solana P. halepensis Bosquetes Repob 5 80 80 - -
ESFP8 612691 4191795 903 MU589 Los Alagueces III Lorca Lorca 707 997 383 Mixta P. halepensis Bosquetes Repob 4 60 60 Latizal Pinus halepensis ESFP9 663459 4193266 544 MU174 El Valle y Carrascoy Murcia Centro 51 607 5228 Umbria P. halepensis Uniforme Repob 10 90 90 Latizal -
ESFP10 594585 4191714 789 MU163 Reverte y Fuente Atocha Lorca Lorca 704 1071 920 Mixta P. halepensis Uniforme Repob 4 80 70 Latizal - ESFP11 595475 4205749 883 MU6 Las Cabezuelas Caravac
a de la Cruz
Noroeste 781 976 357 Mixta P. halepensis Uniforme Repob 8 70 70 Latizal - ESFP12 593600 4216778 864 MU12 Los Enebrales Caravac
a de la Cruz
Noroeste 759 1252 426 Mixta P. halepensis Uniforme Natural 5 50 50 Latizal - ESFP13 591063 4221047 1154 MU23 Solana de Trigueros Caravac
a de la Cruz
Noroeste 1029 1310 87 Umbria P. pinaster Uniforme Natural 10 100 80 Fustal Pinus halepensis ESFP14 584792 4227576 1044 MU10 Loma de Enmedio Moratal
la Noroeste 938 1350 780 Umbria P. halepensis Uniforme Repob 4 70 70 Latizal Juniperus oxycedrus
ESFP15 572332 4224276 1226 MU606 Casa Manta Moratalla
Noroeste 1127 1611 425 Mixta P. nigra Uniforme Natural 6 90 80 Fustal Pinus pinaster ESFP16 646318 4227426 349 MU41 La Navela y el Solan Abarán Centro 131 550 911 Mixta P. halepensis Uniforme Repob 4 50 40 Latizal Rhamnus lycioides ESFP17 649031 4238091 509 MU40 Sierra de la Pila Fortuna Centro 287 1057 4436 Umbria P. halepensis Uniforme Repob 5 80 60 Latizal - ESFP18 671053 4263939 896 MU653 Coto Salinas Yecla Altiplano 640 1233 1252 Umbria P. halepensis Uniforme Natural 8 100 100 Latizal - ESFP19 647053 4273793 927 MU87 Gavilanes Jumilla Altiplano 733 944 1093 Umbria P. halepensis Uniforme Repob 5 80 70 Latizal Pistacia lentiscus ESFP20 642497 4277968 852 MU107 Cenajo de Peñas Blancas Jumilla Altiplano 750 950 1039 Mixta P. halepensis Bosquetes Repob 3 70 60 Latizal Pistacia lentiscus

Anejo 4.4. Características de las parcelas de la Red Muffet de la Comunitat Valenciana.
Id Sp principal X Y Nombre Localidad Comarca Provincia Alt Alt m Alt M Area Estado Origen Ho FFCtot FFCarb Ph Pn Ppstr Pp Ps Qi Qc Euc Jox Jth Jco
Muf1 P.pinea 706008 4219823 Dunas de Guardamar Guardamar La Vega Baja Alicante 11 3 35 106.46 Regular Reforest 4 80 35 0.6 0 0 0.4 0 0 0 0 0 0 0
Muf2 P.halepensis 694260 4239891 Sierra Baja y Lomas Crevillent El Baix Vinalopò Alicante 296 170 395 745.7 Regular Reforest 1 60 20 0.7 0 0 0 0 0 0 0.3 0 0 0
Muf3 P.pinea 698630 4280894 Conjunto de montes de Biar Biar L' Alt Vinalopó Alicante 850 840 1200 522.32 Good Regular 5 85 75 0.7 0 0 0.3 0 0 0 0 0 0 0
Muf4 P.halepensis 715449 4294852 Umbría o Mariola Agres El Comtat Alicante 791 620 1250 478.19 Regular Reforest 7 70 55 0.8 0 0 0 0 0.1 0 0 0 0.1 0
Muf5 P.halepensis 730896 4358246 Devesa de L' Albufera Saler Valencia Valencia 2 0 10 33.64 Good Regular 10 60 40 1 0 0 0 0 0 0 0 0 0 0
Muf6 P.sylvestris 661762 4437505 Puebla de San Miguel Puebla de S. Miguel Rincón de Ademuz Valencia 1733 1650 1836 276.5 Good Regular 10 85 60 0 0.2 0 0 0.7 0 0 0 0 0 0.1
Muf7 P.nigra 659777 4432048 Puebla de San Miguel Puebla de S. Miguel Rincón de Ademuz Valencia 1297 1400 1700 579.83 Good Regular 10 90 85 0 0.7 0 0 0 0 0 0 0 0.2 0.1
Muf8 P.pinaster 653710 4405585 Solana y Campos Herrerias Sinarcas La Plana Utiel - Requena Valencia 877 900 912 702.7 Good Regular 10 85 60 0.4 0 0.5 0 0 0 0 0 0.1 0 0
Muf9 P.pinaster 758753 4443399 Les Santes Pobla Tornesa La Plana Alta Castellón 360 300 500 59.18 Regular Mosaico 8 85 65 0.3 0 0.6 0 0 0 0 0 0.1 0 0
Muf10 P.sylvestris 725750 4457593 San Juan de Peñagolosa Vistabella del Maestrat L'Alcalatén Castellón 1524 1280 1550 358.11 Regular Regular 14 80 65 0 0.5 0 0 0.5 0 0 0 0 0 0
Muf11 P.nigra 729695 4469449 Bovalar y Sabinar Vistabella del Maestrat L'Alcalatén Castellón 1131 1100 1250 218.32 Regular Regular 10 80 50 0 1 0 0 0 0 0 0 0 0 0
Muf12 P.pinaster 724929 4459895 San Juan de Peñagolosa Vistabella del Maestrat L'Alcalatén Castellón 1471 1200 1485 81.83 Regular Regular 20 95 85 0 0 0.75 0 0.25 0 0 0 0 0 0
Muf13 P.nigra 737989 4281927 Serrella y Aitana Confrides La Marina Baixa Alicante 1343 1340 1510 35.78 Good Reforest 6 60 15 0 0 0.7 0 0.3 0 0 0 0 0 0
Muf14 P.halepensis 710475 4416944 La Muela, Roquetillo, Feliciano
Jérica El Alto Palancia Castellón 498 490 669 415.59 Regular Reforesta 8 80 70 0.7 0 0 0 0 0 0.3 0 0 0 0
Muf16 P.nigra 741803 4490223 Carrascals Morella Els Ports Castellón 1005 900 1090 515.52 Bad Mosaico 12 75 65 0 0.2 0 0 0.05 0.7 0 0 0.05 0 0

Anejo 4.5. Listado de especies capturadas en la Red MUFFET en el año 2013. Nivel taxonómico y regímen trófico (RT) de las especies de coleópteros saproxílicos. Símbolos de los regímenes tróficos (RT): xilófaga (X), xilófaga/micófaga (X/M), micófaga (M), micófaga/polinívora (M/P), polinívora (P), carnívora/polinívora (C/P), carnívora (C), carnívora/fitófaga (C/F), fitófaga (F), detritívora/fitófaga (D/F), detritívora (D), saprófaga (S), necrófaga (N), lignívora (L), rizófaga (R) y ectoparásita de larvas de otros coleópteros (EP)
FAMILIA ESPECIE RT
BOSTRICHIDAE Apate monachus Fabricius, 1775 X Bostrichus capucinus (Linnaeus, 1758) Sinoxylon sexdentatum (Olivier, 1790) X Scobicia chevrieri (Villa & Villa, 1835) X Scobicia pustulata (Fabricius, 1801) X Xyloperthella picea (Olivier, 1790) X Xylopertha praeusta (Germar, 1817) DERMESTIDAE Attagenus sp. M Dermestes (Dermestinus) undulatus. Brahm, 1790 N Dermestes sp. N Anthrenus goliath Saulcy, 1868 M/P Anthrenus pimpinellae (Fabricius, 1775) M/P Anthrenus (Florilinus) fuscus Olivier, 1789 M/P Anthrenus sp. M/P Ctesias sp. M Globicornis (Hadrotoma) corticalis (Eichhoff, 1863) M Globicornis sp. M Megatoma undata (Linnaeus, 1758) M Megatoma sp. M Trogoderma sp. Trogoderma granarium Everts, 1898 M Trogoderma versicolor (Creutzer, 1799) M Orphilus sp. M/P PTINIDAE Anobium punctatum (De Geer, 1774) L Stagetus byrrhoides (Mulsant & Rey, 1861) L Ptinomorphus imperialis (Linnaeus, 1767) F Stagetus elongatus (Mulsant & Rey, 1861) L ANOBIDAE Stagetus pilula (Aube, 1861) L Ernobius parens (Mulsant & Rey, 1863) X Ernobius pini (Sturm, 1837) X Ernobius reflexus (Mulsant & Rey, 1863) X Ernobius rufus (Illiger, 1807) X Ernobius sp. X Dignomus irroratus (Kiesenwetter, 1851) Ptinus (Cyphoderes) bidens Olivier, 1790 D Ptinus (Gynopterus) pyrenaeus Pic, 1897 D Ptinus fur (Linnaeus, 1758) D Ptinus latro Fabricius, 1775 D Ptinus perrini (Reitter, 1884) D Ptinus subpilosus Sturm, 1837 D Ptinus clavipes Panzer, 1792 D Ptinus sp. CARABIDAE Harpalus distinguendus (Duftschmid, 1812) C
Pseudomasoreus canigoulensis (Fairmaire & Laboulbene, 1854) C
Calodromius bifasciatus (Dejean, 1825) C Dromius agilis (Fabricius, 1787) C Dromius chobauti Puel, 1924 C Dromius meridionalis Dejean, 1825 C Microlestes abeillei (Brisout de Barneville, 1885) C Microlestes reitteri Holdhaus, 1912 C Philorhizus crucifer (Lucas, 1846) C Philorhizus notatus (Stephens, 1827) C Syntomus fuscomaculatus (Motschulsky, 1844) C CERAMBYCIDAE Cerambyx cerdo Linnaeus. 1758 X Poecilium glabratum (Charpentier 1825) X Chlorophorus pilosus (Forster 1771) X Nathrius brevipennis (Mulsant 1839) X Acanthocinus aedilis (Linnaeus 1758) X Acanthocinus griseus (Fabricius 1792) X Acanthocinus hispanicus Sama & Schurmann 1980 X Monochamus galloprovincialis (Olivier, 1795) X Pogonocherus decoratus Fairmaire, 1855 X Pogonocherus fasciculatus (De Geer, 1775) X Pogonocherus perroudi Mulsant, 1839 X Pogonocherus sturanii Sama & Schurmann, 1982 X Niphona picticornis Mulsant, 1839 X Paracorymbia stragulata (Germar, 1824) Stenurella bifasciata (Muller, 1776) X Rhagium inquisitor Linnaeus, 1758 X Prionus coriarius (Linnaeus, 1758) X Arhopalus ferus (Mulsant, 1839) X Arhopalus rusticus (Linnaeus, 1758) X Arhopalus sp. Arhopalus syriacus (Reitter, 1895) X Nothorhina muricata (Dalman, 1817) X Oxypleurus nodieri Mulsant, 1839 X Spondylis buprestoides (Linnaeus, 1758) X Vesperus xatarti Mulsant, 1839 R CHRYSOMELIDAE Aphthona sp. F Aplocnemus sp. C/P Longitarsus sp. F Psylliodes hyoscyami (Linnaeus, 1758) F Psylliodes sp. F
Lachnaia pseudobarathraea K. Daniel & J. Daniel, 1898 F
Crioceris macilenta Weise, 1880 F Crioceris paracenthesis (Linnaeus 1767) F Cryptocephalus lineellus Suffrian, 1849 F

Stylosomus (Stylomicrus) minutissimus (Germar, 1824) F
CLERIDAE Allonyx quadrimaculatus (Schaller, 1783) C Clerus mutillarius Fabricius, 1775 C Opilo domesticus (Sturm, 1837) C Thanasimus femoralis (Zetterstedt 1828) C Thanasimus formicarius (Linnaeus 1758) C Trichodes leucopsideus (Olivier, 1795) C Denops sp. DASYTIDAE Mauroania bourgeoisi (Pic, 1894) C/P Mauroania hispana (Kiesenwetter, 1859) C/P Dasytes terminalis Jacquelin Du Val, 1861 C/P Dasytes sp. Aplocnemus brevis (Rosenhauer, 1856) C/P Aplocnemus consobrinus (Rosenhauer, 1856) C/P Aplocnemus limbipennis Kiesenwetter, 1865 C/P MALACHIIDAE Attalus (Abrinus) analis (Panzer 1796) C/P Clanoptilus abdominalis (Fabricius 1798) C/P Haplomalachius (Flabellomalachius) hispanus (Perris, 1865) C/P Malachius lusitanicus Erichson, 1840 C/P PRIONOCERIDAE Lobonyx aeneus (Fabricius, 1787) C/P TROGOSSITIDAE Nemozoma elongatum (Linnaeus, 1761) C Temnoschila coerulea Olivier, 1790 C BOTHRIDERIDAE Oxylaemus cylindricus (Panzer 1796) EP COCCINELLIDAE Rhyzobius chrysomeloides (Herbst, 1792) C Rhyzobius sp. C Anatis ocellata (Linnaeus, 1758) C Coccinella septempunctata Linnaeus, 1758 C Myrrha (Metamyrrha) thuriferae (Sicard, 1923) C Myrrha octodecimguttata (Linnaeus, 1758) C Novius cruentatus (Mulsant, 1850) C Nephus (Bipunctatus) bipunctatus (Kugelann, 1794) C CRYPTOPHAGIDAE Atomaria sp. M Caenoscelis ferruginea (C.R.Sahlberg, 1820) M Cryptophagus sp. M ENDOMYCHIDAE Symbiotes gibberosus (Lucas, 1846) M Triplax lacordairii Crotch, 1870 M LAEMOPHLOEIDAE Cryptolestes ferrugineus (Stephens, 1831) C Lathropus sepicola (Muller, 1821) L LATRIDIIDAE Corticaria sp. M Dienerella sp. L Enicmus sp. L MONOTOMIDAE Rhizophagus depressus (Fabricius, 1792) C Rhizophagus ferrugineus (Paykull, 1800) C Rhizophagus sp NITIDULIDAE Glischrochilus quadripunctatus (Linnaeus, 1758) C Pityophagus laevior Abeille, 1872 C Epuraea guttata (Olivier, 1811) M Epuraea sp. M Meligethes sp. P Ipidia binotata Reitter, 1875 L PHALACRIDAE Olibrus sp. P ANTHRIBIDAE Enedreytes hilaris Fahaeus 1839 X CURCULIONIDAE Ceuthorrynchus sp. F Brachytemnus porcatus (Germar, 1824) X Rhyncolus sculpturatus Waltl, 1839 X Brachyderes confusus Viedma, 1967 X Brachyderes grisescens Fairmaire, 1862 F Brachyderes pubescens Boheman, 1833 F Polydrosus sp. F Baryperthes sp. F Otiorhynchus sp. F Magdalis memnonia (Gyllenhal, 1837) F Magdalis rufa Germar, 1824 F Magdalis sp. F Hylobius (Callirus) abietis (Linnaeus, 1758) X/M Pissodes pini (Linnaeus, 1758) X/M Pityogenes calcaratus (Eichhoff 1878) X/M Pityophthorus buyssoni Reitter, 1901 X/M Pityophthorus lichtensteinii (Ratzeburg, 1837) X/M Pityophthorus pubescens (Marsham, 1802) X/M Pityogenes sp. X/M Crypturgus mediterraneus Eichhoff, 1871 X/M Crypturgus numidicus Ferrari, 1867 X/M Crypturgus sp. X/M Xylocleptes bispinus (Duftschmid, 1825) X/M Hylastes ater (Paykull 1800) X/M Hylastes attenuatus Erichson, 1836 X/M Hylastes linearis Erichson, 1836 X/M Hylastes sp. X/M Chaetoptelius vestitus (Mulsant & Rey, 1860) X/M Ips acuminatus (Gyllenhal, 1827) X/M Ips mannsfeldi (Wachtl, 1879) X/M Ips sexdentatus (Börner, 1776) X/M Orthotomicus erosus (Wollaston, 1857) X/M Carphoborus pini Eichhoff, 1881 X/M Hylurgus ligniperda (Fabricius, 1787) X/M Hylurgus miklitzi Wachtl, 1881 X/M Tomicus destruens (Wollaston, 1865) X/M Tomicus piniperda (Linnaeus, 1758) X/M Xyleborinus saxesenii (Ratzeburg, 1837) X/M Xyleborus dispar (Fabricius, 1792) X/M Xyleborus eurygraphus (Ratzeburg, 1837) X/M BUPRESTIDAE Anthaxia (Haplanthaxia) confusa Gory, 1841 X Anthaxia parallela Gory & Laporte, 1839 X Anthaxia (Melanthaxia) godeti. Gory & Laporte, 1839 X Anthaxia lencinai Arnáiz & Bercedo, 2003 X Anthaxia (Melanthaxia) nigritula Ratzeburg, 1837 X Anthaxia (Melanthaxia) nigrojubata Roubal, 1913 X Anthaxia (Melanthaxia) rugicollis Lucas, 1849 X Anthaxia (Melanthaxia) sepulchralis (Fabricius, 1801) X

Anthaxia (Melanthaxia) sp. X Buprestis novemmaculata Linnaeus, 1758 X Buprestis octoguttata Linnaeus, 1758 X Phaenops cyanea (Fabricius 1775) X
Sphenoptera (Chilostetha) laportei Saunders, 1871 R
Chrysobothris solieri Laporte & Gory, 1839 X Chalcophora mariana (Linnaeus, 1758) X Acmaeodera bipunctata (Olivier, 1790) X CANTHARIDAE Cantharis sp. C/F ELATERIDAE Lacon punctatus (Herbst, 1779) C Cardiophorus sp. C Athous sp. C Elathous platiai Zapata & Sánchez-Ruiz, 2007 C Stenagostus laufferi (Reitter, 1904) C Ampedus sp. C Megapenthes lugens (L. Redtenbacher, 1842) C Megapenthes sp. C Melanotus castanipes (Paykull, 1800) C EUCNEMIDAE Anelastidius Jacquelin du Val, 1863 LAMPYRIDAE Nyctophila reichii (Jacquelin du Val, 1859) C THROSCIDAE Trixagus sp. R HISTERIDAE Plegaderus otti Marseul, 1856 C Teretrius (Neotepetrius) parasita Marseul, 1862 C Paromalus parallelepipedus (Herbst, 1792) C Platylomalus complanatus (Panzer, 1796) C Cylister elongatus (Thunberg, 1787) C Cylister filiformis (Erichson, 1834) C APHODIDAE Aphodius sp. S CETONIIDAE Protaetia (Netocia) morio (Fabricius, 1781) D
Protaetia (Netocia) oblonga (Gory & Percheron, 1833) D/P
Tropinota (Tropinota) squalida (Scopoli, 1783) D/P MELOLONTHIDAE Elaphocera sp. R Amadotrogus patruelis (Reiche, 1862) R Rhizotrogus sp. R Paratriodonta alicantina (Reitter, 1890) R EUCINETIDAE Nycteus meridionalis Laporte de Castelnau, 1836 M Nycteus sp. SCIRTIDAE Cyphon sulcicollis Mulsant & Rey, 1865 F LEIODIDAE Choleva sp. M Colon sp. M Agathidium sp. M Leiodes sp. M SILPHIDAE Nicrophorus interruptus Stephens, 1830 N MELANDRYDAE Abdera quadrifasciata (Curtis, 1829) X MELOIDAE Mylabris quadripunctata (Linnaeus, 1767) P MYCETOPHAGIDAE Berginus tamarisci Wollaston, 1854 M Litargus balteatus LeConte, 1856 M Litargus connexus (Geoffroy, 1785) M Mycetophagus quadripustulatus (Linnaeus, 1761) M Mycetophagus salicis Brisout de Barneville, 1862 M MYCTERIDAE Mycterus curculioides (Fabricius, 1781) M OEDEMERIDAE Chrysanthia reitteri Seidlitz, 1899 P RIPIPHORIDAE Ripidius sp. EP SALPINGIDAE Salpingus planirostris (Fabricius, 1787) C Sphaeriestes castaneus (Panzer, 1796) C Sphaeriestes exsanguis (Abeille de Perrin, 1870) C SCRAPTIDAE Anaspis sp. P Scraptia sp. P TENEBRIONIDAE Hymenalia rufipes (Fabricius, 1792) D/F Hymenorus doublieri (Mulsant, 1851) D/F Isomira hispanica Kiesenwetter, 1870 D/F Isomira sp. D/F Diaperis boleti (Linnaeus, 1758) M Penthaphyllus chrysomeloides Rossi, 1792 M Corticeus pini Panzer, 1799 C Probaticus anthracinus (Germar, 1813) D ZOPHERIDAE Aulonium ruficorne (Olivier, 1790) C

Anejo 4.6 Especies capturadas en la Red MUFFET en el año 2013 con frecuencia de captura mayor a 3 individuos
Especies Frecuencias Ips sexdentatus (Börner 1776) 10578 Hylurgus miklitzi (Wachtl 1881) 7642 Rhizophagus (Eurhizophagus) depressus (Fabricius 1792) 3337 Hylastes attenuatus (Erichson 1836) 2864 Corticeus pini (Panzer 1799) 2559 Thanasimus formicarius (Linnaeus 1758) 2313 Aulonium ruficorne (Olivier 1790) 2029 Orthotomicus erosus (Wollaston 1857) 1657 Hylastes ater (Paykull 1800) 1469 Hylurgus ligniperda (Fabricius 1787) 1346 Temnoschelia coerulea (Olivier 1790) 970 Crypturgus mediterraneus (Eichhoff 1871) 768 Rhizophagus (Rhizophagus) ferrugineus (Paykull 1800) 672 Diaperis boleti (Linnaeus 1758) 658 Thanasimus femoralis (Zetterstedt 1828) 490 Oxypleurus nodieri (Mulsant 1839) 471 Megapenthes lugens (L. Redtenbacher 1842) 455 Rhagium (Rhagium) inquisitor (Linnaeus 1758) 455 Monochamus galloprovincialis (Olivier 1795) 421 Acanthocinus griseus (Fabricius 1792) 405 Spondylis buprestoides (Linnaeus 1758) 335 Pityophagus laevior (Abeille 1872) 314 Pityogenes calcaratus (Eichhoff 1878) 277 Arhopalus syriacus (Reitter 1895) 235 Xyleborinus saxesenii (Ratzeburg 1837) 157 Tomicus destruens (Wollaston 1865) 156 Arhopalus ferus (Mulsant 1839) 145 Arhopalus rusticus (Linnaeus 1758) 140 Ips mannsfeldi (Wachtl 1879) 131 Cryptolestes ferrugineus (Stephens 1831) 100 Paromalus (Paromalus) parallelepipedus (Herbst 1792) 92 Stagetus pilula (Aube 1861) 88 Crypturgus numidicus (Ferrari 1867) 71 Acanthocinus hispanicus (Sama & Schurmann 1980) 70 Ips acuminatus (Gyllenhal 1827) 65 Allonyx quadrimaculatus (Schaller 1783) 64 Xyleborus eurygraphus (Ratzeburg 1837) 61 Pogonocherus perroudi (Mulsant 1839) 57 Mycetophagus (Mycetophagus) quadripustulatus (Linnaeus 1761) 56 Cylister elongatus (Thunberg 1787) 49 Hylurgops palliatus (Gyllenhal 1813) 47 Acanthocinus aedilis (Linnaeus 1758) 45 Penthaphyllus chrysomeloides (Rossi 1792) 45 Hylastes linearis (Erichson 1836) 44 Phaenops cyanea (Fabricius 1775) 38 Litargus (Litargus) connexus (Geoffroy 1785) 37 Brachytemnus porcatus (Germar 1824) 36 Lacon punctatus (Herbst 1779) 36 Lobonyx aeneus (Fabricius 1787) 32 Pityophthorus pubescens (Marsham 1802) 32 Dasytes (Dasytes) terminalis (Jacquelin Du Val 1861) 31 Aplocnemus (Aplocnemus) consobrinus (Rosenhauer 1856) 28 Anthrenus (Florilinus) fuscus (Olivier 1789) 27 Chaetoptelius vestitus (Mulsant & Rey 1860) 20 Hymenorus doublieri (Mulsant 1851) 20 Isomira hispanica (Kiesenwetter 1870) 20 Hylobius (Callirus) abietis (Linnaeus 1758) 18 Ernobius pini (Sturm 1837) 16 Salpingus planirostris (Fabricius 1787) 15 Stenagostus laufferi (Reitter 1904) 15 Scobicia chevrieri (Villa & Villa 1835) 14
Especies Frecuencias Acmaeodera (Palaeotethya) bipunctata (Olivier 1790) 13 Cylister filiformis (Erichson 1834) 13 Rhyzobius chrysomeloides (Herbst 1792) 13 Enedreytes hilaris (Fahaeus 1839) 12 Globicornis (Hadrotoma) corticalis (Eichhoff 1863) 12 Berginus tamarisci (Wollaston 1854) 11 Chrysanthia reitteri (Seidlitz 1899) 11 Ptinus (Cyphoderes) bidens (Olivier 1790) 11 Ptinus (Gynopterus) pyrenaeus (Pic 1897) 10 Xyloperthella picea (Olivier 1790) 10 Brachyderes (Brachyderes) pubescens (Boheman 1833) 8 Dermestes (Dermestinus) undulatus (Brahm 1790) 7 Caenoscelis ferruginea (C.R.Sahlberg 1820) 6 Microlestes reitteri (Holdhaus 1912) 6 Pityophthorus lichtensteinii (Ratzeburg 1837) 6 Ptinus (Ptinus) latro (Fabricius 1775) 6 Tomicus piniperda (Linnaeus 1758) 6 Trogoderma versicolor (Creutzer 1799) 6 Dromius (Dromius) chobauti (Puel 1924) 5 Litargus (Alitargus) balteatus (LeConte 1856) 5 Magdalis (Magdalis) memnonia (Gyllenhal 1837) 5 Nemozoma elongatum (Linnaeus 1761) 5 Nyctophila reichii (Jacquelin du Val 1859) 5 Pogonocherus decoratus (Fairmaire 1855) 5 Sinoxylon sexdentatum (Olivier 1790) 5 Sphaeriestes (Sphaeriestes) castaneus (Panzer 1796) 5 Trogoderma granarium (Everts 1898) 5 Xylocleptes bispinus (Duftschmid 1825) 5 Anthrenus (Anthrenus) pimpinellae (Fabricius 1775) 4 Aplocnemus (Aplocnemus) brevis (Rosenhauer 1856) 4 Attalus (Abrinus) analis (Panzer 1796) 4 Lathropus sepicola (Muller 1821) 4 Platylomalus complanatus (Panzer 1796) 4 Psylliodes (Psylliodes) hyoscyami (Linnaeus 1758) 4 Sphaeriestes (Sphaeriestes) exsanguis (Abeille de Perrin 1870) 4 Teretrius (Neotepetrius) parasita (Marseul 1862) 4 Abdera (Abdera) quadrifasciata (Curtis 1829) 3 Anobium punctatum (De Geer 1774) 3 Anthaxia (Melanthaxia) godeti (Gory & Laporte 1839) 3 Anthaxia (Melanthaxia) nigrojubata (Roubal 1913) 3 Chalcophora mariana (Linnaeus, 1758) 3 Chlorophorus pilosus (Forster 1771) 3 Chrysobothris (Chrysobothris) solieri (Laporte & Gory 1839) 3 Dromius (Dromius) meridionalis (Dejean 1825) 3 Epuraea guttata (Olivier 1811) 3 Mauroania bourgeoisi (Pic 1894) 3 Mycterus (Mycterus) curculioides (Fabricius 1781) 3 Niphona picticornis (Mulsant 1839) 3 Nycteus meridionalis (Laporte de Castelnau 1836) 3 Pityophthorus buyssoni (Reitter 1901) 3 Plegaderus (Plegaderus) otti (Marseul 1856) 3 Scobicia pustulata (Fabricius 1801) 3 Stagetus elongatus (Mulsant & Rey 1861) 3 Xyleborus dispar (Fabricius 1792) 3

Anejo 4.7 Especies capturadas en la Red MUFFET en el año 2013 con frecuencia de captura menor a 3 individuos (Singletons/Doubletons)
Species Frequency Amadotrogus patruelis (Reiche 1862) 1 Anatis ocellata (Linnaeus 1758) 1 Anelastidius (Jacquelin du Val 1863) 1 Anthaxia (Haplanthaxia) confusa (Gory 1841) 1 Anthaxia (Haplanthaxia) parallela (Gory & Laporte 1839) 1 Anthaxia (Melanthaxia) lencinai (Arnáiz & Bercedo 2003) 1 Anthaxia (Melanthaxia) nigritula (Ratzeburg 1837) 2 Anthaxia (Melanthaxia) rugicollis (Lucas 1849) 2 Anthaxia (Melanthaxia) sepulchralis (Fabricius 1801) 2 Anthaxia (Melanthaxia) sp. 1 Anthrenus (Anthrenus) goliath (Saulcy 1868) 2 Apate monachus (Fabricius 1775) 1 Aplocnemus (Aplocnemus) limbipennis (Kiesenwetter 1865) 1 Bostrichus capucinus (Linnaeus 1758) 1 Brachyderes (Brachyderes) confusus (Viedma 1967) 2 Brachyderes (Brachyderes) grisescens (Fairmaire 1862) 1 Buprestis (Buprestis) novemmaculata (Linnaeus 1758) 2 Buprestis (Buprestis) octoguttata (Linnaeus 1758) 2 Calodromius bifasciatus (Dejean 1825) 1 Carphoborus pini (Eichhoff 1881) 2 Cerambyx cerdo (Linnaeus 1758) 2 Clanoptilus (Clanoptilus) abdominalis (Fabricius 1798) 2 Clerus mutillarius (Fabricius 1775) 2 Coccinella (Coccinella) septempunctata (Linnaeus 1758) 2 Crioceris macilenta (Weise 1880) 1 Crioceris paracenthesis (Linnaeus 1767) 1 Cryptocephalus (Cryptocephalus) lineellus Suffrian 1849 1 Cyphon sulcicollis (Mulsant & Rey 1865) 1 Dignomus irroratus (Kiesenwetter 1851) 2 Dromius (Dromius) agilis (Fabricius 1787) 1 Elathous platiai (Zapata & Sánchez-Ruiz 2007) 1 Ernobius parens (Mulsant & Rey 1863) 2 Ernobius reflexus (Mulsant & Rey 1863) 1 Ernobius rufus (Illiger 1807) 1 Glischrochilus quadripunctatus (Linnaeus 1758) 1 Haplomalachius (Flabellomalachius) hispanus (Perris 1865) 1 Harpalus (Harpalus) distinguendus (Duftschmid 1812) 1 Hymenalia rufipes (Fabricius 1792) 1 Ipidia binotata (Reitter 1875) 1 Lachnaia (Lachnaia) pseudobarathraea (K. Daniel & J. Daniel 1898) 1 Magdalis (Magdalis) rufa (Germar 1824) 1 Malachius lusitanicus (Erichson 1840) 1 Mauroania hispana (Kiesenwetter 1859) 1 Megatoma (Megatoma) undata (Linnaeus 1758) 1 Melanotus (Melanotus) castanipes (Paykull 1800) 1 Microlestes abeillei (Brisout de Barneville 1885) 1 Mycetophagus (Parilendus) quadriguttatus (Muller 1821) 2 Mycetophagus (Ulolendus) salicis (Brisout de Barneville 1862) 2 Mylabris (Mylabris) quadripunctata (Linnaeus 1767) 1 Myrrha (Metamyrrha) thuriferae (Sicard 1923) 2 Myrrha (Myrrha) octodecimguttata (Linnaeus 1758) 1 Nathrius brevipennis (Mulsant 1839) 1 Nephus (Bipunctatus) bipunctatus (Kugelann 1794) 1 Nothorhina muricata (Dalman 1817) 2 Novius cruentatus (Mulsant 1850) 2 Opilo domesticus (Sturm 1837) 1 Oxylaemus cylindricus (Panzer 1796) 2 Paracorymbia stragulata (Germar 1824) 1 Paratriodonta alicantina (Reitter 1890) 1 Philorhizus crucifer (Lucas 1846) 1 Philorhizus notatus (Stephens 1827) 1 Pissodes (Pissodes) pini (Linnaeus 1758) 2 Poecilium glabratum (Charpentier 1825) 2 Pogonocherus fasciculatus (De Geer 1775) 1 Pogonocherus sturanii (Sama & Schurmann 1982) 1 Prionus (Prionus) coriarius (Linnaeus 1758) 1 Probaticus anthracinus (Germar 1813) 1 Protaetia (Netocia) morio (Fabricius 1781) 2 Protaetia (Netocia) oblonga (Gory & Percheron 1833) 1 Pseudomasoreus canigoulensis (Fairmaire & Laboulbene 1854) 1 Ptinomorphus imperialis (Linnaeus 1767) 1 Ptinus (Ptinus) fur (Linnaeus 1758) 1 Ptinus (Ptinus) perrini (Reitter 1884) 1 Ptinus (Ptinus) subpilosus (Sturm 1837) 1 Ptinus clavipes (Panzer 1792) 2 Rhyncolus (Rhyncolus) sculpturatus (Waltl 1839) 1 Sphenoptera (Chilostetha) laportei (Saunders 1871) 1 Stagetus byrrhoides (Mulsant & Rey 1861) 1 Stenurella bifasciata (Muller 1776) 1 Stylosomus (Stylomicrus) minutissimus (Germar 1824) 1 Symbiotes gibberosus (Lucas 1846) 1 Syntomus fuscomaculatus (Motschulsky 1844) 1 Trichodes leucopsideus (Olivier 1795) 1 Triplax lacordairii (Crotch 1870) 1 Tropinota (Tropinota) squalida (Scopoli 1783) 1 Vesperus xatarti (Mulsant 1839) 2 Xylopertha praeusta (Germar 1817) 1

Anejo 4.8 Lista de especies asociada a cada combinación. Multilevel pattern analysis. Función de asociación: IndVal.g. Nivel de significación (alfa): 0.05. Número de especies total: 93. Número de especies seleccionado: 30. Número de especies asociadas al grupo 1: 13. Número de especies asociadas al grupo 2: 10. Número de especies asociadas al grupo 3: 4. Número de especies asociadas al grupo 4: 1. Número de especies asociadas al grupo 5: 2. Número de especies asociadas al grupo 6: 0.
Grupo stat p.valor
Group 2 Hylurgligniper 0.789 0.038
Hymenodoublier 0.533 0.047
Group 3 Hylastattenuat 0.944 0.001
Hylobiabietis 0.777 0.001 Pityoplaevior 0.773 0.003 Acanthaedilis 0.608 0.021
Group 4 Pityogcalcarat 0.987 0.001
Brachyporcatus 0.677 0.007 Chrysosolieri 0.509 0.028
Group 5 Ipssexdenta 0.918 0.001
Ipsmannsfel 0.735 0.014
Group 6 Arhopasyriacus 0.712 0.027
Laconpunctatu 0.553 0.033
Group 2+3 Xyleboeurygrap 0.654 0.025
Group 3+5 Rhizopferrugin 0.881 0.001
Thanasformicar 0.860 0.001 Hylastater 0.846 0.007 Allonyquadrima 0.637 0.026 Ptinuspyrenaeu 0.565 0.042
Group 3+6 Rhagiuinquisit 0.681 0.007
Oxyplenodieri 0.656 0.023
Group 5+6 Thanasfemorali 0.729 0.013
Group 6+7 Arhopaferus 0.74 0.007
Group 1+3+5 stat p.value Rhizopdepressu 0.788 0.023
Group 2+4+7 Hylurgmiklitzi 0.959 0.001
Xylebosaxeseni 0.699 0.013
Group 2+5+6 stat p.value Spondybupresto 0.808 0.007
Group 2+3+5+6 Temnoscoerulea 0.846 0.001
Group 2+3+4+5+7 stat p.value Orthoterosus 0.941 0.001 Auloniruficorn 0.912 0.018

Anejo 4.9 Lista de especies asociada a cada combinación. Multilevel pattern analysis. Función de asociación: IndVal.g. Nivel de significación (alfa): 1. Número de especies total: 93. Número de especies seleccionado: 93. Número de especies asociadas al grupo 1: 30. Número de especies asociadas al grupo 2: 31. Número de especies asociadas al grupo 3: 18. Número de especies asociadas al grupo 4: 7. Número de especies asociadas al grupo 5: 5. Número de especies asociadas al grupo 6: 2. Grupo stat p.value
Group 2 Hylurgligniper 0.789 0.038
Hymenodoublier 0.533 0.047
Teretrparasita 0.387 0.322
Isomirhispanic 0.354 0.286
Tomicupiniperd 0.250 0.710
Group 3 Hylastattenuat 0.944 0.001
Hylobiabietis 0.777 0.001
Pityoplaevior 0.773 0.003
Acanthaedilis 0.608 0.021
Cylistelongatu 0.571 0.063
Attaluanalis 0.500 0.059
Anthaxnigrojub 0.354 0.317
Ernobipini 0.332 0.414
Group 4 Pityogcalcarat 0.987 0.001
Brachyporcatus 0.677 0.007
Chrysosolieri 0.509 0.028
Litargbalteatu 0.483 0.062
Group 5 Ipssexdenta 0.918 0.001
Ipsmannsfel 0.735 0.014
Caenosferrugin 0.365 0.274
Ptinuslatro 0.258 0.517
Group 6 Arhopasyriacus 0.712 0.027
Laconpunctatu 0.553 0.033
Penthachrysome 0.367 0.237
Group 7 Tomicudestruen 0.378 0.222
Bergintamarisc 0.309 0.527
Niphonpicticor 0.309 0.523
Xylebodispar 0.309 0.525
Stagetelongatu 0.218 1.000
Trogodgranariu 0.218 1.000
Group 1+6 stat p.value
Epuraeguttata 0.408 0.164
Dromiuchobauti 0.374 0.266
Group 1+7 Pogonoperroudi 0.467 0.208
Enedrehilaris 0.392 0.265
Group 2+3 Xyleboeurygrap 0.654 0.025
Lobonyaeneus 0.455 0.244
Chrysareitteri 0.440 0.190
Group 2+6 Chalcomariana 0.361 0.335
Cylistfiliform 0.352 0.702
Group 3+4 Nyctopreichii 0.471 0.088
Group 3+5 Rhizopferrugin 0.881 0.001
Thanasformicar 0.860 0.001
Hylastater 0.846 0.007
Allonyquadrima 0.637 0.026
Ptinuspyrenaeu 0.565 0.042
Anthaxgodeti 0.361 0.325
Abderaquadrifa 0.295 0.586
Group 3+6 Rhagiuinquisit 0.681 0.007
Oxyplenodieri 0.656 0.023
Stagetpilula 0.510 0.097
Aplocnconsobri 0.492 0.091
Group 4+7 Cryptunumidicu 0.498 0.104
Group 5+6 Thanasfemorali 0.729 0.013
Arhoparusticus 0.606 0.072
Stenaglaufferi 0.468 0.174
Anthrefuscus 0.462 0.194
Magdalmemnonia 0.369 0.294
Chloropilosus 0.302 0.535
Group 5+7 Brachypubescen 0.333 0.556
Xyloclbispinus 0.236 0.868
Group 6+7 Arhopaferus 0.74 0.007
Group 1+2+5 Nemozoelongatu 0.333 0.53
Group 1+3+5 Rhizopdepressu 0.788 0.023
Salpinplaniros 0.423 0.214
Group 1+5+6 Pityoplichtens 0.385 0.34
Group 2+3+5 Ipsacuminat 0.555 0.096
Acmaeobipuncta 0.381 0.606
Group 2+3+6 Dasyteterminal 0.44 0.199
Group 2+3+7 Xylopepicea 0.365 0.636
Hylastlinearis 0.354 0.806
Group 2+4+7 Hylurgmiklitzi 0.959 0.001
Xylebosaxeseni 0.699 0.013
Group 2+5+6 Spondybupresto 0.808 0.007
Pogonodecoratu 0.324 0.632
Group 2+5+7 Platylcomplana 0.277 0.931
Group 3+4+6 Scobicchevrier 0.381 0.411
Group 3+5+6 Megapelugens 0.577 0.060
Diaperboleti 0.483 0.298
Mycetoquadripu 0.478 0.167
Group 1+2+5+6 Paromaparallel 0.574 0.558
Acanthhispanic 0.571 0.062
Group 2+3+5+6 Temnoscoerulea 0.846 0.001
Phaenocyanea 0.507 0.238
Group 2+3+6+7 Litargconnexus 0.451 0.547
Group 2+5+6+7 Dermesundulatu 0.319 0.914
Trogodversicol 0.260 0.993
Group 1+2+3+4+7 Cryptumediterr 0.767 0.087
Group 1+2+5+6+7 Corticpini 0.886 0.147
Group 2+3+4+5+7 Orthoterosus 0.941 0.001
Auloniruficorn 0.912 0.018
Group 2+3+4+6+7 Cryptoferrugin 0.652 0.147
Group 1+2+3+5+6+7 Monochgallopro 0.838 0.170
Acanthgriseus 0.794 0.426

Anejos Capítulo 5 Anejo 5.1. Estimación del viento dominante para cada insecto. Se muestra un ejemplo de cada uno de los ensayos tenidos en cuenta
en este análisis.
Exp2010/b La Muela, Jérica (Castellón) Exp2011/a Boqueras, Altura (Castellón)
Exp2011/b Atzeneta del Maestrat (Castellón)
Exp2012/b Atzeneta del Maestrat (Castellón)

Anejo 5.2 Representación gráfica de la regresión entre el viento y el desplazamiento de los insectos durante su periodo en libertad (1ª columna) a las diferentes horas del día de la toma de datos meteorológicos. En la segunda columna, los gráficos muestran las
direcciones e intensidades dominantes de los vientos a las diferentes horas del día.
Exp2010/a (Jérica, Castellón)

Exp2011/a (Altura, Castelló)

Exp2011/b (Atzeneta del Maestrat, Castelló)

Expe2012/b (Atzeneta del Maestrat, Castelló)