parasites as pollution indicators in marine ecosystems: a proposed early warning system
TRANSCRIPT

Parasites as Pollution Indicators inMarine Ecosystems: a Proposed EarlyWarning SystemKEN MACKENZIEDepartment of Zoology, The University of Aberdeen, Tillydrone Avenue, Aberdeen, Scotland AB24 2TZ, UK
There are good reasons for focusing on parasites in thesearch for indicators to monitor the e�ects of pollutantson marine organisms. Firstly, there are more parasiticthan free-living species. Secondly, in parasites with com-plex life cycles, the di�erent stages have widely di�eringrequirements, so that each stage must be assessed sepa-rately, thereby greatly increasing the number of potentialindicators. Thirdly, many parasites have delicate free-living transmission stages which are highly sensitive toenvironmental change. A reduction in their levels of in-fection will serve as an early warning that changes areoccurring. Conversely, other parasites are highly resistantto environmental change and will respond by increasedlevels of infection. As a general rule, infections withendoparasitic helminths tend to decrease, while infectionswith ectoparasites tend to increase, with increasing levelsof pollution. Ó 1999 Elsevier Science Ltd. All rightsreserved.
Keywords: marine ecosystems; parasites; pollution.
Introduction
Good biological indicators to monitor the e�ects ofpollutants on marine organisms must either be excep-tionally sensitive or exceptionally resistant to environ-mental change so that signi®cant changes in theirnumbers can be used as an early warning of deterio-rating conditions before the majority of organisms areseriously a�ected. There are good reasons for focusingon parasites in the search for such indicators.
1. There are more parasitic than free-living species onearth (Windsor, 1998) and parasitic organismsshow enormous biological diversity, re¯ecting ad-aptations to the parasitic way of life in di�erenttypes of hosts and in diverse sites and environ-ments.
2. Add to this the fact that in metazoan parasiteswith complex life cycles, the di�erent developmen-tal stages have widely di�ering biological require-ments, so that each stage must be assessedseparately, thereby greatly increasing the numberof potential indicators.
3. Many parasites have delicate short-lived free-livingtransmission stages which are highly sensitive toenvironmental change. They can therefore be ad-versely a�ected by even minor changes in the envi-ronment and so represent weak links in the lifecycles of the parasites.
While many parasites are extremely sensitive to en-vironmental change, others are more resistant than theirhosts and tend to increase in numbers in polluted con-ditions. As a general rule, infections with endoparasitichelminths with complex indirect life cycles tend to de-crease, while infections with ectoparasites with directsingle-host life cycles tend to increase, with increasinglevels of pollution. Endoparasites with indirect life cy-cles can be a�ected directly or indirectly by pollutants:directly by free-living transmission stages or adult formsin the alimentary tract coming into direct contact withthe pollutant, or indirectly through adverse e�ects of thepollutant on other hosts in the parasite's life cycle. Ec-toparasites with direct life cycles, on the other hand, arein constant direct contact with the external environmentand in the course of their evolution they have developeda ¯exibility and resistance to certain natural changes.Consequently, many ectoparasites have shown that theyare more tolerant than their hosts of certain types ofenvironmental change. However, as La�erty (1997) hasshown, there are enormous variations in the responsesof di�erent parasite taxa to di�erent types of pollution(Table 1). The numbers of trichodinid gill ciliates of ®sh,which have direct single-host life cycles, increase withseveral types of pollution. In contrast, the numbers ofdigeneans, cestodes and acanthocephalans, which have
Marine Pollution Bulletin Vol. 38, No. 11, pp. 955±959, 1999
Ó 1999 Elsevier Science Ltd. All rights reserved
Printed in Great Britain
0025-326X/99 $ - see front matterPII: S0025-326X(99)00100-9
955

complex indirect life cycles, have been found to decreasewith most types of pollution.
The use of parasites as indicators of aquatic pollutionhas been reviewed by M�oller (1987), Khan and Thulin(1991), MacKenzie et al. (1995) and La�erty (1997).Fish parasites as indicators of environmental qualitywas the subject of a special symposium at the VII Eu-ropean Multicolloquium of Parasitology (Paperna,1997). The present article suggests ways of using parasitetransmission processes as sensitive early warning indi-cators of marine pollution.
A Proposed Early Warning System
Until recently, attempts to use parasites as indicatorsof marine pollution focused on the pathology associatedwith parasitic infections. This approach, however, islimited by lack of sensitivity and delayed response(McVicar, 1986). Most obvious pathological conditionsare the end result of a sequence of events that may haveoccurred over a prolonged period and therefore relate tochanges in environmental conditions dating back someconsiderable time. This is particularly true of pathologyresulting from infections with metazoan parasites, whichhave longer generation times than micro-organisms suchas viruses, bacteria and protozoans, and consequentlytake longer to build up their populations to levels likelyto cause disease. Some of the free-living transmissionstages of metazoan parasites, particularly the helminths,are delicate short-lived larvae that are highly sensitive tochanges in environmental conditions. An earlier warn-ing of environmental change is therefore likely to beprovided by monitoring the transmission of selectedmetazoan parasites.
Transmission routes and transmission windowsA transmission route is the pathway through which a
parasite progresses from one developmental stage to thenext and from one host to another. Most parasites havea range of possible host species resulting in a number ofpossible transmission routes. The number of routes isgreatly increased in parasites with life cycles involvingthree or more hosts at di�erent stages of development.
For example, a digenetic trematode will usually infectonly one species of primary mollusc host, or a smallgroup of closely related species, but may have a widerange of intermediate or de®nitive host species. Thesehosts can be arranged along a continuum from those towhich the parasite is best adapted and which provide itwith the highest probability of successful transmission tothe next stage, to `dead-end' hosts or `ecological sinks'through which no further development is possible(Holmes, 1979).
Associated with each transmission route is a trans-mission window. This is the period during which trans-mission of the parasite from one host to another can takeplace. The length of this period depends on the concur-rence of infective stages of the parasite with hosts that aresusceptible to infection. With some transmission routes,the windows are open throughout the entire life span ofthe host so infection can take place at any time. In others,the windows are brief seasonal periods which representweak links in parasite life cycles where routes throughcertain host species may be easily disrupted by changes inenvironmental conditions. Examples of two such narrowtransmission windows are illustrated in Fig. 1. Mac-Kenzie andGibson (1970) showed that infection of plaicePleuronectes platessawith metacercariae of the digeneansStephanostomum baccatum and Rhipidocotyle sp. occursonly in the ®rst year of life, for S. baccatum during Juneand July and for Rhipidocotyle sp. from about the middleof September to the end of October. Transmission ofeach of these parasites to plaice is therefore limited to aperiod of 6±8 weeks in a host species with a maximum lifespan of over 20 years. This limited period is determinedby two things: plaice are susceptible to penetration by thecercariae only when they are very young, and there isonly a short annual period when free-swimming cercariaeemerge from the mollusc host. In a year when emergenceor viability of cercariae or normal growth of plaice isdisrupted by a change in environmental conditions, anentire year class of plaice might show a signi®cantly re-duced level of infection.
Another good example of a narrow transmissionwindow is infection of herring Clupea harengus with thedigeneans Cercaria doricha and C. pythionike. Free-
TABLE 1
E�ects of di�erent types of pollution on di�erent parasite taxa.a
Type of pollution Parasite taxa
Ciliophora Monogenea Nematoda Digenea Cestoda Acanthocephala
Eutrophication + + + + +Thermal e�uent + + ÿPulp-mill e�uent + ÿ ÿ ÿCrude oil + + + ÿ ÿIndustrial e�uent + ÿ ÿ ÿ ÿSewage sludge + ÿAcid precipitation ÿ ÿ ÿ +Heavy metals ÿ ÿ ÿa Plus signs indicate an increase in parasite abundance, minus signs a decrease, and blank spaces indicate where no data are available (after La�erty,1997).
956
Marine Pollution Bulletin

swimming cercariae emerge from the mollusc host dur-ing a period of about three months in spring and earlysummer and herring become infected by ingesting thebrightly coloured cercariae which penetrate the gut walland encyst in the visceral cavity (Wright, 1956). All in-fection of the herring takes place in the ®rst summer oflife, when they are feeding on planktonic organisms ofabout the same size as the cercariae. The fastest growingjuvenile herring apparently progress to feeding on largerorganisms before the end of the period of cercarialemergence (MacKenzie, 1985). Transmission is thereforelimited to a period of one to three months in a hostspecies with a maximum life span of over 20 years. In ayear when conditions favour the rapid growth of juve-nile herring and/or cercarial emergence is delayed orinhibited, levels of infection could be greatly reduced.The above examples feature digeneans, but other
helminths also have narrow and vulnerable transmissionwindows in their life cycles. A good example of howhuman interference with an ecosystem can drasticallya�ect the transmission of a cestode parasite is given byHanzelova (1992). In the year following drainage of afreshwater reservoir in the Czech Republic, the speciescomposition of the copepod community and the sea-sonal dynamics of copepod numbers changed dramati-cally. The copepods serve as intermediate hosts for the®sh cestode Proteocephalus neglectus and the e�ect ontransmission of this parasite was that infection of rain-bow trout Oncorhynchus mykiss in the reservoir de-creased by 97.5% from the year before drainage to theyear after.
Reported transmission failures of marine parasitesNagasawa et al. (1988) described how transmission of
the parasitic copepod Pennella sp. to its ®sh host, the
saury Cololabis saira, in the western North Paci®c failedin 1985. No infected saury were found in this region in1986, but the parasite's reappearance in 1990 was re-ported by Honma and Imai (1991).
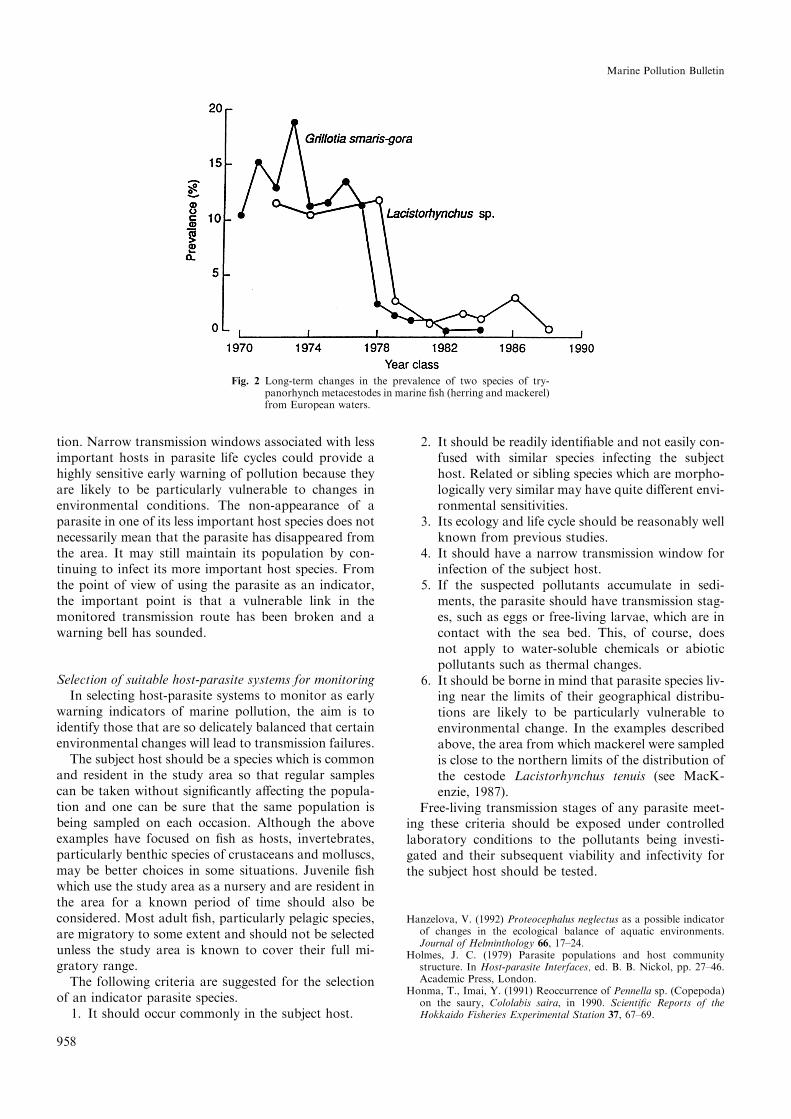
Transmission of Pennella sp. is direct from ®sh to ®shwithout the intervention of an intermediate host, butFig. 2 shows examples of sudden transmission failuresof two marine cestode parasites with complex three-host life cycles. MacKenzie (1987) described transmis-sion failures of the cestodes Grillotia smaris-gora andLacistorhynchus tenuis to their ®sh hosts mackerelScomber scombrus and herring C. harengus, respective-ly, in European waters. Infection of herring begins inthe ®rst, and of mackerel in the second, year of life. Inyear classes of both host species from 1970±1972 to1978 prevalences of both cestodes remained fairly sta-ble, but in 1979 prevalences of both species were dra-matically reduced and had not recovered whensampling stopped in 1984 for mackerel and in 1988 forherring.
The reasons for both the above examples of trans-mission failures are unknown, but they are almostcertainly related to natural environmental changes. Itmay be signi®cant that the sudden change in prevalenceof the two cestodes coincided with the end of a hy-drographic phenomemon in the North Atlantic knownas the 1970s salinity anomaly (Martin et al., 1984), butif the events were indeed linked, it is likely to havebeen through a complex series of causes and e�ectsinvolving changes in a number of biotic and abioticfactors.
In the above examples, plaice, herring and mackerelare each one of many possible second intermediate hostsof these particular parasites, and each is among the lessimportant species for maintaining the parasite popula-
Fig. 1 Changes with time in the prevalence of two species of digeneanmetacercariae infecting juvenile plaice. The transmission win-dows are shaded.
957
Volume 38/Number 11/November 1999
tion. Narrow transmission windows associated with lessimportant hosts in parasite life cycles could provide ahighly sensitive early warning of pollution because theyare likely to be particularly vulnerable to changes inenvironmental conditions. The non-appearance of aparasite in one of its less important host species does notnecessarily mean that the parasite has disappeared fromthe area. It may still maintain its population by con-tinuing to infect its more important host species. Fromthe point of view of using the parasite as an indicator,the important point is that a vulnerable link in themonitored transmission route has been broken and awarning bell has sounded.
Selection of suitable host-parasite systems for monitoringIn selecting host-parasite systems to monitor as early
warning indicators of marine pollution, the aim is toidentify those that are so delicately balanced that certainenvironmental changes will lead to transmission failures.
The subject host should be a species which is commonand resident in the study area so that regular samplescan be taken without signi®cantly a�ecting the popula-tion and one can be sure that the same population isbeing sampled on each occasion. Although the aboveexamples have focused on ®sh as hosts, invertebrates,particularly benthic species of crustaceans and molluscs,may be better choices in some situations. Juvenile ®shwhich use the study area as a nursery and are resident inthe area for a known period of time should also beconsidered. Most adult ®sh, particularly pelagic species,are migratory to some extent and should not be selectedunless the study area is known to cover their full mi-gratory range.
The following criteria are suggested for the selectionof an indicator parasite species.
1. It should occur commonly in the subject host.
2. It should be readily identi®able and not easily con-fused with similar species infecting the subjecthost. Related or sibling species which are morpho-logically very similar may have quite di�erent envi-ronmental sensitivities.
3. Its ecology and life cycle should be reasonably wellknown from previous studies.
4. It should have a narrow transmission window forinfection of the subject host.
5. If the suspected pollutants accumulate in sedi-ments, the parasite should have transmission stag-es, such as eggs or free-living larvae, which are incontact with the sea bed. This, of course, doesnot apply to water-soluble chemicals or abioticpollutants such as thermal changes.
6. It should be borne in mind that parasite species liv-ing near the limits of their geographical distribu-tions are likely to be particularly vulnerable toenvironmental change. In the examples describedabove, the area from which mackerel were sampledis close to the northern limits of the distribution ofthe cestode Lacistorhynchus tenuis (see MacK-enzie, 1987).
Free-living transmission stages of any parasite meet-ing these criteria should be exposed under controlledlaboratory conditions to the pollutants being investi-gated and their subsequent viability and infectivity forthe subject host should be tested.
Hanzelova, V. (1992) Proteocephalus neglectus as a possible indicatorof changes in the ecological balance of aquatic environments.Journal of Helminthology 66, 17±24.
Holmes, J. C. (1979) Parasite populations and host communitystructure. In Host-parasite Interfaces, ed. B. B. Nickol, pp. 27±46.Academic Press, London.
Honma, T., Imai, Y. (1991) Reoccurrence of Pennella sp. (Copepoda)on the saury, Cololabis saira, in 1990. Scienti®c Reports of theHokkaido Fisheries Experimental Station 37, 67±69.
Fig. 2 Long-term changes in the prevalence of two species of try-panorhynch metacestodes in marine ®sh (herring and mackerel)from European waters.
958
Marine Pollution Bulletin

Khan, R. A., Thulin, J. (1991) In¯uence of pollution on parasites ofaquatic animals. Advances in Parasitology 30, 201±238.
La�erty, K. D. (1997) Environmental parasitology, what can parasitestell us about human impacts on the environment? ParasitologyToday 13, 251±255.
MacKenzie, K. (1985) The use of parasites as biological tags inpopulation studies of herring (Clupea harengus L.) in the North Seaand to the north and west of Scotland. Journal du Conseil. Conseilinternational pour l'exploration de la Mer 42, 33±64.
MacKenzie, K. (1987) Long-term changes in the prevalence of twohelminth parasites (Cestoda: Trypanorhyncha) infecting marine®sh. Journal of Fish Biology 31, 83±88.
MacKenzie, K., Gibson, D. I. (1970) Ecological studies of someparasites of plaice Pleuronectes platessa L. and ¯ounder Platichthys¯esus (L). Symposia of the British Society for Parasitology 8, 1±42.
MacKenzie, K., Williams, H. H., Williams, B., McVicar, A. H.,Siddall, R. (1995) Parasites as indicators of water quality and thepotential use of helminth transmission in marine pollution studies.Advances in Parasitology 35, 85±144.
Martin, J. H. A., Dooley, H. D., Shearer, W. (1984) Ideas on the originand biological consequences of the 1970s salinity anomaly. ICESC.M.1984/Gen:18, Minisymposium, p. 7 (mimeo).
McVicar, A. H. (1986) The use of ®sh pathology in programmes tomonitor marine contaminants. In Report of the ICES Workshop onthe Use of Pathology in Studies of the E�ects of Contaminants, ed. J.Thulin, pp. 21±22.
M�oller, H. (1987) Pollution and parasitism in the aquatic environment.International Journal for Parasitology 17, 353±361.
Nagasawa, K., Imai, Y., Ishida, K. (1988) Long-term changes in thepopulation size and geographical distribution of Pennella sp.(Copepoda) on the saury, Cololabis saira, in the western NorthPaci®c Ocean and adjacent seas. In Biology of Copepods, eds. G. A.Boxshall, H. K. Schminke, pp. 571±578. Kluwer AcademicPublishers, Dordrecht, Belgium.
Paperna, I. (1997) Fish Parasites as Indicators of EnvironmentalQuality. VII European Multicolloquium of Parasitology, Parma,Italy, 2±6 September 1996. Parassitologia 39 (3), 255 pp.
Windsor, D. A. (1998) Most of the species on earth are parasites.International Journal for Parasitology 28, 1939±1942.
Wright, C. A. (1956) Studies on the life history and ecology of thetrematode genus Renicola Cohn, 1904. In Proceedings of theZoological Society of London 126, 1±49.
959
Volume 38/Number 11/November 1999