osmotic and mechanical loading of chondrocytes in situ: effect of
TRANSCRIPT

Publications of the University of Eastern Finland Dissertations in Forestry and Natural Sciences
Publications of the University of Eastern Finland
Dissertations in Forestry and Natural Sciences
isbn 978-952-61-1088-2
Siru Turunen
Osmotic and MechanicalLoading of Chondrocytes in SituEffect of the Extracellular and Pericellular Matrix on
Cell Volume and Morphology
Changed chondrocyte deformations in
articular cartilage are thought to be related
to changed cell biosynthesis and viability;
alterations which may lead to and accelerate
the progression of osteoarthritis (OA). How-
ever, it is not known whether the structural
changes of cartilage in early OA are concur-
rent with the changes in the biomechanical
responses of chondrocytes. The aim of this
thesis was to reveal how the structure and
composition of the extra- and pericellular
matrices of cartilage affect chondrocyte
volume and morphology under osmotic and
mechanical loading of the tissue. By under-
standing chondrocyte behavior in situ/in
vivo it might be possible to devise strategies
to slow down, stop or even reverse the devel-
opment of OA.
dissertatio
ns | 103 | S
iru
Tu
ru
nen
| Osm
otic and Mechanical L
oading of Chondrocytes in S
itu - Effect of the E
xtracellular and Pericellular M
atrix...
Siru Turunen
Osmotic and MechanicalLoading of Chondrocytes in Situ
Effect of the Extracellular and Pericellular Matrix on
Cell Volume and Morphology

SIRU TURUNEN
Osmotic and MechanicalLoading of Chondrocytes in
SituEffect of the Extracellular and Pericellular Matrix on
Cell Volume and Morphology
Publications of the University of Eastern FinlandDissertations in Forestry and Natural Sciences
No 103
Academic DissertationTo be presented by permission of the Faculty of Science and Forestry for public
examination in the Auditorium in Mediteknia Building at the University of EasternFinland, Kuopio, on June, 1, 2013, at 12 o’clock noon.
Department of Applied Physics

Kopijyvä OyKuopio, 2013
Editors: Prof. Pertti Pasanen, Prof. Kai-Erik Peiponen, Prof. MattiVornanen, Prof. Pekka Kilpeläinen
Distribution:Eastern Finland University Library / Sales of publications
P.O.Box 107, FI-80101 Joensuu, Finlandtel. +358-50-3058396
http://www.uef.fi/kirjasto
ISBN: 978-952-61-1088-2 (printed)ISSNL: 1798-5668ISSN: 1798-5668
ISBN: 978-952-61-1089-9 (pdf)ISSN: 1798-5676

Author’s address: Department of Applied PhysicsUniversity of Eastern FinlandP.O.Box 1627FI-70211 KUOPIOFINLANDemail: [email protected]
Supervisors: Adjunct Professor Rami Korhonen, Ph.D.Department of Applied PhysicsUniversity of Eastern Finlandemail: [email protected]
Professor Mikko Lammi, Ph.D.Institute of BiomedicineUniversity of Eastern Finlandemail: [email protected]
Adjunct Professor Simo Saarakkala, Ph.D.Department of Medical TechnologyInstitute of BiomedicineUniversity of Ouluemail: [email protected]
Reviewers: Professor Clark T. Hung, Ph.DUniversity of ColumbiaDepartment of Biomedical Engineering351 Engineering Terrace1210 Amsterdam Avenue, Mail Code: 8904New York, NY 10027, USAemail: [email protected]
Assistant Professor Leonidas G. Alexopoulos, Ph.DDepartment of Mechanical EngineeringNational Technical University of AthensHeroon Polytechniou 915780 Zografou, Athens, Greeceemail: [email protected]
Opponent: Professor Alan J. Grodzinsky, Sc.D.Massachusetts Institute of TechnologyCenter for Biological Engineering500 Technology SqCambridge, MA 02139, USAemail: [email protected]


ABSTRACT
Articular cartilage covers the ends of synovial joints and itsfunction is to distribute loads between opposing bones andprovide almost nonfrictional sliding properties for joints. It iscomprised of cells (chondrocytes) embedded in an extracellularmatrix (ECM) containing mainly collagen, proteoglycans (PGs)and interstitial fluid. Chondrocytes are responsible for theproduction of ECM macromolecules and thus, for maintainingthe integrity of the ECM. Alterations in cell biomechanics mightlead to changes in cell biosynthesis and viability.
Degeneration of cartilage in osteoarthritis (OA) results indysfunction and severe pain in joints. Although the onset andprogression of OA have been studied extensively, it is still notknown whether the structural changes related to cartilagedegeneration occur concurrently with changes in chondrocytebiomechanics. More specifically, it is unclear how the integrityof the cartilage matrix, the alterations in the amount of cartilagemacromolecules, and the changes related to cartilage structureand composition in early OA, are interconnected to changes inchondrocyte behavior. Also the detailed explanations behind theobserved phenomena remain unknown.
In this thesis, the effects of the structure and composition ofthe ECM and pericellular matrices (PCM) on chondrocytebiomechanics were studied. Three-dimensional fluorescencemicroscopy was used in order to analyze changes occurring inchondrocyte volume and morphology in bovine and rabbitcartilages before and after osmotic and mechanical loading. Theeffects of sample integrity, collagen degradation, anddegeneration of cartilage in early OA on chondrocyte behaviorwere examined in detail. Collagen content, collagen orientationangle, and PG content were analyzed with Fourier transforminfrared microspectroscopy, polarized light microscopy anddigital densitometry, respectively. Computational modeling wasutilized in order to identify the underlying mechanisms behindthe observed phenomena.

Chondrocyte volume and morphology following osmotic andmechanical loading were found to be dependent on thestructure and integrity of the ECM and PCM. Cells in both intactand explant cartilages swelled in response to hypotonic loading,but volume recovery was only present in cartilage explantswhere sample integrity was compromised. Chondrocytes inenzymatically degraded cartilage, where the amount of collagenwas reduced and the collagen network was severely fibrillated,were also found to recover back to the original, unchallengedvolume. In a rabbit model of early OA, chondrocytes undermechanical loading increased in volume, whereas cell volumesin the contralateral joint cartilage decreased. Detailed structuraland finite element analyses revealed that the decrease in theamount of fixed charge density (FCD) of the tissue and PCM,and the orientation and stiffness of the collagen fibers in theECM explained the changed cell volumetric behavior in earlyexperimentally-induced OA.
To conclude, biomechanical responses of chondrocytes inosmotically and mechanically loaded cartilage were greatlyaffected by their mechanical environment. Tissue integrity,decrease in collagen content, and collagen network fibrillationaltered cell volumetric behavior following osmotic loading. Theloss of FCD in the PCM, and fibrillation of the superficial ECMin the very early stage of experimentally-induced OA explainedthe cell volume increase following mechanical loading ofcartilage. Changes in cell biomechanics have earlier been foundto be related to altered cell biosynthesis and impaired cellviability; all of which may lead to further changes in cartilagestructure and accelerate the cartilage degeneration. Thus, it is ofgreat importance to understand chondrocyte behavior in situ/invivo in order to devise strategies to slow down, stop or evenreverse the development of OA.
National Library of Medicine Classification: QU 55.3, QU 350, WE 103, WE300, WE 348

Medical Subject Headings: Cartilage, Articular; Chondrocytes; ExtracellularMatrix; Collagen; Proteoglycans; Biomechanics; Osmotic Pressure; Stress,Mechanical; Osteoarthritis; Disease Models, Animal; Microscopy; SpectrumAnalysis
Yleinen suomalainen asiasanasto: nivelrusto; kondrosyytit; soluväliaine;kollageenit; proteoglykaanit; biomekaniikka; osmoottinen paine; mekaaninenrasitus; nivelrikko; koe-eläinmallit; mikroskopia; spektroskopia

To Sami, Kiira andbaby Apple

Acknowledgements
This thesis was carried out during the years 2008-2013 in theDepartment of Applied Physics, University of Eastern Finland.
I want to thank my first supervisor Adjunct Professor RamiKorhonen for his guidance and encouragement during theseyears. It has been my privilege to be the first Ph.D. student in hisresearch group.
I also thank my other supervisors Adjunct Professor SimoSaarakkala and Professor Mikko Lammi for their help.Furthermore, I wish thank to my co-authors Dr. Sang-Kuy Han,Professor Walter Herzog, Dr. Arto Koistinen, Adjunct ProfessorPetro Julkunen, and Petri Tanska for their significantcontributions. Furthermore, I would like to thank JaanaMäkitalo, Yevgenia Kobrina and Carolina Mangare for theirassistance in the measurements. I am grateful for being part ofRami’s research group and having close collaboration with theBiophysics of Bone and Cartilage group. I acknowledge theInstitute of Biomedicine for the cooperation provided duringthese years.
I thank the official reviewers of this thesis, Professor Clark T.Hung and Assistant Professor Leonidas G. Alexopoulos for theircomments and constructive criticism. I also thank KimberleyBoisvert and Ewen MacDonald, D.Pharm., for linguistic review.
I am grateful for having received financial support fromAcademy of Finland, European Research Council, EmilAaltonen Foundation, Finnish Cultural Foundation (North-SavoRegional Fund), Kuopio University Hospital, Kuopionyliopistosäätiö, National Doctoral Programme ofMusculoskeletal Disorders and Biomaterials and Sigrid JuseliusFoundation. Atria lihakunta Oyj, Kuopio, is acknowledged forproviding bovine knees for research purposes.
I am grateful to my parents, Airi and Hannu, and my brotherSanttu for encouraging me and believing in me throughout my

life. I thank all my friends for providing me with a life outside ofscience.
Most importantly, I want to thank my loving family, Samiand Kiira, for being there for me every day. With your love andunderstanding, everything is possible.
Kuopio, May 2013
Siru Turunen

LIST OF ABBREVIATIONS
ACLT Anterior cruciate ligament transectionAFM Atomic force microscopyANOVA Analysis of varianceAU Absorption unitCCD Charge-coupled deviseCLSM Confocal laser scanning microscopyDD Digital densitometryDMEM Dulbecco’s modified Eagle’s mediumDVRT Differential variable reluctance transducerD.Z. Deep zoneECM Extracellular matrixEDTA Ethylenediaminetetra-acetic acidFCD Fixed charge densityFE Finite elementFTIR Fourier transform infraredGAG GlycosaminoglycanIR InfraredLPG Lateral patellar grooveM.Z. Middle zoneNA Numerical apertureOA OsteoarthritisOD Optical densityPBS Phosphate buffered salinePCM Pericellular matrixPG ProteoglycanPI Propidium iodidePLM Polarized light microscopyRVD Regulatory volume decreaseSEM Scanning electron microscopySD Standard deviationS.Z. Superficial zoneTEM Transmission electron microscopyZnSe Zinc Selenide

LIST OF SYMBOLS
�� Donnan swelling pressure gradient�int Internal osmotic pressure�ext External osmotic pressure�int Internal osmotic coefficient�ext External osmotic coefficient�int Internal activity coefficient�ext External activity coefficientcext External salt concentrationcF FCDR Gas constantT Absolute temperatureTc Chemical expansion stressc- Concentration of mobile anions�f Fibril stressEf Fibril network modulus�f Fibril strainC Density ratio between the primary and secondary
fibrils�c,tot Fibril density�m The stress of the nonfibrillar phaseK Bulk modulusG Shear modulusF Deformation gradient tensorJ Determinant of FI Unit tensork Permeabilityk0 Initial permeabilitye0 Initial void ratioe Void ratio after compression�t Total stress inside the tissue�fi Stress of the individual fibrilstot f Total amount of fibrilsμf Electrochemical potential of interstitial fluidRlateral Lateral resolution� Wavelength

Emeasured Young’s modulus (calculated directly from theexperiments at equilibrium)
a Radius of the indenterh Thickness of the cartilage Poisson’s ratio Novel scaling factorI Intensity of the emerging lightI0 Intensity of the light illuminating the sampleI(0-90°) Intensity of light with polarizer at position 0-90°S0-4 Stokes parameters that characterize the polarized
state of light� Orientation angle of a polarization ellipsenf Fluid fraction

LIST OF ORIGINAL PUBLICATIONS
This thesis is based on data presented in the following articles,referred to by the Roman numerals I-IV.
I S.M. Turunen, M.J. Lammi, S. Saarakkala, A. Koistinen,and R.K. Korhonen, “Hypotonic challenge modulates cellvolumes differently in the superficial zone of intactarticular cartilage and cartilage explant,” Biomechanics andModeling in Mechanobiology 11, 665-675 (2012).
II S.M. Turunen, M.J. Lammi, S. Saarakkala, S.-K. Han, W.Herzog, P. Tanska, and R.K. Korhonen, “The effect ofcollagen degradation on chondrocyte volume andmorphology in bovine articular cartilage following ahypotonic challenge,” Biomechanics and Modeling inMechanobiology, Epub June 2012.
III S.M. Turunen, S.-K. Han, W. Herzog, and R.K. Korhonen,“Cell deformation behavior in mechanically loaded rabbitarticular cartilage 4 weeks after anterior cruciate ligamenttransection,” Osteoarthritis and Cartilage 21, 505-513 (2013).
IV P. Tanska, S.M. Turunen, S.-K. Han, P. Julkunen, W.Herzog, and R.K. Korhonen, “Superficial collagen fibrilmodulus and pericellular fixed charge density modulatechondrocyte volumetric behaviour in early osteoarthritis,”Computational and Mathematical Methods in Medicine 2013,164146 (2013).
The original articles have been reproduced with the kindpermission of the copyright holders.

AUTHOR’S CONTRIBUTION
The publications selected for this dissertation are originalresearch papers on the effect of the ECM and PCM onchondrocyte biomechanics following osmotic and mechanicalloading. The author was involved in the planning and thedesign of each study and is the main writer in the studies I-III,and a significant co-author in study IV. The author has done allthe measurements and analysis concerning cell volume andmorphology under osmotic and mechanical loading utilizingconfocal and dual-photon microscopy in studies I, III and IV,and participated in their performance in study II. In addition,the author has supervised the use of reference methods andparticipated in the analysis of the Fourier transform infraredmicrospectroscopy, polarized light microscopy and digitaldensitometry data in all studies. Furthermore, the author hasconducted all the statistical analyses and interpretation of datain studies I-III, and participated in these aspects of the work instudy IV.


Contents
1 INTRODUCTION .................................................................... 1
2 STRUCTURE AND FUNCTION OF ARTICULARCARTILAGE ................................................................................... 3
2.1 Extracellular matrix ............................................................. 32.1.1 Collagen ....................................................................... 42.1.2 Proteoglycans .............................................................. 52.1.3 Interstitial fluid ............................................................ 6
2.2 Chondrocytes ....................................................................... 62.3 Osteoarthritis (OA) .............................................................. 8
3 CELL-TISSUE INTERACTIONS IN ARTICULARCARTILAGE ................................................................................. 11
3.1 Cartilage response to osmotic and mechanical load ....... 113.1.1 Osmotic swelling and chemical expansion ............. 113.1.2 Dynamic and static behavior of cartilage ................ 12
3.2 Chondrocyte response to osmotic and mechanical load . 153.2.1 Isolated cells............................................................... 163.2.2 Cells in cartilage explants ......................................... 163.2.3 Cells in intact cartilage .............................................. 18
3.3 Cell-tissue interactions ...................................................... 18
4 IMAGING OF VIABLE CHONDROCYTES WITHMICROSCOPY ............................................................................. 21
4.1 Confocal microscopy ......................................................... 214.2 Dual-photon microscopy ................................................... 23
5 AIMS AND SIGNIFICANCE................................................ 25
6 MATERIALS AND METHODS ........................................... 276.1 Sample collection and processing ..................................... 286.2 Osmotic loading and CLSM microscopy ......................... 31

6.3 Mechanical loading and dual-photon microscopy .......... 336.4 FTIR microspectroscopy .................................................... 346.5 Polarized light microscopy ............................................... 366.6 Digital densitometry .......................................................... 386.7 Biomechanical testing ........................................................ 396.8 Computational modeling .................................................. 406.9 Statistical analysis .............................................................. 43
7 RESULTS ................................................................................. 457.1 Chondrocyte response to osmotic and mechanicalloading in situ .......................................................................... 45
7.1.1 Resting volume and morphology ............................ 457.1.2 Intact cartilage ........................................................... 457.1.3 Cartilage explant ....................................................... 467.1.4 Enzymatically degraded cartilage............................ 467.1.5 Osteoarthritic and contralateral joint cartilage ....... 47
7.2 Structural and mechanical analysis of tissue ................... 487.3 Finite element analysis ...................................................... 51
8 DISCUSSION ......................................................................... 538.1 Chondrocyte response to osmotic loading....................... 538.2 Chondrocyte response to mechanical loading ................. 558.3 Limitations and future prospects...................................... 56
9 SUMMARY AND CONCLUSIONS..................................... 619.1 Next steps toward better understanding of cartilagemechanotransduction .............................................................. 62
BIBLIOGRAPHY .......................................................................... 63

Dissertations in Forestry and Natural Sciences No 103 1
1 Introduction
Articular cartilage is needed in order to maintain thefunctionality of synovial joints [1-3]. It covers the ends ofarticulating bones, and is composed of the extracellular matrix(ECM), and chondrocytes [4]. The main components in the ECMare the collagen network, proteoglycans (PGs) and interstitialfluid [1, 3, 4]. Articular cartilage is subjected to various types ofmechanical loads during daily activities. The main functions ofcartilage are to 1) act as a distributor of the load on tounderlying bone, 2) absorb mechanical energy duringmovements, and 3) provide near-frictionless sliding betweenarticulating surfaces [1-6]. Cartilage degeneration inosteoarthritis (OA), leads to reduced capability of the tissue towithstand loads resulting in an impaired functionality of thejoint, as well as joint pain [2, 7, 8].
Chondrocytes in cartilage are responsible for producing andmaintaining the ECM [1, 2, 9]. In addition to genetic andenvironmental factors, proper mechanical stimulation ofchondrocytes and their normal functioning ensures a balancedsynthesis of the ECM, and thus, this maintains an appropriatestructure and composition for sustainable functioning ofcartilage [10-15]. Although cartilage degeneration in OA hasbeen widely studied and the accompanying structural andcompositional changes are comprehensively known, especiallyin the advanced stage of the disease, less is known about theconcurrent changes taking place in the biomechanical responsesof chondrocytes. In particular, the mechanisms at the cellularlevel in early OA still remain unknown. A profoundunderstanding of chondrocyte biomechanics and its relation tocartilage structure would provide a more detailedunderstanding of the processes involved in cartilagedegeneration during OA.

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
2 Dissertations in Forestry and Natural Sciences No 103
This thesis evaluated the effect of the ECM and pericellularmatrix (PCM) properties on chondrocyte biomechanics byapplying fluorescence microscopy with the tissue under osmoticand mechanical loading. More specifically, the effects of thesample integrity, enzymatic degradation of collagen, andstructural and compositional changes in early experimentally-induced OA on chondrocyte volume and morphology werestudied. The structure and composition of the samples wereanalyzed with quantitative microscopy and spectroscopy. Inaddition, finite element (FE) modeling was used for revealingthe specific phenomena behind the observed changes. Theresults of this study will improve the understanding of theinterrelationship between the structure of cartilage andchondrocyte biomechanics. Importantly, this will deepen theknowledge of the changes in cell behavior during cartilagedegeneration, which is crucial in order to be able to prevent,slow down or reverse the progression of OA.

Dissertations in Forestry and Natural Sciences No 103 3
2 Structure and functionof articular cartilage
Articular cartilage is a thin, hydrated soft tissue covering theends of diarthrodial joints [1, 2]. It provides low-friction glidingproperties and distributes loads applied to the joints [1, 2].Cartilage is an avascular and aneural tissue with a complex,highly organized structure, giving cartilage its uniqueproperties [1, 3, 4]. This chapter reviews the structure,composition, and mechanical properties of healthy articularcartilage. In addition, the structural and compositional changesin OA are also examined with special attention to changes incartilage tissue.
2.1 EXTRACELLULAR MATRIX
Articular cartilage structure is highly organized and it variesdepending on the tissue depth and on the anatomical site [3, 4].The ECM of articular cartilage contains mainly collagen, PGs,and interstitial fluid [1, 3, 4]. Cells (chondrocytes), that occupyonly 1-10 % of the tissue volume (depending on the species), areresponsible for synthesizing, organizing, and the maintenance ofthe matrix macromolecules [3, 4, 16].
Cartilage structure can be divided into three different zonesbased on the cell morphology and structure of the ECM [1, 4, 16].The relative thicknesses of each zone depend on the species, theanatomical site and the age of the subject: 1) superficial zone (3-24 %), 2) middle zone (1-40 %), 3) deep zone (50-94 %) [16-23].The structure and composition of these zones, and theinteraction between the ECM and interstitial fluid gives cartilageits functional properties [1-3, 6].

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
4 Dissertations in Forestry and Natural Sciences No 103
2.1.1 CollagenCollagens comprise 50-80 % of the dry weight and 10-22 % ofthe wet weight of cartilage [1, 6, 24]. Over six different types ofcollagen molecules (III, VI, IX, X, XI, XII, XIV) can be found inmature articular cartilage [24], but the principal form is type IIcollagen (90-95%) [1, 3, 6, 24, 25].
Collagen molecules have a triple helix structure consisting ofthree polypeptide chains (�-chains), each coiled into a left-handed helix, which are further wrapped around each other intoa right handed, super-helix [1-3, 6, 25]. The crosslinks betweenthe �-chains and the collagen super-helix provide strength andhigh tensile stiffness to the tissue [1].
The amount of collagen varies as a function of depth ofcartilage e.g. depending on the species, anatomical site and theage of the subject [23, 24, 26]. Collagen molecules form a fiber-like structure, where collagen fibrils are organized in denselypacked fibers forming a network in cartilage [1, 25]. In thesuperficial zone, collagen fibers are oriented in parallel to thesurface [3, 24, 26]. They give high tensile strength and stiffnessfor the superficial zone cartilage, and also resist shear stresses [1,3]. In the middle zone, the collagen fibers start bending, and inthe deep zone they are organized completely perpendicular tothe surface [6]. Collagen fibrils anchor themselves into calcifiedmatrix in the calcified zone [6].
Collagen has many different ways to affect the mechanicalproperties of cartilage. First, the collagen network providescartilage with its tensile properties. The tensile stiffness of thetissue is dependent on the integrity of the collagen fibrilnetwork, the depth from the cartilage surface and theorientation of the collagen fibrils [27-31]. Second, the structureand integrity of the collagen network contributes significantly tothe dynamic compressive properties of cartilage [31, 32].However, the network itself is fairly weak in compressionbecause of the high slenderness ratio, and thus, the collagenfibers alone have less effect on the compressive stiffness ofcartilage [33]. Instead, the collagen network acts as a cage for thePGs in cartilage, resisting the swelling pressure caused by the

Structure and function of articular cartilage
Dissertations in Forestry and Natural Sciences No 103 5
negative fixed charges. Thus, the collagen network also limitsthe hydration of cartilage and ensures that there is a high PGconcentration inside the tissue [33, 34].
2.1.2 ProteoglycansProteoglycans occupy approximately 4-10 % of the wet weightof cartilage [1, 6]. The core of these macromolecules are proteins,to which glycosaminoglycan (GAG) chains (chondroitin sulfateand keratan sulfate) are covalently attached [1, 3, 6, 35, 36]. SinceGAG chains are negatively charged, they repel other negativelycharged molecules and attract cations and water [1, 3]. Themajority (90 %) of PG aggrecans form large PG aggregates incartilage, where PG aggrecans are bound to a longmonofilament chain of hyaluronan with the help of link proteins[1, 3, 36]. These large aggregates are anchored inside the ECMby the collagen network. Similarly to collagen, the amount of PGvaries as a function of the depth of the cartilage, depending onthe species, site and the age of the subject [37, 38].
The interaction of PGs, interstitial fluid and ions in cartilagecreates a swelling pressure, which has a major impact on themechanical properties of cartilage [1, 6, 36]. The difference in theion concentration between the ECM and the synovial fluidcreates a fluid pressure difference between these sites, calledDonnan osmotic pressure [33, 39]. The total swelling pressure isa sum of the Donnan osmotic pressure and chemical expansionstress. Chemical expansion, on the other hand, results from thePG aggregates, which contain negatively charged groups intheir subunits [6, 35]. When these ions interact with each other,they create repulsive forces and this leads to a tendency ofexpanding [6, 35, 36]. In equilibrium, the total swelling pressureis counteracted by the tensile strength of the collagen network [2,3, 33]. PGs also contribute to the compressive viscoelasticproperties of cartilage by maintaining the balance between theapplied pressure, osmotic pressure inside the tissue, and fluidflow [39-42].

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
6 Dissertations in Forestry and Natural Sciences No 103
2.1.3 Interstitial fluidAround 60-85 % of cartilage wet weight is fluid and itsinteraction with collagens and PGs greatly influences themechanical properties of cartilage [1, 3, 6]. Interstitial fluid ismainly water containing gases, proteins, metabolites, andcations, which balance the negative charges of PGs [3, 33, 43].The majority of the fluid is in the solution domain of the PGs,and the rest is either bound to collagen fibers or is inside thechondrocytes [1, 33].
One of the roles of interstitial fluid flow through cartilagematrix is to be the provider of nutrients [33, 44]. Furthermore,the hydration of cartilage and fluid flow are one of the majorfactors affecting cartilage mechanical properties under dynamicand static loading [39, 40, 45]. This is because the rate at whichthe interstitial fluid flows through the ECM defines thenonlinear behavior of cartilage [33, 39, 45]. This phenomenon isdependent on the osmotic pressure created by the PGs and thepermeability of the tissue [39, 40, 45]. More specifically, when incontact with aqueous solutions, the negatively charged groupsof PGs (fixed charge density, FCD) imbibe fluid until theresulting hydrostatic pressure balances the resisting stressescreated by the collagen network and external loads [2, 33, 46].Thus, the amount of the fluid is dependent on the PG andcollagen contents, and on the relationship between the externalload and the swelling pressure of the tissue [2, 33, 46]. Anequilibrium state is reached when the swelling pressure (therelative amount of PGs) is in balance with external stress andfluid flow has ceased [46]. In addition, interstitial fluidpressurization contributes greatly to the dynamic stiffness ofcartilage [47].
2.2 CHONDROCYTES
The cell density in articular cartilage is very low compared toother soft tissues varying between 1-10 % of the cartilagevolume depending on the species and the age of the subject [3,

Structure and function of articular cartilage
Dissertations in Forestry and Natural Sciences No 103 7
48]. Therefore tissue matrix is responsible for the mechanicalproperties of cartilage under compression, while the role of thechondrocytes is to produce and maintain the structure andcomposition of the matrix [1, 3, 9, 15].
Chondrocyte size, shape and density vary between differentcartilage zones [3, 9, 16, 49]. Superficial zone chondrocytes areflattened, have an ellipsoidal shape with their major axesparallel to the surface. In the middle zone, the cells are roundedin shape and usually dispersed singly or in small groups. In thedeep zone, chondrocytes are usually grouped in columnsperpendicular to the surface, and the cell shape is eitherrounded or column-like.
The cytoskeleton of a chondrocyte comprises of proteins suchas actin, vimentin and tubulin [50], which work togetherdynamically, and are arranged into microfilaments,intermediate filaments and microtubules, respectively. Thebiomechanical properties of chondrocytes are affected by thecytoskeleton [50, 51]. The behavior of a chondrocyte has beenshown to be viscoelastic [50, 52, 53], and specifically themicrofilaments and intermediate filaments in addition to fluid-solid interactions and fluid viscoelasticity are predominantlyresponsible for determining these properties [50]. Thecytoskeleton of the chondrocytes and ECM are connectedthrough integrins [54]. Inside the cell, the integrins are attachedto the actin microfilaments [55].
The chondrocyte membrane is surrounded by a PCMconsisting of a collagen fibril shell (types II, VI and IX),relatively high concentration of PGs, and fluid [3, 16, 56-64].Chondrocyte and PCM together form a chondron [16]. The PCMabsorbs and transduces mechanical loads and provideshydrodynamic protection to chondrocytes. It also regulatesbiomechanical, biophysical and biochemical interactionsbetween the cell and the ECM [65-69].
The metabolic activity of chondrocytes is highly dependenton the environmental factors and mechanical forces experiencedby the cells, and those will affect the maintenance of thecartilage. The loading frequency and magnitude of loading are

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
8 Dissertations in Forestry and Natural Sciences No 103
two important factors affecting the chondrocyte activity [11-15].An increase or a decrease in matrix synthesis has been observed,depending on the duration and magnitude of the load [12-14, 70,71] and the mechanical environment of the cells [72]. In addition,the shape of the cells has been found to have an effect onchondrocyte biochemical activity [73-75].
2.3 OSTEOARTHRITIS (OA)
Osteoarthritis is the most common degenerative joint disease inolder people evoking pain, stiffness, and loss of mobility,consequently decreasing the quality of life [2, 76-78]. Theetiology of OA is still unknown but epidemiological risk factorsfor primary OA are well known; these include systemic (e.g.,age, gender, genetics) and local factors (e.g., obesity, jointinjuries) [79, 80]. Primary OA is the most common type of OA inhumans, whereas post-traumatic or secondary OA is typically aconsequence of a joint injury.
In OA, inadequate repair results in a gradual loss of articularcartilage accompanied by thickening of the subchondral boneand formation of osteophytes [2, 7]. The first signs includecartilage hydration and swelling [81-84], fibrillation of thecollagen network and reduced PG content [8, 81-86]. At laterstages of OA, the fibrillation becomes more profound, the PGdepletion reaches the deep zone, the thickness of the cartilagediminishes, the tissue becomes hypocellular and the integrity ofcollagen network decreases [8, 85, 87].
Collagen fibrillation in the superficial cartilage is one of thespecific features of early cartilage degeneration in OA [85, 87,88]. In addition, the diameter of the collagen fibers becomessmaller [85]. The degeneration of collagen network enables thePGs to escape the cartilage matrix, and this leads to a reductionin the matrix stiffness and a diminished ability to withstandcompressive loads [89, 90]. Collagen degradation also affects theexpelling of the fluid out from the matrix during compression,and it affects the remaining hydration level, which is related to

Structure and function of articular cartilage
Dissertations in Forestry and Natural Sciences No 103 9
contact stresses, and further possible damage to the underlyingbone [89-91].
Even a small change in the amount of PGs in the tissueresults in changes in the cartilage mechanical properties [39, 90,92]. However, even before any changes in the amount of PGscan be detected, PGs become more easily extractable at least inexperimental OA [81]. This is a sign of a loosening of the PG-collagen interaction. The relative amount of PGs in aggregateform also decreases [81, 90, 93, 94] and this further affects themechanical properties of cartilage, since it has been found thatPG aggregates are more viscous and have a higher shearmodulus compared to PG subunits [93]. In the later stages of OA,the amount of PGs decreases also in the deep zone [85].
Since cartilage tissue is not innervated by nerves, thesymptoms of OA are not due to the damage in cartilage tissueitself but due to related phenomena in the adjacent tissues [1, 3,4, 95]. This is one of the reasons why OA diagnosis is usuallymade too late, i.e., when cartilage is already severelydegenerated. Currently OA is diagnosed by a clinicalexamination, and often confirmed with X-ray imaging, magneticresonance imaging or arthroscopy [78].
Although OA has been widely studied in the past, thechanges in cell biomechanics in the early stages of the diseaseand the effect of cartilage degeneration on the cells still remainunclear. If it were possible to clarify the phenomena that occurduring articular cartilage degeneration in both cell and tissuelevels then novel insights could be gained into the processesunderlying in this disease. In the future, this might help in thefight against the progression of OA, or even in preventing thedisease.

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
10 Dissertations in Forestry and Natural Sciences No 103

Dissertations in Forestry and Natural Sciences No 103 11
3Cell-tissue interactions inarticular cartilage
3.1 CARTILAGE RESPONSE TO OSMOTIC AND MECHANICAL
LOAD
The response of articular cartilage to osmotic and mechanicalloads depends on the cartilage structure and composition [32, 92,96-101]. When articular cartilage is mechanically compressed,the interstitial fluid is released into the junction gap of the joint,the tissue becomes dehydrated and there are changes in theosmotic environment in the ECM [15, 39, 102]. The decreasinghydration results in denser packing of PGs and collagen. Whenthe load is removed, the water is imbibed back into the ECMand the osmolarity subsequently decreases. Degradation ofmatrix molecules during the progression of OA has also beenreported to result in decreased osmolarity of cartilage [82]. Thus,chondrocytes are exposed to a changing osmotic environmentduring mechanical compression and also in conditions ofcartilage degeneration, and these changes can be simulated withosmotic loading [40, 103, 104]. In this chapter, osmotic andmechanical behavior of cartilage and chondrocytes arediscussed in detail.
3.1.1 Osmotic swelling and chemical expansionThe swelling pressure of articular cartilage is attributable to thefixed negative charges of PGs due to two different mechanisms.Since the PGs in articular cartilage contain negative charges,positive counter-ions must be present in order for the tissue tobe electronically neutral [39, 105]. However, when there is animbalance between the internal ionic strength of cartilage tissueand the external bath, a swelling pressure is created. This leads

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
12 Dissertations in Forestry and Natural Sciences No 103
to diffusion of ions and fluid flow until an equilibrium isreached. This phenomenon can be described with the Donnanswelling pressure [39, 106]:
extint ��� ��� , (1)
where �int and �ext are the internal and external osmoticpressures. The same osmotic swelling pressure gradient can beexpressed also with the following equation [107]:
� �� � extext
2ext2
int
2ext2
Fint RTc�c�
�cRT��� 24 �
���
�
�
� �
�
�
, (2)
where �int and �ext are internal and external osmotic coefficients,�int and �ext internal and external activity coefficients, cext theexternal salt concentration, cF the FCD, R the gas constant, and Tthe absolute temperature.
The molecular interactions inside the cartilage tissue alsocontribute to the tissue swelling through a chemical expansionstress [6, 35, 36]. The negative charges of the PGs create arepulsive force and the chemical expansion stress can bedescribed with the following equation [108]:
� ����
�
� �� ��
�
�
Fext
Fc ccccaTint
0 exp��
� , (3)
where a0 and are material constants and c- the concentration ofmobile anions.
3.1.2 Dynamic and static behavior of cartilageBecause of the complex structure of cartilage, the compression ofcartilage results in inhomogeneous deformation within thetissue depending on the site and tissue depth [31, 92, 99, 101,109]. In addition, the loading rate and mode affect the responseof cartilage [31, 96].

Cell-tissue interactions in articular cartilage
Dissertations in Forestry and Natural Sciences No 103 13
Under dynamic mechanical load, where the load is appliedrapidly, the fluid inside the matrix has no time to flow throughthe matrix. The tissue can then be considered as beingincompressible and a high fluid pressure is created inside thecartilage [1]. A rapid change in the shape occurs and there is are-arrangement of the collagen network without any alterationsin the tissue volume [31, 110, 111]. The collagen network resiststhe change in shape and is, therefore, the main solid componentcontributing to the cartilage dynamic response [27, 31, 92, 110].
In mathematical form, when the collagen fibrils are assumedto resist only tension, the fibril stress has earlier been defined tobe either elastic or viscoelastic. The elastic fibril stress can bedescribed as:
�f = Ef�f, �f > 0 (4)�f = 0, �f � 0
where Ef is the fibril network modulus and �f the fibril strain.Formulas for the strain-dependent fibril network modulus andviscous properties of the collagen fibrils have also beenpresented [112-115]. In some studies, the collagen fibrils havebeen separated into the organized, primary fibrils andunorganized, secondary fibrils. The primary and secondaryfibril stresses, respectively, are defined as follows:
�f,pri = �c,tot �f (5)�f,sec = �c,tot�f (6)
where C is the density ratio between the primary and secondaryfibrils [116] and �c,tot is the fibril density [112].
In conditions of long term static mechanical compression, theinterstitial fluid is squeezed out of the ECM, resulting in a largedeformation of the tissue under creep loading [1, 39]. Duringloading, the rate of the fluid flow decreases and finally the tissuereaches equilibrium. Then the fluid flow has ceased, tissuedeformation is constant and the applied load is being balancedby the FCD of the PGs and collagen tension [1, 33, 117]. The

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
14 Dissertations in Forestry and Natural Sciences No 103
nonfibrillar matrix with osmotic swelling and chemicalexpansion (contribution of PGs) primarily controls the cartilageresponse at equilibrium. The nonfibrillar phase of the matrix hasbeen typically modeled as a linear Hookean poroelastic or anonlinear Neo-Hookean porohyperelastic material. The stress ofthe Neo-Hookean material can be defined as follows [112, 116]:
)J(JG
Jln(J)K 2/3T
m IFFI �� �� , (7)
where K and G are bulk and shear moduli, J is the determinantof the deformation gradient tensor F and I is the unit tensor.
The permeability of the tissue is dependent on the tissueporosity and void-ratio (ratio of fluid to solid content [118, 119]),and it can be described as:
,eekk
M
00 ��
�
�
�
�11 (8)
where k0 and e0 are the initial permeability and void ratio, e thevoid ratio after compression, and M a positive constantdescribing the depth-dependency of permeability.
Considering the swelling properties (3.1.1) and the fibrillarand nonfibrillar phases, the total stress inside the tissue can thenbe defined as follows:
��
��� �ftot
1ifcm
ift �T III��� , (9)
where �fi is the stress of the individual fibrils [112], tot f theamount of fibrils and μf the electrochemical potential ofinterstitial fluid [108, 120].

Cell-tissue interactions in articular cartilage
Dissertations in Forestry and Natural Sciences No 103 15
3.2 CHONDROCYTE RESPONSE TO OSMOTIC AND
MECHANICAL LOAD
When articular cartilage is mechanically compressed,chondrocytes within the tissue are subjected to a complexenvironment with time-varying changes [15, 15, 39, 102, 121,122]. The stress, strain, osmotic pressure, fluid-flow, fluid-pressure and electric fields affecting chondrocytes change bothspatially and as a function of time. Cartilage deformation undercompressive mechanical load results in deformation and volumechanges in the chondrocytes [21, 109, 122, 123], which should bedirectly affected by cartilage tissue properties as described byequations 1-9.
Chondrocyte volume regulation is vital for the functionalability of these cells [124, 125]. Volume regulation ofchondrocytes occurs in various situations; chondrocyte volumeis increased with aging and exercise, chondrocytes shrinkduring apoptosis, compressive loads flatten cells and thisnecessitates membrane stretch after which the volume is activelyreduced [124, 126]. It is still unclear what kind of role that thechondrocytes exert in the degeneration of cartilage, but thechanges in the volume regulatory processes could be one of thecontributing factors. Furthermore, it remains unknown whetherthe changes in the volume regulation mechanisms occurconcurrently with the cartilage degeneration, or whether theyresult from this process.
The increase in cell volume under hypotonic condition isbased on the permeability of the cell membrane to water, and apassive uptake of water with osmosis through aquaporinchannels [124, 126, 127]. Thereafter an active regulatory volumedecrease (RVD) takes place, where solutes are released fromcells, facilitating an osmotic efflux of water and thus, volumerecovery [125, 128].
Live chondrocytes in both osmotic and mechanical loadingtests are commonly visualized with fluorescence microscopy[129-135], but also stereological studies from tissues fixedhistologically after mechanical compression have been utilized

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
16 Dissertations in Forestry and Natural Sciences No 103
[10, 21, 72, 123]. The majority of the studies have used tissueexplants or isolated chondrocytes [129-132]. There are alsostudies where chondrocytes are left in their native environment[133-135]. Nonetheless, there is a controversy about whether thetissue preparation/integrity or the structure and composition ofthe ECM/PCM affect chondrocyte deformation behavior underosmotic and mechanical loading.
3.2.1 Isolated cellsWhen the response of isolated cells to osmotic loading isstudied, the cells are commonly placed on coverslips and aliquid with a known osmolarity is used as the medium.Hypotonic loading of isolated cells has resulted in an increase inthe cell volume [136], followed by a RVD where the originalvolume is finally reached [129, 131, 137]. It has been proposedthat isolated chondrocytes behave as perfect osmometers andthat the ECM has no effect on the chondrocyte response toosmotic loading [132].
The response of isolated cells to mechanical loading has beenwidely studied using isolated cells e.g. in agarose gels [138-140].The volume of isolated cells under static compression was foundto decrease and cell shape became more flattened [138, 140, 141].Interestingly, when ECM formation was stimulated during thecell culturing, there was a significant decline in the deformationof the cells under static loading [141]. In addition, the recoveryfrom the deformation took significantly longer when matrixaccumulation was induced [141]. The mechanical properties ofisolated cells have also been estimated with unconfinedcompression, micropipette aspiration and nanoindentation [122,142-144].
3.2.2 Cells in cartilage explantsThe chondrocyte response to osmotic loading has beenfrequently studied using cartilage explants with subchondralbone attached [131, 132]. Thin slices of cartilage are usually cutperpendicularly to the cartilage surface and chondrocytes indifferent layers can then be studied three-dimensionally (3D) e.g.

Cell-tissue interactions in articular cartilage
Dissertations in Forestry and Natural Sciences No 103 17
with fluorescence microscopy. Similarly to situation in isolatedcells, chondrocytes in cartilage explants have been shown toinitially increase their volume [132] and thereafter return to theoriginal levels [131]. The rate of RVD after hypotonic challengeusing cartilage explants has been shown to be independent ofthe level of the loading and the cartilage layer studied [131].However, cell behavior without a clear RVD in cartilageexplants has also been detected [145]. In addition, the level ofcartilage degeneration has been found to affect chondrocyteswelling after osmotic loading, i.e., the maximum swelling ofisolated cells and cells in degenerated cartilage explants wasgreater than that of cells in nondegenerated cartilage explants[129].
Mechanical loading of cells in cartilage explants is a commonway to study chondrocyte mechanobiology. Chondrocytes infull-depth disks, plugs or slices of articular cartilage have beenfound to become flattened after static mechanical compressionof the cartilage tissue, i.e., the aspect ratio of the cells issignificantly reduced [10, 21, 72, 146]. After removing thecompression, the chondrocytes have been found to recover backto their original volume and morphology [146]. Experimentaldynamic loading in combination with FE modeling of cartilageplugs has also been shown to deform the chondrocytedimensions principally perpendicularly to the loading direction[147]. Cell volume and height have been found to deformdepending on the cartilage layer under study, with thedeformation being largest close to the surface [72, 109]. Inaddition, the deformation of chondrocytes has been found to begreatest in the lateral direction perpendicular to the local split-line pattern [109, 148]. The nucleus of a chondrocyte has alsobeen shown to compress simultaneously with the cell [10, 148].
Although cartilage explants are frequently used for studyingin situ cell mechanics, there are still justified concerns about theeffect of specimen preparation and cartilage integrity on theresults. In particular, the boundary conditions at the sampleedge due to cutting are expected to be changed, and this couldaffect the cell responses [21].

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
18 Dissertations in Forestry and Natural Sciences No 103
3.2.3 Cells in intact cartilageWhen chondrocytes in fully intact rabbit patellar cartilage underosmotic load were studied, the cell volumes first increased, butthey did not display any subsequent cell volume recovery [133,149]. This was speculated to result from the continuous, long-term tissue swelling which prevented cell volume recovery orallowed the cells to swell into their preferred volume.
Chondrocyte behavior under static mechanical compressiveload has been studied in healthy fully intact cartilage [21, 123,134, 135]. Chondrocytes have been found to primarily expandlaterally under mechanical loading, the deformations beinggreatest close to the cartilage surface and presumablyperpendicular to the split-lines. The cell volume and height havebeen shown to decrease in healthy cartilage under static load[134]. This has also been recently shown for in vivo forchondrocytes inside intact knee joints of mice [150].
3.3 CELL-TISSUE INTERACTIONS
Chondrocytes detect their mechanical environment throughmultiple biological and biophysical interactions with the ECM[122]. Since the PCM directly surrounds the chondrocyte, thisregion can also influence the signals that cells perceive [67, 151-155]. Currently there are only a few experimental studies of theeffect of the ECM/PCM on chondrocyte biomechanics [130, 131,144, 156]. Instead, the effects of articular cartilage composition,structure and mechanical properties on cell mechanics havebeen studied with computational modeling [69, 151, 152, 154,157].
Chondrocytes are physically connected to the ECM viareceptors (e.g., integrins [158, 159]) and thus, alterations in theECM and PCM should also be reflected in the cellularcharacteristics. This is supported by previous theoretical studieswhere the ECM and PCM were found to affect cell deformationbehavior [69, 151, 152, 154, 157]. Thus, it can be hypothesizedthat when cartilage tissue is exposed to osmotic loading,

Cell-tissue interactions in articular cartilage
Dissertations in Forestry and Natural Sciences No 103 19
swelling pressure and chemical expansion of the cartilage tissue(equations 2 and 3) presumably affect the behavior of thechondrocytes. Similarly, under dynamic loading the collagennetwork stress of the tissue (e.g., equation 4) is hypothesized tobe the main component affecting cell level deformations, and instatic compression the swelling pressure caused by the PGs(equations 2 and 3) and the nonfibrillar matrix stress (e.g.,equation 7) presumably accounts for the strains and stressesexperienced by cells.

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
20 Dissertations in Forestry and Natural Sciences No 103

Dissertations in Forestry and Natural Sciences No 103 21
4 Imaging of viablechondrocytes withmicroscopy
Laser scanning microscopy is a 3D tool which can be used toobtain structural and functional data of viable cells in freshbiological tissues [160-163]. In this chapter the principles andadvantages of confocal and dual-photon laser scanningmicroscopies and their applications in cartilage research andchondrocyte imaging are reviewed and discussed.
4.1 CONFOCAL MICROSCOPY
The basis for confocal microscopy lies within conventional lightmicroscopy, where a small focal volume is completelyilluminated and thus, the resulting image contains light comingfrom regions below and above the focal plane [162]. In confocalmicroscopy, any light coming from layers other than the focalplane is physically prevented which results in better axialresolution and allows optical sectioning, and thus, 3D imagedata. The first version of a confocal microscope was built from aconventional microscope, so that a lens identical to the objectivelens replaced the condenser lens [163]. Pinholes were used tolimit both the field of illumination and the field of view. Thesample was scanned by moving it laterally, and the light passingthrough both the specimen and the second pinhole was detectedwith a photoelectric cell. The photoelectric current was thenamplified and displayed on a cathode-ray oscilloscope.
The advantage of using lasers as the light source is that theycan produce light beams with a very high degree of

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
22 Dissertations in Forestry and Natural Sciences No 103
monochromaticity and the intensity can be rather high [163]. Ina common, modern version of a confocal laser scanningmicroscope, the light is first reflected from a dichroic mirror,after which it is directed to a set of scanning mirrors. With thehelp of these motor-driven mirrors, the whole sample can bescanned. The specimen is thus excited by the laser andfluorescence is created. The fluorescent light is then scannedwith the same mirrors, and passed through the dichroic mirror.A pinhole is used in front of the light detector to reject the axialout-of-focus light and the lateral overlapping of the signal. Afterthis, 3D illustrations of the 2D images can be created withappropriate software [161-163].
Fluorescent labeling of the studied samples is common whenutilizing confocal laser scanning microscopy [164]. Fluorescenceis a physical phenomenon where incident light is first absorbedby a molecule and then light of a different color (wavelength) isemitted. In particular, when molecules are hit by a photon, theycan absorb it and, thus, gain energy and cause an electron tomove to a higher discrete excited state. Thereafter the energy isusually dissipated rapidly (e.g., molecular vibrations and heat),and in order to regain the ground state, spontaneous emissionmight occur, i.e., light of a longer wavelength is emitted.However, there are also alternative ways for the molecule to loseits excess energy. The excited electron might undergo a spin flipinto to a triplet state. While the de-excitation from this state isrelatively slow, the molecules might be trapped in this state andthus, to reduce the effective concentration of the fluorophore.Photobleaching, on the other hand, is a phenomenon in whichthe fluorophore irreversibly changes into nonfluorescent species.This limits the total amount of fluorescent information that maybe extracted from the sample. Photobleaching is related toenvironmental factors, e.g., oxygen activity. It can be reducedusing high numerical apertures (NA) that use less excitationlight and a small magnification (a smaller area will be excited).Special fluorescence mounting media have been developed toreduce photobleaching when imaging fixed samples.

Imaging of viable chondrocytes with microscopy
Dissertations in Forestry and Natural Sciences No 103 23
The optical resolution of a microscope is related to the NA ofthe optical components and to the excitation and emissionwavelengths of the light [165]. The highest achievable lateralresolution of a conventional microscope is a function of thewavelength of the emitted light (�) and NA of the objective lens,defined by Ernst Abbe [163]:
NA0.5�Rlateral � . (10)
Clearly, the use of as high as possible NA results in the bestlateral resolution. With confocal microscope this limit can beexceeded. The axial resolution is always poorer than the lateralresolution.
Confocal laser scanning microscopy (CLSM) is a commonlyused method in cartilage research [109, 130-135, 166, 166-170]. Itis suitable for visualizing viable chondrocytes inside cartilagetissue as well as for imaging isolated chondrocytes. Changes inchondrocyte volume, morphology and surface area followingosmotic or mechanical loading can be studied with this method.
4.2 DUAL-PHOTON MICROSCOPY
The optical design of the dual-photon microscope is similar tothe confocal microscope [171, 172]. Dual-photon microscopyutilizes high intensity lasers with low power in order to excite afluorophore with two low-energy photons instead of one high-energy photon [173, 174]. A single low-energy photon does nothave enough energy to excite a fluorophore, but if two of themreach the molecule within 10-16 seconds, then the threshold forexcitation is fulfilled. The probability of this phenomenon is low,and it is proportional to the square of the light intensity [171,172]. This proves to be an advantage in microscopy, sincefluorescence can only happen in the region where the lightintensity is at its highest. Thus, optical sectioning or pinholedetectors are not needed.

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
24 Dissertations in Forestry and Natural Sciences No 103
Dual-photon microscopy has also been used in cartilageresearch [129, 150, 175], although less commonly than confocalmicroscopy, probably due to the higher costs of the laser source.However, there are clear advantages associated with its use infavour of confocal microscopy in cartilage research; it has beenshown to decrease photo-bleaching (accurate focusing of theexcitation light results in a smaller volume of imaging) and toimprove chondrocyte viability, and with optimized conditions,it is possible to achieve an increase in the penetration depth ofthe imaging [175].

Dissertations in Forestry and Natural Sciences No 103 25
5 Aims and significance
Although the responses of chondrocytes to osmotic andmechanical loading have been earlier studied, it is still notknown how the integrity, structure and composition of the PCMand ECM affect cell biomechanics. In this thesis, confocal anddual-photon microscopies were used to study viablechondrocytes in normal, enzymatically degraded andosteoarthritic cartilage. The use of Fourier transform infrared(FTIR) microspectroscopy, polarized light microscopy (PLM)and digital densitometry (DD), made it possible to obtaindetailed and depth dependent data on the structure andcomposition of the samples and this could be related to thechanges detected in cell volume and morphology. In addition,FE modeling was used to provide additional insights into thereasons behind the observed phenomena in early OA.Specifically, the objectives were to answer the followingquestions:
1. Does the sample preparation/integrity affect chondrocyteresponse when cartilage tissue is osmotically loaded?
2. Does collagen degradation have a specific effect on thechondrocyte response following osmotic loading of cartilage?
3. How is the structure of cartilage altered and how are the cellresponses to mechanical loading of the tissue changed in thevery early stage of experimentally-induced OA?
4. What is the role of the ECM and PCM matrices on cellresponses following mechanical loading in the very earlystage of OA?
This study will improve the knowledge on theinterrelationships between cartilage structure and chondrocytebiomechanics in normal, degraded and osteoarthritic tissue.Importantly, in addition to the specific effects of the different

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
26 Dissertations in Forestry and Natural Sciences No 103
constituents of cartilage tissue on the cell biomechanics, thisthesis reveals changes in cell biomechanics in the very earlystage of OA. Hopefully this knowledge will form a platformfrom which to develop methods which can detect these cell levelalterations at the very early stage of OA and perhaps help todevise novel strategies to slow down or prevent the progressionof OA.

Dissertations in Forestry and Natural Sciences No 103 27
6 Materials and methods
This thesis consists of four independent studies. The materialsand methods used are summarized in Table 1. The main focus ofall studies was to reveal changes in chondrocyte volume andmorphology following osmotic (studies I and II) and mechanical(studies III and IV) loading of the tissue, and their relation totissue structure, composition, and biomechanical properties.Chondrocyte volume and morphology were assessed withconfocal and dual-photon laser scanning microscopy. Collagencontent, collagen orientation angle and PG content of thesamples were characterized with FTIR microspectroscopy, PLMand DD, respectively. In addition, biomechanical testing wasused in study II to quantify the biomechanical properties of thetissues. FE modeling was used in study IV in order to reveal thespecific influence of the PCM and ECM on chondrocyte volumeand morphology.
Table 1. Summary of the materials and methods used in studies I-IV.Study Samples Study purpose Methods
I Bovine LPG
The effect of sample
integrity on chondrocyte
biomechanics
CLSM, FTIR
microspectroscopy
II Bovine LPG
The effect of collagen
degradation on
chondrocyte
biomechanics
CLSM, FTIR
microspectroscopy,
PLM, Biomechanical
testing
III Rabbit patella
The effect of early OA
on chondrocyte
biomechanics
Dual-photon
microscopy, FTIR
microspectroscopy,
DD, PLM
IV Rabbit patella
Theoretical study about
the reasons behind
changes in chondrocyte
biomechanics observed
in study III
FE modeling
The abbreviations used in Table 1 can be found in the List of abbreviations.

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
28 Dissertations in Forestry and Natural Sciences No 103
6.1 SAMPLE COLLECTION AND PROCESSING
In studies I and II, bovine knee joints were used (ages 17-21 and17-32 months, respectively). Cartilage samples with theunderlying bone attached were collected from the lateralpatellar groove (LPG) of the joints on the day of slaughter. First,a drill bit (diameter = 21 mm) was used for obtaining cylindricalosteochondral plugs, which were then cut out from the kneejoint with an autopsy saw (Stryker Autopsy Saw 868, StrykerEurope BV, Uden, Netherlands). Two smaller samples werethen further extracted from these plugs using a metallic punch(diameter = 10 mm). Samples were kept in 37°C Dulbecco’smodified Eagle’s medium (DMEM, EuroClone S.p.A., Italy, 285-300 mOsm) until they were measured.
In study I, one of the osteochondral plugs was kept intact,while the other one was cut in half perpendicularly to thecartilage surface (Figure 1). The intact plug was used forstudying cell biomechanics through the intact surface. One halfof the cut plug was used for studying the chondrocytebiomechanics through the cut surface, while the other half wasused for structural analysis (collagen and PG contents), and wasfrozen immediately after the preparation in PBS solution (-20°C).
In study II, collagenase enzyme (30 U/ml, high puritycollagenase type VII (C0773), Sigma-Aldrich, St. Louis, MO,USA) was used in order to generate collagen degradation in oneof the osteochondral plugs (Figure 1). Both samples wereincubated for equal amounts of time; one of the plugs wasimmersed in DMEM and the other placed in a mediumcontaining both DMEM and collagenase.
In studies I and II, calcein-AM was used for fluorescencelabeling of the chondrocytes. Calcein-AM is a nonfluorescentand membrane-permeant chemical substance. Inside the cell, theesterases release the fluorophore, calcein, which is then trappedinside the cell [167]. The samples were immersed in a mediumcontaining DMEM (EuroClone) and 5 μM calcein-AM(Invitrogen, Eugene, OR, USA) for 30 minutes at room

Materials and methods
Dissertations in Forestry and Natural Sciences No 103 29
temperature. In addition, 50-60 μM propidium iodide (PI,Sigma-Aldrich) was added for detecting the dead cells. It ismembrane-impermeant, and it gives a red fluorescence onlywhen bound to nucleic acids [167].
Figure 1. Overall schematic presentation of studies I and II. a) (i) In study I theCLSM measurements were conducted through intact surfaces of cartilage samples(Group 1) and through cut surfaces of cartilage explants (Group 2) before and afterosmotic loading. In study II the CLSM measurements were conducted before and afterosmotic loading using intact (Group 1) and degraded (Group 2) cartilage. (ii) Typicalmicroscopic images of the calcein-AM stained samples, and (iii) a 3D presentation of achondrocyte. Typical (b) FTIR microspectroscopy and (c) PLM images for determiningthe PG and collagen contents and collagen orientation, respectively, in studies I and II(collagen orientation only in study II). S.Z. = superficial zone, M.Z. = middle zone,D.Z. = deep zone.

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
30 Dissertations in Forestry and Natural Sciences No 103
After the measurements (see CLSM microscopy below), thesamples from study II were immersed in isotonic DMEM (285mOsm) for one hour and afterwards cut in half. One half wasused for the estimation of tissue composition (collagen and PGcontents, see below) with FTIR microspectroscopy and tissuestructure (collagen orientation) with PLM, see below. The otherhalf was used for biomechanical testing, see below.
In studies III and IV, anterior cruciate ligament transection(ACLT) of rabbits was used to model OA (Figure 2). UnilateralACLT was conducted on the right knees of 10 female NewZealand white rabbits (age 14 months, weight 5.4 ± 0.6 kg),while the contralateral side was not subjected to any surgicalprocedure. The patellae from both knees were dissected fourweeks after ACLT (29 ± 1 days). These procedures were carriedout according to the guidelines of the Canadian Council onAnimal Care and were approved by the committee on AnimalEthics at the University of Calgary.
The ECM and PCM of the samples were labeled with afluorescent dye (Dextran, 3 kDa molecular weight, Invitrogen,Molecular Probes, OR, USA) suspended in DMEM (Dulbecco’sModified Eagle’s Medium, Gibco, OR, USA) for four hours at4ºC (Figure 2). After staining, the samples were washed in PBStwo times for ten minutes in order to remove the excess dye.Thereafter, the patellae were attached to a sample holder withdental cement and a self tapping bone screw (Fine Science Tools,North Vancouver, BC, Canada). During the entire procedure thesamples were kept moist with PBS.

Materials and methods
Dissertations in Forestry and Natural Sciences No 103 31
Figure 2. Overall schematic presentation of study III. a) (i) Schematic presentation ofthe mechanical indentation system mounted on a dual-photon microscope, (ii) typicalmicroscopic images of the Dextran stained samples, (iii) a 3D presentation of achondrocyte, and typical (b) DD, (c) FTIR microscpectroscopy and (d) PLM imagesfor determining the PG content, collagen content and collagen orientation, respectively,of the samples.
6.2 OSMOTIC LOADING AND CLSM MICROSCOPY
In studies I and II, CLSM was used for studying the chondrocyteresponse to osmotic loading in bovine LPG cartilage (Table 1,Figure 1). The microscope used was a Nikon Eclipse TE-300(Nikon Co., Tokyo, Japan) with an UltraVIEW confocal laserscanner (PerkinElmer, Oxford, UK). The cells were excited witha laser beam of 488 nm wavelength, and the emitted calceinfluorescence (515 nm) was measured with a 500-550 nm bandpass filter. The objective used had a magnification of x40 andx60 in studies I and II, respectively.

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
32 Dissertations in Forestry and Natural Sciences No 103
A reference image stack was first captured as the sampleswere immersed in isotonic DMEM (285-300 mOsm). Thereafterthe solution was rapidly changed to be hypotonic (170-180mOsm), and image stacks were taken after different time points(study I: 2, 20 and 120 min, study II: 20 and 120 min). Theosmolarity of the solutions was checked with a freezing pointosmometer (Halbmikro-osmometer, GWB, Knauer & Co GmbH,Berlin, Germany). The pH of the solutions was alwaysmaintained at 7.4. The chondrocytes were imaged up to ~60-80μm in depth from the measurement surface. The xy-plane was672 x 512 pixels in both studies I and II with the pixel sizes of 0.3and 0.2 μm, respectively. The z-axis increment was 0.25 μm inboth studies. The measurements were conducted at roomtemperature.
The image stacks from each sample/time point wereimported to image analysis software ImageJ (National Instituteof Health, USA), and approximately ten cells from thesuperficial cartilage of each image stack were randomly pickedfor further analysis. The volumes of the cells were calculatedusing 3D reconstructions (Visualization Toolkit 5.2.0, KitwareInc., NY, USA) and a code written with Python programminglanguage. For calibration purposes, fluorescently labeledmicrospheres (7.32 μm in diameter, volume 205.4 μm3) wereimaged in order to determine a threshold that gives correct cellvolumes. A threshold of 40 % of the maximum intensity wasfound to yield correct values (206.4 μm3). In addition,background noise for each image stack was defined from thehistograms and reduced for each individual cell. Cellmorphology (height, width and depth) was analyzed usingMatlab R2007b (MathWorks Inc., Natick, MA, USA). The widthand depth (x- and y-directions) were defined as the minor andmajor axes parallel to the cartilage surface and perpendicular tothe z-axis. This was accomplished by defining the maximumdistances between cell edges in the horizontal plane, and the z-direction in the vertical plane.

Materials and methods
Dissertations in Forestry and Natural Sciences No 103 33
6.3 MECHANICAL LOADING AND DUAL-PHOTON MICROSCOPY
In study III, a dual-photon microscope coupled with amechanical indentation system was used in order to obtainimages of rabbit patellar chondrocytes before and aftermechanical indentation loading of cartilage [135] (Table , Figure2). The unique microscopy indentation system consisted of around light transmissible glass indenter (total diameter 2 mm)mounted on the stage of a dual-photon microscope (ChameleonXR infrared laser, Coherent Inc., Santa Clara, CA, USA) in frontof a water immersion objective (x40 magnification, 0.8 NA)(Figure 2). The xy-plane was 512 x 512 pixels, with a pixel size of0.41 μm. The image stacks were captured with 0.5 μm z-axisincrements up to ~60 μm in depth from the cartilage surface. Theanalyzed cells were located in the superficial cartilage, onaverage at 20 μm depth from the surface.
The calibration of the system was done using Dextran-stainedagarose gel with polystyrene microspheres (diameter 5.93 ± 0.05μm). The gel with microspheres was imaged with the samesetup as the samples, resulting in a correction factor for the z-axis based on the known microsphere diameter (knownmicrosphere diameter divided by the diameter in the images).
In the measurements, a small preload (0.1-0.2 MPa) wasapplied in order to assure good contact between the indenterand the sample. Then, the sample was compressed with a rate of10 μm/s until a contact pressure of 2 MPa was reached.Thereafter the displacement was held constant for 20 minutes,resulting in contact pressures of 0.71 ± 0.1 MPa and 0.78 ± 0.2MPa in the ACLT and contralateral groups, respectively, in theend of the experiment.
The image stacks were imported to ImageJ andapproximately ten cells from each stack were randomly chosenfor the analysis (n = 8 samples, N = 77-79 cells/group).Individual cells were tracked both before and after themechanical loading. Image thresholds were determinedindividually for each cell from intensity histograms by definingthe median value between the cell and the matrix in the middle

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
34 Dissertations in Forestry and Natural Sciences No 103
slice of the cell. The 3D reconstruction and the calculation of cellvolume and morphology were done similarly as in studies I andII, see above.
Cartilage thickness was estimated based on the data obtainedfrom the PLM measurements (see below). The compressioninduced tissue strain was obtained from the differential variablereluctance transducer data (DVRT; measures the distancesduring the mechanical compression). The global tissue strainwas then calculated by dividing the axial compression by thetotal tissue thickness. The local axial and transversal ECMstrains were defined by analyzing the distances of the centers ofof cells from the axial and transversal view, respectively, in bothunloaded and loaded states (individual cells were tracked).Briefly, the distance between recognizable pairs of cells werefirst detected in the unloaded configuration and then in theloaded configuration. The local axial ECM and transversalstrains were then calculated by dividing the difference in thedistances by the original distance. The Young’s modulus of thesamples was calculated using the following equation [176]:
� ���
�� �� 2
measured �
�aEE 12
, (11)
where Emeasured is the modulus calculated directly from theexperiments at equilibrium (stress divided by strain), a is theradius of the indenter, h is the thickness of the cartilage, is thePoisson’s ratio (0.16 [177]) and � is the novel scaling factor [178].
6.4 FTIR MICROSPECTROSCOPY
Fourier transform infrared microspectroscopy has been provento be a powerful tool to assess the composition of cartilage [87,88, 179-182]. In this thesis, FTIR microspectroscopy was used forestimating the spatially varying collagen content in studies I, IIand III, and PG content in studies I and II (Table 1, Figures 1 and2).

Materials and methods
Dissertations in Forestry and Natural Sciences No 103 35
Infrared (IR) spectroscopy is a technique where the structureof a material can be studied at the molecular level based on thevibrations of the atoms of a molecule [183]. Chemical bonds inmolecules absorb IR light at specific wavelengths, thus, differentchemical molecules in the tissue can be recognized with IRspectroscopy [183]. Specifically, when IR radiation is directedthrough a sample, the majority of the energy goes through it,but a fraction is absorbed at specific frequencies/wavelengthsrelated to the chemical composition present in the sample. TheIR light absorption is based on the change in the electric dipolemoment of the molecule during the vibration [183]. Theintensity of the absorption is proportional to the change in theelectric dipole moment e.g. due to bending or stretching of themolecular bonds [183]. Thus, the absorption peaks of thespectrum can be used to quantify the amount of differentchemical components.
In FTIR microspectroscopy, the spectrometer is coupled witha light microscope, which means that it is possible to study theabsorbance spectra at discrete points and create a 2D spectralmap of the compounds of interest in tissue sections. With thismethod, the relative amount of the molecules of interest can bequantified, in addition to their spatial distribution, orientation,and molecular nature. The radiation from the IR source is firstpassed through an interferometer before being transmittedthrough the sample and finally reaching the detector [183]. Thesignal is then amplified, converted from analogical to digitalform, and transferred to a computer for Fourier transformation.
In studies I and II, three 5 μm thick unstained slices fromeach bovine LPG cartilage specimen were prepared from frozensamples (samples were thawed and frozen in liquid nitrogen)with a cryomicrotome (Reichert-Jung Cryostat 2800 Frigocut-E,Nussloch, Germany). In study III, where the samples were fixedin formalin after the dual-photon microscopy, a decalcificationwas conducted with ethylenediaminetetra-acetic acid (EDTA),followed by dehydration in an ascending series of alcoholsolutions (50%, 70%, 80%, 95% and 100%). The remainingalcohol was cleared with xylene. Finally, the samples were

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
36 Dissertations in Forestry and Natural Sciences No 103
infiltrated with paraffin (Paraplast Wax, Lancer division ofSherwood Medical, Kildare, Ireland). Then, three 5 μm thickslices were deparaffinized with xylene and PGs were removedwith a hyaluronidase digestion (1000 U/ml hyaluronidase,Sigma-Aldrich, St. Louis, MO, USA), in order to study thecollagen content in detail [184]. In all studies, the slices wereplaced on Zinc Selenide (ZnSe) slides.
The FTIR equipment used was a PerkinElmer SpectrumSpotlight 300 spectrometer in transmission mode (PerkinElmer,Wellesley, MA, USA). From every sample, a rectangular areafrom the surface to the subchondral bone was imaged (Figures 1and 2, width ~ 500 μm, pixel size 6.25 or 25 μm, spectralresolution 4 cm-1, scans/pixel 2-8 in studies I, II and III).
The spectra were first baselined, i.e., the minimum value wassubtracted in order to correct the offset due to background.Characteristic absorbance bands were detected from the spectraand integrated in order to reveal the amount and distribution ofcollagen and PGs. Collagen concentration of cartilage wasestimated from the integrated area of amide I absorbance (1585-1720 cm-1), while the carbohydrate region (985-1140 cm-1) wasused to estimate the PG content similarly to previous studies [88,180, 185]. The values of each row of data parallel to the surfacewere averaged in order to obtain a depth-dependent profile ofthe cartilage composition (Figures 1 and 2).
6.5 POLARIZED LIGHT MICROSCOPY
Polarized light microscopy has been utilized in cartilageresearch in order to characterize the depth dependentorientation angle of collagen fibrils [19, 100, 186-189]. PLM isbased on the interactions between the matter and light. Light isan electromagnetic wave composed of both electric andmagnetic fields propagating through material perpendicularlyto each other. The molecular composition of a material affectsthe penetration of light, and this can be utilized in PLM in orderto study the structure of a sample [187, 190].

Materials and methods
Dissertations in Forestry and Natural Sciences No 103 37
Conventional PLM usually consists of two linear polarizers.The first polarizer is used for linearizing the incident,nonpolarized light. The second polarizer is perpendicular to thefirst one, and thus, ideally no light can travel through the system.When an anisotropic sample is placed between the twopolarizers, the plane of polarization is slightly shifted and lightis able to pass through the specimen. The amount of lightdetected depends on the material properties of the sample. Theintensity of the emerging light can be determined with Fresnelequation:
I = a + b I0sin2�, 0 �������/2 (12)
where I0 is the intensity of the light illuminating the sample, � isthe rotation angle of the polarization plane, and a and b areconstants dependent on the optical system.
Stokes parameters for each image pixel can be used tocharacterize the polarization phenomenon of a material [100, 187,191].
S0= I0 + I90 (13)S1= I0 - I90 (14)S2= 2I45 - S0 (15)S3= S0 - I(90+�/4), (16)
where I(0-90°) is the intensity of the light with the polarizer atposition 0-90°, S0 the total intensity of the light, S1 the amountlinear polarization, S2 the amount of ±45° polarization and S3 theamount of right or left circular polarization. Subscript �/4 refersto the image taken with a phase shifter.
The orientation angle of the collagen fibrils can then becalculated as follows:
2
���
�
�
� 1
2
SSarctan
� , 0° ����� 90° (17)

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
38 Dissertations in Forestry and Natural Sciences No 103
where � is the orientation angle of a polarization ellipse. Sincethe fibrils in articular cartilage split the light passing through itinto two components (one parallel and one perpendicular to thelong axis of the fibril), � can be used to estimate the averageorientation of collagen fibrils in each pixel.
In study II, frozen bovine LPG cartilage samples werethawed, and fixed in 10 % formalin. Thereafter microscopicslices were prepared similarly as for FTIR microspectroscopy(see above).
A polarized light microscope (Leitz Ortholux II POL, LeitzWetlar, Wetzlar, Germany) and a thermoelectrically cooleddigital camera (Photometrics SenSys, Roper Scientific, Tucson,AZ, USA) with a charge-coupled device (CCD, Kodak,Rochester, NY, USA) were used for the imaging and signaldetection. The system was customized for computation ofStokes parameters [100, 187, 192].
Seven images of each sample were imaged with x4magnification at 0°, 15°, 30°, 45°, 60°, 75°, and 90° polarizer pairpositions (polarizer-analyzer). An additional image wasrecorded at the 90° position with a phase shift plate (�/4) in thepath of light. 0°, 45°, 90° images, and the 90° image with a phaseshift were used in order to calculate the Stokes parameters (S0-S3,
equations 13-16, Figures 1 and 2).
6.6 DIGITAL DENSITOMETRY
Digital densitometry was used in study III for estimating the PGdistribution of the rabbit cartilage samples as a function ofdepth from the cartilage surface to the subchondral bone (Table1, Figure 2). PGs in articular cartilage contain negative groups,in which the cationic dye (Safranin O) binds to. It has beenpreviously shown that there is a linear relationship between theoptical density (OD) and the amount of PGs [193, 194].Therefore the distribution of PGs can be revealed and quantifiedwith this method [100] (Figure 2).

Materials and methods
Dissertations in Forestry and Natural Sciences No 103 39
Samples were processed similarly to FTIR microspectroscopy(see above), excluding deparaffinization and PG digestion. Then,the three micrometer thick slices were stained with safranin Oand placed on standard microscopic slides.
The measurement system consisted of a standard microscopewith an objective of x4 magnification. The calibration of thesystem was conducted with neutral density filters (Schott,Mainz, Germany) with different absorbance units (OD = 1, 1.3,1.6, 2, 2.6 and 3) by fitting a logarithmic curve into the values.Thereafter grayscale images were captured and converted intoabsorbance units. A rectangular area (~400 μm width) wasmanually chosen from the image for further analysis. Theprofiles were then averaged horizontally in order to estimate thedepth dependent PG content of the samples.
6.7 BIOMECHANICAL TESTING
Biomechanical stress-relaxation tests (unconfined compression)were used in study II for the quantification of the biomechanicalchanges in bovine LPG cartilage samples after collagenasetreatment (Table , Figure 3). A custom made high-resolutionmechanical testing device was used for the measurements [195].Briefly, the measurement setup consisted of a load cell unit(Sensotec, Columbus, OH, USA), a precision motion controller(PM500-C, Newport, Irvine, CA, USA) and a data acquisitionsystem.
Cartilage samples were thawed at room temperature beforethe measurements were conducted. First, the measurementchamber was filled with isotonic PBS, and the compression platewas driven to the bottom of the chamber in order to set theoffset position. Then the compression plate was elevated, thecartilage sample was placed in the chamber and an initialcontact was ensured by a change in the force sensor reading. Afull contact was then obtained by further lowering thecompression plate and this was confirmed visually.

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
40 Dissertations in Forestry and Natural Sciences No 103
Stress-relaxation tests were then conducted with 2 x 5 %strain, with a 30 minute relaxation period [115, 196] after bothsteps (2 mm/s compression rate). The reaction forces and axialdeformations were recorded continuously through the entireexperiment. The instantaneous and equilibrium modules werecalculated from the data (stress-strain ratio from the peak andequilibrium forces, respectively) (Figure 3).
Figure 3. a) A schematic presentation of the mechanical testing device and typical b)displacement and c) force curves in a stress-relaxation test.
6.8 COMPUTATIONAL MODELING
An axisymmetric FE model with poroelastic and swellingproperties as well as characterization of the collagen fibrils wasconstructed in order to simulate the experimental indentationmeasurements of study III (equations 1-9). The model was usedto elucidate the reasons behind the experimentally observed cellvolume behavior in rabbit patellar cartilage (Table 1).
The two major components in the model were the nonfibrillar(PGs and interstitial fluid) and fibrillar (collagen network)phases. The osmotic swelling and chemical expansion stresseswere taken into account with equations 2 and 3. The collagen

Materials and methods
Dissertations in Forestry and Natural Sciences No 103 41
network was modeled using elastic fibrils (equation 4) [69, 112,116, 152]: the primary fibrils obey the Benninghoff-like depth-dependent orientation [197], while the secondary fibrils arerandomly organized and also contribute to the crosslinking ofthe primary fibrils (equations 5-6). The nonfibrillar part wasimplemented as Neo-Hookean porohyperelastic material(equation 7), and the permeability was assumed to depend onthe void ratio (equation 8). The total stress in the model wasgiven by the equation 9.
The indenter was assumed to be rigid, and the modelconsisted of 1608 axisymmetric poroelastic elements which eachhad four nodes (Figure 4). The lateral movement of thesymmetry axis was set to zero and the bottom of the sample wasfixed. Free fluid flow was allowed at the edge of the sample andat the surface of cartilage not in contact with the indenter. First,the tissue was allowed to swell freely (osmotic swelling andchemical expansion). Thereafter, the indenter was moved intocontact with the cartilage surface, followed by a 2 MPacompression and a 20 minute relaxation period keeping thetissue strain constant. A submodel, consisting of 398 elements,including elements also for the PCM (80 elements) and cell (102elements), was constructed for the superficial zone. A user-defined material script was used for implementing the materialproperties to the model. All simulations were performed withAbaqus 6.11-2 (Dassault Systemés, Providence, RI, USA).

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
42 Dissertations in Forestry and Natural Sciences No 103
Figure 4. Finite element model consisting of a) a global model (ECM) and b) asubmodel (ECM, PCM and cell), and c) a presentation of the cell shape before and afterthe mechanical loading.
Both experimental data and results from previous studieswere utilized in order to implement the structural,compositional and mechanical parameters in the model (Table 2).The zonal thicknesses (7%, 18% and 75% for superficial, middleand deep zones) were determined from the PLM measurementsof the contralateral joints. Water, collagen and FCD contentswere obtained from previous studies [136, 157]. PCM stiffnesswas 90 % less and FCD content 1.27 times higher than those ofthe ECM [151, 152]. Collagen fibers were assumed to be orientedparallel to the cell surface in the PCM. Permeability wasmodeled as nonlinear and dependent on the void-ratio for theECM, and constant for the PCM and cell.
ACLT induced changes in cartilage samples wereimplemented into the ACLT model. These included reducedcollagen fibril stiffness, decreased FCD and increased fluidfraction. In addition, changes in the properties of the PCM,presumably present in OA, were implemented in the model [8,88, 198, 199]. Finally, cell volume changes following indentationtesting of ACLT and contralateral joint cartilage were simulated.

Materials and methods
Dissertations in Forestry and Natural Sciences No 103 43
Table 2. Mechanical, structural and compositional parameters of the ACLT andcontralateral joint models.
Parameter ACLT model Contralateral
model
Collagen stiffness
Ef(MPa)
ECM
PCM
Cell
7.5
0.1 x Ef (ECM)
-
10
0.1 x Ef (ECM)
-
FCD cF(mEq/ml)
ECM
PCM
Cell
0.08+0.12z
0.11+0.09z
0.08
0.11+0.09z
0.14+0.12z
0.08
Fluid fraction nf
ECM
PCM
Cell
0.9-0.2z
0.9
0.61
0.85-0.15z
0.85
0.61
6.9 STATISTICAL ANALYSIS
Study I: A mixed linear model with a Sidak correction was usedwhen comparing the differences in the cell volume andmorphology between different time points and groups. Themeasurement time point, sample type and their interaction werefixed variables, while the sample number was a randomvariable. The model takes into account that the cells were fromthe same animals, joints and samples, and that different cellsbetween groups and time points were compared. A Pearsoncorrelation coefficient was used for the determination of therelationship between PG/collagen contents and cellbiomechanical behavior. SPSS 14.0 (SPSS Inc., Chicago, IL, USA)was used for the analyses.
Study II: The same mixed linear model as in study I was usedin order to reveal the significant differences between the intactand degraded samples. Differences in the structure andbiomechanical properties were analyzed with a nonparametricWilcoxon signed rank test. SPSS 17.0 was used for the analyses.
Studies III & IV: In the comparison of the cell parametersbetween the groups, one way analysis of variance (ANOVA)was used. The structural differences were evaluated with aWilcoxon signed rank test. SPSS 17.0 was used for the analyses.

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
44 Dissertations in Forestry and Natural Sciences No 103

Dissertations in Forestry and Natural Sciences No 103 45
7 Results
7.1 CHONDROCYTE RESPONSE TO OSMOTIC AND
MECHANICAL LOADING IN SITU
7.1.1 Resting volume and morphologyIn study I, the chondrocyte resting volume, height and depth inthe superficial layer of cartilage explants before osmotic loadingwere significantly larger than the equivalent cells present inintact cartilage (p < 0.05). In addition, cell aspect ratios weresignificantly smaller in intact than in explant samples (p < 0.05).Enzymatic degradation of the collagen network did not affectsignificantly the resting volume or the morphology ofchondrocytes in study II (p > 0.05). In study III, chondrocyteresting volume, height, width and depth were significantlysmaller in ACLT as compared to contralateral samples (p <0.001).
7.1.2 Intact cartilageIn intact cartilage, cell volume and height were significantlygreater 20 minutes after the hypotonic loading compared to thecorresponding values before the loading (p < 0.05, studies I andII, Figure 5). After this, they either fully remained at the elevatedlevel (intact cartilage in study I), or displayed some volumerecovery, although not regaining the original values (intactcartilage in study II, p > 0.05). In study I, cell depth in intactcartilage followed the similar behavior as the volume andheight, while it did not change significantly in study II. Cellwidth did not change significantly following osmotic loading ineither of studies I or II (p > 0.05).

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
46 Dissertations in Forestry and Natural Sciences No 103
Figure 5. Normalized (a) cell volume, (b) height, (c) width and (d) depth before and 20min and 120 min after hypo-osmotic loading in intact cartilage (study I, n = 8, studyII, n = 7), cartilage explants (study I, n = 8) and degraded cartilage (study II, n = 7).*p< 0.05, **p < 0.001.
7.1.3 Cartilage explantIn cartilage explants (study I), cell volume, height and depthincreased significantly immediately after the osmotic loading (2min, p < 0.05). Thereafter, a significant decrease was detected (p< 0.05) and the cell dimensions reached their original valuesduring the 120 min measurement period (Figure 5).
The increase in cell volume after hypotonic loading wassignificantly greater in cartilage explants compared to intactcartilage (2 min, p < 0.05). After 120 minutes, the relative cellvolume and height were significantly greater in intact cartilagecompared to cartilage explants (Figure 5, p < 0.05).
7.1.4 Enzymatically degraded cartilageIn enzymatically degraded cartilage (study II), cell volume andheight had recovered back to their original values 20 minutesafter the hypotonic loading (Figure 5). The cell width decreasedsignificantly from 20 to 120 minutes (p < 0.05), completelyrecovering back to its original value. The cell depth remainedthe same throughout the experiment.

Results
Dissertations in Forestry and Natural Sciences No 103 47
Normalized cell volume and height were significantlysmaller in the degraded cartilage compared to intact cartilage 20minutes after osmotic loading (p < 0.001 and p < 0.05,respectively, Figure 5). Normalized cell volume remainedsignificantly smaller in the degraded cartilage also at 120 minfollowing the hypotonic loading (p < 0.05). Normalized cellwidth was significantly greater in the degraded cartilage ascompared to the intact cartilage samples at the 20 min time point(p < 0.05).
7.1.5 Osteoarthritic and contralateral joint cartilageAs a result of 2 MPa force-relaxation loading of rabbit patellarcartilage, the cell volume and height in the contralateral jointcartilage decreased significantly, while the cell width and depthincreased (Figure 6, p < 0.001). In the ACLT group, there weresignificant increases in cell volume, width and depth of thechondrocytes, while cell height decreased significantly (p <0.001). Importantly, the changes in cell volume (p < 0.001), width(p < 0.05) and depth (p < 0.001) were significantly larger in theACLT group as compared to contralateral group, while thechange in cell height (p < 0.05) was significantly greater in thecontralateral joint cartilage.
Figure 6. Normalized (a) cell volume, (b) height, (c) width and (d) depth before and atsteady state after mechanical loading of cartilage in the experiments (ACLT andcontralateral cartilage) and in the FE model (ACLT model and contralateral model).**p < 0.001.

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
48 Dissertations in Forestry and Natural Sciences No 103
7.2 STRUCTURAL AND MECHANICAL ANALYSIS OF TISSUE
When studying chondrocytes in intact bovine LGP cartilage, theamount of PGs in the superficial zone was found to correlatesignificantly (p < 0.05) with the cell volume at the 120 minutetime point. There were no correlations between the cellvolumetric behavior and the collagen content (p > 0.05).
The structure of the bovine LPG cartilage changed due to theenzymatic treatment in study II (Figure 7). The collagen contentin the superficial tissue was significantly decreased after the 24hours of collagenase digestion (0-10 %, p < 0.05 compared tointact cartilage). The orientation angle of the collagen fibrils wasalso altered: the enzymatic degradation resulted in moreperpendicular fibril orientation (more fibrillated collagennetwork) in the superficial tissue compared to intact,nondegraded cartilage (0-10 %, p < 0.05). Deeper zones remainedunaffected by the treatment (p > 0.05). PG content remainedunaltered through the entire depth (p > 0.05). The enzymatictreatment also affected tissue biomechanics. The instantaneousmodulus of the cartilage was significantly decreased inenzymatically degraded cartilage (p < 0.05). The equilibriummodulus did not change (p > 0.05).
The structure and composition of the rabbit patellar cartilagewas significantly changed 4 weeks after the ACL transection(Figure 8). Specifically, the collagen orientation angle waschanged in the superficial zone of cartilage due to the operation(0-9 %, p < 0.05). The PG content in the superficial cartilage wassignificantly lower in the ACLT group (0-24 %, p < 0.05)compared to the contralateral group. ACL transection did notaffect the collagen content of the samples (p > 0.05).

Results
Dissertations in Forestry and Natural Sciences No 103 49
Figure 7. (a) PG content (AU), (b) collagen content (AU) and (c) collagen orientationangle (degrees) in intact (red line) and degraded (black line) cartilage in study II as afunction of tissue depth, defined using FTIR microspectroscopy and PLM (n =7/group). AU = absorption unit.
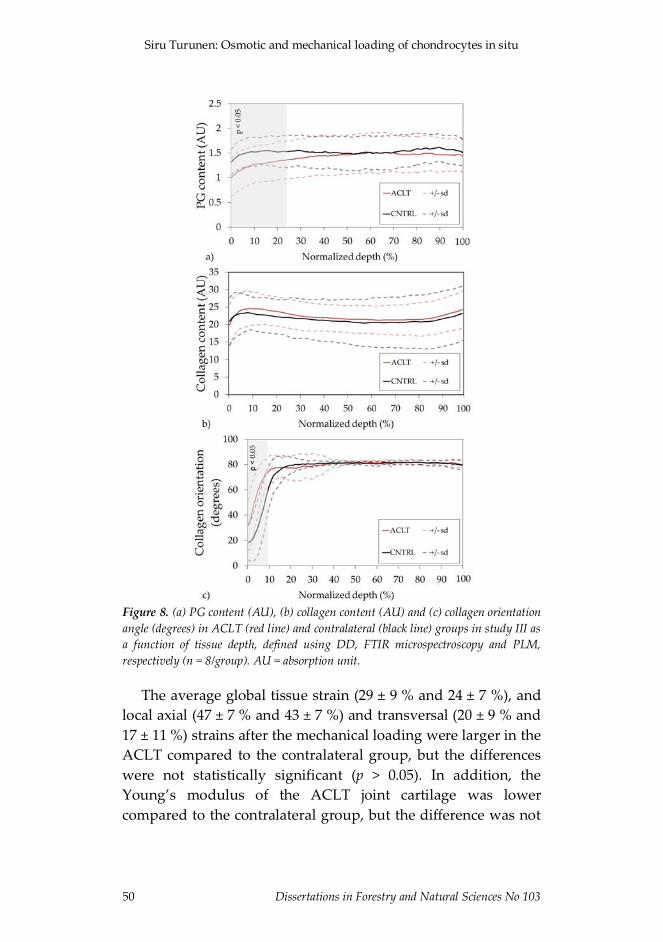
Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
50 Dissertations in Forestry and Natural Sciences No 103
Figure 8. (a) PG content (AU), (b) collagen content (AU) and (c) collagen orientationangle (degrees) in ACLT (red line) and contralateral (black line) groups in study III asa function of tissue depth, defined using DD, FTIR microspectroscopy and PLM,respectively (n = 8/group). AU = absorption unit.
The average global tissue strain (29 ± 9 % and 24 ± 7 %), andlocal axial (47 ± 7 % and 43 ± 7 %) and transversal (20 ± 9 % and17 ± 11 %) strains after the mechanical loading were larger in theACLT compared to the contralateral group, but the differenceswere not statistically significant (p > 0.05). In addition, theYoung’s modulus of the ACLT joint cartilage was lowercompared to the contralateral group, but the difference was not

Results
Dissertations in Forestry and Natural Sciences No 103 51
statistically significant (1.8 ± 0.7 MPa and 2.4 ± 1.2 MPa, p > 0.05,respectively).
7.3 FINITE ELEMENT ANALYSIS
Similarly to the experimental results in study III, cell volumewas found to increase after the mechanical loading in the FEmodel representing the ACLT joint cartilage, while a decrease inthe cell volume in contralateral joint cartilage was detected inboth experiments and simulations (Figure 6). Cell heightdecreased due to the loading in both groups in the experimentsand simulations (Figure 6). Cell width and depth increasedmore in ACLT as compared to contralateral group in bothexperiments and simulations (Figure 6). The local axial andtransversal tissue strains in the FE model were greater in theACLT group compared to the contralateral group, similarly tothe experimental results.

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
52 Dissertations in Forestry and Natural Sciences No 103

Dissertations in Forestry and Natural Sciences No 103 53
8 Discussion
The volume and morphology of chondrocytes in the superficiallayer of articular cartilage were studied under osmotic andmechanical loading in intact, enzymatically degraded andexperimentally-induced OA cartilage. Changes in tissueintegrity, the amount of collagen and PGs as well as fibrillationof the collagen network were found to alter chondrocytebiomechanics in their own distinct manners. In this chapter, theresults are discussed in more detail.
8.1 CHONDROCYTE RESPONSE TO OSMOTIC LOADING
The resting volumes of cells in intact cartilage were smallercompared to those in cartilage explants. Similarly, cellularresting volumes have been previously reported to be increasedin OA cartilage [130, 134]. One explanation for this phenomenonmight be that cells in intact cartilage are squeezed between thehorizontal collagen fibers [133, 149, 157], while cells in explantshave more space in their surroundings. In addition, the cellularresting volumes of enzymatically degraded cartilage were largerthan in intact cartilage, although the difference was notstatistically significant. Thus, it may be that a minor degree ofcollagen degradation alone does not affect cell resting volumes.
There was a significant difference in the cell behaviorfollowing hypotonic loading of cartilage depending on thesample preparation/integrity. In previous studies where eitherisolated cells or cartilage explants have been used, chondrocyteshave been shown to respond to hypotonic loading with anincrease in cell volume [129, 131, 132, 136, 137]. Afterwards avolume decrease has been observed and chondrocytes haveregained their original volumes [129, 131, 131, 137]. In thecurrent experiments, the same behavior was observed for

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
54 Dissertations in Forestry and Natural Sciences No 103
chondrocytes in cartilage explants: at 2 minutes the cell volumeswere highly elevated, but after 20 minutes following osmoticloading they had returned to their original levels. Similarly, ourresults showed that chondrocytes in enzymatically degradedcartilage also regained their original volumes within 20 minutesafter the osmotic loading. In contrast, the response of thechondrocytes to osmotic loading in intact cartilage wassignificantly different. First, an increase in the cell volume wasdetected, however, the subsequent reduction in volume waseither completely missing or not so profound when compared tothe cartilage explants or degraded samples. Importantly,original cell volumes were not reached in intact samples evenafter two hours, similarly to the results from a previous study[133]. Additional morphological analysis revealed that thedifference in cell behavior between intact cartilage and cartilageexplants/degraded cartilage was primarily explained by thechange in cell height. This might be due to the loosening of thecollagen fibril tension, which then allowed the cells toexpand/shrink more easily in the samples where the collagennetwork integrity was compromised.
When the structure of the samples was studied in detail, FTIRand PLM analyses revealed that the enzymatic treatment hadreduced significantly the collagen content in the top 10 % of thecartilage thickness, and that the collagen network was fibrillatedin the superficial layer. These results further reinforce theputative explanation for the difference seen in the chondrocytebehavior between intact and degraded cartilage, i.e., thecollagen content and integrity of the collagen fibril networkprobably contributed to cell biomechanics.
Based on the current results, it is postulated that the recoveryin the cell volume after the initial swelling following hypotonicloading may have been partially restricted by the ECM/PCM inintact samples. One possible explanation could be that since cellmembrane receptors, e.g., integrins [158, 159], are physicallyattached to the ECM/PCM, and osmotic loading is known toresult in swelling of the ECM which can last for hours [133, 200],these physical connections may restrict or slow down

Discussion
Dissertations in Forestry and Natural Sciences No 103 55
chondrocytes to regain their original volumes. This is supportedby the finding that the PG content was found to correlatepositively with the change in the cell volume in intact cartilage,indicating that elevated swelling pressure caused by the PGsmay increase tissue swelling and prevent at least partly the cellvolume recovery. On the contrary, in cartilage explants anddegraded cartilage, these physical connections might be weaker,and consequently volume regulation would happen easier orfaster. This is supported by a former study where the structureof the ECM was found to affect the expression of the adhesiveintegrins [159].
8.2 CHONDROCYTE RESPONSE TO MECHANICAL LOADING
The chondrocyte response to mechanical indentation loadingwas found to be significantly different in the ACLT jointcartilage compared to the contralateral joint cartilage.Chondrocyte volume increased in the ACLT cartilage, while itdecreased in the contralateral cartilage following mechanicalloading. The morphological analysis revealed that the differencein the behavior was due to the significantly larger changes in thecell dimensions parallel to the cartilage surface in the ACLTgroup compared to the contralateral group. Interestingly, cellsin the contralateral group were compressed significantly morethan those in the ACLT group, and at the same time thehorizontal changes were smaller. These results are similar to aprevious study, where chondrocyte volumes were also found toincrease following mechanical compression 9 weeks after ACLT,and cells in normal cartilage were found to decrease in volume[134]. The detailed structural and FE analysis revealed that thedecrease in the amount of FCD of the tissue and PCM and theorientation and stiffness of the collagen fibers in the ECMprimarily explained the changed cell volumetric behavior inearly OA.
As a result of the ACLT, structural and compositionalchanges occurred in the patellar cartilage. While the amount of

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
56 Dissertations in Forestry and Natural Sciences No 103
collagen was found to be similar in both the ACLT andcontralateral groups, the collagen orientation in the superficialcartilage became more perpendicular to the surface as comparedto contralateral cartilage. In addition, the amount of PGs wasreduced in the ACLT group. Despite the clear changes in thestructure of the ACLT joint cartilage, the mechanical propertiesof the tissue were not significantly altered. Although theYoung’s modulus was almost 30 % smaller in the ACLT groupcompared to the contralateral group, and the global and localtissue strains were elevated in the operated knee cartilage, thesechanges were not statistically significant. While the observedstructural and mechanical changes in the ECM might have someeffect on chondrocyte biomechanics, the used FE model revealedthat these changes alone could not account for theexperimentally observed alterations in chondrocyte behavior.
The FE model showed that the increased global and localECM strains were due to fibrillation of the collagen network,reducing the collagen fibril network stiffness. Thus, cells in thefibrillated cartilage can expand more easily in parallel to thecartilage surface, while in the contralateral cartilage, the intactcollagen network effectively restricts the horizontal expansion ofthe cells. The decrease in the amount of FCDs in the PCMresulted in a decrease in the swelling pressure in the PCM [34,105]. This alteration resulted in reduced compressive modulusof the PCM, thus, the protective role of the PCM was diminishedand the cells were able to increase in volume even more.
8.3 LIMITATIONS AND FUTURE PROSPECTS
In this thesis, intact cartilage samples with bone attached wereprimarily used. However, currently in many studies, cartilageexplants are still needed, such as for the measurement of middleand deep zone cells. In order to clarify the usefulness of theexplants, the threshold value (distance from the cut surface)after which the cells in explants begin to behave similarly tothose in intact cartilage should be determined. Furthermore, in

Discussion
Dissertations in Forestry and Natural Sciences No 103 57
the future the utilization of dual-photon microscope [175] forimaging middle and deep zone chondrocytes through intactcartilage surfaces should be enhanced and improved.
There was a significant difference detected in chondrocytebiomechanics between intact and degraded samples (collagendegradation). There was also a significant difference in cellbiomechanical behavior between ACLT and contralateralcartilage. Although the structure and composition of the ECMwas analyzed in the present studies, local structural propertiesof cartilage in the vicinity of cells, especially in the PCM, andtheir relation to chondrocyte biomechanical behavior couldindicate the altered cell responses in more detail. The structureof the collagen network in the PCM could be analyzed withnano- and microscale techniques, such as transmission electronmicroscopy (TEM) and scanning electron microscopy (SEM),respectively [201, 202]. Immunostaining combined with confocallaser scanning could be utilized for studying, e.g., the type VIcollagen distribution in the PCM [203]. The PG content of thePCM could be quantified with DD [151], histochemistry [204] orimmunolabeling combined with fluorescence microscopy [205].
In addition to local structural properties, the determination oflocal material properties around cells would also be one way toobtain more detailed explanations behind changed cellbiomechanics in diseased or degraded cartilage. Thoseproperties could be studied with nanoindentation e.g. by usingatomic force microscopy (AFM) [206]. With AFM-basedmethods, one can obtain detailed information on the materialproperties of the collagen fibers [207], spatially varying tissuematerial properties [208, 209], and even the mechanics of singlemolecules [210, 211]. In addition, microaspiration could be usedfor revealing the mechanical properties of chondrons [155].
It was surprising that in an earlier study where cell responseswere measured 9 weeks after ACLT [134], the cells swelled lessunder mechanical loading compared to those cells measured inthis thesis 4 weeks after ACLT. This could be related to anattempt to repair cartilage, which is supported by the study ofPapaioannou et al. (2004) [212], where a repair phase from 8-16

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
58 Dissertations in Forestry and Natural Sciences No 103
weeks after ACLT was discovered. In both of the studies, theprimary explanations for the altered cell deformation behaviorwere the reduced PG content and increased collagen fibrillationof the ECM. A better explanation for the amplified cell volumeincrease in the 4 week ACLT group could again be found byanalyzing the local structural and material properties of thePCM.
In the present thesis, only collagen was degradedenzymatically. This was intentional because based on somerecent earlier studies [133, 149] the collagen network washypothesized to primarily modulate cell responses. There are noexperimental studies analyzing the specific effect of degradationof PGs on cell biomechanics, nor are there studies where thecombined effect of the collagen and PG degradation would havebeen revealed. Thus, this kind of study should be conducted inthe future. Either enzymatic digestion (e.g., chondroitinase ABCand collagenase together or trypsin) or a physicochemicallyinduced model of OA (natural digestion of the matrix with thehelp of interleukin-1�) could then be used.
For practical reasons the samples in studies I and II had to befrozen and thawed before structural and compositionalmeasurements. One could argue that freezing could haveaffected the results and have a different impact on intactcartilage samples and cartilage explants. In previous studies, theeffect of freezing-thawing on the material properties of cartilagetissue was analyzed, and not found to have any significanteffects on them [213, 214]. On the other hand, the intracellularstructure of chondrocytes in situ [215] and cell membrane [216]have been reported to be affected by freezing-thawing, however,the ultrastructure of the ECM was not found to change [215,216]. Based on the studies by Zheng et al. (2009) [217] andLaouar et al. (2007) [218], there might be some loss of GAGs dueto freezing, but in contrast, Guan et al. (2006) [216] did notobserve any changes in the GAG content. In these previousstudies, different cryopreservation techniques were used, whichfurther complicates the comparison and interpretation of theresults. Previously, in the study of Kääb et al. (1998) [219] SEM

Discussion
Dissertations in Forestry and Natural Sciences No 103 59
did not reveal any significant differences between small excisedsamples and whole knee joints of rabbits fixed with acryotechnique. Importantly, in study I (the only study whereexplants and intact tissues were compared), the structural data(collagen and PG content) was analyzed from whole samplesonly (and correlated with cell volumes of the intact samples), i.e.in that study the possible difference in tissue boundaries(explants, intact) could not affect the results. Furthermore,cartilage tissues with cut surfaces were not used in study II.Nonetheless, in the future, changes in the local structure ofcartilage and in the local material properties due to freezing-thawing should be studied in detail e.g. utilizing TEM or SEMand nanoindentation, respectively.
In the future, engineered cartilage could serve as aninteresting platform to study the effect of modified cartilage onchondrocyte biomechanical properties in a controlled manner.In addition, our methods could also help to define the quality ofengineered cartilage, and provide a method for comparing itwith normal articular cartilage. More specifically, thebiomechanical behavior of the chondrocytes seeded into theengineered cartilage scaffolds could be measured. This wouldprovide detailed information about the quality of the engineeredcartilage in terms of cell responses.

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
60 Dissertations in Forestry and Natural Sciences No 103

Dissertations in Forestry and Natural Sciences No 103 61
9 Summary andconclusions
This thesis investigated the effect of the structure andcomposition of the ECM and PCM on chondrocytebiomechanics. Chondrocyte volume and morphology in bovineand rabbit cartilages before and after osmotic and mechanicalloading were analyzed with confocal and dual-photon laserscanning microscopy, respectively. The effect of sample integrity(sample preparation), collagen degradation (enzymatictreatment) and degeneration of cartilage in early OA (rabbitACLT model) on chondrocyte behavior were studied in detail.Collagen content, collagen orientation angle and PG contentwere analyzed with FTIR, PLM and DD. FE modeling wasutilized in order to find the detailed explanations behind theobserved cell level phenomena occurring in early OA.
The main conclusions can be summarized as follows:
1. Chondrocyte volume and morphology following hypotonicloading is different in cartilage explants and intact cartilage.
2. Enzymatically induced collagen degradation, modulatingthe collagen content and collagen fibril orientation incartilage, alters cell volume and morphology followinghypotonic loading.
3. Experimentally-induced very early stage of OA results insuperficial collagen fibrillation and a reduction in the PGcontent, and this alters the biomechanical response of thechondrocytes to mechanical loading.
4. The loss of FCD in the PCM and the fibrillation of thesuperficial ECM in very early stage of experimentally-induced OA explain the increase in the cell volumefollowing mechanical loading of cartilage.

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
62 Dissertations in Forestry and Natural Sciences No 103
Changes in cell volume and morphology have been earlierproposed to be related to altered cell biosynthesis and viability[72, 75, 220], alterations which may lead to further changes incartilage structure and accelerate cartilage degeneration. Thus, itis of major importance to improve the knowledge of cellularresponses and cell-tissue interactions in normal anddegenerated cartilage in order to develop methods for detectingthese alterations at a very early stage and to create novelstrategies to slow down, stop or even reverse the progression ofOA.
9.1 NEXT STEPS TOWARD BETTER UNDERSTANDING OF
CARTILAGE MECHANOTRANSDUCTION
In this thesis, chondrocyte biomechanics was analyzed primarilyin situ at steady state. This is important for understanding cellbiomechanical behavior in disease and under static loading. Thenext steps should be to conduct measurements in situ or close toin vivo using intact joints and dynamic loading [21, 150]. Thiscould provide a more realistic depiction of the cell responses ina more physiological environment and loading, for instancesimulating walking. Furthermore, in the future, biomechanicalresponses of cells and their alterations in health and diseaseshould be linked to the biological responses of the chondrocytes.This could clarify the importance of the morphological andvolumetrical parameters of cells on cartilagemechanotransduction and progression of OA. Ultimately, thiskind of information would be crucial for incorporation intoadaptive computational models of cartilage permittingprediction of the progression of OA in a patient.

Dissertations in Forestry and Natural Sciences No 103 63
Bibliography
[1] V. C. Mow, A. Ratcliffe, and A. R. Poole, "Cartilage and diarthrodialjoints as paradigms for hierarchical materials and structures," Biomaterials13, 67-97 (1992).
[2] J. A. Buckwalter and J. Martin, "Degenerative joint disease," Clin symp 47,1-32 (1995).
[3] J. A. Buckwalter and H. J. Mankin, "Articular cartilage. Part I: Tissuedesign and chondrocyte-matrix interactions," J Bone Joint Surg Am 79A,600-611 (1997).
[4] G. Meachim and R. A. Stockwell, “The matrix,” in Adult articular cartilage,M. A. R. Freeman (ed), Alden Press, Oxford, England (1973).
[5] A. M. Ahmed and D. L. Burke, "In-vitro measurement of static pressuredistribution in synovial joints-Part I: Tibial surface of the knee," J BiomechEng 105, 216-225 (1983).
[6] V. C. Mow, W. Zhu, and A. Ratcliffe, "Structure and function of articularcartilage and meniscus," in Basic orthopaedic biomechanics, V. C. Mow (ed),Raven Press Ltd, New York (1991).
[7] M. A. R. Freeman and G. Meachim, "Ageing, degeneration andremodeling of articular cartilage," in Adult articular cartilage, M. A. R.Freeman (ed), Alden Press, Oxford, England (1973).
[8] J. A. Buckwalter and H. J. Mankin, "Articular cartilage. Part II:Degeneration and osteoarthritis, repair, regeneration, andtransplantation," J Bone Joint Surg Am 79, 612-632 (1997).
[9] R. A. Stockwell and G. Meachim, "The Chondrocytes," in Adult articularcartilage, M. A. R. Freeman (ed), Alden Press, Oxford, England (1973).
[10] M. D. Buschmann, E. B. Hunziker, Y.-J. Kim, and A. J. Grodzinsky,"Altered aggrecan synthesis correlates with cell and nucleus structure instatically compressed cartilage," J Cell Sci 109, 499-508 (1996).

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
64 Dissertations in Forestry and Natural Sciences No 103
[11] M. D. Buschmann, Y. A. Gluzband, A. J. Grodzinsky, and E. B. Hunziker,"Mechanical compression modulates matrix biosynthesis inchondrocyte/agarose culture," J Cell Sci 108, 1497-1508 (1995).
[12] I. L. Jones, A. Klämfeldt, and T. Sandström, "The effect of continuousmechanical pressure upon the turnover of articular cartilageproteoglycans in vitro," Clin Orthop Relat Res 185, 283-289 (1982).
[13] M. J. Palmoski and K. D. Brandt, "Effects of static and cyclic compressiveloading on articular cartilage plugs in vitro," Arthritis Rheum 27, 675(1984).
[14] R. Schneiderman, D. Keret, and A. Maroudas, "Effects of mechanical andosmotic pressure on the rate of glycosaminoglycan synthesis in thehuman adult femoral head cartilage - an in vitro study," J Orthop Res 4,393-408 (1986).
[15] J. P. G. Urban, "The Chondrocyte - A cell under pressure," Br J Rheumatol33, 901-908 (1994).
[16] C. A. Poole, "Articular cartilage chondrons: form, function and failure," JAnat 191, 1-13 (1997).
[17] H. A. Alhadlaq and Y. Xia, "The structural adaptations in compressedarticular cartilage by microscopic MRI (μMRI) T2 anisotropy,"Osteoarthritis Cartilage 12, 887-894 (2004).
[18] H. A. Alhadlaq, Y. Xia, J. B. Moody, and J. R. Matyas, "Detectingstructural changes in early experimental osteoarthritis of tibial cartilageby microscopic magnetic resonance imaging and polarised lightmicroscopy," Ann Rheum Dis 63, 709-717 (2004).
[19] J. P. Arokoski, M. M. Hyttinen, T. Lapveteläinen, P. Takács, B.Kosztáczky, L. Módis, V. Kovanen, and H. Helminen, "Decreasedbirefringence of the superficial zone collagen network in the canine knee(stifle) articular cartilage after long distance running training, detected byquantitative polarised light microscopy," Ann Rheum Dis 55, 253-264(1996).
[20] A. L. Clark, T. R. Leonard, L. D. Barclay, J. R. Matyas, and W. Herzog,"Opposing cartilages in the patellofemoral joint adapt differently to long-term cruciate deficiency: chondrocyte deformation and reorientationwith compression," Osteoarthritis Cartilage 13, 1100-1114 (2005).

Bibliography
Dissertations in Forestry and Natural Sciences No 103 65
[21] A. L. Clark, L. D. Barclay, J. R. Matyas, and W. Herzog, "In situchondrocyte deformation with physiological compression of the felinepatellofemoral joint," J Biomech 36, 553-568 (2003).
[22] Y. Xia, J. B. Moody, H. Alhadlaq, and J. Hu, "Imaging the physical andmorphological properties of a multi-zone young articular cartilage atmicroscopic resolution," J Magn Reson Imaging 17, 365-374 (2003).
[23] K. A. Athanasiou, M. P. Rosenwasser, J. A. Buckwalter, T. I. Malinin, andV. C. Mow, "Interspecies comparisons of in situ intrinsic mechanicalproperties of distal femoral cartilage," J Orthop Res 9, 330-340 (1991).
[24] D. Eyre, "Collagen of articular cartilage," Arthr Res 4, 30-35 (2002).
[25] D. J. Prockop and K. I. Kivirikko, "Collagens: molecular biology, diseases,and potentials for therapy," Ann Rev Biochem 64, 403-434 (1995).
[26] A. K. Jeffery, G. W. Blunn, C. W. Archer, and G. Bentley, "Three-dimensional collagen architecture in bovine articular cartilage," J BoneJoint Surg Br 73-B, 795-801 (1991).
[27] D. L. Bader, G. E. Kempson, J. Egan, W. Gilbey, and A. J. Barrett, "Theeffects of selective matrix degradation on the short-term compressiveproperties of adult human articular cartilage," Biochim Biophys Acta 1116,147-154 (1992).
[28] R. A. Bank, M. Soudry, A. Maroudas, J. Mizrahi, and J. M. TeKoppele,"The increased swelling and instantaneous deformation of osteoarthriticcartilage is highly correlated with collagen degradation," Arthritis Rheum43, 2202-2210 (2000).
[29] G. E. Kempson, H. Muir, C. Pollard, and M. Tuke, "The tensile propertiesof the cartilage of human femoral condyles related to the content ofcollagen and glycosaminoglycans," Biochim Biophys Acta 297, 456-472(1973).
[30] G. E. Kempson, M. A. R. Freeman, and S. A. V. Swanson, "Tensileproperties of articular cartilage," Nature 220, 1127-1128 (1968).
[31] J. Mizrahi, A. Maroudas, Y. Lanir, I. Ziv, and T. J. Webber, "The"instantaneous" deformation of cartilage: effects of collagen fiberorientation and osmotic stress," Biorheology 23, 311-330 (1986).
[32] S. Park, S. B. Nicoll, R. L. Mauck, and G. A. Ateshian, "Cartilagemechanical response under dynamic compression at physiological stress

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
66 Dissertations in Forestry and Natural Sciences No 103
levels following collagenase digestion," Ann Biomed Eng 36, 425-434(2008).
[33] V. C. Mow, M. H. Holmes, and W. M. Lai, "Fluid transport andmechanical properties of articular cartilage: a review," J Biomech 17, 377-394 (1984).
[34] P. J. Basser, R. Schneiderman, R. A. Bank, E. Wachtel, and A. Maroudas,"Mechanical properties of the collagen network in human articularcartilage as measured by osmotic stress technique," Arch Biochem Biophys351, 207-219 (1998).
[35] I. H. M. Muir, "Biochemistry," in Adult articular cartilage, M. A. R.Freeman (ed), Alden Press, Oxford (1973).
[36] C. Kiani, L. Chen, Y. J. Wu, A. J. Yee, and B. B. Yang. "Structure andfunction of aggrecan," Cell Res 12, 19-32 (2002).
[37] R. Lemperg and S.-E. Larsson, "The glycosaminoglycans of bovinearticular cartilage. I. Concentration and distribution in different layers inrelation to age," Calcif Tissue Res 15, 237-251 (1974).
[38] H. Lipshitz, R. Etheredge, and M. J. Glimcher, "In vitro wear of articularcartilage," J Bone Joint Surg Am 57, 527-534 (1975).
[39] A. Maroudas, "Physico-chemical properties of articular cartilage," inAdult articular cartilage, M. A. R. Freeman (ed), Alden Press, Oxford(1973).
[40] A. Maroudas and C. Bannon, "Measurement of swelling pressure incartilage and comparison with the osmotic pressure of constituentproteoglycans," Biorheology 18, 619-632 (1981).
[41] C. A. McDevitt, "Biochemistry of articular cartilage. Nature ofproteoglycans and collagen of articular cartilage and their role in ageingand in osteoarthrosis," Ann Rheum Dis 32, 364-378 (1973).
[42] M. D. Buschmann and A. J. Grodzinsky, "A molecular model ofproteoglycan-associated electrostatic forces in cartilage mechanics," JBiomech Eng 117, 179-192 (1995).
[43] A. Maroudas, "Distribution and diffusion of solutes in articularcartilage," Biophys J 10, 365-379 (1970).

Bibliography
Dissertations in Forestry and Natural Sciences No 103 67
[44] A. Maroudas, P. Bullough, S. A. V. Swanson, and M. A. R. Freeman, "Thepermeability of articular cartilage," J Bone Joint Surg Br 50-B, 166-177(1968).
[45] G. E. Kempson, "Mechanical properties of articular cartilage," in Adultarticular cartilage, M. A. R. Freeman (ed), Alden Press, Oxford (1973).
[46] J. P. G. Urban and J. F. McMullin, "Swelling pressure of the lumbarintervertebral discs: influence of age, spinal level, composition, anddegeneration," Spine 13, 179-187 (1988).
[47] M. A. Soltz and G. A. Ateshian, "Interstitial fluid pressurization duringconfined compression cyclical loading of articular cartilage," Ann BiomedEng 28, 150-159 (2000).
[48] R. A. Stockwell, "The cell density of human articular and costal cartilage,"J Anat 101, 753-763 (1967).
[49] M. Huber, S. Trattnig, and F. Lintner, "Anatomy, biochemistry, andphysiology of articular cartilage," Invest Radiol 35, 573-580 (2000).
[50] W. R. Trickey, T. P. Vail, and F. Guilak, "The role of the cytoskeleton inthe viscoelastic properties of human articular chondrocytes," J Orthop Res22, 131-139 (2004).
[51] F. Guilak and V. C. Mow, "The mechanical environment of thechondrocyte: a biphasic finite element model of cell-matrix interactionsin articular cartilage," J Biomech 33, 1663-1673 (2000).
[52] W. R. Jones, H. P. Ting-Beall, G. M. Lee, S. S. Kelley, R. M. Hochmuth,and F. Guilak. "Alterations in the Young's modulus and volumetricproperties of chondrocytes isolated from normal and osteoarthritichuman cartilage," J Biomech 32, 119-127 (1999).
[53] W. R. Trickey, G. M. Lee, and F. Guilak, "Viscoelastic properties ofchondrocytes from normal and osteoarthritic human cartilage," J OrthopRes 18, 891-898 (2000).
[54] R. O. Hynes. "Integrins: A family of cell surface receptors," Cell 48, 549-554 (1987).
[55] F. M. Pavalko, C. A. Otey, K. O. Simon, and K. Burridge, "Alpha-actinin:a direct link between actin and integrins," Biochem Soc Trans 19, 1065-1069(1991).

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
68 Dissertations in Forestry and Natural Sciences No 103
[56] C. A. Poole, M. H. Flint, and B. W. Beaumont, "Chondrons extracted fromcanine tibial cartilage: preliminary report on their isolation andstructure," J Orthop Res 6, 408-419 (1988).
[57] C. A. Poole, S. Ayad, and J. R. Schofield, "Chondrons from articularcartilage: I. Immunolocalization of type VI collagen in the pericellularcapsule of isolated canine tibial chondrons," J Cell Sci 90, 635-643 (1988).
[58] C. A. Poole, S. Ayad, and R. T. Gilbert, "Chondrons from articularcartilage. V. Immunohistochemical evaluation of type VI collagenorganisation in isolated chondrons by light, confocal and electronmicroscopy," J Cell Sci 103, 1101-1110 (1992).
[59] C. A. Poole, T. T. Glant, and J. R. Schofield, "Chondrons from articularcartilage. IV. Immunolocalization of proteoglycan epitopes in isolatedcanine tibial chondrons," J Histochem Cytochem 39, 1175-1187 (1991).
[60] R. M. Mason, "Recent advances in the biochemistry of hyaluronic acid incartilage," Connect Tissue Res 54, 87-112 (1981).
[61] G. M. Winter, C. A. Poole, M. Z. Ilic, J. M. Ross, H. C. Robinson, and C. J.Handley, "Identification of distinct metabolic pools of aggrecan and theirrelationship to type VI collagen in the chondrons of mature bovinearticular cartilage explants," Connect Tissue Res 37, 277-293 (1998).
[62] T. Glant, C. Hadházy, K. Mikecz, and A. Sipos, "Appearance andpersistence of fibronectin in cartilage. Specific interaction of fibronectinwith collagen type II," Histochem 82, 149-158 (1985).
[63] C. A. Poole, R. T. Gilbert, D. Herbage, and D. J. Hartmann,"Immunolocalization of type IX collagen in normal and spontaneouslyosteoarthritic canine tibial cartilage and isolated chondrons,"Osteoarthritis Cartilage 5, 191-204 (1997).
[64] S. F. Wotton, R. E. Jeacocke, R. A. Maciewicz, R. J. Wardale, and V. C.Duance, "The application of scanning confocal microscopy in cartilageresearch," Histochem J 23, 328-335 (1991).
[65] F. Guilak, L. G. Alexopoulos, M. L. Upton, I. Youn, J. B. Choi, L. Cao, L.A. Setton, and M. A. Haider, "The pericellular matrix as a transducer ofbiomechanical and biochemical signals in articular cartilage," Ann NYAcad Sci 1068, 498-512 (2006).
[66] L. G. Alexopoulos, G. M. Williams, M. L. Upton, L. A. Setton, and F.Guilak, "Osteoarthritic changes in the biphasic mechanical properties of

Bibliography
Dissertations in Forestry and Natural Sciences No 103 69
the chondrocyte pericellular matrix in articular cartilage" J Biomech 38,509-517 (2005).
[67] L. G. Alexopoulos, L. A. Setton, and F. Guilak, "The biomechanical role ofthe chondrocyte pericellular matrix in articular cartilage," Acta Biomat 1,317-325 (2005).
[68] F. Guilak, W. R. Jones, H. P. Ting-Beall, and G. M. Lee. "The deformationbehavior and mechanical properties of chondrocytes in articularcartilage," Osteoarthritis Cartilage 7, 59-70 (1999).
[69] R. K. Korhonen and W. Herzog, "Depth-dependent analysis of the role ofcollagen fibrils, fixed charges and fluid in the pericellular matrix ofarticular cartilage on chondrocyte mechanics," J Biomech 41, 480-485(2008).
[70] M. L. Gray, A. M. Pizzanelli, A. J. Grodzinsky, and R. C. Lee, "Mechanicaland physiochemical determinants of the chondrocyte biosyntheticresponse," J Orthop Res 6, 777-792 (1988).
[71] R. L.-Y. Sah, Y. J. Kim, J.-Y. H. Doong, A. J. Grodzinsky, A. H. K. Plaas,and J. D. Sandy, "Biosynthetic response of cartilage explants to dynamiccompression," J Orthop Res 7, 619-636 (1989).
[72] M. Wong, P. Wuethrich, M. D. Buschmann, P. Eggli, and E. Hunziker,"Chondrocyte biosynthesis correlates with local tissue strain in staticallycompressed adult articular cartilage," J Orthop Res 15, 189-196 (1997).
[73] P. Newman and F. M. Watt, "Influence of cytochalasin D-inducedchanges in cell shape on proteoglycan synthesis by cultured articularchondrocytes," Exp Cell Res 178, 199-210 (1988).
[74] P. Benya and J. D. Shaffer, "Dedifferentiated chondrocytes reexpress thedifferentiated collagen phenotype when cultured in agarose gels," Cell 30,215-224 (1982).
[75] J. P. G. Urban, A. C. Hall, and K. A. Gehl, "Regulation of matrix synthesisrates by the ionic and osmotic environment of articular chondrocytes," JCell Phys 154, 262-270 (1993).
[76] P. D. Gorevic, "Osteoarthritis. A review of musculoskeletal aging andtreatment issues in geriatric patients," Geriatrics 59, 28-32 (2004).
[77] R. C. Lawrence, D. T. Felson, C. G. Helmick, L. M. Arnold, H. Choi, R. A.Deyo, S. Gabriel, R. Hirsch, M. C. Hochberg, G. G. Hunder, J. M. Jordan,J. N. Katz, H. M. Kremers, and F. Wolfe, "Estimates of the prevalence of

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
70 Dissertations in Forestry and Natural Sciences No 103
arthritis and other rheumatic conditions in the United States. Part II,"Arthritis Rheum 58, 26-35 (2008).
[78] P. A. Dieppe and L. S. Lohmander, "Pathogenesis and management ofpain in osteoarthritis," Lancet 365, 965-973 (2005).
[79] D. T. Felson and Y. Zhang, "An update on the epidemiology of knee andhip osteoarthritis with a view to prevention," Arthritis Rheum 41, 1343-1355 (1998).
[80] D. T. Felson, J. J. Anderson, A. Naimark, A. M. Walker, and R. F. Meenan,"Obesity and knee osteoarthritis. The Framingham Study," Ann Int Med109, 18-18 (1988).
[81] C. A. McDevitt and H. Muir, "Biochemical changes in the cartilage of theknee in experimental and natural osteoarthritis in the dog," J Bone JointSurg Br 58, 94-101 (1976).
[82] A. Maroudas and M. Venn, "Chemical composition and swelling ofnormal and osteoarthrotic femoral head cartilage. II. Swelling." AnnRheum Dis 36, 399-406 (1977).
[83] G. Grushko, R. Schneiderman, and A. Maroudas, "Some biochemical andbiophysical parameters for the study of the pathogenesis ofosteoarthritis: a comparison between the processes of ageing anddegeneration in human hip cartilage," Connect Tissue Res 19, 149-176(1989).
[84] R. A. Stockwell, "Cartilage failure in osteoarthritis: Relevance of normalstructure and function. A review," Clin Anat 4, 161-191 (1991).
[85] C. Weiss and S. Mirow, "An ultrastructural study of osteoarthritischanges in the articular cartilage of human knees," J Bone Joint Surg Am54, 954-972 (1972).
[86] E. Vignon, M. Arlot, P. Meunier, and G. Vignon, "Quantitativehistological changes in osteoarthritic hip cartilage. Morphometricanalysis of 29 osteoarthritic and 26 normal human femoral heads," ClinOrthop Relat Res 103, 269-278 (1974).
[87] X. Bi, X. Yang, M. P. G. Bostrom, and N. P. Camacho, "Fourier transforminfrared imaging spectroscopy investigations in the pathogenesis andrepair of cartilage," Biochim Biophys Acta Biomembr 1758, 934-941 (2006).
[88] S. Saarakkala, P. Julkunen, J. Mäkitalo, J. S. Jurvelin, and R. K. Korhonen,"Depth-wise progression of osteoarthritis in human articular cartilage:

Bibliography
Dissertations in Forestry and Natural Sciences No 103 71
investigation of composition, structure and biomechanics," OsteoarthritisCartilage 18, 73-81 (2010).
[89] M. H. Chen and N. Broom, "On the ultrastructure of softened cartilage: apossible model for structural transformation," J Anat 192, 329-341 (1998).
[90] A. Oloyede and N. Broom, "The biomechanics of cartilage load-carriage,"Connect Tissue Res 34, 119-143 (1996).
[91] J. T. A. Mäkelä, M. R. J. Huttu, and R. K. Korhonen, "Structure–functionrelationships in osteoarthritic human hip joint articular cartilage,"Osteoarthritis Cartilage 20, 1268-1277 (2012).
[92] M. S. Laasanen, J. Töyräs, R. K. Korhonen, J. Rieppo, S. Saarakkala, M. T.Nieminen, J. Hirvonen, and J.S. Jurvelin, "Biomechanical properties ofknee articular cartilage," Biorheology 40, 133-140 (2003).
[93] V. C. Mow, A. F. Mak, W. M. Lai and L. C. Rosenberg, "Viscoelasticproperties of proteoglycan subunits and aggregates in varying solutionconcentrations," J Biomech 17, 325-338 (1984).
[94] R. W. Moskowitz, D. S. Howell, V. M. Goldberg, O. Muniz, and J. C. Pita,"Cartilage proteoglycan alterations in an experimentally induced modelof rabbit osteoarthritis," Arthritis Rheum 22, 155-163 (1979).
[95] R. D. Altman. "Overview of osteoarthritis," Am J Med 83, 65-69 (1987).
[96] P. Julkunen, J. S. Jurvelin, and H. Isaksson, "Contribution of tissuecomposition and structure to mechanical response of articular cartilageunder different loading geometries and strain rates," Biomech ModelMechanobiol 9, 237-245 (2010).
[97] G. E. Kempson, H. Muir, S. A. V. Swanson, and M. A. R. Freeman,"Correlations between stiffness and the chemical constituents of cartilageon the human femoral head," Biochim Biophys Acta 215, 70-77 (1970).
[98] P. Kiviranta, J. Rieppo, R. K. Korhonen, P. Julkunen, J. Töyräs, and J. S.Jurvelin, "Collagen network primarily controls Poisson's ratio of bovinearticular cartilage in compression," J Orthop Res 24, 690-699 (2006).
[99] J. S. Jurvelin, M. D. Buschmann, and E. B. Hunziker, "Optical andmechanical determination of Poisson's ratio of adult bovine humeralarticular cartilage," J Biomech 30, 235-241 (1997).
[100] J. Rieppo, J. Töyräs, M. T. Nieminen, V. Kovanen, M. M. Hyttinen, R. K.Korhonen, J. S. Jurvelin, and H. J. Helminen, "Structure-function

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
72 Dissertations in Forestry and Natural Sciences No 103
relationships in enzymatically modified articular cartilage," Cells TissuesOrgans 175, 121-132 (2003).
[101] J. S. Jurvelin, J. P. Arokoski, E. B. Hunziker, and H. J. Helminen,"Topographical variation of the elastic properties of articular cartilage inthe canine knee," J Biomech 33, 669-675 (2000).
[102] J. P. G. Urban, "Present perspectives on cartilage and chondrocytemechanobiology," Biorheology 37, 185-190 (2000).
[103] W. M. Lai, W. Y. Gu, and V. C. Mow, "On the conditional equivalence ofchemical loading and mechanical loading on articular cartilage," JBiomech 31, 1181-1185 (1998).
[104] C. M. Flahiff, D. A. Narmoneva, J. L. Huebner, V. B. Kraus, F. Guilak, andL. A. Setton, "Osmotic loading to determine the intrinsic materialproperties of guinea pig knee cartilage," J Biomech 35, 1285-1290 (2002).
[105] A. Maroudas, "Balance between swelling pressure and collagen tensionin normal and degenerate cartilage," Nature 260, 808-809 (1976).
[106] A. Maroudas, "Physicochemical properties of cartilage in the light of ionexchange theory," Biophys J 8, 575-95 (1968).
[107] J. M. Huyghe and J. D. Janssen, "Quadriphasic mechanics of swellingincompressible porous media," Int J Eng Sci 35, 793-802 (1997).
[108] W. M. Lai, J. S. Hou, and V. C. Mow, "A triphasic theory for the swellingand deformation behaviors of articular cartilage," J Biomech Eng 113, 245-258 (1991).
[109] F. Guilak, A. Ratcliffe, and V. C. Mow, "Chondrocyte deformation andlocal tissue strain in articular cartilage: a confocal microscopy study," JOrthop Res 13, 410-421 (1995).
[110] D. L. Bader and G. E. Kempson, "The short-term compressive propertiesof adult human articular cartilage," Biomed Mater Eng 4, 245-256 (1994).
[111] A. F. Mak, W. M. Lai, and V. C. Mow, "Biphasic indentation of articularcartilage - I. Theoretical analysis," J Biomech 20, 703-714 (1987).
[112] W. Wilson, C. C. van Donkelaar, B. van Rietbergen, K. Ito, and R.Huiskes, "Stresses in the local collagen network of articular cartilage: aporoviscoelastic fibril-reinforced finite element study," J Biomech 37, 357-366 (2004).

Bibliography
Dissertations in Forestry and Natural Sciences No 103 73
[113] L. P. Li, J. Soulhat, M. D. Buschmann, and A. Shirazi-Adl, "Nonlinearanalysis of cartilage in unconfined ramp compression using a fibrilreinforced poroelastic model," Clin Biomech 14, 673-682 (1999).
[114] P. Julkunen, P. Kiviranta, W. Wilson, J. S. Jurvelin, and R. K. Korhonen,"Characterization of articular cartilage by combining microscopicanalysis with a fibril-reinforced finite-element model," J Biomech 40, 1862-1870 (2007).
[115] R. K. Korhonen, M. S. Laasanen, J. Töyräs, R. Lappalainen, H. J.Helminen, and J. S. Jurvelin, "Fibril reinforced poroelastic model predictsspecifically mechanical behavior of normal, proteoglycan depleted andcollagen degraded articular cartilage," J Biomech 36, 1373-1379 (2003).
[116] W. Wilson, C. C. van Donkelaar, B. van Rietbergen, and R. Huiskes, "Afibril-reinforced poroviscoelastic swelling model for articular cartilage," JBiomech 38, 1195-1204 (2005).
[117] M. W. Cornwall, "Biomechanics of noncontractile tissue: A review," PhysTher 64, 1869-1873 (1984).
[118] W. M. Lai, V. C. Mow, and V. Roth, "Effects of nonlinear strain-dependent permeability and rate of compression on the stress behaviorof articular cartilage," J Biomech Eng 103, 61-66 (1981).
[119] A. F. van der Voet, P. J. Prendergast, W. D. van Driel, and J. H. Kuiper,"A comparison of finite element codes for the solution of biphasicporoelastic problems," Proc Instn Mech Engrs 211, 209-211 (1997).
[120] W. Wilson, C. C. Donkelaar, and J. M. Huyghe, "A comparison betweenmechano-electrochemical and biphasic swelling theories for softhydrated tissues," J Biomech Eng 127, 158-165 (2005).
[121] A. C. Hall, E. R. Horwitz, and R. J. Wilkins, "The cellular physiology ofarticular cartilage," Exp Physiol 81, 535-545 (1996).
[122] F. Guilak, "The deformation behavior and viscoelastic properties ofchondrocytes in articular cartilage," Biorheology 37, 27-44 (2000).
[123] M. J. Kääb, R. G. Richards, K. Ito, I. ap Gwynn, and H. P. Nötzli,"Deformation of chondrocytes in articular cartilage under compressiveload: a morphological study," Cells Tissues Organs 175, 133-139 (2003).
[124] R. Lewis, C. H. Feetham, and R. Barrett-Jolley, "Cell volume regulation inchondrocytes," Cell Physiol Biochem 28, 1111-1122 (2011).

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
74 Dissertations in Forestry and Natural Sciences No 103
[125] E. K. Hoffmann, I. H. Lambert, and S. F. Pedersen, "Physiology of cellvolume regulation in vertebrates," Physiol Rev 89, 193-277 (2009).
[126] F. Wehner, H. Olsen, H. Tinel, E. Kinne-Saffran, and R. K. H. Kinne, "Cellvolume regulation: osmolytes, osmolyte transport, and signaltransduction," Rev Physiol Biochem Pharmacol 148, 1-80 (2003).
[127] A. Mobasheri and D. Marples, "Expression of the AQP-1 water channel innormal human tissues: a semiquantitative study using tissue microarraytechnology," Am J Physiol Cell Physiol 286, 529-537 (2004).
[128] F. Lang, G. L. Busch, M. Ritter, H. Völki, S. Waldegger, E. Gulbins, and D.Häussinger, "Functional significance of cell volume regulatorymechanisms," Physiol Rev 78, 247-306 (1998).
[129] P. G. Bush and A. C. Hall, "Passive osmotic properties of in situ humanarticular chondrocytes within non-degenerate and degenerate cartilage,"J Cell Physiol 204, 309-319 (2005).
[130] P. G. Bush and A. C. Hall, "The volume and morphology of chondrocyteswithin non-degenerate and degenerate human articular cartilage,"Osteoarthritis Cartilage 11, 242-251 (2003).
[131] P. G. Bush and A. C. Hall, "Regulatory volume decrease (RVD) byisolated and in situ bovine articular chondrocytes," J Cell Physiol 187, 304-314 (2001).
[132] P. G. Bush and A. C. Hall, "The osmotic sensitivity of isolated and in situbovine articular chondrocytes," J Orthop Res 19, 768-778 (2001).
[133] R. K. Korhonen, S.-K. Han, and W. Herzog. "Osmotic loading of in situchondrocytes in their native environment," Mol Cell Biomech 7, 125-134(2010).
[134] S.-K. Han, R. Seerattan, and W. Herzog. "Mechanical loading of in situchondrocytes in lapine retropatellar cartilage after anterior cruciateligament transection," J R Soc Interface 7, 895-903 (2010).
[135] S.-K. Han, P. Colarusso, and W. Herzog. "Confocal microscopyindentation system for studying in situ chondrocyte mechanics," Med EngPhys 31, 1038-1042 (2009).
[136] F. Guilak, G. Erickson, and H. P. Ting-Beall, "The effects of osmotic stresson the viscoelastic and physical properties of articular chondrocytes,"Biophys J 82, 720-727 (2002).

Bibliography
Dissertations in Forestry and Natural Sciences No 103 75
[137] C. M. P. Deshayes, A. C. Hall, and J. P. G. Urban, "Effect of extracellularosmolality on porcine articular chondrocyte volume," J Physiol 467, 214P(1993).
[138] P. M. Freeman, R. N. Natarajan, J. H. Kimura, and T. P. Andriacchi,"Chondrocyte cells respond mechanically to compressive loads," J OrthopRes 12, 311-320 (1994).
[139] M. M. Knight, S. A. Ghori, D. A. Lee, and D. L. Bader, "Measurement ofthe deformation of isolated chondrocytes in agarose subjected to cycliccompression," Med Eng Phys 20, 684-688 (1998).
[140] D. A. Lee, M. M. Knight, J. F. Bolton, B. D. Idowu, M. V. Kayser, and D.L. Bader, "Chondrocyte deformation within compressed agaroseconstructs at the cellular and sub-cellular levels," J Biomech 33, 81-95(2000).
[141] D. A. Lee and D. L. Bader, "The development and characterization of anin vitro system to study strain-induced cell deformation in isolatedchondrocytes," In Vitro Cell Dev Biol - Animal 31, 828-835 (1995).
[142] N. D. Leipzig and K. A. Athanasiou, "Unconfined creep compression ofchondrocytes," J Biomech 38, 77-85 (2005).
[143] A. C. Shieh and K. A. Athanasiou, "Biomechanics of single zonalchondrocytes," J Biomech 39, 1595-1602 (2006).
[144] L. Ng, H.-H. Hung, A. Sprunt, S. Chubinskaya, C. Ortiz, and A. J.Grodzinsky, "Nanomechanical properties of individual chondrocytes andtheir developing growth factor-stimulated pericellular matrix," J Biomech40, 1011-1023 (2007).
[145] R. J. Errington, M. D. Fricker, J. L. Wood, A. C. Hall, and N. S. White,"Four-dimensional imaging of living chondrocytes in cartilage usingconfocal microscopy: a pragmatic approach," Am J Physiol 272, C1040-C1051 (1997).
[146] N. D. Broom and D. B. Myers, "A study of the structural response of wethyaline cartilage to various loading situations.," Connect Tissue Res 7, 227-237 (1980).
[147] N. O. Chahine, C. T. Hung, and G. A. Ateshian, "In-situ measurements ofchondrocyte deformation under transient loading," Eur Cell Mater 13,100-111 (2007).

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
76 Dissertations in Forestry and Natural Sciences No 103
[148] F. Guilak, "Compression-induced changes in the shape and volume ofthe chondrocyte nucleus," J Biomech 28, 1529-1541 (1995).
[149] R. K. Korhonen, S.-K. Han, and W. Herzog. "Osmotic loading of articularcartilage modulates cell deformations along primary collagen fibrildirections " J Biomech 43, 783-787 (2010).
[150] Z. Abusara, R. Seerattan, A. Leumann, and W. Herzog, "A novel methodfor determining articular cartilage chondrocyte mechanics in vivo," JBiomech 44, 930-934 (2011).
[151] R. K. Korhonen, P. Julkunen, J. S. Jurvelin, and S. Saarakkala, "Structuraland compositional changes in peri- and extracellular matrix ofosteoarthritic cartilage modulate chondrocyte morphology, " Cell MolBioeng 4, 484-494 (2011).
[152] P. Julkunen, W. Wilson, J. S. Jurvelin, and R. K. Korhonen, "Compositionof the pericellular matrix modulates the deformation behaviour ofchondrocytes in articular cartilage under static loading," Med Biol EngComput 47, 1281-1290 (2009).
[153] T. L. Vincent, C. J. McLean, L. E. Full, D. Peston, and J. Saklatvala, "FGF-2is bound to perlecan in the pericellular matrix of articular cartilage,where it acts as a chondrocyte mechanotransducer," OsteoarthritisCartilage 15, 752-763 (2007).
[154] F. Guilak and V. C. Mow, "The mechanical environment of thechondrocyte: a biphasic finite element model of cell-matrix interactionsin articular cartilage," J Biomech 33, 1663-1673 (2000).
[155] L. G. Alexopoulos, M. A. Haider, T. P. Vail, and F. Guilak, "Alterations inthe mechanical properties of the human chondrocyte pericellular matrixwith osteoarthritis," J Biomech Eng 125, 323-333 (2003).
[156] W. A. Hing, A. F. Sherwin, J. M. Ross, and C. A. Poole, "The influence ofthe pericellular microenvironment on the chondrocyte response toosmotic challenge," Osteoarthritis Cartilage 10, 297-307 (2002).
[157] R. K. Korhonen, P. Julkunen, W. Wilson, and W. Herzog, "Importance ofcollagen orientation and depth-dependent fixed charge densities ofcartilage on mechanical behavior of chondrocytes," J Biomech Eng 130,021003 (2008).
[158] R. F. Loeser, "Chondrocyte integrin expression and function," Biorheology37, 109-116 (2000).

Bibliography
Dissertations in Forestry and Natural Sciences No 103 77
[159] S. J. Kim, E. J. Kim, Y. H. Kim, S. B. Hahn, and W. J. Lee, "The modulationof integrin expression by the extracellular matrix in articularchondrocytes," Yonsei Med J 44, 493-501 (2003).
[160] T. Wilson and C. J. R. Sheppard, Theory and practiceof scanning optical microscopy (Academic Press, London, 1984).
[161] D. Semwogerere and E. Weeks, "Confocal microscopy," Biomed Eng 23, 1-10 (2005).
[162] C. J. R. Sheppard and D. M. Shotton, Confocal laser scanning microscopy(BIOS Scientific publishers, United Kingdom, 1997).
[163] S. Inoue, "Foundations of confocal laser scanned imaging in lightmicroscopy," in Handbook of biological confocal microscopy, J.B. Pawley (ed),Plenum Press, New York (1990).
[164] K. S. Wells, D. R. Sandison, J. Strickler, and W. W. Webb, "Quantitativefluorescence imaging with laser scanning confocal microscopy," inHandbook of biological confocal microscopy, J.B. Pawley (ed), Plenum Press,New York (1990).
[165] H. E. Keller, "Objective lenses for confocal microscope," in Handbook ofbiological confocal microscopy, J.B. Pawley (ed), Plenum Press, New York(1990).
[166] F. Guilak, "Volume and surface area measurement of viable chondrocytesin situ using geometric modelling of serial confocal sections," J Micros173, 245-256 (1994).
[167] P. G. Bush, P. D. Hodkinson, G. L. Hamilton, and A. C. Hall, "Viabilityand volume of in situ bovine articular chondrocytes-changes following asingle impact and effects of medium osmolarity," Osteoarthritis Cartilage13, 54-65 (2005).
[168] S.-K. Han, R. Madden, Z. Abusara, and W. Herzog, "In situ chondrocyteviscoelasticity," J Biomech 45, 2450-2456 (2012).
[169] R. Madden, S.-K. Han, and W. Herzog, "Chondrocyte deformation underextreme tissue strain in two regions of the rabbit knee joint," J Biomech 46,554-560 (2013).
[170] R. Madden and W. Herzog, "Chondrocyte Deformation Under ExtremeTissue Strains," Biophys J 102, 193a (2012).

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
78 Dissertations in Forestry and Natural Sciences No 103
[171] W. Denk, J. H. Strickler, and W. W. Webb, "Two-photon laser scanningfluorescence microscopy," Science 248, 73-76 (1990).
[172] T. J. Holmes and P. C. Cheng. "Basic principles of imaging," in Multi-modality microscopy, H. Yu, P. Cheng, P. Lin, F. Kao (eds.), WorldScientific Pub Co, Singapore (2006).
[173] A. Diaspro, G. Chirico, and M. Collini, "Two-photon fluorescenceexcitation and related techniques in biological microscopy," Quart RevBiophys 38, 97-166 (2005).
[174] F. Bestvater, E. Spiess, G. Stobrawa, M. Hacker, T. Feurer, T. Porwol, U.Berchner-Pfannschmidt, C. Wotzlaw, and H. Acker, "Two-photonfluorescence absorption and emission spectra of dyes relevant for cellimaging," J Microscopy 208, 108-115 (2002).
[175] P. G. Bush, D. L. Wokosin and A. C. Hall, "Two-versus one photonexcitation laser scanning microscopy: Critical importance of excitationwavelength," Front Biosci 12, 2646-2657 (2007).
[176] W. C. Hayes, L. M. Keer, G. Herrmann, and L. F. Mockros, "Amathematical analysis for indentation tests of articular cartilage," JBiomech 5, 541-551 (1972).
[177] R. K. Korhonen, M. S. Laasanen, J. Töyräs, J. Rieppo, J. Hirvonen, H. J.Helminen, and J. S. Jurvelin, "Comparison of the equilibrium response ofarticular cartilage in unconfined compression, confined compression andindentation," J Biomech 35, 903-909 (2002).
[178] P. Julkunen, R. K. Korhonen, W. Herzog, and J. S. Jurvelin, "Uncertaintiesin indentation testing of articular cartilage: A fibril- reinforcedporoviscoelastic study," Med Eng Phys 30, 506-515 (2008).
[179] N. P. Camacho, P. West, P. A. Torzilli, and R. Mendelsohn, "FTIRmicroscopic imaging of collagen and proteoglycan in bovine cartilage,"Biopolymers 62, 1-8 (2001).
[180] X. Bi, X. Yang, M. P. G. Bostrom, D. Bartusik, S. Ramaswamy, K. W.Fishbein, R. G. Spencer, and N. P. Camacho, "Fourier transform infraredimaging and MR microscopy studies detect compositional and structuralchanges in cartilage in a rabbit model of osteoarthritis," Anal BioanalChem 387, 1601-1612 (2007).
[181] E. David-Vaudey, A. Burghardt, K. Keshari, A. Brouchet, M. Ries, and S.Majumdar, "Fourier Transform Infrared Imaging of focal lesions inhuman osteoarthritic cartilage," Eur Cell Mater 10, 51-60 (2005).

Bibliography
Dissertations in Forestry and Natural Sciences No 103 79
[182] M. Kim, X. Bi, W. E. Horton, R. G. Spencer, and N. P. Camacho, "Fouriertransform infrared imaging spectroscopic analysis of tissue engineeredcartilage: histologic and biochemical correlations," J Biomed Opt 10,031105 (2005).
[183] B. H. Stuart, Infrared spectroscopy: Fundamentals and applications (Wiley,Hoboken, USA, 2004).
[184] B. C. Vidal, "The part played by the mucopolysaccharides in the formbirefringence of collagen," Protoplasma 59, 472-479 (1964).
[185] A. Boskey and N. P. Camacho, "FT-IR imaging of native and tissue-engineered bone and cartilage," Biomaterials 28, 2465-2478 (2007).
[186] J. Rieppo, M. M. Hyttinen, E. Halmesmäki, H. Ruotsalainen, A. Vasara, I.Kiviranta, J. S. Jurvelin, and H. J. Helminen, "Changes in spatial collagencontent and collagen network architecture in porcine articular cartilageduring growth and maturation," Osteoarthritis Cartilage 17, 448-455 (2009).
[187] J. Rieppo, J. Hallikainen, J. S. Jurvelin, I. Kiviranta, H. J. Helminen, andM. M. Hyttinen, "Practical considerations in the use of polarized lightmicroscopy in the analysis of the collagen network in articular cartilage,"Micros Res Tech 71, 279-287 (2008).
[188] Y. Xia, J. B. Moody, N. Burton-Wurster, and G. Lust, "Quantitative in situcorrelation between microscopic MRI and polarized light microscopystudies of articular cartilage," Osteoarthritis Cartilage 9, 393-406 (2001).
[189] K. Király, M. M. Hyttinen, T. Lapveteläinen, M. Elo, I. Kiviranta, J. Dobai,L. Módis, H. J. Helminen, and J. P. A. Arokoski, "Specimen preparationand quantification of collagen birefringence in unstained sections ofarticular cartilage using image analysis and polarizing light microscopy,"Histochem J 29, 317-327 (1997).
[190] F. Massoumian, R. Juskaitis, M. A. A. Neil, and T. Wilson, "Quantitativepolarized light microscopy," J Micros 209, 13-22 (2003).
[191] E. Collett, Polarized light: fundamentals and applications (Marcel Dekker,New York, 1992).
[192] D. S. Kliger, J. W. Lewis, and C. E. Randall, Polarized light in optics andspectroscopy (Academic Press, Boston, 1990).
[193] K. Király, T. Lapveteläinen, J. Arokoski, K. Törrönen, L. Módis, I.Kiviranta, and H. J. Helminen, "Application of selected cationic dyes for

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
80 Dissertations in Forestry and Natural Sciences No 103
the semiquantitative estimation of glycosaminoglycans in histologicalsections of articular cartilage by microspectrophotometry," Histochem J28, 577-590 (1996).
[194] I. Kiviranta, J. Jurvelin, M. Tammi, A.-M. Säämänen, and H. J Helminen.,"Microspectrophotometric quantitation of glycosaminoglycans inarticular cartilage sections stained with Safranin O," Histochemistry 82,249-255 (1985).
[195] J. Töyräs, J. Rieppo, M. T. Nieminen, H. J. Helminen, and J. S. Jurvelin."Characterization of enzymatically induced degradation of articularcartilage using high frequency ultrasound," Phys Med Biol 44, 2723-33(1999).
[196] J. S. Jurvelin, M. D. Buschmann, and E. B. Hunziker, "Optical andmechanical determination of Poisson's ratio of adult bovine humeralarticular cartilage," J Biomech 30, 235-241 (1997).
[197] A. Benninghoff, "Form und Bau der Gelenkknorpel in ihren Beziehungenzur Funktion," Zeitschrift für Zellforschung und Mikroskopische Anatomie 2,783-862 (1925).
[198] J. P. A. Arokoski, J. S. Jurvelin, U. Väätäinen, and H. J. Helminen,"Normal and pathological adaptations of articular cartilage to jointloading," Scand J Med Sci Sports 10, 186-198 (2000).
[199] R. L. Sah, A. S. Yang, A. C. Chen, J. J. Hant, R. B. Halili, M. Yoshioka, D.Amiel, and R. D. Coutts, "Physical properties of rabbit articular cartilageafter transection of the anterior cruciate ligament," J Orthop Res 15, 197-203 (1997).
[200] D. A. Narmoneva, J. Y. Wang, and L. A. Setton, "Nonuniform swelling-induced residual strains in articular cartilage," J Biomech 32, 401-408(1999).
[201] I. C. Clarke, "Articular cartilage: a review and scanning electronmicroscope study. II. The territorial fibrillar architecture," J Anat 118, 261-280 (1974).
[202] I. C. Clarke, "Articular cartilage: a review and scanning electronmicroscope study. I. The interterritorial fibrillar architecture," J Bone JointSurg Br 53, 732-750 (1971).
[203] I. Youn, J. B. Choi, L. Cao, L. A. Setton, and F. Guilak, "Zonal variationsin the three-dimensional morphology of the chondron measured in situusing confocal microscopy," Osteoarthritis Cartilage 14, 889-897 (2006).

Bibliography
Dissertations in Forestry and Natural Sciences No 103 81
[204] C. A. Poole, T. Honda, S. J. M. Skinner, J. R. Schofield, K. F. Hyde, and H.Shinkai, "Chondrons from articular cartilage (II): Analysis of theglycosaminoglycans in the cellular microenvironment of isolated caninechondrons," Connect Tissue Res 24, 319-330 (1990).
[205] G. M. Lee, T. A. Paul, M. Slabaugh, and S. S. Kelley, "The incidence ofenlarged chondrons in normal and osteoarthritic human cartilage andtheir relative matrix density," Osteoarthritis Cartilage 8, 44-52 (2000).
[206] L. Han, A. J. Grodzinsky, and C. Ortiz, "Nanomechanics of the cartilageextracellular matrix," Annu Rev Mater Res 41, 133-168 (2011).
[207] S. Strasser, A. Zink, M. Janko, W. M. Heckl, and S. Thalhammer,"Structural investigations on native collagen type I fibrils using AFM,"Biochem Biophys Res Commun 354, 27-32 (2007).
[208] D. M. Ebenstein, A. Kuo, J. J. Rodrigo, A. H. Reddi, M. Ries, and L. Pruitt,"A nanoindentation technique for functional evaluation of cartilagerepair tissue," J Mater Res 19, 273-281 (2004).
[209] C. Li, L. A. Pruitt, and K. B. King, "Nanoindentation differentiates tissue-scale functional properties of native articular cartilage," J Biomed MaterRes A 78, 729-738 (2006).
[210] R. G. Haverkamp, M. A. K. Williams, and J. E. Scott, "Stretching singlemolecules of connective tissue glycans to characterize their shape-maintaining elasticity," Biomacromolecules 6, 1816-1818 (2005).
[211]A. Harder, V. Walhorn, X. Fernandez-Busquets, and D. Anselmetti,"Single-molecule force spectroscopy of cartilage aggrecan self-adhesion,"Biophys J 99, 3498-3504 (2010).
[212] N. Papaioannou, N. Krallis, I. K. Triantafillopoulos, L. Khaldi, I. Dontas,and G. P. Lyritis, "Optimal timing of research after anterior cruciateligament resection in rabbits," Contemp Top Lab Anim Sci 43, 22-27(2004).
[213] A. Changoor, L. Fereydoonzad, A. Yaroshinsky, and M. D. Buschmann,"Effects of refrigeration and freezing on the electromechanical andbiomechanical properties of articular cartilage," J Biomech Eng 132,064502 (2010).
[214] M. Szarko, K. Muldrew, and J. E. A. Bertram, "Freeze-thaw treatmenteffects on the dynamic mechanical properties of articular cartilage," BMCMusculoskeltal Disorders 11, 231 (2010).

Siru Turunen: Osmotic and mechanical loading of chondrocytes in situ
82 Dissertations in Forestry and Natural Sciences No 103
[215] K. Tavakol, R. G. Miller, D. P. Bazett-Jones, W. S. Hwang, L. E. McGann,and N. S. Schachar, "Ultrastructural changes of articular cartilagechondrocytes associated with freeze-thawing," J Orthop Res 11, 1-9(1993).
[216] J. Guan, J. P. G. Urban, and Z. H. Li, "Effects of rapid cooling on articularcartilage," Cryobiology 52, 430-439 (2006).
[217] S. Zheng, Y. Xia, A. Bidthanapally, F. Badar, I. Ilsar, and N. Duvoisin,"Damages to the extracellular matrix in articular cartilage due tocryopreservation by microscopic magnetic resonance imaging andbiochemistry," Magn Reson Imaging 27, 648-655 (2009).
[218] L. Laouar, K. Fishbein, L. E. McGann, W. E. Horton, R. G. Spencer, andN. M. Jomha, "Cryopreservation of porcine articular cartilage: MRI andbiochemical results after different freezing protocols," Cryobiology 54,36-43 (2007).
[219] M. J. Kääb, H. P. Nötzli, J. Clark, and I. ap Gwynn, "Dimensional changesof articular cartilage during immersion-freezing and freeze-substitutionfor scanning electron microscopy," Scanning Microscopy 12, 465-474(1998).
[220] J. D. Szafranski, A. J. Grodzinsky, E. Burger, V. Gaschen, H.-H. Hung,and E. B. Hunziker, "Chondrocyte mechanotransduction: effects ofcompression on deformation of intracellular organelles and relevance tocellular biosynthesis," Osteoarthritis Cartilage 12, 937-946 (2004).

Publications of the University of Eastern Finland Dissertations in Forestry and Natural Sciences
Publications of the University of Eastern Finland
Dissertations in Forestry and Natural Sciences
isbn 978-952-61-1088-2
Siru Turunen
Osmotic and MechanicalLoading of Chondrocytes in SituEffect of the Extracellular and Pericellular Matrix on
Cell Volume and Morphology
Changed chondrocyte deformations in
articular cartilage are thought to be related
to changed cell biosynthesis and viability;
alterations which may lead to and accelerate
the progression of osteoarthritis (OA). How-
ever, it is not known whether the structural
changes of cartilage in early OA are concur-
rent with the changes in the biomechanical
responses of chondrocytes. The aim of this
thesis was to reveal how the structure and
composition of the extra- and pericellular
matrices of cartilage affect chondrocyte
volume and morphology under osmotic and
mechanical loading of the tissue. By under-
standing chondrocyte behavior in situ/in
vivo it might be possible to devise strategies
to slow down, stop or even reverse the devel-
opment of OA.
dissertatio
ns | 103 | S
iru
Tu
ru
nen
| Osm
otic and Mechanical L
oading of Chondrocytes in S
itu - Effect of the E
xtracellular and Pericellular M
atrix...
Siru Turunen
Osmotic and MechanicalLoading of Chondrocytes in Situ
Effect of the Extracellular and Pericellular Matrix on
Cell Volume and Morphology