of the slug, arion lusitanicus ecophysiology and life
TRANSCRIPT

AARHUSUNIVERSTYAU
ECOPHYSIOLOGY AND LIFE HISTORY OF THE SLUG, ARION LUSITANICUS
The slug Arion lusitanicus is an invasive species which has recently become established in many European countries and is considered a serious pest, both in agriculture and private gardens. The aim of this PhD was to describe in detail the ecophysiology of A. lusitanicus and map its life history traits in order to reveal how life history traits respond to major environmental parameters such as temperature and moisture. Knowledge of the species’ biology is neces-sary to generate eff ective pest management plans.
This PhD project contributes with detailed knowledge of the ecophysiology and life history traits of A. lusitanicus. A growth model that enables the prediction of individual slug growth under natural temperature scenarios has been de-veloped. The project has shown how egg developmental time and hatchability are infl uenced by temperature and drought. Furthermore, the cold and drought tolerance limits of A. lusitanicus have been revealed. The results will be a valuable contribution to the development of population growth models to provide a more accurate forecasting of slug abundance as a tool for pest management.
Ecop
hysiolo
gy a
nd life
history of the
slug, A
rion lusitanicus
PhD thesis 2012Stine Slotsbo
ECOPHYSIOLOGY AND LIFE HISTORY OF THE SLUG, ARION LUSITANICUS
ISBN: 978-87-92825-xx-x


ECOPHYSIOLOGY AND LIFE HISTORY OF THE SLUG, ARION LUSITANICUS
Stine Slotsbo
PhD thesis 2012
AARHUSUNIVERSITYAU

Title: Ecophysiology and life history of the slug, Arion lusitanicus Subtitle: PhD thesis Author: Stine Slotsbo Institute: Department of Agroecology and Department of Bioscience, Aarhus University
Publisher: Aarhus University - Denmark URL: http://www.dmu.au.dk
Year of publication: 2012 Editing completed: July 2012
PhD supervisors: Professor Martin Holmstrup, Aarhus University, Department of BioscienceSenior Scientist Lars Monrad Hansen, Aarhus University, Department of Agroecology
Assessment committee: Professor Hans Ramløv, Department of Science, Systems and Models, Roskilde University, DenmarkProfessor William O.C. Symondson, Cardiff School of Biosciences, Cardiff University, UKSenior Scientist Niels Holst (chairman), Aarhus University, Department of Agroecology, Denmark
Please cite as: Slotsbo, S. 2012. Ecophysiology and life history of the slug, Arion lusitanicus. PhD thesis. Aarhus University, Department of Agroecology, Denmark. 80 pp.
Reproduction permitted provided the source is explicitly acknowledged
Abstract: The slug Arion lusitanicus is an invasive species which has recently become established in many European countries and is considered a serious pest, both in agriculture and private gardens. The aim of this PhD was to describe in detail the ecophysiology of A. lusitanicus and map its life history traits in order to reveal how life history traits respond to major envi-ronmental parameters such as temperature and moisture. Knowledge of the species’ bio-logy is necessary to generate eff ective pest management plans.
This PhD project contributes with detailed knowledge of the ecophysiology and life history traits of A. lusitanicus. A growth model that enables the prediction of individual slug growth under natural temperature scenarios has been developed. The project has shown how egg developmental time and hatchability are infl uenced by temperature and drought. Fur-thermore, the cold and drought tolerance limits of A. lusitanicus have been revealed. The results will be a valuable contribution to the development of population growth models to provide a more accurate forecasting of slug abundance as a tool for pest management.
Keywords: Arion lusitanicus, Iberian slug, slug, growth, growth model, freeze tolerance, supercooling, drought, desiccation stress, water conductance, invasive species
Layout: Tinna Christensen Cover photo: Arion lusitanicus. Photo: Stine Slotsbo.
ISBN: 978-87-92825-xx-x
Number of pages: 80
Printed by: Rosendahls – Schultz Grafi sk a/s
Circulation: xx Supplementary notes: xxxxxxxxxxxxxxxxxxxxxxxxxxxxxx. Time: xxxxxxxxxxxxxxxxxxxxxx Place: xxxxxxxxxxxxxxxxxxxxxxx
Data sheet

Content
Preface 5
English summary 6
Dansk resumé 7
REVIEW 9
Abstract 10
Introduction 11
Classifi cation and identifi cation 12
Life history traits 13
Cold tolerance of Arion lusitanicus 19
Drought tolerance of Arion lusitanicus 24
Conclusion and perspectives 27
References 29
PAPER I 35
The infl uence of temperature on life history traits in the Iberian slug, Arion lusitanicus
PAPER II 51
Low temperature survival in diff erent life stages of the Iberian slug,Arion lusitanicus
PAPER III 59
Cold tolerance and freeze-induced glucose accumulation in threeterrestrial slugs
PAPER IV 69
Drought tolerance in eggs and juveniles of the Iberian slug, Arion lusitanicus

Ario
n lu
sita
nicu
s. P
hoto
: Stin
e S
lots
bo
.

5PhD thesis by Stine Slotsbo
Preface
This PhD thesis was submitted to the Graduate School of Science and Technology (GSST), Aarhus University, Denmark. Most of the work has been conducted at the Department of Bioscience, Silkeborg, Aarhus University, Denmark. The project was funded by the Department of Agroecology, Flakkebjerg, and the Department of Bioscience, Silkeborg, Aarhus University, Denmark. The objective of this PhD was to describe in detail the ecophysiology of A. lusitanicus and map its life history traits in order to reveal how life history traits respond to major environmental parameters such as temperature and moisture.
I would like to thank several people, who have contributed to the completion of this project. First of all, sincere thanks to my two supervisors Martin Holmstrup and Lars Monrad Hansen for your supervision, guidance and great support both sci-entifi cally and personally. Thanks Martin for always being there when needed, and for your critical review of this thesis. Thanks to all my colleagues at Biosci-ence Silkeborg for providing an inspiring research community, for unsurpassed technical assistance, and lots of fun. A very special thank you to Dorthe Waagner, Jesper Givskov Sørensen and Karina V. Fisker for sparring, invaluable discussions and many hours of fun. I would also like to thank my former colleagues Kristine Maraldo, Anne Mette Bindelsbøl, Lars-Henrik Heckman and Vibeke Simonsen.
During my stay at the University of Antwerp, Kurt Jordaens introduced me to spe-cies identifi cation of slugs. A special thank you goes to Kurt for always being there with help and guidance, only an email away. I would also like to thank Bjørn Hat-teland and Torstein Solhøj for an inspiring and pleasant stay at the University of Bergen, not to mention Anders Malmendal and Thierry Backeljau for invaluable co-operation.
I am grateful to Vagn and Gerda Nielsen for the many interesting photos of A. lusitanicus, and for letting me use their garden for fi eld work. Also thanks to Sussi Pagh for many useful and fun hours in the fi eld. Several other people have helped - I appreciate all your eff orts.
Last, but not least, I thank my friends and family for their support and encourage-ment. Especially my husband Mads for caring and support, particularly during my travels.
Stine SlotsboStine Slotsbo

6 PhD thesis by Stine Slotsbo
English summary
The slug Arion lusitanicus is an invasive species which has recently become es-tablished in many European countries and is considered a serious pest, both in agriculture and private gardens. The aim of this PhD was to describe in detail the ecophysiology of A. lusitanicus and map its life history traits in order to reveal how life history traits respond to major environmental parameters such as temperature and moisture. Knowledge of the species’ biology is necessary to generate eff ec-tive pest management plans.
This PhD project contributes with detailed knowledge of the ecophysiology and life history traits of A. lusitanicus. A growth model that enables the prediction of individual slug growth under natural temperature scenarios has been developed. The project has shown how egg developmental time and hatchability are infl u-enced by temperature and drought. Furthermore, the cold and drought tolerance limits of A. lusitanicus have been revealed. The results will be a valuable contribu-tion to the development of population growth models to provide a more accurate forecasting of slug abundance as a tool for pest management.

7PhD thesis by Stine Slotsbo
Dansk resumé
Sneglen Arion lusitanicus er en invasiv art, der for nylig har etableret sig i mange Europæiske lande, hvor den anses for at være et alvorligt skadedyr både i land-bruget og i private haver. Formålet med dette PhD projekt var at beskrive Arion lusitanicus’s økofysiologi i detaljer, samt kortlægge dens livshistorietræk, og hvor-dan disse påvirkes af miljøparametre såsom temperatur og fugtighed. For at være i stand til at udvikle en eff ektiv skadedyrsbekæmpelse er det nødvendigt at have et godt kendskab til artens biologi.
Denne PhD bidrager med detaljeret viden om A. lusitanicus´s økofysiologi samt livshistorietræk. En vækstmodel, der kan forudsige den enkelte snegls vækst un-der naturlige temperaturscenarier, er blevet udviklet. Desuden har projektet vist, hvordan ægudviklingen og ægs klækkeevne påvirkes af temperatur og tørke, og fastlagt kulde- og tørkegrænseværdierne for A. lusitanicus. Dette PhD projekt bi-drager med vigtig viden til udviklingen af populationsvækstmodeller og progno-seværktøjer.


9PhD thesis by Stine Slotsbo
REVIEW
Mating. Photo: Vagn Nielsen.

10 PhD thesis by Stine Slotsbo
Abstract
The slug Arion lusitanicus is an invasive species which has recently become established in many Eu-ropean countries and is considered a serious pest, both in agriculture and private gardens. The popu-lation size of A. lusitanicus can vary substantially between years, likely due to climatic factors such as temperature and precipitation. This paper reviews the existing literature on ecophysiology and life his-tory of A. lusitanicus. The review also provides a short introduction to the classifi cation and identifi cation of A. lusitanicus, since this has been a major problem in slug research. A. lusitanicus has an annual life cycle and most adults die in the fall soon after egg-laying. The growth, egg developmental time and egg hatchability of A. lusitanicus are infl uenced by temperature. Rise in temperature will, below 25°C, result in an increase in growth rate. A. lusitanicus has a high degree of plasticity in its thermal biology and can quickly adjust growth with changes in temperature. A. lusitanicus is a freeze-tolerant species both in the juvenile and adult stages, and can survive freezing at temperatures down to -2°C. The egg stage is not freeze-tolerant and survives by supercooling. The winter survival of A. lusitanicus in cold regions, therefore, depends to a high degree on migration to habitats protected from low winter tem-peratures. During drought periods A. lusitanicus has, like other slugs, only little resistance to evaporative water loss, but on the other hand tolerate a substantial water loss. The eggs do not diff er from juveniles in water loss rates, but are slightly more tolerant to water loss. Despite tolerance of substantial water loss, A. lusitanicus depends on humid habitats. Very dry springs and summers will delay the egg-laying period in the given year, and the development time of the eggs increases with increasing desiccation. The challenges in using life-history trait data as a basis for population growth models and forecasting of population dynamics are discussed.
10 PhD thesis by Stine Slotsbo10 PhD thesis by Stine Slotsbo
Pho
to: S
tine
Slo
tsb
o.

11PhD thesis by Stine Slotsbo
Introduction
Slugs are important pests of a wide range of horticultural and arable crops across Europe (South 1992), and the more frequent use of conservation tillage in agri-culture is leading to an increase in slug pest problems around the world (Norris et al. 2003). The reduction of mechanical disturbance of the soil and the retention of litter due to conservation tillage favours slug survival and reproduction (Hunter 1967; Hammond 1985) . Further, the spread of invasive species, including slugs, is an increasing problem worldwide. Along with escalating global trade, transporta-tion and international travel, the introduction and spread of invasive species are increasing in frequency (Meyerson & Mooney 2007; Hulme 2009).
The slug Arion lusitanicus Mabille (Gatropoda: Pulmonata: Arionidae) is an inva-sive species which has spread and become established in many European coun-tries since the 1950ies (Quick 1952; Von Proschwitz 1992; Von Proschiwtz & Winge 1994; Kerney 1999; Kozlowski 2007) and is considered a serious pest, both in agri-culture and private gardens (Frank 1998; Kozlowski 2007). Earlier, it was assumed to originate from the Iberian Peninsula, a misconception that gave rise to its com-mon name, the Iberian slug (Quinteiro et al. 2005). It is possible that A. lusitanicus has spread to the Nordic countries from southern regions, however, the precise region of origin is unknown (Quinteiro et al. 2005).
The fi rst verifi ed record of A. lusitanicus in Scandinavia was reported from Sweden in 1975 (Von Proschwitz 1992). In Norway, the fi rst observation was made in 1988 (Von Proschwitz 1989) and in Denmark, the fi rst identifi cation was made in 1991 (Von Proschwitz 1992), however, it is possible that the species has been in these countries for several years prior to being discovered.
A. lusitanicus seems to cope well with local conditions in the new areas to which it has migrated, it spreads rapidly and can occur in very high densities. The popu-lation size of A. lusitanicus can vary substantially between years (Grimm & Paill 2001) most likely due to climatic factors such as temperature and humidity, but knowledge of A. lusitanicus’ ecophysiology is limited. Nature conservation authori-ties and managers need knowledge of the biology and ecophysiology of this pest in order to generate eff ective pest management plans.
The aim of this PhD was to describe in detail the ecophysiology of A. lusitanicus and map its life history traits in order to reveal how life history traits respond to major environmental parameters such as temperature and moisture. Some of the research questions in the resulting papers were: how does temperature aff ect growth rate, survival, egg developmental time and hatchability in A. lusitanicus (Paper I)? Can A. lusitanicus survive sub-zero temperatures and by which cold tolerance strategy (Paper II)? Are there diff erences in cold tolerance between A. lusitanicus and the two closely related species A. ater and A. rufus (Paper III)? How sensitive to drought are eggs and juveniles of A. lusitanicus and does drought af-fect egg development (Paper IV)?
The purpose of the present review is to synthesize the existing literature on eco-physiology and biology of A. lusitanicus, including my own work. This review be-gins with a short introduction to the classifi cation and identifi cation of A. lusitanicus, because one major problem in slug research is the diffi culties of slug classifi cation.

PhD thesis by Stine Slotsbo12
Classifi cation and identifi cation
Slugs and snails belong to the class Gastropoda; snails have a fully developed external shell while slugs have a reduced or a small internal shell that sometimes only consists of a few granules (South 1992). Slugs are not a natural group of closely related gastropods, but have evolved several times from diff erent, strongly diverged snail groups (Barker 2001). Terrestrial snails and slugs belong, with few exceptions, to the subclass pulmonata (South 1992).
Because the taxonomy of pulmonates is largely based on characteristics of the shell, the classifi cation of slug species continues to be a point of debate. Slugs are often distinguished on the basis of their body pigmentation and the structure of their reproductive organs, but body pigmentation may depend on age, diet or environment, and the reproductive organs can be subjected to considerable ontogenetic, seasonal and physiological variation (Barker 2001; Jordaens et al. 200 1; 2002 and references therein). Moreover, several European slug “species” ap-pear rather to be species complexes that are diffi cult, sometimes even impossible, to distinguish using morphology alone (e. g. Backeljau et al. 1996; Noble & Jones 1996; Pinceel et al. 2005; Engelke et al. 2011). Therefore, molecular techniques have become an important part in species identifi cation.
A case in point is the genus Arion that currently comprises 30-50 species, of which many belong to complexes of cryptic species and whose identifi cation remains problematic. Especially the ‘large arionids’ continue to provoke debate. In NW Eu-rope, the large arionids comprise Arion rufus (Linnaeus, 1758), A. ater (Linnaeus, 1758), A. lusitanicus (Mabille, 1868) and A. fl agellus (Collinge, 1893). Currently, the taxonomic status of A. ater and A. rufus is still unclear, and there is no consensus as to whether they should be regarded as species or subspecies (e.g. Quinteiro et al. 2005; Engelke et al. 2011 and references therein). In this review, I treat A. rufus and A. ater as two diff erent taxa. The taxonomic status of A. lusitanicus remains unresolved. The invasive A. lusitanicus occurring in NW Europe diff ers from A. lusi-tanicus of the Iberian Peninsula in its internal anatomy and the shape of the sper-matophore (Castillejo 1997). Some authors (e.g. Anderson 2005) use the name A. vulgaris (Moquin-Tandon, 1855) for the former, whereas others prefer the name A. lusitanicus (e.g. Quinteiro et al. 2005; Hatteland 2010). Throughout this review I will follow Bank et al. (2007) and use the name A. lusitanicus when referring to the non-Iberian A. lusitanicus.

13PhD thesis by Stine Slotsbo
Life history traits
Lifecycle
All terrestrial slugs are hermaphrodites; some reproduce by self-fertilization, some by out-crossing, and others do a combination of both (South 1992). A. lusitanicus is mainly out-crossing, but is able to self-fertilize, even though this does not com-monly occur (Hagnell et al. 2006; Slotsbo et al. 2012b).
A. lusitanicus has an annual life cy cle (Davies 1987; Kozlowski & Kozkowski 2000; Grimm 2001; Slotsbo et al. 2012b), but occasionally large overwintered specimens can be found in the sp ring (Davies 1987; Kozlowski 2007), indicating that semivolt-ine life-cycles also occur. Laboratory studies show that A. lusitanicus may potentially live for up to three years, as long as they do not reproduce (Slotsbo et al. 2012b). Field observations from Poland, Great Britain and Switzerland (Davies 1987; Briner & Frank 1998) indicate that the majority of A. lusitanicus die soon after egg-laying and this is confi rmed by laboratory studies (Kozlowski & Sionek 2000; Slotsbo et al. 2012b). Kozlowski and Sionek (2000) found that 75 % of the slugs kept in laboratory cultures died after egg-laying before the winter season and Slotsbo et al. (2012b) confi rmed that A. lusitanicus dies after egg-laying and showed that the mortality was not infl uenced by ambient temperature or age of the slugs. Post-reproductive mortality has also been found in the related slug A. rufus (Rollo 1983), and therefore an annual lifecycle seems to be the most common in these species.
A description of the mating behaviour of A. lusitanicus has been reported by Davies (1987). Egg-laying may begin in late June and in some years continues until De-cember if the temperature is not too low; the exact time varies depending on geo-graphical region and also fl uctuates between years within an area. However, egg deposition most often peaks in August and Sept ember (Kozlowski 2000; Kozlowski & Sionek 2000; Grimm 2001; Slotsbo unpublished). Eggs are laid in clutches of up to 225, and one slug can lay several cl utches (Briner & Frank 1998; Kozlowski 2000; Kozlowski & Sionek 2000; Slotsbo unpublished). Slugs may produce more than 500 eggs, but a single slug normally lays 200-400 eggs (Kozlowski 2000). The eggs are deposited in a variety of places; occasionally unprotected on the surface of bare soil, but most often in more protected places, such as under vegetation or objects lying on the ground or in the soil to a depth of 10 cm. Eggs are often laid in places ensuring humid conditions (Kozlowski 2000).
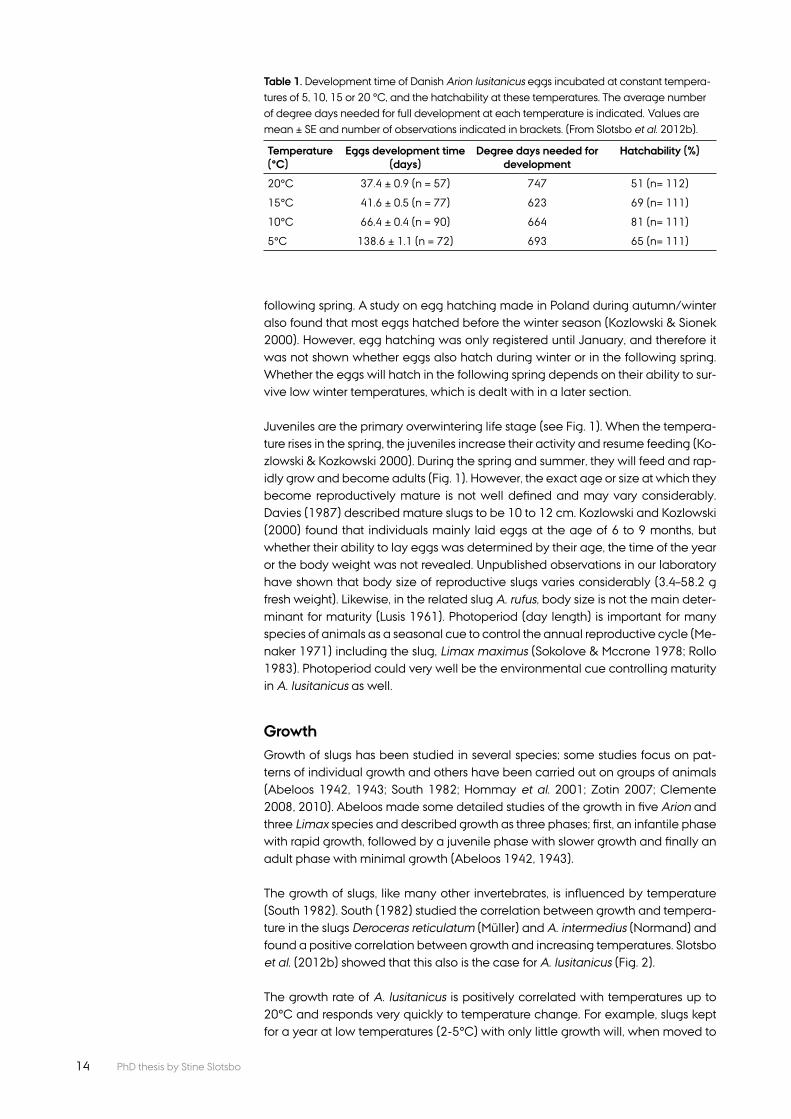
The egg developmental time and hatching success are infl uenced by tempera-ture and may vary between regions (Kozlowski 2000; Slotsbo et al. 2012b). Thus, eggs from Denmark developed fastest at 20°C and hatched in 37 days (Slotsbo et al. 2012b) (Table 1). Eggs laid by slugs from Poland also developed fastest at 20°C, but hatched within 30 days (Kozlowski 2000). Not surprisingly, high tem-perature increases egg development up to a certain limit. Kozlowski (2000) found no embryonic development or hatching at 25°C, which appears to be the upper temperature limit for A. lusitanicus. Even though 20°C resulted in the most rapid development, this temperature was not the optimum for hatching success (Table 1). The optimum temperature appears to be between 10 and 15°C.
On a degree-day scale, egg development is completed in 625-750 degree days (Slotsbo et al. 2012b). Using this information together with the average tempera-tures of Denmark, it can be shown that most eggs will hatch before the winter sea-son. Eggs laid as late as mid-September in Denmark are expected to hatch in the beginning of December. Eggs laid after October 1 will probably not hatch until the
14 PhD thesis by Stine Slotsbo
following spring. A study on egg hatching made in Poland during autumn/winter also found that most eggs hatched before the winter season (Kozlowski & Sionek 2000). However, egg hatching was only registered until January, and therefore it was not shown whether eggs also hatch during winter or in the following spring. Whether the eggs will hatch in the following spring depends on their ability to sur-vive low winter temperatures, which is dealt with in a later section.
Juveniles are the primary overwintering life stage (see Fig. 1). When the tempera-ture rises in the spring, the juveniles increase their activity and resume feeding (Ko-zlowski & Kozkowski 2000). During the spring and summer, they will feed and rap-idly grow and become adults (Fig. 1). However, the exact age or size at which they become reproductively mature is not well defi ned and may vary considerably. Davies (1987) described mature slugs to be 10 to 12 cm. Kozlowski and Kozlowski (2000) found that individuals mainly laid eggs at the age of 6 to 9 months, but whether their ability to lay eggs was determined by their age, the time of the year or the body weight was not revealed. Unpublished observations in our laboratory have shown that body size of reproductive slugs varies considerably (3.4–58.2 g fresh weight). Likewise, in the related slug A. rufus, body size is not the main deter-minant for maturity (Lusis 1961). Photoperiod (day length) is important for many species of animals as a seasonal cue to control the annual reproductive cycle (Me-naker 1971) including the slug, Limax maximus (Sokolove & Mccrone 1978; Rollo 1983). Photoperiod could very well be the environmental cue controlling maturity in A. lusitanicus as well.
GrowthGrowth of slugs has been studied in several species; some studies focus on pat-terns of individual growth and others have been carried out on groups of animals (Abeloos 1942, 1943; South 1982; Hommay et al. 2001; Zotin 2007; Clemente 2008, 2010). Abeloos made some detailed studies of the growth in fi ve Arion and three Limax species and described growth as three phases; fi rst, an infantile phase with rapid growth, followed by a juvenile phase with slower growth and fi nally an adult phase with minimal growth (Abeloos 1942, 1943).
The growth of slugs, like many other invertebrates, is infl uenced by temperature (South 1982). South (1982) studied the correlation between growth and tempera-ture in the slugs Deroceras reticulatum (Müller) and A. intermedius (Normand) and found a positive correlation between growth and increasing temperatures. Slotsbo et al. (2012b) showed that this also is the case for A. lusitanicus (Fig. 2).
The growth rate of A. lusitanicus is positively correlated with temperatures up to 20°C and responds very quickly to temperature change. For example, slugs kept for a year at low temperatures (2-5°C) with only little growth will, when moved to
Table 1. Development time of Danish Arion lusitanicus eggs incubated at constant tempera-tures of 5, 10, 15 or 20 °C, and the hatchability at these temperatures. The average number of degree days needed for full development at each temperature is indicated. Values are mean ± SE and number of observations indicated in brackets. (From Slotsbo et al. 2012b).
Temperature (°C)
Eggs development time (days)
Degree days needed for development
Hatchability (%)
20°C 37.4 ± 0.9 (n = 57) 747 51 (n= 112)
15°C 41.6 ± 0.5 (n = 77) 623 69 (n= 111)
10°C 66.4 ± 0.4 (n = 90) 664 81 (n= 111)
5°C 138.6 ± 1.1 (n = 72) 693 65 (n= 111)

15PhD thesis by Stine Slotsbo
higher temperatures (15°C), quickly resume growth rates similar to specimens kept at constant 15°C (Slotsbo et al. 2012b) (Fig. 3). Being able to survive for long peri-ods with low temperature and halted growth must be an advantage as it means that metabolic requirements are low during winter when food is limited. Further, it is quite remarkable and demonstrates a high degree of plasticity that slugs even after a year of very slow growth, without any delay, are able to obtain rapid growth rates when temperatures increase. In the spring, this means that overwintering ju-veniles immediately start growing when the temperature rises, provided that food is not a limiting factor. Due to this plasticity of the species, the population size of A. lusitanicus can vary substantially between years (Grimm & Paill 2001), most likely due to climatic factors such as temperature and precipitation.
Figure 1. Life cycle of Arion lusitanicus.
Winter
A majority of the adults
die
Summer
A
eer
Mating Laying of eggs
Hatching
Juvenile
Adult
Juvenile
Age (days)0 100 200 300 400
Fre
sh w
eigh
t (g)
0
5
10
15
20
25
30 2°C 5°C 10°C 15°C 20°C
Figure 2. Growth of Arion lusitani-cus kept at constant temperatures of 2°C, 5°C, 10°C, 15°C or 20°C.Values are mean ± SE (N = 6-30).(From Slotsbo et al. 2012b)

16 PhD thesis by Stine Slotsbo
Being a pest species of economic importance, D. reticulatum is well described with respect to growth (South 1982; Zotin 2007; Clemente 2008, 2010). However, the models developed to describe population growth of D. reticulatum (Shirley et al. 2001; Choi et al. 2004; Willis et al. 2006) cannot be used to describe the population dynamics of A. lusitanicus because the two species diff er in growth pattern and life cycle. For example, D. reticulatum is assigned to animals with fi nite growth, where A. lusitanicus seems to have infi nite growth (Slotsbo et al. 2012b). Furthermore, D. reticulatum normally has several generations each year, whereas A. lusitanicus only has one (South 1982; Kozlowski & Kozlowski 2000; Grimm 2001; Willis et al. 2008).
A deterministic model of individual growth using ambient temperature as the driving factor has been developed for A. lusitanicus (Slotsbo et al. 2012b). This model was developed using laboratory data, but a fi eld survey found that the predicted growth matched the observed with reasonable accuracy (Slotsbo et al. 2012b) (Fig. 4).
Cold tolerance Cold tolerance of terrestrial invertebrates in temperate and cold regions has been widely studied, as these organisms often face temperatures during winter that may cause freezing of their body fl uids. The protective strategies of cold hardy in-vertebrates are often divided into two major strategies; freeze tolerance or freeze av oidance (Zachariassen 1985; Ramløv 2000). Freeze tolerant species seek to establish controlled, protective freezing of the extracellular body fl uids at high sub-zero temperatures, whereas freeze avoiding species have developed mech-anisms to stay unfrozen even at temperatures much below the melting point of bod y fl uids (Sømme 1982; Zachariassen 1985; Storey & Storey 1996).
Fre
sh w
eigh
t (g)
Age (days)0 100 200 300 400 500 600 700
0
5
10
15
20
2515°C
Figure 3. Growth of juvenile Arion lusitanicus raised at a constant temperature of either 2°C or 5°C for 358 days and then moved to 15°C and kept there. The triangles show growth of slugs raised at constant 15°C, from the time that they had reached the same fresh weight as slugs kept for 358 days at 2°C and 5°C, respectively (see Fig. 2). Values are mean ± SE. (From Slotsbo et al. 2012b)
Fre
sh w
eigh
t (g)
1st Apr 1st May 1st Jun 1st Jul 1st Aug 1st Sep 1st Oct
0
4
8
12
16
Figure 4. The average fresh weight (g) of Arion lusitanicus captured in the fi eld at 20 diff erent locations in East Jutland in the period from 1 April to 1 October, 2009. Each black dot represents the average fresh weight (±SE; N = 1-36) of slugs captured at that day. The solid line shows the calculated growth using the growth model developed in Slotsbo et al. (2012b) and the dai-ly mean temperature in East Jut-land, Denmark, in the given period. The data within the black triangle (lower right corner) are most likely juveniles that hatched in late Sep-tember/October (From Slotsbo et al. 2012b).

17PhD thesis by Stine Slotsbo
Freeze toleranceFreeze tolerant animals face several challenges when undergoing freezing of their body fl uids. Non-cold-adapted cells will get injuries at temperatures below zero due to both low temperature and ice formation. Low temperature can aff ect the cell metabolism and the viscosity of the cell membrane. Growing ice crystals may pene-trate and rupture tissues and cell membranes (Roberts et al. 1987). Ice formation will naturally reduce the amount of free water in the organisms. Because ice crystals only consist of water, freezing of body fl uid results in an increase in the solute con-centrations in the extracellular fl uid, which leads to an osmotic outfl ow of water from the cells and, consequently, dehydration of the cells (Zachariassen 1985). The in-creases in solute concentrations, both extra- and intracellular, may lead to changes in enzyme activity and denaturation of proteins (Ramløv 2000). Extensive dehydra-tion of cells may cause phase change in the membranes (Hazel 1995).
Freeze tolerant organisms therefore require physiological and biochemical mecha-nisms that protect them against these challenges. The induction of ice crystals at relatively high sub-zero temperatures helps control the development of ice, ensur-ing protective extracellular freezing and preventing injurious freezing in other fl uid compartments (Storey & Storey 1992). Early crystallisation can be achieved by ice nucleating agents, which induce freezing at high sub-zero temperatures (Zacha-riassen 1985). Another well-known protection mechanism is the accumulation of compatible osmolytes, such as sugars and polyols. These osmolytes, called cryo-protectants, act colligatively and reduce the amount of ice at a given temperature and protect cells against dehydration (Storey & Storey 1988). Further, they can help stabilize membranes and proteins when the body fl uids freeze (Crowe et al. 1987; Storey & Storey 1992). Lastly, a very important protective mechanism is the ability to change the composition of cellular lipids, which aid to preserve an appropriate fl uidity of the cell membrane under changing temperatures (Hazel 1995).
Freeze avoidanceFreeze avoiding ectotherms have the challenge of staying unfrozen when the surroundings freeze. When unable to escape frost, a common strategy for freeze avoiding species is prolonged supercooling. The temperature to which a fl uid can be cooled before it spontaneously freezes is called the supercooling point. Freeze avoiding ectotherms must lower the supercooling point well below the ambient temperature and stabilize the supercooled state (Zachariassen 1985). Supercool-ing is promoted by removal of ice nucleating particles and production of cryopro-tectants. Accumulation of high concentrations of cryoprotectants such as sugars and polyols depress the supercooling point and stabilize membranes and cellu-lar proteins at low temperature (Zachariassen 1980). Furthermore, some freeze avoiding ectotherms produce antifreeze proteins that depress the freezing point by inhibiting the growth of seeding ice crystals (Duman et al. 1982; Ramløv 2000).
Some small permeable freeze avoiding ectotherms are able to use a diff erent cold hardiness strategy, namely cryoprotective dehydration. Small permeable or-ganisms will lose water if the water vapour pressure in equilibrium with their body fl uid is larger than the water vapour pressure in the surrounding atmosphere. The vapour pressure of supercooled water is higher than that in equilibrium with ice (Salt 1963; Weast 1989). Therefore, in a frozen environment, supercooled perme-able organisms will lose water until equilibrium in water vapour pressure is at-tained. The melting point of the body fl uid is then in equilibrium with the surround-ing temperature, so that no freezing can occur. Organisms using this strategy must be small in order for the dehydration process to occur rapidly enough to avoid ice formation, and to be successful, they must tolerate extensive d ehydration (Holm-strup et al. 2002).

18 PhD thesis by Stine Slotsbo
Cold tolerance of slugsA wide range of invertebrates, especially the arthropods, have been the focus of cold tolerance research, but cold tolerance of terrestrial molluscs has received less attention (Ansart & Vernon 2003), and only few studies are concerned with ter-restrial slugs (Getz 1959; Mellanby 1961; Cook 2004; Storey et al. 2007; Udaka et al. 2008; Berman et al. 2011; Slotsbo et al. 2011b, 2012a). Both freeze tolerance and freeze avoidance strategies occur within terrestrial Mollusca. Several species of land snails survive winters using freeze avoidance as a cold hardiness strat-egy (Riddle 1981; Riddle & Miller 1988). However, in contrast to land snails, slugs lack physical protection such as shell, epiphragm and operculum. The lack of this protection likely makes them susceptible to inoculative freezing, and therefore adoption of a freeze avoidance strategy seems inappropriate. Hence, their winter survival may depend on the ability to fi nd frost-free hibernation sites or their use of freeze tolerance as a strategy. Recent studies show that some terrestrial slugs are indeed free ze tolerant (Storey et al. 2007; Berman et al. 2011; Slotsbo et al. 2011b, 2012a) (Table 2).
Freeze tolerance is a relative term, since survival in many species depends on both temperature and time in the frozen state. Some authors use the term partially freeze tolerant about species that can survive a low fraction of internal ice over short periods of time (e.g. hours) (Sinclair 1999; Ansart et al. 2001; Cook 2004). It is unclear, however, how ecologically relevant partial freeze tolerance is as a strat-egy. Ecologically relevant freeze tolerance usually implies that a species is able to survive in the frozen state for l onger periods (Storey & Storey 1988; Storey et al. 2007), and often such species endure the conversion of more than 50% of their body water into ext racellular ice (Storey & Storey 1988; Storey et al. 2007).
Table 2. Overview of experiments of cold tolerance in slugs.
Species Exposure regime Population Reference
Freeze tolerant
Deroceras laeve Frozen for 5 h at –8°C North America near Michigan Getz 1959
Deroceras laeve Frozen for 1 h at –2°C Southern parts of Canada Storey et al. 2007
Deroceras laeve Frozen for 3–4 months at –10°CLowest temperature survived was –28°C
Siberian near Magadan Berman et al. 2011
Arion lusitanicus Frozen for 3 days at –2°C Denmark Slotsbo et al. 2012a
Arion rufus Frozen for 3 days at –2°C Denmark Slotsbo et al. 2012a
Arion ater Frozen for 2 days at –1°C Denmark Slotsbo et al. 2012a
Partially freeze tolerant
Deroceras reticulatum Frozen for 30 min at –4.8°C Southeast England Cook 2004
Deroceras reticulatum Frozen for 1 h at –1.5°CTolerate ice content of max 44 %
Southern parts of Canada Storey et al. 2007
Arion circumscriptus Frozen for 1 h at –1.2°C, Tolerate ice content of only 23 %
Southern parts of Canada Storey et al. 2007
Deroceras agreste Frozen for 1 day at –1°C (only 2 out of 5 survive this)
Siberian near Magadan Berman et al. 2011
Freeze intolerant
Deroceras reticulatum Do not survive freezing North America near Michigan Getz 1959
Arion circumscriptus Do not survive freezing North America near Michigan Getz 1959
Deroceras reticulatum Do not survive freezing Siberian near Magadan Berman et al. 2011
Deroceras altaicum Do not survive freezing Siberian near Magadan Berman et al. 2011
Unknown strategy
Lehmania valentiana Survive 1 h at – 8°C (but no information on whether slugs were frozen)
Osaka Japan Udaka et al. 2008

19PhD thesis by Stine Slotsbo
The freeze tolerance within a species can vary extensively between diff erent geo-graphical populations. Populations of the same species from colder regions might be more freeze tolerant than populations from less cold regions. One example is the species D. laeve, in which freeze survival down to –28°C has been reported for a Siberian population collected near Magadan, where winter temperatures are extremely low (Berman et al. 2011). Studies of D. laeve collected elsewhere do not show the same degree of freeze tolerance. D. laeve collected in Michigan survived only –8°C (Getz 1959) and D. laeve collected in south-eastern Ontario survive only –2°C, fo r short periods (Storey et al. 2007) (Table 2).
Cold tolerance of Arion lusitanicus
Survival of freezing
A. lusitanicus primarily overwinters as juveniles, but adults and eggs can also be found during winter. Cold tolerance of A. lusitanicus has been studied in some Danish popula-tions, showing that A. lusitanicus is indeed freeze-tolerant, although these slugs only tolerate moderate sub-zero tem-peratures. In a study by Slotsbo et al. (2011b), most juveniles and some adult slugs survived being frozen at –1.3°C for three days with an estimated ice content of 60-70 %. This large amount of ice corroborates that A. lusitanicus is in-deed surviving ecologically realistic freezing conditions by the freeze tolerance strategy. Another study of A. lusitanicus showed that some survival also occurred at –2°C (Slotsbo et al. 2012a).
Juvenile A. lusitanicus survive freezing better than adults (Slotsbo et al. 2011b). This link between small size and bet-ter freeze survival has also proved to be true for freeze toler-ant earthworms (Holmstrup et al. 2007). The melting point of
the body fl uid of A. lusitanicus is positively correlated with their body weight (Slots-bo et al. 2011b). This could explain the better survival of smaller slugs, because a lower melting point will result in a lower amount of ice at a given temperature.
A. lusitanicus typically overwinters deep in the litter layer or even buried deep in the soil (Kozlowski 2000). Danish A. lusitanicus does not endure very low tempera-tures, but the temperatures met in their overwintering habitat correspond well to the lower lethal temperature, which is not much below –2°C. Several studies show that soil temperatures in subnivean and subterranean Danish habitats are highly buff ered and seldom reach more than a few degrees below zero (Rundgren 1975; Overgaard et al. 2009).
In response to freezing, A. lusitanicus accumulates cryoprotectants. Metabolite pro-fi les of frozen A. lusitanicus have shown that glucose, lactate, and succinate accu-mulate during freezing of body fl uids (Slotsbo et al. 2012a). Glucose accumulation indicated that glycogen is catabolized as a response to freezing of body fl uids. The mobilisation of cryoprotectants resembles the response seen in frogs and earth-worms, although glucose of A. lusitanicus is accumulated in much lower concentra-tions. Thus, frozen A. lusitanicus only accumulated about 15 μmol glucose/g fresh weight (Slotsbo et al. 2012a) and other freeze tolerant slugs accumulate similar concentrations of glucose (Slotsbo et al. 2012a; Storey et al. 2007). In freeze tolerant
Arion lusitanicus.Photo: Stine Slotsbo.

20 PhD thesis by Stine Slotsbo
frogs and earthworms, glucose may reach concentrations of 200-400 mmol/l and, thereby, stabilize membranes and proteins during freeze-induced dehydration of cells (Crowe et al. 1987; Storey 1997; Holmstrup et al. 1999; Slotsbo et al. 2008). It may be disputed whether the same eff ects are of importance in slugs, where concentrations were appar-ently below 30 mmol/l (Storey et al. 2007; Slotsbo et al. 2012a). Again, freeze tolerant slugs from colder regions such as Siberia may synthesize much higher concentrations of cryoprotectants to survive temperatures below –20°C (Berman et al. 2011), but such studies have not been published yet.
Freeze tolerance is of-ten linked to anoxia tolerance, since freez-ing halts the circulation of blood or hemolymph and, thereby, supply of oxy-gen to tissues (Storey & Sto-rey 1996). Therefore, survival in the frozen state depends on ATP production via fermenta-tive pathways. The most common fer-mentation pathway in animal tissues is that of anaerobic glycolysis. A common end product of this pathway is lactate, but other end products, such as succinate and alanine, are known (Hochachka & Somero 2002). The accumulation of lactate and succinate in frozen A. lusitanicus shows that anaerobic metabolism occurs, as has also been reported for other terrestrial slugs when frozen (Storey et al. 2007).
Supercooling A. lusitanicus are generally poor supercoolers under humid conditions, but if un-frozen both juveniles and adults can survive down to –3°C. However, the risk of spontaneous freezing is high, especially for adults, due to a high supercooling point (–3°C). Smaller size classes of A. lusitanicus have lower supercooling points (Slotsbo et al. 2011b) (Table 3). The lower supercooling capacity with higher body mass of A. lusitanicus can be explained by physical phenomena (Bigg 1953) and is often observed in ectothermic animals, from arthro pods to reptiles (Lee & Cos-tanzo 1998). Larger animals have higher water volume, which increases the prob-ability of spontaneous freezing and also increases the probability that an eff ective ice nucleator is present in body fl uids.
Groups Fresh weight (mg) Supercooling point (°C)
Adult 8 000 – 17 000 –3.0 ± 0.2 (N=8)
Juvenile size 2 1800 ± 0.3 –3.8 ± 0.3 (N=12)
Juvenile size 1 30 – 54 –5.7 ± 0.2 (N=24)
Eggs 28 – 34 –6.2 ± 0.3 (N=23)
Table 3. Fresh weight and supercooling point of cold acclimated Arion lusitanicus. Values are mean ± SE. (Data from Slotsbo et al. 2011b and 2012a).

21PhD thesis by Stine Slotsbo
An experiment showed that small juveniles (16-26 mg) were able to stay su-percooled for 20 days at –3°C. During the experiment, the juveniles lost
a signifi cant amount of water (approx. 37 % after 20 days). Despite this water loss, the melting point did not change, indicating that small A. lusitanicus do not use cryoprotective dehydration as a cold tolerance strategy (Slotsbo et al. 2011b).
The supercooling point of A. lusitanicus is in congruency with other studies on terrestrial slugs from temperate regions, generally show-
ing that spontaneous freezing of slugs kept under dry conditions typically occurs at temperatures around –4°C (Cook 2004; Storey
et al. 2007; Slotsbo et al. 2012a). Measurements of super-cooling capacities under “dry conditions” provide basic
information of cold tolerance, but it may be discussed how relevant such measurements are for soil dwell-ing ectotherms, since ice crystals in the soil may inoculate the freezi ng of body fl uids (Holmstrup & Zachariassen 1996; Costanzo et al. 1997). If migra-tion to frost-free micro-sites is not possible, it is evi-
dent that survival of sub-zero temperatures must be based on freeze tolerance as strategy. Even though A.
lusitanicus cannot rely on supercooling as a cold hardiness strategy, juveniles have been observed being active at –3°C, which would transiently enable them to migrate to deeper and frost-free micro-habitats even at temperatures below 0°C.
Cold tolerance of eggsThe eggs of A. lusitanicus generally have poor cold tolerance, only surviving tem-peratures down to –2°C. The eggs do not survive by being freeze tolerant, but sur-vive cold periods by supercooling, and they are less prone to inoculative freezing than the slugs (Slotsbo et al. 2011b). The eggs are primarily laid in the late sum-mer and fall (Kozlowski 2000) and most of them hatch before winter . Therefore, it could be argued that winter survival of the population does not to any large de-gree depend on cold tolerance of eggs, but the ability of eggs to supercool could be important during cold spells in the late fall. The survival of overwintering eggs will, to a large degree, depend on the deposit place and how well the eggs are protected against low temperature.
Methodological considerations on cold tolerance experiments When performing cold tolerance experiments, it is important that the exposed organisms have been acclimated to low temperatures simulating as much as possible the gradual decrease in temperature in the autumn. This optimizes the chances that experimental animals have been adequately acclimatized to winter conditions before experiments and measurements are carried out. Thus, the slugs and eggs used in Paper II and III were cold acclimated prior to the experiments by keeping them for one week at 10°C, one week at 5°C, and lastly 4 weeks at 2°C. The cooling rate of cold tolerance experiments is also of great importance. Again, simulating natural conditions is important, and this was ensured in the freeze toler-ance assays where the temperature was lowered by just 1°C per day.
At present, only few species of terrestrial slugs have been found to be freeze tolerant, but employment of natural cooling and freezing rates may reveal that freeze tolerance is much more widespread in terrestrial slugs than previously anticipated.
Eggs of Arion lusitanicus. Photo: Vagn Nielsen.

22 PhD thesis by Stine Slotsbo
Drought tolerance
Terrestrial animals living in the soil or on the soil surface are occasionally exposed to drought and have adapted to this. One survival strategy, perhaps the most im-portant one, is to migrate to moist microhabitats during drought periods. When evading drought is not possible, two survival strategies have generally evolved: one is to reduce the water loss as much as possible and the other is to increase the tolerance of water loss (Hadley 1994).
Reduce water lossChanges in behaviour are one of the most used strategies to reduce water loss; this could be in terms of reducing the activity level or escaping from the drought in time or space. Escaping drought may, besides migration, also be achieved by changing to a more desiccation tolerant life stage. For example, some soil inverte-brates survive severe drought periods in the more desiccation tolerant egg stage (Holmstrup & Westh 1995). Some earthworms enter diapause if the soil water po-tential gets too low (Gerard, 1967; Nordström, 1975; Holmstrup, 2001), and land snails survive hot and dry periods by entering a stage of dormancy, called aestiva-tion. In aestivation, the snail is completely withdrawn into its shell so only a limited area of the mantle tissue remains exposed to the air. Further, the mouth of the shell is closed by thin fi lms of dried mucus, running either across the shell mouth or from the rim of the shell to some hard object to which the snail has attached itself (Howes & Wells 1934).
Other morphologically or physiologically based adaptations may reduce the wa-ter loss, such as reducing the water conductance of the skin or cuticle, hereby lowering the water loss by evaporation (Edney 1977). Evidence has been found for a “water barrier” in the mantel tissue of snails (Machin 1974). The water con-ductance of active specimens of the common garden snail Helix aspersa is nearly identical to that from a free water surface, whereas inactive snails are able to regulate evaporation markedly. The water conductance of the mantel tissue of inactive H. aspersa is 39 μg cm-2 h-1 mmHg-1 (Machin 1964, 1966). Accumula-tion of compatible osmolytes (like the cryoprotectants) will reduce the gradient in water vapour pressure across the skin and reduce evaporative water loss. Some collombolans are even able to accumulate compatible osmolytes to a degree that makes them hyperosmotic to their surroundings, enabling them to passively re-absorb water vapour from the atmosphere (Bayley & Holmstrup 1999).
Increase tolerance of water lossSome drought adapted animals have the ability to tolerate extensive loss of body water and have evolved physiological adaptations to overcome the challenges involved. Dehydration is harmful for non-adapted animals, as extensive water loss may cause severe cellular shrinkages, phase change in the membranes (Ha-zel 1995) and increases in solute concentrations which may potentially lead to changes in enzyme activity and denaturation of proteins. Accumulation of com-patible osmolytes can protect cells during dehydration and help stabilize mem-branes (Yancey 2005).
Drought tolerance of slugsTerrestrial molluscs are generally susceptible to desiccation, and several studies have found habitat selection to be well correlated with the ava ilability of water (Getz 1959; Prior 1985; Carne-Cavagnaro et al. 2006). The moist permeable in-tegument of terrestrial molluscs requires adaptations to avoid or cope with exces-
Eggs of Arion lusitanicus in a grass fi eld. Photo: Stine Slotsbo.

23PhD thesis by Stine Slotsbo
sive dehydration. The most obvious water regulating behaviour of slugs and snails is their preference for moist habitats ( Prior et al. 1983; Prior 1985). In contrast to snails, slugs lack the physical protection of a shell and must fi nd other ways to reduce water loss
Water loss resistance of slugsIn reducing the water loss, the water conductance of the skin is an important factor. The water conductance of slugs has been studied by Thomson (2006), who found the water conductance of the slug species Philomycus carolinianus, D. leave and Lehmannia valentiana to be 210, 348 and 378 μg cm-2 h-1 mmHg-1, respectively. This shows that there are diff erences between species of slugs, but that slugs in general are very permeable and, not surprisingly based on their water conduct-ance, confi ned to hygric habitats. In comparison, insects of mesic habitats typi-cally have water conductance values ranging from 20 to 60 μg cm-2 h-1 mmHg-1 (Edney 1977); almost one order of magnitude lower. The ability to survive drought cannot be explained by water conductance alone, but is also infl uenced by body size and magnitude of water reserves. Large animals lose a smaller proportion of their original weight per time unit than small ones with the same water conduct-ance. Because of surface-to-volume relationships, adult slugs would be able to withstand drought for longer periods than juveniles of the same species.
There are diff erent behavioural adaptations to reduce the water loss. Juvenile and adult slugs have a tendency to cluster if they are exposed to desiccation stress. This behaviour is called huddling behaviour (Prior et al. 1983) in which they pack closely together, thereby reducing the surface area and generating a high humidity micro-environment around the huddled slugs. Huddling behaviour (also applying to clusters of eggs) reduces the vapour pressure defi cit and, thereby, evaporation (Prior 1985).
Tolerance of water loss in slugsA high degree of dehydration tolerance seems to be widespread among gastro-pods. The aquatic snail Australorbis glabratus tolerates losing up to 70 % of its initial water content (Von Brand et al. 1957), and in the intertidal snail, Melampus Biden-tatus, even 80 % loss of body water is tolerated by some individuals (Price 1980). Among terrestrial slugs, P. carolinianus, L. valentiana and D. leave survive losses up to 89 %, 64 % and 68 % of their initial water content, respectively (Thompson et al. 2006). An increased osmolality as a consequence of water loss has been observed in both slug and snail species (Prior 1985), but dehydrated snails and slugs are able to recover rapidly from dehydration by absorbing water through their integument; this process is termed c ontact re-hydration (Dainton 1954; Prior 1984, 1985).
Drought tolerance of slug eggsWater balance is of importance for population dynamics of terrestrial slugs (Carne-Cavagnaro et al. 2006; Willis et al. 2008). As stated, their eggs are often laid at pro-tected and humid sites, e.g. in humid soil, in crevices or beneath debris, microhabi-tats that help ensure a high humidity. However, since the development of eggs can last several months in the fi eld, the eggs may from time to time be exposed to desiccation conditions which may delay development or cause mortality. The fact that eggs are often laid in clusters and that one slug lays several clusters of eggs will increase the chances that at least some eggs survive drought conditions and hatch when conditions are again favourable (Carmichael & Rivers 1932; Carrick 1942; Bayne 1969). Eggs of many soil-living invertebrates are often more drought resistant than post-embryonic stages, but our knowledge of drought tolerance of eggs of terrestrial slugs is limited.

24 PhD thesis by Stine Slotsbo
Carmichael and Rivers (1932) have studied the eggs of the terrestrial slug Limax fl avus and found that some of the eggs were able to survive as much as 85 % weight loss. In early literature (Binney 1878), there are reports of even greater drought tolerance of slug eggs, but those results have been disproven by more recent authors (Carmichael & Rivers 1932; Carrick 1942; Bayne 1969).
Drought tolerance of Arion lusitanicus
Like other slugs, A. lusitanicus is with its permeable and humid skin highly suscepti-ble to desiccation. Both eggs and juveniles begin losing water when the ambient humidity falls below 99.8 and 99.5 %, respectively (Slotsbo et al. 2011a).
Drought tolerance of juvenilesThe water conductance of small juvenile A. lusitanicus has been estimated to be 242 μg cm-2 h-1 mmHg-1 (Slotsbo et al. 2011a), which shows that the skin of A. lusi-tanicus, like the skin of other slugs (Thompson et al. 2006), only provides little resist-ance against water loss. The rate of water loss increases exponentially with increas-ing desiccation stress (Fig. 5). The hemolymph osmolality of fully hydrated juveniles is 230 mOsmol (Slotsbo et al. 2011a), which corresponds to a relative humidity of 99.5 %, below which they will lose water. The osmolality during drought at tempera-tures above zero has not been measured, but at sub-zero temperatures osmolality of unfrozen juveniles did not increase despite considerable water loss (Slotsbo et al. 2011b). It seems, therefore, that A. lusitanicus relies solely on behavioural responses (and not physiological) to reduce water loss in terms of homing to humid resting places, huddling behaviour, and a nocturnal activity pattern. As a consequence, A. lusitanicus will, inevitably, often experience water loss in nature. Therefore, to a cer-tain degree, its drought tolerance relies on its ability to tolerate dehydration.
Indeed, juveniles tolerate considerable water loss. Exposure for 7 days to a range of relative humidities showed that 50 % mortality occured when juveniles lose about 72 % of their initial water content (Fig. 6). Moreover, no mortality occurred if water loss did not exceed 56 %, but no juveniles survived water loss above 80 % (Slotsbo et al. 2011a). Due to lower surface-to-volume ratio and a larger water reserve, it is likely that adults would be able to withstand desiccation for longer periods and, presumably, would survive drought better than the juveniles. During longer drought periods, slugs can seek deeper and more humid micro-habitats
100 80 60 40 20 0
Relative humidity (%)
Wat
er lo
ss (
% o
f WC
initi
al/1
00 m
in)
0
10
20
30
40Figure 5. The water loss rate of ju-venile Arion lusitanicus exposed to seven diff erent relative humidi-ties in the range from 7.5–98 %. The water loss rate is expressed as percent water loss of the initial water content per 100 min. Values are mean ± SE. (From Slotsbo et al. 2011a).

25PhD thesis by Stine Slotsbo
in the soil; it therefore seems reasonable to conclude that summer drought under Danish conditions is unlikely to cause any substantial mortality in populations of A. lusitanicus. This being said, the activity of slugs is undoubtedly reduced during droughts (Grimm et al. 2000; Sternberg 2000; Kozlowski 2001), reducing forag-ing of the slugs and therefore also growth, development and reproduction. Field observations support this notion; according to Grimm (2001), a very dry spring and summer will delay the egg-laying period in the given year.
Drought tolerance of eggsThe outer layer of A. lusitanicus eggs is fairly thick and contains crystals of calcium carbonate (South 1992) and below this calcareous coating are inner shell mem-branes (Bayne 1966, 1968). Nevertheless, the shell and membranes of the eggs are permeable and provide little resistance against water loss. The osmolality of the fully hydrated eggs is very low (130 mOsmol), suggesting that water loss will begin at relative humidities below 99.8 % (Slotsbo et al. 2011a).
The eggs can tolerate losing a considerable amount of water. Thus, hatchability is largely unaff ected if water loss does not exceed 60 % (Fig. 7). Even a loss of 81 % of the initial water content caused only a 50 % reduction in hatching success, and some eggs were able to survive a loss of 86 % of their initial water content.
Clustering of eggs has a marked eff ect on the desiccation rates of eggs, especially eggs in the middle of a cluster exper ience reduced water loss (Bayne 1969; Willis et al. 2008).
-40 0-20 20 40 60 80 100
Water loss (% of WCinitial)
Sur
viva
l (%
)0
20
40
60
100
80
Figure 6. The relationship be-tween survival and water loss of juveniles of Arion lusitanicus ex-posed to diff erent relative humidi-ties for 7 days. Data from an ex-periment, where water loss and survival were measured on sepa-rate individuals, are shown as cir-cles. Data from an experiment, where water loss and survival were measured on the same in-dividuals, are shown as squares. Sigmoidal dose-response curves were fi tted to the survival data. Values are mean ± SE survival. (From Slotsbo et al. 2011a)
0 20 40 60 80 100
Water loss (% of WCinitial)
Sur
viva
l (%
)
0
20
40
60
100
80
Figure 7. The relationship be-tween survival and water loss of Arion lusitanicus eggs exposed to diff erent relative humidities for 7 days. A sigmoidal dose-re-sponse curve was fi tted to the sur-vival data. Values are mean ± SE. (From Slotsbo et al. 2011a)

26 PhD thesis by Stine Slotsbo
Desiccation eff ects on the development time of eggs
The development time of A. lusitanicus eggs increases with increasing desiccation (Fig. 8). The same has been found in the pond snail Lymnaea humilis, where partial drying of egg masses prolonged the hatching time (McCrawn 1961). For eggs of A. lusitanicus, a 7 day exposure to 95 % relative humid-ity resulted in a 7 day delay in hatching compared to controls, meaning that the embryonic development was probably rap-idly arrested due to water loss. In a study of Bayne (1969), no increase in the development time of eggs of the terrestrial slug D. reticulatum was observed as a consequence of drought ex-posure. However, eggs in that experiment were dehydrated for only 14 hours before they were allowed to rehydrate, which did not allow good assessment of delays in development time.
Methodological considerations on drought experiments It is extremely diffi cult to create and control a precise drought stress levels in soil under laboratory conditions due to the of-ten very steep retention curves of various soil types. For this reason, I used a closed system and controlled relative hu-midities using aqueous solutions of NaCl or glycerol in the drought experiments performed in Paper IV. Silica gel was used for (almost) dry air. Juveniles or eggs were then placed in small plastic tubes, placed in the centre of sealed plas-
tic beakers. The air in this small closed system quickly equilibrated with water vapour from either the solutions or the silica gel, creating a range of precisely controlled relative humidities (Holmstrup & Westh 1995). The relative humidity can be aff ected by temperature because the water vapour pressure is highly de-pendent on temperature, however, if temperature fl uctuations are prevented this system has proven to be easily controllable. A similar system has been used for drought studies of earthworm cocoons and collembolans (Holmstrup & Westh 1995; Holmstrup 1997).
In the desiccation tolerance experiment in Paper IV, I use relative humidities be-tween 100 and 90%, although it may seem a very mild desiccation level for an animal living on the soil surface. But as shown, these relative humidities cause dramatic water loss in slugs and their eggs.
Relative humidity (%)
Dev
elop
men
t tim
e (d
ays)
32
34
36
38
40
42
44
100 99 98 97 96 9495
Figure 8. The development time of Arion lusitanicus eggs exposed for 7 days to a range of relative humidities between 95–100 %. Values are mean ± SE. (From Slotsbo et al. 2011a).
Arion lusitanicus. Photo: Stine Slotsbo.

27PhD thesis by Stine Slotsbo
Conclusion and perspectives
This PhD project contributes detailed knowledge of the ecophysiology and life his-tory of A. lusitanicus. The growth, egg developmental time and egg hatchability of A. lusitanicus are infl uenced by temperature. Rise in temperature will, below 25°C, result in an increase in growth rate. A. lusitanicus has a high degree of plasticity in its thermal biology and can quickly adjust its growth with changes in temperature.
A. lusitanicus is a freeze-tolerant species both in the juvenile and adult stages, but only tolerates freezing at relatively high subzero temperatures (–2°C). The eggs also only survive relatively high subzero temperatures, but probably by supercool-ing. Juveniles and adults may survive in a supercooled state, but are generally poor supercoolers. Therefore, the winter survival of A. lusitanicus in very cold re-gions to a high degree depends on migration to habitats protected from low win-ter temperatures. A. lusitanicus, like other slugs, has little resistance to evaporative water loss, but on the other hand tolerates substantial water loss. The eggs are slightly more tolerant to water loss than juveniles. The development time of the eggs was found to increase with increasing desiccation because embryonic de-velopment is arrested due to dehydration. Despite tolerance of substantial water loss, A. lusitanicus depends on humid habitats.
In NW Europe, the large arionids comprise A. rufus (Linnaeus, 1758), A. ater (Lin-naeus, 1758), A. lusitanicus (Mabille, 1868) and A. fl agellus (Collinge, 1893). A. lusitanicus was not discovered in Denmark until recently (1991) (Von Proschwitz and Winge 1994), whereas A. rufus was introduced to Denmark from Germany about a century ago and A. ater is native to Denmark (Engelke et al. 2011). A. lusitanicus is the only Arion species regarded as a serious pest in Denmark. But, as mentioned, the three species are easily confused (Cameron et al. 1983; Noble 1992), and identifi cation using external morphology is almost impossible. Person-ally, I believe that some of our “A. lusitanicus problems” could in reality be “A. rufus problems”. Species identifi cation of fi eld collected specimens during my studies showed that this could be the case, but further work is needed to reveal whether this is true. In Poland, A. rufus is, like A. lusitanicus, known to cause serious damage to crops, especially in gardens (Kozlowski & Kozlowska 2004).
The reason for the mass occurrences and consequent pest status of A. lusitanicus remains unrevealed, but anecdotal reports fl ourish in public media stating that the success of the species is related to its superior resistance to natural stresses such as drought and low winter temperatures, and that it in this way may outcompete other closely related Arion species. Paper III suggests that there are only subtle diff erences in cold tolerance of the three Arion species, if any at all. This PhD pro-ject was not able to reveal any ecophysiological diff erences between the three species that could give rise to diff erences in pest status. In order to search for such diff erences, a thorough investigation of the life history traits of A. ater and A. rufus would be a natural next step. A survey from Switzerland has found that adult A. rufus in agricultural landscapes have reduced survival rates compared to A. lusi-tanicus (Ryser et al. 2011).
This PhD project provides a number of basic parameters which may contribute to the understanding of the population dynamics of A. lusitanicus. There is, however, a need for further studies of fi eld populations, which together with data from this PhD project could form the basis of a population growth model to provide a more accurate forecasting of slug abundance. Such a forecast model could be a useful tool in the pest control of A. lusitanicus.

28 PhD thesis by Stine Slotsbo
The growth model developed in Paper I enables the prediction of individual slug growth under natural temperature scenarios. But to be able to create a population growth model forecasting population dynamics, some challenges still need to be overcome.
This PhD project has revealed the cold and drought tolerance limits of A. lusitanicus, but spatial variations of microclimatic conditions must also be taken into account together with behavioural responses of A. lusitanicus. In very dry periods, some areas, e.g. borders of water bodies or fl ood plains, can still be very humid and can create refuges from where the species can re-colonize drought-impacted areas when conditions become favourable again. In very cold periods, opportunities for protected overwintering sites are of importance. To be able to predict the eff ect of cold or dry periods on the population dynamics, we need to have more knowl-edge on the behaviour of A. lusitanicus during overwintering periods, and this must be generated by further fi eld studies.
Besides the eff ects of climatic conditions, biotic factors such as predation and competition with other slug species for food and space may also have eff ects on the population dynamics of A. lusitanicus. Lastly, it has to be taken into account that A. lusitanicus probably still is unintentionally dispersed between regions due to human activity.
Arion lusitanicus. Photo: Stine Slotsbo.

29PhD thesis by Stine Slotsbo
References
Abeloos, M. 1942. The stages of growth of the red slug ( Arion rufus L.). – Comptes Rendus Hebdomadaires des Seances de l Academie des Sciences 215:38-39.
Abeloos, M. 1943. Form of the growth of species of the Limax genus. – Comptes Rendus Hebdomadaires des Seances de l Academie des Sciences 217:159-161.
Anderson, R. 2005. An annotated list of non-marine Mollusca of Britain and Ireland. – Jour-nal of Conchology 38:607-637.
Ansart, A., Vernon, P. and Daguzan, J. 2001. Freezing tolerance versus freezing susceptibil-ity in the land snail Helix aspersa (Gastropoda: Helicidae). – Cryoletters 22:183-190.
Ansart, A. and Vernon, P. 2003. Cold hardiness in molluscs, Acta Oecologica-International. – Journal of Ecology 24:95-102.
Backeljau, T., Jordaens, K., Parejo, C. and Rodríguez, T. 1996. Protein electrophoresis in Arionid taxonomy. – British Crop Protection Council symposium proceedings 66:21-28.
Bank, R.A., Falkner, G. and Von Proschiwtz, T. 2007. A revised checklist of the non-marine Mollusca of Britain and Ireland. – Heldia 5:41-72.
Barker, G.M. 2001. The Biology of Terrestrial Molluscs. – CABI Publishing, New York.
Bayley, M. and Holmstrup, M. 1999. Water vapor absorption in arthropods by accumulation of myoinositol and glucose. – Science 285:1909-1911.
Bayne, C.J. 1966. Observations on composition of layers of egg of Agrio Limax reticulatus grey fi eld slug (Pulmonata stylomatophora). – Comparative Biochemistry and Physiology 19:317-337.
Bayne, C.J. 1968. A study of desiccation of egg capsules of 8 gastropod species. – Journal of Zoology 155:401-411.
Bayne, C.J. 1969. Survival of the embryos of the grey fi eld slug Agrio Limax reticulatus, fol-lowing desiccation of the egg. – Malacologia 9:391-401.
Berman, D.I., Meshcheryakova, E.N. and Leirikh, A.N. 2011. Cold hardiness, adaptive strategies and invasion of slugs of the Genus Deroceras (Gastropoda, Pulmonata) in Northeastern Asia. – Zoologichesky Zhurnal 90:387-401.
Bigg, E.K. 1953. The supercooling of water. Proceedings of the Physical Society of London Section B, 66: 688-694.
Binney, W.G. 1878. The terrestrial air-breathing mollusks, 1 edn. Bulletin of the museum of comparative zoology at Harvard college, Cambridge University Press.
Briner, T. and Frank, T. 1998. Egg laying activity of the slug Arion lusitanicus Mabille in Swit-zerland. – Journal of Conchology 36:9-15.
Cameron, R.A.D., Eversham, B. and Jackson, N. 1983. A fi eld key to the slugs of the British Isles. – Field Studies 5:807-824.
Carmichael, E.B. and Rivers, T.D. 1932. The eff ects of dehydration upon the hatchabilty of Limax fl avus eggs. – Ecology 13:375-380.
Carne-Cavagnaro, V.L., Keller, M.A. and Baker, G.H. 2006. Soil moisture and soil type infl u-ence the breeding behavior of the pest snail Cernuella virgata (da Costa). – Applied Soil Ecology 33:235-242.
Carrick, R. 1942. The grey fi eld slug Agrio Limax agrestis L, and its environment. – Annals of Applied Biology 29:43-55.
Castillejo, J. 1997. The slugs of the family Arionidae Grey, 1840 from Iberian Peninsula and Balearic Islands. Morphology and distribution (Gastropoda, Pulmonata, Terrestria nuda).– Revista Real Academia Galega de Ciencias 16:51-118.

30 PhD thesis by Stine Slotsbo
Choi, Y.H, Bohan, D.A., Powers, S.J., Wiltshire, C.W., Glen, D.M. and Semenov, M.A. 2004. Modelling Deroceras reticulatum (Gastropoda) population dynamics based on daily tem-perature and rainfall. – Agriculture, Ecosystems and Enviroment 103:519-525.
Clemente, N.L., López, A.N., Monterubbianesi, M.G., Cazzaniga, N.J. and Manetti, P.L. 2008. Biological studies and phenology of slug Deroceras reticulatum (Müller, 1774) (Pulmonata: Stylommatophora). – Invertebrate Reproduction and development 52:23-30.
Clemente, N.L., Faberi, A.J., Salvio, C. and López, A.N. 2010. Biology and individual growth of Milax gagates (Draparnaud, 1801) (Pulmonata: Stylommatophora). – Invertebrate Re-production and development 54:163-168.
Cook, R.T. 2004. The tolerance of the fi eld slug Deroceras reliculatum to freezing tempera-tures. – Cryoletters 25:187-194.
Costanzo, J.P., Moore, J.B., Lee, R.E., Kaufman, P.E. and Wyman, J.A. 1997. Infl uence of soil hydric parameters on the winter cold hardiness of a burrowing beetle, Leptinotarsa de-cemlineata (Say). – Journal of Comparative Physiology B 167:169-176.
Crowe, J.H., Crowe, L.M. Carpenter, J.F. and Wistrom, C.A. 1987. Stabilization of dry phospho-lipid bilayers and proteins by sugars. – Biochemical Journal 242:1-10.
Dainton, B.H. 1954. The activity of slugs.1. The induction of activity by changing tempera-tures. – Journal of Experimental Biology 31:165-187.
Davies, S.M. 1987. Arion-Flagellus Collinge and Arion-Lusitanicus Mabille in the British-Isles – A Morphological, Biological and Taxonomic Investigation. – Journal of Conchology 32:339-354.
Duman, J.G., Horwarth, K.L., Tomchaney, A. and Patterson, J.L. 1982. Antifreeze agents of terrestrial arthropods. – Comparative Biochemistry and Physiology A 73:545-555.
Edney, E.B. 1977. Water balance in Land Arthropods, 1 edn. – Springer-Verlag, Berlin.
Engelke, S., Kompf, J., Jordaens, K., Tomiuk, J. and Parker, E.D. 2011. The genetic dynamics of the rapid and recent colonization of Denmark by Arion lusitanicus (Mollusca, Pulmonata, Arionidae). – Genetica 139:709-721.
Frank, T. 1998. Slug damage and numbers of the slug pests, Arion lusitanicus and Deroceras reticulatum, in oilseed rape grown beside sown wildfl ower strips. – Agriculture, Ecosystems and Environment 67:67-78.
Gerard, B. 1967. Factors aff ecting earthworms in pastures. – Journal of Animal Ecology 36:235-252.
Getz, L.L. 1959. Notes on the ecology of slugs: Arion circumscriptus, Deroceras reticukatum, and D. laeve. – American Midland Naturalist 61:485-498.
Grimm, B. 2001. Life cycle and population density of the pest slug Arion lusitanicus Mabille (Mollusca:Pulmonata) on grassland. – Malacologia 43:25-32.
Grimm, B. and Paill, W. 2001. Spatial distribution and home-range of the pest slug Arion lusi-tanicus (Mollusca Pulmonata). – Acta Oecologica-International Journal of Ecology 22:219-227.
Grimm, B., Paill, W. and Kaiser, H. 2000. Daily activity of the pest slug Arion lusitanicus Mabille. – Journal of Molluscan Studies 66:125-128.
Hadley, N. 1994. Water Relations of Terrestrial Arthropods. – Academic Press, San Diego.
Hagnell, J., Von Proschiwtz, T. and Schander, C. 2006. Self-fertilising observed in the inva-sive Iberian slug Arion lusitanicus, Mabille 1868. – Journal of Conchology 39:107.
Hammond, R.B. 1985. Slugs as a new pest of soybeans. – Journal of the Kansas Entomo-logical Society 58:364-366.
Hatteland, B.A. 2010. Predation by carabid beetles (Coleoptera, Carabidae) on the inva-sive Iberian slug Arion lusitanicus. PhD Thesis. – University of Bergen, Norway.
Hazel, J.R. 1995. Thermal adaptation in biological membranes: Is homeoviscous adapta-tion the explanation. – Annual Review of Physiology 57:19-42.

31PhD thesis by Stine Slotsbo
Hochachka, P.W. and Somero, G.N. 2002. Biochemical adaptation: Mechanism and pro-cess in physiological evolution. – Oxford University Press, New York.
Holmstrup, M. 1997. Drought tolerance in Folsomia candida Willem (Collemhola) after exposure to sublethal concentrations of three soil-polluting chemicals. – Pedobiologia 41:361-368.
Holmstrup, M. 2001. Sensitivity of life history parameters in the earthworm Aporrectodea cali-ganosa to small changes in water potential. – Soil Biology and Biochemistry 33:1217-1223.
Holmstrup, M., Bayley, M. and Ramløv, H. 2002. Supercool or dehydrate? An experimental analysis of overwintering strategies in small permeable arctic invertebrates. – Proceedings of the National Academy of Sciences of the United States of America 99:5716-5720.
Holmstrup, M., Overgaard, J. and Bayley, M. 2007. Small Dendrobaena earthworms survive freezing better than large worms. – Cryobiology 54:298-300.
Holmstrup, M. and Westh, P. 1995. Eff ects of dehydration on water relations and survival of lumbricid earthworm egg capsules. – Journal of Comparative Physiology B 165:377-383.
Holmstrup, M. and Zachariassen, K.E. 1996. Physiology of cold hardiness in earthworms. – Comparative Biochemistry and Physiology A 115:91-101.
Holmstrup, M., Costanzo, J.P. and Lee, R.E. 1999. Cryoprotective and osmotic responses to cold acclimation and freezing in freeze-tolerant and freeze-intolerant earthworms. – Jour-nal of Comparative Physiology B 169:207-214.
Howes, N.H. and Wells, G.P. 1934. The Water relations of snails and slugs. I. Weight rhythms in Helix pomatia L. – The Journal of Experimental Biology 11:327-343.
Hulme, P.E. 2009. Trade, transport and trouble: managing invasive species pathways in an era of globalization. – Journal of Applied Ecology 46:10-18.
Hommay, G., Kienlen, J.C., Gertz, C. and Hill, A. 2001. Growth and reproduction of the slug Limax valentianus Férussac in experimental conditions. – Journal of Molluscan Studies 67:191-207.
Hunter, PJ. 1967. Eff ect of cultivations on slugs of arable land. – Plant Pathology 16:153-156.
Jordaens, K., Van Dongen, S., Van Riel, P., Geenen, S., Verhagen, R. and Backeljau, T. 2002. Multivariate morphometrics of soft body parts in terrestrial slugs: comparison between two datasets, error assessment and taxonomic implications. – Biological Journal of the Linnean Society 75:533-542.
Jordaens, K., Van Riel, P., Geenen, S., Verhagen, R. and Backeljau, T. 2001. Food-induced body pigmentation questions the taxonomic value of colour in the self-fertilizing slug Cari-narion spp. – Journal of Molluscan Studies 67:161-167.
Kerney, M. 1999. Atlas of the Land and Freshwater Molluscs of Britain and Ireland. – Harley Books, Colchester.
Kozlowski, J. 2000. Reproduction of Arion lusitanicus Mabille, 1868 (Gastropoda: Pulmo-nata: Arionidae) introduced in Poland. – Folia Malacologica 8:87-94.
Kozlowski, J. 2001. Daily activity of Arion lusitanicus Mabille, 1868 (Gastropoda: Pulmonata: Arionidae). – Journal of Plant Protection Research 41:279-286.
Kozlowski, J. 2007. The Distribution, biology, population dynamics and harmfulness of Arion lusitanicus Mabille, 1868 (Gastropoda: Pulmonata: Arionidae) in Poland. – Journal of Plant Protection Research 47:219-230.
Kozlowski, J. and Kozlowska, M. 2004. Food preferences of Deroceras reticulatum, Arion lusitanicus and Arion rufus for various medicinal herbs and oilseed rape. – ournal of Plant Protection Research 44:239-249.
Kozlowski, J. and Kozkowski, R.J. 2000. Periods of occurrence and fecundity of Arion lusi-tanicus Mab. (Gastropoda:Pulmonata) in crop plant habitats in Poland. – Journal of Plant Protection Research 40:260-266.
Kozlowski, J. and Sionek, R. 2000. The rate of egg laying and hatching of the slug Arion lusi-tanicus Mabille, a pest of arable crops. – Journal of Plant Protection Research 40:160-167.

32 PhD thesis by Stine Slotsbo
Lee, R.E. and Costanzo, J.P. 1998. Biological ice nucleation and ice distribution in cold-hardy ectothermic animals. – Annual Review of Physiology 60:55-72.
Lusis, O. 1961. Postembryonic changes in the reproductive system of the slug Arion Ater Ru-fus L. – Proceedings of the Zoological Society of London 137:433-468.
Machin, J. 1964. The evaporation of water from Helix aspersa. I. The nature of the evapo-rating surface. – The Journal of Experimental Biology 41:759-769.
Machin, J. 1966. The evaporation of water from Helix aspersa. IV. Loss from the mantle of the inactive snail. – The Journal of Experimental Biology 45:269-278.
Machin, J. 1974. Gradients across snail epidermis: Evidence for a Water Barrier. – Science 183:759-760
McCrawn, B.M. 1961. Life history and growth of the snail, Lymnaea humilis Say. – Transac-tions of the American Microscopical Society 80:16-27.
Mellanby, K. 1961. Slugs at Low Temperatures. – Nature 189:944.
Menaker, M. 1971. Rhythms, Reproduction, and Photoreception. – Biology of Reproduction 4:295-308.
Meyerson, L.A. and Mooney, H.A. 2007. Invasive alien species in an era of globalization.– Frontiers in Ecology and the Environment 5:199-208.
Noble, L.R. 1992. Diff erentiation of large arionid slugs (Mollusca, Pulmonata) using ligula morphology. – Zoologica Scripta 21:255-263.
Noble, L.R. and Jones, C.S. 1996. A molecular and ecological investigation of the large arionid slugs of North-West Europe: the potential for a new pest species. In The Ecology of Agricultural Pest: Biochemical approaches. – Systematics association special volume series (Eds. Symondson, W.O.C. and Liddell, J.E.) Chapman & Hall, London.
Nordström, S. 1975. Seasonal activity of lumbricids in southern Sweden. – Oikos 26:307-315.
Norris, R.F., Caswell-Chen, E.P. and Kogan, M. 2003. Concepts in integrated pest manage-ment. Prentice Hall. – Upper Saddle River, New Jersey.
Overgaard, J., Tollarova, M., Hedlund, K., Petersen, S.O. and Holmstrup, M. 2009. Seasonal changes in lipid composition and glycogen storage associated with freeze-tolerance of the earthworm, Dendrobaena octaedra. – Journal of Comparative Physiology, B 179:569-577.
Pinceel, J., Jordaens, K. and Backeljau, T. 2005. Extreme mtDNA divergences in a terrestrial slug (Gastropoda, Pulmonata, Arionidae): accelerated evolution, allopatric divergence and secondary contact. – Journal of Evolutionary Biology 18:1264-1280.
Price, C.H. 1980. Water relations and physiological ecology of the salt marsh snail, Melampus bidentatus Say. – Journal of Experimental Marine Biology and Ecology 45:51-67.
Prior, D.J. 1984. Analysis of contact-rehydration in terrestrial gastropods - osmotic control of drinking behavior.– Journal of Experimental Biology 111:63-73.
Prior, D.J. 1985. Water-regulatory behavior in terrestrial gastropods. – Biological Reviews of the Cambridge Philosophical Society 60:403-424.
Prior, D.J., Hume, M., Varga, D. and Hess, S.D. 1983. Physiological and behavioral aspects of water balance and respiratory function in the terrestrial slug, Limax maximus. – Journal of Experimental Biology 104:111-127.
Quick, H.E. 1952. Rediscovery of Arion lusitanicus Mabille in Britain. – Proceedings of the malacological society 29:93-101.
Quinteiro, J., Rodriguez-Castro, J., Castillejo, J., Iglesias-Pineiro, J. and Rey-Mendez, M. 2005. Phylogeny of slug species of the genus Arion: evidence of Iberian endemics and of the existence of relict species in Pyrenean refuges. – Journal of Zoological Systematics and Evolutionary Research 43:139-148.
Ramløv, H. 2000. Aspects of natural cold tolerance in ectothermic animals. – Human Re-production 15:26-46.
Riddle, W.A. 1981. Cold hardiness in the woodland snail, Anguispira alternata (Say) (Endo-dontidae). – Journal of Thermal Biology 6:117-120.

33PhD thesis by Stine Slotsbo
Riddle, W.A. and Miller, V.J. 1988. Cold hardiness in several species of land snails. – Journal of Thermal Biology 13:163-167.
Roberts, S., Grout, B.W.W. and Morris, G.J. 1987. Consecutive observations of a frozen cell sample by cryogenic light-microscopy and cryogenic scanning electron-microscopy. – Cryo-Letters 8:122-129.
Rollo, C.D. 1983. Consequences of competition on the reproduction and mortality of 3 spe-cies of terrestrial slugs. – Researches on Population Ecology 25:20-43.
Rundgren, S. 1975. Vertical distribution of lumbricids in southern Sweden. – Oikos 26:299-306.
Ryser, S., Rindlisbacher, N., Grüebler, M.U. and Knop, E. 2011. Diff erential survival rates in a declining and an invasive farmland gastropod species. – Agriculture, Ecosystems and Envi-ronment 144:302-307.
Salt, R.W. 1963. Delayed inoculative freezing of insects.– The Canadian Entomologist 95:1190-1202.
Shirley, M.D.F., Rushton, S.P., Young, A.G. and Port, G.R. 2001. Simulating the long-term dy-namics of slug populations: a process-based modelling approach for pest control. – Journal of Applied Ecology 38:401-411.
Sinclair, B.J. 1999. Insect cold tolerance: How many kinds of frozen? – European Journal of Entomology 96:157-164.
Slotsbo, S., Maraldo, K., Malmendal, A., Nielsen, N.C. and Holmstrup, M., 2008. Freeze toler-ance and accumulation of cryoprotectants in the enchytraeid Enchytraeus albidus (Oli-gochaeta) from Greenland and Europe. – Cryobiology 57:286-291.
Slotsbo, S., Fisker, K., Hansen, L. and Holmstrup, M. 2011a. Drought tolerance in eggs and ju-veniles of the Iberian slug, Arion lusitanicus. – Journal of Comparative Physiology B 175:1-9.
Slotsbo, S., Hansen, L.M. and Holmstrup, M. 2011b. Low temperature survival in diff erent life stages of the Iberian slug, Arion lusitanicus. – Cryobiology 62:68-73.
Slotsbo, S., Hansen, L.M., Jordaens, K., Backeljau, T. Malmendal, A., Nielsen, N.C. and Holm-strup, M. 2012a. Cold tolerance and freeze-induced glucose accumulation in three terres-trial slugs. – Comparative Biochemistry and Physiology A 161:443-449.
Slotsbo, S., Damgaard, C., Hansen, L.M. and Holmstrup, M. 2012b. The infl uence of tem-perature on growth and egg development time of Arion lusitanicus (submitted to Annals of Applied Biology).
Sokolove, P.G. and Mccrone, E.J. 1978. Reproductive maturation in slug, Limax maximus, and eff ects of artifi cial photoperiod. – Journal of Comparative Physiology 125:317-325.
Sømme, L. 1982. Supercooling and winter survival in terrestrial arthropods. – Comparative Biochemistry and Physiology A 73:519-543.
South, A. 1982. A comparison of the life cycles of Deroceras reticulatum (Muller) and Arion intermedius Normand (Pulmonata, Stylommatophora) at diff erent temperatures under laboratory conditions. – Journal of Molluscan Studies 48:233-244.
South, A. 1992. Terrestrial slugs. – Biology, Ecology and Control., 1 edn. Chapman & Hall, London.
Sternberg, M. 2000. Terrestrial gastropods and experimental climate change: A fi eld study in a calcareous grassland. – Ecological Research 15:73-81.
Storey, K.B. and Storey, J.M. 1988. Freeze tolerance in animals. – Phsiological Reviews 68:27-84.
Storey, K.B. and Storey, J.M. 1992. Biochemical adaptions for winter survival in insects. In: Advances in Low-Temperature Biology. (Eds. Steponkus, P.L.) – JAI Press, London, pp. 101-140.
Storey, K.B. and Storey, J.M. 1996. Natural freezing survival in animals. – Annual Review of Ecology and Systematics, 27: 365-386.
Storey, K. B. 1997. Organic solutes in freezing tolerance. Comparative Biochemistry and Physiology A 117:319-326.
Storey, K.B., Storey, J.M. and Churchill, T.A. 2007. Freezing and anoxia tolerance of slugs: a metabolic perspective. – Journal of Comparative Physiology B 177:833-840.

34 PhD thesis by Stine Slotsbo
Thompson, J.M., Appel, A.G., Sibley, J.L., Keever, G.J. and Foshee, W.G. 2006. Comparative water relations of three sympatric terrestrial slugs: (Stylommatophora: Agriolimacidae, Limacidae, and Philomycidae. – Journal of Alabama Academy of Sciences 77:181-192.
Udaka, H. and Numata, H. 2008. Short-day and low-temperature conditions promote reproductive maturation in the terrestrial slug, Lehmannia valentiana. – Comparative Bio-chemistry and Physiology A 150:80-83.
Von Brand, T., McMahon, P. and Nolan, M.O. 1957. Observations on starvation and desicca-tion of snail Australorbis glabratus. – Biological Bulletin 113:89-102.
Von Proschiwtz, T. and Winge, K. 1994. Iberia skogsnegl - en art pä spredning i Norge (Arion lusitanicus Mabille - en anthropochorous slug spreading in Norway). – Fauna 47:195-300.
Von Proschwitz, T. 1989. Arion lusitanicus Mabille - en för Sverige ny snigelart. – Göteborgs Naturhistoriska Museum Årstryk, 43-53.
Von Proschwitz, T. 1992. Spanska skogsnigeln - Arion lusitanicus Mabille - en art i snabb spridning med människan i Sverige. – Göteborgs Naturhistoriska Museum Årstryk, 35-42.
Weast, R.C. 1989. Handbook of Chemistry and Physics. – CRC Press, Cleveland.
Willis, J.C., Bohan, D.A., Choi, Y.H., Conrad, K.F. and Semenov, M.A. 2006. Use of an individual-based model to forecast the eff ect of climate change on the dynamics, abundance and geographical range of the pest slug Deroceras reticulatum in the UK. – Global Change Bio-logy 12:1643-1657.
Willis, J.C., Bohan, D.A., Powers, S.J., Choi, Y.H., Park, J. and Gussin, E. 2008. The importance of temperature and moisture to the egg-laying behaviour of a pest slug, Deroceras reticu-latum. – Annals of Applied Biology 153:105-115.
Yancey, P.H. 2005. Organic osmolytes as compatible, metabolic and counteracting cytoprotectants in high osmolarity and other stresses. – Journal of Experimental Biology 208:2819-2830.
Zachariassen, K.E. 1985. Physiology of cold tolerance in insects. – Physiological Reviews 65:799-832.
Zachariassen, K.E. 1980. The role of polyols and nucleating agents in cold-hardy beetles. – Journal of Comparative Physiology B 140:227-234.
Zotin, A.A. 2007. Patterns of individual growth of grey garden slug Deroceras reticulatum. – Biology Bulletin 34:457-462.

35PhD thesis by Stine Slotsbo
Submitted to Annals of Applied Biology
Stine Slotsbo1,2, Christian Damgaard2, Lars Monrad Hansen1 and Martin Holmstrup2
1Aarhus University, Department of Agroecology, Forsøgsvej 1, DK-4200 Slagelse, Denmark2Aarhus University, Department of Bioscience, Vejlsøvej 25, DK-8600 Silkeborg, Denmark
PAPER I
The infl uence of temperature on life history traits in the Iberian slug, Arion lusitanicusRunning page heading: Thermal biology of A. lusitanicus
Juvenile Arion lusitanicus. Photo: Stine Slotsbo.


37PhD thesis by Stine Slotsbo
Abstract
The slug Arion lusitanicus is an invasive species which has recently become established in many Euro-pean countries and is considered a serious pest, both in agriculture and private gardens. The eff ect of temperature on growth rate, survival, egg developmental time and hatchability of the slug are report-ed. The growth rate was positively correlated with temperatures up to 20°C and A. lusitanicus seems to have infi nite growth. A model describing how growth depends on temperature was developed, and a fi eld experiments found that the predicted growth matched the observed with reasonable accuracy.
The present study confi rms the assumption that A. lusitanicus die after egg-laying. Free-ranging A. lusitanicus has an annual life cycle, but our data shows that slugs also possess the ability to live for up to 3 years.
The egg developmental time was like growth positive correlated with temperature and was shortest at 20°C. But the optimum temperature for egg development, when regarded as the temperature with highest hatchability was found to be 10°C. In conclusion, the present study has shown that growth and the life history traits egg developmental time and hatchability of A. lusitanicus are highly infl uenced by temperature. However, data also show that temperatures within the range 2-20°C did not infl uence survival rates to any large degree, and that A. lusitanicus can quickly adjust to changes in temperature. The study provides a number of basic parameters which contribute to the understanding of the popula-tion dynamics of A. lusitanicus.
Key words: Arion lusitanicus, slugs, growth, growth model.
The infl uence of temperature on life history traits in the Iberian slug, Arion lusitanicus
Running page heading: Thermal biology of A. lusitanicus
Stine Slotsbo1,2*, Christian Damgaard2, Lars Monrad Hansen1 and Martin Holmstrup2
1 Aarhus University, Department of Agroecology, Forsøgsvej 1, DK-4200 Slagelse, Denmark2 Aarhus University, Department of Bioscience, Vejlsøvej 25, DK-8600 Silkeborg, Denmark
*Corresponding author: Stine Slotsbo, Aarhus University, Department of Bioscience, Vejlsøvej 25, DK-8600 Silkeborg, Denmark.

38 PhD thesis by Stine Slotsbo
Introduction
Slugs are important pests of a wide range of horticultural and arable crops across Europe (South, 1992), and the more frequent use of conservation tillage in agri-culture is leading to an increase in slug pest problems (Norris, 2003). This is be-cause the reduction of mechanical disturbance of the soil and the retention of litter favours slug survival and reproduction (Hunter, 1967; Hammond, 1985). The slug Arion lusitanicus Mabille (Gatropoda: Pulmonata: Arionidae) is an invasive species which has recently become established in many European countries and is con-sidered a serious pest, both in agriculture and private gardens (Kozlowski, 2007). The region of origin is unknown; earlier it was assumed to originate from the Iberi-an Peninsula, a misconception that gave rise to its common name, the Iberian slug (Quinteiro et al., 2005). A. lusitanicus seems to cope well with the local conditions in the new areas to which it has migrated, it spreads rapidly and often occurs in high densities. The population size of A. lusitanicus can vary substantially between years (Grimm & Paill, 2001) likely due to climatic factors such as temperature and precipitation. Low winter temperatures are unlikely to limit the populations under Danish conditions (Slotsbo et al., 2011a) whereas drought periods will constrain activity and reproduction, but are unlikely to cause substantial mortality (Grimm et al., 2000; Sternberg, 2000; Kozlowski 2001; Slotsbo et al., 2011b). There is a need for better knowledge of the ecophysiology of A. lusitanicus in order to be able to predict the eff ect of climatic factors, such as temperature on the population dy-namics and abundance.
Arion lusitanicus occurring in NW Europe are out-crossing hermaphrodites that is assumed to have an annual life cycle (Kozlowski & Kozlowski, 2000). Egg pro-duction starts in the summer period and continues in some years until December (Kozlowski & Kozlowski, 2000; Grimm, 2001). Field observations and laboratory studies indicates that the majority of A. lusitanicus die after egg-laying (Davies, 1987; Briner & Frank, 1998; Kozlowski & Kozlowski, 2000). The egg developmental time and hatching success are infl uenced by temperature (Kozlowski, 2000) and humidity (Slotsbo et al., 2011b), and may vary geographically.
The growth of slugs, like many other invertebrates, is infl uenced by temperature (South, 1982). Detailed studies of A. lusitanicus growth rates and longevity do not exist, although the overall life cycle is well described (Kozlowski & Kozlowski, 2000; Grimm, 2001).
Abeloos (1943,1944) made some detailed studies of the growth in fi ve Arion and three Limax species and described growth as three phases; First an infantile phase with rapid growth, followed by a juvenile phase with slower growth and fi nally an adult phase with minimal growth. South (1982) studied the correlation between growth and temperature in the species Deroceras reticulatum (Müller) and Arion intermedius Normand, and found a positive correlation between growth and in-creasing temperatures.
A huge challenge in pest slug management today, is to predict timing of the pest problem. The lack of reliable predictive models often results in frequent and inef-fective use of molluscicides. Species specifi c growth models enable pest manag-ers to predict the abundance and timing of the pest slugs in question. For a model to be aff ective it has to take into account how the life history traits of a particular species are aff ected by climate condition such as daily temperature, rainfall and winter conditions. Several growth models have been developed for the serious agricultural pest Deroceras reticulatum (Shirley et al., 2001; Choi et al., 2004; Wil-

39PhD thesis by Stine Slotsbo
lis et al., 2006), but there have not been developed a model for A. lusitanicus. Development of a good model for this species would result in higher effi ciency and reduced application of molluscicides which would have both economic and environmental benefi ts.
The aims of this study were to determine how temperature aff ects growth rate, survival, egg developmental time and hatchability of A. lusitanicus. Further, we wanted to test the hypothesis of post-reproductive mortality. These studies pro-vide some basic parameters of population dynamics, which would be the fi rst step required to develop integrated management strategies for the control of A. lusitanicus.
Materials and Methods
Test animals
Adult specimens of A. lusitanicus were collected in a public park near Randers, Denmark (N56° 26’; E10° 03’), in September 2008. To ascertain correct identifi ca-tion, 24 representative specimens were identifi ed to taxon by oviduct morphology and the position of the ligula (e.g.; Moquin-Tandon, 1855; Kerney et al., 1979; No-ble, 1992). In addition, they were also identifi ed using DNA sequences of a 410bp fragment of the mitochondrial 16S rRNA as described in Slotsbo et al. (2012). All sequenced specimens corresponded to the species A. lusitanicus and the identifi -cation was confi rmed by the morphology of the reproductive organs.
The collected slugs were kept at 15°C in plastic boxes (17×22×6 cm) and fed carrots, lettuce, oatmeal and dog food pellets ad libitum. The bottom of each box was covered with wet paper towel to ensure high humidity. Five slugs were kept in each box so that they had the opportunity to mate, if they had not already been fertilized in the fi eld. The collected slugs produced eggs; some eggs were used in egg development experiments, whereas others were allowed to hatch by in-cubating the eggs at 15°C in Petri dishes lined with wet fi lter paper. These newly hatched juveniles were then used in growth experiments.
Development time of eggs
The fi rst experiment examined the development time (time elapsed from laying to hatching) of eggs at diff erent temperatures. The eggs were laid at 15°C, and were within 24 h moved to the experimental temperatures, which were either 5, 10, 15 or 20 °C. The eggs were kept in Petri dishes lined with wet fi lter paper. Five Petri dishes, each containing 20-32 eggs, were placed at each experimental tempera-ture. Hatching was recorded every second to third day.
Growth and mortality rates of slugs
Newly hatched juveniles (0-48 h old) were moved to experimental tempera-tures of 2, 5, 10, 15 or 20°C. The newly hatched juveniles were placed individu-ally in plastic boxes (17×22×6 cm) and kept as described above. The boxes were cleaned and new food added twice a week.
The growth and surv ival of 30 juveniles was monitored at each experimental tem-perature. Mortality of slugs was checked twice a week and their fresh weight was determined 15 times during the fi rst year of the experiment (358 days). In order to

40 PhD thesis by Stine Slotsbo
study changes of growth rate when shifting temperature from low to higher, the specimens raised at 2 and 5°C for a year were moved to 15°C and kept at this tem-perature for another 259 days.
Growth model
The growth of the slugs during the fi rst 358 days of the growth experiment was de-scribed using a structural equation modeling approach (Damgaard, in press) link-ing growth to temperature and time by following modifi ed Richards growth model:
110 1)()(
1)( −
⎟⎠⎞⎜
⎝⎛ −
−+
=δ
αττ
δκκ
ττ vvtemp
dvd
(1)
where v(τ) is the weight at time τ, temp10 κκ + is the initial relative growth rate, which is assumed to be a linear function of temperature, α is the asymptotic mean of the fresh weight, and δ is a shape parameter that control the infl ection point of the growth curve (Seber, 1989).
The conditional likelihood functions of the structural equation models describes the growth between the census times, t, for each temperature by latent variables,
temptx , , using a re-parameterized gamma distribution ),;( pxGamma νμ where μ is the expected weight and νp is a scale parameter.
),)(;(1
,1, p
t
ttempttempt dvxxGamma νττ∫
−− + (2),
where temptxtv ,1)1( −=− . The gamma distribution was used to model the vari-ance of the fresh weight since the slope in a log (variance) - log (mean) plot was approximately two (McCullagh & Nelder, 1989). The measurement equation link the latent variables to the observed values by the following likelihood function.
),;( ,, mtempttempt xyGamma ν (3),
where tempty , is the observed fresh weight of the slugs at the census times, t, for temperature temp, and νm is a scale parameter.
The likelihood function of a full structural equation model was found by multiplying structural equation (2) and measurement equation (3) for all size increments and temperatures.
The structural equation model was parameterized using numerical Bayesian methods, where the joint posterior distribution of the parameters and the latent variables was calculated using MCMC (Metropolis-Hastings) simulations (Carlin & Louis, 1996) with a multivariate normal candidate distribution, and using a MCMC run of 50 000 iterations with a burn-in period of 100 iterations. The prior distribu-tions of all parameters and the latent variables were assumed to be uniformly distributed in their assumed domains, i.e. δ > 1, νp < 20, νm < 25, and the latent vari-ables were assumed to be positive. The domain of the scale parameter of the gamma distributions were constrained in order to assure realistic estimates of the simulated variance compared to the observed variance. Plots of the sampling chains of all parameters and latent variables were inspected in order to check the mixing properties of the used sampling procedure.

41PhD thesis by Stine Slotsbo
The median values of the marginal posterior distribution of the parameters were inserted into growth equation (1) and the predicted growth was calculated using information of the actual temperatures.
Post-reproductive mortality
An experiment was set up to test the hypothesis that slugs die after egg laying. Of the surviving slugs raised in isolation at 10, 15 and 20°C for one year, 6 to 8 slugs from each temperature were combined in pairs, so having an opportunity to mate. Remaining survivors were kept singly as controls. Pairs and individuals were kept at the temperatures by which they where raised. During the following 690 days their egg laying and survival was registered.
Longevity
The survival of slugs raised at 10, 15 and 20°C that did not lay eggs was followed for almost 3 years (1048 days). The growth of these slugs was also registered by weighing them at 10 occasions in the period from day 358 to day 1048.
Field experiment
Growth in the fi eld was estimated based on the development in average body weight of trapped slugs under the assumption that the majority of overwintering slugs were small juveniles (Kozlowski & Kozlowski, 2000; Kozlowski, 2007; Slotsbo et al., 2011a). Slugs were sampled using pitfall traps (Eb rør, Tyholm, Denmark), placed in 20 diff erent private gardens located in east Jutland, Denmark, in the period from April to October, 2009. The traps were placed in trimmed lawns with a distance of 50 cm from kitchen garden, herb garden, fl ower bed or other similar herb vegetation in which slugs had been observed.
The trap consisted of a closed black plastic cylinder (diameter 16 cm, height 27 cm), with two openings, one on each side of the cylinder. The traps were buried to 15 cm depth. The openings of the pitfall traps were 4.5 cm in diameter and placed 7 cm above ground. Pilsner type beer (4.8 % vol. ethanol) was used to attract the slugs, as beer is attractive for snails and slugs (Edwards, 1991). The traps were checked for slugs daily and were recharged with beer weekly. All collected slugs were brought to the laboratory for species identifi cation and fresh weight determination.
Statistical analysis
The relationship between developmental time of eggs and incubation temperature was tested using non-linear regression. Diff erences in hatchability between diff erent incubation temperatures were tested using Chi-square tests. All statistical tests were performed using Sigmaplot for Windows Version 11 (SPSS Inc., Chicago, IL, USA).
Modeling of growth was performed using Mathematica for Windows Version 7 (Wolfram Research, Inc., Champaign, IL, USA).
The slug mortality during the fi rst 358 days of the growth experiment was ana-lyzed using a two-parameter Weibull distribution that was fi tted to the censored survival data. The shape parameter of the Weibull distribution can be interpreted directly as follows; a value of k < 1 indicates that the mortality decreases with ages. This happens if there is signifi cant “infant mortality”. A value of k = 1 indicates that the mortality is constant at all ages and a value of k > 1 indicates that the mortal-ity increases with ages. Diff erences in mortality distribution between the diff erent temperatures were tested using likelihood ratio tests

42 PhD thesis by Stine Slotsbo
Results
Development time of eggs
Developmental time was shortest at 20°C (37 days). The correlation between developmental time D(T) and incubation temperature (T) is non-linear and can be described by the following equation: D(T) = 32.9+340.6 exp(-0.23T) (R2= 0.98; P < 0.0001). The developmental time calculated as degree days was on the aver-age 676 days under the assumption that the developmental null point was 0°C (Table 1).
The hatchability of the eggs diff ered signifi cantly between temperatures (Chi-square; P < 0.005), with the lowest hatchability at 20°C and the highest at 10°C (Table 1).
Growth and mortality rates of slugs
The slugs gained weight at all temperatures throughout the whole experiment (358 days) (Fig. 1). The growth rate of the slugs depended on temperature, shown by κ1
being signifi cantly larger than 0 (κ1= 6.35×10-4; 95% credibility interval (5.99×10-4 - 6.71×10-4)). Higher temperatures resulted in higher growth rates (Fig. 1).
The survival curves calculated for each temperature group displayed an ade-quate fi t to the data, and mortality was not signifi cantly diff erent between the dif-ferent temperatures (P = 0.24). The shape parameter k was 0.727, indicating that the mortality rate in this experiment decreased with age, meaning that younger slugs had a signifi cantly higher mortality rate than older slugs (Fig. 2; P < 0.0001).
Temperature (°C) Eggs development time (days)
Degree days needed for development
Hatchability (%)
20°C 37.4 ± 0.9 (n = 57) 747 51 (n= 112)
15°C 41.6 ± 0.5 (n = 77) 623 69 (n= 111)
10°C 66.4 ± 0.4 (n = 90) 664 81 (n= 111)
5°C 138.6 ± 1.1 (n = 72) 693 65 (n= 111)
Table 1. Development time of Arion lusitanicus eggs incubated at constant temperatures of 5, 10, 15 or 20°C, and the hatchability at these temperatures. The average number of de-gree days at each temperature is indicated. Values are mean ± SE and number of obser-vations indicated in brackets.
Age (days)0 100 200 300 400
Fre
sh w
eigh
t (g)
0
5
10
15
20
25
30 2°C 5°C 10°C 15°C 20°C
Figure 1. Growth of Arion lusi-tanicus raised at constant tem-peratures of 2°C, 5°C, 10°C, 15°C or 20°C. Values are mean ± SE (N = 6-30)

43PhD thesis by Stine Slotsbo
When the slugs raised at 2°C or 5°C for one year, were moved to 15°C, they quick-ly resumed the same growth rate as the slugs raised at constant 15°C (Fig. 3), meaning that the growth rate was determined by the temperature and size of the slug, and not by the age.
Post-reproductive mortality and longevity
Of the 26 pairs of slugs (combined in pairs at age 358 days) only 3 pairs (six slugs) laid eggs. All the egg laying slugs died within 38-52 days after they had been combined in pairs. In comparison, the paired slugs that did not lay eggs survived much longer, with the fi rst slug dying 144 days after they had been combined in pairs and some slugs survived until the experiment was terminated, i.e. 690 days after they were combined in pairs. Of the 15 slugs that were not combined in pairs, 2 produced eggs, but the fi rst egg clutch was laid when the slugs were 458 days old. Eggs of one clutch hatched, showing that A. lusitanicus was able to self fer-tilize. At the termination of the experiment some slugs were still alive (1048 days old). Most slugs continued to grow throughout their life (Fig. 4), but some individu-als lost weight shortly before they died.
Field experiment and growth model
The average fresh weight of the fi eld collected slugs increased over the season from April to October (Fig. 5). The growth model (Equation 1) was fi tted to the laboratory-based growth data and using the actual fi eld temperatures (daily average, in the pe-
Sur
viva
l (nu
mbe
rs o
f slu
gs)
0
5
10
15
20
25
35
30
Age (days)0 100 200 300 400
2°C5°C
10°C15°C20°C
Fre
sh w
eigh
t (g)
Age (days)0 100 200 300 400 500 600 700
0
5
10
15
20
2515°C
Figure 2. Survival of juvenile Arion lusitanicus raised at constant tem-peratures of 2°C, 5°C, 10°C, 15°C or 20°C. Survival is shown as num-ber of live slugs at a given age. Since the mortality of the slugs was not signifi cantly diff erent be-tween the diff erent temperatures only one common survival curve is shown; this curve is described by the Weibull distribution with k = 0.727 and λ = 285.798.
Figure 3. Growth of juvenile Arion lusitanicus raised at a constant temperature of either 2°C or 5°C for 358 days, and then moved to 15°C and kept there. The triangles show growth of slugs raised at constant 15°C, from the time that they had reached the same fresh weight as slugs kept for 358 days at 2°C and 5°C, respectively (see Fig. 1). Values are mean ± SE.

44 PhD thesis by Stine Slotsbo
Figure 4. Growth of Arion lusi-tanicus raised at constant tem-peratures of 10°C (A), 15°C (B) or 20°C (C). Black dots are slug fresh weight and each line represents one slug.
Figure 5. (A) The daily mean temperature in East Jutland, Denmark in 2009. (B) The average fresh weight (g) of Arion lusitanicus captured in the fi eld at 20 diff erent locations in East Jutland in the period from 1 April to 1 October, 2009. Each black dot repre-sents the average fresh weight of slugs captured at that day. Values are mean ± SE (N = 1-36). The solid line shows the calculated growth using Equation (1) and the actual temperature given in Panel A. The parameters used in Equation (1) is as follows; α = 16.39, κ = 6.34×10-4, δ = 1.004.The data within the black triangle (lower right corner of Panel B) are most likely juveniles that hatched in late September/October
Fre
sh w
eigh
t (g)
Fre
sh w
eigh
t (g)
Fre
sh w
eigh
t (g)
Time (days)0 200 400 600 800 1000 1200
0
20
40
60
80
0
20
40
60
80
0
20
40
60
80
A) 10°C
B) 15°C
C) 20°C
Tem
pera
ture
(°C
)
Fre
sh w
eigh
t (g)
1st Apr 1st May 1st Jun 1st Jul 1st Aug 1st Sep 1st Oct1st Apr 1st May 1st Jun 1st Jul 1st Aug 1st Sep 1st Oct
0
5
10
15
20
25
0
4
8
12
16A B

45PhD thesis by Stine Slotsbo
riod from April 12 to October 1, 2009) and the initial weight set to the weight of a fi eld collected slug (collected 12th of April 2009). The growth model was used to predict the growth of Danish slugs under the assumption that growth is only regulated by tem-perature (Fig. 5). We found that the predicted weight during the season matched the observed increase in average body weight with satisfactory accuracy (Fig. 5).
Discussion
In the present study the developmental time of the eggs was shortest at 20°C (37.4 days). A study by Kozlowski (2000) also found the shortest developmental time (mean 30.3 days) at 20°C. Not surprisingly, high temperature increases egg devel-opment up to a certain limit. Kozlowski (2000) found no eggs development and hatching at 25°C which appears to be the upper temperature limit for A. lusitani-cus. Even though 20°C result in the most rapid development this temperature was not the optimum temperature for A. lusitanicus eggs. The optimum temperature, regarded as the temperature with the highest number of eggs surviving, was in Kozlowski’s study found to be 15°C and in the present study found to be 10°C. This diff erence may indicate that thermal relationships may diff er between geographi-cally separate populations.
Arion lusitanicus occurring in NW Europe is assumed to have an annual life cycle (Kozlowski & Kozlowski, 2000; Grimm, 2001). Adults begin egg-laying in late sum-mer, the exact time vary depending on geographical region and also fl uctuate between years within an area (Kozlowski & Kozlowski, 2000; Grimm, 2001; Slots-bo, unpublished). The last egg laying may occur as late as December depending on temperature (Kozlowski & Kozlowski, 2000; Slotsbo, unpublished).
In the present study the developmental time of eggs was in range 623-747 degree days. Using this information together with the average temperatures in Denmark at the time of egg-laying, it shows that most eggs will hatch before winter. Eggs laid as late as mid September are expected to hatch in the beginning of December. A study on egg hatching made in Poland in the autumn/winter 1999 also found that most of the eggs hatched before winter (Kozlowski & Sionek, 2000). However, egg hatching was only registered until January, and therefore it was not shown wheth-er eggs hatch during winter or in the following spring. Calculating the time of egg hatching using the development time found in this study together with average Danish temperatures, fi nds that eggs laid after October 1 will probably not be able to hatch before the following spring. Likewise Kozlowski and Sionek, (2000) found that eggs deposited in late September and November did not hatch before win-ter. Whether the eggs will hatch in the following spring depend on their ability to survive the winter. A previous study found that the eggs of A. lusitanicus were not particularly cold resistant suggesting that egg survival during winter depends on frost-free microhabitats (Slotsbo et al., 2011a).
Field observations obtained in Poland, Great Britain and Switzerland (Davies, 1987; Briner & Frank, 1998) indicate that the majority of A. lusitanicus die after egg-laying. Laboratory observation by Kozlowski and Kozlowski (2000) supported this assumption by fi nding that 75 % of the slugs kept in laboratory died after egg-laying before winter. Our study confi rms that A. lusitanicus die after eggs-laying and that the mortality was not infl uenced by ambient temperature or age of the slugs. Post-reproductive mortality has also been found in the related slug A. rufus (Rollo, 1983), and overwintering of adult slugs of these species seems therefore to be a rare phenomenon.

46 PhD thesis by Stine Slotsbo
Growth of slugs has been studied in several species; some studies focus on pat-terns of individual growth, and others have been carried out on groups of animals (Abeloos, 1943; Abeloos, 1944; South, 1982; Hommay et al., 2001; Zotin, 2007; Clemente, 2008; Clemente, 2010). One slug species is well described with re-spect to growth, namely the pest species D. reticulatum. (South, 1982; Zotin, 2007; Clemente, 2008; Clemente, 2010). However the models developed to describe population growth of D. reticulatum (Shirley et al., 2001; Choi et al., 2004; Willies et al., 2006) cannot be used to describe the population dynamics of A. lusitanicus, because the two species diff er in growth pattern and life cycle. For example, D. reticulatum is assigned to animals with fi nite growth, were A. lusitanicus seems to have infi nite growth. Further on, D. reticulatum normally has several generations each year, where A. lusitanicus has only one (South, 1982; Kozlowski & Kozlowski, 2000; Grimm, 2001; Willis, 2006).
The growth rate of D. reticulatum is, like many other invertebrates, positively cor-related to temperatures (South, 1982; Clemente, 2008). Our study showed that this also is the case for A. lusitanicus. We found that our growth model (Equation (1)) provide a useful tool to estimate growth of A. lusitanicus under constant or fl uctu-ating temperatures. The growth model enabled the prediction of individual slug growth under natural thermal scenarios, suggesting that food availability appar-ently is not a limiting factor under these conditions. The parameters of our model were found by fi tting the model to data of the fi rst year of growth at all fi ve diff er-ent temperatures used in the experiment. The parameter a is the asymptotic mean of the maximum fresh weight to be described by the model and was estimated to 16 gram. However, the estimation of a is only used for curve fi tting and has in fact no biological relevance knowing that A. lusitanicus has no asymptotic maxi-mum body weight when kept in the laboratory, but instead exhibit indeterminate growth. Our model is therefore restricted to predict slug growth until the average slug weight in the population reaches a maximum of 16 gram. In practise, this is not a marked problem for predicting growth in the fi eld since most of the slugs in the fi eld will lay eggs and then die before the average slug weight in the popula-tion reach 16 g. Our fi eld data corroborate this assumption since slugs trapped in the fi eld in 2009 did not attain a mean weight above 16 g.
In order to construct a population growth model for A. lusitanicus, we need data describing the diff erent life history traits of importance for population growth. Most of these data exist, many of them have been quantifi ed through the present study, but one important trait have not been described; namely, at which size does A. lusitanicus become reproductively mature. Kozlowski and Kozlowski (2000) found that the individuals mainly laid eggs at an age of 6 to 9 month, but whether their ability to lay egg was caused by their age, the time of the year or the body weight was not revealed. Unpublished observations in our laboratory have shown that body size of reproductive slugs varies considerably. In the related slug, A. rufus, body size is neither the main determinant for maturity (Lusis, 1961). Photoperiod (day length) is employed by many species of animals as a seasonal cue to control the annual reproductive cycle (Menaker, 1971) including the slug, Limax maximus (Sokolove & McCrone, 1978; Rollo, 1983). Photoperiod could very well be the cue controlling maturity in A. lusitanicus as well.
Even though free-ranging A. lusitanicus has an annual life cycle, our data shows that they also posses the ability to live for much longer. Large specimens have on occasions been found in nature in the spring (Kozlowski and Kozlowski, 2000), which indicate that semivoltine life-cycles also occurs in nature. The fact that A. lusitanicus is able to live for more than a year, and hereby have the ability to wait and reproduce the following season if they do not succeed in the fi rst year, makes this species very robust against constraining climatic conditions, and perhaps also

47PhD thesis by Stine Slotsbo
in situations such as introductions to new regions where mating may be restricted due to low population density. The present study showed that A. lusitanicus was indeed able to self-fertilize, but that this was not a very common phenomenon. It could be speculated that self-fertilization could be an advantage in situations where mating is not possible, however, our study have not investigated if such progeny is in fact fertile.
When the slugs raised at 2°C and 5°C were moved to 15°C, they quickly obtained growth rates similar to the specimens raised at constant 15°C. It is an advan-tage for the slugs that their growth rate decreases when the temperature drops; it means that their metabolic requirements are low during winter when food may be limited. Further, it is quite remarkable and demonstrating a high degree of plastic-ity that slugs even after a year of very slow growth, without any delay, are able to obtain rapid growth rates when exposed to higher temperatures. In the spring this means that overwintering juveniles immediately starts growing when temperature rises provided that food is not a limiting factor.
In conclusion, the present study has shown that growth and the life history traits egg developmental time and hatchability of A. lusitanicus are highly infl uenced by temperature. However, data also show that temperatures within the range 2-20°C did not infl uence survival rates to any large degree, and that A. lusitanicus can quickly adjust to changes in temperature. Our study provides a number of basic parameters which may contribute to the understanding of the population dynamics of A. lusitanicus. There is, however, a need for further studies of fi eld populations, which together with data from this study could form the basis of a model that could provide a more accurate forecasting of slug abundance.
Acknowledgments
Thanks go to my colleagues that have allowed me to use their gardens for the fi eld survey collection of trapped slugs every day for six months. I also thank Kurt Jordanes for help with species identifi cation of the collected slugs.

48 PhD thesis by Stine Slotsbo
References
Abeloos, M. (1943). Forme de la croissance des espéces du genre Limax. Comptes Rendus de l’ Académie des Sciences, Paris 217:159-161.
Abeloos, M. (1944). Recherches expérimentales sur la croissance. La croissance des mol-lusques Arionidés. Bulletin Biologique de la France et de la Belgique. 78:215-256.
Briner, T. & Frank, T. (1998). Egg laying activity of the slug Arion lusitanicus Mabille in Swit-zerland. Journal of Conchology 36:9-15.
Carlin, B.P. & Louis, T.A. (1996). Bayes and empirical Bayes methods for data analysis. Chapman & Hall, London.
Choi, Y.H, Bohan, D.A., Powers, S.J., Wiltshire, C.W., Glen, D.M. & Semenov, M.A. (2004). Mod-elling Deroceras reticulatum (Gastropoda) population dynamics based on daily tempera-ture and rainfall. Agriculture, Ecosystems and Enviroment 103:519-525.
Clemente, N.L., López, A.N., Monterubbianesi, M.G., Cazzaniga, N.J. & Manetti, P.L. (2008). Biological studies and phenology of slug Deroceras reticulatum (Müller, 1774) (Pulmonata: Stylommatophora). Invertebrate Reproduction and development 52:23-30.
Clemente, N.L., Faberi, A.J., Salvio, C. & López, A.N. (2010). Biology and individual growth of Milax gagates (Draparnaud, 1801) (Pulmonata: Stylommatophora). Invertebrate Reproduc-tion and development 54:163-168.
Damgaard, C. (in press). Trend analyses of hierarchical pin-point cover data. Ecolog
Davies, S.M. (1987). Arion-Flagellus Collinge and Arion-Lusitanicus Mabille in the British-Isles - A morphological, biological and taxonomic investigation. Journal of Conchology, 32:339-354.
Edwards, C.A. (1991). The assessment of populations of soil-inhabiting invertebrates. Agri-culture, Ecosystems & Environment 34:145-176.
Grimm, B. (2001). Life cycle and population density of the pest slug Arion lusitanicus Ma-bille (Mollusca: Pulmonata) on grassland. Malacologia 43:25-32.
Grimm, B. & Paill, W. (2001). Spatial distribution and home-range of the pest slug Arion lusitani-cus (Mollusca: Pulmonata). Acta Oecologica-International Journal of Ecology 22:219-227.
Grimm, B., Paill, W., & Kaiser, H. (2000). Daily activity of the pest slug Arion lusitanicus Ma-bille. Journal of Molluscan Studies, 66, 125-128.
Hammond, R. B. (1985). Slugs as a New Pest of Soybeans. Journal of the Kansas Entomo-logical Society, 58, 364-366.
Hommay, G., Kienlen, J.C., Gertz, C. and Hill, A. (2001). Growth and reproduction of the slug Limax valentianus Férussac in experimental conditions. Journal of Molluscan Studies 67:191-207.
Hunter, PJ. (1967). Eff ect of Cultivations on Slugs of Arable Land. Plant Pathology 16:153-156.
Kerney, M.P., Cameron, R.A.D., & Riley, G. (1979). A fi eld guide to the land snails of Britain and Northwest Europe. Wm. Collins Sons & Co., London.
Kozlowski, J. (2000). Reproduction of Arion lusitanicus Mabille, 1868 (Gastropoda: Pulmo-nata: Arionidae) introduced in Poland. Folia Malacologica 8:87-94.
Kozlowski, J. (2001). Daily activity of Arion lusitanicus Mabille, 1868 (Gastropoda: Pulmo-nata: Arionidae). Journal of Plant Protection Research 4:279-286.
Kozlowski, J. (2007). The Distribution, biology, population dynamics and harmfulness of Arion lusitanicus Mabille, 1868 (Gastropoda: Pulmonata: Arionidae) in Poland. Journal of Plant Protection Research 47:219-230.
Kozlowski, J. & Kozlowski, R.J. (2000). Periods of occurrence and fecundity of Arion lusitani-cus Mab. (Gastropoda: Pulmonata) in crop plant habitats in Poland. Journal of Plant Protec-tion Research 40:260-266.

49PhD thesis by Stine Slotsbo
Kozlowski, J. & Sionek, R. (2000). The rate of egg laying and hatching of the slug Arion lusi-tanicus Mabille, a pest of arable crops. Journal of Plant Protection Research 40:160-167.
Lusis, O. (1961). Postembryonic changes in the reproductive system of the slug Arion Ater Rufus L. Proceedings of the Zoological Society of London 137:433-468.
McCullagh P. & Nelder J.A. (1989. Generalized Linear Models, 2 edn, Chapman & Hall/CRC, Boca Raton.
Menaker, M. (1971). Rhythms, reproduction, and photoreception. Biology of Reproduction 4:295-308.
Moquin-Tandon, A. (1855). Histoire Naturelle des Mollusques Terrestres et Fluviatiles de France, Tome Second. Chez J.-B.Bailliére, Libraire de L’ Académie Impériale de Médecine, rue Hautefeuille 19, Paris 2:1-17.
Noble, L.R. (1992). Diff erentiation of Large arionid slugs (Mollusca, Pulmonata) using ligula morphology. Zoologica Scripta 21:255-263.
Norris, R.F., Caswell-Chen, E.P., & Kogan, M. (2003). Concepts in integrated pest manage-ment. Prentice Hall, New Jersey.
Quinteiro, J., Rodriguez-Castro, J., Castillejo, J., Iglesias-Pineiro, J., & Rey-Mendez, M. (2005). Phylogeny of slug species of the genus Arion: evidence of Iberian endemics and of the existence of relict species in Pyrenean refuges. Journal of Zoological Systematics and Evo-lutionary Research 43:139-148.
Rollo, C.D. (1983). Consequences of competition on the reproduction and mortality of 3 species of terrestrial slugs, Researches on Population Ecology 25:20-43.
Seber, G.A.F & Wild, C.J. (1989). Nonlinear regression. John Wiley & Sons. New York.
Shirley, M.D.F., Rushton, S.P., Young, A.G. & Port, G.R. (2001). Simulating the long-term dy-namics of slug populations: a process-based modelling approach for pest control. Journal of Applied Ecology 38:401-411.
Slotsbo, S., Hansen, L.M., & Holmstrup, M. (2011a). Low temperature survival in diff erent life stages of the Iberian slug, Arion lusitanicus. Cryobiology 62, 68-73.
Slotsbo, S., Fisker, K., Hansen, L., & Holmstrup, M. (2011b). Drought tolerance in eggs and juveniles of the Iberian slug, Arion lusitanicus. Journal of Comparative Physiology B: Bio-chemical, Systemic, and Environmental Physiology 181:1001-1009.
Slotsbo, S., Hansen, L., Jordaens, K., Backeljau, T., Malmendal, A., Nielsen N. & Holmstrup, M. (2012). Cold tolerance and freeze-induced glucose accumulation in three terrestrial slugs. Comparative Biochemistry and Physiology Part A. 161:443-449.
Sokolove, P. G. & McCrone, E. J. (1978). Reproductive maturation in slug, Limax-maximus, and eff ects of artifi cial photoperiod. Journal of Comparative Physiology 125:317-325.
South, A. (1982). A comparison of the life cycles of D. reticulatum (Müller) and A. interme-dius Normand (Pulmnata: Stylommatophora) at diff erent temperatures under laboratory conditions. Journal of Molluscan Studies 48:233-244.
South, A. (1992). Terrestrial Slugs. Biology, Ecology and Control., 1 edn, Chapman & Hall, London.
Sternberg, M. (2000). Terrestrial gastropods and experimental climate change: A fi eld study in a calcareous grassland. Ecological Research 15:73-81.
Willis, J.C., Bohan, D.A., Choi, Y.H., Conrad, K.F. & Semenov, M.A. (2006). Use of an individ-ual-based model to forecast the eff ect of climate change on the dynamics, abundance and geographical range of the pest slug Deroceras reticulatum in the UK. Global Change Biology 12:1643-1657.
Zotin, A.A. (2007). Patterns of individual growth of gray garden slug Deroceras reticulatum. Biology Bulletin 34:457-462.


51PhD thesis by Stine Slotsbo
Cryobiology 62 (2011) 68–73, doi:10.1016/j.cryobiol.2010.12.005
Stine Slotsboa,b, Lars Monrad Hansena & Martin Holmstrupb
aFaculty of Agricultural Sciences, Aarhus University, Department of Integrated Pest Management, Forsøgsvej 1, DK-4200 Slagelse, DenmarkbNational Environmental Research Institute, Aarhus University, Department of Terrestrial Ecology, P.O. Box 314, Vejlsøvej 25,
DK-8600 Silkeborg, Denmark
PAPER II
Low temperature survival in diff erent life stages of the Iberian slug,Arion lusitanicus
Photo: Bresson Thomas/Wikimedia Commons.


53PhD thesis by Stine Slotsbo
Author's personal copy
Low temperature survival in different life stages of the Iberian slug,Arion lusitanicusq
Stine Slotsbo a,b,⇑, Lars Monrad Hansen a, Martin Holmstrup b
a Faculty of Agricultural Sciences, Aarhus University, Department of Integrated Pest Management, Forsøgsvej 1, DK-4200 Slagelse, DenmarkbNational Environmental Research Institute, Aarhus University, Department of Terrestrial Ecology, P.O. Box 314, Vejlsøvej 25, DK-8600 Silkeborg, Denmark
a r t i c l e i n f o
Article history:Received 1 September 2010Accepted 13 December 2010Available online 17 December 2010
Keywords:Arion lusitanicusSlugsFreeze toleranceSupercooling
a b s t r a c t
The slug Arion lusitanicus Mabille (Gastropoda: Pulmonata: Arionidae) is an invasive species which hasspread to most parts of Europe. The area of origin is unknown, but A. lusitanicus seems to cope well withthe local conditions in the countries to which it has migrated. It spreads rapidly, occurs often in high den-sities and has become a serious pest in most European countries. Therefore there is an urgent need forbetter knowledge of the ecophysiology of A. lusitanicus, such as the influence of climatic conditions, inorder to develop prognostic models and strategies for novel pest management practises.The aim of our study was to investigate the influence of subzero temperatures in relation to winter
survival. A. lusitanicus is shown to be freeze-tolerant in some life stages. Most juveniles and some adultslugs survived being frozen at �1.3 �C for 3 days, but none of the slugs survived freezing at �3 �C. Theeggs survived subzero temperatures (down to �2 �C) probably by supercooling. Juveniles and adultsmay also survive in a supercooled state (down to �3 �C) but are generally poor supercoolers. Therefore,the winter survival of A. lusitanicus depends to a high degree on migration to habitats protected from lowwinter temperatures, e.g. under plant litter, buried in the soil or in compost heaps.
� 2010 Elsevier Inc. All rights reserved.
Introduction
Slugs are important pests of a wide range of horticultural andarable crops across Europe [30]. The slug Arion lusitanicus Mabille(Gastropoda: Pulmonata: Arionidae) is an invasive species whichhas spread tomost parts of Europe,where it often has become a seri-ous pest [16]. The country of origin is unknown; earlier it was as-sumed to originate from the Iberian Peninsula, a misconceptionthat gave rise to its common name, the Iberian slug [22].A. lusitanicus spreads rapidly and often occurs in high densities. Itseems to cope well with the local conditions in the countries towhich it hasmigrated, but knowledge of its ecophysiology is limited.
Ectothermic animals living in habitats where they are exposedto frost have evolved different physiological and biochemicaladaptations to survive. These adaptations can be divided into threemajor strategies: freeze-tolerance, freeze avoidance and cryopro-tective dehydration. Freeze tolerance, is to establish controlled,protective freezing of the extracellular body fluids at high sub-zerotemperatures. Freeze avoidance, is based on the ability to stay in asupercooled state even at temperatures much below the melting
point of body fluids [23,36]. For small permeable soil invertebratesthere is a third option, cryoprotective dehydration. Through dehy-dration such organisms rapidly equilibrate their melting point tothe surrounding temperature in a frozen environment [10].
Cold hardiness of invertebrates has been widely studied[13,29,31], but literature on cold hardiness of the terrestrial snailsand slugs is more limited. Cold tolerance in land snails is primarilybased on freeze avoidance [1,3,24,25]. They are protected againstinoculative freezing by their impermeable shell and a tough winterepiphragm [6,25]. Most land snails bury in the soil or hide in cre-vices where temperatures are milder during winter [1].
Only few studies on the survival of terrestrial slugs at low tem-peratures exist [8,18,32,33]. Terrestrial slugs lack, in contrast toland snails, physical protection such as shell and epiphragm. Thelack of protection and their moist skin likely make them suscepti-ble to inoculative freezing. Hence, their winter survival will prob-ably depend on migration to frost-free hibernation sites, freezetolerance, cryoprotective dehydration or overwintering as coldtolerant eggs only.
As a cold hardiness strategy, cryoprotective dehydration wouldprobably only be an option for small juveniles or eggs. Organismsusing this strategy must be small for the dehydration process tooccur rapidly enough to avoid ice formation. Small permeableinvertebrates such as juvenile slugs lose water if the water vapourpressure in equilibrium with their body fluid is larger than thewater vapour pressure in the surrounding atmosphere. The vapour
0011-2240/$ - see front matter � 2010 Elsevier Inc. All rights reserved.doi:10.1016/j.cryobiol.2010.12.005
q Statement of funding: This study did not receive any external funding.⇑ Corresponding author at: National Environmental Research Institute, Aarhus
University, Department of Terrestrial Ecology, P.O. Box 314, Vejlsøvej 25, DK-8600Silkeborg, Denmark. Fax: +45 8920 1481.
E-mail address: [email protected] (S. Slotsbo).
Cryobiology 62 (2011) 68–73
Contents lists available at ScienceDirect
Cryobiology
journal homepage: www.elsevier .com/locate /ycryo

54 PhD thesis by Stine Slotsbo
Author's personal copy
pressure of supercooled water is higher than that in equilibriumwith ice [27,34]. Therefore slugs placed in a closed system withice could, if not frozen, lose water until equilibrium in water va-pour pressure is attained.
In many invertebrates, eggs or cocoons are very resistant to lowtemperatures [7,9,13], but there is no literature on the cold hardi-ness of slug eggs which, because of their shell, could utilize freezeavoidance as a strategy to survive low temperature.
The aim of the present study was to investigate if A. lusitanicuscan survive sub-zero temperatures and which cold tolerance strat-egy they use. Further, we wanted to clarify whether the ability tocope with low temperature is dependent on life-stage.
Materials and methods
Test animals
Adult specimens of A. lusitanicus were collected in a public parknear Randers, Denmark (N56� 260 E10� 030), in September 2008. Theslugs were kept at 15 �C, with a 12:12 h light:dark photoperiod.They were kept in plastic boxes (17 � 22 � 6 cm), with five indi-viduals in each box. The bottom of each box was covered withwet paper towel to ensure high humidity. The slugs were fed withcarrots, lettuce, oatmeal and dog food pellets ad libitum. The boxeswere cleaned and new food added once a week. The collected slugsproduced eggs; some of these were used in the experiment,whereas others were allowed to hatch by incubating eggs at15 �C in petri dishes lined with wet filter paper. The juveniles werekept under the same conditions as the adults.
The slugs and eggs were cold acclimated prior to the experi-ments by keeping them for one week at 10 �C, one week at 5 �C,and lastly 4 weeks at 2 �C.
Five life stages were used in experiments: adults, two differentsize classes of juveniles (juvenile 2 and juvenile 1), newly hatchedjuveniles and eggs. The age, weight and size of the different groupsat the start of the cold acclimation period are shown in Table 1.After cold acclimation slugs and eggs were sorted and only healthylooking specimens were used in the experiments.
Supercooling point, melting point and water content
Supercooling point, melting point of body fluids and the watercontent was determined for cold acclimated specimens. Supercool-ing point was measured using copper-constantan thermocouplesas previously described [20]. The slugs were placed in plastic tubesfitting so they were unable to move, hereby ensuring constantcontact between the thermocouple and the slug. The eggs weresurface dried with filter paper and attached to the thermocoupleby means of vaseline. The temperature of the sample was loweredby 1 �C min�1 from +5 to �10 �C. Adults were cooled more slowly(0.1 �C min�1) because of their larger size.
The body fluid melting point of cold acclimated specimens wasdetermined by measuring the osmolality using a Wescor HR 33 T
Dewpoint Microvoltmeter (Wescor, Logan, Utah) connected toC-52 sample chambers as previously described by Bayley andHolmstrup [4]. To this end, slugs were opened dorsally usingscissors to expose the hemolymph, which was absorbed by filterpaper discs fitting directly into sample chambers. Measurementsof egg fluids were done by opening (using a pair of needles) awhole egg placed in the sample holder. The measured osmolalitieswere transformed to melting points using the osmolal meltingpoint depression constant of �1.86 �C Osmol�1.
The water content of eggs and slugs was determined. Slugs andeggs were blotted with filter paper and their fresh weight deter-mined to the nearest 0.1 mg. After drying for 24 h at 60 �C theirdry weight (dw) was determined and water content calculated asg water g�1 dw.
Survival of freezing and supercooling
The first experiment examined whether A. lusitanicus toleratesubzero temperatures and by which strategy; freeze tolerance orsupercooling. The slugs were placed individually in 160 ml plasticbeakers with lids (4.2 cm high, 7 cm diameter), and the eggs wereplaced in Petri dishes containing 20 eggs each. Both eggs and slugswere placed on wet filter paper to ensure a high humidity. Thecontainers and Petri dishes were placed in a walk-in freezer at�1.3 ± 0.2 �C, and held at this temperature for 24 h. Then somecrushed ice was added to half of the beakers and half of the Petridishes, to induce inoculative freezing. Thus, the ones without icehad the opportunity to supercool. Half of the beakers and Petridishes (equal number with and without ice) were kept at�1.3 ± 0.2 �C for further 3 days. The other half was moved to afreezer cabinet (WTB Binder Labortechnik, Tüttlingen, Germany)preset at �1.5 ± 0.2 �C. The temperature in this freezing cabinetwas then gradually lowered by �0.042 �C/h until the temperaturehad reached �3 ± 0.2 �C. The temperature was held at �3 �C forfurther 24 h. After exposure to sub-zero temperatures it was as-sessed whether the slugs were frozen (by gentle prodding) andthe containers were transferred to a cold room (+2 �C), where theywere allowed to thaw for 24 h before survival of the slugs was as-sessed. The slugs were scored as survived if they had no visibledamages and were able to move normally. The eggs were trans-ferred to 15 �C where they were allowed to hatch and the hatchingpercentage was recorded.
Cryoprotective dehydration
Newly hatched and fully hydrated juvenile slugs were placedindividually in small plastic vials (3 cm high, 1.6 cm diameter) cov-ered with net to prevent slugs from escaping. Each vial was placedin the centre of a 160-ml plastic beakers (4.2 cm high, 7 cm diam-eter) containing either 25 ml demineralised water or crushed ice,sealed with a tightly fitting plastic lid and Parafilm. The beakerscontaining pure water were kept at 2 �C as a control treatment.Beakers containing ice were kept at �3 ± 0.2 �C. The air in thissmall closed system quickly equilibrated with water vapour fromeither the water or the ice. The slugs were kept under these condi-tions for up to 20 days and slugs were sampled over this period.
The water content during the experiment was determined in or-der to evaluate if the slugs became dehydrated. At each measure-ment six slugs were sampled, blotted with filter paper and theirfresh weight determined to nearest 0.1 mg. After drying for 24 hat 60 �C their dry weight (dw) was determined and water contentcalculated as g water g�1 dw. Melting point was determined for sixother juveniles as described.
For assessing survival at the end of the experiment, 30 juvenilesfrom each treatment were scored after rehydration at 2 �C for 24 h.
Table 1The range of age, fresh weight and size of Arion lusitanicus groups used in theexperiments at the start of the cold acclimation period.
Groups Age (days) Fresh weight (mg) Size* (cm)
Adult Not known 8000–17000 –Juvenile size 2 64–66 115–269 2Juvenile size 1 43–45 30–54 1Newly hatched juvenile 2–5 16–26 –Eggs 3–7 28–34 0.3–0.4
* The size is approximate and measured as the length when crawling for slugs, andthe diameter of eggs, respectively.
S. Slotsbo et al. / Cryobiology 62 (2011) 68–73 69

55PhD thesis by Stine Slotsbo
Author's personal copy
Supercooling of eggs
Six eggs were placed in a small plastic vial placed in the centreof a 160-ml plastic beaker (4.2 cm high, 7 cm diameter) containingeither 25 ml demineralised water or crushed ice, sealed with atightly fitting plastic lid and Parafilm. The beakers containingwater were kept at 2 �C whereas those containing ice were keptat either �2 ± 0.2 or �4 ± 0.2 �C. After 20 days of exposure, ninerandomly chosen replicates from each temperature were trans-ferred to 15 �C where they were allowed to hatch and the hatchingpercent was recorded. Another five replicates from �2 ± 0.2 to�4 ± 0.2 �C, respectively, were examined for ice formation, byopening the eggs using preparation needles.
Statistical methods
Temperature and ice effects on hatching success, and tempera-ture and time effects on osmolality and water content were testedusing a two-way ANOVA. Differences in supercooling point, melt-ing point and water content between age groups were tested usinga one-way ANOVA and pair-wise comparisons by Tukey post hoctests. Differences in survival of slugs and their ability to supercoolwere tested by Chi-square tests. T-test was used to test eggs’ abilityto supercool. Data were transformed prior to statistical analysis toimprove normality where necessary. All statistical analyses wereperformed using Sigmaplot for Windows Version 11.0 (SPSS Inc.,Chicago, IL). All results are presented as mean ± SE.
Results
Supercooling point, melting point, water content and estimated icecontent
The average supercooling point of the adult slugs was �3.0 ±0.2 �C which was significantly higher (P < 0.001; Tukey) than thesupercooling point of eggs and juvenile size 1 (Table 2). Themelting point of the slugs rose with rising size/ages of the slugs.The newly hatched juveniles had the lowest melting point at�0.44 �C (Table 2). The eggs and adults had relatively high meltingpoints (Table 2).
There were no significant difference in water content, calculatedas g water g�1 dw, between the age groups of slugs (P = 0.26;one-wayANOVA; Table 2). Thewater content of eggswas lower thanthat of slugs.
The ice content was not measured in this study but wasestimated as percentage of total body water [35], under theassumptions that only 80% of the water can be regarded as osmot-ically active [12]. The estimated ice content in frozen specimens at�1.3 �C was 68%, 63% and 58% for adults, juvenile size 2 andjuvenile size 1, respectively.
Survival of freezing and supercooling
Not all juveniles exposed to �1.3 �C in contact with ice froze asexpected. About 53% of the juvenile size 1, and 80% of the juvenile
size 2, froze. Most of the juveniles and some adult slugs survivedbeing frozen at �1.3 �C for three days, but none of the slugs sur-vived freezing at �3 �C (Table 3). The juveniles survived freezingat -1.3 �C significantly better than the adult slugs (P < 0.001; v2-test).
If freezing of body fluids did not occur, juveniles and adults sur-vived exposures at �3 �C (Table 3). However, of the individuals ex-posed to �3 �C without being in contact with ice, 27% of thejuvenile size 1, 73% of the juvenile size 2 and 85% of the adults,respectively, froze spontaneously. The juvenile size 1 class had asignificantly better ability to stay supercooled at �3 �C than thelarger juvenile size 2 class and the adults (P = 0.003; v2-test).
Eggs survived being exposed to �1.3 �C, and the hatching suc-cess was more than 79%, even for those being in contact with ice(Fig. 1). There was a clear effect of temperature since eggs exposedto �3 �C had significantly lower (Fig. 1) hatching success than eggsexposed to �1.3 �C. Only few eggs (3%) survived exposure to �3 �Cfor 1 day (Fig. 1). There was no significant effect of contact with iceat any of these temperatures (P = 0.087, two-way ANOVA). How-ever, it was in this experiment not possible to determine whichof the eggs froze and which did not, so it is unclear if the eggs ex-posed to ice froze or stayed unfrozen.
Cryoprotective dehydration
Small juveniles were able to stay supercooled for 20 days at�3 �C, and no juveniles froze during this experiment. The meltingpoint of the juveniles did not change significantly (P = 0.441;two-way ANOVA) through the experiment, and there were no sig-nificant difference in melting point (P = 0.119; two-way ANOVA)between the controls and the juveniles kept at �3 �C (Fig. 2).
Table 2Water content, melting point and supercooling point of cold acclimated Arion lusitanicus used in the experiments. Values are mean ± SE.Different letters indicate significant differences.
Groups Water content (g/g DW) Melting point (�C) Supercooling point (�C)
Adults 4.8 ± 0.2 (n = 7)a �0.20 ± 0.02 (n = 6)a �3.0 ± 0.2 (n = 8)a
Juvenile size 2 – �0.28 ± 0.01 (n = 6) bd –Juvenile size 1 4.7 ± 0.1 (n = 7)a �0.35 ± 0.02 (n = 7)b �5.7 ± 0.2 (n = 24)b
Newly hatched juveniles 4.4 ± 0.2 (n = 6)ac �0.44 ± 0.02 (n = 6)c –Eggs 3.9 ± 0.2 (n = 6)bc �0.25 ± 0.02 (n = 6)ad �6.2 ± 0.3 (n = 23)b
Table 3The survival rates of Arion lusitanicus (various size classes) frozen or supercooled ateither �1.3 or �3 �C. Different letters indicate significant difference in survivalbetween age groups within the same treatment.
Treatment Age group Exposed* Survivors Survival (%)
Control, 2 �C Adults 10 10 100Juveniles size 2 15 15 100Juveniles size 1 15 15 100
�1.3 �C, supercooled Adults 14 13 93Juveniles size 2 18 18 100Juveniles size 1 22 22 100
�1.3 �C, frozen Adults 14 3 21a
Juveniles size 2 12 10 83b
Juveniles size 1 8 7 88b
�3 �C, supercooled Adults 2 2 100Juveniles size 2 4 4 100Juveniles size 1 11 10 91
�3 �C, frozen Adults 25 0 0Juveniles size 2 26 0 0Juveniles size 1 19 0 0
* 10–15 Specimens were exposed in each treatment, but those that were unin-tentionally frozen or the opposite (observed by inspection; see text), respectively,were removed and included in the appropriate treatment.
70 S. Slotsbo et al. / Cryobiology 62 (2011) 68–73
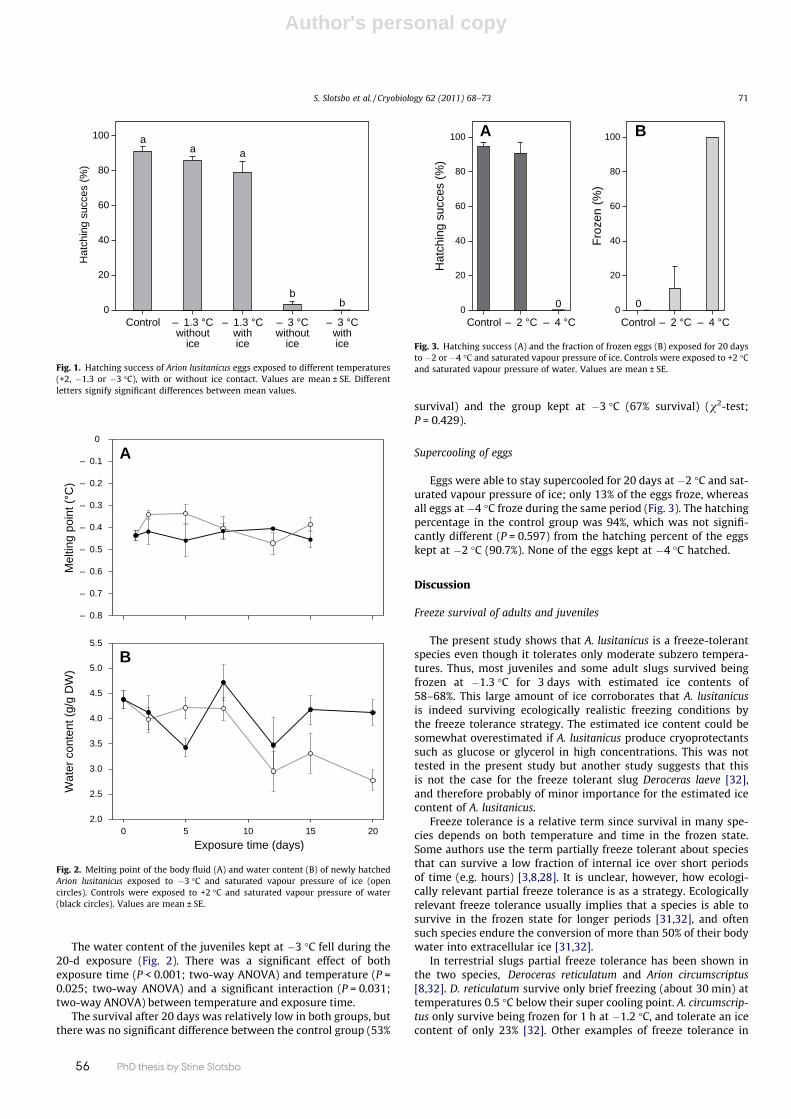
56 PhD thesis by Stine Slotsbo
Author's personal copy
The water content of the juveniles kept at �3 �C fell during the20-d exposure (Fig. 2). There was a significant effect of bothexposure time (P < 0.001; two-way ANOVA) and temperature (P =0.025; two-way ANOVA) and a significant interaction (P = 0.031;two-way ANOVA) between temperature and exposure time.
The survival after 20 days was relatively low in both groups, butthere was no significant difference between the control group (53%
survival) and the group kept at �3 �C (67% survival) (v2-test;P = 0.429).
Supercooling of eggs
Eggs were able to stay supercooled for 20 days at �2 �C and sat-urated vapour pressure of ice; only 13% of the eggs froze, whereasall eggs at �4 �C froze during the same period (Fig. 3). The hatchingpercentage in the control group was 94%, which was not signifi-cantly different (P = 0.597) from the hatching percent of the eggskept at �2 �C (90.7%). None of the eggs kept at �4 �C hatched.
Discussion
Freeze survival of adults and juveniles
The present study shows that A. lusitanicus is a freeze-tolerantspecies even though it tolerates only moderate subzero tempera-tures. Thus, most juveniles and some adult slugs survived beingfrozen at �1.3 �C for 3 days with estimated ice contents of58–68%. This large amount of ice corroborates that A. lusitanicusis indeed surviving ecologically realistic freezing conditions bythe freeze tolerance strategy. The estimated ice content could besomewhat overestimated if A. lusitanicus produce cryoprotectantssuch as glucose or glycerol in high concentrations. This was nottested in the present study but another study suggests that thisis not the case for the freeze tolerant slug Deroceras laeve [32],and therefore probably of minor importance for the estimated icecontent of A. lusitanicus.
Freeze tolerance is a relative term since survival in many spe-cies depends on both temperature and time in the frozen state.Some authors use the term partially freeze tolerant about speciesthat can survive a low fraction of internal ice over short periodsof time (e.g. hours) [3,8,28]. It is unclear, however, how ecologi-cally relevant partial freeze tolerance is as a strategy. Ecologicallyrelevant freeze tolerance usually implies that a species is able tosurvive in the frozen state for longer periods [31,32], and oftensuch species endure the conversion of more than 50% of their bodywater into extracellular ice [31,32].
In terrestrial slugs partial freeze tolerance has been shown inthe two species, Deroceras reticulatum and Arion circumscriptus[8,32]. D. reticulatum survive only brief freezing (about 30 min) attemperatures 0.5 �C below their super cooling point. A. circumscrip-tus only survive being frozen for 1 h at �1.2 �C, and tolerate an icecontent of only 23% [32]. Other examples of freeze tolerance in
Hat
chin
g su
cces
(%)
0
20
40
60
80
100
Control – 1.3 °Cwithout
ice
– 1.3 °Cwithice
– 3 °Cwithout
ice
– 3 °Cwithice
aa a
bb
Fig. 1. Hatching success of Arion lusitanicus eggs exposed to different temperatures(+2, �1.3 or �3 �C), with or without ice contact. Values are mean ± SE. Differentletters signify significant differences between mean values.
Wat
er c
onte
nt (g
/g D
W)
2.0
2.5
3.0
3.5
4.0
4.5
5.0
5.5
0 5 10 15 20Exposure time (days)
Mel
ting
poin
t (°C
)
– 0.8
– 0.7
– 0.6
– 0.5
– 0.4
– 0.3
– 0.2
– 0.1
0
A
B
Fig. 2. Melting point of the body fluid (A) and water content (B) of newly hatchedArion lusitanicus exposed to �3 �C and saturated vapour pressure of ice (opencircles). Controls were exposed to +2 �C and saturated vapour pressure of water(black circles). Values are mean ± SE.
Hat
chin
g su
cces
(%)
0
20
40
60
80
100
Froz
en (%
)
0
20
40
60
80
100
Control – 2 °C – 4 °C Control – 2 °C – 4 °C
00
A B
Fig. 3. Hatching success (A) and the fraction of frozen eggs (B) exposed for 20 daysto �2 or �4 �C and saturated vapour pressure of ice. Controls were exposed to +2 �Cand saturated vapour pressure of water. Values are mean ± SE.
S. Slotsbo et al. / Cryobiology 62 (2011) 68–73 71

57PhD thesis by Stine Slotsbo
Author's personal copy
terrestrial slugs have only been shown in one species, namely D. laeve,which survive �2 �C and an ice content of 65%; but exposure timesabove 1 h were not tested [32].
Supercooling of adults and juveniles
Juvenile and adult A. lusitanicus are generally poor supercoolersunder humid conditions. When not frozen, survival of juvenilesand adults was ensured down to �3 �C, but with a mean SCP of�3 �C, not surprisingly, most of the adults (85%) froze spontane-ously under these conditions, whereas smaller size classes couldstay supercooled. The differences in supercooling ability betweenthe different sizes of slugs can be explained by physical phenom-ena alone. Bigg [5] proposed a simple linear relationship betweennucleation temperature and volume of water (logarithm of waterdroplet diameter), with larger volumes having higher nucleationtemperatures. This relationship has also been shown to be truefor whole organisms [37]. Lee and Costanzo [17] found, in animalsrepresenting Arthropoda and Vertebrata, a positive correlation be-tween supercooling point and body mass, probably because a lar-ger volume of body fluid also will have a higher probability ofcontaining effective ice nucleators [17]. The same relationshiphas also been found in land snails, where smaller snails have lowersupercooling points than large individuals [2].
Influence of size on freeze tolerance
The juveniles survived freezing at �1.3 �C better than the adultslugs. This link between small size and better survival has alsoshown to be true for freeze tolerant earthworms [11]. But in freezetolerant intertidal molluscs the opposite relationship has been ob-served [19]. The amount of latent heat of fusion generated with iceformation is influenced by the body weight. The heat will slowdown the freezing process, and the freezing rate is negatively cor-related with survival in intertidal molluscs [19]. In the presentstudy the melting point was positively correlated with bodyweight of the slugs. The lower melting point in small slugs will re-sult in lower amount of ice at a given temperature, and this mayexplain the better survival of the smaller slugs.
Cryoprotective dehydration
The small juveniles were able to stay supercooled for 20 days at�3 �C and under these conditions they lost a significant amount ofwater (approximately 37% after 20 days). Despite this water loss,the melting point did not change during the experiment and itmust therefore be concluded that small A. lusitanicus do not usecryoprotective dehydration as a cold tolerance strategy.
Maintaining of the melting point (osmolality) despite water lossis contrary to earlier studies of dehydration in slugs. Prior et al. [21]found that 33% water loss in Limax maximus lead to a fall in meltingpoint from �0.26 to �0.41 �C, and Hughes and Kerkut [14] foundthat the melting point of D. reticulatum fell from �0.30 to�0.80 �C when they lost 35% of their body water.
The observation that A. lusitanicus had an unchanged meltingpoint despite a significant drop in water content indicates that theyare able to osmoregulate. If not, the water loss would have causedthe melting point to fall to �0.69 �C instead of staying unchangedat �0.44 �C. The dehydration of A. lusitanicus took place over a per-iod of 20 days, contrary to the studies of L. maximus and D. reticul-atumwhere the slugs were dehydrated over a period of only 2–3 h.This brief desiccation is maybe not sufficient time for sequesteringof salts and hereby osmoregulation, whereas the longer-lastingexposures used in the present study allowed osmoregulation. Thewater content of slugs collected in nature often varies considerably[30] and there is a high variation in an individual’s water content
from day to day [26]; the ability to osmoregulate and hereby avoidhuge fluctuations in the osmolality of the hemolymph could beadvantageous.
Cold tolerance of eggs
The eggs of A. lusitanicus generally have a poor cold toleranceonly surviving temperatures down to �2 �C. The cold toleranceseemed not be influenced by contact with ice but it was not deter-mined whether freezing actually occurred at high subzero temper-ature (�1.3 �C). Under dry conditions most eggs were able to staysupercooled for at least 20 days at �2 �C but at �4 �C all eggs frozeduring this period. Our results therefore suggest that the eggs donot survive by being freeze tolerant, and they are less prone toinoculative freezing than the slugs. The eggs are primarily laid inthe late summer and fall [15] and hatch before winter (Slotsbo,unpublished data). Therefore, it could be argued that winter sur-vival of the population is not to any large degree depending on coldtolerance of eggs, but the ability of eggs to supercool could beimportant during cold spells in the late fall.
Conclusion
A. lusitanicus is shown to be a freeze-tolerant species, however,only in the juvenile and adult stages. The eggs survive relativelyhigh subzero temperatures, probably by supercooling. Juvenilesand adults may also survive in a supercooled state (down to�3 �C) but are generally poor supercoolers. The winter survival ofA. lusitanicus in cold areas depends therefore to a high degree onmigration to habitats protected from low winter temperatures.
Acknowledgments
We are very grateful to Kurt Jordaens for help with correct spe-cies identification of the slugs.
References
[1] A. Ansart, P. Vernon, Cold hardiness in molluscs, Acta Oecologica-InternationalJournal of Ecology 24 (2003) 95–102.
[2] A. Ansart, P. Vernon, Cold hardiness abilities vary with the size of the land snailCornu aspersum, Comparative Biochemistry and Physiology A-Molecular andIntegrative Physiology 139 (2004) 205–211.
[3] A. Ansart, P. Vernon, J. Daguzan, Freezing tolerance versus freezingsusceptibility in the land snail Helix aspersa (Gastropoda: Helicidae),Cryoletters 22 (2001) 183–190.
[4] M. Bayley, M. Holmstrup, Water vapor absorption in arthropods byaccumulation of myoinositol and glucose, Science 285 (1999) 1909–1911.
[5] E.K. Bigg, The supercooling of water, Proceedings of the Physical Society ofLondon Section B 66 (1953) 688–694.
[6] W.C. Blinn, Ecology of the land snails Mesodon thyroidus and Allogona profunda,Ecology 44 (1963) 498–505.
[7] W. Block, Cold Hardiness in invertebrate Poikilotherms, ComparativeBiochemistry and Physiology 73A (1982) 581–593.
[8] R.T. Cook, The tolerance of the field slug Deroceras reticulatum to freezingtemperatures, Cryoletters 25 (2004) 187–194.
[9] H.V. Danks, The roles of insect cocoons in cold conditions, European Journal ofEntomology 101 (2004) 433–437.
[10] M. Holmstrup, M. Bayley, H. Ramlov, Supercool or dehydrate? An experimentalanalysis of overwintering strategies in small permeable arctic invertebrates,Proceedings of the National Academy of Sciences of the United States ofAmerica 99 (2002) 5716–5720.
[11] M. Holmstrup, J. Overgaard, M. Bayley, Small Dendrobaena earthworms survivefreezing better than large worms, Cryobiology 54 (2007) 298–300.
[12] M. Holmstrup, P. Westh, Effects of dehydration on water relations and survivalof lumbricid earthworm egg capsules, Journal of Comparative PhysiologyB-Biochemical Systemic and Environmental Physiology 165 (1995) 377–383.
[13] M. Holmstrup, K.E. Zachariassen, Physiology of cold hardiness in earthworms,Comparative Biochemistry and Physiology A-Physiology 115 (1996) 91–101.
[14] G.M. Hughes, G.A. Kerkut, Electrical activity in a slug ganglion in relation to theconcentration of locke solution, Journal of Experimental Biology 33 (1956)282–294.
[15] J. Kozlowski, Reproduction of Arion Lusitanicus Mabille, 1868 (Gastropoda:Pulmonata: Arionidae) introduced in Poland, FoliaMalacologica 8 (2000) 87–94.
72 S. Slotsbo et al. / Cryobiology 62 (2011) 68–73

58 PhD thesis by Stine Slotsbo
Author's personal copy
[16] J. Kozlowski, The Distribution, biology, population dynamics and harmfulnessof Arion lusitanicus Mabille, 1868 (Gastropoda: Pulmonata: Arionidae) inPoland, Journal of Plant Protection Research 47 (2007) 219–230.
[17] R.E. Lee, J.P. Costanzo, Biological ice nucleation and ice distribution in cold-hardy ectothermic animals, Annual Review of Physiology 60 (1998)55–72.
[18] K. Mellanby, Slugs at low temperatures, Nature 189 (1961) 944.[19] D.J. Murphy, L.C. Johnson, Physical and temporal factors influencing the
freezing tolerance of the marine snail Littorina-littorea (L), Biological Bulletin158 (1980) 220–232.
[20] P.G. Pedersen, M. Holmstrup, Freeze or dehydrate: only two options for thesurvival of subzero temperatures in the arctic enchytraeid Fridericia ratzeli,Journal of Comparative Physiology B-Biochemical Systemic and EnvironmentalPhysiology 173 (2003) 601–609.
[21] D.J. Prior, M. Hume, D. Varga, S.D. Hess, Physiological and behavioral aspects ofwater balance and respiratory function in the terrestrial slug, Limax maximus,Journal of Experimental Biology 104 (1983) 111–127.
[22] J. Quinteiro, J. Rodriguez-Castro, J. Castillejo, J. Iglesias-Pineiro, M. Rey-Mendez,Phylogeny of slug species of the genus Arion: evidence of Iberian endemics andof the existence of relict species in Pyrenean refuges, Journal of ZoologicalSystematics and Evolutionary Research 43 (2005) 139–148.
[23] H. Ramløv, Aspects of natural cold tolerance in ectothermic animals, HumanReproduction 15 (2000) 26–46.
[24] W.A. Riddle, Cold hardiness in the woodland snail, Anguispira alternata (Say)(Endodontidae), Journal of Thermal Biology 6 (1981) 117–120.
[25] W.A. Riddle, V.J. Miller, Cold hardiness in several species of land snails, Journalof Thermal Biology 13 (1988) 163–167.
[26] D.K. Roach, Analysis of the haemolymph of Arion ater L. (Gastropoda:Pulmonata), Journal of Experimental Biology 40 (1963) 613–623.
[27] R.W. Salt, Delayed inoculative freezing of insects, The Canadian Entomologist95 (1963) 1190–1202.
[28] B.J. Sinclair, Insect cold tolerance: how many kinds of frozen?, EuropeanJournal of Entomology 96 (1999) 157–164
[29] L. Sømme, Supercooling and winter survival in terrestrial arthropods,Comparative Biochemistry and Physiology A-Physiology 73 (1982) 519–543.
[30] A. South, Terrestrial Slugs. Biology, Ecology and Control, Chapman and Hall,London, 1992.
[31] K.B. Storey, J.M. Storey, Freeze tolerance in animals, Physiological Reviews 68(1988) 27–84.
[32] K.B. Storey, J.M. Storey, T.A. Churchill, Freezing and anoxia tolerance of slugs: ametabolic perspective, Journal of Comparative Physiology B-BiochemicalSystemic and Environmental Physiology 177 (2007) 833–840.
[33] H. Udaka, H. Numata, Short-day and low-temperature conditions promotereproductive maturation in the terrestrial slug, Lehmannia valentiana,Comparative Biochemistry and Physiology A-Molecular and IntegrativePhysiology 150 (2008) 80–83.
[34] R.C. Weast, Handbook of Chemistry and Physics, CRC Press, Cleveland, 1989.[35] K.E. Zachariassen, Mechanism of the cryoprotective effect of glycerol in beetles
tolerant to freezing, Journal of Insect Physiology 25 (1979) 29–32.[36] K.E. Zachariassen, Physiology of cold tolerance in insects, Physiological
Reviews 65 (1985) 799–832.[37] K.E. Zachariassen, E. Kristiansen, S.A. Pedersen, H.T. Hammel, Ice nucleation in
solutions and freeze-avoiding insects – homogeneous or heterogeneous?,Cryobiology 48 (2004) 309–321
S. Slotsbo et al. / Cryobiology 62 (2011) 68–73 73

59PhD thesis by Stine Slotsbo
Comparative Biochemistry and Physiology, Part A 161 (2012) 443–449, doi:10.1016/j.cbpa.2012.01.002
Stine Slotsboa,b, Lars Monrad Hansena, Kurt Jordaensc, Thierry Backeljauc,d, Anders Malmendale,Niels Chr. Nielsene, Martin Holmstrupb
aDepartment of Agroecology, Aarhus University, Forsøgsvej 1, DK-4200 Slagelse, DenmarkbDepartment of Bioscience, Aarhus University, Vejlsøvej 25, DK-8600 Silkeborg, DenmarkcEvolutionary Ecology Group, Department of Biology, University of Antwerp, Groenenborgerlaan 171, B-2020 Antwerp, BelgiumdRoyal Belgian Institute of Natural Sciences, Vautierstraat 29, B-1000 Brussels, BelgiumeCenter for Insoluble Protein Structures, Interdisciplinary Nanoscience Center and Department of Chemistry, Aarhus University,
Langelandsgade 140, DK-8000 Aarhus, Denmark
PAPER III
Cold tolerance and freeze-induced glucose accumulation in three terrestrial slugs
Representatives of the Arion species: A. rufus (orange), A. ater (white and black) and A. lusitanicus (brown). Photo: Stine Slotsbo.


61PhD thesis by Stine Slotsbo
Cold tolerance and freeze-induced glucose accumulation in three terrestrial slugs
Stine Slotsbo a,b,⁎, Lars Monrad Hansen a, Kurt Jordaens c, Thierry Backeljau c,d, Anders Malmendal e,1,Niels Chr. Nielsen e, Martin Holmstrup b
a Department of Agroecology, Aarhus University, Forsøgsvej 1, DK-4200 Slagelse, Denmarkb Department of Bioscience, Aarhus University, Vejlsøvej 25, DK-8600 Silkeborg, Denmarkc Evolutionary Ecology Group, Department of Biology, University of Antwerp, Groenenborgerlaan 171, B-2020 Antwerp, Belgiumd Royal Belgian Institute of Natural Sciences, Vautierstraat 29, B-1000 Brussels, Belgiume Center for Insoluble Protein Structures, Interdisciplinary Nanoscience Center and Department of Chemistry, Aarhus University, Langelandsgade 140, DK-8000 Aarhus, Denmark
a b s t r a c ta r t i c l e i n f o
Article history:Received 30 September 2011Received in revised form 21 December 2011Accepted 3 January 2012Available online 9 January 2012
Keywords:ArionidaeArion lusitanicusFreeze toleranceMetabolomePulmonata
Cold tolerance and metabolic responses to freezing of three slug species common in Scandinavia (Arion ater,Arion rufus and Arion lusitanicus) are reported. Autumn collected slugs were cold acclimated in the laboratoryand subjected to freezing conditions simulating likely winter temperatures in their habitat. Slugs spontaneouslyfroze at about−4 °Cwhen cooled under dry conditions, but freezing of body fluidswas readily induced at−1 °Cwhen in contact with external ice crystals. All three species survived freezing for 2 days at −1 °C, and some A.rufus and A. lusitanicus also survived freezing at −2 °C. 1H NMR spectroscopy revealed that freezing of bodyfluids resulted in accumulation of lactate, succinate and glucose. Accumulation of lactate and succinate indicatesthat ATP production occurred via fermentative pathways, which is likely a result of oxygen depletion in frozentissues. Glucose increased from about 6 to 22 μg/mg dry tissue upon freezing in A. rufus, but less so in A. aterand A. lusitanicus. Glucose may thus act as a cryoprotectant in these slugs, although the concentrations are notas high as reported for other freeze tolerant invertebrates.
© 2012 Elsevier Inc. All rights reserved.
1. Introduction
Cold tolerance of terrestrial invertebrates in temperate and coldregions has been widely studied, as these organisms often facetemperatures during winter that may cause freezing of their bodyfluids. Two major protective physiological strategies are found: freezetolerant species seek to establish controlled, protective freezing of theextracellular body fluids at high sub-zero temperatures, whereasfreeze avoiding species have developed mechanisms to stay unfrozeneven at temperatures much below the melting point of body fluids(Sømme, 1982; Zachariassen, 1985; Storey and Storey, 1996). Awide range of invertebrates, especially arthropods, have been thefocus of this research, but cold tolerance of terrestrial molluscs hasreceived less attention (Ansart and Vernon, 2003), and only fewstudies are concerned with terrestrial slugs (Mellanby, 1961; Cook,2004; Storey et al., 2007; Udaka et al., 2008; Berman et al., 2011;Slotsbo et al., 2011). Both freeze tolerance and freeze avoidancestrategies occur within terrestrial Mollusca. However, in contrast toland snails, slugs lack physical protection such as shell, epiphragm
and operculum. The lack of this protection likely makes themsusceptible to inoculative freezing and, therefore, adoption of a freezeavoidance strategy seems inappropriate. Hence, their winter survivalmay depend on the ability to find frost-free hibernation sites or theiruse of freeze tolerance as a strategy. Recent studies have shown thatsome terrestrial slugs are, indeed, freeze tolerant (Berman et al.,2011; Slotsbo et al., 2011).
Freeze tolerant animals undergoing freezing of their body fluids facethe problem of dehydration of cells; therefore, they requirephysiological and biochemical mechanisms that protect them againstdehydration injuries (Storey, 1997). A well-known protectionmechanism is the accumulation of compatible osmolytes, such as sugarsand polyols. These osmolytes (cryoprotectants) act colligatively andprotect cells against intracellular dehydration. Furthermore, they canhelp stabilize membranes and proteins when the body fluids freeze(Crowe et al., 1987).
The aim of the present study was to compare the cold tolerance ofthree slug species common in Scandinavia: Arion ater L, 1758, A. rufusL, 1758 and A. lusitanicusMabille, 1868. The species A. ater is native toScandinavia and A. rufus is native to central Europe, but has beenintroduced to Scandinavia. Neither A. ater nor A. rufus seem toconstitute a pest problem in Scandinavia, whereas A. lusitanicus isan invasive species that often occurs in high numbers and is regardeda serious pest in most of Europe (Kozlowski, 2007). The three speciesare easily confused (Cameron et al., 1983; Noble, 1992), and identifi-cation of these species is complex, where using external morphology
Comparative Biochemistry and Physiology, Part A 161 (2012) 443–449
⁎ Corresponding author at: Department of Bioscience, Aarhus University, Vejlsøvej25, DK-8600 Silkeborg, Denmark. Tel.: +45 87 15 88 13; fax: +45 87 15 89 01.
E-mail address: [email protected] (S. Slotsbo).1 Present address: Department of Biomedical Sciences, Faculty of Health Sciences,
University of Copenhagen, Blegdamsvej 3, DK-2200 Copenhagen N, Denmark.
1095-6433/$ – see front matter © 2012 Elsevier Inc. All rights reserved.doi:10.1016/j.cbpa.2012.01.002
Contents lists available at SciVerse ScienceDirect
Comparative Biochemistry and Physiology, Part A
j ourna l homepage: www.e lsev ie r .com/ locate /cbpa

62 PhD thesis by Stine Slotsbo
is almost impossible. The reasons for the different degree of success ofthese species are unknown.
Specimens were collected during the autumn months, coldacclimated in the laboratory and subjected to freezing conditionssimulating likely winter temperatures in their habitat. Slugs werescreened for the accumulation of cryoprotectants, and survival of bodyfluid freezing was tested.
2. Materials and methods
2.1. Test animals
Specimens of A. lusitanicus, A. rufus and A. ater were collected in2009 and 2010 (September–October) in eastern parts of Jutland,Denmark. The slugs were kept at 5 °C, with a 12:12 h light:darkphotoperiod, until onset of experimentation. They were kept inplastic boxes (17×22×6 cm), with five individuals in each box. Thebottom of each box was covered with wet paper towel to ensurehigh humidity. The slugs were fed carrots, lettuce, oatmeal and dogfood pellets ad libitum. Once a week, the boxes were cleaned andnew food was added.
The slugs were cold acclimated prior to the experiments bykeeping them for at least 1 week at 5 °C, and then for 4 weeks at2 °C, to simulate early winter temperatures and to induce freezetolerance (Slotsbo et al., 2011). The slugs were active and eating at2 °C, but eating was limited compared to higher temperatures. Aftercold acclimation, slugs were sorted and only healthy lookingspecimens were used in cold tolerance experiments and metabolicprofiling. The fresh masses of the slugs in 2009 were as follows; A.lusitanicus: 1.0–19.4 g (mean±SE; 7.4±0.7 g; N=40); A. rufus:0.8–43.2 g (mean±SE; 9.0±2.4 g; N=21) and A. ater: 1.5–26.9 g(mean±SE; 9.3±1.3 g; N=40). In 2010, the fresh masses were asfollows; A. lusitanicus: 2.7–6.0 g (mean±SE; 4.2±0.2 g; N=32); A.rufus: 2.1–19.3 g (mean±SE; 10.2±1.3 g; N=15) and A. ater: 1.0–25.7 g (mean±SE; 4.1±0.8 g; N=40).
2.2. Species identification
The three species are easily confused (Cameron et al., 1983; Noble,1992), and identification of the species using external morphology isdifficult. However, species can be identified by their genitalmorphologycombined with sequence data of mitochondrial 16S rRNA (Barr et al.,2009; Jordaens et al., unpublished results). The individuals in thisstudy were identified to species by oviduct morphology and theposition of the ligula (e.g. Kerney et al., 1979; Noble, 1992). In addition,and because the genitalia of some specimens were too small to allowaccurate identification, we also identified a number of representativespecimens of each species (10–20 individuals) using DNA sequencesof a 410 bp fragment of the mitochondrial 16S rRNA. The speciesidentification was conducted after the experiment. To this end, a pieceof the foot was digested for 4 h in 500 μL of CTAB buffer (2% CTAB,1.4 MNaCl, 20 mMEDTA and 100 mMTris/HCl)with 0.1 mg proteinaseK at 56 °C. Total genomicDNAwas extracted using theQIAmpDNAminikit (Qiagen), resuspended in 200 μL of sterile water and stored at−20 °C. The 16S rRNA gene was amplified using the primer pair16SAR (CGCCTGTTTATCAAAAACAT) and 16SBR (CCGGTCTGAACTCA-GATCACGT) (Palumbi, 1996). The PCR reaction buffer contained1.5 mMMgCl2, 0.2 mM dNTPs, 0.2 μM of each primer and nine units ofTaq (Promega, Inc.). The PCR profile was as follows: 5 min at 95 °C, 30cycles of 1 min at 95 °C, 1 min at 46 °C and 2 min at 72 °C and a finalextension of 5 min at 72 °C. PCR products were purified using the GFXPCR DNA Purification Kit (Amersham Biosciences) and diluted in100 μL of sterile water. Purified PCR products were sequenced usingthe ABI PRISM BigDye Terminator Cycle Sequencing Kit and run on acapillary Applied Biosystems 3730 DNA Analyser.
Sequenceswere aligned using ClustalX version 1.81with default gapopening/gap extension costs of 15/6.66 (Thompson et al., 1997). ANeighbor-Joining (NJ) tree (Saitou and Nei, 1987) was constructed onthe basis of Kimura 2-Parameter distances calculated with completedeletion of sites with alignment gaps in MEGA version 4.0.2 (Tamuraet al., 2007). jModeltest version 0.1.1 (Posada, 2008) was used to selectthemost appropriate substitutionmodel forMaximumParsimony (MP)and Maximum Likelihood (ML) analysis, which was the TIM2+Gmodel. MP and ML trees were made in PAUP* version 4.0b10(Swofford, 2002) using a heuristic search with the tree-bisection–reconnection branch-swapping algorithm and random addition oftaxa. Relative branch support was evaluated with 1000 bootstrap repli-cates (Felsenstein, 1985) for the NJ and MP trees, and 200 for the MLtree. In all analyses we included several sequences of the three species,and of A. flagellus that could be confused with especially A. lusitanicus,that were available from GenBank, viz. AY947349 and EU541904–EU541907 for A. ater, FJ896811, AY947376–AY947378 and EU541919–EU541927 for A. rufus, AY947369–AY947370 and EU541909–EU541915 for A. lusitanicus and AY947359–AY947361 and EU541929–EU541931 for A. flagellus. Phylogenetic relationships were inferredwith Arion hortensis (AY947364) as outgroup.
2.3. Melting point and temperature of crystallization (Tc)
The temperature of crystallization (Tc) of the slugs andmelting pointof their hemolymph were determined for cold acclimated specimens.The Tc was measured using copper–constantan thermocouples asdescribed earlier (Pedersen and Holmstrup, 2003). The slugs wereplaced in plastic tubes so that they were unable to move, herebyensuring constant contact between the thermocouple and the slug.The temperature of the plastic tubes was lowered by 0.1 °C min−1
from +5 to −10 °C, and the Tc was taken as the lowest temperaturereading before the occurrence of the freezing exotherm.
The hemolymph melting point was determined by measuring theosmolality of 5–6 specimens of each species. This was done using aWescor HR 33T Dewpoint Microvoltmeter (Wescor, Logan, Utah)connected to C-52 sample chambers as previously described byHolmstrup et al. (1999). Slugs were opened dorsally using scissorsto expose the hemolymph, which was absorbed by filter paper discsfitting directly into sample chambers. The measured osmolalitieswere transformed to melting points using the osmolal melting pointdepression constant of −1.86 °C Osmol−1.
2.4. Survival of freezing
Freeze tolerance was tested in two similar experiments; oneperformed in November 2009, and one in November 2010. Slugswere placed individually in 160 mL plastic beakers with lid (4.2 cmhigh, 7 cm diameter), of which the bottom was covered with wetfilter paper to ensure a high humidity. The beakers were placed in awalk-in freezer at approximately −1 °C (for exact temperature seeFig. 1). To induce freezing, a piece of crushed ice was added to eachbeaker after 6 h. The beakers were kept at this temperature for2 days. It was assessed whether all slugs were frozen by gentle prod-ding. In the 2009 experiment, randomly chosen frozen and controlslugs (+2 °C) were sampled for metabolic profiling at this point inthe protocol. The sampled slugs were transferred to liquid nitrogenand stored at −80 °C until analysis. The remaining slugs weretransferred to a cold room (+2 °C), where they were allowed tothaw for 24 h before survival was assessed. Slugs were scored assurvivors if they had no visible damages and were able to movenormally.
To test if the slugs tolerated freezing at temperatures below−1 °C, allthe survivorsweremoved to thewalk-in freezer at approximately−1 °Cand crushed ice was added after 6 h. The beakers were kept at thistemperature for 3 days prior to lowering the temperature to
444 S. Slotsbo et al. / Comparative Biochemistry and Physiology, Part A 161 (2012) 443–449

63PhD thesis by Stine Slotsbo
approximately −2 °C (Fig. 1), and were held at this temperature for2 days. It was assessedwhether all slugswere frozen by gentle prodding.The slugs were thawed and their survival scored as described above. Theslugs were kept at +2 °C, and after another 48 h survival was scoredagain. Differences in survival rates between species were assessedusing a Chi-square test.
2.5. Metabolic profiling
Nuclear magnetic resonance spectroscopy (1H NMR) was used toscreen for metabolites (e.g. sugars, polyols and free amino acids).The slugs were freeze dried for 24 h and crushed to powder. A sampleof 23–50 mg of the powder was used for analysis. To extract water-soluble metabolites, the sample was homogenized in 500 μL cold50% acetonitrile using a TissueLyser (Qiagen, Copenhagen, Denmark).Samples were then centrifuged at 10,000 g for 10 min and thesupernatant was stored at −80 °C until 1H NMR analysis. Thesupernatant was transferred to new tubes, lyophilized and storedagain at −80°. Immediately before the NMR measurements, sampleswere rehydrated in 650 μL of 50 mM phosphate buffer (pH 7.4) inD2O, and 600 μL of the solution was transferred to a 5 mm NMRtube. The buffer contained 50 mg/L of the chemical shift reference3-(trimethylsilyl)-propionic acid-D4, sodium salt (TSP) and 100 mg/L NaN3.
The NMR measurements were performed at 25 °C on a BrukerAvance-2 500 spectrometer (Bruker Biospin) operating at a 1Hfrequency of 500.13 MHz and equipped with a 5 mm HCN tripleresonance probe. 1H NMR spectra were acquired using a single-90°-pulse experiment with a Carr–Purcell–Meiboom–Gill (CPMG) delayadded in order to attenuate broad signals from high-molecular-weight components. The total CPMG delay was 40 ms and the spin-echo delay was 200 μs. The water signal was suppressed by presatura-tion of the water peak during the relaxation delay of 1 s. A total of 256transients of 8 K data points spanning a spectral width of 24 ppmwere collected, corresponding to a total experiment time of 10 min.For assignment purposes, a 2-dimensional 1H-1H TOCSY(Braunschweiler and Ernst, 1983; Bax and Davis, 1985) spectrumwith 80 ms mixing was acquired.
Metabolite signals were identified from a two-dimensional 1H-1HTOCSY spectrum, STOCSY spectra (Cloarec et al., 2005) and fromcomparison with known metabolite chemical shifts (Fan, 1996;Lindon et al., 1999; Malmendal et al., 2006; Overgaard et al., 2007a).
NMR data were scaled using VAST scaling (Keun et al., 2003) toobtain unit variance within each selection regime (i.e. data for eachbin was divided by the average standard deviation of the valueswithin the selection regimes) and then centered. This scaling reducesthe weight of variations that are not related to the treatments, i.e.
random variations between “identical” samples are reduced, and theanalysis is not biased towards metabolites present in high concentra-tions. Principal component analysis (PCA) was carried out on themetabolite data. PCA was performed using in house R-scripts (RDevelopment Core Team 2003) and the number of significant PCswas assessed by cross validation (Wold, 1978). Scores from the PCAwere subjected to two-way MANOVA to test for effects of speciesand freezing and the interaction between both.
Significant spectral differences were measured between unfrozenand frozen slugs. Bonferroni correction was applied for multiple tests(significance level 0.05). Since the number of detected metabolitesusing NMR is typically in the order of 100 we used an adjusted P-value of 0.0005.
2.6. Glucose quantification
Glucose analysis was carried out as described by Overgaard et al.(2007b) using a spectrophotometrically based enzymatic test kit(Glucose Gluc-DH, Diagnostic systems GmbH, Holzheim, Germany).Prior to statistical analysis, data was log transformed to improvenormality. A two-way ANOVA was used to test for treatment andspecies effects and interaction between the two factors. Pairwisecomparisons were made using a Tukey post hoc test.
3. Results
3.1. Species identification
The genetic analysis based on 16S rRNA confirmed that thespecimens used in the experiments corresponded to the species A.ater, A. rufus and A. lusitanicus, respectively, and that none of thespecimens clustered with or within A. flagellus. Even though A. rufuswas not monophyletic (Fig. 2) the specimens with an A. rufus oviductmorphology grouped well within one of the A. rufus clades arisingfrom the 16S rRNA results. Also for A. ater and A. lusitanicus, theidentification based on the morphology of the reproductive organs(whenever the reproductive organs were fully developed) wasconfirmed by genetic analysis.
3.2. Melting point and temperature of crystallization (Tc)
The melting point of slug hemolymph was about −0.25 °C, and Tcwas about −4 °C (Table 1). There was no significant differences inmelting point (ANOVA; P=0.475) or in Tc (ANOVA; P=0.544)between the species. The mean fresh weights of the specimens usedfor the Tc measurements were not significantly different (ANOVA;P=0.140).
3.3. Survival of freezing
The survival rate was between 95% and 100% for all three specieswhen frozen at −1 °C for 2 days, and there was no species differencein survival at this treatment (Fig. 3). In the first freeze toleranceexperiment (performed in 2009), the survival rate after the −2 °Ctreatment was 73, 25 and 0% for A. rufus, A. lusitanicus and A. ater,respectively (Fig. 3). The survival rate in this year differed significantlybetween the three species (Chi-square; Pb0.001). In the second freezetolerance experiment (performed in 2010), the temperature decreasedmore rapidly to −2 °C (Fig. 1). None of the individuals survived thistreatment (Fig. 3).
3.4. Metabolic profiling
A typical NMR spectrum of A. lusitanicus is shown in Fig. 4A. Therelative similarities between the metabolomes of the different species(cold acclimated and frozen specimens) were characterized by
90 1 2 3 4 5 6 7 8
Days
Tem
pera
ture
(°C
)
–2
–1
0
1
2 20092010
Fig. 1. The exposure temperatures during freezing experiments with slugs (grey line:2009; black line: 2010). Arrows indicate the time where ice crystals were added to in-duce freezing of slugs.
445S. Slotsbo et al. / Comparative Biochemistry and Physiology, Part A 161 (2012) 443–449

64 PhD thesis by Stine Slotsbo
principal component analysis (PCA). The seven significant principalcomponents used in this analysis explained 78% of the variationbetween the samples. MANOVA showed that the metabolomes
differed significantly between species and between unfrozen andfrozen specimens (Table 2). There was, however, no significantinteraction between the effects of freezing and species, meaning thatthe response to freezing of each species was similar when consideringthe total metabolome (Table 2).
To identify differences in metabolite levels due to freezing (i.e.comparing unfrozen and frozen individuals), spectral differenceswere measured between control and frozen slugs. Three metabolitesresponded significantly to freezing (Bonferroni; Pb0.05). Theresponse included increases in glucose (signals at 4.63, 3.91, 3.46and 3.23 ppm), lactate (4.09 and 1.32 ppm) and succinate concentra-tions (2.39 ppm) (Fig. 4B–D).
Glucose was quantified in greater detail, showing that concentra-tions of cold acclimated slugs ranged from 5 to 7 μg mg−1 dry mass(dw) in all three species (Fig. 5). The glucose concentration increasedsignificantly (ANOVA; Pb0.001) in response to freezing in A. rufus(21.6 μg/mg dw) and A. lusitanicus (13.4 μg/mg dw), but the increase
Fig. 2. Phylogenetic tree of 16S rDNA haplotypes of large arionids and an outgroup haplotype of Arion hortensis. Individuals used in this study are labeled as ‘Denmark’ followed by num-bers. The numbers 3–19 indicate individuals identified by external morphological characters as A. rufus. The numbers 22–60 are individuals identified as A. ater. The numbers 64–101 areindividuals identified as A. lusitanicus. Individuals that were also identified using the proximal genitalia are indicated with an asterisk. Values for node support are: bootstrap value for NJtree based on the Kimura-2-Parameter model (1000 replicates) / bootstrap value based on the MP tree (1000 replicates)/bootstrap value for the ML tree (200 pseudo-replicates). Onlybootstrap values≥70 are shown. Branch lengths are according to the NJ tree. When the node was not supported by one of the analyses, this is indicated with “-”.
Table 1The melting point of hemolymph and temperature of crystallization (Tc) of Arion rufus,A. ater and A. lusitanicus acclimated at 2 °C. Values are mean±SE. No significantdifferences between species were observed.
Species Melting point(°C)
Temperature of crystallization(Tc) (°C)
Fresh massa
(g)
Arionlusitanicus
−0.20±0.02 (N=6) −3.8±0.3 (N=12) 1.8±0.3
Arion rufus −0.24±0.02 (N=5) −4.1±0.4 (N=8) 1.8±0.6Arion ater −0.23±0.03 (N=5) −4.2±0.3 (N=8) 2.6±0.3
a Average fresh mass of the specimens used for Tc measurements.
446 S. Slotsbo et al. / Comparative Biochemistry and Physiology, Part A 161 (2012) 443–449
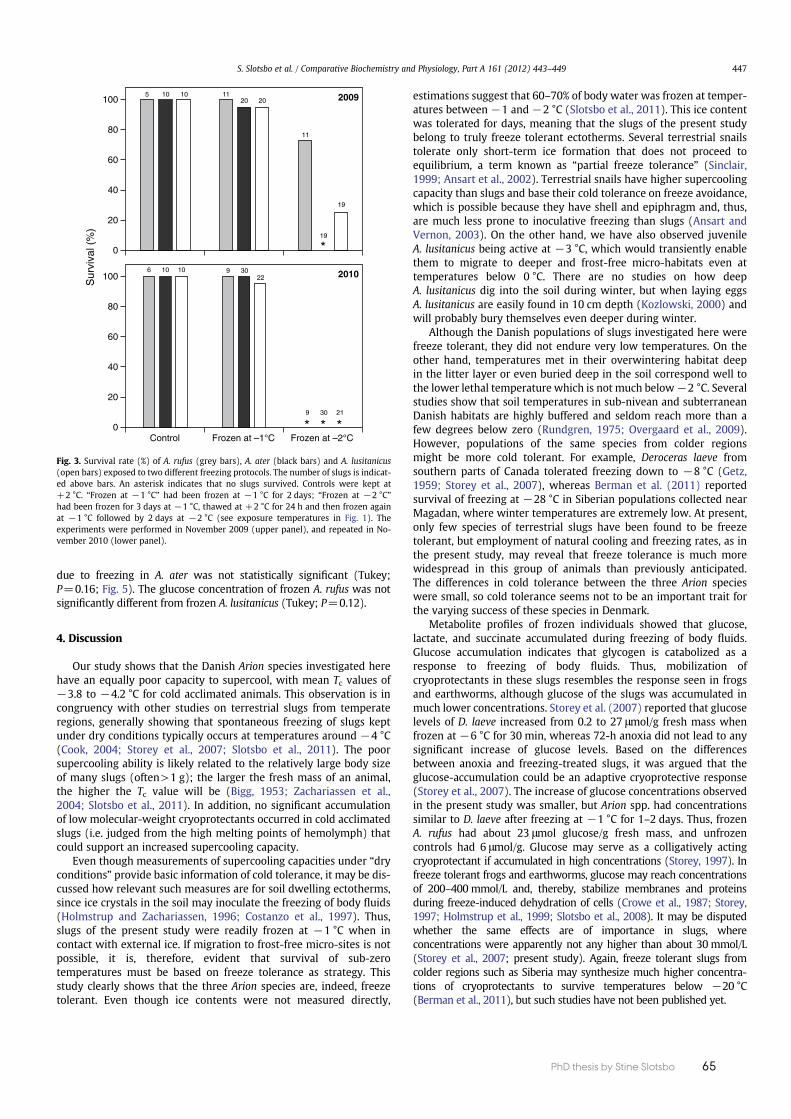
65PhD thesis by Stine Slotsbo
due to freezing in A. ater was not statistically significant (Tukey;P=0.16; Fig. 5). The glucose concentration of frozen A. rufus was notsignificantly different from frozen A. lusitanicus (Tukey; P=0.12).
4. Discussion
Our study shows that the Danish Arion species investigated herehave an equally poor capacity to supercool, with mean Tc values of−3.8 to −4.2 °C for cold acclimated animals. This observation is incongruency with other studies on terrestrial slugs from temperateregions, generally showing that spontaneous freezing of slugs keptunder dry conditions typically occurs at temperatures around −4 °C(Cook, 2004; Storey et al., 2007; Slotsbo et al., 2011). The poorsupercooling ability is likely related to the relatively large body sizeof many slugs (often>1 g); the larger the fresh mass of an animal,the higher the Tc value will be (Bigg, 1953; Zachariassen et al.,2004; Slotsbo et al., 2011). In addition, no significant accumulationof low molecular-weight cryoprotectants occurred in cold acclimatedslugs (i.e. judged from the high melting points of hemolymph) thatcould support an increased supercooling capacity.
Even though measurements of supercooling capacities under “dryconditions” provide basic information of cold tolerance, it may be dis-cussed how relevant such measures are for soil dwelling ectotherms,since ice crystals in the soil may inoculate the freezing of body fluids(Holmstrup and Zachariassen, 1996; Costanzo et al., 1997). Thus,slugs of the present study were readily frozen at −1 °C when incontact with external ice. If migration to frost-free micro-sites is notpossible, it is, therefore, evident that survival of sub-zerotemperatures must be based on freeze tolerance as strategy. Thisstudy clearly shows that the three Arion species are, indeed, freezetolerant. Even though ice contents were not measured directly,
estimations suggest that 60–70% of body water was frozen at temper-atures between −1 and −2 °C (Slotsbo et al., 2011). This ice contentwas tolerated for days, meaning that the slugs of the present studybelong to truly freeze tolerant ectotherms. Several terrestrial snailstolerate only short-term ice formation that does not proceed toequilibrium, a term known as “partial freeze tolerance” (Sinclair,1999; Ansart et al., 2002). Terrestrial snails have higher supercoolingcapacity than slugs and base their cold tolerance on freeze avoidance,which is possible because they have shell and epiphragm and, thus,are much less prone to inoculative freezing than slugs (Ansart andVernon, 2003). On the other hand, we have also observed juvenileA. lusitanicus being active at −3 °C, which would transiently enablethem to migrate to deeper and frost-free micro-habitats even attemperatures below 0 °C. There are no studies on how deepA. lusitanicus dig into the soil during winter, but when laying eggsA. lusitanicus are easily found in 10 cm depth (Kozlowski, 2000) andwill probably bury themselves even deeper during winter.
Although the Danish populations of slugs investigated here werefreeze tolerant, they did not endure very low temperatures. On theother hand, temperatures met in their overwintering habitat deepin the litter layer or even buried deep in the soil correspond well tothe lower lethal temperature which is not much below−2 °C. Severalstudies show that soil temperatures in sub-nivean and subterraneanDanish habitats are highly buffered and seldom reach more than afew degrees below zero (Rundgren, 1975; Overgaard et al., 2009).However, populations of the same species from colder regionsmight be more cold tolerant. For example, Deroceras laeve fromsouthern parts of Canada tolerated freezing down to −8 °C (Getz,1959; Storey et al., 2007), whereas Berman et al. (2011) reportedsurvival of freezing at −28 °C in Siberian populations collected nearMagadan, where winter temperatures are extremely low. At present,only few species of terrestrial slugs have been found to be freezetolerant, but employment of natural cooling and freezing rates, as inthe present study, may reveal that freeze tolerance is much morewidespread in this group of animals than previously anticipated.The differences in cold tolerance between the three Arion specieswere small, so cold tolerance seems not to be an important trait forthe varying success of these species in Denmark.
Metabolite profiles of frozen individuals showed that glucose,lactate, and succinate accumulated during freezing of body fluids.Glucose accumulation indicates that glycogen is catabolized as aresponse to freezing of body fluids. Thus, mobilization ofcryoprotectants in these slugs resembles the response seen in frogsand earthworms, although glucose of the slugs was accumulated inmuch lower concentrations. Storey et al. (2007) reported that glucoselevels of D. laeve increased from 0.2 to 27 μmol/g fresh mass whenfrozen at −6 °C for 30 min, whereas 72-h anoxia did not lead to anysignificant increase of glucose levels. Based on the differencesbetween anoxia and freezing-treated slugs, it was argued that theglucose-accumulation could be an adaptive cryoprotective response(Storey et al., 2007). The increase of glucose concentrations observedin the present study was smaller, but Arion spp. had concentrationssimilar to D. laeve after freezing at −1 °C for 1–2 days. Thus, frozenA. rufus had about 23 μmol glucose/g fresh mass, and unfrozencontrols had 6 μmol/g. Glucose may serve as a colligatively actingcryoprotectant if accumulated in high concentrations (Storey, 1997). Infreeze tolerant frogs and earthworms, glucose may reach concentrationsof 200–400 mmol/L and, thereby, stabilize membranes and proteinsduring freeze-induced dehydration of cells (Crowe et al., 1987; Storey,1997; Holmstrup et al., 1999; Slotsbo et al., 2008). It may be disputedwhether the same effects are of importance in slugs, whereconcentrations were apparently not any higher than about 30 mmol/L(Storey et al., 2007; present study). Again, freeze tolerant slugs fromcolder regions such as Siberia may synthesize much higher concentra-tions of cryoprotectants to survive temperatures below −20 °C(Berman et al., 2011), but such studies have not been published yet.
Control Frozen at –1°C Frozen at –2°C
Sur
viva
l (%
)
20
0
40
60
80
100
20
0
40
60
80
100
2009
20103022
9 30 21
10
6 10 10 9
5 10 1120 20
11
19
19
Fig. 3. Survival rate (%) of A. rufus (grey bars), A. ater (black bars) and A. lusitanicus(open bars) exposed to two different freezing protocols. The number of slugs is indicat-ed above bars. An asterisk indicates that no slugs survived. Controls were kept at+2 °C. “Frozen at −1 °C” had been frozen at −1 °C for 2 days; “Frozen at −2 °C”had been frozen for 3 days at −1 °C, thawed at +2 °C for 24 h and then frozen againat −1 °C followed by 2 days at −2 °C (see exposure temperatures in Fig. 1). Theexperiments were performed in November 2009 (upper panel), and repeated in No-vember 2010 (lower panel).
447S. Slotsbo et al. / Comparative Biochemistry and Physiology, Part A 161 (2012) 443–449

66 PhD thesis by Stine Slotsbo
Glucose may also serve as fuel for anaerobic metabolism in frozentissues (Storey and Storey, 1990; Calderon et al., 2009). Freezetolerance is often linked to a great anoxia tolerance, since freezinghalts the circulation of blood or hemolymph and, thereby, circulationof oxygen (Storey and Storey, 1996). Therefore, survival in the frozenstate depends on ATP production via fermentative pathways. Themost common fermentation pathway in animal tissues is that ofanaerobic glycolysis. A common end product of this pathway islactate, but other end products such as succinate and alanine areknown (Hochachka and Somero, 2002). The accumulation of lactateand succinate (although not quantified directly in the present study)in frozen tissues shows that anaerobic metabolism occurred in allthree Arion species, as has also been reported for other terrestrialslugs in the frozen state (Storey et al., 2007).
Arion lusitanicus is an invasive species, which is considered as aserious pest in several European countries, including Denmark,because it sometimes occurs in very high numbers (Kozlowski,
2007). It has only been found in Denmark very recently (1991) (vonProschwitz and Winge, 1994), whereas A. rufus was introduced fromGermany about a century ago and A. ater is native to Denmark(Engelke et al., 2011). The reason for the mass occurrences andconsequent pest status of A. lusitanicus is unrevealed, but anecdotalreports flourish in public media stating that the success of the speciesis related to its superior resistance to natural stresses such as droughtand low winter temperatures, and that it in this way may outcompete
012345678910
A
B
chemical shift (ppm)
C
D
glucose lactate
012345678910
012345678910
012345678910
succinate
Fig. 4. NMR spectra and spectral differences of water soluble metabolites extracted from slug tissues by acetonitrile/water (50/50). (A) Typical NMR spectrum of Arion lusitanicus.(B–D) Mean spectral differences between unfrozen and frozen A. lusitanicus (B; N=10) A. ater (C; N=10) and A. rufus (D; N=5) (positive signals (i.e., going up) have higherintensity in frozen than in unfrozen slugs). In (B–D) signals which are significantly different when comparing unfrozen and frozen spectra for all species together are highlightedin black (Pb0.05, Bonferroni). Intensities are scaled as I/√|I| where I is the intensity. The vertical scale is identical in (B–D).
Table 2Two-way MANOVA for freezing and species on the PCA scores of metabolite dataobtained by 1H NMR spectroscopya.
Effect Significance (P-value)
Freezing 1.9×10−12
Species 6.4×10−7
Freezing×Species 0.16
a The scores from all seven principal components were included.
a
a
a
b
a
a
A. rufus A. ater A. lusitanicus
µg g
luco
se m
g-1 d
ry w
eigh
t
0
5
10
15
20
25
30
Fig. 5. Glucose concentration (mean±SE) of Arion rufus (N=5), A. ater (N=10) and A.lusitanicus (N=10) acclimated at 2 °C (black bars) or frozen at −1 °C for 2 days (greybars). Different letters indicate significant differences between species within a giventreatment. An asterisk indicates significant differences between treatments within aspecies.
448 S. Slotsbo et al. / Comparative Biochemistry and Physiology, Part A 161 (2012) 443–449

67PhD thesis by Stine Slotsbo
other closely related Arion species. Our study suggests that there areonly subtle differences in cold tolerance of the species investigatedhere, if any at all.
Acknowledgements
Dr. A. P. S. Svane assisted in the NMR-based identification ofmetabolites. We thank N. Van Houtte for her help in the molecularidentification of the slugs.
References
Ansart, A., Vernon, P., Daguzan, J., 2002. Effects of a freezing event during hibernationon further survival, reproduction and growth in the partially freezing tolerantland snail Helix aspersa Muller (Gastropoda: Helicidae). Cryo Lett. 23, 269–274.
Ansart, A., Vernon, P., 2003. Cold hardiness in molluscs. Acta Oecol. 24, 95–102.Barr, N.B., Cook, A., Elder, P.,Molongoski, J., Prasher, D., Robinson, D.G., 2009. Application of
a DNA barcode using the 16S rRNA gene to diagnose pest Arion species in the USA. J.Moll. Stud. 75, 187–191.
Bax, A., Davis, D.G., 1985. Mlev-17-based two-dimensional homonuclear magnetizationtransfer spectroscopy. J. Magn. Reson. 65, 355–360.
Berman, D.I., Meshcheryakova, E.N., Leirikh, A.N., 2011. Cold hardiness, adaptive strategiesand invasion of slugs of the genusDeroceras (Gastropoda, Pulmonata) inNortheasternAsia. Zool. Zh. 90, 387–401.
Bigg, E.K., 1953. The supercooling of water. Proc. Phys. Soc. Lond. B 66, 688–694.Braunschweiler, L., Ernst, R.R., 1983. Coherence transfer by isotropic mixing—application
to proton correlation spectroscopy. J. Magn. Reson. 53, 521–528.Calderon, S., Holmstrup, M., Westh, P., Overgaard, J., 2009. Dual roles of glucose in the
freeze-tolerant earthworm Dendrobaena octaedra: cryoprotection and fuel formetabolism. J. Exp. Biol. 212, 859–866.
Cameron, R.A.D., Eversham, B., Jackson, N., 1983. A field key to the slugs of the BritishIsles. Field Stud. 5, 807–824.
Cloarec, O., Dumas,M.E., Craig, A., Barton, R.H., Trygg, J., Hudson, J., Blancher, C., Gauguier, D.,Lindon, J.C., Holmes, E., Nicholson, J., 2005. Statistical total correlation spectroscopy: anexploratory approach for latent biomarker identification frommetabolic 1H NMR datasets. Anal. Chem. 77, 1282–1289.
Cook, R.T., 2004. The tolerance of the field slug Deroceras reliculatum to freezingtemperatures. Cryo Lett. 25, 187–194.
Costanzo, J.P., Moore, J.B., Lee, R.E., Kaufman, P.E., Wyman, J.A., 1997. Influence of soilhydric parameters on the winter cold hardiness of a burrowing beetle, Leptinotarsadecemlineata (Say). J. Comp. Physiol. B 167, 169–176.
Crowe, J.H., Crowe, L.M., Carpenter, J.F., Wistrom, C.A., 1987. Stabilization of dryphospholipid bilayers and proteins by sugars. Biochem. J. 242, 1–10.
Engelke, S., Kompf, J., Jordaens, K., Tomiuk, J., Parker, E.D., 2011. The genetic dynamicsof the rapid and recent colonization of Denmark by Arion lusitanicus (Mollusca,Pulmonata, Arionidae). Genetica 139, 709–721.
Fan, W.M.T., 1996. Metabolite profiling by one- and two-dimensional NMR analysis ofcomplex mixtures. Prog. Nucl. Mag. Res. Sp. 28, 161–219.
Felsenstein, J., 1985. Confidence-limits on phylogenies—an approach using thebootstrap. Evolution 39, 783–791.
Getz, L.L., 1959. Notes on the ecology of slugs: Arion circumscriptus, Deroceras reticulatum,and D. laeve. Am. Midl. Nat. 61, 485–498.
Hochachka, P.W., Somero, G.N., 2002. Biochemical Adaptation: Mechanism and Processin Physiological Evolution. Oxford University Press, New York.
Holmstrup, M., Zachariassen, K.E., 1996. Physiology of cold hardiness in earthworms.Comp. Biochem. Physiol. A 115, 91–101.
Holmstrup, M., Costanzo, J.P., Lee, R.E., 1999. Cryoprotective and osmotic responses tocold acclimation and freezing in freeze-tolerant and freeze-intolerant earthworms.J. Comp. Physiol. B 169, 207–214.
Kerney, M.P., Cameron, R.A.D., Riley, G., 1979. A field guide to the land snails of Britainand Northwest Europe. William Collins Sons and Co., London.
Keun, H.C., Ebbels, T.M.D., Antti, H., Bollard, M.E., Beckonert, O., Holmes, E., Lindon, J.C.,Nicholson, J.K., 2003. Improved analysis of multivariate data by variable stabilityscaling: application to NMR-based metabolic profiling. Anal. Chim. Acta 490, 265–276.
Kozlowski, J., 2000. Reproduction of Arion lusitanicus Mabille, 1868 (Gastropoda:Pulmonata: Arionidae) introduced in Poland. Folia Mal. 8, 87–94.
Kozlowski, J., 2007. The distribution, biology, population dynamics and harmfulness ofArion lusitanicus Mabille, 1868 (Gastropoda: Pulmonata: Arionidae) in Poland. J.Plant Prot. Res. 47, 219–230.
Lindon, J.C., Nicholson, J.K., Everett, J.R., 1999. NMR spectroscopy of biofluids. Annu.Rep. NMR Spectrosc. 38, 1–88.
Malmendal, A., Overgaard, J., Bundy, J.G., Sørensen, J.G., Nielsen, N.C., Loeschcke, V.,Holmstrup, M., 2006. Metabolomic profiling of heat stress: hardening and recoveryof homeostasis in Drosophila. Am. J. Physiol. 291, R205–R212.
Mellanby, K., 1961. Slugs at low temperatures. Nature 189, 944.Noble, L.R., 1992. Differentiation of large arionid slugs (Mollusca, Pulmonata) using
ligula morphology. Zool. Scr. 21, 255–263.Overgaard, J., Malmendal, A., Sørensen, J.G., Bundy, J.G., Loeschcke, V., 2007a. Metabolomic
profiling of rapid cold hardening and cold shock in Drosophila melanogaster. J. InsectPhysiol. 53, 1218–1232.
Overgaard, J., Slotsbo, S., Holmstrup, M., Bayley, M., 2007b. Determining factors forcryoprotectant accumulation in the freeze-tolerant earthworm, Dendrobaenaoctaedra. J. Exp. Zool. 307A, 578–589.
Overgaard, J., Tollarova, M., Hedlund, K., Petersen, S.O., Holmstrup, M., 2009. Seasonalchanges in lipid composition and glycogen storage associated with freeze-tolerance of the earthworm, Dendrobaena octaedra. J. Comp. Physiol. B 179,569–577.
Palumbi, S.R., 1996. Nucleic acids II: the polymerase chain reaction. In: Hillis, D.M.,Mable, B., Moritz, C. (Eds.), Molecular Systematics. Sinauer Associates, Inc., USA,pp. 205–247.
Pedersen, P.G., Holmstrup, M., 2003. Freeze or dehydrate: only two options for thesurvival of subzero temperatures in the arctic enchytraeid Fridericia ratzeli. J.Comp. Physiol. B 173, 601–609.
Posada, D., 2008. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 25,1253–1256.
Rundgren, S., 1975. Vertical distribution of lumbricids in southern Sweden. Oikos 26,299–306.
Saitou, N., Nei, M., 1987. The neighbor-joining method: a new method for reconstructingphylogenetic trees. Mol. Biol. Evol. 4, 406–425.
Sinclair, B., 1999. Insect cold tolerance: How many kinds of frozen? Eur. J. Entomol. 96,157–164.
Slotsbo, S., Maraldo, K., Malmendal, A., Nielsen, N.C., Holmstrup,M., 2008. Freeze toleranceand accumulation of cryoprotectants in the enchytraeid Enchytraeus albidus(Oligochaeta) from Greenland and Europe. Cryobiology 57, 286–291.
Slotsbo, S., Hansen, L.M., Holmstrup, M., 2011. Low temperature survival in differentlife stages of the Iberian slug, Arion lusitanicus. Cryobiology 62, 68–73.
Sømme, L., 1982. Supercooling and winter survival in terrestrial arthropods. Comp.Biochem. Physiol. A 73, 519–543.
Storey, K.B., Storey, J.M., 1990. Facultativemetabolic rate depression:molecular regulationand biochemical adaptation in anaerobiosis, hibernation and estivation. Quart. Rev.Biol. 65, 145–174.
Storey, K.B., 1997. Organic solutes in freezing tolerance. Comp. Biochem. Physiol. A 117,319–326.
Storey, K.B., Storey, J.M., 1996. Natural freezing survival in animals. Annu. Rev. Ecol.System. 27, 365–386.
Storey, K.B., Storey, J.M., Churchill, T.A., 2007. Freezing and anoxia tolerance of slugs: ametabolic perspective. J. Comp. Physiol. B 177, 833–840.
Swofford, D.L., 2002. PAUP*: Phylogenetic analysis using parsimony (*and othermethods), version 4.0. Sinauer Associates Inc, Sunderland, Massachusetts.
Tamura, K., Dudley, J., Nei, M., Kumar, S., 2007. MEGA4: molecular evolutionary geneticsanalysis (MEGA) software version 4.0. Mol. Biol. Evol. 24, 1596–1599.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins, D.G., 1997. The ClustalXwindows interface: flexible strategies for multiple sequence alignment aided byquality analysis tools. Nucleic Acids Res. 24, 4876–4882.
Udaka, H., Goto, S.G., Numata, H., 2008. Effects of photoperiod and acclimation temperatureon heat and cold tolerance in the terrestrial slug, Lehmannia valentiana (Pulmonata:Limacidae). Appl. Entomol. Zool. 43, 547–551.
von Proschwitz, T., Winge, K., 1994. Iberiaskogsnegl—en art på spredning i Norge.Fauna 47, 195–203.
Wold, S., 1978. Cross-validatory estimation of number of components in factor andprincipal components models. Technometrics 20, 397–405.
Zachariassen, K.E., 1985. Physiology of cold tolerance in insects. Physiol. Revs. 65,799–832.
Zachariassen, K.E., Kristiansen, E., Pedersen, S.A., Hammel, H.T., 2004. Ice nucleation insolutions and freeze-avoiding insects—homogeneous or heterogeneous? Cryobiology48, 309–321.
449S. Slotsbo et al. / Comparative Biochemistry and Physiology, Part A 161 (2012) 443–449


69PhD thesis by Stine Slotsbo
J Comp Physiol B, doi: 10.1007/s00360-011-0594-y
Stine Slotsbo • Karina Vincents Fisker • Lars Monrad Hansen • Martin Holmstrup
S. Slotsbo • L.M. HansenDepartment of Integrated Pest Management, Faculty of Agricultural Sciences, Aarhus University, Forsøgsvej 1, DK-4200 Slagelse, Denmarke-mail: [email protected]
S. Slotsbo • K.V. Fisker • M. HolmstrupDepartment of Terrestrial Ecology, National Environmental Research Institute, Aarhus University, Vejlsøvej 25, P.O. Box 314,
DK-8600 Silkeborg, Denmark
PAPER IV
Drought tolerance in eggs and juveniles of the Iberian slug, Arion lusitanicus
Photo: Leyo/Wikimedia Commons.


71PhD thesis by Stine Slotsbo
ORIGINAL PAPER
Drought tolerance in eggs and juveniles of the Iberian slug,Arion lusitanicus
Stine Slotsbo • Karina Vincents Fisker •
Lars Monrad Hansen • Martin Holmstrup
Received: 9 February 2011 / Revised: 27 May 2011 / Accepted: 7 June 2011
� Springer-Verlag 2011
Abstract The spread of invasive species is an increasing
problem world wide. The invasive slug Arion lusitanicus
has spread to most parts of Europe, where it often is con-
sidered as a serious pest. There is a need for better
knowledge of its ecophysiology to be able to predict the
effect of climatic factors, such as temperature and humidity
on the population dynamics and abundance. The aim of the
present study was to assemble data on the water balance
and drought tolerance of eggs and juveniles of A. lusita-
nicus. Both eggs and juveniles had little capacity to prevent
evaporative water loss and lost water when the ambient
humidity fell below 99.8 and 99.5%, respectively. The
water conductance of the cuticle of juveniles was
242 lg cm-2 h-1 mmHg-1 and resembles that of other
slug species. Both eggs and juveniles of A. lusitanicus
tolerate a substantial water loss. There was no difference in
water loss resistance between eggs and juveniles, but the
eggs were slightly more tolerant to water loss than the
juveniles. The percent water loss causing 50% mortality
was 72% for the juveniles and 81% for the eggs. Despite
A. lusitanicus’ tolerance of substantial water loss, its sur-
vival depends on humid habitats.
Keywords Arion lusitanicus � Slugs � Drought �Desiccation stress � Water conductance � Invasive species
Introduction
The spread of invasive species is an increasing problem
world wide. Along with escalating global trade, transpor-
tation and international travel, the introduction and spread
of invasive species are increasing in frequency (Hulme
2009; Meyerson and Mooney 2007). This also applies to
the invasive slug Arion lusitanicus Mabille (Gatropoda:
Pulmonata: Arionidae), which has recently become estab-
lished in many European countries (including Denmark)
and is considered as a serious pest (Kozlowski 2007). It is
possible that A. lusitanicus has spread to Nordic countries
from more southern regions, however, the precise region
of origin (despite the name of the species) is unknown
(Quinteiro et al. 2005). A. lusitanicus seems to cope well
with the local conditions in the new areas to which it has
migrated, it spreads rapidly and often occurs in high den-
sities. The population size of A. lusitanicus can vary sub-
stantially between years (Grimm and Paill 2001) likely due
to climatic factors such as temperature and humidity, but
knowledge of A. lusitanicus’ ecophysiology is limited. Low
winter temperatures are unlikely to limit the populations
under Danish conditions (Slotsbo et al. 2011) whereas
drought periods are probably constraining performance;
therefore knowledge of the drought tolerance of A. lusita-
nicus is needed.
Nature conservation authorities and managers need
knowledge of the effect of temperature and rainfall on the
population dynamics and abundance of A. lusitanicus to be
able to predict the pest problem of the species at a certain
time or place. This knowledge will enable them to decide
Communicated by I.D. Hume.
S. Slotsbo (&) � L. M. Hansen
Department of Integrated Pest Management,
Faculty of Agricultural Sciences, Aarhus University,
Forsøgsvej 1, 4200 Slagelse, Denmark
e-mail: [email protected]
S. Slotsbo � K. V. Fisker � M. Holmstrup
Department of Terrestrial Ecology, National Environmental
Research Institute, Aarhus University, Vejlsøvej 25,
P.O. Box 314, 8600 Silkeborg, Denmark
123
J Comp Physiol B
DOI 10.1007/s00360-011-0594-y

72 PhD thesis by Stine Slotsbo
whether to institute management programmes in a given
year or in a given area.
Terrestrial animals living in the soil or on the soil sur-
face are occasionally exposed to drought and have adapted
to these drought periods. When coping with drought, two
strategies have generally evolved: one is to reduce the
water loss, and the other is to increase the tolerance of
water loss. To achieve these, soil-living organisms have
developed many different types of adaptation (Holmstrup
2002). Changes in behaviour are one of the most used
strategies to reduce water loss; this could be in terms of
reducing the activity level or escaping from the drought in
time or space. Other morphologically or physiologically
based adaptations may reduce the water loss or enhance the
organisms’ tolerance of water loss, such as reducing the
water conductance of the skin or cuticle, hereby lowering
the water loss by evaporation (Edney 1977).
Terrestrial molluscs are generally susceptible to desic-
cation, and several studies have found habitat selection to
be well correlated with the availability of water (Carne-
Cavagnaro et al. 2006; Getz 1959; Prior 1985). Suitable
micro-habitats offering protection against daily and sea-
sonal variation in availability of water are of importance
(Prior 1985).
The moist permeable integument of terrestrial molluscs
demands adaptations to avoid or cope with excessive
dehydration. The most obvious water regulating behaviour
of slugs and snails is their preference for moist habitats
(Prior et al. 1983; Prior 1985). However, dehydrated snails
and slugs are able to recover rapidly from dehydration by
absorbing water through their integument; this process is
termed contact re-hydration (Dainton 1954; Prior 1984,
1985). In contrast to land snails, terrestrial slugs lack
physical protection such as shell and epiphragm. During
drought periods, snails have the opportunity to withdraw
into their shells and become dormant, whereas slugs have
to respond differently.
Water balance is of importance for population processes
of terrestrial slugs (Carne-Cavagnaro et al. 2006;Willis et al.
2008). Their eggs are often laid at protected and humid sites,
e.g. in humid soil, in crevices or beneath debris, microhab-
itats that help ensure a high humidity. Since the development
time of eggs may be several months depending on temper-
ature, the eggs may from time to time be exposed to desic-
cation stress. The eggs are often laid in clusters, and one slug
lays several clusters of eggs,which increases the chances that
at least some eggs will survive and hatch under drought
conditions. Eggs of many soil-living invertebrates are
drought resistant, but little is known of the drought tolerance
of eggs of terrestrial slugs (Bayne 1969; Carmichael and
Rivers 1932; Carrick 1942). A. lusitanicus typically lays
eggs in summer and fall (Kozlowski 2000) and the eggs hatch
before winter (Slotsbo, unpublished data).
The aim of the present study was to investigate water
balance and drought tolerance of eggs and juveniles of
A. lusitanicus, including measurement of the water con-
ductance of the juveniles.
Materials and methods
Animals
Adult specimens of A. lusitanicus were collected in Sep-
tember 2008, 2009 and 2010, from several localities in east
Jutland, Denmark. The collected slugs were kept at 15�C in
plastic boxes (17 9 22 9 6 cm) and fed carrots, lettuce,
oatmeal and dog food pellets ad libitum. The bottom of
each box was covered with wet paper towel to ensure high
humidity. The collected slugs produced eggs which were
kept at 15�C in Petri dishes lined with wet filter paper.
Eggs laid in 2008 and 2009 were allowed to hatch, and the
juveniles were kept under the same conditions as the
adults. For experimentation, small juveniles with an aver-
age fresh weight of 23–53 mg were used. Eggs for the
experiment were obtained in 2010 and used 14–17 days
after they had been laid.
Osmolality and water content
The osmolality of body fluid was measured using a Wescor
HR 33 T Dewpoint Microvoltmeter (Wescor Inc., Logan,
Utah, USA) connected to C-52 sample chambers, as pre-
viously described by Bayley and Holmstrup (1999). To this
end, slugs were opened dorsally using scissors to expose
the hemolymph, which was absorbed by filter paper discs
fitting directly into sample chambers. Measurements of egg
fluids were done by opening (using a pair of needles) a
whole egg placed in the sample holder.
The water content of eggs and slugs was determined.
Slugs and eggs were blotted with filter paper and their fresh
weight determined to the nearest 0.1 mg. After drying for
24 h at 60�C, their dry weight (dw) was determined and
water content calculated as g water g-1 dw.
Water loss rate
The water loss rate was determined for juveniles exposed
for 100 min to a range of relative humidities (RH). Fully
hydrated juvenile slugs were placed individually in small
plastic tubes covered with net to prevent them from
escaping. Each tube was placed in a vial (3 cm high,
1.6 cm diameter), which was placed in the centre of a
160-ml plastic beaker (4.2 cm high, 7 cm diameter). The
beaker contained an aqueous NaCl solution, a glycerol
solution or grains of silica gel. Each beaker was sealed with
J Comp Physiol B
123

73PhD thesis by Stine Slotsbo
a tightly fitting plastic lid and Para film. The air in this
small closed system quickly equilibrated with water vapour
from either the solutions or the silica gel, creating a range
of RHs (Table 1; Holmstrup and Westh 1995). There were
five to six replicates of each RH. The experiment was
conducted at approximately 20�C.The water loss was measured by recording the weight
loss of each juvenile after 100 min of desiccation exposure.
At the start of the experiment, the slugs were blotted with
filter paper, placed in the plastic tube, and their fresh
weight determined to nearest 0.1 mg. After desiccation
exposure the water content of the slugs were determined as
before.
Water conductance
The water conductance was calculated as lg water lost
per cm2 surface per hour per unit saturation deficit (mmHg)
in the desiccation beakers. The saturation deficit was
17.535 mmHg at 0% RH and 20�C, which was used to
calculate the saturation deficit at different RHs. The water
conductance was assumed to be equal from all body parts
(Machin 1975). Surface area was estimated for each
specimen using Meeh’s formula: S = 12 M2/3, where S is
surface area (cm2) and M is initial mass (g) (Meeh 1897
cited by Thompson et al. 2006).
Desiccation tolerance
Survival and water loss after a 7-day exposure to a range of
RHs were determined for both juveniles and eggs. Fully
hydrated juveniles and eggs were placed in small plastic
vials (3 cm high, 1.6 cm diameter), with one juvenile or
four eggs in each vial. The vials with juveniles were cov-
ered with net to prevent them from escaping. Each vial was
placed in the centre of a 160-ml plastic beaker (4.2 cm
high, 7 cm diameter) containing an aqueous NaCl solution.
Different concentrations of NaCl solutions were used to
establish a range of desiccation levels between 100 and
90% RH (Table 1). The experiment was conducted at a
constant temperature 15 ± 1�C.The water loss during the experiment was determined as
described. After 7 days of exposure, the slugs and eggs
were transferred to Petri dishes lined with wet filter paper
to rehydrate. The survival of the slugs was determined after
24 h of rehydration, and slugs were scored as survivors if
they were able to move normally. The eggs were kept at
15 ± 1�C and their development time to hatching and
hatching success was recorded.
The experiment with juveniles was conducted two
times, first in 2008, and then again in 2009. In the
experiment conducted in 2008, the juveniles were
exposed to desiccation levels between 99.9 and 95.1%
RH. Water loss was determined for five juveniles at each
RH. The survival was determined using 20 juveniles for
each RH. In the experiment conducted in 2009, the
juveniles were exposed to desiccation levels between 99.9
and 90% RH. In this experiment, the survival and water
loss was determined for each individual, with 11–20
replicates for each RH.
Statistical methods
Differences in osmolality and water content between
juveniles and eggs were tested by t tests. The linear and
non linear regressions were, like the t tests, performed
using Sigmaplot for Windows Version 11 (SPSS Inc.,
Chicago, IL, USA).
Survival data from the desiccation tolerance experi-
ments were analysed by a three parameter model describ-
ing the sigmoid dose–response curve (Damgaard et al.
2002). The three parameters were as follows; control
mortality (k), the shape parameter (b) and the x value in the
point of inflexion (x0).
f ðx) ¼ 1þ e�b�x0
1þ ebðx�x0Þ � ð1� kÞ ð1Þ
The calculation and solving of the equation was
performed using Mathematica for Windows Version 7
(Wolfram Research, Inc., Champaign, IL, USA).
Table 1 Concentration of NaCl and Glycerol in aqueous solutions
with corresponding relative humidity (RH) and vapour pressure in air
equilibrated with the solutions
RH
(%)
Vapour pressure
at 15�C (kPa)
Vapour pressure
at 20�C (kPa)
g NaCl L-1
0.5 99.9 1.703 2.335
17 99.0 1.688 2.314
36 98.0 1.671 2.291
45 97.5 1.662 2.279
54 97.0 1.654 2.268
63 96.5 1.645 2.256
71 96.1 1.638 2.247
72 96.0 1.637 2.244
89 95.1 1.621 2.223
90 95.0 1.620 2.221
107 94.0 1.603 2.198
124 93.0 1.586 2.174
174 90.0 1.534 2.104
g glycerol L-1
566.9 73.4 1.251 1.716
884.4 44.3 0.755 1.036
1009.2 33.8 0.576 0.790
1141.2 16.9 0.288 0.395
J Comp Physiol B
123

74 PhD thesis by Stine Slotsbo
Results
Osmolality and water content
The water content, calculated as g water g-1 dw, was
significantly higher in juveniles than in eggs (P\ 0.001;
t test). The osmolality of fully hydrated juveniles was
significantly higher than the osmolality of fully hydrated
eggs (P\ 0.001; t test) (Table 2).
Water conductance of juvenile slugs
The rate of water loss increased with decreasing RH, cor-
responding to increasing desiccation stress (Fig. 1). The
correlation between RH and water loss rate was exponen-
tial and could be described by the following equation
f ðxÞ ¼ 39:36� expð�0:023� xÞ. The nonlinear regressionused had a correlation coefficient (r2) at 0.87 (P\ 0.0001).
The water conductance for the juveniles was estimated to
be 242 ± 68 lg cm-2 h-1 mmHg-1.
Desiccation tolerance of eggs
Seven-day drought exposure at a RH above 97% did not
affect the hatching success of eggs (Fig. 2a). Using Eq. 1,
the parameters of the dose–response curve were estimated
to be: k = 0.11, b = 2.38 and x0 = 95.48. The RH causing
a 50% reduction in the hatching success was, thus, esti-
mated to be 95.5% RH. After 7 days’ exposure at this RH,
eggs had lost more than 80% of their initial water content
(Fig. 2b).
The hatching success was plotted against water loss, and
it became clear that eggs tolerated the loss of a consider-
able amount of water (Fig. 2c). A survival curve was fitted
using Eq. 1 and the parameters were estimated to be:
k = 0.12, b = 0.26 and x0 = 81.30. Eggs exposed to
drought could lose up to 63–70% of the initial water con-
tent before it affected the hatching success. The percentage
water loss causing a 50% reduction in the hatching success
was estimated to be 81% of the eggs’ initial water content.
In the treatment resulting in a loss of 86% of the initial
water content, 18% of the eggs still hatched.
The development time of the eggs increased signifi-
cantly with decreasing relative humidity (r2 = 0.85;
P[ 0.0011) (Fig. 3). Seven-day exposure to 95% relative
humidity resulted in a 7-day increase in development time
compared with controls.
Desiccation tolerance of juveniles
The survival of the juvenileswas unaffectedwhen exposed to
humidities above 97% RH for 7 days (Fig. 4a) despite con-
siderable water loss of their initial water content (Fig. 4b).
Slugs already began losing water at humidities around 99%
RH. At humidities below 97% RH the survival decreased,
and no juveniles survived 7 days’ exposure to RH below
94% (Fig. 4a). Equation 1 fitted to the data yielded the
parameters: k = 0.04, b = 1.80 and x0 = 95.82. Thus, 50%
mortality occurred approximately at 96% RH.
When survival was plotted against water loss, it became
clear that the juveniles tolerated considerable water loss
(Fig. 4c). Equation 1 fitted to the data gave the following
parameters: k = 0.08, b = 0.20 and x0 = 71.75, meaning
that 50% mortality occurred when juveniles had lost about
72% of the initial water content. Survival was unaffected if
water loss did not exceed 56%, and no juveniles survived a
water loss above 80%.
Discussion
Desiccation tolerance of eggs
The outer layer of eggs of A. lusitanicus is fairly thick and
contains crystals of calcium carbonate (South 1992). Below
Table 2 The dry weight (DW), water content and osmolality of juveniles and eggs of Arion lusitanicus used in this study
Groups DW (mg) Water content (g g-1 DW) Osmolality (mOsmol)
Juvenile 7.3 ± 1.1 (n = 5) 4.1 ± 0.1 (n = 5) 230 ± 1.0 (n = 6)
Eggs 4.6 ± 0.2 (n = 10) 3.1 ± 0.1 (n = 10) 130 ± 1.0 (n = 6)
Values are mean ± SE
100 80 60 40 20 0
Relative humidity (%)
Wat
er lo
ss (
% o
f WC
initi
al/1
00 m
in)
0
10
20
30
40
Fig. 1 The water loss rate of juveniles of Arion lusitanicus exposedto seven different relative humidities in the range from 7.5 to 98%.
The water loss rate is expressed as percent water loss of the initial
water content per 100 min. Values are mean ± SE
J Comp Physiol B
123

75PhD thesis by Stine Slotsbo
this calcareous coating are some inner shell membranes.
The innermost membrane of the egg forms a sac enclosing
the egg albumen (Bayne 1966, 1968). In the present study,
it was observed that eggs were able to hatch despite loss of
some of their shell, as long as the inner sac was intact. The
shell and membranes of the eggs were permeable and
provided little resistance against water loss. The osmolality
of the fully hydrated eggs was 130 mOsmol, which cor-
respond to a RH of 99.8%, below which they lost water.
Results of the present study show that A. lusitanicus
eggs were largely unaffected by water loss below 60%, and
even loss of 81% of the initial water content caused only a
50% reduction in hatching success. Comparable results
were reported by Carmichael and Rivers (1932), who found
that some eggs of the terrestrial slug Limax flavus were able
to survive as much as 85% weight loss, which corresponds
to a loss of 94% of the initial water content (Bayne 1969).
In early literature (Binney 1878), there are reports of even
greater drought tolerance of slug eggs, but those results
have been disproven by more recent authors (Bayne 1969;
Carmichael and Rivers 1932; Carrick 1942). In the present
study, some eggs survived a loss of up to 86% of their
initial water content. Effects of even higher water loss were
not tested in this study, but the survival under these con-
ditions would, indeed, have been very low.
In nature, A. lusitanicus, like most other terrestrial slugs,
lays its eggs in clusters rather than singly. Clustering has a
marked effect on the drying rates of eggs, especially eggs
in the middle of a cluster experience reduced water loss
(Bayne 1969; Willis et al. 2008). In the present study, the
95100 99 98 97 96 94
0 20 40 60 80 100
100 99 98 97 96 9495
Relative humidity (%)
Water loss (% of WCinitial)
Relative humidity (%)
Sur
viva
l (%
)S
urvi
val (
%)
Wat
er lo
ss (
% o
f WC
initi
al)
0
20
40
60
100
80
0
20
40
60
100
80
0
20
40
60
100
80
a
c
b
Fig. 2 The survival (a) and water loss (b) of Arion lusitanicus eggs exposed to different relative humidities for 7 days. c The relationship
between survival and water loss. Sigmoid dose–response curves were fitted to the survival data. Values are mean ± SE
Relative humidity (%)
Dev
elop
men
t tim
e (d
ays)
32
34
36
38
40
42
44
100 99 98 97 96 9495
Fig. 3 The development time of Arion lusitanicus eggs exposed for
7 days to a range of relative humidities between 95 and 100%. Values
are mean ± SE
J Comp Physiol B
123

76 PhD thesis by Stine Slotsbo
desiccation exposure was performed on batches of only
four eggs, which did not form a clutch during the exposure
period. It is, therefore, likely that the water loss at the given
RH reported here was higher than for eggs laid in clusters.
The desiccation tolerance of eggs can be influenced by
the developmental stage of the eggs. In the present study,
only one developmental stage (about midway in develop-
ment) was investigated, but the literature does not provide
consistent conclusions on which developmental stage is the
most or least tolerant. Carmichael and Rivers (1932) found
fully developed embryos of L. flavus to be least tolerant, in
contrast Bayne (1969) found the early stages of embryos of
Deroceras reticulatum to be least tolerant.
Desiccation effects on the development time of eggs
The development time of the eggs in the present study
increased significantly with increasing desiccation. The
same has been found in the pond snail Lymnaea humilis,
where partial drying of egg masses prolonged the hatching
time (McCraw 1961). For eggs of A. lusitanicus, a 7-day
exposure to 95% RH resulted in a 7-day delay in hatching
compared with controls, meaning that the embryonic
development was probably rapidly arrested due to water
loss.
This arrest of embryonic development as a consequence
of desiccation has also been observed in cocoons of the
earthworm Aporrectodea caliginosa subjected to desicca-
tion (Holmstrup 2001). In a study of Bayne (1969), no
increase in the development time of eggs of the terrestrial
slug D. reticulatum was observed as a consequence of
drought exposure. However, eggs in that experiment were
dehydrated for only 14 h before they were allowed to
rehydrate, which did not allow for any good assessment of
delays in development time.
Water loss resistance of juveniles
There are two ways for slugs to cope with drought, one is
to reduce the water loss, and the other is to increase the
tolerance of water loss. In reducing the water loss, the
water conductance of the skin is an important factor. The
9094969810092 92100 98 96 94 90
-40 0-20 20 40 60 80 100
Relative humidity (%)
Water loss (% of WCinitial)
Relative humidity (%)
Sur
viva
l (%
)S
urvi
val (
%)
Wat
er lo
ss (
% o
f WC
initi
al)
0
20
40
60
100
80
0
20
40
60
100
80
-40
20
0
-20
40
60
100
80
a
c
b
Fig. 4 The survival (a) and water loss (b) of juveniles of Arionlusitanicus exposed to different relative humidities for 7 days. c The
relationship between survival and water loss. Data from the exper-
iment performed in 2008, where water loss and survival were
measured on separate individuals, are shown as circles. Data from the
experiment preformed in 2009, where water loss and survival were
measured on the same individuals, are shown as triangles. Sigmoid
dose–response curves were fitted to the survival data. Values are
mean ± SE
J Comp Physiol B
123

77PhD thesis by Stine Slotsbo
water conductance of juvenile A. lusitanicus was estimated
to 242 lg cm-2 h-1 mmHg-1, which corresponds fairly
well with data from the study by Thompson et al. (2006),
who found the water conductance of the slug species
Philomycus carolinianus, D. leave and L. Valentiana to be
210, 348 and 378 lg cm-2 h-1 mmHg-1, respectively.
Not surprisingly, these water conductance values show that
slugs, based on their water conductance, are confined to
hygric habitats. In comparison, insects of mesic habitats
typically have water conductance values ranging from 20
to 60 lg cm-2 h-1 mmHg-1 (Edney 1977); almost one
order of magnitude lower.
Snails cope with drought differently than slugs, mainly
because snails can withdraw into the shell so that only a
limited area of the mantle tissue remains exposed to the air.
Further, evidence has been found for a ‘‘water barrier’’ in
the mantel tissue of snails (Machin 1975). The water
conductance of active specimens of the common garden
snail Helix aspersa is nearly identical to that from a free
water surface. This is in sharp contrast to inactive snails
that are able to regulate evaporation markedly (Machin
1964). The water conductance of the mantel tissue of
inactive H. aspersa is 39 lg cm-2 h-1 mmHg-1 (Machin
1966), which is six times lower than the water conductance
of juvenile A. lusitanicus.
The ability to survive drought cannot be explained by
water conductance alone. The water conductance tells
nothing about the size and, therefore, the magnitude of
water reserves. Large animals lose a smaller proportion of
their original weight per time than small ones with the
same water conductance. Comparing the slugs mentioned
above, reveals that the slug with the lowest water con-
ductance, P. carolinianus, is also the largest, with a weight
of 2.87 g, and therefore perhaps well adapted to withstand
long periods of desiccating conditions. The juveniles used
in the present study weighed only 37 mg. Because of sur-
face–volume relationships, adult slugs would be able to
withstand drought for longer periods than the juveniles of
the same species. The water loss and survival reported in
this study are, therefore, true for the most drought sensitive
size class of A. lusitanicus. Older and larger individuals of
A. lusitanicus would, presumably, survive drought better.
There are different behavioural adaptations to reduce the
water loss. Juvenile and adult slugs have a tendency to
cluster if they are exposed to desiccation stress. This
behaviour is called huddling behaviour (Prior et al. 1983)
in which they pack closely together, thereby reducing the
surface area and generating a high humidity microenvi-
ronment around the huddled slugs. This huddling behav-
iour (also applying to clusters of eggs) reduces the vapour
pressure deficit and, thereby evaporation (Prior 1985).
Another way of reducing water loss is to reduce the
water vapour pressure deficit between the animal and its
surroundings. Some animals raise their internal osmolality
in response to drought, either passively as a consequence of
water loss or actively by the production of osmolytes
(Bayley and Holmstrup 1999). An increased osmolality as
a consequence of water loss has been observed in both slug
and snail species (Prior 1985). The osmolality during
drought was not measured in the present study, but in an
earlier investigation it was found that the osmolality did not
rise in A. lusitanicus despite considerable water loss
(Slotsbo et al. 2011). It seems, therefore, that A. lusitanicus
relies solely on behavioural responses to reduce water loss
in terms of homing to humid resting places, huddling
behaviour, and nocturnal activity pattern.
Dehydration tolerance of juveniles
As indicated by the water conductance of 242 lgcm-2 h-1 mmHg-1, the skin of the juveniles was quite
permeable and provided little resistance against water loss.
The osmolality of the fully hydrated juveniles was
230 mOsmol, which corresponds to a relative humidity of
99.5%, below which they lost water. As a consequence,
A. lusitanicus will, inevitably, often experience water loss
in nature. Therefore, to a certain degree its drought toler-
ance relies on its ability to tolerate dehydration. A high
degree of dehydration tolerance seems to be widespread
among gastropods. The aquatic snail Australorbis glabra-
tus tolerates losing up to 70% of its initial water content
(Von Brand et al. 1957), and in the intertidal snail,
Melampus Bidentatus, an 80% loss of body water resulted
in 64% mortality (Price 1980). Among terrestrial slugs,
L. tenellus survives the loss of 80% of its weight (Kunkel
1916 cited by Bayne 1969), and P. carolinianus approxi-
mately 65% (Thompson et al. 2006).
Implications for population dynamics
The results of the present study show that both eggs and
juveniles of A. lusitanicus have little resistance to evapo-
rative water loss under desiccating conditions, but on the
other hand tolerate a substantial water loss. The tolerance
limits established in the present study could be used to
predict range limits of the species given that detailed
knowledge of soil pore air humidity conditions of the
concerned area are available. For example, the abundance
of the invasive slug, D. panormitanum, on Marion Island
was associated with habitats, where lower lethal tempera-
tures were not occurring, but also the species preference for
habitats with a high humidity below the vegetation (Lee
et al. 2009).
Soil pore air humidity data are scarce in the literature
but a few studies exist for Danish habitats. Thus, field
experiments have shown that even under extreme drought
J Comp Physiol B
123

78 PhD thesis by Stine Slotsbo
conditions soil water potentials seldom drop below
-40 bar (RH 97.1%) at 5 cm depth (Maraldo and
Holmstrup 2009). Juveniles of A. lusitanicus did lose water
at this RH, but due to their dehydration tolerance survival
was unaffected after 1 week’s exposure. During longer
drought periods, slugs can seek deeper and more humid
layers; it seems therefore reasonable to conclude that
summer drought under Danish conditions is unlikely to
cause any substantial mortality in populations of A. lusi-
tanicus. This being said, activity of slugs is undoubtedly
reduced during droughts (Grimm et al. 2000; Kozlowski
2001; Sternberg 2000). A decrease in activity will reduce
foraging of the slugs, which can reduce the development
and thereby delay the reproduction of the slugs.
In order to use tolerance limits for prediction of range
limits, one has to be aware that factors other than climate
influence the distribution of the species (Lee et al. 2009).
Thus, understanding the general processes of climatic
factors’ implications for the abundance structure and range
limits of a species, the species has to be investigated in a
relatively isolated part of the species range, to avoid dis-
persal from or to other populations (Lee et al. 2009). Since
A. lusitanicus is probably still unintentionally dispersed
between regions due to human activity this analysis is
difficult. Moreover, biotic factors such as predation (which
was not present in the Marion Island case) and competition
with other slug species for food and space may confound
the effects of climatic conditions. Lastly, predicting the
range limits of a species on the basis of the species’ tol-
erance of climatic factors, one has to take spatial variations
of microclimatic conditions into account. In very dry
periods some areas, e.g. borders of water bodies or flood
plains, can still be very humid and can create refuges from
where the species can re-colonize when conditions become
favourable again. Further work is therefore needed to fully
exploit the results on desiccation tolerance of A. lusitanicus
to understand why this species has spread rapidly to Nordic
countries over the last decades.
References
Bayley M, Holmstrup M (1999) Water vapor absorption in arthropods
by accumulation of myoinositol and glucose. Science
285:1909–1911
Bayne CJ (1966) Observations on composition of layers of egg of
Agriolimax reticulatus grey field slug (Pulmonata stylomato-
phora). Comp Biochem Physiol 19:317–337
Bayne CJ (1968) A study of desiccation of egg capsules of 8
gastropod species. J Zool 155:401–411
Bayne CJ (1969) Survival of the embryos of the grey field slug
Agriolimax reticulatus, following desiccation of the egg. Mal-
acol 9:391–401
Binney WG (1878) The terrestrial air-breathing mollusks, 1st edn.
Bull Mus Comp Zool Harv Coll, Cambridge
Carmichael EB, Rivers TD (1932) The effects of dehydration upon
the hatchabilty of Limax flavus eggs. Ecol 13:375–380Carne-Cavagnaro VL, Keller MA, Baker GH (2006) Soil moisture
and soil type influence the breeding behavior of the pest snail
Cernuella virgata (da Costa). Appl Soil Ecol 33:235–242
Carrick R (1942) The grey field slug Agriolimax agrestis L, and its
environment. Ann Appl Biol 29:43–55
Dainton BH (1954) The activity of slugs. 1. Induction of activity by
changing temperatures. J Exp Biol 31:165–187
Damgaard C, Hojer R, Bayley M, Scott-Fordsmand JJ, Holmstrup M
(2002) Dose-response curve modeling of excess mortality caused
by two forms of stress. Environ Ecol Stat 9:195–200
Edney EB (1977) Water balance in land Arthropods, 1st edn.
Springer, Berlin
Getz LL (1959) Notes on the ecology of slugs: Arion circumscriptus,Deroceras reticulatum, and D. laeve. Am Midl Nat 61:485–498
Grimm B, Paill W (2001) Spatial distribution and home-range of the
pest slug Arion lusitanicus (Mollusca: Pulmonata). Acta Oecol
Int J Ecol 22:219–227
Grimm B, Paill W, Kaiser H (2000) Daily activity of the pest slug
Arion lusitanicus Mabille. J Molluscan Stud 66:125–128
Holmstrup M (2001) Sensitivity of life history parameters in the
earthworm Aporrectodea caliginosa to small changes in soil
water potential. Soil Biol Biochem 33:1217–1223
Holmstrup M (2002) Strategies for cold and drought tolerance in
permeable soil invertebrates. Doctor’s dissertation (DSc).
National Environmental Research Institute, Silkeborg, Denmark
Holmstrup M, Westh P (1995) Effects of dehydration on water
relations and survival of lumbricid earthworm egg capsules.
J Comp Physiol B 165:377–383
Hulme PE (2009) Trade, transport and trouble: managing invasive
species pathways in an era of globalization. J Appl Ecol
46:10–18
Kozlowski J (2000) Reproduction of Arion Lusitanicus Mabille, 1868
(Gastropoda: Pulmonata: Arionidae) introduced in Poland. Folia
Malacol 8:87–94
Kozlowski J (2001) Daily activity of Arion lusitanicus Mabille, 1868
(Gastropoda: Pulmonata: Arionidae). J Plant Prot Res
41:279–286
Kozlowski J (2007) The Distribution, biology, population dynamics
and harmfulness of Arion lusitanicusMabille, 1868 (Gastropoda:
Pulmonata: Arionidae) in Poland. J Plant Prot Res 47:219–230
Lee JE, Janion C, Marais E, van Vuuren BJ, Chown SL (2009)
Physiological tolerances account for range limits and abundance
structure in an invasive slug. Proc Royal Soc B Biol Sci
276:1459–1468
Machin J (1964) Cutaneous regulation of evaporative water loss in the
common garden snail Helix aspersa. Nat Wiss 53:18
Machin J (1966) Evaporation of water from Helix aspersa.4. Loss
from mantle of inactive snail. J Exp Biol 45:269–278
Machin J (1975) Osmotic gradient across snail epidermis evidence for
a water barrier. Science 183:759–760
Maraldo K, Holmstrup M (2009) Recovery of enchytraeid populations
after severe drought events. Appl Soil Ecol 42:227–235
McCraw (1961) Life history and growth of the snail, Lymnaea humilisSay. Trans Am Microsc Soc 80:16–27
Meeh K (1897) Oberflachenmessungen des menschlichen korpers.
Z Biol 15:425–458
Meyerson LA, Mooney HA (2007) Invasive alien species in an era of
globalization. Front Ecol Environ 5:199–208
Price CH (1980) Water relations and physiological ecology of the salt
marsh snail, Melampus bidentatus Say. J Exp Mar Biol Ecol
45:51–67
Prior DJ (1984) Analysis of contact-rehydration in terrestrial gastro-
pods—osmotic control of drinking behavior. J Exp Biol
111:63–73
J Comp Physiol B
123

79PhD thesis by Stine Slotsbo
Prior DJ (1985) Water-regulatory behavior in terrestrial gastropods.
Biol Rev Camb Philos Soc 60:403–424
Prior DJ, Hume M, Varga D, Hess SD (1983) Physiological and
behavioral aspects of water balance and respiratory function in
the terrestrial slug, Limax maximus. J Exp Biol 104:111–127
Quinteiro J, Rodriguez-Castro J, Castillejo J, Iglesias-Pineiro J, Rey-
Mendez M (2005) Phylogeny of slug species of the genus Arion:evidence of Iberian endemics and of the existence of relict
species in Pyrenean refuges. J Zool Syst Evol Res 43:139–148
Slotsbo S, Hansen LM, Holmstrup M (2011) Low temperature
survival in different life stages of the Iberian slug, Arionlusitanicus. Cryobiol 62:68–73
South A (1992) Terrestrial slugs. Biology, ecology and control, 1st
edn. Chapman & Hall, London
Sternberg M (2000) Terrestrial gastropods and experimental climate
change: a field study in a calcareous grassland. Ecol Res
15:73–81
Thompson JM, Appel AG, Sibley JL, Keever GJ, Foshee WG (2006)
Comparative water relations of three sympatric terrestrial slugs:
(Stylommatophora: Agriolimacidae, Limacidae, and Philomyci-
dae). J Ala Acad Sci 77:181–192
Von Brand T, McMahon P, Nolan MO (1957) Observations on
starvation and desiccation of snail Australorbis glabratus. BiolBull 113:89–102
Willis JC, Bohan DA, Powers SJ, Choi YH, Park J, Gussin E (2008)
The importance of temperature and moisture to the egg-laying
behaviour of a pest slug, Deroceras reticulatum. Ann Appl Biol
153:105–115
J Comp Physiol B
123


AARHUSUNIVERSTYAU
ECOPHYSIOLOGY AND LIFE HISTORY OF THE SLUG, ARION LUSITANICUS
The slug Arion lusitanicus is an invasive species which has recently become established in many European countries and is considered a serious pest, both in agriculture and private gardens. The aim of this PhD was to describe in detail the ecophysiology of A. lusitanicus and map its life history traits in order to reveal how life history traits respond to major environmental parameters such as temperature and moisture. Knowledge of the species’ biology is neces-sary to generate eff ective pest management plans.
This PhD project contributes with detailed knowledge of the ecophysiology and life history traits of A. lusitanicus. A growth model that enables the prediction of individual slug growth under natural temperature scenarios has been de-veloped. The project has shown how egg developmental time and hatchability are infl uenced by temperature and drought. Furthermore, the cold and drought tolerance limits of A. lusitanicus have been revealed. The results will be a valuable contribution to the development of population growth models to provide a more accurate forecasting of slug abundance as a tool for pest management.
Ecop
hysiolo
gy a
nd life
history of the
slug, A
rion lusitanicus
PhD thesis 2012Stine Slotsbo
ECOPHYSIOLOGY AND LIFE HISTORY OF THE SLUG, ARION LUSITANICUS
ISBN: 978-87-92825-xx-x