leaf anatomy and its relation to the ecophysiology of … · leaf anatomy and its relation to the...
TRANSCRIPT

Leaf anatomy and its relation to the ecophysiology ofsome non-succulent desert plants from Egypt
Gamal M. Fahmy
Department of Botany, Faculty of Science, University of Cairo, Giza12613, Egypt
(Received 1 April 1995, accepted 15 July 1996)
A quantitative study of leaf anatomy of 20 species of some non-succulentperennial desert plants belonging to 12 families of the Dicotyledoneae isprovided. The species were collected from natural desert habitats belongingto three phytogeographical regions of Egypt. Features representing xer-omorphy include small leaf or leaflet area (18 species), pubescent surfaces (16species), amphistomaticy, iso-bilateral leaf construction and multilayeredpalisade of two to four layers (18 species). Particular attention is given toquantify the palisade properties as well as the hydraulic conductance (Kh) ofthe xylem tissue supplying the transpiring areas. The ratio of the palisade cellsurface area/the leaf or leaflet area (Apal/A) ranges from 22·6 in Astragalusspinosus to 49·6 in Zilla spinosa. Moreover, Apal/A is highly correlated with thepalisade thickness (r = 0·541, N = 100, p = 0·0001) and with the thicknessof lamina. The predicted Kh of the xylem is calculated from the conduitlumen diameters using the Hagen-Poiseuille relation. Eighteen species showlow predicted Kh values ranging from 0·6 to 300·5 3 10–13 m4 MPa–1 s–1.The highest values of predicted Kh occur in Gomphocarpus sinaicus andChrozophora obliqua, which have the highest leaf areas. Predicted Khcorrelates very well with leaf or leaflet area supplied by the petiole or petiolule(r = 0·685, N = 100, p = 0·0001). The efficiency of xylem conductance isfurther identified by calculations of the leaf specific conductance (LSC = Kh/distal leaf or leaflet area) and the predicted maximum pressure gradients(MPa m–1). The measured anatomical parameters and their correlations arediscussed in terms of the ecophysiological adjustments of the desert plants totheir habitat.
©1997 Academic Press Limited
Keywords: anatomy; xeromorphy; desert plants; non-succulents; Egypt
Introduction
Heat and drought are the commonest characteristics of deserts, but the two are notalways combined and deserts may, in fact, be either hot or cold, according to theirlatitude and altitude. In all, however, there is, for one reason or another, a deficiencyof available moisture, and this is usually due to the lack of rain (Good, 1953; Fahn &Cutler, 1992).
Journal of Arid Environments (1997) 36: 499–525
0140–1963/97/030499 + 27 $25.00/0/ae960217 © 1997 Academic Press Limited

The Sahara is a hot desert straddling the Tropic of Cancer and occupying nearly 10million km2 of Africa north of the Equator, roughly 20% of the world’s total arid andsemi-arid land surface (Larmuth, 1984). This area, within which the mean annualprecipitation is 100 mm or less and the mean annual temperature exceeds 30°C, is oneof the hottest regions of the world (Cloudsley-Thompson, 1984). The Sahara istraversed by only one river, the Nile, which crosses the entire length of Egypt on itsnorthward course to the Mediterranean sea. The vast desert areas represent more than97% of the total area of Egypt (Zahran & Willis, 1992).
In Egypt, as in the majority of arid lands, the survival of desert plants depends ontheir capacity to maintain a favourable balance between water uptake and water lossunder conditions of severe climatic and atmospheric drought (Kassas & Batanouny,1984). The structural and physiological characteristics typical of plants of arid regionsare not exclusive to desert species (Evenari, 1985). Through natural selection andevolution, characters constitutionally present in the various taxonomic groups areamplified, intensified and combined in various ways yielding adaptation(s) to aparticular habitat. This is manifested by the existence of particular adaptive featurescommon or shared between desert plants and other species commonly found inphysiologically or physically dry habitats, e.g. areas of high salinity, steppes of the semi-desert, grasslands, sclerophyllous scrub, coniferous forest, and alpine and arctic tundra(Oppenheimer, 1960; Fahn, 1964; Pyykko, 1966; Bocher, 1979; Abd El Rahman etal., 1980; Ibrahim & Fahmy, 1985; Hegazy, 1987; Korner et al., 1989; and others).Moreover, the above-mentioned studies involve species growing in different soil types,i.e. saline, serpentine, mineral poor, gypsum, calcareous, etc. (Shields, 1951; Small,1973). The existence of similar morpho-anatomical and sometimes physiologicalfeatures among the plant groups of different types of vegetation inhabiting dry habitatsleads one to predict that a degree of adaptive convergence between such diverse speciesis likely.
At least three photosynthetic carbon assimilation types (Laetsch, 1974) occur in allgreen plants, i.e. C3, C4 and CAM (Crassulacean Acid Metabolism). When takentogether, it is apparent that C4 and CAM plants have a higher water-use efficiency(ratio of net assimilation to water loss) in respect to photosynthesis (Jones, 1992). Inthe Egyptian desert, the percentage of C3 grasses decreases with decreasing latitude(Batanouny et al., 1988). The summer annual and other perennial grasses are mainlyC4 species. The CAM pathway is linked to succulence, though not all succulents areCAM plants, and this is in turn linked, at least to a certain degree, to highenvironmental salinity (Evenari, 1985).
Clearly, the non-succulent perennial dicots constitute the permanent framework ofvegetation in the Sahara Desert (Kassas & Batanouny, 1984). Although a highproportion of these species lack the C4 and CAM pathways and even lack a non-CAMsucculence (Ziegler et al., 1981; Kassas & Batanouny, 1984), it is apparent that theability of such species to withstand the adverse conditions of the desert environment isclearly attributed to particular traits.
Previous studies indicate that the long vertically extended roots of desert perennialsare sufficient to supply the shoots with their water requirements. This explains theexistence of such species in a green state during summer without showing wilting(Migahid, 1962). The sandy depressions in the gravel desert along the Cairo–Suezroad permanently contain 2·5% water at a depth of only 75 cm (wilting point, 0·8%),so that they never dry out and are capable of supporting a sparse perennial vegetation(cf. Walter, 1979, p. 105). This reflects the importance of xeromorphic characters ofthe perennial species, particularly those of the non-succulents, in minimising water lossduring the hot, dry rainless season.
The present study has been undertaken to examine how the structure andorganisation of the leaf or leaflet tissues of some non-succulent perennials affectvarious ecophysiological processes. This will shed light on the role of xeromorphic
G. M. FAHMY 500

features of such species in enabling them to exist and survive under arid desertconditions.
Material and methods
Investigated species and their sites
Specimens from 20 species belonging to 12 families of Dicotyledonae (Table 1) werecollected from the natural desert vegetation in Egypt. Identification and nomenclatureof the species followed Tackholm (1974) and Boulos (1995). The species are non-succulent perrenial chamaephytes which grow in different areas belonging to thefollowing phytogeographical regions of Egypt: (1) southern Sinai (S) in an areaextending from latitude 28° to 28° 25' N and longitude 33° 55' to 34° 25' E; (2) easterndesert in northern Sinai known as the Galalah Desert (Dg) in an area extending fromthe desert of Cairo near Helwan eastwards to that of Suez (latitude 29° 45' to 30° Nand longitude 31° 20' to 32° 20' E); and (3) western desert (Dl) near Faiyium (latitude29° 15' N, longitude 30° 55' E).
Sampling
Ten mature, fully exposed sun leaves were collected from five different individuals(two leaves from each individual plant) of each species growing in its specific regionduring the drought period (April–September) or at its end. Stalks of the laminae (thepetioles of simple leaves or the petiolules or leaflets) or the bases of sessile laminae andsmall segments of laminae were fixed in FAA (O’Brien & McCully, 1981).
Preparation
For light microscopy five leaves or leaflets were randomly obtained from the fixed plantmaterial previously sampled. The fixed parts of leaves or leaflets were dehydratedfollowing standard methods, embedded in Paraplast, sectioned in transverse andparadermal (longitudinal) directions at 10–15 µm and stained with 0·05% ToluidineBlue O in distilled water (Sakai, 1973). Microtome or freehand transverse sectionswere made in that part of the petiole or petiolule or in the base of transpiring part (inthe case of sessile leaves) proximal to the leaf or leaflet.
For SEM, samples from the FAA solution were dehydrated in a graded ethanolseries, critical point dried and coated with 200 A of gold–paladium in a Hummer ISputter Coater and viewed with an E.T.E.C. Autoscan scanning electron microscopeat 10 kV.
Measurements
The area of leaf or leaflet (one side only) was obtained by weighing their tracings onhigh quality paper, and comparing them with a paper of known area and weight.Optical micrometers (linear and square) were used to measure the densities (numberper mm2) of stomata and trichomes and the length of guard cells in epidermal stripsof leaves or leaflets. The number of palisade cells per unit area was obtained fromparadermal sections of leaves or leaflets at the adaxial (upper) and abaxial (lower)sides. The ratio of palisade cell area/leaf or leaflet area ( = Apal/A) was determinedfrom paradermal sections (obtained for light microscopy and for SEM) taken from
LEAF ANATOMY AND ECOPHYSIOLOGY 501

Table 1. A list of taxa of the investigated desert plants (from 1 to 20) and thephytogeographical regions and sites from which they have been collected for this study.
S=southern Sinai; Dg=Galalah Desert; Dl=Libyan Desert. See Materials andmethods section for details of the regions
Species Phytogeographical Site ofNo. Taxa region collection
Asclepiadaceae1 Gomphocarpus sinaicus Boiss. S Wadi Al-Arbaeen
Asteraceae2 Achillea fragrantissima (Forssk.)
Schultz Dg Wadi HofAsteraceae
3 Seriphidiom herba-alba (Asso)Y. R. Li S Wadi El-Raha
Asteraceae4 Artemisia judaica L. S St Katherine Village
Asteraceae5 Launaea spinosa (Forssk.) Sch. Bip. Dg Wadi Hagoul
Boraginaceae6 Heliotropium arbainense Fresen. Dg Wadi Hof
Boraginaceae7 Moltkiopsis ciliata (Forssk.) Johnston Dl Faiyium Desert
Brassicaceae8 Farsetia aegyptiaca Turra Dg Wadi Al-Gafra
Brassicaceae9 Zilla spinosa (Turra) Prantl Dg Wadi Hagoul
Capparidaceae10 Cleome droserifolia (Forssk.) Del. Dg Wadi Hagoul
Convolvulaceae11 Convolvulus lanatus Vahl Dg Wadi Al-Gafra
Euphorbiaceae12 Chrozophora obliqua (Vahl)
A. Juss ex Spreng. S Near Wadi FiranFabaceae
13 Acacia tortilis subsp. raddiana (Savi) Dg The Suez DesertFabaceae
14 Astragalus spinosus (Forssk.) Muschl. S Wadi Al-ArbaeenFabaceae
15 Crotalaria aegyptiaca Benth. Dg Wadi Al-GafraLamiaceae 40 km to the north of
16 Salvia deserti Decne. S St Katherine monastryLamiaceae
17 Teucrium pilosum (Decne.)Aschers. & Schweinf. S Wadi Al-Arbaeen
Resedaceae18 Ochradenus baccatus Del. Dg Wadi Al-Gafra
Solanaceae19 Lycium shawii Roem. & Schult. Dg Wadi Hof
Zygophyllaceae20 Fagonia arabica L. Dg Wadi Al-Gafra
G. M. FAHMY 502

both sides (adaxial and abaxial) of the leaf or leaflet from which the number of palisadecells per unit area was determined. Transverse sections 100 µm thick were then usedto construct a three-dimensional model from which lengths and diameters of palisadecells were determined (Nobel et al., 1975). Cell surface area of palisade (Apal) wascalculated assuming that the cells are cylindrical.
The thickness of epidermis, palisade and spongy tissues was measured fromtransections. The types of trichomes (glandular or non-glandular) and the surfacefeatures of cuticle were obtained from light microscope examinations and SEMphotomicrographs.
The predicted hydraulic conductance of the xylem of the petiole, petiolule or basalpart of the sessile leaf nearest to the transpiring area was determined according toGibson et al. (1984). The following equation was employed:
Kh = π∑di4
128η
where di is the diameter of the ith vessel element (m) and η is the solution viscosity(MPa s). The vessel elements were counted and their diameters were measured witha linear optical micrometer. The species studied differ in leaf or leaflet area supplied bythe vessel elements. In order to adjust hydraulic conductivities of such structuresaccording to the leaf area supplied, the leaf specific conductivity (LSC) wasdetermined according to Tyree et al. (1983) as follows:
LSC =Kh
distal leaf or leaflet area
Clearly, as the LSC decreases, the pressure drop (dp/dx) required for transportingwater through the axis of the transpiring area increases and consequently the steeperthe water potential gradient will be (Zimmermann, 1978). In order to predict thepressure gradient in the leaf or leaflet axes of the studied desert species, the LSC mustbe combined with the maximum transpiration rates (E max) as in the followingequation:
dp/dx (MPa m–1) =E max (10–7 m s–1)
LSC (10–7 MPa m–2 s–1)
Statistical analysis
Unless otherwise stated, data are means of five replicates obtained from measurementsperformed on five leaves sampled from five individual plants of each species. Means,differences among several means of the 20 species or between the means of eachcharacter on the adaxial and abaxial sides of leaf or leaflet were analysed by analysis ofvariance (Snedecor & Cochran, 1962). Least significant difference (LSD) was appliedto verify the significance of the difference between the means at 0·05 level ofprobability. Linear correlations were calculated between various parameters.
Results
Investigated species and their sites
The majority of the species studied (14 out of 20) are restricted to the silt terraces and
LEAF ANATOMY AND ECOPHYSIOLOGY 503

to channels and runnels of the main wadis of the eastern (Dg) and western deserts (Dl)which receive alluvial material of fine soil particles of diameters less than 0·7 mm(Kassas & Imam, 1954). Such habitats constitute substrate suitable for moisturestorage in deep soil layers which provide the roots of perennials with a continuouswater supply (Migahid, 1962). Species collected from southern Sinai (S) are membersof the perennial vegetation which inhabits the short wadis (Wadi Al-Raha and WadiAl-Arbaeen). In such habitats, the species are confined to the wadi bed crevices ofsmooth-faced outcrops, e.g. Gomphocarpus sinaicus, Astragalus spinosus and Teucriumpilosum. Likewise, the limestone country of Dg harbours some species (Achilleafragrantissima and Cleome droserifolia) which grow in the wadis covered by pebblycoarse textured soil with rock detritus of various sizes.
From the map of the distribution of different bioclimatic provinces in Egypt,southern Sinai (S), the Galalah (Dg) and the western deserts (Dl) belong to thehyperarid climatic provinces with a mild rainy winter and a hot summer (UNESCO,1979). Data extracted from Ayyad & Ghabbour (1985) showed that rainfall in suchregions is characterised by scantiness and irregularity. The rainy season extends fromDecember to April. Annual rainfall is less than 30 mm in El-Tor (S) and is 23·6 and10·8 mm in Wadi Hagoul (Dg) and Faiyium desert (Dl), respectively (average of 35years at least; data from Anon (1979)). Mean minimum temperature of the coldestmonth ranges from 5·9°C in Faiyium to 9°C in El-Tor region. The maximumtemperature of the hottest month ranges from 34·8 to 36·8°C in El-Tor and in FaiyiumDesert, respectively.
Morphology and surface area of simple leaf or leaflet
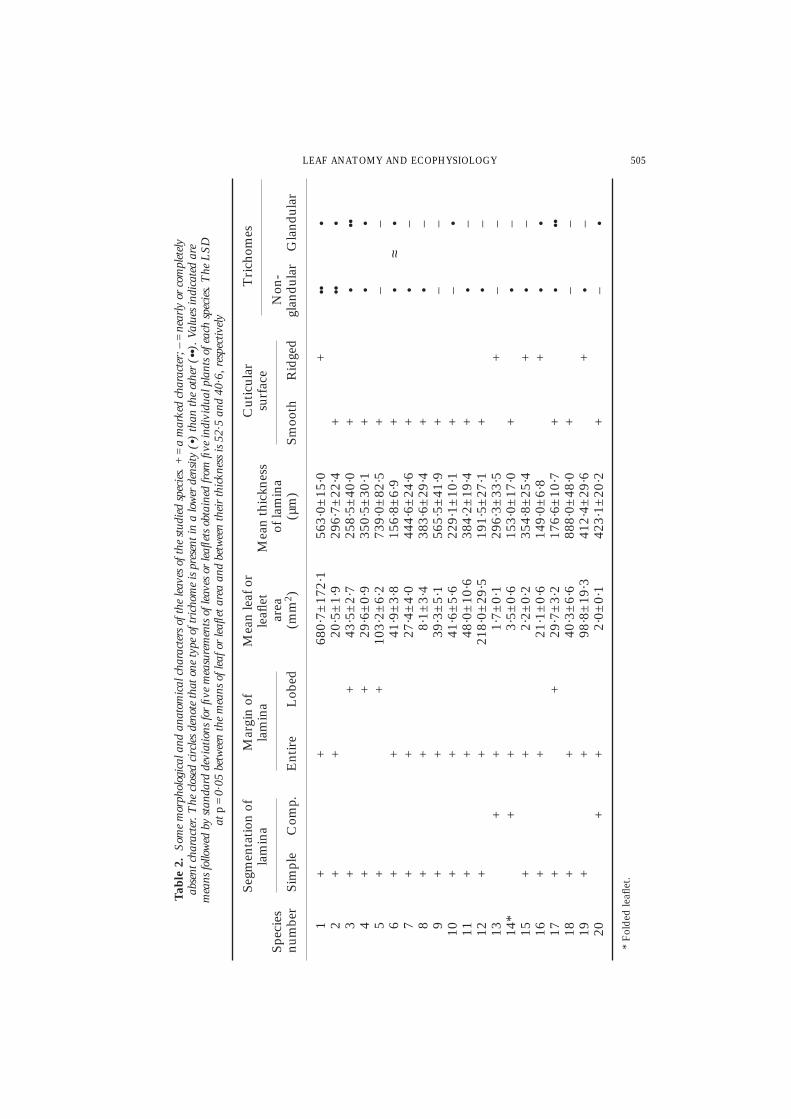
Seventeen of the 20 species investigated have simple leaves (Table 2) with prominentshort petioles. Seriphidium herba-alba, Artemisia judaica and Launea spinosa have deeplydivided leaf laminae (pinnatisect) while the leaves of Salvia deserti are pinnatifid withbullate crenate lobes. Compound leaves occur in Acacia, Astragalus and Fagonia. InAcacia, the leaf is bipinnate, while in Astragalus the leaf is imparipinnate. The leaf ofFagonia is trifoliate. The highest leaf areas are those of Gomphocarpus (680·7mm2 ± 172; mean ± standard deviation, N = 5) and Chrozophora (218 mm2 ± 29·5).The most prominent character of the majority of the studied species is the small leafletor leaflet size (Table 2) which ranges from 2·2 mm2 ± 0·2 in Crotalaria to 98·8mm2 ± 19·3 in Lycium. Folding of leaflets is a character observed in Astragalus (Table2).
The epidermal tissue system
Examination of the outer cuticular surface of the epidermis by SEM indicates that themajority of the pubescent leaves have a smooth or slightly rough appearance. A ridgedouter cuticular surface is observed in Gomphocarpus, Cleome and Salvia (Table 2).
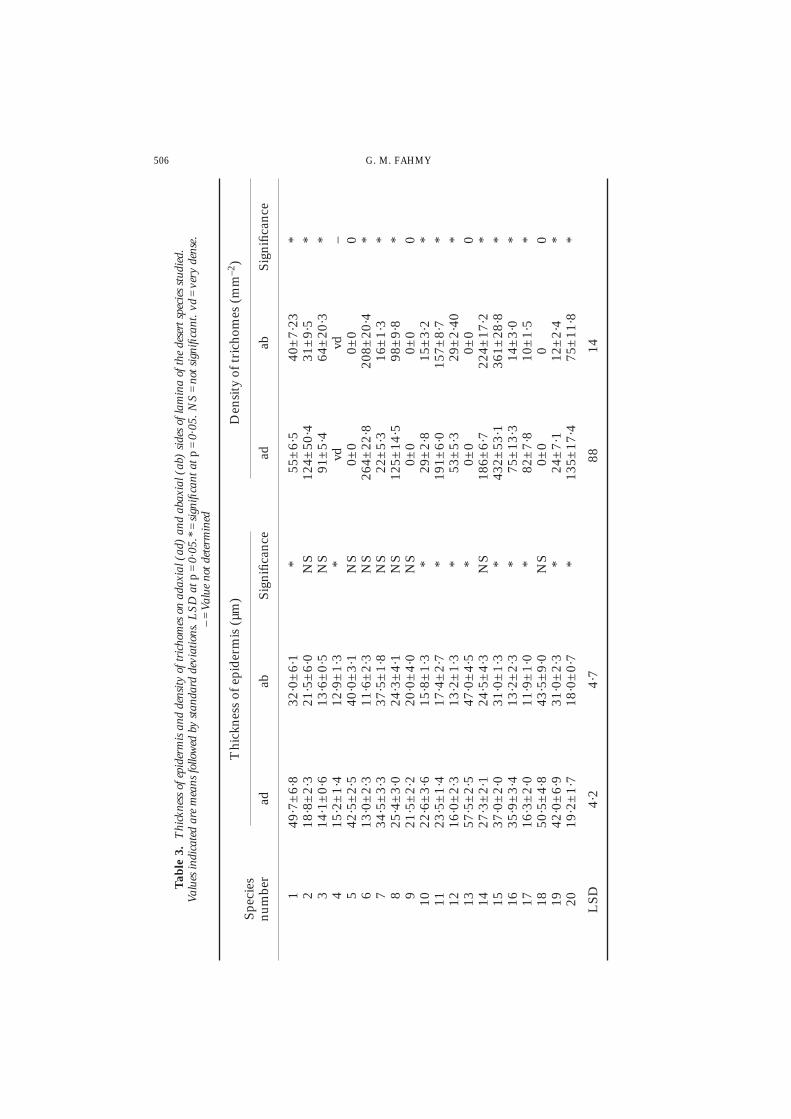
All the species studied have an adaxial and an abaxial uniseriate epidermis. In 10 ofthe 20 species, the thickness of the adaxial epidermis is significantly greater than theabaxial one (Table 3). Species which have thick laminae (Gomphocarpus, Launea andOchradenus) are characterised by a thick epidermis. On the other hand, the thin leafletsof Acacia have a significantly thicker epidermis than the other species.
Epidermal trichomes of non-glandular, glandular or mixed types occur in 16 of the20 species examined (Tables 2 and 3). Trichomes are entirely absent in the cases ofsmall leaflets of Acacia, and in the thick leaves of Launea, Zilla and Ochradenus. Thetrichomes may be very dense and interlocked as in Artemisia judaica (Table 3) or mayoccur in low density as in Moltkiopsis, Cleome and Lycium.
G. M. FAHMY 504
Tab
le 2
.S
ome
mor
phol
ogic
al a
nd a
nato
mic
al c
hara
cter
s of
the
leav
es o
f the
stu
died
spe
cies
. +=
a m
arke
d ch
arac
ter;
–=
near
ly o
r co
mpl
etel
yab
sent
cha
ract
er. T
he c
lose
d ci
rcle
s de
note
that
one
type
of t
rich
ome
is pr
esen
t in
a lo
wer
den
sity
( •)
than
the
othe
r ( •
•). V
alue
s in
dica
ted
are
mea
ns fo
llow
ed b
y st
anda
rd d
evia
tions
for
five
mea
sure
men
ts o
f lea
ves
or le
aflet
s ob
tain
ed fr
om fi
ve in
divi
dual
pla
nts
of e
ach
spec
ies.
The
LS
Dat
p=
0·05
bet
wee
n th
e m
eans
of l
eaf o
r le
aflet
are
a an
d be
twee
n th
eir
thic
knes
s is
52·5
and
40·
6, r
espe
ctiv
ely
Seg
men
tati
on o
fM
argi
n of
Mea
n le
af o
rC
utic
ular
Tri
chom
esla
min
ala
min
ale
aflet
Mea
n th
ickn
ess
surf
ace
Spe
cies
area
of la
min
aN
on-
num
ber
Sim
ple
Com
p.E
ntir
eL
obed
(mm
2 )(µ
m)
Sm
ooth
Rid
ged
glan
dula
rG
land
ular
1+
+68
0·7±
172·
156
3·0±
15·0
+••
•2
++
20·5
±1·
929
6·7±
22·4
+••
•3
++
43·5
±2·
725
8·5±
40·0
+•
••4
++
29·6
±0·
935
0·5±
30·1
+•
•5
++
103·
2±6·
273
9·0±
82·5
+–
–6
++
41·9
±3·
815
6·8±
6·9
+•
ť
7+
+27
·4±
4·0
444·
6±24
·6+
•–
8+
+8·
1±3·
438
3·6±
29·4
+•
–9
++
39·3
±5·
156
5·5±
41·9
+–
–10
++
41·6
±5·
622
9·1±
10·1
+–
•11
++
48·0
±10
·638
4·2±
19·4
+•
–12
++
218·
0±29
·519
1·5±
27·1
+•
–13
++
1·7±
0·1
296·
3±33
·5+
––
14*
++
3·5±
0·6
153·
0±17
·0+
•–
15+
+2·
2±0·
235
4·8±
25·4
+•
–16
++
21·1
±0·
614
9·0±
6·8
+•
•17
++
29·7
±3·
217
6·6±
10·7
+•
••18
++
40·3
±6·
688
8·0±
48·0
+–
–19
++
98·8
±19
·341
2·4±
29·6
+•
–20
++
2·0±
0·1
423·
1±20
·2+
–•
* F
olde
d le
aflet
.
LEAF ANATOMY AND ECOPHYSIOLOGY 505
Tab
le 3
.T
hick
ness
of e
pide
rmis
and
dens
ity o
f tri
chom
es o
n ad
axia
l (ad
) an
d ab
axia
l (ab
) sid
es o
f lam
ina
of th
e de
sert
spe
cies
stu
died
.Va
lues
indi
cate
d ar
e m
eans
follo
wed
by
stan
dard
dev
iatio
ns. L
SD
at p
=0·
05.*
=sig
nific
ant a
t p=
0·05
. NS
=no
t sig
nific
ant.
vd=
very
den
se.
–=
Valu
e no
t det
erm
ined
Thi
ckne
ss o
f epi
derm
is (
µm)
Den
sity
of t
rich
omes
(m
m–2
)S
peci
esnu
mbe
rad
abS
igni
fican
cead
abS
igni
fican
ce
149
·7±
6·8
32·0
±6·
1*
55±
6·5
40±
7·23
*2
18·8
±2·
321
·5±
6·0
NS
124±
50·4
31±
9·5
*3
14·1
±0·
613
·6±
0·5
NS
91±
5·4
64±
20·3
*4
15·2
±1·
412
·9±
1·3
*vd
vd–
542
·5±
2·5
40·0
±3·
1N
S0±
00±
00
613
·0±
2·3
11·6
±2·
3N
S26
4±22
·820
8±20
·4*
734
·5±
3·3
37·5
±1·
8N
S22
±5·
316
±1·
3*
825
·4±
3·0
24·3
±4·
1N
S12
5±14
·598
±9·
8*
921
·5±
2·2
20·0
±4·
0N
S0±
00±
00
1022
·6±
3·6
15·8
±1·
3*
29±
2·8
15±
3·2
*11
23·5
±1·
417
·4±
2·7
*19
1±6·
015
7±8·
7*
1216
·0±
2·3
13·2
±1·
3*
53±
5·3
29±
2·40
*13
57·5
±2·
547
·0±
4·5
*0±
00±
00
1427
·3±
2·1
24·5
±4·
3N
S18
6±6·
722
4±17
·2*
1537
·0±
2·0
31·0
±1·
3*
432±
53·1
361±
28·8
*16
35·9
±3·
413
·2±
2·3
*75
±13
·314
±3·
0*
1716
·3±
2·0
11·9
±1·
0*
82±
7·8
10±
1·5
*18
50·5
±4·
843
·5±
9·0
NS
0±0
00
1942
·0±
6·9
31·0
±2·
3*
24±
7·1
12±
2·4
*20
19·2
±1·
718
·0±
0·7
*13
5±17
·475
±11
·8*
LS
D4·
24·
788
14
G. M. FAHMY 506

In all the species with pubescent leaves, the trichome density on the adaxial side issignificantly higher than that on the abaxial side (Table 3). The outer surface of thenon-glandular trichomes and the multicellular uniseriate stalks of the glandular onesmay show particular sculpturing. Micropapillate surface sculpturing is observed in thetwo armed trichomes of Farsetia, as well as on the surface of the multicellular uniseriateglandular trichomes of Gomphocarpus and Salvia. A smooth surface is observed on theglandular trichomes of Teucrium, Cleome and in Convolvulus and Astragalus which havelong simple unbranched trichomes. In mature, fully expanded sun leaves of the speciesinvestigated, it is found that the protoplasm of the non-glandular trichomes was notdetected and the lumen of the hair cell(s) contains some irregular inclusions. Thus, thenon-glandular trichomes covering the leaf surface of Moltkiopsis, Farsetia, etc., appeardead and air-filled. Conversely, the glandular trichomes covering the mature leaves ofSeriphidium herba-alba, Artemisia judaica, Cleome, Salvia, etc. may have vital intact cellswith rich protoplasm or may appear old with shrunken, collapsed and broken-downwalls.
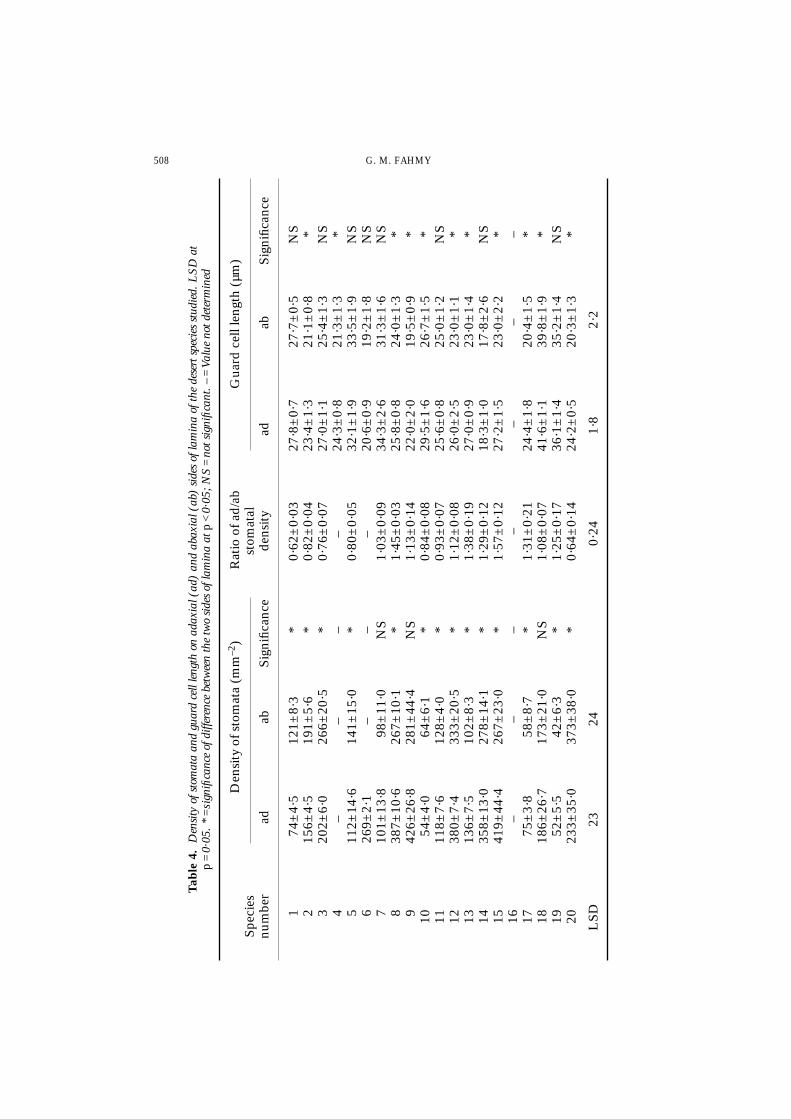
All the species investigated in the present study possess stomata on both surfaces ofthe leaf or leaflet (amphistomatous leaves). The highest densities of stomata occur inZilla and Crotalaria (Table 4). In seven species, the density of stomata is significantlyhigher on the abaxial surface than on the adaxial (Table 4). The rest of the speciesshowed either non-significant differences between the two surfaces (as in Moltkiopsis,Zilla and Ochradenus) or high stomatal density on the adaxial surface (in seven species).In the latter seven species the ratio of adaxial to abaxial stomatal density equals morethan unity. In 11 species, the guard cell length on the adaxial surface is significantlyhigher than that on the abaxial (Table 4).
Thickness of leaf or leaflet and their mesophyll properties
The thickness of leaf or leaflet ranges from 149 µm for Salvia to 888 µm for Ochradenuswhich has the thickest lamina (Table 2). It is apparent that the thickness is largely dueto the presence of palisade tissue which consists of cells arranged on each side, forminga unifacial or isobilateral leaf or leaflet. The number of layers beneath each epidermisis equal in the majority of species, ranging from one or two layers to four layers in Zilla,Ochradenus and Lycium which have thick laminae. In paradermal section, thesubstomatal chambers are cavities bounded by mesophyll cells.
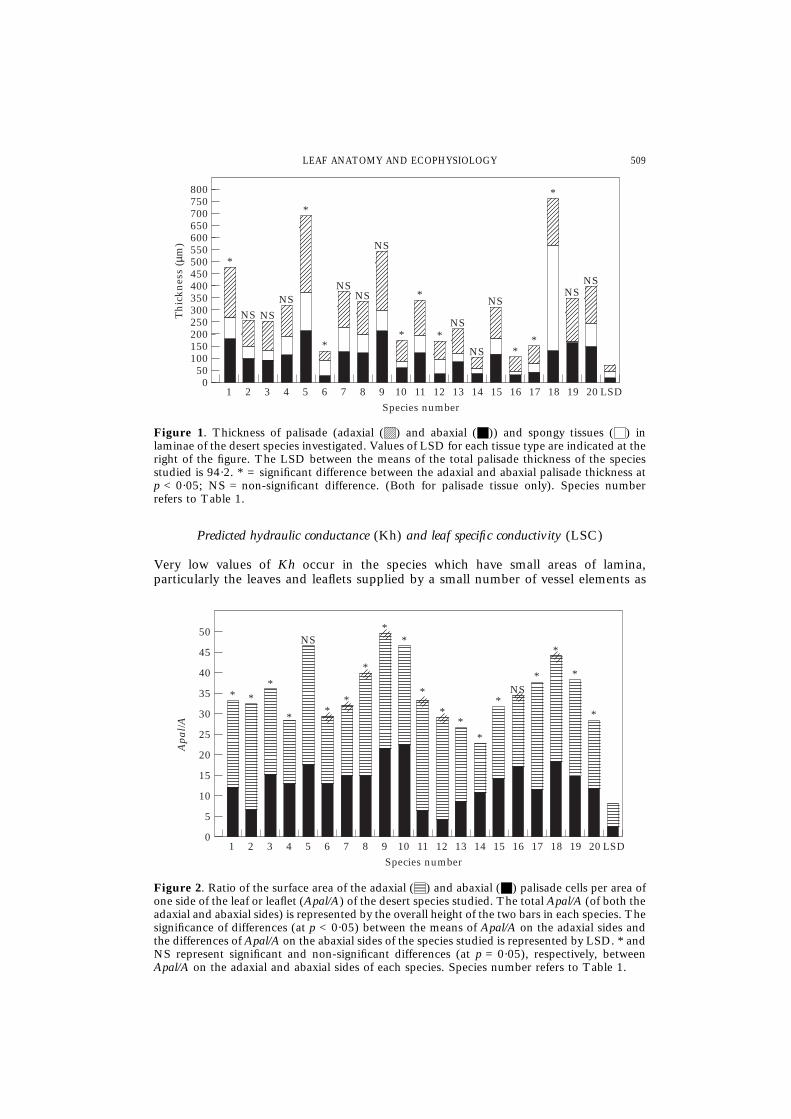
In Salvia, the outermost palisade layers facing the adaxial and abaxial epidermisconsist of cells which often taper towards the epidermis, resulting in a small contactarea. Except for Lycium leaf, the adaxial and abaxial palisade layers enclose a uni- ormulti-layered spongy tissue of rounded (in cross-section) compact cells with smallintercellular spaces. The ratio of thickness of palisade layer/spongy layer ranges from0·7 for the thick leaf of Ochradenus to 6·5 for Salvia (Fig. 1) which has the thinnestlamina.
In the leaves of 11 species, there are no statistically significant differences betweenthe thickness of adaxial and abaxial palisade (Fig. 1). The rest of the species showedsignificantly greater thickness of palisade adaxially than abaxially. The total palisadethickness follows a similar trend to that of the lamina thickness, being high in the thickleaves (Table 2, Fig. 1).
For 18 species the ratio of Apal/A adaxially is significantly higher than abaxially (Fig.2). Total Apal/A of both adaxial and abaxial palisade regions ranges from the lowestvalues in the thin leaflets of Astragalus (Apal/A = 22·6) to the highest values in thecases of Launea (46·7), Zilla (49·6) and Ochradenus (43·8, LSD = 11·8) leaves whichhave thick blades (Table 2, Fig. 2). It is worth indicating that Cleome has a low totalpalisade thickness of 148·2 µm (Fig. 1) and an Apal/A of 46·7 (Fig. 2), while Launeahas an Apal/A of 46·7 and a significantly higher palisade thickness (528·4 µm).
LEAF ANATOMY AND ECOPHYSIOLOGY 507
Tab
le 4
.D
ensit
y of
sto
mat
a an
d gu
ard
cell
leng
th o
n ad
axia
l (ad
) an
d ab
axia
l (ab
) sid
es o
f lam
ina
of th
e de
sert
spe
cies
stu
died
. LS
D a
tp
=0·
05. *
=si
gnifi
canc
e of
diff
eren
ce b
etw
een
the
two
sides
of l
amin
a at
p<
0·05
; NS
=no
t sig
nific
ant.
–=
Valu
e no
t det
erm
ined
Den
sity
of s
tom
ata
(mm
–2)
Rat
io o
f ad/
abG
uard
cel
l len
gth
(µm
)S
peci
esst
omat
alnu
mbe
rad
abS
igni
fican
cede
nsit
yad
abS
igni
fican
ce
174
±4·
512
1±8·
3*
0·62
±0·
0327
·8±
0·7
27·7
±0·
5N
S2
156±
4·5
191±
5·6
*0·
82±
0·04
23·4
±1·
321
·1±
0·8
*3
202±
6·0
266±
20·5
*0·
76±
0·07
27·0
±1·
125
·4±
1·3
NS
4–
––
–24
·3±
0·8
21·3
±1·
3*
511
2±14
·614
1±15
·0*
0·80
±0·
0532
·1±
1·9
33·5
±1·
9N
S6
269±
2·1
––
–20
·6±
0·9
19·2
±1·
8N
S7
101±
13·8
98±
11·0
NS
1·03
±0·
0934
·3±
2·6
31·3
±1·
6N
S8
387±
10·6
267±
10·1
*1·
45±
0·03
25·8
±0·
824
·0±
1·3
*9
426±
26·8
281±
44·4
NS
1·13
±0·
1422
·0±
2·0
19·5
±0·
9*
1054
±4·
064
±6·
1*
0·84
±0·
0829
·5±
1·6
26·7
±1·
5*
1111
8±7·
612
8±4·
0*
0·93
±0·
0725
·6±
0·8
25·0
±1·
2N
S12
380±
7·4
333±
20·5
*1·
12±
0·08
26·0
±2·
523
·0±
1·1
*13
136±
7·5
102±
8·3
*1·
38±
0·19
27·0
±0·
923
·0±
1·4
*14
358±
13·0
278±
14·1
*1·
29±
0·12
18·3
±1·
017
·8±
2·6
NS
1541
9±44
·426
7±23
·0*
1·57
±0·
1227
·2±
1·5
23·0
±2·
2*
16–
––
––
––
1775
±3·
858
±8·
7*
1·31
±0·
2124
·4±
1·8
20·4
±1·
5*
1818
6±26
·717
3±21
·0N
S1·
08±
0·07
41·6
±1·
139
·8±
1·9
*19
52±
5·5
42±
6·3
*1·
25±
0·17
36·1
±1·
435
·2±
1·4
NS
2023
3±35
·037
3±38
·0*
0·64
±0·
1424
·2±
0·5
20·3
±1·
3*
LS
D23
240·
241·
82·
2
G. M. FAHMY 508
800
01
Species number
Th
ickn
ess
(µm
)
50
700
600
400
300
200150100
350
250
750
650
450500550
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 LSD
*
NSNS
*
*
NSNS
NS
*
*
*
NS
NS
NS
**
*
NSNS
NS
01
Species number
Apa
l/A
5
40
30
20
15
10
35
25
45
50
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 LSD
**
*
NS
**
*
**
*
**
*
*NS
*
*
*
**
Predicted hydraulic conductance (Kh) and leaf specific conductivity (LSC)
Very low values of Kh occur in the species which have small areas of lamina,particularly the leaves and leaflets supplied by a small number of vessel elements as
Figure 1. Thickness of palisade (adaxial (C) and abaxial (G)) and spongy tissues (F) inlaminae of the desert species investigated. Values of LSD for each tissue type are indicated at theright of the figure. The LSD between the means of the total palisade thickness of the speciesstudied is 94·2. * = significant difference between the adaxial and abaxial palisade thickness atp < 0·05; NS = non-significant difference. (Both for palisade tissue only). Species numberrefers to Table 1.
Figure 2. Ratio of the surface area of the adaxial (E) and abaxial (G) palisade cells per area ofone side of the leaf or leaflet (Apal/A) of the desert species studied. The total Apal/A (of both theadaxial and abaxial sides) is represented by the overall height of the two bars in each species. Thesignificance of differences (at p < 0·05) between the means of Apal/A on the adaxial sides andthe differences of Apal/A on the abaxial sides of the species studied is represented by LSD. * andNS represent significant and non-significant differences (at p = 0·05), respectively, betweenApal/A on the adaxial and abaxial sides of each species. Species number refers to Table 1.
LEAF ANATOMY AND ECOPHYSIOLOGY 509

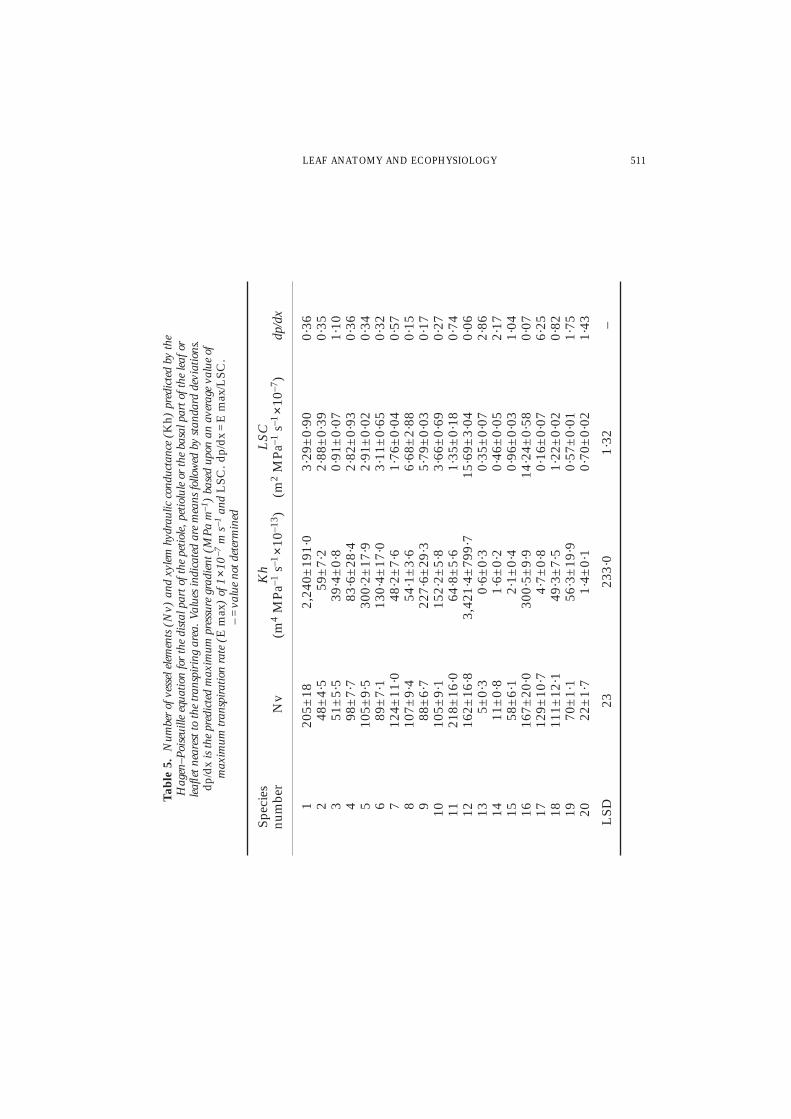
found in the leaflets of Acacia, Astragalus and Fagonia and in the leaves of Achillea(Tables 2 and 5). Although the number of vessel elements in the petioles ofChrozophora and Salvia is nearly equal, the Kh of the former is about 11 times higherthan that of the latter (Table 5). Such variation in Kh of those species is also valid fortheir LSC but with narrow difference where the highest value of LSC obtained inChrozophora (15·69 m2 MPa–1 s–1 3 10–7) is slightly higher than that of Salvia (14·20m2 MPa–1 s–1 3 10–7, LSD = 1·32). In other words, the wide difference of Khbetween the two species is nearly cancelled in the cases of their LSC (Table 5). Thisis attributed to their leaf area which is 10·3 times higher in Chrozophora than in Salvia(Table 2). Such explanation is also valid for Gomphocarpus which shows a low LSC(3·29 m2 MPa–1 s–1 3 10–7) despite the high value of its Kh and the high number ofvessel elements in its petiole.
Correlations
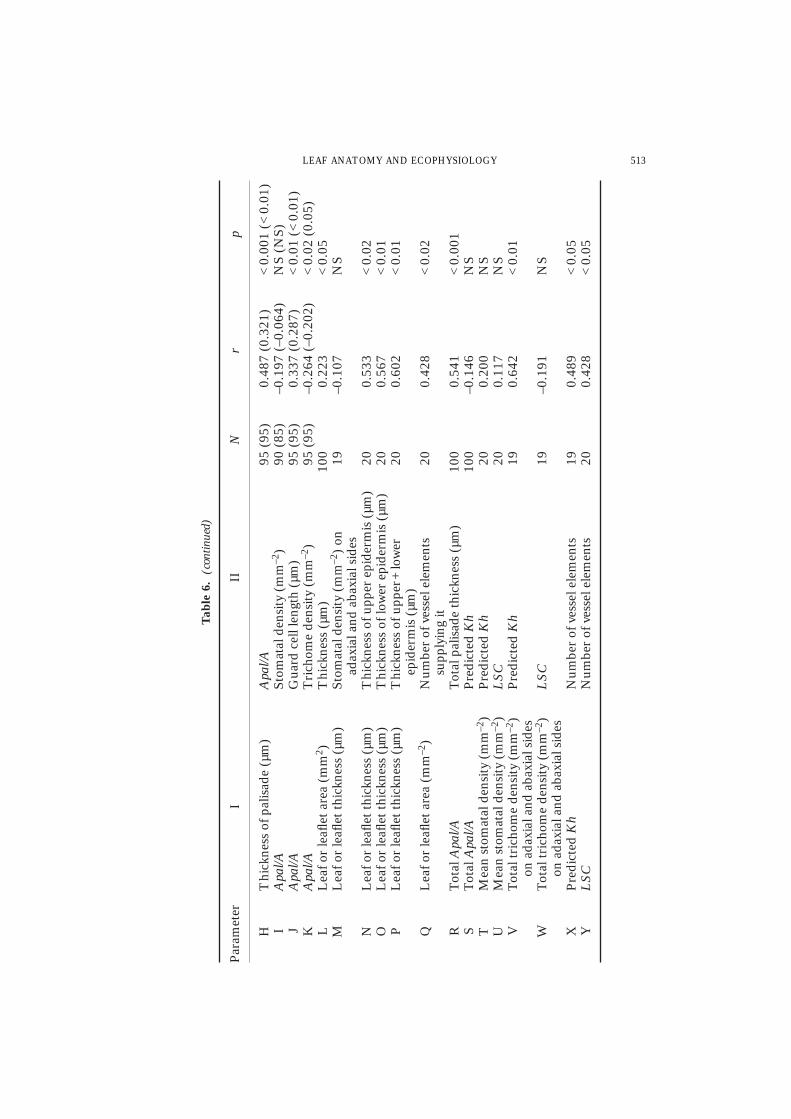
On both leaf surfaces significant correlations existed between the majority ofanatomical variables (Table 6). On the adaxial surface, the percentage of significantcorrelations is lower (64%) than that on the abaxial one (73%). On both surfaces, theincrease in stomatal density is paralleled by an increase in trichome density. However,the correlation coefficient is relatively low but still significant (r = 0·259, N = 85,p < 0·02) for the abaxial surface. Negative correlations exist between stomatal densityand guard cell length. Correlation of such variables is higher on the adaxial surface(r = –0·440, N = 90) than on the abaxial one (r = –0·298, N = 85). Trichome densityon the adaxial surface correlates negatively with the thickness of epidermis(r = –0·283, N = 95, p < 0·01).
The relationship between palisade layer thickness and the measured variables shownin Table 6 (from D to H) is more significant from the abaxial surface than on theadaxial one. However, when the significant relationships are compared on both sides,the values of the correlation coefficients on the abaxial surfaces are relatively low. Onthe other hand, the characteristic most highly correlated with the palisade thickness isApal/A on the adaxial surface (r = 0·487, N = 95, p < 0·001). Guard cell lengthappears to be the variable most highly correlated with the thickness of abaxial palisade(r = 0·414, N = 95, p < 0·001) and with Apal/A on the adaxial side (r = 0·337,p < 0·01).
Apal/A is most highly correlated with total palisade thickness (r = 0·541, N = 100,p < 0·0001) and with the thickness of lamina (r = 525, Fig. 3). Measurements of leafthickness and/or total palisade thickness are therefore an easy means of estimatingApal/A. The correlation coefficient between the laminal area and its thickness isrelatively low (r = 0·223) but still significant (N = 100, p < 0·05, Table 6).
The leaf or leaflet thickness is more highly correlated with the total thickness ofadaxial and abaxial epidermises (r = 0·602, N = 20, p < 0·01) than with the thicknessof either of them singly.
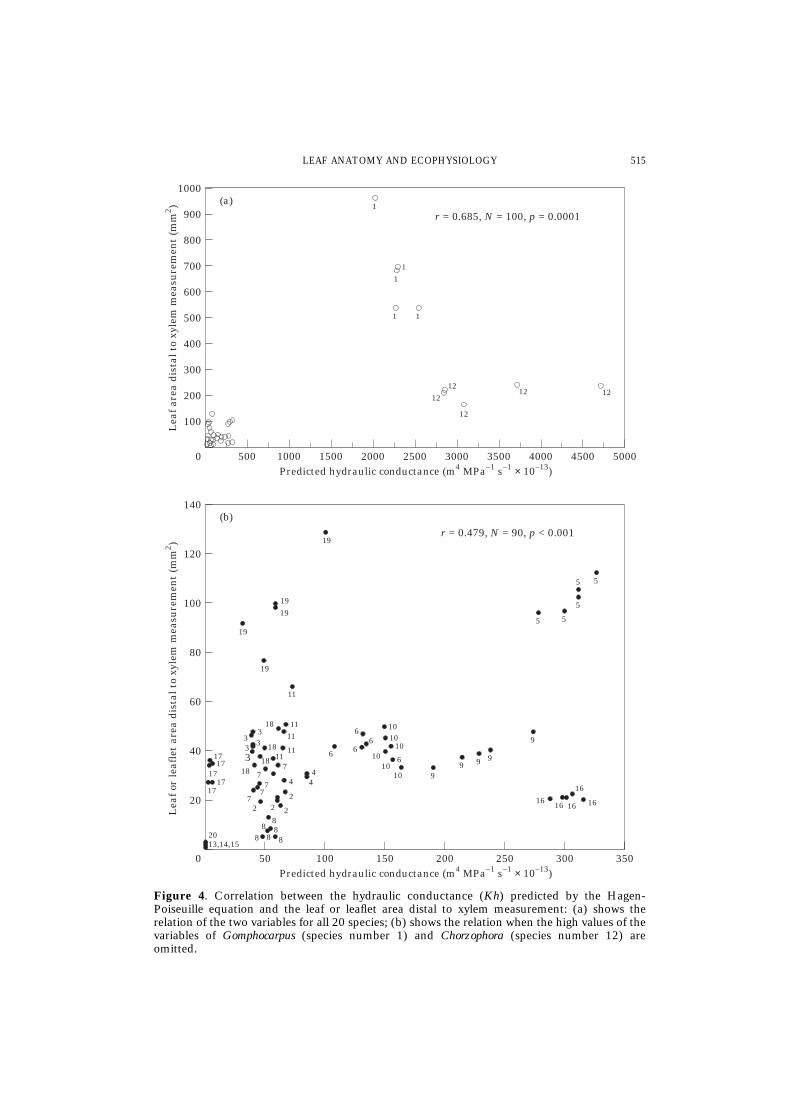
The predicted Kh correlates very well with the leaf or leaflet area supplied by thepetioles or petiolules (Fig. 4(a, b)). When considering the 20 species, a significantcorrelation coefficient is obtained (r = 0·685, N = 100, p < 0·0001) but the pointsplotted for 18 species cluster and overlap each other at the lower left corner of Fig.4(a). At the upper right corner, the high values of the variables of Gomphocarpus andChrozophora show greater departures than those of the rest of the species. Because ofthis, a correlation analysis was done for 18 species excluding Gomphocarpus andChrozophora. Still, the predicted Kh is correlated with the area supplied (r = 0·749,N = 90, p < 0·001, Fig. 4(b)). The second characteristic correlated with the predictedKh is the total trichome density (mm–2) of adaxial and abaxial sides (r = 0·642,N = 20, p < 0·01). There is no correlation between the values of the predicted Kh and
G. M. FAHMY 510
Tab
le 5
.N
umbe
r of
ves
sel e
lem
ents
(N
v) a
nd x
ylem
hyd
raul
ic c
ondu
ctan
ce (
Kh)
pre
dict
ed b
y th
eH
agen
–Poi
seui
lle e
quat
ion
for
the
dist
al p
art o
f the
pet
iole
, pet
iolu
le o
r th
e ba
sal p
art o
f the
leaf
or
leafl
et n
eare
st to
the
tran
spir
ing
area
. Val
ues
indi
cate
d ar
e m
eans
follo
wed
by
stan
dard
dev
iatio
ns.
dp/d
xis
the
pred
icte
d m
axim
um p
ress
ure
grad
ient
(M
Pa m
–1)
base
d up
on a
n av
erag
e va
lue
ofm
axim
um tr
ansp
iratio
n ra
te (
E m
ax)
of 1
×10
–7m
s–1
and
LS
C. d
p/dx
=E
max
/LS
C.
–=
valu
e no
t det
erm
ined
Spe
cies
Kh
LS
Cnu
mbe
rN
v(m
4M
Pa–
1s–
1×1
0–13
)(m
2M
Pa–
1s–
1×1
0–7 )
dp/d
x
120
5±18
2,24
0±19
1·0
3·29
±0·
900·
362
48±
4·5
59±
7·2
2·88
±0·
390·
353
51±
5·5
39·4
±0·
80·
91±
0·07
1·10
498
±7·
783
·6±
28·4
2·82
±0·
930·
365
105±
9·5
300·
2±17
·92·
91±
0·02
0·34
689
±7·
113
0·4±
17·0
3·11
±0·
650·
327
124±
11·0
48·2
±7·
61·
76±
0·04
0·57
810
7±9·
454
·1±
3·6
6·68
±2·
880·
159
88±
6·7
227·
6±29
·35·
79±
0·03
0·17
1010
5±9·
115
2·2±
5·8
3·66
±0·
690·
2711
218±
16·0
64·8
±5·
61·
35±
0·18
0·74
1216
2±16
·83,
421·
4±79
9·7
15·6
9±3·
040·
0613
5±0·
30·
6±0·
30·
35±
0·07
2·86
1411
±0·
81·
6±0·
20·
46±
0·05
2·17
1558
±6·
12·
1±0·
40·
96±
0·03
1·04
1616
7±20
·030
0·5±
9·9
14·2
4±0·
580·
0717
129±
10·7
4·7±
0·8
0·16
±0·
076·
2518
111±
12·1
49·3
±7·
51·
22±
0·02
0·82
1970
±1·
156
·3±
19·9
0·57
±0·
011·
7520
22±
1·7
1·4±
0·1
0·70
±0·
021·
43
LS
D23
233·
01·
32–
LEAF ANATOMY AND ECOPHYSIOLOGY 511

Tab
le 6
.C
orre
latio
n co
effic
ient
s am
ong
som
e an
atom
ical
par
amet
ers
(I &
II)
of t
he le
aves
and
leafl
ets
of th
e st
udie
d de
sert
spe
cies
. The
num
ber
of o
bser
vatio
ns (
N),
the
corr
elat
ion
coef
ficie
nt (
r) a
nd th
e pr
obab
ility
leve
l (p)
for
each
pai
r of
par
amet
ers
(I &
II)
in th
e se
t fro
m A
to K
are
thos
e fo
r th
e ad
axia
l sid
e fo
llow
ed b
y th
e ab
axia
l one
whi
ch is
incl
uded
bet
wee
n pa
rent
hese
s. A
pal/A
=th
e ra
tio o
f tot
al a
rea
of p
alisa
de c
ells
to le
afar
ea, K
h=
hydr
aulic
con
duct
ance
(in
m4
MPa
–1s–
1 ×10
–13 )
, LS
C=
leaf
or
leafl
et s
peci
fic c
ondu
ctan
ce (
in m
2M
Pa–1
s–1×1
0–7
).N
S=
not s
igni
fican
t at t
he 0
·05
leve
l of p
roba
bilit
y
Par
amet
erI
IIN
rp
AS
tom
atal
den
sity
(m
m–2
)T
rich
ome
dens
ity
(mm
–2)
90 (
85)
0.47
5 (0
.259
)<
0.00
1 (<
0.02
)B
Sto
mat
al d
ensi
ty (
mm
–2)
Gua
rd c
ell l
engt
h (µ
m)
90 (
85)
–0.4
40 (
–0.2
98)
<0.
001
(<0.
01)
CT
rich
ome
dens
ity
(mm
–2)
Thi
ckne
ss o
f epi
derm
is (
µm)
95 (
95)
–0.2
83 (
–0.1
31)
<0.
01 (
NS
)D
Thi
ckne
ss o
f pal
isad
e (µ
m)
Thi
ckne
ss o
f epi
derm
is (
µm)
100
(100
)0.
077
(0.4
40)
NS
(<
0.00
1)E
Thi
ckne
ss o
f pal
isad
e (µ
m)
Sto
mat
al d
ensi
ty (
mm
–2)
90 (
85)
–0.1
51 (
0.04
9)N
S (
NS
)F
Thi
ckne
ss o
f pal
isad
e (µ
m)
Gua
rd c
ell l
engt
h (µ
m)
95 (
95)
0.11
8 (0
.414
)N
S (
<0.
001)
GT
hick
ness
of p
alis
ade
(µm
)T
rich
ome
dens
ity
(mm
–2)
95 (
95)
–0.3
57 (
–0.2
31)
<0.
001
(<0.
05)
G. M. FAHMY 512
Tab
le 6
.(c
ontin
ued)
Par
amet
erI
IIN
rp
HT
hick
ness
of p
alis
ade
(µm
)A
pal/A
95 (
95)
0.48
7 (0
.321
)<
0.00
1 (<
0.01
)I
Apa
l/AS
tom
atal
den
sity
(m
m–2
)90
(85
)–0
.197
(–0
.064
)N
S (
NS
)J
Apa
l/AG
uard
cel
l len
gth
(µm
)95
(95
)0.
337
(0.2
87)
<0.
01 (
<0.
01)
KA
pal/A
Tri
chom
e de
nsit
y (m
m–2
)95
(95
)–0
.264
(–0
.202
)<
0.02
(0.
05)
LL
eaf o
r le
aflet
are
a (m
m2 )
Thi
ckne
ss (
µm)
100
0.22
3<
0.05
ML
eaf o
r le
aflet
thi
ckne
ss (
µm)
Sto
mat
al d
ensi
ty (
mm
–2)
on19
–0.1
07N
Sad
axia
l and
aba
xial
sid
esN
Lea
f or
leafl
et t
hick
ness
(µm
)T
hick
ness
of u
pper
epi
derm
is (
µm)
200.
533
<0.
02O
Lea
f or
leafl
et t
hick
ness
(µm
)T
hick
ness
of l
ower
epi
derm
is (
µm)
200.
567
<0.
01P
Lea
f or
leafl
et t
hick
ness
(µm
)T
hick
ness
of u
pper
+lo
wer
200.
602
<0.
01ep
ider
mis
(µm
)Q
Lea
f or
leafl
et a
rea
(mm
–2)
Num
ber
of v
esse
l ele
men
ts20
0.42
8<
0.02
supp
lyin
g it
RT
otal
Apa
l/AT
otal
pal
isad
e th
ickn
ess
(µm
)10
00.
541
<0.
001
ST
otal
Apa
l/AP
redi
cted
Kh
100
–0.1
46N
ST
Mea
n st
omat
al d
ensi
ty (
mm
–2)
Pre
dict
ed K
h20
0.20
0N
SU
Mea
n st
omat
al d
ensi
ty (
mm
–2)
LS
C20
0.11
7N
SV
Tot
al t
rich
ome
dens
ity
(mm
–2)
Pre
dict
ed K
h19
0.64
2<
0.01
on a
daxi
al a
nd a
baxi
al s
ides
WT
otal
tri
chom
e de
nsit
y (m
m–2
)L
SC
19–0
.191
NS
on a
daxi
al a
nd a
baxi
al s
ides
XP
redi
cted
Kh
Num
ber
of v
esse
l ele
men
ts19
0.48
9<
0.05
YL
SC
Num
ber
of v
esse
l ele
men
ts20
0.42
8<
0.05
LEAF ANATOMY AND ECOPHYSIOLOGY 513
50
1000
0 20Total Apal/A
Th
ickn
ess
of le
af o
r le
afle
t (µ
m)
500
900
800
700
600
400
300
200
100
30 40
r = 0.525, N = 100, p = 0.0001
2020
2020
201314
4 154
15
4 4
15
15
15
13 13
13 4
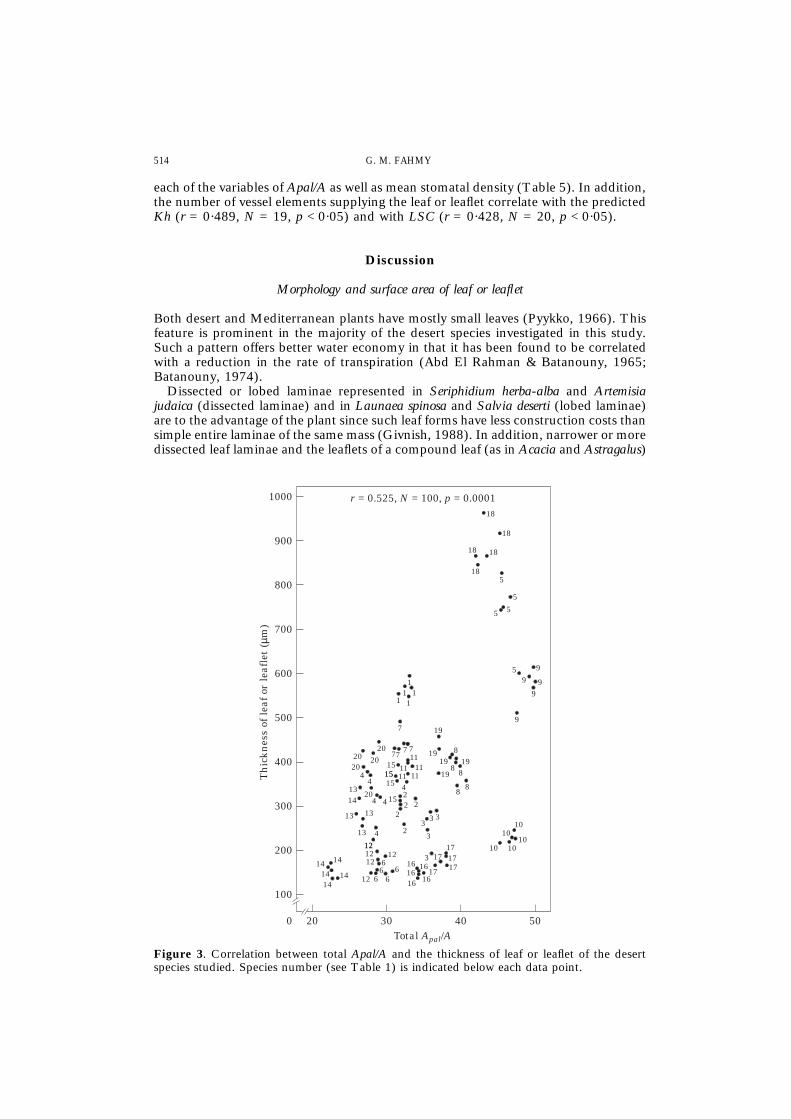
4
2
2
1212 12
2
12
22
333
3
3
8
171717
17
1712
126
66
6 6 1616
16 1616
1414
1414
14
10
1010 10
10
7
7
77
1111
111111
19
1919
19
19
8
88
87
11 11
1
5
5
5
5
18
18
18
18 18
59
9
99
9
each of the variables of Apal/A as well as mean stomatal density (Table 5). In addition,the number of vessel elements supplying the leaf or leaflet correlate with the predictedKh (r = 0·489, N = 19, p < 0·05) and with LSC (r = 0·428, N = 20, p < 0·05).
Discussion
Morphology and surface area of leaf or leaflet
Both desert and Mediterranean plants have mostly small leaves (Pyykko, 1966). Thisfeature is prominent in the majority of the desert species investigated in this study.Such a pattern offers better water economy in that it has been found to be correlatedwith a reduction in the rate of transpiration (Abd El Rahman & Batanouny, 1965;Batanouny, 1974).
Dissected or lobed laminae represented in Seriphidium herba-alba and Artemisiajudaica (dissected laminae) and in Launaea spinosa and Salvia deserti (lobed laminae)are to the advantage of the plant since such leaf forms have less construction costs thansimple entire laminae of the same mass (Givnish, 1988). In addition, narrower or moredissected leaf laminae and the leaflets of a compound leaf (as in Acacia and Astragalus)
Figure 3. Correlation between total Apal/A and the thickness of leaf or leaflet of the desertspecies studied. Species number (see Table 1) is indicated below each data point.
G. M. FAHMY 514
5000
1000
0 500Predicted hydraulic conductance (m4 MPa–1 s–1 × 10–13)
Lea
f ar
ea d
ista
l to
xyle
m m
easu
rem
ent
(mm
2 )
500
900
800
700
600
400
300
200
100
3000 4000
r = 0.685, N = 100, p = 0.0001
12
(a)
45003500250020001000 1500
1212
12
12
1 1
11
1
350
140
0 50Predicted hydraulic conductance (m4 MPa–1 s–1 × 10–13)
Lea
f or
leaf
let
area
dis
tal t
o xy
lem
mea
sure
men
t (m
m2 )
120
80
60
40
20
300
(b)
250200100 150
100
20 88
8
88
2
22 2
4
7
777
71818
18
18 1111
1111
1717
17
1717
33
33
34
4
1010
10
1010
106
66
6
9
69 9 9
9
1616 1616
16
5 5
5 5
5
11
19
19
19
19
19
13,14,158
r = 0.479, N = 90, p < 0.001
Figure 4. Correlation between the hydraulic conductance (Kh) predicted by the Hagen-Poiseuille equation and the leaf or leaflet area distal to xylem measurement: (a) shows therelation of the two variables for all 20 species; (b) shows the relation when the high values of thevariables of Gomphocarpus (species number 1) and Chorzophora (species number 12) areomitted.
LEAF ANATOMY AND ECOPHYSIOLOGY 515

have thinner air boundary layers, maximizing heat convection and thus coupling leaftemperature closely to ambient air temperature (Lewis, 1972; Nobel, 1975).Minimizing the area of the transpiring surface by folding of leaflets is a featurerepresented in Astragalus. Six species of the desert plants studied are characterised byspine formation, a feature which can reduce the transpiring surface. Shoots (Zilla,Convolvulus and Lycium), axes of the compound pinnate leaves (Astragalus) or stipules(Acacia and Fagonia) are metamorphosed into spines.
The high leaf area of Gomphocarpus sinaicus possibly reflects its natural distributionin a less arid habitat than that of the other 19 species; it grows in the mountainousregions of southern Sinai where the conditions are wetter than in the desert (Zahran &Willis, 1992).
Epidermal tissue system
Reduction of leaf absorptance over the entire solar band (400–3000 nm) by increasingsurface reflectance through epidermal modifications is one of the main factors in thereduction of leaf temperature in plants from arid habitats (Ehleringer, 1981). Thesmooth or slightly rough outer cuticular surface is a feature observed by SEM in themajority of the non-pubescent species of this study. This is suggested to play a role inreflecting less than 10% of the incident light (McClendon, 1984). A combination oflight reflectance and water and secretion repellency of the surface are assumed to beachieved by the presence of trichomes and cuticular ridges on the pubescent leaves,e.g. Gomphocarpus, Cleome and Salvia. Juniper & Jeffree (1983) indicate that thepresence of ornamented cuticle results in an increased contact angle where the droplet(in the present case it may be water or a fluid excreted by glandular trichomes) is heldaway from the surface. It is suggested that the possible occurrence of the above-mentioned phenomenon in desert species having glandular leaf surface may protect thestomatal pores from clogging by the excretion of the glands.
The majority of the desert species of this investigation (16 out of 20) have pubescentleaves. Largely based on previous ecological studies, it is apparent that leaf hairs areadaptive features of plants to arid conditions (Fahn & Culter, 1992). A characteristicfeature of the pubescent species of this study is the predominance of those havingpurely non-glandular trichomes or a mixture of glandular and non-glandular types.Reflectance of solar radiation in the range 700–3000 nm wave band (near infraredradiation) increases when the trichomes are non-glandular or air-filled (Ehleringer &Björkman, 1978). This will directly reduce the heat load and consequently leaftemperature. The occurrence of a statistically significant higher trichome density onthe adaxial leaf side than on the abaxial one increases the reflectance on the upper sidewhich is more exposed to intensive solar radiation. On the lower side, the trichomesmay act as a near infrared reflector (Larcher, 1983, p. 10). An exception is found in theleaflets of Astragalus where the trichome density in the upper side is significantly lowerthan on the lower side. This may be attributed to leaflet folding, where the upper sideis less exposed.
The occurrence of cuticular covering on the outer surface of the epidermis, and theglandular and non-glandular trichomes of the desert species investigated is acharacteristic feature (Volkens, 1887, pp. 45–46; Uphof, 1962). This cuticularcovering is considered as a mechanism to minimize transpirational water loss throughthe epidermis and trichomes.
The occurrence of thickened anticlinal walls in the basal cells of the non-glandulartrichomes and in the stalk cells of the glandular ones is observed by Fahn (1986) in thetrichomes of some xeromorphic leaves. He considered these thickenings to be similarto endodermal cells in preventing apoplastic water flow into the trichomes. In deadtrichomes (as the non-glandular trichomes in the desert species studied) evaporation of
G. M. FAHMY 516

water from their walls is thus avoided (Fahn & Cutler, 1992, pp. 62–63). Lightmicroscope observations indicate that in the leaves with thick indumentum, the hairsare dead and air-filled (Ehleringer, 1984).
The presence of trichomes, even as a thick layer, have been considered as exertinglittle effect on the boundary layer resistance to water vapour loss (Gibson, 1983;Ehleringer, 1984). Boundary layer resistance due to hairs is commonly quantified bydividing the thickness of the pubescent layer (expressed in m) by the diffusioncoefficient of water vapour in the air (Nobel, 1991). Let us examine the thickness ofthe indumentum on leaves of varying pubescence. We will consider the highlypubescent leaf of Moltkiopsis in which the trichome thickness reaches 200 µm ± 18(N = 15) and the low pubescent leaf of Lycium in which the height of the pubescentlayer is 33 µm ± 4·7 (N = 15) (data are not shown in this study). Assuming low windvelocity (0·1 m s–1) and that the characteristic leaf dimension in the direction of thewind is 0·004 m for Moltkiopsis and 0·006 m for Lycium, and using the diffusioncoefficient of water vapour under desert conditions at 30°C (2·57 3 10–5 m2 s–1), theresistance to water vapour diffusion caused by the hairs is 7·8 s m–1 for Moltkiopsis andonly 0·9 s m–1 for Lycium. These calculated values represent almost 25·1 and 2·4% ofthe boundary layer resistance of Moltkiopsis and Lycium, respectively. In xerophyteswith open stomata, the boundary layer resistances are smaller than their respectivevalues for diffusion along stomatal pores which ranged from 250 to 1000 s m–1 (Nobel,1991). Previous studies on non-succulent desert perennials of North America indicatethat the boundary layer resistance due to trichomes covering the stem of Salazariamixicana Torr, and the leaf of Encelia farinosa Gray ranges from 4 s m–1 for a hair layerthickness of 110 µm in the former (Gibson, 1983) to c. 17 s m–1 for a hair layerthickness of 400 µm in the latter (Ehleringer, 1984). Functionally, the effect ofboundary layer resistance due to hair cover on transpiration will be very small(Ehleringer, 1984; Nobel, 1991).
The thin leaves or leaflets of the desert species studied are characterised by relativelysmall palisade thickness and consequently low Apal/A. Under desert conditions, it isbelieved that a thin non-pubescent leaf will possibly be vulnerable to wilting. Thisimplies the importance of dense cover of trichomes particularly on the thin leaves. Onthe contrary, it is suggested that the high palisade packing of the thick glabrous leavesprotects them from rapid wilting during water stress. These interpretations aresupported by the significant negative correlations between trichome density and bothpalisade thickness and Apal/A.
The existence of a significant negative correlation (r = –0·283, N = 95, p < 0·01)between trichome density on the adaxial side and thickness of the epidermis indicatesthat the epidermal cells may be considered as a buffer layer. They may partiallycontribute to the reflectance properties of pubescent leaves, or totally perform a dualfunction (reflection and absorption) in the case of the relatively thick glabrous leaves.This conclusion is further supported by the significant positive correlations (atp < 0·02) between thicknesses of the leaf and epidermis (adaxial, abaxial and both). Insuch glabrous leaves, the epidermis consists of thick uniseriate cells with high watercontent. At the near infrared radiation (1300–3000 nm), absorption by water increases(Carter, 1991), leading to the thermal effect (Larcher, 1983). Since the possibility ofheat storage increases in thick fleshy organs (cf. Lewis & Nobel, 1977), it wouldtherefore be to the advantage of the relatively thick glabrous leaves of Launaea, Zillaand Ochradenus to increase the absorption of the near infrared radiation by their thickepidermal cells before the radiation transmits to the internal mesophyll of the leaf.
The feature of amphistomatic leaves observed in the species of the presentinvestigation is characteristic of plants growing in strongly lighted habitats. In NorthAmerican deserts, over 90% of the species have amphistomatic leaves (Mott et al.,1982). The significant negative correlation between stomatal density and guard celllength of the investigated desert species is a characteristic feature (Willmer, 1983).
LEAF ANATOMY AND ECOPHYSIOLOGY 517

Sundberg (1986) found a similar relationship in non-succulent species in the desertsof North America. This relationship between stomatal density and guard cell lengthwill result in low diffusive resistance ( = high stomatal conductance) and consequentlyhigher rates of transpiration and photosynthesis (Stocker, 1976). The significantpositive relationship between Apal/A and the length of the guard cell indicates that theleaf is built for maximum photosynthetic efficiency; a suggestion that is supported bytheoretical and physical considerations (Nobel & Walker, 1985).
When the anatomical measurements of Zilla and Launaea of this study are combinedwith their transpiration measurements given by Abd El Rahman et al. (1966, 1976) forLaunaea and for Zilla it is found that although Launaea has a high guard cell length(33·1 µm) it had a lower transpiration rate (280 mg g–1 h–1) than Zilla (400 mg g–1 h–1)owing to its low stomatal density (100 mm–2). Preliminary assessments indicate thatthe mean maximum width of elliptical pores of Zilla and Launaea is 6 µm. It is evidentthat the perimeter of the stomatal opening of Launaea would be higher than that ofZilla owing to the larger guard cell length of the former than the latter. According toBrown & Escombe (1900) and Sayer (1926) the rate of diffusion of water vapourthrough smaller pores is more than through larger ones and the rate of diffusion ismore nearly proportional to the perimeter rather than its area. Thus under identicalconditions the rate of water vapour diffusion through the stomatal pores of Launaeawould be lower than that of Zilla owing to the large pore perimeter of the former inaddition to the low stomatal density in Launaea. Moreover, when a number of smallevaporating surfaces are close together, there is a mutual interference between themand the rate of evaporation from each area is reduced (Bange, 1953), which is alsoknown as the diffusion shell antagonism. On the abaxial surface, the stomatal spacingis 67·33 µm ± 5·5 (N = 18) in Zilla and 91·75 µm ± 10·1 (N = 20) in Launaea(unpublished data). Since the maximum diameter (width) of open stomata is onlyabout 6 µm and whereas the interference of diffusion shells is effective if the pores aresituated less than ten times away in relation to their diameter (Bange, 1953), it isevident that the spacing values on the abaxial leaf surface is 11·2 in Zilla and 15·3 inLaunaea. Accordingly, the mutual interference between the stomata is possible in Zillaand is most likely ineffective in Launaea.
Thickness of leaf or leaflet and their mesophyll properties
The similar arrangement of palisade cells on each side of the lamina is a commoncharacter found in plants growing in full sun, often high temperature habitats such asdeserts (Böcher, 1979; Nobel & Walker, 1985). Such leaves are known as unifacial oriso-bilateral. In the majority of the studied desert species, the palisade tissue is welldeveloped and formed at the expense of the spongy tissue which has compact cells. Ahigh ratio of palisade tissue thickness/spongy tissue thickness has been proposed as oneof the characters increasing the photosynthetic efficiency of leaves (Shields, 1951).
Compared to several literature sources, it is apparent that the range of leaf thicknessof the desert plants studied, although wide, is consistent with other species inhabitingsunny habitats, and all of them showed higher leaf thickness than shade plants. Leavesfrom perennial high altitude mountain plants, including tropics (Körner et al., 1989)and those from the alpine cushion plants from Canada (Hegazy, 1987) show widevariation from 141 to 696 µm in the former and from 125 to 682 µm in the latter.Although leaves from the chapparal sclerophyllous xerophytes from North America(Krause & Kummerow, 1977) show narrower ranges of leaf thickness (290–530 µm)than the above-mentioned groups, they are consistent with those from some perennialnon-succulent desert species (243–571 µm) from the eastern desert of Egypt whichhave a higher range of leaf thickness than the same species growing naturally undershaded habitats (291–416 µm) of desert wadi (Emad El-Deen, 1990). In crop plants,
G. M. FAHMY 518

such as sunflower, Dengler (1980) reported that the leaves which expand under lowlight intensities (25% daylight) have smaller leaf thickness (222·2 µm ± 8·5) thanleaves expanding under full sunlight (354·9 µm ± 9·1). Numerous investigators haveshown that ‘sun’ plants do produce thicker leaves than ‘shade’ plants (see review byLewis, 1972). This correlates with greater mesophyll thickness, mesophyll cells surfacearea, amphistomaty and stomatal density. Increases in maximum rates of photo-synthesis has been correlated with increased leaf thickness and increased area ofmesophyll cells (Nobel & Walker, 1985).
The fairly equal correlations between total Apal/A and thickness of lamina orbetween total Apal/A and total palisade thickness suggest the usefulness of measuringeither of these criteria as a convenient method of clarifying the photosyntheticefficiency of the desert species studied. Nobel et al. (1975) established a relationshipbetween the mesophyll area/leaf area (Ames/A) and leaf thickness. The palisadeparenchyma has a much higher amount of surface per unit volume than does thespongy mesophyll, from 1·6 to 3·5 times more (Turrell, 1936). Palisade cells areusually separated from one another by small but numerous air spaces. This gives a verylarge total cell surface/intercellular space and increase the internal gas and waterexchange surface of cells to intercellular space (Nobel & Walker, 1985). The highAmes/A is probably one of the reasons why, besides the increase in photosyntheticactivity, the rate of transpiration of xerophytes is high under conditions of favourablewater supply (Fahn, 1982). Previous studies (Turrell, 1936; Fahn, 1982) indicate thatAmes/A is small in shade leaves (6·8 to 9·9) and large in mesomorphic leaves (11·6 to19·2). Moreover, xerophytes tend to have a somewhat more highly developed palisaderegion than do mesophytes, which leads to values of 20 to 50 for Ames/A (Nobel,1991). The values of Apal/A for the desert species of the present investigation lie withinthe range of other xeromorphic ‘sun’ leaves previously reported.
In the cross-section of Salvia leaf, the tapering of mesophyll cells towards theepidermis results in a small contact area which, presumably, hampers the transport ofwater and other substances to the epidermal cells. A small contact area between thehypodermis and epidermis is considered a factor which reduces the loss of water to theepidermis from the internal cells of the assimilating stems of Anabasis articulata(Forssk.) Moq. and Calligonum crinitum Boiss. subsp. comosum (L’ Her.) Sosk.(Lyshede, 1977).
Predicted hydraulic conductance (Kh) and leaf specific conductivity (LSC)
A low Kh limits water transport to the transpiring parts (Woodhouse & Nobel, 1982)which may be advantageous for desert species. This pattern is consistent with 18species of this investigation. Since high Kh is an indication of less arid conditions, it issuggested that both Gomphocarpus and Chrozophora which show the highest predictedKh live under wetter conditions than the other 18 desert species. This conclusion isvalid in the case of Gomphocarpus which has restricted geographical and ecologicalrange in mountainous regions of southern Sinai where the environment is less arid(Zahran & Willis, 1992) than in the other desert areas from which the studied desertspecies were collected. However, such explanation is not valid for Chrozophora whichhas a wide geographical range in the deserts of Egypt. Owing to its predicted high Kh,it is assumed that Chrozophora has high transpiration rates, which may be severeenough to cause drying up of the leaves. This assumption agrees with Zahran & Willis(1992) who indicated that Chrozophora obliqua flowers in summer and undergoesdrying up of the branches at the end of the flowering phase in winter. In the petiolesof six species, the Kh values are lower than Gomphocarpus and Chrozophora and rangefrom 83·6 to 300·5 m4 MPa–1 s–1 3 10–13 for A. judaica, Launaea, Heliotropium, Zilla,Cleome and Salvia. The Kh values in the above-mentioned range are similar to those
LEAF ANATOMY AND ECOPHYSIOLOGY 519

reported by Gibson et al. (1984) for pinna and petiolule of the mesic fern Cyrtomiumfalcatum which ranged from 97 to 232 m4 MPa–1 s–1 3 10–13 in pinna and petiolule,respectively.
The remaining 12 species of the present study have the lowest values of predictedKh, ranging from 0·6 to 64·8 m4 MPa–1 s–1 3 10–13. The values for Acacia, Astragalus,Crotalaria, Teucrium and Fagonia are small (from 0·6 to 4·7 m4 MPa–1 s–1 3 10–13)and are consistent with the few vessels in the axes of their small laminae. The lowpredicted Kh values of the above-mentioned five species are much lower than the rangefound by Woodhouse & Nobel (1982) for the Kh of six fern species (13–59 m4 MPa–1
s–1 3 10–13) from southern California deserts. Moreover, the petioles of the remainingseven species (Achillea, Seriphidium herba-alba, Moltkiopsis, Farsetia, Ochradenus,Convolvulus and Lycium) have predicted Kh values similar to the stipes of the desertfern species reported previously. These Kh values are two to three orders of magnitudeless than for dicotyledons and six orders less than for conifers (Hellkvist et al.,1974).
The existence of a significant positive correlation between the number of vesselelements of the leaf or leaflet axis and the predicted Kh (r = 0·489, N = 19, p < 0·05;Table 6) is in agreement with similar measurements in the minor branches of somedicotyledons reported in previous studies (Tyree, 1989). The correlation of predictedKh with transpiring area (r = 0·685, N = 100, p < 0·0001; Fig. 4(a, b)) agrees withother studies on woody angiosperms (Waring et al., 1977) and fern species (Gibson etal., 1985; Veres, 1990).
The LSC is a usual measure of the hydraulic efficiency of the stem to supply waterto leaves distal to it (Tyree & Ewers, 1991). The stem with the larger LSC is able tosupply water more efficiently to leaves than a stem with smaller LSC (Zimmermann,1978). Although, the values of LSC for 16 species of the present investigation are forleaf or leaflet axes, they are low (0·16–3·66 m2 MPa–1 s–1 3 10–7) and lie within therange of measurements on the raches, stems and stipes supplying the pinnae of somemesic and greenhouse ferns, as well as the petioles and minor stem axes of some trees.The low LSC values (0·16 to 1·76 m2 MPa–1 s–1 3 10–7) which occur in nine speciesare similar to the rachis sections diverted to the pinnal area of the mesic fernPhanerophlebra falcata (L.f.). Copel., syn., Cyrtomium falcatum L.f. (0·2–0·3 m2 MPa–1
s–1 3 10–7; Gibson et al., 1984) and to the greenhouse Blechnum ferns (Veres, 1990)in which the LSC varied from 0·37 to 1·4 m2 MPa–1 s–1 3 10–7 for stems and elongateleaf stipes, respectively. The remaining seven species have LSC values similar to thepetioles of sugar maple trees (3·9 m2 MPa–1 s–1 3 10–7; Zimmermann, 1978) and theprimary stem segments of olive (2·9 m2 MPa–1 s–1 3 10–7; Thompson et al., 1983).
The highest LSC values (in m2 MPa–1 s–1 3 10–7) which occur in Zilla (14·24),Farsetia (6·68), Salvia (14·24) and Chrozophora (15·64) are similar to the LSC valuesin the stem segments of some shrubs (Ewers et al., 1991) as in Bauhinia aculeata(LSC = 6·2 m2 MPa–1 s–1 3 107) and to the stem segments of lianas (LSC = 11·0 m2
MPa–1 s–1 3 10–7 for Tylosema fassoglense (Schweinf.) Torre & Hillc. syn. Bauhiniafassoglensis Schweinf.). The similarities between the high LSC values in the leaf petiolesof the four desert species above-mentioned and those in the stem segments of lianas(which have wide vessels and high transpiration rates) is unusual because the petiolesof the desert species investigated are much smaller than the stem segments of lianas.This may be attributed to the occurrence of a high amount of xylem conduitssupplying the small leaves of the desert species, i.e. the lamina may be over-vascularised as in Salvia which has very small area (21·1 3 10–6 m2), relatively manyxylem conduits (167) in the petiole and a high LSC (14·24 m2 MPa–1 s–1 3 10–7).
In conclusion, it is apparent that the combination of water retaining features in theleaves of the majority of the desert species investigated, together with the relatively lowKh and LSC of their petioles or petiolules (low Kh and LSC occur in 16 and 18 species,respectively, out of the 20 species studied), may be some of the adaptive features of the
G. M. FAHMY 520

species to living under desert conditions where climatic and edaphic aridity areprevailing. Gartner et al. (1990) found that the vines and trees of the dry sites havelower conductivities than plants of the same growth form living in wetter seasons orwetter sites.
A mean value of E max of 1 3 10–7 m s–1 is based on the maximum rates of desertplants measured by Evenari & Richter (1938). The values of predicted dp/dx are shownin Table 5. They range from 0·06 MPa m–1 in Chrozophora to 6·25 MPa m–1 inTeucrium. Zimmermann (1978) reported a theoretical dp/dx of about 0·02 MPa m–1 inthe trunks of many dicotyledonous trees, but much greater values occur in lateral andminor branches. Values less than 1 MPa m–1 (calculated in 13 species of the presentinvestigation) are of a magnitude reported to be significant in inducing stomatalclosure in dicot trees and lianas (Ewers et al., 1989, 1991). Stomatal closure mayprotect the shoots from excessive embolism which results in the dysfunction of xylemconduits (Sperry & Tyree, 1988). In some crop plants, the lowest embolism-inducingpressures were c. 0·2 MPa for tomato, 0·5 MPa for castor bean (cf. Jones, 1992, p. 94)and below c. 1·0 MPa for irrigated corn plants (Tyree et al., 1986). In trees, theembolism-inducing pressure is higher ( = becomes more negative) than in crop plants.It reached 2·9 MPa for Rhapis palm (Sperry, 1985) and 3·00 MPa for both Fraxinus(cf. Zimmermann & Milburn, 1982) and Acer saccharum K. Koch trees (Tyree &Dixon, 1986).
The value of calculated dp/dx of Teucrium (6·25 MPa m–1) is much higher than thosecalculated for other desert species of this study which have nearly similar shoot size toTeucrium. Therefore, such a value is not accepted to be applied to Teucrium but couldbe only valid for plants having extensive shoot system. Salleo et al. (1982) measured Emax indoors with a potometer and used the above-mentioned equation to predict dp/dxin Vitis vinifera L: from 0·01 MPa m–1 near the base of 1-year-old stems to 6·5 MPam–1 near the apical nodes.
Conclusions
Desert perennials have developed a great variety of morphological and anatomicaladaptive characteristics. Structural adaptations to xeric environments, the xer-omorphic characteristics, may occur in leaves, stems and roots (Fahn & Cutler, 1992).In the present investigation, two major characteristics of xeromorphic leaf adaptationscan be distinguished; one is present in the lamina and possibly leads to the reductionin rates of transpiration and photosynthesis, and the other mainly depends ontracheary elements and affects their hydraulic conductance. The characteristicconcerned with the lamina includes a group of ‘water retaining features’ which can besummerised as follows: (1) Morphological features: these include small leaf or leafletsize and dissected or lobed narrow lamina which lower the boundary layerconductances and which are most likely to be advantageous to the plant underconditions of high temperatures and large humidity deficits; (2) Anatomical features:these include either non-stomatal features (pubescent surfaces, smooth or slightlyrough outer cuticular surface) or stomatal-related characteristics. The formers affectthe leaf temperature while the laters respond to metabolic and environmental factorsand affect the water retaining features by regulating water loss.
The efficiency of the conducting tissue was demonstrated from the calculation ofhydraulic conductance (Kh) of the xylem of the petiole, petiolule or the basal part ofthe sessile leaf or leaflet using the Hagen-Poiseuille relation (Gibson et al., 1984). Forthe majority of species (18 out of 20) the Kh was low, particularly in the species whichhave small areas of lamina supplied by a small number of vessel elements. Predicted Khvalues of the majority of the desert species studied lie within the range of the stipes ofsome ferns (Woodhouse & Nobel, 1982) and are two to three orders of magnitude less
LEAF ANATOMY AND ECOPHYSIOLOGY 521

than for dicotyledons (Hellkvist et al., 1974). A low Kh limits water transport to thetranspiring parts (Woodhouse & Nobel, 1982) and this led us to conclude that it maybe advantageous for desert plants living under conditions of limited water supply. Inorder to adjust the Kh values according to the leaf or leaflet area supplied, the leafspecific conductivity (LSC) was determined (LSC = Kh/distal leaf or leaflet area)according to Tyree et al. (1983). The majority of the desert species investigated havelow LSC which indicates that they have a low hydraulic efficiency. Zimmermann(1978) pointed out that the stem with a larger LSC is better able to supply water toleaves than a stem with a smaller LSC.
It is apparent that the combination of water retaining features in the leaves of themajority of the desert species investigated together with the relatively low Kh and LSCvalues may be some of the adaptive features of the species to live under desertconditions where climatic and edaphic aridity are prevailing.
Compared to several literature sources it is apparent that the ranges of laminathickness and the ratio of the palisade cells surface area/the leaf or leaflet area (Apal/A),although wide, are consistent with other species inhabiting sunny habitats (Lewis,1972; Nobel, 1991). Fahn (1982) pointed out that a high ratio of mesophyll area/leafarea is probably one of the reasons why, besides the increase in photosynthetic activity,the rates of transpiration of xerophytes is high under conditions of favourable watersupply.
I thank my colleagues Dr A.K. Hegazy and Dr H. El-Tantawy who accompanied me on somefield trips to the desert. Thanks are also extended to the colleagues of the Cairo UniversityHerbarium (CAI); in particular to Dr M.M. Abd El-Ghani, who confirmed the identificationand geographical distribution of the studied species. I am thankful to Prof. Dr A.C. Gibson,University of California, Los Angeles, for help in SEM and for suggestions. Words cannotadequately describe the humanistic support, encouragement and advice of Prof. Dr A.A. AbdEl-Rahman, Department of Botany, Faculty of Science, Cairo University and Prof. Dr H.Ziegler, The Technical University of Munich, Germany. The comments of two anonymousreviewers have cleared many ideas. Thanks also to Dr G.E. Wickens, formerly of the RoyalBotanic Gardens, Kew, England for advice and suggestions. I am also thankful to Dr DarwishS. Darwish, Faculty of Agriculture, University of Cairo and to Prof. Dr B. Lewis, Departmentof Statistical Analysis, Agricultural Research Centre, Giza for comments and help.
References
Abd El Rahman, A.A. & Batanouny, K.H. (1965). The water output of desert vegetation in thedifferent microhabitats of Wadi Hoff. Journal of Ecology, 53: 139–145.
Abd El Rahman, A.A., Ayyd, M.A. & El-Monayeri, M.M. (1966). Hydroecology of the sanddunes habitat at Burg El Arab. Bulletin of the Faculty of Science, Cairo University, 40: 29–54.
Abd El Rahman, A.A., Ezzat, N.H. & Hassan, A.H. (1976). Comparative hydroecologicalstudies on some hydrophytes, wet and dry halophytes and xerophytes. Flora, 165: 1–16.
Abd El Rahman, A.A., Ibrahim, A. Amal & Hassan, H.T. (1980). Contributions to theanatomical characters of some xerophytes. Bulletin of the Faculty of Science, Cairo University,49: 139–162.
Anon. (1979). Climatological Normals for the Arab Republic of Egypt, pp. 188, 210, 242. Cairo,Egypt: Ministry of Civil Aviation-Meterological Authority. 433 pp + map.
Ayyad, M.A. & Ghabbour, S. (1986). Hot deserts of Egypt and the Sudan. In: Evenari, M.,Noy-Meir, I. & Goodall, D. (Eds), Ecosystems of the World 12B: hot deserts and arid shrublands,pp. 149–202. Amsterdam: Elsevier Science Publishers B.V. 451 pp.
Bange, G.G.J. (1953). On the quantitative explanation of stomatal transpiration. Acta BotanicaNeerlandica, 2: 255–296.
Batanouny, K.H. (1974). Eco-physiological studies on desert plants. IX: Types of transpirationcurves of Zilla spinosa Prantl under natural conditions. Flora, 163: 1–6.
Batanouny, K.H., Stichler, W. & Ziegler, H. (1988). Photosynthetic pathways, distribution andecological characteristics of grass species in Egypt. Oecologica (Berlin), 75: 539–548.
G. M. FAHMY 522

Böcher, T.W. (1979). Xeromorphic leaf types: Evolutionary strategies and tentative semi-phyletic sequences. Biologiske Skrifter Danske Videnskabernes Selskab, 22: 1–71.
Boulos, L. (1995). Flora of Egypt Checklist. Cairo: Al Hadra Publishing. 283 pp.Brown, W.V. & Escombe, F. (1900). Static diffusion of gases and liquids in relation to the
assimilation of carbon and translocation in plants. Philosophical Transactions of the RoyalSociety (London), Botany, 193: 223–291.
Carter, G.A. (1991). Primary and secondary effects of water content on the spectral reflectanceof leaves. American Journal of Botany, 78: 916–924.
Cloudsley-Thompson, J.L. (1984). Introduction. In: Cloudsley-Thompson, J.L. (Ed.), SaharaDesert (Key Environments), pp. 1–15. Oxford: Pergamon Press (published in collaborationwith the IUCN). 348 pp.
Dengler, N.G. (1980). Comparative histological basis of sun and shade leaf dimorphism inHelianthus annuus. Canadian Journal of Botany, 58: 717–730.
Ehleringer, J.R. (1980). Leaf morphology and reflectance in relation to water and temperaturestress. In: Turner, N.C. & Kramer, P.J. (Eds), Adaptation of Plants to Water and HighTemperature Stress, pp. 295–308 New York: John Wiley & Sons. 482 pp.
Ehleringer, J.R. (1981). Leaf absorptances of Mohave and Sonoran Desert plants. Oecologia(Berlin), 49: 366–370.
Ehleringer, J.R. (1984). Ecology and ecophysiology of leaf pubescence in North Americandesert plants. In: Rodriguez, E., Healey, P.L. & Mehta, I. (Eds), Biology and Chemistry of PlantTrichomes, pp. 113–132. New York: Plenum Press. 255 pp.
Ehleringer, J.R. & Björkman, O. (1978). A comparison of photosynthetic characteristics ofEncelia species possessing glabrous and pubescent leaves. Plant Physiology, 62: 185–190.
Emad El-Deen, H.M. (1990). Some Aspects of Drought Resistance Mechanisms of DesertPlants. M.Sc. thesis, Faculty of Science, Cairo University, Egypt. 177 pp + Appendix.
Evenari, M. (1985). Adaptations of plants and animals to the desert environment. In: Evenari,M., Noy-Meir, I. & Goodal, D.W. (Eds), Ecosystems of the World 12A: hot deserts and aridshrublands, pp. 79–92. Amsterdam: Elsevier Science Publishers B.V. 365 pp.
Evenari, M. & Richter, R. (1938): Physiological – ecological investigations in the wilderness ofJudaea. Journal of Linnean Society (Botany), 51: 333–381.
Ewers, F.W., Fisher, J.B. & Chiu, S.-T. (1989). Water transport in the liana Bauhinia fassoglensis(Fabaceae). Plant Physiology, 91: 1625–1631.
Ewers, F.W., Fisher, J.B. & Fichtner, K. (1991). Water flux and xylem structure in vines. In:Putz, F.E. & Mooney, H.A. (Eds), The Biology of Vines, pp. 127–160. Cambridge: CambridgeUniversity Press. 526 pp.
Fahn, A. (1964). Some anatomical adaptations of desert plants. Phytomorphology, 14: 93–102.Fahn, A. (1982). Plant Anatomy. Oxford: Pergamon Press. 544 pp.Fahn, A. (1986). Structural and functional properties of trichomes of xeromorphic leaves.
Annals of Botany, 57: 631–637.Fahn, A. & Cutler, D.F. (1992). Xerophytes. Encyclopedia of plant anatomy, Band III, Teil 3.
Berlin: Gebruder Borntraeger. 176 pp.Gartner, B.L., Bullock, S.H., Mooney, H.A., By Brown, V. & Whitbeck, J.L. (1990). Water
transport properties of vine and tree stems in a tropical deciduous forest. American Journal ofBotany, 77: 742–749.
Gibson, A.C. (1983). Anatomy of photosynthetic old stems of nonsucculent dicotyledons fromNorth American deserts. Botanical Gazette, 144: 347–362.
Gibson, A.C., Calkin, H.W. & Nobel, P.S. (1984). Xylem anatomy, water flow, and hydraulicconductance in the fern Cyrtomium falcatum. American Journal of Botany, 71: 564–574.
Gibson, A.C., Calkin, H.W. & Nobel, P.S. (1985). Hydraulic conductance and xylem structurein tracheid-bearing plants. International Association of Wood Anatomists Bulletin (New Series),6: 293–302.
Givnish, T.J. (1988). Adaptation to sun and shade: a whole plant perspective. Australian Journalof Plant Physiology, 15: 63–92.
Good, R. (1953). The Geography of the Flowering Plants (2nd Edn). London: Longman, Greenand Co. 452 pp.
Hegazy, A.K. (1987). Perspectives on Autecology and Population Biology of Alpine CushionPlants. Ph.D. thesis, The University of Alberta, Canada. 285 pp. + Appendix.
Hellkvist, J., Richards, G.P. & Jarvis, P.G. (1974). Vertical gradients of water potential andtissue water relations in Stika spruce trees measured by the pressure character. Journal ofApplied Ecology, 11: 637–667.
LEAF ANATOMY AND ECOPHYSIOLOGY 523

Ibrahim, A.A. & Fahmy, G.M. (1985). The significance of anatomical characters in the watereconomy of some desert plants. Annals of Agricultural Science, Moshtohor (Egypt), 23:145–162.
Jones, H.G. (1992). Plants and Microclimate: a quantitative approach to environmental plantphysiology (2nd Edn). Cambridge: Cambridge University Press. 428 pp.
Juniper, B.E. & Jeffree, C.E. (1983). Plant Surfaces. London: Edward Arnold. 93 pp.Kassas, M. & Batanouny, K.H. (1984). Plant ecology. In: Cloudsley-Thompson, J.L. (Ed.),
Sahara Desert (Key Environments), pp. 77–90. Oxford: Pergamon Press (Published incollaboration with the IUCN). 348 pp.
Kassas, M. & Imam, M. (1954). Habitat and plant communities in the Egyptian desert. III. Thewadi bed ecosystem. Journal of Ecology, 42: 425–441.
Körner, C., Neumayer, M., Menedez-Riedel, S.P. & Smeets-Scheel, A. (1989). Functionalmorphology of mountain plants. Flora, 182: 353–383.
Krause, D. & Kummerow, J. (1977). Xeromorphic structure and soil moisture in the chaparral.Oecologia Plantarum, 12: 133–148.
Laetsch, W.M. (1974). The C4 syndrome: a structural analysis. Annual Review of PlantPhysiology, 25: 27–52.
Larcher, W. (1983). Physiological Plant Ecology (2nd Edn). Berlin, Heidelberg, New York:Springer-Verlag. 303 pp.
Larmuth, J. (1984). Microclimates. In: Cloudsley-Thompson, J.L. (Ed.), Sahara Desert (KeyEnvironments), pp. 57–66. Oxford: Pergamon Press (Published in collaboration with theIUCN). 348 pp.
Lewis, M.C. (1972). The physiological significance of variation in leaf structure. Science Progress(Oxford), 60: 25–51.
Lewis, D.A. & Nobel, P.S. (1977). Thermal energy exchange model and water loss of a barrelcactus, Ferocactus acanthodes. Plant Physiology, 60: 609–616.
Lyshede, O.B. (1977). Anatomical features of some stem assimilating desert plants of Israel.Botanisk Tidsskrift, 71: 255–230.
McClendon, J.H. (1984). The micro-optics of leaves. I. Patterns of reflection from theepidermis. American Journal of Botany, 71: 1391–1397.
Migahid, A.M. (1962). The drought resistance of Egyptian desert plants. Proceedings of AridZone Symposium on Plant Water Relations in Arid and Semi-Arid Conditions, pp. 213–233. Paris:UNESCO.
Mott, K.A., Gibson, A.C. & O’Leary, J.W. (1982). The adaptive significance of amphistomaticleaves. Plant Cell and Environment, 5: 455–460.
Nobel, P.S. (1975). Effective thickness and resistance of the air boundary layer adjacent tospherical plant parts. Journal of Experimental Botany, 26: 120–130.
Nobel, P. (1991). Physicochemical and Environmental Plant Physiology. San Diego: AcademicPress. 635 pp.
Nobel, P.S. & Walker, D.B. (1985). Structure of leaf photosynthetic tissue. In: Barber, J. &Baker, N.R. (Eds), Photosynthetic Mechanisms and Environment, pp. 501–536. Amsterdam:Elsevier Science Publishers (Biomedical Division). 565 pp.
Nobel, P.S., Zaragoza, L.J. & Smith, W.K. (1975). Relation between mesophyll surface area,photosynthetic rate, and illumination level during development for leaves of Plectranthusparviflorus Henckel. Plant Physiology, 55: 1067–1070.
O’Brien, T.P. & McCully, M.E. (1981). The Study of Plant Structure: principles and selectedmethods. Melbourne: Termarcarphi. 328 pp + Appendix xxvi pp.
Oppenheimer, H.R. (1960). Adaptation to drought: Xerophytism. Arid Zone Research, 15:105–138. Paris: UNESCO.
Pyykko, M. (1966). Leaf anatomy of East Patagonican xeromorphic plants. Annales BotaniciFennici, 3: 453–622.
Sakai, W. (1973). Simple method for differential staining of paraffin embedded plant materialusing toluidine blue O. Stain Technology, 48: 247–249.
Salleo, S., Rosso, R. & Lo Gullo, M.A. (1982). Hydraulic architecture of Vitis vinifera L. andPopulus deltoides Bartr. 1-year-old twigs: I. Hydraulic conductivity (LSC) and water potentialgradients. Giornale Botanico Italiano, 116: 15–27.
Sayer, J.D. (1926). Physiology of stomata of Rumex patienta. Ohio Journal of Science, 26:233–266.
Shields, L.M. (1951). Leaf xeromorphy in dicotyledon species from a gypsum sand deposit.American Journal of Botany, 38: 175–190.
G. M. FAHMY 524

Small, E. (1973). Xeromorphy in plants as a possible basis for migration between arid andnutritionally–deficient environments. Botaniska Notiser, 126: 534–539.
Snedecor, G.W. & Cochran, W.G. (1962). Statistical Methods (5th Edn). Iowa: Iowa StateCollege Press. 535 pp.
Sperry, J.S. (1985). Xylem embolism in the palm Rhapis excelsa. International Association of WoodAnatomists Bulletin (New Series), 6: 283–292.
Sperry, J.S. & Tyree, M.T. (1988). Mechanism of water stress-induced xylem embolism. PlantPhysiology, 88: 581–587.
Stocker, O. (1976). Water–photosynthesis syndrome and geographical plant distribution in theSahara. In: Lange, O.L., Kappen, L. & Schulze, E.D. (Eds), Water and Plant Life, pp.506–522. Berlin: Springer-Verlag. 536 pp.
Sundberg, M.D. (1986). A comparison of stomatal distribution and length in succulent andnon-succulent desert plants. Phytomorphology, 36: 53–66.
Tackholm, V. (1974). Students’ Flora of Egypt (2nd Edn). Cairo: Cairo University Press. 888pp.
Thompson, R.G., Tyree, M.T., Lo Gullo, M.A. & Salleo, S. (1983). The water relations ofyoung olive trees in a Mediterranean winter: Measurements of evaporation from leaves andwater conduction in wood. Annals of Botany, 52: 399–406.
Turrell, F.M. (1936). The area of internal exposed surface of dicotyledon leaves. AmericanJournal of Botany, 23: 255–264.
Tyree, M.T. (1989). Cavitation in trees and the hydraulic sufficiency of woody stems. Annalesdes Sciences Forestieres, 46s: 330s–337s.
Tyree, M.T. & Dixon, M.A. (1986). Water stress induced cavitation and embolism in somewoody plants. Physiologia Plantarum, 66: 397–405.
Tyree, M.T. & Ewers, F. (1991). The hydraulic architecture of trees and other woody plants.(Tansley Review No. 34). New Phytologist, 119: 345–360.
Tyree, M.T., Graham, M.E.D., Cooper, K.E. & Bazos, L.J. (1983). The hydraulic architectureof Thuja occidentalis. Canadian Journal of Botany, 61: 2105–2111.
Tyree, M.T., Fiscus, E.L., Wullschleger, S.D. & Dixon, M.A. (1986). Detection of xylemcavitation in corn under field conditions (A communication). Plant Physiology, 82:597–599.
UNESCO (1979). Map of the World Distribution of Arid Regions. Explanatory Note. MABTechnical Notes 7. Paris: UNESCO. 54 pp. + map.
Uphof, J.C. Th. (1962). Plant Hairs. Encyclopedia of plant anatomy, Band IV, Teil 5. Berlin:Gebruder Borntraeger. 292 pp.
Veres, J.S. (1990). Xylem anatomy and hydraulic conductance of Costa Rican Blechnum ferns.American Journal of Botany, 77: 1610–1625.
Volkens, G. (1887). Die Flora der Aegyptisch-arabischen Wüste auf Grundlage anatomisch-physiologischer Forschungen dargestellt. Berlin: Gebruder Borntraeger. 156 pp + xviii
Walter, H. (1979). Vegetation of the Earth and Ecological Systems of the Geo-biosphere (2nd Edn)(translated from the 3rd revised German Edn). Berlin, New York, Heidelberg: Springer-Verlag. 274 pp.
Waring, R.H., Gholz, H.L., Grier, C.C. & Plummer, M.L. (1977). Evaluating stem conductingtissue as an estimator of leaf area in four woody angiosperms. Canadian Journal of Botany, 55:1474–1477.
Willmer, C.M. (1983). Stomata. London: Longman. 166 pp.Woodhouse, R.M. & Nobel, P.S. (1982). Stipe anatomy, water potentials, and xylem
conductances in seven species of ferns (Filicopsida). American Journal of Botany, 69:135–140.
Zahran, M.A. & Willis, A.J. (1992). The Vegetation of Egypt. London: Chapman & Hall. 424pp.
Ziegler, H., Batanouny, K.H., Sankhla, N., Vyas, O.P. & Stichler, W. (1981). Thephotosynthetic pathway types of some desert plants from India, Saudia Arabia and Iraq.Oecologia (Berlin), 48: 93–99.
Zimmermann, M.H. (1978). Hydraulic architecture of some diffuse porous trees. CanadianJournal of Botany, 56: 2286–2295.
Zimmermann, M.H. & Milburn, J.A. (1982). Transport and storage of water. In: Lange, O.L.,Nobel, P.S., Osmond, C.B. & Ziegler, H. (Eds), Encyclopedia of Plant Physiology, New Series,Vol. 12B, pp. 135–151. New York: Springer-Verlag. 747 pp.
LEAF ANATOMY AND ECOPHYSIOLOGY 525