late eocene to oligocene events: molino de cobo, betic...
TRANSCRIPT

REVISTA EspAÑOLA DE MICROPALEONTOLOOIA
Vol. XX, mlm. 3, 1988, pp. -491.51-4
EUSTaQUIO MaLINA
Departamento de PaleontologíaUniversidad de Zaragoza50009 Zaragoza, España
GERTA KELLER
Departmellt 01 Geological and Geophysical SciencesPrinceton University
Princeton, NJ 08544, U.S.A.
MARINA MADILE
Dipartimento Sciem.:e del/a TenaUniversitá di Firenze50121 Firenze, Italia
LATE EOCENE TO OLIGOCENE EVENTS:MOLINO DE COBO, BETIC CORDILLERA, SPAIN
Quantitative analysis of upper Eocene to Oligocene planktonic foraminifers andcalcareous nannofossils in the Molino de Cobo section, Betic Cordillera, Spain indicate three major extinction evenLS: 1) in the upper Eocene at the extinction ofGlobigerapsis index, 2) at the Eocene-Oligocene boundary and 3) al tbe lower/upperOligocene boundary. The Globigerapsis index extinction event in Ibis area coincideswith the dramatic abundance decline of the discshaped discoasters (D. saipanensis,D. barbadiensis). This faunal and floral assemblage change coincides with a carbonate dissolution interval.
The Eocene-Oligocene boundary extinction event involves five planktonic foraminiferal species, which contrary to common belief did not go extinct simulta~
neously, but stretched out over a 3m intervalo This extinction event is probably related to the isotopic enrichment that signals the development of the psychrosphere,or two layer ocean with cold bottom and warm surface water. The lower/upperO1igocene faunal tumover event involves the extinction of surviving Eocene speciesand the evolulion of late Oligocene to Miocene species. A short hiatos may be present at this intervalo This faunal turnover is most likely related to global coolingand a major sea level drop.
El análisis cuantitativo de los foraminíferos planctónicos y nannofosiles calcáreos del Eoceno superior y Oligoceno del corte. de Molino de Cobo (Cordillera Bética,España). pone de manifiesto tres importantes eventos de extinción; en el Eocenosuperior, en el límite Eocene/Oligoceno y en el límite Oligoceno inferior/superior,respectivamente. El evento del Eoce.no superior implica la súbita desaparición delgenero Globigerapsis. el cual constituia el 20 por 100 del tolal de la población. Losdiscoaster en roseta (D. saipanensis y D. barbadiensis) también declinan dramáticamente en este momento. Estos cambios en las asociaciones de foraminíferos y nannofosiles, coinciden estratigráficamente con un intervalo de disolución.
El evento de. extinción del límite Eoceno/Oligoceno involucra a cinco especies deforaminíferos planctónicos, las cuales contrariamente a la creencia más generalizadano se extinguen simultáneamente, sino que se producen en un intervalo de 3 metros.Este evento de extinción está probablemente ligado al desarrollo definitivo de lapsicrosfera.
El último evento coincide con el límile Oligoceno inferior/superior y supone unimportante relevo; la extinción de especies supervivientes del Eoceno y la evoluciónde especies del Oligoceno superior y Mioceno. Este relevo está seguramente ligadoa un enfriamiento global y a una importante caída del nivel del mar, habiéndosedetectado además, un breve hiato dentro de este intervalo.
491

MOLINA, KELLER, MADILE
INTRODUCTlON
The late middlc Eocene 10 Oligocene represcnts a time of majar paleoenvironmental andpaleoclimatic changes as observed by a seriesof stepwise extinctions (Keller. 1983. 1986)accompanied by a longlerm c1imatic coolingtrend and a permanent dTay in bollom watertemperatures ncal' the Eocene·Oligocene houndary (Shackleton and Kennclt, 1976; Keigwin,1980; Keigwin and Keller. 1984; Millcr andThomas. 1985; Kcigwin and Corliss, 1986;Oberhansli and Toumark.ine, 1985; Williamsel aL, 1985). 'Lowering of sea level and widespread hiatuses are frenquently associated withthese faunal and climalic cvenls (MilJer el aL,I98S; Keller el aL, 1986, 1987; Haq el al., 1987).
Faunal changes gene rally parallcl climalictrends. Late middle Eoeenc tO Oligoeene wanowater asscmblages are sueeessively replaeedby planktonic foraminifera (Ke!lcr, 1983, 1985,1986), ealcareous nannoplanklon (Haq and Lohman, 1976; Haq el al., 1977; Pereh-Nielsen etal., 1986), ostraeod faunas (Steincck et al., 1984)and mollusks (Hut el al., 1987). The climatieeooling trend is gene rally eonsidered lO be related lo Ihe developmenl of the drcum-Anlarelie eurrenl and subsequent eooling of Antareliea initialed by lhe northward movement ofTasmania and Auslralia by mi~dle Eoeene time (Wcissel and Hayes, 1972; MeGowran, 1973;Kennctt, 1977).
The recent diseovcry of t.hree micrOlektiteand relatcd mierospherule layers in low latilude upper Eocene marine sediments (Kelleret al., 1983, 1987; Glass et al., 1985), however,has suggested that some faunal turnoversand climalic coolings may have becn triggcred01- acc!cl-alcd by cxtraterrest,·ial impacl events.Invcsligation of microplanklon in marine sectians containing one 01' more of these microtcklite or micl"Ospherule layers re"caled lhalno planktonie foraminiferal specics extinetionscoincide prccisely wilh these cvents, but fiveradiolarian spccies cxlinction al·e associalcdwith one lnycr (Maurasse and Glass, 1976; San-
492
filippo et al., 1985). One olher microspherulelayer is associated with a catastrophic declinein the planklOnic foraminiFer Cenus Glabigerapsis fe. semiínvolutus, e. luterbacheri, e. howei) and t.he extinction of lhis group occursshortly aboye lhis layer (Keller, 1986; Kelleret al., 1987).
These investigations indieate that species exlinetions aione may be a pOOl' measure of theeffect on a population after a sudden environmental jolt such as ao impacI by an eXlraterres·lria! body. Species popuJatioos are geoeraUyrare at time oC evolution and extinetion andreaeh Iheir apex sometime in between. ·Sudden adverse environmental eonditions may decimate a species population lO the point where only Cew individuals survive, bUI do not reeover as a stable population, and eventuallybecome extinel. Quantitative population sludiesbased on relative abundances of individuals ina species may thcrefore provide a more accurate measure oC the covironmental effects(Keller, 1986). It is therefore important tosearch for good physical (microtektites) andbiological (fossil) evidenee in late Eocene seclions, to document whether species eXlinetionsas well as major fauna! tumovers and climaticchanges are triggered by largc body Eanh impacts.
The upper Eocene sedirncnts of the Molinode Coba scction of the Betic CordilJera of Spain(Figure 1) offer a unique opportunity to iovestigale biotic cvents during late Eocene to earlyOügoeene time. We have studicd the planktonie foraminifers and ealcareous nannofossilsof this scction quamitatively lO determine theslratigraphy and population changcs in lermsoC relative abundanees of dominant species. Wehave been able lO document three majar faunalevcnts which apparcnlly correlate with a lateEocene evcnt, Ihe Eocene-Oligocene boundaryevenl and the carly/late Oligocenc sea-Ieveldrop. Faunal evenls al thcse lime have alsobecn obscrved in lo\V lalilude seetions (Kcller,1983, 1985, 1986) and are lhus believcd to represenl global events.

LOCATION AND LITHOLOGY
GeographicaJly, thc Molino de Cobo sectionis located in (he Cañada de Jaen Ravine, inthe lownship cf El Gobernador (Granada Province), 2 km northeast from the village and200 ro west froro the Molino de Cobo farmhouse. The section is exposed a]ong the Granada-Madrid railway tracks accessible by apath fTom the Gobernador viUage lo the PedroMartinez train stalion.
Geologically, the Molino de Cobo section islocated in the Subetic Zone of the Betic Cordillera. The section is part of lhe Cañada FormalioÍl {Eocene-Aquitanian} of the CardelaGraup. Sediments of the Cañada Formation
out<. ,...
Figure I
Location map oC the Molino de Cobo sectioni!'l Andalucia. southcrn Spain.
MOUNA, KELLER, MADILE
are over SOO ro thick and consist of detriticIimestone of turbidite origin, interbedded withtbid hemipelagic mar!s.
Upper Eocene to Oligoeene sedimeots of theMolino de Cobo section eonsist of 194 m of interbedded bioclaslic calcarenites and macisthat dip 30 degrees north forming the southernf1ank of a wide syncline. The eontinuation lowards the oucleus was named Cañada de Jaen(El Gobernador) seetion and biostratigraphically studied by meaos ef planktonic forami·nifers (Malina, 1979). The sediments are lithologically similar lo tbe nearby coeval FuenteCaldera section (Comas et al., 1984-85; Molina,1986), excepl lhat tbe turbidite facies are.lessextensive and 00 olithostromes are present inthe Molino de Cobo section. The bioclastiecalcareoites are 30-90 cm lhick and range frompackestones to pseudograinslones with frequent marly inclusions. The bioclasts consistmainly of smaller and larger bentbonic foraminifera and calcareous algae primari1y of pe·necontemporaoeous platform origin, althoughsorne middle Eocene species have been found.The autochthonous hemipelagic Olarly interbeds mnge between 1-5 ro thiek and do noteontain displaced larger benthonic foraroini·fers. Reworked rniddle Eocene planktonic foraminifers and calcareaus nannafossils are fewto cornman in sorne mar! beds.
METHOD5
Sediment samples were disaggregated by soaking in water ayer night with the addition afa small amOUnl or 10 OJo hydrogen peroxide. Iri·durated samples wece also heated to aid thedjsaggregalion process. Samples were thenwashed over 63 and 150 rnieron screens withtap water. Quantitative eounls for faunal anaIysis wece obtained from aliquots (using a Olodified Otto microsplitter) of approximately300-500 specimens of the size fraetion greater(han 150 microns. AH spccimens were thenpicked from the aliquot and mountcd on a microslide for identification and 'pennanent re-
493

MOLINA, KELLER, MADlLE
cardo The sample was a150 scanned for rarespecies and the smaU size fraclion was exa.mined far smaller species.
Thin sections were marle cf the calcarenitielimes tone beds lo study the larger foraminiferaand examine the lithology. Calcareous Dannofossils were studied from the marly beds. Asmall quantity af a c1ean rack chip was crus·herl with a pestle in a mortar contain.ing disoti1led water (ph 6/7). A drop cf the suspensionwas then smeared on a cover slide, dried andemhedded on a slide with Pyccolite. A semi·quantitative analyses cf the abunrlance cf dominante species was estimated using the Back.mano and Shackleton (1983) method. Abun.dance is expressed in number of specimensper mmJ al a magnification cf x 1250.
BIOSTRATlGRAPHY
PLANKTONIC FORAMINIFERA
A planktonic foraminiferal biostratigraphicanalysis of the Molino de Cobo section wasearlier published by Martinez-Gallego (1974).We have resampled the section at closer intervals and studied tbe planktonie foraminiferaand caleareous nannofossils based on bothstandard biostratigraphic techniques (first andlast appearances of index species) and quantitative faunal analysis. These combined methodspennit a higher resolution time control andin addition provide paleoecologic and paleoclimatic 'information. Quantitative faunal ana·Iysis can also minimize problems of reworkedolder faunas and contamination by isolatingthese species. II also permits a more accurateassessment of the magnitude of an cxtinctionevent by providing information 00 the relativeportion of individuals of the populationaffected.
Our faunal counts are shown in Table 1.Figure 2 shows the stratigraphie ranges ofspecies indieating where they are abundant,common or few. Figure 3 i1ustrates relative pereeot abuodances of dominant species. We haveuscd the standard low Iatitudc zonations of
494
BoIli (1966) and Blow (1979) as well as a revised middle latitude zonatioo for this regionby Molina (1979, 1986). BolJi's 10w latit~dezonation is applicable, except tbat the boundary between Cassigerinella cllipolensis-Pseudo-hastigerilla micra and Globigerina ampliapertura Zones could not be recognized dueto absence or rarity of P. micra. Blow's zonation was a1so sometimes diffieuh to apply because of absenee or rarity of species and difficulty in identification of sorne index species.In sorne cases (Zones PIS-PI6) the species ranges also appear diaehronous. We therefore prefer Bolli's (1966) zonation to correlate to lowlalilude sections and Molina's (1979, 1986) ronatioo for middle latitudes.
Although the aim of this paper is nol lodiscuss the taxonomic problems of tbe planktonie foraminifers, sorne enteria of identificalion should be clarified. The species eonceptthat has been followed in this work is lhesarne as proposed by the European WorkingGroup on Planktonie Forarnioifera (Robasynski et al., 1984). Thus, no subspecies have beencoosidered. The species are general1y definedby typological eriteria. and are often morpbotypes with rather arbitrary boundaries due lotheir interspecific variations. For stratigrapbicpurposes morphotypes with bioslratigraphicalsignifieance are choscn. Tbus, sorne of thescspecies may not be tme biological species.Other speeies rnay inelude a wide variety ofmorphotypes of uncertain affinities such asCatapsydrax unicavus 5_ 1. which ¡neludesC. pera and eertain .Clobigerinasa with smallabortive final chambers.
Taxonomic problems in this paper a1so eoneero the genus Pseudohastigeril1a.. Pseudohastigerina. micra is a laterally eompresscd forrowhich spans the Eocene/Oligocene boundaryand is an important stratigraphic marker. In1979 Blow proposed the name P. danvillensisfor this form which we eonsider a junior synonym. Two similar but much smaller morphotypes of the species P. naguewichiensis andP. barbadoensis are also present, but can bedistinguished only with the scanning electron

microscope: We have IUlhped these two smaJlspecies as P. naguewichiensis. Also because oftheir small size and difficulties in identification with the stereomicroscope, biserial Chiloguembelinas have been lumped as Ch. cubensis s. 1.
Upper Eocene
The Jowennost 20 m (up to sample 2'4, Figure 2) oF the Molino de Cobo section correspondto the Gfobigerapsis semiinvolurus Zone. Thetop of this zone is defined by the extinctionof G. semiinvolwus. Blow's (1979) PIs/PI6Zone boundary based in the first appearanceof Cribrohantkemna inlfara occurs at 13mfrom the base of the section where also Globigerina gartanii which defines Blow's Zone P17was found. Both these species first appearances seem diachronous between middle and lowlatítudes (Boersma and Premoli-Silva, 1986).
Species charactenstic oF the Gfobigerapsissemiinvolutus Zone are G. semiinvolutus, G.howei, G. index, G. luterbacheri, GlaborataliacerrOl1l.ufensis, GI. pomeroli, Globigerina rransdanubicá, and abundant G. linaperta (Figures 2, 3, Plates 1,2).
The uppennost pari of the Eocene sediments(20 m· 68 m) correspond to me Globororaliacerroazulensis Zone. The top of this zone coincides wilh the extínction of the GI. cerroazulensis group. Molina (1979, 1986) subdividedtbis interval into Cribrohantkenina inflata andC. lazzarii Zones based on the last occurrencesof these species respectively. These specieshave only been rarely observed in low latitudesections (KeUer, 1983, 1985). Because the lastappearance of Gl. cerroazufensis and C. lazzariido not occur at the same time, there is a sligbtdifference in age of the Eocene/Oligoceneboundary between MoJina's (1979, 1980) andBolH's (1966) zonal systems. Characteristicspecies of me uppermost Eocene are Glaborotalia cocoaensis, Gl. cuniaJensis, GI. increbescens, Gl. nana, Cribrohantkenina lazzarii, Hant~kenina alabamensis, H. brevispina, Pseudohastigerina micra larger than 150 microns, Globi-
MOLINA, KELLER, MADILE
gerina ampliapertura and G. gnlavisi (Figures 2, 3, Plates 1, 2).
The maio biostratigraphic event io the upperEocene occurs at the top of the Globigerapsissemiinvalutus Zone (sample 2, 4) where thisspecies goes extinct. In the middle latitudeMolino de Cobo section G. semiinvolutus is notas abundant as in low latitude sections (Keller,1986), therefore, the population change associated wilh tbe extinction of this species isless pronounced. Tbe Globigerapsis group goesextinct 23 m aboye the last appearance of G.semiinvolutus in the Molino de Cobo sectionand is associated with carbonate dissolution.This is considerably' higher than observed. inlow latitude sections where G. index, the lastsurviving Globigerapsis species, Frequently disappears sbortly above G. semiinvolutus (Keller,1986). Oiachroneity of last appearances of thesespecies must be assumed between middle andlow latitudes.
In sorne low latitude sections (DSDP Sites216 and 292) a microspberule layer of impactorigin is found near the extinction of G. semiinvolwus and tbe Globigerapsis group disappears shortly thereafter (Keller, 1986; Kelleret al., 1987). In OUT original sample analysisglassy microspberuJes were also found in tbeMolino de Cobo section in sample 4 near theextinction of the Globigerapsis group (Kel1eret al., 1987). However, in samples coIlectedsubsequently at the same locality we failed tofind microspheruJes. It is possible that wemissed coUection the ¡ayer, or lhat the microspherules carne from surface contamination ofa yet unknowo source.
Above the decline in tbe Globigerapsis groupand the dissolution interval in ¡he Molino deCoba section there is a major decrease, relative to other species, in tbe abundance of Globigerina linaperta and an increase in G. galavisi, G. ampliapertura, G. officinalis, G. ouachitaensis, Gfoborotalia nana and Gl. increbescens(Figure 3). This suggests a major paleoecologicshift possibly toward cooler climatic conditionswhich has also been observed in low latitudesection (Keller, 1986). A general cooling trend
495

MüLINA, KELLER, MADILE
during the late Encene is indicated in the oxygen isotope record (Corliss el aL, 1984; Oberhansli and Toumarkine, 1985; Williams el aL,1985; Keigwin and Corliss, 1986).
Eocenc-Oligocene Bounda!")'
The Eocene-Oligocene Boundary is definedby the simultaneous extinction of the GlaboroTafia cerroaz.ulensis eroup (GI. cerraoazulensis,GI. cocoaensis, Gl. crmialensis) and the genusHantkenilla and Cribrohanlkenina. To determine the nature of these extinctions in theMolino de Cobo section a 3 m interval acrossthe boundary was sampled al approximately25 cm intervals. This expanded bounclary section revealed that these species extinctions didnol occur simultaneously (Martinez·Gallegoand Molina, 1975; Molina, 1979, 1986). The G/oborotalia cerroaztllensis group disappeared before the genus Hantkenina (H. alabamensis,H. brevispilTa) 'which disappeared before thegenus Cribrohanlkenj¡1Q re. fazzariO (Table 1,Figure 2). Thus, Ihe Eocene-Oligocene boundaryextinctions appear to have occurred more gradually than prcviously assumed.
Although five species extinctions occurredacross the Eocene-Oligocene boundary, theoverall effect on the faunal assemblage appearsnot to have been very dramatic. This is bestillustraled by the relative abundances of thedominanl species in Figur~ 3. The' spedes goingextinct were general1y less than 10 % of thetotal fauna at the time of their demise; thesame' has been observed in low latitude sections (Kel1cr, 1983, 1985, 1986). No new speciesevolved at lhis time (Figure 2). No majar spedes abundance ehanges are observed (Figure 3), although lhere is a general increase inthe deeper water spedes Calapsydrax and Gioboquadrina venezuelana. This inerease in the
deeper water dwellers has also been observedin low lalitudes (Keller, 19B3, 1985, 1986).
There is, however, a cucious morphologicehange in the size of Psel/dohastigerina micraat the Eocene-Oligoccne houndary which hasalso been ohserved in the nearby Fuente Caldera seclion (Molina, 1986) and in low latitudesections (Kel1er, 1983, 1985). Psetldohastigerinamicra is common to abundant in upper Eocenesedimenls in the size fraction greatcr than 150mierons, hui disappcars from lhis size fraction aboye the Eocene-Oligocene boundary(Figure 3). In low latitudes this species is stillcommon lO abundant in the smaller than 150micron size fraction along with P. barbadoensis (Keller, 1983, 1985), but in Ihe Molino deCobo section P. micra is very rare whereasP. nagtlewichiensis s. L (which ineludes P. barbadoel1sis) is common (Table 1, Figure 2). Thecommon presence or absence of P. micra inIhe larger than ISO micron size fraclion is aseconclary indicator lo differentiate upper Eocene and lower Oligocene sediments.
Oligocene
According lo Bolli's (1966) zonation theOligocene is divided inlo the Cassigerhwllacllipolel1sis-Pselldohasfigeril7a micra Zone defined by the extinction of P. micra, and theGloberigeril1a amplil1.perll1Ya Zone defined bythe first appearance of Globorotalia opima opima. As noted earlier P. micra is very rare inthe Oligocene of the Molino de Cobo sectionand is therefore not useful as index species.The altemative middle lalitude zonation of Mo·lina (1979, 1986) provides better control here.The lowermost Oligocene Globigeril1a gorta¡¡¡iIG. faptlriemis Zone boundary is definedby the firsl appearance of G. taptlriel1sis andcorrelates to the P17/P18 Zone boundary of
Figure 2Range chart of planktonic foraminifers from the Molino de Cabo section.Relative thickness of range lines indica tes abundant, common or few specimens.
496

MQLINA KELLER MADILEUPPER EOCEHE
~LOWER OLlGQCENf UPPER OUGQCEHE EPOCHS
PAIABOHIAH ~ RUPELIAH " CHATTIAH AGES
G. 5EM- G.GOIITA G. TAPU-G. SEU.ii G. AHWLisUTURAli$ MOLINA • •C. INFLAIA <. l-.zv,..,¡ "lN\IOlUTil "" ~jEH:ois 1919. 86 il •>
.- .- > , • BLOW O
".- .,!• • , • ~ • , 1919
~G. SEJ4-
'.m~"'i~¡~.r~"'· ---::.~::;80<." "G.Of>i .... "l~ >
~iNWW¡A /N. lllellA~ ... PUI 19" ~
m 1:1$ ~ ~~~ • N ~ ~e ¡flól" w "i"~I; SAMPLES• 5PEClES
--- -- - G. urTUllACHERI---r- -- G. "'*EI- G. SEI'IIIKVOLlITA
- :'~"':11 ,. INDO· ..f- : e ". H. ALAI1N'IE"SIS_u - --- -- - - -- W"- -~ - - ~ -- - - --- ·~
H. BREVISI'INAn_ e. INFLAT"
-- · . C. LAZZARllu _
",:.:",:- ":". <5':''''-:-- ---, -- f- .... 0 ".0'P. MleR" S.L.• ;: O'u_ ------------ -.~ P. ~GUEWleHIENSIS S.L.- 1>. I'IlMUlDLl_u1-"'- ---- G. CERflO"ZULENSIS- --- G. eOCOAENSIS
- n - - -- --- - G. eUNIALENSIS
- --- -1~ ---=-::,. 1IICIlEBESeENS-,- f-' --,- G. ~N"
- 6. OPI"'"uu ___
G. SI"kENSIS-- --- G. PSEUOOCOflTlNUOSA
- - - n G. Ge-
- -- - G. el.Ll\EIICl.o.E
-- 6. eF. l"BESA
+ G. PSEUDONtPUN'ERTURA
- ¡. •~ • 6. llNAPERTA-- u 6. TF'.ANS!lANUlICA--~- - - - ------ - - u_1-- f-::: -" ~. G. ANGlPOROIDU.- --- - - - - G. ll"LAVISI
---1- __ u_I-u- -- -- --~--- n- -- - ,. TRIP"RTITA
-- - .-- - - 6. TAPURlENSISu -- ---------- G. SELlll
- - •• IINAJENSls--f-- ----~ ___ n
n --- -- ~- G. EOC"EN"-- ---1------- f- -r-- - -- - G. CORPULENTA-- -- u _ - - - - - - - ---o .- u _n -- --- --- G. GORT"Nll
- - - - - - 1- - •• PIl'''ETURRlT ILI NA--- ----- - 6. ",",PLlAPERTURil-- - n - - - - ---- - - - 1- - ---- - G. EU"PERTURA..-~ -- - • - 6. VEtlHUELANA
---- ---- - -- + - -" - - u - - - G. OFFIClNAlIS
- u - 6. "NGUl!OFFICINAl IS
~t:'. ANGUl! SUTURAlI S-- • - -- -- - 6. ANGUSTl~ILICATA
- -- • -- -- • - - - -- G. OUAOUTAENSIS
- - - G. CIPEflOEIlSlS
- --f-- - -- - -- - - - G. PP.AEBUUOlDES. f- - -o -- - • 6. OCCLUSA-- f- • 1-=" -- - ti. UROYI-- -- • _u 1- - -----1- - - - - - 6. SOlERI• • C. UNlCAVU$ S.L.
- - - - - n u - C. DISSIlUllSh_ - C. CIPEROENSlS
-- ---.r----- - G. GLOIUUR lS
- - - G. CF. GLOBOSA- 1--- - • - - - - (tI. CtrBENSIS S.L.
- • - - - - C. CHlPOLE~SIS
Figure 2 497

MOLINA, KELLER, MADILE
Blow (1979). The first appearance of Globige·rina aflgulisuturalis defines lhe C. selli/G. anogulisuturalis Zone boundary and correspondslo Lbe base of Zone P21 (Figures 2. 3). All thes~
species are rare or absent in low latitudes secliaos (KeUer, 1983, 1985).
The lower Oligocene is marked by a relatively low diversity bUl stable faunal assemblageconsisting primarily of Late Eocene survivors.There are no majar species extinclions or abundance changes apparent. Faunal assemblagesare charaeterized by the coa! water speciesGlobigeriJ1a linaperta, G. galavisi, Globorota.lianana and the surface dwellers Globigerina amopliapertura and Gl. increbescens.. Except forGI. nana, these species ahruptly disappear between samples 11 and 11'3 marking the lower/upper Oligocene boundary (Figures 2, 3). Theevolving species Globigerina angulisuturalis,G. binaiensis, Gl. siakensis, Gl. pseudocontinuosa, and common typical Globigerina cipe·roensis appear aboye lrus interval (Figure 2).A shon hiatus of about 1 m.y. (31.5-32.5 Ma)may be presenl at this faunal change as suggested by sediment accumulation rates. Themajar faunal turnover and short hiatus coincide with a major drop in sea level postulatedby Vail and Hardenbol (1979) and Haq et al.(1987). Widespread erosion or nondepositionoccurred at lhis time globally as observed bythe widespread distribution of Lhis hiatus(Keller el al., 1986).
CALCAREOUS NANNOFOSSILS
Calcareoux nannofossils from Lhe marly intervals are generally abundant and well diver·sified. Preservation varies from poor lo mode·raleo Reworked Cretaceous and Eocene nannofossils have been identified lhroughout thisseetion (Figure ~).
As a resul t of reworked nannofossils tbe positian of the zonal boundaries based on lastoccurrences are often uncertain. To circumvent this problem a semiquantilative analysis(Figure 4) lo estimate the relative abundancesof the domioant species has heen roade· (Fi-
498
gure 5). Nevertheless, lhe zonal'boundaries arefrequenlly not in tbe standard relative positionto lhe planktonic foraminiferal zones as repor·ted by Beruren et al. (1986). Despite theseproblems a complete sequence of Late EoceneSubzone CP15a to Late Oligocene SubzoneCP19a has becn identified using the biozonation of Bukry (1973, 1975) and Okada andBukry (1980).
Late Eocene
The late Eocene assemblages are characte·rized by abundant Cyclicargolithus Iloridanus,Coccolilhus pelagicus, Dictyococciles bisectus,Ericsonia formosa, Discoaster btirbadiensisand D. saipanensis (Figures 4, 5). The genusSpllenolilhus ranges from the late Eocene tothe early Oligoccne and is represented byS. moriformis and S. predistenlus. Sphenolitifus pseudoradians used by Martini (1971) todefine the base of Zone NP20 was nol found.Tbe genus Chiasmolilhus is very rare and itis lherefore not possible to cakulate the Discoaster/Chiasmolitlzus ratio used by Bukry(1973) as indicator of water temperature. Zygrhablithus bijugatus, Lallternithus minutusand a few solution resistanl Pedinocyclus larvalis were also found; all these species areconsidered as indicative of ncar shore environments.
The first appearance of lsrhmolithus recurvus which defines the base of Subzone CP15boccurs near the base of the Molino de Cabosection (sample 1,4 m). The extinction of Cribroce'ltrum reticulatum occurs just below theextinction of the disc shaped Discoasters asalso reported by Nocchi et al. (1986) andPerch-Nielsen et al. (1986).
A major biostratigraphic event is lhe declinein abundance and extinction oC Discoaster barbadiensis and D. saipanensis. Relative speciesabundance analyses indicates that Discoasterbarbadiensis is always more abundant thanD. saipanel1sis and both appear to decline simultaneous·ly. The top of Zone CP15b hasbeen placed at sample 4 where a strong de-

MülINA, KEllER, MADIlE
...
FORAMINIFERA
ABUNDANCES
PLANKTONIC
DOMINANT SPECIES
•~•
"1~1- l'\§-Jt5 ... ,.
"•
"
•
•••ª ~, u
i•u
> U•"• •,~• uiu
" "u,•
"
•,§
,i!•
Figure 3
Relative percent abundances of dominant Illanktonic foraminifcra! species inthe Molino de Cobo section. Note short hiatus al the LowerjUpper üligocene
boundary. 499

MOLINA, KELLER, MADlLE
crease of D. barbadiensis and D. saipanensisis observed. The number of specimens/mml ofD. barbadiensis decreases from 43 in sample3 lo 13 in sample 4 and lO 2 in sample 4, 5.Likewise D. saipanensis decreases from 16 spe·cimens in sample 3. 5 to 3 in sample 4 andlo 1 in sample 4, S. A similar decrease in thesespecies abundances was a150 observed in lhenearby Fuente Caldera section by Monechi(1986) and Perch-Nielsen el al. (1986). The fewspecimens of Diseoasters above sample 4 areconsidered reworked (Figure 4, 5).
Imerestingly. the strong decrease and probable extinction of D. barbadiensis and D. sajopanensis in t..he Molino de Cobo seclion coin·cides Wilh the decline and extinction of theplanklOnic foraminifer genus Globigerapsisand [he dissolution interval. No quantitativeanalysis of nannofossils are available fram lowlatitudes at this time. Therefore, it is notknown whether the decline in lhe nannofossilspecies con-eiates preciscly with the micraspherule layer.
The uppermost Eocene layers between theextinction of D. barbadiensis and D. saipanensis, and the Eocene/Oligocene boundary (basedon planktonic foraminirers) are characterizedby the rollowing variations in nannofossils.
1. A slight increase in lhe abundance ofISll1molilhus recurvus, indicative of relative1ycaoler walcrs. This increase has also been observed in lhc nearby Fuente Caldera sectionby Monechi (1986) and Pcrch Nielsen et al.(1986), in lhe Umbro-Marchean Apennines (Ita·Iy) by Monechi (l986). Nocchi et al. (1986, inpress), Premoli Silva et al. (in press). as wellas in DSDP Site 522·522A (Soulh Atlantic) byBackmann (1986), in Sitc 362-363 (Angola Basin) by Proto Decima (pers. comm.) and inseveral si tes of the Pacific Oeean by PerchNiclsco (1986).
2. A slight decrease in lhe abundance ofEricsonia far/nasa ..
3. An increase in abundance or L. miltUfusand Z. bijugatus ncar the Eoccne/Oligocene
500
boundary. This event has also b....---en observedin rtaly (Nocchi et al.. 1986) and in Hungary(Baldi el al., 1984).
Early Ollgocene
rt was not possible to subdivide the earlyOligocene Zone CPI6 inla subzones CPlóa/CPI6b (Ericsonia subdisficha/E. formosa) because the acme of E. subdisficha which definesthis subzonal boundary (Okada and Bukry,1980) cannot be recognized. Ncither could theacme of E. obrufa be recorded which was usedby Madile and Monechi in the Contessa Highway section to identify the same boundary(Premoli Silva et aL, in press). The. extinclionof EricSO'lia formosa which defines the top ofSubzone CP16b is placed in sample 8 (89 m),although variable abundance is presenl throughout lhe earIy Oligocene.
In subzone CPlóc an increase in abundanceof L. mimltus is observed. A similar increasein abundance of L. minufus and Z. bijugatuswas observed in CP16b/c Subzones of the Conlessa Highway of Italy by Premoli Silva et al.(in press) and in Hungary by Baldi el al. (1984)who interpreled this abundance change as adecrease in surface water temperature.
The extinction of Reficulofenestra umbilicadefines the top of Subzone CPI6c (R. hillae).As wilh all nannofossil last occurrences in theMolino de Cabo section, reworking of oldersediments makes it difricuh lo determine thetrue extinction of index fossils. It is assumedthat lhe extinctions of, R. wnbilica and L. mi·nutus occur in sample 9'4 (112 m) near theextinction of l. recurvus, altbough rare specimeos intcrpreted as reworked occur above trusinterval. In sorne sections where R. umbilicais rare (e.g. Contessa Quarry in Italy, Lowrieel al., 1982), the extinclion of l. recurvus hasbecn used to mark the top of Subzone CPI6c.This suggests that the top of Subzone CPI6cis inded c10se to sample 9'4 of the Molino deCobo section.
The first occurrence or S. dislenlus in sample 10'6 (137 ro) marks the base of Zone CPI8,

MülINA, KEllER, MADlLE
MOLLNO DE COSO Cale, nannofosslls
. '.
. . O
. O.
·00·onO'U
:On·oU
OOO
OO O, .o· O·0
l ~
O·"
O'
..
:ó~·O::. .. .. .. . .
~Ó.O ~
~....
o·
·0
: ~ I1I : : U'l:' ....
. O.
. O.
~~:nH-'U
••
•
·.: O·.
..·.
..
. O.
. O.
O
• • •
1::::. .....
: :n: . .O' , :.:
O ' . . U. . O : . O: : : ~ :•
~' ; O• : ~ • • : ¡:O; : ~ ~ ..o· .... n •..•n ... U +,0,'" ++¡ 1 ;~~:O ;O~· ~ .~ ~~IIII!li• .¡. .... .. I
O -0+ '+0- +
~ -'---"-'-'~~-L:-'-':~L-'-':~,-,-,:~'-'-'''-'-~. : "-"-~-'--':~:-'UiI..lLlJ~·--,,-"--,--,''-''--'-
·...·.
·.·.'0. ..·.·.
, ., .., ..
:~O·.··0...• O·.·.
O
O
·.·.lO - ... -
•
•••
••
c.
"
c.
"
c.m
•c. •'" •
H- •!ce- •
c. ~ •".,.
•
c.oo,
1IlW
1Il ...J
~ ~o <:
~~
Figure 4
Range chart óf calcareous nannofossils fmm the Molino de Cobo section.The assemblages were called abundant (A) ir the relative abundanceS exceeded60 % af all componcnts, common (C) it there were more lhan 40 % coccoliths,few (F) tor more lhan 10 % coccolilhs and rore (R) tor a relaLive abundanceaf identifjable coccolith af less than 10 %.Relative ahundancc af calcareous nannofossils is based on the rrequency aflhe species per ficld of view in the Light microscope al a magnificaLion of 1500.The species was considered: A • I species x neld of view
e • 0,5-1 species x field of view+ Reworkcd species F ~ 1-4 species x 10 field of view
R o 0,2-0.9 species x 10 field of view
501

MOLINA, KELLER, MADILE
, , , ,7h b ' o , i , '7J
•¡,•
•,,",
,•••••o••o
, i 'j '70 ~ ¿'
Thc Globigerapsis exlinction 'event in theMolino de Cobo section is associated with adissolution intcrval and involvcs thc cxtinctionor a1l species of the Genus Glubigerapsis aswell as Globigerillll Iral1sdtlllllbica and Globorowlia pUl'l1eroli and coincides with a majordecline in disc shaped discoasters. A catastrophjc decline in thc dise shaped diseoaslers atIhis leve! has al so been observed in the nearbyFuente Caldera seelion (Perch-Nielsen et al.,1986) and in Umbrian sequcnees or haly (Nocchi et al., 1986). These species extinctions andfaunal abundance changes imply a majar paleoeeanagraphic event al Ihis time.
Oxygen isolape dala summarized by Keigwin and Carliss (1986) show that a globalcooling Irend aecurred during middle to lateEocenc lime. This coo!ing trend resu1ted ingradual replaecmcnt of warm waler species.
-..--,
r~, ..
"i~ r~.. ;-
-=:::::
l~~I~
~It15'
, 1~0 'o I
,••••,••, , , j' 'j , I , ,
N.\1lNOrOSS1LS
" zmlES~
SAIIPHS
< 0_,MAl\TIMl
.U~aY
,,~ 1111
" "... " " "u " ..z ..u " '0u " 13.~
"o c-- " n "~ 10.6H " 'o~ " 11 ,J 11,7
o m ---- ,., ,.." " • O.',~
_R~ O,, 8,2n- O, '. ,f-- o.' ..,
" " " , 6, ~91h/b " o.'• O••• ¡.•
",'---- ---- • ,..U
" ",
Z '.'" '" 19/20U '."O ,"
... ," "
o.; O
"" ".
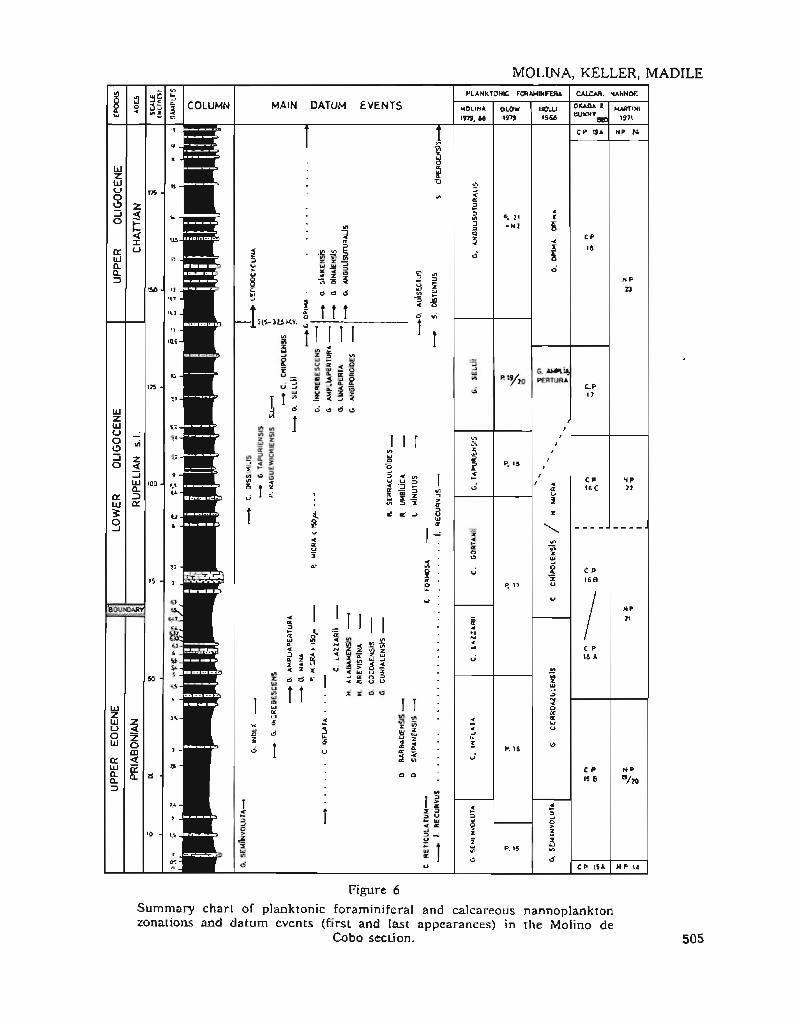
The correlation between planktonic faraminifers and calcareous nannofossils is il1ustrated in Figure 6 along wilh the con-cspondingEW'opean stages. Over 40 first appearances andextinctions uf stratigraphically significant species (see Plates 1-3) have becn irlenlified as usefui dalum events. Mast uf tbese datum eventscluster around three time periods: the lateEocene, al ¡he Eocene-Oligocene boundary, andal the early lo late Olígoccnc boundary. Severa}species extinctions and first appearances a1sooccur scattered in the early Oligocene, bu!these species are general1y very rare (Table 1).
DJSCUSSION
::md the top of lhis Zone is marked by thefirst appearance uf Sphcl10lirhus ciperoensisin sample 18 (191 m).
Figure 5
Relative abundances of domimml ea1careous nannofossils in the Molino deCobo seclion. Abundance cxpresscd in numbcr of spccimcns pcr mm2 at a
magnif.ication of x 1250.
502

MüLINA, KELLER, MADILE
Eocene-Oligocene boundary exlinctionevent is not associated \Vilh any calcareousnannofossil specics cxlinctions (Nocchi et al.,1986) and no majar spccics abundance changes
lerreslrial impact evenl. TI is nol yet clear
whclher lhc extinction of Ihis gmup in IheMolino de Coba scction con-elates lO this evenl01' is diachronoLls, espedal1y since we failed lo
reconfirm our earlier microspherule discovery(Keller et aL, 1987).
The
The abundance decline <lnd subsequent extinclion of Ihe Globigerapsis group could Ihereforehave becn a consequence of this global coolingtrend_ However, Lhis inlerpretation does notcxplain lhe near inslanlaneous decline in theGlobigerapsis group from a high of more Ihan60 % lo I % in low latitudes al Lhe time ofdeposition of a microspherulc layer. or from20 % to zem in ¡he Molino de Coba section.The de mise of Ihe Globigerapsis group in lowlatitudes \Vas probably has tened by an extra-
.,~ ~ ~ ~ ~
": ~
~ ~ ~ ~ ~o........_,a"""o. .,,."~f(;.'..\;.o......" ..t ...h.Q, _'......l_O. _h......- .O•• ,.. , ..•.•1...-'••. d. ,,__'o'.01. ,'.-..'_•. "",,"l~'"
".. _t.....•. ,.1_101•. ,lobol.".a.,_a.....t_l,a._'a......'a."",_;.a.l_'
, ••• I , . ,
··• 3!l ti" ..... 11,.
• •'" •
" •, , •·. ,
' .. " • ,," •• .
"
".,•
·..· , ·· , • • • •" '" • , 'lI,.lO •, , , • ,
• • • • •, , , · ·, , • , • • , • • • , ", • , liza , ""lO" , "
11 o I)t~ ~ .. c21 .>lID
.'0 r 5 '15"'Z14'Z"15,""
a. "_"O. "_t~w". 0011.a • ..." ...O••"'d•• ll.O. _,,_.... _"_'.....•. 11.'_a t '''l''.•. _]ll.. '-''''''•. ',_l~.... 1<'_""Oo, _1_01. .--..1_'.01. _1.01. _ ••,_,.01.' _
al .".d._O, ....l_ •.••
• 1. _,.Il01.""--"_01. ., "01. ••, ..c.. ,_.... ......,Co.l.t._.."". -,,--,.,.o.t........... __ ,.1Il.o.•,., ••"., cl__..•. _'ro ..O. _''''' loo Ooll.l...l_l,........'''''l_
,•,
,• ••
•1> ."
" _. ,li .,. Il~
" •• 1
'" o •I r ,•
"'"
•• T • 3 •• 1 • ,
""'lIltll.
",
'",", , , ,
•'" .. • •
. • •., , • , • •.. ... • ,," "
• • • • " .• •• · '", • ,," ,n, • ,...... ., • ...
, , ,. • ,, · • lO • • .. • , •
• , · , • ,, • • ." .. " .... " • • ,
'" • , , ,
.. ,.rl.,. ... ,~ ...." · , ·r. _'"'''''' •. 1. • • • , •...,... • • • "'o • ."o. _lO'. ,. • • • • • • • , • , • •-- , • • • • • • • ,lIo'''''lfl... .. • •, • •...,.1 -.o-t.
~ ¡ , , ,,i' ¡ - --. ,, ! -¡ ,-i ¡ ! •, ! ¡ i , - i i , i, ,~ ¡ ~ i i ; ~ ¡ , ¡ ~ ¡ ,
Table 1
Percent abundance data of specics in Lhe Molino de Coba scclion bascd onpopulation counts of 300-500 individuals. X = lcss than 2 %.
503

MaLINA, KELLER, MADlLE
are evidcnt among planktonic foraminifers(Keller, 1983. 1985, 1986). However, large(> 150 microns) Pseudohasrigerina micradisappears shortly after the boundary. ln addj·tion. lhe five foraminifer species extinctionsoccur more gradually Ihan previously thougbt.These faunal changes may be related to pro·gressively adverse environmental condilionswhich may be related lO the maximum developmeDt of lhe psychrosphere, or two layer oceanwith cold bollom water and warm surfacewater al this time (Keonett and Shackleton.1976; Keigwin, 1980).
The earIy to late Oligocene extinction even t
is somewhat obscured due to a possible 1 m.y.hiatus. Nevertheless. a majar faunal turnoveroccurs during Ihis interval with four speciesextinclions and seven firsl appearanees including the nannofossil Dictyococcites abisectusand Spheno/ithus distentus and the larger benthic foraminifer genus Lepidocyclina whichappears at the 10p of lhe early Oligocene (Figure 6). The four planktonic foraminiferal spedes Globorotalia increbescel1s, Globigerina ampliapertura, G. linapena and G. angiporoidesgoing extinct are Eocene survivors which constitute between 30-40 % of the foraminiferalassemblage. Their clemisc occurs globally althis time ·(KeIler, 1983, 1985). This major faunal tumover may be related to the globalcooling recognized by Keigwin and Keller(l984) at this time and to the drastic sea leveldrop noted by Vail and Hardenbol (1979) andHaq el al. (1987).
CONCLUSIONS
Three major faunal event are observed inlhe upper Eoccne to Oligocene strata of theMolino de Cobo section:
1. Extinction of Globigerapsis group anddisc shaped discoasters. This Faunal event involves the demisc of the Globigerapsis group(e. il1dex survives in lo the lalest Eocene inhigh lalilude sections). Species of the elobige·rapsL~ group constitute 20 % of the lotal foraminifcral populalion at the time of eXlinction.
504
The dise shaped discoasters D. saipanensis andD. barbadiensis also decline dramatically atthis time. This faunal turnover is associatedwith a dissolution event and a microspherulelayer in low latitude deep-sea sections.
2. The Eocene-Oligocene Boundary extinction cvent involves S planktonic foraminiferalspecies, which did not go extinct simultaneousIy, but stretched oul over a 3 m interval. TheGloborotalia ccrroazulensis group (el. cocoaen<;is, GI. CL/nialensis) disappears before the ex·tinction of the Genus Hantkenina (H. alaba·mensis, 1/. brevispinaJ, which in tum preceedsthe extinction of Cribrohant kel1ina (e. lau.arii)and Pseudohastigerina micra > 150 microns.This gradual extinction event appears to berelated to adverse environmental conditionsassociated with the maximum development ofI he psychrosphere.
3. The early/latc Oligocene faunal turnoverevent involves thc extinclion of Eocene survivors (Globorotalia increbescens, Globigerinaampliapertura, G. /inaperra, and G. angiporoides) and the evolution of late Oligocene to Miocene spedes (Globif!,eri/la binaiensis, G. angtl
Iisuturalis, and el. siakensisJ as wel1 as twonannofossi¡ spccies (Dictyococcites abisectusand Spllenolitllus distentus). This faunal turnover appears to be rclatcd lO paleoceanogra·phic changes associatcd with a global coolingand majar sea leve! drop.
ACKNOWLEDGEMENT
We would Iike to thank I. Premoli Silva andS. Monechi for their comments. This researchwas supporled by the U.S. - Spain Joint Committcc for scientifie and Technological Coope·ration and by M.P.1. 60 % (S. MoneehO.
REFERENCES
BACKMAN J., SIlACKU:rON N. .l.
1983 Quulllilalive bioc/¡ronology 01 PUocenc. mldEarly PleiSlOcc.J1I: calcareOI/S nUIlIJOlossils Iro/11AtltJnlic, Jndian alld Pacific Dcealls. Mar. Mi·cropalcont., vol. 8. pp. 141-170.
o.n
o.n
e" lO"l' e 11
'""
",..
,...
e" ...."' Illro
"...I
1!•¡•
C" '" .... l'
,~,
,!~•
io•u
" I----+---j,,,,,,
".15
p. "00'
... "
.."
MaLINA, KELLER, MAOlLE
'3•
ª,,
•;!,
Io
OOOU"''' lllOW lIOlU''''. M 1m 11M
l'.,, u• u..g.¡
1i
o •
•
1 I I.jla .~ ~~ ,
~ ~-i
EVENTS
~ I r r r• l! •3~ ~ g~u!.:~~
"'.oc o:>... u u
~ :It .,; ..
t
u
DATUM
•
MAlN
t
Ji~ '" d e
~ tI•~•
.i•
l
!~1•
'"
•
B•
Figure 6
Surnmary chart of planktonic foraminife.ral and calcareous nannoplanklonzonalions and datum cvents (rirSI and lasl appcarances) in the Molino de
Cobo seclion. 505

MOLINA, KELLER, MADILE
flAWI T. el al.
1984 Tlle Eocene Oligocene hall/ldar", in HWlgary:Tlle Kiscellian Stuge. Acta Ceo!. Hung., vol. n.pp. 41-65.
IIERGGREN W. A., KEt-..,- D. v., FLYN J. J.
1985 Po/cagene geocJ¡rollofogy olla c/lrOIlOslraligrtl'p1zy. Geol. Soc. Amer. Bull., vol. 96. No. JI.pp. 1419·1427.
BLOW W. li.
1979 The CailJuzuic Globigerinida. Leiden, Brill, ] vol.
BOERSM" A. o PRUIOL.l-SILVA l.
1986 Terminal Eocene Evctl/s: plollktollic (orami.,¡¡{era ami isotopic evidenCI!. Developments inPale. and Slnuig., \'01. 9, pp. 213-224.
BOLL.¡ H • .\\.
1966 Zorra/ion of CreraceOl4s lo Pliorenc marine se·diments based 011 pfallktollic lorumillifera. Bol.1nfor. AsSQC. Venez. Ceo!. Mine. y Petrol., \'0
lume 9, pp. ]·31.
DUKRY n.1973a Coccolilll olld Silicofragellale slroligrapllY Tas.
mml sea alld sOlltll\vcstem l'ucific Ocean D$DPleg 2/. In R. E. Burns et al., ¡nitial rcports ofDSDP, vol. 21, pp. 885-893.
1973b Luw lat;flldcs coccofitll bioslratigrapJty ~olla
tiOll. In Edgar N. T. et al., Inilia1s Reports oCthe DSDP vol. 21. pp. 685-703.
1975 Coceofi", and 5;lico{lagellare stratigrap1Jy.Northll'cSlerl1 Paci{ic OceuII DSDP Leg 32. InLarson R. L el al. ¡nitial RcpoTls oC the DSDP,vol. 32 pp. ón.701.
CO.\lAS M. c., .\IARTlNEZ-GAU.ECO J., .\IDUNA E.
1984--85 Lito{acie.{)' sucesiÓn cslraliráfica de/ Eoce110 y Oligoceno al Norll! del cerro Menca/(10110 Stlbbit;cu Proll. de Grallada). Cuad.Cco!., vol. 12 pp. 145·155.
CORI.ISS O. 11., ¡\UBREY M. P., BIlRCGREN W. A.,
FENNER J. M., KEIGWIN L. D. Jr., KELt.ER G.
1984 Tlle Eocene Dligocem:: boundary Eve", in theDeep Sea. Sciencc, vol. 226, pp. 806-810.
O'IIONDT S. L.. KELLER G., STAl.L\RD R.
1987 Ma;or element composiliollal variarioll \Villlinalld betll't!en diflerell/ late Eocellt!' ",icrolektite streIV {ields. Mclcorilics, vol. 22, No. 1,pp. 61-79.
GL\SS O. P., IlURNS C. A., CROSIlll! J. R., lIUUOIS D. L.
1985 Late Eocene Norl/r American microteklile.\· alldcli'IOpY"oxe¡¡c-bcar¡',g sp/rerulcs. Proee. 16thLunar and Planct Sei. ConC. Jour. Gcophys.Res., vol. 90. pp. 175·196.
HAQ B. U., t.OH,\IAN G. P.
1976 Early Cellol.u;c IIalllloplollklon b;ogeograpl,yu{ file Allanlic Dccan. Mar. Micropalcont., vol. 1,pp. 119-194.
PLATE 1
Scale bar = 100 ¡.lm
1. Hatllke"ina aloballlf!lIsis (Cushman). sample 1.5. lop of Gfobigerapsissemiinvolutus Zonc.
2_ Globigerop!is semiinvolulUS (Kcijcr), samplc 2. G. semiinvolutus Zone.3, 4. Gfobigerapsis index (Finlay), sample 2, G. semiirlvo/utus Zone.
5. G/obigerapsis interbacJlCri (Bolli), samplc 2, C. sf!miinvolutl/s Zone.6. Globigeropsis 1Io\Vci (Blow and Banncr), samplc O, G. semiirlvolr41us
Zonc.7. '8. Gfoborotofia pol1leroli (Toumarkinc and Bolli). samplc 1,5, C. semi
illvoluiHS Zonc.9, 10. Globororolia cerrOGl,ulellsis (Cole). sample 2. G. sellliillvolutlls Zonc.
11. 12. Cloborotalia cocoaellsis (Cushman). samplc S, GI. cerroazulellsis orC. inllala Zone.
13, 14. CribruluUllkenilla i/Jflala (Howc), sólmplc 2, C. sell'liiHvo/u/tls Zonc.14, 15. Cribrollalllk·enina lazzadi (Pcrieo1i), samplc 6.4, Globoro/lllia cerro
al.ulellsis or C. laZl,a,.ii Zone.
506
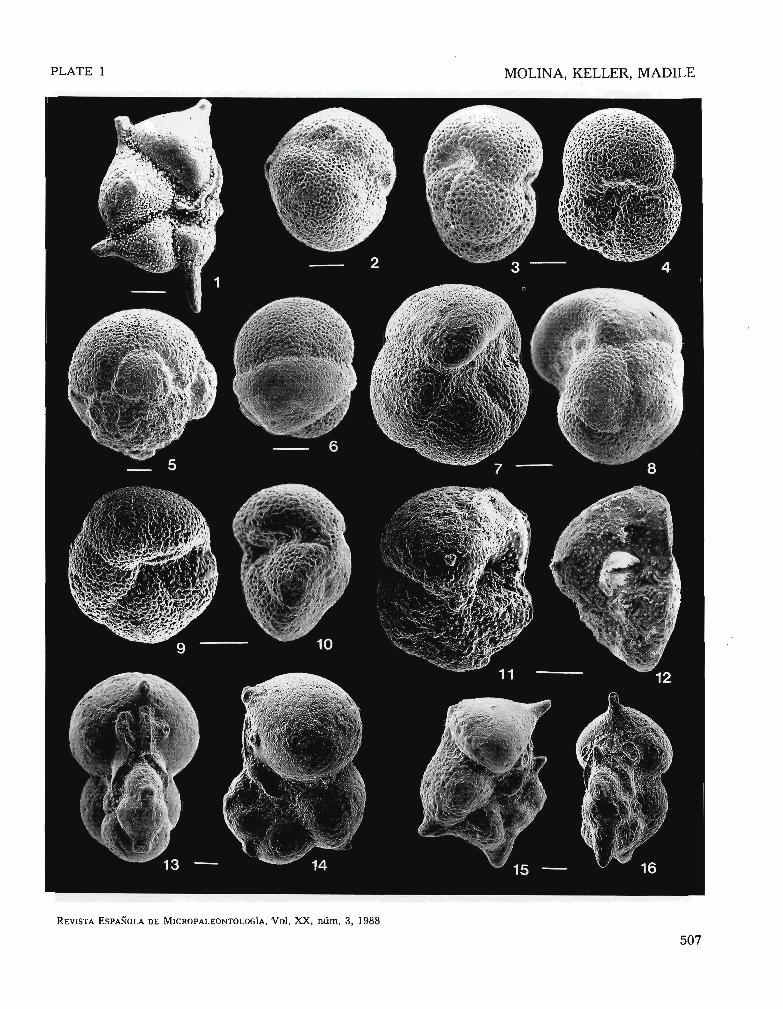
PLATE 1 MüLINA, KELLER, MADILE
REVISTA ESPAÑOLA DE MICRO PALEONTOLOGÍA, Vol. XX, núm. 3, 1988
507

MaLINA, KELLER, MADILE PLATE 2
REVISTA ESPAÑOLA DE MICRQPALEONTOLOGÍA, Vol. XX, núm. 3, 1988
508

HAO B. H., PREMOLl-SH.VA l., LOHMAN C. P.
19TI CalcareollS pfankto" biogeograpllic l!vidence for/IIajor e/imafic flllCtllatiOIlS in lile earl)! Ce1lo-
HAO B. 1-1., llARDENBOL J., VAIL P. R.
1987 C/tremology of fluclllaling sea levels since lileTriassic. Science, vol. 235, pp. 1156-1167.
HUT P., ALVAREZ W., EUIER W. P., l-IAN5EN T.,
lCAUFFMAN E. G., KELLER G., SH OEUAKER E. M.,
WEISSMAN P. R.
1987 Comel showers as a passlble cauSe uf massextillcli01ls. Naturc, vol. 329, pp. 118-126.
K.E..lGWIN L O. J r.1980 Paleoc:eal1ographic cllange in lIle Pacific al lile
Eocen¡;·Oligocene Boundary. Nature, vol. 287,pp. 722-725.
KEIGWIN LO., KElJ..ER G.
1984 Middle Oligoccne e/imolie ellonge frotll equotoriol Pacific DSDP Sile 77B. Geology, vol. 12,No. 1, pp. 16--20.
MOLlNA, KELLER, MADILE
ltI!.lGW1N LO., CORUSS 8. H.
1986 Slabfe isOIOpes in Lale Eocelle to Ofigocel1eForomini/era. Geol. Soc. Amer. Bull., vol. 97,pp. 335·345.
KELLER G.
1983 Bioc1lrollology olld Palcoclimalic implicolionsof middle Eocene lO Oligocelle planklOllic /oro.mini/eral faunas. Marine Micropal., vol. 7, pa·ges 464-486.
1985 Eocelle and Oligocelle slraligraplry olla erosiollal uncullformities in lhe GIIJ/ of Mexicoond GIIJf Cuasi. Jour. Paleo., vol. 59, No. 4,pp. 882·903.
1986 Stepwise Mass Extillction ond ¡/IIpaCI Evenls:Lale Eocene lO Early Oligocene. Mar. Micropal.,vol. 10, pp. 267·293.
KEU.ER e., U'HONUT S., VIILLlER T. L
1983 MII/liple microlektite 110riZOllS ill upper EocelleMarine Sedimenls: No evidence lor Mass ExIinctions. Seicnc!:, vol. 221, pp. 150-152_
PLATE 2
Scale bar = 100 um
1. 2. Globigerina cocaella (Gumbel), samplc 6,49, Cribrohantkeniru1 lauarUor Globormalia cerruazulensis Zonc.
3. 4. PseudolrllStigerina micra (Cole), sample 2,4, Gtobiguapsis semiinvolu/us Zone.
5. Pseudollastigerina barbadoensis (Blo",,), sample 9, Globigerina /apu·riensis Zone.
6. Clliloguembelina cllbensis S. L. (Palmer). sample 9, Globigeril1a lapu-riellsis Zone. '
7. Globigerina galavisi (Bermudez), sample 6,48, C. lazzarii or Gl. cerroazulensis Zonc.
8. Globigerina tapuriensis (Blo"" and Banncr), sample 9,7, GlobigerinaseW or G. ampliaperlura lonc.
9. Globigerina selli (BorsclIi), sample 12, GtobigerirTa angulisuturalis orGloboro(alia opin¡a Zone.
10. Globigerina bil1aiensis (Koch) , sample 11.3, G. angulisuturalis or Gl.opima Zonc.
11,12_ Globorotalia nana (Bolli), samplc 5,6, C. lazzar;i or Gl. cerroazulensisZo~nc.
13. Globigerina gorlmlii (Borsctli), sample 13,5, G. al1gulisuturalis or GI.opillla Zone.
14. Globorolalia siakensis (Le Roy). sample 12, C. al1gulisLHuralis or Gl.opima Zonc.
15. Globiger;na angulislltllralis (BolJi), samplc 12, G. angulisulllralis orGl. opima Zone.
16. G/oborolalia opima (Bolli), sample 11,7, G. allgulisuturalis or Gl. opima Zone.
509

MOLlNA, KELLER, MADll.E
KEU.ER G., HER8F.RT T., DORSEY R., JDHNSSON M.,
CHI W. R.
1986 Global distributi011 o( lAte Paleogene Hiatuse.s.Gcology, vol. 15, No. J. pp. 199-203.
KEU.ER c., D'HONDT S. L. ORTH C. J., GIL\'ORE J. s.,OUVER P. o, SIiOEMAKER E. M., MOUNA E.
1987 lAte Eocene ImpaCI Microspherules: Slratígraphy, Age and Geochemistry. Meteo;'ilics, vol. 22,No. 1, pp. 25-60.
KENNETT J. P.
1977 Ceno~ojc EWJlution o( Anlarclic g/adalion, Ihecircum hllarctic Ocean and their impac/ onglobal Paleoceanography. Jaur. Geophys. Re!.,vol. 82, No. 27, pp. 3843·3860.
LOWRIE W. el al.
1982 Paleogellf~ magnelic s/ratigraplly in Umbrianpelagic carbonate rocks in tJu: Conlessa secorions, Gubbio. Ceol. Soc. Amer. Bull., vol. 93,pp. 321-325.
MARTINEZ-cAllECO J.
1977 Estudio micropaleontof6gico del Nummuliticode un sectOr comprendido entre Mort!da Pina,Pedro Martinet. (Zo"a SlIbbilica). Tesis Doct.Univ. Granada, No. 175, 264 pp.
MARTlNEZ-GAll.F:CO J.• ),lOUNA E-
1975 ESl/(dio del lrdnsito Eouno-oUgoceno con roraminiferos plallct6nicos al Sur de Torre Car·dela (Prov. de Granada. Zona Subbilica). Cuad.Geol.• vol. 6. pp. 1TI-195.
MARTlNI E.
1971 Standard Tt!rfiary and Quaternary calcareousnannoplankton lOna/io". Proc. Second Plank.Canf. Roma, 1971, vol. 2, pp. 739--785.
MAURASSE F.• CIASS, B. P,
1976 Radiolarian stra/igraphy and Norlh Americanmicro/ektiles itl Caribbean Core RC9-58:¡;;jplication cotlcernin, La/e Eocene radiolarian
PLATE 3
The abbrevialions XN and QL de,note cross-poIarized and trasmited light.
1. Discoaster de/landrei Bramlelte and Riedel, sample 3,5 (OL) x 2200.2. Discoasler lani nodifer Bramlette and Riedcl, sampIe 3,5 (Ol) x 2200.
3, 4. Cruciplacolill1us crux (Deflandre and Fert) ROlh, samplc 3,5. (3) OL,(4) XN. x 2200.
5, 6. HelicospJtaera euphratis Haq, sample O. (5) Ol, (6) XN. ¡( 2200.7, 8. HelicosplJaera bramlettei Mullcr.
9, 10. Pontospllaera plana (Bramlelte and Sullivan) Haq, sample O. (9) Ol,,(lO) XN. x 2200.
11, 12. Pontosphaera multipora (Kamptner) Roth, sample 9. (11) OL, (12) XN.x 2200.
13, 14. Transversopon/is obliquipons (Deflandre) Hay, Mohlcr and Wade,sample 3,5. (13) OL, (14) XN. x 2200.
15. 16. Ericsonia fOTmosa (Kamplner) Romein, sample 6,37. (15) OL, (16) XN.x 2200.
17, 18_ Diclyococciles abiseclus (Muller) Bu"-ry and Pcrcival, sample 10,6.(17) Ol, (18) XN. x 2200.
19. 20. ZygThablilhus bijugalus (Deflandre) Deflandre. sample 18. (19) Ol,(20) XN. x 2200.
~1. '22. Coronocyclus nilescens (Kamptner) Bramlette and Wilcoxon, sampIe 2,6. (21) OL. (22) XN. x 2200.
23. Corannulus germanicus Slradner, sample 6,5, Ol. x 2200.24_ Discoaster sublodoensis BramIette and Sullivan, sample 5, OL. x 2200.
25, 26. Chiasmolilhus oamaruePfsis (Deflandre) Hay, MohJer and Wade, sampie 5,6. (25) OL. (26) XN. x 2200.
27. Discoasler baTbadiensis Tan Sin Hok, sample 3.5. OL. x 2800.
510
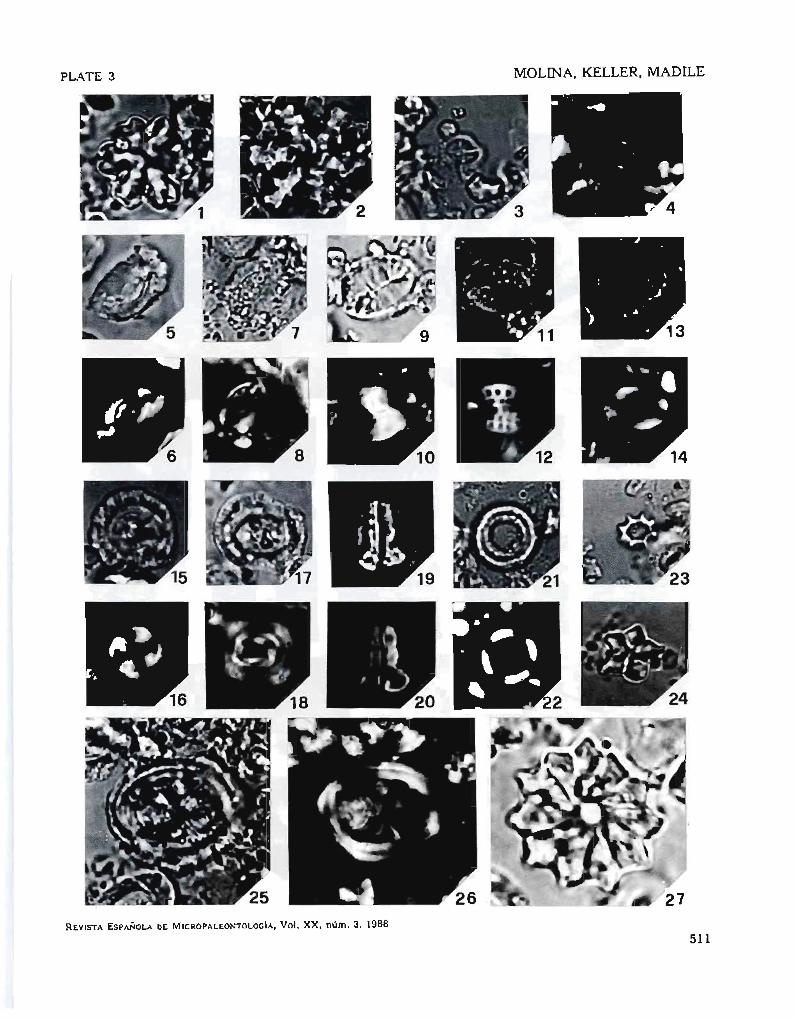
PLATE 3 MüUNA. KELLER, MADlLE
, ., .~ .-I
..•.¡
•••.,~ .'.'
•• •, .... .••
REVISTA ESPAÑOl~ DE M1CROPAlEOmOlOO1A. Vol. XX. mlm. 3, 1988
511

MOLlNA, KELLER, MADlLE PLATE 4
REVISTA EspAÑOLA DE MlCao'AL[ONTUlOGlA, Vol. XX, nOmo J, 1988
512

chronology and tlle ape 01 l/le Eoeene-Oligo.eene bOlmdary. 7th Caribean Gcol. Conf. Proc.1974, Guadeloupc, pp. 205-212.
MCGQWRAN B.
1973 Rifling and drifl af Australia and Ihe migralían 01 mamma/s. Sciencc, vol. 180, pp. 759.
MIu..ER K. G., THOMAS E.
1985 Lale Eoeene lO O/igocene bcnlhíc /uramíni/eralisotope record, Sile 574, equaloria/ Paci/ic. Inilials Rcports oC Ihe DSDP, vol. 85; pp. 771-7&0.
MIl.l.ER K. G., MOUNTAlN G. s., TUCHOLKE B. E.
1985 Oligoeene glacioellsla.sy and erosion on Ihemargín of Ihe Norrh AI/maie. Geology, vol. 13,pp. 1-13.
MOUNA E.
1979 O/igoceno-Mioeeno inferior por medio de foramini/eros plunclónicos en el seclor cenlral delas Cordilleras Belieas (Espalia). Tesis Doct.Public. Univ. Granada y Zaragoza, 342 pp.
1986 Descriplion alld bioslratigraphy 01 lhe maillreference seclioll o/ l/le Eocene Oligocene botlndary hl Spai,l.· Flleme Caldera seClioll. Develop.in Paleont. and Slralig., No. 9, pp. 49-52.
MOLlNA E. el al.1986 Biostraligrap/¡ic corre/alioli belll'eell LIle cell
Iru/ Sllbbelic (Spaill) mld Umbro-Marc!leUfl(1Ialy) pelagic seqw:l1t:es al l!le E/O bOllndaryIlSillg farwlli/lilera. Develop. in Paleont. andStratig., No. 9, pp. 75-86.
MüLINA, KELLER, MADILE
MONECHI s.1986 Calcareous nannofossils of lhe Fuente Caldera
seclion. Develop. in Paleont. and Stratig., vol. 9,pp. 65-70.
In press Cu/careollS nannofossil cvellls around IheEocene Oligocl;lle bOlmdar)! in lhe Umbrian secliolls (Ita/y). Submitted to Palco. Palco. Paleo.
NOCCHI M., PARISI e., MONACO P., MONECHI S.,
Mo\tHLE M., NAPOLEONE G., RIPEPE M., ORLANDO M.,
PREMDLI-SILVA l., lIlCE D. M.
1986 T/lI.' Eocene Oligocene boundar)! in the Umbriall pelagic seqtlenccs, Ila/y. Devclop. in Paleont. and Stratigr. vol. 9, pp. 25-40.
NOCCHI M., PARlSt G., MONACO P., MONECHI S.,
MADILE M.
In press Middle Eocene lO Ear/)! O/lgocenc faraminiferal and calcareous namlOlossils biostratigrapJlY and paleoenvironmental c/lallges in IheSoul/¡ Eas/crll seqlleflces, Ilaly. Submitted toPalco. Paleo. Paleo.
OBERHANSLI 1-1., TOUMARKINE M.
1985 T}¡e Puleogene oxygen and carbon is%pe hislury of Sites 522, 523 and 524 Irom /he cefl/ralSOllt!J Atlalllic. In: Hsu K. J., Weissert H. J.eds. South Atlantic Paleoceanography. Cambrigc Universily Prcss, pp. 125-148.
o KADA H., BUKRY D.
1980 SlIpp/emc'l/ary modificalion al/d inlrodllction(J/ code IIl1mbers ro l!Je lolV lariLUdes cOCCOlitll
1, 2.
3, 4.S, 7.
8, 9.lO, 11.
12.13.
14, 15.
16.I7, 18.
19.
PlATE 4
Reticlllofel1estra lIlIlbilica (Levin) Martini and Ritzkowsky, samplc 3,5.(1) Ol, (2) XN. x 2800.EricsOIlia sp., sample 6,4. (3) Ol, (4) XN. x 2800.Spllel101illlUS disretl!us (Martini) Bramlette and X ilcoxon, sample 16.(5) Ol, (6) XN, (7) XN 45". x 2800.Eriesollia ObTUra Perch Nielsen, sample 5,4. (8) Ol, (9) XN. x 2200.SpllcllolitJltIs predislcnlllS Bramlcllc and Wilcoxon, sample 5,6. (ID) al(11) XN. x 2200.Hulodiscolillws solidlls (Deflandre) Roth, sample 3,5, aL. x 2200.Diseoas/er saipallensis Bramlette and Riedcl, and Braarudospllaerabigelowii (Gran and 8raarud) Deflandrc, sample 3,5, OL. x 2800.CyclococcolitJJilla killgi (Roth) Roth, sample 1,5. (14) al, (15) XN.x 2200.Isllil/loli/llUS recllrvus Deflandre, sample 5,6, Ol. x 2200.Cribrocen/rwll reliculalwl1 (Gartner and Smith) Perch Niclsen, sampIe 1,5. (17) Ol, (18) XN. x 1200.Pedillocyclus larvalis (Bukry and Bram1cttc) Loeblich and Tappan,sample 5,6, aL. x 2800.
513

MOLlNA, KELLER, MAOILE
bioslrotigro.ph¡c W"lJ.lioll, Mar. Micropalcont.,vol. 3, pp. 32J-325.
PERe 1I NIElSEN K.
1986 CalcareollS NOlltlofossil elle'1ll al Ihe Eocelle/Ofigocerre bOll1ldafY. Develop. in Palcont. andStraLig.. vol. 9, pp. 215-282.
¡'RUIOU SILVA l., MOS:ECIlI s., MAUlLE: M., SA.POLEONE u.,ORLANDO M., RIPEI'E M.
In press CalcareollS plallkto" bioslratigrap/,)' alldmagllt!lDslraligraplly lIf fhe Eoccnc 01igoccIleIrtll/siliOI! fmm ti/e G/lbbio area Early Paleocene fJ¡rougJi Earl)' Oligocen/!o Submiltcd lOPalco. Palco. Pako.
Il.OI:IASZ\'NSKI 1'., CARUN M., GONZALEZ-DONOSO J. M.,
WONDEllS .\. H.
1984 Atlas 01 /(lIe Creluceolls globolnmCtlllids. Revue de Micropalcontologic, vol. 26, pp. 145-305.
SANFIUPPO A., RIEOEl. W. R., GUSS B. P., KYTI': F. T.
1985 LA/e Eoccue microtekliles 0",1 radia/ariall ex·tillClioll U/I Barbados. Nalure. vol. 314, pa·ges 613-615.
SAUSOERS J. R.• IIERNOULLl O. ,MUU,.EK-MERZ E.,
OIlERHA/'IlSU 1-1., PERII NII,ISEN K., RIEDH.. W. R.,
SANFII.IPI'O A., TORRINI R. Jr.1984 StrOligroplly 01 tl,e lAte Middle Eocene lO
Eorly O/igocene in tllC Batll Cliff Section, Bar·bodas. Ulest IlIdies. Micropaleontology, vol. lO,pp. 390-425.
514
SHACKl..lrrON N. J., KENNETT J. P •
1976 Paleotemperalllre I-listory 01 lile Cenoz.oic andtlle ;"itiaríon of Antarctic g/aciation: oxigenalld carbOIl istope allalysis in DSDP Sites 277,n9 al/d 181.. IniEia! Rcports of the D5DP, vol. 29,pp. 743-755.
STE.lNECK P. L., BREF_" M•• NEVlNS N., O'I-IAR-' r.
1984 Middle Eocenc mld Dligocelle dl!ep sea ostra·coda Iro", lile oceallic formation Barbados.J. Palconlology, vol. 58, No. 6, pp. 1463.. 1496.
VAII.. P. R, II,\ROENBOL J.1979 Sea level ellallges dllring tlJe Terliary. Occanus,
vol. 22, No. 3, pp. 7J.80.
WEISSEL J. K., Il",VES D. 1:"
1972 Magl1elic ulIol11111ies ¡" flre sOIlI}¡ea..s1 IndianDccan. In Hayes O. E. ed. Antarctic Oceanology JI. Anlarclic Res. Ser., vol. 19, p. 165.
WILLIAMS O. F., TIIUNELL R. e .• 1I01)EI.l. 11. A.,
VERGN... UlI-GItAZZINI c.1985 SYlltcsis of IAle CretaceollS, Tertiary alld Qua
ternary sluble isolope records of Ole SOIlIfI
Atlamic basea 011 Leg 7Z DSDP core Material.m Hsu K. J., Weissen H. J. cds. Soulh Allantic Palcoccanogrnphy. Cambridge UniveTSilYPress, pp. 205-242.
MallflScrito recibido el S-VI-1987