kinetics of larval gregarius behavior in the bark beetle
TRANSCRIPT

Journal iif Insecl Bchavior, Vol. 3, No. 2, 1990
Kinetics of Larval Gregarius Behavior in the Bark Beetle Dendroctonus micans (Coleoptera: Scolytidae)
J . -L . Deneubourg, ' J . -C . Grégoire , ' and E. Le For t '
Acceptée! 8 Mardi 1989: reviscd 24 April 1989
This paper provides a simple mathematical model of the aggregation behavior ofthe gregarious intracortical-feediitg larvae of the bark beetle, Dendroctonus micans. The model's assumptions are that each individiuil produces phero-mones at a constant rate and reacts positively to a pheromone gradient. Thèse hypothèses have been tested by comparing experiinents and simulations, bot h ofwhich showed that (I) homogeneously distributed individuals aggregate rap-idly at the center of the expérimental or theoretical arena and (2) excenrric. prefonned groups succeed in attracting dispersed individuals, provided that the initial size of thèse groups is sufficiently high. TItere is good agreement between most ofthe expérimental and theoretical results, providing a link among chein-ical communication, density of larvae, and random events occurring during the development of a spatial structure.
KEY WORDS: aggregation pheromones; aggregalive behavior, collective fccding; Dciiilni< -lonus micans; mathematical model.
INTRODUCTION
Dendroctonus micans (Kug.) is a primary pest of spruce, Picea spp., in ihe Palearctic Région (Grégoire, 1988a). It attacks green trees, which usually romain alive during the pest's entire iife cycle. Although most other bark beeties col-onizing living trees weaken or kill their host by fungus inoculation, there is no évidence of pathogenic fungi associated with D. micans (F. Lieutier, personal communication). The eggs of D. micans are laid in batches in intracortical chambers, and immediately upon hatching, the young larvae aggregate and feed
'Uni t of Behavioural Ecology, C.P . 231, Université libre de Bnixelles, 50 av. F. D. RiM).scvclt. B-IOSO Bivxelle.s, Belgium.
U9

170 Deneubourg, Grégoire, and Le Fort
side by side upon the living host phloem (Fig. 1). The feeding groups can split around an obstacle and merge afterward or fuse with other groups if two adjacent brood Systems meet. Even though pathogenic associated fungi are absent, induced host reaction occurs in the tissues surrounding the larval chambers (F. Lieutier, personal communication), and communal feeding could constitute a way for the larvae to outrun the tree's reaction (Grégoire, 1988a).
The effects of dumping have been inVestigated in many insect species. Group feeding often improves individual survival, growth, and development (Ghent, 1960; Nakamura, 1980; Tsubaki, 1981), allows better exploitation of food resources (Tsubaki and Shiotsu, 1982), and provides better protection against enemies, by reinforcing the effects of chemical défense and aposematic waming signais (Pasteels et al., 1983).
In Dendroctonus micans, group feeding is a highly coordinated behavior, mediated by the production of and response to aggregation pheromones, trans-and ds-verbenol, verbenone, and myrtenol. Thèse compounds apparently resuit from the oxydation of the host-tree monoterpene, a-pinene. They are emitted when the insect eats host tissues or simply after it has been exposed several hours to a-pinene vapors (Grégoire et al., 1982). Several other Dendroctonus species display such a capability in the adult stage (see, e .g. , Hughes, 1973) and during larval life (Hughes, 1975). D. micans is remarkably tolérant to a-pinene, a compound toxic to many insects (Everaerts et al., 1988); this immunity is perhaps partly achieved by oxydizing the a-pinene into less dan-gerous derivatives such as verbenols, verbenone, and myrtenol.
While it seems clear that volatile chemicals are the agents responsible for larval aggregation in D. micans, the mechanisms of this behavior remain hypo-thetical. How do larvae feeding in a favorable place attract the other individu-als? What makes feeding front lines move according to the local heterogeneities of the bark, dissolving here and forming again there?
A simple model, backed by expérimental results, is presented here. It sug-
Fig. 1. Larval group-feeding in Dendroctonus micans.
Kinetics of Larval Gregarious Behavior In D. micans 171
gests that very simple mechanisms are sufficient for eliciting the paltcms of aggregation in the larvae of D. micans.
T H E T H E O R E T I C A L APPROACH
The Model
Previous expérimental work (Grégoire et al., 1982) suggcsted that clunip formation in the larvae of D. micans results from the compétition bctween two opposing forces: the random movement of the larvae and their positive reaction to a pheromone.
The model's assumptions are as follows. (1) Each larva emits pheromones continuousiy and at a constant rate, sim-
ilar for each individual. (2) The pheromones diffuse in the brood chamber, and a gradient is fomied. (3) The larvae move along this gradient to areas of high concentration. (4) Thigmotaxis has a negligible effect on group formation.
The model is developed within a two-dimensional universe, similar to the natural one, but with several simplifications: it is homogeneous, without obstacles, and open to free pheromone diffusion at its edges.
x{r, t) and c ( r , t) are, respectively, the larval density and the pheromone concentration at point r and time t. The temporal development o{ .x{r, t) is given by the équation
dx/dt = D{dh/dr^) + G[d {x{dc/dr)}/dr] ( l a )
(I) (II)
I represents the random walk of the larvae, and II, the positive reaction of the animais to the gradient (c / r ) . D is proportional to the spced of the random walk and has been estimated from expérimental observations where the time necessary for a larva to leave a 1-cm^ square was measured (D = 1 to 2 c n r / min). G is proportional to the attractiveness of the gradient. Its numerical value ( = 2 or 3 cm'*/min) has been obtained by comparisons between model and experiments. Equation ( la) does not take into account any threshold value in the gradient perception. This simplification induces a higher sensitivity of the équations. In our open expérimental arena, the steep gradients were probably higher than the threshold.
The pheromone concentration is govemed by the équation
de / dt = Ax + D, (d^c / dr') ( I b )
A is the amount of pheromone emitted per animal and per time unit. The second term is the diffusion of the pheromone (D,. is its diffusion rate). Its numerical value has been approximated by comparison with organic cyclic compounds
172 IK'iiciiltmn);, (IH');!!!!c, uiul l.c i iiil
such as toluène and benzène (Wilson et al., 1969) and has becn maintained constant in ail the simulations. By dividing Eqs. ( la) and (Ib) by the coefficient A, c is measured in terms of the individual émission rate. With this change of unit, the value for A does not appear in the model and its estimation is not required anymore.
The boundary conditions for jc are nul flux. For c, we assume a zéro concentration outside the arena. Thèse conditions correspond to those in our expérimental setup (see Materials and Methods underThe Expérimental Approach).
This System behaves in an autocatalytic manner, since as the larvae gather in a cluster (we use "cluster" to name a group where physical contact is per-manently maintained between the individuals), they contribute to enhance the attractiveness of the corresponding région. The higher the larval density level in this région, the stronger the gradient, and the greater the tendcncy to move loward the crowdcd area. The final stationary distribution of the larvae results from the interplay between this positive feedback and the individual random walk.
Simulations of différent situations have been made on a 22 x 22-cm area. The finite différence method was used; the area was divided into 121 squares.
The model tested the efîects of différent larval densities and of several initial conditions: homogeneous larval distribution with or without a preconstituted nucleus.
Results of the Model
Initially Homogeneous Distnbution
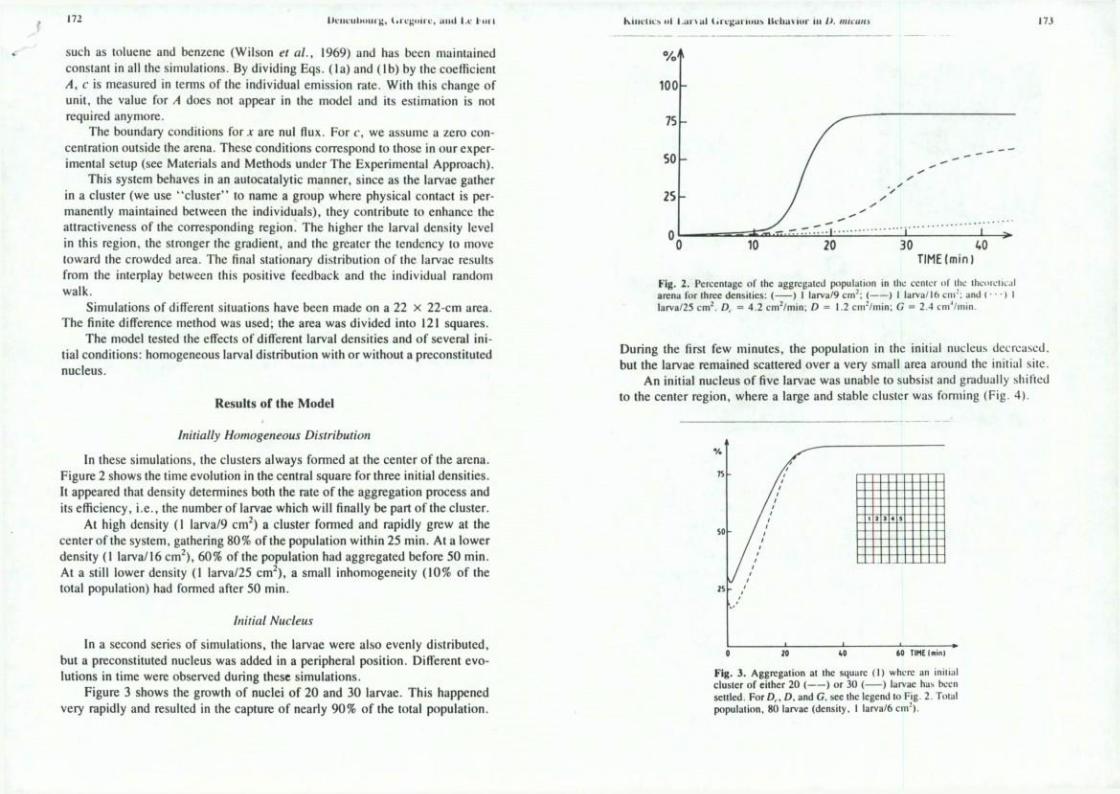
In thèse simulations, the clusters aiways formed at the center of the arena. Figure 2 shows the time évolution in the central square for three initial densities. It appeared that density détermines both the rate of the aggregation process and its efficiency, i.e., the number of larvae which will finally be part of the cluster.
At high density (I larva/9 cm^) a cluster formed and rapidly grew at the center of the system, gathering 80% of the population within 25 min. At a lower density (I larva/16 cm^), 60% of the population had aggregated before 50 min. At a still lower density (1 larva/25 cm^), a small inhomogeneity (10% of the total population) had formed after 50 min.
Initial Nucleus
In a second séries of simulations, the larvae were aiso evenly di.stributed, but a preconstituted nucleus was added in a peripheral position. Différent évolutions in time were observed during thèse simulations.
Figure 3 shows the growth of nuclei of 20 and 30 larvae. This happcned very rapidly and resulted in the capture of nearly 90% of the total population.
Kiiiclic.s 1)1 l .urvul drcguii<iu.'> I t i l iuvior iii O. imcain
100
I7 J
so
251-
10 20 30 W TIME(mln)
Fig. 2. Perccntagc of the aggrcgaled population in llic ccnicr ol ihc thcoichc.il arena for Ihrce densities: ( ) I larva/9 c m ' ; ( ) I larva/16 ci i r ; and ( • • • ) I larva/25 cm^. D, = 4 .2 cm^/min; £) = 1.2 cm ' /min ; G = 2.4 cm' /ni in .
During the first few minutes, the population in the initial nucleus dccrcascd, but the larvae remained scattered over a very small area around the initial site.
An initial nucleus of five larvae was unable to subsist and gradually shitied to the center région, where a large and stable cluster was fomiing (Fig. 4).
(0 M riHEIminl
Fig. 3. Aggregation at the square (1) where an initial cluster of eithcr 20 ( ) or 30 ( ) larvae has been setlled. For D, , D, and C, see the legend to Fig. 2. Total population, 80 larvae (density, I larva/6 cm").

174
20 U 60 M TIM£(mnl
Fig. 4. Time évolution of the number of larvae in squaix:s 3 and 4, wiih an initial clusierof five larvae iniroduced in square 1. For D, , D, and G, see the legend to Fig. 2. Total population, 80 larvae (density, I larva/6 cm^).
For values of the initial nucleus intermediate between thèse extrême values (20-5 larvae), the final cluster formed midway between the position of the initial nucleus and the center of the arena, being doser to the center of the setup for smaller values of the initial nucleus.
In the latter two cases, as in the first one, about 90% of the larvae end up in the final cluster.
THE EXPERIMENTAL A P P R O A C H
Materials and Methods
The observations and experiments were made with fourth- to fifth-instar larvae of D. micans, excised from logs kept in the laboratory.
The expérimental arena consisted of two square glass plates (30 X 30 cm), laid one above the other, separated by a 3-mm space allowing free movement of the insects between them. The "floor-plate" was covered with a circular pièce of filter paper, 24 cm in diameter. This expérimentai arena was similar to the theoretical one described previously, except that it was circular instead of square. The larvae were released on the filter paper, either homogeneously or with an excentric, preconstituted nucleus. The "cei l ing" glass plate was then replaced, and photographs were taken at regular intervais, every 2 or 5 min. No larva ever ieft the moist paper; this is équivalent to the boundary conditions in the model.
75 -
Kinetics of Larval Gregarious Behavior in D. micans m
I Results
Initially Homogeneous Distribution
In the first group of experiments, larvae were evcnly distributcd in the aicna, their density varying from one experiment to another (from I larva/40 cm ' to 1 larva/3 cm^).
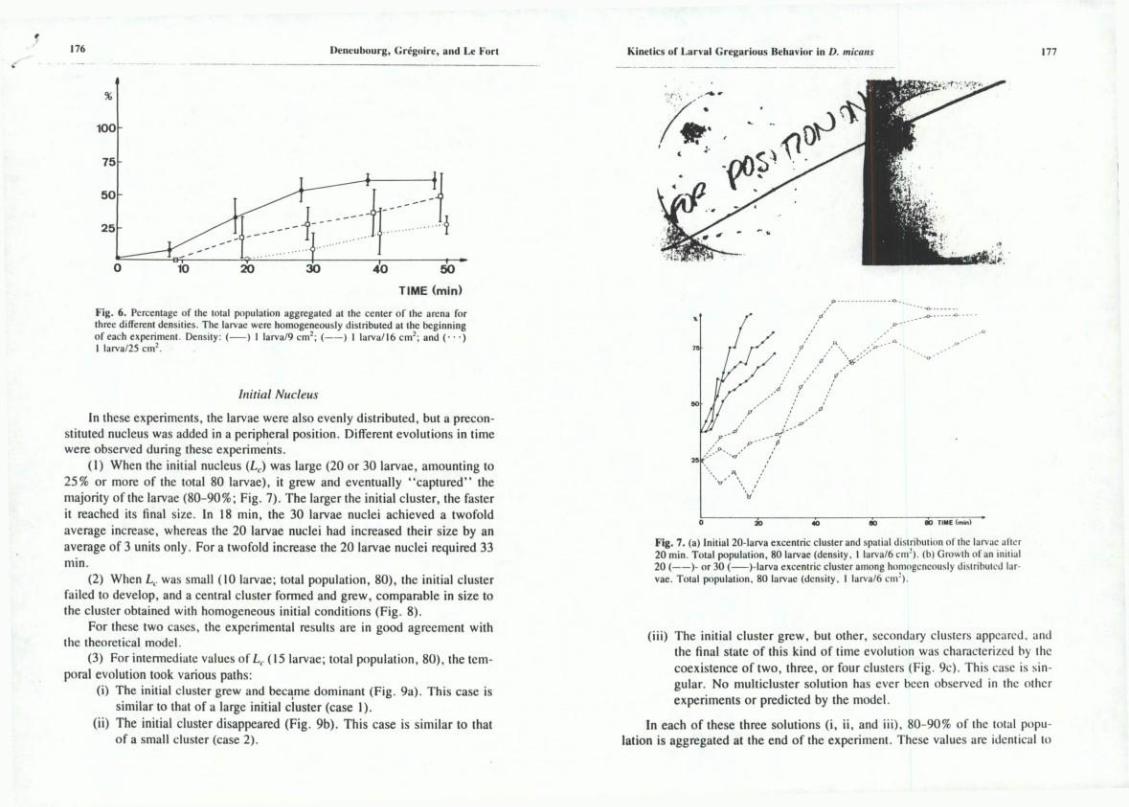
Throughout this whole set of experimenis, one single cluster formed ut the center of the arena, or very close to it, as was observed in the theoretical setup. Figure 5 shows this phenomenon at very high density (I larva/6 cnr) . Within 5 min, more or less 50% of the larvae had aiready clustercd at the center of the arena. Ninety percent of the larvae had clustered within 20 min. For lowcr densities, the larvae clustered more slowly and the clusters were proportionaily smaller (Fig. 6). At the low density of I larva/25 cm^, a loose cluster appcared very slowly, gathering only 25% of the population (Fig. 6). Thèse différent results are in agreement with the model's prédictions (compare Figs. 2 and 6).
Fig. 5. Cluster formation at high density (I larva/6 cm"). Total population, 80 lariae. ; = 0, 5, 10, and 20 min.
Denvubourg, Grégitire, and Le Forl
%
1 0 0 -
7S-
TIME (min)
Fig. 6. Percenlage of Ihe total population aggregated at the center of the arena for three différent densilies. The larvae were homogeneousiy distributed at the beginnlng of each experiment. Density: ( ) I larva/9 cm^; ( ) 1 larva/16 cm"; and (• • •) 1 larva/25 cm".
Initial Nucleus
In thèse experiments, the larvae were also evenly distributed, but a precon-stituted nucleus was added in a peripheral position. Différent évolutions in time were observed during thèse experiments.
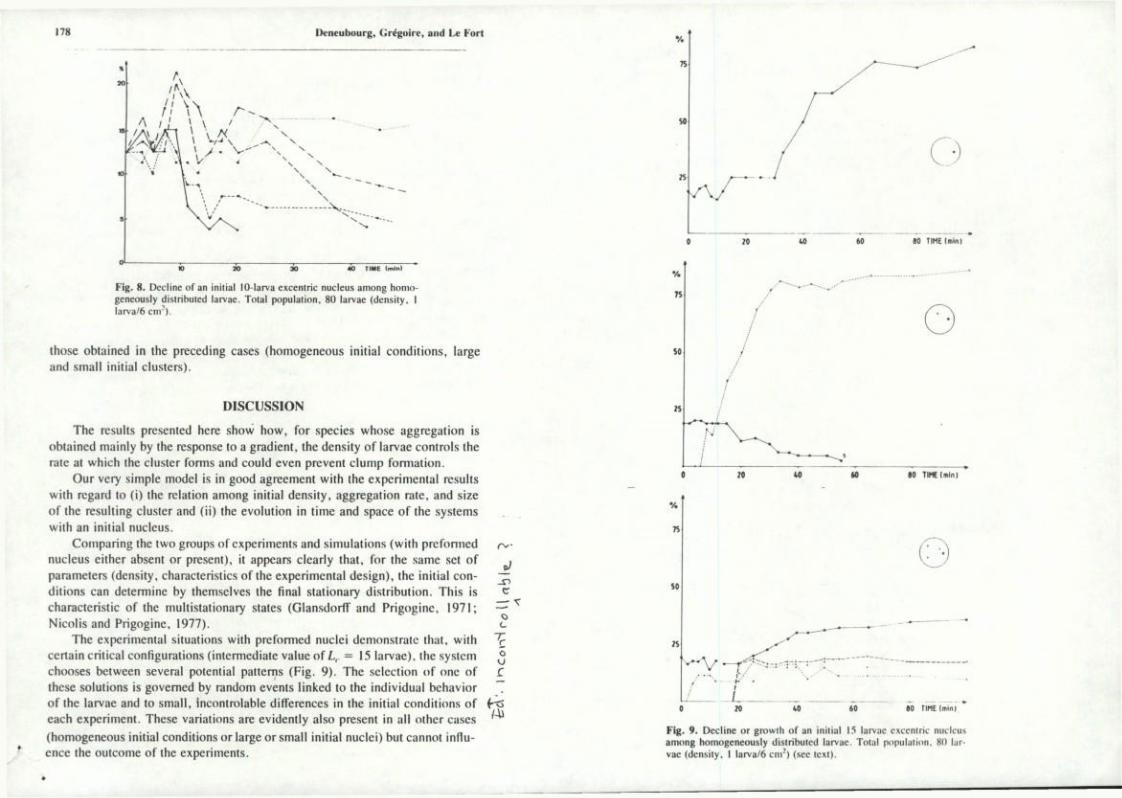
(1) When the initial nucleus (L^) was large (20 or 30 larvae, amounting to 25% or more of the total 80 larvae), it grew and eventually "captured" the majority of the larvae (80-90%; Fig. 7). The larger the initial cluster, the faster it reached its final size. In 18 min, the 30 larvae nuclei achieved a twofold average increase, whereas the 20 larvae nuclei had increased their size by an average of 3 units only. For a twofold increase the 20 larvae nuclei required 33 min.
(2) When L. was small (10 larvae; total population, 80), the initial cluster failed to develop, and a central cluster formed and grew, comparable in size to the cluster obtained with homogeneous initial conditions (Fig. 8).
For thèse two cases, the expérimental results are in good agreement with the theoretical model.
(3) For intermediate values of L, (15 larvae; total population, 80), the temporal évolution took various paths:
(i) The initial cluster grew and became dominant (Fig. 9a). This case is similar to that of a large initial cluster (case 1).
(ii) The initial cluster disappeared (Fig. 9b). This case is similar to that of a small cluster (case 2).
Kinetics of Larval Gregarious Behavior in D. micans 177
0 10 40 a o 80 TIME (mini
Fig. 7. (a) Initial 20-larva excentric cluster and spatial distribution of the larvae aller 20 min. Total population, 80 larvae (density, 1 larva/6 cnr ) . (b) Growlh of an initial 20 ( )- or 30 ( )-larva excentric cluster among homogcncousiy distributed larvae. Total population, 80 larvae (density, 1 larva/6 cm").
(iii) The initial cluster grew, but other, secondary clusters appearcd, und the final state of this kind of time évolution was characterized by the coexistence of two, three, or four clusters (Fig. 9c). This case is sin-gular. No multicluster solution has ever bcen observed in the other experiments or predicted by the model.
In each of thèse three solutions (i, ii, and iii), 80-90% of the total population is aggregated at the end of the experiment. Thèse values are idcntical lo
10 90 30 40 TIME (min)
Fig. 8. Décline of an initial lO-larva excentric nucleus among homo-geneously disiribuled larvae. Total population, 80 larvae (densily, I larva/6 ciiv).
those obtained in the preceding cases (homogeneous initiai conditions, large and small initial clusters).
DISCUSSION
The results presented here show how, for species whose aggregation is obtained mainly by the response to a gradient, the density of larvae controls the rate at which the cluster forms and could even prevent clump formation.
Our very simple model is in good agreement with the expérimental results with regard to (i) the relation among initial density, aggregation rate, and size of the resulting cluster and (ii) the évolution in time and space of the Systems with an initial nucleus.
Comparing the two groups of experiments and simulations (with prefoniied nucleus eilher absent or présent), it appears clearly that, for the same set of parameters (density, characteristics of the expérimental design), the initial conditions can détermine by themselves the final stationary distribution. This is c: characteristic of the multistationary states (Glansdorff and Prigogine, 1971; — Nicolis and Prigogine, 1977). c
The expérimental situations with preformed nuclei demonstratc that, with ~Ç certain critical configurations (intermediate value of L, = 15 larvae), the systcm ^ chooses between several potential patterns (Fig. 9). The sélection of one of r thèse solutions is govemed by random events linked to the individual behavior T of the larvae and to small, incontrôlable différences in the initial conditions of fxs each experiment. Thèse variations are evidently also présent in ail other cases '"^ (homogeneous initial conditions or large or small initial nuclei) but cannot influence the outcome of the experiments.
Fig. 9. Décline or growth of an initial 15 larvae excentric nucleus among homogeneousiy distribuled larvae. Total population. 80 larvae (density, I larva/6 c n r ) (,see Icxl).

180 Deneiibourg, Grégoire, and Le Fort
Another mechanism could possibly be considered responsible for group formation in D. micans larvae: random walk followed by thigmotactic arrestment. This hypothesis relies upon the observation that, once formed, the groups do not move anymore. This potential mechanism is quitc unlikely. A model of aggregation in mollusks developed by Focardi et al. (1985) has been adapted to the présent situation. The results were that random walk and thigmotactic arrestment would lead to the formation of large numbers of very small groups (mostly two to four individuals).
Thigmotaxis may, however, have a function complemcntary to that of chi-miotaxis. It may explain the discrepancy between our model and our expérimental results with initial nuclei of intermediate size (15 larvae). In the range of the parameter values used for the simulation, the model does not exhibit stable multicluster solutions as found during the experiments (Fig. 9c). The muhicluster solutions are only transient states. Thigmotaxis, which has not been taken into account in our model, may have a stabilizing function within the small clusters. However, the good gênerai agreement between our model and expérimental data confirm, in most of the expérimental conditions, the validity of our hypothèses and the secondary rôle of thigmotaxis.
Very little is known about the kinetics of cluster formation and models have not been developed in this field, except in some isolated cases (see Okubo, 1980). In a séries of interesting experiments. Farine and Lobreau (1984) studied the kinetics of gregarious behavior in the Heteropteran Dysdercus cingulatus. They demonstrated a strong relation, between the population density and the level of aggregation.
Another notable exception concems the classical model of Keller and Segel (1970) about social amebae. In this case the aggregation mechanisms are char-acterized by the présence of an enzyme destroying the aggregation signal, thus preventing its concentration from reaching levels at which orientation is impossible (Dannon et al., 1978).
Our model is clo.se to that of Keller and Segel, except that, in our System, the signal's decay through a chemical reaction is not necessary. It has been observed (Grégoire and Meriin, 1984) that the larvae of D. micans regularly bore ventilation holes in the proximity of their feeding front line. This would allow the pheromone to escape, thus maintaining its concentration at low level. A similar model has been used to describe the successive steps in the building behavior of termites (Deneubourg, 1977).
Other conditions prevail for leaf-eating insect larvae, e.g. , .sawflies (Ghcnt, 1960; Nakamura, 1980), Lepidoptera (Tsubaki, 1981), and leaf beetles (Grégoire, 1988b). Air turbulence around the leaf would limit any chemical communication based on gradient perception. Although little information is available from the literature, other group cohésion mechanisms have been described such as thigmo'gtaxis (Demolin, 1962) and the use of sonic signais (Meyer-Rochow, 1972).
Kinetics of Larval Gregarious Behavior in D. micans 181
Aggregation is very fréquent in arthropods (Wilson, 1971) and in other invertebrate groups, such as mollusks (Focardi et al., 1984, 1985). In spitc ol the great différences among the communication vectors involved, the functions achieved, and the environmental conditions, the varions instances of aggregation have many common features, particulariy regarding the rôle of animal density, fluctuations, perturbations, and multiplicity of solutions.
Because of the complexity of thèse Systems and the scalc dillercnce between individual and collective levels, mathematical models arc ncedcd as a link.
ACKNOWLEDGMENTS
We would like to thank Professor J. M. Pasteels and Dr. S. Goss Ibr niany helpful discussions and Professors A. A. Berryman, T. L. Payne, and M. Sabelis and Dr. M. C. De Jong for their comments on an early draft. We are indebted to Professors I. Prigogine and G. Nicolis for their interest.
REFERENCES
Darmon, M. , Barra, J . , and Brachet, P. (1978). The rôle of phosphodicsicrase in iiggrLjjalion ol' Dictyostelium discoideum. J. Cell Sci. 31: 233.
Demolin, G. (1962). ComportemenI des chenilles de Thaumeiopocti piiyoccimpci Schlll au cours des processions de nymphose. C.R. Acad. Sci. 254(4): 733-734.
Deneubourg, J .-L. (1977). Application de l 'ordre par fluctuations à la description de Lcnaincs étapes de la construction du nid chez les termites. Soc. Inscci.s 24: 117-130.
Everaerts, C Grégoire, J . - C , and Merlin, J. (1988). The toxicity of spnicc nionolcriicncs against bark beetles and their associâtes. In Mattson, W. J. , Lévicux, J . . and Bemard Daiian, C. (eds.), Mechanisms of Woody Plant Défenses Aj>ainsl Insecis and Palhoacns. Springcr Vcriag. New York, pp. 331-340.
Farine, J . -P . , and l^obreau, J .-P. (1984). Le grégarisme chez Dysdercus ciiii^uhitus l ahr (llclcr-optera, Pyrrhocoridac): Une nouvelle méthode d'interprétation statistique. Sac ln\cci\ 31: 277-290.
Focardi, S., Deneubourg, J . -L . , and Chclazzi, G. (1985a). The cxtcmal nicniory of inlcrtidal molluscs: A thcoretical study of trail-following. In Caspa.s.so. V. . Gosso, E.. and Pavcri-F-'on-tana, L. (eds.), Malbemalics in Biologv and Medicine, Springcr Vcriag, Hcidclhcrg. pp. .'>4-59.
Focardi, S., Deneubourg, J . -L . , and Chclazzi, G. (1985b). How shorc niorphnlogy aiul oiioiilalion mechanisms can afléct the spatial organization of intcrtidal molluscs. J. Tlieai: Binl. 112: 771-782.
Ghent, A. W. ( i960) . A study of the group feeding behavior of the lar\'ae of the jack pine sawtly. Nendiprion pratli banksianac Rho. Behavioiir 16( 1-2): 110-148.
Glansdorir, P. , and Prigogine, \. (1971). Tltennodynuniics of Siniciiire. Stahiliiy ami l lm niaiions. Wiley-lnierscicncc, New York.
Grégoire. J . -C. (1988a). The greater European sprucc bcctie. In Berryman. A. A. (cd I. l'apidniiim Dynamics of Foresl Insecis, Plénum, New York, pp. 455-478.
Grégoire, J . -C. (1988b). Larval gregariousness in the Chrysomelidae. In Jolivct, P.. IViilpicrrc. E. , and Hsiao, T . (eds.), Tlie Biology of Chrysomelidae. Junk. DordrechI, pp. 251-258.
Grégoire, J . - C . and Merlin, J. (1984). Dendroctonus micans: The évolution of a brood .sysicm. In Grégoire. J . - C , and Pa.stccis, J. M. (eds.). Proceedinits of ihe EEC Scniinar on ihe Bio-logical Conirol of Bark Beelles (Dendroctonus micans). Brii.<iscls. 3-4/I0/W84. pp. 80-96.
Grégoire, J . - C , Braekman, J . - C , and Tondeur, A. (1982). Chemical communication bclwccn the
I

182 Deneubourg, Grégoire, and Le Fort
larvae of Dcndroclomis muons Kug. Les Colloques de l'INRA. 7. Les Médiateurs chimiques, pp. 253-257.
Hughes, P. R. (1973). Demirocioims: Production of pheromoncs and relatcd coinpounds In rcsponse to host monoterpenes. Z. angew. Emomnl. 73: 294-312.
Hughes, P. R. (1975). Pheroinones of Dendroclonus: Origin of a-pincne oxydation producis présent in émergent adulls. J. Inseci Physiol. 21: 687-691.
Kcller, E. F., and Segcl, L. A. (1970). Initiation of slinie niold aggrcgalion vicwcd as an insta-billty. J. Vu'or. Biol. 26: 399-415.
Mcycr-Rochow, B. V. (1972). Vcrstiindingungswciscn bci kolonicbildcndcn Blattwcspcn- und Kafcr-Larvcn. Z Tierpsychol. 30(5): 451-455.
Nakamura, H. (1980). Ecological studics of the europcan pinc sawfly, Neodiprion serlifer (Geoffroy) (Hymcnoptera: Diprionidae). 1. The elTect of larval aggrcgation and ils forni. ./<;/». J. Appt. Enlomol. Zool. 24: 137-144.
Nicolis, G. , and Prigoginc, I. (1977). Self-Organisation in Non-Equilihrium Systems. Wiley-lnter-sciencc, New York.
Okubo, A. (1980). Diffusiim and Ecological Prohiems: Mathematical Models. Biomatheniatics Séries, Springer-VeHag, Hcidelberg.
Pasteels, J. M. . Grégoire, J . - C , and Rowcll-Rahier, M. (1983). The chcniical ecology of défense in arthropods. Annu. Rev. Entomol. 28: 263-289.
Tsubaki, Y. (1981). Some bénéficiai efTccts of aggrcgation in young larvae o( Pryeria sinica Moore (Lcpidoptera: Zygaenidae). Res. Pnpul. Ecol. 23: 156-167.
Tsubaki, Y., and Shiotsu, Y. (1982). Group feeding as a strategy for cxploiling food rcsources in the Bumct molh Pryeria sinica. Oecologia (Berl.) 55: 12-20.
Wilson, E. O. (1971). llie Insect Societies. Bclknap Press of Harvard University Press, Cambridge, Mass.
Wilson, E. O. , Bossen, W. H., and Régnier, F. E. (1969). 7. In.seci Physiol. 15: 597-610.