do bark beetle remains in lake sediments correspond to ... · pdf filedo bark beetle remains...
TRANSCRIPT

Do bark beetle remains in lake sediments correspond to severeoutbreaks? A review of published and ongoing research
Jesse L. Morris a,b,*, Colin J. Courtney Mustaphi c,1, Vachel A. Carter d, Jennifer Watt e,Kelly Derr a, Michael F.J. Pisaric f, R. Scott Anderson g, Andrea R. Brunelle d
aDepartment of Forest, Rangeland, and Fire Sciences, College of Natural Resources, 875 Perimeter Drive MS1133, University of Idaho, Moscow, ID 83844, USAbDepartment of Geography, Kansas State University, 118 Seaton Hall, Manhattan, KS 66506, USAcOttawa-Carleton Geoscience Centre, Department of Earth Sciences, Carleton University, 1125 Colonel By Drive, Ottawa, Ontario K1S 5B6, CanadadDepartment of Geography, University of Utah, 260 S. Central Campus Drive, Room 270, Salt Lake City, UT 84112, USAe Environmental and Sustainability Studies, University of Utah, 260 S. Central Campus Drive, Room 152B, Salt Lake City, UT 84112, USAfDepartment of Geography, Brock University, 500 Glenridge Avenue, St. Catharines, Ontario L2S 3A1, Canadag School of Earth Sciences & Environmental Sustainability, Northern Arizona University, Building 19, Room 100 (Box 5694), Flagstaff, AZ 86011, USA
a r t i c l e i n f o
Article history:Available online xxx
Keywords:DendroctonusLake sedimentsMountain pine beetleScolytinaeSpruce beetleTaphonomy
a b s t r a c t
The recent continental-scale outbreak of native bark beetles in western North America is unprecedentedat least since Euro-American settlement. Observational and modeling evidence suggest that warmtemperatures observed during the late 20th century altered beetle population dynamics by acceleratingbeetle reproductive cycles leading to exponential population growth. The linkage between beetle out-breaks and climate warming has motivated efforts to reconstruct these disturbances using long-termenvironmental records using lake sediments. Here, we present data from across western North Amer-ica in an effort to understand how beetle remains retrieved from lake sediments may be used as a proxyfor reconstructing severe outbreaks and ecosystem response over centennial to millennial timescales. We(1) review existing literature related to beetle taphonomy; (2) present previously unpublished data ofbeetle remains in lake sediments; (3) comment on the development of a methodology to retrieveterrestrial beetle remains from lake sediments; (4) discuss potential controls on beetle carcasstaphonomy into the sediment matrix; and lastly (5) speculate on the use of primary and secondary attackbeetle remains as indicators of past outbreak episodes. Our synthesis suggests that the remains of pri-mary attack beetles are rarely preserved in lake sediments, at least using small-diameter piston devicescommon in multi-proxy studies. Alternatively, remains of secondary attach beetles may be common butfurther work is required to understand how these insects can be used to aid in interpreting past forestdisturbances, including bark beetle outbreaks and wildfire. A number of factors may influence whetheror not bark beetle remains become entrained in the area of sediment focusing including lake waterchemistry, fish predation and scavenging, and weather conditions during peak beetle emergence.
! 2014 Elsevier Ltd and INQUA. All rights reserved.
1. Introduction
The recent continental-scale epidemics of native bark beetles inwestern North America has reorganized ecosystem nitrogen pools(Morris et al., 2013a; Rhoades et al., 2013), altered terrestrial carbonfluxes (Pfeifer et al., 2011; Hicke et al., 2012a), changed forest
structure and composition (DeRose and Long, 2012), generallyreduced landscape flammability and rearranged fuel structure(Jenkins et al., 2008; Hicke et al., 2012b; Harvey et al., 2013), whilealso diminishing air andwater quality (Amin et al., 2012; Mikkelsonet al., 2013), devaluing real estate (Hanson and Naughton, 2013),increasing winter and spring surface albedo (Vanderhoof et al.,2013), and reducing the duration of snow cover (Biederman et al.,2014). In recent decades, the cumulative economic and aestheticlosses from bark beetle outbreaks (i.e. Dendroctonus spp.) exceedthose of wildfire and other forest disturbances (Logan et al., 2003).Anthropogenic climate warming is widely acknowledged as theprimary driver promoting the irruptive populations of bark beetles(Raffa et al., 2008). Observational and modelling evidence
* Corresponding author. Department of Forest, Rangeland, and Fire Sciences,College of Natural Resources, 875 Perimeter Drive MS1133, University of Idaho,Moscow, ID 83844, USA.
E-mail addresses: [email protected], [email protected] (J.L. Morris).1 Current address: Environment Department, York Institute for Tropical Ecosys-
tems, University of York, Heslington, York YO10 5DD, United Kingdom.
Contents lists available at ScienceDirect
Quaternary International
journal homepage: www.elsevier .com/locate/quaint
http://dx.doi.org/10.1016/j.quaint.2014.03.0221040-6182/! 2014 Elsevier Ltd and INQUA. All rights reserved.
Quaternary International xxx (2014) 1e15
Please cite this article in press as: Morris, J.L., et al., Do bark beetle remains in lake sediments correspond to severe outbreaks? A review ofpublished and ongoing research, Quaternary International (2014), http://dx.doi.org/10.1016/j.quaint.2014.03.022

demonstrate that increasing temperatures in the lower atmospherealter beetle population dynamics in two primary ways: (1)temperature-induced acceleration of bark beetle fecundity in-creases the likelihood of exponential population growth (Hansenet al., 2001a,b; Cudmore et al., 2010; Mitton and Ferrenberg,2012); and (2) reductions in cold-induced mortality allow greatersurvival of over-wintering bark beetles, which is particularly pro-nounced at high-elevations and high-latitudes locales (Reid, 1963;Reid and Gates, 1970; Bentz et al., 2010).
Affecting 47,000,000 ha during the past two decades, the scaleand severity of the current outbreak, is unprecedented at least sincethe arrival of Euro-American settlers in the 19th century (Taylor andCarroll, 2004; Raffa et al., 2008). During outbreaks, primary attackbeetles (Dendroctonus spp.) first infest the boles of downed and/orstressed trees (Fig.1). As the outbreak progresses, even healthy hosttrees succumb to attack. As host trees die, they become susceptibleto infestation from secondary bark beetles (e.g. Pityophthorus spp.,Ips spp.), which may attack both the bole and branches of dead host
Fig. 1. Conceptual diagram of the progression of a spruce beetle (Dendroctonus rufipennis) disturbance through endemic (A), incipient or attack phase (B), red or epidemic phase (C),and grey or collapse phase (Panel D). Secondary attack beetles generally appear during red (C) and/or grey phase (D). Reproduced from Morris (2013). (For interpretation of thereferences to colour in this figure legend, the reader is referred to the web version of this article.)
J.L. Morris et al. / Quaternary International xxx (2014) 1e152
Please cite this article in press as: Morris, J.L., et al., Do bark beetle remains in lake sediments correspond to severe outbreaks? A review ofpublished and ongoing research, Quaternary International (2014), http://dx.doi.org/10.1016/j.quaint.2014.03.022

trees. In subalpine and mid-montane environments, recent conifermortality has been caused primarily by the mountain pine beetle(Dendroctonus ponderosae), spruce beetle (Dendroctonus rufipennis),and Douglas-fir beetle (Dendroctonus pseudotsugae) (Fig. 2). Ourbaseline of knowledge about the past dynamics of forest damagingbeetles has been established primarily through dendroecologicaland historical records that span the last few centuries (Veblen et al.,1994; Hebertson and Jenkins, 2008; Sherriff et al., 2011; DeRose andLong, 2012; Donato et al., 2013).
The spatial and temporal synchrony of outbreaks during recentcenturies is not as widespread or abrupt in comparison to thepresent outbreaks. For example, spruce beetle is the most impor-tant biotic disturbance agent in Engelmann spruce (Picea engel-mannii) e subalpine fir (Abies lasiocarpa) forests, and is animportant driver of landscape dynamics in this expansive forestecosystem (Lindgren and Lewis, 1997; Jenkins et al., 2014). Out-breaks of spruce beetle occurred during the 1920s on the AquariusPlateau in Utah and during the 1940s on the White River Plateau inColorado. These landforms are separated spatially by approxi-mately 600 km and these outbreaks were temporally discrete bytwo decades (Dixon, 1935; Mielke, 1950; Schmid and Frye, 1977;Hebertson and Jenkins, 2008). Mortality associated with thesetwo outbreaks was reported to be less than 80% in mature Engel-mann spruce. In contrast, recent spruce beetle outbreaks haveoccurred on the Markagunt Plateau and Wasatch Plateau, both insouth-central Utah, as well as those in Rocky Mountain NationalPark and the San Juan Mountains, both in Colorado. These regionsare separated by more than 700 km, but outbreaks were synchro-nous during the 1990s, with spruce mortality observed at >90%(Dymerski et al., 2001; DeRose and Long, 2007; Raffa et al., 2008).
There exist many centennial- to millennial-timescale re-constructions of wildfire disturbance using charcoal fragmentsrecovered from lake sediments (e.g. Carter et al., 2013; CourtneyMustaphi and Pisaric, 2013; Morris et al., 2013b). However, thereare noticeably fewer examples of reconstructions focused on non-fire disturbances, despite their obvious ecological and societalsignificance (Morris, 2013; Waller, 2013). Efforts to reconstructnon-fire disturbances utilize sediment-based proxy assemblages ofsubfossil pollen and preserved plant and biotic remains (Davis,1981; Anderson et al., 1986). The linkage between beetle out-breaks and a warming climate has motivated efforts to reconstructoutbreaks from longer environmental archives, such as lake sedi-ments, in an effort to examine the interactions among disturbancetypes (Morris, 2013). Recent work documenting bark beetleewildfire interactions underscores the need to develop a broadlyaccessible methodology for detecting past bark beetle disturbancesover longer temporal scales (Bigler et al., 2005; Simard et al., 2011;Jolly et al., 2012; Harvey et al., 2013). Accordingly, lake sediment-based reconstructions have sought to provide longer-term ecolog-ical context for bark beetle disturbances (Anderson et al., 2010;Morris et al., 2010; Morris and Brunelle, 2012; Morris et al.,2013a). These studies are focused on developing a methodologyfor detecting bark beetle disturbances in the recent geologic past(i.e. the Holocene). However, currently few paleoenvironmentalreconstructions from lake sediments have definitely detected out-breaks prior to the historical period in western North America(Brunelle et al., 2008).
In general, lake sediment reconstructions of bark beetle out-breaks examine for direct evidence of beetle presence, includingpreserved beetle elytra, thoracic parts, and to a lesser degree, head
Fig. 2. Panels of light micrographs of the head capsules and elytra from lab specimens of primary attack beetles (spruce beetle AeC and mountain pine beetle DeF).
J.L. Morris et al. / Quaternary International xxx (2014) 1e15 3
Please cite this article in press as: Morris, J.L., et al., Do bark beetle remains in lake sediments correspond to severe outbreaks? A review ofpublished and ongoing research, Quaternary International (2014), http://dx.doi.org/10.1016/j.quaint.2014.03.022

capsules (Brunelle et al., 2008; Watt, 2008; Morris et al., 2010).Recently, indirect indicators of past beetle dynamics, includingshifts in the ratio of host to non-host pollen (Anderson et al., 2010;Morris and Brunelle, 2012; Morris et al., 2013a) and alterations tobiogeochemical cycling assessed from stable light isotopes andelemental concentrations (Morris et al., 2013a) have also shownpromise for tracking past beetle disturbances. Brunelle et al. (2008)found mountain pine beetle remains in sediments correspondingtemporally to the early 20th-century and early Holocene. The au-thors proposed that when bark beetle populations reach outbreakstatus, their remains are so ubiquitous that they may be used todetect past outbreak episodes, akin to sedimentary charcoal (e.g.Whitlock and Larsen, 2001). However, recent papers present find-ings frommultiple sites that have been unable to reproduce similarresults despite significant efforts to do so (Anderson et al., 2010;Morris and Brunelle, 2012). Jackson (2012) reiterates the impor-tance of understanding taphonomic processes in paleoecology. Inthat spirit, here we: (1) review existing literature related to barkbeetle taphonomy; (2) present previously unpublished data on barkbeetle remains in lake sediments; (3) propose a methodology toisolate and preserve terrestrial chitinous beetle remains fromsediments; (4) discuss potential controls on bark beetle carcasstaphonomy and deposition to the sediment matrix; and lastly (5)speculate on the use of primary and secondary attack beetle re-mains as robust indicators of past outbreak episodes.
It is important to note that the sediment sampling strategiesused in the studies reviewed here contrast with Coleopteran-focused research found in the paleoenvironmental literature (e.g.Coope, 1959; Ashworth, 1977; Ashworth et al., 1981; Elias, 1985,2001). This difference stems from a dissimilarity of approach un-dertaken to acquire subfossil and fossil material for paleoenvir-onmental interpretation. For example, Coleopteran- orTrichopteran-based paleoenvironmental reconstructions seek toassemble a large (1 L), diverse sample with preserved insect re-mains that does not specifically target any genera or sub-family ofColeoptera (Elias, 2001). By identifying as many taxa as possible,the beetle assemblage is then used to reconstruct paleoclimateconditions and/or provide a landscape classification (e.g. forest vs.tundra) based on the known thermal tolerances and/or habitatpreferences of modern Coleopteran (Elias, 2001). Specifically,presence/absence is emphasized over abundances of any particulartype (Elias, 2001). This approach emphasizes the collection of largevolumes of sediment from deltaic or littoral sites and does notnecessarily require a continuous depositional sequence or high-resolution chronology to the same degree as lake sediment re-cords (see Ashworth, 1977; Elias, 1985). The primary aim of theirsampling strategy is to maximize the recovery of the largestnumber of beetle types. Given the considerable amount of sedi-ment harvested in these sampling approaches, it is not surprisingthat, in some instances the presence of several Dendroctonus spp.remains were noted using this strategy, among hundreds of otherColeopteran taxa (e.g. Ashworth, 1977; Ashworth et al., 1981; Elias,1985).
The sampling protocols of Coleopteran-based paleoclimatic re-constructions (e.g. Elias, 2001) have been largely disregarded inrecent attempts to reconstruct beetle disturbances in favor ofsampling protocols that emphasize high-resolution temporalanalysis (e.g. Morris and Brunelle, 2012). Specifically, recent effortsto reconstruct Dendroctonus outbreaks diverge from the afore-mentioned assemblage-focused studies in several important ways.First, outbreak reconstructions were focused solely on one Cole-opteran genus, namely Dendroctonus spp. The focus on Den-droctonus spp. is motivated by the need to test whether the resultsof Brunelle et al. (2008) are repeatable using similar field andlaboratory procedures. Second, the primary aim of outbreak
reconstructions is to develop a methodology that can be readilyincorporated into multi-proxy approaches, that also analyze formacroscopic charcoal, preserved pollen, stable light isotopes (d15N,d13C), among others. Most multi-proxy sampling approaches utilizeLivingstone devices and other coring apparatuses of similar di-ameters (w5 cm), which contrasts with coleopteran-focused sam-pling procedures (Elias, 2001). We acknowledge that the ‘bestpractice’ for retrieving sufficient quantities of Coleopteran remainsto support paleoclimate reconstructions does not necessarily reflectthe needs of multi-proxy paleoenvironmental reconstructions thatmotivated the work we present here. Specifically, we used a samplesize of 1e5 cc because this volume reflects standard paleolimno-logical methodology that enables the development of high-resolution chronologies. A third difference is that while the link-age between bark beetles and warming climate is unequivocal, thebark beetle remains recovered from lake sediments outlined hereare intended for the sole purpose of reconstructing disturbanceevents and not necessarily to understand the past climate condi-tions under which the disturbance occurred.
2. Study area
2.1. Western North America
The recent bark beetle outbreaks began during the late 1980sand have persisted into the new millennium. Conifer-dominatedforests from Alaska to the American southwest have beenaffected during this period (Raffa et al., 2008). Other notable barkbeetle epidemics occurred during the early and middle 20th cen-turies, including major outbreaks in Utah during the 1920s (e.g.Dixon, 1935), Colorado during the 1940s (e.g. Schmid and Frye,1977), and Idaho in the 1920s and 1940s (e.g. Perkins andSwetnam, 1996). Beginning in the mid-1990s, a large irruptiveoutbreak of mountain pine beetle exceeded any previously knownoutbreak in severity and spatial extent in central British Columbia(Aukema et al., 2006). By 2002, the outbreak expanded northwardin British Columbia and eastward into Alberta (Nealis and Peter,2008; Robertson et al., 2009; de la Giroday et al., 2012). In recentyears, the mountain pine beetle has been observed to attack andcausemortality in jack pine (Pinus banksiana), which it has not beenknown to infest historically (Cullingham et al., 2011). This unprec-edented interaction is significant because Jack pine is an econom-ically important conifer and is widespread throughout theCanadian boreal forest (Safranyik et al., 2010). Throughout westernNorth American, the extreme nature of the widespread beetleoutbreak has resulted in a shift of these forests from net carbonsinks towards net carbon sources, providing a positive feedback toclimate warming (Kurz et al., 2008).
The 20 lake basins reviewed in this study generally satisfied thefollowing five conditions described by Morris and Brunelle (2012):(1) small surface area (<15 ha); (2) minimal (in)outflow surfacewater; (3) close proximity to beetle-killed forests; (4) catchmentsunmodified by large impoundments; (5) no large-scale post-outbreak logging operations; and (6) an absence of post-outbreakfire. In southeastern British Columbia, we investigated six lakesites occurring in watersheds that were affected by mountain pinebeetle during the early 2000s. Specifically, the forests surroundingthe lakes NEL01 and Sasquatch Lake experienced mixed-severitymortality while the higher elevation NEL03 site was little affectedby the recent epidemic (Fig. 3; Table 1). We also studied two siteson the White River Plateau in western Colorado. The White RiverPlateau is among of the more extensively studied bark beetle out-breaks (e.g. Love, 1955; Bethlahmy, 1974, 1975; Schmid and Frye,1977; Veblen et al., 1991; Kulakowski and Veblen, 2006). The sub-alpine landscape surrounding both Antler Pond and Bison Lake
J.L. Morris et al. / Quaternary International xxx (2014) 1e154
Please cite this article in press as: Morris, J.L., et al., Do bark beetle remains in lake sediments correspond to severe outbreaks? A review ofpublished and ongoing research, Quaternary International (2014), http://dx.doi.org/10.1016/j.quaint.2014.03.022
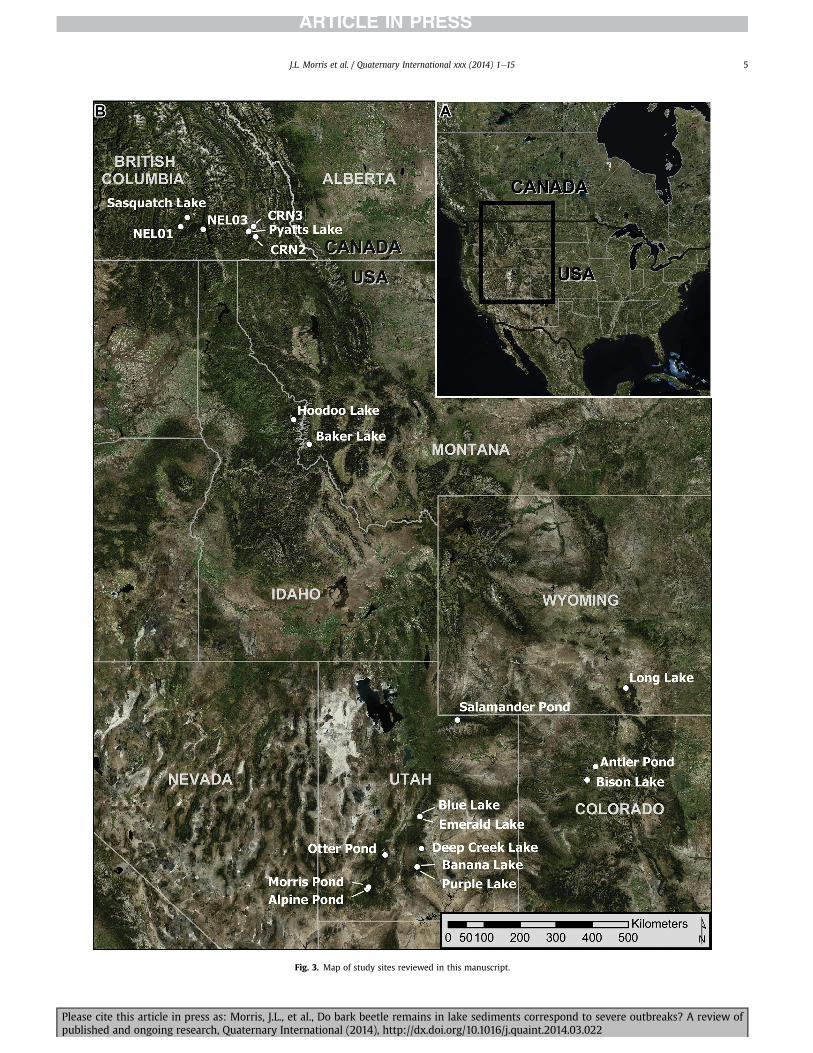
Fig. 3. Map of study sites reviewed in this manuscript.
J.L. Morris et al. / Quaternary International xxx (2014) 1e15 5
Please cite this article in press as: Morris, J.L., et al., Do bark beetle remains in lake sediments correspond to severe outbreaks? A review ofpublished and ongoing research, Quaternary International (2014), http://dx.doi.org/10.1016/j.quaint.2014.03.022
experienced a severe spruce beetle outbreak during the 1940s,resulting in <80% mortality in mature Engelmann spruce (Schmidand Frye, 1977). In Idaho, we report results from Hoodoo Lake,located on the western slope of the Bitterroot Range that occupies acutoff stream channel. Engelmann spruce and subalpine fir domi-nate the modern forest surrounding Hoodoo Lake. No historicalbark beetle activity has been reported in the watershed. In Mon-tana, we examined Baker Lake located on the eastern slope of theBitterroot Range, which is surrounded by a subalpine forestdominated by whitebark pine (Pinus albicaulis) and subalpine larch(Larix lyallii). The forest surrounding Baker Lake experienced anoutbreak of mountain pine beetle in 1920e1930s. In Utah, wepresent information on bark beetle outbreaks from the ColoradoPlateau and Uinta Mountains. The highlands of the ColoradoPlateau include a number of subalpine ranges, three of which wepresent here: the Wasatch Plateau, Aquarius Plateau, and
Markagunt Plateau. These forests are comprised of Engelmannspruce and subalpine fir, with lesser components of quaking aspen(Populus tremuloides) and limber pine (Pinus flexilis). Throughoutthe 1990s, spruce beetle outbreaks resulted in >90% mortality inEngelmann spruce across the Wasatch and Markagunt (Dymerskiet al., 2001; DeRose and Long, 2007). During the 1920s and 30s,spruce beetle killed <80% of mature Engelmann spruce on theAquarius Plateau (Dixon, 1935; Mielke, 1950). Data from six lakesaffected by these outbreaks are presented here: Blue and Emeraldlakes (Wasatch Plateau), Alpine and Morris ponds (MarkaguntPlateau), and Banana and Purple lakes (Aquarius Plateau). On thenorth-slope of the Uinta Mountains, mountain pine beetle out-breaks occurred during the 1920s, 1980s, and 2000s (Thorne, 1935;Utah DNR, 2003). Salamander Pond was cored to investigate themost recent of these outbreaks. In Wyoming, an unprecedentedmountain pine beetle outbreak affected w1.6 million ha of
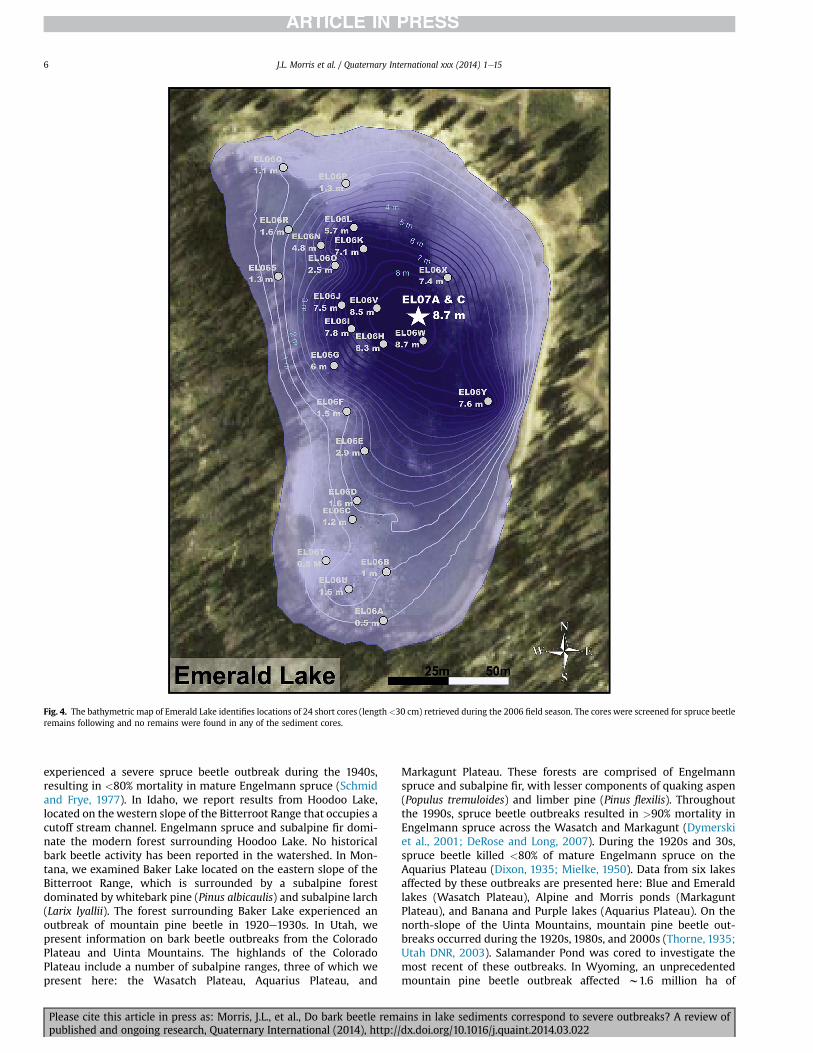
Fig. 4. The bathymetric map of Emerald Lake identifies locations of 24 short cores (length <30 cm) retrieved during the 2006 field season. The cores were screened for spruce beetleremains following and no remains were found in any of the sediment cores.
J.L. Morris et al. / Quaternary International xxx (2014) 1e156
Please cite this article in press as: Morris, J.L., et al., Do bark beetle remains in lake sediments correspond to severe outbreaks? A review ofpublished and ongoing research, Quaternary International (2014), http://dx.doi.org/10.1016/j.quaint.2014.03.022
lodgepole pine forest in southernWyoming and northern Coloradoin the late 2000s (Dennison et al., 2010; USFS, 2011). We presentdata from Long Lake, Wyoming, which is located in the MedicineBow River drainage (Atwood, 1937). The forest canopy at Long Lakeis dominated by lodgepole pine with interspersed subalpine fir(Abies bifolia) and Engelmann spruce. In sum, we discuss 43 sedi-ment cores, including 24 surface sediment cores that werecollected from Emerald Lake, Utah (Figs. 3 and 4, Tables 1 and 2).
3. Methods
3.1. Sediment core retrieval
The uppermost lake sediments that correspond to the 20thcentury are typically unconsolidated due to high water content. Tocapture the uppermost flocculent depositional layer, sediments areretrieved using one of a variety of devices. These coring apparatuses
Table 1Site characteristics.
Coring site Landform Designation Province or state Coordinates Elevation (m) Lake size (ha) Primary attack beetle Decade ofoutbreak
CRN2 Rocky Mountain Trench British Columbia 49! 220 4400 N115! 350 5200 W
1250 1 n/a None known
CRN3 Rocky Mountain Trench British Columbia 49! 330 0300 N115! 380 5800 W
818 3.5 n/a None known
Pyatts Lake Rocky Mountain Trench British Columbia 49! 280 1100 N115! 460 1300 W
1080 8.8 n/a None known
NEL01 Columbia Mountains British Columbia 49! 320 3000 N117! 270 5000 W
1526 0.86 Mountain pine beetle 2000-2010s
NEL03 Columbia Mountains British Columbia 49! 290 4700 N116! 540 1800 W
2074 0.35 n/a None known
Sasquatch Lake Columbia Mountains British Columbia 49! 410 3000 N117! 180 2700 W
1054 2 Mountain pine beetle 2000-2010s
Antler Pond White River Plateau Routt NF Colorado 40! 010 2200 N107! 070 0900 W
3128 0.1 Spruce beetle 1940s
Bison Lake White River Plateau Routt NF Colorado 39! 450 4700 N107! 200 8700 W
3280 15.3 Spruce beetle 1940s
Hoodoo Lake Bitteroot Mountains Clearwater NF Idaho 46! 190 1400 N114! 390 0100 W
1770 3.5 Mountain pine beetle 1920s
Baker Lake Bitteroot Mountains Bitteroot NF Montana 45! 530 3100 N114! 150 4300 W
2300 3.1 Mountain pine beetle 1920s
Blue Lake Wasatch Plateau Manti-LaSal NF Utah 39! 30 2000 N111! 300 1700 W
3129 3.2 Spruce beetle 1990s
Emerald Lake Wasatch Plateau Manti-LaSal NF Utah 39! 40 2600 N111! 290 5000 W
3090 3.3 Spruce beetle 1990s
Banana Lake Aquarius Plateau Dixie NF Utah 38! 40 3000 N111! 350 1200 W
3128 10.6 Spruce beetle 1930s
Purple Lake Aquarius Plateau Dixie NF Utah 38! 40 2800 N111! 340 1600 W
3226 6.2 Spruce beetle 1930s
Alpine Pond Markagunt Plateau Cedar Breaks NM Utah 37! 380 1100 N112! 490 2600 W
3172 0.1 Spruce beetle 1990s
Morris Pond Markagunt Plateau Dixie NF Utah 37! 400 2500 N112! 460 4900 W
3126 0.7 Spruce beetle 1990s
Salamander Pond Uinta Mountains Ashley NF Utah 40! 540 0700 N110! 340 1200 W
3074 1 Mountain pine beetle 2000s
Deep Creek Lake Thousand Lakes Mountain Fishlake NF Utah 38! 260 0000 N111! 270 5100 W
3222 2.3 n/a None known
Otter Pond Tuschar Mountains Fishlake NF Utah 38! 180 2900 N112! 220 0900 W
2914 1 Spruce beetle 1970s
Long Lake Medicine Bow Mountains Medicine Bow NF Wyoming 41! 300 0600 N106! 220 0500 W
2700 12 Mountain pine beetle 2000s
Table 2Coring and sampling specifics.
Site Coring device Subsampleresolution
Subsamplevolume (cc)
Disaggregate Citation
Antler Pond Modified Livingstone 0.5 cm 15 cc None Anderson et al., 2010; Watt, 2008Bison Lake Glew gravity corer, Livingston piston corer 0.5 cm 5 cc Na6P6O18 Anderson, 2011Hoodoo Lake Klein corer 1 cm 5 cc Na6P6O18 Brunelle et al., 2005; Brunelle et al., 2008Baker Lake Klein corer 1 cm 5 cc Na6P6O18 Brunelle et al., 2005, 2008Blue Lake Gravity core 1 cm 5 cc Na6P6O18 Morris et al., 2010Emerald Lake Klein corer 1 cm 5 cc Na6P6O18 Morris and Brunelle, 2012; Morris et al., 2013aBanana Lake Klein corer 1 cm 5 cc Na6P6O18 Morris and Brunelle, 2012; Morris et al., 2013aPurple Lake Klein corer 1 cm 5 cc Na6P6O18 Morris and Brunelle, 2012; Morris et al., 2013aAlpine Pond Klein corer 1 cm 5 cc Na6P6O18 Morris and Brunelle, 2012; Morris et al., 2013aMorris Pond Klein corer 1 cm 5 cc Na6P6O18 Morris and Brunelle, 2012; Morris et al., 2013aDeep Creek Lake Klein corer 1 cm 5 cc Na6P6O18 Morris unpublished dataOtter Pond Klein corer 1 cm 5 cc Na6P6O18 Morris unpublished data
(continued on next page)
J.L. Morris et al. / Quaternary International xxx (2014) 1e15 7
Please cite this article in press as: Morris, J.L., et al., Do bark beetle remains in lake sediments correspond to severe outbreaks? A review ofpublished and ongoing research, Quaternary International (2014), http://dx.doi.org/10.1016/j.quaint.2014.03.022

can be generally described as falling into of two categories: (1)piston and (2) box devices. Piston corers are deployed either viagravity (i.e. a gravity core) or are manually driven into the sedimentcolumn (e.g. Klein corer). Glew gravity corers (Glew et al., 2001)employ the use of a plunger positioned at the top of a tube to holdand lift the topmost sediments. Box devices use a hand-triggered‘grab’ mechanism that encloses sediments within a chamber (e.g.Wright, 1991). Other box-type devices utilize a freezing techniqueachieved via a combination of ethyl alcohol (C2H6O) and dry ice thatcauses the sediments to freeze to the outside of the box (e.g.Anderson et al., 2010). Table 2 provides a list of coring devices thatwere used to retrieve the upper most sedimentary layers for thesites discussed here.
3.2. Macrofossil analysis
Sediment cores were subsampled contiguously at volumes of 1e5 cm3. Subsamples were disaggregated in Whirl-Pak! bags in asolution of distilled water and sodium hexametaphosphate(Na6P6O18) (Bamber, 1982). Table 2 summarizes subsampling stra-tegies for the cores we review here. Disaggregated samples werescreened through nested sieves with mesh sizes of 125 mm and250 mm. These dimensions are commonly used in macroscopiccharcoal analysis (e.g. Whitlock and Larsen, 2001) and these meshsizes are sufficient to retain adult Dendroctonus spp. elytra and headcapsules (Fig. 2). However, some studies suggest that larger meshsizes (300 mm) are also sufficient to retain adult Dendroctonusbeetles (e.g. Elias, 1985, 2001).
3.3. Insect remains confirmation
For elytra and head capsules to be identified to the species-specific level required for taxonomic assignment, bark beetle re-mains should be submitted to a qualified entomologist. Dependingon the state of preservation, it may not be possible to diagnoseelytra and/or head capsules confidently (Brunelle et al., 2008;Morris et al., 2010). To our knowledge, intact bark beetle car-casses have yet to be observed in Holocene lacustrine sediments.Therefore, elytra, which are more common (e.g. Elias, 2001), andhead capsules, which are less common, are the most useful barkbeetle remains to be submitted for taxonomic identification (Fig. 2).Specimens from the British Columbia sites were submitted to Dr.Hume Douglas at the Canadian Food Inspection Agency, OttawaPlant Laboratory. Suspected bark beetle remains from Idaho andMontana were submitted to Dr. Stephen Wood at Brigham YoungUniversity (Wood, 1982; Brunelle et al., 2008). Antler Pond barkbeetle remains were submitted to Drs. Richard Hofstetter (NorthernArizona University), Anthony Cognato (Michigan State University),and Kerry Katovitch (University of Wisconsin-Whitewater) (Watt,2008). Potential bark beetle remains recovered from the Utah andWyoming sites were submitted to Dr. James Pitts at Utah StateUniversity (Carter, 2010; Morris and Brunelle, 2012).
3.4. Laboratory testing
During fieldwork in 2005, lake water conditions at Blue Lake,Utah were observed to have a pH of 9.5. Morris et al. (2010) re-ported that diagnostic insect remains were not recovered from BlueLake sediment subsamples. However, translucent chitinous resi-dues were observed in sediments within the same stratigraphiccontext as significant declines in spruce pollen attributed to thespruce beetle outbreak (Morris et al., 2010). Morris et al. (2010)provided a brief literature synthesis suggesting that bark beetlechitin is susceptible to chemical dissolution, particularly in alkalineenvironments. To test this hypothesis, we exposed spruce beetlecarcasses to a strong alkaline solution of potassium hydroxide(KOH) (pH 9.5) for two weeks to assess how conditions similar tothose observed at Blue Lake might influence bark beetle chitinpreservation. A period of two weeks was selected to mimic theduration of spruce beetle emergence (Dyer, 1969), while alsoensuring a manageable time to conduct a laboratory experiment.The pH of the KOH solution was monitored daily using aMilwaukee! pH meter that was manually calibrated before eachreading using standard buffer solutions.
In a separate experiment to test the likelihood of the depositionof beetle carcasses into the lake system, 100 live spruce beetles,represented equally between male and female, were introduced toa 5-L beaker of tap water in the laboratory. This crude experimentwas designed to assess whether beetle carcasses would eventuallysink into the water column or whether they would remainbuoyant following mortality. The spruce beetles were observedover a two-week period (similar to the duration of spruce beetleemergence).
4. Results
4.1. British Columbia
Well-preserved Scolytinae remains were observed in sixHolocene-aged lake sediment cores, although no primary attackbeetles were confirmed (Fig. 3; Tables 3 and 4). While no fullyintact individual beetle remains were identifiable, the elytra,prothorax, and head capsules of secondary beetles were posi-tively identified (Fig. 5). Secondary attack beetles (e.g. Pseudo-hylesinus) were identified in the sediments collected from CRN3and Sasquatch Lake. The latter site also contained the remains ofa twig beetle (Pityophthorus) as well as the secondary barkbeetles Hylastes, Scolytus, and Carphoborusi. At NEL03, Carpho-borusi and Phloeosinus were recovered and positively identified.Images of the secondary bark beetles that were recovered fromCRN3, Sasquatch Lake, and NEL03 are shown in Fig. 5 and aresummarized in Table 3. In addition, potential bark beetle remainsthat compare favorably with Scolytinae were recovered fromCRN2, Pyatts Lake, and NEL01 and are awaiting taxonomicconfirmation.
Table 2 (continued )
Site Coring device Subsampleresolution
Subsamplevolume (cc)
Disaggregate Citation
Long Lake Klein corer 1 cm 5 cc Na6P6O18 Carter, 2010; Dennison et al., 2010Salamander Pond Klein corer 1 cm 5 cc Na6P6O18 Carter in prepNEL01 Glew gravity corer, Livingston piston corer 0.25e0.5 1 cc Na6P6O18 Courtney Mustaphi and Pisaric, 2013NEL03 Glew gravity corer, Livingston piston corer 0.25e0.5 1 cc Na6P6O18 Courtney Mustaphi and Pisaric, 2013, in pressSasquatch Lake Glew gravity corer, Livingston piston corer 0.25e0.5 1 cc Na6P6O18 Courtney Mustaphi, 2013CRN2 Glew gravity corer, Livingston piston corer 0.25e0.5 1 cc Na6P6O18 Courtney Mustaphi, unpublished dataCRN3 Glew gravity corer, Livingston piston corer 0.25e0.5 1 cc Na6P6O18 Courtney Mustaphi, unpublished dataPyatts Lake Glew gravity corer, Livingston piston corer 0.25e0.5 1 cc Na6P6O18 Courtney Mustaphi, unpublished data
J.L. Morris et al. / Quaternary International xxx (2014) 1e158
Please cite this article in press as: Morris, J.L., et al., Do bark beetle remains in lake sediments correspond to severe outbreaks? A review ofpublished and ongoing research, Quaternary International (2014), http://dx.doi.org/10.1016/j.quaint.2014.03.022

4.2. Colorado
No primary attack beetle (i.e. Dendroctonus spp.) remains wererecovered, however secondary bark beetles were recovered andidentified from the Antler Pond sediments. The confirmed remainsincluded several species from the following genera: Pityophthorous,Carphoborus, Phloeosinus, Scolytus, and others only identifiable tothe sub-family Scolytinae. Images of the identified taxa are pro-vided in Fig. 6 and are summarized in Tables 3 and 4.
4.3. Idaho
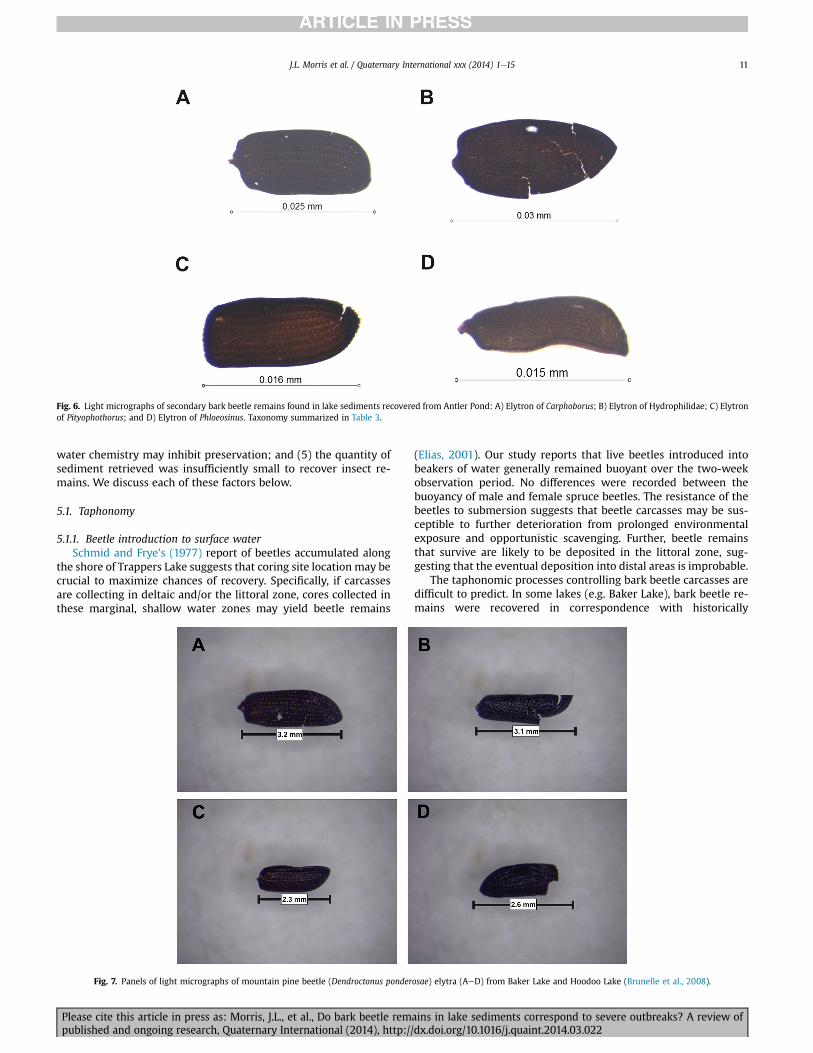
Primary attack beetle remains were recovered and identifiedfrom Hoodoo Lake (Fig. 7). Brunelle et al. (2008) provide evidencethat the recovered mountain pine beetle remains correspond to the
years 7954 and 8163 cal BP. Although the forest composition thatsurrounds Hoodoo Lake is currently dominated by Engelmannspruce and subalpine fir, a pollen- and macrofossil-based recon-struction from Hoodoo Lake indicates that during the time periodcorresponding tomountain pine beetle remains, thewatershedwasdominated by whitebark pine (P. albicaulis) (Brunelle et al., 2008).
4.4. Montana
At Baker Lake, primary attack beetle remains were recovered(Fig. 7). Brunelle et al. (2008) demonstrated that these remains dateto 8331 cal BP, 8410 cal BP, and 8529 cal BP. Additionally, Brunelleet al. (2008) recovered primary attack beetle remains (mountainpine beetle) that also corresponded to the 1920e1930s historicmountain pine beetle outbreak. The specimens that were recoveredfrom the early Holocene correspond temporally to those recoveredfrom Hoodoo Lake, Idaho.
4.5. Utah
No primary bark beetle remains were recovered from any of theColorado Plateau and Uinta sites. No diagnostic remains of primaryor secondary attack beetles were recovered from any of the 24 shortsediment cores analyzed from Emerald Lake (Fig. 4). Of the elytrathat were recovered, the remains of secondary bark beetles werenot encountered.
4.6. Wyoming
Suspected bark beetle elytra were recovered from Long Lake.However, these specimens were determined not to be Scolytinaebeetles.
4.7. Laboratory experiments
Monitored exposure of spruce beetle carcasses to alkaline so-lution confirms that bark beetle chitin preservation is sensitive tolake water pH. Times-series light micrographs are provided inFig. 8, which depicts a rubbery trace protein (i.e. reselin). Reselin isa chitinous residue that persists following exposure to moderatelyalkaline solution that has various applications in the pharmaceu-tical industry (Morris et al., 2010 and references therein).
Of the 100 spruce beetles introduced into water, less than 5% ofbeetle carcasses became fully submerged in the 5-L beaker over thetwo-week period in two replicate experiments (n ! 5, n ! 3).Mortality was observed for 100% of all beetles utilized in thisexperiment.
5. Discussion
To date, our efforts to understand how bark beetles impactedpast ecosystems have been moderately successful. Of the 20 sitespresented here, only two sites, Hoodoo Lake and Baker Lakerecovered primary bark beetle (Dendroctonus spp.) remains. Threesites, Antler Pond, CRN3, and NEL02 recovered secondary barkbeetles. The remaining sites either did not preserve any bark beetleremains, or remains (other than primary attack beetles) were notsubmitted for identification.
Schmid and Frye (1977) report that during a spruce beetleoutbreak in July 1949, accumulations of spruce beetle carcasseswere observed in drifts for at least 1 km along the shoreline ofTrappers Lake. Trappers Lake is located on the White River Plateauin western Colorado, proximal to Antler Pond and Bison Lake re-ported on here. If these carcass accumulations are indicative ofsevere outbreaks, based on this report it is hardly surprising that
Table 3Secondary beetle taxa found in lake sediment stratigraphies.
Lake Order Family Sub-family Genus Species
Antler Pond Coleoptera Curculionidae Scolytinae Pityophthorus sp. 1Antler Pond Coleoptera Curculionidae Scolytinae Pityophthorus sp. 2Antler Pond Coleoptera Curculionidae Scolytinae CarphoborusAntler Pond Coleoptera Curculionidae Scolytinae ScolytusAntler Pond Coleoptera Curculionidae Scolytinae PhloeosinusAntler Pond Coleoptera HydrophilidaeAntler Pond Coleoptera Curculionidae ScolytinaeAntler Pond Coleoptera (terrestrial)Antler Pond OdonataAntler Pond Coleoptera HydrophilidaeCRN3 Coleoptera Curculionidae Scolytinae PseudohylesinusNEL01 Coleoptera Curculionidae Scolytinae Pityophthorus plagiatus
knechteliNEL01 Coleoptera Curculionidae Scolytinae PseudohylesinusNEL01 Coleoptera Curculionidae Scolytinae PseudohylesinusNEL01 Coleoptera Curculionidae Scolytinae Carphoborus
carriNEL01 Coleoptera Curculionidae Scolytinae ScolytusNEL01 Coleoptera Curculionidae Scolytinae PseudohylesinusNEL01 Coleoptera Curculionidae Scolytinae HylastesNEL03 Coleoptera Curculionidae Scolytinae Carphoborus
carriNEL03 Coleoptera Curculionidae Scolytinae Phloeosinus
Table 4Summary of presence/absence results for primary and secondary attack beetles.
Lake Primary beetlesrecovered
Secondary beetlesrecovered
CRN2 No NoCRN3 No YesPyatts Lake No NoNEL01 No YesNEL03 No NoSasquatch Lake No NoAntler Pond No YesBison Lake No NoHoodoo Lake Yes NoBaker Lake Yes NoBlue Lake No NoEmerald Lake No NoBanana Lake No NoPurple Lake No NoAlpine Pond No NoMorris Pond No NoSalamander Pond No NoDeep Creek Lake No NoOtter Pond No NoLong Lake No No
J.L. Morris et al. / Quaternary International xxx (2014) 1e15 9
Please cite this article in press as: Morris, J.L., et al., Do bark beetle remains in lake sediments correspond to severe outbreaks? A review ofpublished and ongoing research, Quaternary International (2014), http://dx.doi.org/10.1016/j.quaint.2014.03.022

Brunelle et al. (2008) recoveredmountain pine beetle remains fromthe Baker Lake sediments that are coeval with the 1920s outbreak.Furthermore, mountain pine beetle remains were also recoveredfrom Hoodoo Lake and Baker Lake that were deposited during theearly Holocene, which suggests that remains are a reliable distur-bance indicator. The site characteristics and coring locations atHoodoo Lake and Baker Lake are similar to other sites reviewedhere. Therefore, it is surprising that no remains of primary attackbeetles were recovered in any of the sediment cores analyzed fromBritish Columbia, Colorado, Utah, and Wyoming. It is noteworthythat secondary bark beetles were successfully recovered andidentified from Antler Pond (Colorado), CRN3, Sasquatch Lake,NEL01, and NEL03 (British Columbia). Specifically, elytra and pro-thorax remains were observed in sediments from these lakes. Thegreatest abundance of secondary beetle remains was found in
Sasquatch Lake, a small lakewith a steep catchment situatedwithina dense, mid-elevation forest. Inconsistent recovery of both pri-mary and secondary beetles indicates that the taphonomic pro-cesses controlling bark beetle deposition and preservation in lakesediments are complex and require further study.
All 20 lake basins reviewed here followed the site selectionprotocol outlined in the methods section above (Morris andBrunelle, 2012). Despite conformance to protocol, inconsistent re-covery of bark beetle remains was observed. We propose that thiscould be due to a number of factors, including: (1) beetle intro-duction to surface water occurs unpredictably; (2) current samplepreparation protocols fail to adequately preserve sensitivechitinous remains; (3) other biotic entities may be consumingbeetle carcasses within the water column before deposition andpreservation in sediment can occur (e.g. fish, microbes); (4) lake
Fig. 5. Light photomicrographs of secondary attack beetles recovered from lake sediment cores in British Columbia: A) Posterior fragment of Pseudohylesinus elytron from at CRN3;B) Pseudohylesinus elytron collected from Sasquatch Lake; C) Left-lateral view of Pityophthorus plagiatus knechteli prothorax found at Sasquatch Lake; D) Left-lateral view of pro-thorax found at Sasquatch Lake; E) Elytron of Carphoborus carri from Sasquatch Lake; and F) Elytron of Hylastes from Sasquatch Lake. Scale bar represents 0.5 mm. Taxonomysummarized in Table 3.
J.L. Morris et al. / Quaternary International xxx (2014) 1e1510
Please cite this article in press as: Morris, J.L., et al., Do bark beetle remains in lake sediments correspond to severe outbreaks? A review ofpublished and ongoing research, Quaternary International (2014), http://dx.doi.org/10.1016/j.quaint.2014.03.022
water chemistry may inhibit preservation; and (5) the quantity ofsediment retrieved was insufficiently small to recover insect re-mains. We discuss each of these factors below.
5.1. Taphonomy
5.1.1. Beetle introduction to surface waterSchmid and Frye’s (1977) report of beetles accumulated along
the shore of Trappers Lake suggests that coring site location may becrucial to maximize chances of recovery. Specifically, if carcassesare collecting in deltaic and/or the littoral zone, cores collected inthese marginal, shallow water zones may yield beetle remains
(Elias, 2001). Our study reports that live beetles introduced intobeakers of water generally remained buoyant over the two-weekobservation period. No differences were recorded between thebuoyancy of male and female spruce beetles. The resistance of thebeetles to submersion suggests that beetle carcasses may be sus-ceptible to further deterioration from prolonged environmentalexposure and opportunistic scavenging. Further, beetle remainsthat survive are likely to be deposited in the littoral zone, sug-gesting that the eventual deposition into distal areas is improbable.
The taphonomic processes controlling bark beetle carcasses aredifficult to predict. In some lakes (e.g. Baker Lake), bark beetle re-mains were recovered in correspondence with historically
Fig. 6. Light micrographs of secondary bark beetle remains found in lake sediments recovered from Antler Pond: A) Elytron of Carphoborus; B) Elytron of Hydrophilidae; C) Elytronof Pityophothorus; and D) Elytron of Phloeosinus. Taxonomy summarized in Table 3.
Fig. 7. Panels of light micrographs of mountain pine beetle (Dendroctonus ponderosae) elytra (AeD) from Baker Lake and Hoodoo Lake (Brunelle et al., 2008).
J.L. Morris et al. / Quaternary International xxx (2014) 1e15 11
Please cite this article in press as: Morris, J.L., et al., Do bark beetle remains in lake sediments correspond to severe outbreaks? A review ofpublished and ongoing research, Quaternary International (2014), http://dx.doi.org/10.1016/j.quaint.2014.03.022

documented outbreaks. At other sites, only secondary beetles wererecovered in correspondence with historical outbreaks (AntlerPond and British Columbia lakes). Some sites yielded no diagnosticremains despite systematic and considerable sampling across var-iable depths and locations, including littoral zones (Emerald Lake;Fig. 4), while in others suspected beetle remains (i.e. Dendroctonusspp.) were degraded beyond taxonomic confirmation (Blue Lake).
Morris and Brunelle (2012) suggested that the significantcarcass accumulations cited by Schmid and Frye (1977) werefacilitated by stormy conditions with frequent and/or abundantprecipitation at the time of beetle flight (when adults emerge fromhost trees). It is likely unusual that beetles would come into contactwith surface water because bark beetles spend the majority of theirlives in subcortical chambers of the host tree and emerge for flightonly briefly (approximately two weeks) during their 1e2 yearlifespan (Holsten et al., 1999). Beetles emerge to seek new hostswhen subcortical and ambient air temperatures reach 9 and 16 !C,respectively (Dyer, 1969; Wood, 1982). In much of the native rangeof bark beetles, including the US Rocky Mountains, temperaturesconducive for beetle flight may occur in late May or June. For theJune 1949 event of extensive spruce beetle carcass drifts at TrappersLake reported by Schmid and Frye (1977), the National Climate DataCenter recorded the single wettest June during the 20th century(National Climate Data Center, 2010). Abundant, frequent precipi-tation would increase the probability that beetle flight would bedisrupted and these weather conditions would also increase the
likelihood that downed beetles would come into contact with thelake surface and be deposited into the sediment matrix.
Although Watt (2008) did not recover primary attack beetles atAntler Pond, numerous secondary beetles were recovered in a stateof preservation sufficient for taxonomic confirmation. Interestingly,the period of the Antler Pond record with the greatest amount ofbeetle recovery occurred at a time of reduced lake level (Watt,2008). Because the Antler Pond core was collected in the modernlittoral zone of a small basin, during the time period when sec-ondary beetle remains were recovered, this coring area was likelyto have been the forest floor. This interpretation is supported by anabundance of conifer needles and other organic detritus that ap-pears compositionally similar to a duff layer. Given the comparativeabundance of secondary bark beetle remains recovered from thispart of the core, it is possible that soil cores may yield bark beetlestratigraphies useful for deriving disturbance chronologies.
5.1.2. Subsample preparation and reference materialWe have not explicitly tested the effects of commonly applied
disaggregating solutions on the perseveration of the chitinous re-mains of bark beetles. In general, disaggregation of bark beetles andother Coleopteran remains from sediment matrix may be achievedthrough floatation of bulk sediment using apolar liquids (i.e.Kerosene), which has yielded successful results (Elias, 2001). Pre-vious studies have also shown that chitin is insoluble in dilute acids(e.g. HCL) and weak bases (e.g. KOH), deionized water, and ethylalcohol (ETOH) (Elias, 2010). Chemical treatment may or may not,cause deproteinization of chitin and/or alter d18O isotopic values,based on studies of chitinous chironomid head capsules (Woolleret al., 2004; Verbruggen et al., 2010). To our knowledge this con-dition has not been explicitly tested on bark beetle remains. Thesepotential effects should be considered prior to sediment processing,depending upon the research objectives of the study. Elias (2010)lists the use of Calgon! (a sodium hexametaphosphate-baseddetergent) and warm water for disaggregating clay-rich sedi-ments. Once insect remains are isolated, recommended storage is inethyl alcohol until analysis (Elias, 2010).
For identification of sub-fossil remains, it is essential to build amodern reference collection of taxonomically identified species. Ofparticular importance are elytra, and to a lesser degree head cap-sules (Fig. 2). Investigators can pursue one of several potentialstrategies to obtain intact bark beetle carcasses, including phero-mone baited traps, bark removal and beetle extraction from infes-ted trees, and preservation of elytra and beetles recovered fromlake sediments (e.g. Figs. 5e7). To preserve diagnosed beetles and/or elytra for referencematerial, insect remains must first bewashedwith alcohol, then dried. Elytra and head capsules can then beplaced into a depression slide. Identification of beetle remainsrecovered from lake sediments should be confirmed by a qualifiedColeopteran taxonomist. Clear epoxy/resin (e.g. acrylic) can then beapplied to encase the beetle remains. Slides should be stored in acool, dark environment similar to other slide-mounted referencematerials. Reference insect slides can be examined using light mi-croscopy at 10 " magnification similar to sedimentary charcoalpreparations (e.g. Whitlock and Larsen, 2001).
5.1.3. Potential role of fishAnthropogenic impacts on lake ecosystems during the 20th
century may have inadvertently influenced the taphonomy of barkbeetle carcasses. Introduced salmonid fish, such as trout, into highelevation lakes have significant effects on benthic micro-invertebrates and dytiscid beetles (aquatic beetles), and abun-dances of these communities decrease after fish introduction(Knapp et al., 2001). Stocking of game fish can then conceivablyaffect terrestrial bark beetle deposition and preservation, as
Fig. 8. Degraded chitinous remains were observed in sediments from Blue Lakecontemporaneous with the 1990s outbreak. Laboratory testing suggests preservationof spruce beetle carcasses may be inhibited by lake water chemistry, microbialdegradation, and/or ingestion by salmonid fish (Morris et al., 2010).
J.L. Morris et al. / Quaternary International xxx (2014) 1e1512
Please cite this article in press as: Morris, J.L., et al., Do bark beetle remains in lake sediments correspond to severe outbreaks? A review ofpublished and ongoing research, Quaternary International (2014), http://dx.doi.org/10.1016/j.quaint.2014.03.022

introduced fish also prey on terrestrial beetles (Allen, 1981). Forexample, a recent synthesis concludes that many types of salmonidfish feed exclusively on terrestrial insects (Keeley and Grant, 2001).Bark beetles would be an enticing addition to the diets of pelagicfish, an effect that is likely enhanced in lakes where fish pop-ulations are re-stocked annually. A review of state-level stockingreports suggests that many subalpine lakes are stocked annuallysince the 1950s (e.g. Utah Division of Wildlife Resources, 2013).Given the likelihood that unnaturally high fish populations exist inmany subalpine lakes across the west, it is not entirely surprisingthat beetle remains have not been recovered in recent outbreaks.Assuming that salmonid fish introduction in many subalpine sys-tems is unique to the 20th century, it is presumable that fish pop-ulations during the recent bark beetle outbreaks are greater thanprior to the mid-20th century. Therefore, we speculate that barkbeetle remains in sediments prior to the introduction of salmonidsmay not be reproducible in lakes that have recently been stockedwith game fish.
5.1.4. DegradationChitin is potentially susceptible to degradation from aerobic
decay, dissolution in moderate to strong alkaline and acidic water,fungal growth, and bacterial decomposition or interactions amongthese agents (Allison, 1986, 1988; Baas et al., 1995; Flintoft, 2004).Fig. 8 depicts time lapse photographs of chitin exposed to alkalinesolution for two weeks at pH 9.5. This pH is within the range ofnatural variability observed in small lakes occurring in calcareoussubstrate (Morris et al., 2010). The dissolution of diagnostic featureson the sclerorized exoskeleton evident in these photos suggeststhat positive identification of remains can be hampered by limno-logic conditions. Thesemicrographs provide images that are similarin appearance to the reselin smears observed in the Blue Lakesediments by Morris et al. (2010). Seasonal variations in lake waterpH fluctuations have been documented and can be considerable insmall lake basins (Tank et al., 2009; Xu et al., 2010). While seasonalfluctuations in lake water chemistry are challenging to assessduring site selection, pH is an important control on chitin preser-vation to be considered. Future laboratory research should alsoinvestigate the role of acidic conditions and potentially use NaHCO3(instead of KOH), which may more closely mimic alkaline lakewater conditions.
5.1.5. Sediment quantityOur synthesis suggests that the remains of primary attack bee-
tles are rare in lake sediments, at least using small-diameter pistondevices common in multi-proxy studies. Future studies may benefitfrom utilizing sampling approaches that retrieve greater volumesof sediment (w1 L).
6. Conclusions
Amplified by warming climate conditions and pervasivedroughts, the current continental-scale bark beetle outbreak maybe unprecedented in scale and severity when considered overcentennial- to millennial-timescales. Irruptive native bark beetleoutbreaks are part of an expanding positive climate feedback sys-tem, where increasingly warmer climates encourage bark beetlepopulations, affecting ecosystem processes and forest composition.To date, our efforts to understand how bark beetles impactedecosystems in the past have been moderately successful. Of the 20sites presented, only two sites, Hoodoo Lake and Baker Lake hadprimary bark beetle (Dendroctonus spp.) remains. Three sites,Antler Pond, CRN3, and NEL02 had secondary bark beetles. Theremaining sites either did not have any bark beetle remains, orremains (other than primary attack beetles) were not submitted for
identification. Secondary beetles may also be useful in providingsupporting information about past forest health conditions, how-ever these insects presumably infest dead trees that could havedied from a variety of disturbances including wildfire, windthrow,and avalanche.
Several generalizations result from our synthesis, which areinstructive for future research. The presence of standing-deadbeetle-killed timber in proximity to the lake margin is likelyimportant. Endorheic lakes without outflowing streams are ideal,but we did not explore the importance of inflowing streams, whichcould increase the likelihood of beetle carcass influx into the lake,particularly in deltaic settings (Elias, 2001). Laboratory observa-tions suggest that live beetle bodies do not immediately becomesubmerged in the water column, although our laboratory tests onlyexamined spruce beetle. Future laboratory experiments testing thebuoyancy of other primary beetles as well as secondary beetleswould be appropriate. The influence of waves and surface tensionsin lakes exposed to ambient atmospheric conditions has yet to beexplored, but presumably could influence the buoyancy of insectcarcasses. Laboratory experiments utilizing wave machines withsimulated precipitation are needed to better mimic ambient envi-ronmental conditions to test the buoyancy of carcasses. It may alsobe advantageous to concentrate on obtaining sediment cores in thelittoral zone along the margins of lakes (Elias, 2001), even thoughthere is no guarantee of recovering primary attack beetles (e.g. thesystematic sampling of Emerald Lake). Lakes that are not stockedwith fish and/or those lakes that do not have fish populations mayor may not increase chances of finding beetles. Similarly, lakes withpH neutral to weakly acidic may be preferable over moderatelyalkaline lakes.
Sampling efforts reviewed in this paper focused on the deepdistal areas of lakes as well as across transects that include thelittoral zone. These efforts have yielded few remains of primaryattack beetles. This is a surprising result considering the results ofBrunelle et al. (2008) and the number of beetles that may come incontact with the surface of lakes during large outbreaks (e.g.Schmid and Frye, 1977). Transport, deposition, taphonomic, anddiagenetic processes may be responsible for the lack of remainsrecovered. Additionally, the volume of sediment collected may beinsufficiently small to reliably yield beetle remains. Remains maybe more prevalent in portions of lake basins that are not oftentarget locations for multi-proxy studies that retrieve sediment ar-chives for distal areas of lakes. Entomologists seeking to collectspecimens frequently sieve littoral zone sediments to collectspecimens. As for other macrofossils, insect remains may concen-trate where aquatic plants and macrophytes collect material. Pre-vailing water and air currents may force floating and suspendedbeetle carcasses to specific areas of a lake. Future studies shouldinvestigate areas of lakes that tend to concentrate buoyant materialand possibly consider other natural archives, such as peat, uncon-solidated terrestrial sediments, and soil as insect remains alsosometimes preserve at such sites (Elias, 1985; Andersen, 1993).Sampling greater volumes of sediment (w1 L) may produce lessrobust chronologies, however overlapping coarse archives withhigh-resolution lake sediment analyses may be worthy of explo-ration in future work. Analysis of multiple terrestrial and epilittoralarchives in conjunction with high-resolution palynological studiesof lake sediments may provide robust records of irruptive beetles inforested ecosystems that exhibit a strong host-non-host signal(Morris and Brunelle, 2012). Peatlands may also be worthy ofinvestigation, as they are known to collect insect remains, areacidic, and fish predation would not be an issue, though terrestrialscavengers may present similar challenges.
Our synthesis suggests that the remains of primary attack bee-tles are rare in lake sediments, at least using small-diameter piston
J.L. Morris et al. / Quaternary International xxx (2014) 1e15 13
Please cite this article in press as: Morris, J.L., et al., Do bark beetle remains in lake sediments correspond to severe outbreaks? A review ofpublished and ongoing research, Quaternary International (2014), http://dx.doi.org/10.1016/j.quaint.2014.03.022

devices common in multi-proxy studies. Reconstructing insect andpathogen disturbances remains an important challenge for paleo-ecology (Waller, 2013; Seddon et al., 2014). The scale and severity ofrecent outbreaks and the high probability of outbreaks moving intoregions where the thermal habitat has been historically unsuitablefor brood reproduction and overwintering survival underscores thenecessity of long-term understanding of these disturbance agents,their ecological role, and interactionwith other disturbance agents.Future research is needed to understand the potential controls onbeetle carcass taphonomy and deposition that we have discussedhere. Reconstructing past beetle disturbances using opportunisticsecondary beetles must be tested across a diversity of ecosystems.The linkage between primary beetle outbreaks and climatewarming is extensively studied. However, the relationship betweensecondary beetles and climate has not been explicitly explored andprovides an important opportunity for future research.
Acknowledgements
We thank Dr. Barbara Bentz for providing spruce beetle speci-mens that were analyzed in our laboratory experiments. We aregrateful to Dr. Lesleigh Anderson for enabling the analysis of theBison Lake sediments. We are indebted to Corey Unger and Dr.Phoebe McNeally for producing the maps in Figs. 2 and 3, respec-tively. We also recognize the entomologists that assisted in classi-fying insect remains. We thank Drs. Hume Douglas and Mitchell J.Power for use of photographic equipment at their respective labs.We are grateful to Drs. Scott Staratt and Jeannine St. Jacques fortheir efforts in compiling this special issue of Quaternary Interna-tional as well as their comments and suggestions that lead to sig-nificant improvements in our paper. This manuscript alsobenefitted from critiques submitted by two anonymous reviewers.
References
Allen, J.D., 1981. Determinants of diet of brook trout (Salvelinus fontinalis) in amountain stream. Canadian Journal of Fisheries and Aquatic Sciences 38, 184e192.
Allison, P.A., 1986. Soft-bodied animals in the fossil record: the role of decay infragmentation during transport. Geology 14, 979e981.
Allison, P.A., 1988. The role of anoxia in the decay and mineralization of proteina-ceous macro-fossils. Paleobiology 14, 139e154.
Amin, H., Atkins, P.T., Russo, R.S., Brown, A.W., Sive, B., Hallar, A.G., Hartz, K.E.H.,2012. Effect of bark beetle infestation on secondary organic aerosol precursoremissions. Environmental Science and Technology 46, 5696e5703.
Andersen, J., 1993. Beetle remains as indicators of the climate in the Quaternary.Journal of Biogeography 20, 557e562.
Anderson, L., 2011. Holocene record of precipitation seasonality from lakecalcite d18O in the central Rocky Mountains, United States. Geology 39,211e214.
Anderson, R.S., Davis, R.B., Miller, N.G., Stuckenrath, R., 1986. History of late-andpost-glacial vegetation and disturbance around Upper South Branch Pond,northern Maine. Canadian Journal of Botany 64, 1977e1986.
Anderson, R.S., Smith, S.J., Lynch, A.M., Giels, B.W., 2010. The pollen records of a 20th
century spruce beetle (Dendroctonus rufipennis) outbreak in a Colorado subal-pine forest, USA. Forest Ecology Management 260, 448e455.
Ashworth, A.C., 1977. A late Wisconsin Coleopterous assemblage from southernOntario and its environmental significance. Canadian Journal of Earth Science14, 1625e1634.
Ashworth, A.C., Schwert, D.P., Watts, W.A., Wright, H.E., 1981. Plant and insect fossilsat Norwood in south-central Minnesota: a record of late-glacial succession.Quaternary Research 16, 66e79.
Atwood Jr., W.W., 1937. Records of Pleistocene glaciers in the Medicine Bow andPark Ranges. Journal of Geology 45, 113e140.
Aukema, B.H., Carroll, A.L., Zhu, J., Raffa, K.F., Sickley, T.A., Taylor, S.W., 2006.Landscape level analysis of mountain pine beetle in British Columbia, Canada:spatiotemporal development and spatial synchrony within the presentoutbreak. Ecography 29, 427e441.
Baas, M., Briggs, D.E.G., Van Heemst, J.D.H., Kear, A.J., De Leeuw, J.W., 1995. Selectivepreservation of chitin during the decay of shrimp. Geochimica et CosmochimicaActa 59, 945e951.
Bamber, R.N., 1982. Sodium hexametaphosphate as an aid in benthic sample sort-ing. Marine Environmental Research 7, 251e255.
Bentz, B.J., Régnière, J., Fettig, C.J., Hansen, E.M., Hayes, J.L., Hicke, J.A., Kelsey, R.G.,Negrón, J.F., Seybold, S.J., 2010. Climate change and bark beetles of the westernUnited States and Canada: direct and indirect effects. BioScience 60, 602e613.
Bethlahmy, N., 1974. More streamflow after a bark beetle epidemic. Journal of Hy-drology 23, 185e189.
Bethlahmy, N., 1975. A Colorado episode: beetle, ghost forests and more streamflow.Northwest Science 49, 95e105.
Biederman, J.A., Brooks, P.D., Harpold, A.A., Gochis, D.J., Gutmann, E., Reed, D.E.,Pendall, E., Ewers, B.E., 2014. Multiscale observations of snow accumulation andpeak snowpack following widespread, insect-induced lodgepole pine mortality.Ecohydrology 7, 150e162.
Bigler, C., Kulakowski, D., Veblen, T.T., 2005. Multiple disturbance interactions anddrought influence fire severity in Rocky Mountain subalpine forests. Ecology 86,3018e3029.
Brunelle, A.R., Rehfeldt, J., Bentz, B.J., Munson, A.S., 2008. Holocene records ofMountain Pine Beetle infestation in the US Northern Rocky Mountains. ForestEcology and Management 255, 836e846.
Brunelle, A., Whitlock, C., Bartlein, P., Kipfmueller, K., 2005. Holocene fire andvegetation along environmental gradients in the Northern Rocky Mountains.Quaternary Science Reviews 24, 2281e2300.
Carter, V.A., 2010. A Paleoecological Fire and Vegetation History of SoutheasternWyoming (M.S. thesis). University of Utah, USA.
Carter, V.A., Brunelle, A.R., Minckley, T.A., Dennison, P.E., Power, M.J., 2013.Regionalization of fire regimes in the Central Rocky Mountains, USA. Quater-nary Research 80, 406e416.
Coope, G.R., 1959. A Late Pleistocene insect fauna from Chelford, Cheshire. Pro-ceedings of the Royal Society of London. Series B, Biological Sciences 151, 70e86.
Courtney Mustaphi, C.J., 2013. A Landscape-scale Assessment of Holocene FireRegime Controls in South-eastern British Columbia, Canada (PhD dissertation).Carleton University, Canada.
Courtney Mustaphi, C.J., Pisaric, M.F.J., 2013. Varying influence of climate and aspectas controls of montane forest fire regimes during the late Holocene, south-eastern British Columbia, Canada. Journal of Biogeography 40, 1983e1996.
Courtney Mustaphi, C.J., Pisaric, M.F.J., 2014. Holocene climate-fire-vegetation in-teractions at a subalpine watershed in southeastern British Columbia, Canada.Quaternary Research 81, 228e239.
Cudmore, T.J., Björklund, N., Carroll, A.L., Lindgren, B.S., 2010. Climate change andrange expansion of an aggressive bark beetle: evidence of higher reproductivesuccess in naïve host tree populations. Journal of Applied Ecology 47, 1036e1043.
Cullingham, C.I., Cooke, J.E., Dang, S., Davis, C.S., Cooke, B.J., Coltman, D.W., 2011.Mountain pine beetle host-range expansion threatens the boreal forest. Mo-lecular Ecology 20, 2157e2171.
Davis, M.B., 1981. Outbreaks of forest pathogens in Quaternary history. In:Bharadwaj, D.C., Vishnu-Mittre, Maheshwari, H.K. (Eds.), Fourth InternationalPalynological Conference Proceedings. Birbal Sahni Institute of Paleobotany,Lucknow, pp. 216e228.
de la Giroday, H.C., Carroll, A.L., Aukema, B.H., 2012. Breach of the northern RockyMountain geoclimatic barrier: initiation of range expansion by the mountainpine beetle. Journal of Biogeography 39, 1112e1123.
Dennison, P.E., Brunelle, A.R., Carter, V.A., 2010. Assessing canopy mortality during amountain pine beetle outbreak using GeoEye-1 high spatial resolution satellitedata. Remote Sensing of Environment 114, 2431e2435.
DeRose, R.J., Long, J.N., 2007. Disturbance, structure, and composition: spruce beetleand Engelmann spruce forests on the Markagunt Plateau, Utah. Forest Ecologyand Management 244, 16e23.
DeRose, R.J., Long, J.N., 2012. Factors influencing the spatial and temporal dynamicsof Engelmann spruce mortality during a spruce beetle outbreak on the Mar-kagunt Plateau, Utah. Forest Science 58, 1e14.
Dixon, H., 1935. Ecological studies on the high plateaus of Utah. Botanical Gazette97, 272e320.
Donato, D.C., Harvey, B.J., Romme, W.H., Simard, M., Turner, M.G., 2013. Bark beetleeffects on fuel profiles across a range of stand structures in Douglas-fir forests ofGreater Yellowstone. Ecological Applications 23, 3e20.
Dyer, E.D.A., 1969. Influence of temperature inversion on development of sprucebeetle Dendroctonus obesus (Mannerheim) (Coleoptera: Scolytidae). Journal ofthe Entomological Society of British Columbia 66, 41e45.
Dymerski, A.D., Anhold, J.A., Munson, A.S., 2001. Spruce beetle (Dendroctonus rufi-pennis) outbreak in Engelmann spruce (Picea engelmannii) in central Utah,1986e1998. Western North American Naturalist 61, 19e24.
Elias, S.A., 1985. Paleoenvironmental interpretations of Holocene insect fossil as-semblages from four high-altitude sites in the Front Range, Colorado, USA.Arctic and Alpine Research 17, 31e48.
Elias, S.A., 2001. Coleoptera and Trichoptera. In: Smol, J.P., Birks, H.J.B., Last, W.M.(Eds.), Tracking Environmental Change Using Lake Sediments, Zoological In-dicators, vol. 4. Kluwer Academic Publishers, pp. 67e80.
Elias, S., 2010. Advances in Quaternary Entomology. In: Developments in Quater-nary Science, vol. 12. Elsevier online. http://www.sciencedirect.com/science/bookseries/15710866/12.
Flintoft, L., 2004. Unlocking chitin degradation. Nature Reviews Microbiology 2, 90.Glew, J.R., Smol, J.P., Last, W.M., 2001. Sediment core collection and extrusion. In:
Last, W.M., Smol, J.P. (Eds.), Tracking Environmental Change Using Lake Sedi-ments, Basin Analysis, Coring, and Chronological Techniques, vol. 1. KluwerAcademic Publishers, Dordrecht, Netherlands, pp. 73e105.
J.L. Morris et al. / Quaternary International xxx (2014) 1e1514
Please cite this article in press as: Morris, J.L., et al., Do bark beetle remains in lake sediments correspond to severe outbreaks? A review ofpublished and ongoing research, Quaternary International (2014), http://dx.doi.org/10.1016/j.quaint.2014.03.022

Hansen, E.M., Bentz, B.J., Turner, D.L., 2001a. Physiological basis for flexible vol-tinism in the spruce beetle (Coleoptera: Scolytidae). Canadian Entomologist133, 805e817.
Hansen, E.M., Bentz, B.J., Turner, D.L., 2001b. Temperature-based model for pre-dicting univoltine brood proportions in spruce beetle (Coleoptera: Scolytidae).Canadian Entomologist 133, 827e841.
Hanson, W.D., Naughton, H.T., 2013. The effects of a spruce bark beetle outbreak andwildfires on property values in the wildlandeurban interface of south-centralAlaska, USA. Ecological Economics 96, 141e154.
Harvey, B.J., Donato, D.C., Romme, W.H., Turner, M.G., 2013. Influence of recent barkbeetle outbreak on fire severity and post-fire tree regeneration in montaneDouglas-fir forests. Ecology 94, 2475e2486.
Hebertson, E.G., Jenkins, M.J., 2008. Climate factors associated with historic sprucebeetle (Coleoptera: Curculionidae) outbreaks in Utah and Colorado. Environ-mental Entomology 37, 281e292.
Hicke, J.A., Allen, C.D., Desai, A.R., Dietze, M.C., Hall, R.J., Hogg, E.H., Kashian, D.M.,Moore, D., Raffa, K.F., Sturrock, R.N., Vogelmann, J., 2012a. Effects of biotic dis-turbances on forest carbon cycling in the United States and Canada. GlobalChange Biology 18, 7e34.
Hicke, J.A., Johnson, M.C., Hayes, J.L., Preistler, H.K., 2012b. Effects of bark beetle-caused tree mortality on wildfire. Forest Ecology and Management 271, 81e90.
Holsten, E.H., Thier, R.W., Munson, A.S., Gibson, K.E., 1999. The Spruce Beetle. USDAForest Service, Forest Insect and Disease Leaflet 127.
Jackson, S.T., 2012. Representation of flora and vegetation in Quaternary fossil as-semblages: known and unknown knowns and unknowns. Quaternary ScienceReviews 49, 1e14.
Jenkins, M.J., Hebertson, E.G., Page, W., Jorgensen, C.A., 2008. Bark beetles, fuels,fires, and implications for forest management in the Intermountain West.Forest Ecology and Management 254, 16e34.
Jenkins, M.J., Hebertson, E.G., Munson, A.S., 2014. Spruce beetle biology, ecology andmanagement in the Rocky Mountains: an addendum to spruce beetle in therockies. Forests 5, 21e71.
Jolly, W.M., Parsons, R.A., Hadlow, A.M., Cohn, G.M., McAllister, S.S., Popp, J.B.,Hubbard, R.M., Negrón, J.F., 2012. Relationships between moisture, chemistry,and ignition of Pinus contorta needles during the early stages of mountain pinebeetle attack. Forest Ecology and Management 269, 52e59.
Keeley, E.R., Grant, J.W.A., 2001. Prey size of salmonid fishes in streams, lakes, andoceans. Canadian Journal of Fisheries and Aquatic Sciences 58, 1122e1132.
Knapp, R.A., Matthews, K.R., Sarnelle, O., 2001. Resistance and resilience of alpinelake fauna to fish introductions. Ecological Monographs 71, 401e421.
Kulakowski, D., Veblen, T.T., 2006. The effect of fires on susceptibility of subalpineforests to a 19th century spruce beetle outbreak in western Colorado. CanadianJournal of Forest Research 36, 2974e2982.
Kurz, W.A., Dymond, C.C., Stinson, G., Rampley, G.J., Neilson, E.T., Carroll, A.L.,Ebata, T., Safranyik, L., 2008. Mountain pine beetle and forest carbon: feedbackto climate change. Nature 454, 987e990.
Lindgren, S., Lewis, K.J., 1997. Ecological Processes in the McGregor Model Forest:Interactions with Insects and Pathogens. McGregor Model Forest Association,Prince George, B.C.
Logan, J.A., Règnière, J., Powell, J.A., 2003. Assessing the impacts of global warmingon forest pest dynamics. Frontiers in Ecology and the Environment 1, 130e137.
Love, L.D., 1955. The effect on stream flow of the killing of spruce and pine by theEnglemann spruce beetle. Transactions of the American Geophysical Union 36,113e118.
Mielke, J.L., 1950. Rate of deterioration of beetle-killed Engelmann spruce. Journal ofForestry 48, 882e888.
Mikkelson, K.M., Bearup, L.A., Maxwell, R.M., Stednick, J.D., McCray, J.E., Sharp, J.O.,2013. Bark beetle infestation impacts on nutrient cycling, water quality andinterdependent hydrological effects. Biogeochemistry 15, 1e21.
Mitton, J.B., Ferrenberg, S.M., 2012. Mountain pine beetle develops an unprece-dented summer generation in response to climate warming. American Natu-ralist 179, 163e171.
Morris, J.L., Brunelle, A.R., Munson, A.S., 2010. Pollen evidence of historical forestdisturbance on the Wasatch Plateau, Utah. Western North American Naturalist70, 175e188.
Morris, J.L., Brunelle, A.R., 2012. Pollen records of historic spruce beetle (Den-droctonus rufipennis) disturbance from the subalpine ranges of southern Utah,USA. The Holocene 22, 961e974.
Morris, J.L., 2013. Using lake sediments to reconstruct bark beetle disturbances inwestern North America. Frontiers of Biogeography 5, 219e226.
Morris, J.L., le Roux, P.C., Macharia, A.N., Brunelle, A.R., Hebertson, E.G., Lundeen, Z.J.,2013a. Organic, elemental, and geochemical contributions to lake sedimentdeposits during severe spruce beetle (Dendroctonus rufipennis) disturbances.Forest Ecology and Management 289, 78e89.
Morris, J.L., Brunelle, A., DeRose, R.J., Seppӓ, H., Power, M.J., Carter, V., Bares, R.,2013b. Using fire regimes to delineate zones in a high-resolution lake sedimentrecord from the western United States. Quaternary Research 79, 24e36.
National Climate Data Center, 2010. Climate Division Data Base. National ClimaticData Center. Available at: http://www.ncdc.noaa.gov.
Nealis, V., Peter, B., 2008. Risk Assessment of the Threat of Mountain Pine Beetle toCanada’s Boreal and Eastern Pine Forests. Canadian Forest Service InformationReport BC-X-417. Natural Resources Canada, Canadian Forest Service, PacificForestry Centre, Victoria, BC.
Perkins, D.L., Swetnam, T.W., 1996. A dendroecological assessment of whitebarkpine in the Sawtooth-Salmon River region, Idaho. Canadian Journal of ForestResearch 26, 2123e2133.
Pfeifer, E.M., Hicke, J.A., Meddens, A.J., 2011. Observations and modeling of above-ground tree carbon stocks and fluxes following a bark beetle outbreak in thewestern United States. Global Change Biology 17, 339e350.
Raffa, K.F., Aukema, B.H., Bentz, B.J., Carroll, A.L., Hicke, J.A., Turner, M.G.,Romme, W.H., 2008. Cross-scale drivers of natural disturbances prone toanthropogenic amplification: the dynamics of bark beetle eruptions. BioScience58, 501e517.
Reid, R.W., 1963. Biology of the mountain pine beetle, Dendroctonus monticolaeHopkins, in the east Kootenay region of British Columbia. Interaction betweenthe beetle and its host, with emphasis on brood mortality and survival. Cana-dian Entomologist 95, 225e238.
Reid, R.W., Gates, H., 1970. Effect of temperature and resin on hatch of eggs of themountain pine beetle (Dendroctonus ponderosae). Canadian Entomologist 102,617e622.
Rhoades, C.C., McCutchan, J.H., Cooper, L.A., Clow, D., Detmer, T.M., Briggs, J.S.,Stednick, J.D., Veblen, T.T., Ertz, R.M., Likens, G.E., Lewis, W.M., 2013. Biogeo-chemistry of beetle-killed forests: explaining a weak nitrate response. Pro-ceedings of the National Academy of Sciences 110, 1756e1760.
Robertson, C., Nelson, T.A., Jelinski, D.E., Wulder, M.A., Boots, B., 2009. Spatialetemporal analysis of species range expansion: the case of the mountain pinebeetle, Dendroctonus ponderosae. Journal of Biogeography 36, 1446e1458.
Safranyik, L., Carroll, A.L., Régnière, J., Langor, D.W., Riel, W.G., Shore, T.L., Cooke, B.J.,Nealis, V.G., Taylor, S.W., 2010. Potential for range expansion of mountain pinebeetle into the boreal forest of North America. The Canadian Entomologist 142,415e442.
Schmid, J.M., Frye, J., 1977. Spruce Beetle in the Rockies. USDA Forest Service, RockyMountain and Range Experiment Station, General Technical Report RM-49.
Seddon, A.W.R., et al., 2014. Looking forward through the past: identification of 50priority research questions in palaeoecology. Journal of Ecology 102, 256e267.
Sherriff, R., Berg, E.E., Miller, A.E., 2011. Climate variability and spruce beetle(Dendroctonus rufipennis) outbreaks in south-central and southwest Alaska.Ecology 92, 1459e1470.
Simard, M., Romme, W.H., Griffin, J.M., Turner, M.G., 2011. Do mountain pine beetleoutbreaks change the probability of active crown fire in lodgepole pine forests?Ecological Monographs 81, 3e24.
Tank, S., Lesack, L., McQueen, D., 2009. Elevated pH regulates bacterial carboncycling in lakes with high photosynthetic activity. Ecology 90, 1910e1922.
Taylor, S.W., Carroll, A.L., 2004. Disturbance, forest age, and mountain pine beetleoutbreak dynamics in BC: a historical perspective. In: Shore, T.L., Brooks, J.E.,Stone, J.E. (Eds.), Mountain Pine Beetle Symposium: Challenges and Solutions.Report BC-X-399, Canadian Forest Service, Pacific Forestry Centre, Victoria, BC,pp. 67e94.
Thorne, G., 1935. Nemic parasites and associates of the mountain pine beetle(Dendroctonus monticolae) in Utah. Journal of Agricultural Research 51, 131e144.
United States Forest Service, 2011. http://www.fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5340736.pdf (accessed 13.08.13.).
Utah Division of Wildlife Resources, 2013. Annual Fish Stocking Reports. http://wildlife.utah.gov/dwr/fishing/stocking.html (accessed 10.08.13.).
Utah Department of Natural Resources, 2003. Forest Health in Utah. http://www.ffsl.utah.gov/foresthealth/fhgov4a.pdf (accessed 13.08.13.).
Vanderhoof, M., Williams, C.A., Ghimire, B., Rogan, J., 2013. Impact of mountain pinebeetle outbreaks on forest albedo and radiative forcing, as derived from Mod-erate Resolution Imaging Spectroradiometer, Rocky Mountains, USA. Journal ofGeophysical Research: Biogeosciences 118, 1461e1471.
Veblen, T.T., Hadley, K.S., Reid, M.S., Rebertus, A.J., 1991. The response of subalpineforests to spruce beetle outbreak in Colorado. Ecology 72, 213e231.
Veblen, T.T., Hadley, K.S., Nel, E.M., Kitzberger, T., Reid, M., Villalba, R., 1994.Disturbance regime and disturbance interactions in a Rocky Mountain subal-pine forest. Journal of Ecology 82, 125e135.
Verbruggen, F., Heiri, O., Reichart, G.J., Lotter, A.F., 2010. Chironomid d18O as a proxyfor past lake water d18O: a Lateglacial record from Rotsee (Switzerland). Qua-ternary Science Reviews 29, 2271e2279.
Waller, M., 2013. Drought, disease, defoliation and death: forest pathogens asagents of past vegetation change. Journal of Quaternary Science 28, 336e342.
Watt, J.H., 2008. A Late-Holocene Reconstruction of Spruce Beetle (Dendroctonusrufipennis) Disturbance from Antler Pond (M.S. thesis). Northern Arizona Uni-versity, USA.
Whitlock, C., Larsen, C.P.S., 2001. Charcoal as a fire proxy. In: Smol, J.P., Birks, H.J.B.(Eds.), Tracking Environmental Change Using Lake Sediments, Terrestrial, Algal,and Siliceous Indicators, vol. 3. Kluwer Academic Publishers, Dordrecht, TheNetherlands, pp. 75e97.
Wood, S.L., 1982. The bark and ambrosia beetles of North and Central America(Coleoptera: Scolytidae), a taxonomic monograph. Great Basin NaturalistsMemoirs, No. 13.
Wooller, M.J., Francis, D., Fogel, M.L., Miller, G.H., Walker, I.R., Wolfe, A.P., 2004.Quantitative paleotemperature estimates from d18O of chironomid head cap-sules preserved in arctic lake sediments. Journal of Paleolimnology 31, 267e274.
Wright Jr., H.E., 1991. Coring tips. Journal of Paleolimnology 6, 37e49.Xu, H., Paerl, H.W., Qin, B., Zhu, G., Gao, G., 2010. Nitrogen and phosphorus inputs
control phytoplankton growth in eutrophic Lake Taihu, China. Limnology andOceanography 55, 420e432.
J.L. Morris et al. / Quaternary International xxx (2014) 1e15 15
Please cite this article in press as: Morris, J.L., et al., Do bark beetle remains in lake sediments correspond to severe outbreaks? A review ofpublished and ongoing research, Quaternary International (2014), http://dx.doi.org/10.1016/j.quaint.2014.03.022