increases in concentrations of potassium and bicarbonate ions promote acquisition of motility in...
TRANSCRIPT

THE JOURNAL OF EXPERIMENTAL ZOOLOGY 277:171–180 (1997)
© 1997 WILEY-LISS, INC.
JEZ 730
Increases in Concentrations of Potassium andBicarbonate Ions Promote Acquisition of Motility InVitro by Japanese Eel Spermatozoa
HIROMI OHTA,1* KAZUO IKEDA,1 AND TOSHIO IZAWA2
1National Research Institute of Aquaculture, Nansei, Mie 516-01, Japan2Hokkaido Fish Hatchery, Eniwa, Hokkaido 061-14, Japan
ABSTRACT We examined the effects in vitro of the ionic composition and pH of the mediumon the acquisition of motility by testicular spermatozoa of the Japanese eel, Anguilla japonica. Weobtained milt from the genital pore and testicular milt from the testes of artificially maturedcultivated eels. About 45% of milt spermatozoa were motile when they were diluted withhyperosmotic solution. By contract, 1.4% of testicular spermatozoa were motile. We produced anartificial seminal plasma (ASP), which corresponded in terms of ionic constituents to the seminalplasma obtained from the milt, and incubated immotile testicular spermatozoa with the ASP. Thespermatozoa showed increased motility (44.0–60.7%) after a 60-min incubation at all pH valuesexamined (pH 7.5–8.7). Absence of bivalent cations from the ASP did not impede the acquisition ofmotility at pH 8.1. However, testicular spermatozoa incubated with potassium-free ASP (pH 7.8–8.4) did not become motile, and the motility increased in proportion to the concentrations of potas-sium ions (0–30 mM) in ASP at pH 8.1. When testicular spermatozoa were incubated with HCO3
–-freeASP at pH 8.1 (the ASP including 15.2 mM KCI), spermatozoa did not become motile. Motility of thespermatozoa increased in proportion to the concentrations of HCO3
– ions in ASP. Thus, acquisition ofmotility required potassium and bicarbonate ions and their concentrations regulated the percentageof motile spermatozoa. The pH of testicular milt (pH7.5) was significantly lower than that of milt (pH8.0), and the concentration of potassium ions in testicular plasma (44.3 mM) was significantly higherthan that in seminal plasma (15.2 mM). Thus, it appears that immotile testicular spermatozoa be-come motile upon transfer from the testicular lumen to the sperm duct, with increases in pH and inthe concentration of bicarbonate ions. J. Exp. Zool. 277:171–180, 1997. © 1997 Wiley-Liss, Inc.
Contract Grant sponsor: Ministry of Agriculture, Forestry and Fish-eries, Japan; Contract Grant number BMP 96-II-2-11.
*Correspondence to: Dr. Hiromi Ohta, National Research Instituteof Aquaculture, Nansei, Mie 516-01, Japan.
Received 28 July 1996; Revision accepted 13 September 1996
Fish spermatozoa are immotile in the testis andalso, in many species, in the seminal plasma(Stoss,’83; Morisawa, ’85). Motility is initiatedwhen they are spawned into the environmentalwater. Morisawa and Suzuki (’80) divided the fac-tors that initiate the motility of teleost spermato-zoa into those that increase (marine teleosts) ordecrease (fresh water teleosts) osmolality, andthose that change the concentration of ions (salmo-nids and viviparous teleost). After spermiogenesis,teleost spermatozoa acquire motility, as a finalstep in their maturation, in the testis (Bedford,’79) or in the sperm duct (Morisawa and Mori-sawa, ’86).
In salmonids, spermatozoa acquire motility dur-ing passage from the testes along the sperm duct(Morisawa and Morisawa, ’86) and high concen-trations of bicarbonate ions and a higher pH (7.8–8.15) are important factors that promote theacquisition of motility (Morisawa and Morisawa,’88). Injections of 17,20β-dihydroxy-4-pregnen-3-one, a maturation-inducing hormone in salmonids
(Nagahama, ’87), into male masu salmon (Miuraet al., ’92) and Japanese eel (Miura et al., ’91)accelerate the acquisition of sperm motility. Inboth cases, acquisition of motility is accompaniedby increases in the pH in the sperm duct. Theseresults strongly suggest that increases of environ-mental pH are essential for acquisition of motil-ity by spermatozoa.
Lahnsteiner et al. (’96) reported that motilityof milt spermatozoa of Alburnus alburnus (Cyp-rinidae) was closely related to the concentrationsof Na+ or K+ ions, in addition to the pH of theseminal plasma. Redond-Müller et al. (’91) con-firmed that carp spermatozoa acquire motilityupon incubation in isotonic solutions with highconcentrations of KCl or NaCI, and this process

172 H. OHTA ET AL.
is not dependent of pH. These reports suggest thatthe ionic constituents of testicular fluid might beinvolved in the acquisition of motility of teleosteansperm, in addition to pH.
Although cultivated Japanese eels, Anguilasjaponica, are sexually immature, repeated injec-tions of gonadotropin can readily induce matura-tion and spermiation (Yamamoto et al., ’72; Ohtaet al., ’96a), and the testicular spermatozoa areimmotile when they are diluted with hyperosmoticsolution (Miura et al., ’95). Therefore, the eel iswell suited for studies of the mechanism that regu-lates the acquisition of sperm motility. In pre-liminary experiments, we found that immotiletesticular spermatozoa of the eel acquired motil-ity within 3 hours when incubated with seminalplasma obtained from milt that contained motilespermatozoa. This observation suggests that theseminal plasma provides the factors necessary forthe acquisition of sperm motility. In the presentstudy, we examined the effects in vitro of the ioniccomposition and pH of seminal plasma on the ac-quisition of motility by immotile testicular sper-matozoa of the eel.
MATERIALS AND METHODSFish and hormonal treatment
Cultivated male eels (body weight, 278–325 g)that had been reared under freshwater conditionswere purchased from a commercial supplier inMarch 1994. They were gradually acclimated toseawater over the course of 4 days. During ex-periments, they were held without feeding underthe natural photoperiod at 20±1°C, in 1,000-literflow-through seawater aquariums.
Intraperitoneal injections of 300 IU of humanchorionic gonadotropin (hCG; Gonatropin, TeikokuZoki Co. Ltd., Tokyo) per fish were given once aweek to induce maturation. After the fourth to sixthinjection, milt could be obtained by striping fromall the males. Within 7 days after the eighth weeklyinjection, treated fish were weighed and killed.
Collection of milt and testicular miltMilt was obtained by the application of gentle
pressure of the abdomen. The area around thegenital pore was rinsed with distilled water anddried with a soft towel. Care was taken to avoidcontamination of the milt with seawater or urine.Milt was collected with a Pasteur pipette andstored in a micro-tube on crushed ice prior to mea-surements of sperm motility and the spermatocrit(see below for definition).
The paired testes of the Japanese eel are com-posed of numerous seminiferous lamellae sus-pended from the dorsal peritoneal wall (Yamamotoet al., ’72). These lamellae are arranged along thewall, and a sperm duct is located at the roots ofthe lamellae. After the collection of milt, fish weresacrificed and the lower halves of the lamellaewere cut and minced into small pieces with scis-sors. The mixture was then filtered through a ny-lon mesh (mesh size, 250 µm) to separate thetesticular milt from the testicular fragments.
Analysis of pH, spermatocrit, andconcentrations of ions
The pH of the milt and of the testicular miltwas measured with a pH meter (PHI 100; Beck-man Instruments, Inc., Porterville). One drop ofmilt was placed on a probe (Smart ISFET Probe;Beckman Instruments, Inc. The measurement ofthe pH of the milt was completed within 1 min ofthe collection of the milt from the genital pore.For the measurement of the pH of testicular milt,a small incision was made on the surface of thetesticular lamellae and the probe was inserted intothe incision.
The spermatocrit [(packed cell volume/total miltvolume) × 100] was determined after centrifuga-tion of the milt or the testicular milt in a hemat-ocrit tube (12,000 rpm for 15 min).
For analysis of ionic concentrations, milt or tes-ticular milt was centrifuged at 3,500g for 10 minand the resulting surpernatant was then recen-trifuged at 9,500g for 10 min. This supernatantwas designated the seminal plasma (milt) or thetesticular plasma (testicular milt). Concentrationsof ions were determined with an atomic absorp-tion spectrophotometer (Z-6000 or Z-6100; Hitachi,Tokyo).
Incubation of spermatozoaSpermatozoa of the Japanese eel are immotile
in isotonic solutions, and the motility is initiatedwhen osmolality is increased (Ohta and Izawa,’96). Before the incubation tests, 2 µl of testicularmilt were mixed with 2,000 µl of 450 mM NaCl(buffered with 20 mM HEPES NaOH at pH 7.5)in a 10-ml polyethylene tube on a vortex mixer.Twenty microliters of the mixture was immedi-ately transferred onto a glass slide, and the per-cent of motile sperm was measured as an initialcontrol value.
Five microliters of testicular milt was incubatedin 995 µl of various isotonic media at 20°C in a2.2-ml cylindrical micro-tube (8.0 mm in diameter;

ACQUISITION OF MOTILITY BY EEL SPERMATOZOA 173
Multi-safe seal tubes; Sorenson Bioscience, Inc.,Salt Lake City, UT) and each tube was placed ina tube-incubator (CHT100; Iwaki Glass Co., Ltd,Funabashi, Japan) at 20 ± 0.1°C for 60 min. Theincubation media was based on the ionic compo-sition of seminal plasma obtained from artificiallymatured male eels; 149.3 mM NaCl, 15.2 mM KCl,1.3 mM CaCl2, 1.6 mM MgCl2, 20 mM NaHCO3plus 20 mM HEPES (pH 7.5–7.9)-NaOH or 20 mMTAPS-NaOH (pH 8.1–8.7). Some modifications ofthe constituents were made in each case (see Re-sults). The dilution ratio for incubation (200-fold)was chosen to minimize the changes in incuba-tion conditions upon addition of testicular milt.Fifteen, 30, and 60 min after the start of incuba-tion, the diluted suspension of testicular milt wasgently mixed on a vortex mixer, and 20 µl of themixture was taken from the tube and further di-luted with 80 µl of 450 mM NaCl (final dilution,1,000-fold) in a polyethylene tube. Twenty micro-liters of the mixture was immediately transferredonto a glass slide and the percent motility ofsperm was determined.
Before and after the incubation, the pH of eachmedium was measured. All changes in pH duringthe incubations were within 0.13 pH units of therespective pre-incubation value.
Measurement of sperm motilitySperm motility (percent motility) was observed
at room temperature (approximately 20°C) witha videotape recorder through a video camera thatwas connected to a light microscope (×200). Sper-matozoa were considered to have acquired motilitywhen the sperm head showed forward movement15 sec after dilution with 450 mM NaCl (bufferedwith 20 mM HEPES-NaOH at pH 7.5). Percent-age motility was determined by assessing the mo-tility of at least 50 randomly chosen spermatozoafor each measurement. Measurements for each di-lution were repeated twice, and averages wereused in the analysis of data.
StatisticsAll data are represented as means ± SEM. Means
were examined by one-way analysis of variance andBartlett’s test for uniformity of variances. When dif-ferences in variance were significant, data were sub-jected to the Cochran-Cox test in order to detectsignificant differences. Other data were subjectedto Student’s unpaired t-test. The calculations wereperformed with the computer programs of Waka-bayashi (’84). A value of P<0.05 was considered torepresent a significant difference.
RESULTSComparison between milt and
testicular miltSperm motility, spermatocrit, and the pH of milt
and of testicular milt from nine artificially ma-tured males are shown in Table 1. The ionic con-stituents of seminal plasma and testicular plasmaare also presented in Table 1. The percentage ofmotile milt spermatozoa obtained from the geni-tal pore was 45.8 ± 6.2%, while testicular sper-matozoa were almost immotile (1.4 ± 1.1%). Thespermatocrit of the milt was significantly lower(P < 0.001) than that of the testicular milt. ThepH of the milt was higher than that of testicularmilt (P < 0.01). Concentrations of Na+ and Ca2+
ions in the seminal plasma were higher than thosein the testicular plasma; by contrast, the concen-tration of K+ ions of the former was lower thanthat in the latter (P < 0.001, respectively). In par-ticular, concentrations of Na+ and K+ ions werevery different in seminal plasma and testicularplasma.
Acquisition of motility by testicularspermatozoa in vitro
Testicular spermatozoa were incubated with ar-tificial seminal plasma (ASP) that correspondedin terms of ionic constituents to the seminalplasma and changes in motility with increasedduration of incubation were monitored (Fig.1).Since the pH of the milt from artificially maturedJapanese eels ranged from 7.5 to 8.7 and the meanpH was 8.1 ± 0.1 (n = 117; Ohta et al., unpub-lished data), we prepared five samples of ASP withdifferent pH values, from pH 7.5 to 8.7, and ex-amined the effects of the pH of the ASP on theacquisition of sperm motility (Fig. 1). Initial mo-tility of the testicular milt used was 0% (n = 5).
TABLE 1. Comparison between milt and testicular miltfor the Japanese eel (n = 9)
Milt Testicular milt
Sperm motility (%) 45.8 ± 6.2 1.4 ± 1.1*Spermatocrit (%) 71.5 ± 2.0 91.8 ± 1.1*pH 8.0 ± 0.1 7.5 ± 0.1**Concentrations of ions in theseminal plasma and testicularplasma (mM)
Na+ 149.3 ± 3.2 118.2 ± 6.2*K+ 15.2 ± 0.8 44.3 ± 2.6*Ca2+ 1.3 ± 0.1 0.6 ± 0.1*Mg2+ 1.6 ± 0.3 2.2 ± 0.2
*Indicates a significant difference (P < 0.001) to the value for the milt.**Indicates a significant difference (P < 0.01) to the value for milt.

174 H. OHTA ET AL.
After a 15-min incubation, the spermatozoa thathad been diluted in all ASPs became motile. Inparticular, spermatozoa at pH 8.4 and 8.7 showedthe highest motility (35.1 ± 7.9% and 54.6 ± 3.3%,respectively). Finally, the percentages of motilespermatozoa in all ASPs tested were high (from44.0 ± 4.6% at pH 7.8 to 60.7 ± 4.7% at pH 8.7)after a 60-min incubation. Thus, immotile testicu-lar spermatozoa acquired motility within 60 minwhen incubated in ASP from pH 7.5 to 8.7. InASPs at lower pH more time was needed for theacquisition of sperm motility.
The result in Figure 1 suggest that the constitu-ents of the ASP promote the acquisition of motil-ity by testicular spermatozoa. Therefore, weexamined the effects of various cations and an-ions in the ASP at pH 8.1, namely, K+ Ca2+, Mg2+,and/or HCO3
– ions, on the acquisition of sperm mo-tility (Fig. 2). Spermatozoa incubated in ASP orin Ca2+- or Mg2+-free ASP acquired motility, andno significant differences in percent motility were
found between them. By contrast, most sperma-tozoa incubated in K+-free or HCO3-free ASP failedto become motile within 60 min.
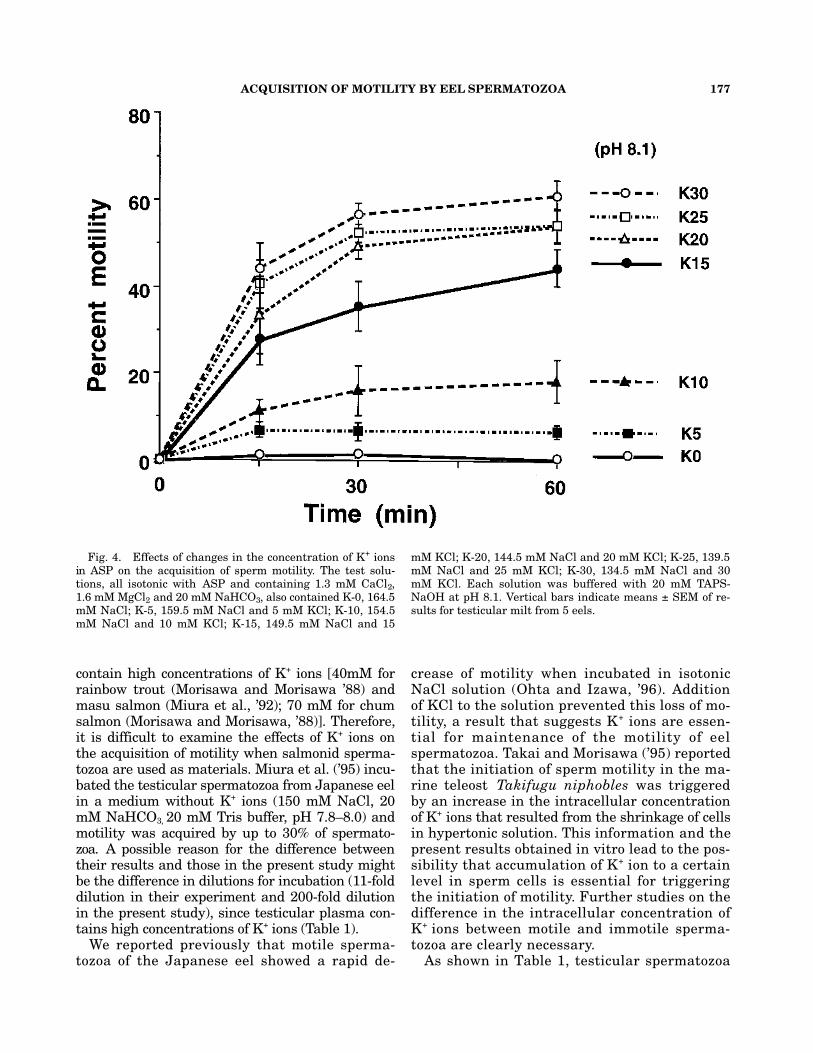
To determine whether K+ ions are necessary forthe acquisition of motility, irrespective of pH, tes-ticular spermatozoa were incubated in K+-freeASP at different pH values (Fig. 3). Spermatozoaincubated in K+-free ASP failed to acquire motil-ity within 60 min at all tested pH values. We thenexamined the effects of changes in the concen-tration of K+ ions in ASP at pH 8.1 on the acqui-sition of motility. As shown in Figure 4, thepercent motility increased as the concentrationsof K+ ions increased. The percent motility of sper-matozoa incubated in ASP with more than 15 mMK+ ions was significantly higher than that in ASPwith lower potassium concentrations of K+ ionsafter a 15-min incubation.
When testicular spermatozoa were incubated inHCO3
–-free ASP at various pH values, motility de-veloped in ASP at pH values above 8.25, and the
Fig. 1. Effects of the pH of isotonic medium (artificial semi-nal plasma, ASP) on the acquisition of sperm motility. TheASP contained: 149.3 mM NaCl, 15.2 mM KCl, 1.3 mM CaCl21.6 mM MgCl2 and 20 mM NaHCO3. Each solution was buff-
ered with 20 mM HEPES-NaOH (pH 7.5–7.8) or TAPS-NaOH(Ph 8.1–8.7). Motiltiy was examined 15 sec after dilution with450 mM NaCl. Vertical bars indicate means ± SEM of resultsfrom 5 eels.

ACQUISITION OF MOTILITY BY EEL SPERMATOZOA 175
percent motility increased with increases in pH(Fig.5). After a 60-min incubation, spermatozoaincubated in HCO3
–-free ASP at pH 8.7 or 8.4yielded significantly higher values of percent mo-tility than those at pH values below 8.25. We ex-amined the effects of the concentration of HCO3
–
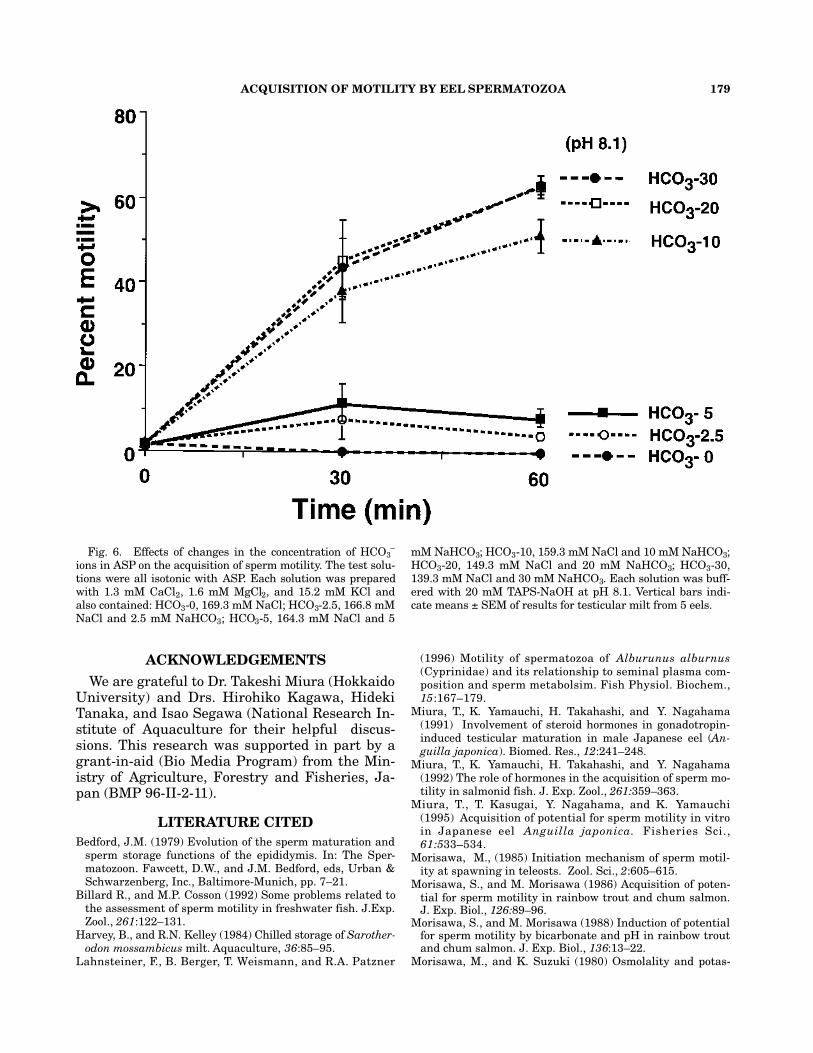
ions in ASP at pH 8.1. The percent motility of sper-matozoa that had been incubated for 30 and 60min increased in proportion to the concentrationof HCO3-ions in ASP (Fig.6).
DISCUSSIONOur study demonstrates that acquisition of mo-
tility be eel spermatozoa requires K+ and HCO3–
ions or K+ ions and a high pH. Increases in per-cent motility were proportional to the concentra-tions of both K+ and HCO3 ions in the medium.
Morisawa and Morisawa (’86) reported thatspermatozoa of chum salmon acquired the capac-ity for movement during their passage from thetestis along the sperm duct. This process shouldreflect the increase in pH that results from theincrease in the seminal concentration of HCO3
–
ions in the duct (Morisawa and Morisawa,’88). Ac-quisition of sperm motility in response to an in-crease in pH was also confirmed by studies in vitroin chum salmon, rainbow trout (Morisawa andMorisawa, ’88), and masu salmon (Miura et
Fig. 2. Effects of cations or anions in the ASP on the ac-quisition of sperm motility. The four test solutions usedwere as follows: (1) ASP containing 149.3 mM NaCl, 15.2 mMKCl, 1.3 mM CaCl2, 1.6 mM MgCl2, and 20 mM NaHCO3; (2)K+-free ASP containing 164.5 mM NaCl (equal in osmolalityto ASP with 149.3 mM NaCl and 15.2 mM KCl), 1.3 mMCaCl2, 1.6 mM MgCl2, and 20 mM NaHCO3; (3) Ca2+- andMg2+-free ASP containing 153.7 mM NaCl (equal in osmolal-
ity to ASP with 149.3 mM NaCl, 1.3 mM CaCl2, and 1.6 mMMgCl2), 2.0 mM EDTA, 15.2 mM KCl and 20 mM NaHCO3;(4) HCO3
–-free ASP containing 169.3 mM NaCl (equal in os-molality to ASP with 149.3 mM NaCl and 20 mM NaHCO3),15.2 mM KCl, 1.3 mM CaCl2, and 1.6 mM MgCl2. Eachsolution was buffered with 20 mM TAPS-NaOH at pH 8.1.Vertical bars indicate means ± SEM of results for testicu-lar milt from 5 eels.

176 H. OHTA ET AL.
al.,’92). Miura et al. (’92) and Morisawa et al. (’93)found that acquisition of motility by salmonidspermatozoa was accompanied by increases in in-tracellular levels of cAMP. Morisawa et al. (’93)suggested that elevated external pH stimulatesthe membranous adenyl cyclase that synthesizescAMP and this stimulation might be a factor thatinduces the acquisition of sperm motility in salmo-nid fishes. By contrast, Redond-Müller et al. (’91)reported that milt spermatozoa from carp, with alow potential for movement, acquired motility dur-ing incubation in K+-rich or Na+-rich solutions.Moreover, the activation by K+ ions was not pH-dependent between pH 6.04 and 9.03. In thepresent study, testicular immotile spermatozoaacquired motility within 60 min, irrespective ofthe pH between pH 7.5 and 8.7 in medium thatcontained 15.2 mM KCl and 20 mM NaHCO3 al-though the time required for acquisition of motil-ity was shorter at higher pH. At pH 8.1 theaverage pH of the milt, spermatozoa failed to ac-
quire motiltiy in HCO3–-free medium and the ac-
quisition of motility occurred in direct proportionto the amount of HCO3
– ions added to the medium.Only at very high pH (above pH 8.4) did they ac-quire motility without addition of HCO3
– ions to themedium. In other words, an increase in the concen-tration of HCO3
– ions an essential factor for the ac-quisition of motility under physiological conditions.
The present study demonstrated that K+ ions arealso essential for acquisition of sperm motility byJapanese eel spermatozoa, and the effect was con-centration-dependent in medium that included 20mM NaHCO3 at pH 8.1. To date, mechanisms ofthe acquisition of sperm motility in teleosts havebeen investigated exclusively in salmonids, and theimportance of an increase in pH has been empha-sized (Morisawa and Morisawa, ’88; Miura et al.,’92). Since salmonid spermatozoa begin to move inK+-free medium (for review see Scott and Baynes,’80, Morisawa, ’85) as soon as they acquire motility,incubation media for acquisition of motility must
Fig. 3. Effects of the pH of K+-free ASP on the acquisi-tion of sperm motility. K+-free ASP was prepared with 164.5mM NaCl, 1.3 mM CaCl2, 1.6 mM MgCl2, 20 mM NaHCO3
and 20 mM HEPES-NaOH (pH 7.8) or 20 mM TAPS-NaOH(pH 8.1 and 8.4). Vertical bars indicate means ± SEM of re-sults for testicular milt from 5 eels.
ACQUISITION OF MOTILITY BY EEL SPERMATOZOA 177
contain high concentrations of K+ ions [40mM forrainbow trout (Morisawa and Morisawa ’88) andmasu salmon (Miura et al., ’92); 70 mM for chumsalmon (Morisawa and Morisawa, ’88)]. Therefore,it is difficult to examine the effects of K+ ions onthe acquisition of motility when salmonid sperma-tozoa are used as materials. Miura et al. (’95) incu-bated the testicular spermatozoa from Japanese eelin a medium without K+ ions (150 mM NaCl, 20mM NaHCO3, 20 mM Tris buffer, pH 7.8–8.0) andmotility was acquired by up to 30% of spermato-zoa. A possible reason for the difference betweentheir results and those in the present study mightbe the difference in dilutions for incubation (11-folddilution in their experiment and 200-fold dilutionin the present study), since testicular plasma con-tains high concentrations of K+ ions (Table 1).
We reported previously that motile sperma-tozoa of the Japanese eel showed a rapid de-
crease of motility when incubated in isotonicNaCl solution (Ohta and Izawa, ’96). Additionof KCl to the solution prevented this loss of mo-tility, a result that suggests K+ ions are essen-tial for maintenance of the motility of eelspermatozoa. Takai and Morisawa (’95) reportedthat the initiation of sperm motility in the ma-rine teleost Takifugu niphobles was triggeredby an increase in the intracellular concentrationof K+ ions that resulted from the shrinkage of cellsin hypertonic solution. This information and thepresent results obtained in vitro lead to the pos-sibility that accumulation of K+ ion to a certainlevel in sperm cells is essential for triggeringthe initiation of motility. Further studies on thedifference in the intracellular concentration ofK+ ions between motile and immotile sperma-tozoa are clearly necessary.
As shown in Table 1, testicular spermatozoa
Fig. 4. Effects of changes in the concentration of K+ ionsin ASP on the acquisition of sperm motility. The test solu-tions, all isotonic with ASP and containing 1.3 mM CaCl2,1.6 mM MgCl2 and 20 mM NaHCO3, also contained K-0, 164.5mM NaCl; K-5, 159.5 mM NaCl and 5 mM KCl; K-10, 154.5mM NaCl and 10 mM KCl; K-15, 149.5 mM NaCl and 15
mM KCl; K-20, 144.5 mM NaCl and 20 mM KCl; K-25, 139.5mM NaCl and 25 mM KCl; K-30, 134.5 mM NaCl and 30mM KCl. Each solution was buffered with 20 mM TAPS-NaOH at pH 8.1. Vertical bars indicate means ± SEM of re-sults for testicular milt from 5 eels.

178 H. OHTA ET AL.
Fig. 5. Effects of pH of HCO3–-free ASP on the acquisi-
tion of sperm motility. HCO3–-free ASP contained 169.3 mM
NaCl, 15.2 mM KCl, 1.3 mM CaCl2, 1.6 mM MgCl2, and 20
mM HEPES-NaOH (pH 7.8) or 20 mM TAPS-NaOH (pH 8.1–8.7). Vertical bars indicate means ± SEM of results for tes-ticular milt from 5 eels.
are maintained in an aqueous enviornment ata low pH (pH 7.5) and a high concentration ofK+ ions (44.3 mM). They experience an increasein pH (to pH 8.0) and a decrease in the concen-tration of K+ ions (to 15.2 mM) as they movefrom the seminal lobules to the sperm duct. Invivo 45.8% of spermatozoa acquired the poten-tial for motility after such a change. At pH 8.1,testicular spermatozoa did not become motilein HCO3
–-free ASP. The results shown in Fig-ure 1 indicate that immotile spermatozoa canacquire motility at pH values above 7.5 whensufficient HCO3
–-ions are suplied in addition to15.2 mM KCl. At pH 8.1, motility is acquiredin media that include more than 10 mM HCO3
–-ions (Fig.6). Thus, a major cause for the absenceof motility in the testicular spermatozoa mightbe the low concentration of HCO3
–-ions in thetesticular plasma. We did not measure levelsof HCO3
–-ions in the testicular and seminalplasma in this study. Morisawa and Mroisawa(’88) compared concentrations of total inorganic
carbon in testicular plasma and seminal plasmain rainbow trout and chum salmon. The con-centrations were 4.9 and 9.2 mM in rainbowtrout and 14.7 and 23.5 mM in chum salmon,respectively, suggesting an increase in the con-centration of bicarbonate ions from the testicu-lar lumen to the sperm duct in these salmonids.
A relationship between motility and capacityfor the fertilization of teleost spermatozoa hasbeen confirmed by several authors (Harvey andKelley, ’84; Billard and Cosson, ’92; Ohta et al.,’95). Our recent preliminary studies haveshown that testicular spermatozoa, which adbeen rendered motile by incubation in artifi-cial seminal plasma that included 30 mM KCland 20 mM NaHCO3, were as capable of fer-tilization as milt spermatozoa (Ohta et al., un-published data). Techniques for obtaining largequantities of motile spermatozoa from eels bythis method should compensate for the short-age of milt at the time of artificial fertiliza-tion (Ohta et al., ’96b).
ACQUISITION OF MOTILITY BY EEL SPERMATOZOA 179
ACKNOWLEDGEMENTSWe are grateful to Dr. Takeshi Miura (Hokkaido
University) and Drs. Hirohiko Kagawa, HidekiTanaka, and Isao Segawa (National Research In-stitute of Aquaculture for their helpful discus-sions. This research was supported in part by agrant-in-aid (Bio Media Program) from the Min-istry of Agriculture, Forestry and Fisheries, Ja-pan (BMP 96-II-2-11).
LITERATURE CITEDBedford, J.M. (1979) Evolution of the sperm maturation and
sperm storage functions of the epididymis. In: The Sper-matozoon. Fawcett, D.W., and J.M. Bedford, eds, Urban &Schwarzenberg, Inc., Baltimore-Munich, pp. 7–21.
Billard R., and M.P. Cosson (1992) Some problems related tothe assessment of sperm motility in freshwater fish. J.Exp.Zool., 261:122–131.
Harvey, B., and R.N. Kelley (1984) Chilled storage of Sarother-odon mossambicus milt. Aquaculture, 36:85–95.
Lahnsteiner, F., B. Berger, T. Weismann, and R.A. Patzner
(1996) Motility of spermatozoa of Alburunus alburnus(Cyprinidae) and its relationship to seminal plasma com-position and sperm metabolsim. Fish Physiol. Biochem.,15:167–179.
Miura, T., K. Yamauchi, H. Takahashi, and Y. Nagahama(1991) Involvement of steroid hormones in gonadotropin-induced testicular maturation in male Japanese eel (An-guilla japonica). Biomed. Res., 12:241–248.
Miura, T., K. Yamauchi, H. Takahashi, and Y. Nagahama(1992) The role of hormones in the acquisition of sperm mo-tility in salmonid fish. J. Exp. Zool., 261:359–363.
Miura, T., T. Kasugai, Y. Nagahama, and K. Yamauchi(1995) Acquisition of potential for sperm motility in vitroin Japanese eel Anguilla japonica. Fisheries Sci.,61:533–534.
Morisawa, M., (1985) Initiation mechanism of sperm motil-ity at spawning in teleosts. Zool. Sci., 2:605–615.
Morisawa, S., and M. Morisawa (1986) Acquisition of poten-tial for sperm motility in rainbow trout and chum salmon.J. Exp. Biol., 126:89–96.
Morisawa, S., and M. Morisawa (1988) Induction of potentialfor sperm motility by bicarbonate and pH in rainbow troutand chum salmon. J. Exp. Biol., 136:13–22.
Morisawa, M., and K. Suzuki (1980) Osmolality and potas-
Fig. 6. Effects of changes in the concentration of HCO3–
ions in ASP on the acquisition of sperm motility. The test solu-tions were all isotonic with ASP. Each solution was preparedwith 1.3 mM CaCl2, 1.6 mM MgCl2, and 15.2 mM KCl andalso contained: HCO3-0, 169.3 mM NaCl; HCO3-2.5, 166.8 mMNaCl and 2.5 mM NaHCO3; HCO3-5, 164.3 mM NaCl and 5
mM NaHCO3; HCO3-10, 159.3 mM NaCl and 10 mM NaHCO3;HCO3-20, 149.3 mM NaCl and 20 mM NaHCO3; HCO3-30,139.3 mM NaCl and 30 mM NaHCO3. Each solution was buff-ered with 20 mM TAPS-NaOH at pH 8.1. Vertical bars indi-cate means ± SEM of results for testicular milt from 5 eels.

180 H. OHTA ET AL.
sium ion: Their roles in initiation of sperm motility in te-leosts. Science, 210:1145–1147.
Morisawa, S., K. Ishida, M.Okuno, and M. Morisawa (1993)Roles of pH and cyclic adenosine monophosphate in the ac-quisition of potential for sperm motility during migrationfrom the sea to the river in chum salmon. Mol. Repord.Dev., 34:420–426.
Nagahama, Y. (1987) 17α,20β-dihydroxy-4-pregnen-3-one: Ateleost maturation-inducing hormone. Dev. Growth Differ.,29:1–12.
Ohta, H., and T. Izawa (1996) Diluent for cool storage of theJapanese eel (Anguilla japonica) spermatozoa. Aquaculture,142:107–118.
Ohta, H., H. Shimma, and K. Hirose (1995) Relationship be-tween fertility and motility of cryopreserved spermatozoaof the amago salmon Oncorhynchus masou ishikawe. Fish-eries Sci., 61:886-887.
Ohta, H., H. Kagawa, H. Tanaka, K. Okuzawa, and K. Hirose(1996a) Milt production in the Japanese eel Anguillajaponica induced by repeated injections of human chorionicgonadotropin. Fisheries Sci., 62:44–49.
Ohta, H., H. Kagawa, H. Tanaka, K. Okuzawa, N. Iinuma,
and K. Hirose (1996b) Artificial induction of maturation andfertilization in the Japanese eel, Anguilla japonica. FishPhysiol. Biochem. (in press).
Redond-Müller, C., M.P. Cosson, J. Cosson, and R. Billard(1991) In vitro maturation of the potential for movement ofcarp spermatozoa. Mol. Reprod. Dev., 29:259–270.
Scott, A.P., and S.M. Baynes (1980) A review of the biology,handling and storage of salmonid spermatozoa. J. Fish Biol.,17:707–739.
Stoss, J. (1983) Fish gamete preservation and spermatozoanphysiology. In: Fish Physiology, Vol., 9, Part B W.S. Hoar etal., eds. Academic Press, New York, pp. 305–350.
Takai, H., and M. Morisawa (1995) Change in intracellularK+ concentration caused by external osmolality change regu-lates sperm motility of marine and freshwater teleosts. J.Cell Sci., 108:1175–1181.
Wakabayashi, K. (1984) Arrangement of results. Life SciencePersocom Series 5. Baifukan, Tokyo (in Japanese).
Yamamoto, K., O. Hiroi, T. Hirano, and T. Morioka (1972)Artificial maturation of cultivated male Japanese eelsby synahorin injection. Nippon Suisan Gakkaishi 38:1083–1090.