cryopreservation of equine and porcine spermatozoa …
TRANSCRIPT

CRYOPRESERVATION OF EQUINE AND PORCINE SPERMATOZOA
WITH A UNIQUE FREEZING TECHNOLOGY (UFT)
by
HEATHER A. VARTORELLA, B.S., M.S.
A DISSERTATION
IN
ANIMAL SCIENCE
Submitted to Uie Graduate Faculty
of Texas Tech University in Partial Fulfilhnent of the Requirement
for the Degree of
DOCTOR OF PHILOSOPHY
Approved
Chairperson of the Committee
Accepted
Dean of the Graduate School
May, 2003

2003, Heather Ann Vartorella

ACKNOWLEDGEMENTS
From an academic standpoint, this dissertation represents an organized, written
collection of research and knowledge. In the real world though, this dissertation
represents a conglomeration of late nights, experiences, relationships, deadlines,
education, and emotions. By no means was this research project purely a feat of my own.
With that in mind, I would like to acknowledge a few people who added to the following
pages.
First off, I would like to acknowledge the people in my life that are close to my
heart. To my family in Michigan and my friends, in Texas and abroad, I am always
gratefiil for their continuous love and support. A special thanks to my future husband,
J.W. Goolsby, for helping me keep my sanity during my various academic challenges.
In regards to the actual production of this research project, there are several
people I need to mention. In appreciation of their collaboration with Texas Tech
University, I would like to recognize Brian Wood and Supachill USA, Inc., for
supporting this project. I also need to acknowledge all the staff at the TTUHSC
OB/GYN Laboratory, especially Susan Chavez and Roland Pirtle, thank-you for your
encouragement and help. 1 am also very grateftti for the support 1 received from Pete
Cotter, since two sets of hands are definitely quicker than just one. Also 1 need to thank
the management and staff at the TTU Ranch Horse Center and TTU Swine Unit for the
extra time taken to facilitate this project. To my committee. Dr. Kevin Pond, Dr. Sam

Jackson, Dr. Gail Comwall, and Dr. John Blanton; I would like to thank-you for your
time and supervision.
Finally, I need to convey my admiration and gratitude for Dr. Sam Prien, my
committee chair, mentor, and above all else, my friend. Without your guidance, my
graduate career would have not been possible.
i l l

TABLE OF CONTENTS
ACKNOWLEDGEMENTS Ji
ABSTRACT vh
LIST OF TABLES ix
LIST OF FIGURES x
CHAPTER
I. n^TRODUCTION 1
n. LITERATURE REVIEW 4
Introduction 4
Current Status of Equine Cryopreservation 5
Current Status of Porcine Cryopreservation 6
Cellular Effects of Cryopreservation 7
Diluents 9
Thermal Gradients 13
Pre-Freezing Protocols 15
Packaging Systems for Cryopreservation 18
Summary 20
111. EFFECTS OF SEMEN HANDLING ON POST-THAW MOTILITY
OUTCOMES OF CRYOPRESERVED EQUINE SPERMATOZOA 22
Abstract 22
Introduction 23
Materials and Methods 24
iv

Results 28
Discussion 28
IV. COMPARISONS OF A UNIQUE FREEZING TECHNOLOGY (UFT) TO TRADITIONAL LIQUID NITROGEN METHODOLOGY FOR CRYOPRESERVATION OF EQUINE SPERMATOZOA 31
Abstract 31
Infroduction 32
Materials and Methods 34
Results 37
Discussion 42
V. EFFECTS OF SEMEN HANDLING ON POST-THAW MOTILITY
OUTCOMES OF CRYOPRESERVED PORCINE SPERMATOZOA 45
Abstract 45
Introduction 46
Materials and Methods 47
Results 51
Discussion 52 VI. COMPARISONS OF A UNIQUE FREEZING TECHNOLOGY (UFT)
TO TRADITIONAL LIQUID NITROGEN METHODOLOGY FOR CRYOPRESERVATION OF PORCINE SPERMATOZOA 55
Abstract 55
Introduction 56
Materials and Methods 57
Results 60
Discussion 64

VII. LIPID PEROXIDATION EFFECTS FROM SEMEN HANDLING
DURUSIG CRYOPRESERVATION OF PORCINE SPERMATOZOA 66
Abstract 66
Infroduction 67
Materials and Methods 68
Results 73
Discussion 80 Vin. CONCLUSIONS 83
LITERATURE CITED 86
VI

ABSTRACT
The cryopreservation of sperm offers several benefits to various biological
sciences including clinical medicine, biotechnology and agriculture. This conservation
process requires a reduction in cellular metabolism while maintaining viability and
stractural stability through freezing and eventually thawing thus allowing for an
extension of viability past normal limitations. However, the application of sperm
cryopreservation has not been successfiil in all species. For example in the livestock
sector, advancements in semen technology have been made in the dairy industry, but not
as significantiy within the equine and swine areas.
Recently, a new Unique Freezing Technology (UFT) was developed to freeze
foodstuffs at a quick rate while maintaining freshness. This dissertation represents the
initial series of trials utilizing this new technology on cellular specimens not of meat
origin. From August 2001 to March 2003, numerous ejaculates from five different
Quarter Horse stallions and seven boars were collected. From these samples, over 1000
treatments were created to test various cryopreservation procedures and techniques
including extenders, freezing rates, thawing rates, male influence, and storage containers.
The design of this study was to improve post-thaw motility rates of stallion and boar
spermatozoa.
Several factors were found to affect the post-thaw motility outcome. In the
stallion trials significant factors included preparative handling, stallion influence and
freezing technique. The results of the boar trials also revealed the significant influences
vu

of the pre-freezing protocols, boar influence and freezing techniques. Further testing was
conducted on the boar sperm samples to evaluate the stability of the phospholipid
membrane post-thaw. Large variations were noted in the amounts of lipid free radicals
and lipid peroxidation from the fresh samples compared to those found from the post-
thaw samples. This idea was ftirther demonstrated by an analysis of the progressive
sfress accumulated during pre-freezing preparation.
The unique freezing technology could be potentially considered to be used for
spermatozoa cryopreservation for swine. However, ftirther studies need to be
implemented to standardize techniques and find alterative extenders.
Vlll

LIST OF TABLES
3.1 Comparison of Post-Thaw Motilities from Four Pre-Freezing Protocols 28
4.1 Post-Collection Evaluation Values of Stallion Semen 41
4.2 Comparison of Initial, Post-Thaw and Retum Motilities from Four StaUions 41
4.3 Egg Yolk and Glycerol Composition of Biladyl AB and Test Yolk Buffer Extenders 43
5.1 Comparison of Pre-Freezing Procedural Trials on Boar Spermatozoa 52
6.1 Post-Thaw Motility Outcomes from Cryopreserved Boar Semen 64
7.1 Averaged Lipid Peroxidation and Free Radical Amounts from Three Freezing Treatments in Experiment #1 74
7.2 Averaged Lipid Peroxidation Amounts from Fresh and Extended Boar Samples in Experiment #2 74
7.3 Averaged Lipid Peroxidation Amounts from Procedural Steps 76
7.4 Averaged Lipid Free Radical Amounts from Procedural Steps 78
7.5 Motility and Lipid Stability Trends from the Samples of Experiment #3 80
IX

LIST OF FIGURES
4.1 Comparison of Post-lliaw Motilities from Stallion Semen Frozen with Various Freezing Treatments 37
4.2 Comparison of Post-Thaw Motilities of Semen from Acceptable Breeding StaUions Frozen with Various Freezing Treatments 38
4.3 Comparison of Human and Bovine Freezing Extenders on the Post-Thaw Motility of Frozen Stallion Spermatozoa 39
4.4 Comparison of Freezing Techniques used with the Human Extender on
the Post-Thaw Motility of Frozen Stallion Spermatozoa in Experiment #2 40
6.1 Cooling Rates of the UFT 61
6.2 Post-Thaw Motility Percentages of Cryopreserved Boar Semen 62
6.3 Original Motility Percentage Retums of Cryopreserved Boar Semen 63
7.1 Procedural Lipid Peroxidation Trends 77
7.2 Procedural Lipid Free Radical Trends 79

CHAPTER I
INTRODUCTION
Mammalian mating requires a series of specific events with various sequences
and timing ratios to lead to a successfiil fertilization. In simple terms, fertilization is
commonly referred to as an interaction between an ova and a sperm. However, in the
complicated world we live in, it is not that simple, especially in the area of timing. In
response to infertility issues and genetic preservation where timing is the key, assisted
reproductive techniques (ARTs) have been developed and advanced. Though several
techniques have been utilized, including; in vitro fertilization (FVF) and intracytoplasmic
sperm injection ICSI, no other ART offers the availability for sperm conservation other
than cryopreservation. Cryopreservation of sperm requires a reduction in cellular
metabolism while maintaining viability and stractural stability through freezing and
eventually thawing. Depending on the mammalian species, sperm can live up to 120
hours (Hafez and Hafez, 2000) once ejaculated. Holding a sample in a frozen state until
needed allows the useftil lifespan of a sample to be extended up to 50 years (Yoshida,
2000). This process would allow genetic preservation in a variety of scenarios including:
intemational exchange, availability of laboratory evaluation, conservation of neiir-
extinction, transgenic or superior genetic lines, or supply in response to illness, death or
unavailability of the male. This conservation would benefit various biological sciences
such as clinical medicine, biotechnology and agriculture.

The use of assisted reproductive techniques (ARTs) has become increasingly
widespread in the livestock industry. The acceptability of artificial insemination, embryo
fransfer, and cryopreservation of sperm has lead to the establishment of breeding
organizations, semen banks, ART courses, and sire stations. This is especially tine in the
dairy cattle industry, where semen technological advancements have been the most
successful (Vishwanath and Shannon, 2000) and conception rates from frozen semen are
comparable to natural mating (Watson, 2000). However, the resulting quality of
cryopreservation in many species is not remotely similar to that of the initial specimen
collected. In the swine industry, the cryopreservation of bozir semen has repeatedly
produced low pregnancy rates and reduced litter size (Yoshida, 2000). The estimated
average farrowing rate from frozen sperm was 50% with seven piglets to a litter (Johnson
et al., 2000). The acceptability of this technique is newer to the equine industry,
especially within two of the major American breed organizations. Yet a survey of 1999
and 2000 revealed that commercial program conception rates from frozen semen
averaged was only 51.3% during first estras cycle attempts emd 74.7% for the season
(Loomis, 2001). The different success rates between species are not understood, even
though variations in cold shock sensitivity and the molecular composition differences
among sperm types have been confirmed (Parks and Lynch, 1992).
Since the first endeavors of freezing in 1776 by Spallanzani, the techniques
involving cryopreservation of sperm have improved. Although the expansion within the
past few decades has been progressively slow (Yoshida, 2000). Several factors involved
with the handling process during the pre-freeze, freezing and thawing phases have been

suggested as the cause. Nevertheless, the key to answer has not been found and
conceptive rates from spermatozoa following cryopreservation rates remain less than
optimal. Currently, it is estimated that 40-50% of a semen sample will be lost during
cryopreservation, even when following what are considered to be optimum protocols
(Watson, 2000). Thus, previous investigations of cryopreservation concerning the
reductions of fertility have focused on sperm viability and stractural integrity and suggest
significant room for improvements. The objective of the present study was to improve
post-thaw freezing rates while evaluating the effects of thermal gradients, packaging
systems, male specimen variation, membrane stability, and cryoprotectants on equine and
porcine spermatozoa.

CHAPTER II
LITERATURE REVIEW
Introduction
The preservation of sperm through cryopreservation offers a varied of advantages
to the biological sciences, yet the full potential has yet to be realized. Current estimations
project that 40-50% of a semen sample will be lost during cryopreservation even with
optimum protocols (Watson, 2000). Therefore, it is well understood that as the
temperature declines, there may be a reduction in the functionality, biochemical
comp>osition and stracture of the sperm cell. Though a various specie cell lines seem
more susceptible to cold shock, such as the pig, the mechanisms are not totally
understood (Holt, 2000; Johnson et al., 2000). It is assumed cryostress is due to the
variations of composition and the varying functions of homologous component parts
could be the cause for species-specific cryopreservation differences (Calvete et al., 1997).
Affecting factors can be separated into two groups, external and internal. The
external group refers to the alterations of stress due to preparative handling including
diluents, thermal gradient changes during freezing and thawing, cryoprotectants, and
holding periods. The internal factors of cryopreservation are involved with the
biochemistry and composition of the sperm cell itself; including protein stability,
membrane integrity, and oxidative damage. The following is a literature review to briefly
summarize and address these topics.

It should be noted that there is a lack of standard handling and freezing
techniques. Several of the reports reviewed use different practices and various markers
as a means for measuring success (Day, 2000; Love et al., 1988). Due to the absence of
standardized procedures, the findings of some published reports may not be presented as
completely as intended (Samper et al., 2002).
Current Status of Equine Cryopreservation
With the acceptance of cooled, shipped semen, artificial insemination (Al) has
become routine practice in most commercial equine operations (Sieme et al., 2002). The
utilization of frozen semen however has not progressed as quickly (Squires et al., 2002).
This is due in part to breed registry regulations and limitations. Though the European
based breed organizations such as the Warmblood and Trakehner have embraced this
technology, the American associations have recently amended their breeding rales. For
example in 2001, the American Quarter Horse Association and American Paint Horse
Association, two of the world's largest breed associations, permitted the artificial
insemination of frozen-thawed semen (Loomis, 2001). Yet with this new opportunity and
its benefits of genetic transport and decreased venereal disease risk, horse breeders have
not fully adopted this technique. Reports of lower pregnancies rates per cycle have been
commonly observed compared to natural mating or cooled semen (Samper et al., 2002).
Theses reported outcomes makes the utilization of this reproductive technique less
desirable to producers. It is estimated that only 24% of stallions are able to produce an
ejaculate suitable for cryopreservation (Linfor et al., 2002; Vidament, et al., 1997). In a

recent worid-wide survey of commercial breeding centers, Samper and colleagues (2002)
found pregnancy rates from frozen semen ranged from 12.5-60%, with an average of
45.9%. Loomis (2001) reported that pregnancy rate averages are 81.9% outside North
America and only 65.6% within with frozen semen.
Though offering long-term benefits, the possible financial losses from frozen
semen without a fertilization guarantee are a concern. Frozen semen also requfres a
stringent adherence to mare management and specialized personnel fraining. Samper et
al. (2002) noted breeding centers that bred more than 15 mares had a 10% higher
pregnancy rate than those that bred less. However, errors in semen processing,
equipment, and management have also been reported as a cause of reduced fertility rates
(Loomis, 2001). To improve fertility chances, increased inseminations have
demonstrated higher pregnancy rates. In spite of this, stallion owners usually limit frozen
availability since increased use reduces semen reserves (Squires et al., 2002). Regardless
of cause, the equine industry does not currently have a standardized cryopreservation
technique. This variability makes improvements, trends and comparisons hard to
investigate at this time (Samper et al., 2002). Furthermore in the United States, there is a
lack of a unified breeding certification program for facilities, technicians and sire registry
as seen in other countries (Loomis, 2001).
Current Status of Porcine Cryopreservation
In the swine industry, the use of artificial insemination (Al) of sows with
preserved semen has tripled within the past 15 years. It is estimated that more than 99%

of the inseminations performed worldwide are performed with extended liquid state
semen. Semen cryopreservation has been commercially available since 1975. In the last
15 years, the utilization of frozen semen in Europe has significantly increased (Johnson et
al., 2000). Yet with the increased utilization, the fertility performance of cryopreserved
samples is not efficient as emphasized by low litter rates (Yoshida, 2000). The estimated
average farrowing rate from frozen semen is 50% with seven piglets to a litter (Johnson
et. al., 2000).
Porcine semen has been recognized for its sensitivity to cold shock, though the
mechanisms are not totally understood (Holt, 2000; Johnson et al., 2000; Yoshida, 2000).
Even with successful motility levels, fertilization and embryonic development are not
guaranteed. As a litter bearing animal, the sow presents a unique challenge to the ART
through prolonged ovulations requiring sperm viability to encompass 40 hours. Though
fresh semen is able to maintain this longevity, frozen sperm by this point commonly
show a rapid decline (Watson, 2000). Thus, advances in the understanding of the porcine
physiology and development of reproductive techniques are paramount (Yoshida, 2000).
Cellular Effects of Cryopreservation
Mammalian spermatozoa are very temperature sensitive. This sensitivity, known
as cold shock, refers to the irreversible alterations commonly observed during
cryopreservation (Medeiros et al., 2002). Rapid thermal stress on spermatozoa can cause
abnormal swimming pattems, loss of motility, acrosomal and/or plasma membrane
damage, altered metabolism, and infracellular component loss (Amann and Graham,

1993). Once this damage has occurred, it cannot not be overcome since sperm lose the
ability to self repair, grow and biosynthesis after spermatogenesis (Yoshida, 2000). Plus,
this damage can occur in any region of the sperm either independently or collectively
(Zhu and Liu, 2000).
During the cooling process, the plasma membrane will enter a transition phase
changing from a selectively permeable fluid state to one with increased rigidity by
changing the arrangement of the membrane constituents. This will limit the lateral
movement of phospholipids and cause the proteins to become irreversibly clustered.
Functionality of protein-lipid complexes forming membrane receptors and ion pores may
be lost (Amann and Graham, 1993). As the fransition continues, unstable gel-fluid
portions form to create ion gaps. These conformational changes to the inner membrane
can lead to an increased permeability of exfracellular calcium (McLaughlin and Ford,
1994) possibly triggering calcium-dependent processes associated with capacitation
(Buhr et al., 1989; Green and Watson, 2001; Johnson et al., 2000; Watson, 2000). If tiie
membrane is leaking, ATP levels may decrease from the mid-piece signaling cryolysis
(Bosman et al., 1994). While the solutes and cryoprotectants increase outside the sperm,
phospholipid damage increases, protein-lipid complexes weaken and the sperm
dehydrates (Gao et al., 1993). If ice crystal formation occurs, the membrane will most
likely rapture. As the phase fransition completes, protein function may cease due to
denaturation (Amann and Graham, 1993). Thus, protein and DNA conformations may be
altered in the head portion.

Lipid peroxidation, which leads to lipid radical formation, is a normal mechanism
in cellular aerobic metabolism. Reactive oxygen species (ROS) including the superoxide
anion (O2") and hydrogen peroxide (H2O2) are involved in capacitation of the sperm
(Medeiros et al., 2002). However at toxic levels, peroxidation can have degradative
effects on the phospholipid membrane of spermatozoa. To briefly review, during the
oxidation of various metals including iron, an electron is transferred to oxygen (Huang et
al., 2001). This single electron on the oxygen produces a negative charge and unstable
molecule, free oxygen radical or anion. With addition of superoxide dismutase and
hydrogen molecules, the oxygen radicals can convert to peroxide. During the reduction
of iron, peroxide can fransform in to radical hydroxyls (OH). In the presence of
hydroxyl radicals, the stable polyunsaturated fatty acids (LH) of the membrane will lose
its hydrogen to become a lipid radical (L). In an aerobic environment, the lipid radicals
will begin to accelerate a cascade of lipid degradation. To reduce the accumulation of
peroxide, cells possesses catalase and gluthathione, to convert peroxide to water.
However, mammalian sperm have veuious, reduced levels of protective enzymes (Alvarez
and Storey, 1984; Bilodeau et al., 2000; Mennella and Jones, 1980).
With the possibility of any one of these events occurring, the ability of
cryopreserved sperm to sustain embryonic development is questionable (Watson, 2000).
Diluents
In an attempt to create the ideal environment for sperm viability, several studies
have been conducted to determine the effects of the diluents used in cryopreservation.

Diluents for cryopreservation are very similar to those used for storage at a liquid state.
The basic requirements for a cryopreservation solution must include an energy source,
cryoprotectant, osmolarity buffers, cold shock preventative substances, enzymes and
antibiotics (Vishwanath and Shannon, 2000).
When released from the male reproductive tract, sperm are submerged in a natural
buffering nutrient source known as seminal plasma. The presence of seminal fluid in the
female reproductive tract plays a role in the fertilization process both directly and
indirectly. As the sperm swims to its destination, the seminal plasma is an aid as a
nutrition source, buffering agent, and motility regulation while stimulating enhancement
of uterine contractions and the relaxation of the oviductal opening (Johnson et al., 2000).
Seminal plasma also down-regulates the uterine inflammation response to the presence of
sperm (Troedsson et al., 2002). Commonly seminal plasma is removed to increase
insemination rates and increase fertility of frozen-thawed sperm. However, some recent
studies have shown the possible positive influences of seminal fluid used as a diluent.
Using this concept, studies have demonstrated improved motility rates (Katila et al.,
2002; Pickett et al., 1975) and promotion of sperm viability normally inhibited by uterine
inflammation (Troedsson et al., 2002). Catalase activity, an enzymatic converter of
hydrogen peroxide, has been observed in bovine seminal fluid (Bilodeau et al., 2000,
LaPointe et al., 1998). It does need to be recognized that there are contradicting papers.
It has been suggested that the ferrous iron (Fe ^ ) within seminal plasma, is the driving
factor for malondialdehyde (MDA) production during incubation (Huang et al., 2001).
Verma and Kanwar (1999) confirmed increases in lipid peroxidation rates from samples
10

incubated with seminal plasma. Though Braemmer et al. (2002) found seminal plasma
had no effect on the post-thaw motilities of epididymal sperm. Love et al. (2002)
discovered detrimental motility and chromatin condensing effects.
At certain stages of the cryopreservation process, sperm cells may undergo
fluctuating environments due to the amounts of solutes inside or outside the cell. This
process is referred to as osmotic stress. During the freezing stage, the cells enter into a
hyperosmotic environment. As exfracellular ice crystals form, the solutes outside of the
cell increase in concenfration. In response, the sperm cell will dehydrate and shrink in
volume by losing infracellular water in attempt to equilibrate to the outer environment. A
hyperosmotic environment can also induce a reduction in motility while increasing
tyrosine phosphorylation, a step in the capacitation process (Linfor et al., 2002). During
the thawing stage, the sperm cell enters into a hypotonic envfronment where water will
passively enter into cell. This increase in volume can cause the cell to swell. Osmotic
sfress thus can cause irreversible damage, rendering the sperm cell incapable of
fertilization (Curry and Watson, 1994; Pommer et al., 2002).
More than one-half century ago, egg yolk was discovered to possess protective
attributes when added to spermatozoa during cryopreservation (Medeiros et al., 2002). It
has been proposed that the success of egg yolk extenders is due to the low density
lipoproteins that are found in two-thirds of the composition (Moussa et al., 2002).
Although egg yolk has been a successfiil addition to bull freezing extender (Vishwanath
and Shannon, 2000), the results have not been as successful for boars (Pursel et al, 1972).

Egg yolk extenders are still more commonly utilized, yet milk components have also
been developed into extenders (Vishwanath and Shannon, 2000).
To attempt to further reduce ice formation and osmotic situations, cryoprotectants
are usually added to freezing extenders. There are two types of cryoprotective agents,
permeable (i.e., glycerol, dimethyl sulfoxide, 1,2-propanediol) or non-permeable (i.e.
sucrose, sodium chloride, raffinose) (Liu and Foote, 1998; Sztein et al., 2001). In most
mammalian cryopreservation processes, glycerol is considered the standard
cryoprotectant (Silva et al., 2003; Watson, 2000). Glycerol is a permeating agent that
enters into the cell to replace the water that leaves during cooling, thus reducing the
osmotic sfress (Medeiros et al., 2002). The levels of glycerol added are dependant on the
cooling velocity. At each glycerol level, sperm can tolerate a variety of temperature
ranges without viability losses (Johnson et al., 2000). Glycerol levels are also dependant
on specie type. Glycerol is usually added at concentrations of 2.5 to 6% for stallions
(Samper and Morris, 1998; Vidament et al., 2002), 2.25 to 9% for bulls (Vishwanatii and
Shannon, 2000), 5 to 6% for goat bucks (Amoah and Gelaye, 1996), and 13-16% in
roosters (Gill et al., 1996).
However, permeating cryoprotective agents, such as glycerol, can be toxic to the
cell and cause membrane instability and motility loss (Medeiros et al., 2002). The toxic
effects are due to high glycerol concenfrations or too rapid of an addition and removal
from the cell (Katkov et al., 1998). Also, it has been observed that higher glycerol
concentrations cause an increase in acrosomal damage (Almid and Johnson, 1988).
Several articles have tested other cryoprotective agents in various species. A few of the
12

otiier cryoprotective agents evaluated include dimethyl sulfoxide (Amoah and Gelaye,
1996; Ball and Vo, 2001; Gilmore et al., 1998, 1995; Johnson et al., 2000; Sztein et al.,
2001), ethyl glycol (Ball and Vo, 2001; Gilmore et al., 1998, 1995; Henry et al., 2002),
and various amides (Blanco et al., 2000; Henry et al., 2002; Johnson et al., 2000;
Medeiros et al., 2002; Sztein et al., 2001; Vidament et al., 2002). Results vary
depending on the species, concentrations levels, and utilization of the agents in
combination or alone.
Thermal Gradients
The process of cryopreservation involves a range of temperatures during the
freezing and thawing process. A sperm cell is ejaculated close to body temperature,
cooled to below freezing, and returned to body temperature during thawing. The stress
upon the spermatozoa during these thermal insults can create irreversible damage to the
ftmctionality of the cell. In general terms, this damage is referred to as cold shock.
Though spermatozoa from any mammalian species can demonsfrate cold shock, the
thermal sensitivity or resistance varies between species (Medeiros et al., 2001). This is
due to the different of ultrastractural changes that occur and composition differences
between sperm (Curry and Watson, 1994; DeLeeuw et al., 1990; Parks and Lynch, 1992).
During the cooling process, several stractural and metabolic alterations occur
within the sperm cell at various temperatures. As the temperature nears 20-25°C, the
membrane fluidity will decrease in the samples from bulls, stallions, boars and roosters
(Parks and Lynch, 1992). This alteration is due to the stiffening of the hydrocarbon tails
13

of lipid within the bilipid layer to shift the membrane into a gel-like solid. Lowering the
temperature further from 22°C to 4°C, oxygen free radical amounts increase in bull sperm
(Chatterjee and Gagnon, 2001). This indicates an increase in lipid instability through
lipid peroxidation rates. Once the temperature falls to 4°C, there is also depression in
metabolic cellular activity (Medeiros et al., 2002) including the ftmctionality of sodium-
potassium (Na/K) ion pumps (Vishwanath and Shannon, 2000). Green and Watson
(2001) observed capacitation-like events in boar sperm cooled to 5*'C. This capacitation-
like event is related to flux of calcium, involved in hyperosmotic environments (Watson,
2000). At the freezing point, the latent heat radiated from the sperm cell during ice
formation must also be considered (Thurston et al., 2003). Past the freezing point, ice
formation occurs at about -10°C, depending on the cryoprotectant (Medefros et al., 2002).
Freezing rates must be fast enough to reduce infracellular ice formation, yet slow
enough to avoid osmotic alterations. As discussed in a previous section, glycerol is
commonly used as a cryoprotectant. However, there is a relation between the cooling
velocity and glycerol concentrations. The faster the cooling rate, the less glycerol needed
(Johnson et al., 2000).
Mammalian sperm are frozen at range of a rate of-15°C to -60°C/minute
(Watson, 2000). Various studies have reported a wide range of freezing rates. For
example, sperm viabilities were maintained in equine sperm at rates of-20°C to
-130°C/minute (Devireddy et al., 2002) and in swine sperm at -3°C to -80°C/minute
(Medrano et al., 2002). Recommended optimal freezing rates for each species are more
limited. Suggested rates include -5°C/minute in rams (Byme et al., 2000), -30°C/minute
14

in boars (Fiser et al., 1993; Johnson et al., 2000), and between -80°C and-120°C/minute in
bulls (Vishwanath and Shannon, 2000). Though the equine cryopreservation protocols
vary, most use liquid nitrogen vapor rate of-5°C/minute (Samper and Morris, 1998).
Stractural integrity due to thermal stress is not limited to the freezing rate but also
the thawing rate. Osmotic stresses have been recognized during the re-warming process
(Holt and North, 1994; Pommer et al., 2002; Watson, 2000). It has been suggested that
more damage occurs in hyposmotic environment than in a hyperosmotic environment.
These frends have been observed in the sperm of stallion (Ball and Vo, 2001; Pommer et
al., 2002), human male (Curry and Watson, 1994; Gao et al., 1993), boar (Gihnore et al.,
1996; Medrano et al., 2002; Zeng et al., 2001), mouse (Willoughby et al., 1996) and ram
(Curry and Watson, 1994; Holt and North, 1994). An exception though would be the bull
(DeJamettee et al., 2000). Regardless of these reports, the relation between freezing and
thawing rates must be taken in consideration (Henry et al., 1993). Post-thaw rates of
motility and acrosomal stability from fast freezing rates, 30°C/minute, were superior to
those from slow-freeze, l°C/minute, after a rapid thawing process (Fiser et al., 1993). In
general, faster rates of thawing are considered to be more beneficial. At slower rate of
thaw, the risk of ice reformation is more likely (Vishwanath and Shannon, 2000).
Pre-Freezing Protocols
Artificial insemination with the use of liquid-state semen has become common
procedure in several breeding facilities. Developments in this area of reproduction have
extended the life of semen from a few hours to several days. Yet, a few days is still
15

limiting in comparison to the potential genetic preservation gained through
cryopreservation. In attempt to refine cryopreservation protocols, these developments
have been integrated into the preparative handling techniques being utilized today.
Seminal plasma can only sustain sperm viability for a few hours after ejaculation
(Johnson et al., 2000). Boar semen incubated at 37°C in seminal plasma developed high
rates of peroxidative degradation (Comaschi et al., 1989). Though keeping a portion of
the seminal fluid amount may have beneficial properties, as discussed earlier, it is
common to remove the fluid through centrifugation. Samper and Morris (1998) found
that all horse breeders surveyed centrifuged samples after collection. Centrifugation
creates a more concenttated specimen while allowing for the limiting nutrient source to
be replaced. Removal of all seminal plasma also created a higher rate of chromatin
stability (Love et al., 2002) and higher post-thaw motilities (Martin et al., 1979) in
stallion samples. Nevertheless it has been suggested that centrifugation may cause undue
sfress to sperm (Gil et al., 1999). With dog semen, a higher amount of sperm was lost in
the discarded supemant at lower centrifiigal speeds. At faster centrifiigation speeds,
viability losses were higher (Rijsselaere et al., 2002). Neild and colleagues (2003) also
reported a capacitation-like phenomenon after centrifugation of stallion sperm. As a
compromise, Johnson and colleagues (2000) suggested the use of one of the most utilized
protocols for boar cryopreservation in a commercial setting, the Beltsville method. Initial
cooling involves incubating a sample in seminal fluid for 2 hours before centrifugation as
part of the protocol.
16

It has been suggested that post-thaw success may be achieved through a series of
osmotic steps (Curry and Watson, 1994; Gao et al., 1993). There is evidence of this idea
through the increases in cold shock resistance observed in sperm after incubation or
holding periods at cooler temperatures. Epididymal spermatozoa of the stallion held at
5°C for 24 hours demonstrated similar post-thaw motilities to those from samples
processed immediately (Braemmer et al., 2002). Similar trends in ejaculated sperm were
observed after 12 hours of incubation at 5°C (Crockett et al., 2001). Boar sperm
appeared to be more cold resistance after a 5 hour incubation period versus two or seven
hour periods (Pursel et al., 1972). More recentiy, Eriksson et al. (2001) demonstrated
higher retum rates with incubation periods of 10 and 20 hours for boar semen cooled to
17°C.
To create an optimal cryopreservation protocol, a cooling rate compatible with
osmotic environment must be considered (Watson, 2000). Glycerolized diluents have
reported to have varying success rates based on time to equilibration. Initially it was
thought that extended semen exposed to glycerol did not show any survival differences
post-thaw based on exposure time. However, a trial utilizing 0.5 mL straws demonsfrated
that an incubation period of 4 hours produced higher motilities and improved morphology
(Johnson et al., 2000). Temperature in which the glycerol is added also affects the post-
thaw outcomes. Vidament and colleagues (2000) found the addition of glycerol at 22°C
generated a higher retum rate than glycerol added at 4°C.
17

Packatzing Systems for Cryopreservation
In agriculture, frozen sperm offers an alterative solution to the labor and expense
of shipping live animals with the intent of spreading superior genetics. This advantage
not only allows for extension of travel distances and availability of semen supply, but
also helps ensure the health of a herd involved with breeding practices. As stated
previously, maximum fertility levels have not been reached for several species with
regard to cryopreservation. Aside from the challenges of optimum temperature gradients
and diluents, there is also a quest for the perfect containment system. Various types of
packaging systems have utilized in attempt to overcome the problems of identification,
contamination, thermal gradients and insemination amounts available associated with
some packaging units.
During the initial onset of cryopreservation use in the livestock industry, pelleting
semen on dry ice was a technique initially used in bulls (Martin et al., 1979; Merkt et al.,
1975). It has been discovered however that this methodology has several disadvantages.
Concems with identification, contamination and exchange of cellular material have been
recognized (Pickett et al., 1978). Straws with various volumes have also been utilized.
Though the 0.25 mL and 0.15 mL straws do not have a large volume capacity, there is a
higher rate of heat transfer allowing for a more uniformity in the freeze. From a
management standjwint, the 5 mL offers more desirable volume due to the required
eunount needed for an insemination dose.
Several studies have evaluated the efficiency of each package. Though the
seminal processing was not performed consistently, the results still show variation in
18

between species. Using bull (Johnson et al., 1995) and rooster (Duplaix and Sexton,
1983) semen, there was little difference in the post-thaw results between the 0.25 mL and
0.5 mL straws. However, Thomas and colleagues (1993) found higher rates of
progressive motile sperm post-thaw in canine semen with pellets when compared to
either of these sfraws. In lieu of the reduced capacity available from pellets and straws,
larger volume packages have also been investigated. Increased fertility rates were
reported in stallion semen was using 4 mL straws (Martin et al., 1979) and also in boars
semen using 5 mL straws (Cordova et al., 2002). Cordova and colleagues also observed a
fiigher rate of chromatin condensation with an increased stability in the larger straws.
Yet, aluminum tubes with larger volumes showed potential in stallion semen (Tischner,
1979) and later outperformed the enhanced straws with stallion sperm (Love et al., 1989)
and pelleting with canine semen (Ivanova-Kicheva et al., 1997). In the swine industry, 5
mL FlatPacks have shown potential in both the laboratory (Eriksson et al., 2001) and in
field trials (Eriksson et al., 2002). Several studies have evaluated the efficiency of each
package. Though the seminal processing was not performed consistentiy, the results still
show variation in between species. Using bull (Johnson et al., 1990) and rooster
(Duplaix and Sexton, 1983) semen, there was little difference in the post-thaw results
between the 0.25 mL and 0.5 mL straws. However, Thomas and colleagues (1993) found
higher rates of progressive motile sperm post-thaw in canine semen with pellets when
compared to either of these straws. In lieu of the reduced capacity available from pellets
and straws, larger volume packages have also been investigated.

The straw is considered to be the most accepted technique for cryopreservation of
many cryopreservation protocols. Species included in this group include bovine (Johnson
et al., 1995), canine (Pena and Linde-l'orsberg, 2000), and equine (Samper et al., 2002).
In the swine industry, the most utilized commercial packaging unit is the 5 mL Maxi-
straw (Cordova et al., 2002; Eriksson et al., 2000), even though all three packaging
variations are still used (Eriksson et al., 2002).
Summary
The potential benefits of cryopreservation of sperm could provide ways to
overcome the limits of time involving reproduction by increasing the long-term storage
and use of the genetic materials of the male. However, as presented in this section, the
cryopreservation process is multi-faceted with interactions of several factors to take in to
consideration, including the alterations of stractural integrity due to the effects of
preparative handling (Comaschi et al., 1989; Crockett et al.. 2001; Eriksson et al., 2001;
Love et al., 2002), extenders (Arrada et al., 2002; Ball and Vo, 2001; Wilhelm et al.,
1996) and thermal insults (Devireddy et al., 2002; Lagares et al., 2000; Linfor et al.,
2002; Pommer and Meyers, 2002). In several species, the optimal combination has not
yet been found. Two species included in this grouping are the boar and stallion. Both
have been shown to have significant problems with cold shock (Holt, 1999; Johnson et
al., 2000; Pursel et al., 1972; Yoshida, 2000) as well as sensitivity to cryoprotectants
(Henry et al., 2002; Johnson et al., 2000; Medeiros et al., 2002). The objective of tiie
present study was to determine if many of the problems described within this review
20

could be overcome in the boar and stallion using a variety of pre-freezing techniques in
conjunction with a new freezing technology (UFT).
21

CHAPTER 111
EFFECTS OF SEMEN HANDLING ON POST-THAW
MOTILITY OUTCOMES OF CRYOPRESERVED
EQUINIi SPERMATOZOA
Abstract
The equine industry current lacks a standard protocol for handling semen for
cryopreservation. Due to this variability, success of extenders, incubations periods,
cryoprotectant amount and thermal alterations are difficult to assess. The objectives of
this trial were to assess the post-thaw outcomes for various protocols. This study was
conducted in four trials. In trial #1, an extended post-collection sample was diluted with
cryoprotectant (0%-10%) and held at one of two incubation periods (10 minutes or 1
hour) before being frozen in a unique freezing technology (UFT) system. Trial #2
evaluated similar cryoprotectant concentrations with extended incubations periods (0, 2,4
or 6 hours) at 5* 0. In trial #3, glycerol levels were limited to 2%, 5%, or 10% with a two
hour 5°C incubation. These samples were divided ftirther for freezing in a unique
freezing technology (UFT) system, controlled freezing unit or liquid nifrogen mist. The
fourth trial, added the effects of centrifiigation, step-down cooling rate during various
incubations, and the addition of two freezing extenders. From the results, all of the post-
thaw motilities for trials #1, #2, and #3 were 0%. However, trial #4 produced post-thaw
motilities ranging from 2% to 43%, averaging 21%. Comparing the outcomes from each
trial, there was a significant difference in the results between the first three trials and #4
22

(P<0.001). Though there was some consistency between the initial extender and freezing
treatments utilized for this study, our results suggest that incubation periods with
decreasing temperatures, centrifugation and the addition of freezing extenders
demonstrated potential. Further research is needed to investigate the influences of these
specific factors.
Introduction
Artificial insemination of mares with cooled; shipped semen has become common
procedure in several breeding facilities (Sieme et al., 2002). The acceptance of this
artificial reproductive technique has grown since the 1980's with the elimination of breed
organization regulations and the development of an effective transport container fLoomis,
2001). The utilization of frozen semen however has not progressed as quickly (Squires et
al.. 2002). However, the acceptance of cryopreservation within the equine industry has
only occurred within the last few years (Loomis, 2001). As a consequence, there is a lack
of standardized techniques (Samper et al., 2002; Samper and Morris, 1998; Thurston et
al., 2003) unlike the more established programs involving boars (Johnson et al., 2000) or
bulls (Vishwanath and Shannon, 2000).
The objective of this study was to evaluate the effects of several different
preparative handling factors while implementing the UFT.
23

Materials and Methods
Post-Collection Processing
Several ejaculates from four Quarter Horse stallions were collected during the
2001 and 2002 horse breeding season. All of the stallions were collected at the Texas
Tech University Ranch Horse Center in New Deal, TX, by trained personnel. After each
collection, the ejaculate was initially evaluated for total volume, gel-free volume,
concentration and initial motility. The original motility of all samples needed to be equal
or greater than 60% to be considered acceptable for this project. A sample was extracted
and extended 2:1 with pre-warmed E-Z Mixin CST Extender (Animal Reproductive
Systems; Chino, CA) and packaged for transport to one of two off-site laboratories for
ftirther processing.
Freezing Technology
A new unique freezing technology (UFT) was utilized in this study. SupaChill®
(SupaChill, Inc.; Lubbock, TX) incorporates an organic freezing fluid with a heat index
similar to water; thus allowing for a high rate of heat dissipation. The freezing
technology of UFT has the unique ability to be set at temperature levels below 0°C
(freezing point) with out solidifying.
24

Procedural Methodology
Trial #1
Upon arrival at the off-site laboratory, two stallion samples were divided into six
5 mL amounts in pre-labeled tubes. Samples were then diluted with glycerol at
concentrations of 0%, 2%, 4%, 6%, 8% or 10%. Two aliquots of 1.5 mL were removed
from each tube and placed in 1.8 mL cryo-tubes (Nalge Nunc Intemational; Denmark)
witii an "A" or "B" label. The A tubes were held at room temperature (22°C) for ten
minutes before being plunged into the -25°C UFT. The B tubes were cooled in a 5°C
refrigerator for one hour before being frozen in the -25*'C UFT. After freezing, the 24
vials were stored in liquid nifrogen until post-thawing analysis could be completed.
Trial #2
Upon arrival at the off-site laboratory, a sample was divided into five 7 mL
amounts in pre-labeled tubes. Samples were then diluted with glycerol at concentrations
of 2%, 4%, 6%, 8% or 10%. One mL aliquots were removed from each tube and placed
in 1.8 mL cryo-tubes with a number label ranging from 0 to 6. These numbers
represented the number of hours in which the samples were held at 5°C before freezing.
Vials were frozen in the -25*'C UFT, after holding for one hour. After fi-eezing, the 35
vials were stored in liquid nitrogen until post-thawing analysis could be completed.
25

Trial #3
A third trial similar to the procedure of the second was also completed using two
stallion samples. However, only 2%, 5%, and 10% glycerol concentrations were used
with a two hour incubation in the 5°C. The freezing treatments were expanded to include
the UFT, a control freezing unit (Planar Unit, Planar Products Ltd.; United Kingdom) or
nittogen mist vapor. After freezing, the 18 vials were stored in liquid nifrogen until post-
thawing analysis could be completed.
Trial #4
Upon arrival at either laboratory, each sample was placed into a 17°C refrigeration
unit. TTie sample was held for one hour at this temperature after which a pre-cooled 17**C
wash buffer was added. The sample was held at this temperature for an additional 1 Vi
hour to equilibrate with the wash buffer. The sample then was centrifiiged to create a
sperm-composed pellet. The pellet was then dissipated in a pre-cooled 25% egg-yolk
extender at a 1:1 ratio with the original sample size. The sample was placed in a 5°C
refrigeration unit for 1 '/j hour before a secondary freezing extender was added in three
increments over 30 minutes. This 25% egg-yolk extender with 6% glycerol was also pre-
cooled to 5°C. Following the addition of the last increment, the sample was held at 5°C
for one hour before freezing. The sample was not broken into confrol and freatment
groups until immediately before freezing to ensure consistent handling. One ml aliquots
were extracted from the extended sample and placed into 1.8 ml labeled cryo-tubes for
freezing. Four freezing treatments were tested for this trial, three UFT and one confrol:
26

B: 4 minutes at-25°C
C: 4 minutes at -25°C with a proprietary filter #1
E: 4 minutes at -25°C with a proprietary filter #3
Control: 10 minutes in liquid nitrogen mist.
After freezing, the 24 vials were stored in a liquid nitrogen tank until the post-thaw
analysis could be completed.
Thawing Procedure
In Trials #1 and #2, a slow thaw protocol was utilized. The samples were
extracted from the nifrogen storage tank and placed in a 5°C refrigerator for 30 minutes.
This was followed by 5 minutes in a 37*'C water bath. For trials #3 and #4, a faster
thawing protocol was utilized. The samples were placed held in the air (22*'C) for 45
seconds before warming in a 37°C water bath for 5 minutes. All samples were
subjectively evaluated on a random and individual basis for motility.
Statistical Analysis
The post-thaw motilities from the four trials were compared using analysis of
variance (ANOVA). ANOVA was also used to compare the post-thaw outcomes from
the four freezing freatments (B, C, E, Control) within Trial #4. All statistics were
analyzed using the SPSS Version 8.0 Software (SPSS, Inc.; Chicago, IL).
27

Results
All samples from trials 1#, 2#, and #3 had 0% post-tiiaw motilities. Post-thaw
motilities from samples of trial #4 ranged from 2% to 43%. Consequentiy, a significant
difference was observed when comparing the post-thaw motilities of Trials #1, #2, and #3
to tiiose of Trial #4 (P<0.001). This is presented in Table 3.1.
Table 3.1 Comparison of Post-Thaw Motilities from Four Pre-Freezing Protocols
Motility
Trial n Avg Post-Thaw, % SD
#1 24 0%" 0.0
#2 35 0%" 0.0
#3 18 0%" 0.0
#4 24 21%** 9^4
Different superscripts represent significant differences in the post-thaw motihty percentages between the four trials as determined by P<0.001
Variations in between the four freezing treatments used in Trial #4 were also
investigated. The average post-thaw motilities for B (19%), C (18%). E (22%). and
Confrol (20%) groups were not significantly different.
Discussion
From our post-thaw comparison, the higher retums produced in Trial #4
emphasizes the need for a pre-freezing protocol with focus on osmotic equilibrium.
28

Though temperature shifts were utilized in the first three trials, trial #4 was the only
protocol with a two phase pre-freeze temperature shift, centrifiigation, and the addition of
an egg yolk freezing extenders.
Seminal plasma can only sustain sperm viability for a few hours after ejaculation
(Johnson et al., 2000). Higher rates of lipid peroxidation were observed in boar semen
incubated at 37°C in seminal plasma (Comaschi et al., 1989). This coincides with the
findings of Trials #1, #2, and #3, where seminal fluid was maintained in the sample
during the cooling process. Removal of the seminal plasma by centrifiigation created a
higher rate of chromatin stability (Love et al., 2002) and higher post-thaw motilities
(Martin et al., 1979) in stallion samples. The beneficial effects of removing the supemant
were observed in Trial #4.
It has been suggested that successfiil post-thaw results may be achieved through a
series of osmotic steps (Curry and Watson, 1994; Gao et al., 1993). Evidence of this idea
has been observed through the increases in cold shock resistance observed in sperm after
incubation or holding {jeriods at cooler temperatures. Species in which this included was
the stallion (Braemmer et al., 2002; Crockett et al., 2001) and boar sperm (Eriksson et al.
2001; Pursel et al., 1972). In several commercial boar settings, protocols involves
cooling a sample in seminal fluid for 2 hours before centrifiigation and cooling further
before freezing (Johnson et al., 2000).
Glycerol is usually added at concenttations of 2.5 to 6% for stallions (Samper and
Morris, 1998; Vidament et al., 2002). All four trials had glycerol concenfrations that fell
into this range. However glycerolized diluents have reported to have varying success
29

rates based on time to equilibration. Initially it was thought that extended semen exposed
to glycerol did not show any survival differences post-thaw based on exposure time.
However, a trial utilizing 0.5 mL straws demonstrated that an incubation period of 4
hours produced higher motilities and improved morphology (Johnson et al., 2000).
Temperature in which the glycerol is added also affects the post-thaw outcomes.
Vidament and colleagues (2000) found the addition of glycerol at 22°C generated a
higher retum rate than glycerol added at 4°C. Though trials #2 and #3 had longer
incubation periods at 5°C, the lack of an additional extender may lead to a metabolism
deficiency. Also egg yolk extenders possess protective attributes when added to
spermatozoa during cryopreservation (Medeiros et al., 2002).
In conclusion, refinement to preparative handling techniques of stallion sperm is
required for the development of an optimal protocol. Our results suggest that incubation
periods with decreasing temperatures, centrifiigation and the addition of freezing
extenders demonstrated potential. Further research is needed to investigate the influences
from these factors and the repeatability of this protocol with other stallions
30

CHAPTER IV
COMPARISONS OF A UNIQUE FREEZflvlG
TECHNOLOGY (UFT) TO TRADITIONAL
LIQUID NITROGEN METHODOLOGY FOR
CRYOPRESERVATION OF EQUINE SPERMATOZOA
Abstract
Several breed organizations in the equine industry have recently accepted the use
of frozen semen. However, cryopreserved stallion semen is commonly associated with
poor post-thaw semen quality and decreased pregnancy rates as compared to those
produced during normal mating or with cooled semen techniques. Therefore our
objective was to evaluate a new unique freezing technique (UFT) with the intent of
improving fertility outcomes. A series of experiments tested the UFT compared to
traditional liquid nitrogen methodology in combination with influence of the extenders
and stallions used. In Experiment #1, post-thaw motility results of UFT variations were
compared to those from liquid nitrogen methods. The averaged post-thaw motility
percentages of the four UFT treatments were similar when compared to the traditional
liquid nitrogen control group (P=0.845). In experiment #2, two egg-yolk based freezing
extenders, Biladyl AB® (Minitub GmbH; Germany) intended for bovine samples and
Freezing Medium Test Yolk Buffer® extender (Irving Scientific; San Jose, CA) used
human samples, were compared. A significant difference in the average post-thaw
motilities was found in between the Biladyl AB® (17%) as compared to the Test Yolk
31

Medium Buffer® extender (25%) (P<0.002). In the final experiment, we compared
variability between stallions using the UFT with the intensions of creating a more
consistent outcome. Post-thaw motilities and percent of original motility retum between
the four stallions were significantiy different (P<0.001).
In conclusion, the UFT may be potentially used as an alterative freezing method
to replacement current liquid nitrogen methodology. However, further investigation is
needed to refine techniques.
Infroduction
Until recently, freezing of equine spermatozoa was limited due to poor post-thaw
results (Devireddy et al., 2002; Samper et al., 2002) and breed regulations. Though the
reproductive capacity of equine frozen semen is still low, the acceptance of this
technology is slowly growing due to the elimination of other breed registry barriers
(Loomis, 2001).
Traditional freezing methodology of equine semen involved placing a sample in
liquid nitrogen vapor for 10 to 20 minutes before plunging. Though still used today, the
post-thaw results are less than optimal. Within the last few years, a high speed freezing
system has emerged for freezing foodstuffs with an increased ability of preserving
freshness (SupaChill®, SupaChill USA; Lubbock, TX). This Unique Freezing
Technique (UFT) contains an organic fluid with a heat capacity similar to water with a
freezing rate of-6.1°C/minute. This technique has been applied to various cell samples
and tissue biopsies with successful thawing results in preliminary trials. Our objective
32

was to compare the Ul i to traditional nitrogen methods while investigating various
cryopreservation factors.
Two cryopreservation factors were addressed in this study while studying the
UFT. More than one-half century ago, egg yolk was discovered to possess protective
attributes when added to spermatozoa during cryopreservation (Medeiros et al., 2002).
At least two egg-yolk cryoprotectant have demonstrated successfiil results in two other
species; Biladyl AB® (Minitub GmbH; Germany), a cryoprotectant is commonly used
for bovine semen cryopreservation, and freezing Medium Test Yolk Buffer® (Irving
Scientific; San Jose, Califomia), a cryoprotectant used on human samples. As no
standard cryoprotectant system exists in equine, the intention of this study was to assess
the post-thaw motilities of stallion semen frozen with these two extenders while studying
tiie UFT.
Several reports have discussed the variability seen in the cooling and freezing of
equine semen. Although several factors including health, season and age have been
known to affect semen quality, lower pregnancy rates from various stallions have been
related to the tolerance of semen sample (Samper et al., 2002). Our evaluation of the use
of the UFT in cryopreservation of stallion spermatozoa focused on the performance of
post-thaw motility, and comparing consistency between stallions.
33

Materials and Methods
Semen Collection
Several ejaculates from four Quarter Horses stallions were collected for this trial
during the 2002 breeding season. All stallions were collected off a breeding phantom
using a Missouri artificial vagina at the Texas Tech University New Deal Farm, TX.
After each collection, the ejaculate was initially evaluated for total volume, gel-free
volume, concentration and initial motility. The original motility of all samples needed to
be at least 60% to be considered acceptable for this project. A 30 ml sample was
exfracted and extended 2:1 with Equine E-Z Mixin CST Extender (Animal Reproductive
Systems; Chino, CA) and packaged for transport to one of two off-site laboratories for
ftirther processing.
Semen Processing
Up)on arrival to either laboratory, each sample was placed into a 17°C
environment. After holding for one hour at this temperature, a wash buffer pre-cooled to
the same temperature was added. The sample was maintained at this temperature for an
additional 1 '/2 hour, after which the sample was centrifuged. The supernatant fluid was
extracted and discarded. The remaining pellet was dissipated in 30 mL of a pre-cooled
freezing egg-yolk based extender (17°C). The sample was placed in 5°C refrigeration
unit for 1 '/z hours before 30 mL of the second pre-cooled freezing extender with 6%
glycerol was added. The second extender was added in increments over 30 minutes.
Following the addition of the final increment, the sample was held at 5°C for one hour
34

before freezing. The sample was not broken into control and treatment groups until
immediately before freezing to ensure consistent handling.
Unique Freezing Technology
A new unique freezing technology (UFT) was utilized in this study. SupaChill®
(SupaChill USA; Lubbock, TX) incorporates an organic freezing fluid with a heat index
similar to water; thus allowing for a high rate of heat dissipation. The freezing fluid of
UFT has the unique ability to be set at temperature levels below O C (freezing point) with
out solidifying. Preliminary trials freezing foodstuffs and various cell lines with UFT
showed improved post-thaw retums versus other fraditional freezing methodology.
Freezing Methodology
One mL increments were extracted from the extended sample and placed into 1.8
mL cryo-tubes (Nalge Nunc Intemational; Denmark) for freezing. Preliminary trials
demonstrated higher post-thaw outcome with the cryo-tube than with the fraditional 0.5
ml straw. Cryo-tubes were grouped and labeled in sets of three for freezing. Preliminary
trials on boar semen determined the UFT freezing freatments selected. Freezing and
labeling for the UFT were as follows:
B: 4 minutes at-25°C
F: 5 minutes at -10°C, tiien 4 minutes at -25°C
G: 5 minutes at -10°C then 4 minutes at -25°C with a proprietary filter #1
35

J: 5 minutes at -10°C witii a proprietary filter #1 then 4 minutes at -25°C with a
proprietary filter #1
Control: 10 minutes in the liquid nitrogen mist.
Confrol vials were frozen using traditional liquid nitrogen methodology. Once initial
freezing was complete, all vials were stored in a nitrogen tank for at least 48 hours before
being evaluated for post-thaw values.
Thawing Process
Randomly selected samples were thawed individually. After extraction from the
liquid nifrogen tank, samples were warmed for 45 sec at room temperature (22°C) before
being placed in a 37°C water bath for 5 minutes. Samples were assessed manually by the
same observer for motility values.
Statistical Analysis
All data was analyzed using the SPSS Version 8.0 Software (SPSS, Inc.; Chicago,
IL). In Experiment #1, the statistical significance of the post-thaw motility values
between the UFT treatment groups and control group was analyzed using analysis of
variance (ANOVA). In experiments #2 and #3, ANOVA was used to statistical
determine the differences between extenders and stallions. In the case that the
comparison was significantly different, such as in Experiment #3 the stallion comparison,
the stallions were separated using Fischer's Least Square Difference (LSD) procedure.
36
In each experiment, the effects of the specific UFT techniques used were also evaluated
by ANOVA.
Results
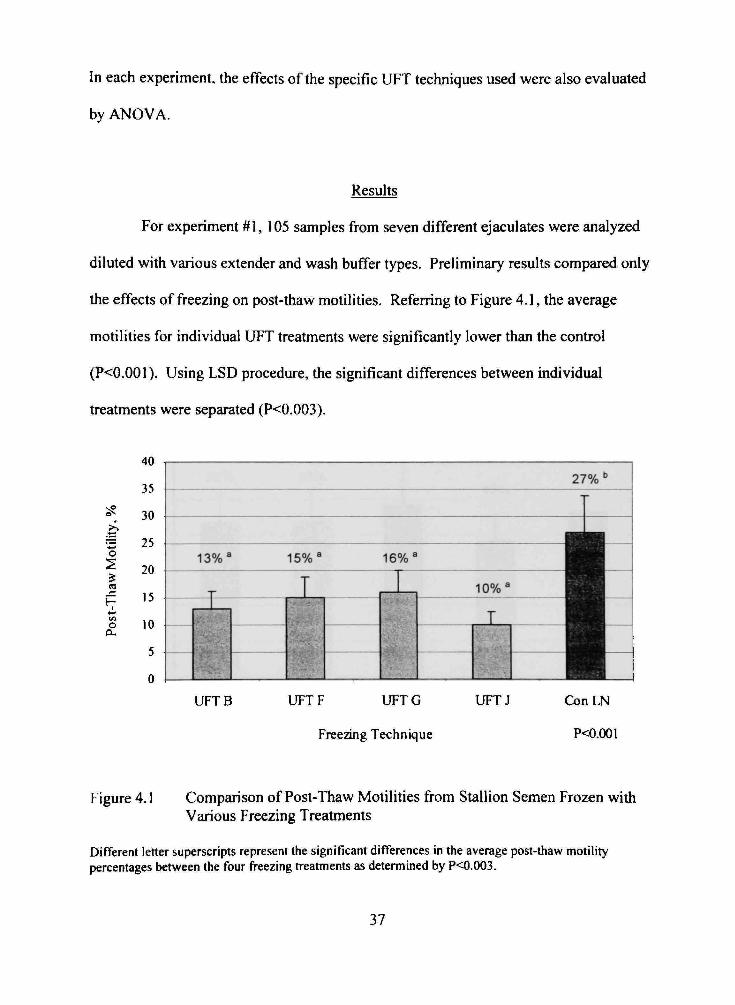
For experiment #1, 105 samples from seven different ejaculates were analyzed
diluted with various extender and wash buffer types. Preliminary results compared only
the effects of freezing on post-thaw motilities. Referring to Figure 4.1, the average
motilities for individual UFT treatments were significantly lower than the control
(P<0.001). Using LSD procedure, the significant differences between individual
treatments were separated (P<0.003).
CO
o Q-
40
35
30
25
20
15
10
5
27% "
13%" 1 5 % ' 16%'
^H
- m-'~ rrr. _, ^H
^1
— 1
UFT B UFT F UFT G
Freezing Technique
UFTJ ConLN
P<0.001
Figure 4.1 Comparison of Post-Thaw Motilities from Stallion Semen Frozen with Various Freezing Treatments
Different letter superscripts represent the significant differences in the average post-thaw motility percentages between the four freezing treatments as determined by P<0.003.
37
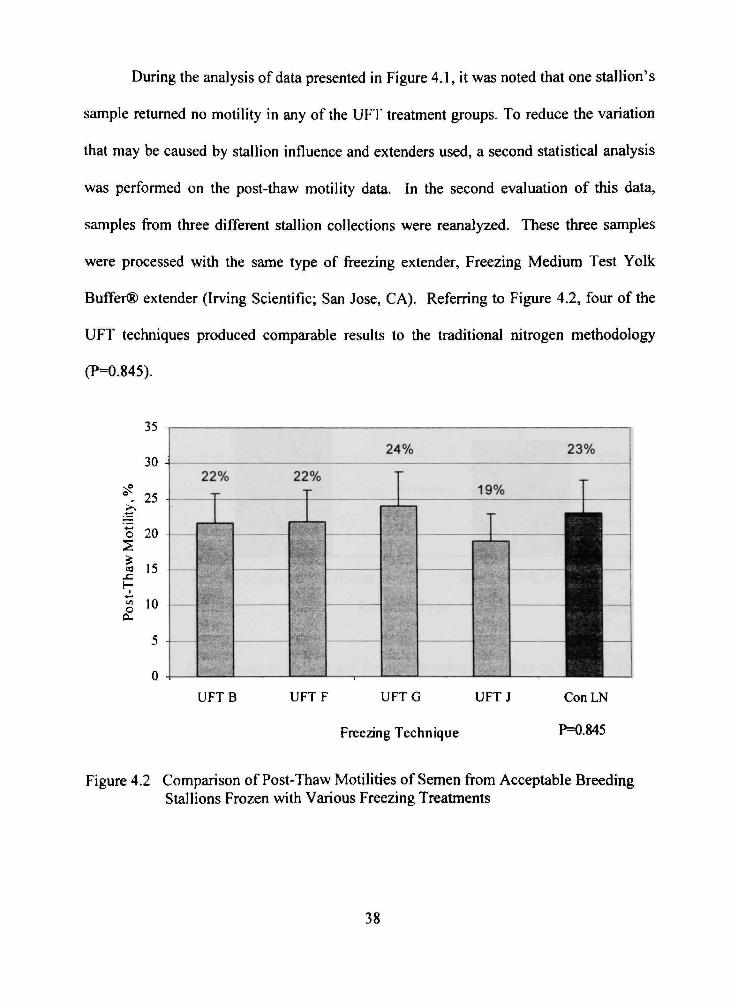
During the analysis of data presented in Figure 4.1, it was noted that one stallion's
sample returned no motility in any of the UFT treatment groups. To reduce the variation
that may be caused by stallion influence and extenders used, a second statistical analysis
was performed on the post-thaw motility data. In the second evaluation of this data,
samples from three different stallion collections were reanalyzed. These three samples
were processed with the same type of freezing extender. Freezing Medium Test Yolk
Buffer® extender (Irving Scientific; San Jose, CA). Referring to Figure 4.2, four of the
UFT techniques produced comparable results to the traditional nitrogen methodology
(P=0.845).
UFTB UFTF UFT G UFTJ
Freezing Technique
ConLN
P=0.845
Figure 4.2 Comparison of Post-Thaw Motilities of Semen from Acceptable Breeding Stallions Frozen with Various Freezing Treatments
38
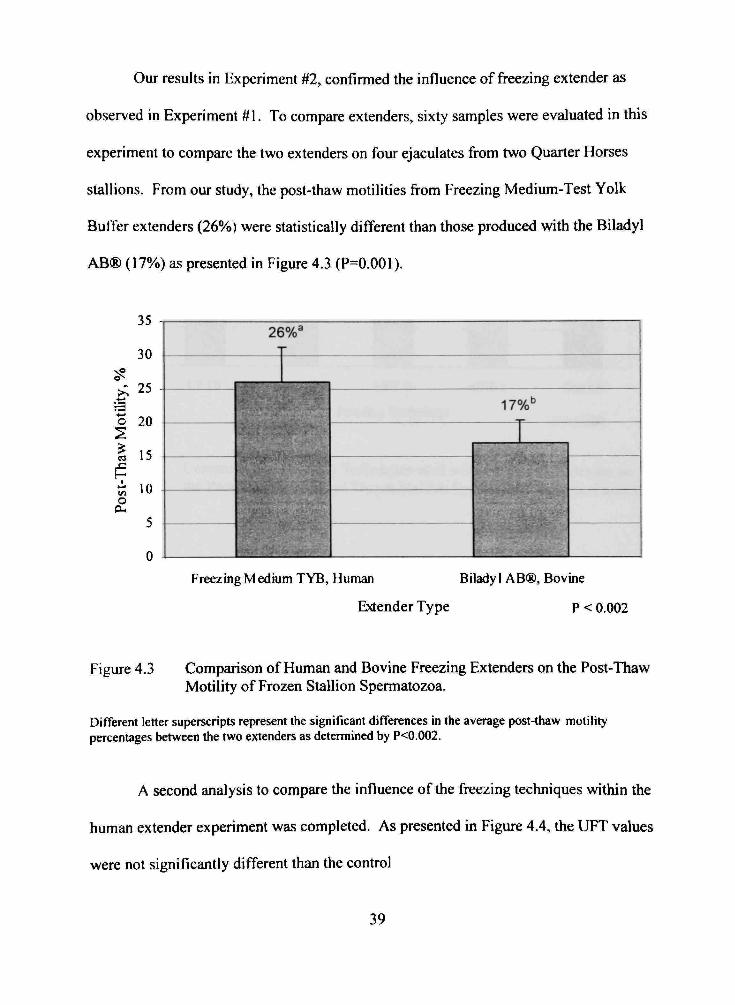
Our results in Experiment #2, confirmed the influence of freezing extender as
observed in Experiment #1. To compare extenders, sixty samples were evaluated in this
experiment to compare the two extenders on four ejaculates from two Quarter Horses
stallions. From our study, the post-thaw motilities from Freezing Medium-Test Yolk
Buffer extenders (26%) were statistically different than those produced with the Biladyl
AB® (17%) as presented in Figure 4.3 (P=0.001).
35
30
25 •*- .»
a 0 20 4
1 15 H t); 10 o
O H
5
26%^
17%"
1
Freezing M edium TYB, Human Bilady i AB®, Bovine
Extender Type p < 0.002
Figure 4.3 Comparison of Human and Bovine Freezing Extenders on the Post-Thaw Motility of Frozen Stallion Spermatozoa.
Different letter superscripts represent the significant differences in the average post-thaw motility percentages between the two extenders as determined by P<0.002.
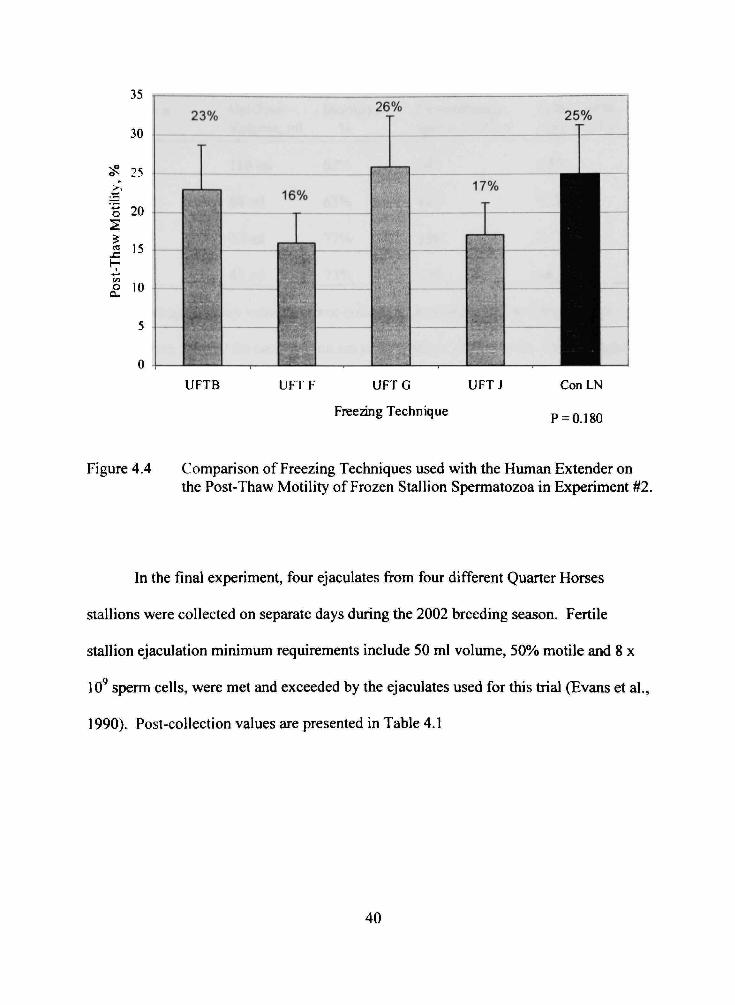
A second analysis to compare the influence of the freezing techniques within the
human extender experiment was completed. As presented in Figure 4.4, the UFT values
were not significantly different than the control
39
UFT F UFT G UFT J Freezing Technique
ConLN
P = 0.180
Figure 4.4 Comparison of Freezing Techniques used with the Human Extender on the Post-Thaw Motility of Frozen Stallion Spermatozoa in Experiment #2.
In the final experiment, four ejaculates from four different Quarter Horses
stallions were collected on separate days during the 2002 breeding season. Fertile
stallion ejaculation minimum requirements include 50 ml volume, 50% motile and 8 x
lO' sperm cells, were met and exceeded by the ejaculates used for this trial (Evans et al.,
1990). Post-collection values are presented in Table 4.1
40
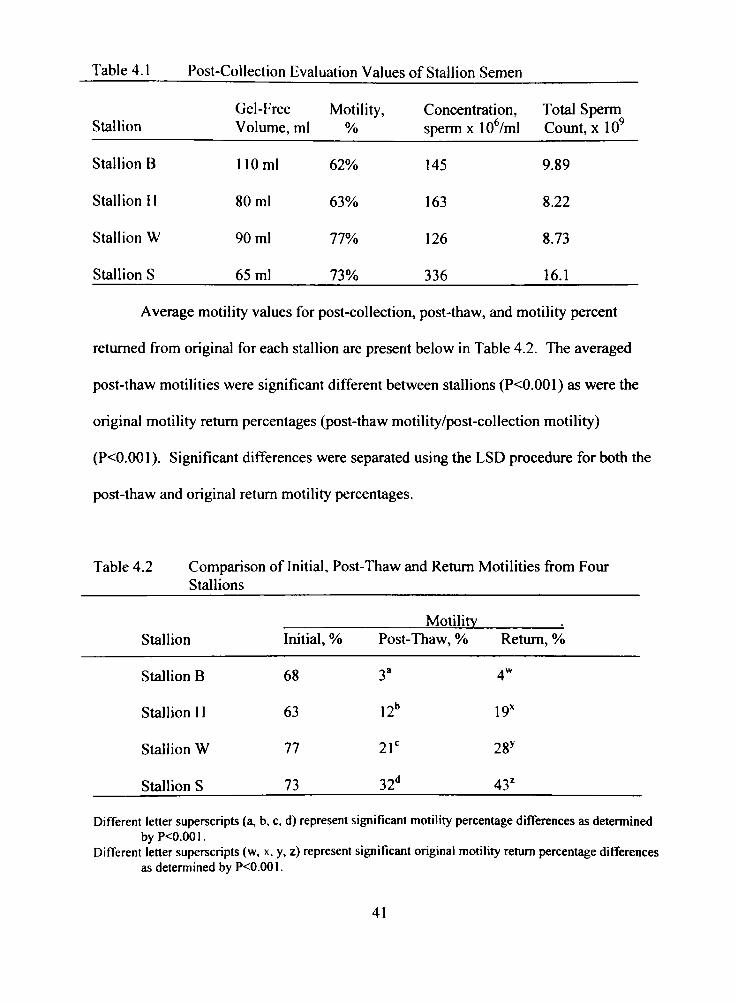
Table 4.1 Post-Collection Evaluation Values of Stallion Semen
Gel-Free Motility, Concentration, Total Sperm
Stallion Volume, ml % sperm xlO^/ml Count, x lO '
Stallion B 110 ml 62% 145 9.89
Stallion 11 80 ml 63% 163 8.22
Stallion W 90 ml 77% 126 8.73
Stallion S 65 ml 73% 336 16J
Average motility values for post-collection, post-thaw, and motility percent
returned from original for each stallion are present below in Table 4.2. The averaged
post-thaw motilities were significant different between stallions (P<0.001) as were the
original motility retum percentages (post-thaw motility/post-coUection motility)
(P<0.001). Significant differences were separated using the LSD procedure for both the
post-thaw and original retum motility percentages.
Table 4.2 Comparison of Initial, Post-Thaw and Retum Motilities from Four Stallions
Stallion
Stallion B
Stallion H
Stallion W
Stallion S
Initial,
68
63
77
73
% Motility
Post-Thaw,
3a
12"
2,c
32"
% Retum, %
4W
19"
28^
43^
Different letter superscripts (a, b, c, d) represent significant motility percentage differences as determined byPO.OOl.
Different letter superscripts (w, x, y, z) represent significant original motility retum percentage differences as determined by P<0.001.
41

Variability caused by the UFT freezing treatments was also investigated. The
post-thaw motility averages of the UFT treatments, B (16%), F (19%), G(l 8%), and J
(15%), were found not to be significantly different (P=0.832). The average original
motility retum percentages for B (23%), F (27%), G (25%), and J (20%) were also not
found to be statistically different (P=0.782).
Discussion
The data has shown that the UFT has demonstrated post-thaw motility potential
comparable to that produced by liquid nitrogen methodology. In preliminary studies,
UFT freezing rate was determined to be -4.3°C/minute, which is vary similar to that of
liquid nitrogen.
It has also been suggested that an optimal freezing procedure would include
osmotic changes or temperature decreases in a series of steps (Curry and Watson, 1994;
Gao et al., 1993). The UFT has the unique ability to cool samples to set temperatures
below the point of freezing (0°C). This point is emphasized by the most successfiil UFT
treatment of this trial, G, in Experiments #1 and #2. In this freezing freatment, the
sample was lowered in steps to -10°C, -25°C, and then -196°C in the liquid nifrogen.
During the initial freeze, the sample enters a hyperosmotic environment where the
extracellular fluid freezes before the intracellular water at -10°C (Medrano et al., 2002).
Limiting the freezing rate to -5°C/minute through this critical temperature range before
increasing the freezing rate, has previously proven successful in rams (Byme et al.,
2000).
42

It should also be recognized that the fluid used within the UFT has a heat
dissipation rate similar to that of water. Thus the issue of non-uniform freezing involving
larger volume samples would not be a concern.
From our results, the Freezing Medium Test Yolk Buffer® extender appeared to
be more effective. This extender may have a cryoprotectant concenfration may be more
appropriate for the rate of freezing used (Table 4.3). Glycerol is usually added at
concenfrations of 2.5 to 6% to stallion samples (Samper and Morris, 1998; Vidament et
al., 2002). The bovine extender not only had a higher glycerol amount, but also
contained more active antibiotics such as Gentamycin, Spectinomycin and Linomycin.
Table 4.3 Egg Yolk and Glycerol Composition of Biladyl AB and Test Yolk Buffer Extenders
Biladyl AB®
20%
7%
Component
Egg Yolk
Glycerol
Test Yolk Buffer®
20%
6%
Reference: Insert instructions from Biladyl® (Minitub GmbH; Germany) and Freezing Medium-Test Yolk Buffer Freezing extender (Irving Scientific; San Jose, Califomia)
Male individual viuiation has been noted in several other mammalian species such
as tiie mouse (Holt, 1999), bull (Iritani, 1988; Vidament et al.. 2000), and ram (Byrne et
al., 2000) with regard to cryopreservation success. Therefore, some breeding stallions
may produce acceptable samples for natural or artificial cooled breeding standards; but
not all stallions produce samples acceptable for freezing. It is estimated that only 24% of
43

all stallions are to produce an ejaculate suitable for cryopreservation (Linfor et al., 2002;
Vidament et al., 1997). Our results also reflected an inconsistency of seminal retum
between the four different stallions using our new freezing technique. However, two of
the stallions had post-thaw motilities considered to at acceptable levels suggested from a
survey of commercial equine breeding stations (Samper and Morris, 1998). Thus, there is
potential for this new technique.
In summary, the UFT produced comparable results to those from the traditional
liquid nifrogen methodology. Due to the potential of this technology, ftirther
investigations involving stallion semen are merited.
44

CHAPTER V
EFl ECTS OF SEMEN HANDLING ON POST-THAW
MOTILITY OUTCOMES OF CRYOPRESERVED
PORCUME SPERMATOZOA
Abstract
Cryopreservation of boar semen within the swine industry has commonly been
correlated to lower piglet numbers per litter produced. However, swine reproduction
offers various challenges compared to other livestock animals including large ejaculate
volumes, litter bearing sows, and temperature sensitive semen. Due to this variability of
success, the aim of the present study was to assess the post-thaw outcomes of various
preparative protocols. This study was conducted in four trials. In trial #1, an extended
post-collection sample was diluted with cryoprotectant (0%-10%) and held at one of two
incubation periods (30 minutes or 5 hour) before being frozen in a unique freezing
technology (UFT). Trial #2 evaluated similar cryoprotectant concenfrations with
extended incubations periods (0, 2, 4, 6, 8 hours) at 5°C before UFT freezing. In trial #3,
extended samples are incubated at either 22°C or 17°C before centrifiigation and addition
of glycerolized media prior to UFT or nitrogen mist freezing. In the last trial, trial #4, the
effects of centrifiigation, step-down cooling rates during various incubations, and the
addition of two freezing extenders were examined. All of the post-thaw motilities for
trials #1 and #2 were 0%. Though trial #3 produced a few samples with significant
motility, the average was also 0%. However, there was a significant difference between
45

each trial with #4 (P<0.00l). Though there was some consistency between the initial
extender and freezing treatments utilized for this study, the variation observed between
the four trials emphasizes the need for a cooling incubation periods and centrifugation as
part of a freezing protocol for boar semen.
Introduction
Semen cryopreservation for boar semen has been commercially available since
1975. Yet, the fertility performance of cryopreserved samples is not considered efficient
as demonsfrated by lower average farrowing rates of seven piglets to a litter (Johnson et.
al., 2000). Swine reproduction, though, is unique compared to other livestock sjjecies.
Boars produce large ejaculate amounts with temperature sensitive sperm (Pursel et al.,
1972). Sows, as a litter bearing animal, create a fertilizing challenge through prolonged
ovulations (Johnson et al., 2000). Thus, insemination amounts must also be large (Holt,
2000).
Currently, two cryopreservation protocols, the Westendorf and Beltsville
methods, are being utilized in commercial boar settings. As part of the Westendorf
method, semen is diluted and then frozen in straws. As for the Beltsville method, a
sample is cooled for 2 hours prior to centrifugation and then cooled ftirther for 3 hours
before pelleting (Johnson et al., 2000). Even though the swine industry has established
protocols, as recognized with bulls (Vishwanath and Shannon. 2000), the procedures
need refinement to produce satisfactory results. Therefore, the objective of this study
46

was to evaluate the effects of several different preparative handling factors while
implementing a new UFT.
Materials and Methods
Post-Collection Processing
Several ejaculates from three boars were collected during the fall of 2001. All the
boars were collected at the Texas Tech University Farm at New Deal, TX, by frained
personnel. After each collection, the ejaculate was evaluated for total gel-free volume,
semen concenfration and initial motility. The original motility of all samples utilized had
60% or greater and meet concentration requirements for established breeding protocols.
A sample was extracted, extended 2:1 with pre-warmed X Cell Extender (IVM, Maple
Grove, MN), and packaged for transport to one of two off-site laboratories for further
processing.
Freezing Technology
A new unique freezing technology (UFT) was utilized in this study. SupaChill®
(SupaChill, Inc.. Lubbock, TX) incorporates an organic freezing fluid with a heat index
similar to water; thus allowing for a high rate of heat dissipation. The freezing fluid of
UFT has the unique ability to be set at temperature levels below 0°C (freezing point) with
out solidifying. This technology was utilized in all four of the trials within this study.
47

Trial #1
Upon arrival at the off-site laboratory, a sample was divided into six 5 mL
amounts in pre-labeled tubes. Samples were then diluted with glycerol at concentrations
of 0%, 2%, 4%, 6%, 8% or 10%. Two aliquots of 1.5 mL were removed from each ttibe
and placed in 1.8 mL cryo-tubes (Nalge Nunc Intemational, Denmark) with an "A" or
"B" label. The A tubes were held at cooled in a 5°C refrigerator for thirty minutes while
the B tubes were held at 5°C for one hour. Upon completion of this incubation, the
samples were plunged into the -25°C UFT and maintained in this environment for one
hour.
Trial #2
Upon arrival at the off-site laboratory, a sample was divided into four 6 mL
amounts in pre-labeled tubes. Samples were then diluted with glycerol at concentrations
of 4%, 6%, 8% or 10%. One mL aliquots were removed from each tube and placed in 1.8
mL cryo-tubes with a number label ranging from 0 to 10. These numbers represented the
number of hours in which the samples were held at 5°C before freezing. Vials were
frozen and held in the -25°C UFT for one hour.
Trial #3
At the off-site laboratory, two samples were individually divided into two equal
parts. One half was placed in a 17°C refrigeration unit, the other held at room
temperature (22°C). Samples remained in this environment for one hour. After
48

centrifuging the sample and removing the supemant, a 6% glycerol media was added to
each sample. Cryo-tubes and sfraws with 0.5 mL volume were filled with samples from
both incubation groups. Upon freezing, the samples were divided again. One half of
each group was frozen in the liquid nifrogen mist for 30 minutes. The other half was
frozen in the UFT at -25°C for two minutes.
Trial #4
Upon arrival at either laboratory, each sample was divided and placed into a 17°C
refrigeration unit or held at room temperature (22°C). The sample was held for one hour
at this temperature after which a pre-cooled 17*'C wash buffer (IVM, Maple Grove, MN)
was added. The sample was held at this temperature for an additional 1.5 hour to
equilibrate with the wash buffer. The sample then was centrifiiged to create a sperm-
composed pellet. The pellet was then dissipated in a pre-cooled 25% egg-yolk extender
(Boarchipos A, IVM, Maple Grove, MN) at a 1:1 ratio with the origmal sample size. The
sample was placed in a 5° C refrigeration unit for 1.5 hour before a secondary freezing
extender was added in three increments over 30 minutes. This 25% egg-yolk extender
with 6% glycerol (Boarcipheos B, IVM, Maple Grove, MN) was also pre-cooled to 5° C.
Following the addition of the last increment, the sample was held at 5° C for one hour
before freezing. The sample was not broken into confrol and treatment groups until
immediately before freezing to ensure consistent handling.
49

One ml aliquots were exfracted from the extended sample and placed into 1.8 ml
labeled cryo-tubes for freezing. Only UFT freezing treatments were tested for this trial:
A: 10 minutes at-10°C
B: 4 minutes at -25° C
C: 4 minutes at -25° C with a proprietary filter #1
D: 4 minutes at -25° C with a proprietary filter #2
E: 4 minutes at -25° C with a proprietary filter #3
F: 5 minutes at -10° C, then 4 ntinutes at -25° C
G: 5 minutes at -10° C, then 4 minutes at -25° C with a proprietary filter #1
H: 5 minutes at -10° C, then 4 minutes at -25° C with a proprietary filter #2
1: 5 minutes at -10° C, then 4 minutes at -25° C with a proprietary filter #3
J; 5 minutes at -10° C with a proprietary filter #1, then 4 minutes at -25° C with a
proprietary filter #1
K: 5 minutes at -10° C with a proprietary filter #2, then 4 minutes at -25° C with a
proprietary filter #2
L: 5 minutes at -10° C with a proprietary filter #3, then 4 minutes at -25° C with a
proprietary filter #3.
Thawing Procedure
All samples were held in a liquid nitrogen tank for at least 48 hours before
assessment. In Trials #1 and #2, the thawing protocol included 2 minutes at ambient
temperature (22°C) before placement into a 37°C water bath for 2 minutes. For trial #3,
50

five different thawing procedures were utilized. The procedures involved warming at
room temperature for 10 or 45 seconds before incubation in the 37°C water bath for 5 or
10 minutes. The samples from Trial #4 were thawed using three thawing rates. Either
warming for 45 seconds at 22°C before 5 minutes in a 37°C water bath, 45 seconds at
22°C before 5 minutes in a 37°C water bath and 15 seconds in a 55°C water bath, or 15
seconds in a 55°C water bath before 5 minutes in a 37°C water bath.
Statistical Analysis
On the data from the four trials was analyzed using analysis of variance
(ANOVA). All statistics were analyzed using the SPSS Version 8.0 Software (SPSS,
Inc., Chicago, IL).
Results
Post-thaw motilities from trials #1 and #2 were 0%. Post thaw motilities from
samples of trial #3 ranged from 0% to 15%. However the majority were 0% (8 motile
samples: 140 total samples). Trial #4 produced the highest post-thaw motilities,
averaging 24%. Consequently, a significant difference was observed between the
treatments (P<0.001). This is emphasized in Table 5.1.
51

Table 5.1 Comparison of Pre-Freezing Procedural Trials on Boar Spermatozoa
Motility
Trial Avg Post-Thaw, % SD Range
#1 0%" 0.0 0
#2 0%" 0.0 0
#3 0%' 0.0 0-15
#4 24%" n j 0-45
Different superscripts represent significant motility percentage differences as determined by P<0.001
Discussion
The higher motilities reported in trial #4 reflect the importance of incubation
cooling periods and centrifugation of the semen sample as part of the boar
cryopreservation protocol. Trial #4 had a similar procedure to the Beltsville method,
used in commercial; breeding facilities (Johnson et al., 2000). Trials #1 and #2, lacked
centrifugation, as does the Westendorf method. However, these procedures did not prove
to be successful.
Seminal plasma cannot sustain sperm viability beyond a few hours after
ejaculation (Johnson et al., 2000). In boars, higher rates of lipid peroxidation were
observed in semen when incubated at 37°C within this native plasma (Comaschi et al.,
1989). These findings concur with the poor performance of trials #1 and #2, where
seminal fluid was maintained in the sample during the cooling process. In stallion semen
samples, removal of the seminal plasma by centrifugation created a higher rate of
chromatin stability (Love et al., 2002) and higher post-thaw motilities (Martin et al..
52

1979). This beneficial effect of centrifugation was observed in trial #3, but more
significantly in trial #4.
Boar spermatozoa are very temperature sensitive (Watson, 2000). The Beltsville
Method involves cooling a sample in seminal fluid for 2 hours before centrifiigation and
cooling further before freezing (Johnson et al., 2000). It has been suggested that
successful post-thaw results may be achieved through a series of osmotic steps (Curry
and Watson, 1994; Gao et al., 1993) such as thermal reductions. Evidence of this trend
has been observed in sperm after holding periods at cooler temperatures. Improvements
in cold shock resistance post-incubation have been reported in seminal samples from
stiillions (Brummer et al., 2002; Crockett et al., 2001) and boars (Eriksson et al. 2001;
Pursel etal., 1972).
Due to the sensitivity of boar sperm, the recommended glycerol concenfration is
only 3% (Johnson et al., 2002). Of the four trials tested in this study, all four trials had
glycerol concenfrations that were similar to this percentage. In reviewing the total
sample composition, trial #4 had a 3% glycerol concentration. However, not only is the
amount of glycerol important, so is the way in which it is added. Incubation timing
involving glycerol is necessary to reduce osmotic stress from the cryoprotectant, itself. It
was suggested initially it was thought that extended semen exposed to glycerol did not
show any survival differences post-thaw based on exposure time. Higher motilities and
improved morphology were observed after incubating samples for 4 hours (Johnson et al.,
2000). The temperature in which the glycerol is added also affects the post-thaw
outcomes. Vidament and colleagues (2000) found the addition of glycerol at 22°C
53

generated a higher return rate than glycerol added at 4°C. Though trials #1 and #2 had
longer incubation periods at 5°C. the lack of an additional extender may lead to a
metabolism deficiency. Also egg yolk extenders possess protective attributes when
added to spermatozoa during cryopreservation (Medeiros et al., 2002).
In conclusion, refinement to preparative handling techniques of boar spermatozoa
is recommended for the development of an optimal protocol. It should be noted that some
variation within the results may be due to varying thawing procedures. However, our
results suggest that incubation periods with decreasing temperatures, centrifugation and
the addition of freezing extenders demonstrated post-thaw potential.
54

CHAPTER VI
COMPARISONS OF A UNIQUE FREEZING
TIXIJNOLOGY (UFT) TO TRADITIONAL
LIQUID NITROGEN METHODOLOGY FOR
CRYOPRESERVATION OF PORCINE SPERMATOZOA
Abstract
Due to the poor results from current cryopreservation methodology in several
species, especially the boar, new techniques need to be investigated. Therefore our
objective was to investigate newly developed freezing techniques using a new Unique
Freezing Technology (UFT) with the intent of improving current fertility outcomes. Four
ejaculates from four different boars were used to compare the results of a UFT to those
from traditional liquid nitrogen methods on the cryopreservation of boar semen. Though
treatment and control groups were handled similarly during pre- and post-freezing
procedures, the samples were divided into freatment groups immediately before freezing.
The effects of the freezing freatments demonstrated significantly different post-thaw
motilities (P<0.02). As expected there was a significant difference between freezing
treatments as evaluated through original motility percent retums (P<0.05). When
assessing the individual UFT techniques, all of six UFT techniques had significantly
higher post-thaw motilities (P<0.05) and original motility percent retums (P<0.05)
compared to either the control vial or straw groups. The preliminary data suggests that
the UFT could improve current cryopreservation fertility results of boar semen.
55

Introduction
The use of artificial reproductive techniques in the swine industry has increased
greatly within the past twenty years. It is estimated that 45-50% of all sows in the United
States are bred by artificial insemination, and of those inseminations 99% are extended
for same day use or cooled for 1-5 days (Johnson et al., 2000). However, the capacity of
cryopreservation is very limited due to porcine sperm's high vulnerability to cold shock
(Buhr et al.. 1989; Holt, 1999).
Freezing rates must be fast enough to reduce intracellular ice formation, yet slow
enough to avoid osmotic alterations (Watson, 2000). Currentiy, most boar samples are
frozen as pellets on dry-ice or in maxi-straws in nitrogen vapor (Johnson et al., 2000).
The procedures are very similar to those used thirty years ago (Larsson, 1978). This
produces a dramatic temperature change from the holding temperature of 5°C degrees to
that of liquid nitrogen, -196°C. Medrano and colleagues (2002) demonstrated higher
rates of membrane damage at a faster freeze rate of 24°C/minute versus 3°C-12°C/minute
while cooling to -5**C. Thurston et al. (2003) showed higher post-thaw viabilities and
motilities in boar sperm which were cooled at 6°C/minute to -5°C. then froze at higher
rate of-40°C/minute to -80°C.
Within the last few years, a new freezing system has developed for freezing
foodstuffs with an increased ability to dissipate heat. This unique freezing technology
(UFT) has a relatively high dissipation rate which appears to vitrify water and preserve
freshness (SupaChill®, SupaChill USA; Lubbock, TX). Due to the unique freezing
capacity, the temperature can be adjusted at range of freezing levels below 0°C.
56

Preliminary trials on various cell and tissue types have shown very promising results.
Our objective was to compare the effects of freezing swine spermatozoa using this UFT
and traditional nitrogen methods in attempt to create a more optimal standardized
cryopreservation technique.
Materials and Methods
Freezing Technology
A new unique freezing technology (UFT) was utilized in this study. SupaChill®
(SupaChill USA; Lubbock, TX) incorporates an organic freezing fluid with a heat index
similar to water; thus allowing for a high rate of heat dissipation. The freezing fluid of
UFT has the unique ability to be set at temperature levels below 0°C (freezing point) with
out solidifying. To evaluate the rate of freezing of the UFT used, samples of water and
the two egg-yolk freezing extenders used in this study were placed in 1.8 mL cryo-tubes
(Nalge Nunc Intemational; Denmark) and froze in the -25°C UFT unit. Temperatures
inside each vial were monitored using Thermocouple Thermometer (Cole-Parker
Instrument Company; IL). Data was collected every 10 seconds for 9 minutes.
Post-collection Processing
Four ejaculates from four boars were collected during four separate occasions.
All the boars were collected at the Texas Tech University Farm at New Deal, Texas by
trained personnel. After each collection, the ejaculate was evaluated for total volume,
gel-free volume, concentration and initial motility. To meet the standards of the
57

experiments the original motility of all samples needed to be equal or greater than 60%
and the concentrations had to be consistent with farm breeding practices. A sample was
extt^cted and extended 2:1 with a tris-egg yolk extender (X Cell Extender, FVM; Maple
Grove. MN) and packaged for transport to one of two off-site laboratories for ftirther
processing.
Cryopreservation Processing
Upon arrival at either laboratory, each sample was placed into a 17°C refrigeration
unit. The sample was held for one hour at this temperature after which a pre-cooled 17*'C
wash buffer was added. The sample was held at this temperature for an additional 1.5
hour to equilibrate with the wash buffer. The sample then was centrifiiged to create a
sperm-composed pellet. TTie pellet was then dissipated in a pre-cooled 25% egg-yolk
extender, Boarchipos A (IVM; Maple Grove, MN (Ext #1)) at a 1:1 ratio with the original
sample size. The sample was placed in a 5°C refrigeration unit for 1.5 hour before a
secondary freezing extender, Boarchipos B (FVM; Maple Grove, MN (Ext #2)), was
added in three increments over 30 minutes. This 25% egg-yolk extender with 6%
glycerol was also pre-cooled to 5°C. Following the addition of the last increment, the
sample was held at 5°C for one hour before freezing. The sample was not broken into
confrol and treatment groups until immediately before freezing to ensure consistent
handling.
58

Freezing Methodology
Preliminary trials produced significant improvements in post-thaw outcome when
comparing the 1.8 ml cryo-tubes (Nalge Nunc Intemational; Denmark) to the 0.5 ml
straw in the UFT. Therefore, one ml aliquots were extracted from the extended sample
and placed into cryo-tubes for freezing in the UFT and control C groups. Additional
samples were also exfracted and placed in 0.5 mL straws for control S group. Eight
freezing treatments were tested for this trial, six UFT and two controls. Preliminary trials
demonstrated higher results with the selected six UFT treatments:
B: 4 minutes at-25°C
C: 4 minutes at -25°C with a proprietary filter #1
E: 4 minutes at -25°C with a proprietary filter #3
F: 5 minutes at -10°C. tiien 4 minutes at -25°C
G: 5 minutes at -10°C then 4 minutes at -25°C with a proprietary filter #1
J: 5 minutes at -10°C with a proprietary filter #1 then 4 minutes at -25°C with a
proprietary filter #1
Con C (Cryo-tubes): 10 minutes in the liquid nitrogen mist
Con S (Straws): 10 minutes in the liquid nifrogen mist.
After initial freezing was complete, all cryo-tubes and straws were stored in liquid
nitrogen.
59

Thawing Methodology
The thawing method used is based on previous trials performed in our laboratory.
Samples were stored at -196 °C for at least 48 hours before being evaluated for post-thaw
values. Each sample was thawed on an individual, random basis by a trained observer.
The thawing protocol involved a 45 second duration at room temperature, then 5 minutes
in a 37°C water bath. Each vial was manually assessed immediately following thaw.
Statistical Analysis
All data from the UFT thermal gradient evaluation was analyzed using a linear
regression model. Based on a best fit linear estimate, the average slope was considered to
represent the rate of temperature change.
Tbe analyses of the motilities, both post-thaw and original retum percentages,
from the eight freezing treatments were analyzed by ANOVA using the general linear
model (GLM) of tiie Statistical Program of the Social Sciences (SPSS Version 8.0; SPSS,
Inc.; Chicago, IL). Post-thaw motility averages and percentages of original motility
retumed averages of the individual freezing techniques were separated through the Post
Hoc fimction of Fisher's Least Squared Differences (LSD).
Results
UFT Thermal Gradient
Data from the three medias froze are represented in Figure 6.1. The average
temperature change was -6.1°C/minute.
60
• Water
-Ext# l
• Ext #2 E
B H
1 4 7 10 13 16 19 22 25 28 31 34 37 40 43 46 49 52 55
Time (in 10 sec increments)
Figure 6.1 Cooling Rates of the UFT
Freezing Treatment Comparison
These preliminary results reflect the effects of freezing techniques on the post-
thaw motilities from each treatment group. Referring to Figure 6.2, the significant
differences between post-thaw motilities of freezing technique treatments is shown.
There was a significant difference between freezing techniques (P<0.02). Using the LSD
procedure, averages post-thaw motilities of the individual freezing techniques were
separated.
61
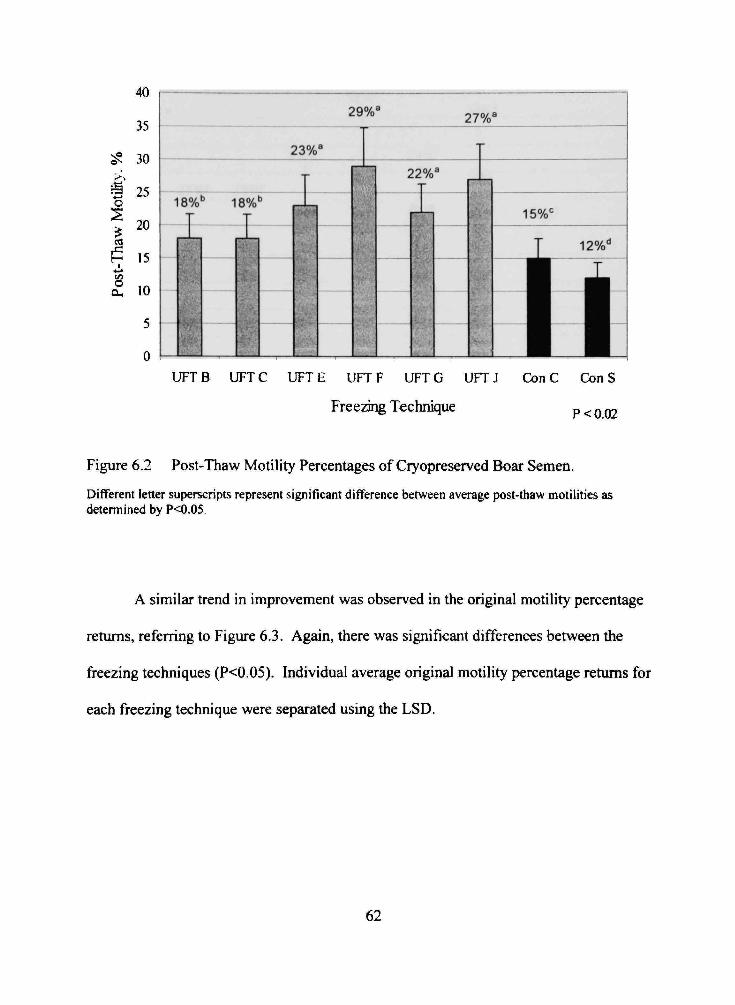
UFTB UFTC UFTE UFTF UFT G UFTJ Con C Con S
Freezing Technique p < o 02
Figure 6.2 Post-Thaw Motility Percentages of Cryopreserved Boar Semen.
Different letter superscripts represent significant difference between average post-thaw motilities as determined by P<0.05
A similar trend in improvement was observed in the original motility percentage
retums, referring to Figure 6.3. Again, there was significant differences between the
freezing techniques (P<0.05). Individual average original motility percentage retums for
each freezing technique were separated using the LSD.
62
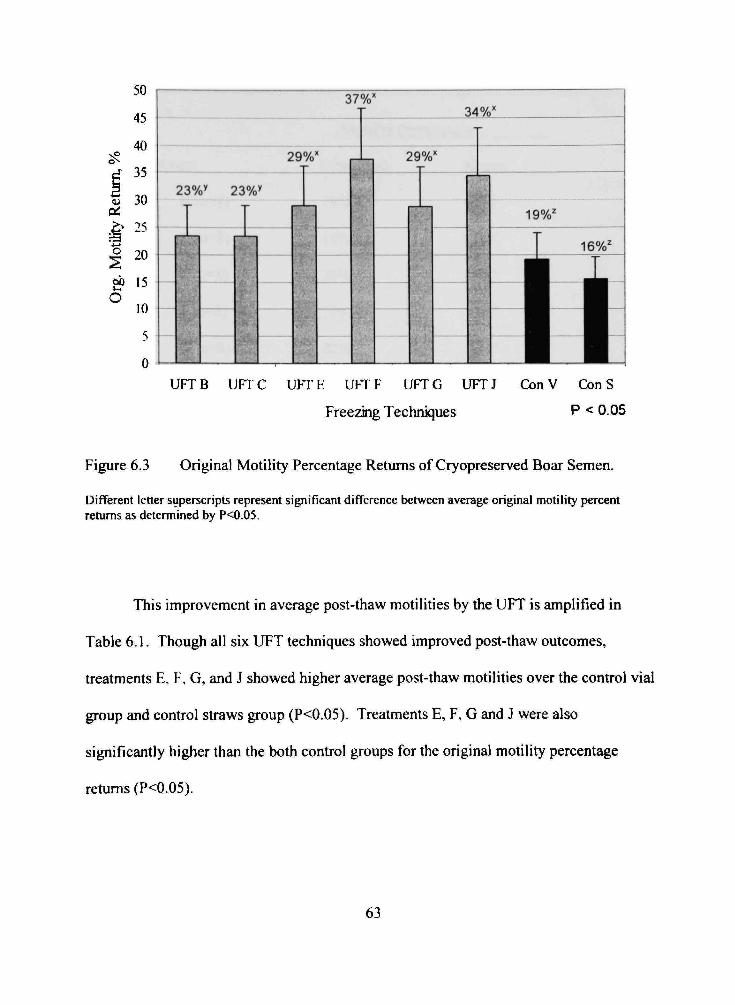
50
45
40
35
u 30
I 20 w) 15 O
10
23%>' 23%>'
29%"
19%'
UFTB UFTC UFTE UFTF UFFG UFTJ Con V Con S
Freezing Techniques P < 0.05
Figure 6.3 Original Motility Percentage Retums of Cryopreserved Boar Semen.
Different letter superscripts represent significant difference between average original motility percent retums as determined by P<0.05.
This improvement in average post-thaw motilities by the UFT is amplified in
Table 6.1. Though all six UFT techniques showed improved post-thaw outcomes,
treatments E, F, G, and J showed higher average post-thaw motilities over the control vial
group and control straws group (P<0.05). Treatments E, F, G and J were also
significantly higher than the both control groups for the original motility percentage
rehims (P<0.05).
63

Table 6.1 Post-Thaw Motility Outcomes from Cryopreserved Boar Semen.
Treatment
B (n=8)
C (n=8)
E (n=8)
F(n=8)
G (n=8)
J (n=8)
Con Vials (n= =6)
Con Straws (n=6)
Motility Percentages
Post-Thaw
18%"
18%"
22%"
29%^
22%''
27%'
15%'
12%"
Retum
23%''
23%y
29%"
37%"
29%"
34%"
19%'
15%'
Different letter superscripts (a,b,c,d) represent significant difference between average post-thaw motilities as determined by P<0.05 Different letter superscripts (x,y,z) represent significant difference between average original motility percent retums as determined by P<0.05.
Discussion
With the low success of cryopreservation in the swine industry, new techniques must
be investigated. Commonly, damage is observed through reduced pregnancy and motility
rates produced in part to the freezing rate of liquid nifrogen vapor at 20°C/minute. Our
preliminary data suggests a slower pre-freeze phase below the freezing point before
acceleration to liquid nitrogen temperatures. Our data is in agreement with two other
reports demonstrating a step-down cooling below freezing from 5°C (Medrano, 2002;
64

Thurston et al., 2002). By nearing the point of intracellular ice formation at a slower rate
of 4°C/minute. the water is cooled to a stable transition point. Then the freezing rate is
accelerated by the liquid nitrogen mist, thus reducing ice formation.
Therefore, the UFT could potentially be considered as an altemative to replace the
current traditional liquid nifrogen methods. These improvement trends on post-thaw
results using the UFT have been also reported in similar UFT trials involving other
species and cell lines. However, ftirther studies need to be conducted to refine a standard
technique. Additional testing is needed to evaluate the effects of UFT on boar sperm
viability and structural integrity as well as pregnancy rates.
65

CHAPTER VII
LIPID PEROXIDATION EFFECTS FROM SEMEN
HANDLING DURING CRYOPRESERVATION OF
PORCUSIE SPERMATOZOA
Absfract
The potential advantages of sperm cryopreservation have not been fiiUy
accomplished due to the limiting detrimental effects on sperm stmcture and composition.
Previous studies have suggested that cells suffer lipid peroxidation damage during the
cryopresevation process, specifically suggesting the damage results from mechanical
stress during the preparatory and freezing processes. In this present study, sperm samples
were analyzed for lipid stability throughout sample processing through evaluations for
lipid peroxidation and lipid free radical amounts. Our analysis was completed in three
experiments. In Experiment 1, lipid stability levels were gained from five separate boar
ejaculates frozen using three different freezing methods to compare cryopreservation
techniques. In Experiment 2, lipid p)eroxidation amounts for fresh post-ejaculate and
albumin extended boar samples were compared. Experiment 3 involved evaluations of
the semen processing to evaluate sample and seminal fluid alterations. Samples tested
from the freezing protocol included fresh, extended, addition of a wash buffer, cooling to
]7°C, centrifugation, addition of two egg-yolk extenders, cooling to 5°C and post-thaw
values. Though there was no difference between the three freezing freatments, significant
differences were noted between the fresh and extended samples (P<0.001). These
66

findings were exemplified by the step by step analysis of the processing and freezing
protocol. The lipid peroxidation amounts accumulated after each the procedural step
(P<0.001). Significant differences were also observed in the lipid radical levels
(P<0.001). The largest gains of both lipid parameters developed after the addition of an
egg-yolk freezing extender. The results of the pre-freezing protocol, alterations in lipid
stability do not appear to be due to thermal or mechanical sfress. The results suggest
ftirther studies in alterative extenders need to be explored.
Introduction
In lieu of the different success rates of the varying freezing methodologies, this
study was performed to investigate the possible source of damage on a molecular level.
More specifically, we evaluated the lipid stability of the phospholipid membrane during
the freezing and pre-freezing process.
Several articles have been published based on lipid peroxidation and oxygen free
radicals in human and murine samples. Negative correlations have between associated
between increased levels of malondialdehyde (MDA) reduced motility (Engel et al.,
1999; Huang et al., 2001; Verma and Kanwar, 1999) and poor fertilization capability
(Zabludovsky et al., 1999) in human sperm. Though conclusions are made from trials
based on other species, species differences in composition have demonstrated the
presence of varying protective enzymatic mechanisms against peroxidation (Alvarez and
Storey, 1984; Medeiros et al., 2002; Mennella and Jones, 1980).
67

Very few articles have been published in relation to boar semen. Comaschi and
colleagues (1989) suggested the lipid radicals were more detrimental than the radical
hydroxyls. MDA levels have been identified in samples incubated at 37*'C (Comaschi et
al., 1989), cooled liquid storage (Cerolini et al., 2000) and post-thaw (Cerolini et al.,
2001).
Though peroxidation levels have been analyzed in previous reports specifically
studying the post-collection and post-thawed samples, the evaluation of the possible rate
during pre-freezing procedures has not been investigated. The objective was to
investigate the rate of lipid peroxidation of the porcine spermatozoa and amount of lipid
peroxidation and lipid free radicals during the cryopreservation process.
Materials and Methods
Semen Preparation
Semen specimens were collected artificially from boars at the Texas Tech
University New Deal Farm by trained staff. During the post-collection evaluation,
standard measurements including total gel-free volume, concentration, and motility were
obtained. To fulfill standard breeding practices, all samples used possessed a minimum
motility rate of 60% post-collection and met the breeding requirements of the swine urtit.
From the filtered ejaculate, a 30 mL sample was retrieved and extended with 60 mL of a
pre-heated X-Cell extender (IVM; Maple Grove, MN). The sample was then packaged
for transport to one of two possible laboratories.
68

Experiment #1
Upon arrival, the extended sample was placed in a refrigeration unit (17°C) for
one hour. At the end of the hour, 210 mL of a wash buffer maintained at 17°C was added
before continuing the storage in refrigeration unit for another 1.5 hours. The samples
were tiien centrifuge for 20 minutes at 1900-2000 rpm in a 15°C setting. The supemant
fluid was removed and discarded. The remaining pellets were reconstituted in 30 mL of a
25% egg-yolk extender, Boarciphos A (IVM; Maple Grove, MN), also maintained at
17°C. The sample was allowed to equilibrate for 1.5 hours in a secondary refiigeration
unit set at 5°C. With 10 minute intervals between, a second 25% egg-yolk extender with
6% glycerol, Boarciphos B (FVM; Maple Grove, MN), cooled to 5°C, was added three
times in 10 mL increments. After all of the 30 mL were added, the sample was
maintained at 5°C for one hour. One ml aliquots of the extended sample were placed in
1.8 mL labeled cryo-vials (Nalge Nunc Intemational; Denmark) before being
cryopreserved. Through labeling, specimens were separated into three freezing treatment
groups:
Control - liquid nifrogen mist for 10 minutes
F: 5 minutes at -10°C followed by 4 minutes at -25°C
J: 5 minutes at -10°C followed by 4 minutes at -25°C within a proprietary fluid
filter in each unit.
Freezing treatments "F" and "J" were two optimal freezing freatments of the UFT
(SupaChill®, SupaChill USA; Lubbock, TX) justified in previous trials within our
69

laboratory. Samples were plunged into liquid nitrogen and stored until testing began for
lipid peroxidation rates and lipid free radical amounts.
Experiment #2
For Experiment #2, we used fresh post-ejaculate boar samples and extended post-
ejaculate samples for lipid stability comparisons of five samples from five boars. The
above protocol discussed in semen processing was utilized and completed until arrival at
the off-site lab. One mL aliquots were exfracted from all ten samples and placed in 1.8
mL cryo-tubes (Nalge Nunc Intemational; Denmark). Immediately afterwards, each vial
was plunged in liquid nitrogen for freezing and storage.
Experiment #3
The third phase of this trial utilized the same protocol discussed above in
Experiment #1 on three semen samples from separate boars. However, the proportions of
original semen sample and the various extenders used were increased. This ensured
sufficient amounts for three 1 mL aliquots of semen samples and fluid to be extracted
during each major step of the protocol without influencing final ratios. The semen
samples represent the whole fractions available at that step. Fluid samples were created
by centrifuging addition samples during each step specified below. The procedural steps
represented include:
1. Ejaculation samples post-transport,
2. Extended samples post-transport.
70

3. Extended samples cooled to 17°C for 1 hour,
4. Samples equilibrated with a wash buffer at 17°C for 1 Vi hours,
5. Samples after centrifiigation for 20 minutes at 2500 rpm,
6. Samples equilibrated with the first freezing extenders at 5°C for 1 '/z hours,
7. Samples equilibrated with the second freezing extenders at 5°C for 1 Vi hours,
8. Post-thaw values of optimal freezing treatment.
Due to the density of the semen pellet after centrifugation and concems of altering final
concentrations, a semen sample was not exfracted for this step. While thawing the
samples, fluid specimens were not removed for analysis. After exfraction during each
step, the aliquots were plunged and stored in liquid nifrogen.
Quantification of Lipid Peroxidation
After thawing at room temperature and then being vortexed, a 200 pL sample was
extracted from each aliquot and utilized for this test. The semen samples were mixed
witii 200 pL of SDS solution, 1.5 mL of 20% AA solution (at 3.5 pH), 1.5 mL TBAR
solution (1.6 g 2-thiobaritric acid with 200 mL of deionized water) and 600 pL of
deionized water. After vortexing each sample, the samples were incubated in a 100**C
water bath for 2 hours. The samples were then cooled for 10 minutes at room
temperature before centrifuged at 2000 rpm for 10 min. A 0.5 mL sample of fluid was
again extracted from each tube and mixed with 2 mL of butanol. After a second
centrifugation at 2500 rpm for 10 minutes, each fluid sample was transferred into a glass
71

absorbance box one at a time for analysis at 532 nm wavelength in a spectrophometer
(Coleman 575, Perkin-Elmer; Oak Brook, IL).
Quantification of Free Lipid Radicals
A 250 pL sample was extracted from each specimen and placed into a pre-labeled
12 X 75 mm glass tube. After diluting each sample in 2 mL of a 2:1 chloroform:heptane
mixture, each tube was thoroughly vortexed. The samples were then centrifiiged for 10
minutes at 2500 rpm. Fluid in the amount of 700 |jL was extracted from each specimen
and transfer to a new set of glass tubes. Samples were dried under a flow of normal afr at
ambient temperatures for 15-20 minutes. The dried extracts were reconstituted in 2 mL
of heptane before performing a spectrophotometer (Coleman 575, Perkin-Elmer; Oak
Brook, IL) analysis at a 233 nm wavelength. The lipid radical confrol, requfred for
proper calculations, was processed the same as a sample except the confrol contained 250
pL of water instead of a sample.
Statistical Analysis
The analysis of the lipid peroxidation and lipid free radical amounts for the three
freezing freatments was analyzed using analysis of variance (ANOVA). Boar and
freezing treatment were both used as part of the statistical model. The means were
separated through Fisher's Least Squared Differences (LSD).
72

ANOVA was also used to compare the results of the fresh and extended values for
lipid peroxidation. Though the results were significant, the sample size is too small to
complete a LSD analysis.
On the procedural analysis in Experiment #3, the lipid peroxidation and lipid free
radical amounts were also analyzed using the ANOVA model. Since these values
represent accumulations of lipid instability over time, a linear regression model function
was used to analyze each parameter.
All statistical analyses were performed using the SPSS Version 8.0 Software
(SPSS, Inc.; Chicago, IL).
Results
Experiment # 1
When comparing the three freezing treatments using five separate semen samples,
the differences between the treatments were not significant for either lipid evaluations.
However, there was an interaction between the boar and lipid peroxidation amounts
(P<0.005). These differences, shown in Table 7.1, concur with the variability of semen
quality in-between boars.
73
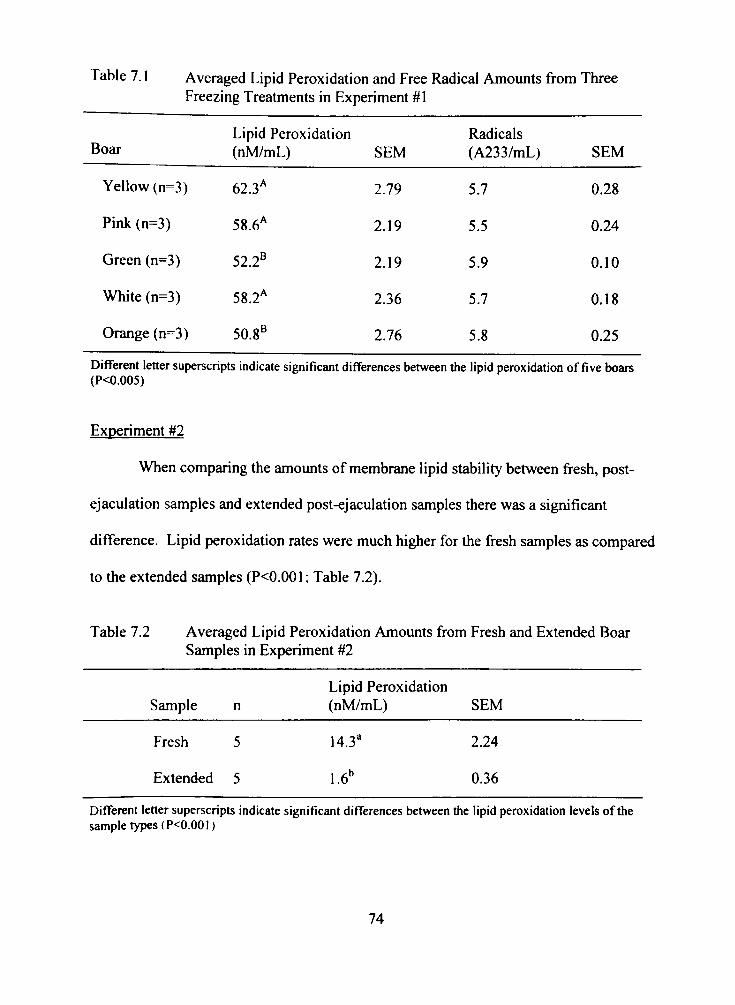
Table 7.1 Averaged Lipid Peroxidation and Free Radical Amounts from Three Freezing Treatments in Experiment #1
Boar
Yellow (n=3)
Pink (n=3)
Green (n=3)
White (n=3)
Orange (n=3)
Lipid Peroxidation (nM/mL)
62.3^
58.6^
52.2^
58.2^
50.8^
SEM
2.79
2.19
2.19
2.36
2.76
Radicals (A233/mL)
5.7
5.5
5.9
5.7
5.8
SEM
0.28
0.24
0.10
0.18
0.25
Different letter superscripts indicate significant differences between the lipid peroxidation of five boars (P<0.005)
Experiment #2
When comparing the amounts of membrane lipid stability between fresh, post-
ejaculation samples and extended post-ejaculation samples there was a significant
difference. Lipid peroxidation rates were much higher for the fresh samples as compared
to tiie extended samples (P<0.001; Table 7.2).
Table 7.2 Averaged Lipid Peroxidation Amounts from Fresh and Extended Boar Samples in Experiment #2
Lipid Peroxidation Sample n (nM/mL) SEM
Fresh 5 14.3' 2.24
Extended 5 1.6" 0.36
Different letter superscripts indicate significant differences between the lipid peroxidation levels of the sample types(P<0.001)
74

Experiment #3
Referring to Table 7.3, lipid peroxidation levels accumulated during the handling
procedure in both specimen types evaluated. The levels of peroxidation were
significantly different within the semen samples (P<0.001) and fluid samples (P<0.001).
Using the LSD procedure, significant differences were observed at various fransition
points of the procedure including steps 1 to 2, steps 4 to 6, and steps 7 to 8 in the sample
(P<0.05) and fluid (P<0.001).
Significant changes were found during the duration of the cryopreservation
protocol. Amounts of MDA were correlated between the semen and fluid levels for both
the lipid peroxidation (r=0.96). These increases in peroxidation rates are emphasized in
Figure 7.1.
75
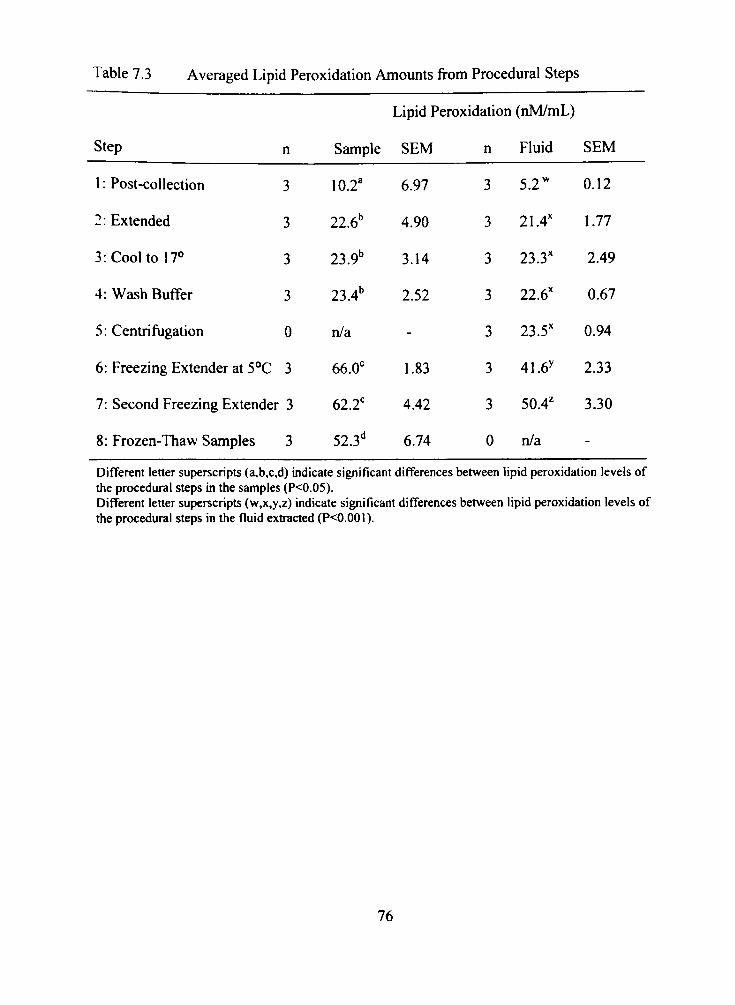
Table 7.3 Averaged Lipid Peroxidation Amounts from Procedural Steps
Lipid Peroxidation (nM/mL)
Step n Sample SEM n Fluid SEM
1: Post-collection
2: Extended
3: Cool to 17"
4: Wash Buffer
5: Centrifugation
6: Freezing Extender at 5°C
3
3
3
3
0
3
7: Second Freezing Extender 3
8: Frozen-Thaw Samples 3
10.2'
22.6"
23.9"
23.4"
n/a
66.0'
62.2'
52.3"
6.97
4.90
3.14
2.52
-
1.83
4.42
6.74
3
3
3
3
3
3
3
0
5.2^
21.4"
23.3"
22.6"
23.5"
41.6^
50.4"
n/a
0.12
1.77
2.49
0.67
0.94
2.33
3.30
-
Different letter superscripts (a,b,c,d) indicate significant differences between lipid peroxidation levels of the procedural steps in the samples (P<0.05). Different letter superscripts (w,x,y,z) indicate significant differences between lipid peroxidation levels of the procedural steps in the fluid extracted (P<0.001).
76
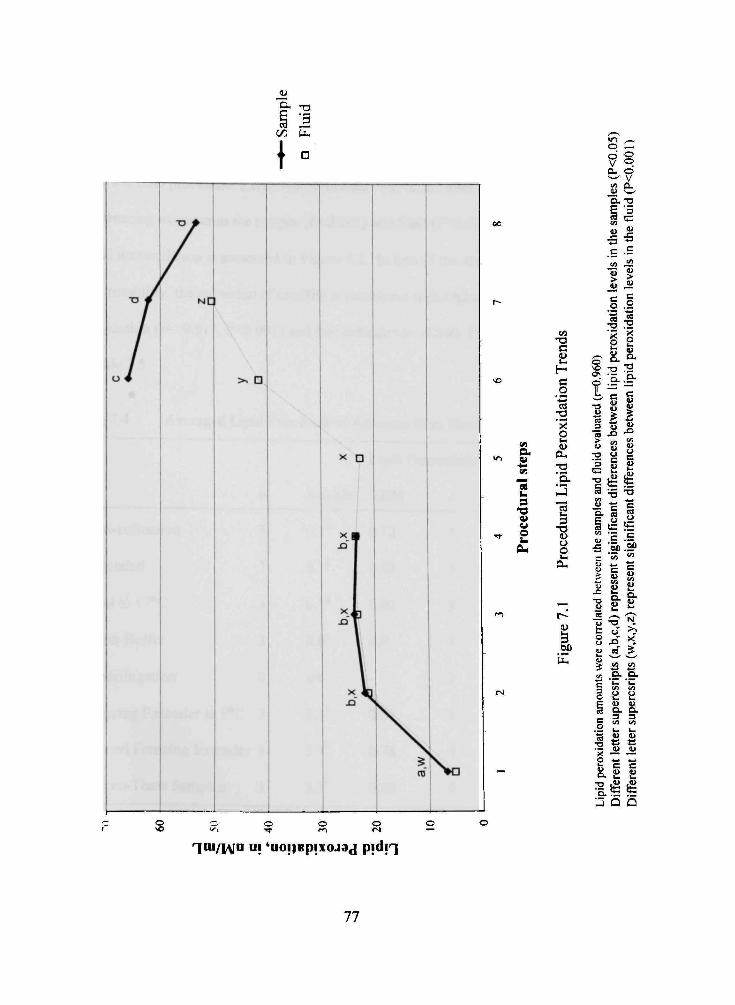
_1>
I D
NCI
>. D
X D
00
r
\o
en
a en
^ U 3
- £ a.
r*
CM
—~
CO
•a c
tion
T
CO
•o 'S o
Per
•;a
Lip
ra
l oc
edu
i i
a.
_
r-
&p tl!
S'
(r=0
.96
• o
ate
3 a
idev
3
and
pies
th
esam
c
>
ted
bet
ela
corr
u
wer
Vi
c 3 9U
JBI
o
X o u D.
' a J
m p O V CL
C/1
a>
^ . r~m
o O O V Cu •..-•
Q. ia E ra
leve
ls i
n tli
e s
datio
n iro
xi
5en
lipi
d et
w(
.£1
nces
V
i •o s
igin
ific
en
c u
repr
es
T3^
xT CO
tn
g. *C (n u
uper
en b .
u ts u 4_(
c
3 C
leve
ls i
n th
e id
atio
n
X
ero
o.
en l
ipid
V
B 4>
^
:nce
s
•5
;ant
;ig
inifi
c
VJ
C
in
repr
e:
x"
£ en
Q . ' i -en o
uper
en t -V
c u 4_(
c £
j£iS •Q Q
o
i u / | ^ a Ul *uo!|Bp!XOJ3j pfdiq
77
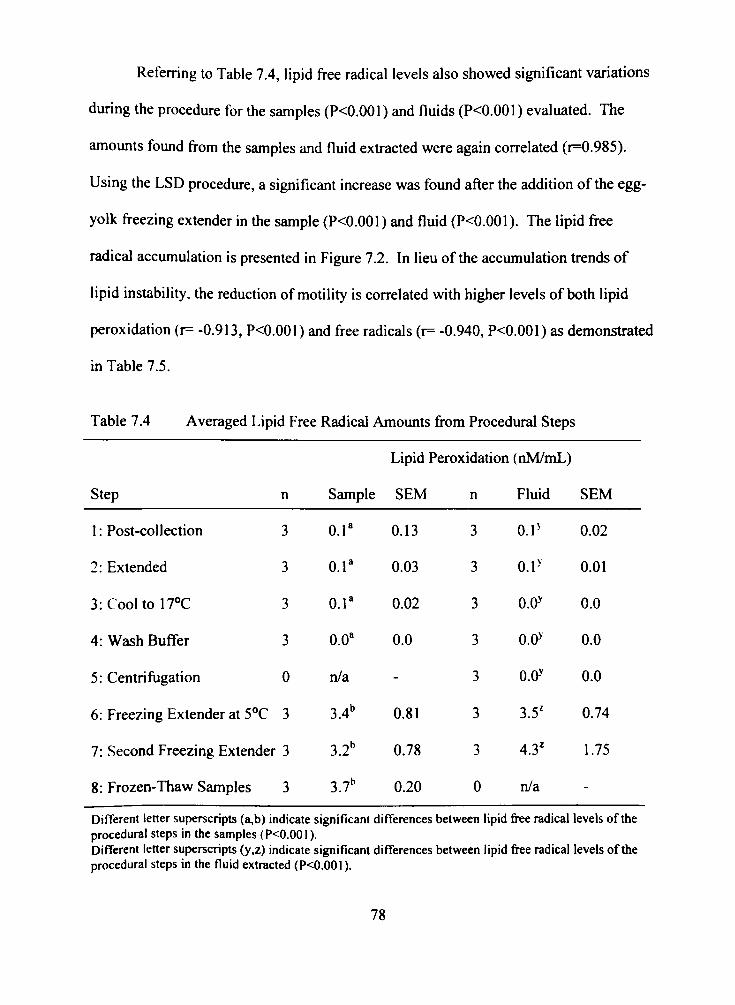
Referring to Table 7.4, lipid free radical levels also showed significant variations
during the procedure for the samples (P<0.001) and fluids (P<0.001) evaluated. The
amounts found from the samples and fluid extracted were again correlated (r=0.985).
Using the LSD procedure, a significant increase was found after the addition of the egg-
yolk freezing extender in the sample (P<0.001) and fluid (P<0.001). The lipid free
radical accumulation is presented in Figure 7.2. In lieu of the accumulation trends of
lipid instability, the reduction of motility is correlated with higher levels of both lipid
peroxidation (r= -0.913, P<0.001) and free radicals (r= -0.940, P<0.001) as demonstrated
in Table 7.5.
Table 7.4 Averaged Lipid Free Radical Amounts from Procedural Steps
Step
Lipid Peroxidation (nM/mL)
Sample SEM n Fluid SEM
1: Post-collection
2: Extended
3:Cooltol7*'C
4: Wash Buffer
5: Centrifugation
6: Freezing Extender at 5°C
7: Second Freezing Extender
8: Frozen-Thaw Samples
3
3
3
3
0
3
3
3
0.1"
0.1"
0.1"
0.0"
n/a
3.4"
3.2"
3.7"
0.13
0.03
0.02
0.0
-
0.81
0.78
0.20
3
3
3
3
3
3
3
0
O.P
O.P
0.0^
0.0^
0.0''
3.5^
4.3^
n/a
0.02
0.01
0.0
0.0
0.0
0.74
1.75
-
Different letter superscripts (a,b) indicate significant differences between lipid fi^e radical levels of the procedural steps in the samples (P<O.OOI). Different letter superscripts (y,z) indicate significant differences between lipid free radical levels of the procedural steps in the fluid extracted (P<0.001).
78
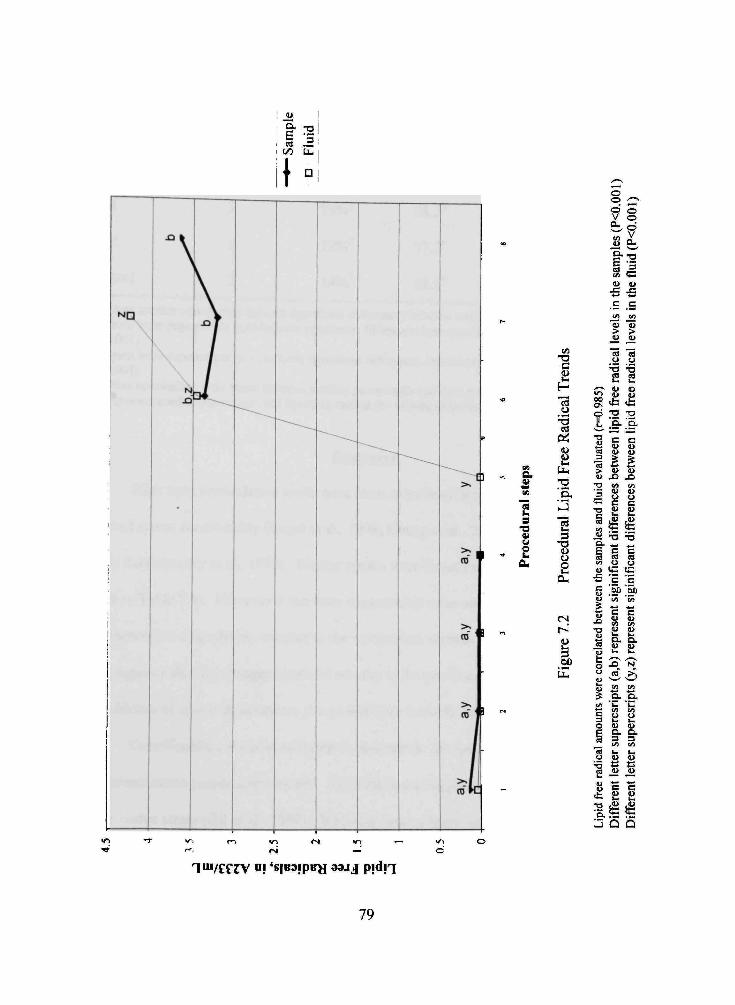
D.
ee
ral
edu
w p La
su
C/1 T3
ren
H •3 o
^ CO
D!i u
Fre
T3 • Q .
J
lora
l -a u o 2
CL,
<N
r
op
^..v
985
o l^
ted
(S
valu
u ;a 'S
les
and
fl
o. p n en U
J =
c u s u
X i
elat
i co
n
u
5 en C 3 o
"5 _o • a CS 1 -
u 4> *
pid
- ]
o o ^ ?? «= V o - ^ OJ CL
sam
p fl
uid
u u •S S •S .£ en en
u a> > > <U V
"to "5 u u
radi
ra
di
u u <£<£ T 3 T3
!§• !§• C C U 1> U U
a> o ^ X ) CA CO
diff
e di
ffe
•fc- 4-t
c c n es o t>
<a «= 's S '5b "ab en en * j 4..1
c c eo en
& . CL
X> N
V3 CA 4—* ^ J
. £ • .£ • *c *c en en U U »- b. u u CL CL 3 3 en en b. b. U V
ts e 9> U
C B
£ £ ^^ Q Q
Tui/ff jv "! *s|B3!PBy aajj pidiq
79
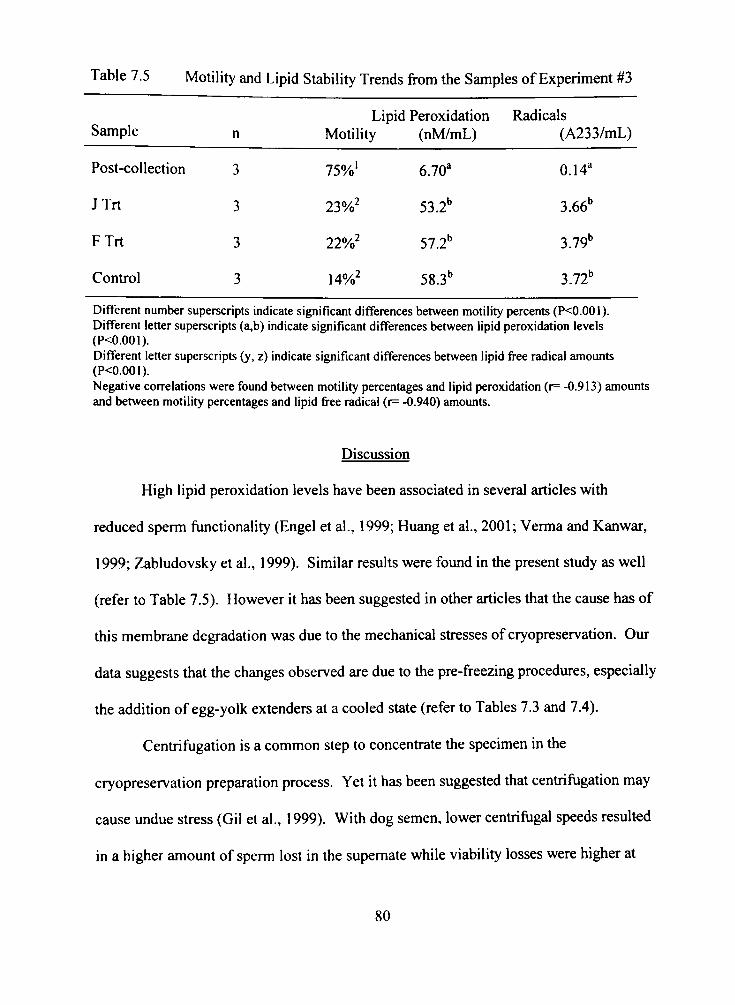
Table 7.5 Motility and Lipid Stability Trends from the Samples of Experiment #3
Sample
Post-collection
JTrt
FTrt
Control
n
3
3
3
3
Lipid Peroxidation Motility (nM/mL)
75%'
23%^
22%^
14%^
6.70"
53.2"
57.2"
58.3"
Radicals (A233/mL)
0.14^
3.66"
3.79"
3.72"
Different number superscripts indicate significant differences between motility percents (P<0.001). Different letter superscripts (a,b) indicate significant differences between lipid peroxidation levels (P<0.001). Different letter superscripts (y, z) indicate significant differences between lipid free radical amounts (P<0.001). Negative correlations were found between motility percentages and lipid peroxidation (r= -0.913) amounts and between motility percentages and lipid free radical (r= -0.940) amounts.
Discussion
High lipid peroxidation levels have been associated in several articles with
reduced sperm ftmctionality (Engel et al., 1999; Huang et al., 2001; Verma and Kanwar,
1999; Zabludovsky et al., 1999). Similar results were found in the present study as well
(refer to Table 7.5). However it has been suggested in other articles that the cause has of
this membrane degradation was due to the mechanical stresses of cryopreservation. Our
data suggests that the changes observed are due to the pre-freezing procedures, especially
the addition of egg-yolk extenders at a cooled state (refer to Tables 7.3 and 7.4).
Centrifugation is a common step to concenfrate the specimen in the
cryopreservation preparation process. Yet it has been suggested that centrifugation may
cause undue stress (Gil et al., 1999). With dog semen, lower centrifugal speeds resulted
in a higher amount of sperm lost in the supemate while viability losses were higher at
80

increased centrifugational velocities. Yet, membrane integrity was maintained at the
various speeds (Rijsselaere et al., 2002). Our evaluation of the seminal fluid extracted
before and after centrifugation, show very little difference in lipid stability (refer to Table
7.3 and 7.4). Neild and colleagues (2003) reported a capacitation-like phenomenon after
centrifugation of stallion sperm. Thus, permeability alterations may be due to other
sources than lipid peroxidation.
Protein degradation due to peroxidation has been observed (Medeiros et al.,
2002). Baumber et al. (2002) correlated high peroxidation levels in post-thawed semen
with DNA fragmentation. This may actually be the cause for the reduced fertilization
capability recognized in sperm after cryopreservation (Dobrinski et al., 1995).
As observed in bovine semen, our results showed higher levels of peroxidation in
frozen-thawed semen versus fresh or cooled (refer to Tables 7.3 and 7.4). However, due
to the trends at various points in the procedural analysis, our results do not demonstrate
the stress in relation to cooling and thawing as suggested by Chatterjee and Gagnon
(2001). Instead the largest accumulation of both lipid peroxides and lipid radicals was
observed during the addition of the freezing egg yolk extender (refer to Tables 7.3 and
7.4). Egg yolk contains phosphatidylcholine, similar to that already present in the
membrane of sperm. Though studies have demonstrated the protective effects of egg
yolk (Vishwanath and Shannon, 2000), our results project a negative side. Misik and
colleagues (1994) demonstrated the amplification in MDA levels in egg yolk held at 22°C
compared to 50*'C. The introduction of a diluent with ongoing lipid peroxidation or the
addition of a product, specifically lipid peroxides (LOOH), will enhance lipid
81

peroxidation (Tang et al., 2000). Thus, altemative extenders need to be investigated.
Braun and colleagues (1995) observed an increase in the membrane mtegrity and motility
of frozen stallion semen with the exclusion of egg-yolk. Dacaranhe and Terao (2001)
observed reduced iron-peroxidation levels with the presence of phosphatidylserine. A
trial on frozen stallion semen has already verified an improvement in motility with the
addition of phosphatidylserine (Wilhem et al., 1996).
The addition of antioxidants, such as Vitamin E, may need to be incorporated into
breeding schemes to reduce lipid peroxidation rates. Increases in semen quality have
been observed in turkey (Donoghue and Donoghue, 1997), boar (Brzeezinska-
Slebodzinska et al., 1995; Cerolini et al., 2000) and horse samples (Baumber et al., 2002)
with antioxidant supplementation. Dietary antioxidant supplementation has also
increased semen quality in boars (Brzeezinska-Slebodzinska et al., 1995) and roosters
(Neuman etal., 2002).
82

CHAPTER Vlll
CONCLUSIONS
Numerous groups have searched for an optimal freezing protocol for semen
cryopreservation. Their studies have combination of factors, including: cooling rates,
extenders, and packaging systems. Conversely, these comparisons are performed on a
limited basis with small animal samples. Though the final solution will include
interactions between cooling rates and cryoprotectant amounts and osmotic pressures,
consideration must be taken for what we do and do not understand. With the large
amounts of variation currently in both the literature and commercially practiced
cryopreservation protocols, this assessment is difficult to ascertain (Day, 2000; Samper,
2002; Samper and Morris).
Cryopreservation procedures are currently riddled with long standing tradition.
For example, pelleting initial began in the cattle industry over thirty years ago (Larsson,
1978) and has been replaced by liquid nitrogen and freezing straw storage (Vishwanath
and Shannon, 2000). Even though contamination and identification problems have
become a concern (Pickett et al., 1978), this technique is still used today in the swdne
industry (Johnson et al., 2000). In an attempt to improve boar freezing techniques, we
tested a unique freezing technology (UFT). Our investigation revealed a higher post-
thaw motility yields from UFT than those produced by traditional nitrogen freezing
methods on boar sperm.
83

Membrane integrity problems associated to cryopreservation have been attributed
to mechanical stresses such as rapid centrifugational velocities (Gil et al., 1999; Neild et
al.. 2003; Rijsselaere et al., 2002) or rapid freezing rates (Curry and Watson, 1994;
Pommer et al.. 2002). At extreme rates, we admit these areas can cause significant
damage. However, our lipid peroxidation and lipid radical data instead suggests that egg
yolk extender may actually cause the weakening of the membrane while using acceptable
levels of mechanical stress. Yet, egg yolk is the mostly commonly used a freezing
extender for mammalian sperm (Medeiros et al., 2002; Vishwanath and Shaimon, 2000).
Attention also needs to be given to preparative handling techniques. There have
been recommendations for a series of osmotic steps (Curry and Watson, 1994; Gao et al.,
1993) such as thermal reductions to be included in a protocol. Our step-down technique
utilized incubation periods in cooling environments demonstrated significant
improvements on both stallion and boar samples. This is in agreement with other studies
involving stallions (Bruemmer et al., 2002; Crockett et al., 2001) and boars (Eriksson et
al. 2001; Pursel et al., 1972). In spite of these findings, Johnson and colleagues (2000)
reported that one of the two most used protocols in commercial boar settings utilizes only
the addition of a diluent before freezing.
Tliough certain factors are specie specific, our trials applied techniques normally
used for species other than on those that were tested. Both the stallion and boar have
been shown to have significant problems with cold shock (Holt, 1999; Johnson et al.,
2000; Pursel et al., 1972; Yoshida, 2000). Yet, we attempted to utilize techniques used on
less thermally sensitive species, such as the human and bull (Vishwanath and Shannon,
84

2000; Watson, 2000). Our results actually showed superior trends using both medias and
packaging systems. Regardless of all attempts, certain challenges may not be overcome
such as infertility. The variability reported in stallions (Katila et al., 2002; Medeiros et
al., 2002; Pattie and Dowsett, 1982) also was apparent in our results.
Several alterations in our studies to current methodology demonstrated higher
trends or significant improvements. The utilization of these outdated techniques has
produced lower pregnancy rates. Though these techniques are not ideal, the blame for
poor fertility results are often placed upon the handlers (Holt,, 2000; Loomis, 2001).
However, without a set of standards to work from, the variation in results will remain to
be high (Day, 2000; Samper, 2002; Samper and Morris, 1998).
The potential preservation offered by semen cryopreservation has not been ftilly
grasped thus far. Currently, cryopreservation is commonly associated with poor fertility
performance, especially in boars (Thurston et al., 2003; Yoshida, 2000) and stallions
(Loomis, 2001; Medeiros et al., 2002). However, our results reflect the potential for
improvement. Additional studies will be necessary to establish the ftill potential of the
UFT in the cryopreservation of semen and to determine if this technology can be
combined with newer preparatory techniques to improve semen quality in species which
have previously been reported as poor candidates for cryopreservation.
85

LITERATURE CITED
Alvarez, J.G. and B.T Storey. 1984. Lipid peroxidation and the reactions of superoxide and hydrogen peroxide in mouse spermatozoa. Biol Repro 30: 833-841.
Amanda, R.P. ad Graham. Spermatozoal Function. 1993. In Equine Reproduction. Edited by A.O. McKinnon and J.L. Voss. Philadelphia: Lea and Febiger. pp. 715-740.
Amoah, E.A. and S. Gelaye. 1996. Biotechnological advances in goat reproduction. J Ani Sci. 75: 578-585.
Arruda, R.P., B.A. Ball, C.G. Gravance, A.R. Garcia and I.K.M Liu. 2002. Effects of extenders and cryoprotectants on stallion sperm head morphometry. Therio 58: 253-256.
Ball, B.A. and A. Vo. 2001. Osmotic tolerance of equine spermatozoa and the effects of soluble cryoprotectants on equine sperm motility, viability, and mitochondrial membrane potential. J Andro. Nov-Dec 22(6): 1061-1069.
Baumber, J., B.A. Ball, J.J. Linfor and S.A. Meyers. 2002. Reactive oxygen species and cryopreservation promote deoxyribonucleic acid (DNA) damage in equine sperm. Therio 58: 301-302.
Bilodeau, J.F., S. Chatterjee, M.A. Sirard and C. Gagnon. 2000. Levels of antioxidant defenses are decreasing in bovine spermatozoa after a cycle of freezing and thawing. Molec Repro Develop 55: 282-288.
Braun, J.. S. Hochi, N. Oguri, K. Sato, and F. Torres-Boggino. 1995. Effect of different protein supplements on motility and plasma membrane integrity of frozen-thawed stallion spermatozoa. Cryobiology. Oct; 32(5):487-492.
Bruemmer, J.E., H. Reger, G. Zibinski, and E.L. Squires. 2002. Effect of storage at 5''C on the motility and cryopreservation of stallion epididymal spermatozoa. Therio. 58: 405-407.
Brzeezinska-Slebodzinska, E., A.B. Slebodzinski, B. Piettas and G. Wieczorek. 1995. Antioxidant effect of vitamin E and gluthathione on lipid peroxidation in boar semen plasma. Biol Trace Elem Res 47: 69-74.
Buhr, M.M., A.T. Canvin, and J.L. Bailey. 1989.Effects of semen preservation on boar spermatozoa head membranes. Gamete Res. Aug: 23(4): 441-449.
86

Byme, G.P.. P. Lonergan, M. Wade, P Duffy, A. Donovan, J.P. Hanarahan and M.P. Boland. 2000. Effect of freezing rate of ram spermatozoa on subsequent fertility in vivo and in vitro. Ani Repro Sci 62: 265-275.
Calvete, J.J., M. Raidia, M. Gentzel, C. Urbanke, L. Sanz, and E. Topfer-Petersen. 1997. Isolation and characterization of heparin- and phosphorylcholine-binding proteins of boar and stallion seminal plasma. Primary structure of porcine pBl. FEBS Lett, Apr 28; 407(2): 201-206.
Cerolini, S., A. Maldjian, F. Pizzi and T.M. Gliozzi. 2001. Changes in sperm quality and lipid composition during cryopreservation of boar semen. Reprod. 121: 395-401.
Cerolini, S., A. Maldjian, P. Sural, and R. Noble. 2000. Viability, susceptibility to peroxidation and fatty acid composition of boar semen during liquid storage. Ani Repro Sci. 58:99-111.
Chatterjee, S. and C. Gagnon. 2001. Production of reactive oxygen species by spermatozoa undergoing cooling, freezing and thawdng. Molec Repro Develep. 59:451-458.
Comaschi. V., L. Linder, G. Farruggin, N. Gesmudo, L. Colombi and L. Masotti. 1989. An investigation on lipoperoxidation mechanisms in boar spermat02x>a. Biochem Biophys Res Commun 158(3): 769-775.
Cordova, A., J.F. Perez-Gutierrez, B. Lleo, C. Garcia-Artiga, A. Alvarez, V. Drobchak, and S. Martin-Rillo. 2002. In vifro fertilizing capacity and chromatin condensation of deep frozen boar semen packaged in 0.5 and 5 ml sfraws. Therio. 57:2119-2128.
Crockett, E.C., J.K. Graham, J.E. Bruemmer and E.L. Squires. 2001. Effect of cooling of equine spermatozoa before freezing on post-thaw motility: preliminary results. Therio. Feb 1 55(3): 793-803.
Curry, M.R. and P.F. Watson. 1994. Osmotic effects on ram and human sperm membranes in relation to thawing injury. Cryobiol 31:39-46.
Day, B.N. 2000. Reproductive biotechnologies: current status in porcine reproduction. Ani Repro Sci. 60-61: 161-172.
Dacaranhe, CD. and J. Terao. 2001. A unique antioxidant activity of phosphatidylserine on iron induced lipid peroxidation of phospholipid bilayers. Lipids 36 (10): 1105-1110.
87

DeLeeuw, F.E., ll.C. Chen, B. Colenbrander and A.J. Verkleij. 1990. Cold induced ultrastructural changes in bull and boar sperm plasma membranes. Cryobiol 27(2): 171-83.
DeJamette, J.M., D.A. Barnes and C.E. Marshall. 2000. Effects of pre and post thaw thermal insults on viability characteristics of cryopreserved bovine semen. Therio 53: 1225-1238.
Devireddy, R.V., D.J. Swanlund, T.LOlin, W. Vincente, M.H.T. Troedsson, J.C. Bischof and K.P. Roberts. 2002. Cryopreservation of equine sperm: optimal cooling rates in the presence and absence of cryoprotective agents determined using differential scanning calorimetry. Biol Repro 66: 222-231.
Dobrinski, 1., P.G. Thomas, and B.A. Ball. 1995. Cryopreservation reduces the ability of equine spermatozoa to attach to the oviductal epithelial cells and zonae pellucida in vitro. J Andro. Nov-Dec; 16(6):536-542.
Donoghue, A.M. and D.J. Donoghue. 1997. Effects of water and lipid soluble antioxidants on turkey sperm viability, membrane integrity and motility during liquid storage. Poul Sci. 76: 1440-1445.
Duplaix, M. and T.J. Sexton. 1984. Effects of type of freeze straw and thae temperature on the fertilizing capability of frozen chicken semen. Poul Sci. 63: 775-780.
Engel, S., T. Schreiner and R. Petzoldt. 1999. Lipid peroxidation in human spermatozoa and maintenance of progressive sperm motility. Andrologia 31: 17-22.
Eriksson, B.M., J.M. Vazquez, E.A. Martinez, J. Roca, X. Lucas, and H. Rodriquez-Martinez. 2000. Effects of holding time during cooling and type of package on plasma membrane integrity, motility and in vitro oocyte penetration ability of frozen-thawed boar spermatozoa. Therio. May; 55(8): 1593-1605.
Eriksson, B.M.. H. Petersson, and H. Rodriquez-Martinez. 2002, Field fertility with exported boar smeen frozen in the new FlatPack container. Therio. 58: 1065-1079.
Fiser, P.S., R.W. Fairfull, C. Hansen, P.L. Panich, J.N.B Shrestha and L. Underbill. 1993. The effect of warming velocity on motility and acrosomal integrity of boar sperm as influenced by the rate of freezing and glycerol level. Mole Repro Devel. 34: 190-195.
Gao, D.Y., E. Ashworth, P.F. Watson, F.W. Kleinhaus, P. Mazur and J.K. Critser. 1993. Hyperosmotic tolerance of human spermatozoa: separate effects of glycerol, sodium chloride and sucrose on spermolysis. Biol Repro. 49(1): 112-123.
88

Gil, J. L. Soderquist and H. Rodriquez-Martinez. 1999. Influence of centrifiigation and different extenders on post-thaw sperm quality of ram semen. Therio 54: 93-108.
Gill, S.P.S., E. Buss and R.J. Mallis. 1996. Cryopreservation of rooster semen in thirteen and sixteen percent glycerol. Poul Sci. 75: 254-256.
Gilmore, J.A., J. Du, J. Tao, A.T. Peter and J.K. Critser. 1996. Osmotic properties of boar spermatozoa and their relevance to cryopreservation. J Repro Fert. May; 107(1): 87-95.
Gilmore, J.A., J. Liu, A.T. Peter, and J.K. Critser. 1998. Determination of plasma membrane characteristics of boar spermatozoa and their relevance to cryopreservation. Biol Rep. 58: 28-36.
Gilmore, J.A., L.E. McGann, J. Lui, D.Y. Gao, A.T. Peter, F.W. Kleinhaus, and J.K. Critser. 1995. Biol Repro 53(5): 985-995.
Green, C.E. and P.F. Watson. 2001. Comparison of capacitation-like state of cooled boar spermatozoa with true capitation. Repro. 122: 889-898.
Hafez, E.S.E and B. Hafez Transport and Survival of Gametes. 2000. In Reproduction in Farm Animals. Seventh Edition. Edited by E.S.E Hafez and B. Hafez. Philadelphia: Lippincott Williams and Wilkens. pp. 82-95.
Henry, M., P.P.N. Snoeck, A.C.P. Cottorello. 2002. Post-thaw spermatozoa plasma membrane integrity and motility of stallion semen frozen with different cryoprotectants. Therio. 58: 245-248.
Henry, M.A., E.E. Noiles, D. Gao, P. Mazur and J.K. Critser. 1993. Cryopreservation of human spermatozoa. FV. The effects of cooling rate and warming rate on the maintenance of motility, plasma membrane integrity, and mitochondrial function. Fert Steril 60(5): 911-918.
Holt, W.V. 2000. Fundamental aspects of sperm cryobiology: the importance of species and individual differences. Therio. 53:47-58.
Holt, W.V., and R.D. North. 1994. Effects of temperature and restoration of osmotic equilibrium during thawing on induction of plasma membrane damage in cryopreserved ram spermatozoa. Biol Repro. 51: 414-424.
Huang, Y.L., W.C. Tseng and T.H. Lin. 2001, In vifro effects of metal ions (Fe, Mn, Pb) on sperm motility and lipid peroxidation in human sf)erm. J Toxic Environ Health 62, part A: 259-267.
89

Iritani, A. 1988. Current status of biotechnological studies in mammalian reproduction. lertSter50(4): 543-551.
Ivanova-Kicheva, M.G., N. Bobadov, and B. Solev. 1997. Cryopreservation of canine semen in pellets and 5 ml aluminum tubes using three extenders. Therio 48: 1343-1349.
Johnson, L.A., K.F. Weitze, P. Fiser, and W.M.C. Maxwell. 2000. Storage of boar semen. Ani Repro Sci. 62: 143-172.
Katila, T., M. Andersson, T. Reilas and E. Koskinen. 2002. Post-thaw motility and viability of fractioned and frozen stallion ejaculates. Therio 58: 241-244.
Katkov, I.I., N. Katkova, J. K. Critser, and P. Mazur. 1998. Mouse spermatozoa in high concentrations of glycerol: chemical toxicity vs. osmotic shock at normal and reduced oxygen concentrations. Cryobiol 37: 325-338.
Lagares, M.A., R. Petzoldt, H. Sieme, and E. Klug. 2000. Assessing equine sperm -mebreme integrity. Andrologia 32: 163-167.
Lapointe, S., R. Sullivan, M.A. Sirard. 1998. Binding of a bovine oviductal fluid catalase to mammalian spermatozoa. Biol Repro. 58: 747-753.
Larsson, K. Deep-freezing of boar semen. Cryobio. 1978; 15: 352-354.
Linfor, J.J., A.C. Pommer and S.A. Meyers. 2002. Osmotic sfress induces tyrosine phosphorylation of equine sperm. Therio 58: 355-358.
Liu, Z. and R.H. Foote. 1998. Osmotic effects on volume and motility of bull sperm exposed to membrane permeable and non-permeable agents. Cryobiol 37: 207-218.
Loomis, P.R. 2001. The equine frozen semen industry. Ani Repro Sci. 68: 191-200.
Love, C C , J.A. Thompson, S.P. Brinsko, S.L. Rigby. T.L. Blanchard and D.D. Vamer. 2002. Relationship of seminal plasma level and extender type to sperm motility and DNA integrity. Therio 58: 221-224.
Martin, J.C, E. Klug, and A.R Gunzel. 1979. Cenfrifugation of stallion semen and its storage in large volume sfraws. J Repro Fert Suppl. (27): 47-51.
Maxwell, W.M.C, and L.A. Johnson. 1997. Chlorotett-acycline analysis of boar spermatozoa after incubation, flow cytometric sorting, cooling and cryopreservation. Molec Repro Develop. 46: 408-418.
90

McLaughlin, E.A. and W.C. Ford. 1994. Effects of cryopreservation on the intracellular calcium concentration of human spermatozoa and its response to progesterone. Mol Repro Develop 37(2): 241-246.
Medeiros, A.S.L., G.M. Gomes, M.T. Carmo, F.O. Papa and M.A. Alvarenga. 2001. Cryopreservation of stallion sperm using different amides. Therio 58: 273-276.
Medeiros, C.M.O., F. Forell, A.T. Oliveira and J.L. Rodriquez. 2002. Current status of sperm cryopreservation: why isn't it better? Therio 57: 327-344.
Medrano, A. P.F. Watson, and W.V. HoU. 2002. Importance of cooling rate and animal viability for boar sperm cryopreservation: insights from the cryomicroscope. Repro. 123:315-322.
Mennella, M.R. and R. Jones. 1980. Properties of spermatozoal superoxide dismutase and lack of involvement of superoxides in metal-ion catalyzed lipid peroxidation reactions in semen. Biochem J. 191: 289-297.
Merkt, H., E. Klug, D. Krause, and H. Bader. 1975. Results of long-term storage of stallion semen frozen by pellet form. J Repro Fert Suppl. 23 :105-106.
Misik, v., D. Gergel, P. Alov, K. Ondrias. 1994. An unusual temperature dependence of malondialdehyde formation in Fe ' /H202" Initiated Lipid Peroxidation of Phosphatidylcholine Liposomes. Physiol Res. 43: 163-167.
Moussa, M., V. Martinet, A. Trimeche, D. Tainturier and M. Anton. 2002. Low density lipoproteins extracted from hen egg yolk by an easy method: cryoprotective effect on frozen-thawed bull semen. Therio 57: 1695-1706.
MuUer, Z. 1982. Fertility of frozen equine semen. J Repro Fert Suppl 32: 47-51.
Neild, D.M., B.M Gadella, M.G. Chaves, M.H. Miragaya, B. Colenbrander and A. Aguero. 2003. Membrane changes during different stages of a freez-thaw protocol for equine semen cryopreservation. 59: 1693-1705.
Neild, D.M., B.M. Gadella, B. Colenbrander, A. Aguero and J.F.H.M. Brouwers. 2002. Lipid peroxidation in stallion spermatozoa. Therio 58: 295-298.
Neuman, S.L., T.L. Lin, and P.Y. Hester. 2002. The effect of dietary carnitine on semen fraits of white leghorn roosters. Poul Sci 81: 495-503.
Parks, J.E. and D.V. Lynch. 1992. Lipid composition and thermofropic phase behavior of boar, bull, stallion, and rooster sperm membranes. Cryobiol 29: 255-266.
91

Pattie, W.A. and K.F. Dowsett. 1982. The repeatability of seminal characteristics for stallions. J repro Fert Suppl 32: 9-13.
Pena, A. and CB. Linde-Forsberg. 2000. Effects of spermatozoal concenfration and post-thaw dilution rate on survival after thawing of dog spermatozoa. Therio 54: 703-718.
Pickett B.W., J.J. Sullivan, W.W. Byers, M.M. Pace, and E.E. Remmenga. 1975. Effects of centriftigation and seminal plasma on motility and fertility of stallion and bull spermatozoa. Fertil Steril. Feb; 26(2): 167-74.
Pommer, A.C. and S.A. Meyers. 2002. Tyrosine phosphorylation is an indicator of capacitation status in fresh and cryopreserved stallion spermatozoa. Therio 58: 351-354.
Pommer, A.C, J. Rutllant, and S.A. Meyers. 2002. The role of osmotic resistance on equine spermatozoal function. Therio 58: 1373-1384.
Pursel, V.G., L.A. Johnson, and L.L. Schulman. 1972. Interaction of extender composition and incubadon period on cold shock susceptibility of boar spermatozoa. J Ani Sci. Sep; 35(3): 580-584.
Rijsselaere, T., A. Van Soom, D. Maes and A. de Kruif 2002. Effect of centrifugation on in vitro survival of fresh diluted canine spermatozoa. Therio 57: 1669-1681.
Samper, J.C, M. Vidament, T. Katila, J. Newcombe, A. Estrada and J. Sargeant. 2002. Analysis of some factors associated with pregnancy rates of frozen semen: a multi-center smdy. Therio 58: 647-650.
Samper, J.C. and CA. Morris. 1998. Current methods for stallion semen cryopreservation: a survey. Therio 49(5): 895-903
Sieme, H. A. Echte and E. Klug. 2002. Effect of frequency and interval of semen collection on seminal parameters and fertility of stallions. Therio 58: 313-316.
Silva, A.R., R.C.S. Cardoso, D.C Uchoa and L.D.M. Silva. 2003. Quality of canine semen submitted to single or fractionated glycerol addition diuing the freezing process. Therio. 59: 821-829.
Squires, E.L., H.P Reger, L.J. Maclellan and J.E. Bruemmer. 2002. Effect of time of insemination and site of insemination on pregnancy rates with frozen semen. Therio 58: 655-658.
92

Sztein, J.M., K. Noble, S. Farley, and L.E. Mobraaten. 2001. Comparison of permeating and nonpermeating cryoprotectants for mouse sperm cryopreservation. Cryobiol 41:28-39.
Tang, L., Y. Zhang, Z. Oian and X. Shen. 2000. The mechanism of Fe initiated lipid peroxidation in liposomes: the dual function of ferrous ions, the roles of the pr-existing lipid peroxides and the lipid peroxyl radicals. Biochem J 352, part 1: 27-36.
Thomas, P.G.A., R.E. Larsen, J.M. Bums and C.N. Hahn. 1993. A comparison of three packaging techniques using two extenders for the cryopreservation of canine semen. Therio 40: 119-1205.
Thurston, L.M., W.V. Holt, and P.F. Watson. 2003. Post-thaw fiinctional status of boar spermatozoa cryopreserved using three controlled rate freezers: a comparison. 60: 101-113.
Tischner, M. 1979. Evaluation of deep frozen semen in stallions. J Repro Fert Suppl 27: 53-59.
Troedsson, M.H.T., A.S. Alghamdi and J. Mattisen. 2002. Equine seminal plasma protects the fertility of spermatozoa in an inflamed uterine environment. Therio 58: 453-456.
Verma, A. and K.C Kanwar. 1999. Human spermatozoal motility and lipid peroxidation. Ind J Exp Biol 37: 83-85.
Vidament, M., P. Ecot, P. Noue, C Bourgeious, M. Magistrini, E. Pahner. 2000. Centrifugation and addition of glycerol at 22 degrees C instead of 4 degrees C improve post-thaw motility and fertility of stallion spermatozoa. Therio 54(6): 907-19.
Vidament, M., C Daire, J.M. Yvon, P. Doligez, B. Bruneau, M. Magistrini, and P. Ecot. 2002. Motility and fertility of stallion semen frozen with glycerol and/or dimethyl formamide. Therio 58: 249-251.
Vishwanath, R. and P. Shannon. 2000. Storage of bovine semen in liquid and frozen state. Ani Repro Sci. 63: 23-53.
Watson, P.F. 2000. The causes of reduced fertility with cryopreserved semen. Ani Repro Sci60-61: 481-492.
93

Wilhelm, K.M., J.K. Graham, E.L. Squires. 1996. Comparison of the fertility of cryopreserved stallion spermatozoa with sperm motion analyses, flow cytometric evaluation and zona free hamster oocyte penetration. Therio 46: 559-578.
Willoughby, C.i;., P. Mazur, A.T. Peter and J.K. Critser. 1996. Osmotic tolerance limits and properties of murine spermatozoa. Biol Repro. 55(3): 715-727.
Yoshida,M. 2000. Conservation of sperms: current status and new trends. Ani Repro Sci 60-61:349-355.
Zabludovsky, N., F. Elites, E. Geva, E. Berkovitz, A. Amit, Y. Barak, D. Har-Even, and B. Bartoov. 1999. Relationship between human sperm lipid peroxidation, comprehensive quality parameters and fVF outcome. Andrologia 31: 91-98.
Zeng, W.X., M. Shimada, N. Isobe, and T. Terada. 2001. Survival of boar spermatozoa in diluents of varying osmolarity. Therio. 56: 447-458.
94