human anti-cd40 antibody and poly ic:lc adjuvant combination
TRANSCRIPT

of March 16, 2018.This information is current as
PrimatesResponses in the Lung of Nonhuman
CellAdjuvant Combination Induces Potent T Human Anti-CD40 Antibody and Poly IC:LC
LoréRichard A. Koup, Robert A. Seder, Ross M. Kedl and KarinKerrie J. Sandgren, Kylie M. Quinn, Patricia A. Darrah, Elizabeth A. Thompson, Frank Liang, Gustaf Lindgren,
ol.1500078http://www.jimmunol.org/content/early/2015/06/26/jimmun
published online 29 June 2015J Immunol
MaterialSupplementary
8.DCSupplementalhttp://www.jimmunol.org/content/suppl/2015/06/26/jimmunol.150007
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Errata
/content/195/7/3497.full.pdfor:
next pageAn erratum has been published regarding this article. Please see
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2015 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on March 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on March 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on March 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on March 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Human Anti-CD40 Antibody and Poly IC:LC AdjuvantCombination Induces Potent T Cell Responses in the Lung ofNonhuman Primates
Elizabeth A. Thompson,*,† Frank Liang,*,† Gustaf Lindgren,* Kerrie J. Sandgren,*
Kylie M. Quinn,† Patricia A. Darrah,† Richard A. Koup,† Robert A. Seder,†
Ross M. Kedl,‡ and Karin Lore*,†
Nonlive vaccine platforms that induce potent cellular immune responses inmucosal tissue would have broad application for vaccines
against infectious diseases and tumors. Induction of cellular immunity could be optimized by targeted activation of multiple innate
and costimulatory signaling pathways, such as CD40 or TLRs. In this study, we evaluated immune activation and elicitation of T cell
responses in nonhuman primates after immunization with peptide Ags adjuvanted with an agonistic anti-CD40Ab, with or without
the TLR3 ligand poly IC:LC. We found that i.v. administration of the anti-CD40Ab induced rapid and transient innate activation
characterized by IL-12 production and upregulated costimulatory and lymph node homing molecules on dendritic cells. Using
fluorescently labeled Abs for in vivo tracking, we found that the anti-CD40Ab bound to all leukocytes, except T cells, and dissem-
inated to multiple organs. CD4+ and CD8+ T cell responses were significantly enhanced when the anti-CD40Ab was coadministered
with poly IC:LC compared with either adjuvant given alone and were almost exclusively compartmentalized to the lung. Notably,
Ag-specific T cells in the bronchoalveolar lavage were sustained at ∼5–10%. These data indicate that systemic administration of
anti-CD40Ab may be particularly advantageous for vaccines and/or therapies that require T cell immunity in the lung. The
Journal of Immunology, 2015, 195: 000–000.
Most vaccines currently available primarily elicit Ab-based protection. In contrast, vaccines for infectionssuch as HIV-1/AIDS, tuberculosis, malaria, as well as
therapeutic cancer vaccines, will likely require potent cellularimmune responses, with or without humoral responses. Immune-stimulatory vaccine adjuvants provide a powerful approach tomodulate the type of adaptive response a vaccine elicits by usingearly innate immune activation. Adjuvants can stimulate vaccineresponses by targeting APCs through multiple pathways, includingengaging TLRs or other activating cell-surface receptors, such asCD40. CD40 is a major regulatory receptor in the TNF-R familywidely expressed on APCs, including dendritic cells (DCs),
monocytes, and B cells. CD40L (CD154) is expressed on ac-tivated T cells, and CD40 ligation on DCs increases costimu-latory molecule expression required for stimulation of T cellresponses (1, 2). The importance of this pathway is high-lighted by the fact that Ag presentation in the absence of CD40can lead to immune tolerance (3). By contrast, CD40 activationpromotes production of IL-12 and the costimulatory functionof APCs, which enhances both CD4+/Th1 and CD8+ T cellimmunity (4). In mouse models, CD40 stimulation can be de-livered to DCs without T cell engagement by administering ag-onistic anti-CD40Abs, soluble CD40L, or an adenovirus vectorexpressing CD40L, all of which have been shown to enhanceCD8+ T cell responses (1, 5, 6).In addition to targeting CD40, TLR ligands are effective acti-
vators of innate immunity and are critical for optimizing T cellresponses. In our previous work, we demonstrated that combin-ing CD40 activation with TLR ligands synergistically enhancedcellular immunity in mice, with increased CD4+ and CD8+ T cellresponses .10-fold compared with either adjuvant alone (7–9).T cell responses with this vaccine regimen were shown to haveantitumor effects in mice, as assessed by tumor reduction (10–12).Poly IC:LC for a combinatorial TLR ligand is particularlypromising because it is already tested in clinical cancer immu-notherapy and can target multiple innate pathways: TLR3 in DCsand MDA-5 in stromal cells, inducing production of IL-12 andtype I IFN, respectively. Mouse models have shown extensiveevidence for potential clinical use of CD40Abs in combinationwith TLR ligands as a versatile vaccine adjuvant to induce robustadaptive immunity.The partially agonistic anti-CD40Ab clone SGN-40 or huS2C6,
dacetuzumab, was tested in patients with chronic lymphocyticleukemia and non-Hodgkin’s lymphoma and showed it was welltolerated, although none of the patients achieved an objective re-
*Clinical Immunology and Allergy Unit, Department of Medicine, Karolinska Insti-tutet, 171 76 Stockholm, Sweden; †Vaccine Research Center, National Institute ofAllergy and Infectious Diseases, National Institutes of Health, Bethesda, MD 20892;and ‡Department of Immunology and Microbiology, University of Colorado Denver,Aurora, CO 80045
Received for publication January 13, 2015. Accepted for publication June 2, 2015.
This work was supported by the Vetenskapsradet (Grant 521-2012-3377 to K.L.), theSwedish Governmental Agency for Innovation Systems (Vinnova) (Grant 2010-00999 to K.L.), intramural funds as U.S. National Institutes of Health investigators(to R.A.K. and R.A.S.), and Small Business Innovation Research Phase 2 Grant5R44AI080030 to ImmuRx, Inc.
Address correspondence and reprint requests to Dr. Ross M. Kedl or Dr. Karin Lore,Department of Immunology and Microbiology, University of Colorado Denver,12800 E. 19th Avenue, Aurora, CO 80045 (R.M.K.) or Clinical Immunology andAllergy Unit, L2:04, Karolinska Institutet, Karolinska University Hospital Solna, 17176 Stockholm, Sweden (K.L.). E-mail addresses: [email protected] (R.M.K.)or [email protected] (K.L.)
The online version of this article contains supplemental material.
Abbreviations used in this article: BAL, bronchoalveolar lavage; DC, dendritic cell;Env, envelope glycoprotein; LN, lymph node; MDC, myeloid DC; MFI, medianfluorescence intensity; NHP, nonhuman primate; NIH, National Institutes of Health;SLP, synthetic long peptide.
Copyright� 2015 by The American Association of Immunologists, Inc. 0022-1767/15/$25.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1500078
Published June 29, 2015, doi:10.4049/jimmunol.1500078 by guest on M
arch 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

sponse (13, 14). Further, the agonistic anti-CD40Ab, clone CP-870,893, was tested in patients with advanced solid tumors andshowed that a single dose can confer antitumor activity (15–17), butimproved dosing intervals for optimal immune pharmacodynamicsand clinical effect are required. These results highlight the need foran improved understanding of the mechanisms by which this classof Abs mediates their effects in vivo to facilitate the use of anti-CD40Ab as an adjuvant for inducing T cell immunity in humans.Therefore, in this study, we investigated the adjuvanticity of
a novel human agonistic anti-CD40Ab (clone 341G2) together withpoly IC:LC in nonhuman primates (NHPs). NHPs provide a morepredictable model than mice for how immunomodulation can beachieved in humans based on their greater similarity in immunecell subsets, TLR distribution among APCs with humans, andtheir outbred nature. Moreover, the ability to obtain multipletissues from NHPs facilitates an extensive characterization of theinnate and adaptive immune responses mediated by human anti-CD40Abs, not possible in clinical trials, which are aspects thatmay be critical for protection against infection or tumors.
Materials and MethodsSample material
Approval for this animal study was granted by the Animal Care and UseCommittees of the Vaccine Research Center, National Institutes of Health(NIH). Indian rhesus macaques were housed at Bioqual and handledaccording to the standards of the American Association for the Accredi-tation of Laboratory Animal Care. Human PBMCs were obtained fromindividuals participating in the NIH research apheresis program. Signedinformed consent was obtained in accordance with the Declaration ofHelsinki and approved by the relevant Institutional Review Board.
Human CD40Ab screening
A variety of human anti-CD40Ab clones, including well-known and novelsequences, were screened for their ability to induce DC activation and Bcell proliferation in both human and rhesus macaque PBMCs in vitro(Supplemental Fig. 1A–C). The highest cell activation was found by theclone 341G2, which was designed based on the sequence developed byKyowa Hakko Kirin (Tokyo, Japan) (18). The clone was therefore chosento investigate potential synergy of CD40 and TLR signaling in vivo.
Immunizations
All doses used for immunizations were based on prior dose-response in vivoexperiments and previously published ranges (14, 19, 20). For innate activity,rhesus macaques received i.v. administration of 1 mg/kg anti-CD40Ab (clone341G2 IgG2), 1 mg poly IC:LC (Oncovir, Washington, DC) or the combi-nation of the two. For Ab tracking studies, anti-CD40Ab or isotype controlAb (human IgG2 DNP) was first conjugated to Alexa 680 according tomanufacturer’s protocol (Molecular Probes, Carlsbad, CA). The conjugatedAb was then treated with Triton X-114 to remove residual endotoxin and wasvalidated at ,0.1 endotoxin units with an Endpoint Chromogenic LALAssay (Lonza, Basel, Switzerland), as has been performed for prior studies(21, 22). The envelope glycoprotein (Env) peptides (Biomatik, Wilmington,DE) were resuspended to 50 mg/ml in 30% DMSO before immunization. Atotal of 1.5 mg/kg Ax680-conjugated Ab was mixed with 1 mg poly IC:LCimmediately before immunization. The formulation was delivered i.v. andwas immediately followed with 1 mg/kg Env peptides delivered i.v. Forimmunogenicity studies, animals were immunized with 1.5 mg/kg anti-CD40Ab, 1 mg poly IC:LC, and/or 4–8 mg/kg Env peptide pool (as indi-cated in Supplemental Figs. 2A, 4A), all delivered i.v. as previously de-scribed. Control animals received i.m. rAd5 HIV-1 Gag (1 3 1010 PU).Complete blood counts and liver function tests were performed 48 h after theimmunization (Idexx, Westbrook, ME) (Supplemental Fig. 1D). Animalswere first boosted with 1 mg poly IC:LC and 1 mg/kg Env peptides or rAd5HIV-1 Gag (1 3 1010 PU) and where indicated received a second boost of1.5 mg/kg anti-CD40Ab and 1 mg/kg Env peptides. Notably, endogenousAbs against the administered anti-CD40Ab were not detected until afterthe second immunization with anti-CD40Ab (data not shown).
Rhesus tissue and blood sample processing
Blood PBMCs were isolated using a density gradient with Ficoll–Paque(GE Healthcare, Fairfield, CT) according to standard procedures. All tissue
samples were collected at euthanasia and processed to a single-cell sus-pension following standard protocol. In brief, lymph nodes (LNs), spleen,and liver samples were manually disrupted and filtered through a 70-mmcell strainer. Liver samples were further purified with Ficoll–Paque. Lungand gut tissues were digested with collagenase treatment for 30 min andmechanically disrupted using the gentleMACS Dissociator (Miltenyi,Auburn, CA). PBMCs and single-cell suspensions were washed andmaintained in complete media (R10; RPMI 1640/10% FCS/100 Upenicillin/0.1 mg streptomycin; Sigma-Aldrich, St. Louis, MO) or frozenin 90% heat-inactivated FBS and 10% DMSO (Sigma-Aldrich) and storedin liquid nitrogen. Experiments were performed on both fresh and frozencells.
Cytokine secretion analysis
Rhesus serum samples were evaluated for IL-12 p40/p70 and IFN-g levelsusing NHP ELISA kits (Invitrogen). All assays were performed accordingto manufacturer’s protocol.
Phenotypic analysis
For innate studies and Ab dissemination studies, 6–24 h after immunization,5 3 106 cells were stained with LIVE/DEAD Fixable Aqua Dead Cell kitaccording to manufacturer’s protocol (Invitrogen) and blocked with Fcreceptor–blocking reagent (Miltenyi). Samples were then surfaced stainedwith a panel of fluorescently labeled Abs (Supplemental Table I) to de-termine cell distribution and maturation.
Ag recall assay
For assessment of Ag-specific cytokine production, PBMCs or single-cellsuspensions from tissues were restimulated in vitro. A total of 1.5 3 106
cells were cultured in 200 ml R10 per stimulation in a 96-well plate.Samples were stimulated as previously described (23) with 2 mg/ml 9- to13-mer overlapping HIV-1 Env peptides (matched to immunization) in thepresence of 10 mg/ml brefeldin A (Sigma-Aldrich) overnight. Sampleswere stained the following morning to evaluate IFN-g, IL-2, and TNFproduction. Cells were first stained with LIVE/DEAD Fixable AquaBlueviability dye and then surface stained and intracellularly stained (SupplementalTable I).
Multiparameter flow cytometry
Samples were resuspended in 1% paraformaldehyde before acquisitionusing a modified LSRII flow cytometer (BD Biosciences). Results wereanalyzed using FlowJo version 9.7.5, Pestle version 1.7 and Spice version5.3. Background cytokine staining was subtracted, as defined by staining insamples incubated without peptide. Where indicated, numbers of specificcell subsets are normalized to 1 3 105 viable cells (percent of gated cellsubset out of AquaBlue low cells multiplied by 105).
CD40 stimulation
Human MDDCs were prepared as previously described (24) and culturedovernight at 1 3 106 cells/ml in R10 with 5 mg/ml anti-CD40Ab clones,5 mg/ml isotype control Ab (human IgG2 DNP), or 5 mg/ml polyinosinic:polycytidylic acid (Invitrogen). Supernatants were saved for cytokineanalysis, and cells were stained for maturation markers CD70 and CD80.
ResultsDynamics and activation of leukocytes after anti-CD40Abadministration
We first determined the level of CD40 expression on differentleukocyte populations in blood of rhesus macaques. As expected,B cells showed the highest expression of CD40 compared with theother cell populations (Fig. 1A). DC subsets, monocytes, andneutrophils expressed moderate levels of CD40, whereas NK cellsand T cells did not have detectable CD40 expression. To highlightthe similarity with humans, we assessed CD40 expression in thesame cell populations in human PBMCs and showed a similarhierarchy of expression (Fig. 1A). We thereafter determined theearly innate activity of an anti-CD40Ab in vivo by evaluating howits administration affected mobilization of circulating cell pop-ulations and their activation. Human anti-CD40Ab clones werefirst screened in vitro for cell activation to select an agonistic clonesuitable for in vivo investigation (Supplemental Fig. 1A–C). In
2 ANTI-CD40 AS ADJUVANT ENHANCES T CELL RESPONSES IN THE LUNG
by guest on March 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

addition to anti-CD40Ab, the adjuvant poly IC:LC was also usedin these studies based on its potency for inducing T cell immunity.Animals received either i.v. anti-CD40Ab alone, anti-CD40Ab in
combination with poly IC:LC, or poly IC:LC alone. Blood leu-kocytes were monitored at 6 h, 24 h, 48 h, and 8 d (192 h) for cellfrequencies and phenotypes using multiparameter flow cytometry.After immunization, leukocytes quickly mobilized from the bloodin all three groups (Fig. 1B). There was a rapid decline of DCs,B cells, and T cells, but all cells returned to baseline levels by day8 in all groups (Fig. 1C). Monocytes and neutrophils and in somecases plasmacytoid DCs appeared to first increase at 6 h afteradministration, before declining and returning to baseline levels(Fig. 1C). Because there was a transient decline of all cell pop-ulations regardless of adjuvant group, this is unlikely dependenton Ab interaction with the CD40 receptor on the cells, and mayinstead be an effect of systemic immune stimulation.The DCs remaining in circulation, particularly at 6 h after ad-
ministration, exhibited phenotypic differentiation associated withupregulated CD70 and CCR7 expression. CD11c+ myeloid DC(MDC) maturation was most pronounced in the anti-CD40Ab/poly IC:LC group (Fig. 1D). We have previously shown thatCD70, a costimulatory receptor on APCs that binds to CD27 onT cells (8, 25–27), was required for enhancement of CD4+ andCD8+ T cell responses in mice after immunization with anti-CD40Ab in combination with TLR agonists (8, 9, 28). In ourNHP model, administration of anti-CD40Ab alone also inducedhigh levels of IL-12p40/p70 in the serum at 6–48 h, whichreturned back to baseline by day 8 (Fig. 1E). There was minimalIL-12 induced by poly IC:LC alone and there was no notablesynergy of anti-CD40Ab and poly IC:LC. IFN-g was also detected
in the serum, peaking at 24 h after administration of anti-CD40Ab,suggesting that early IL-12 may augment subsequent IFN-gresponses (Fig. 1E). Finally, consistent with the in vivo observa-tions, both human and NHP DCs produced IL-12 when exposed toanti-CD40Ab in vitro and upregulated CD70 and other costimu-latory molecules such as CD80 on DCs (Supplemental Fig. 1A,1B). In vitro exposure also induced proliferation of B cells(Supplemental Fig. 1C). Altogether, these data demonstrate thatanti-CD40Ab alone induces robust innate immune activation interms of cell mobilization, differentiation, and Th1-type cytokineproduction.
Systemic detection of anti-CD40Ab after i.v. administration
To investigate cell- and organ-specific targeting of the anti-CD40Ab after i.v. administration, we labeled the anti-CD40Abwith the fluorescent dye Alexa 680 to track its disseminationin vivo. To mimic the conditions used in the synergistic adjuvantmodel reported in mice, the labeled Ab was delivered with poly IC:LC and Ag (synthetic long peptides [SLPs]) (10, 12). Using flow-cytometric analysis, we assessed the median fluorescence intensity(MFI) of Alexa 680 at necropsy 24 h after vaccine administration(Fig. 2A). Labeled anti-CD40Ab was readily detected in multipleorgans and on several cell subsets at levels above background, de-fined by animals that received unlabeled anti-CD40Ab (Fig. 2B).Thus, anti-CD40Ab distributed systemically after i.v. administrationand did not target a specific organ. The highest Ab signal abovebackground was detected in the spleen and liver (Fig. 2C), con-sistent with the typical pharmacodynamics of i.v. delivery and thelarge number of immune cells, which express CD40, in the spleen.Anti-CD40Ab binding and uptake was evident in B cells, DCs,
FIGURE 1. In vivo innate activity of agonistic CD40Ab. (A) Baseline levels of CD40 expression were determined for rhesus macaque and human PBMC
subsets by CD40 MFI with florescence minus one levels subtracted (white bars) and percent of CD40+ cells (gray bars), n = 3. Refer to Fig. 2A for cell
subset gating. (B and C) Animals were split into three groups (n = 2/group) and immunized i.v. with anti-CD40Ab (1 mg/kg), poly IC:LC (1 mg), or the
combination. Circulating PBMCs were monitored for innate activity by flow cytometry. (B) Representative flow-cytometry plots from an animal receiving
anti-CD40Ab showing rapid and transient decline of blood leukocytes, including B cells, T cells, and DCs. (C) Time course (0–48 h and day 8) for each cell
type as number of cells per 105 viable cells (mean 6 SEM, n = 2). (D) Histograms show expression of differentiation makers CD70 and CCR7 on MDCs
remaining in the blood at time points 0, 6, 24, and 48 h. Maturation peaks at 6 h after administration. (E) Bar graphs show serum cytokines IL-12p40/p70
and IFN-g after administration (mean 6 SEM, n = 2).
The Journal of Immunology 3
by guest on March 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

monocytes, and neutrophils (Fig. 2B, 2C). Although neutrophilshave modest expression of CD40 at steady-state, they exhibited thehighest binding of anti-CD40Ab, which suggests they may also bindAb via Fc receptors. In contrast, T cells express neither CD40 norFc receptors and did not appreciably bind anti-CD40Ab. We per-formed similar experiments tracking the dissemination of an Alexa680–labeled IgG2 isotype control Ab. Several cell subsets andorgans showed detectable levels of isotype Ab, although at muchlower levels than the anti-CD40Ab. This suggests that CD40-specific interactions are responsible for the majority of anti-CD40Ab binding in vivo (Fig. 2D).
In vivo cell targeting and DC maturation after anti-CD40Abadministration
As described earlier (Fig. 1A), the level of CD40 expression be-tween cell types in circulation differs substantially, with B cellshaving the highest CD40 expression. Consistent with this obser-
vation, we found that tissue-resident B cells also expressed higherCD40 levels compared with other tissue-matched leukocytes.Despite B cells showing the highest expression of CD40, theAlexa 680–labeled anti-CD40Ab did not preferentially targetthem. To demonstrate this, we correlated the MFI of CD40 ex-pression, using the 5C3 clone as a staining CD40Ab, with the cell-matched MFI of Alexa 680–labeled anti-CD40Ab (Fig. 3A). Ingeneral, the leukocytes displayed the same hierarchy of CD40expression regardless of the tissue they were derived from, butCD40 expression did not consistently correlate with anti-CD40Abbinding. The Alexa 680–labeled anti-CD40Ab showed the highestsignal on neutrophils and monocytes, despite their moderate tolow CD40 expression. The same analysis using the labeled isotypecontrol Ab showed a striking reduction in binding to neutrophilsand a modest reduction in binding to monocytes and B cells(Fig. 3B). Thus, expression level of CD40 on cells or in tissuesdid not dictate their level of binding/uptake of the Ab, but it
FIGURE 2. In vivo dissemination
of anti-CD40Ab after i.v. adminis-
tration. The anti-CD40Ab or IgG
isotype control Ab were conjugated
to Alexa 680 and administered i.v.
with poly IC:LC and a pool of HIV-1
Env peptides to rhesus macaques to
determine Ab dissemination. Blood
and tissues were harvested at 24 h
and evaluated by flow cytometry for
Alexa 680 signal on a variety of cell
subsets. (A) Gating scheme used to
analyze cell-specific Alexa 680
signal. (B) Representative plots of
Alexa 680 signal from different cell
populations, as indicated in the key,
from one anti-CD40Ab–treated ani-
mal compared with PBMCs from an
unexposed animal (gray). Bar graphs
show (C) anti-CD40Ab Alexa 680
MFI on different cell subsets orga-
nized by tissue (mean 6 SEM, n =
3) and (D) isotype Ab Alexa 680
MFI on different cell subsets orga-
nized by tissue (mean 6 SEM, n = 2).
4 ANTI-CD40 AS ADJUVANT ENHANCES T CELL RESPONSES IN THE LUNG
by guest on March 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
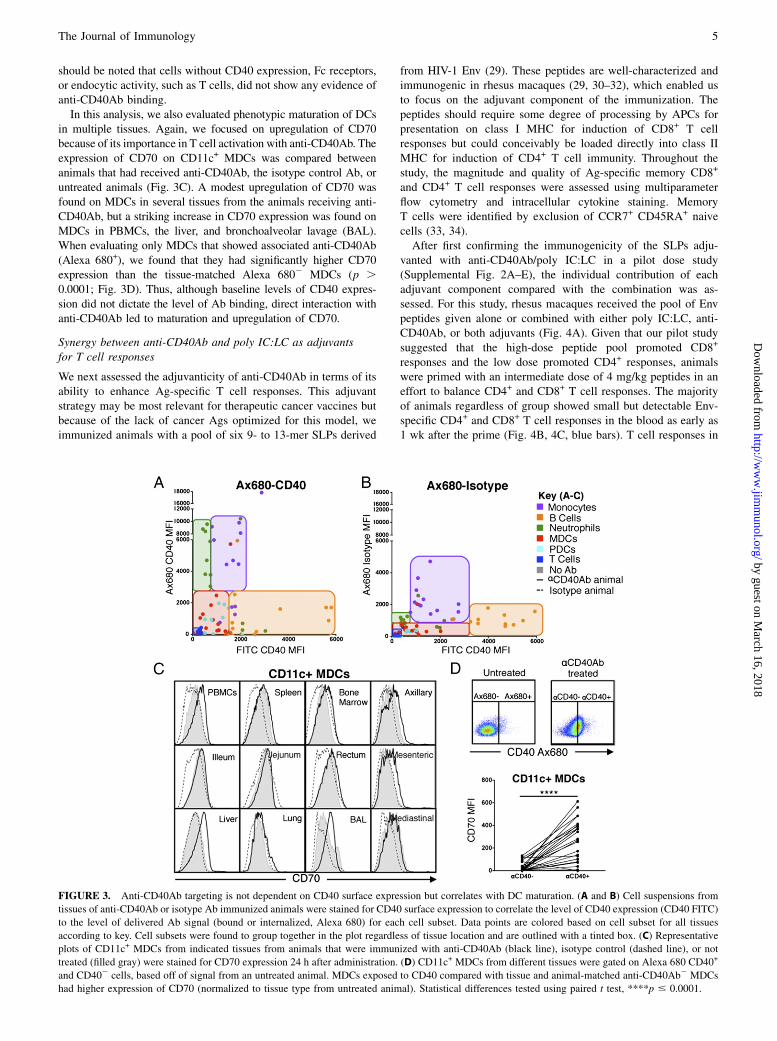
should be noted that cells without CD40 expression, Fc receptors,or endocytic activity, such as T cells, did not show any evidence ofanti-CD40Ab binding.In this analysis, we also evaluated phenotypic maturation of DCs
in multiple tissues. Again, we focused on upregulation of CD70because of its importance in T cell activation with anti-CD40Ab. Theexpression of CD70 on CD11c+ MDCs was compared betweenanimals that had received anti-CD40Ab, the isotype control Ab, oruntreated animals (Fig. 3C). A modest upregulation of CD70 wasfound on MDCs in several tissues from the animals receiving anti-CD40Ab, but a striking increase in CD70 expression was found onMDCs in PBMCs, the liver, and bronchoalveolar lavage (BAL).When evaluating only MDCs that showed associated anti-CD40Ab(Alexa 680+), we found that they had significantly higher CD70expression than the tissue-matched Alexa 6802 MDCs (p .0.0001; Fig. 3D). Thus, although baseline levels of CD40 expres-sion did not dictate the level of Ab binding, direct interaction withanti-CD40Ab led to maturation and upregulation of CD70.
Synergy between anti-CD40Ab and poly IC:LC as adjuvantsfor T cell responses
We next assessed the adjuvanticity of anti-CD40Ab in terms of itsability to enhance Ag-specific T cell responses. This adjuvantstrategy may be most relevant for therapeutic cancer vaccines butbecause of the lack of cancer Ags optimized for this model, weimmunized animals with a pool of six 9- to 13-mer SLPs derived
from HIV-1 Env (29). These peptides are well-characterized andimmunogenic in rhesus macaques (29, 30–32), which enabled usto focus on the adjuvant component of the immunization. Thepeptides should require some degree of processing by APCs forpresentation on class I MHC for induction of CD8+ T cellresponses but could conceivably be loaded directly into class IIMHC for induction of CD4+ T cell immunity. Throughout thestudy, the magnitude and quality of Ag-specific memory CD8+
and CD4+ T cell responses were assessed using multiparameterflow cytometry and intracellular cytokine staining. MemoryT cells were identified by exclusion of CCR7+ CD45RA+ naivecells (33, 34).After first confirming the immunogenicity of the SLPs adju-
vanted with anti-CD40Ab/poly IC:LC in a pilot dose study(Supplemental Fig. 2A–E), the individual contribution of eachadjuvant component compared with the combination was as-sessed. For this study, rhesus macaques received the pool of Envpeptides given alone or combined with either poly IC:LC, anti-CD40Ab, or both adjuvants (Fig. 4A). Given that our pilot studysuggested that the high-dose peptide pool promoted CD8+
responses and the low dose promoted CD4+ responses, animalswere primed with an intermediate dose of 4 mg/kg peptides in aneffort to balance CD4+ and CD8+ T cell responses. The majorityof animals regardless of group showed small but detectable Env-specific CD4+ and CD8+ T cell responses in the blood as early as1 wk after the prime (Fig. 4B, 4C, blue bars). T cell responses in
FIGURE 3. Anti-CD40Ab targeting is not dependent on CD40 surface expression but correlates with DC maturation. (A and B) Cell suspensions from
tissues of anti-CD40Ab or isotype Ab immunized animals were stained for CD40 surface expression to correlate the level of CD40 expression (CD40 FITC)
to the level of delivered Ab signal (bound or internalized, Alexa 680) for each cell subset. Data points are colored based on cell subset for all tissues
according to key. Cell subsets were found to group together in the plot regardless of tissue location and are outlined with a tinted box. (C) Representative
plots of CD11c+ MDCs from indicated tissues from animals that were immunized with anti-CD40Ab (black line), isotype control (dashed line), or not
treated (filled gray) were stained for CD70 expression 24 h after administration. (D) CD11c+ MDCs from different tissues were gated on Alexa 680 CD40+
and CD402 cells, based off of signal from an untreated animal. MDCs exposed to CD40 compared with tissue and animal-matched anti-CD40Ab2 MDCs
had higher expression of CD70 (normalized to tissue type from untreated animal). Statistical differences tested using paired t test, ****p # 0.0001.
The Journal of Immunology 5
by guest on March 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

PBMCs were modest, and there was no significant differencebetween the groups (Fig. 4B, 4C). In contrast, T cell responses inBAL were robust and groups that received anti-CD40Ab exhibitedsignificantly higher Env-specific T cell responses in BAL afterprime compared with the other groups, especially when codeliv-ered with poly IC:LC (p , 0.0001 for all groups; Fig. 4D, 4E).High CD8+ T cell responses were found in the animals receivinganti-CD40Ab with or without poly IC:LC, whereas CD4+
T cell responses were detected only when both adjuvants werecombined.In addition to the magnitude of T cell response, high proportions
of multifunctional Ag-specific T cells, that is, the ability to producemultiple cytokines at a single-cell level (IFN-g, IL-2, and TNF),correlates with protection against infection (35). When multi-functionality was assessed in our study, groups that had receivedanti-CD40Ab had higher proportions of CD8+ T cells that pro-duced more than one cytokine (Fig. 4B–E, pie charts, white or
gray portions). Differences in multifunctionality were most pro-nounced in CD8+ T cells in the BAL, where a high proportionproduced all three cytokines (Fig. 4D, pie charts, white portions),but only in groups that received anti-CD40Ab. Taken together,these data illustrate that anti-CD40Ab acts as an adjuvant to en-hance both the magnitude and the quality of Ag-specific CD8+
T cell responses.Given the qualitative and quantitative differences in Ag-specific
CD8+ T cell responses induced by anti-CD40Ab during priming,we assessed whether such responses would be differentiallyboosted. All animals were boosted at week 6 with a common boostof poly IC:LC and Env peptides, thereby controlling for the effectsof the different formulations on priming only. Surprisingly, wefound that this formulation provided a minimal boost of T cellresponses (Fig. 4B–E, purple bars), but groups primed with anti-CD40Ab maintained higher proportions of multifunctional T cellsafter boosting (Supplemental Fig. 3). At 15 wk after priming and
FIGURE 4. Development of Ag-specific T cells in mucosal tissue is dependent on anti-CD40Ab. Each component of the anti-CD40Ab/poly IC:LC
immunization was tested to see individual contributions to the development of Env-specific T cell responses. (A) Experimental design, rhesus macaques
were divided into four groups (n = 5/group) and immunized i.v. with one of the four indicated combinations of Env peptide pool (4 mg/kg), poly IC:LC
(1 mg), and anti-CD40Ab (1.5 mg/kg). At week 6, animals received a boost of the Env peptide pool (1 mg/kg) with poly IC:LC (1 mg) and at week 15 with
the peptide pool (1 mg/kg) and anti-CD40Ab (1.5 mg/kg). Blood and BAL samples were collected as indicated in (A) and used in an intracellular cytokine
staining–based assay for T cell responses. (B–E) PBMC and BAL samples were stimulated overnight with the immunizing peptide pool to recall total
memory (Live/CD3+, CD45RA+, and CCR7+ naive cells gated out) Env-specific CD8 T cells (B and D) or CD4 T cells (C and E) as determined by IFN-g
production. Pie charts show the week 5 postprime proportion of total memory CD4/CD8 T cells producing IFN-g, IL-2, and TNF (white), 2+ cells
producing any two of IFN-g, IL-2, and TNF (gray), and 1+ cells producing IFN-g, IL-2, or TNF alone (black). The black arc represents cells that produce
IFN-g. Statistical differences between the groups (all time points combined) are indicated in bar graphs, ***p , 0.0001 using two-way ANOVA. Statistical
differences for pie graphs are represented as #p # 0.05 compared with peptide alone.
6 ANTI-CD40 AS ADJUVANT ENHANCES T CELL RESPONSES IN THE LUNG
by guest on March 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

9 wk after the poly IC:LC boost, we administered a second boostusing a formulation containing peptides with anti-CD40Ab.Groups that had received anti-CD40Ab during priming exhibiteda boosting of CD8+ T cell responses in BAL (Fig. 4B–E, greenbars). The CD8+ T cell responses were boosted much more effi-ciently than the CD4+ T cell responses. After the anti-CD40Abboost, the proportion of multifunctional T cells in the BAL of allgroups increased as compared with before the boost, althoughgroups primed with anti-CD40Ab again possessed the highestproportions of such cells (Supplemental Fig. 3). Of note, groupsthat had not received anti-CD40Ab in the prime but received anti-CD40Ab in the second boost exhibited a noticeable boost of T cellresponses in PBMCs, but not in the BAL. This suggests that afterAg-specific T cell responses had been primed, the inclusion ofanti-CD40Ab in the boost was not able to reprogram T cells tobecome BAL-resident.
Anti-CD40Ab induces effector T cell responses predominatelyin the lung
Considering the widespread dissemination of anti-CD40Ab and thehigh levels of Ag-specific T cells in the BAL, we examined T cell
responses in a variety of additional tissues. The two animals with
the highest level of Ag-specific T cells in the BAL were euthanized
4 wk after the third boost (anti-CD40Ab and peptides). As a
comparison, tissues from two animals primed with peptide alone
were also evaluated. Remarkably, high frequencies of Ag-specific
T cells were exclusively detected in the lung (Fig. 5A). The
responding T cells were primarily CD8+ and were only found in
animals primed with anti-CD40Ab, poly IC:LC, and peptides, but
not in animals primed with peptide alone. Most of the responding
cells in the lung expressed the activation marker CD69 and about
half of them were CD103+, indicating that they were tissue-
resident T cells (Fig. 5B).We also assessed proliferative capacity by restimulating cells
from tissues with the Env peptides in vitro for 5 d and were able to
detect responding Env-specific CD4+ and CD8+ T cells in all
animals, including those primed with peptide alone (Fig. 5C).
Proliferating T cells were found in the spleen, PBMCs, lung, and
liver, but the most robust proliferation was observed in cells de-
rived from the bone marrow and the lung, especially cells from
animals primed with anti-CD40Ab and poly IC:LC. This illus-
trates the synergistic effect of targeting CD40 and TLR3/MDA-5
simultaneously and suggests that although immunization induced
effector CD4+ and CD8+ Env-specific T cells in the BAL, it also
established an expandable pool of memory cells in a variety of
tissues.Because of the small number of animals examined for the T cell
distribution and phenotype, an additional three animals from the
dose pilot study were analyzed. These animals were primed with
high-dose Env peptides, anti-CD40Ab, and poly IC:LC, then
boosted once with poly IC:LC and peptides. The three animals
received an additional boost with anti-CD40Ab and peptides 3 wk
before euthanasia. Again there was a striking increase in the fre-
quency of IFN-g–producing Env-specific T cells, but only in BAL
(Fig. 5D). We were unable to assess the lung responses in these
animals because the lung samples had low viability after in vitro
restimulation, potentially reflecting ongoing T cell activation.
Responding T cells in the BAL had a similar phenotype to those
found in the lung tissue above, expressing high levels of CD103
(Fig. 5E). Altogether, these data demonstrate that detectable Ag-
specific memory T cells can be identified from a variety of tissues
after expansion, but Ag-specific effector T cells are preferentially
resident in the lung and BAL.
DiscussionIn this article, the immune mechanisms leading to T cell responsesin vivo after administering an agonistic anti-CD40Ab in an NHP
model were assessed. We show that anti-CD40Ab rapidly spreads
to multiple organs after i.v. administration. This is accompanied by
mobilization of circulating leukocytes, induction of high levels of
serum IL-12, phenotypic maturation of DCs, and development of
robust T cell responses, particularly compartmentalized in the lung
when anti-CD40Ab was systemically codelivered with poly IC:LC.An NHP model provides several advantages for performing
comprehensive mechanistic and immunological evaluation in mul-
tiple tissues compared with mouse models, which can have dif-
ferent tissue-specific distribution of innate signaling pathways.
Moreover, using a human Ab provides useful data for translating
these findings to human trials. In addition to clear differences
between the mouse and human immune systems, recent studies
have also evaluated the role of anti-CD40Ab cross-linking and Fc
receptor engagement and found that differences in mouse Ab
isotypes that mediate differential engagement of activating and
inhibitory Fcg receptors did not translate to human Abs (36, 37).
When studying the commonly used murine agonistic anti-CD40Ab,
clone FGK45, it was found that specific Fc receptor engagement
was necessary for optimal activity. In contrast, the human ago-
nistic Ab clone CP-870,893 did not show the same dependence
on Fc receptor engagement (36, 37). CP-870,893 and the anti-
CD40Ab used in this study are both of the IgG2 isotype, which
typically is less likely to bind Fc receptors or activate complement
as compared with the IgG1 isotype. Despite this, the Abs clearly
induced a robust innate immune activity, further highlighting that
human anti-CD40Abs can act independently of Fc receptors.
However, altering the Fc portion of the Ab to improve affinity to
activating Fc receptors could still potentially further enhance the
function (38). It is therefore important to evaluate human Ab clones
in vivo to fully understand the mechanisms behind Ab targeting in
vaccination.Despite some promising data on CD40mAb therapy, there is also
evidence of negligible or negative clinical effect (13, 15, 17, 39).
However, the clinical trials only evaluate anti-CD40Ab as a single
agent, neglecting its potential as an adjuvant. In a phase I clinical
trial, patients treated with clone CP-870,893 showed a strikingly
similar innate profile to our animals, and the Ab was well tolerated
overall (15–17, 40). In both the clinical trial and our study there
was a brief cytokine release, activation of circulating cells, and
a transient decline returning to baseline at day 8. In this study, the
data suggest that leukocytes were mobilized to tissues rather than
being depleted. For example, whole blood counts correlated with
cell counts from flow cytometry and there were no differences in
viability staining after administration (data not shown). In addi-
tion, DCs remaining in the blood had upregulated CCR7 expres-
sion, which directs them to LNs. Cell mobilization was also not
unique to anti-CD40Ab administration, because poly IC:LC alone
induced a similar decline and cell types such as T cells, which do
not express CD40, were also affected. The redistribution of cells
may therefore be a consequence of systemic delivery of immune-
stimulatory agents (41–43). The similarities between innate pro-
files of humans and NHPs treated with agonistic anti-CD40Ab
clones demonstrate the clinical relevance of the model.Although the blood offers some insight into the innate activation
induced by anti-CD40Ab administration, the ability to track the Ab
and examine multiple tissues after administration offered a unique
opportunity to further characterize immune responses that was not
possible in clinical trials. We found that multiple cell types bound
the anti-CD40Ab, but the binding and internalization did not
The Journal of Immunology 7
by guest on March 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

correlate with the level of CD40 expression on the cells. However, itis clear that some level of CD40-specific interaction is necessary foranti-CD40Ab targeting considering the IgG isotype control Abshowed a lower Alexa 680 signal. Interestingly, there may also bea role for indirect CD40 interaction, as demonstrated by the un-expected finding that in vivo binding of isotype Ab was greatlyreduced in cell subsets such as neutrophils, which express relativelylow levels of CD40. However, neutrophils have the ability toupregulate costimulatory molecules, including CD40, as a result ofimmune activation (44–46). The isotype Ab was also deliveredwith poly IC:LC, which leads to systemic immune activation in-cluding neutrophil mobilization (Fig. 1C). However, DC differ-entiation and in particular IL-12 production were much morepronounced with anti-CD40Ab administration as compared withpoly IC:LC alone. In tissues we found that CD70 upregulation wasdirectly induced by anti-CD40Ab binding, and the highest levelswere seen in BAL and liver, potentially reflecting a better envi-
ronment for priming. The anti-CD40Ab was therefore superiorand unique in directly stimulating DCs and inducing the strongTh1-type stimulatory immune milieu, likely leading to the adju-vant effect found in this study. Even though we did not find anyevidence of obvious synergy between anti-CD40Ab and poly IC:LC with regard to innate immune activity, the combination of thetwo adjuvants clearly led to improved adaptive responses com-pared with either adjuvant alone.Our immunization strategy induced remarkably high frequencies
of sustainable Ag-specific T cells (5–10%) in the BAL, despitemodest responses in the periphery. This suggests that there wascompartmentalized priming and development of T cell responsesin the lung. Prior mouse studies evaluating anti-CD40Ab asa vaccine adjuvant have also shown high levels of Ag-specificT cells in the lung (47). However, most mouse studies only re-port on the responses in peripheral blood and spleen, which appearto be readily detectable, whereas such responses were modest in
FIGURE 5. Immunization with anti-
CD40Ab elicits tissue-resident T cells com-
partmentalized to the lung. (A) Tissues from
animals primed with anti-CD40Ab, poly IC:
LC, and Env peptide pool (filled bars) and
animals primed with peptide only (open
bars) were evaluated for Env-specific T cells
for weeks after the final boost (see Fig. 4)
as assessed by IFN-g production (mean 6SEM, n = 2/group). Tissue sample T cells
additionally had NKg2a+ and TCRgd+ gated
out. (B) Env-specific T cells in the lung were
predominantly tissue-resident T cells, based
on their CD103 and CD69 expression. (C)
CFSE-labeled cell suspensions from tissues
that were harvested from the same animals
were cultured for 5 d with immunizing
peptides and Env-specific proliferation was
analyzed (mean 6 SEM, n = 2/group). (D)
Additional animals (see Supplemental Fig.
2) that were primed with anti-CD40Ab,
poly IC:LC, and Env peptide pool that had
previously demonstrated high levels of T cell
responses were boosted with anti-CD40Ab,
poly IC:LC, and Env peptide pool. Tissues
were evaluated 3 wk later for Env-specific T
cell responses as assessed by IFN-g pro-
duction (mean 6 SEM, n = 3). (E) Env-
specific T cell responses that were highest in
the lung were predominantly tissue-resident
T cells, based on their CD103 and CD69
expression. Statistical differences compared
with all other tissues are indicated in bar
graphs, *p# 0.05 and ****p# 0.0001 using
two-way ANOVA.
8 ANTI-CD40 AS ADJUVANT ENHANCES T CELL RESPONSES IN THE LUNG
by guest on March 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

our NHP study. It is possible that the responses in mouse are moreefficiently disseminated and/or that the doses of model Ags suchas OVA, rather than HIV Ags, are not comparable between theanimal models.The lung-compartmentalized responses could be a result of the
i.v. route of delivery and/or are due to inherent characteristics ofthe anti-CD40Ab. We found that systemic administration of theanti-CD40Ab efficiently targeted lung DCs, which could lead topreferential generation of lung resident T cells. Lung resident DCsin mice were shown to imprint T cell homing to the lung throughinduction of CCR4 expression on the T cells (48). Although wewere not able to evaluate CCR4 expression on T cells in rhesustissues, the responding T cells appeared to be largely CD103+,indicating that they were tissue-resident memory cells (49). Wealso found that APCs sorted from the lung enhanced T cell pro-liferation and induced CD103 expression when pretreated with theanti-CD40Ab in vitro (data not shown). As mentioned earlier, thehigh DC maturation detected in the lung may indicate a superiorenvironment for DC presentation and priming of T cell responsesafter anti-CD40Ab administration. Mouse data also show thatalthough T cells imprinted by lung DCs preferentially home to thelung, they are more flexible in their peripheral distribution thanT cells imprinted by skin or gut DCs (48). This may explain ourobservations that Ag-specific effector T cells were preferentiallyin the lung but could also be found systemically after expansionfor 5 d in a proliferation assay. These results indicate that theimmunization targeted and activated lung DCs because of theroute of delivery, but they do not rule out the possibility of in-trinsic signaling of the anti-CD40Ab to imprint lung homingT cells. In a pilot study when we administered Ag and poly IC:LCs.c. and anti-CD40Ab i.v., the T cell responses were still modest inperipheral blood and higher in BAL, suggesting that priming re-stricted to the skin draining LNs in the presence of systemicCD40Ab still induces some degree of compartmentalized lungresponses (Supplemental Fig. 2F–I).Future studies are needed to elucidate whether anti-CD40Ab
would be particularly potent for vaccine formulations to patho-gens and tumors specific to the lung and warrants investigation ofother therapeutic Abs, for example, anti-PD1 and anti-CD20 alsodelivered i.v. It will also be important to test the functionality ofthese T cells with tumor Ags and investigate the potential oftargeting other organs with this Ab with alternative routes of de-livery. Although it is clear that successful next-generation vaccinesto both infectious diseases and cancer will require potent anddurable T cell responses, a much better understanding of howstrong vaccine responses can be elicited and maintained is es-sential. These studies demonstrate the ability of an agonistic anti-CD40Ab to induce high levels of DC targeting and activation,presumably leading to substantial lung-specific CD8+ T cell re-sponses. Altogether, such data contribute to the understanding ofhow T cell responses can be tailored via innate immune stimula-tion to be more efficacious and to improving the design of futurevaccine formulations.
AcknowledgmentsWe thank John-Paul Todd, Vaccine Research Center, NIH; the animal care
personnel at Bioqual; Oncovir, Inc. for providing poly IC:LC; and Linnea
Haeggblom for technical assistance.
DisclosuresR.M.K. is a founder of ImmuRx, Inc. whose intellectual property is based
on the combined TLR agonist/anti-CD40Ab immunization platform. The
other authors have no financial conflicts of interest.
References1. Hanyu, K., T. Iida, H. Shiba, T. Ohashi, Y. Eto, and K. Yanaga. 2008. Immu-
nogene therapy by adenovirus vector expressing CD40 ligand for metastatic livercancer in rats. Anticancer Res. 28(5A): 2785–2789.
2. van Kooten, C., and J. Banchereau. 2000. CD40-CD40 ligand. J. Leukoc. Biol.67: 2–17.
3. Buhlmann, J. E., T. M. Foy, A. Aruffo, K. M. Crassi, J. A. Ledbetter,W. R. Green, J. C. Xu, L. D. Shultz, D. Roopesian, R. A. Flavell, et al. 1995. Inthe absence of a CD40 signal, B cells are tolerogenic. Immunity 2: 645–653.
4. Diehl, L., A. T. Den Boer, E. I. van der Voort, C. J. Melief, R. Offringa, andR. E. Toes. 2000. The role of CD40 in peripheral T cell tolerance and immunity.J. Mol. Med. 78: 363–371.
5. Davis, I. D., Q. Chen, L. Morris, J. Quirk, M. Stanley, M. L. Tavarnesi,P. Parente, T. Cavicchiolo, W. Hopkins, H. Jackson, et al. 2006. Blood dendriticcells generated with Flt3 ligand and CD40 ligand prime CD8+ T cells efficientlyin cancer patients. J. Immunother. 29: 499–511.
6. Gladue, R. P., T. Paradis, S. H. Cole, C. Donovan, R. Nelson, R. Alpert,J. Gardner, E. Natoli, E. Elliott, R. Shepard, and V. Bedian. 2011. The CD40agonist antibody CP-870,893 enhances dendritic cell and B-cell activity andpromotes anti-tumor efficacy in SCID-hu mice. Cancer Immunol. Immunother.60: 1009–1017.
7. Ahonen, C. L., C. L. Doxsee, S. M. McGurran, T. R. Riter, W. F. Wade,R. J. Barth, J. P. Vasilakos, R. J. Noelle, and R. M. Kedl. 2004. Combined TLRand CD40 triggering induces potent CD8+ T cell expansion with variable de-pendence on type I IFN. J. Exp. Med. 199: 775–784.
8. McWilliams, J. A., P. J. Sanchez, C. Haluszczak, L. Gapin, and R. M. Kedl.2010. Multiple innate signaling pathways cooperate with CD40 to induce potent,CD70-dependent cellular immunity. Vaccine 28: 1468–1476.
9. Sanchez, P. J., J. A. McWilliams, C. Haluszczak, H. Yagita, and R. M. Kedl.2007. Combined TLR/CD40 stimulation mediates potent cellular immunity byregulating dendritic cell expression of CD70 in vivo. J. Immunol. 178: 1564–1572.
10. Barrios, K., and E. Celis. 2012. TriVax-HPV: an improved peptide-based ther-apeutic vaccination strategy against human papillomavirus-induced cancers.Cancer Immunol. Immunother. 61: 1307–1317.
11. Hailemichael, Y., Z. Dai, N. Jaffarzad, Y. Ye, M. A. Medina, X. F. Huang,S. M. Dorta-Estremera, N. R. Greeley, G. Nitti, W. Peng, et al. 2013. Persistentantigen at vaccination sites induces tumor-specific CD8⁺ T cell sequestration,dysfunction and deletion. Nat. Med. 19: 465–472.
12. Assudani, D., H. I. Cho, N. DeVito, N. Bradley, and E. Celis. 2008. In vivoexpansion, persistence, and function of peptide vaccine-induced CD8 T cellsoccur independently of CD4 T cells. Cancer Res. 68: 9892–9899.
13. Advani, R., A. Forero-Torres, R. R. Furman, J. D. Rosenblatt, A. Younes,H. Ren, K. Harrop, N. Whiting, and J. G. Drachman. 2009. Phase I study of thehumanized anti-CD40 monoclonal antibody dacetuzumab in refractory or re-current non-Hodgkin’s lymphoma. J. Clin. Oncol. 27: 4371–4377.
14. Furman, R. R., A. Forero-Torres, A. Shustov, and J. G. Drachman. 2010. A phaseI study of dacetuzumab (SGN-40, a humanized anti-CD40 monoclonal antibody)in patients with chronic lymphocytic leukemia. Leuk. Lymphoma 51: 228–235.
15. Beatty, G. L., D. A. Torigian, E. G. Chiorean, B. Saboury, A. Brothers, A. Alavi,A. B. Troxel, W. Sun, U. R. Teitelbaum, R. H. Vonderheide, and P. J. O’Dwyer.2013. A phase I study of an agonist CD40 monoclonal antibody (CP-870,893) incombination with gemcitabine in patients with advanced pancreatic ductal ad-enocarcinoma. Clin Cancer Res. 19: 6286–6295.
16. Vonderheide, R. H., J. M. Burg, R. Mick, J. A. Trosko, D. Li, M. N. Shaik,A. W. Tolcher, and O. Hamid. 2013. Phase I study of the CD40 agonist antibodyCP-870,893 combined with carboplatin and paclitaxel in patients with advancedsolid tumors. OncoImmunology 2: e23033.
17. Vonderheide, R. H., K. T. Flaherty, M. Khalil, M. S. Stumacher, D. L. Bajor,N. A. Hutnick, P. Sullivan, J. J. Mahany, M. Gallagher, A. Kramer, et al. 2007.Clinical activity and immune modulation in cancer patients treated with CP-870,893, a novel CD40 agonist monoclonal antibody. J. Clin. Oncol. 25: 876–883.
18. Takahashi, N., T. Miura, Y. Kitagawa, and A. Matsushima, inventors; KyowaHakka Kirin Co., Ltd., assignee. Anti-cd40 antibody mutants. United States patentapplication 14/017,789, Publication No. US 2014/0105907 A1. 2014 Apr 17.
19. Byrd, J. C., T. J. Kipps, I. W. Flinn, M. Cooper, O. Odenike, J. Bendiske,J. Rediske, S. Bilic, J. Dey, J. Baeck, and S. O’Brien. 2012. Phase I study of theanti-CD40 humanized monoclonal antibody lucatumumab (HCD122) in relapsedchronic lymphocytic leukemia. Leuk. Lymphoma 53: 2136–2142.
20. Park, H., L. Adamson, T. Ha, K. Mullen, S. I. Hagen, A. Nogueron,A. W. Sylwester, M. K. Axthelm, A. Legasse, M. Piatak, Jr., et al. 2013.Polyinosinic-polycytidylic acid is the most effective TLR adjuvant for SIV Gagprotein-induced T cell responses in nonhuman primates. J. Immunol. 190: 4103–4115.
21. Wille-Reece, U., B. J. Flynn, K. Lore, R. A. Koup, R. M. Kedl, J. J. Mattapallil,W. R. Weiss, M. Roederer, and R. A. Seder. 2005. HIV Gag protein conjugatedto a Toll-like receptor 7/8 agonist improves the magnitude and quality of Th1and CD8+ T cell responses in nonhuman primates. Proc. Natl. Acad. Sci. USA102: 15190–15194.
22. Wille-Reece, U., B. J. Flynn, K. Lore, R. A. Koup, A. P. Miles, A. Saul,R. M. Kedl, J. J. Mattapallil, W. R. Weiss, M. Roederer, and R. A. Seder. 2006.Toll-like receptor agonists influence the magnitude and quality of memory T cellresponses after prime-boost immunization in nonhuman primates. J. Exp. Med.203: 1249–1258.
23. Quinn, K. M., A. Da Costa, A. Yamamoto, D. Berry, R. W. Lindsay, P. A. Darrah,L. Wang, C. Cheng, W. P. Kong, J. G. Gall, et al. 2013. Comparative analysis of
The Journal of Immunology 9
by guest on March 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

the magnitude, quality, phenotype, and protective capacity of simian immuno-deficiency virus gag-specific CD8+ T cells following human-, simian-, andchimpanzee-derived recombinant adenoviral vector immunization. J. Immunol.190: 2720–2735.
24. Lore, K., A. Sonnerborg, A. L. Spetz, U. Andersson, and J. Andersson. 1998.Immunocytochemical detection of cytokines and chemokines in Langerhanscells and in vitro derived dendritic cells. J. Immunol. Methods 214: 97–111.
25. Bullock, T. N., and H. Yagita. 2005. Induction of CD70 on dendritic cellsthrough CD40 or TLR stimulation contributes to the development of CD8+T cell responses in the absence of CD4+ T cells. J. Immunol. 174: 710–717.
26. Keller, A. M., A. Schildknecht, Y. Xiao, M. van den Broek, and J. Borst. 2008.Expression of costimulatory ligand CD70 on steady-state dendritic cells breaksCD8+ T cell tolerance and permits effective immunity. Immunity 29: 934–946.
27. Dong, H., N. A. Franklin, D. J. Roberts, H. Yagita, M. J. Glennie, andT. N. Bullock. 2012. CD27 stimulation promotes the frequency of IL-7 receptor-expressing memory precursors and prevents IL-12-mediated loss of CD8(+)T cell memory in the absence of CD4(+) T cell help. J. Immunol. 188: 3829–3838.
28. Kurche, J. S., C. Haluszczak, J. A. McWilliams, P. J. Sanchez, and R. M. Kedl.2012. Type I IFN-dependent T cell activation is mediated by IFN-dependentdendritic cell OX40 ligand expression and is independent of T cell IFNR ex-pression. J. Immunol. 188: 585–593.
29. Nehete, P. N., B. P. Nehete, L. Hill, P. R. Manuri, V. Baladandayuthapani,L. Feng, J. Simmons, and K. J. Sastry. 2008. Selective induction of cell-mediatedimmunity and protection of rhesus macaques from chronic SHIV(KU2) infectionby prophylactic vaccination with a conserved HIV-1 envelope peptide-cocktail.Virology 370: 130–141.
30. Nehete, P. N., B. P. Nehete, P. Manuri, L. Hill, J. L. Palmer, and K. J. Sastry.2005. Protection by dendritic cells-based HIV synthetic peptide cocktail vaccine:preclinical studies in the SHIV-rhesus model. Vaccine 23: 2154–2159.
31. Koopman, G., N. Beenhakker, I. Nieuwenhuis, G. Doxiadis, P. Mooij,J. W. Drijfhout, J. Koestler, T. Hanke, Z. Fagrouch, E. J. Verschoor, et al. 2013.DNA/long peptide vaccination against conserved regions of SIV induces partialprotection against SIVmac251 challenge. AIDS 27: 2841–2851.
32. Rosario, M., A. Bridgeman, E. D. Quakkelaar, M. F. Quigley, B. J. Hill,M. L. Knudsen, V. Ammendola, K. Ljungberg, N. Borthwick, E. J. Im, et al.2010. Long peptides induce polyfunctional T cells against conserved regions ofHIV-1 with superior breadth to single-gene vaccines in macaques. Eur. J.Immunol. 40: 1973–1984.
33. Darrah, P. A., D. T. Patel, P. M. De Luca, R. W. B. Lindsay, D. F. Davey,B. J. Flynn, S. T. Hoff, P. Andersen, S. G. Reed, S. L. Morris, et al. 2007.Multifunctional TH1 cells define a correlate of vaccine-mediated protectionagainst Leishmania major. Nat. Med. 13: 843–850.
34. Betts, M. R., M. C. Nason, S. M. West, S. C. De Rosa, S. A. Migueles,J. Abraham, M. M. Lederman, J. M. Benito, P. A. Goepfert, M. Connors, et al.2006. HIV nonprogressors preferentially maintain highly functional HIV-specificCD8+ T cells. Blood. 107: 4781–4789.
35. Seder, R. A., P. A. Darrah, and M. Roederer. 2008. T-cell quality in memory andprotection: implications for vaccine design. Nat. Rev. Immunol. 8: 247–258.
36. Richman, L. P., and R. H. Vonderheide. 2014. Role of crosslinking for agonisticCD40 monoclonal antibodies as immune therapy of cancer. Cancer Immunol.Res. 2: 19–26.
37. Vonderheide, R. H., and M. J. Glennie. 2013. Agonistic CD40 antibodies andcancer therapy. Clin. Cancer Res. 19: 1035–1043.
38. Nimmerjahn, F., and J. V. Ravetch. 2005. Divergent immunoglobulin g subclassactivity through selective Fc receptor binding. Science 310: 1510–1512.
39. R€uter, J., S. J. Antonia, H. A. Burris, R. D. Huhn, and R. H. Vonderheide. 2010.Immune modulation with weekly dosing of an agonist CD40 antibody in a phaseI study of patients with advanced solid tumors. Cancer Biol. Ther. 10: 983–993.
40. Carpenter, E. L., R. Mick, J. R€uter, and R. H. Vonderheide. 2009. Activation ofhuman B cells by the agonist CD40 antibody CP-870,893 and augmentation withsimultaneous toll-like receptor 9 stimulation. J. Transl. Med. 7: 93.
41. Januszkiewicz, A., K. Lore, P. Essen, B. Andersson, M. A. McNurlan,P. J. Garlick, O. Ringden, J. Andersson, and J. Wernerman. 2002. Response ofin vivo protein synthesis in T lymphocytes and leucocytes to an endotoxinchallenge in healthy volunteers. Clin. Exp. Immunol. 130: 263–270.
42. Kamphuis, E., T. Junt, Z. Waibler, R. Forster, and U. Kalinke. 2006. Type Iinterferons directly regulate lymphocyte recirculation and cause transient bloodlymphopenia. Blood 108: 3253–3261.
43. Gunzer, M., H. Riemann, Y. Basoglu, A. Hillmer, C. Weishaupt, S. Balkow,B. Benninghoff, B. Ernst, M. Steinert, T. Scholzen, et al. 2005. Systemic ad-ministration of a TLR7 ligand leads to transient immune incompetence due toperipheral-blood leukocyte depletion. Blood 106: 2424–2432.
44. Fanger, N. A., C. Liu, P. M. Guyre, K. Wardwell, J. O’Neil, T. L. Guo,T. P. Christian, S. P. Mudzinski, and E. J. Gosselin. 1997. Activation of humanT cells by major histocompatability complex class II expressing neutrophils:proliferation in the presence of superantigen, but not tetanus toxoid. Blood 89:4128–4135.
45. Reinisch, W., W. Tillinger, C. Lichtenberger, A. Gangl, M. Willheim,O. Scheiner, and G. Steger. 1996. In vivo induction of HLA-DR on humanneutrophils in patients treated with interferon-gamma. Blood 87: 3068.
46. Beauvillain, C., Y. Delneste, M. Scotet, A. Peres, H. Gascan, P. Guermonprez,V. Barnaba, and P. Jeannin. 2007. Neutrophils efficiently cross-prime naiveT cells in vivo. Blood 110: 2965–2973.
47. Lee, S., K. L. Stokes, M. G. Currier, K. Sakamoto, N. W. Lukacs, E. Celis, andM. L. Moore. 2012. Vaccine-elicited CD8+ T cells protect against respiratorysyncytial virus strain A2-line19F-induced pathogenesis in BALB/c mice. J.Virol. 86: 13016–13024.
48. Mikhak, Z., J. P. Strassner, and A. D. Luster. 2013. Lung dendritic cells imprintT cell lung homing and promote lung immunity through the chemokine receptorCCR4. J. Exp. Med. 210: 1855–1869.
49. Farber, D. L., N. A. Yudanin, and N. P. Restifo. 2014. Human memory T cells:generation, compartmentalization and homeostasis. Nat. Rev. Immunol. 14:24–35.
10 ANTI-CD40 AS ADJUVANT ENHANCES T CELL RESPONSES IN THE LUNG
by guest on March 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Corrections
Thompson, E. A., F. Liang, G. Lindgren, K. J. Sandgren, K.M.Quinn, P. A. Darrah, R. A.Koup, R. A. Seder, R.M.Kedl, andK. Lore. 2015.Human anti-CD40 antibody and poly IC:LC adjuvant combination induces potent T cell responses in the lung of nonhuman primates.J. Immunol. 195: 1015–1024.
The graphs from Fig. 2C were inadvertently duplicated in Fig. 2D in the published article. The corrected Fig. 2 is shown below. Thefigure legend is correct as published and is shown below for reference.
Copyright � 2015 by The American Association of Immunologists, Inc. 0022-1767/15/$25.00
The Journal of Immunology

www.jimmunol.org/cgi/doi/10.4049/jimmunol.1501682
FIGURE 2. In vivo dissemination of anti-CD40Ab after i.v. administration. The anti-CD40Ab or IgG isotype control Ab were conjugated to Alexa
680 and administered i.v.with poly IC:LC and a pool ofHIV-1Env peptides to rhesusmacaques to determineAbdissemination.Blood and tissueswere harvested at
24 h and evaluated by flow cytometry for Alexa 680 signal on a variety of cell subsets. (A) Gating scheme used to analyze cell-specific Alexa 680 signal.
(B) Representative plots of Alexa 680 signal from different cell populations, as indicated in the key, fromone anti-CD40Ab–treated animal comparedwith PBMCs
from an unexposed animal (gray). Bar graphs show (C) anti-CD40Ab Alexa 680 MFI on different cell subsets organized by tissue (mean6 SEM, n5 3) and
(D) isotype Ab Alexa 680 MFI on different cell subsets organized by tissue (mean6 SEM, n5 2).
3498 CORRECTIONS