anti-cd40 monoclonal antibody synergizes with ctla4-ig … · receive free email-alerts when new...
TRANSCRIPT

of July 16, 2018.This information is current as
TransplantationGraft Survival in Murine Models ofwith CTLA4-Ig in Promoting Long-Term Anti-CD40 Monoclonal Antibody Synergizes
L. Ford and Christian P. LarsenGangappa, Diane Hollenbaugh, Thomas C. Pearson, Mandy Christopher R. Gilson, Zvonimir Milas, Shivaprakash
http://www.jimmunol.org/content/183/3/1625doi: 10.4049/jimmunol.0900339July 2009;
2009; 183:1625-1635; Prepublished online 10J Immunol
Referenceshttp://www.jimmunol.org/content/183/3/1625.full#ref-list-1
, 30 of which you can access for free at: cites 72 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2009 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Anti-CD40 Monoclonal Antibody Synergizes with CTLA4-Ig inPromoting Long-Term Graft Survival in Murine Modelsof Transplantation
Christopher R. Gilson,* Zvonimir Milas,* Shivaprakash Gangappa,* Diane Hollenbaugh,†
Thomas C. Pearson,* Mandy L. Ford,* and Christian P. Larsen1*
Blockade of the CD40/CD154 signaling pathway using anti-CD154 Abs has shown promise in attenuating the alloimmune responseand promoting long-term graft survival in murine model systems, although side effects observed in humans have hampered itsprogression through clinical trials. Appropriately designed anti-CD40 Abs may provide a suitable alternative. We investigated twoisoforms of a novel monoclonal rat anti-mouse CD40 Ab (7E1) for characteristics and effects mirroring those of anti-CD154:7E1-G1 (an IgG1 isotype); and 7E1-G2b (an IgG2b isotype). In vitro proliferation assays to measure the agonist properties of thetwo anti-CD40 Abs revealed similar responses when plate bound. However, when present as a soluble stimulus, 7E1-G1 but not7E1-G2b led to proliferation. 7E1-G2b was as effective as anti-CD154 when administered in vivo in concert with CTLA4-Ig inpromoting both allogeneic bone marrow chimerism and skin graft survival, whereas 7E1-G1 was not. The protection observedwith 7E1-G2b was not due to depletion of CD40-bearing APCs. These data suggest that an appropriately designed anti-CD40 Abcan promote graft survival as well as anti-CD154, making 7E1-G2b an attractive substitute in mouse models of costimulationblockade-based tolerance regimens. The Journal of Immunology, 2009, 183: 1625–1635.
T he generation of an optimal T cell response requires sig-nals delivered via TCR engagement (signal 1) as well asaccessory signals in the form of costimulatory molecules
(signal 2; Ref. 1). When signal 1 is delivered in the absence ofsignal 2, T cell activation can be severely altered, resulting inanergy, apoptosis, abortive proliferation, or immunoregulation (2–4). The potential of inducing these states using agents that interruptthese signaling events has captivated the interest of the transplantcommunity. Several signaling pathways have been targeted forblockade with some measure of success including ICOS, OX40,4-1BB, and CD70 (5–8). In particular, inhibition of the earliestidentified and most well-studied costimulatory pathways, CD28and CD40, has shown impressive synergy in mouse models for theprevention of graft rejection (9). Blockade of the CD28 pathwaywith CTLA4-Ig (abatacept) and LEA29Y (belatacept), a second-generation CD28 blocker with an increased affinity for CD80/86,proved efficacious in nonhuman primate models (10–12), pavingthe way for the use of belatacept in human clinical trials of kidneytransplantation (13).
Blockade of the CD40 signaling pathway also has shown greatpromise in nonhuman primates (14, 15). However, clinical trialsusing anti-CD154 mAbs were halted because of evidence of un-anticipated thromboembolic side effects (16–18). It now is appre-ciated that CD154, expressed by platelets as well as present in
soluble form, is intimately involved in the formation and stabili-zation of thrombi (19, 20). Clinical development of traditional anti-CD154 mAbs has been largely suspended. To continue exploringthe inhibition of the CD40 pathway as a therapeutic modality, al-ternative means must be sought to interrupt this pivotal costimu-lation pathway without the potential for precipitating dysregulatedthrombostasis.
The CD40 cascade provides essential signals to many aspects ofimmunity. Studies in vitro and using CD40� mice have describedits role in, among others, T cell-dependent Ig class switching andgerminal center formation, CD8 T cell priming, dendritic cell lon-gevity and cytokine production, and endothelial activation (21–25). In fully allogeneic models of cardiac and islet, but not skin,transplantation, genetic deficiency of CD40 in both the donor andthe recipient results in long-term graft survival (26–28). Investi-gations outside of the field of transplantation using anti-CD40mAb have generally focused on their agonistic properties to aug-ment immune responses to thymus-independent Ags or to boostantitumor and antivirus activity (22, 29–32). There are, however,reports of successful immune attenuation achieved in mouse andnonhuman primate kidney allograft models using anti-CD40 mAb(33–36). These anti-CD40 Abs have shown synergy with CD28pathway blockers (both a CTLA-4 Ig fusion protein and anti-CD86mAbs) in promoting renal and islet allograft survival in nonhumanprimates while also preserving established antiviral immunity (33,35). The mechanisms behind anti-CD40-mediated immunosup-pression and how it compares with anti-CD154 mediated suppres-sion remain unclear.
Herein we report two novel murine anti-CD40-binding Abs:7E1-G1, a rat IgG1 isotype; and its natural isotype switch variant7E1-G2b, a rat IgG2b isotype. After standard hybridoma selectionand purification techniques, the clone 7E1-G1 was selected as thelead candidate based on its specificity for murine CD40, ability toinhibit ligand binding, and paucity of agonism (37). It was hypoth-esized that an Ab that strongly induced both complement fixation
*Department of Surgery and Emory Transplant Center, Emory University, Atlanta,GA 30322; and †Bristol-Myers Squibb Pharmaceutical Research Institute, Princeton,NJ 08543
Received for publication February 2, 2009. Accepted for publication June 2, 2009.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 Address correspondence and reprint requests to Dr. Christian P. Larsen, WoodruffMemorial Building Suite 5105, Emory University, 101 Woodruff Circle, Atlanta, GA30322. E-mail address: [email protected]
Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00
The Journal of Immunology
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0900339
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

and FcR interactions would be the most favorable for promotinggraft survival. Such an Ab would potentially deplete CD40-ex-pressing APCs before the priming of an effective response couldoccur. However, rat IgG1 Ab isotypes have been shown in variousmodels to poorly fix complement and interact weakly with FcRs(38–40). Thus, an Ab having the same specificity as 7E1-G1 butpossessing an IgG2b isotype was sought. To this end, the isotypeswitch variant 7E1-G2b was generated by the sib selection tech-nique (37, 41).
In this study, we found that when used in concert with CTLA4-Ig, the anti-CD40 Ab 7E1-G2b, but not 7E1-G1, was as effectiveas anti-CD154 in promoting allogeneic bone marrow chimerismand skin graft survival. The mechanism underlying this prolonga-tion did not involve depletion of CD40-expressing cell subsets,given that numbers of APCs remained unchanged among the treat-ment groups. Furthermore, 7E1-G1 but not 7E1-G2b functioned asa potent CD40 agonist in vivo, suggesting the involvement of FcRsin mediating these effects. However, in our system, proinflamma-tory signals delivered by activating FcRs were not responsible forthe immunostimulatory properties of 7E1-G1, nor were inhibitorysignals through Fc�RIIb found to be involved in the immunosup-pressive effects observed with 7E1-G2b.
Materials and MethodsAb generation
A recombinant mouse CD40 Ig fusion protein consisting of the extracel-lular region of mouse CD40 fused to the hinge zinc-binding domains 2 and3, based on relative positions of cysteine (C) and histidine (H) of a mouseIgG2a Ab was used to immunize Lewis rats. Three days after the lastimmunization, leukocytes were fused with X63-Ag8.653 mouse myelomacells to create rat/mouse heterohybridomas. Supernatants were tested forreactivity with mouse CD40 and for the ability to inhibit the binding ofsoluble mouse CD40 to its ligand CD154. Stimulatory properties weremeasured by proliferation of splenic B cells in the presence of anti-IgM.Suppression of Ab production was measured by immunization of mice withSRBC. Clone 7E1-G1 was selected as the lead anti-CD40 mAb candidatefor murine studies (37).
A natural isotype switch variant of 7E1-G1, from an IgG1 to an IgG2b,was generated by the sib selection technique (41). Briefly, an anti-CD40mAb of the IgG2b isotype was identified by ELISA among supernatants of96-well plates that had been seeded at 1000 cells/well with the original7E1-G1 hybridoma. Subsequent rounds of plating and identification ofIgG2b-positive wells at seeding densities of 200 and then 20 cells/wellfollowed by two rounds of cloning by limiting dilution led to the isolationof the clonal IgG2b switch variant, 7E1-G2b.
Mice
Adult male 6- to 8-wk-old C57BL/6, BALB/c, C3H/HeJ, B6.129S2-Igh-6tm1Cgn/J, and B6.129P2-Fcgr3tm1Sjv/J, were obtained from The JacksonLaboratory. Act-mOVA mice were generously provided by Dr. M. Jenkins(University of Minnesota, Minneapolis, MN). B6.129P2-Fcer1gtm1Rav N12and B6.129S4-Fcgr2btm1TtK N12 mouse strains were obtained from Tac-onic Farms. Animals received humane care in specific pathogen-free hous-ing conditions in accordance with Emory University Institutional AnimalCare and Use Committee guidelines.
Competitive binding and affinity assays
The M12 B cell lymphoma cell line was generously provided by Dr. N.Iwakoshi (Emory University, Atlanta, GA). To compare the binding affin-ities of the Abs, M12 B cells were incubated at 4°C for 30 min with 8 pg/mlto 1 mg/ml 7E1-G1, 7E1-G2b, isotype-matched control, or FGK4.5. Iso-type control Abs and FGK4.5, a rat IgG2a isotype (42), were purchasedfrom BioXCell. Cells were washed with PBS supplemented with 0.5%BSA and 1 mM EDTA, pH 7.2 (FACS buffer), and bound Ab was thendetected with PE-conjugated F(ab�)2 goat anti-rat IgG (Jackson Immuno-Research Laboratories) at 4°C for 30 min. For the competitive bindingassay, cells were incubated with the same titrations of Abs as noted pre-viously, along with 10 �g/ml FLAG-tagged soluble recombinant mouseCD154 (Axxora) at 4°C for 30 min. Cells were washed with FACS buffer,and bound CD154 was detected with FITC-conjugated anti-FLAG M2 Ab(Sigma-Aldrich) at 4°C for 30 min. Nonviable cells were excluded by the
addition of 7-aminoactinomycin D (7-AAD)2 10 min before sample acqui-sition (BD Biosciences).
In vitro proliferation assay
To test the 7E1 variants as an immobilized stimulus, Immulon 4 HBXplates (Daigger) were incubated overnight with 1 ng/ml to 1 mg/ml of7E1-G1, 7E1-G2b, isotype-matched control, or FGK4.5. For provision ofa soluble stimulus, Costar cell culture plates (Sigma-Aldrich) were blockedwith 10% BSA in PBS before the addition of the same titrated concentra-tions of Abs as noted before. B cell responders from C57BL/6 mice wereenriched from spleen and lymph node with Lymphocyte Separation Media(Mediatech) and then purified using the Mouse B cell Isolation Kit andVarioMACS Separator (Miltenyi Biotec). Purity was confirmed to be�98% B220� B cells. Purified B cells were added to both immobilized andsoluble stimulus plates in parallel and, after 72 h in culture, were pulsed for12 h with 0.5 �Ci/well [methyl-3H]thymidine. Cells were harvested, and[methyl-3H]thymidine incorporation was measured on a standard betacounter microplate reader (Biotek).
Bone marrow chimerism
BALB/c donor bone marrow was flushed from tibiae and femora, andC57BL/6 recipient mice were transfused on days 0 and 6 with 2 � 107
donor marrow cells i.v, along with a single 500-mg dose of busulfan on day5. On days 0, 2, 4, and 6, mice received 500 �g i.p. of human CTLA4-Ig(Bristol-Myers Squibb) and 7E1-G1, 7E1-G2b, isotype-matched control, orhamster anti-mouse CD154 mAb (MR1: BioXCell). Chimerism was mon-itored by flow cytometric analysis of H-2Kd-expressing donor cells.
Skin grafting
Full-thickness ear and tail skin grafts (�1 cm2) were transplanted onto thedorsal thorax of recipient mice and secured with a plastic adhesive bandagefor 6 days. As indicated, recipients received 500 �g i.p. of CTLA4-Ig and7E1-G1, 7E1-G2b, isotype-matched control, or anti-CD154 on days 0, 2, 4,and 6. Graft survival was monitored by daily visual inspection and rejec-tion was defined as the complete loss of viable epidermal tissue.
BrdU labeling for in vivo proliferation
Mice were treated on days 0, 2, 4, and 6 with 500 �g i.p. of 7E1-G1,7E1-G2b, isotype-matched control, or FGK4.5. On day 7, mice werepulsed with 2 mg i.p. of BrdU and sacrificed 4 h later. BrdU incorporationwas detected using the allophycocyanin-BrdU flow kit according to themanufacturer’s protocol (BD Biosciences).
Flow cytometry for phenotype and absolute number
At the indicated time points, animals were sacrificed, spleens were har-vested, and single-cell suspensions were prepared in FACS buffer. Unlesspreviously stated, Abs obtained from BD Biosciences included the follow-ing: anti-CD8-, -CD11b-, -CD19-, -CD43-, -CD44-, -CD80-, -CD86-, -H-2Kd-, -I-Ab-FITC; -CD4-, -CD19-, -CD43-, -CD45R-, -CD70-, -CD86-,4-1BBL-, -V�5-, -V�8-, -V�11-PE; -CD3-, -CD4-, -CD11c-, -CD19-,-CD25-, -CD44-, and -CD80-allophycocyanin; anti-CD3-, anti-CD4-Pacific Blue; anti-CD8-PerCP; anti-CD11b-, -CD45R-, anti-CD69-PerCP-Cy5.5; and I-Ab-biotin.
Abs obtained from Invitrogen included: anti-CD8-, anti-CD45R-, andstreptavidin-Pacific Orange. Abs obtained from eBioscience included anti-CD19-Pacific Blue. Where noted, absolute numbers of splenocyte subsetswere determined by TruCount bead analysis according to the manufactur-er’s instructions (BD Biosciences). Flow cytometric data were analyzedusing FlowJo Software (Treestar).
Statistical analysis
Time to skin graft rejection was represented by Kaplan-Meier survivalcurves, and comparison of graft survival was calculated using log-rankassessment. Titration curves were compared by nonlinear regression bestcurve fit using a sigmoidal dose-response (variable slope) equation over theexponential portion of each data set. Additional statistical analysis wasperformed using a two-tailed unpaired Student t test. A p value of �0.05was considered significant for all tests. (GraphPad Software).
2 Abbreviations used in this paper: 7-AAD, 7-aminoactinomycin D; sCD154, solubleCD154 fusion protein; mOVA, murine OVA; MST, median survival time.
1626 EFFICACY OF ANTI-CD40 mAbs IN TRANSPLANTATION
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Results7E1-G1 and 7E1-G2b possess equivalent affinities for murineCD40 and both effectively inhibit the interaction of CD154with CD40
Preliminary characterization during the development of the 7E1variants suggested that 7E1-G2b was a legitimate isotype switchvariant of 7E1-G1 and thus should possess the same specificity andaffinity for CD40 (37). To confirm this, three sets of tests wereperformed. First, N-terminal peptide sequencing of the H chainshowed that both Abs were identical for the first 35 aas. Second,PCR primers specific for the variable H chain CDRs of 7E1-G1yielded a band of the appropriate size from cDNA obtained fromeither 7E1 Ab, but not from two unrelated Abs. Finally, ELISA-based measurement of the binding activity of purified 7E1-G1 and7E1-G2b to immobilized murine CD40 gave essentially identicaltitration curves (37). To corroborate this finding, we comparedtheir binding affinities to CD40 in vitro, using the B cell line M12that constitutively expresses a high level of murine CD40 on itssurface (data not shown). Cultured M12 cells were incubated withincreasing concentrations of 7E1-G1, 7E1-G2b, or FGK4.5, astrong mouse CD40 agonist (42), and bound Ab was subsequentlydetected with a fluorescently labeled polyclonal anti-rat IgG sec-ondary Ab. The resulting titration curves for the 7E1 variants wereessentially overlapping (EC50 81.63 ng/ml), whereas the titrationcurve for the unrelated Ab FGK4.5 was distinct from either of the7E1 variants (EC50 95.00 ng/ml; Fig. 1A). These results suggestedthat any potential alterations in the idiotypic region of the 7E1variants did not significantly affect its affinity for mouse CD40.
We next sought to assess the ability of the 7E1 variants to com-pete with soluble CD154 for binding to cell-bound CD40. Wehypothesized that both 7E1-G1 and 7E1-G2b would inhibit thisinteraction to a similar degree. To test this, M12 B cells wereincubated with increasing concentrations of 7E1-G1, 7E1-G2b, orFGK4.5, along with a constant amount of FLAG-tagged solubleCD154 fusion protein (sCD154). Bound sCD154 then was de-tected with a fluorescently labeled anti-FLAG secondary Ab. Themean fluorescence intensity observed at each concentration ofcompetitor Ab was divided by the mean fluorescence intensity ofM12 cells cultured with sCD154 alone. The competitive inhibitioncurves generated for the 7E1 variants were again essentially over-lapping (EC50 0.238 �g/ml) and distinct from FGK4.5 (EC50 1.362�g/ml; Fig. 1B). These results further confirmed that the idiotypicregion of 7E1-G2b had not undergone significant changes that im-pacted CD154 binding and also showed that both 7E1-G1 and7E1-G2b had the same ability to block the interaction betweenCD40 and CD154.
Anti-CD40 treatment combined with CTLA4-Ig promotesallogeneic chimerism and skin graft survival
Previous murine studies from our laboratory and others have dem-onstrated the efficacy of costimulatory blockade-based protocolsfor promoting hemopoietic chimerism and skin graft survival (9,43–45). These have involved the use of an anti-CD154-bindingmAb paired with the CD28 pathway-blocking reagent CTLA4-Ig.Here we sought to determine whether an anti-CD40 mAb couldeffectively substitute for anti-CD154 in costimulation blockade-based tolerance induction regimens. First, groups of C57BL/6mice were grafted with allogeneic BALB/c skin and treated withCTLA4-Ig and one of 7E1-G1, 7E1-G2b, anti-CD154, or isotype-matched control on days 0, 2, 4, and 6. Untreated recipientspromptly rejected their grafts with a median survival time (MST)of 11 days. Recipients treated with CTLA4-Ig/7E1-G2b or anti-CD154 showed a modest prolongation of skin graft survival (MST
27 and 23 days, respectively) compared with CTLA4-Ig/7E1-G1(MST 18 days) in the stringent BALB/c to C57BL/6 model, in-dicative of the costimulation blockade-resistant phenotype (46)seen with this particular strain combination of donor and recipient(Fig. 2A). In the more permissive BALB/c to C3H to model (47),graft survival also was prolonged with CTLA4-Ig combined with7E1-G2b or anti-CD154 treatment (MST �75 days), as comparedwith CTLA4-Ig/7E1-G1 treatment (MST 29 days) or untreated re-cipients (MST 19 days; Fig. 2B).
Next, we sought to test the 7E1 variants in a model of allogeneicchimerism induction. C57BL/6 mice received BALB/c bone mar-row (2 � 107 cells i.v. on days 0 and 6) and BALB/c skin graftson day 0, as well as busulfan, an alkylating agent toxic to hemo-poietic stem cells, on day 5. Mice received CTLA4-Ig and 7E1-G1,7E1-G2b, anti-CD154, or isotype-matched control on days 0, 2, 4,and 6. Peripheral blood was monitored for the presence of donor-derived B and T cells. The percentage of donor B220� B cells(Fig. 2C) and CD4� T cells (Fig. 2D) reached similar levels in7E1-G2b- and anti-CD154-treated groups, whereas 7E1-G1 andisotype-treated animals completely failed to sustain significant do-nor chimerism. The successful induction of hemopoietic chimer-ism directly correlated to skin graft survival, as both 7E1-G2b- andanti-CD154-treated recipients enjoyed prolonged graft survival(MST �75 days). In contrast, 7E1-G1-treated recipients rapidlyrejected their skin grafts (MST 16 days; Fig. 2E).
We also tracked the frequency of donor reactive V�11� andV�5.1/5.2�CD4� T cells in these chimeric recipients. T cells pos-sessing these V� segments are normally deleted by negative se-lection in the thymus of BALB/c mice due to their high affinity forendogenous retroviral superantigens bound to I-E MHC molecules.Wild-type C57BL/6 mice do not express I-E; V�11�, and V�5.1/
FIGURE 1. 7E1-G1 and 7E1-G2b have an equivalent affinity for murineCD40 and effectively inhibit the binding of CD154. A, M12 B cells wereincubated with the indicated concentrations of 7E1-G1, 7E1-G2b, orFGK4.5. Bound Ab was detected with a PE-labeled polyclonal anti-rat IgGsecondary Ab by flow cytometry. The values are presented as the percent-age of maximum fluorescence observed for each Ab. B, M12 B cells wereincubated with the indicated concentrations of 7E1-G1, 7E1-G2b, orFGK4.5 along with 10 �g/ml FLAG-tagged soluble CD154 fusion protein.A FITC-labeled anti-FLAG secondary Ab was used to detect boundCD154. The values represent a percentage of the maximum fluorescenceobserved with sCD154 alone. Data points represent four independent ex-periments with triplicate samples.
1627The Journal of Immunology
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
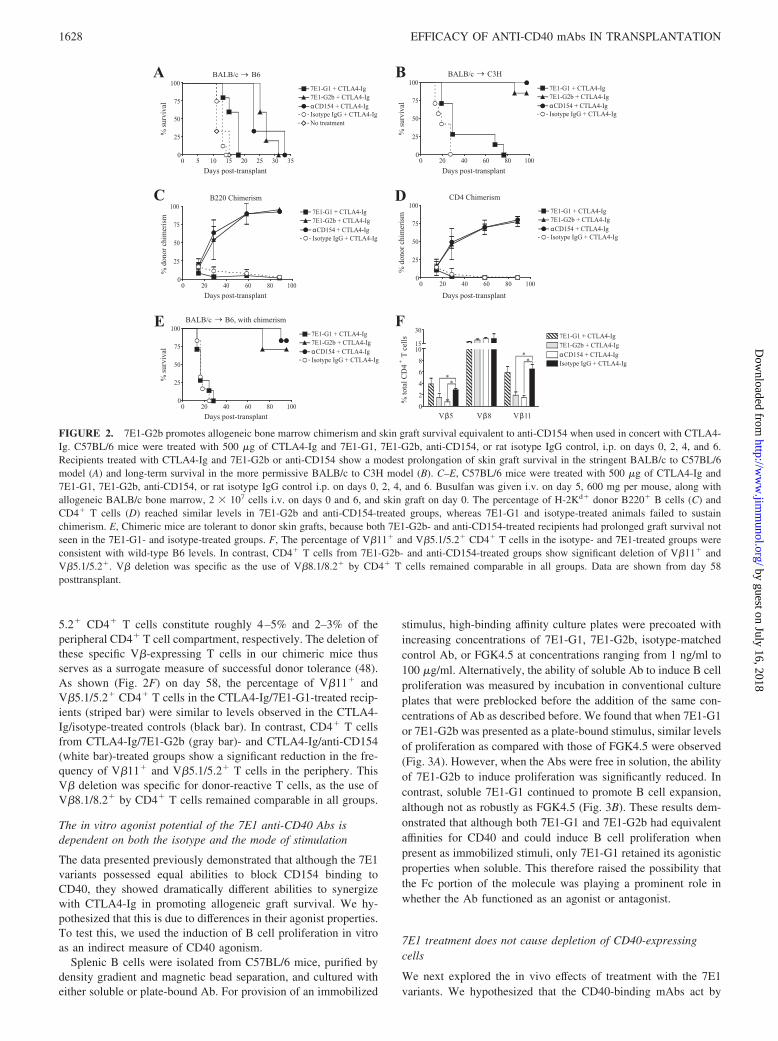
5.2� CD4� T cells constitute roughly 4–5% and 2–3% of theperipheral CD4� T cell compartment, respectively. The deletion ofthese specific V�-expressing T cells in our chimeric mice thusserves as a surrogate measure of successful donor tolerance (48).As shown (Fig. 2F) on day 58, the percentage of V�11� andV�5.1/5.2� CD4� T cells in the CTLA4-Ig/7E1-G1-treated recip-ients (striped bar) were similar to levels observed in the CTLA4-Ig/isotype-treated controls (black bar). In contrast, CD4� T cellsfrom CTLA4-Ig/7E1-G2b (gray bar)- and CTLA4-Ig/anti-CD154(white bar)-treated groups show a significant reduction in the fre-quency of V�11� and V�5.1/5.2� T cells in the periphery. ThisV� deletion was specific for donor-reactive T cells, as the use ofV�8.1/8.2� by CD4� T cells remained comparable in all groups.
The in vitro agonist potential of the 7E1 anti-CD40 Abs isdependent on both the isotype and the mode of stimulation
The data presented previously demonstrated that although the 7E1variants possessed equal abilities to block CD154 binding toCD40, they showed dramatically different abilities to synergizewith CTLA4-Ig in promoting allogeneic graft survival. We hy-pothesized that this is due to differences in their agonist properties.To test this, we used the induction of B cell proliferation in vitroas an indirect measure of CD40 agonism.
Splenic B cells were isolated from C57BL/6 mice, purified bydensity gradient and magnetic bead separation, and cultured witheither soluble or plate-bound Ab. For provision of an immobilized
stimulus, high-binding affinity culture plates were precoated withincreasing concentrations of 7E1-G1, 7E1-G2b, isotype-matchedcontrol Ab, or FGK4.5 at concentrations ranging from 1 ng/ml to100 �g/ml. Alternatively, the ability of soluble Ab to induce B cellproliferation was measured by incubation in conventional cultureplates that were preblocked before the addition of the same con-centrations of Ab as described before. We found that when 7E1-G1or 7E1-G2b was presented as a plate-bound stimulus, similar levelsof proliferation as compared with those of FGK4.5 were observed(Fig. 3A). However, when the Abs were free in solution, the abilityof 7E1-G2b to induce proliferation was significantly reduced. Incontrast, soluble 7E1-G1 continued to promote B cell expansion,although not as robustly as FGK4.5 (Fig. 3B). These results dem-onstrated that although both 7E1-G1 and 7E1-G2b had equivalentaffinities for CD40 and could induce B cell proliferation whenpresent as immobilized stimuli, only 7E1-G1 retained its agonisticproperties when soluble. This therefore raised the possibility thatthe Fc portion of the molecule was playing a prominent role inwhether the Ab functioned as an agonist or antagonist.
7E1 treatment does not cause depletion of CD40-expressingcells
We next explored the in vivo effects of treatment with the 7E1variants. We hypothesized that the CD40-binding mAbs act by
FIGURE 2. 7E1-G2b promotes allogeneic bone marrow chimerism and skin graft survival equivalent to anti-CD154 when used in concert with CTLA4-Ig. C57BL/6 mice were treated with 500 �g of CTLA4-Ig and 7E1-G1, 7E1-G2b, anti-CD154, or rat isotype IgG control, i.p. on days 0, 2, 4, and 6.Recipients treated with CTLA4-Ig and 7E1-G2b or anti-CD154 show a modest prolongation of skin graft survival in the stringent BALB/c to C57BL/6model (A) and long-term survival in the more permissive BALB/c to C3H model (B). C–E, C57BL/6 mice were treated with 500 �g of CTLA4-Ig and7E1-G1, 7E1-G2b, anti-CD154, or rat isotype IgG control i.p. on days 0, 2, 4, and 6. Busulfan was given i.v. on day 5, 600 mg per mouse, along withallogeneic BALB/c bone marrow, 2 � 107 cells i.v. on days 0 and 6, and skin graft on day 0. The percentage of H-2Kd� donor B220� B cells (C) andCD4� T cells (D) reached similar levels in 7E1-G2b and anti-CD154-treated groups, whereas 7E1-G1 and isotype-treated animals failed to sustainchimerism. E, Chimeric mice are tolerant to donor skin grafts, because both 7E1-G2b- and anti-CD154-treated recipients had prolonged graft survival notseen in the 7E1-G1- and isotype-treated groups. F, The percentage of V�11� and V�5.1/5.2� CD4� T cells in the isotype- and 7E1-treated groups wereconsistent with wild-type B6 levels. In contrast, CD4� T cells from 7E1-G2b- and anti-CD154-treated groups show significant deletion of V�11� andV�5.1/5.2�. V� deletion was specific as the use of V�8.1/8.2� by CD4� T cells remained comparable in all groups. Data are shown from day 58posttransplant.
1628 EFFICACY OF ANTI-CD40 mAbs IN TRANSPLANTATION
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

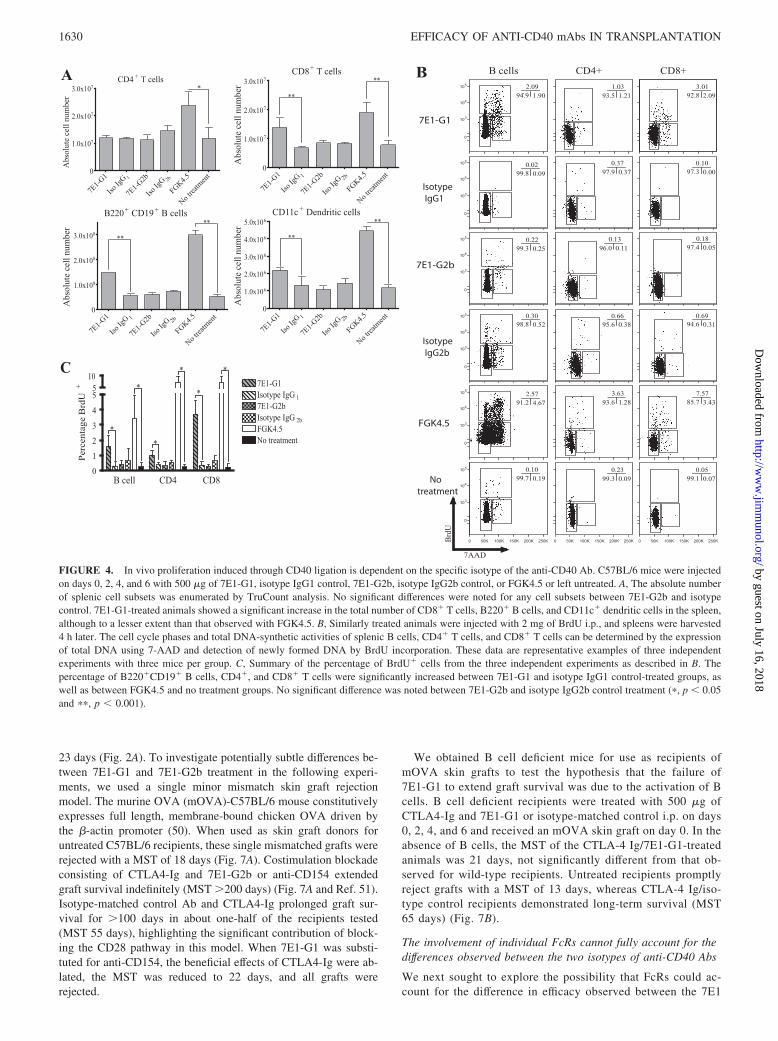
depleting CD40-expressing cells, resulting in a dearth of cells re-maining to stimulate graft rejection. To test this, mice were in-jected on days 0, 2, 4, and 6 either 7E1-G1, 7E1-G2b, isotype-matched control Ab, or FGK4.5 or were left untreated. On day 7,mice were sacrificed, and the absolute numbers of various cellsubsets were enumerated from the spleen by TruCount analysis.We found that treatment with 7E1-G2b did not lead to B cell,dendritic cell, or T cell depletion, because the absolute numbers ofeach did not significantly differ from isotype control or untreatedanimals (Fig. 4A). Assessment of cell numbers in the lymph nodes,bone marrow, and blood showed trends similar to those observedin the spleen (data not shown). However, mice treated with7E1-G1 showed a statistically significant 3-fold increase in CD8�
T cells and B cells and a �2-fold increase in dendritic cells, withnumbers close to those seen with FGK4.5.
To determine whether these increases in cell numbers were dueto increased trafficking to the spleen or de novo cell division,groups of mice were given the same treatment as described pre-viously and pulsed on day 7 with 2 mg of BrdU for 4 h. BrdU,when paired with the DNA stain 7-AAD, can reveal both the pres-ence of dividing cells and their specific state in the cell cycle. Bcells, CD8� T cells, and, to a lesser extent, CD4� T cells show aclear population of cells progressing through the cell cycle with7E1-G1 and FGK4.5 treatment, but not in 7E1-G2b or isotypecontrol-treated animals (Fig. 4B). This brief snapshot of divisionrepresents a 5-fold increase in proliferating B cells, a 3-fold in-crease in proliferating CD4� T cells, and an 11-fold increase inproliferating CD8� T cells with 7E1-G1 as compared with isotypecontrol treatment (Fig. 4C). Thus, the ability of 7E1-G2b to pro-mote graft survival is not due to the depletion of CD40-expressingcells. In contrast, 7E1-G1 treatment leads to a significant increase
in the total number of T cells, B cells, and dendritic cells in thespleen.
7E1-G1, but not 7E1-G2b, induces the expression of activationmarkers on APCs
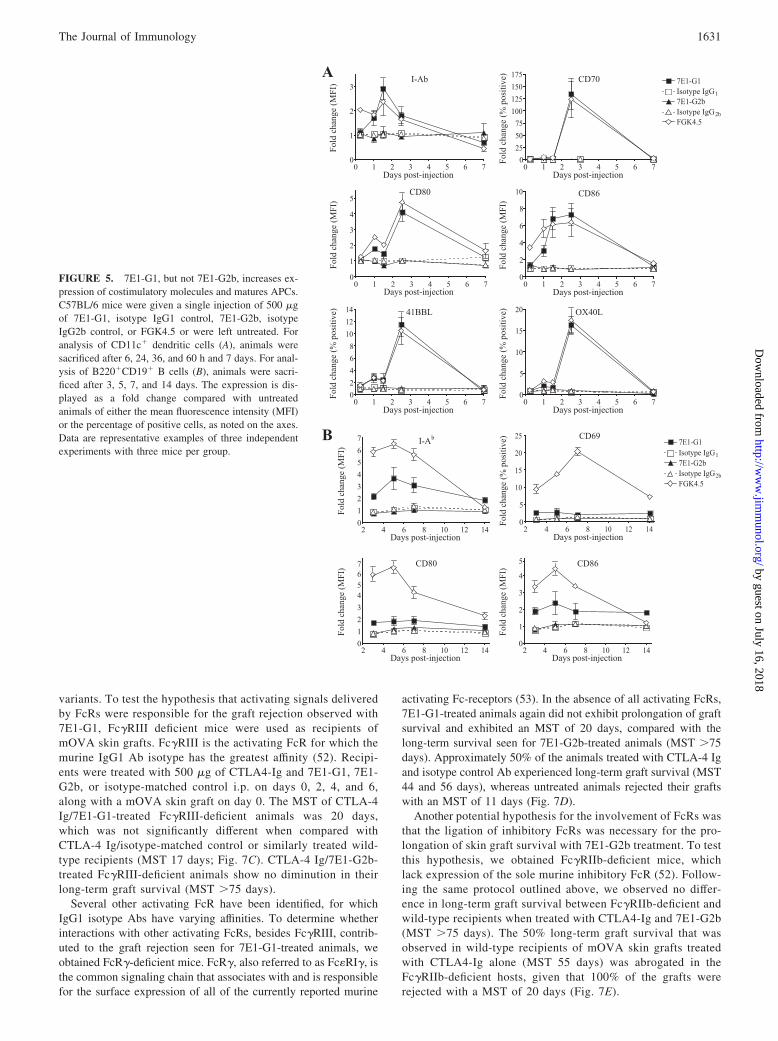
Due to the findings that 7E1-G1 induced proliferation in vitro andin vivo, we hypothesized that administration of 7E1-G1, but not7E1-G2b, results in the maturation of APCs. To examine this,C57BL/6 mice were given a single injection of 7E1-G1, 7E1-G2b,isotype-matched control Ab, or FGK4.5. Mice were sacrificed atvarious time points, and splenocytes were stained for the presenceof activation markers on CD11c� dendritic cells. 7E1-G1 treat-ment led to a 3-fold increase in MHC class II expression, a 4-foldincrease in CD80 expression and a 7-fold increase in the expres-sion of CD86 on dendritic cells, closely mirroring the level ofexpression observed for FGK4.5-treated animals (Fig. 5A). Thepercentage of dendritic cells expressing CD70 increased by �100-fold, 41BBL increased by 12-fold, and OX40L increased by 15-fold in 7E1-G1 and FGK4.5 but not 7E1-G2b-treated animals.
B220�CD19� B cells also were measured for activation mark-ers in these same treated animals. As noted for dendritic cells, Bcells in 7E1-G1-treated animals showed a 3-fold increase in theexpression of MHC class II, and an almost 2-fold increase in bothCD80 and CD86 (Fig. 5B). The percentage of CD69-expressing Bcells increased roughly 2-fold immediately after 7E1-G1 treat-ments. This was in contrast to what was observed in 7E1-G2b-treated animals, in which expression patterns did not differ fromisotype-treated animals.
Increased expression of activation markers on T cells aftertreatment with 7E1-G1, but not 7E1-G2b
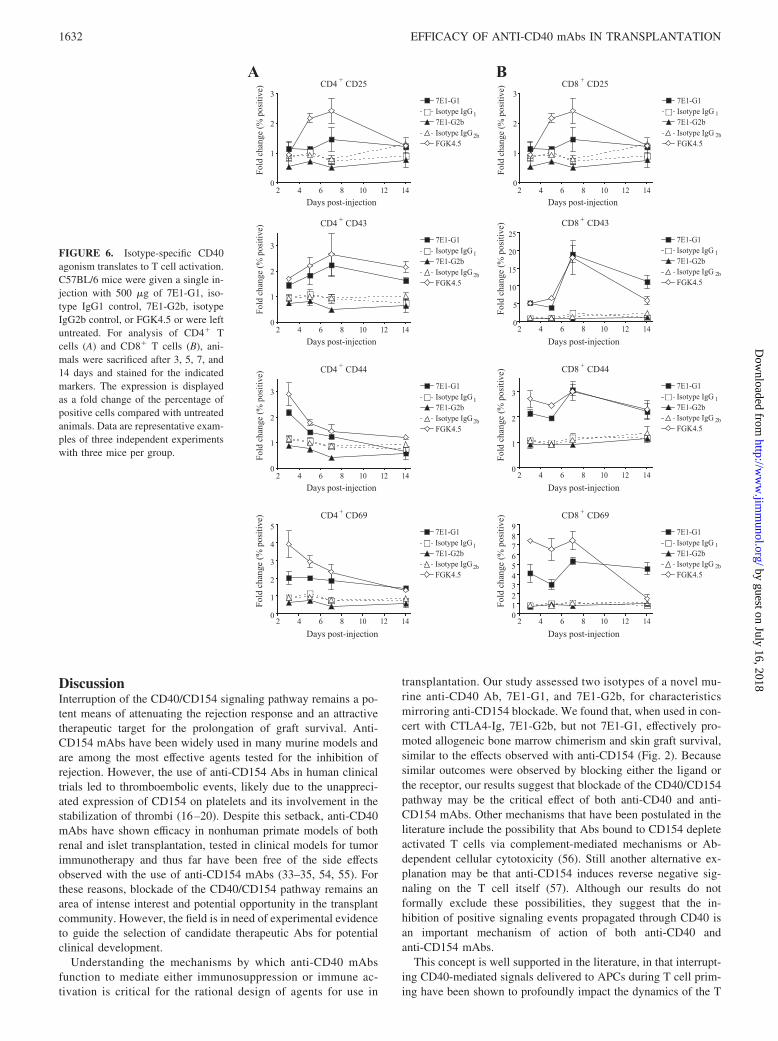
We hypothesized that the effects of the anti-CD40 Abs on APCswould subsequently cause T cell activation as well. To test this,C57BL/6 mice were given a single injection of either 7E1-G1,7E1-G2b, isotype-matched control Ab, or FGK4.5. Mice were sac-rificed at the indicated time points and stained for the expression ofactivation markers on CD3� T cell subsets. CD4� T cells from7E1-G1-treated mice showed an almost 2-fold increase in the per-centage of cells expressing CD25 and CD43, which peak by day 7(Fig. 6A). The percentage of cells expressing CD44 and CD69increased by 2-fold, which can be seen as early as day 3. In con-trast, the percentage of CD4� T cells expressing each of theseactivation markers for 7E1-G2b-treated animals was at or belowwhat was observed for isotype control-treated animals. The per-centage of CD8� T cells from 7E1-G1-treated animals expressingCD25 increased by 4-fold, CD43 by almost 20-fold, CD44 by3-fold, and CD69 by 5-fold, all of which peak by day 7 (Fig. 6B).7E1-G2b did not induce any notable activation marker expressionon CD8� T cells beyond that seen for isotype-treated animals.
The presence of activated B cells alone cannot explain thedifferences observed between the two isotypes of anti-CD40 Abs
Resting B cells make up a large percentage of the total cellspresent in the spleen and express a high basal level of CD40 (49).As we have shown, B cells aggressively responded to 7E1-G1 bothin vivo and in vitro, up-regulating activation markers and robustlyproliferating. We therefore hypothesized that the prominent acti-vation of B cells by 7E1-G1 created an overall stimulated envi-ronment that supercedes the effects of CTLA4-Ig, thus leading toprompt graft rejection. To test this, we made use of a differentmodel of skin graft rejection than previously presented herein. Un-treated C57BL/6 mice reject BALB/c skin grafts with a MST of 11days, whereas costimulation blockade modestly extends survival to
FIGURE 3. 7E1-G2b induces B cell proliferation in vitro only whenpresent as an immobilized stimulus. A, Purified B cells were added tohigh-binding-affinity 96-well plates that had been coated with the indicatedconcentrations of 7E1-G1, isotype IgG1 control, 7E1-G2b, isotype IgG2bcontrol, or FGK4.5. B, In parallel, purified B cells were added to conven-tional 96-well plates, preblocked with BSA, to which the same Ab con-centrations were added. Following 72 h of incubation at 37°C, all plateswere pulsed for an additional 18 h with 0.5 �Ci/well of [methyl-3H]thy-midine. Actively dividing cells that had taken up the radioactive probewere quantified by scintillation counter. Representative data with triplicatesamples, and experiments were repeated three times.
1629The Journal of Immunology
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
23 days (Fig. 2A). To investigate potentially subtle differences be-tween 7E1-G1 and 7E1-G2b treatment in the following experi-ments, we used a single minor mismatch skin graft rejectionmodel. The murine OVA (mOVA)-C57BL/6 mouse constitutivelyexpresses full length, membrane-bound chicken OVA driven bythe �-actin promoter (50). When used as skin graft donors foruntreated C57BL/6 recipients, these single mismatched grafts wererejected with a MST of 18 days (Fig. 7A). Costimulation blockadeconsisting of CTLA4-Ig and 7E1-G2b or anti-CD154 extendedgraft survival indefinitely (MST �200 days) (Fig. 7A and Ref. 51).Isotype-matched control Ab and CTLA4-Ig prolonged graft sur-vival for �100 days in about one-half of the recipients tested(MST 55 days), highlighting the significant contribution of block-ing the CD28 pathway in this model. When 7E1-G1 was substi-tuted for anti-CD154, the beneficial effects of CTLA4-Ig were ab-lated, the MST was reduced to 22 days, and all grafts wererejected.
We obtained B cell deficient mice for use as recipients ofmOVA skin grafts to test the hypothesis that the failure of7E1-G1 to extend graft survival was due to the activation of Bcells. B cell deficient recipients were treated with 500 �g ofCTLA4-Ig and 7E1-G1 or isotype-matched control i.p. on days0, 2, 4, and 6 and received an mOVA skin graft on day 0. In theabsence of B cells, the MST of the CTLA-4 Ig/7E1-G1-treatedanimals was 21 days, not significantly different from that ob-served for wild-type recipients. Untreated recipients promptlyreject grafts with a MST of 13 days, whereas CTLA-4 Ig/iso-type control recipients demonstrated long-term survival (MST65 days) (Fig. 7B).
The involvement of individual FcRs cannot fully account for thedifferences observed between the two isotypes of anti-CD40 Abs
We next sought to explore the possibility that FcRs could ac-count for the difference in efficacy observed between the 7E1
FIGURE 4. In vivo proliferation induced through CD40 ligation is dependent on the specific isotype of the anti-CD40 Ab. C57BL/6 mice were injectedon days 0, 2, 4, and 6 with 500 �g of 7E1-G1, isotype IgG1 control, 7E1-G2b, isotype IgG2b control, or FGK4.5 or left untreated. A, The absolute numberof splenic cell subsets was enumerated by TruCount analysis. No significant differences were noted for any cell subsets between 7E1-G2b and isotypecontrol. 7E1-G1-treated animals showed a significant increase in the total number of CD8� T cells, B220� B cells, and CD11c� dendritic cells in the spleen,although to a lesser extent than that observed with FGK4.5. B, Similarly treated animals were injected with 2 mg of BrdU i.p., and spleens were harvested4 h later. The cell cycle phases and total DNA-synthetic activities of splenic B cells, CD4� T cells, and CD8� T cells can be determined by the expressionof total DNA using 7-AAD and detection of newly formed DNA by BrdU incorporation. These data are representative examples of three independentexperiments with three mice per group. C, Summary of the percentage of BrdU� cells from the three independent experiments as described in B. Thepercentage of B220�CD19� B cells, CD4�, and CD8� T cells were significantly increased between 7E1-G1 and isotype IgG1 control-treated groups, aswell as between FGK4.5 and no treatment groups. No significant difference was noted between 7E1-G2b and isotype IgG2b control treatment (�, p � 0.05and ��, p � 0.001).
1630 EFFICACY OF ANTI-CD40 mAbs IN TRANSPLANTATION
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
variants. To test the hypothesis that activating signals deliveredby FcRs were responsible for the graft rejection observed with7E1-G1, Fc�RIII deficient mice were used as recipients ofmOVA skin grafts. Fc�RIII is the activating FcR for which themurine IgG1 Ab isotype has the greatest affinity (52). Recipi-ents were treated with 500 �g of CTLA4-Ig and 7E1-G1, 7E1-G2b, or isotype-matched control i.p. on days 0, 2, 4, and 6,along with a mOVA skin graft on day 0. The MST of CTLA-4Ig/7E1-G1-treated Fc�RIII-deficient animals was 20 days,which was not significantly different when compared withCTLA-4 Ig/isotype-matched control or similarly treated wild-type recipients (MST 17 days; Fig. 7C). CTLA-4 Ig/7E1-G2b-treated Fc�RIII-deficient animals show no diminution in theirlong-term graft survival (MST �75 days).
Several other activating FcR have been identified, for whichIgG1 isotype Abs have varying affinities. To determine whetherinteractions with other activating FcRs, besides Fc�RIII, contrib-uted to the graft rejection seen for 7E1-G1-treated animals, weobtained FcR�-deficient mice. FcR�, also referred to as Fc�RI�, isthe common signaling chain that associates with and is responsiblefor the surface expression of all of the currently reported murine
activating Fc-receptors (53). In the absence of all activating FcRs,7E1-G1-treated animals again did not exhibit prolongation of graftsurvival and exhibited an MST of 20 days, compared with thelong-term survival seen for 7E1-G2b-treated animals (MST �75days). Approximately 50% of the animals treated with CTLA-4 Igand isotype control Ab experienced long-term graft survival (MST44 and 56 days), whereas untreated animals rejected their graftswith an MST of 11 days (Fig. 7D).
Another potential hypothesis for the involvement of FcRs wasthat the ligation of inhibitory FcRs was necessary for the pro-longation of skin graft survival with 7E1-G2b treatment. To testthis hypothesis, we obtained Fc�RIIb-deficient mice, whichlack expression of the sole murine inhibitory FcR (52). Follow-ing the same protocol outlined above, we observed no differ-ence in long-term graft survival between Fc�RIIb-deficient andwild-type recipients when treated with CTLA4-Ig and 7E1-G2b(MST �75 days). The 50% long-term graft survival that wasobserved in wild-type recipients of mOVA skin grafts treatedwith CTLA4-Ig alone (MST 55 days) was abrogated in theFc�RIIb-deficient hosts, given that 100% of the grafts wererejected with a MST of 20 days (Fig. 7E).
FIGURE 5. 7E1-G1, but not 7E1-G2b, increases ex-pression of costimulatory molecules and matures APCs.C57BL/6 mice were given a single injection of 500 �gof 7E1-G1, isotype IgG1 control, 7E1-G2b, isotypeIgG2b control, or FGK4.5 or were left untreated. Foranalysis of CD11c� dendritic cells (A), animals weresacrificed after 6, 24, 36, and 60 h and 7 days. For anal-ysis of B220�CD19� B cells (B), animals were sacri-ficed after 3, 5, 7, and 14 days. The expression is dis-played as a fold change compared with untreatedanimals of either the mean fluorescence intensity (MFI)or the percentage of positive cells, as noted on the axes.Data are representative examples of three independentexperiments with three mice per group.
1631The Journal of Immunology
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
DiscussionInterruption of the CD40/CD154 signaling pathway remains a po-tent means of attenuating the rejection response and an attractivetherapeutic target for the prolongation of graft survival. Anti-CD154 mAbs have been widely used in many murine models andare among the most effective agents tested for the inhibition ofrejection. However, the use of anti-CD154 Abs in human clinicaltrials led to thromboembolic events, likely due to the unappreci-ated expression of CD154 on platelets and its involvement in thestabilization of thrombi (16–20). Despite this setback, anti-CD40mAbs have shown efficacy in nonhuman primate models of bothrenal and islet transplantation, tested in clinical models for tumorimmunotherapy and thus far have been free of the side effectsobserved with the use of anti-CD154 mAbs (33–35, 54, 55). Forthese reasons, blockade of the CD40/CD154 pathway remains anarea of intense interest and potential opportunity in the transplantcommunity. However, the field is in need of experimental evidenceto guide the selection of candidate therapeutic Abs for potentialclinical development.
Understanding the mechanisms by which anti-CD40 mAbsfunction to mediate either immunosuppression or immune ac-tivation is critical for the rational design of agents for use in
transplantation. Our study assessed two isotypes of a novel mu-rine anti-CD40 Ab, 7E1-G1, and 7E1-G2b, for characteristicsmirroring anti-CD154 blockade. We found that, when used in con-cert with CTLA4-Ig, 7E1-G2b, but not 7E1-G1, effectively pro-moted allogeneic bone marrow chimerism and skin graft survival,similar to the effects observed with anti-CD154 (Fig. 2). Becausesimilar outcomes were observed by blocking either the ligand orthe receptor, our results suggest that blockade of the CD40/CD154pathway may be the critical effect of both anti-CD40 and anti-CD154 mAbs. Other mechanisms that have been postulated in theliterature include the possibility that Abs bound to CD154 depleteactivated T cells via complement-mediated mechanisms or Ab-dependent cellular cytotoxicity (56). Still another alternative ex-planation may be that anti-CD154 induces reverse negative sig-naling on the T cell itself (57). Although our results do notformally exclude these possibilities, they suggest that the in-hibition of positive signaling events propagated through CD40 isan important mechanism of action of both anti-CD40 andanti-CD154 mAbs.
This concept is well supported in the literature, in that interrupt-ing CD40-mediated signals delivered to APCs during T cell prim-ing have been shown to profoundly impact the dynamics of the T
FIGURE 6. Isotype-specific CD40agonism translates to T cell activation.C57BL/6 mice were given a single in-jection with 500 �g of 7E1-G1, iso-type IgG1 control, 7E1-G2b, isotypeIgG2b control, or FGK4.5 or were leftuntreated. For analysis of CD4� Tcells (A) and CD8� T cells (B), ani-mals were sacrificed after 3, 5, 7, and14 days and stained for the indicatedmarkers. The expression is displayedas a fold change of the percentage ofpositive cells compared with untreatedanimals. Data are representative exam-ples of three independent experimentswith three mice per group.
1632 EFFICACY OF ANTI-CD40 mAbs IN TRANSPLANTATION
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

cell:APC interaction. Among these effects are limitation of theup-regulation of activation markers such as CD80 and CD86 (58)and a decrease in the production of proinflammatory cytokinessuch as IL-8, IL-12, MIP-1�, and TNF-� (59, 60). CD40 signalingis also important for APC survival, mediating the up-regulation ofthe antiapoptotic molecules Bcl-2 and Bcl-xL (61–63). Althoughwe did not observe active depletion of CD40-expressing APCsfollowing treatment with 7E1-G2b, the blockade of survival sig-nals normally provided by the CD40 pathway may be detrimentalto APC longevity and the duration of Ag presentation (23, 64, 65).The contributions of these effects may culminate in the deviationof responding T cells from an effector phenotype and direct themtoward an alternative program of anergy (66), abortive prolifera-tion (67), apoptosis (68), or a regulatory cell phenotype (69). Theextent to which these mechanisms play a role in transplant toler-ance mediated by either anti-CD154 or 7E1-G2b remains an im-portant area of future investigation.
We found that the anti-CD40 mAb 7E1-G2b effectively syner-gized with CTLA-4 Ig to prolong graft survival (Fig. 2). In con-trast, treatment with 7E1-G1 failed to prolong graft survival (Fig.2), despite the fact that these Abs possess identical idiotypic re-gions. These results highlight the ability of an appropriately de-signed anti-CD40 mAb to be used therapeutically as either immu-nosuppression/tolerance induction agents or immunostimulatoryadjuvants. Both Abs have equivalent affinity for their Ag and areable to inhibit the interaction between CD40 and CD154 to thesame extent (Fig. 1). Thus, the dichotomy in their functionalitymay be explained by the fact that whereas both Abs behaved asagonists when plate bound (Fig. 3A), 7E1-G1 but not 7E1-G2bacted as a CD40 agonist to increase B cell proliferation and acti-vation when present in soluble form both in vitro and in vivo (Figs.3B and 4–6). Prior studies have focused for the most part on dif-ferences in idiotypic regions as being the primary determinant thatconferred either activating or antagonistic properties upon a par-
ticular mAb; however, our results suggest that monoclonals withidentical epitope specificity can possess agonist or antagonist prop-erties in vivo depending on the isotype alone. Our results alsosuggest that assessment of the behavior of an anti-CD40 mAb asan agonist with regard to B cell proliferation in soluble form maybe a useful in vitro screening assay to help guide the selection ofcandidate anti-CD40 mAbs likely to function as blocking agents invivo. We would propose that any candidate anti-CD40 mAbsshould be evaluated based on their 1) ability to prevent ligandbinding in vitro and 2) failure to induce B cell proliferation whenused in soluble form in vitro. These results therefore demonstratethe therapeutic potential of an appropriately designed anti-CD40blocking agent in synergizing with CTLA-4 Ig to promote long-term graft survival. Although these results help to establish theefficacy of anti-CD40 mAbs in transplantation and provide aspringboard for mechanistic studies of these Abs, mouse modelsare sometimes limited in their ability to predict side effects relatedto Ab administration in human subjects (17, 70–72). Detailedsafety studies of each individual Ab clonotype are warranted be-fore clinical translation.
Despite the findings that 7E1-G1 but not 7E1-G2b induced Bcell proliferation and activation both in vitro and in vivo, we foundthat B cells were not required for the prorejection phenotype as-sociated with the 7E1-G1 mAb, as 7E1-G1 still exhibited this pro-rejection phenotype in B cell-deficient recipients (Fig. 7). Like-wise, we found no evidence of significant depletion of B cells (orother CD40-expressing cells) with either 7E1 variant; therefore,our data further suggest that depletion of CD40-expressing APC isnot the major mechanism by which 7E1-G2b promotes long-termgraft survival (Fig. 4). Although studies investigating the effects ofChi220, a chimeric anti-human CD40 mAb, did demonstrate sig-nificant depletion of B cells in nonhuman primates, those studiesalso indicated that B cell depletion was not the primary mechanismof immunosuppression in their model (73).
FIGURE 7. FcR and B cell in-volvement in mediating the differencesseen between 7E1-G1 and 7E1-G2b.A, Wild-type C57BL/6 mice weretransplanted with minor Ag-mis-matched mOVA (C57BL/6 back-ground) skin grafts and treated with500 �g of CTLA4-Ig and with 7E1-G1, isotype IgG1 control, 7E1-G2b,isotype IgG2b control, or anti-CD154i.p on days 0, 2, 4, and 6. B, B cell-deficient mice were used as recipientsof mOVA skin grafts with the sametreatment regimen as noted in A andshowed graft survival similar to that ofwild-type recipients. C, Fc�RIII-defi-cient recipients did not show signifi-cant prolongation of mOVA skin graftsurvival when treated with CTLA4-Igand 7E1-G1. D, Fc�R�I-deficient re-cipients also did not show significantdifferences in the prolongation ofmOVA skin graft survival as com-pared with wild-type recipients. E,Fc�RIIb-deficient recipient mice,lacking the inhibitory FcR, exhibitedno diminution in the long-term graftsurvival observed in 7E1-G2b-treatedmice, whereas all other treatmentgroups promptly rejected their grafts.
1633The Journal of Immunology
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Due to the fact that the idiotypic regions are identical betweenthese two Abs, their differences in functionality likely lie withinthe Fc portion of the Ab. One potential mechanism that couldunderlie these differences in isotype is their differential ability tofix complement and mediate the depletion of CD40-bearing cells.However, we reject this hypothesis on the basis of two observa-tions: 1) no depletion of CD40-bearing cells was observed in vivo;and 2) the differential effects of the Abs on B cell proliferationwere observed in an in vitro, complement-free system. An alter-nate hypothesis for how isotype might impact functionality isthrough their binding to FcRs.
With regard to FcR binding, one might envision that there aretwo possibilities to explain the immunostimulatory propertiesobserved with 7E1-G1: either the mAb binds activating Fc�RIIIreceptors and induces positive FcR-mediated signaling into theAPC or the mAb binding to any FcR simply acts as a scaffold toinduce cross-linking of CD40 molecules on the surface of theAPC, resulting in the delivery of CD40-mediated signals. Al-though the role of FcRs in conferring either immunosuppressive orimmunostimulatory effects on a given anti-CD40 mAb remains tobe fully explored, in our system, proinflammatory signals deliv-ered by activating FcRs were not responsible for immunostimula-tory properties of 7E1-G1, as Fc�R�I-deficient mice exhibitedsimilar graft survival as compared with wild-type mice (Fig. 7D).Likewise, one also could imagine two analogous scenarios to ex-plain the immunosuppressive effects of 7E1-G2b: the mAb is func-tioning primarily to block CD40 engagement of CD154; or themAb is binding to inhibitory Fc�RIIb receptors to induce negativesignaling to B cells or other APCs. We favor the former, given thatinhibitory signals through Fc�RIIb were not found to be involvedin the immunosuppressive effects observed with 7E1-G2b, becausegraft survival was unchanged between Fc�RIIb-deficient and wild-type animals (Fig. 7E). Our results are consistent with the hypoth-esis that FcRs cross-link surface-bound 7E1-G1 mAb to a greaterextent than the 7E1-G2b mAb and thus contribute to the agonisticsignal delivered uniquely by 7E1-G1. Future experiments using invivo treatment with Fc-blocking Abs or delivery of F(ab�)2 frag-ments could further delineate the role of FcRs in the in vivo effectsof these Abs.
In conclusion, we have demonstrated that CD28 blockade caneffectively synergize with CD40 blockade using either anti-CD154or an appropriately designed anti-CD40 mAb. Although clinicaltranslation of anti-CD154 has encountered difficulties, this shouldnot belie the clear utility that can be gained by further studying theinhibition of the CD40 pathway in transplantation. Understandingthe cellular and subcellular mechanisms that underlie the ability ofblockade of the CD40/CD154 interaction to attenuate donor-reac-tive T cell responses and promote long-term graft survival is cru-cial to the development of novel therapeutics to target thispathway.
DisclosuresThe authors have no financial conflict of interest.
References1. Jenkins, M. K. 1994. The ups and downs of T cell costimulation. Immunity 1:
443–446.2. Wood, K. J., and S. Sakaguchi. 2003. Regulatory T cells in transplantation tol-
erance. Nat. Rev. Immunol. 3: 199–210.3. Jenkins, M. K., P. S. Taylor, S. D. Norton, and K. B. Urdahl. 1991. CD28 delivers
a costimulatory signal involved in antigen-specific IL-2 production by human Tcells. J. Immunol. 147: 2461–2466.
4. Schwartz, R. H. 1990. A cell culture model for T lymphocyte clonal anergy.Science 248: 1349–1356.
5. Yamada, A., A. D. Salama, M. Sho, N. Najafian, T. Ito, J. P. Forman,R. Kewalramani, S. Sandner, H. Harada, M. R. Clarkson, et al. 2005. CD70
signaling is critical for CD28-independent CD8� T cell-mediated alloimmuneresponses in vivo. J. Immunol. 174: 1357–1364.
6. Ozkaynak, E., W. Gao, N. Shemmeri, C. Wang, J. C. Gutierrez-Ramos, J. Ama-ral, S. Qin, J. B. Rottman, A. J. Coyle, and W. W. Hancock. 2001. Importance ofICOS-B7RP-1 costimulation in acute and chronic allograft rejection. Nat Immu-nol. 2: 591–596.
7. Demirci, G., F. Amanullah, R. Kewalaramani, H. Yagita, T. B. Strom,M. H. Sayegh, and X. C. Li. 2004. Critical role of OX40 in CD28 and CD154-independent rejection. J. Immunol. 172: 1691–1698.
8. Wang, J., Z. Guo, Y. Dong, O. Kim, J. Hart, A. Adams, C. P. Larsen,R. S. Mittler, and K. A. Newell. 2003. Role of 4-1BB in allograft rejectionmediated by CD8� T cells. Am. J. Transplant. 3: 543–551.
9. Larsen, C. P., E. T. Elwood, D. Z. Alexander, S. C. Ritchie, R. Hendrix,C. Tucker-Burden, H. R. Cho, A. Aruffo, D. Hollenbaugh, P. S. Linsley,K. J. Winn, and T. C. Pearson. 1996. Long-term acceptance of skin and cardiacallografts after blocking CD40 and CD28 pathways. Nature 381: 434–438.
10. Kirk, A. D., D. M. Harlan, N. N. Armstrong, T. A. Davis, Y. Dong, G. S. Gray,X. Hong, D. Thomas, J. H. Fechner, and S. J. Knechtle. 1997. CTLA4-Ig andanti-CD40L prevent renal allograft rejection in primates. Proc. Natl. Acad. Sci.USA 94: 8789–8794.
11. Levisetti, M. G., P. A. Padrid, G. L. Szot, N. Mittal, S. M. Meehan,C. L. Wardrip, G. S. Gray, D. S. Bruce, J. R. Thistlethwaite, Jr., andJ. A. Bluestone. 1997. Immunosuppressive effects of human CTLA4Ig in a non-human primate model of allogeneic pancreatic islet transplantation. J. Immunol.159: 5187–5191.
12. Larsen, C. P., T. C. Pearson, A. B. Adams, P. Tso, N. Shirasugi, E. Strobertm,D. Anderson, S. Cowan, K. Price, J. Naemura, et al. 2005. Rational developmentof LEA29Y (belatacept), a high-affinity variant of CTLA4-Ig with potent immu-nosuppressive properties. Am. J. Transplant. 5: 443–453.
13. Vincenti, F., C. Larsen, A. Durrbach, T. Wekerle, B. Nashan, G. Blancho,P. Lang, J. Grinyo, P. F. Halloran, K. Solez, et al. 2005. Costimulation blockadewith belatacept in renal transplantation. N. Engl. J. Med. 353: 770–781.
14. Kirk, A. D., L. C. Burkly, D. S. Batty, R. E. Baumgartner, J. D. Berning,K. Buchanan, J. H. Fechner, Jr., R. L. Germond, R. L. Kampen, N. B. Patterson,et al. 1999. Treatment with humanized monoclonal antibody against CD154 pre-vents acute renal allograft rejection in nonhuman primates. Nat. Med. 5:686–693.
15. Kenyon, N. S., L. A. Fernandez, R. Lehmann, M. Masetti, A. Ranuncoli,M. Chatzipetrou, G. Iaria, D. Han, J. L. Wagner, P. Ruiz, et al. 1999. Long-termsurvival and function of intrahepatic islet allografts in baboons treated with hu-manized anti-CD154. Diabetes 48: 1473–1481.
16. Kanmaz, T., J. J. Fechner, Jr., J. Torrealba, H. T. Kim, Y. Dong, T. D. Oberley,J. M. Schultz, D. D. Bloom, M. Katayama, et al. 2004. Monotherapy with thenovel human anti-CD154 monoclonal antibody ABI793 in rhesus monkey renaltransplantation model. Transplantation 77: 914–920.
17. Kawai, T., D. Andrews, R. B. Colvin, D. H. Sachs, and A. B. Cosimi. 2000.Thromboembolic complications after treatment with monoclonal antibody againstCD40 ligand. Nat. Med. 6: 114.
18. Weaver, T. A., A. H. Charafeddine, and A. D. Kirk. 2008. Costimulation block-ade: towards clinical application. Front. Biosci. 13: 2120–2139.
19. Inwald, D. P., A. McDowall, M. J. Peters, R. E. Callard, and N. J. Klein. 2003.CD40 is constitutively expressed on platelets and provides a novel mechanism forplatelet activation. Circ. Res. 92: 1041–1048.
20. Andre, P., K. S. Prasad, C. V. Denis, M. He, J. M. Papalia, R. O. Hynes,D. R. Phillips, and D. D. Wagner. 2002. CD40L stabilizes arterial thrombi by a�3 integrin–dependent mechanism. Nat. Med. 8: 247–252.
21. Kawabe, T., T. Naka, K. Yoshida, T. Tanaka, H. Fujiwara, S. Suematsu,N. Yoshida, T. Kishimoto, and H. Kikutani. 1994. The immune responses inCD40-deficient mice: impaired immunoglobulin class switching and germinalcenter formation. Immunity 1: 167–178.
22. Bennett, S. R. M., F. R. Carbone, F. Karamalis, R. A. Flavell, J. F. A. P. Miller,and W. R. Heath. 1998. Help for cytotoxic-T-cell responses is mediated by CD40signalling. Nature 393: 478–480.
23. Miga, A. J., S. R. Masters, B. G. Durell, M. Gonzalez, M. K. Jenkins,C. Maliszewski, H. Kikutani, W. F. Wade, and R. J. Noelle. 2001. Dendritic celllongevity and T cell persistence is controlled by CD154-CD40 interactions. Eur.J. Immunol. 31: 959–965.
24. Lazarevic, V., A. J. Myers, C. A. Scanga, and J. L. Flynn. 2003. CD40, but notCD40L, is required for the optimal priming of T cells and control of aerosol M.tuberculosis infection. Immunity 19: 823–835.
25. Hollenbaugh, D., N. Mischel-Petty, C. P. Edwards, J. C. Simon, R. W. Denfeld,P. A. Kiener, and A. Aruffo. 1995. Expression of functional CD40 by vascularendothelial cells. J. Exp. Med. 182: 33–40.
26. Phillips, N. E., T. G. Markees, J. P. Mordes, D. L. Greiner, and A. A. Rossini.2003. Blockade of CD40-mediated signaling is sufficient for inducing islet but notskin transplantation tolerance. J. Immunol. 170: 3015–3023.
27. Raisky, O., B. M. Spriewald, K. J. Morrison, S. Ensminger, T. Mohieddine,J. F. Obadia, M. H. Yacoub, and M. L. Rose. 2003. CD8� T cells induce graftvascular occlusion in a CD40 knockout donor/recipient combination. J. HeartLung Transplant. 22: 177–183.
28. Nathan, M. J., J. E. Mold, S. C. Wood, K. Csencsits, G. Lu, E. J. Eichwald, andD. K. Bishop. 2004. Requirement for donor and recipient CD40 expression incardiac allograft rejection: induction of Th1 responses and influence of donor-derived dendritic cells. J. Immunol. 172: 6626–6633.
29. Schoenberger, S. P., R. E. M. Toes, E. I. H. van der Voort, R. Offringa, andC. J. M. Melief. 1998. T-cell help for cytotoxic T lymphocytes is mediated byCD40-CD40L interactions. Nature 393: 480–483.
1634 EFFICACY OF ANTI-CD40 mAbs IN TRANSPLANTATION
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

30. Staveley-O’Carroll, K., T. D. Schell, M. Jimenez, L. M. Mylin, M. J. Tevethia,S. P. Schoenberger, and S. S. Tevethia. 2003. In vivo ligation of CD40 enhancespriming against the endogenous tumor antigen and promotes CD8� T cell ef-fector function in SV40 T antigen transgenic mice. J. Immunol. 171: 697–707.
31. Maxwell, J. R., J. D. Campbell, C. H. Kim, and A. T. Vella. 1999. CD40 acti-vation boosts T cell immunity in vivo by enhancing T cell clonal expansion anddelaying peripheral T cell deletion. J. Immunol. 162: 2024–2034.
32. Dullforce, P., D. C. Sutton, and A. W. Heath. 1998. Enhancement of T cell-independent immune responses in vivo by CD40 antibodies. Nat. Med. 4: 88–91.
33. Haanstra, K. G., J. Ringers, E. A. Sick, S. Ramdien-Murli, E. M. Kuhn, L. Boon,and M. Jonker. 2003. Prevention of kidney allograft rejection using anti-CD40and anti-CD86 in primates. Transplantation 75: 637–643.
34. Haanstra, K. G., E. A. Sick, J. Ringers, J. A. Wubben, E. M. Kuhn, L. Boon, andM. Jonker. 2005. Costimulation blockade followed by a 12-week period of cy-closporine A facilitates prolonged drug-free survival of rhesus monkey kidneyallografts. Transplantation 79: 1623–1626.
35. Adams, A. B., N. Shirasugi, T. R. Jones, M. M. Durham, E. A. Strobert,S. Cowan, P. Rees, R. Hendrix, K. Price, N. S. Kenyon, et al. 2005. Developmentof a chimeric anti-CD40 monoclonal antibody that synergizes with LEA29Y toprolong islet allograft survival. J. Immunol. 174: 542–550.
36. Masunaga, T., K. Yamashita, H. Sakihama, T. Hashimoto, N. Hua, A. Imai,M. Inobe, T. Miyazaki, S. Todo, and T. Uede. 2005. Dimeric but not monomericsoluble CD40 prolongs allograft survival and generates regulatory T cells thatinhibit CTL function. Transplantation 80: 1614–1622.
37. Aruffo, A. A., D. Hollenbaugh, A. W. Siadak, K. K. Berry, L. Harris,B. A. Thorne, and J. Bajorath. 1998. Methods of using antibodies against humanCD40. USP Office, ed. Bristol-Myers Squibb, Princeton, NJ.
38. Hale, G., M. Clark, and H. Waldmann. 1985. Therapeutic potential of rat mono-clonal antibodies: isotype specificity of antibody-dependent cell-mediated cyto-toxicity with human lymphocytes. J. Immunol. 134: 3056–3061.
39. Bruggemann, M., C. Teale, M. Clark, C. Bindon, and H. Waldmann. 1989. Amatched set of rat/mouse chimeric antibodies. J. Immunol. 142: 3145–3150.
40. Nimmerjahn, F., and J. V. Ravetch. 2005. Divergent immunoglobulin � subclassactivity through selective Fc receptor binding. Science 310: 1510–1512.
41. Hale, G., S. P. Cobbold, H. Waldmann, G. Easter, P. Matejtschuk, andR. R. Coombs. 1987. Isolation of low-frequency class-switch variants from rathybrid myelomas. J. Immunol. Methods 103: 59–67.
42. Rolink, A., F. Melchers, and J. Andersson. 1996. The SCID but not the RAG-2gene product is required for S�-S � heavy chain class switching. Immunity 5:319–330.
43. Adams, A. B., M. M. Durham, L. Kean, N. Shirasugi, J. Ha, M. A. Williams,P. A. Rees, M. C. Cheung, S. Mittelstaedt, A. W. Bingaman, et al. 2001. Co-stimulation blockade, busulfan, and bone marrow promote titratable macrochi-merism, induce transplantation tolerance, and correct genetic hemoglobinopathieswith minimal myelosuppression. J. Immunol. 167: 1103–1111.
44. Salomon, B., and J. A. Bluestone. 2001. Complexities of CD28/B7: CTLA-4costimulatory pathways in autoimmunity and transplantation. Annu. Rev. Immu-nol. 19: 225–252.
45. Wekerle, T., J. Kurtz, S. Bigenzahn, Y. Takeuchi, and M. Sykes. 2002. Mecha-nisms of transplant tolerance induction using costimulatory blockade. Curr. Opin.Immunol. 14: 592–600.
46. Trambley, J., A. W. Bingaman, A. Lin, E. T. Elwood, S. Y. Waitze, J. Ha,M. M. Durham, M. Corbascio, S. R. Cowan, T. C. Pearson, and C. P. Larsen.1999. Asialo GM1� CD8� T cells play a critical role in costimulation blockade-resistant allograft rejection. J. Clin. Invest. 104: 1715–1722.
47. Williams, M. A., J. Trambley, J. Ha, A. B. Adams, M. M. Durham, P. Rees,S. R. Cowan, T. C. Pearson, and C. P. Larsen. 2000. Genetic characterization ofstrain differences in the ability to mediate CD40/CD28-independent rejection ofskin allografts. J. Immunol. 165: 6849–6857.
48. Tomita, Y., A. Khan, and M. Sykes. 1994. Role of intrathymic clonal deletion andperipheral anergy in transplantation tolerance induced by bone marrow transplan-tation in mice conditioned with a nonmyeloablative regimen. J. Immunol. 153:1087–1098.
49. Grewal, I. S., and R. A. Flavell. 1998. CD40 and CD154 in cell-mediated im-munity. Annu. Rev. Immunol. 16: 111–135.
50. Ehst, B. D., E. Ingulli, and M. K. Jenkins. 2003. Development of a novel trans-genic mouse for the study of interactions between CD4 and CD8 T cells duringgraft rejection. Am. J. Transplant. 3: 1355–1362.
51. Ford, M. L., M. E. Wagener, S. S. Hanna, T. C. Pearson, A. D. Kirk, andC. P. Larsen. 2008. A critical precursor frequency of donor-reactive CD4� T cellhelp is required for CD8� T cell-mediated CD28/CD154-independent rejection.J. Immunol. 180: 7203–7211.
52. Ravetch, J. V., and S. Bolland. 2001. IgG Fc receptors. Annu. Rev. Immunol. 19:275–290.
53. Nimmerjahn, F., and J. V. Ravetch. 2006. Fc� receptors: old friends and newfamily members. Immunity 24: 19–28.
54. Robak, T. 2008. Novel monoclonal antibodies for the treatment of chronic lym-phocytic leukemia. Curr. Cancer Drug Targets 8: 156–171.
55. Fanale, M. A., and A. Younes. 2007. Monoclonal antibodies in the treatment ofnon-Hodgkin’s lymphoma. Drugs 67: 333–350.
56. Monk, N. J., R. E. Hargreaves, J. E. Marsh, C. A. Farrar, S. H. Sacks, M. Millrain,E. Simpson, J. Dyson, and S. Jurcevic. 2003. Fc-dependent depletion of activatedT cells occurs through CD40L-specific antibody rather than costimulation block-ade. Nat. Med. 9: 1275–1280.
57. Blair, P. J., J. L. Riley, D. M. Harlan, R. Abe, D. K. Tadaki, S. C. Hoffmann,L. White, T. Francomano, S. J. Perfetto, A. D. Kirk, and C. H. June. 2000. CD40ligand (CD154) triggers a short-term CD4� T cell activation response that resultsin secretion of immunomodulatory cytokines and apoptosis. J. Exp. Med. 191:651–660.
58. Hancock, W. W., M. H. Sayegh, X. G. Zheng, R. Peach, P. S. Linsley, andL. A. Turka. 1996. Costimulatory function and expression of CD40 ligand, CD80,and CD86 in vascularized murine cardiac allograft rejection. Proc. Natl. Acad.Sci. USA 93: 13967–13972.
59. Cella, M., D. Scheidegger, L. K. Palmer, P. Lane, A. Lanzavecchia, and G. Alber.1996. Ligation of CD40 on dendritic cells triggers production of high levels ofinterleukin-12 and enhances T cell stimulatory capacity: T-T help via APC ac-tivation. J. Exp. Med. 184: 747–752.
60. Koch, F., U. Stanzl, P. Jennewein, K. Janke, C. Heufler, E. Kampgen, N. Romani,and G. Schuler. 1996. High level IL-12 production by murine dendritic cells:upregulation via MHC class II and CD40 molecules and downregulation by IL-4and IL-10. J. Exp. Med. 184: 741–746.
61. Chen, M., L. Huang, Z. Shabier, and J. Wang. 2007. Regulation of the lifespanin dendritic cell subsets. Mol. Immunol. 44: 2558–2565.
62. Dallman, C., P. W. Johnson, and G. Packham. 2003. Differential regulation of cellsurvival by CD40. Apoptosis 8: 45–53.
63. Nopora, A., and T. Brocker. 2002. Bcl-2 controls dendritic cell longevity in vivo.J. Immunol. 169: 3006–3014.
64. Obst, R., H. M. van Santen, R. Melamed, A. O. Kamphorst, C. Benoist, andD. Mathis. 2007. Sustained antigen presentation can promote an immunogenic Tcell response, like dendritic cell activation. Proc. Natl. Acad. Sci. USA 104:15460–15465.
65. Obst, R., H. M. van Santen, D. Mathis, and C. Benoist. 2005. Antigen persistenceis required throughout the expansion phase of a CD4� T cell response. J. Exp.Med. 201: 1555–1565.
66. Kurtz, J., H. Ito, T. Wekerle, J. Shaffer, and M. Sykes. 2001. Mechanisms in-volved in the establishment of tolerance through costimulatory blockade andBMT: lack of requirement for CD40L-mediated signaling for tolerance or dele-tion of donor-reactive CD4� cells. Am. J. Transplant. 1: 339–349.
67. Quezada, S. A., K. Bennett, B. R. Blazar, A. Y. Rudensky, S. Sakaguchi, andR. J. Noelle. 2005. Analysis of the underlying cellular mechanisms of anti-CD154-induced graft tolerance: the interplay of clonal anergy and immune reg-ulation. J. Immunol. 175: 771–779.
68. Wells, A. D., X. C. Li, Y. Li, M. C. Walsh, X. X. Zheng, Z. Wu, G. Nunez,A. Tang, M. Sayegh, W. W. Hancock, T. B. Strom, and L. A. Turka. 1999.Requirement for T-cell apoptosis in the induction of peripheral transplantationtolerance. Nat. Med. 5: 1303–1307.
69. Graca, L., K. Honey, E. Adams, S. P. Cobbold, and H. Waldmann. 2000. Cuttingedge: anti-CD154 therapeutic antibodies induce infectious transplantation toler-ance. J. Immunol. 165: 4783–4786.
70. Ford, M. L., M. E. Wagener, S. Gangappa, T. C. Pearson, and C. P. Larsen. 2007.Antigenic disparity impacts outcome of agonism but not blockade of costimula-tory pathways in experimental transplant models. Am. J. Transplant. 7:1471–1481.
71. Yu, X. Z., M. H. Albert, P. J. Martin, and C. Anasetti. 2004. CD28 ligationinduces transplantation tolerance by IFN-�-dependent depletion of T cells thatrecognize alloantigens. J. Clin. Invest. 113: 1624–1630.
72. Suntharalingam, G., M. R. Perry, S. Ward, S. J. Brett, A. Castello-Cortes,M. D. Brunner, and N. Panoskaltsis. 2006. Cytokine storm in a phase 1 trial ofthe anti-CD28 monoclonal antibody TGN1412. N. Engl. J. Med. 355:1018–1028.
73. Pearson, T. C., J. Trambley, K. Odom, D. C. Anderson, S. Cowan, R. Bray,A. Lin, D. Hollenbaugh, A. Aruffo, A. W. Siadak, et al. 2002. Anti-CD40 therapyextends renal allograft survival in rhesus macaques. Transplantation 74:933–940.
1635The Journal of Immunology
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from