high production of squalene using a newly isolated yeast...
TRANSCRIPT

High Production of Squalene Using a Newly Isolated Yeast-like StrainPseudozyma sp. SD301Xiaojin Song,*,†,‡ Xiaolong Wang,∥ Yanzhen Tan,†,‡ Yingang Feng,†,‡ Wenli Li,∥ and Qiu Cui*,†,‡,§
†Shandong Provincial Key Laboratory of Energy Genetics, Qingdao Institute of Bioenergy and Bioprocess Technology, ‡QingdaoEngineering Laboratory of Single Cell Oil, Qingdao Institute of Bioenergy and Bioprocess Technology, and §Key Laboratory ofBiofuels, Qingdao Institute of Bioenergy and Bioprocess Technology, Chinese Academy of Sciences, 189 Songling Road, Qingdao,Shandong 266101, People’s Republic of China∥Key Laboratory of Marine Drugs, Ministry of Education of China, School of Medicine and Pharmacy, Ocean University of China, 5Yushan Road, Qingdao, Shandong 266003, People’s Republic of China
*S Supporting Information
ABSTRACT: A yeast-like fungus, termed strain SD301, with the ability to produce a high concentration of squalene, wasisolated from Shuidong Bay, China. The nucleotide sequence analysis of the internal transcribed spacer (ITS) region of SD301indicated the strain belonged to Pseudozyma species. The highest biomass and squalene production of SD301 were obtainedwhen glucose and yeast extracts were used as the carbon and nitrogen sources, respectively, with a C/N ratio of 3. The optimalpH and temperature were 6 and 25 °C, with 15 g L−1 of supplemented sea salt. The maximum squalene productivity reached0.039 g L−1 h−1 in batch fermentation, while the maximum squalene yield of 2.445 g L−1 was obtained in fed-batch fermentation.According to our knowledge, this is the highest squalene yield produced thus far using fermentation technology, and the newlyisolated strain Pseudozyma sp. SD301 is a promising candidate for commercial squalene production.
KEYWORDS: marine fungi, optimization, Pseudozyma, squalene
■ INTRODUCTION
Squalene (2,6,10,15,19,23-hexamethyltetracosa-2,6,10,14,18,22-hexane, C30H50) is a linear polyunsaturated aliphatic hydro-carbon, which is an intermediate for the biosynthesis of bileacids, ergosterol, and steroids.1 In addition, squalene has anatural antioxidant effect in a model of lipid peroxidation ofliposomes that protects cells from free radicals and reactiveoxygen species (ROS).2,3 Recent epidemiological studies haveindicated that squalene has effective antitumor activities, whichcan strengthen the immunity of humans and decrease the riskof various cancers, such as colon, lung, and skin tumori-genesis.4,5 In experiments with rats, a significant reduction inthe level of lipid peroxidation in the heart tissue is observed bya given combined administration of squalene and polyunsatu-rated fatty acids.6 Moreover, squalene also has a cardiopro-tective effect on experimentally induced myocardial infarc-tion.7,8 Currently, squalene is used as a moisturizing agent andan emollient in the cosmetic industry, and it has garnered moreand more attention for its potential applications inpharmaceuticals and medical sectors.3
The extensive use of squalene in industry is hampered bylimited resources and its relatively high price. Currently, thecommercial sources of squalene are the liver oil of deep seasharks and vegetable oils.9 Squalene is the main ingredient inthe liver oil of sharks, which could reach up to a percentage of60%.10 Some plant seed oils also have high squalene levels, suchas peanut oil (1.28 g kg−1),2 amaranth oil (2.15 g kg−1),11 andolive oil (5.99 g kg−1).12 However, the continuous supply andfuture availability of squalene are uncertain because of thedecreasing number of sharks, increasing environmentalpollutants as well as an unpleasant fishy smell,13,14 and the
instability of production in certain plant species.9 Recently,major efforts have been made to identify alternative sources ofsqualene in microalgae and microorganisms. The yield ofsqualene is approximately 0.43−340.5 mg L−1 dry cell weight(DCW) from yeasts15−17 and a lower productivity than 0.72 mgg−1 DCW from thraustochytrids.18,19 Currently, severalAurantiochytrium strains, such as strain 18W-13a20−22 andstrain PQ6,23 can produce squalene at a yield of approximately1 g L−1. However, the squalene productivity is still very low forcommercial use.In this study, we screened various environmental samples for
squalene-producing microorganisms with high yield andisolated several squalene producers. Among them, we identifieda novel yeast-like strain that possessed the highest squaleneproductivity thus far. This isolate, termed Pseudozyma sp.SD301, proved its strong potential in commercial squaleneproduction.
■ MATERIALS AND METHODSIsolation and Screening of High-Squalene-Producing
Strains. Pseudozyma strains were obtained from seawater, soil, andleaf samples of the mangrove ecosystem in Shuidong Bay, Guangdong,China. The samples were incubated on KMV+-medium-containingagar plates (glucose, 1.0 g L−1; gelatin hydrolysates, 1.0 g L−1; yeastextract, 0.1 g L−1; peptone, 0.1 g L−1; and agar, 12 g L−1; in 1 L ofnatural seawater containing penicillin G, 50 ppm; and streptomycin
Received: July 11, 2015Revised: September 6, 2015Accepted: September 9, 2015Published: September 9, 2015
Article
pubs.acs.org/JAFC
© 2015 American Chemical Society 8445 DOI: 10.1021/acs.jafc.5b03539J. Agric. Food Chem. 2015, 63, 8445−8451

sulfate, 50 ppm) at 25 °C in the dark. After 90 h of incubation, themonocolonies on the surface of the plates were transferred ontoKMV+ medium several times to purify the strains. These isolatedstrains were kept on a KMV+ medium (without antibiotics) asmonoclonal cultures, and then we determined their squalene contentusing gas chromatography.Molecular and Phylogenetic Analyses. The genomic DNA of
selected strains was extracted using a Universal Genomic DNAextraction kit (Takara DV811A, Dalian, China), according to theprotocol of the manufacturer. The internal transcribed spacer (ITS)region was amplified using the ITS4 primer P1 (5 ′ -TCCTCCGCTTATTGATATGC-3′) and the ITS5 primer P2 (5′-GGAAGTAAAAGTCGTAACAAGG-3′). The polymerase chain re-action (PCR) products were visualized by agarose gel electrophoresisand purified using the TIANgel Midi Purification Kit (TIANGENDP209, Beijing, China). The purified PCR products were cloned intothe pGEM-T Easy vector (Promega) and then were sequenced byTakara Co., Ltd.The resulting sequence data were searched using NCBI BLAST,
and the sequences were aligned with the ClustalX program. Aphylogenetic tree was inferred using the Minimum Evolution methodof MEGA, version 5.0. The tree reliability was evaluated by bootstrapanalysis of 1000 replicates.Strains and Cultivation. Strain SD301 was isolated by the
methods mentioned above and cultivated in 50 mL of GPY medium(6% glucose, 1% peptone, and 1% yeast extract) containing 1.5%artificial sea salt in a 250 mL flask shaken at 200 rpm and 25 °C.Optimization of Culture Conditions. To investigate the effects
of various cultivation media, eight carbon sources (6% of glucose, D-fructose, D-xylose, maltose, sucrose, lactose, glycerol, and starch) andseven nitrogen sources (2% of yeast extract, peptone, tryptone, urea,ammonium sulfate, sodium glutamate, and sodium nitrate) weretested. Tryptone (2%) was used as the nitrogen source for the carbonsource testing, while glucose (6%) was consistently used as the carbonsource when the nitrogen source testing was carried out. To investigatethe C/N ratio effect on the cell growth and squalene yield of strainSD301, the constant concentration of the C source of 6% and variousN source concentrations were used.The effects of varying concentrations (0−60 g L−1) of sea salt were
also studied using the optimized medium. To examine the effects ofvarious growth parameters, various pH values (pH 4.0, 5.0, 6.0, 7.0,8.0, and 9.0) and different temperatures (15, 20, 25, 30, and 35 °C)were used to cultivate Pseudozyma sp. SD301.All tests were carried out in 250 mL flasks with 50 mL medium
shaken at 200 rpm and 25 °C for 5 days.Batch and Fed-Batch Fermentation Experiments for
Squalene Production. A batch fermentation experiment wasperformed in a 5 L Biostat B plus bioreactor, which was equippedwith controllers for pH, temperature, agitation, and dissolved oxygen(DO) concentration. Batch cultures were carried out in 3.5 L ofproduction medium. The temperature was maintained at 25 °C. Theagitation speed automatically varied from 300 to 800 rpm at a fixed airflow rate of 1.2 vessel volumes per minute (vvm) to maintain the DOat 20% air saturation. The pH was maintained at 6.0 by adding 2 molL−1 NaOH or 2 mol L−1 HCl. To control foam formation, 1 mL ofantifoam was added at the beginning of the run. Samples for off-linedetermination of the glucose concentration, biomass, and squaleneyield were withdrawn every 6 h until the end of the fermentation (48h).Another fed-batch fermentation experiment was also performed in
the 5 L Biostat B plus bioreactor, with the same conditions of thebatch fermentation, except that intermittent glucose feeding wassupplied to maintain the residual glucose concentration atapproximately 5 to 25 g L−1 by feeding a 40% (w/v) glucose stocksolution. The samples were withdrawn every 10 h until the end of thefermentation (100 h).The initial fermentation medium contained 60 g L−1 glucose and 20
g L−1 yeast extract in artificial seawater (artificial sea salt concentrationof 15 g L−1). This medium also contained a vitamin solution (2 mLL−1). The vitamin solution was filter-sterilized (0.22 μm) and
contained 100 mg/L thiamine, 10 mg/L biotin, and 50 mg/Lcyanocobalamin.
Biomass Determination and Glucose Assay. The biomass ofPseudozyma sp. SD301 was expressed as DCW. Samples (5 mL) werecollected and centrifuged at 7000g at 4 °C for 10 min, and they werethen freeze-dried to a constant weight at −50 °C for approximately 60h. The residual glucose in the fermentation medium was analyzed by abiosensor equipped with a glucose oxidase electrode (SBA-40E,Institute of Biology, Shandong Academy of Sciences, Jinan, China).
Observation Using Confocal Laser Scanning Microscopy(CLSM). To investigate the intracellular localization of squalene inPseudozyma sp. SD301, the observation of in vivo fluorescence imagingof the cells were carried out using CLSM (FluoView FV1000,Olympus, Tokyo, Japan). First, the cells were dyed by nile red usingthe method mentioned by Morita et al.24 In details, 500 μg mL−1 nilered was prepared in acetone and stored at 4 °C as the stock solution.Then, the stock solution was added to the culture with a finalconcentration of 0.1 μg mL−1 and incubated at 37 °C for 15 min.Second, the stained cells were observed by CLSM, and the excitationand emission wavelengths were 559 and 612 nm, respectively. Thefluorescent images were obtained at 1000× magnification.
Lipid Extraction and Fatty Acid and Squalene ContentAnalyses. The total lipid content was estimated using a modifiedminiaturized Bligh−Dyer method.25 The harvested cells were extractedinto 30 mL of chloroform/methanol (2:1, v/v) at room temperature.The lipid extract was dried over anhydrous Na2SO4, and the solventwas removed by evaporation.
The total lipid was converted into fatty acid methyl esters (FAMEs),and the fatty acid composition was analyzed using an Agilent 7890-5975 gas chromatography−mass spectrometry (GC−MS) system(Agilent Technologies, Inc., Santa Clara, CA).26
Squalene was isolated from the total lipid by saponification,27 withsome modifications. Samples (5 mL, V1) were freeze-dried and used toextract the total lipid by the methods mentioned above. The extractedtotal lipid was transferred into 10 mL of 10% KOH−ethanol solutionand incubated at 60 °C for 1 h with refluxing for saponification. Then,4 mL of distilled water was added, and the unsaponifiablecompositions were extracted with 2 mL (V2) of n-hexane. Afterward,squalene was separated by Agilent 7890A GC with a 7650A automaticliquid sampler using a HP-5, 30 m × 320 μm × 0.25 μm column;meanwhile, nitrogen was used as a carrier gas. The initial oventemperature was 100 °C and was maintained for 1 min, and then thetemperature was raised to 300 °C for 15 °C min−1 and, finally,maintained at 300 °C for another minute. Peak detection wasperformed by a flame ionization detector. The temperature of theinjection port and flame ionization port was 300 °C. The injectionvolume was 1 μL, and the split ratio was 1:10. The squaleneconcentration (X, μg mL−1) was further quantified using an externalstandard method. The standard curve (Figure S1) was made from a setof concentrations of commercial squalene standard (>99% squalene,J&K Chemical) by GC analysis. The squalene production (P, g L−1)was calculated by eq 1.
= × −P XVV
102
1
3
(1)
■ RESULTS AND DISCUSSIONIsolation and Identification of Pseudozyma sp. SD301.
Samples obtained from a mangrove ecosystem were screenedfor squalene-producing microorganisms. Almost 100 purifiedstrains were isolated as colonies on the KMV+ medium platesand microscopically observed. The biomass (DCW) of thestrains ranged from 5.29 to 27.61 g L−1 when cultivated in theGPY medium, and the squalene contents (percent of DCW)and squalene yields ranged from 0.4 to 5.6% DCW and from0.1 to 0.9 g L−1, respectively.Among these tested strains, SD301 showed the highest levels
of squalene content and yield. On the basis of morphological
Journal of Agricultural and Food Chemistry Article
DOI: 10.1021/acs.jafc.5b03539J. Agric. Food Chem. 2015, 63, 8445−8451
8446
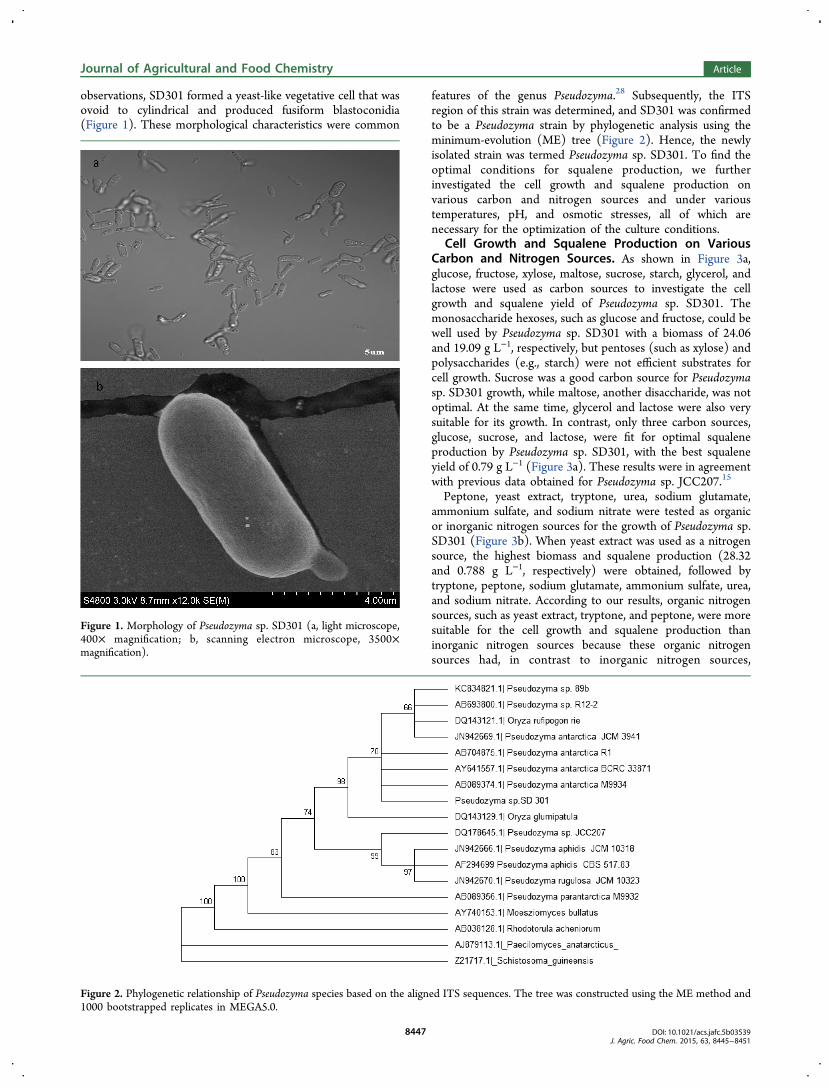
observations, SD301 formed a yeast-like vegetative cell that wasovoid to cylindrical and produced fusiform blastoconidia(Figure 1). These morphological characteristics were common
features of the genus Pseudozyma.28 Subsequently, the ITSregion of this strain was determined, and SD301 was confirmedto be a Pseudozyma strain by phylogenetic analysis using theminimum-evolution (ME) tree (Figure 2). Hence, the newlyisolated strain was termed Pseudozyma sp. SD301. To find theoptimal conditions for squalene production, we furtherinvestigated the cell growth and squalene production onvarious carbon and nitrogen sources and under varioustemperatures, pH, and osmotic stresses, all of which arenecessary for the optimization of the culture conditions.
Cell Growth and Squalene Production on VariousCarbon and Nitrogen Sources. As shown in Figure 3a,glucose, fructose, xylose, maltose, sucrose, starch, glycerol, andlactose were used as carbon sources to investigate the cellgrowth and squalene yield of Pseudozyma sp. SD301. Themonosaccharide hexoses, such as glucose and fructose, could bewell used by Pseudozyma sp. SD301 with a biomass of 24.06and 19.09 g L−1, respectively, but pentoses (such as xylose) andpolysaccharides (e.g., starch) were not efficient substrates forcell growth. Sucrose was a good carbon source for Pseudozymasp. SD301 growth, while maltose, another disaccharide, was notoptimal. At the same time, glycerol and lactose were also verysuitable for its growth. In contrast, only three carbon sources,glucose, sucrose, and lactose, were fit for optimal squaleneproduction by Pseudozyma sp. SD301, with the best squaleneyield of 0.79 g L−1 (Figure 3a). These results were in agreementwith previous data obtained for Pseudozyma sp. JCC207.15
Peptone, yeast extract, tryptone, urea, sodium glutamate,ammonium sulfate, and sodium nitrate were tested as organicor inorganic nitrogen sources for the growth of Pseudozyma sp.SD301 (Figure 3b). When yeast extract was used as a nitrogensource, the highest biomass and squalene production (28.32and 0.788 g L−1, respectively) were obtained, followed bytryptone, peptone, sodium glutamate, ammonium sulfate, urea,and sodium nitrate. According to our results, organic nitrogensources, such as yeast extract, tryptone, and peptone, were moresuitable for the cell growth and squalene production thaninorganic nitrogen sources because these organic nitrogensources had, in contrast to inorganic nitrogen sources,
Figure 1. Morphology of Pseudozyma sp. SD301 (a, light microscope,400× magnification; b, scanning electron microscope, 3500×magnification).
Figure 2. Phylogenetic relationship of Pseudozyma species based on the aligned ITS sequences. The tree was constructed using the ME method and1000 bootstrapped replicates in MEGA5.0.
Journal of Agricultural and Food Chemistry Article
DOI: 10.1021/acs.jafc.5b03539J. Agric. Food Chem. 2015, 63, 8445−8451
8447

additional growth factors, such as vitamins and coenzymes,which might be necessary for squalene biosynthesis. Theseresults were slightly different from the results of Pseudozyma sp.JCC207,15 which could also produce squalene with inorganicnitrogen sources.It is well-known that the C/N ratio of the culture medium
dramatically affects the growth of microorganisms. A high C/Nratio is effective for increasing the lipid content in cells.29
Therefore, the effects of the C/N ratio on the cell growth andsqualene yield of strain SD301 were examined (Figure 3c). Thecell growth increased to the maximum level by increasing theC/N ratio to 3. The highest squalene yield of 0.756 g L−1 wasalso obtained at this C/N ratio. A further increase of the C/Nradio showed a negative effect on both the cell growth andsqualene yield. When the C/N ratio reached 7.5, the biomassand squalene production decreased to 19.53 and 0.632 g L−1,respectively. This phenomenon could be explained by the fact
that the high C/N ratio might lead to relative nitrogen hunger(or other nutrient hunger, e.g., phosphorus and sulfur), whichlimits cell growth and increases the biosynthesis of other lipids(such as fatty acids and triglyceride).
Effects of the Sea Salt Concentration and CultureTemperature on Cell Growth. The effects of the sea saltconcentration on cell growth and squalene production wereexamined in a range of 0−6%. Pseudozyma sp. SD301 showed awide salinity tolerance. As shown in Figure 4a, the optimum salt
concentration was 15 g L−1, with a biomass and squalene yieldof 26.25 and 0.847 g L−1, respectively. A slight change in thecell growth was observed in the range of 0.75−3%. Manymicroorganisms living in the mangrove ecosystem showeuryhalinity30,31 because the salinity of this area often sharplychanges as a result of the afflux of fresh water and evaporation.The effects of the temperature on biomass and squalene yield
were tested from 15 to 35 °C. As shown in Figure 4b, therewere only slight fluctuations in biomass and squalene yieldbetween the temperatures of 25 and 30 °C. The biomassproduction was the highest (26.23 g L−1) at 25 °C. However,when the temperature was raised to 35 °C, the biomass
Figure 3. Effects of (a) carbon sources, (b) nitrogen sources, and (c)C/N ratio on the cell growth and squalene yield in Pseudozyma sp.SD301.
Figure 4. Effect of (a) artificial sea salt concentration, (b) temperature,and (c) initial pH on cell growth and squalene yield in Pseudozyma sp.SD301.
Journal of Agricultural and Food Chemistry Article
DOI: 10.1021/acs.jafc.5b03539J. Agric. Food Chem. 2015, 63, 8445−8451
8448

decreased to 21.37 g L−1 and the squalene yield also decreasedsignificantly, from 0.811 g L−1 at 25 °C to 0.555 g L−1 at 35 °C.The variation in squalene yields may be the result of thebiosynthesis of ergosterol and steroids, which are vital structuraland regulatory components in eukaryotic cells.Effects of Initial pH on Cell Growth. Because the pH
value of the medium profoundly affects cell membranefunctions, cell metabolism, and the uptake of nutrients andproduct biosynthesis, the initial pH values of the medium testedin this study ranged from 4.0 to 9.0. The results showed thatstrain SD301 had a wide adaptability of initial pH (4.0−9.0) forgrowth and squalene accumulation (Figure 4c). The variationof biomass and squalene yields in the tested pH ranges rangedfrom 21.70 g L−1 (pH 9.0) to 26.90 g L−1 (pH 6.0) and from0.625 g L−1 (pH 4.0) to 0.820 g L−1 (pH 6.0), respectively. Theoptimal pH for biomass and squalene yield was 6.0. The widepH adaptability of Pseudozyma sp. SD301 may due to itsadaptation to the mangrove ecosystem, in which the pH isconstantly changing with tidal actions.Microscopic Observation and the Fatty Acid Compo-
sition of Pseudozyma sp. SD301. From the fluorescenceimaging of the cells (Figure 5), the lipid droplets containingsqualene were distinctly observed at the both ends of the cellsand were accumulated along with the cell growth. Thisphenomenon was coincident with that in Saccharomycescerevisiae.16,17 The fatty acid composition of SD301 wasshown in Table 1, and the main ingredients were the fatty
acids with 16 or 18 carbons, such as C16:0, C18:0, C18:1, andC18:2. Along with the cell growth, the content of saturatedfatty acids in cells decreased and the content of unsaturatedfatty acids reached up to over 51%.
Batch and Fed-Batch Fermentation Experiments forSqualene Production. Using the optimized culture con-ditions described above, the biomass, squalene production, andglucose consumption were examined during batch and fed-batch cultivation.Glucose was exhausted after 42 h of batch fermentation,
reaching the highest biomass and squalene production of 30.24and 1.649 g L−1, respectively (Figure 6a). The conversion ratefrom glucose to biomass was approximately 50.4%, and thesqualene productivity reached 0.039 g L−1 h−1. In contrast, forthe fed-batch fermentation, a total glucose concentration of 143g L−1 was consumed and the maximum DCW was 71.53 g L−1
with a squalene yield of 2.445 g L−1 at 80 h (Figure 6b). Incomparison to the batch fermentation, both the glucoseconversion rate and the squalene productivity were slightlydecreased. This result might be caused by the increasing C/Nratio during glucose feeding, which could influence cell growthand increase fatty acid and triglyceride biosynthesis.Some strains of Aurantiochytrium (Schizochytrium or
Thraustochytrium) were used to produce squalene with thefinal yield of approximately 1 g during 96 h of fermentation. Incomparison to these strains, Pseudozyma sp. SD301 hadsuperior squalene productivity and higher squalene yield(Table 2). The squalene yield of 2.445 g L−1 by Pseudozymasp. SD301 was 2 times greater than the reported highest yieldof Aurantiochytrium sp. 18W-13a (1.29 g L−1), and the squaleneproductivity was approximately 3 times higher than that ofAurantiochytrium strains.In summary, this study characterized a newly isolated yeast-
like strain, Pseudozyma sp. SD301, which is a promisingcandidate for commercial squalene production. The resultsillustrated that the optimized medium for squalene productionshould contain 60 g L−1 glucose, 20 g L−1 yeast extract, and 15g L−1 artificial sea salt concentration, with the appropriatecultivation conditions of pH 6.0 and 25 °C.
Figure 5. Observation of Pseudozyma sp. SD301 using CLSM for locating the lipid droplets and squalene accumulation during cultivation.
Table 1. Fatty Acid Profiles of Pseudozyma sp. SD301
content (% TFA)
fatty acid 24 h 72 h 120 h
C14:0 2.03 1.57 1.34C16:0 30.06 26.46 24.89C16:1 2.32 2.09 1.45C17:0 2.44 2.77 1.51C18:0 24.12 20.99 15.97C18:1 23.21 29.22 35.94C18:2 13.87 14.58 16.74C20:0 1.83 2.28 2.14
Journal of Agricultural and Food Chemistry Article
DOI: 10.1021/acs.jafc.5b03539J. Agric. Food Chem. 2015, 63, 8445−8451
8449

■ ASSOCIATED CONTENT*S Supporting InformationThe Supporting Information is available free of charge on theACS Publications website at DOI: 10.1021/acs.jafc.5b03539.
Standard curve of squalene concentrations (Figure S1)(PDF)
■ AUTHOR INFORMATIONCorresponding Authors*Telephone: +86-532-80662705. Fax: +86-532-80662707. E-mail: [email protected].*Telephone: +86-532-80662706. Fax: +86-532-80662707. E-mail: [email protected].
FundingThis work was financially supported by the National NaturalScience Foundation of China (41306132) and the NationalHigh Technology Research and Development Program ofChina (863 Program, 2014AA021701).
NotesThe authors declare no competing financial interest.
■ REFERENCES(1) Smith, T. J. Squalene: potential chemopreventive agent. ExpertOpin. Invest. Drugs 2000, 9, 1841−1848.(2) Amarowicz, R. Squalene: A natural antioxidant? Eur. J. Lipid Sci.Technol. 2009, 111, 411−412.(3) Spanova, M.; Daum, G. Squalene − biochemistry, molecularbiology, process biotechnology, and applications. Eur. Eur. J. Lipid Sci.Technol. 2011, 113, 1299−1320.(4) Rao, C. V.; Newmark, H. L.; Reddy, B. S. Chemopreventive effectof squalene on colon cancer. Carcinogenesis 1998, 19, 287−290.(5) Smith, T. J.; Yang, G. Y.; Seril, D. N.; Liao, J.; Kim, S. Inhibitionof 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone-induced lung tu-morigenesis by dietary olive oil and squalene. Carcinogenesis 1998, 19,703−706.(6) Storm, H. M.; Oh, S. Y.; Kimler, B. F.; Norton, S.Radioprotection of mice by dietary squalene. Lipids 1993, 28, 555−559.(7) Aguilera, Y.; Dorado, M. E.; Prada, F. A.; Martinez, J. J.; Quesada,A.; Ruiz-Gutierrez, V. The protective role of squalene in alcoholdamage in the chick embryo retina. Exp. Eye Res. 2005, 80, 535−543.(8) Chan, P.; Tomlinson, B.; Lee, C. B.; Lee, Y. S. Effectiveness andsafety of low-dose pravastatin and squalene, alone and in combination,in elderly patients with hypercholesterolemia. J. Clin. Pharmacol. 1996,36, 422−427.(9) Newmark, H. L. Squalene, olive oil, and cancer risk - review andhypothesis. Ann. N. Y. Acad. Sci. 1999, 889, 193−203.
Figure 6. Fermentation experiments of Pseudozyma sp. SD301 in a 5 L bioreactor: (a) batch fermentation experiment and (b) fed-batchfermentation experiment: (open squares) residual glucose, (closed diamonds) biomass, and (closed triangles) squalene yield.
Table 2. Comparison of the Squalene Yield by the StrainsMentioned in This Study
strainsqualene yield
(g L−1)
squaleneproductivity(g L−1 h−1) reference
Aurantiochytrium sp.18W-13a (2011)
1.29 0.014 20
Aurantiochytrium sp.18W-13a (2012)
0.9 21
Thraustochytrium(2013)
≈1 0.012 22
Schizochytriummangrovei PQ6(2014)
0.992−1.019 0.011 23
Pseudozyma sp. JCC207(2008)
0.341 0.004 15
Pseudozyma sp. SD301 2.445 0.039 thisstudy
Journal of Agricultural and Food Chemistry Article
DOI: 10.1021/acs.jafc.5b03539J. Agric. Food Chem. 2015, 63, 8445−8451
8450

(10) Pietsch, A.; Jaeger, P. Concentration of squalene from shark liveroil by short-path distillation. Eur. J. Lipid Sci. Technol. 2007, 109,1077−1082.(11) He, H. P.; Corke, H. Oil and squalene in amaranthus grain andleaf. J. Agric. Food Chem. 2003, 51, 7913−7920.(12) García-Gonzalez, D. L.; Aparicio-Ruiz, R.; Aparicio, R. Virginolive oil - Chemical implications on quality and health. Eur. J. Lipid Sci.Technol. 2008, 110, 602−607.(13) Storelli, M. M.; Ceci, E.; Storelli, A.; Marcotrigiano, G. O.Polychlorinated biphenyl, heavy metal and methylmercury residues inhammerhead sharks: contaminant status and assessment. Mar. Pollut.Bull. 2003, 46, 1035−1039.(14) Turoczy, N. J.; Laurenson, L. J.; Allinson, G.; Nishikawa, M.;Lambert, D. F.; Smith, C.; Cottier, J. P.; Irvine, S. B.; Stagnitti, F.Observations on metal concentrations in three species of shark(Deania calcea, Centroscymnus crepidater, and Centroscymnus owstoni)from southeastern Australian waters. J. Agric. Food Chem. 2000, 48,4357−4364.(15) Chang, M. H.; Kim, H. J.; Jahng, K. Y.; Hong, S. C. Theisolation and characterization of Pseudozyma sp. JCC 207, a novelproducer of squalene. Appl. Microbiol. Biotechnol. 2008, 78, 963−972.(16) Garaiova, M.; Zambojova, V.; Simova, Z.; Griac, P.; Hapala, I.Squalene epoxidase as a target for manipulation of squalene levels inthe yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2014, 14, 310−323.(17) Mantzouridou, F.; Naziri, E.; Tsimidou, M. Z. Squalene versusergosterol formation using Saccharomyces cerevisiae: combined effect ofoxygen supply, inoculum size, and fermentation time on yield andselectivity of the bioprocess. J. Agric. Food Chem. 2009, 57, 6189−6198.(18) Fan, K. W.; Aki, T.; Chen, F.; Jiang, Y. Enhanced production ofsqualene in the thraustochytrid Aurantiochytrium mangrovei by mediumoptimization and treatment with terbinafine. World J. Microbiol.Biotechnol. 2010, 26, 1303−1309.(19) Chen, G.; Fan, K. W.; Lu, F. P.; Li, Q.; Aki, T.; Chen, F.; Jiang,Y. Optimization of nitrogen source for enhanced production ofsqualene from thraustochytrid Aurantiochytrium sp. New Biotechnol.2010, 27, 382−389.(20) Kaya, K.; Nakazawa, A.; Matsuura, H.; Honda, D.; Inouye, I.;Watanabe, M. M. Thraustochytrid Aurantiochytrium sp. 18W-13aAccummulates High Amounts of Squalene. Biosci., Biotechnol., Biochem.2011, 75, 2246−2248.(21) Nakazawa, A.; Matsuura, H.; Kose, R.; Kato, S.; Honda, D.;Inouye, I.; Kaya, K.; Watanabe, M. M. Optimization of cultureconditions of the thraustochytrid Aurantiochytrium sp. strain 18W-13afor squalene production. Bioresour. Technol. 2012, 109, 287−291.(22) Nakazawa, A.; Kokubun, Y.; Matsuura, H.; Yonezawa, N.; Kose,R.; Yoshida, M.; Tanabe, Y.; Kusuda, E.; Van Thang, D.; Ueda, M.;Honda, D.; Mahakhant, A.; Kaya, K.; Watanabe, M. M. TLC screeningof thraustochytrid strains for squalene production. J. Appl. Phycol.2014, 26, 29−41.(23) Hoang, M. H.; Ha, N. C.; Thom, L. T.; Tam, L. T.; Anh, H. T.;Thu, N. T.; Hong, D. D. Extraction of squalene as value-added productfrom the residual biomass of Schizochytrium mangrovei PQ6 duringbiodiesel producing process. J. Biosci. Bioeng. 2014, 118, 632−639.(24) Morita, T.; Konishi, M.; Fukuoka, T.; Imura, T.; Kitamoto, D.Physiological differences in the formation of the glycolipidbiosurfactants, mannosylerythritol lipids, between Pseudozyma antarc-tica and Pseudozyma aphidis. Appl. Microbiol. Biotechnol. 2007, 74,307−315.(25) Bligh, E. G.; Dyer, W. J. A rapid method of total lipid extractionand purification. Can. J. Biochem. Physiol. 1959, 37, 911−917.(26) Xiao, Y.; Zhang, J.; Cui, J.; Feng, Y.; Cui, Q. Metabolic profilesof Nannochloropsis oceanica IMET1 under nitrogen-deficiency stress.Bioresour. Technol. 2013, 130, 731−738.(27) Dhara, R.; Bhattacharyya, D. K.; Ghosh, M. Analysis of steroland other components present in unsaponifiable matters of mahua, saland mango kernel oil. J. Oleo Sci. 2010, 59, 169−176.
(28) Boekhout, T. PseudozymaBandoni emend. Boekhout, a genusfor yeast-like anamorphs of Ustilaginales. J. Gen. Appl. Microbiol. 1995,41, 359−366.(29) Converti, A.; Casazza, A. A.; Ortiz, E. Y.; Perego, P.; Del Borghi,M. Effect of temperature and nitrogen concentration on the growthand lipid content of Nannochloropsis oculata and Chlorella vulgaris forbiodiesel production. Chem. Eng. Process. 2009, 48, 1146−1151.(30) Gao, M.; Song, X. J.; Feng, Y. G.; Li, W. L.; Cui, Q. Isolation andcharacterization of Aurantiochytrium species: high docosahexaenoicacid (DHA) production by the newly isolated microalga, Aurantiochy-trium sp SD116. J. Oleo Sci. 2013, 62, 143−151.(31) Yokochi, T.; Honda, D.; Higashihara, T.; Nakahara, T.Optimization of docosahexaenoic acid production by Schizochytriumlimacinum SR21. Appl. Microbiol. Biotechnol. 1998, 49, 72−76.
Journal of Agricultural and Food Chemistry Article
DOI: 10.1021/acs.jafc.5b03539J. Agric. Food Chem. 2015, 63, 8445−8451
8451