genetics of sex-linked yellow in the syrian hamster
TRANSCRIPT

Copyright � 2009 by the Genetics Society of AmericaDOI: 10.1534/genetics.108.095018
Genetics of Sex-linked yellow in the Syrian Hamster
Azita Alizadeh,* Lewis Z. Hong,* Christopher B. Kaelin,* Terje Raudsepp,†
Hermogenes Manuel* and Gregory S. Barsh*,1
*Departments of Genetics and Pediatrics, Stanford University, Stanford, California 94305 and †Department of Veterinary IntegrativeBiosciences, Texas A&M University, College Station, Texas 77843
Manuscript received August 28, 2008Accepted for publication February 1, 2009
ABSTRACT
Alternating patches of black and yellow pigment are a ubiquitous feature of mammalian color variationthat contributes to camouflage, species recognition, and morphologic diversity. X-linked determinants ofthis pattern—recognized by variegation in females but not in males—have been described in the domesticcat as Orange, and in the Syrian hamster as Sex-linked yellow (Sly), but are curiously absent from othervertebrate species. Using a comparative genomic approach, we develop molecular markers and a linkagemap for the euchromatic region of the Syrian hamster X chromosome that places Sly in a regionhomologous to the centromere-proximal region of human Xp. Comparison to analogous work carried outfor Orange in domestic cats indicates, surprisingly, that the cat and hamster mutations lie in nonhomolo-gous regions of the X chromosome. We also identify the molecular cause of recessively inherited black coatcolor in hamsters (historically referred to as nonagouti) as a Cys115Tyr mutation in the Agouti gene. Animalsdoubly mutant for Sly and nonagouti exhibit a Sly phenotype. Our results indicate that Sly represents amelanocortin pathway component that acts similarly to, but is genetically distinct from, Mc1r and that hasimplications for understanding both the evolutionary history and the mutational mechanisms of pigment-type switching.
THE genetics of coat color is a longstanding and richmodel system for studying gene action and cell
signaling. Like classical genetic systems in invertebratemodel organisms, the cell types and tissues that give riseto mammalian hair color are well characterized andexperimentally accessible, and alterations in gene act-ivity can be easily detected. Coat color mutations areespecially useful for studying genes and pathwaysunique to vertebrate genomes and have led to a deeperunderstanding of diverse biological processes becausemuch of the molecular machinery used by the pigmen-tary system is either shared by or homologous to mole-cules used in other physiological pathways ( Jackson
1997; Bennett and Lamoreux 2003; Steingrimsson
et al. 2006).This approach has proven particularly useful for
dissecting the molecular mechanisms of pigment-typeswitching, a phenomenon in which melanocytes choosebetween synthesizing eumelanin (a relatively insolubleblack or brown pigment) or pheomelanin (a cysteine-rich red or yellow pigment that is soluble in dilute alkali)(Silvers 1979). A focal point for pigment-type switch-ing is the Agouti–melanocortin 1 receptor (Mc1r) pathway.Mc1r is a G-protein coupled receptor expressed inmelanocytes, whereas Agouti protein is a paracrine sig-
naling molecule secreted by specialized dermal cells thatinhibits Mc1r signaling (Cone et al. 1996; reviewed inBarsh 2006). In laboratory mice, gain-of-function muta-tions that constitutively activate the Mc1r (e.g., somber,Mc1rso) cause exclusive production of eumelanin, whereasloss-of-function Mc1r mutations (e.g., recessive yellow, Mc1re)cause exclusive production of pheomelanin (Robbins
et al. 1993). On the other hand, because Agouti is a Mc1rantagonist, gain-of-function Agouti mutations (e.g., lethalyellow, Ay) cause exclusive production of pheomelanin,whereas loss-of-function mutations (e.g., extreme nonagouti,ae) cause exclusive production of eumelanin (Siracusa
1994; Hustad et al. 1995).In the early 1900s, Sewall Wright (Wright 1917)
concluded that genetic mechanisms that control pigmen-tary variation have been largely conserved during mamma-lian evolution. In particular, the Agouti phenotype—in which individual hairs display a subapical band ofpheomelanin on an otherwise eumelanic background—is observed across a wide range of mammalian phyla. Inthe past decade, gain- and loss-of-function Mc1r mutationshavebeenidentifiedinmanydomesticanimals(Andersson
2003; Klungland and Vage 2003), and Agouti and/orMc1r variation has been shown to contribute to pigmen-tary variation in several natural populations (Eizirik
et al. 2003; Nachman et al. 2003; Mundy et al. 2004;Hoekstra et al. 2006; Steiner et al. 2007). Mostexperimental work on the Agouti–Mc1r pathway hasbeen carried out in laboratory mice. Nonetheless,
1Corresponding author: Beckman Center B271A, Stanford UniversitySchool of Medicine, Stanford, CA 94305. E-mail: [email protected]
Genetics 181: 1427–1436 (April 2009)

comparative zoologic studies by Wright (1918) andothers (Little 1957; Searle 1968) suggested that somecomponents of mammalian pigment-type switching arenot represented as coat color mutations in laboratorymice. In addition to a regular black and yellow stripepattern found in many carnivores that is usually attributedto the Tabby gene (Searle 1968), irregular black andorange patches due to the activity of an X-linked gene arealso present in the domestic cat and the Syrian hamster;neither of these phenomena have been described inlaboratory mice.
In the domestic cat, female-specific variegation ofblack and orange coat color patches has been appreci-ated for more than a century and helped support theinitial hypothesis that random and epigenetically heri-table X inactivation is a universal feature of placentalmammals (Lyon 1962). A similar phenotype thoughtto be caused by mutation of a homologous X-linkedgene was described in Syrian hamsters .40 years ago(Robinson 1966). As in cats, hemizygous male hamstersand homozygous female hamsters are completely yel-low, while heterozygous females exhibit a characteristictortoiseshell pattern of yellow patches on a black back-ground. (The terms tortoiseshell and calico are often usedto refer to the phenotype, the gene, and/or the allele inboth cats and hamsters; here we use Orange, O, to describethe gene in cats and Robinson’s original designation, Sex-linked yellow, Sly, to describe the gene in hamsters.)
More than 100 mouse coat color mutations have beencharacterized over the past century, and multiple allelesexist for those directly implicated in pigment-type switch-ing (Agouti, Mc1r, Atrn, Mgrn, Sox18), none of which lie onthe X chromosome (Bennett and Lamoreux 2003). Theapparent absence from laboratory mice of an X-linkedpigment-type switching mutation is perplexing in theface of Ohno’s law, which predicts conservation ofsynteny on the X chromosome across different mamma-lian species due to selection against altered gene dosage(Ohno 1969). Genetic characterization of Sex-linked yellowin Syrian hamsters could shed light on this paradox andidentify additional components of the pigment-typeswitching pathway. However, by contrast to laboratorymice, very little of the classical work on coat color varia-tion in Syrian hamsters (Nixon et al. 1970) has beendeveloped at the molecular genetic level.
We have established a laboratory-based colony ofSyrian hamsters to study the biology and genetics ofSex-linked yellow together with other coat color variantsand confirmed that the mutation behaves similarly towhat was originally reported (Robinson 1966, 1972).Here we use a comparative genomic approach togenerate a molecular genetic map of the Syrian hamsterX chromosome that includes Sex-linked yellow, whichprovides a basis for molecular genetic and epistasisstudies revealing that Sly represents a component ofthe melanocortin pathway similar to, but independentof, the Mc1r. Comparison to analogous work carried out
for Orange in domestic cats (Schmidt-Kuntzel et al.2009, accompanying article, this issue) indicates that thecat and hamster mutations likely lie in nonhomologousregions of the X chromosome, which has implicationsfor understanding both evolutionary history and themutational mechanisms of pigment-type switching.
MATERIALS AND METHODS
Animal husbandry: Sly and other coat color variants in theSyrian hamster are well recognized in the hobbyist community,but are not maintained in an academic- or a research-orientedsetting; none of the laboratory animal vendors we contactedwere aware of a sex-linked and/or variegated phenotype. Weobtained animals with a diverse set of coat color phenotypesfrom San Joaquin Valley Fisheries (Fresno, CA), includingbelted, black, cream, golden, red-eyed dilute, tortoiseshell,and tricolor (analogous to calico in the domestic cat). Forphenotypic characterization and linkage mapping of Sly, weused tortoiseshell females and black males and confirmedthat tortoiseshell segregated as an X-linked trait in accordwith Mendelian expectations as indicated in Figures 1 and2 and Table 2. All animal work was carried out underan Asia Pacific Laboratory Accreditation Cooperation-ap-proved protocol.
Genetic markers: We first developed simple sequencelength polymorphism (SSLP) markers using a comparativegenomic strategy in which aligned regions of the mouse andhuman X chromosomes were used to identify potential SSLPtargets for PCR amplification using Primer 3 (Rozen andSkaletsky 2000). Precomputed alignments of the human andmouse genomes were obtained from ftp://hgdownload.cse.ucsc.edu/goldenPath/hg16/vsMm4/axtTight/. From 39,889regions of human–mouse X chromosome alignments, we useda custom perl script to identify 7204 pairs of alignments inwhich each pair was separated by a 100- to 300-bp gap (in whichno alignment was recorded and therefore may have containedan SSLP). We filtered the 7204 pairs of alignments by theextent of similarity, allowing only those alignments in whichthere was a perfect human–mouse match of at least 20 bp,yielding 943 alignment pairs. Among these, suitable primers(that met default criteria for Primer3) could be designed for452. Among the 452 alignment pairs, the intervening (to beamplified) sequence contained one or more SSLP targets inboth the mouse and the rat genomic sequence in �30 align-ments, all of which were used for PCR of hamster genomicDNA. This approach yielded seven amplicons that containedpolymorphic SSLPs; we then designed new internal primersdirectly from the hamster sequence for subsequent amplifica-tion of hamster genomic DNA.
We supplemented the SSLP framework map with SNP-basedmarkers ascertained by resequencing introns or between pairsof conserved noncoding sequences. For resequencing introns,exon-based primer pairs were chosen using a perl scriptdesigned to target oligonucleotides corresponding to littleor no codon degeneracy (and therefore likely to be conservedbetween mouse and hamster). For resequencing betweenpairs of conserved noncoding sequences, we modified thestrategy described above to start with precomputed mouse–human alignments (Kent et al. 2003; Brudno et al. 2004).From �300 amplicons from hamster DNA, we identified nineSNPs used to supplement the SSLP-based linkage map. Forseven of the nine SNP markers, primers based on mousesequence yielded robust and specific amplification productsfrom hamster genomic DNA; in two cases (DBarX111 and
1428 A. Alizadeh et al.
DBarX118) we developed new internal primers directly fromthe hamster sequence.
Identity of the amplicons for all 16 primer pairs was in-vestigated by aligning the amplified hamster sequence to thepredicted mouse amplicon. In four cases (DXBar53, DXBar
54, DXBar DXBar 57, and DXBar 119), no significantsimilarity was observed between mouse and hamster ampli-cons, probably due to internal repetitive sequence and/or lackof evolutionary constraint; therefore the identity of theseamplicons relies on comparative mapping results (Figure 3).
Molecular genetic and linkage analysis: SSLP-based markerswere amplified with dye-labeled primers; SNP-based markerswere sequenced with dye terminators after amplification; allmarkers were separated on a capillary instrument equippedfor fluorescence detection. Genotypes for SSLP markers werecalled manually; genotypes for SNPs were called with anautomated commercial detection platform (CodonCode).Prior to linkage analysis, Mendelian error checking was per-formed; apparent instances of non-Mendelian transmissionwere inspected for potential genotype errors and either cor-rected or dropped from the analysis. An initial map order wasestablished by minimizing the number of double crossoversand subsequently refined with a regression-based analysis forintermarker distance implemented in JoinMap (Stam 1993).
Molecular cytogenetics: Fluorescence in situ hybridization(FISH) of bacterial artificial chromosome (BAC) probes tometaphase chromosomes from cultured mouse and ham-ster fibroblast cells was carried out as described previously(Raudsepp and Chowdhary 2008). We selected three mouseBACs (RP24-77C5, RP24-186P1, and RP24-96H13) on the basisof their gene content as inferred from the July 2007 (Build 37)assembly on the UCSC genome browser, http://genome.cse.ucsc.edu/ (Karolchik et al. 2003). Each BAC contains oneor more genes that are found in segments whose order isphysically conserved among mice, humans, dogs, and cats; thegenes (and BACs) were chosen to lie close to, and possiblyflanking, the cat Orange gene as depicted in Figure 5. BACswere labeled by nick translation with digoxigenin (77C5 and196H13)- or biotin (186P1)-coupled UTP, hybridized to nu-clear chromatin in pairwise combinations (77C5 1 186P1 or196H13 1 186P1), and detected with indirect immunofluo-rescence (Raudsepp and Chowdhary 2008). For cross-speciesdetection (mouse probes hybridized to hamster chromosomes),probe concentration was increased by �5- to 10-fold, andhybridization was carried out for 72 rather than 24 hr.
RESULTS
Sex-linked yellow in hamsters—phenotype and genet-ics: Commercial suppliers of Syrian hamsters to re-search laboratories carry few, if any, coat color variants.We identified a major pet store supplier in the Californiacentral valley, imported several different coat colorvariants, and established standardized pedigrees fromseven tortoiseshell females and six black males to con-firm and characterize the phenotype and inheritance
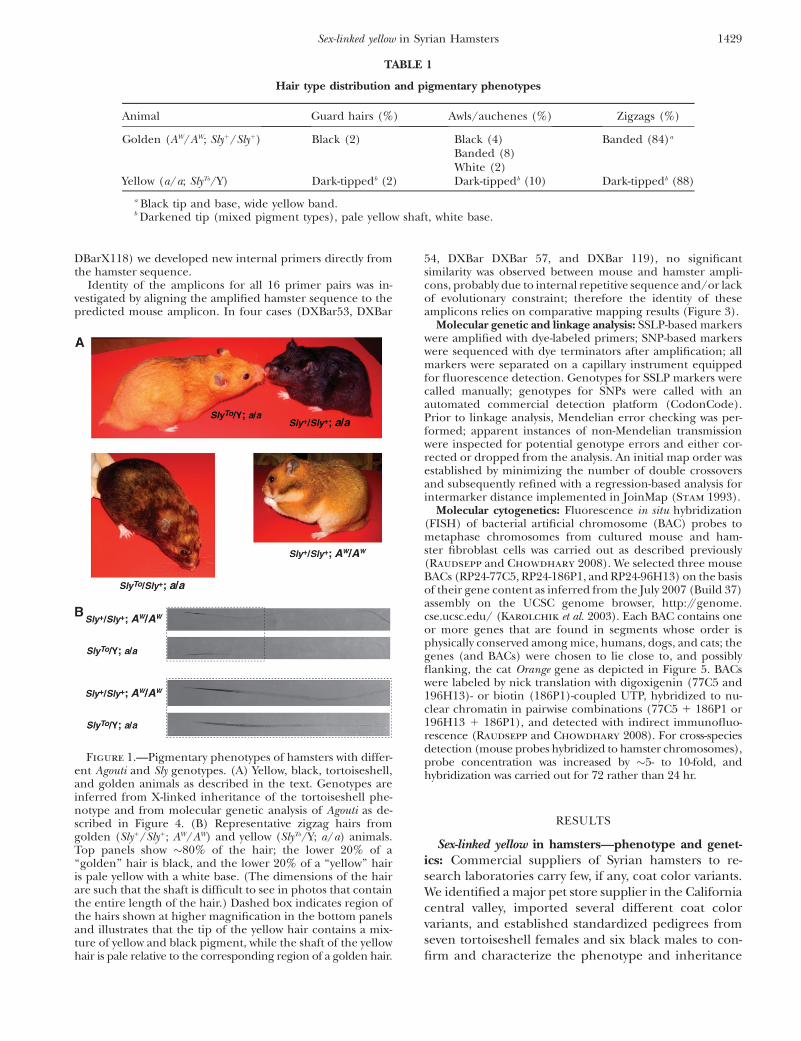
Figure 1.—Pigmentary phenotypes of hamsters with differ-ent Agouti and Sly genotypes. (A) Yellow, black, tortoiseshell,and golden animals as described in the text. Genotypes areinferred from X-linked inheritance of the tortoiseshell phe-notype and from molecular genetic analysis of Agouti as de-scribed in Figure 4. (B) Representative zigzag hairs fromgolden (Sly1/Sly1; AW/AW) and yellow (SlyTo/Y; a/a) animals.Top panels show �80% of the hair; the lower 20% of a‘‘golden’’ hair is black, and the lower 20% of a ‘‘yellow’’ hairis pale yellow with a white base. (The dimensions of the hairare such that the shaft is difficult to see in photos that containthe entire length of the hair.) Dashed box indicates region ofthe hairs shown at higher magnification in the bottom panelsand illustrates that the tip of the yellow hair contains a mix-ture of yellow and black pigment, while the shaft of the yellowhair is pale relative to the corresponding region of a golden hair.
TABLE 1
Hair type distribution and pigmentary phenotypes
Animal Guard hairs (%) Awls/auchenes (%) Zigzags (%)
Golden (AW/AW; Sly1/Sly1) Black (2) Black (4) Banded (84)a
Banded (8)White (2)
Yellow (a/a; SlyTo/Y) Dark-tippedb (2) Dark-tippedb (10) Dark-tippedb (88)
a Black tip and base, wide yellow band.b Darkened tip (mixed pigment types), pale yellow shaft, white base.
Sex-linked yellow in Syrian Hamsters 1429

pattern. In what follows, we refer to mutant and non-mutant alleles for Sly as SlyTo and Sly1, respectively.
As described originally (Robinson 1966), mutantanimals (SlyTo/Y or SlyTo/SlyTo) are orange–yellow in colorwith a darker dorsum than ventrum and a sooty ap-pearance, whereas nonmutant animals (Sly1/Sly1 orSly1/Y) are ‘‘golden,’’ presumably due to the presenceof an Agouti allele that promotes an extended band ofpheomelanin. However, in the hobbyist communitytoday, SlyTo is almost always described on a presumptivenonagouti (a/a) background such that nonmutantanimals (Sly1/Sly1 or Sly1/Y) are black.
We examined dorsal hair pigment distribution fromyellow (SlyTo/Y; a/a), black (Sly1/Y; a/a), and golden(Sly1/Y; AW/AW) animals. As in the mouse (Sundberg
1994), the hamster coat contains three major types ofhair: long guard hairs and shorter hairs, awls and zigzags,that constitute the underfur and whose relative propor-tions are similar among yellow, black, and goldenhamsters (Table 1). Pigment-type switching phenotypesare most evident in the zigzag hairs, which compose themajority of the underfur. Hairs from golden animals ex-hibit a dark tip and base separated by a wide pheome-lanic band that extends over �80% of the hair length
Figure 2.—Transmission and linkage of Sly. (A) Structure of pedigrees used for segregation and linkage analysis. Shaded,hatched, and yellow symbols represent black, tortoiseshell, and yellow animals, respectively, with associated Sly genotype inferredon the basis of coat color phenotype and sex. As described in the text, 17 such pedigrees yielded 155 F3 progeny, who were thengenotyped for the molecular markers as indicated. (B and C) Haplotype segregation diagram, with gray or yellow indicatingwhether the chromosome of origin carried Sly1 or SlyTo, respectively, and haplotypes organized into single (B) and multiple(C) recombinants. Gene order is based on multiple-regression analysis of intermarker distances; minimizing the number of dou-ble crossovers yields an alternative order for the first three markers, DXBar112–DXBar52–DXBar51. Recombination frequency(RF) in centimorgans between each marker and Sly is given on the right together with the number of informative meioses.
1430 A. Alizadeh et al.

(thereby accounting for a golden rather than a typicalbrushed Agouti appearance). Hairs from yellow animalsexhibit a dark tip (but less dark than in golden animals),a pale yellow shaft (less intense than the pheomelanicband of golden hairs), and a white base (Figure 1).
To characterize the inheritance of Sly, we used a three-generation breeding scheme in which phase could beinferred unambiguously for X-linked markers; tortoise-shell females were always mated to black males, suchthat only a single Sly allele was segregating among F3
progeny (Figure 2A). A total of 155 F3 progeny wereobtained from 17 pedigrees. X-linked inheritance of Slywas confirmed by the absence of yellow females in eachpedigree and a distribution of F3 progeny that did notdeviate significantly from that expected for X linkage(Table 2). These observations confirmed that eachkindred is segregating a single X-linked allele that resultsin constitutive pheomelanin production and provided asufficient number of meiotic events to carry out a linkagescan of the hamster X chromosome.
Linkage scan of the X chromosome: We used acombination of comparative genomic and PCR-basedapproaches to develop molecular genetic markers forthe hamster X chromosome. Among all possible combi-nations of markers (seven SSLP markers and nine SNPmarkers) with 155 F3 individuals, 1077 assays yieldedinformative results and were used to establish a geneticmap (Figures 2B and 3, Table 3). The hamster Xchromosome map covers 46 cM with a median inter-marker distance of 4.1 cM; by comparison, the mouse X-specific region is 72 cM in length (Blake et al. 2003).The hamster map includes markers that map to bothends of the human X chromosome, but lacks markerscorresponding to two key regions in mouse (0–20 Mband 85–129 Mb) and one key region in human (54–98 Mb). Thus, we cannot determine whether the shortergenetic length of the hamster X chromosome relative tothe mouse represents a reduced ratio of genetic/physical distance in the hamster relative to the mouseor a terminal region on the hamster X that has not beencaptured by our genetic markers.
Comparing the relative position of homologousmarkers between the mouse, hamster, human, and
dog X chromosomes reveals a considerable degree ofintrachromosomal rearrangement between hamstersand mice and between hamsters and humans or dogs(Figure 3A). However, the relative position of markersin the hamster is almost identical to that in the rat(Figure 3B), consistent with previous molecular cytoge-netic work (Kuroiwa et al. 2001a,b,c), suggesting thatgene order on the X chromosome is largely conservedbetween several rodent species (including the rat andthe Syrian hamster), although not in the laboratorymouse.
The Syrian hamster X chromosome is metacentric,but with the long arm composed entirely of constitutiveheterochromatin (DiPaolo and Popescu 1973); thus,our molecular map applies to hamster Xp. Comparisonof our results to molecular cytogenetic studies suggestsan orientation in which DXBar111 and DXBar51 liemost centromere proximal and centromere distal, re-spectively. Sly lies in a 9.6-cM region at the centromere-proximal end, flanked by DXBar111 and DXBar56(Figure 3). Transposed against the human and dog Xchromosomes, this region defines 8- and 5-Mb intervals,respectively, that also lie close to the human and dogcentromeres.
Candidate genes for Sly—Agouti and Mc1r : As in-dicated above, although SlyTo was originally described onan Agouti (golden) background, most animals carryingSlyTo are presumably nonagouti, such that Sly1/Y andSly1/Sly1 animals display a black coat, whereas Sly1/SlyTo
animals display patches of yellow hair on a black back-ground, presumably in regions where melanocyte cloneshave undergone epigenetic inactivation of the wild-type Xchromosome (Figure 1A). However, coat color genetics inhamsters are based almost entirely on the similarity ofphenotypes and genetic interactions to those observed inlaboratory mice, and there are at least two hamstergenes that can yield recessive inheritance of a black coat(Kuramoto et al. 2002).
To investigate the underlying molecular basis forblack coat color (and, indirectly, the epistasis relation-ship between black coat color and Sly), we used a PCR-based strategy to determine the coding sequence forthe hamster Agouti gene. Within exon 4, which encodesthe majority of the Agouti protein-coding region, wedetected a G to A mutation that predicts a Cys115Tyrsubstitution (Figure 4A) found in both black hamstersand yellow hamsters in our colony, but not in goldenhamsters. This residue is conserved among all knownAgouti homologs and helps stabilize the key active loopresponsible for melanocortin receptor antagonism(McNulty et al. 2005). Furthermore, a Cys115Ser muta-tion in a mouse Agouti transgene has the same effect as anull allele (Perry et al. 1996). Taken together, theseresults indicate that loss-of-function in Agouti is re-sponsible for the black coat color in our colony.
Similar to what has been described for Orange intortoiseshell and calico cats (Searle 1968), the patches
TABLE 2
Phenotype of F3 progeny from black 3
tortoiseshell intercross
Phenotype Sex No. progeny
Black Male 39Yellow Male 33Tortoiseshell Male 0Black Female 46Yellow Female 0Tortoiseshell Female 37
Based on cross as depicted in Figure 2A and described in thetext.
Sex-linked yellow in Syrian Hamsters 1431

of black and yellow pigment in Sly1/SlyTo hamstersbecome larger in the presence of white spottingmutations (Robinson 1972), suggesting that SlyTo actsin a melanocyte-autonomous manner, similar to Mc1re inlaboratory mice. To investigate the possibility that SlyTo
might represent an Mc1r loss-of-function allele that hadbeen transposed to the hamster X chromosome, weused oligonucleotide primers based on the mouse andrat genome to PCR amplify the hamster Mc1r gene andflanking sequences from yellow and black hamsters. Weidentified several synonymous and/or flanking se-quence SNPs that were heterozygous in at least onemale animal (data not shown), but did not observe any
alterations expected to impair Mc1r function; thus, asin all other mammals, including cats (Eizirik et al.2003), the hamster Mc1r appears to be autosomal. Weconclude that, similar to Orange in cats, Sly is epistatic toAgouti but genetically distinct from Mc1r (Figure 4B).
Relationship to Orange in domestic cats: The puz-zling similarities between cat Orange and hamsterSly—puzzling given the evolutionary distance betweenphyla and the absence of a similar mutation in any othermammal—prompted us to investigate whether the twomutations lie in homologous locations. In an accompa-nying article, Schmidt-Kuntzel et al. (2009) demon-strate that Orange lies in a 6-cM, �11-Mb interval that
Figure 3.—ComparativeX chromosome maps formouse, hamster, human,dog, and rat. (A) Physicalposition (rounded to thenearest megabase) is givenfor mouse, human, anddog markers, and the loca-tion of the pseudoautoso-mal regions (PAR or PAR1)is indicated for mouse andhuman. As described inthe text, the hamster maprefers to Xp (the long armis heterochromatic) withthe position of the centro-mere based on comparisonto molecular cytogeneticresults (Kuroiwa et al.2001a,b,c). A homologyblock on human (46–54Mb) and dog (40–45 Mb)Xp corresponds to the posi-tion of Sly in the hamster.(B) Position of markers onthe hamster X chromosomerelative to the rat.
1432 A. Alizadeh et al.

corresponds to 96–106 Mb and 120–131 Mb in the dogand human genomes, respectively. By contrast, flankingmarkers for hamster Sly delineate a different interval, onthe short arm of the dog (40–45 Mb) and human (46–54 Mb) X chromosomes (Figure 5C).
A potential caveat to the conclusion that the hamsterSly and cat Orange mutations lie in nonhomologouslocations is that the large number of evolutionary break-points apparent from comparing the mouse and ham-ster X chromosomes to those of other mammals mightmask one or more chromosomal rearrangements in
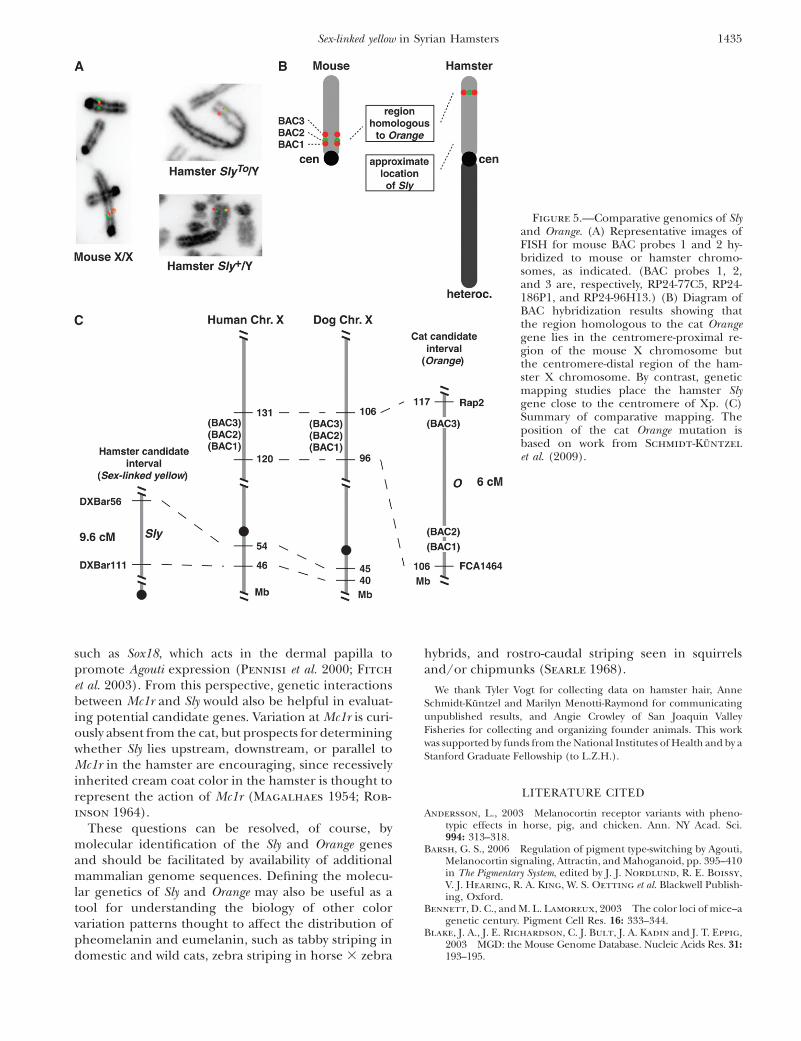
which segments harboring the Orange gene were trans-ferred to what we currently recognize as the Sly region.To investigate this possibility, we carried out cytogeneticFISH experiments to evaluate the location of the catOrange interval in mouse and hamster chromosomes.We selected three BAC clones from a mouse genomicDNA library that each carried genes found in homolo-gous locations in humans, dogs, and cats and whosepositions were likely to flank the Orange mutation(Figure 5C). Each of the three BACs was hybridized tometaphase spreads from an X/X mouse, an Sly1/Y
TABLE 3
Genetic markers
Marker name Type Primersa Coordinatesb
DBarX111 2 (Rbm10) GTCCTGAAAGGCCCGTGT 20212996–20214673GAACCAAAGGCCAAAACTGA
DBarX 51 1 GGAGGCTTAGCTTACCCCTTT 40781605–40782207ACGTGAAATTGTGTGCGTGT
DBarX 52 1 CACATGAGCTCTGTTGCATCT 53759564–53760159AAAGTCACCCTCATGGGAAG
DBarX 112 2 (Bgn) GACCACAACAAAATCCAGGCa 70738334–70738684CTCTCACCTGGAGGAGCTTGa
DBarX 113 2 (Tbl1x) GGACGCTCACACAGGAGAAGa 74895226–74895824GTCCTTGGAAGGTTTTGACTGa
DBarX 114 2 (CNS) CCAGCTTTGGTCATTTGACAGa 84947982–84948629AGGGACTTGGAAAGCTCACAa
DBarX 53 1 AAGGCAATGGCTTCATTGTT 129415830–12941667TGCTTCTGACCTTTGAGCAG
DBarX 54 1 CCACCTCTGTGAGCACATTC 131291877–131292275TGTCTAAATGTGTTTCTCACACTCC
DBarX 55 1 ATGCACCCCTCCAGTTCTGC 136812895–136813493TGGATCTGCAGTAATGTGCAGT
DBarX 56 1 GAGCGTCCCGGGAGCTCCTT 147389767–147390410CCACCTCTTACAGGAAGCC
DBarX 115c 2 (CNS) ATTGCCTGCCAAATCACACTa 156649400–156649723TGATTAAGTGACCCCAGAAATCa
DBarX 57 1 TCCATTGTTATTCAAGGAAGGAA 158286467–158286797GCTTTTGTGCAGATTCCCAG
DBarX 119c 2 (CNS) AGGTGATGTCAGCGGCTCTa 158595286–158595666TCTTTCCAGAACTCATCCCCa
DBarX 116 2 (CNS) GTCGATTAGCATTGGCATTTa 162665068–162665558AAAGCATAGCACACACAAGGAa
DBarX 118c 2 (Prps2) GCTCATTGGTCAGCCAATCT 163801537–163803099CCATGTGCTTTGTGATTCCA
DBarX 117 2 (CNS) GAACAGCAAGGAGGAAATTGa 164867514–164867994CCTGAAGCCTCCCACTCCa
Marker names correspond to those in Figures 2 and 3; types ‘‘1’’ and ‘‘2’’ refer to SSLP and SNP markers,respectively, the latter followed either by the gene name or conserved noncoding sequence (CNS) dependingon how the primers were derived. Oligonucleotide primers were originally designed from mouse genome se-quence, but unless otherwise stated (see footnote a), new primers were generated from the amplified hamstersequence and used for subsequent genotyping.
a All of the type 1 (SSLP) primers represent hamster sequence. However, for seven of the nine type 2 (SNP)primers (those indicated with a superscript a), oligonucleotides based on the mouse sequence yielded robustproducts when amplifying hamster genomic DNA, and we did not redesign new hamster primers.
b Physical position of amplicons on the mouse X chromosome based on coordinates from the July 2007 (build37) release.
c Not included in Figures 2 and 3 because they added no new map information. Genotypes from DXBar115and DXBar119 were identical to those obtained with DXBar57; genotypes from DXBar118 were identical tothose obtained with DXBar117.
Sex-linked yellow in Syrian Hamsters 1433

hamster, and an SlyTo/Y hamster. As expected, thecytogenetic locations for the three BACs on the mouseX chromosome correspond to their positions on thephysical map, at �38 Mb (BAC1: 77C5), 39 Mb (BAC2:186P1), and 47 Mb (BAC3: 196H13) (Figure 5A).Hybridization of the same BAC probes to hamsterchromosomes yielded weaker signals, and it was notpossible to order the probes in double-labeling experi-ments. However, all three probes hybridized to the sameregion at the telomeric end of hamster Xp for both theSly1- and the SlyTo-bearing chromosomes (Figure 5, Aand B). These results suggest that the X-chromosomalregion that contains Orange (as defined by geneticmapping experiments carried out by Schmidt-Kuntzel
et al. 2009) remained intact during rodent evolution andis distinct from the region where Sly maps in hamsters.
DISCUSSION
In most respects, the effects of Sly in hamsters andOrange in domestic cats are similar to those caused byMc1r loss-of-function mutations in other mammals.Both Sly and Orange yield a uniformly pheomelanic pel-age whose effects are epistatic to those of nonagouti, asis the case for Mc1r loss-of-function in mice (Silvers
1979), horses (Marklund et al. 1996; Rieder et al.2001), and dogs (Newton et al. 2000; Kerns et al. 2004).Furthermore, both Sly and Orange interact with white-spotting mutations in a manner that suggests they aremelanocyte autonomous, as is the case for mosaic Mc1rmutations in pigs (Kijas et al. 2001) and mice (Lamoreux
and Mayer 1975) and presumptive Mc1r mutations in
guinea pigs and rabbits (Searle 1968). However, bothSly and Orange are genetically distinct from the Mc1r andpossibly from each other. In what follows, we consider twoexplanations that could account for these observations.
Sly and Orange may be orthologs, representing loss-of-function mutations in the same (previously unrecog-nized) component of melanocortin signaling that actsin a manner and cell type similar to that of the Mc1r, butlies genetically upstream, such as a transcription factorrequired for Mc1r expression or a receptor-associatedprotein required for proper cell surface expressionand/or targeting of the Mc1r. Absence of a similar X-linked phenotype in mammals other than hamsters andcats could be explained by redundancy, e.g., if the Sly/Orange gene was duplicated early in mammalian evolu-tion, and the duplicated genes were redundant withregard to pigmentary function in most mammals. Acco-rding to this scenario, most mammals would then havetwo Sly/Orange genes, but one of the duplicated geneswould have been lost, by chance, from the hamster and,independently, from the cat lineage.
If Sly and Orange are orthologs, the apparent lack of‘‘homology’’ in their respective chromosomal locations(homology as assessed with the comparative mappingapproach in Figure 5) likely reflects additional chromo-somal rearrangements during rodent evolution, in whichthe Orange gene in an ancestral mammal translocated tothe centromere-proximal region of hamster Xp. Al-though the FISH-based cytogenetics do not support sucha rearrangement, the sequences tested (three BACs in an�10-Mb region) represent only a fraction of the intervaland cannot exclude the possibility of small chromosomalrearrangements that will become apparent only fromadditional genome sequences.
Alternatively, Sly and Orange could represent unusualgain-of-function mutations in which a (perhaps pre-viously known) component of melanocortin signalinghas been activated in melanocytes. For example, retro-transposition of Agouti into a melanocyte-specific locuson the X chromosome or a point mutation of amelanocyte-specific X-linked Gi-coupled receptor thatcauses constitutive activation would be expected to yielda phenotype and set of genetic interactions similar tothose observed for Sly and Orange. Both sorts of eventswould be very rare—much less frequent than loss-of-function mutations—and therefore might explain whyX-linked pheomelanism has not been observed inspecies where very large numbers of animals have beensubjected to a natural screen, i.e., laboratory mice andhumans. This idea is also consistent with preliminaryanalyses of the cat and dog X chromosome sequences,in which the dog region homologous to that whichcarries Orange also harbors several pseudogenes whoseorigin is autosomal.
A consideration of SlyTo candidate genes must alsoaccount for the epistatic relationship between Sly andAgouti; for example, Sly is unlikely to represent a gene
Figure 4.—Genetics and epistasis of Agouti and Sly. (A) Asdescribed in the text, black hamsters carry a missense mutationthat predicts a Cys115Tyr substitution, predicted to disruptAgouti function. (B) Sly is epistatic to Agouti because SlyTo/SlyTo;a/a animals exhibit the same phenotype as SlyTo/SlyTo; AW/AW
animals.
1434 A. Alizadeh et al.
such as Sox18, which acts in the dermal papilla topromote Agouti expression (Pennisi et al. 2000; Fitch
et al. 2003). From this perspective, genetic interactionsbetween Mc1r and Sly would also be helpful in evaluat-ing potential candidate genes. Variation at Mc1r is curi-ously absent from the cat, but prospects for determiningwhether Sly lies upstream, downstream, or parallel toMc1r in the hamster are encouraging, since recessivelyinherited cream coat color in the hamster is thought torepresent the action of Mc1r (Magalhaes 1954; Rob-
inson 1964).These questions can be resolved, of course, by
molecular identification of the Sly and Orange genesand should be facilitated by availability of additionalmammalian genome sequences. Defining the molecu-lar genetics of Sly and Orange may also be useful as atool for understanding the biology of other colorvariation patterns thought to affect the distribution ofpheomelanin and eumelanin, such as tabby striping indomestic and wild cats, zebra striping in horse 3 zebra
hybrids, and rostro-caudal striping seen in squirrelsand/or chipmunks (Searle 1968).
We thank Tyler Vogt for collecting data on hamster hair, AnneSchmidt-Kuntzel and Marilyn Menotti-Raymond for communicatingunpublished results, and Angie Crowley of San Joaquin ValleyFisheries for collecting and organizing founder animals. This workwas supported by funds from the National Institutes of Health and by aStanford Graduate Fellowship (to L.Z.H.).
LITERATURE CITED
Andersson, L., 2003 Melanocortin receptor variants with pheno-typic effects in horse, pig, and chicken. Ann. NY Acad. Sci.994: 313–318.
Barsh, G. S., 2006 Regulation of pigment type-switching by Agouti,Melanocortin signaling, Attractin, and Mahoganoid, pp. 395–410in The Pigmentary System, edited by J. J. Nordlund, R. E. Boissy,V. J. Hearing, R. A. King, W. S. Oetting et al. Blackwell Publish-ing, Oxford.
Bennett, D. C., and M. L. Lamoreux, 2003 The color loci of mice–agenetic century. Pigment Cell Res. 16: 333–344.
Blake, J. A., J. E. Richardson, C. J. Bult, J. A. Kadin and J. T. Eppig,2003 MGD: the Mouse Genome Database. Nucleic Acids Res. 31:193–195.
Figure 5.—Comparative genomics of Slyand Orange. (A) Representative images ofFISH for mouse BAC probes 1 and 2 hy-bridized to mouse or hamster chromo-somes, as indicated. (BAC probes 1, 2,and 3 are, respectively, RP24-77C5, RP24-186P1, and RP24-96H13.) (B) Diagram ofBAC hybridization results showing thatthe region homologous to the cat Orangegene lies in the centromere-proximal re-gion of the mouse X chromosome butthe centromere-distal region of the ham-ster X chromosome. By contrast, geneticmapping studies place the hamster Slygene close to the centromere of Xp. (C)Summary of comparative mapping. Theposition of the cat Orange mutation isbased on work from Schmidt-Kuntzel
et al. (2009).
Sex-linked yellow in Syrian Hamsters 1435

Brudno, M., A. Poliakov, A. Salamov, G. M. Cooper, A. Sidow et al.,2004 Automated whole-genome multiple alignment of rat,mouse, and human. Genome Res. 14: 685–692.
Cone, R. D., D. Lu, S. Koppula, D. I. Vage, H. Klungland et al.,1996 The melanocortin receptors: agonists, antagonists, andthe hormonal control of pigmentation. Recent Prog. Horm.Res. 51: 287–318.
DiPaolo, J. A., and N. C. Popescu, 1973 Distribution of chromo-some constitutive heterochromatin of Syrian hamster cells trans-formed by chemical carcinogens. Cancer Res. 33: 3259–3264.
Eizirik, E., N. Yuhki, W. E. Johnson, M. Menotti-Raymond, S. S.Hannah et al., 2003 Molecular genetics and evolution of mela-nism in the cat family. Curr. Biol. 13: 448–453.
Fitch, K. R., K. A. McGowan, C. D. van Raamsdonk, H. Fuchs, D.Lee et al., 2003 Genetics of dark skin in mice. Genes Dev. 17:214–228.
Hoekstra, H. E., R. J. Hirschmann, R. A. Bundey, P. A. Insel andJ. P. Crossland, 2006 A single amino acid mutation contrib-utes to adaptive beach mouse color pattern. Science 313: 101–104.
Hustad, C. M., W. L. Perry, L. D. Siracusa, C. Rasberry, L. Cobb
et al., 1995 Molecular genetic characterization of six recessiveviable alleles of the mouse agouti locus. Genetics 140: 255–265.
Jackson, I. J., 1997 Homologous pigmentation mutations in hu-man, mouse and other model organisms. Hum. Mol. Genet. 6:1613–1624.
Karolchik, D., R. Baertsch, M. Diekhans, T. S. Furey, A. Hinrichs
et al., 2003 The UCSC genome browser database. Nucleic AcidsRes. 31: 51–54.
Kent, W. J., R. Baertsch, A. Hinrichs, W. Miller and D. Haussler,2003 Evolution’s cauldron: duplication, deletion, and rear-rangement in the mouse and human genomes. Proc. Natl. Acad.Sci. USA 100: 11484–11489.
Kerns, J. A., J. Newton, T. G. Berryere, E. M. Rubin, J. F. Cheng
et al., 2004 Characterization of the dog Agouti gene and a non-agouti mutation in German Shepherd dogs. Mamm. Genome 15:798–808.
Kijas, J. M., M. Moller, G. Plastow and L. Andersson, 2001 Aframeshift mutation in MC1R and a high frequency of somaticreversions cause black spotting in pigs. Genetics 158: 779–785.
Klungland, H., and D. I. Vage, 2003 Pigmentary switches in do-mestic animal species. Ann. NY Acad. Sci. 994: 331–338.
Kuramoto, T., T. Nomoto, A. Fujiwara, M. Mizutani, T. Sugimura
et al., 2002 Insertional mutation of the Attractin gene in theblack tremor hamster. Mamm. Genome 13: 36–40.
Kuroiwa, A., F. Suto, H. Fujisawa and Y. Matsuda, 2001a Chro-mosome assignment of four plexin A genes (Plxna1, Plxna2,Plxna3, Plxna4) in mouse, rat, Syrian hamster and Chinese ham-ster. Cytogenet. Cell Genet. 92: 127–129.
Kuroiwa, A., K. Tsuchiya, K. Matsubara, T. Namikawa and Y.Matsuda, 2001b Construction of comparative cytogeneticmaps of the Chinese hamster to mouse, rat and human. Chromo-some Res. 9: 641–648.
Kuroiwa, A., K. Tsuchiya, T. Watanabe, H. Hishigaki, E. Takahashi
et al., 2001c Conservation of the rat X chromosome gene orderin rodent species. Chromosome Res. 9: 61–67.
Lamoreux, M. L., and T. C. Mayer, 1975 Site of gene action in thedevelopment of hair pigment in recessive yellow (e/e) mice. Dev.Biol. 46: 160–166.
Little, C. C., 1957 The Inheritance of Coat Color in Dogs. Comstock,Ithaca, NY.
Lyon, M. F., 1962 Sex chromatin and gene action in the mammalianX-chromosome. Am. J. Hum. Genet. 14: 135–148.
Magalhaes, H., 1954 Cream and tawny, coat color mutations ingolden hamster, Mesocricetus auratus. Anat. Rec. 120: 752.
Marklund, L., M. J. Moller, K. Sandberg and L. Andersson,1996 A missense mutation in the gene for melanocyte-stimulat-ing hormone receptor (MC1R) is associated with the chestnutcoat color in horses. Mamm. Genome 7: 895–899.
McNulty, J. C., P. J. Jackson, D. A. Thompson, B. Chai, I. Gantz
et al., 2005 Structures of the agouti signaling protein. J. Mol.Biol. 346: 1059–1070.
Mundy, N. I., N. S. Badcock, T. Hart, K. Scribner, K. Janssen et al.,2004 Conserved genetic basis of a quantitative plumage trait in-volved in mate choice. Science 303: 1870–1873.
Nachman, M. W., H. E. Hoekstra and S. L. D’Agostino, 2003 Thegenetic basis of adaptive melanism in pocket mice. Proc. Natl.Acad. Sci. USA 100: 5268–5273.
Newton, J. M., A. L. Wilkie, L. He, S. A. Jordan, D. L. Metallinos
et al., 2000 Melanocortin 1 receptor variation in the domesticdog. Mamm. Genome 11: 24–30.
Nixon, C. W., J. H. Beaumont and M. E. Connelly, 1970 Gene in-teraction of coat patterns and colors in the Syrian hamster.J. Hered. 61: 221–228.
Ohno, S., 1969 Evolution of sex chromosomes in mammals. Annu.Rev. Genet. 3: 495–524.
Pennisi, D., J. Bowles, A. Nagy, G. Muscat and P. Koopman,2000 Mice null for sox18 are viable and display a mild coat de-fect. Mol. Cell. Biol. 20: 9331–9336.
Perry, W. L., T. Nakamura, D. A. Swing, L. Secrest, B. Eagleson
et al., 1996 Coupled site-directed mutagenesis/transgenesisidentifies important functional domains of the mouse agouti pro-tein. Genetics 144: 255–264.
Raudsepp, T., and B. P. Chowdhary, 2008 FISH for mapping singlecopy genes. Methods Mol. Biol. 422: 31–49.
Rieder, S., S. Taourit, D. Mariat, B. Langlois and G. Guerin,2001 Mutations in the agouti (ASIP), the extension (MC1R),and the brown (TYRP1) loci and their association to coat colorphenotypes in horses (Equus caballus). Mamm. Genome 12:450–455.
Robbins, L. S., J. H. Nadeau, K. R. Johnson, M. A. Kelly, L. Roselli-Rehfuss et al., 1993 Pigmentation phenotypes of variant ex-tension locus alleles result from point mutations that alter MSHreceptor function. Cell 72: 827–834.
Robinson, R., 1964 Genetic studies of the Syrian hamster. Genetica35: 241–250.
Robinson, R., 1966 Sex-linked yellow in the Syrian hamster. Nature212: 824–825.
Robinson, R., 1972 Genetic studies of the Syrian hamster. 8. Thesex-linked gene tortoiseshell. Genetica 43: 239–243.
Rozen, S., and H. Skaletsky, 2000 Primer3 on the WWW for gen-eral users and for biologist programmers. Methods Mol. Biol.132: 365–386.
Schmidt-Kuntzel, A., G. Nelson, V. A. David, A. A. Schaffer,E. Eizirik et al., 2009 A domestic cat X chromosome linkagemap and the sex-linked orange locus: mapping of orange, multipleorigins and epistasis over nonagouti. Genetics 181: 1415–1425.
Searle, A. G., 1968 Comparative Genetics of Coat Color in Mammals. Ac-ademic Press, New York.
Silvers, W. K., 1979 The agouti and extension series of alleles,umbrous and sable, pp. 6–44 in The Coat Colors of Mice. Springer-Verlag, New York.
Siracusa, L. D., 1994 The agouti gene: turned on to yellow. TrendsGenet. 10: 423–428.
Stam, P., 1993 Construction of integrated genetic linkage mapsby means of a new computer package: Join Map. Plant J. 3: 739–744.
Steiner, C. C., J. N. Weber and H. E. Hoekstra, 2007 Adaptive var-iation in beach mice produced by two interacting pigmentationgenes. PLoS Biol. 5: e219.
Steingrimsson, E., N. G. Copeland and N. A. Jenkins, 2006 Mousecoat color mutations: from fancy mice to functional genomics.Dev. Dyn. 235: 2401–2411.
Sundberg, J., 1994 Handbook of Mouse Mutations With Skin and HairAbnormalities: Animal Models and Biomedical Tools. CRC Press,Cleveland, OH/Boca Raton, FL.
Wright, S., 1917 Color inheritance in mammals: results of experi-mental breeding can be linked up with chemical researches onpigments—coat colors of all mammals classified as due to varia-tions in action of two enzymes. J. Hered. 8: 224–235.
Wright, S., 1918 Color inheritance in mammals: X. The cat–curious association of deafness with blue-eyed white color andof femaleness with tortoise-shelled color, long known-variationsof tiger pattern present interesting features. J. Hered. 9: 139–144.
Communicating editor: L. Siracusa
1436 A. Alizadeh et al.