expression of matrilin-2 in human skin
TRANSCRIPT

Expression of Matrilin-2 in Human Skin
Dorothea Piecha, Karin Hartmann, Birgit Kobbe,* Ingo Haase, Cornelia Mauch, Thomas Krieg, andMats Paulsson*Department of Dermatology and *Institute for Biochemistry, Medical Faculty, University of Cologne, Germany
The extracellular matrix is composed of a largenumber of different modular proteins. Matrilin-2 is anewly described member of the protein superfamilywith von Willebrand factor A-like modules. Toexamine the expression of matrilin-2 in human skin,the distribution of protein and mRNA was studied byimmunohistochemistry and in situ hybridization. Inaddition, immunoblotting and real-time reverse tran-scription polymerase chain reaction were used toinvestigate the expression of matrilin-2 in keratino-cyte and ®broblast cultures. In vivo, keratinocytes and®broblasts were both found to express matrilin-2mRNA and deposit the protein at the basal side of the
dermal±epidermal basement membrane. Matrilin-2molecules synthesized by the two cell types in vitroappeared to be processed differently by cell-associatedproteases. Transcription of matrilin-2 mRNA inkeratinocytes was enhanced by a diffusible factor pro-duced by ®broblasts, suggesting a regulatory mechan-ism for the production of extracellular matrix at thedermal±epidermal junction. These ®ndings demon-strate that matrilin-2 is expressed in normal skin bykeratinocytes and ®broblasts and may thus contributeto cutaneous homeostasis. Key words: cutaneous/extra-cellular matrix/laminin/matrilin/skin. J Invest Dermatol119:38±43, 2002
The extracellular matrix (ECM) is composed of a largenumber of different molecules, which determine thehistoarchitecture speci®c to every organ and providecells with crucial information on migration, adhesion,and differentiation (Krieg and LeRoy, 1998).
Molecules of the ECM are produced by adjacent cells such as®broblasts, osteocytes, and epithelial cells and can ful®ll bothstructural and regulatory functions. The expression and assembly ofECM molecules is often determined by complex regulatoryprocesses, including transcriptional and translational events as wellas post-translational modi®cations. Most ECM molecules sharehomologous modules that consist of speci®c protein domainsderived from a common evolutionary origin.
The family of matrilins are typical modular proteins belonging toa superfamily characterized by von Willebrand factor A-likedomains (Deak et al, 1999; Paulsson et al, 1999). To date, fourdifferent matrilins have been identi®ed, but their biologic role isstill largely unknown. Matrilin-1 (Paulsson and HeinegaÊrd, 1979;Kiss et al, 1989), formerly also named cartilage matrix protein, andmatrilin-3 (Wagener et al, 1997; Belluoccio et al, 1998; Klatt et al,2000) are mainly expressed in cartilage, whereas matrilin-2 (Deaket al, 1997; Piecha et al, 1999) and matrilin-4 (Wagener et al, 1998a,b; Klatt et al, 2001) have been found in a number of differenttissues. Mice lacking matrilin-1 show no overt phenotype, but anabnormal cartilage collagen ®brillogenesis and ®bril organizationhas been described (Huang et al, 1999; AszoÂdi et al, 1999). Inhumans, immunoreactivity against matrilin-1 has been shown to beinvolved in relapsing polychondritis (Buckner et al, 2000).
Matrilin-2 consists of 956 amino acids and is composed of twovon Willebrand factor A-like domains, 10 epidermal growth factor-
like modules, one unique sequence, and a coiled-coil domain(Deak et al, 1997). Via the coiled-coil domain, matrilin-2 assemblesinto oligomers, and monomers, dimers, trimers, and tetramers ofthe matrilin-2 subunit can be detected in tissue extracts and in themedium of cultured cells (Piecha et al, 1999). A similarly complexband pattern is seen with matrilin-4 and, in this case, it has recentlybeen shown that the protein becomes proteolytically processedbefore or at secretion (Klatt et al, 2001). The cleavage of matrilin-4occurs C-terminal from two glutamic acid residues found in thehinge region between the second von Willebrand factor A domainand the coiled-coil region and results in the release of an almostcomplete subunit from the rest of the molecule. In the homologousregion of the matrilin-2 sequence, two pairs of glutamic acidresidues occur and, by analogy, it is likely that the complex bandpattern seen with extracellular matrilin-2 (Piecha et al, 1999) is dueto an extensive proteolytic processing at these sites. The genecoding for matrilin-2 has recently been mapped to chromosomeposition 8q22 (Muratoglu et al, 2000). Expression of matrilin-2 hasbeen demonstrated in the ECM of different murine tissues,including kidney, heart, lung, and esophagus (Piecha et al, 1999).Matrilin-2 has also been shown to be present in murine skin, with aprominent expression throughout the papillary dermis and aroundskin appendages such as vessels, nerves, muscles, and sweat glands.The human matrilin-2 precursor displays 93% similarity to themouse protein and gene expression has been demonstrated inseveral human cell lines of ®broblastic and epithelial origin(Muratoglu et al, 2000).
In order to elucidate the expression of matrilin-2 in human skin,its protein and mRNA distribution was studied by immuno-histochemistry and in situ hybridizaton. We show that matrilin-2protein is mainly expressed at the dermal side of the basementmembrane and around skin appendages. Keratinocytes and®broblasts both express matrilin-2 mRNA and protein, but appearto process the secreted protein differently. Fibroblasts were foundto enhance expression of matrilin-2 mRNA in keratinocytes.
Manuscript received July 11, 2001; revised January 23, 2002; acceptedfor publication February 4, 2002.
Reprint requests to: Dr. Karin Hartmann, Department of Dermatology,University of Cologne, Joseph-Stelzmann-Str. 9, 50931 Cologne,Germany; Email: [email protected]
0022-202X/02/$15.00 ´ Copyright # 2002 by The Society for Investigative Dermatology, Inc.
38
MATERIALS AND METHODS
Tissue Normal skin was obtained with written consent of the patients.Donors included an infant of a few weeks of age and adults between theage of 40 and 60 y. Biopsies were either snap-frozen and stored at±80°C until analysis or ®xed in 4% paraformaldehyde and embedded inparaf®n.
Cell culture Primary human ®broblasts were established by outgrowthfrom biopsies of normal skin as previously described (Krieg et al, 1980).Fibroblasts were maintained in Dulbecco's modi®ed Eagle's medium(DMEM) supplemented with 10% fetal bovine serum, 50 mg per mlsodium ascorbate, 2 mM glutamine, and antibiotics. Primary humankeratinocytes were isolated as previously described (Watt, 1998) andgrown on feeder layers of 3T3 cells in FAD medium supplemented with0.5 mg per ml insulin, 0.5 mg per ml hydrocortisone, 0.1 nM choleraenterotoxin, and 10 ng per ml epidermal growth factor. For someexperiments, ®broblasts and keratinocytes were also cultured together for48 h in a transwell system, where the two cell types were separated by aporous membrane allowing the passage of supernatants but no directcontact between the two cell types.
Immunohistochemistry Indirect immunoperoxidase staining of 6 mm-thin cryosections was performed as previously described (Hauser andPaulsson, 1994) using polyclonal af®nity-puri®ed antisera against matrilin-2 (Piecha et al, 1999) or laminin-1, a swine antirabbit horseradishperoxidase-conjugated IgG (Dako), and 3-amino-9-ethyl-carbazole(Sigma) as reagents. To further investigate deposition of matrilin-2 at thedermal±epidermal junction, indirect immunohistochemistry was alsoperformed on salt-split skin treated with 1 M NaCl at 4°C for 72 h aspreviously described (Gammon et al, 1984; Zillikens et al, 1996).
Preparation of RNA probes Total RNA of human skin was isolatedusing the RNeasy Mini Kit (Qiagen). Using 1 mg of cutaneous RNA astemplate, reverse transcription was carried out using SuperscriptÔ IIRNase H± Reverse Transcriptase (Life Technologies). Polymerase chainreaction (PCR) was performed using the following primers: for matrilin-2, 5¢-AATGGAAAAGATGCTCGCAGG-3¢ (mat2h1) and sense 5¢-TGATGCAGAAGTGGGCACAG-3¢ (mat2h2); for keratin 14, 5¢-ATG-GCAGAGAAGAACCGCAAG-3¢ (kera1) and sense 5¢-GATCTT-CCAGTGGGATCTGTG-3¢ (kera4); and AmpliTaq DNA Polymerase(Perkin Elmer). The resulting 763 bp PCR product coding for matrilin-2 as well as the 603 bp product coding for keratin 14 were cloned in thepCRII Vector (Invitrogen). In vitro transcription of nonradioactiveantisense and sense RNAs was performed using the DigoxigeninLabeling Kit (Roche Molecular Biochemicals). Nonradioactive antisenseand sense RNA probes transcribed from a 206 bp cDNA styI-KpnIfragment coding for the 5¢ untranslated region of the a1(I) collagen werekindly provided by Dr. N. Miosge (Department of Anatomy, Universityof Goettingen, Germany) (Miosge et al, 1998).
In situ hybridization Four-micron-thin paraf®n-embedded sectionswere dewaxed in xylene, rehydrated, and washed in phosphate-bufferedsaline (PBS). After digesting with 10 mg per ml proteinase K, sectionswere post®xed and acetylated with 0.25% acetic anhydride. Sectionswere then hybridized overnight at 46°C with digoxigenin-labeledriboprobes. As controls, serial sections were treated with a sense RNAprobe instead of the antisense probe. After hybridization, sections werewashed in 50% formamide, 2 3 sodium citrate/chloride buffer (SSC)(0.3 M NaCl containing 0.03 M sodium citrate) at 42°C (matrilin-2 andcollagen type I) or 46°C (keratin 14) for 30 min, digested with RNaseA, and washed once with 2 3 SSC and twice with 0.2 3 SSC at 42°C(matrilin-2 and collagen type I) or 46°C (keratin 14) for 20 min each.Color detection of the digoxigenin-labeled sections was carried out asrecommended by the manufacturer (Roche Molecular Biochemicals)with the additional use of polyvinyl alcohol in the detection buffer (DeBlock and Debrouwer, 1993).
Northern hybridization In order to investigate epidermal and dermalRNA separately, epidermis and dermis were separated by incubationwith 0.5 mg per ml thermolysin in 10 mM HEPES buffer containing150 mM NaCl, 6 mM KCl, 1 mM CaCl2, and 1 mM MgCl2, pH 7.4,overnight at 4°C. Total RNA and mRNA of epidermis and dermis wereprepared from guanidinium thiocyanate extracts using the isolationRNeasy Kit and Oligotex mRNA Kit (Qiagen). For RNA blot analysis,10 mg total RNA or 2 mg mRNA were electrophoresed, blotted toHybond N ®lters (Perkin Elmer), and hybridized with probes speci®c formatrilin-2, glyceraldehyde-3-phosphate dehydrogenase (GAPDH), orcollagen type I.
Immunoblot analysis Cells were washed in PBS and cultured inserum-free DMEM, and supernatants were harvested after 48 h. In thecase of keratinocytes, feeder cells were ®rst removed by trypsin/ethylenediamine tetraacetic acid and cells were then cultured with orwithout ®broblasts in serum-free medium for 48 h. Supernatants wereanalyzed by sodium dodecyl sulfate polyacrylamide gel electrophoresis(SDS-PAGE) without prior reduction as previously described (Piecha
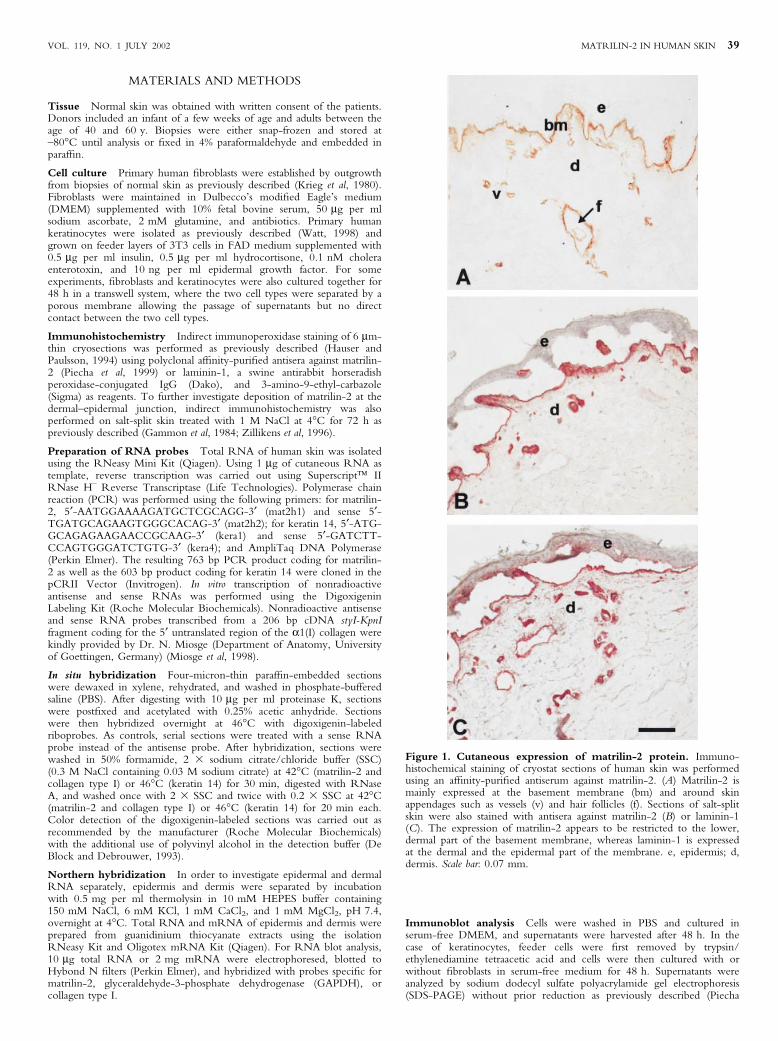
Figure 1. Cutaneous expression of matrilin-2 protein. Immuno-histochemical staining of cryostat sections of human skin was performedusing an af®nity-puri®ed antiserum against matrilin-2. (A) Matrilin-2 ismainly expressed at the basement membrane (bm) and around skinappendages such as vessels (v) and hair follicles (f). Sections of salt-splitskin were also stained with antisera against matrilin-2 (B) or laminin-1(C). The expression of matrilin-2 appears to be restricted to the lower,dermal part of the basement membrane, whereas laminin-1 is expressedat the dermal and the epidermal part of the membrane. e, epidermis; d,dermis. Scale bar: 0.07 mm.
VOL. 119, NO. 1 JULY 2002 MATRILIN-2 IN HUMAN SKIN 39

et al, 1999). Brie¯y, proteins were electrophoresed on gradient gelscontaining 4%±15% polyacrylamide, transferred to nitrocellulose ®lters,and visualized using af®nity-puri®ed antiserum against matrilin-2, swineantirabbit horseradish peroxidase-conjugated IgG (DAKO), and the ECLchemiluminescence reagent (Amersham Pharmacia Biotech).
Real-time quantitative reverse transcription PCR (RT-PCR)Fibroblasts and keratinocytes were cultured alone or in a transwellcoculture system, where the two cell types were separated by amembrane, for 48 h. Total RNA was then isolated separately from eachcell type using the RNeasy Kit (Qiagen). One microgram of total RNAwas reversely transcribed with MuLV reverse transcriptase (LifeTechnologies) using random hexamers. Pairs of primers for ampli®cationof matrilin-2 and ribosomal protein s26 were designed using the PrimerExpress Software (Applied Biosystems). In all experiments, primerconcentrations were ®rst optimized in order to avoid unspeci®c bindingof primers, and after running the PCR a dissociation curve analysis wasperformed to verify the speci®city of the ampli®cation products. Primersequences used were as follows: 5¢-GACGGACGGGCTCAGGAT-3¢(373mtr2) and 5¢-GATACCATTGGCCTTGGCTTTA-3¢ (432mtr2)for matrilin-2, and sense 5¢-GCAGCAGTCAGGGACATTTCTG-3¢and reverse 5¢-TTCACATACAGCTTGGGAAGCA-3¢ for ribosomalprotein s26.
Real-time quantitative PCR was performed using the GeneAmp 5700system (Applied Biosystems) with the double-stranded DNA-binding dyeSYBR Green I and the AmpliTAQ gold polymerase (AppliedBiosystems). Quanti®cation of the samples was performed by measure-ment of the threshold cycle using a standard curve that determines thestarting copy numbers of nucleic acid targets. To normalize the samplesfor differences in the amounts of total RNA added to each cDNAtranscription, s26 ribosomal protein was used as endogenous control. Toavoid contamination, all assays were performed using the universalthermal cycling parameters (Applied Biosystems) with AmpErase UNG.All experiments were performed in duplicate.
RESULTS
Cutaneous expression of matrilin-2 protein To study theexpression of matrilin-2 in normal human skin, indirectimmunohistochemistry was performed (Fig 1A). Deposition ofmatrilin-2 protein was restricted to a narrow zone close to thebasement membrane at the dermal±epidermal junction and to areasaround vessels and appendages. Similar results were obtained fromindividuals of different age. Sections of salt-split skin demonstratedthat the deposition of matrilin-2 is restricted to the lower, dermalpart of the basement membrane zone (Fig 1B). In contrast,laminin-1, which is known to be an integral protein of thebasement membrane, was found at the lower as well as the upperpart of the basement membrane (Fig 1C).
Cutaneous expression of matrilin-2 mRNA In order toinvestigate the cellular source of matrilin-2, in situ hybridization ofhuman skin was performed (Fig 2). In contrast to the proteindistribution, matrilin-2 mRNA was found to be expressed in boththe epidermis and dermis (Fig 2A). Keratinocytes, ®broblasts, andendothelial cells all showed expression of matrilin-2 mRNA. Thespeci®city of the method used was demonstrated by the fact that anantisense riboprobe for keratin 14 hybridized speci®cally to basalkeratinocytes (Fig 2C) and a probe for the a1 chain of collagentype I to dermal ®broblasts (Fig 2E).
To quantify the expression of matrilin-2 mRNA in differentcutaneous layers, northern blot analysis was additionally performedusing separated dermal and epidermal tissue after treatment withthermolysin (Fig 3). Total RNA was isolated from dermis andepidermis and hybridized with a probe speci®c for matrilin-2(Fig 3A). Matrilin-2 mRNA showed nearly equal expression in
Figure 2. In situ hybridization of matrilin-2cRNA to human skin. Nonradioactive antisense(A) and sense (B) riboprobes speci®c for matrilin-2 mRNA were hybridized to paraf®n-embeddedsections of human skin. Matrilin-2 mRNA isfound to be expressed in the epidermis anddermis. The speci®city of the nonradioactivemethod was demonstrated using antisense (C) andsense (D) riboprobes for keratin 14 and antisense(E) and sense (F) riboprobes for the a1 chain ofcollagen type I that hybridized speci®cally to basalkeratinocytes or dermal ®broblasts, respectively.Scale bar: 0.1 mm.
40 PIECHA ET AL THE JOURNAL OF INVESTIGATIVE DERMATOLOGY
dermis and epidermis. Equal loading of RNA was con®rmed byGAPDH hybridization (Fig 3B). To exclude contamination fromother cutaneous layers, hybridization of dermis-speci®c collagentype I mRNA was also performed (Fig 3C).
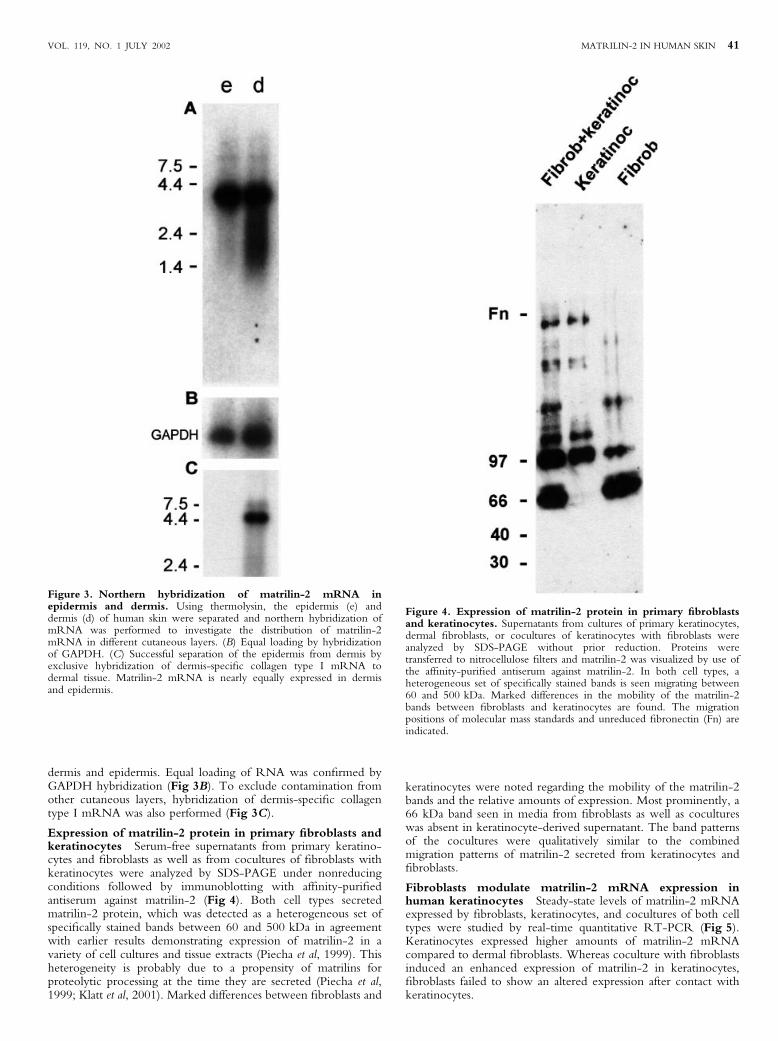
Expression of matrilin-2 protein in primary ®broblasts andkeratinocytes Serum-free supernatants from primary keratino-cytes and ®broblasts as well as from cocultures of ®broblasts withkeratinocytes were analyzed by SDS-PAGE under nonreducingconditions followed by immunoblotting with af®nity-puri®edantiserum against matrilin-2 (Fig 4). Both cell types secretedmatrilin-2 protein, which was detected as a heterogeneous set ofspeci®cally stained bands between 60 and 500 kDa in agreementwith earlier results demonstrating expression of matrilin-2 in avariety of cell cultures and tissue extracts (Piecha et al, 1999). Thisheterogeneity is probably due to a propensity of matrilins forproteolytic processing at the time they are secreted (Piecha et al,1999; Klatt et al, 2001). Marked differences between ®broblasts and
keratinocytes were noted regarding the mobility of the matrilin-2bands and the relative amounts of expression. Most prominently, a66 kDa band seen in media from ®broblasts as well as cocultureswas absent in keratinocyte-derived supernatant. The band patternsof the cocultures were qualitatively similar to the combinedmigration patterns of matrilin-2 secreted from keratinocytes and®broblasts.
Fibroblasts modulate matrilin-2 mRNA expression inhuman keratinocytes Steady-state levels of matrilin-2 mRNAexpressed by ®broblasts, keratinocytes, and cocultures of both celltypes were studied by real-time quantitative RT-PCR (Fig 5).Keratinocytes expressed higher amounts of matrilin-2 mRNAcompared to dermal ®broblasts. Whereas coculture with ®broblastsinduced an enhanced expression of matrilin-2 in keratinocytes,®broblasts failed to show an altered expression after contact withkeratinocytes.
Figure 3. Northern hybridization of matrilin-2 mRNA inepidermis and dermis. Using thermolysin, the epidermis (e) anddermis (d) of human skin were separated and northern hybridization ofmRNA was performed to investigate the distribution of matrilin-2mRNA in different cutaneous layers. (B) Equal loading by hybridizationof GAPDH. (C) Successful separation of the epidermis from dermis byexclusive hybridization of dermis-speci®c collagen type I mRNA todermal tissue. Matrilin-2 mRNA is nearly equally expressed in dermisand epidermis.
Figure 4. Expression of matrilin-2 protein in primary ®broblastsand keratinocytes. Supernatants from cultures of primary keratinocytes,dermal ®broblasts, or cocultures of keratinocytes with ®broblasts wereanalyzed by SDS-PAGE without prior reduction. Proteins weretransferred to nitrocellulose ®lters and matrilin-2 was visualized by use ofthe af®nity-puri®ed antiserum against matrilin-2. In both cell types, aheterogeneous set of speci®cally stained bands is seen migrating between60 and 500 kDa. Marked differences in the mobility of the matrilin-2bands between ®broblasts and keratinocytes are found. The migrationpositions of molecular mass standards and unreduced ®bronectin (Fn) areindicated.
VOL. 119, NO. 1 JULY 2002 MATRILIN-2 IN HUMAN SKIN 41

DISCUSSION
Most murine cells producing matrilin-2 are of mesodermal origin(Piecha et al, 1999). Epithelial cells have also been shown to expressmatrilin-2 mRNA, but protein deposition was only observed inadjacent subepithelial tissues. In murine skin, matrilin-2 has beendemonstrated to be prominently expressed throughout the upperdermis and around vessels, nerves, muscles, and sweat glands(Piecha et al, 1999).
In this study, the expression of matrilin-2 in human skin clearlydiffered from the expression in murine skin. In humans, matrilin-2protein was con®ned to a narrow zone close to the basementmembrane at the dermal±epidermal junction and to areas aroundblood vessels. Staining of salt-split skin and comparison with thedistribution of the ubiquitous basement membrane protein laminindemonstrated that matrilin-2 is only deposited at the lower part ofthe basement membrane or the upper part of the dermis adjacent tothe basement membrane. In situ hybridization and northern blotanalysis also revealed that matrilin-2 mRNA in humans was foundin both dermis and epidermis. Although matrilin-2 protein was notabundant in resting epidermis, keratinocytes in culture were alsoclearly capable of expressing matrilin-2. It may therefore be thattranslation of matrilin-2 mRNA is determined by extracellularin¯uences. Alternatively, matrilin-2 may be produced by epidermalkeratinocytes, but would then rapidly be transported to subepi-thelial tissues. A similar mechanism of protein processing has beensuggested for laminin (Smola et al, 1998) and collagen type VII(KoÈnig and Bruckner-Tuderman, 1994), although it is not knownhow these proteins are transported across the basement membraneand how they are then incorporated into the mesenchymal stroma.The presence of speci®c binding ligands or interacting partners maybe associated with the restricted deposition of matrilin-2 protein inthe basement membrane zone.
In vitro studies also showed that primary dermal ®broblasts andkeratinocytes are able to secrete matrilin-2 protein. As has beendescribed, the secreted matrilin-2 displayed a large structuralheterogeneity (Piecha et al, 1999), presumably corresponding topost-translational modi®cations speci®c for each cellular origin. Inanalogy with the better studied matrilin-4 (Klatt et al, 2001), it islikely that this heterogeneity is related to proteolytic processing,
with ®broblasts yielding smaller fragments than keratinocytes.When the pooled media from cocultures were analyzed, the bandpattern observed largely corresponded to the sum of the patternsobtained with the two cell types in separate cultures and the largercomponents produced by keratinocytes were not further degradedby ®broblast proteases. It can thus be concluded that the proteasescontributing to the processing must be cell-associated and are notable to act on matrilin-2 secreted by another cell type. Further, thepronounced cleavage of matrilin-2 by ®broblasts (Fig 4) mayhamper the matrix deposition of protein secreted by these cells.
The transcription of matrilin-2 mRNA in keratinocytes isenhanced by coculture with ®broblasts, whereas the same process in®broblasts is not affected by the presence of keratinocytes.Accordingly, ®broblasts must release a diffusible factor regulatingmatrilin-2 gene expression. It is possible that such interplaybetween ®broblasts and keratinocytes is of importance in regulatingECM production during wound healing.
We thank C. Berns, Department of Dermatology, University of Cologne,
Germany, for excellent technical assistance and Dr. N. Miosge, Department of
Anatomy, University of Goettingen, Germany, for donating riboprobes speci®c for
a1 collagen type I. This work was supported by a grant from the German Research
Foundation (Deutsche Forschungsgemeinschaft) to M.P. (FOR 265/2-1).
REFERENCES
AszoÂdi A, Bateman JF, Hirsch E, et al: Normal skeletal development of mice lackingmatrilin 1: redundant function of matrilins in cartilage? Mol Cell Biol 19:7841±7845, 1999
Belluoccio D, Schenker T, Baici A, Trueb B: Characterization of human matrilin-3(MATN3). Genomics 53:391±394, 1998
Buckner JH, Wu JJ, Reife RA, Terato K, Eyre DR: Autoreactivity against matrilin-1in a patient with relapsing polychondritis. Arthritis Rheum 43:939±943, 2000
Deak F, Piecha D, Bachrati C, Paulsson M, Kiss I: Primary structure and expressionof matrilin-2, the closest relative of cartilage matrix protein within the vonWillebrand factor type A-like module superfamily. J Biol Chem 272:9268±9274,1997
Deak F, Wagener R, Kiss I, Paulsson M: The matrilins: a novel family of oligomericextracellular matrix proteins. Matrix Biol 18:55±64, 1999
De Block M, Debrouwer D: RNA-RNA in situ hybridization using digoxigenin-labeled probes: the use of high-molecular-weight polyvinyl alcohol in thealkaline phosphatase indoxyl-nitroblue tetrazolium reaction. Anal Biochem215:86±89, 1993
Gammon WR, Briggaman RA, Inman AO, Queen LL, Wheeler CE: Differentiatinganti-lamina lucida and anti-sublamina densa anti-BMZ antibodies by indirectimmuno¯uorescence on 1.0 M sodium chloride-separated skin. J InvestDermatol 82:139±144, 1984
Hauser N, Paulsson M: Native cartilage matrix protein (CMP). A compact trimer ofsubunits assembled via a coiled-coil alpha-helix. J Biol Chem 269:25747±25753,1994
Huang X, Birk DE, Goetinck PF: Mice lacking matrilin-1 (cartilage matrix protein)have alterations in type II collagen ®brillogenesis and ®bril organization. DevDyn 216:434±441, 1999
Kiss I, Deak F, Holloway RG, et al: Structure of the gene for cartilage matrix protein,a modular protein of the extracellular matrix. J Biol Chem 264:8126±8134, 1989
Klatt AR, Nitsche DP, Kobbe B, Morgelin M, Paulsson M, Wagener R: Molecularstructure and tissue distribution of matrilin-3, a ®lament-forming extracellularmatrix protein expressed during skeletal development. J Biol Chem 275:3999±4006, 2000
Klatt A, Nitsche DP, Kobbe B, Macht M, Paulsson M, Wagener R: Molecularstructure, processing, and tissue distribution of matrilin-4. J Biol Chem276:17267±17275, 2001
KoÈnig A, Bruckner-Tuderman L: Transforming growth factor-beta promotesdeposition of collagen VII in a modi®ed organotypic skin model. Lab Invest70:203±209, 1994
Krieg T, LeRoy EC: Diseases of the extracellular matrix. J Mol Med 76:224±225,1998
Krieg T, Aumailley M, Dessau W, Wiestner M, MuÈller P: Synthesis of collagen byhuman ®broblasts and their SV40 transformants. Exp Cell Res 125:23±30, 1980
Miosge N, Waletzko K, Bode C, Quondamatteo F, Schutz W, Herken R: Light andelectron microscopic in-situ hybridization of collagen type I and type II mRNAin the ®brocartilaginous tissue of late-stage osteoarthritis. Osteoarthritis Cartilage6:278±285, 1998
Muratoglu S, Krysan K, Balazs M, et al: Primary structure of human matrilin-2,chromosome location of the MATN2 gene and conservation of an AT-ACintron in matrilin genes. Cytogenet Cell Genet 90:323±327, 2000
Paulsson M, HeinegaÊrd D: Matrix proteins bound to associatively preparedproteoglycans from bovine cartilage. Biochem J 183:539±545, 1979
Figure 5. Quantitative RT-PCR of RNA from primary human®broblasts, keratinocytes, and cocultures. Total RNA of ®broblast(F) and keratinocyte (K) monolayers as well as ®broblasts cocultured withkeratinocytes (F(co-K)) and keratinocytes cocultured with ®broblasts(K(co-F)) was isolated and matrilin-2 mRNA was quanti®ed by real-timeRT-PCR. Keratinocytes express higher amounts of matrilin-2 mRNAcompared to ®broblasts. Matrilin-2 expression in keratinocytes isenhanced by coculture with ®broblasts, whereas keratinocytes fail to alterexpression of matrilin-2 in ®broblasts.
42 PIECHA ET AL THE JOURNAL OF INVESTIGATIVE DERMATOLOGY

Paulsson M, Piecha D, Segat D, Smyth N, Wagener R: The matrilins: a growingfamily of A-domain-containing proteins. Biochem Soc Trans 27:824±826, 1999
Piecha D, Muratoglu S, MoÃrgelin M, et al: Matrilin-2, a large, oligomeric matrixprotein, is expressed by a great variety of cells and forms ®brillar networks. J BiolChem 274:13353±13361, 1999
Smola H, Stark HJ, Thiekotter G, Mirancea N, Krieg T, Fusenig NE: Dynamics ofbasement membrane formation by keratinocyte±®broblast interactions inorganotypic skin culture. Exp Cell Res 239:399±410, 1998
Wagener R, Kobbe B, Paulsson M: Primary structure of matrilin-3, a new memberof a family of extracellular matrix proteins related to cartilage matrix protein(matrilin-1) and von Willebrand factor. FEBS Lett 413:129±134, 1997
Wagener R, Kobbe B, Paulsson M: Matrilin-4, a new member of the matrilin familyof extracellular matrix proteins. FEBS Lett 436:123±127, 1998a
Wagener R, Kobbe B, Paulsson M: Genomic organisation, alternative splicing andprimary structure of human matrilin-4. FEBS Lett 438:165±170, 1998b
Watt FM: Cultivation of human epidermal keratinocytes with a 3T3 feeder layer. In:Celis JE, ed. Cell Biology: a Laboratory Handbook. London: Academic Press,1998:pp 113±118
Zillikens D, Kawahara Y, Ishiko A, et al: A novel subepidermal blistering disease withautoantibodies to a 200-kDa antigen of the basement membrane zone. J InvestDermatol 106:1333±1338, 1996
VOL. 119, NO. 1 JULY 2002 MATRILIN-2 IN HUMAN SKIN 43