expression of cyclooxygenase-2 (cox-2) in hepatocellular...
TRANSCRIPT

Expression of Cyclooxygenase-2 (COX-2) in HepatocellularCarcinoma and Growth Inhibition of Hepatoma CellLines by a COX-2 Inhibitor, NS-3981
Si Hyun Bae, Eun Sun Jung, Young Min Park,2
Boo Sung Kim, Byung Kee Kim, Dong Goo Kim,and Wang Shick RyuDepartments of Internal Medicine [S. H. B., Y. M. P., B. S. K.],Pathology [E. S. J., B. K. K.], and General Surgery [D. K. K.], andWHO Collaborating Center for Reference and Research on ViralHepatitis, College of Medicine, Catholic University of Korea, andDepartment of Biochemistry, College of Science, Yonsei University,Seoul, Korea [W. S. R.]
ABSTRACTCyclooxygenase-2 (COX-2) has been suggested to be
associated with carcinogenesis. In hepatocellular carcinoma(HCC), the expression pattern of COX-2 protein has beenwell correlated with the differentiation grade, suggestingthat abnormal COX-2 expression plays an important role inhepatocarcinogenesis. We investigated the expression pat-tern and clinical significance of COX-2 in HCC tissues. Inaddition, we evaluated the efficacy of a selective COX-2inhibitor, NS-398, in three hepatoma cell lines. Thirty-sixHCC tissues, 15 hepatoma cell lines, 1 colorectal cell line(HT-29), and 1 fibroblast cell line (SV80) were included inthe study. We evaluated serological tests and histologicaland radiological evaluations of HCC tissues. Immunohisto-chemical staining for COX-2 was performed on 36 HCCtissues and 17 cancer cell lines. A cell viability assay forgrowth inhibition of NS-398 in five cell lines was performed.Immunohistochemically, all six well-differentiated HCCswere positive, whereas 83% (10 of 12) of the poorly differ-entiated HCCs were negative. There was no significant re-lationship between the intensity of COX-2 expression andthe level ofa-fetoprotein, tumor size, presence of portal veinthrombosis, tumor capsule and metastasis, Tumor-Node-Metastasis staging, and growth types (P > 0.05). Accordingto the cell viability assay, NS-398 suppressed the growth ofall cell lines, independent of the degree of COX-2 expression.
The inhibitory effect on each cell line was identified in 10mM
NS-398 and was significantly strong in 100mM NS-398. All celllines exhibited apoptosis, which was identified by 4*-6-diamidino-2-phenylindole staining. In conclusion, COX-2 maybe a determinant of the differentiation grade of HCC, and theinhibition of COX-2 can induce growth suppression of hepa-toma cell lines via induction of apoptosis.
INTRODUCTIONHCC3 is the leading cause of malignant cancer death in the
world (1). However, the exact molecular mechanism of hepato-carcinogenesis is still unclear. COX-2, a rate-limiting enzyme inthe pathway of PG synthesis, is one of the interesting cellularfactors and has been suggested to be associated with carcino-genesis in colorectal cancer (2–5). In addition, expression ofCOX-2 has also been found to be high in other types of cancersincluding prostate (6), breast, gastric (7, 8), pancreatic (9), lung(10), and head and neck cancers (11). Empirically, inhibition ofCOX-2 with NSAIDs such as aspirin could induce the regres-sion of colon polyps in patients with familial adenomatouspolyposis (12). Experimentally, the size and frequency of intes-tinal polyps were markedly reduced in mice with a doubleknockout ofadenomatous polyposis coliandCOX-2genes (13).Furthermore, a selective inhibition of COX-2 brought the nearlycomplete suppression of azoxymethane-induced colon cancer(14).
COX, known as PG synthase, catalyzes the metabolism ofarachidonic acid to PGs and thromboxanes (15), and two iso-forms, COX-1 and COX-2, have been identified. COX-1 isconstitutively present in many cell types and is responsible forvarious cytoprotective prostanoids in a number of organs, suchas the gastric mucosa and the kidneys, whereas COX-2 isusually absent under basal conditions but inducible in certaincells by mitogens, cytokines, and other factors (16, 17). Aspirin,which acetylates a serine on the substrate binding site of COX-1and COX-2, completely blocks the substrate use of COX-1 andPGH2 formation by COX-2 (18, 19). Thus, the ability ofNSAIDs like aspirin to inhibit COX-2 may well explain thetherapeutic utility. In contrast, inhibition of COX-1 may explainthe unwanted side effects of nonspecific NSAIDs, such asgastric toxicity and renal dysfunction (20). Recently, newNSAIDs, which are selective inhibitors of COX-2, were discov-ered, and these exert therapeutic efficacy without the toxic
Received 9/11/00; revised 1/8/01; accepted 1/11/01.The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisementin accordance with 18 U.S.C. Section 1734 solely toindicate this fact.1 Supported by Molecular Medicine Research Group Program Grant2000-J-MM-01-B-10 from the Ministry of Science and Technology andthe St. Vincent Hospital Research Fund.2 To whom requests for reprints should be addressed, at Departmentof Internal Medicine, Kangnam St. Mary’s Hospital, College ofMedicine, Catholic University of Korea, # 505 Banpo-dong, Seocho-ku,137-040 Seoul, Korea, or 134 Shinchon-Dong, Seodaemun-ku, 120-752,Seoul, Korea. Phone: 82-2-590-2388; Fax: 82-2-536-9559; E-mail:[email protected].
3 The abbreviations used are: HCC, hepatocellular carcinoma; COX-1,-2, cyclooxygenase-1, -2; NSAIDs, nonsteroidal anti-inflammatorydrugs; HBV, hepatitis B virus; HCV, hepatitis C virus; PG, prostag-landin; AFP,a-fetoprotein; TNM, Tumor-Node-Metastasis staging sys-tem; DAPI, 49-6-diamidino-2-phenylindole; T, tumor; NT, nontumor.
1410Vol. 7, 1410–1418, May 2001 Clinical Cancer Research
Research. on August 19, 2019. © 2001 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

effects from the inhibition of COX-1. A recent study has dem-onstrated that a selective COX-2 inhibitor reduced the growth ofcolon cancer in carcinogen-treated rats and also inhibited thevarious cancer cell lines (21–23).
In HCC, the expression pattern of COX-2 protein wasfound to be well correlated with the differentiation grade, sug-gesting that abnormal COX-2 expression plays an important rolein hepatocarcinogenesis (24, 25). In the present study, the ex-pression pattern of COX-2 was investigated in HCC tissues forevaluation of its clinical significance. Furthermore, we demon-strated that a selective COX-2 inhibitor, NS-398, strongly in-hibited the proliferation of hepatoma cell lines.
MATERIALS AND METHODSMaterials
Human Tissues. A total of 36 paired specimens of tumorand surrounding nontumor liver tissues were obtained fromHCC patients, who were treated surgically in Kangnam St.Mary’s Hospital, The Catholic University of Korea, Seoul,
Korea, between 1994 and 1999. Nontumor tissues, which werecomprised of 5 chronic active hepatitis-based livers and 31chronic active hepatitis tissues with cirrhosis, were examined.Before surgical resection of HCC, 15 patients were treated withtransarterial chemotherapy. All specimens for this study werefresh tissues without necrosis from transarterial chemotherapy.The characteristic features of the patients are summarized inTable 1.
Cancer Cell Lines. A total of 15 hepatoma cell lines(HepG2, HepG2.2.15, HLE, HLF, HuH-7, Hep3B, PLC/PRF-5,SNU182, SNU354, SNU368, SNU387, SNU398, SNU423,SNU449, and SNU475), one colorectal cell line (HT-29), andone fibroblast cell line (SV80) were all obtained from the KoreaCell Bank (Seoul, Korea).
MethodsSerological Tests. HBV surface antigen and anti-HCV
antibody were measured by commercially available enzyme
Table 1 Patient characteristics of HCC and COX-2 expression in tumor and nontumor tissue according to histological degree of differentiation
Patientno. Age Sex Cause Size (cm)
Morphologicalpattern TNM stage
Histologicalgrading
Intensity of COX-2a
NTb T
1 59 M NBNCc 6 1 II WDd 1 12 60 M HBV 2 1 IVa WD, MD 1 2, 1, 113 58 M HBV 8 1 II WD 1 114 52 F NBNC 1.5 1 I WD 2 15 73 F NBNC 3 1 II WD 2 16 41 M Alcohol 8 1 III WD 2 17 41 M HBV 5.5 1 II MD 2 18 51 M HBV 5 1 II MD 1 29 38 M HBV 4 4 III MD 1 1
10 68 M HCV 3.5 4 III MD 1 1111 53 M HBV 1.2 1 I MD 1 212 68 M HBV 8.3 2 II MD 2 213 63 M HBV 4.2 1 II MD 2 214 58 M HBV 4.5 4 III MD 2 215 58 M Not available 3.5 3 II MD 1 216 59 M HBV 1.5 4 II MD 1 1117 48 F HBV 1 1 II MD 1 118 64 F HCV 4.7 1 II MD 1 119 71 M Alcohol 12 1 II MD 2 220 52 M HBV 1.2 4 II MD 1 221 72 F NBNC 5 4 II MD 2 222 46 F HBV 2.5 2 II MD 2 123 52 M Alcohol 8 1 III MD 1 1124 55 M HCV 2.5 4 III MD 1 1125 72 M Alcohol 12 3 II MD, PD 1 2, 1, 1126 66 F HCV 2 1 II PD 1 227 77 M Alcohol 5 1 II PD 1 228 65 M Alcohol 4 5 II PD 1 229 54 M NBNC 4.2 1 II PD 1 130 55 M Not available 3 1 I PD 1 231 58 M HBV 1.5 1 1 PD 1 232 43 M HBV 15 6 III PD 1 233 64 M NBNC 3.7 1 II PD 1 134 58 M HBV 1 HCV 4.8 1 II PD 1 235 45 M HBV 7.5 4 IVb PD 1 236 49 M Not available 16 6 IVb PD 1 2a Intensity of COX-2 staining:11, strongly positive;1, positive;2, negative.b NT, nontumor; T, tumor.c NBNC, non-B, non-C virus.d WD, well-differentiated; MD, moderately differentiated; PD, poorly differentiated.
1411Clinical Cancer Research
Research. on August 19, 2019. © 2001 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
immunoassay kits (Abbott Laboratories, Abbott Park, IL). Se-rum AFP titer was measured by a RIA kit (Abbott Laboratories).
Histological Interpretation (H&E Staining). The dif-ferentiation grade was classified into three types according tothe criteria proposed by the Liver Cancer Study Group of Japan:(a) well-differentiated; (b) moderately differentiated; and (c)poorly differentiated types (26).
Radiological Evaluation of HCC. Radiological featuresfor the TNM staging of HCC before operation, such as size andgrowth types, were evaluated by computed tomography scan.The growth types were evaluated for gross histological inter-pretation according to the method suggested by the Liver CancerStudy Group of Japan: type I, single nodular type; type II, singlenodular with perinodal type; type III, multinodular confluent
type; type IV, multinodular discrete type; type V, massive type;and type VI, diffuse type (26).
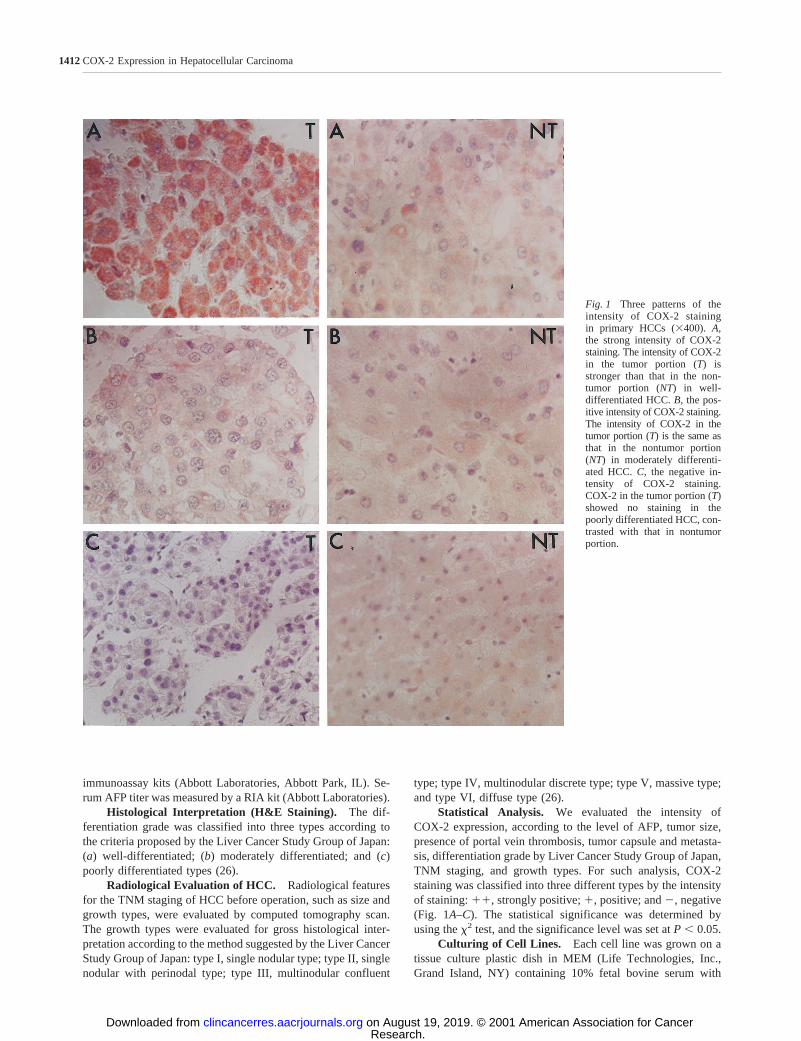
Statistical Analysis. We evaluated the intensity ofCOX-2 expression, according to the level of AFP, tumor size,presence of portal vein thrombosis, tumor capsule and metasta-sis, differentiation grade by Liver Cancer Study Group of Japan,TNM staging, and growth types. For such analysis, COX-2staining was classified into three different types by the intensityof staining:11, strongly positive;1, positive; and2, negative(Fig. 1A–C). The statistical significance was determined byusing thex2 test, and the significance level was set atP , 0.05.
Culturing of Cell Lines. Each cell line was grown on atissue culture plastic dish in MEM (Life Technologies, Inc.,Grand Island, NY) containing 10% fetal bovine serum with
Fig. 1 Three patterns of theintensity of COX-2 stainingin primary HCCs (3400).A,the strong intensity of COX-2staining. The intensity of COX-2in the tumor portion (T) isstronger than that in the non-tumor portion (NT) in well-differentiated HCC.B, the pos-itive intensity of COX-2 staining.The intensity of COX-2 in thetumor portion (T) is the same asthat in the nontumor portion(NT) in moderately differenti-ated HCC.C, the negative in-tensity of COX-2 staining.COX-2 in the tumor portion (T)showed no staining in thepoorly differentiated HCC, con-trasted with that in nontumorportion.
1412COX-2 Expression in Hepatocellular Carcinoma
Research. on August 19, 2019. © 2001 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
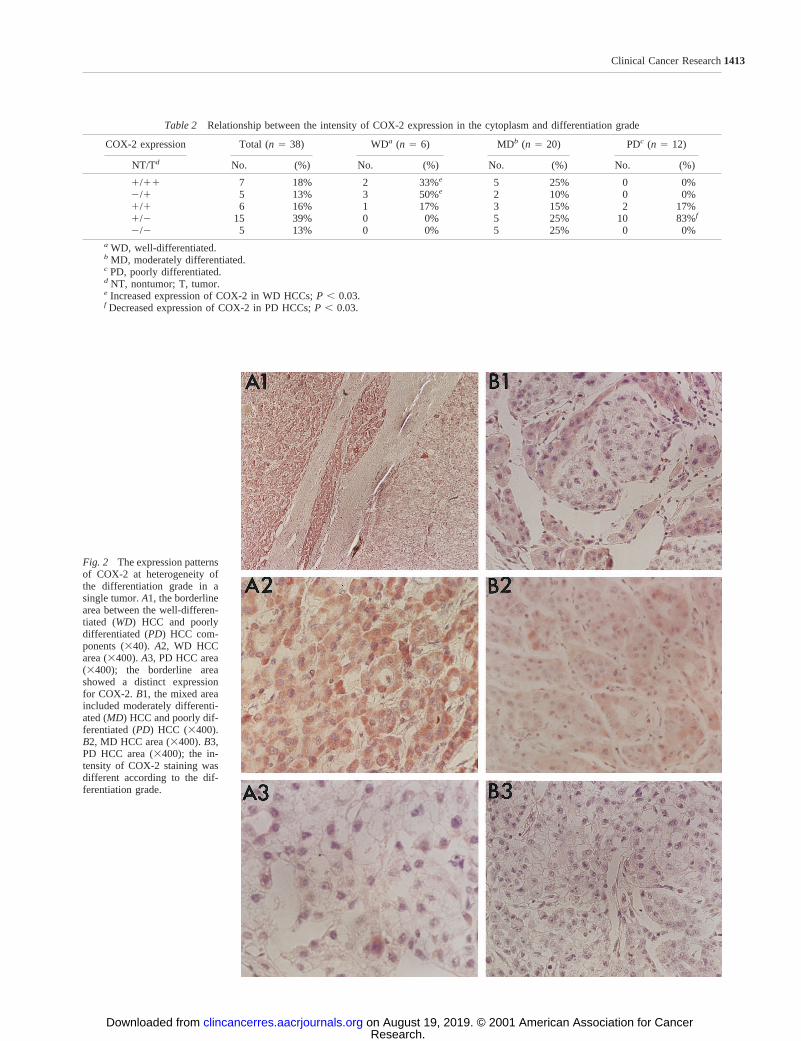
Fig. 2 The expression patternsof COX-2 at heterogeneity ofthe differentiation grade in asingle tumor.A1, the borderlinearea between the well-differen-tiated (WD) HCC and poorlydifferentiated (PD) HCC com-ponents (340).A2, WD HCCarea (3400).A3, PD HCC area(3400); the borderline areashowed a distinct expressionfor COX-2. B1, the mixed areaincluded moderately differenti-ated (MD) HCC and poorly dif-ferentiated (PD) HCC (3400).B2, MD HCC area (3400).B3,PD HCC area (3400); the in-tensity of COX-2 staining wasdifferent according to the dif-ferentiation grade.
Table 2 Relationship between the intensity of COX-2 expression in the cytoplasm and differentiation grade
COX-2 expression Total (n 5 38) WDa (n 5 6) MDb (n 5 20) PDc (n 5 12)
NT/Td No. (%) No. (%) No. (%) No. (%)
1/11 7 18% 2 33%e 5 25% 0 0%2/1 5 13% 3 50%e 2 10% 0 0%1/1 6 16% 1 17% 3 15% 2 17%1/2 15 39% 0 0% 5 25% 10 83%f
2/2 5 13% 0 0% 5 25% 0 0%a WD, well-differentiated.b MD, moderately differentiated.c PD, poorly differentiated.d NT, nontumor; T, tumor.e Increased expression of COX-2 in WD HCCs;P , 0.03.f Decreased expression of COX-2 in PD HCCs;P , 0.03.
1413Clinical Cancer Research
Research. on August 19, 2019. © 2001 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
essential amino acid, 100 units/ml penicillin (Life Technologies,Inc.), and 100mg/ml streptomycin (Life Technologies, Inc.) andwas maintained in a CO2 incubator at 37°C in a humidifiedatmosphere containing 5% CO2 and 95% humidity.
Immunohistochemical Staining for COX-2. For liverspecimens, tissue sections (4mm) mounted on silanized slides(DAKO Japan, Kyoto, Japan) were deparaffinized with xyleneand dehydrated in a graded series of ethanol. After rehydrationin absolute ethanol for 15 s, the slides were heated by micro-wave in 10 mmol/liter citrate buffer (pH 6.0; Zymed Lab Inc.,San Francisco, CA) for 8 min. After washing in a PBS bath for10 min at 4°C, the specimens were preblocked for 10 min in anautoblocker (Research Genetics, Huntsville, AL). Then, theywere incubated overnight with mouse antihuman COX-2 mono-clonal antibody (dilution, 1:100; Transduction Lab Inc., Frank-lin Lakes, NJ) at room temperature. After three washes in PBS,the sections were incubated with biotinylated antimouse IgG(Zymed Lab Inc., San Francisco, CA) for 10 min and then,incubated with a labeled-avidin-biotin method (Zymed Lab Inc.,San Francisco, CA). After washing in PBS, 3-amino-9-ethylcar-bazole (Lab Vision, Fremont, CA) solution was added. Counter-staining was done with hematoxylin (Research Genetics).
Growth Inhibition of Hepatoma Cell Lines by NS-398.Some cell lines were selected to evaluate the inhibitory growtheffect of NS-398 (Cayman Chemicals, Ann Arbor, MI) accord-
ing to the staining patterns of COX-2. The cells were plated at5 3 105 cells/well in six-well plastic dishes with 2 ml of 10%fetal bovine serum-supplemented medium for 24 h. The mediawere changed with serum-free (0.1% BSA) and phenol red-freeones containing various concentrations of NS-398, and cellswere incubated for 5 days with refeeding at day 3.
For concentration-dependent experiments, media contain-ing various concentrations of NS-398 (0, 1, 10, 50, and 100mM)were used (27, 28). For time course experiments, media con-taining 100mM NS-398 were used, and the cells were harvesteddaily for 5 days.
The cells were detached with 800ml of trypsin-EDTA for10 min at 37°C, washed, and resuspended in the medium. Analiquot was combined with an equal volume of trypan blue dye(Life Technologies, Inc.). The surviving cells were countedusing a hemocytometer.
DAPI Staining. DAPI, a DNA-binding fluorescent dye,was used to determine whether the mechanism of growth inhi-bition of these cell lines by NS-398 was apoptosis. After treat-ment with 100mM NS-398 for 5 days, the cells were washedthree times with PBS, fixed in a solution of 3.7% formaldehydefor 10 min, fixed once in 1 ml of methanol, and then stainedwith 4 mg/ml DAPI (Oncor, Gaithersburg, MD) for 10 min (29).The results were determined with visual observation of thenuclear morphology by fluorescence microscopy.
Table 3 Relationship between the intensity of COX-2 expression in the cytoplasm and TNM staging (P. 0.05)
COX-2 expressiona
Stage I (n5 4) Stage II (n5 11) Stage III (n5 20) Stage IVa (n5 1) Stage IVb (n5 2)
No. % No. % No. % No. % No. %
Type 1 2 50% 7 63% 8 40% 1 100% 2 100%Type 2 1 25% 1 10% 4 20%Type 3 1 25% 3 27% 8 40%
a Type 1, weaker; type 2, equal; type 3, stronger staining of COX-2 in tumor tissue compared with that in nontumor tissue.
Table 4 Relationship between the intensity of COX-2 expression in the cytoplasm and growth typesa (P . 0.05)
Single nodular(n 5 22)
Single nodule withperinodular (n5 2)
Multinodularconfluent (n5 3)
Multinodulardiscrete (n5 8) Massive (n5 1) Diffuse (n5 2)
No. % No. % No. % No. % No. % No. %
Type 1 9 41% 1 50% 1 33% 4 50% 1 100% 2 100%Type 2 6 27% 1 33% 1 12%Type 3 7 32% 1 50% 1 33% 3 38%
a Type 1, weaker; type 2, equal; type 3, stronger staining of COX-2 in tumor tissue compared with that in nontumor tissue.
Table 5 Relationship between the intensity of COX-2 expression in the cytoplasm and the level of AFP, size, presence of portal veinthrombosis, tumor capsule, and metastasis (P. 0.05)
COX-2expressiona
AFP Size-grade PVTb Capsule Metastasis
,100(n 5 19)
.100(n 5 17)
,5 cm(n 5 23)
.5 cm(n 5 13)
Yes(n 5 6)
No(n 5 30)
Yes(n 5 24)
No(n 5 12)
Yes(n 5 2) No
Type 1 9 9 10 7 4 14 13 3 2Type 2 4 2 5 2 6 5 3Type 3 6 6 8 4 2 10 6 6a Type 1, weaker; Type 2, equal; Type 3, stronger staining of COX-2 in tumor tissue compared with that in nontumor tissue.b PVT, portal vein thrombosis.
1414COX-2 Expression in Hepatocellular Carcinoma
Research. on August 19, 2019. © 2001 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

RESULTSClinicopathological Features. Thirty-six subjects con-
sisted of 29 males and 7 females; their mean age was 57 yearsof age (range, 38–77 years). Sixteen were HBV surface antigen-positive, four were anti-HCV antibody-positive, one had bothHBV and HCV, six had alcohol-related liver diseases, and sixhad other non-B, non-C liver diseases. Three patients wereundetermined (Table 1).
The mean tumor size was 5.1 cm; 23 patients had HCC#5.0 cm, and 13 patients had HCC$5.0 cm. As determined bymorphological types on the computed tomography scan, therewere 21 of type I, 2 of type II, 2 of type III, 8 of type IV, 1 oftype V, and 2 of type VI (Table 1).
Among the 36 HCC specimens, two cases had hetero-
geneity in a single tumor, showing a mixture of two cellgroups in which the differentiation grades differed: well andmoderately differentiated in one; and moderately and poorlydifferentiated in the other. Therefore, as determined by theLiver Cancer Study Group of Japan, there were 6 well-differentiated, 20 moderately differentiated, and 12 poorlydifferentiated types (Table 1).
COX-2 Expression in Tumor and Nontumor LiverTissues. In 36 nontumor specimens, COX-2 staining was pos-itive in 26 (72%) and negative in 10 (28%). The nontumorspecimens expressed mild intensity of COX-2 staining, becausemost cases had mild to moderate inflammatory activity. In 36HCC specimens, COX-2 staining was positive to various de-grees in 17 samples (47%) and negative in 19 (53%) samples.
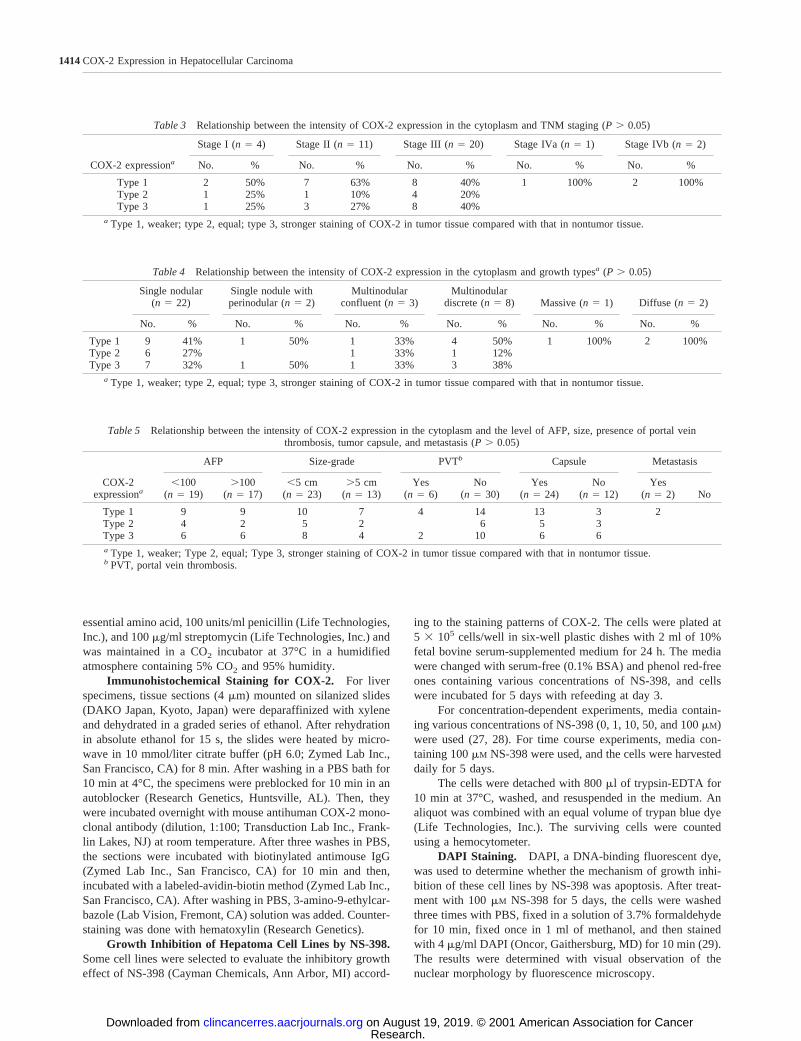
Fig. 3 Immunohistochemicalstaining for COX-2 in hepa-toma cells (3400). COX-2 pro-tein was found in the cytoplasmof hepatoma cells (A,HLE; B,HuH-7; andC, HepG2).D, HT-29, which is a positive control,showed COX-2 expressionstrongly in the perinuclear area.E, SV80, a fibroblast cell line,did not show COX-2 protein inthe cytoplasm.
1415Clinical Cancer Research
Research. on August 19, 2019. © 2001 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Among the 17 COX-2-positive HCC specimens, 5 were stronglypositive (Table 1).
Compared with that of the surrounding nontumor livertissues, five different patterns of COX-2 staining could beidentified in the tumor tissues: NT/T,1/11; NT/T, 2/1;NT/T, 1/1; NT/T, 1/2; NT/T, 2/2 (Table 2). When ana-lyzed according to an individual differentiation grade, all ofthe 6 well-differentiated HCCs were positive, whereas 83%(10 of 12) of the poorly differentiated HCCs were negative.In the 20 moderately differentiated HCCs, 50% were negativein COX-2 expression and 50% were positive (Table 2 andFig. 1).
Among the two HCC specimens that demonstrated heter-ogeneity in differentiation grade in a single tumor, one showedpositive COX-2 expression in the area of well-differentiatedHCC and negative in the area of moderately differentiated HCC.In the other, COX-2 expression was positive in the area ofmoderately differentiated HCC and negative in the area ofpoorly differentiated HCC (Fig. 2).
There was no significant relationship between the intensityof COX-2 expression and the level of AFP, tumor size, presenceof portal vein thrombosis, tumor capsule and metastasis, TNMstaging, and growth types (P. 0.05; Tables 3, 4, and 5).
COX-2 Expression in Hepatoma Cell Lines. In 12 hep-atoma cell lines, except PLC/PRF-5, SNU398, and SNU423,COX-2 protein was expressed in the cytoplasm. Unexpectedly,SV80, a fibroblast cell line, did not show COX-2 protein in thecytoplasm. HT-29, a colon cancer cell line, which is used as apositive control, showed a strong COX-2 expression in theperinuclear area (Fig. 3).
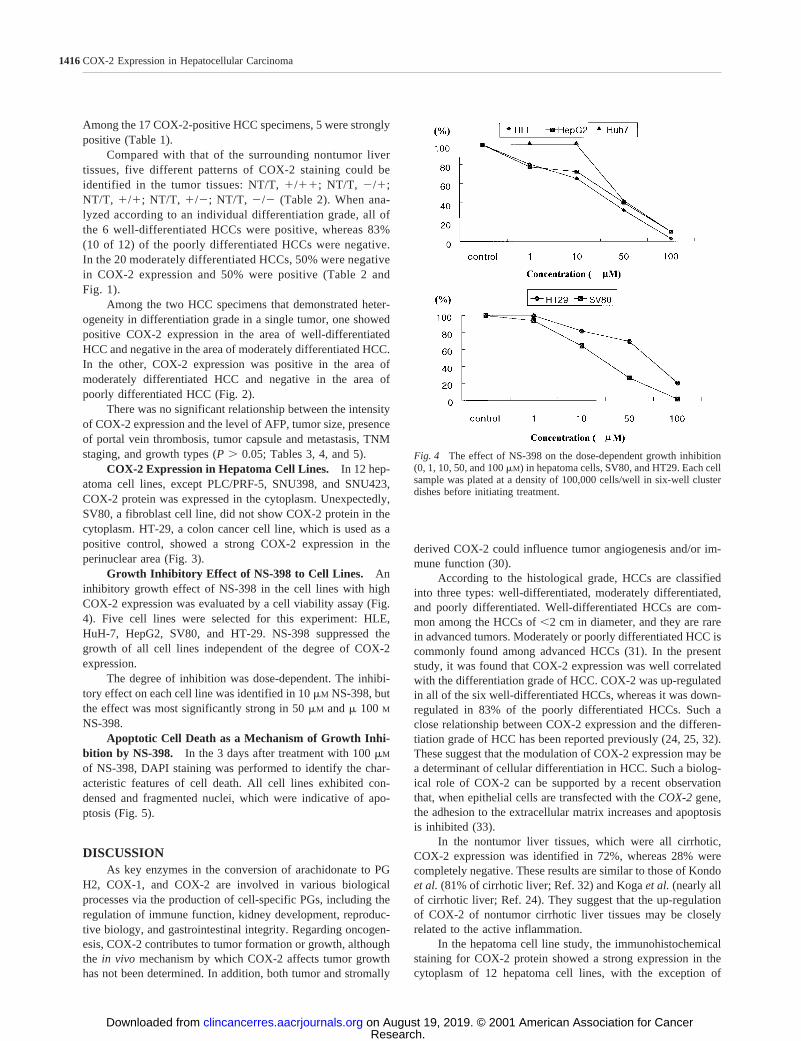
Growth Inhibitory Effect of NS-398 to Cell Lines. Aninhibitory growth effect of NS-398 in the cell lines with highCOX-2 expression was evaluated by a cell viability assay (Fig.4). Five cell lines were selected for this experiment: HLE,HuH-7, HepG2, SV80, and HT-29. NS-398 suppressed thegrowth of all cell lines independent of the degree of COX-2expression.
The degree of inhibition was dose-dependent. The inhibi-tory effect on each cell line was identified in 10mM NS-398, butthe effect was most significantly strong in 50mM andm 100 M
NS-398.Apoptotic Cell Death as a Mechanism of Growth Inhi-
bition by NS-398. In the 3 days after treatment with 100mM
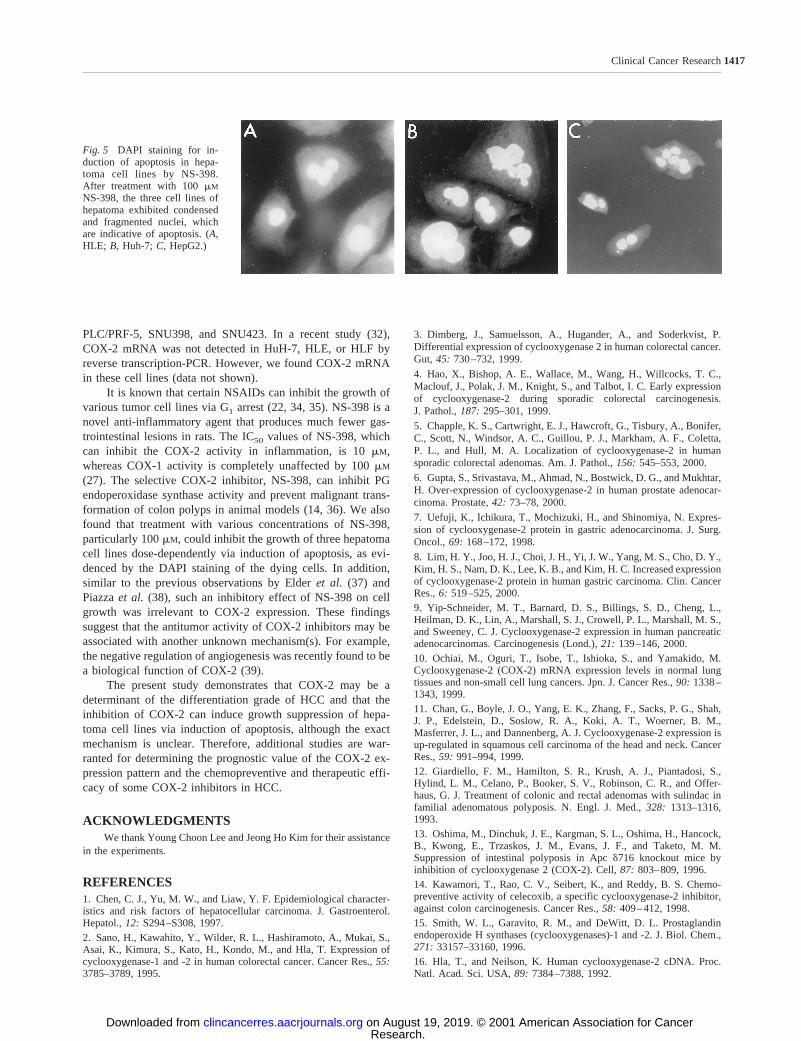
of NS-398, DAPI staining was performed to identify the char-acteristic features of cell death. All cell lines exhibited con-densed and fragmented nuclei, which were indicative of apo-ptosis (Fig. 5).
DISCUSSIONAs key enzymes in the conversion of arachidonate to PG
H2, COX-1, and COX-2 are involved in various biologicalprocesses via the production of cell-specific PGs, including theregulation of immune function, kidney development, reproduc-tive biology, and gastrointestinal integrity. Regarding oncogen-esis, COX-2 contributes to tumor formation or growth, althoughthe in vivo mechanism by which COX-2 affects tumor growthhas not been determined. In addition, both tumor and stromally
derived COX-2 could influence tumor angiogenesis and/or im-mune function (30).
According to the histological grade, HCCs are classifiedinto three types: well-differentiated, moderately differentiated,and poorly differentiated. Well-differentiated HCCs are com-mon among the HCCs of,2 cm in diameter, and they are rarein advanced tumors. Moderately or poorly differentiated HCC iscommonly found among advanced HCCs (31). In the presentstudy, it was found that COX-2 expression was well correlatedwith the differentiation grade of HCC. COX-2 was up-regulatedin all of the six well-differentiated HCCs, whereas it was down-regulated in 83% of the poorly differentiated HCCs. Such aclose relationship between COX-2 expression and the differen-tiation grade of HCC has been reported previously (24, 25, 32).These suggest that the modulation of COX-2 expression may bea determinant of cellular differentiation in HCC. Such a biolog-ical role of COX-2 can be supported by a recent observationthat, when epithelial cells are transfected with theCOX-2gene,the adhesion to the extracellular matrix increases and apoptosisis inhibited (33).
In the nontumor liver tissues, which were all cirrhotic,COX-2 expression was identified in 72%, whereas 28% werecompletely negative. These results are similar to those of Kondoet al. (81% of cirrhotic liver; Ref. 32) and Kogaet al. (nearly allof cirrhotic liver; Ref. 24). They suggest that the up-regulationof COX-2 of nontumor cirrhotic liver tissues may be closelyrelated to the active inflammation.
In the hepatoma cell line study, the immunohistochemicalstaining for COX-2 protein showed a strong expression in thecytoplasm of 12 hepatoma cell lines, with the exception of
Fig. 4 The effect of NS-398 on the dose-dependent growth inhibition(0, 1, 10, 50, and 100mM) in hepatoma cells, SV80, and HT29. Each cellsample was plated at a density of 100,000 cells/well in six-well clusterdishes before initiating treatment.
1416COX-2 Expression in Hepatocellular Carcinoma
Research. on August 19, 2019. © 2001 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
PLC/PRF-5, SNU398, and SNU423. In a recent study (32),COX-2 mRNA was not detected in HuH-7, HLE, or HLF byreverse transcription-PCR. However, we found COX-2 mRNAin these cell lines (data not shown).
It is known that certain NSAIDs can inhibit the growth ofvarious tumor cell lines via G1 arrest (22, 34, 35). NS-398 is anovel anti-inflammatory agent that produces much fewer gas-trointestinal lesions in rats. The IC50 values of NS-398, whichcan inhibit the COX-2 activity in inflammation, is 10mM,whereas COX-1 activity is completely unaffected by 100mM
(27). The selective COX-2 inhibitor, NS-398, can inhibit PGendoperoxidase synthase activity and prevent malignant trans-formation of colon polyps in animal models (14, 36). We alsofound that treatment with various concentrations of NS-398,particularly 100mM, could inhibit the growth of three hepatomacell lines dose-dependently via induction of apoptosis, as evi-denced by the DAPI staining of the dying cells. In addition,similar to the previous observations by Elderet al. (37) andPiazzaet al. (38), such an inhibitory effect of NS-398 on cellgrowth was irrelevant to COX-2 expression. These findingssuggest that the antitumor activity of COX-2 inhibitors may beassociated with another unknown mechanism(s). For example,the negative regulation of angiogenesis was recently found to bea biological function of COX-2 (39).
The present study demonstrates that COX-2 may be adeterminant of the differentiation grade of HCC and that theinhibition of COX-2 can induce growth suppression of hepa-toma cell lines via induction of apoptosis, although the exactmechanism is unclear. Therefore, additional studies are war-ranted for determining the prognostic value of the COX-2 ex-pression pattern and the chemopreventive and therapeutic effi-cacy of some COX-2 inhibitors in HCC.
ACKNOWLEDGMENTSWe thank Young Choon Lee and Jeong Ho Kim for their assistance
in the experiments.
REFERENCES1. Chen, C. J., Yu, M. W., and Liaw, Y. F. Epidemiological character-istics and risk factors of hepatocellular carcinoma. J. Gastroenterol.Hepatol.,12: S294–S308, 1997.2. Sano, H., Kawahito, Y., Wilder, R. L., Hashiramoto, A., Mukai, S.,Asai, K., Kimura, S., Kato, H., Kondo, M., and Hla, T. Expression ofcyclooxygenase-1 and -2 in human colorectal cancer. Cancer Res.,55:3785–3789, 1995.
3. Dimberg, J., Samuelsson, A., Hugander, A., and Soderkvist, P.Differential expression of cyclooxygenase 2 in human colorectal cancer.Gut, 45: 730–732, 1999.
4. Hao, X., Bishop, A. E., Wallace, M., Wang, H., Willcocks, T. C.,Maclouf, J., Polak, J. M., Knight, S., and Talbot, I. C. Early expressionof cyclooxygenase-2 during sporadic colorectal carcinogenesis.J. Pathol.,187: 295–301, 1999.
5. Chapple, K. S., Cartwright, E. J., Hawcroft, G., Tisbury, A., Bonifer,C., Scott, N., Windsor, A. C., Guillou, P. J., Markham, A. F., Coletta,P. L., and Hull, M. A. Localization of cyclooxygenase-2 in humansporadic colorectal adenomas. Am. J. Pathol.,156: 545–553, 2000.
6. Gupta, S., Srivastava, M., Ahmad, N., Bostwick, D. G., and Mukhtar,H. Over-expression of cyclooxygenase-2 in human prostate adenocar-cinoma. Prostate,42: 73–78, 2000.
7. Uefuji, K., Ichikura, T., Mochizuki, H., and Shinomiya, N. Expres-sion of cyclooxygenase-2 protein in gastric adenocarcinoma. J. Surg.Oncol.,69: 168–172, 1998.
8. Lim, H. Y., Joo, H. J., Choi, J. H., Yi, J. W., Yang, M. S., Cho, D. Y.,Kim, H. S., Nam, D. K., Lee, K. B., and Kim, H. C. Increased expressionof cyclooxygenase-2 protein in human gastric carcinoma. Clin. CancerRes.,6: 519–525, 2000.9. Yip-Schneider, M. T., Barnard, D. S., Billings, S. D., Cheng, L.,Heilman, D. K., Lin, A., Marshall, S. J., Crowell, P. L., Marshall, M. S.,and Sweeney, C. J. Cyclooxygenase-2 expression in human pancreaticadenocarcinomas. Carcinogenesis (Lond.),21: 139–146, 2000.10. Ochiai, M., Oguri, T., Isobe, T., Ishioka, S., and Yamakido, M.Cyclooxygenase-2 (COX-2) mRNA expression levels in normal lungtissues and non-small cell lung cancers. Jpn. J. Cancer Res.,90: 1338–1343, 1999.11. Chan, G., Boyle, J. O., Yang, E. K., Zhang, F., Sacks, P. G., Shah,J. P., Edelstein, D., Soslow, R. A., Koki, A. T., Woerner, B. M.,Masferrer, J. L., and Dannenberg, A. J. Cyclooxygenase-2 expression isup-regulated in squamous cell carcinoma of the head and neck. CancerRes.,59: 991–994, 1999.12. Giardiello, F. M., Hamilton, S. R., Krush, A. J., Piantadosi, S.,Hylind, L. M., Celano, P., Booker, S. V., Robinson, C. R., and Offer-haus, G. J. Treatment of colonic and rectal adenomas with sulindac infamilial adenomatous polyposis. N. Engl. J. Med.,328: 1313–1316,1993.13. Oshima, M., Dinchuk, J. E., Kargman, S. L., Oshima, H., Hancock,B., Kwong, E., Trzaskos, J. M., Evans, J. F., and Taketo, M. M.Suppression of intestinal polyposis in Apcd716 knockout mice byinhibition of cyclooxygenase 2 (COX-2). Cell,87: 803–809, 1996.14. Kawamori, T., Rao, C. V., Seibert, K., and Reddy, B. S. Chemo-preventive activity of celecoxib, a specific cyclooxygenase-2 inhibitor,against colon carcinogenesis. Cancer Res.,58: 409–412, 1998.15. Smith, W. L., Garavito, R. M., and DeWitt, D. L. Prostaglandinendoperoxide H synthases (cyclooxygenases)-1 and -2. J. Biol. Chem.,271: 33157–33160, 1996.16. Hla, T., and Neilson, K. Human cyclooxygenase-2 cDNA. Proc.Natl. Acad. Sci. USA,89: 7384–7388, 1992.
Fig. 5 DAPI staining for in-duction of apoptosis in hepa-toma cell lines by NS-398.After treatment with 100mM
NS-398, the three cell lines ofhepatoma exhibited condensedand fragmented nuclei, whichare indicative of apoptosis. (A,HLE; B, Huh-7;C, HepG2.)
1417Clinical Cancer Research
Research. on August 19, 2019. © 2001 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

17. Jones, D. A., Carlton, D. P., McIntyre, T. M., Zimmerman, G. A.,and Prescott, S. M. Molecular cloning of human prostaglandin endoper-oxide synthase type II and demonstration of expression in response tocytokines. J. Biol. Chem.,268: 9049–9054, 1993.18. Marnett, L. J. Aspirin and the potential role of prostaglandins incolon cancer. Cancer Res.,52: 5575–5589, 1992.19. Eberhart, C. E., and Dubois, R. N. Eicosanoids and the gastroin-testinal tract. Gastroenterology,109: 285–301, 1995.20. Pairet, M., and Engelhardt, G. Distinct isoforms (COX-1 andCOX-2) of cyclooxygenase: possible physiological and therapeutic im-plications. Fundam. Clin. Pharmacol.,10: 1–17, 1996.21. Reddy, B. S., Rao, C. V., and Seibert, K. Evaluation of cyclooxy-genase-2 inhibitor for potential chemopreventive properties in coloncarcinogenesis. Cancer Res.,56: 4566–4569, 1996.22. Qiao, L., Hanif, R., Sphicas, E., Shiff, S. J., and Rigas, B. Effect ofaspirin on induction of apoptosis in HT-29 human colon adenocarci-noma cells. Biochem. Pharmacol.,55: 53–64, 1998.23. Crew T. E., Elder, D. J., Paraskeva, C. A cyclooxygenase-2(COX-2) selective non-steroidal anti-inflammatory drug enhances thegrowth inhibitory effect of butyrate in colorectal carcinoma cells ex-pressing COX-2 protein: regulation of COX-2 by butyrate. Carcinogen-esis (Lond.),21: 69–77, 2000.24. Koga, H., Sakisaka, S., Ohishi, M., Kawaguchi, T., Taniguchi, E.,Sasatomi, K., Harada, M., Kusaba, T., Tanaka, M., Kimura, R., Na-kashima, Y., Nakashima, O., Kojiro, M., Kurohiji, T., and Sata, M.Expression of cyclooxygenase-2 in human hepatocellular carcinoma:relevance to tumor dedifferentiation. Hepatology,29: 688–696, 1999.25. Shiota, G., Okubo, M., Noumi, T., Noguchi, N., Oyama, K., Ta-kano, Y., Yashima, K., Kishimoto, Y., and Kawasaki, H. Cyclooxyge-nase-2 expression in hepatocellular carcinoma. Hepatogastroenterology,46: 407–412, 1999.26. Liver Cancer Study Group of Japan. Primary liver cancer in Japan.Clinicopathologic features and results of surgical treatment. Ann. Surg.,211: 277–287, 1990.27. Futaki, N., Yoshikawa, K., Hamasaka, Y., Arai, I., Higuchi, S.,Iizuka, H., and Otomo, S. NS-398, a novel non-steroidal anti-inflam-matory drug with potent analgesic and antipyretic effects, which causesminimal stomach lesions. Gen. Pharmacol.,24: 105–110, 1993.28. Liu, X. H., Yao, S., Kirschenbaum, A., and Levine, A. C. NS398,a selective cyclooxygenase-2 inhibitor, induces apoptosis and down-regulates bcl-2 expression in LNCaP cells. Cancer Res.,58: 4245–4249,1998.29. Shono, M., Shimizu, I., Omoya, T., Hiasa, A., Honda, H., Tomita,Y., and Ito, S. Differences in ethidium bromide and 49-6-diamidino-2-
phenylindole staining profiles with regard to DNA fragmentation duringapoptosis. Biochem. Mol. Biol. Int.,46: 1055–1061, 1998.
30. Williams, C. S., Mann, M., and DuBois, R. N. The role of cyclooxy-genases in inflammation, cancer, and development. Oncogene,18:7908–7916, 1999.
31. Okuda, K., and Tabor, E. Liver cancer.In: Z. Schaff, and P. Nagy(eds.), Pathology Techniques and Grading Systems in the Diagnosis ofHCC. pp. 111–133. Edinburgh: Churchill Livingstone Publishers, B. V.,1997.
32. Kondo, M., Yamamoto, H., Nagano, H., Okami, J., Ito, Y., Shimizu,J., Eguchi, H., Miyamoto, A., Dono, K., Umeshita, K., Matsuura, N.,Wakasa, K., Nakamori, S., Sakon, M., and Monden, M. Increasedexpression of COX-2 in nontumor liver tissue is associated with shorterdisease-free survival in patients with hepatocellular carcinoma. Clin.Cancer Res.,5: 4005–4012, 1999.
33. Tsujii, M., and DuBois, R. N. Alterations in cellular adhesion andapoptosis in epithelial cells overexpressing prostaglandin endoperoxidesynthase 2. Cell,83: 493–501, 1995.
34. Bayer, B. M., Kruth, H. S., Vaughan, M., and Beaven, M. A. Arrestof cultured cells in the G1 phase of the cell cycle by indomethacin.J. Pharmacol. Exp. Ther.,210: 106–111, 1979.
35. Hanif, R., Pittas, A., Feng, Y., Koutsos, M. I., Qiao, L., Staiano-Coico, L., Shiff, S. I., and Rigas, B. Effects of nonsteroidal anti-inflammatory drugs on proliferation and on induction of apoptosis incolon cancer cells by a prostaglandin-independent pathway. Biochem.Pharmacol.,52: 237–245, 1996.
36. Sheng, H., Shao, J., Kirkland, S. C., Isakson, P., Coffey, R. J.,Morrow, J., Beauchamp, R. D., and DuBois, R. N. Inhibition of humancolon cancer cell growth by selective inhibition of cyclooxygenase-2.J. Clin. Investig.,99: 2254–2259, 1997.
37. Elder, D. J., Halton, D. E., Hague, A., and Paraskeva, C. Inductionof apoptotic cell death in human colorectal carcinoma cell lines by acyclooxygenase-2 (COX-2)-selective nonsteroidal anti-inflammatorydrug: independence from COX-2 protein expression. Clin. Cancer Res.,3: 1679–1683, 1997.
38. Piazza, G. A., Rahm, A. K., Finn, T. S., Fryer, B. H., Li, H., Stou-men, A. L., Pamukcu, R., and Ahnen, D. J. Apoptosis primarily ac-counts for the growth-inhibitory properties of sulindac metabolites andinvolves a mechanism that is independent of cyclooxygenase inhibition,cell cycle arrest, and p53 induction. Cancer Res.,57: 2452–2459, 1997.
39. Tsujii, M., Kawano, S., Tsuji, S., Sawaoka, H., Hori, M., andDuBois, R. N. Cyclooxygenase regulates angiogenesis induced by coloncancer cells. Cell,93: 705–716, 1998.
1418COX-2 Expression in Hepatocellular Carcinoma
Research. on August 19, 2019. © 2001 American Association for Cancerclincancerres.aacrjournals.org Downloaded from

2001;7:1410-1418. Clin Cancer Res Si Hyun Bae, Eun Sun Jung, Young Min Park, et al. COX-2 Inhibitor, NS-398Carcinoma and Growth Inhibition of Hepatoma Cell Lines by a Expression of Cyclooxygenase-2 (COX-2) in Hepatocellular
Updated version
http://clincancerres.aacrjournals.org/content/7/5/1410
Access the most recent version of this article at:
Cited articles
http://clincancerres.aacrjournals.org/content/7/5/1410.full#ref-list-1
This article cites 38 articles, 15 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/7/5/1410.full#related-urls
This article has been cited by 17 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/7/5/1410To request permission to re-use all or part of this article, use this link
Research. on August 19, 2019. © 2001 American Association for Cancerclincancerres.aacrjournals.org Downloaded from