effects of procaine on pharmaco-mechanical coupling mechanisms
TRANSCRIPT

356
Effects of Procaine on Pharmaco-MechanicalCoupling Mechanisms Activated by Acetylcholine
in Smooth Muscle Cells of PorcineCoronary Artery
Hikaru Ueno, Kotoko Sumimoto, Toshihiko Hashimoto, Masato Hirata,
and Hirosi Kuriyama
The action of procaine on pharmaco-mechanical coupling activated by application of acetylcholine(ACh) was investigated using collagenase-treated dispersed intact and skinned smooth muscle cellsand intact muscle tissues of the porcine coronary artery. ACh reduced stored 45Ca2+, and this actionwas prevented by procaine in intact dispersed cells. The maximum reduction in the level of stored 45Cainduced by caffeine (25 mM) or inositol 1,4,5-trisphosphate (InsP,; 3 /JLM) was also prevented byprocaine in the skinned muscle cells in the presence or absence of ATP. However, inhibitions of thelatter required higher concentrations of procaine than the former. Release by 10 fiM ACh of Ca2+
from its store site in the presence or absence of extracellular Ca2+ was also inhibited by procaine andwas detected using the quin2 fluorescence method. In these smooth muscle tissues, ACh (above 10 nM)reduced the amount of phosphatidylinositol 4,5-bisphosphate (PI-P2) and dose dependently increasedthe amount of phosphatidic acid. Procaine inhibited the hydrolysis of PI-P2 activated by ACh, thusreducing the amount of InsP3 and the release of Ca2+ from the store site. It is concluded that procainehas multiple actions on the porcine coronary artery, and one of the actions related with pharmaco-mechanical coupling appears through inhibition of hydrolysis of PI-P2 induced by ACh. (CirculationResearch 1987;60:356-366)
I n the porcine coronary artery, Ito et al12 found that10 (JLM acetylcholine (ACh) neither modifies themembrane potential nor the membrane resistance
of the smooth muscle cells, but produces a larger con-traction than that evoked by 128 mM K+ . Further-more, in Ca2+-free solution containing 2 mM EGTA[ethylene glycol bis (/3-aminoethyl ether)-N,N,N',N'-tetraacetic acid], ACh produces contraction in this vas-cular tissue, with an amplitude similar to that observedin Ca2+-containing solution (0.86 times the control).This indicates that in smooth muscles of the porcinecoronary artery, ACh releases Ca2+ from the intracel-lular store site (usually sarcoplasmic reticulum),2"4 asdoes caffeine.2 While the source of Ca2+ in the storesite is presumably the same, the mechanisms underly-ing the release of Ca2+ by each agent seem to differbecause in skinned muscle tissues of the porcine coro-nary artery caffeine, but not ACh, releases Ca2+ fromthe store site.2
Suematsu et al5 studied the effects of inositol 1,4,5-trisphosphate (InsP3) on the release of Ca2+ from intra-cellular store sites in saponin-treated dispersed smooth
From the Department of Pharmacology, Faculty of Medicine,Department of Biochemistry (MH), and the Faculty of Dentistry,Kyushu University, Fukuoka, Japan.
Supported in part by a grant from the Ministry of Education ofJapan.
Address for reprints: Dr. Hikaru Ueno, Department of Pharma-cology, Faculty of Medicine, Kyushu University, Fukuoka 812,Japan.
Received April 1, 1986; accepted November 7, 1986.
muscle cells of the porcine coronary artery. Theyfound that this agent releases Ca2+ from nonmitochon-drial store sites and that the mechanism differs fromthat induced by a calcium-ionophore, A23187. AChincreases the production of InsP3 and diacylglyerol invarious tissues.6"" In the porcine coronary and rabbitmesenteric arteries, ACh and norepinephrine, respec-tively, increased the amounts of InsP3 and phospha-tidic acid (PA) and reduced the amount of phosphati-dylinositol 4,5-bisphosphate (PI-P2). InsP3 releasedCa2+ accumulated in saponin-treated dispersed singlecells and also produced contraction in saponin-treatedskinned vascular smooth muscle strips.51213 There-fore, InsP3 synthesized by activation of the muscarinicreceptor or a,-adenoceptor may provoke pharmaco-mechanical coupling.14
Procaine, a local anesthetic, possessed multiple ac-tions on smooth muscle of the porcine coronary artery,i.e., this agent depolarized and reduced ionic conduc-tance mainly by inhibition of the K+ conductance ofthe smooth muscle membrane. Furthermore, in thissmooth muscle tissue, outward current pulse evokedan action potential only in the presence of procaine ortetraethylammonium.1 Procaine also inhibited the caf-feine-induced release of Ca2+ from the store site. Thisrelease by caffeine involves mechanisms similar to therelease mechanism in cardiac muscles.315"17
Furthermore, Itoh et al2 also reported that in Ca2+-free solution containing 2 mM EGTA, 1 mM procaineblocks contraction in the porcine coronary artery
by guest on February 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

Ueno et al Procaine in Smooth Muscle Cells 357
evoked by 10 FXM ACh, but 5 mM procaine is neces-sary to block contraction evoked by 20 mM caffeine,i.e., procaine inhibits to a greater extent the ACh-induced contraction than the caffeine-induced con-traction.
The present study attempted to clarify the action ofprocaine on mechanisms involved in the pharmaco-mechanical coupling provoked by ACh in smoothmuscle cells of the porcine coronary artery. For thispurpose, intact and skinned dispersed smooth musclecells were prepared by application of collagenase formeasurements of effluxes and releases of Ca2+. Sincethe mutual relation between synthesis of InsP3 andhydrolysis of PI-P2 has been clarified,13 the amount ofInsP3 hydrolyzed by activation of phosphodiesterase(phospholipase C) through activation of the muscarinicreceptor was estimated from the reduction in theamount of PI-P2 and the increase in the amount of PAin intact smooth muscle tissues.
Materials and MethodsPreparation of Single Smooth Muscle Cells
Suspensions of isolated smooth muscle cells wereprepared by enzymic digestion of the porcine coronaryartery as previously described.418 Large branches ofthe right and left coronary arteries of the adult pig(approximately 2-3 mm diameter) were dissected freeof connective tissue. The adventitia was carefullystripped off under a binocular microscope, and theendothelium was rubbed with a cotton ball. The mus-cle layers were cut longitudinally and fixed at a slightlyextended length on a plastic plate in Ca2+-free HEPES(N - 2 - hydroxyethylpiperazine - N' - 2 - ethanesulfonic
acid) solution (EGTA was not present). After equili-bration for 1 hour at 36° C, this solution was replacedby Ca2+-free HEPES solution containing collagenase 1mg/ml (Worthington Biochem. Co., Freehold, N.J.,CLS type III), trypsin inhibitor 0.5 mg/ml (SigmaChemical Co., St. Louis, Mo., type I-S), and bovineserum albumin 2 mg/ml (Sigma, fraction V). Follow-ing digestion for 1 hour at 36° C, loosened musclestrips were chopped into small pieces with a razorblade and digested in a polyethylene tube with thesame solution, under gentle agitation through a wide-bore polyethylene pipette, for 1 hour at 36° C. Thesuspension was filtered through double nylon gauze toseparate single muscle cells from undigested residue.Isolated muscle cells were collected by centrifugationat 30g for 5 minutes and then washed twice with 20volumes of Ca2+-free HEPES solution at about 20° C.The prepared cells were spindle shaped (about 200-300 /xm in length and 5-10 /xm in width) and relaxed,even in the Ca2+-containing solution. About 90-95%of dispersed cells contracted in response to 10 /JLMACh in the Ca2+-containing solution (Figure 1). Itcould not be excluded that the cell suspension wascontaminated with nonmuscle cells such as fibroblasts.However, almost all of the cells were similar in mor-phology under phase-contrast light microscopy, andblood cells and macrophages were not evident. Over95% of cells had the ability to exclude trypan blue.19
Furthermore, after completion of experimental proce-dures described in "Results," more than 90% of thecells still retained resistance to trypan blue inclusion.When the cells were stored at 5° C, the viability as-sessed by trypan blue exclusion was retained for 48
FIGURE 1. Shapes of dispersedsingle smooth muscle cells of theporcine coronary artery. PanelA: Cells suspended in Krebs solu-tion. Panel B: Three minutes aftertreatment with 10 fiM acetylcho-line.
100/jm iOOjum
by guest on February 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

358 Circulation Research Vol 60, No 3, March 1987
hours. However, the cells were used within 4 hours ofisolation.4
45Ca2+ Efflux Experiments With Intact DispersedMuscle Cells
The amount of Ca2+ in the dispersed intact smoothmuscle cells was measured by the filtration method,using 45Ca2+ as described by Ueno.4 The results wereexpressed as nanomoles of Ca2+/2 x 105 cells. Thecontent of 43Ca2+ described in "Results" refers to theexchangeable Ca2+ as estimated from the amount of45Ca2+. The efflux of Ca2+ was estimated from the45Ca2+ remaining in dispersed smooth muscle cells.
The dispersed smooth muscle cells were first incu-bated in physiological salt solution containing 45Ca2+
(2.5-2.7 /xCi/ml) for 60 minutes at 35° C. Sampleswere then transferred to the ice-cold Ca2+-chelatingsolution (without ATP; see "Solutions"). The 45Ca2+
efflux was measured after application of a small vol-ume of concentrated EGTA (final concentration 4mM) to the incubation solution at time zero. After thedesired time, a sample of the cell suspension was treat-ed with caffeine, ACh, or procaine and then filtered,washed, and counted. Briefly, this suspension waspassed through a glass microfiber membrane filter(Whatman GF/C, pore size 1.2 fim), and the filter wasimmediately washed 4 times with 2-3 ml of the coldCa2+-free solution to remove nonspecific 45Ca2+ bind-ing to the membrane filter. The washed filter was thenplaced in a counting vial (Wheaton Scientific, Mill-ville, N.J.) and dried at 80° C for 1 hour. A scintilla-tion cocktail consisting of 4 g 2,5-diphenyloxazole and0.3 g 1,4-bis [2-(4-methyl-5-phenyloxazolyl)] ben-zene in 1 liter toluene was added to the vial, and thesample was counted for radioactivity in a Packard Tri-Carb liquid scintillation counter (Packard InstrumentCo., Downers Grove, 111.). During these procedures,damage to the dispersed cells was minimized, and thefiltered medium contained almost no fragments of da-maged cells.
Preparation of Saponin-Treated Muscle CellsTo study the effects of caffeine, procaine, or InsP3
on intracellular Ca2+ store sites, skinned dispersed sin-gle muscle cells were prepared by treatment with sapo-nin,5.20,21 This treatment leaves intact the membranes ofcytosolic organdies and contractile systems as esti-mated from the caffeine-induced contraction inskinned muscle preparations.31522"24 Saponin-treatedmicrosomes are capable of accumulating Ca2+ into theintracellular storage with no marked change in ATPaseactivity42326(see "Results"). Dispersed smooth musclecells were suspended in 30 ml of a relaxing solution(see "Solutions") used for skinned muscle cells con-taining saponin 15 /ng/ml and 1 mM-EGTA. Afterskinning for 10 minutes at 35° C, the cells were centri-fuged, then washed twice with 20 volumes of freshsolution and suspended in fresh solution at 5° C. Theconcentration of saponin and the skinning time weredetermined in preliminary experiments as the mini-mum required for skinning all cells.4
4SCa2+ Accumulation and45Co2+ ReleaseExperiments With Skinned Dispersed Muscle Cells
The 45Ca2+ release from the store site in dispersedskinned muscle cells was assayed by the dilution meth-od described by Hirata et al27 and Ueno.4 The 45Ca2+
was accumulated in the store site of skinned dispersedmuscle cells suspended in 0.5 ml of the relaxing solu-tion (see "Solutions") at 37° C. After 20 minutes, 0.1ml of the suspension was passed through a glass fiberfilter. The filter was then treated with the Ca2+-chelat-ing solution as described for the Ca2+ efflux experi-ment, and the amount of 45Ca2+ uptake was deter-mined. At the same time, the remaining cellsuspension (0.4 ml) was diluted tenfold with a solutioncontaining KC1 130 mM, Tris-maleate 20 mM (pH6.8), NaN3 5 mM, MgCl2 5 mM, and EGTA 4 mM. Atthe desired time, 1 ml of the mixture containing caf-feine or InsP3 in the Ca2+-chelating solution was fil-tered, and the remaining 45Ca2+ was determined. Toprevent the action of ATP remaining in the solutionthat was required for Ca2+ uptake, 10 mM glucose and4.5 units of hexokinase (Sigma, type II) were added 1minute before dilution (Figure 8).
To measure the accumulation and release of 4SCa2+
from the store site simultaneously in skinned dispersedcells, the filtration method was used in the presence ofATP (relaxing solution; see "Solutions"). After accu-mulation of 45Ca2+ reached a steady level, InsP3 orA23187 alone or together with procaine was applied(Figure 3).
Preparation oflnositol Trisphosphate (InsP3)InsP3 was prepared by incubating human erythro-
cyte ghosts with CaCl2 followed by a Dowex formatecolumn separation and was desalted by elution from aDowex chloride column with 1 M LiCl, followed byremoval of the LiCl with ethanol.28 For estimation ofthe purity of InsP3, cellulose thin layer chromatog-raphy was used. The Rf value was estimated at 0.20,which corresponded with the value obtained by Gradoand Ballou.29
Analysis of lnositol Phospholipids and PhosphatidicAcid in Smooth Muscle Tissue
The contents of inositol phospholipids (PI-P2 andphosphatidylinositol 4-monophosphate [PI-P]) and PAin the coronary artery were measured before and aftertreatment with ACh, according to the procedures de-scribed by Hashimoto et al.13 For this purpose, musclestrips (2-3 mm x 10 mm) were labelled in a phos-phate-free, HEPES-buffered Krebs solution (pH 7.4)containing 40 ^iCi/ml of [32P] Pi (specific radioactiv-ity, 30-40 Ci/mol; Japan Atomic Energy ResearchInstitute) at 37° C for 3 hours. This time was adequatefor isotopic equilibrium in terms of the inositol phos-pholipids because a longer incubation did not yieldhigher radioactivity. The strips were then washed threetimes with the above solution without [32P] Pi and wereincubated with ACh for 10-120 seconds. The reactionwas halted by adding an organic solvent containingchloroform, methanol, and HC1 (100:200:2, v/v), and
by guest on February 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

Ueno et al Procaine in Smooth Muscle Cells 359
the strips were homogenized in a glass homogenizer.Crude lipid extracts in the solvent were chromato-graphed on Silica Gel 60 plates (Merck Sharp &Dohme, West Point, Penn.), according to the methodof Billah and Lapetina,30 and were then autoradio-graphed with Sakura X-ray film for 12-15 hours. Thefractions corresponding to PI-P, PI-P2, and PA on theplate were cut out and counted for radioactivity in aliquid scintillation counter.
Quin2 Loading and Fluorescence MeasurementsThe dispersed smooth muscle cells were loaded with
quin2 in the presence of 15-100 /iM quin2/AM (tetra-acetoxymethyl ester) for 120 minutes at 37° C, as pre-viously described.31"33 Hydrolysis of quin2/AM wasmonitored by the gradual shift in the emission spec-trum from a peak at 430 nm for the ester to a peak at490 nm for the final indicator. The spectral shift wasnearly complete in 120 minutes. The loaded cells werethen washed twice, resuspended in fresh Ca2+-contain-ing HEPES buffer solution, and kept at room tempera-ture. Intracellular quin2 concentration was determinedby comparing the Ca2+-dependent fluorescence ofquin2 loaded cells with the quin2 unloaded cells in thepresence of quin2-free acid in a standard solution.Quin2 concentration and the intensity of fluorescenceshowed a linear relation.33 Assuming that the cell vol-ume was 3,000 firri3,34 quin2 concentrations obtainedby incubating cells with 15-100 /xM quin2/AM for 120minutes were 0.7-3 mM. After quin2 loading, theviability of the muscle cells, assessed by the trypanblue exclusion test and the contractility in response toagonists, was well preserved.33
Before measurement, the cell suspension wasequilibrated in HEPES buffer solution containing 1mM Ca2+ for 30 minutes and then incubated at 37° Cfor 10 minutes.
Fluorescence of quin2 loaded and unloaded cellswas measured at 37° C in a Hitachi 650-40 fluores-cence, spectrophotometer with a thermostated cellholder at a wave length for excitation of 339 nm (slit 4nm) and for emission of 490 nm (slit 10 nm). The cellswere suspended in 2 ml HEPES buffer solution (NaCl140 mM, KC1 4 mM, CaCl2 1 mM, MgCl2 1 mM,K2HPO41 mM, glucose 10 mM, HEPES 20 mM) at aconcentration of 106 cells/ml in a 1-cm square quartzcuvette and continuously stirred.
Following completion of the fluorescence measure-ment, the cells were dialyzed with 0.2% Triton Xsaturated with dye at 1.0 mM to give the maximumfluorescence of the sample (Fmu). Fmin was the fluores-cence obtained after addition of 0.5 mM MnCl2 todisplace the bound Ca2+ and to quench the quin2 emis-sion.35 Mn2+ also quenches the Ca2+-independent flu-orescence. However, this value was under 5% of theCa2+-dependent fluorescence. Therefore, this factorwas omitted from the calculation.
The concentration of intracellular free Ca2+
([Ca2+]i) was calculated from the equation [Ca2+]j =Kd (F - Frain)/(FmM - F),31 where K,,, the effectivedissociation constant for Ca2+ — quin2 is 115 nM,
assuming that intracellular free Mg2+ is about 1 mM.36
The increase rate of fluorescence was expressed as thepercent of total fluorescence (Fmax — F ^ ) . Extracellu-lar dye or dye in leaky cells was estimated by addingMnCl2 (0.5 mM) to an aliquot of suspension of theintact, quin2 loaded cells. The immediate decrease influorescence caused by extracellular dye was 10-15%.There was little variability in the amount of leakagebetween different aliquots of cell suspensions. Thecalculation of [Ca2+]j was corrected by subtracting thefluorescence caused by extracellular dye. Addition ofACh, EGTA, A23187, or procaine did not affect theautofluorescence of the unloaded cells. Triton Xquenched the autofluorescence, but this was negligi-ble. Conformation changes of cells during contractionby ACh had no effect on the quin2-induced fluores-cence, as estimated from measurements of the controlaliquot.
SolutionsAll solutions were prepared with deionized double-
glass-distilled water.The Ca2+-free HEPES solution (for preparation of
single cells) contained (in mM) 140 NaCl, 5 KC1, 2MgCl2, 15 glucose, and 20 HEPES (pH 7.3). Thephysiological salt solution for45Ca2+ equilibration andthe quin2 experiments contained 140 NaCl, 5 KC1, 1MgCl2, 1 CaCl2, 15 glucose, and 20 HEPES (pH 7.3).The mM concentration for the Ca2+-chelating solutionwas 130 NaCl, 5 KC1, 2 MgCl2, 10 EGTA (disodiumsalt), and 20 HEPES (pH 7.3); and for the high [K]o
solution 102.5 KC1, 42.5 NaCl, 1 MgCl2, 1 CaCl2, 15glucose, and 20 HEPES (pH 7.3). All solutions ofintact cells were bubbled.with 95% O2 and 5% CO2.The mM concentration of the solution for the skinneddispersed smooth muscle cells (relaxing solution) was130 KC1, 5 MgCl2, 5 ATP (disodium salt), 20 Tris-maleate, and pH 6.8. For 45Ca2+-uptake experiments,0.3 mM Ca/EGTA buffer and 5 mM NaN3 were added(except for45Ca2+ efflux experiments). CaCl2 was pre-pared from dried CaCO3 and HC1.
DrugsChemicals used in the present experiments were
acetylcholine chloride (ACh-Cl; Sigma Chemical Co.,St. Louis, Mo.), saponin (ICN Pharmaceuticals, Inc.,Cleveland, Ohio), caffeine and sodium azide (WakoPharmaceutical Co., Kyoto, Japan), procaine (DaiichiPharmaceutical Co., Tokyo), A 23187 (free acid; Cal-biochemical, La Jolla, Calif.) and EGTA (DojindoLaboratory, Kumamoto, Japan). 45CaCl2 was obtainedfrom New England Nuclear Co., Boston, Mass.Quin2/AM and quin2 were obtained from DojindoLaboratory, Kumamoto, Japan.
StatisticsThe mean and standard deviation (SD) are given.
Statistical evaluations were made by Student's t test forpaired and unpaired observations.
by guest on February 5, 2018http://circres.ahajournals.org/
Dow
nloaded from
360 Circulation Research Vol 60, No 3, March 1987
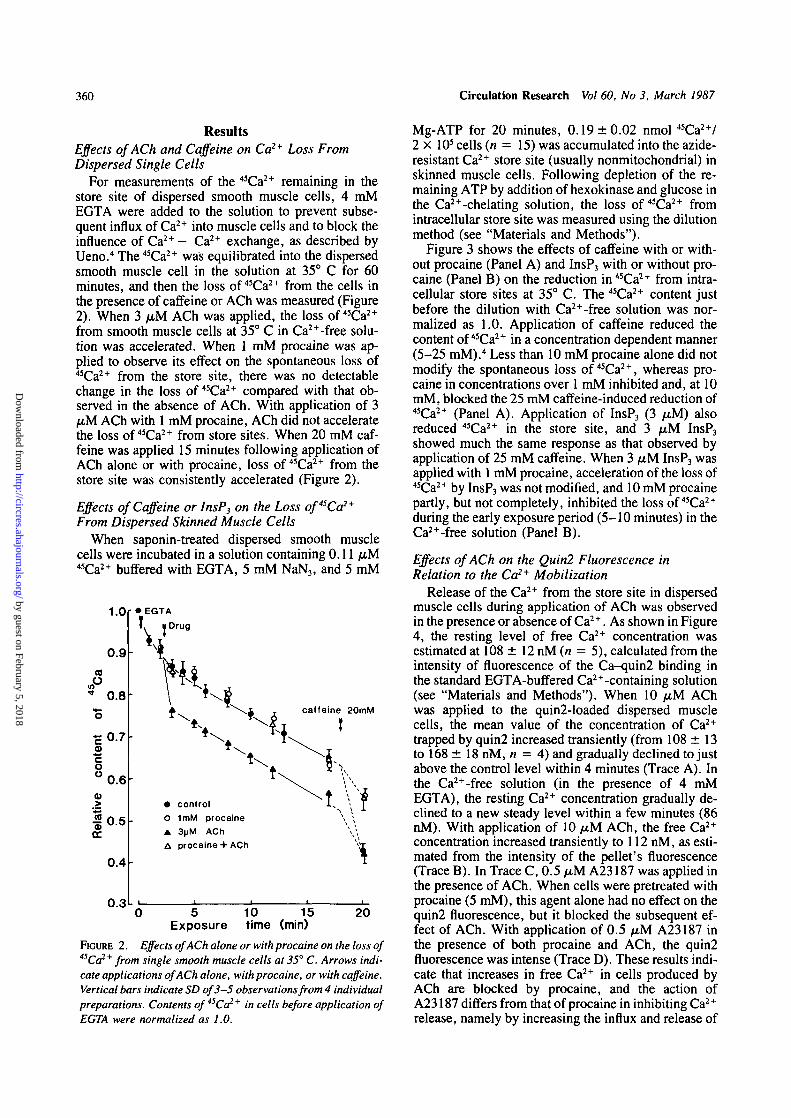
ResultsEffects of ACh and Caffeine on Ca2+ Loss FromDispersed Single Cells
For measurements of the 45Ca2+ remaining in thestore site of dispersed smooth muscle cells, 4 mMEGTA were added to the solution to prevent subse-quent influx of Ca2+ into muscle cells and to block theinfluence of Ca2+ - Ca2+ exchange, as described byUeno.4 The 45Ca2+ was equilibrated into the dispersedsmooth muscle cell in the solution at 35° C for 60minutes, and then the loss of 45Ca2+ from the cells inthe presence of caffeine or ACh was measured (Figure2). When 3 /AM ACh was applied, the loss of 45Ca2+
from smooth muscle cells at 35° C in Ca2+-free solu-tion was accelerated. When 1 mM procaine was ap-plied to observe its effect on the spontaneous loss of45Ca2+ from the store site, there was no detectablechange in the loss of 45Ca2+ compared with that ob-served in the absence of ACh. With application of 3fiM ACh with 1 mM procaine, ACh did not acceleratethe loss of 45Ca2+ from store sites. When 20 mM caf-feine was applied 15 minutes following application ofACh alone or with procaine, loss of 45Ca2+ from thestore site was consistently accelerated (Figure 2).
Effects of Caffeine or InsP3 on the Loss of4SCa2+
From Dispersed Skinned Muscle CellsWhen saponin-treated dispersed smooth muscle
cells were incubated in a solution containing 0.11 /xM45Ca2+ buffered with EGTA, 5 mM NaN3, and 5 mM
1.0
CO
o
0.9
0.8
0.7
0.6
0.5
0.4
0.3
caffeine 20mM
• controlO 1mM procaine• 3)jM AChA procaine + ACh
5Exposure
10time
15(min)
20
FIGURE 2. Effects of ACh alone or with procaine on the loss of45Ca2+ from single smooth muscle cells at 35° C. Arrows indi-cate applications of ACh alone, with procaine, or with caffeine.Vertical bars indicate SD of 3-5 observations from 4 individualpreparations. Contents of 45Ca2+ in cells before application ofEGTA were normalized as 1.0.
Mg-ATP for 20 minutes, 0.19 ±0 .02 nmol 45Ca2+/2 x 105 cells (it = 15) was accumulated into the azide-resistant Ca2+ store site (usually nonmitochondrial) inskinned muscle cells. Following depletion of the re-maining ATP by addition of hexokinase and glucose inthe Ca2+-chelating solution, the loss of 45Ca2+ fromintracellular store site was measured using the dilutionmethod (see "Materials and Methods").
Figure 3 shows the effects of caffeine with or with-out procaine (Panel A) and InsP3 with or without pro-caine (Panel B) on the reduction in 45Ca2+ from intra-cellular store sites at 35° C. The 45Ca2+ content justbefore the dilution with Ca2+-free solution was nor-malized as 1.0. Application of caffeine reduced thecontent of 45Ca2+ in a concentration dependent manner(5-25 mM).4 Less than 10 mM procaine alone did notmodify the spontaneous loss of 45Ca2+, whereas pro-caine in concentrations over 1 mM inhibited and, at 10mM, blocked the 25 mM caffeine-induced reduction of45Ca2+ (Panel A). Application of InsP3 (3 PM) alsoreduced 45Ca2+ in the store site, and 3 juM InsP3
showed much the same response as that observed byapplication of 25 mM caffeine. When 3 /u,M InsP3 wasapplied with 1 mM procaine, acceleration of the loss of45Ca2+ by InsP3 was not modified, and 10 mM procainepartly, but not completely, inhibited the loss of 45Ca2+
during the early exposure period (5-10 minutes) in theCa2+-free solution (Panel B).
Effects of ACh on the Quin2 Fluorescence inRelation to the Ca2+ Mobilization
Release of the Ca2+ from the store site in dispersedmuscle cells during application of ACh was observedin the presence or absence of Ca2+. As shown in Figure4, the resting level of free Ca2+ concentration wasestimated at 108 ± 12 nM (n = 5), calculated from theintensity of fluorescence of the Ca-quin2 binding inthe standard EGTA-buffered Ca2+-containing solution(see "Materials and Methods"). When 10 /u,M AChwas applied to the quin2-loaded dispersed musclecells, the mean value of the concentration of Ca2+
trapped by quin2 increased transiently (from 108 ± 13to 168 ± 18 nM, n = 4) and gradually declined to justabove the control level within 4 minutes (Trace A). Inthe Ca2+-free solution (in the presence of 4 mMEGTA), the resting Ca2+ concentration gradually de-clined to a new steady level within a few minutes (86nM). With application of 10 fxM ACh, the free Ca2+
concentration increased transiently to 112 nM, as esti-mated from the intensity of the pellet's fluorescence(Trace B). In Trace C, 0.5 /i-M A23187 was applied inthe presence of ACh. When cells were pretreated withprocaine (5 mM), this agent alone had no effect on thequin2 fluorescence, but it blocked the subsequent ef-fect of ACh. With application of 0.5 /JM A23187 inthe presence of both procaine and ACh, the quin2fluorescence was intense (Trace D). These results indi-cate that increases in free Ca2+ in cells produced byACh are blocked by procaine, and the action ofA23187 differs from that of procaine in inhibiting Ca2+
release, namely by increasing the influx and release of
by guest on February 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

Ueno et al Procaine in Smooth Muscle Cells 361
0.6
0.5
0.4
Io0,0.3
L
controlcaffeine 10mMcaffeine 25mMprocaine 10mMcaffeine 25mM
+ procaine 10mMcaffeine 25mM
+ procaine 1mM
" I.
B1.0
0.6
0.5
0.4
0.3
0.2
3 5Exposure time
10(min)
15
controlInsRj 3uMlnsP3 3uM+procaine 10mMInsPj 3wM+procaine 1mM
3 5 10Exposure time (min)
15
FIGURE 3. Lossof45Ca2+ fromsaponin-treated skinned musclecells in the presence of caffeinealone or with procaine (Panel A)and InsP3 alone or with procaine(Panel B). After accumulation ofCa2+ in 0.11 yM Ca2+-contain-ing solution (see "Materials andMethods"), loss of 4SCa2+ wasmeasured by the dilution methodin the presence of caffeine aloneor with procaine and InsP3 aloneor with procaine in the absence ofATP. Individual points indicatethe mean value of 3-6 experi-ments. Panel A: Effects of caf-feine alone or with procaine onthe loss ofCa2 + stored in skinnedmuscle cells. The amount of Ca2+stored in cells just before appli-cation of drugs was normalizedas 1.0 and indicated as 0 in thetime scale. Panel B: Effects ofInsP3 alone or with procaine onthe loss ofCa2* stored in skinnedmuscle cells. The experimentalprocedures were the same as de-scribed in A. Dotted lines in Aand B indicate presence of pro-caine.
Ca2+.37 Furthermore, ACh produced only a transientincrease in the intensity of fluorescence observed onsmooth muscle tissues of the ferret portal vein, usingaequorin.38
Effects of ACh on Phosphatidylinositol 4,5-Bisphosphate Breakdown in Smooth Muscle Tissues
Assuming that the first step of ACh-induced Ca2+
release from the store site is the breakdown of PI-P2 byactivation of phosphodiesterase as described by Ber-ridge and Irvine,10 we measured the amounts of PI-P2
and PA. Figure 5 shows the effects of ACh on hydroly-sis of PI-P2 in the lipid fraction extracted from thesmooth muscle tissue. The amount of PI-P2 beforeapplication of ACh was normalized as 100% (the abso-lute value was 38,360 ± 5,826 cpm/mg protein, n =8). Application of 10 jitM ACh rapidly reduced theamount of PI-P2 within 10 seconds (76.5 ± 3.1%, n =5), and the amount gradually reverted to the controllevel in the continued presence of ACh (after 120 sec-onds, the content of PI-P2 was recovered to 92 ± 3 . 2 %of the control, n = 5). This decrease in the amount ofPI-P2 induced by ACh was observed to the same extentin Ca2+-free solution containing EGTA. To determinewhether or not the hydrolysis of PI-P2 by ACh is due tophosphodiesteric cleavage, the amount of PA wasmeasured after application of 10 fiM ACh. Theamount of PA before application of ACh was normal-ized as 100% (the absolute value was 13,280 ± 1,923
cpm/mg protein, n = 7); 10 fiM ACh increased it to252 ± 14% (n = 5) after 120 seconds. The increase inthe amount of PA was delayed over the reduction in theamount of PI-P2. However, the amount of PI-P re-mained the same as that observed before application ofACh up to 120 seconds. Thus, reduction in the amountof PI-P2 may be due to phosphodiesteric hydrolysis forthe synthesis of InsP3 activated by ACh.
Figure 6 shows the dose-response relation betweenACh and PI-P2 hydrolysis in smooth muscle tissues ofthe porcine coronary artery. The minimum concentra-tion of ACh required for PI-P2 hydrolysis was 10 nM,and the maximum effects were observed at 10 /xM,such being comparable to the minimum and maximumamplitudes of the ACh-induced contraction in thistissue.2
The effects of procaine on the PI-P2 breakdown acti-vated by ACh were observed. As shown in Figure 7,when 10 /AM ACh and 1 mM procaine were simulta-neously applied, reductions in the amount of PI-P2
were markedly inhibited after 10 and 30 seconds(77.2 ±3 .4% vs. 92.4 ± 2 . 8 % in the presence of 1mM procaine at 10 seconds and 79.2 ± 2.5% vs.91.2 ± 4 .1% in the presence of 1 mM procaine at 30seconds, n = 5). When 3 mM procaine was appliedtogether with 10 FXM ACh, the amount of PI-P2 wasreduced after 10 or 30 seconds, but the values were notsignificantly different from controls. Application ofACh with 10 mM procaine had no effect on the amount
by guest on February 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

362
A
Circulation Research Vol 60, No 3, March 1987
ACh
\
[C"],
ACh A23187
300
— 200
— 100
i— 200
— 100
— 50
— 300
— 200
— 100
nM
FIGURE 4. Effects of ACh in the presence and ab-
sence of Ca2+ on the quin2-induced fluorescence indispersed smooth muscle cells. Quin2IAM (30 fiM)was loaded as described in "Materials and Meth-ods." Trace A: Effects of 10 fiM ACh on the Carelease from the store site as estimated from the inten-sity of fluorescence trapped by quin2. Trace B: Effectsof 10 fxM ACh in Ca2 + -free solution containing 4 mMEGTA. Trace C: Effects of 10 /iM ACh and subse-quently applied 0.5 pM A23187 on the Ca2+ releasedfrom the store site. A23187 was applied in the pres-ence of ACh. Trace D: Effects of 10 fxM ACh and 0.5liM A23187 on the Co2* from the store site underconditions of treatment with 5 mM procaine. Arrowsin the figure indicate application of various agents,and hereafter, drug is present in the medium through-out an experiment. Scales indicate the concentrationof intracellular free Ca2* calculated from the equa-tion described in "Materials and Methods."
Procaine ACh A23187I— 300
— 200
— 100
of PI-P2 up to 120 seconds. These results indicate thatprocaine inhibits the hydrolysis of PI-P2 induced byACh.
Effects of InsP3 on Ca2+ Store Sites in DispersedSaponin-Treated Skinned Muscle Cells
When saponin-treated skinned muscle cells were in-cubated in solution containing 0.27 fiM Ca2+ (5 mMATP with 5 mM MgCl2) for 20 minutes, Ca2+ accumu-lated at maximum levels in the store sites45 (0.44nmol/2 x 103 cells, n = 8). With application of 1 or 3/iM InsP3 in the presence of 5 mM ATP and 5 mMMgCl2, the amount of stored Ca2+ was reduced dosedependently, as estimated from the Ca2+ remaining incells (Figure 8). However, mere was a gradual de-crease in the amount of Ca2+ released from the storesite, presumably due to reaccumulation of Ca2+ in thepresence of ATP. These results confirmed the observa-tions made by Suematsu et al5 using the same prepara-tion. On the other hand, as expected from the mechani-cal response, 10 ^iM ACh had no effect on Ca2+
release from the store site in skinned muscles. As seenin Figure 8A, A23187 (5 fiM) completely released theATP-dependently accumulated Ca2+ from the storesite, and the residual amount of Ca2+ was similar to the
Ca2+ accumulated into the cell ATP independently.When 10 mM procaine was added together with 3
fiM InsP3, the reduction of 45Ca2+ in stores was partlyinhibited (Figure 8B), i.e., when the amount of Ca2+,accumulated in cells using the same procedure as de-scribed in Figure 8A for 20 minutes, was normalizedas 100%, application of 3 fxM InsP3 reduced theamount of Ca2+ stored in cells to about 60% of thecontrol within 3 minutes, and there was a gradual reac-cumulation of stored Ca2+ within 10 minutes. Withaddition of 10 mM procaine to the 3 fiM InsP3-contain-ing solution, the release of Ca2+ was reduced to halfthat observed in the absence of procaine (about 80% ofCa2+ in the control was stored). The results on theeffects of InsP3 on releases of Ca2+ in the presence orabsence of procaine using the present methods anddilution methods were in agreement. Therefore, pro-caine inhibits the release of Ca2+ from intracellularsites but also inhibits the hydrolysis of PI-P2 activatedby ACh.
DiscussionTo measure the mobilization of Ca2+ induced by
ACh or InsP, in intact and skinned muscle cells of the
by guest on February 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

Ueno et al Procaine in Smooth Muscle Cells 363
%110
100
90Q.
a.
,80
70
60
280
260
240
220
200
180
160
140
120
100
0 10 30 60Time
120sec
FIGURE 5. Effects of ACh on breakdown of phosphatidylinosi-tol 4,5-bisphosphate in muscle strips of the porcine coronaryartery as estimated from the amount of nP labelled in thesubstrates as described in "Materials and Methods." Amountsof phosphatidylinositol bisphospnate (PI-P2), phosphatidylmonophosphate (PI-P), and phosphatidic acid (PA) observedbefore application of ACh were normalized as 100%, (left verti-cal axis for PI-P 2 and PI-P, right axis for PA). Mean ± SD, n= 5. The absolute values ofPl-P2 and PA are indicated in thetext.
porcine coronary artery, dispersed single smooth mus-cle cells prepared by treatment with collagenase wereused. In these cells, the physiological functions re-quired to measure the agonist action, ion flux, andCa2+ mobilization in cells were well preserved, and theresults obtained agreed well with the extent of Ca2+
mobilization as estimated from the contraction evokedin intact and skinned muscle tissues.2"4
Procaine inhibited the release of Ca2+ activated byACh, but not by A23187, in dispersed cells as estimat-ed from the fluorescence of Ca-quin2. However, theamount of free Ca2+ released by 10 fiM ACh as esti-mated from the intensities of the fluorescence of Ca-quin2 binding in the present experiment was too low(<0.2 /u,M Ca2+) in comparison to that estimated fromthe minimum concentration of Ca2+ required to gener-ate contraction in skinned muscle tissues (about 0.1/uM C a ) . In this muscle tissue, 10 jiiM ACh pro-duced the maximum contraction of much the sameamplitude as that seen with 10 /xM Ca2+-induced con-traction in skinned muscle tissue.2 Presumably, buffer-ing actions of quin2 may2+ prevent measurement of theabsolute value of free Ca in dispersed cells. Howev-er, comparable intensities of fluorescence enabled us
110
JCM
50 - * » -
10 9 10 8 10 7 10 6 10 5
ACh (M)FIGURE 6. Dose-response relationship of the ACh-inducedbreakdown of phosphatidylinositol 4,5-bisphosphate (PI-P2).The amount ofPI-P2 was measured 30 seconds after applicationof various concentrations of ACh. The amount of Pl-P2 mea-sured in the absence of ACh was normalized as 100%. Mean ±SD, n = 5.
to estimate the release of Ca2+ from cellular storageby ACh in Ca2+-free solution and also actions ofprocaine.3339
The pharmaco-mechanical coupling process14 ob-served in the response of vascular tissue to severalagonists appears with no detectable change in themembrane properties.1214'40^2 Thus, activation of re-
%100
90
80
70
60
• 10JUM ACh
O ACh + imM procaineA ACh + 3mM procainex ACh + 10mM procaine
0 10 30 60Time
120 sec
FIGURE 7. Effects of ACh alone or with procaine on the pro-duction of inositol 1,4,5-trisphosphate (lnsP3) as estimatedfrom the reduction in the amount of32P incorporated in phos-phatidyl inositol 4,5-bisphosphate (PI-P2) in smooth musclestrips of the porcine coronary artery. The amount of PI-P2
before application of ACh was normalized as 100%. Individualpoints indicate the mean ± SD, n = 5.
by guest on February 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

364 Circulation Research Vol 60, No 3, March 1987
x 5UM A23187o ATP-indepondent
Ca uptake
0 5<°> <20) Time
10mm
B
1.0
0.9
0.8
0.7
0.6
0.5
(0)0
(20)
3uM lnsP3
imM procaineInsPj + SmM procainelnsP3+ 10mM procaine
5Time
10
FIGURE 8. Effects of inositol.1,4,5-trisphosphate (lnsP3) aloneor with procaine on reductionin the amount of calcium storedin saponin-treated dispersedskinned muscle cells in the pres-ence of ATP and MgCl2. Whenthe cells were incubated in 0.27fiM Ca2+ for 20 minutes, theamount of stored Ca2 * reacheda steady value of 0.44nmol/2 X 10s cells (n = 5), andthis value was normalized as100%. Panel A: Effects of ACh(10 yM), InsP3 (1 and 3 fiM),andA23187 (5 yM) on the Ca2*stored in dispersed skinned mus-cle cells. As an indication of theATP-independent Ca2* accumu-lation, Ca2* was accumulatedinto the cell in the absence ofATP. A23187 completely re-leased the ATP-dependent accu-mulated Ca*. ACh had no effecton the Ca * release in dispersedskinned muscles. Panel B: Effectsof lnsP3 with and without pro-caine on the Ca * release fromthe stored site in dispersedskinned muscles. Experimentalprocedures were the same as de-scribed in A. Concentrations ofdrugs are indicated in the figure.
ceptors may increase the influx of Ca2+ with no detect-able change in the membrane properties. However, inthe porcine coronary artery, ACh produced contractionin the absence of extracellular Ca2+ with little reduc-tion in the amplitude.2 Therefore, receptor-operatedCa2+ release may be the main source of Ca2+ in theproduction of the ACh-induced contraction rather thanreceptor-activated Ca2+ influx.
Reduction in the amount of PI-P2 activated by AChaccompanied increases in the synthesis of PA in aproportional manner. This means that activity of phos-phodiesterase on PI-P2 is preserved in the present ex-perimental procedures, and InsP3 would be synthe-sized by activation of PI-P2 phosphodiesterase. Theseobservations confirmed our previous observationsmade on vascular tissues in which the reduction in theamount of PI-P2 was accompanied by increases in theamount of InsP3.
12 Norepinephrine had much the sameeffect on the amounts of PI-P2 and InsP3 observed withthe rabbit mesenteric artery.13 In the present experi-ments, intact smooth muscle tissue was used to mea-sure the amounts of PI-P2 and PA following applicationof ACh. Preliminary results obtained using intact dis-persed smooth muscle cells showed much the sameresults as obtained using intact smooth muscle tissues.
It was apparent in the porcine coronary artery that
release of Ca2+ from the store site by ACh in dispersedintact muscle cells and by InsP3 in dispersed skinnedmuscle cells has a causal relation. Suematsu et al5
reported that the Km value for InsP3 release of Ca2+
from the store site is 0.7 [iM measured from dispersedskinned smooth muscle cells in this tissue. The resultsobtained in Figures 3B and 8B for the InsP3-inducedCa2+ release from the store site as estimated from theamount of 45Ca2+ remaining in skinned dispersed mus-cle cells differed, presumably due to the absence (Fig-ure 3B) or presence (Figure 8B) of ATP in the solutionbecause ATP possessed an ability to accelerate thereuptake of released Ca2+ into the store site. An InsP3-induced Ca2+ release in visceral smooth muscle tissueshas also been reported by Somlyo et al43 and Yama-moto and van Breemen.44
Procaine inhibits the K+-conductance of the mem-brane, depolarizes the membrane,1 and inhibits theCa2+-induced Ca+ release mechanism in contractiletissues.215 This agent (above 1 mM) inhibited the con-tractions evoked by action potentials, electrical de-polarization, excess K+ , caffeine, and agonists in thepresence of Ca2+ and the contractions evoked by caf-feine and agonists in Ca2+-free solution.12 Further-more, procaine also inhibited the caffeine- and InsP3-induced contractions in skinned muscle tissues of the
by guest on February 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

Ueno et al Procaine in Smooth Muscle Cells 365
rabbit mesenteric artery.13 However, the mechanismunderlying the release of Ca2+ from the store site in-duced by A23187 may differ from that induced bycaffeine or InsP3. A23187 increased the influx of Ca2+
and released Ca2+ from the store site in the presence ofprocaine, as estimated from the quin2 fluorescencemethod (Figure 3) and also released more Ca2+ fromstore sites than caffeine or InsP3 (Figure 7). Itoh et al37
reported that lower concentrations of A23187 releaseCa2+ from the storage site only once. Following reap-plication of Ca2+, additional application of A23187 nolonger released Ca2+ from the store site, presumablybecause the ability to accumulate Ca2+ is lost. On theother hand, InsP3 repeatedly produced contractionsfollowing application of Ca2+.13 This means thatmechanisms of release of Ca2+ induced by InsP3 andA23187 differ, and procaine may act on the InsP3- orcaffeine-induced Ca2+ release but not on the A23187-induced Ca2+ release.
Procaine prevented the hydrolysis of PI-P2. Howev-er, it is not certain whether procaine inhibits phospho-diesterase, the ACh-receptor, or some other unknownprocess. In concentrations over 1 mM, procaine inhib-ited the potassium permeability of the membrane,leading to a depolarization.1 In the porcine coronaryartery, synthesis of InsP3 by phosphodiesterase in thepresence of ACh required the presence of guanosine5'-triphosphate (GTP).12 If actions of procaine are re-lated to Ca2+ mobilization in the sarcolemmal mem-brane, it may be that steps of the ACh-receptor-phos-phodiesterase coupling through binding with theGTP-dependent subunit10 are prevented via inhibitionof the membrane fluidity by procaine or unknownmechanisms, thereby reducing the InsP3 synthesis.Procaine also inhibited the norepinephrine-inducedcontraction in the rabbit mesenteric artery in Ca2+-freesolution.4445 Presumably, the agonist-induced contrac-tion may have a causal relation to production of InsP3.Although InsP3 and caffeine released Ca2+ from thesame store site,13 the underlying mechanisms for re-lease of Ca2+ from the store site may be differentbecause caffeine releases more Ca2+ than InsP3, but theconcentration of procaine required to inhibit release ofCa2+ by InsP3 was higher than by caffeine.
It is proposed that the action of ACh on pharmaco-mechanical coupling in smooth muscle cells of theporcine coronary artery is as follows: ACh activatesPI-P2 phosphodiesterase after binding with the musca-rinic receptor, which produces InsP3. This productmay play a role as the first step of the pharmaco-mechanical coupling process since it releases Ca2+
from the store site. Procaine possesses multiple actionson vascular smooth muscle tissues, and the inhibitoryaction of procaine on the ACh-induced contraction inthe presence or absence of Ca2+ may be due to inhibi-tion of the hydrolysis of PI-P2 and in part to inhibitionof the InsP3-induced Ca2+ release from the store site.
AcknowledgmentWe thank Dr. K. Creed for reading the manuscript.
References1. Ito Y, Kitamura K, Kuriyama H: Effects of acetylcholine and
catecholamines on the smooth muscle cells of the porcine coro-nary artery. J Physiol (Lond) 1979;294:595-611
2. Itoh T, Kajiwara M, Kitamura K, Kuriyama H: Roles of storedcalcium on the mechanical response evoked in smooth musclecells of the porcine coronary artery. J Physiol (Lond)1982;322:107-125
3. Itoh T, Kuriyama H, Suzuki H: Excitation-contraction cou-pling in smooth muscle cells of the guinea-pig mesenteric ar-tery. J Physiol (Lond) 1981;321:513-535
4. Ueno H: Calcium mobilization in enzymatically isolated singleintact and skinned muscle cells of the porcine coronary artery.J Physiol (Lond) 1985;363:103-117
5. Suematsu E, Hirata M, Kuriyama H: Inositol 1,4,5-trisphos-phate releases Ca2+ from intracellular store sites in skinnedsingle cells of porcine coronary artery. Biochem Biophys ResCommun 1984;120:481-485
6. Hokin MR, Hokin LE: Enzyme secretion and the incorporationof P32 into phospholipides of pancreas slices. J Biol Chem1953;203:967-977
7. Michell RH: Inositol phospholipids and cell surface receptorfunction. Biochem Biophys Ada 1975;415:81-147
8. Abdel-Latif AA, Akhtar RA, Hawthorne JN: Acetylcholineincreases the breakdown of triphosphoinositide of rabbit irismuscle prelabelled with (32P) phosphate. Biochem J 1977;162:61-73
9. Berridge MJ: Inositol trisphosphate and diacylglycerol as sec-ond messengers. Biochem J 1984;220:345-360
10. Berridge MJ, Irvine RF: Inositol trisphosphate, a novel secondmessenger in cellular signal transduction. Nature 1984;312:315-321
11. Nishizuka Y: The role of protein kinase C in cell surface signaltransduction and tumour promotion. Nature 1984;308:693-698
12. Sasaguri T, Hirata M, Kuriyama H: Dependence on Ca2+ ofthe activities of phosphatidyl inositol 4,5-bisphosphate phos-phodiesterase and inositol 1,4,5-trisphosphate phosphatase insmooth muscles of the porcine coronary artery. Biochem J1985;231:497-503
13. Hashimoto T, Hirata M, Itoh T, Kanmura Y, Kuriyama H:Inositol 1,4,5-trisphosphate activates pharmaco-mechanicalcoupling in smooth muscle of the rabbit mesenteric artery. JPhysiol (Lond) 1986;370:605-618
14. Somlyo AP, Somlyo AV: Vascular smooth muscle. I. Normalstructure, pathology, biochemistry, and biophysics. Pharma-col Rev 1968;20:197-272
15. Endo M: Calcium release from the sarcoplasmic reticulum.Physiol Rev 1977;57:71-108
16. Endo M, Yagi S, lino M: Tension-pCa relation and sarcoplas-mic reticulum responses in chemically skinned smooth musclefibers. FedProc 1982;41:2245-2250
17. Itoh T, Ueno H, Kuriyama H: Calcium-induced calcium re-lease mechanism in vascular smooth muscles: Assessmentsbased on contractions evoked in intact and saponin-treatedskinned muscles. Experientia 1985;41:989-996
18. Hirata M, Itoh T, Kuriyama H: Effects of external cations oncalcium efflux from single cells of the guinea-pig taenia coliand porcine coronary artery. J Physiol (Lond) 1980;310:321-336
19. Dow J, Harding NG, Powell T: Isolated cardiac myocytes I.Preparation of adult myocytes and their homology with theintact tissue. Cardiovasc Res 1981 ;15:483—514
20. Seeman P: Transient holes in the erythrocyte membrane duringhypotonic hemolysis and stable holes in the membrane afterlysis by saponin and lysolecithin. J Cell Biol 1967;32:55-70
21. Ohtsuki I, Manzi RM, Palade GE, Jamieson JD: Entry ofmacromolecular tracers into cells fixed with low concentra-tions of aldehydes. Biol Cell 1978;31:119-126
22. Endo M, KitazawaT, Yagi S, lino M, Kakuta Y: Some proper-ties of chemically skinned smooth muscle fibers, in Casteels R,Godfraind T, Ruegg JC (eds): Excitation-Contraction Cou-pling in Smooth Muscle. Amsterdam, Elsevier/North-Holland,1977, pp 199-209
by guest on February 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

366 Circulation Research Vol 60, No 3, March 1987
23. Saida K, Nonomura Y: Characteristics of Ca2+ and Mg2+-induced tension development in chemically skinned smoothmuscle fibers. J Gen Physiol 1978;72:1-14
24. lino M: Tension responses of chemically skinned fibre bundlesof the guinea-pig taenia caeci under varied ionic environments.J Physiol (Lond) 1981;320:449-467
25. Martonosi A: Sarcoplasmic reticulum. V. The structure of sar-coplasmic reticulum membranes. Biochem Biophys Acta1968;150:694-704
26. Suematsu E, Hirata M, Kuriyama H: Effects of cAMP- andcGMP-dependent protein kinase, and calmodulin on Ca2+ up-take by highly purified sarcolemmal vesicles of vascularsmooth muscle. Biochem Biophys Acta 1984;773:83-90
27. Hirata M, Suematsu E, Hashimoto T, Hamachi T, Koga T:Release of Ca2+ from a non-mitochondrial store site in perito-neal macrophages treated with saponin. Biochem J 1984;223:229-236
28. Downes CD, Mussat MC, Michell PH: The inositol trisphos-phate phosphoesterase of the human erythrocyte membrane.Biochem J 1982;203:169-177
29. Grado C, Ballou CE: Myo-inositol phosphates obtained byalkaline hydrolysis of beef brain phosphoinositide. J BiolChem 1961;236:54-60
30. Billah MM, Lapetina EG: Degradation of phosphatidyl inosi-tol-4,5-bisphosphate is insensitive to Ca2+ mobilization instimulated platelets. Biochem Biophys Res Commun 1982;109:217-222
31. Tsien RY, Pozzan P, Pink TJ: Calcium homeostasis in intactlymphocytes: Cytoplasmic free calcium monitored with a new,intracellularly trapped fluorescent indicator. J Cell Biol1982;94:325-334
32. Hirata M, Hashimoto T, Hamachi T, Koga T: Changes ofintracellular free Ca2+ in macrophages following N-formylchemotactic peptide stimulation: Direct measurement by theloading of quin 2. J Biochem 1984;96:9-16
33. Sumimoto K, Kuriyama H: Mobilization of free Ca2+ mea-sured during contraction-relaxation cycles in smooth musclecells of the porcine coronary artery using quin2. Pflugers Arch1986;406:173-180
34. Gabella G: Structure of smooth muscles, in Bulbring E, Brad-ing AF, Jones AW, Tomita T (eds): Smooth Muscle: An As-sessment of Current Knowledge. London, Arnold, 1981,pp 1-46
35. Hesketh TR, Smith GA, Moore JP, Taylor MV, Metcalfe JC:Free cytoplasmic calcium concentration and the mitogenicstimulation of lymphocyte. J Biol Chem 1983;258:4876-4882
36. Baylor SM, Chandler WK, Marshall MW: Optical measure-ments of intracellular pH and magnesium in frog skeletal mus-cle fibres. J Physiol (Lond) 1982;331:105-137
37. Itoh T, Kanmura Y, Kuriyama H: A23187 increases calciumpermeability of store sites more than of surface membranes inthe rabbit mesenteric artery. J Physiol (Lond) 1985;359:467-484
38. Morgan JP, Morgan KG: Stimulus-specific patterns of intracel-lular calcium levels in smooth muscles. Nature 1984;288:585-587
39. DeFeo TT, Morgan KG: A comparison of two different indica-tors: quine 2 and aequorin in isolated single cells and intactstrips of ferret portal vein. Pflugers Arch 1986;406:427-429
40. Su C, Bevan JA, Ursillo RC: Electrical quiescence of pulmo-nary artery smooth muscle during sympathomimetic stimula-tion. CircRes 1964;15:20-27
41. Droogmans G, Raeymaekers L, Casteels R: Electro- and phar-maco-mechanical coupling in the smooth muscle cells of therabbit ear artery. J Gen Physiol 1977;70:129-148
42. Casteels R, Kitamura K, Kuriyama H, Suzuki H: The mem-brane properties of the smooth muscle cells of the rabbit mainpulmonary artery. J Physiol (Lond) 1977;271:41—61
43. Somlyo AV, Bond M, Somlyo AP, Scarpa A: Inositol trisphos-phate-induced calcium release and contraction in vascularsmooth muscle. Proc Natl Acad Sci USA 1985;82:5341-5345
44. Yamamoto H, van Breemen C: Inositol-1,4,5-trisphosphaterelease calcium from skinned cultured smooth muscle cells.Biochem Biophys Res Commun 1985; 130:270-274
45. Itoh T, Kuriyama H, Ueno H: Mechanisms of the nitroglycer-ine-induced vasodilation in vascular smooth muscles of therabbit and pig. J Physiol (Lond) 1983;343:233-252
46. Itoh T, Kanmura Y, Kuriyarha H, Sasaguri T: Nitroglycerine-and isoprenaline-induced vasodilation: assessment from theaction of cyclic nucleotides. BrJ Pharmacol 1985,84:393-406
KEY WORDS • porcine coronary artery • acetycholine• inositol trisphosphate • procaine • pharmaco-mechanicalcoupling
by guest on February 5, 2018http://circres.ahajournals.org/
Dow
nloaded from

H Ueno, K Sumimoto, T Hashimoto, M Hirata and H Kuriyamaacetylcholine in smooth muscle cells of porcine coronary artery.
Effects of procaine on pharmaco-mechanical coupling mechanisms activated by
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 1987 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.60.3.3561987;60:356-366Circ Res.
http://circres.ahajournals.org/content/60/3/356World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Research Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on February 5, 2018http://circres.ahajournals.org/
Dow
nloaded from