
CENTRO DE INVESTIGACIONES MARINAS (CIMA)
CONSELLERÍA DO MAR
Memoria 2017 m
m

2
Foto portada: "Scanning electron micrograph of a mature cercaria showing surface ultrastructure and body details." M. Ruiz1 et al (2012) Epidemiological survey of digenean trematodes affecting razor clams, Ensis arcuatus, from Galicia (NW Spain). Bull. Eur. Ass. Fish Pathol., 32(1) 2012, 3 1Centro de Investigacións Mariñas (CIMA). Consellería do Mar. Xunta de Galicia Ganadora del concurso 2011-2015 de la EAFP (European Association of Fish Pathologists) Bulletin Picture.

CIMA Memoria 2017 3
CENTRO DE
INVESTIGACIONES MARINAS (CIMA)
CONSELLERÍA DO MAR
Memoria 2017
m
m
m

4

CIMA Memoria 2017 5
1 Presentación
EL CENTRO
2 Estructura y funciones
3 Personal
4 Servicios
INVESTIGACIÓN
5 Acuicultura
- Exploring the biological and socio-economic potential of new/emerging candidate fish
species for the expansión of the European aquaculture industry.
- Inducción a la vitelogénesis, maduración ovocitaria, ovulación y espermiogénesis en
cherna (Polyprion americanus).
- Contribución genética para la recuperación de los bancos naturales de coquina (Donax
spp.) en Galicia.
- Inducción de la fijación y metamorfosis en almeja fina, babosa y japonesa.
- Procedimientos de cultivo intensivo de especies de interés marisquero.
6 Patología
- Estudio epidemiológico de la almeja rubia (Venerupis rhomboides) en Galicia afectada
por colonias procariotas intracelulares (CPI). Identificación de las CPI. Seguimiento de la
estructura de tallas de poblaciones de almeja rubia de 4 bancos de la ría de Vigo.
- Estudio de la eliminación de virus entéricos patógenos en ostra mediante procesos de
reinstalación y depuración.
- Poblaciones bacterianas en criadero de bivalvos. Procesos de patogénesis.
- Epidemiología de la enfermedad causada por Marteilia cochillia en las poblaciones de
berberecho Cerastoderma edule de Galicia.
- Determinación de la posible resistencia a la marteiliosis de una cohorte de berberecho
Cerastoderma edule detectada en el banco de O Ariño (Vilanova de Arousa).
- Evaluación del incremento de supervivencia en el engorde de ostra rizada Crassostrea
gigas por el uso de semilla descendiente de supervivientes a la exposición prolongada al
virus herpes de ostreidos tipo 1. Identificación de marcadores moleculares de resistencia
para su aplicación en un programa de selección de estirpes resistentes al virus.
- Co-operation for restoring cockle shellfisheries and its ecosystem-services in the Atlantic
Area (AA).
7 Procesos oceanográficos costeros
- Mejora analítica de la detección de toxinas en moluscos bivalvos para maximizar el flujo
de trabajo en sistemas de control.
8 Recursos marinos
- Evaluación y seguimiento de las Reservas Marinas de Interés Pesquero de Galicia.
- Determinación de las causas de la disminución de los bancos de coquina.
Condicionantes naturales y antropogénicos.
- Engorde en batea de semilla de berberecho producida en criadero. Mejora de la
metodología de cultivo.
- Estudio de la presencia de Rapana venosa en la ría de Arousa, expansión de Stramonita
hemastoma en la costa lucense y litoral de Galicia y Callinectes sapidus, en base a la
evolución de su distribución actual, potencial reproductivo, crecimiento y capacidad
alimentaria. Análisis de riesgos.

6
DIVULGACIÓN
9 Artículos en revistas SCI
10 Artículos en revistas no SCI
11 Libros
12 Ponencias y paneles
13 Edición de publicaciones
ASESORAMIENTO
14 Informes
15 Reuniones
16 Otros
16.1. Asesoramiento. Otros
16.2. Revisión de artículos en publicaciones científicas
FORMACIÓN
17 Tesis doctorales
17.1. Dirección/tutorización de tesis
17.2. Participación en tribunales de tesis
18 Becarios FP 2º grado
19 Cursos
19.1. Cursos impartidos
19.2. Asistencia a cursos
20 Estancias
20.1. Estudiantes en prácticas
20.2. TFM. Máster Interuniversitario Gallego de Acuicultura
CUENTAS
21 Gastos
22 Fuentes de financiación

CIMA Memoria 2017 7
1 PRESENTACIÓN
El Centro de Investigacións Mariñas (CIMA) depende de la Consellería do Mar, está constituido por el Centro de
Investigacións Mariñas de Corón, en Vilanova de Arousa (Pontevedra) y el Centro de Cultivos Mariños de Ribadeo
(Lugo). La función básica de estos centros es el desarrollo de investigaciones encaminadas a conseguir una gestión
racional y eficiente de los recursos marinos renovables en el ámbito geográfico de competencia de la Administración
autonómica de Galicia.
El CIMA actúa como organismo asesor de la Administración pesquera gallega en las cuestiones de su competencia
que demandan los diferentes sectores productivos a través de las direcciones generales y oficinas territoriales de la
Consellería do Mar. También, mediante bolsas, se ocupa de la formación de nuevos investigadores (con tutorías y
dirección de tesis de doctoramiento) y de técnicos de laboratorio. La difusión de la investigación se realiza a través de
la asistencia y participación en reuniones científicas de diverso carácter (congresos, seminarios, foros, workshops, etc.)
y en numerosas publicaciones científicas de diferentes ámbitos.
Durante 2017 el Centro de Investigacións Mariñas (CIMA) continuó su labor de investigación científica y tecnológica en
temas marinos, orientada principalmente a la acuicultura de moluscos y peces, patología de moluscos bivalvos,
bioacumulación y eliminación de toxinas fitoplanctónicas y a la dinámica de poblaciones de invertebrados marinos con
interés comercial. Un trabajo cuyos resultados se muestran resumidos en la presente memoria anual de actividades
(número 19 de la serie iniciada en 1999).

8

CIMA Memoria 2017 9
EL CENTRO
CENTRO DE INVESTIGACIÓNS MARIÑAS
Dirección postal:
Pedras de Corón s/n
Apartado de Correos 13
36620 Vilanova de Arousa
Pontevedra
Teléfonos:
Teléfono: +34 886 20 63 64
Fax: +34 886 20 63 72
Correo electrónico:
CENTRO DE CULTIVOS MARIÑOS
Dirección postal:
Muelle de Porcillán s/n
27700 Ribadeo
Lugo
Teléfonos:
Teléfono: +34 982 88 94 03
Fax: +34 982 88 94 19
Correo electrónico:

10
2 ESTRUCTURA Y FUNCIONES
La actividad científica del CIMA se reparte en cuatro áreas:
Área de Acuicultura
Su objetivo es desarrollar y mejorar los procedimientos de cultivo de las especies
marinas consideradas de interés comercial. El ámbito de investigación abarca dos
grandes líneas: cultivo de moluscos y cultivo de peces.
Área de Patología
El objetivo es estudiar las alteraciones patológicas que afectan a los moluscos bivalvos
con interés comercial, para establecer estrategias eficaces de lucha que permitan
minimizar los efectos patogénicos.
Área de Procesos Oceanográficos Costeros
El objetivo del área es el conocimiento de los aspectos de la oceanografía relacionados
con la producción de los recursos pesqueros y marisqueros de Galicia. Las líneas de
investigación se relacionan principalmente con los episodios de toxicidad originados por
el fitoplancton.
Área de Recursos Marinos
El objetivo es conocer la biología, ecología y dinámica de población de las especies
marinas con interés comercial para mejorar la gestión sostenible de los recursos
marinos vivos.

CIMA Memoria 2017 11
Director: D. Gabín Sánchez, Carlos (desde 22/10/2017)
Gerente: D. Villanueva Domínguez, Rafael
Centro de Investigacións Mariñas de Vilanova de Arousa
Investigadores/as
Dr. Blanco Pérez, Juan Carlos
Dra. Cao Hermida, Asunción
asunció[email protected]
Dra. Carballal Durán, Mª Jesús
D. de Coo Martín, Alberto
D. Cuiñas Olmedo, Pedro
D. Febrero Mayor, Fernando
Dr. Fuentes González, José Miguel
D. García Fernández, Antonio
Dr. Iglesias Estepa, David
Dña. Linares Cuerpo, Fátima
Dra. López Gómez, Mª del Carmen
Dr. Montes Pérez, Jaime
3 PERSONAL

12
Dña. Pérez Acosta, Carmen
Dr. Villalba García, Antonio
Analistas de laboratorio
Dña. Fernández Besada, Mercedes
D. Giráldez Rivero, Ramón
Dña. Solla Vicente, Mª del Carmen
(contrato de sustitución desde 23/10//2017)
Dña. González Fontela, Ana Isabel
Dña. Gregorio Chenlo, Mª Victoria
Dña. Mariño Cadarso, Mª Carmen
Dña. Martín Sánchez, Helena
Dña. Martínez Verde, Gemma
Dña. Méndez Pérez, Belén
Auxiliares de laboratorio
Dña. Brianes Beiras, Mª José
Dña. Campaña Ferro, Emilia
Dña. Cores González, Mª José
Dña. Camino Saco, Mª de los Angeles
(contrato de sustitución desde 10/07/2017)
Dña. Meléndez Ramos, Isabel
Dña. Rúa Santervas, Patricia Margarita
(contrato de sustitución desde 05/06/2017)
Dña. Penas Pampín, Elena
Contratados por proyectos
D. Fernández Abuín, Isidro
Dña. Pazos Sieira, Gema
Becarios/as
Dña. Acuña López, Iria
Dña. Alonso Costas, Juana Beatriz
D. Méndez Montenegro, José Carlos
Dña. Riomayor Cernadas, Alicia
Dña. Ulla Refojo, Laura
Dña.Fernández González, Jenifer
D. Grande Ocampo, Oscar
Dña. Pena Thomas, Guadalupe
Dña. Viturro García, Raquel
D. Araújo Sandín, Alexis
Dña. Fuentes Villar, Noelia
Encargado de mantenimiento
D. López Ares, Hermes
Jefe de negociado administrativo
Dña. Fernández Sanmartin, Inmaculada, C.
Sistemas de información
D. Carballal Fortes, Félix Manuel
Biblioteca
Dña. Álvarez Francisco, Mª Esther
Auxiliares administrativos
D. Lago Torrado, José
D. Martínez Paz, Angel Rubén
Subalternos-ordenanzas
D. Cubiella Granda, Rubén
D. Martínez Crespo, José Manuel
Limpiador
D. García Gamallo, Luis

CIMA Memoria 2017 13
Centro de Cultivos Mariños. Ribadeo
Responsable de planta
Dra. Martínez Patiño, Dorotea
Investigadoras
Dña. Fernández Álvarez, Aurora
Dra. Nóvoa Vázquez, Susana
Dra. Ojea Martínez, Justa
Analista de laboratorio
Dña. Bouzamayor Yánez, Mª Victoria
Auxiliares de laboratorio
Dña. Cotarelo Jardón, Mª José
D. Páez Pérez, Francisco Javier
Dña. Ruíz Misioné, Mercedes
Becarios/as
Dña. Castilla del Río, Sonia
D. Matesanz Salgado, Diego (hasta 16/05/18)
D. Granjel Piñeiro, Eduardo (desde 01/07/17)
Contratados/as por proyectos
D. González Cid, Emilio Jesús
Dña. Couso da Costa, Myriam Séfora
Oficial 1º de mantenimiento
D. Fernández Escudero, Antonio
Oficial administrativa
Dña. Neira Páez, Concepción
Dña. Vilar Varela, Adelaida
(contrato de sustitución desde 25/09/2017)
Administrativa
Dña. Cruzado Estévez, Ana
Subalterno/a
Limpiadora
Dña. Fernández Goás, Carmen
14
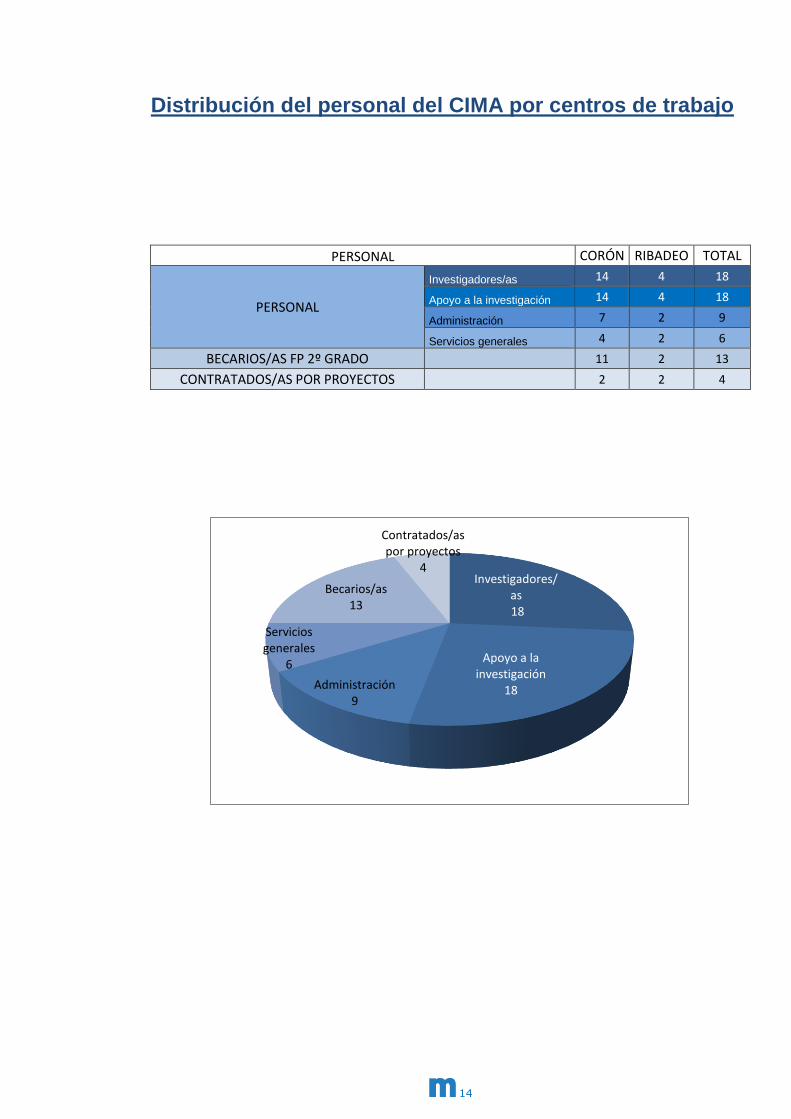
Distribución del personal del CIMA por centros de trabajo
PERSONAL CORÓN RIBADEO TOTAL
PERSONAL
Investigadores/as 14 4 18
Apoyo a la investigación 14 4 18
Administración 7 2 9
Servicios generales 4 2 6
BECARIOS/AS FP 2º GRADO 11 2 13
CONTRATADOS/AS POR PROYECTOS
2 2 4
Investigadores/as 18
Apoyo a la investigación
18 Administración 9
Servicios generales
6
Becarios/as 13
Contratados/as por proyectos
4

CIMA Memoria 2017 15
4 SERVICIOS
Biblioteca
La Biblioteca del CIMA es una biblioteca especializada en información
científica sobre recursos marinos, oceanografía, acuicultura… que atiende las
necesidades de información del personal del Centro de Investigacións Mariñas
de Corón (CIMA) y del Centro de Cultivos Mariños (CECUMAR) de Ribadeo,
así como consultas puntuales de otro personal de la Consellería. El acceso
para otros usuarios es público pero de carácter restringido (previa
autorización).
Instalaciones:
En el Centro de Investigacións Mariñas de Corón, la Biblioteca se encuentra ubicada en la segunda planta, en una sala
única que cumple la triple función de despacho, depósito, y espacio de consulta/lectura. La biblioteca pone a
disposición de los usuarios un par de mesas de trabajo individual/en grupo y un ordenador de consulta.
En el Centro de Cultivos Mariños de Ribadeo, se encuentra ubicada en la tercera planta, en una sala dedicada a
depósito, consulta y lectura.
Horario: de lunes a viernes de 9 a 14 horas.
Contacto:
CIMA-Corón: Telf.: 886 20 63 60 Correo electrónico: [email protected]
CIMA-Ribadeo Telf.: 982 88 94 02
Colección:
- Fondo bibliográfico: en el CIMA-Corón está compuesto por aproximadamente 1.500 volúmenes, sin posibilidad de
búsqueda por no haber un catálogo informatizado. En el CIMA-Ribadeo está compuesto por aproximadamente 700
volúmenes inventariados en una base de datos.
- Publicaciones periódicas: en 2015 el CIMA y el Consorcio de Bibliotecas Universitarias de Galicia (CBUG), firman un
convenio de colaboración que posibilita el acceso consorciado a tres colecciones de grandes editores (Elsevier,
Springer y Wiley), sumando un total que ronda los 5000 títulos de revistas científicas accesibles. El CIMA mantiene a
mayores suscripción directa a un reducido número de títulos especializados que no forman parte de los paquetes
consorciados.
- Bases de Datos: en la actualidad, el CIMA cuenta con acceso a la Web of Science (WoS) de la empresa Thomson
Scientific, gracias a la licencia contratada por el Ministerio de Educación y Ciencia y gestionada por la FECyT, con el
fin de dar acceso a estos recursos de información científica a las instituciones públicas de investigación.

16
A mayores de los recursos disponibles mediante adquisición/contratación, desde la biblioteca se hace una labor de
localización y clasificación de fuentes de información en acceso abierto, tratando de visibilizar sus posibilidades de uso
al tiempo que se complementa la colección de base.
Servicios
Sala de lectura.
Información sobre los recursos disponibles y acceso a los mismos http://www.netvibes.com/bibliotecacima.
Préstamo interno.
Servicio de obtención de documentos.
Asesoramiento en planteamiento de búsquedas y consulta de fuentes de información secundarias.
Orientación en el uso de herramientas de gestión de información.
Formación en el uso de gestores bibliográficos (EndNote Desktop / Web, Mendeley, Zotero): durante 2014-2015
se impartieron sesiones de formación de EndNote X7.
Atención a usuarios / sesiones de formación mediante videollamadas (Skype / Hangouts).
Salón de conferencias
El Centro de Investigacións Mariñas de Corón, cuenta con un salón de
conferencias con capacidad para 84 personas.
El acceso es público pero de carácter restringido (presta servicio a usuarios
internos y a usuarios externos autorizados).
Sala de juntas
El CIMA-Corón cuenta con tres salas de juntas dos con capacidad para 10
personas y una con capacidad para 15 personas.
Y la del Centro de Cultivos Mariños de Ribadeo tiene capacidad para 15
personas.
El acceso es público pero de carácter restringido (prestan servicio a usuarios
internos y a usuarios externos autorizados).

CIMA Memoria 2017 17
INVESTIGACIÓN

18

CIMA Memoria 2017 19
5 ACUICULTURA

20
RESUMEN
El proyecto Diversify aborda el estudio de los aspectos
clave para su desarrollo en la acuicultura europea de 6
especies: la corvina (Argyrosomus regius), el halibut
(Hippoglossus hippoglossus), la perca (Sanders
lucioperca), la seriola (Seriola dumerili), la lisa (Mugil
cephalus) y la cherna (Polyprion americanus).
Los centros CIMA e IGAFA de la Consellería do Mar
están involucrados en el desarrollo del cultivo de la
cherna (Polyprion americanus) conjuntamente con el IEO
y el Aquarium Finisterrae de A Coruña (AF) así como el
Acuario de O Grove como colaborador.
Otros centros de investigación españoles y extranjeros
participan en el desarrollo de esta especie: Hellenic
Center for Marine Research (HCMR), Universidad de las
Palmas de Gran Canaria (ULPGC), Institut de Recerca i
Tecnologia Agroalimentaries (IRTA), Institut Francais de
Recherche Pour L'Exploitation de la Mer (IFREMER) y
Universidad de La Laguna (ULL).
La cherna (Polyprion americanus) perteneciente a la
familia Serranidae está en fase de investigación y no
existe producción comercial, es una especie muy
atractiva para la acuicultura ya que tiene un crecimiento
rápido, madurez reproductiva tardía, alto precio en el
mercado, fácil manipulación en cautividad etc. Además
su pesquería se ha visto reducida drásticamente en los
últimos años.
El control de la reproducción y el establecimiento de
protocolos de cultivo larvario son considerados los
principales retos para el desarrollo del cultivo de la
cherna. Asimismo, la obtención de dietas óptimas
específicas para la cherna es de gran importancia para
su desarrollo en cultivo intensivo.
Las tareas desarrolladas están incluidas en tres áreas de
trabajo (WPs):
WP6 Reproducción de la cherna
WP12 Nutrición de la cherna
WP18 Cultivo larvario de la cherna
WP6. Reproducción de la cherna
Tarea 6.1 Captura de ejemplares salvajes para
establecer nuevos stocks de reproductores
Como se ha resaltado en informes anteriores presenta
gran dificultad la consecución de ejemplares del medio
natural para incrementar los stocks existentes, dada la
brusca caída de la pesquería de la cherna en los últimos
años (Fig.1).
PROYECTO DE INVESTIGACIÓN EQUIPO
TÍTULO:
Exploring the biological and socio-economic potential
of new/emerging candidate fish species for the
expansión of the European aquaculture industry.
ACRÓNIMO: DIVERSIFY
INICIO: diciembre, 2013
FINALIZACIÓN: diciembre, 2018
ENTIDAD FINANCIADORA: 7º Programa Marco.
KBBE2013.1.2.09 Diversification of fish species and
products in European Aquaculture Area 2.1.2.
Increased Sustainability of all Production Systems
(agriculture, forestry, fisheries and aquaculture
INVESTIGADOR PRINCIPAL:
Coordinador del proyecto: Constantinos C. Mylonas,
HCMR. GREECE
INVESTIGADORA PRINCIPAL EN EL CIMA:
Linares Cuerpo, F.
INVESTIGADORES/AS:
IGAFA: Rodriguez Villanueva, J.L.
Investigadores pertenecientes a 20 centros de investigación,
9 pequeñas y medianas empresas, 3 grandes empresas, 5
asociaciones profesionales y 1 de consumidores, de 12
países diferentes.
APOYO A LA INVESTIGACIÓN:
Méndez Pérez, B.; Cores González, M.J. (hasta 09/07/17),
Camino Saco, A. (desde10/07/17)
BECARIOS/AS:
Araujo Sendín, A.; Fuentes Villar, N.

CIMA Memoria 2017 21
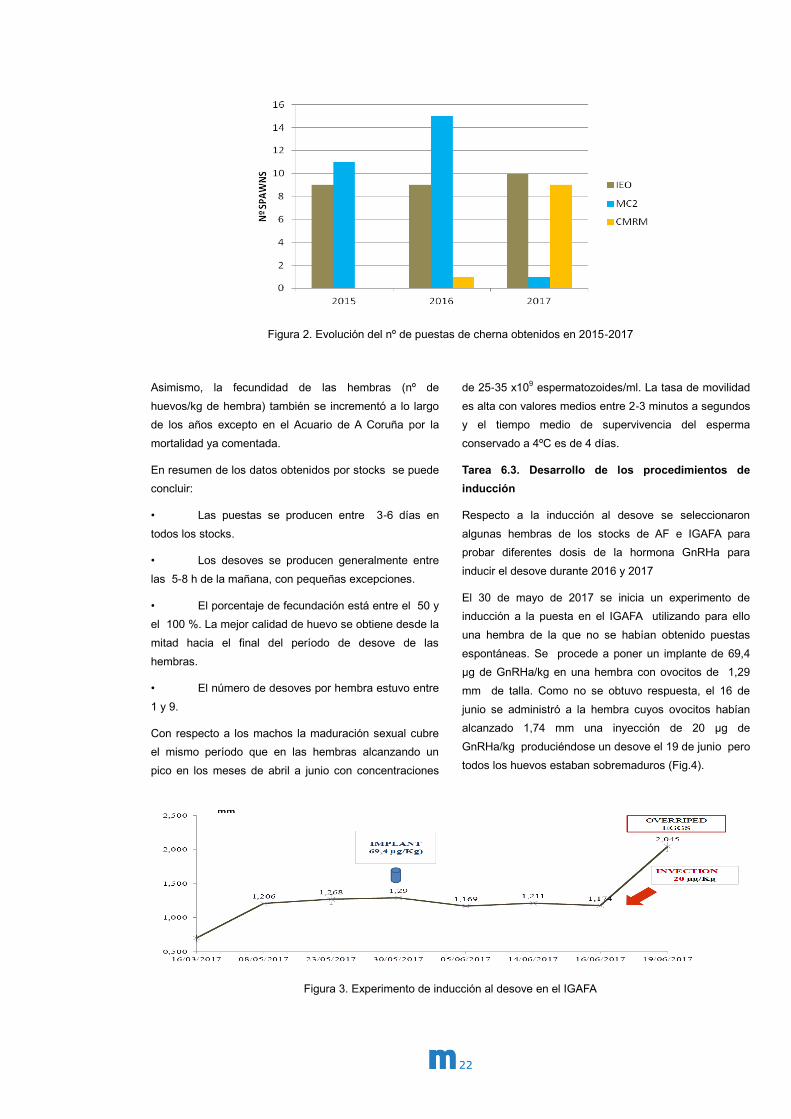
Figura1. Evolución anual de las capturas de cherna en Galicia (Kg) años 2000-2017
Se siguió la evolución de los 5 ejemplares capturados en
2014 y 2015. 4 se mantuvieron en las instalaciones del
IEO y 1 en el Acuario de O Grove (peso y longitud total:
6.05 kg y 73 cm, 11.25 kg y 79 cm, 10.58 kg y 77 cm, 2
kg y 46 cm y 3.99 kg y 83 cm respectivamente).
A pesar del esfuerzo realizado contactando con
pescadores de diversas localidades durante 2017 solo se
capturó 1 pez el 09/07/2017 (peso 4 kg y talla 55cm) que
se introdujo en un tanque de cuarentena (02 =95%,
salinidad= 35‰, pH=7.84 y Tª17.2) en las instalaciones
del Acuario de A Coruña (AF). Debido al stress sufrido
en la pesca y a pesar de haber sido sometido a
tratamiento antiinflamatorio el pez murió el 20/07/2017 y
la necropsia mostró numerosas alteraciones de la piel,
pérdida de escamas, hemorragias desde la mitad del
cuerpo hasta la aleta caudal así como aletas
deshilachadas. Las branquias tenían una coloración
normal sin presencia de parásitos.
Además, el stock de cherna del Acuario de O Grove se
controló mediante muestreos hasta abril de 2017, 7
peces: 2 hembras, 3 machos, 1 indeterminado y 1
inmaduro, el peso medio de los 6 ejemplares maduros
fue 13.39 ±1.40 kg, la longitud standard y la total fueron
de 76.33±3.67 cm y 88.17±4.17 cm. El ejemplar
inmaduro tenía un peso de 3.99 kg y 73 y 83 cm de
longitud estándard y total respectivamente. El
10/07/2017, 29/08/2017 y 19/09/ 2017, 3 peces murieron
en el Acuario de O Grove: 2 hembras (pesos 12.5 y 11.6
kg) y 1 macho (peso 8.8 Kg). Se realizó la necropsia de
estos peces en el CIMA observándose que todos ellos
presentaban una gran cantidad de grasa perivisceral . Se
mandaron muestras de hígado, riñón, cerebro y bazo al
Departamento de Ictiopatología de la Universidad de
Santiago para la realización de análisis bacteriológicos y
virológicos. No se encontró infección por virus y en
relación al control bacteriano los resultados fueron
positivos en Vibrio sp., pero no estaban claramente
asociados a la mortalidad de los peces.
Se tomaron también muestras de hígado, músculo y
gónada de los peces muertos para la realización de
análisis bioquímicos (proteínas, lípidos y ácidos grasos).
Debido a la gran cantidad de grasa encontrada en estos
especímenes que pensamos puede ser debida a la
alimentación suministrada en el Acuario se recomienda
la utilización de una dieta más adecuada para la cherna.
Tarea 6.2. Descripción del ciclo reproductivo
Se continúa el seguimiento exhaustivo de los stocks de
reproductores de cherna (IEO, AF e IGAFA) con la
finalidad de seguir avanzando en la descripción del ciclo
reproductivo en cautividad.
El principal objetivo fue conseguir suficientes puestas
para confirmar los intervalos de desove en las puestas
naturales y valorar la calidad de los huevos obtenidos
procedentes de hembras con diferentes tipos de
alimentación (WP12).
En 2017 las puestas tuvieron lugar de marzo a mayo y el
número de desoves se incrementó en todos los stocks a
lo largo de los años (Fig.2) excepto en el Acuario de A
Coruña debido a la mortalidad de parte las hembras
maduras en 2017. En el año 2017 el 81,5% de los
desoves fueron espontáneos y el 18,5% por inducción.
22
Figura 2. Evolución del nº de puestas de cherna obtenidos en 2015-2017
Asimismo, la fecundidad de las hembras (nº de
huevos/kg de hembra) también se incrementó a lo largo
de los años excepto en el Acuario de A Coruña por la
mortalidad ya comentada.
En resumen de los datos obtenidos por stocks se puede
concluir:
• Las puestas se producen entre 3-6 días en
todos los stocks.
• Los desoves se producen generalmente entre
las 5-8 h de la mañana, con pequeñas excepciones.
• El porcentaje de fecundación está entre el 50 y
el 100 %. La mejor calidad de huevo se obtiene desde la
mitad hacia el final del período de desove de las
hembras.
• El número de desoves por hembra estuvo entre
1 y 9.
Con respecto a los machos la maduración sexual cubre
el mismo período que en las hembras alcanzando un
pico en los meses de abril a junio con concentraciones
de 25-35 x109 espermatozoides/ml. La tasa de movilidad
es alta con valores medios entre 2-3 minutos a segundos
y el tiempo medio de supervivencia del esperma
conservado a 4ºC es de 4 días.
Tarea 6.3. Desarrollo de los procedimientos de
inducción
Respecto a la inducción al desove se seleccionaron
algunas hembras de los stocks de AF e IGAFA para
probar diferentes dosis de la hormona GnRHa para
inducir el desove durante 2016 y 2017
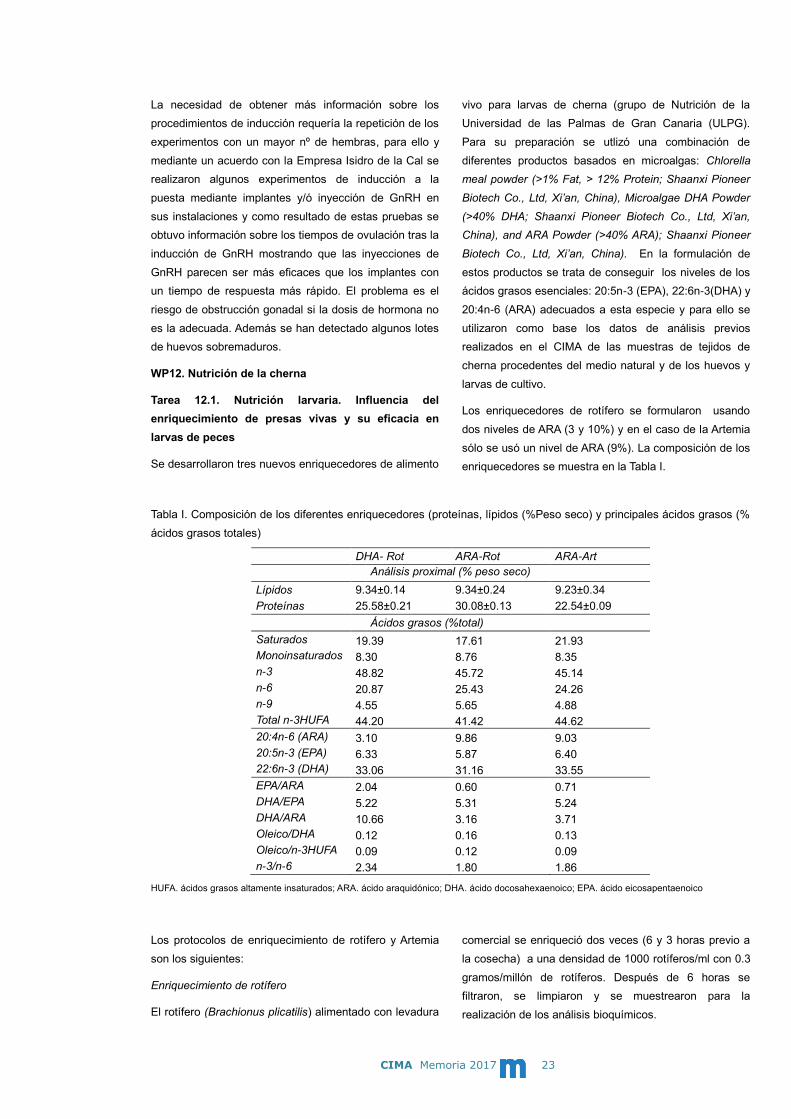
El 30 de mayo de 2017 se inicia un experimento de
inducción a la puesta en el IGAFA utilizando para ello
una hembra de la que no se habían obtenido puestas
espontáneas. Se procede a poner un implante de 69,4
µg de GnRHa/kg en una hembra con ovocitos de 1,29
mm de talla. Como no se obtuvo respuesta, el 16 de
junio se administró a la hembra cuyos ovocitos habían
alcanzado 1,74 mm una inyección de 20 µg de
GnRHa/kg produciéndose un desove el 19 de junio pero
todos los huevos estaban sobremaduros (Fig.4).
Figura 3. Experimento de inducción al desove en el IGAFA
CIMA Memoria 2017 23
La necesidad de obtener más información sobre los
procedimientos de inducción requería la repetición de los
experimentos con un mayor nº de hembras, para ello y
mediante un acuerdo con la Empresa Isidro de la Cal se
realizaron algunos experimentos de inducción a la
puesta mediante implantes y/ó inyección de GnRH en
sus instalaciones y como resultado de estas pruebas se
obtuvo información sobre los tiempos de ovulación tras la
inducción de GnRH mostrando que las inyecciones de
GnRH parecen ser más eficaces que los implantes con
un tiempo de respuesta más rápido. El problema es el
riesgo de obstrucción gonadal si la dosis de hormona no
es la adecuada. Además se han detectado algunos lotes
de huevos sobremaduros.
WP12. Nutrición de la cherna
Tarea 12.1. Nutrición larvaria. Influencia del
enriquecimiento de presas vivas y su eficacia en
larvas de peces
Se desarrollaron tres nuevos enriquecedores de alimento
vivo para larvas de cherna (grupo de Nutrición de la
Universidad de las Palmas de Gran Canaria (ULPG).
Para su preparación se utlizó una combinación de
diferentes productos basados en microalgas: Chlorella
meal powder (>1% Fat, > 12% Protein; Shaanxi Pioneer
Biotech Co., Ltd, Xi’an, China), Microalgae DHA Powder
(>40% DHA; Shaanxi Pioneer Biotech Co., Ltd, Xi’an,
China), and ARA Powder (>40% ARA); Shaanxi Pioneer
Biotech Co., Ltd, Xi’an, China). En la formulación de
estos productos se trata de conseguir los niveles de los
ácidos grasos esenciales: 20:5n-3 (EPA), 22:6n-3(DHA) y
20:4n-6 (ARA) adecuados a esta especie y para ello se
utilizaron como base los datos de análisis previos
realizados en el CIMA de las muestras de tejidos de
cherna procedentes del medio natural y de los huevos y
larvas de cultivo.
Los enriquecedores de rotífero se formularon usando
dos niveles de ARA (3 y 10%) y en el caso de la Artemia
sólo se usó un nivel de ARA (9%). La composición de los
enriquecedores se muestra en la Tabla I.
Tabla I. Composición de los diferentes enriquecedores (proteínas, lípidos (%Peso seco) y principales ácidos grasos (%
ácidos grasos totales)
DHA- Rot ARA-Rot ARA-Art
Análisis proximal (% peso seco)
Lípidos 9.34±0.14 9.34±0.24 9.23±0.34
Proteínas 25.58±0.21 30.08±0.13 22.54±0.09
Ácidos grasos (%total)
Saturados 19.39 17.61 21.93
Monoinsaturados 8.30 8.76 8.35
n-3 48.82 45.72 45.14
n-6 20.87 25.43 24.26
n-9 4.55 5.65 4.88
Total n-3HUFA 44.20 41.42 44.62
20:4n-6 (ARA) 3.10 9.86 9.03
20:5n-3 (EPA) 6.33 5.87 6.40
22:6n-3 (DHA) 33.06 31.16 33.55
EPA/ARA 2.04 0.60 0.71
DHA/EPA 5.22 5.31 5.24
DHA/ARA 10.66 3.16 3.71
Oleico/DHA 0.12 0.16 0.13
Oleico/n-3HUFA 0.09 0.12 0.09
n-3/n-6 2.34 1.80 1.86
HUFA. ácidos grasos altamente insaturados; ARA. ácido araquidónico; DHA. ácido docosahexaenoico; EPA. ácido eicosapentaenoico
Los protocolos de enriquecimiento de rotífero y Artemia
son los siguientes:
Enriquecimiento de rotífero
El rotífero (Brachionus plicatilis) alimentado con levadura
comercial se enriqueció dos veces (6 y 3 horas previo a
la cosecha) a una densidad de 1000 rotíferos/ml con 0.3
gramos/millón de rotíferos. Después de 6 horas se
filtraron, se limpiaron y se muestrearon para la
realización de los análisis bioquímicos.
24
Los análisis de los rotíferos enriquecidos con los dos
enriquecedores mencionados así como de los rotíferos
sin enriquecer se muestran en la Tabla II.
Tabla II. P. Composición del rotífero sin enriquecer y enriquecido con los dos enriquecedores (proteínas, lípidos (%Peso
seco)) y principales ácidos grasos (% ácidos grasos totales)
Rot-Yeast Rot-DHA Rot-ARA
Análisis proximal (% peso seco)
Lípidos 10.21±0.06 12.76±0.43 13.08±0.85
Proteínas 58.20±0.37 60.66±0.03 60.91±0.41
Ácidos grasos (%total)
Saturados 23.29 20.73 18.67
Monoinsaturados 53.48 40.41 38.71
n-3 5.45 16.50 18.37
n-6 13.42 16.45 18.55
n-9 24.24 20.54 20.10
Total n-3HUFA 3.68 13.52 15.27
20:4n-6 (ARA) 1.15 1.35 3.53
20:5n-3 (EPA) 1.58 3.12 3.23
22:6n-3 (DHA) 0.68 8.00 9.49
EPA/ARA 1.37 2.31 0.91
DHA/EPA 0.43 2.56 2.94
DHA/ARA 0.59 5.92 2.69
Oleico/DHA 31.88 2.00 1.65
Oleico/n-3HUFA 5.88 1.18 1.03
n-3/n-6 0.41 1.00 0.99
HUFA. ácidos grasos altamente insaturados; ARA. ácido araquidónico; DHA. ácido docosahexaenoico; EPA. ácido
eicosapentaenoico
Enriquecimiento de Artemia
Se utilizó Artemia salina (EG Inve). Artemia recién
eclosionada se enriqueció (0.4 gramos/litro) a una
densidad de 500 Art/ml en una sala iluminada con aire
acondicionado (12L:12D) durante 12 horas. La salinidad
del agua se mantuvo a 37 gramos/litro y la temperatura a
24 ± 0.01ºC. Tras el enriquecimiento se cosechó y se
muestreó para la realización de los análisis (Tabla III).
Tabla III. Proteínas, lípidos (%Peso seco) y ácidos grasos (% ácidos grasos totales) de Artemia sin enriquecer y
enriquecida.
Art-No enriquecida
Art-ARA
Análisis proximals (% peso seco)
Lípidos 21.42±0.18 19.48±0.24
Proteínas 54.59±0.87 68.53±2.21
Ácidos grasos (%total)
Saturados 21.41 20.73
Moninsaturados 32.35 40.41
n-3 35.56 16.50
n-6 8.14 16.45
n-9 21.55 20.54
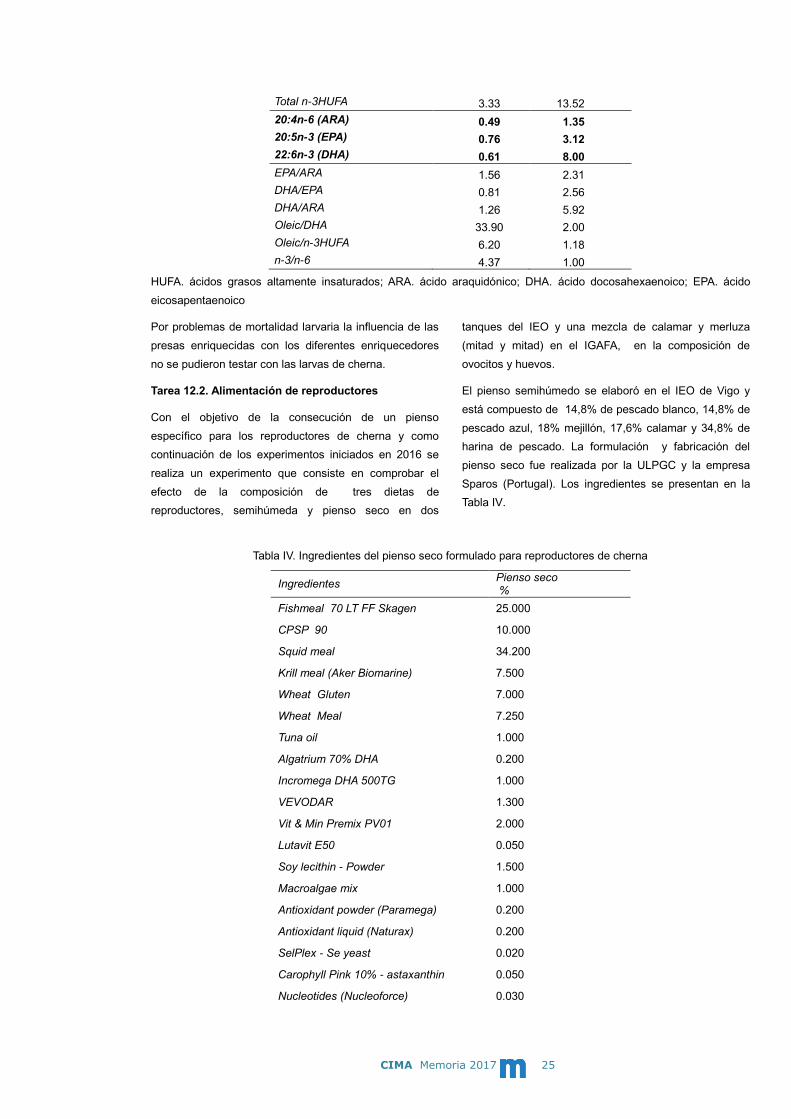
CIMA Memoria 2017 25
Total n-3HUFA 3.33 13.52
20:4n-6 (ARA) 0.49 1.35
20:5n-3 (EPA) 0.76 3.12
22:6n-3 (DHA) 0.61 8.00
EPA/ARA 1.56 2.31
DHA/EPA 0.81 2.56
DHA/ARA 1.26 5.92
Oleic/DHA 33.90 2.00
Oleic/n-3HUFA 6.20 1.18
n-3/n-6 4.37 1.00
HUFA. ácidos grasos altamente insaturados; ARA. ácido araquidónico; DHA. ácido docosahexaenoico; EPA. ácido
eicosapentaenoico
Por problemas de mortalidad larvaria la influencia de las
presas enriquecidas con los diferentes enriquecedores
no se pudieron testar con las larvas de cherna.
Tarea 12.2. Alimentación de reproductores
Con el objetivo de la consecución de un pienso
específico para los reproductores de cherna y como
continuación de los experimentos iniciados en 2016 se
realiza un experimento que consiste en comprobar el
efecto de la composición de tres dietas de
reproductores, semihúmeda y pienso seco en dos
tanques del IEO y una mezcla de calamar y merluza
(mitad y mitad) en el IGAFA, en la composición de
ovocitos y huevos.
El pienso semihúmedo se elaboró en el IEO de Vigo y
está compuesto de 14,8% de pescado blanco, 14,8% de
pescado azul, 18% mejillón, 17,6% calamar y 34,8% de
harina de pescado. La formulación y fabricación del
pienso seco fue realizada por la ULPGC y la empresa
Sparos (Portugal). Los ingredientes se presentan en la
Tabla IV.
Tabla IV. Ingredientes del pienso seco formulado para reproductores de cherna
Ingredientes Pienso seco %
Fishmeal 70 LT FF Skagen 25.000
CPSP 90 10.000
Squid meal 34.200
Krill meal (Aker Biomarine) 7.500
Wheat Gluten 7.000
Wheat Meal 7.250
Tuna oil 1.000
Algatrium 70% DHA 0.200
Incromega DHA 500TG 1.000
VEVODAR 1.300
Vit & Min Premix PV01 2.000
Lutavit E50 0.050
Soy lecithin - Powder 1.500
Macroalgae mix 1.000
Antioxidant powder (Paramega) 0.200
Antioxidant liquid (Naturax) 0.200
SelPlex - Se yeast 0.020
Carophyll Pink 10% - astaxanthin 0.050
Nucleotides (Nucleoforce) 0.030
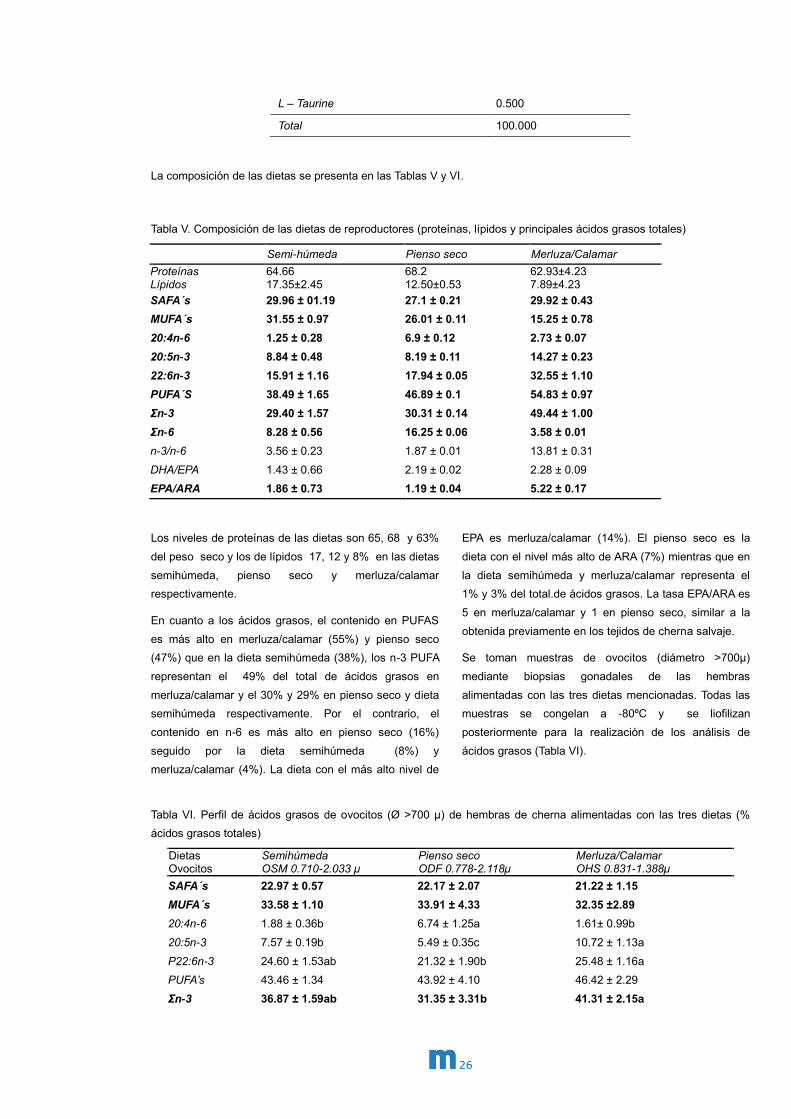
26
L – Taurine 0.500
Total 100.000
La composición de las dietas se presenta en las Tablas V y VI.
Tabla V. Composición de las dietas de reproductores (proteínas, lípidos y principales ácidos grasos totales)
Semi-húmeda Pienso seco Merluza/Calamar
Proteínas Lípidos
64.66 17.35±2.45
68.2 12.50±0.53
62.93±4.23 7.89±4.23
SAFA´s 29.96 ± 01.19 27.1 ± 0.21 29.92 ± 0.43
MUFA´s 31.55 ± 0.97 26.01 ± 0.11 15.25 ± 0.78
20:4n-6 1.25 ± 0.28 6.9 ± 0.12 2.73 ± 0.07
20:5n-3 8.84 ± 0.48 8.19 ± 0.11 14.27 ± 0.23
22:6n-3 15.91 ± 1.16 17.94 ± 0.05 32.55 ± 1.10
PUFA´S 38.49 ± 1.65 46.89 ± 0.1 54.83 ± 0.97
Σn-3 29.40 ± 1.57 30.31 ± 0.14 49.44 ± 1.00
Σn-6 8.28 ± 0.56 16.25 ± 0.06 3.58 ± 0.01
n-3/n-6 3.56 ± 0.23 1.87 ± 0.01 13.81 ± 0.31
DHA/EPA 1.43 ± 0.66 2.19 ± 0.02 2.28 ± 0.09
EPA/ARA 1.86 ± 0.73 1.19 ± 0.04 5.22 ± 0.17
Los niveles de proteínas de las dietas son 65, 68 y 63%
del peso seco y los de lípidos 17, 12 y 8% en las dietas
semihúmeda, pienso seco y merluza/calamar
respectivamente.
En cuanto a los ácidos grasos, el contenido en PUFAS
es más alto en merluza/calamar (55%) y pienso seco
(47%) que en la dieta semihúmeda (38%), los n-3 PUFA
representan el 49% del total de ácidos grasos en
merluza/calamar y el 30% y 29% en pienso seco y dieta
semihúmeda respectivamente. Por el contrario, el
contenido en n-6 es más alto en pienso seco (16%)
seguido por la dieta semihúmeda (8%) y
merluza/calamar (4%). La dieta con el más alto nivel de
EPA es merluza/calamar (14%). El pienso seco es la
dieta con el nivel más alto de ARA (7%) mientras que en
la dieta semihúmeda y merluza/calamar representa el
1% y 3% del total.de ácidos grasos. La tasa EPA/ARA es
5 en merluza/calamar y 1 en pienso seco, similar a la
obtenida previamente en los tejidos de cherna salvaje.
Se toman muestras de ovocitos (diámetro >700µ)
mediante biopsias gonadales de las hembras
alimentadas con las tres dietas mencionadas. Todas las
muestras se congelan a -80ºC y se liofilizan
posteriormente para la realización de los análisis de
ácidos grasos (Tabla VI).
Tabla VI. Perfil de ácidos grasos de ovocitos (Ø >700 µ) de hembras de cherna alimentadas con las tres dietas (%
ácidos grasos totales)
Dietas Ovocitos
Semihúmeda OSM 0.710-2.033 µ
Pienso seco ODF 0.778-2.118µ
Merluza/Calamar OHS 0.831-1.388µ
SAFA´s 22.97 ± 0.57 22.17 ± 2.07 21.22 ± 1.15
MUFA´s 33.58 ± 1.10 33.91 ± 4.33 32.35 ±2.89
20:4n-6 1.88 ± 0.36b 6.74 ± 1.25a 1.61± 0.99b
20:5n-3 7.57 ± 0.19b 5.49 ± 0.35c 10.72 ± 1.13a
P22:6n-3 24.60 ± 1.53ab 21.32 ± 1.90b 25.48 ± 1.16a
PUFA’s 43.46 ± 1.34 43.92 ± 4.10 46.42 ± 2.29
Σn-3 36.87 ± 1.59ab 31.35 ± 3.31b 41.31 ± 2.15a
CIMA Memoria 2017 27
Σn-6 6.34 ± 0.84b 12.32 ± 1.21ª 4.86 ± 0.50b
n-3/n-6 5.90 ± 0.93b 2.58 ± 0.20c 8.63 ± 0.90a
DHA/EPA 3.25 ± 0.15b 3.91 ± 0.27ª 2.40 ± 0.18c
EPA/ARA 4.16 ± 0.59b 0.86 ± 0.10c 6.71 ± 0.38a
No hay diferencias significativas en SAFAS’s, MUFA’s y PUFA’s en los ovocitos de hembras alimentadas con dieta
semihúmeda (OSM), pienso seco (ODF) y merluza/calamar (OHS) con valores de SAFA´s entre 21-23%, MUFA 32-34% y
PUFA 43-46%.
El contenido más alto en n-3 se observa en OHS (41%),
mientras OSM tenía 37% y ODF 31% del total de ácidos
grasos. En el caso de los n-6 PUFA los valores son
mucho más altos (12%) en ODF que en OHS y OSM (5-
6%). En lo que se refiere a los PUFA n-3, los valores de
20:5n-3 (EPA) y 22:6n-3 (DHA) presentan diferencias
significativas en los ovocitos procedentes de las hembras
alimentadas con las tres dietas. EPA tiene el valor más
alto en OHS, 11% del total, 8% en OSM y 5% en ODF.
Los valores de DHA son significativamente diferentes
entre OHS y ODF (25 y 21% respectivamente) y no hay
diferencias significativas entre OSM y ODF y OSM y
OHS. Con referencia a los n-6 PUFA, el nivel de 20:4n-6
(ARA) es más alto en ODF representando el 7% del total
de ácidos grasos, frente a OSM y OHS con el 2%. El
18:2n-6 presenta valores del 6% (ODF), 4% (OSM) y 3%
en OHS. La tasa n-3/n-6 es alta en OHS (9) debido a la
gran cantidad de EPA y tiene valores de 6 y 3 en OSM y
ODF respectivamente. DHA/EPA ratio tiene valores de 4,
3 y 2 en ODF, OSM y OHS respectivamente, mientras
que la tasa EPA/ARA es más alta en OHS (7) que en
OSM (4) y un valor mucho más bajo se obtuvo en
ODF(1) debido al alto valor de ARA.
Por otro lado, se tomaron muestras para análisis de
huevos viables procedentes de hembras alimentadas
con las tres dietas en los stocks de reproductores del
IEO e IGAFA. El contenido en lípidos (%P.seco) y la
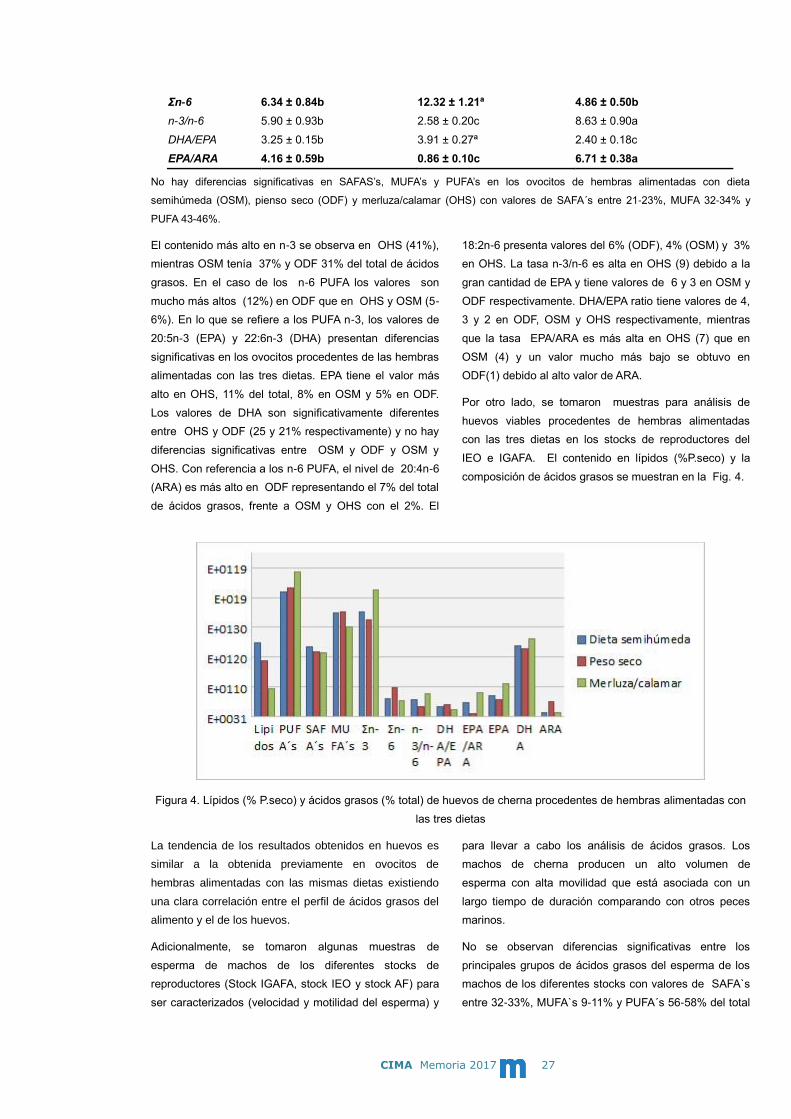
composición de ácidos grasos se muestran en la Fig. 4.
Figura 4. Lípidos (% P.seco) y ácidos grasos (% total) de huevos de cherna procedentes de hembras alimentadas con
las tres dietas
La tendencia de los resultados obtenidos en huevos es
similar a la obtenida previamente en ovocitos de
hembras alimentadas con las mismas dietas existiendo
una clara correlación entre el perfil de ácidos grasos del
alimento y el de los huevos.
Adicionalmente, se tomaron algunas muestras de
esperma de machos de los diferentes stocks de
reproductores (Stock IGAFA, stock IEO y stock AF) para
ser caracterizados (velocidad y motilidad del esperma) y
para llevar a cabo los análisis de ácidos grasos. Los
machos de cherna producen un alto volumen de
esperma con alta movilidad que está asociada con un
largo tiempo de duración comparando con otros peces
marinos.
No se observan diferencias significativas entre los
principales grupos de ácidos grasos del esperma de los
machos de los diferentes stocks con valores de SAFA`s
entre 32-33%, MUFA`s 9-11% y PUFA´s 56-58% del total

28
de ácidos grasos. El contenido en n-3 PUFA varia entre
49-51% y el de n-6 PUFA es más alto en el esperma
procedente de los machos del stock del IEO (tanque S2)
suponiendo el 9% del total que en el IGAFA (5%) y AF
(6%) probablemente debido también a la influencia de la
dieta de los reproductores.
Como resumen del área de nutrición es de destacar:
- Se diseñaron los enriquecedores de presas
vivas (rotíferos y Artemia). Se utilizaron dos niveles de
ARA en el enriquecedor de rotífero y un nivel de ARA en
el de Artemia. y se evaluó el efecto del enriquecimiento
con los nuevos productos sobre la composición
bioquímica de rotíferos y Artemia.
- Se encontró una clara relación entre el perfil de
ácidos grasos de las dietas de reproductores probadas
(semihúmeda, pienso seco y merluza/calamar) y el perfil
de ácidos grasos de ovocitos y huevos procedentes de
hembras alimentadas con esas dietas.
- Los resultados obtenidos con pienso seco
demostraron que la dieta de la cherna debe contener
gran cantidad de proteínas, bajo nivel de lípidos, una alta
cantidad de n-3 PUFA y la tasa EPA/ARA debe ser
similar a la obtenida en las gónadas de las hembras del
medio natural (1-1.5 aproximadamente). No obstante la
dieta de merluza/calamar (mitad y mitad) parece tener
una buena calidad ya que presenta una gran cantidad de
proteínas y de n-3 PUFA (EPA y DHA) aunque la tasa
EPA/ARA obtenida en ovocitos y huevos de hembras
alimentadas con esta dieta es alta comparada con la
obtenida en gónadas de hembras del medio natural.
- Se han obtenido los primeros datos del perfil de
ácidos grasos del esperma de machos de cherna de los
diferentes stocks de reproductores.
WP18. Cultivo larvario de cherna
Durante abril de 2017 se llevó a cabo un experimento de
incubación de huevos de cherna con tres rangos de
temperatura 13-14ºC, 16-17ºC y 19-20ºC con la finalidad
de validar los resultados obtenidos en 2016.
El experimento se realiza con huevos flotantes que
proceden de tres desoves (uno de ellos fue rechazado
por contener huevos de baja calidad). Se llevó a cabo
por triplicado con una densidad entre 350-450
huevos/litro en vasos de 2.5 litros en cuya base tenían
una malla 500 μm el agua se filtró a 1 μm, y el flujo fue
continuo de modo que el agua se renueva cada 2-3
horas con aireación suave. La temperatura se reguló con
una bomba de calor y se registró cada 3 horas entre las
9:00 am y las 9:00 pm.
Diariamente se sacó una muestra de 10 huevos para
identificar el estado de desarrollo embrionario y se retiran
los huevos muertos mediante sifonado. Al final del
proceso se cuentan las larvas eclosionadas
diferenciando las de buena calidad de las que presentan
alguna deformidad.
Se calcula la tasa de eclosión y la de deformidad
mediante las siguientes ecuaciones:
% Eclosión = NL/NH x 100
% Deformes = ND/NL x 100
NL es el nº total de larvas eclosionadas, NH el nº de
huevos eclosionados y ND el nº de larvas deformes.
La temperatura media obtenida a lo largo del
experimento fue de 13.7 ± 0.2ºC, 16.6 ± 0.4ºC y 19.5 ±
0.4ºC para cada uno de los rangos de temperatura.
La duración del desarrollo embrionario varió según la
temperatura: 4 días a 19.5ºC, 5 días a 16.6ºC y 7 días a
13.7ºC.
Se observaron diferencias significativas (p<0.05) en tasa
de eclosión y deformidad de las larvas entre 13.7ºC y
19ºC mientras que las diferencias no son significativas
con respecto a la temperatura de 16.6ºC. Estos
resultados sugieren que las bajas temperaturas
producen bajas tasas de eclosión y alto % de deformes.
Se calcularon los días considerados críticos para la
viabilidad de la embriogénesis, para ello se recogieron
los huevos del fondo de los vasos durante los tres
primeros días de incubación calculándose el % respecto
al total de huevos incubados, obteniéndose que a 13.7ºC
la mortalidad de los huevos fue del 30% en los tres
primeros días de incubación, a 16.6ºC, alrededor del
75% de la mortalidad se concentra en los dos primeros
días y a 19.5ºC la mayor mortalidad (60%) se produjo en
el primer día de incubación.
Los parámetros de incubación deben de ajustarse y las
instalaciones y los sistemas deben ser optimizados para
incrementar la calidad de la embriogénesis y la
supervivencia.
Se han realizado avances en lo que se refiere a la
comprensión de la ontogenia y el desarrollo larvario de la
cherna. De este estudio se concluye que la ontogénesis
del sistema digestivo de la cherna es un proceso lento en
comparación con otras especies. La ontogénesis de los

CIMA Memoria 2017 29
órganos relacionados con el sistema digestivo y la visión
no se completaron hasta los 23 días de vida de las
larvas. Las estructuras como las glándulas gástricas y el
ciego pilórico que son características de un desarrollo
completo del sistema digestivo no fueron identificadas en
este estudio.
La ontogenia de la retina de la cherna es similar a la de
la mayor parte de las especies de peces. En la eclosión,
la retina era un tejido indiferenciado y no funcional, como
ocurre en los primeros estadios de vida pelágica de la
mayoría de peces marinos. El estudio también muestra
que la agudeza visual, la distancia a la que el ojo puede
diferenciar entre dos puntos mejoraba con el tiempo
como muestran los análisis histológicos. Aunque según
el estudio el sistema digestivo y visual estaban
desarrollados para permitir que las larvas comiesen no
se detectó ningún alimento en el canal digestivo lo que
demuestra que la dieta de las larvas estaba basada
exclusivamente en las reservas del saco vitelino.
El estudio de la ontogénesis y de la formación de los
sistemas básicos en los organismos de cultivo, como se
ha probado en otros casos, parece ser la base para
lograr la optimización del protocolo de cultivo larvario de
las larvas de cherna. Ya que los principales órganos
como las glándulas gástricas no aparecen hasta que las
larvas tiene una longitud de 5.5 mm, una combinación de
presas fácilmente capturables y digeribles como los
rotíferos o copépodos tienen que ser incluidas en el
protocolo de alimentación de las larvas de cherna. Ello
en combinación con la optimización de las condiciones
de cultivo, como tipo e hidrodinámica de los tanques de
cultivo larvario, temperatura, etc. son imprenscindibles
para el desarrollo de un protocolo de cultivo larvario.
Información detallada de este proyecto en la web:
www.diversifyfish.eu/

30

CIMA Memoria 2017 31
RESUMEN
Esta acción se presenta como un complemento de las
actuaciones que se están realizando en el ámbito de la
reproducción de la cherna en el marco del proyecto
Diversify.
El objetivo fundamental es definir un protocolo de la
maduración gonadal en cherna, Polyprion
americanus mediante la administración de FSH y LH
recombinantes con el objetivo final de la obtención
de puestas regulares en cautividad.
Objetivos específicos:
Valoración de la viabilidad de la utilización
de la hormona FSH en ejemplares de cherna.
Determinación de la dosis adecuada en
cherna µg/Kg (FSH) probando diferentes dosis
utilizadas en otras especies.
Influencia de la administración de FSH en la
evolución del desarrollo reproductivo de las
hembras de cherna.
Influencia de la administración de LH en la
evolución del desarrollo reproductivo de las hembras
de cherna.
En el desarrollo de esta acción además del CIMA e
IGAFA participan la Planta de Cultivos del CO de Vigo y
la Empresa Rara Avis. Biotec, S.L.
La estrategia de esta acción de investigación tiene como
fundamento fisiológico la maduración gonadal en cherna
mediante la administración de FSH y LH recombinantes.
Dado que estas hormonas actúan directamente sobre las
células gonadales induciendo su maduración, su
utilización permite obviar la intervención del eje
hipotálamo-hipofisario y, por tanto, todas las variables
ambientales que influyen sobre el ciclo reproductivo
espontáneo.
Esta metodología de inducción del ciclo es la que se
lleva a cabo regularmente en reproducción asistida
humana y en ganadería, y se ha demostrado eficaz en
aves y reptiles, y actualmente ya se está experimentando
con especies de peces que presentan disfunción en su
reproducción como el lenguado senegalés, la lubina y la
anguila.
Descripción de los experimentos:
1. Obtención de FSH y LH recombinantes.
Esta tarea es relizada por la empresa “Rara Avis.Biotec
S.L.” que es la encargada de la producción de hormonas.
No existe información acerca de las secuencias
aminoacídicas específicas para FSH y LH del género
Polyprion. Sin embargo, dada la similitud de otras
secuencias sí conocidas (citocromo b, rodopsina, G
protein-coupled receptor 61, etc.) con perciformes como
Dicentrarchus labrax, se considera que la utilización de
FSH y LH recombinantes con las secuencias conocidas
de esta última especie será satisfactoria.
El proceso para la obtención de FSH y LH
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Inducción a la vitelogénesis, maduración ovocitaria,
ovulación y espermiogénesis en cherna (Polyprion
americanus).
INICIO: marzo, 2018
FINALIZACIÓN: diciembre, 2018
ENTIDAD FINANCIADORA:
Dirección Xeral de Desenvolvemento Pesqueiro
Consellería do Mar
Código: PGIDIT-CIMA
INVESTIGADORA PRINCIPAL:
Linares Cuerpo, F.
INVESTIGADORES/AS:
IGAFA: Rodriguez Villanueva, J.L.
IEO: Álvarez Blázquez, B.; Pérez Rial, E.
Rara Avis. Biotec S.L.: Giménez Nebot, I.
APOYO A LA INVESTIGACIÓN:
Méndez Pérez, B.; Cores González, M.J. (hasta agosto
2017); Camino Saco, A. (desde septiembre 2017)
BECARIOS/AS:
Araujo Sendín, A.; Fuentes Villar, N.

32
recombinantes en cantidades suficientes para la
inducción de la maduración gonadal tiene una duración
aproximada de dos a tres meses y se realiza en cultivos
celulares CHO.
2. Inducción de maduración gonadal en
Polyprion.
Se utilizan las hormonas disponibles para trabajar en
cherna, que fueron usadas anteriormente en lubina
(Dicentrarchus labrax). Las hormonas FSH y LH que se
utilizan habitualmente en peces suelen proceder de
hormonas producidas en Pichia pastoris (hongo).
Los resultados obtenidos en 2016 nos indicaron que una
de las hembras del IGAFA (7B19) responde
positivamente al tratamiento con FSH/LH consiguiendo
completar la vitelogénesis evolucionando los ovocitos
desde 0,480 mm a 1,139 mm en un período de tres
meses. Los análisis de hormonas esteroideas: Extradiol
(E2), Ketotestosterona (KT) y Testosterona (T) muestran
la tendencia a incrementarse del E2 y KT en la hembra
7B19 tras la inyección con FSH/LH, pero los resultados
deben considerarse preliminares por la gran variabilidad
existente en las muestras.
Asimismo, se tomaron muestras de ovocitos para realizar
análisis de ácidos grasos de todas las hembras del
IGAFA del tanque (T2) donde se realizaron los
experimentos de inducción y del tanque T3:
♀4BDB- control. No presenta desarrollo ovocitario
♀7B19-hormonada con FSH/LH y que presenta
evolución de ovocitos
♀3FF2- control con evolución ovocitaria y primera puesta
espontánea en 2016
y de dos hembras del T3:
♀6D01- evolución ovocitaria y puestas espontáneas en
2017
♀499A- sin evolución ovocitaria
Estos datos se procesaron en 2017 con los siguientes
resultados:
En primer lugar con las muestras del tanque T2 se trata
de conocer si existen diferencias en el perfil de ácidos
grasos entre los ovocitos obtenidos mediante inducción
con FSH/LH y los obtenidos de manera natural
procedentes de las hembras ♀7B19 y ♀3FF2 (Tabla I).
Tabla I. Análisis de ácidos grasos de los ovocitos (% total ácidos grasos) de las hembras con y sin tratamiento hormonal
♀7B19
Ø 0,484-1,255
♀3FF2
Ø 0,831-1,388
14:0 1,38 1,17
16:0 16,09 14,05
17:0 0,80 0,73
18:0 5,01 4,67
SAFA´s 23,50 20,83
16:1n-7 4,66 3,85
18:1n-9 15,21 15,15
18:1n-7 7,03 6,93
20:1n-9 1,68 1,71
MUFA´s 31,13 30,30
18:2n-6 2,64 3,39
20:4n-6 1,70 1,76
20:5n-3 11,38 11,68
22:5n-3 3,29 4,25
22:6n-3 26,41 27,52
PUFA´S 46,85 50,27
Σn-3 42,38 44,98
Σn-6 4,34 5,14
n-3/n-6 9,80 8,75
DHA/EPA 2,33 2,36
EPA/ARA 6,82 6,67
CIMA Memoria 2017 33
Los perfiles de ácidos grasos de los ovocitos
procedentes de las dos hembras son bastante similares
en lo que se refiere al porcentaje respecto al total de
ácidos grasos por lo que no parecen estar afectados por
el tratamiento hormonal con FSH/LH de las hembras.
Adicionalmente, se realizaron los perfiles de ácidos
grasos de los ovocitos procedentes de todas las
hembras del IGAFA con distinto grado de desarrollo
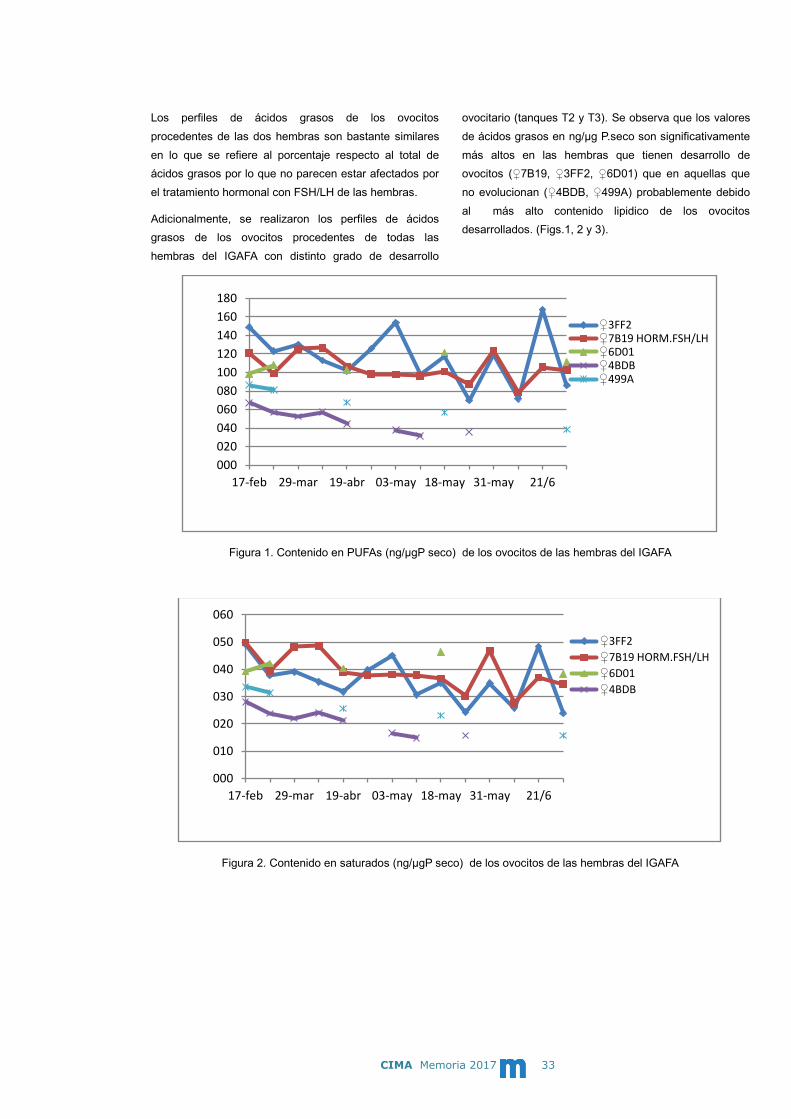
ovocitario (tanques T2 y T3). Se observa que los valores
de ácidos grasos en ng/µg P.seco son significativamente
más altos en las hembras que tienen desarrollo de
ovocitos (♀7B19, ♀3FF2, ♀6D01) que en aquellas que
no evolucionan (♀4BDB, ♀499A) probablemente debido
al más alto contenido lipidico de los ovocitos
desarrollados. (Figs.1, 2 y 3).
Figura 1. Contenido en PUFAs (ng/µgP seco) de los ovocitos de las hembras del IGAFA
Figura 2. Contenido en saturados (ng/µgP seco) de los ovocitos de las hembras del IGAFA
000
020
040
060
080
100
120
140
160
180
17-feb 29-mar 19-abr 03-may 18-may 31-may 21/6
♀3FF2 ♀7B19 HORM.FSH/LH ♀6D01 ♀4BDB ♀499A
000
010
020
030
040
050
060
17-feb 29-mar 19-abr 03-may 18-may 31-may 21/6
♀3FF2
♀7B19 HORM.FSH/LH
♀6D01
♀4BDB
34
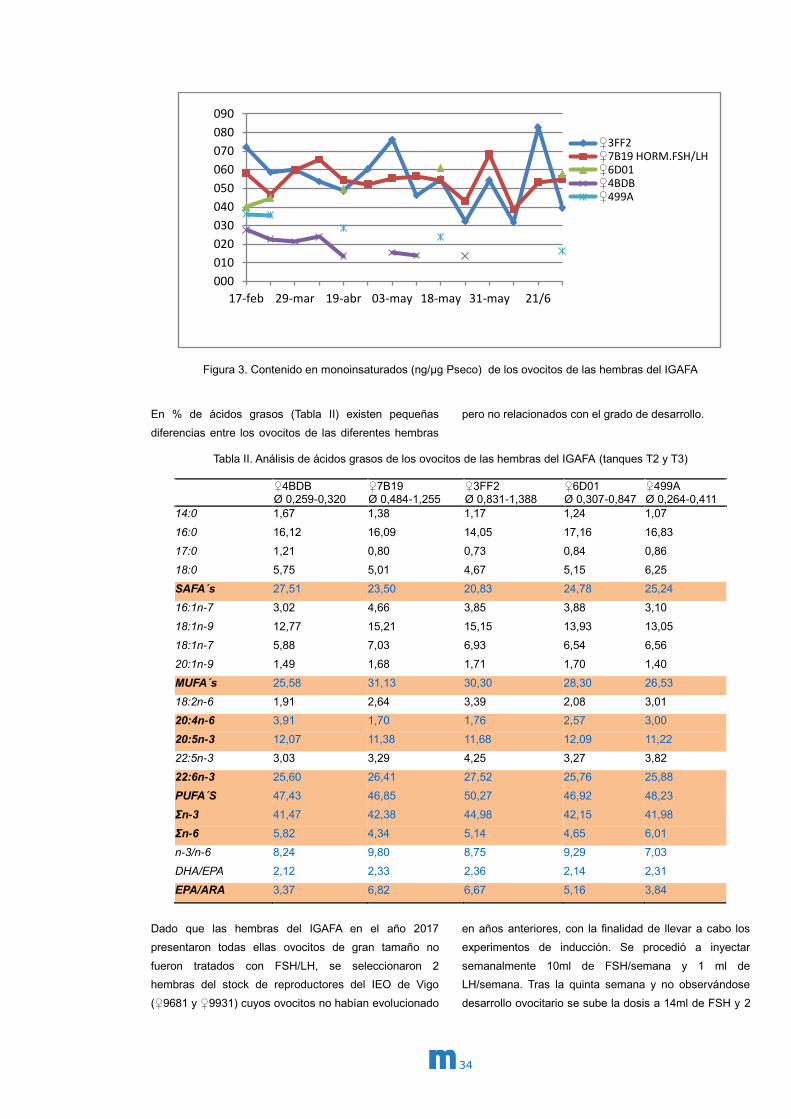
Figura 3. Contenido en monoinsaturados (ng/µg Pseco) de los ovocitos de las hembras del IGAFA
En % de ácidos grasos (Tabla II) existen pequeñas
diferencias entre los ovocitos de las diferentes hembras
pero no relacionados con el grado de desarrollo.
Tabla II. Análisis de ácidos grasos de los ovocitos de las hembras del IGAFA (tanques T2 y T3)
♀4BDB
Ø 0,259-0,320
♀7B19
Ø 0,484-1,255
♀3FF2
Ø 0,831-1,388
♀6D01
Ø 0,307-0,847
♀499A
Ø 0,264-0,411
14:0 1,67 1,38 1,17 1,24 1,07
16:0 16,12 16,09 14,05 17,16 16,83
17:0 1,21 0,80 0,73 0,84 0,86
18:0 5,75 5,01 4,67 5,15 6,25
SAFA´s 27,51 23,50 20,83 24,78 25,24
16:1n-7 3,02 4,66 3,85 3,88 3,10
18:1n-9 12,77 15,21 15,15 13,93 13,05
18:1n-7 5,88 7,03 6,93 6,54 6,56
20:1n-9 1,49 1,68 1,71 1,70 1,40
MUFA´s 25,58 31,13 30,30 28,30 26,53
18:2n-6 1,91 2,64 3,39 2,08 3,01
20:4n-6 3,91 1,70 1,76 2,57 3,00
20:5n-3 12,07 11,38 11,68 12,09 11,22
22:5n-3 3,03 3,29 4,25 3,27 3,82
22:6n-3 25,60 26,41 27,52 25,76 25,88
PUFA´S 47,43 46,85 50,27 46,92 48,23
Σn-3 41,47 42,38 44,98 42,15 41,98
Σn-6 5,82 4,34 5,14 4,65 6,01
n-3/n-6 8,24 9,80 8,75 9,29 7,03
DHA/EPA 2,12 2,33 2,36 2,14 2,31
EPA/ARA 3,37 6,82 6,67 5,16 3,84
Dado que las hembras del IGAFA en el año 2017
presentaron todas ellas ovocitos de gran tamaño no
fueron tratados con FSH/LH, se seleccionaron 2
hembras del stock de reproductores del IEO de Vigo
(♀9681 y ♀9931) cuyos ovocitos no habían evolucionado
en años anteriores, con la finalidad de llevar a cabo los
experimentos de inducción. Se procedió a inyectar
semanalmente 10ml de FSH/semana y 1 ml de
LH/semana. Tras la quinta semana y no observándose
desarrollo ovocitario se sube la dosis a 14ml de FSH y 2
000
010
020
030
040
050
060
070
080
090
17-feb 29-mar 19-abr 03-may 18-may 31-may 21/6
♀3FF2 ♀7B19 HORM.FSH/LH ♀6D01 ♀4BDB ♀499A

CIMA Memoria 2017 35
ml de LH.
Diariamente se midieron los parámetros de temperatura,
pH, oxígeno, etc. Periódicamente se controló el
crecimiento de los ejemplares (talla, peso). Para el
control de los ejemplares y previamente a la inyección
con FSH se procedió a su anestesia con 2-fenoxietanol
(0,4 mg/litro).
La duración del experimento fue de 10 semanas y se
tomaron muestras a tiempo 0 y en las semanas 4, 8 y 10.
En cada muestreo se tomó muestra de sangre para
analizar hormonas esteroideas y se realizó biopsia
ovárica para extracción de ovocitos para control del
desarrollo y para análisis de ácidos grasos. El
experimento finalizó el 12 de julio no observándose
evolución del tamaño de ovocitos tras la inducción con
FSH/LH. Se considera que la elevada temperatura del
agua de mar durante los meses de mayo- julio pudo
haber influido negativamente en cuanto a los efectos
producidos por la inducción con FSH/LH, por lo que se
decide el año próximo comenzar los experimentos
durante el invierno para evitar la subida de temperaturas.

36

CIMA Memoria 2017 37
PROYECTO DE INVESTIGACIÓN EQUIPO
TÍTULO:
Contribución genética para la recuperación de los
bancos naturales de coquina (Donax spp.) en Galicia.
INICIO: junio, 2017
FINALIZACIÓN: diciembre, 2019
ENTIDAD FINANCIADORA:
MINECO (Ministerio de Economía y Competitividad)
Código: AGL-2016-75288-R
INVESTIGADORA PRINCIPAL:
UDC: Méndez Felpeto, J.
INVESTIGADORES/AS:
CIMA: Martínez Patiño, D.; Nóvoa Vázquez, S.
UDC: Insua Pombo, A.; Nantón, A.; Fernández Pérez,
J.; Avecilla, F.
APOYO A LA INVESTIGACIÓN:
Bouzamayor Yáñez, V.; Cotarelo Jardón, M.J.; Páez
Pérez, F.J.; Ruíz Misioné, M.
BECARIOS/AS:
Castilla del Río, S.; Granjel Piñeiro, E.
RESUMEN
Ante la disminución de las capturas de coquina en Galicia
se inició un estudio genético que aporte conocimientos
científicos que ayuden a identificar el problema existente
en los bancos naturales de este molusco bivalvo.
El objetivo general del proyecto consiste en la evaluación
genética de la coquina Donax trunculus en el mayor
número de localidades de la Península Ibérica con el fin
de conocer la diversidad genética y la existencia o no de
diferenciación poblacional con el propósito de conservar
y gestionar este recurso de manera eficaz.
Durante este año hemos evaluado, en parte, la
diversidad genética y la diferenciación poblacional en la
coquina Donax trunculus de la Península Ibérica
mediante marcadores moleculares.
Asimismo, dada la existencia de otras especies del
género Donax (D. vittatus, D. semistriatus y D.
variegatus) en el área de distribución a analizar,
caracterizaremos el gen de ADNr 5S y los espaciadores
transcritos internos, ITS1 y ITS2, y secuenciaremos el
genoma mitocondrial en las cuatro especies objeto de
estudio con el fin de obtener marcadores especie-
específicos para la identificación de estas especies en
fresco o procesadas, lo que mejoraría los métodos de
identificación. Nuestro trabajo ha permitido desarrollar
nuevos marcadores para diferenciar especies del género
Donax basados en ADN nuclear.
Además, hemos procedido a la secuenciación del
mitogenoma de las especies del género Donax presentes
en la Península Ibérica.
Ante la presencia de al menos dos especies (D.
trunculus y D. vittatus) en la costa atlántica de la
Península Ibérica, se hace necesario desarrollar
marcadores moleculares para caracterizar los bancos
naturales de D. vittatus como otra especie a conservar
y/o gestionar.
Para este objetivo, nos planteamos desarrollar loci
microsatélite a través de un nuevo enfoque mediante
herramientas de secuenciación de nueva generación
(NGS) y evaluar la diversidad genética y estructura
poblacional en localidades de la costa atlántica de la
Península Ibérica mediante marcadores microsatélite de
ADN mitocondrial (gen COI). Trabajo en realización.
En el criadero del CIMA de Ribadeo se realizó la
estabulación, acondicionamiento de reproductores y
cultivo de Donax trunculus. Los progenitores se
obtuvieron a partir de muestreos realizados en el banco
de Vilarrube (ría de Cedeira, NO Galicia), en las
siguientes fechas: 20 septiembre 2016, 15 diciembre
2016, 29 marzo 2017, 25 abril 2017 y 24 mayo 2017. Se
acondicionaron los individuos de diciembre y se
obtuvieron desoves espontáneos de todos los adultos
estabulados en el criadero (Tabla I).

38
A B
Tabla I.- Muestreos realizados en Vilarrube en los que se capturaron progenitores de cuyos desoves: CQ1, CQ2, CQ3, CQ4, CQ5, CQ6, CQ7, CQ8, CQ9, CQ10, CQ11, CQ12, CQ14, se obtuvo semilla para sembrar en el medio natural.
Los lotes de los progenitores se mantienen separados y los
individuos que van muriendo gradualmente a lo largo del
tiempo, se almacenan en botes con alcohol de 96º para su
posterior análisis genotípico y de su contribución genética a la
semilla.
Con fecha 29 de agosto de 2017 se hace un recuento de la
semilla obtenida en los desoves CQ7 y CQ10 (del 25 de mayo
y del 1 de junio de 2017), de progenitores obtenidos en el
muestreo de Vilarrube del 29 de marzo de 2017. A esta semilla
también se le realizó el seguimiento genético, analizando los
individuos que quedan retenidos en tamices de 800, 1000 y
1500 µm (Fig. 1A).
Figura 1.- Semilla obtenida de los desoves CQ7 y CQ10 analizada genéticamente y sembrada en la playa de Abrela. A: semilla retenida en tamices de 800, 1000 y 1500 µm; B: semilla de 12 mm marcada y sembrada en mayo de 2018.
La primera siembra de esta semilla (unos 10.000
individuos), se realiza el 3 de mayo de 2018 en la playa
de Abrela (ría de Viveiro, NO Galicia), cuando ya tienen
una talla de 12 mm (Fig. 1B).Se están haciendo pruebas
de marcaje de la semilla, con diferentes métodos, con el
objetivo de realizar un seguimiento periódico a la playa
de Abrela y localizar posibles individuos pertenecientes a
las siembras realizadas. Esta semilla también será
analizada por el grupo de genética, cerrando el ciclo del
estudio.
Muestreos=Progenitores Desoves
Muestreo 15 diciembre 2016 15/02/2017 CQ1
09/03/2017 CQ2
Muestreo 29 marzo 2017
03/04/2017 CQ3
24/04/2017 CQ4
16/05/2017 CQ6
25/05/2017 CQ7
01/06/2017 CQ10
Muestreo 25 abril 2017
27/04/2017 CQ5
26/05/2017 CQ8
02/06/2017 CQ9
12/06/2017 CQ12
21/06/2017 CQ14
Muestreo 24 mayo 2017 01/06/2017 CQ11

CIMA Memoria 2017 39
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Inducción de la fijación y metamorfosis en almeja fina,
babosa y japonesa.
INICIO: junio, 2016
FINALIZACIÓN: diciembre, 2018
ENTIDAD FINANCIADORA:
Consellería do Mar. Xunta de Galicia.
Código: CIMA 16/05
INVESTIGADORA PRINCIPAL:
Nóvoa Vázquez, S.
INVESTIGADORES/AS:
CIMA: Martínez Patiño, D.; Ojea Martínez, J.
USC: Pazos Castelos, A.J.; Pérez-Parallé Mera, M.L.;
Sánchez López, J.L.
APOYO A LA INVESTIGACIÓN:
Bouzamayor Yáñez, V.; Cotarelo Jardón, M.J.; Páez
Pérez, F.J.; Ruíz Misioné, M.
BECARIOS/AS:
Castilla del Río, S.; Granjel Piñeiro, E.; Matesanz
Salgado, D.
RESUMEN
Los dos equipos de trabajo que forman parte de la
acción son el CIMA de Ribadeo y el Instituto de
Acuicultura (IA), de la Universidad de Santiago de
Compostela (USC).
En esta acción se estudia la fijación y metamorfosis en
especies de almeja de interés marisquero en Galicia:
almeja japonesa Ruditapes philippinarum (Adams &
Reeve, 1850), almeja babosa Venerupis corrugata
(Gmelin, 1791) y almeja fina Ruditapes decussatus
(Linnaeus, 1758). Su principal objetivo es establecer un
método rutinario, efectivo y de bajo coste para inducir a
su fijación y metamorfosis y de esta forma, mejorar su
producción en criadero industrial.
Los objetivos que se pretenden abordar en los tres años
que dura el proyecto son:
1. Efecto de iones y compuestos neuroactivos sobre la
fijación y metamorfosis de larvas de almeja.
2. Efecto de “biofilms” naturales sobre la fijación y
metamorfosis de larvas de almeja.
3. Efecto de diferentes substratos sobre la fijación y
metamorfosis de larvas de almeja.
Los trabajos realizados durante el año 2016 a 2017,
abordaron el estudio de la fijación y metamorfosis de la
almeja japonesa y babosa. Se realizaron experimentos
de laboratorio en los que no solamente se probaron
distintas sustancias inductoras (el ion cloruro cálcico) y
compuestos neuroactivos (ácido γ-amino-n-butírico
(GABA), epinefrina, norepinefrina, L-DOPA, IBMX,
serotonina y acetilcolina), sino que además se testaron
diferentes sustratos para estudiar la fijación de las larvas.
Para complementar estos experimentos, también se hizo
su seguimiento durante varios días, a nivel de cultivo
estándar, y así conocer más los procesos de
metamorfosis de cada especie. Y dada la circunstancia
de poder disponer en el CIMA de Ribadeo de la navaja
Ensis magnus, Schumacher, 1817 y de la coquina Donax
trunculus, Linnaeus, 1758, se hicieron estudios
preliminares de estas especies de igual interés
marisquero.
Las tareas del cronograma de la acción realizadas en
este año fueron la T0, T1 y T3 que se describen a
continuación.
T0. Acondicionamiento de progenitores y cultivos
larvarios
En las instalaciones del CIMA de Ribadeo se realizó el
cultivo larvario hasta que las larvas eran competentes, es
decir, hasta que se pone de manifiesto la aparición del
pie en la mayoría de las larvas y en este momento, se
iniciaban las pruebas.
Para iniciar el cultivo de estas especies, los
reproductores se recogieron del medio natural en la
época que coincidía con su madurez sexual. En el caso
de la almeja japonesa, desde mayo hasta octubre. La
almeja babosa presenta gametos maduros durante todo
el año, no existe un período de reposo. Una vez
estabuladas en el criadero, en tanques de 150 litros con
circuito abierto y a una temperatura similar a la del medio

40
natural, se obtuvieron los desoves unas veces de forma
espontánea y otras, mediante inducciones con choques
térmicos.
Después del desarrollo embrionario se inició el cultivo
larvario en tanques de fibra de vidrio de 500 litros, con
agua filtrada a una temperatura entre 18-20ºC,
renovación total del agua cada dos días y alimentación
con una mezcla de especies microalgales cultivadas en
sistema de producción continua.
Cada especie tiene una duración diferente del cultivo
larvario, es decir, desde larva D hasta pediveliger. La
almeja japonesa inicia la fijación y proceso de
metamorfosis a los 15 días con un tamaño de 180-230
µm y las larvas quedan retenidas en tamiz de 150 µm; en
el caso de la almeja babosa, la fijación tiene lugar a los
16 días con un tamaño de 250 µm y también quedan
retenidas en tamiz de 150 µm.
Cuando la mayoría del total de individuos del cultivo
llegaron a esta etapa, se iniciaron las pruebas de
laboratorio. Se recogieron los individuos necesarios para
las pruebas del resto, se continuó su cultivo en los
mismos tanques de 500 litros pero en cilindros con malla
de 150 µm y a los que se les aplica un flujo forzado por
“air-lift” descendente.
Obtenidas las larvas competentes, con una parte se
iniciaban las pruebas de laboratorio en estas
instalaciones y otra parte, era necesario su transporte en
las condiciones más óptimas posibles, hasta el
laboratorio del Instituto de Acuicultura (IA) de Santiago
de Compostela.
Se consiguió optimizar un método de transporte con el
que las larvas llegaron al laboratorio del IA, en perfecto
estado a las 12 o hasta 24 horas, pudiendo iniciar de
forma inmediata los ensayos.
Las larvas fueron transportadas en un pequeño tamiz de
malla de 140 µm dentro de una placa Petri de vidrio. Iban
ligeramente humedecidas mediante una gasa mojada en
agua salada y conservaron la humedad y temperatura, al
ir embaladas herméticamente en una pequeña caja de
porexpán (poliestireno expandido, EPS) con enfriadores
de hielo.
T1 y T3. Efecto de iones y diferentes substratos
sobre la fijación de larvas de almeja
Se realizó un experimento de laboratorio con almeja
japonesa (Ruditapes philippinarum). La sustancia
inductora empleada fue el cloruro cálcico (CaCl2.2H2O)
en distintas concentraciones y tres sustratos distintos:
placas de poliestireno (empleadas anteriormente en otro
ensayo de laboratorio); placas de vidrio y placas de vidrio
pero con un tamiz con malla de nylon de 140 µm en su
interior.
El experimento fue por triplicado en placas Petri de 90
mm pero las larvas fueron mantenidas en un volumen
final de 25 ml de agua de mar filtrada (FSW). Cada
experimento incluyó un control con FSW sin inductor
(Control: 0 mM). La fijación y metamorfosis fue
monitorizada con un microscopio Nikon SMZ-2T
transcurridas 24 y 48 horas.
Sustratos empleados:
Placas Petri de poliestireno
Placas Petri de vidrio
Placas Petri de vidrio con tamiz de malla de nylon de 140 µm
Preparación de las concentraciones de cloruro cálcico
STOCK
Cloruro cálcico (30 mM) 2,2 gr 500 mL
AGUA MAR FILTRADA
CIMA Memoria 2017 41
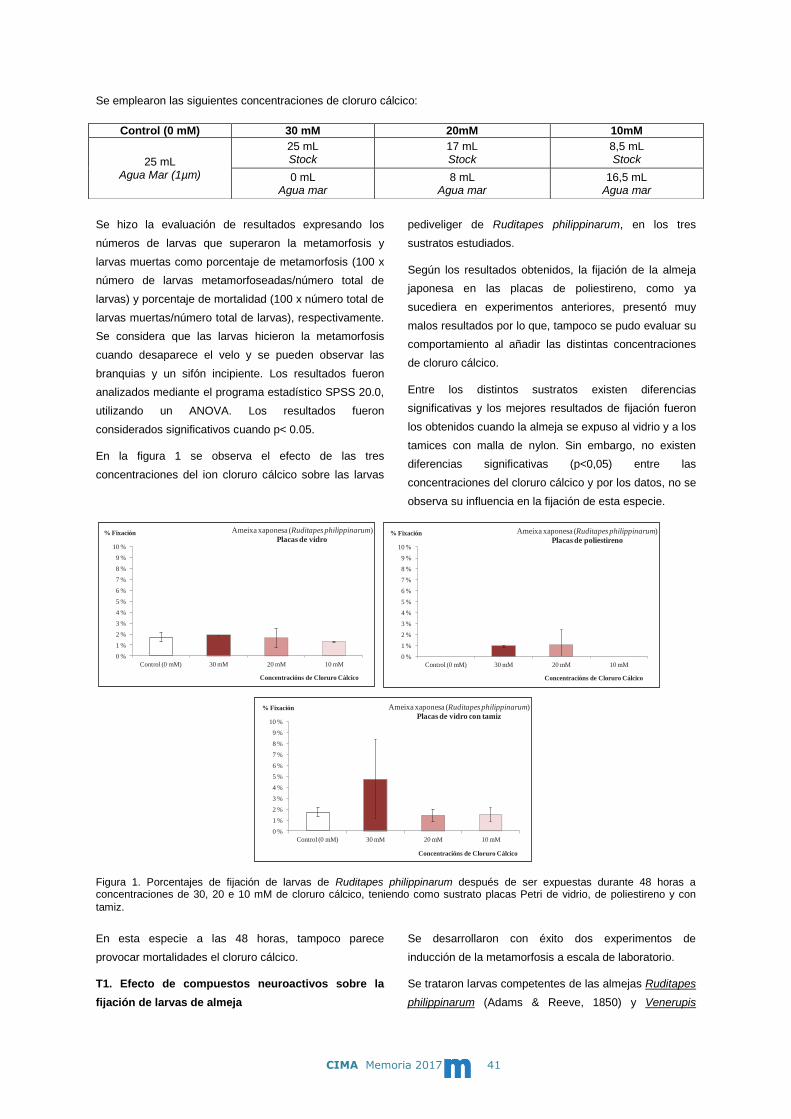
Se emplearon las siguientes concentraciones de cloruro cálcico:
Se hizo la evaluación de resultados expresando los
números de larvas que superaron la metamorfosis y
larvas muertas como porcentaje de metamorfosis (100 x
número de larvas metamorfoseadas/número total de
larvas) y porcentaje de mortalidad (100 x número total de
larvas muertas/número total de larvas), respectivamente.
Se considera que las larvas hicieron la metamorfosis
cuando desaparece el velo y se pueden observar las
branquias y un sifón incipiente. Los resultados fueron
analizados mediante el programa estadístico SPSS 20.0,
utilizando un ANOVA. Los resultados fueron
considerados significativos cuando p< 0.05.
En la figura 1 se observa el efecto de las tres
concentraciones del ion cloruro cálcico sobre las larvas
pediveliger de Ruditapes philippinarum, en los tres
sustratos estudiados.
Según los resultados obtenidos, la fijación de la almeja
japonesa en las placas de poliestireno, como ya
sucediera en experimentos anteriores, presentó muy
malos resultados por lo que, tampoco se pudo evaluar su
comportamiento al añadir las distintas concentraciones
de cloruro cálcico.
Entre los distintos sustratos existen diferencias
significativas y los mejores resultados de fijación fueron
los obtenidos cuando la almeja se expuso al vidrio y a los
tamices con malla de nylon. Sin embargo, no existen
diferencias significativas (p<0,05) entre las
concentraciones del cloruro cálcico y por los datos, no se
observa su influencia en la fijación de esta especie.
Figura 1. Porcentajes de fijación de larvas de Ruditapes philippinarum después de ser expuestas durante 48 horas a concentraciones de 30, 20 e 10 mM de cloruro cálcico, teniendo como sustrato placas Petri de vidrio, de poliestireno y con
tamiz.
En esta especie a las 48 horas, tampoco parece
provocar mortalidades el cloruro cálcico.
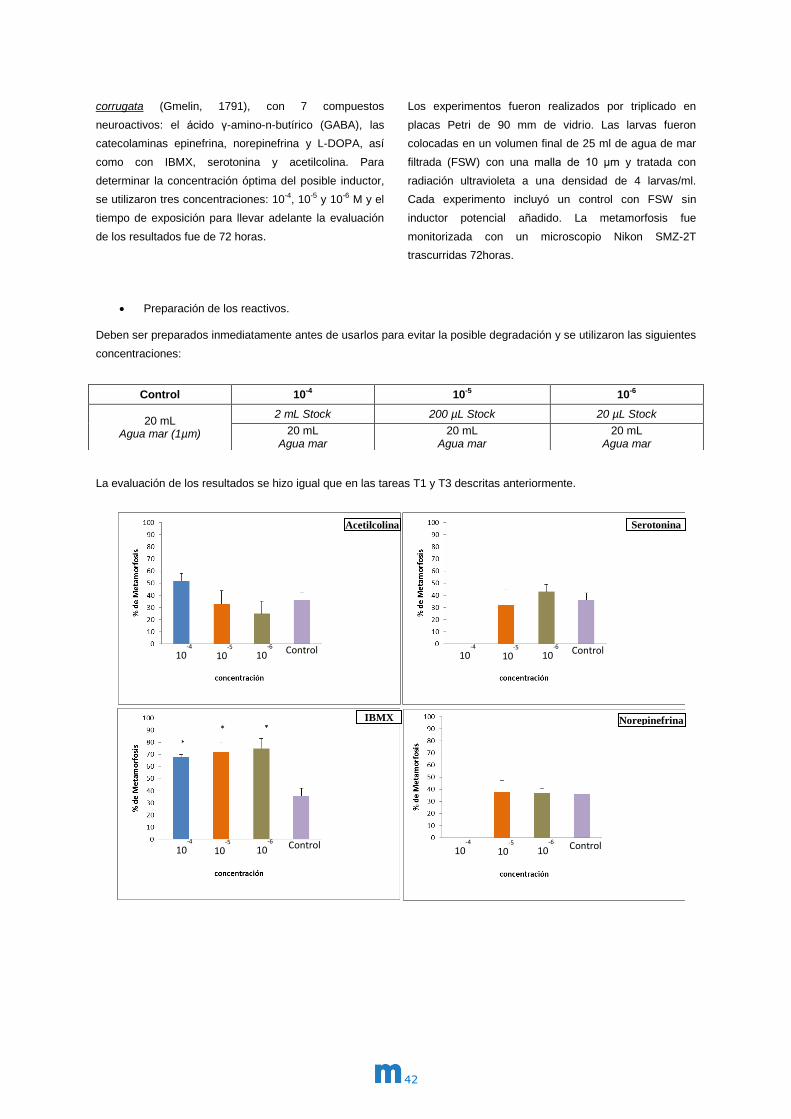
T1. Efecto de compuestos neuroactivos sobre la
fijación de larvas de almeja
Se desarrollaron con éxito dos experimentos de
inducción de la metamorfosis a escala de laboratorio.
Se trataron larvas competentes de las almejas Ruditapes
philippinarum (Adams & Reeve, 1850) y Venerupis
Control (0 mM) 30 mM 20mM 10mM
25 mL Agua Mar (1µm)
25 mL Stock
17 mL Stock
8,5 mL Stock
0 mL Agua mar
8 mL Agua mar
16,5 mL Agua mar
0 %
1 %
2 %
3 %
4 %
5 %
6 %
7 %
8 %
9 %
10 %
Control (0 mM) 30 mM 20 mM 10 mM
% Fixación
Concentracións de Cloruro Cálcico
Ameixa xaponesa (Ruditapes philippinarum)
Placas de poliestireno
0 %
1 %
2 %
3 %
4 %
5 %
6 %
7 %
8 %
9 %
10 %
Control (0 mM) 30 mM 20 mM 10 mM
% Fixación
Concentracións de Cloruro Cálcico
Ameixa xaponesa (Ruditapes philippinarum)
Placas de vidro
0 %
1 %
2 %
3 %
4 %
5 %
6 %
7 %
8 %
9 %
10 %
Control (0 mM) 30 mM 20 mM 10 mM
% Fixación
Concentracións de Cloruro Cálcico
Ameixa xaponesa (Ruditapes philippinarum)
Placas de vidro con tamiz
42
corrugata (Gmelin, 1791), con 7 compuestos
neuroactivos: el ácido γ-amino-n-butírico (GABA), las
catecolaminas epinefrina, norepinefrina y L-DOPA, así
como con IBMX, serotonina y acetilcolina. Para
determinar la concentración óptima del posible inductor,
se utilizaron tres concentraciones: 10-4, 10-5 y 10-6 M y el
tiempo de exposición para llevar adelante la evaluación
de los resultados fue de 72 horas.
Los experimentos fueron realizados por triplicado en
placas Petri de 90 mm de vidrio. Las larvas fueron
colocadas en un volumen final de 25 ml de agua de mar
filtrada (FSW) con una malla de 10 μm y tratada con
radiación ultravioleta a una densidad de 4 larvas/ml.
Cada experimento incluyó un control con FSW sin
inductor potencial añadido. La metamorfosis fue
monitorizada con un microscopio Nikon SMZ-2T
trascurridas 72horas.
Preparación de los reactivos.
Deben ser preparados inmediatamente antes de usarlos para evitar la posible degradación y se utilizaron las siguientes
concentraciones:
La evaluación de los resultados se hizo igual que en las tareas T1 y T3 descritas anteriormente.
Control 10-4 10-5 10-6
20 mL Agua mar (1µm)
2 mL Stock 200 µL Stock 20 µL Stock
20 mL Agua mar
20 mL Agua mar
20 mL Agua mar
10-4
10-5
10-6
Control
Acetilcolina
10-4
10-5
10-6
Control
Serotonina
10-4
10-5
10-6
Control
* IBMX
10-4
10-5
10-6
Control
Norepinefrina
CIMA Memoria 2017 43
Figura 2.- Porcentaje de metamorfosis de larvas de Ruditapes philippinarum tras ser expuestas 72 h a las concentraciones 10-
4, 10
-5 y 10
-6 M de: acetilcolina, serotonina, IBMX, norepinefrina, epinefrina, GABA y L-DOPA.
Los asteriscos indican diferencias significativas (*p<0,05) con el control, que carece de la presencia de moléculas inductoras.
En la figura 2 se puede observar el efecto de la
concentración de los diferentes inductores en el
porcentaje de metamorfosis en larvas pedivelíger de
Ruditapes philippinarum.
En los experimentos con larvas de almeja japónica,
Ruditapes philippinarum, se puede observar como
GABA, epinefrina e IBMX son inductores efectivos de la
metamorfosis. En los dos primeros casos para una
concentración de 10-5 M el porcentaje de éxito en cuanto
a superación de este proceso va desde
aproximadamente un 30% en el control hasta más de un
60% en presencia de estos inductores. En el caso del
IBMX se encontraron incrementos semejantes tanto
para concentraciones de 10-5 M como de 10-6 M.
Norepinefrina, acetilcolina, serotonina y L-DOPA no
fueron efectivas en las concentraciones estudiadas o
incluso afectaron de forma negativa la metamorfosis de
las larvas.
Es importante señalar que tanto GABA como epinefrina
no se mostraron tóxicas para las larvas en la
concentración antes señalada y aún disminuyeron la
mortalidad a la mitad de la observada en los controles. El
inductor IBMX no mostró diferencias significativas con los
controles en cuanto a mortalidad larvaria.
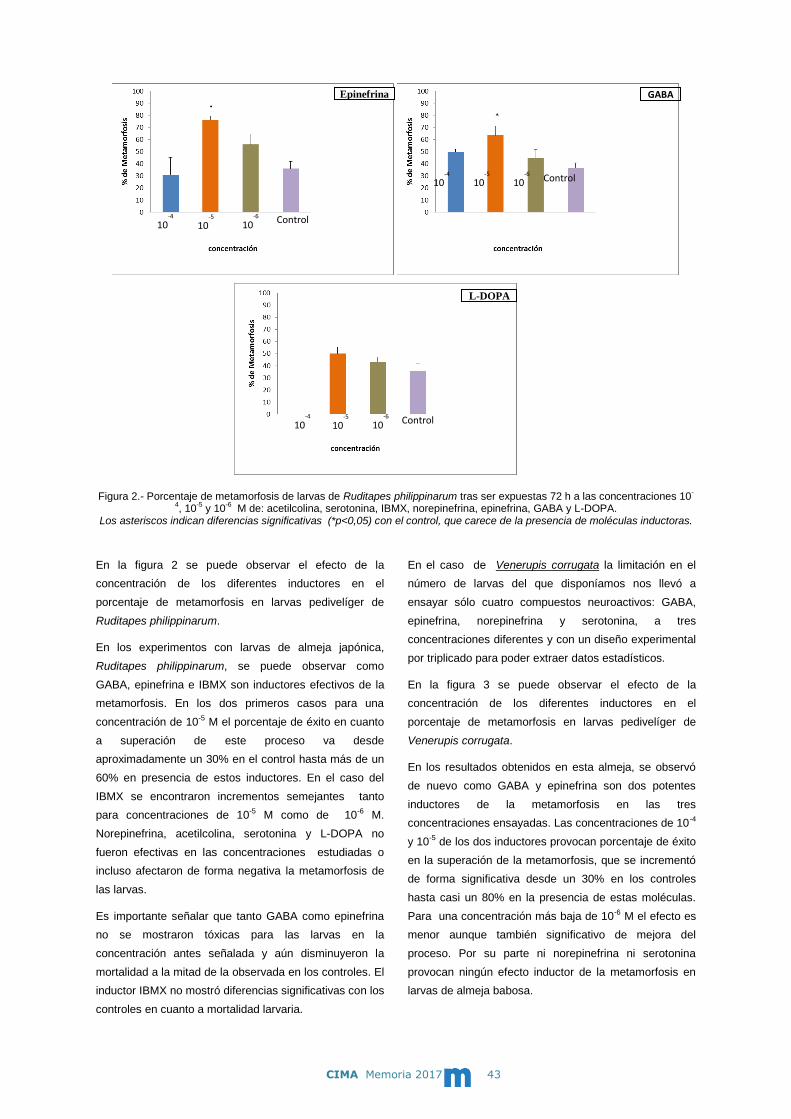
En el caso de Venerupis corrugata la limitación en el
número de larvas del que disponíamos nos llevó a
ensayar sólo cuatro compuestos neuroactivos: GABA,
epinefrina, norepinefrina y serotonina, a tres
concentraciones diferentes y con un diseño experimental
por triplicado para poder extraer datos estadísticos.
En la figura 3 se puede observar el efecto de la
concentración de los diferentes inductores en el
porcentaje de metamorfosis en larvas pedivelíger de
Venerupis corrugata.
En los resultados obtenidos en esta almeja, se observó
de nuevo como GABA y epinefrina son dos potentes
inductores de la metamorfosis en las tres
concentraciones ensayadas. Las concentraciones de 10-4
y 10-5 de los dos inductores provocan porcentaje de éxito
en la superación de la metamorfosis, que se incrementó
de forma significativa desde un 30% en los controles
hasta casi un 80% en la presencia de estas moléculas.
Para una concentración más baja de 10-6 M el efecto es
menor aunque también significativo de mejora del
proceso. Por su parte ni norepinefrina ni serotonina
provocan ningún efecto inductor de la metamorfosis en
larvas de almeja babosa.
10-4
10-5
10-6
Control
L-DOPA
10-4
10-5
10-6
Control
Epinefrina GABA
10-4
10-5
10-6
Control
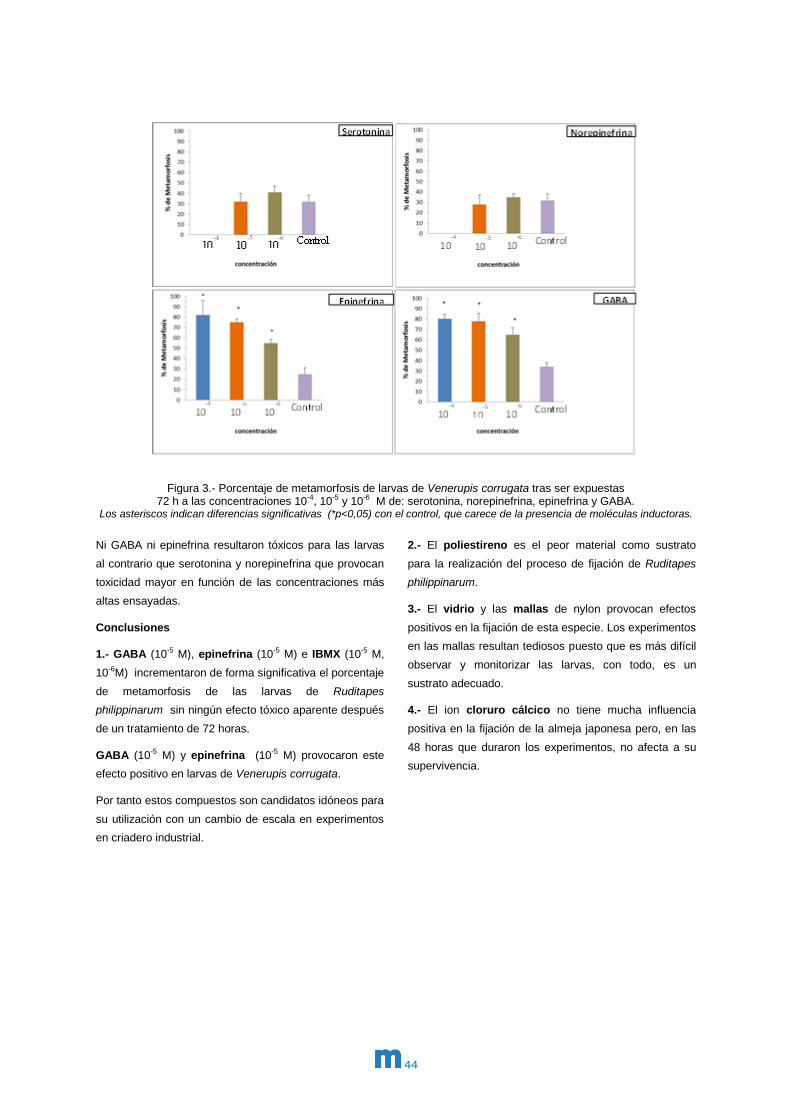
44
Figura 3.- Porcentaje de metamorfosis de larvas de Venerupis corrugata tras ser expuestas 72 h a las concentraciones 10-4, 10-5 y 10-6 M de: serotonina, norepinefrina, epinefrina y GABA.
Los asteriscos indican diferencias significativas (*p<0,05) con el control, que carece de la presencia de moléculas inductoras.
Ni GABA ni epinefrina resultaron tóxicos para las larvas
al contrario que serotonina y norepinefrina que provocan
toxicidad mayor en función de las concentraciones más
altas ensayadas.
Conclusiones
1.- GABA (10-5 M), epinefrina (10-5 M) e IBMX (10-5 M,
10-6M) incrementaron de forma significativa el porcentaje
de metamorfosis de las larvas de Ruditapes
philippinarum sin ningún efecto tóxico aparente después
de un tratamiento de 72 horas.
GABA (10-5 M) y epinefrina (10-5 M) provocaron este
efecto positivo en larvas de Venerupis corrugata.
Por tanto estos compuestos son candidatos idóneos para
su utilización con un cambio de escala en experimentos
en criadero industrial.
2.- El poliestireno es el peor material como sustrato
para la realización del proceso de fijación de Ruditapes
philippinarum.
3.- El vidrio y las mallas de nylon provocan efectos
positivos en la fijación de esta especie. Los experimentos
en las mallas resultan tediosos puesto que es más difícil
observar y monitorizar las larvas, con todo, es un
sustrato adecuado.
4.- El ion cloruro cálcico no tiene mucha influencia
positiva en la fijación de la almeja japonesa pero, en las
48 horas que duraron los experimentos, no afecta a su
supervivencia.

CIMA Memoria 2017 45
RESUMEN
Desarrollo de las tareas de cultivo de semilla de
moluscos bivalvos realizadas en las instalaciones del
minicriadero situado en el IGAFA.
En noviembre de 2016 se incorporaron los nuevos
becarios a las instalaciones del minicriadero del IGAFA
para llevar a cabo la mejora en el aprendizaje de
técnicas de cultivo de semilla de moluscos bivalvos y
para la realización de tareas de control microbiológico de
las distintas zonas de la instalación.
Los dos primeros meses, se llevó a cabo la puesta a
punto de las instalaciones para poder comenzar la
actividad de cultivo de semilla en enero de 2017.
Mejoras realizadas:
.- Puesta a punto de la zona del tanque de decantación.
.- Reparación y mantenimiento del filtro rotativo.
.- Puesta a punto de la climatización de la sala larvaria.
.- Puesta a punto y mejora del circuito de agua caliente
en la sala de reproductores.
.- Reparación del pasteurizador, cambio de la resistencia
y puesta a punto del intercambiador de placas para tratar
el agua que entra en el circuito continuo de fitoplancton.
.- Cambio del equipo U.V. en el circuito continuo de
fitoplancton que queda como sistema de seguridad por si
fallase el autoclave.
.- Introducción de un sistema de vaporización portátil
para la limpieza de los distintos circuitos de fitoplancton.
.- Introducción de una lámpara U.V. para mejorar la
calidad del agua del semillero en caso de necesidad.
.- Reparación y mantenimiento del climatizador del
semillero.
.- Compra de una nevera para el mantenimiento de los
medios y placas que se utilizan en microbiología.
.- Introducción de una cabina para llevar a cabo todas las
tareas de microbiología.
.- Reparación, mantenimiento y puesta a punto del
autoclave.
Una vez realizados estos ajustes, comenzamos la
actividad de cultivo de 4 especies de moluscos bivalvos:
almeja babosa (Venerupis corrugata), almeja japonesa
(Ruditapes philippinarum), almeja bicuda (Venerupis
aurea) y almeja fina (Ruditapes decussatus).
Los lotes de reproductores que utilizamos para la
producción de semilla, fueron adquiridos en diferentes
cofradías de la ría de Arousa (Illa de Arousa, Vilanova de
Arousa y Cambados).
Una vez la semilla alcanzó la talla de entrega, se
facilitaron los ejemplares a diferentes cofradías que las
solicitaran previamente.
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Procedimientos de cultivo intensivo de especies de
interés marisquero.
ACRÓNIMO: PESCIM
INICIO: enero, 2017
FINALIZACIÓN: diciembre, 2018
ENTIDAD FINANCIADORA:
Consellería do Mar. Xunta de Galicia.
Código: CIMA 17/02
INVESTIGADORA PRINCIPAL:
Martínez Patiño, D.
INVESTIGADORES/AS:
CIMA: Montes Pérez, J.; Nóvoa Vázquez, S.; Ojea Martínez, J.
IGAFA: Andrés Rivas, M.C.; Lastres Couto, M.A.
USC: Barja Pérez, J.L.; Prado Plana, S.
APOYO A LA INVESTIGACIÓN:
Bouzamayor Yáñez, V.; Cotarelo Jardón, M.J.; Meléndez
Ramos, I.; Páez Pérez, F.J.; Penas Pampín, E.; Ruíz Misioné,
M.
BECARIOS/AS:
Castilla del Río, S.; Granjel Piñeiro, E.; Matesanz Salgado, D.
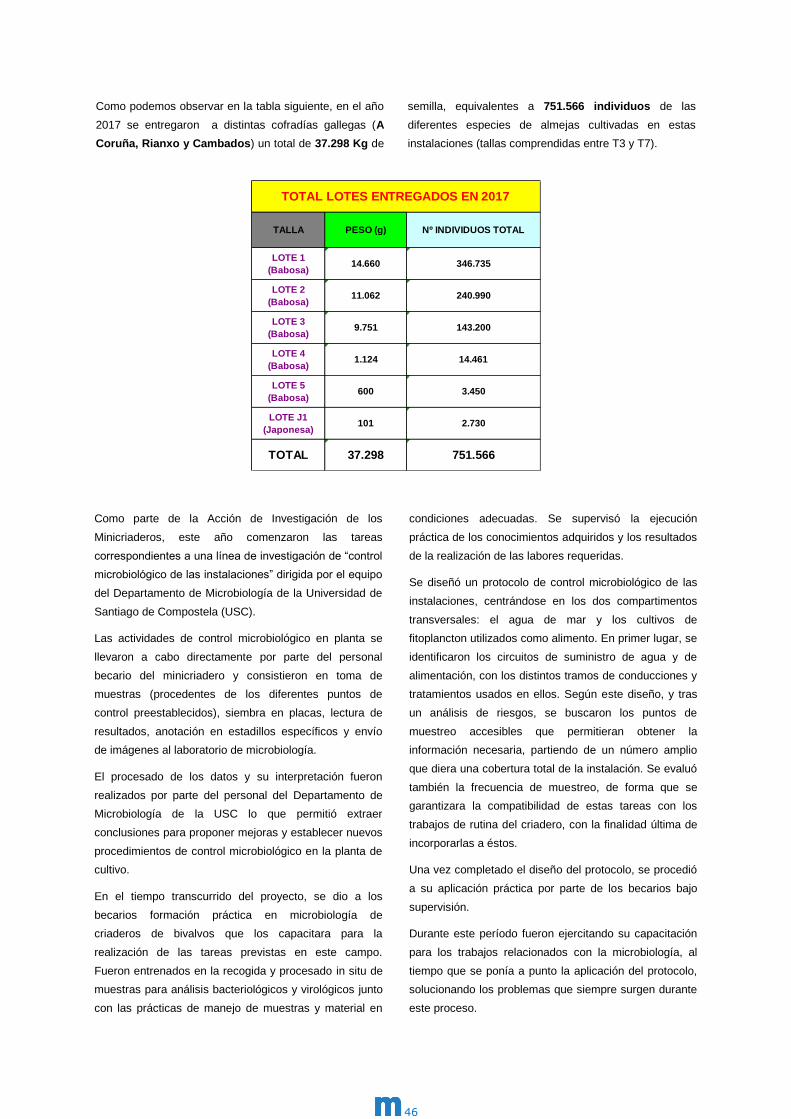
46
Como podemos observar en la tabla siguiente, en el año
2017 se entregaron a distintas cofradías gallegas (A
Coruña, Rianxo y Cambados) un total de 37.298 Kg de
semilla, equivalentes a 751.566 individuos de las
diferentes especies de almejas cultivadas en estas
instalaciones (tallas comprendidas entre T3 y T7).
Como parte de la Acción de Investigación de los
Minicriaderos, este año comenzaron las tareas
correspondientes a una línea de investigación de “control
microbiológico de las instalaciones” dirigida por el equipo
del Departamento de Microbiología de la Universidad de
Santiago de Compostela (USC).
Las actividades de control microbiológico en planta se
llevaron a cabo directamente por parte del personal
becario del minicriadero y consistieron en toma de
muestras (procedentes de los diferentes puntos de
control preestablecidos), siembra en placas, lectura de
resultados, anotación en estadillos específicos y envío
de imágenes al laboratorio de microbiología.
El procesado de los datos y su interpretación fueron
realizados por parte del personal del Departamento de
Microbiología de la USC lo que permitió extraer
conclusiones para proponer mejoras y establecer nuevos
procedimientos de control microbiológico en la planta de
cultivo.
En el tiempo transcurrido del proyecto, se dio a los
becarios formación práctica en microbiología de
criaderos de bivalvos que los capacitara para la
realización de las tareas previstas en este campo.
Fueron entrenados en la recogida y procesado in situ de
muestras para análisis bacteriológicos y virológicos junto
con las prácticas de manejo de muestras y material en
condiciones adecuadas. Se supervisó la ejecución
práctica de los conocimientos adquiridos y los resultados
de la realización de las labores requeridas.
Se diseñó un protocolo de control microbiológico de las
instalaciones, centrándose en los dos compartimentos
transversales: el agua de mar y los cultivos de
fitoplancton utilizados como alimento. En primer lugar, se
identificaron los circuitos de suministro de agua y de
alimentación, con los distintos tramos de conducciones y
tratamientos usados en ellos. Según este diseño, y tras
un análisis de riesgos, se buscaron los puntos de
muestreo accesibles que permitieran obtener la
información necesaria, partiendo de un número amplio
que diera una cobertura total de la instalación. Se evaluó
también la frecuencia de muestreo, de forma que se
garantizara la compatibilidad de estas tareas con los
trabajos de rutina del criadero, con la finalidad última de
incorporarlas a éstos.
Una vez completado el diseño del protocolo, se procedió
a su aplicación práctica por parte de los becarios bajo
supervisión.
Durante este período fueron ejercitando su capacitación
para los trabajos relacionados con la microbiología, al
tiempo que se ponía a punto la aplicación del protocolo,
solucionando los problemas que siempre surgen durante
este proceso.
TALLA PESO (g) Nº INDIVIDUOS TOTAL
LOTE 1
(Babosa)14.660 346.735
LOTE 2
(Babosa)11.062 240.990
LOTE 3
(Babosa)9.751 143.200
LOTE 4
(Babosa)1.124 14.461
LOTE 5
(Babosa)600 3.450
LOTE J1
(Japonesa)101 2.730
TOTAL 37.298 751.566
TOTAL LOTES ENTREGADOS EN 2017

CIMA Memoria 2017 47
Después de las etapas iniciales de ajustes, los
resultados obtenidos permitieron aportar información
para la mejora de la instalación en sí y de los protocolos
de manejo. El objetivo en todo caso es la evaluación
continua de los circuitos, que garantice su correcto
funcionamiento. Dado que la mayor parte de los
tratamientos utilizados en criaderos de bivalvos están
enfocados a un control de la microbiota, es
imprescindible, para evitar gastos inútiles y asegurar la
sostenibilidad, comprobar su eficacia. La información
obtenida resulta también de utilidad para la
comprobación interna de los protocolos de limpieza, de
tanta importancia en estas instalaciones. Además, aporta
información de los riesgos de problemas de producción
vinculados a potenciales patógenos bacterianos.
Avanzando un paso más, una vez en marcha el
protocolo de los circuitos de agua y microalgas se
comenzó con controles microbiológicos de los desoves
obtenidos en las instalaciones, apartado este en
desarrollo.
Se realizaron controles histopatológicos de diversos lotes
de almeja babosa (Venerupis corrugata), utilizados como
reproductores en el minicriadero del IGAFA.
Estos controles se efectuaron sobre un total de 4 lotes
de almeja babosa, 3 procedentes de la zona de O
Bohido, y 1 lote también procedente de O Bohido, pero
con un periodo de unos 3 meses en la batea de
experimentación del IGAFA.
En cuanto al número total de almejas procesadas fue de
110 ejemplares. Este procesado de la muestra, conlleva
las mediciones y pesos correspondientes. A continuación
tras ser abiertas con el bisturí, se retira de cada una de
las almejas, un fragmento de vianda que contenga
manto, gónada, masa visceral, branquia y pie. Este
fragmento se fija con solución Davidson, se deshidrata
en alcoholes de gradación creciente, se aclara con xileno
y se realiza la impregnación e inclusión en parafina. Los
cortes de microtomo de 5 micras se tiñen con
hematoxilina de Harris y eosina. Las preparaciones
histológicas se examinan en microscopio óptico, para
determinar la posible presencia de organismos parásitos
y alteraciones patológicas.
En la actualidad, nos encontramos en una fase
intermedia de muestreos y de la observación
microscópica, de momento el principal problema
patológico detectado fue el G. Perkinsus, con
prevalencias de parasitación, que en algunos lotes que
pueden llegar al 40%, pero siempre con intensidad de
infestación baja.
Otras parasitologías entendidas como menores
detectadas son: ciliados, gregarinas del G. Nematopsis y
casos estrictamente puntuales de organismos tipo
Rickettsia.
Desarrollo de las tareas realizadas en el Centro de
Cultivos Marinos de Ribadeo
Durante el año 2017 se hizo el estudio del ciclo
gametogénico de dos especies de bivalvos como son la
almeja bicuda (Venerupis aurea) y berberecho
(Cerastoderma edule). De la almeja bicuda se recogieron
muestras de reproductores durante todo un año con una
periodicidad mensual para procesarlas y hacer el estudio
histológico de la gónada para determinar su ciclo
reproductivo. Las muestras proceden de Cangas do
Morrazo y los resultados indican que la etapa de
madurez de esta especie corresponde con los meses de
primavera y verano, coincidiendo esto con los resultados
alcanzados por otros autores para esta especie.
Por lo que se refiere al berberecho, no se pudo
completar el ciclo. Las muestras de los reproductores
procedían de Noia pero sólo se pudieron conseguir
muestras durante cuatro meses, con lo cual fue
imposible completar un año de muestreo que sería lo
mínimo necesario para elaborar el esquema del ciclo
reproductivo.
Durante el año 2017 se realizó el cultivo de las diferentes
especies de bivalvos que se llevan a cabo en el
desarrollo de esta acción; se trabajó con las siguientes
especies: almeja babosa, fina, japonesa, rubia y bicuda;
coquina y navaja.
Se trata de establecer un protocolo de cultivo estándar
estrapolable a cualquier criadero de moluscos bivalvos
en el que se intenta diversificar las especies, para
mantener activo el criadero a lo largo de todo el año. Por
eso especies como la almeja babosa, bicuda, rubia y
navaja se cultivaron sobre todo en los meses de otoño,
invierno y primavera; la coquina durante la primavera y el
verano y dejando sólo para los meses de verano el
cultivo de almeja fina y japonesa.
Aunque según los resultados del ciclo gametogénico de
la almeja bicuda la madurez es sobre todo en primavera
y verano, en el criadero conseguimos desoves ya desde
el mes de febrero y hasta el mes de mayo. La duración
media del cultivo larvario hasta la fijación para esta
especie fue de 15 días y la supervivencia media del 55%.

48
El cultivo postlarvario se realizó en el criadero (hasta un
tamaño de 2 mm) y en el invernadero (hasta un tamaño
de 5 mm). El preengorde de semilla entre 5 y 10 mm se
llevó a cabo en una batea experimental que está
fondeada en el puerto de Ribadeo y la semilla obtenida
se llevó para Cangas do Morrazo ya que la cofradía tenía
interés en recuperar un banco natural con esta especie.
En el año 2017 se entregaron 72.000 unidades con una
talla media de 10 mm. La mayor parte de esta entrega
fue semilla obtenida de los desoves de 2016, ya que esta
especie precisa de varios meses para alcanzar la talla de
10 mm y en los meses de noviembre hasta abril el
crecimiento en la batea fue solo del 5-10%, mientras que
durante la primavera y verano el aumento en talla está
entre un 30-40% mensual.
El cultivo de la navaja se llevó a cabo entre los meses de
febrero y abril, la supervivencia durante el cultivo larvario
osciló entre un mínimo del 29% y un máximo del 78%. El
preengorde de semilla a partir de 8-10 mm se hace
colocando la semilla en un sustrato arenoso y cuando
alcanzan la talla media de 15 mm ya se llevan al medio
natural, la siembra la llevaron a cabo buzos-navajeros de
la cofradía de Cangas, para reforzar la producción de los
bancos naturales, se entregaron 63.000 unidades en
2017. Esta especie alcanza la talla de 15 mm en unos
seis meses desde el desove así que al desovar entre
febrero y abril se pueden trasladar al medio natural a
finales del verano o principios de otoño del mismo año.
El cultivo de la almeja rubia resulta más complicado que
las otras especies, siendo ya más difícil obtener
desoves. En el mes de febrero hicimos el seguimiento de
un cultivo larvario y aunque la supervivencia del cultivo
larvario fue del 50%, tuvimos altas mortalidades en el
preengorde de la semilla.
Para los cultivos de coquina empleamos reproductores
de Vilarrube (Valdoviño-A Coruña), los desoves se
llevaron a cabo entre los meses de febrero y junio, esta
especie tiene también un cultivo difícil con altas
mortalidades ya que es especialmente sensible a los
efectos de bacterias del género vibrio. Los episodios de
mortalidades están siempre asociados con la
proliferación de estas bacterias. Para minimizar estos
efectos se llevan a cabo estrictos controles
microbiológicos de todo el criadero (agua, fitoplancton y
larvas) haciendo siembras en placas con medio de
cultivo específico para los vibrios todas las semanas.
De los cultivos llevados a cabo de las almejas fina,
babosa y japonesa en el año 2016 se entregaron durante
el año 2017 a diferentes cofradías, partidas de semilla de
estas almejas para su engorde en el medio natural. De
almeja babosa fueron 450.000 unidades repartidas en
las cofradías de A Coruña, Noia y San Cosme de
Barreiros.
De almeja fina 400.000 unidades para las cofradías de
San Cosme, Baldaio, Noia, A Coruña, Vicedo, Cedeira,
Barqueiro y Espasante. De almeja japonesa fueron
400.000 unidades a las cofradías de Ribadeo, San
Cosme, Vicedo, Barqueiro y Espasante.
Parte de esta semilla fue preengordada en el
minicriadero de O Vicedo, en sistema de cubos con
aireación forzada que están colocados en un pantalán
del puerto. La semilla se trasladó desde el criadero de
Ribadeo y una vez que alcanzó la talla necesaria para la
siembra en el medio natural fue entregada a las
cofradías de la zona Vicedo, Barqueiro, Espasante y
Cedeira.

CIMA Memoria 2017 49
6 PATOLOGÍA

50
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Estudio epidemiológico de la almeja rubia (Venerupis
rhomboides) en Galicia afectada por colonias procariotas
intracelulares (CPI). Identificación de las CPI.
Seguimiento de la estructura de tallas de poblaciones de
almeja rubia de 4 bancos de la ría de Vigo.
INICIO: octubre, 2016
FINALIZACIÓN: octubre, 2017
ENTIDAD FINANCIADORA:
Consellería do Mar. Xunta de Galicia.
Código: CIMA 16/01/13/04
INVESTIGADORA PRINCIPAL:
López Gómez, C.
INVESTIGADORES/AS:
CIMA: Febrero Mayor, F.M.; No Couto, E.
Xefatura Territorial CM Vigo: Rodríguez González,
L.M.
INTECMAR: Darriba Couñago, S.
APOYO A LA INVESTIGACIÓN:
Brianes Veiras, M.J.; Meléndez Ramos, M.I.
BECARIO:
Grande Ocampo, O.
RESUMEN
En la presente acción se planteó el desarrollo de un
patológico que pretendía la identificación de las colonias
de procariotas intracelulares (CPI) detectadas en los
estudios anteriores (Darriba et al., 2012; Informe CIMA
2012) como agente patógeno asociado a episodios de
mortalidad y continuar el estudio epidemiológico iniciado
con posterioridad a las mortalidades en la ría de Vigo y
Cabío (ría de Arousa). Así como el seguimiento de la
estructura de tallas de poblaciones de almeja rubia de 4
bancos de esta ría (Cabo de Mar, Tirán, Liméns e Islas
Cíes).
Se presenta, a continuación, un resumen de los
resultados obtenidos por objetivos.
Objetivo 1. Análisis de la población
En otoño de 2013, sólo los bancos de Cabo de Mar e
Islas Cíes presentaron una estructura de tallas
aparentemente equilibrada, entendida esta como aquella
en la que la abundancia de individuos menores de 40
mm y la frecuencia de individuos de las nuevas cohortes
es mayor que la de individuos comerciales (>= 40 mm) y
que, por lo tanto, permita una renovación eficaz de la
fracción de la población objeto de explotación. El resto
de los bancos estudiados presentaba una estructura de
tallas no equilibrada. A partir del otoño de 2015, en cada
uno de los distintos bancos de la ría de Vigo, la
estructura de tallas empieza a estar equilibrada, lo que
permitiría pensar en una mejora en la dinámica de la
población, si no existiese explotación de tallas ilegales
para alcanzar los topes.
En la figura 1, donde la totalidad de la población de
almeja rubia de la ría de Vigo se considera como un
único banco, la estructura de tallas se observa también
equilibrada a partir del otoño de 2015.
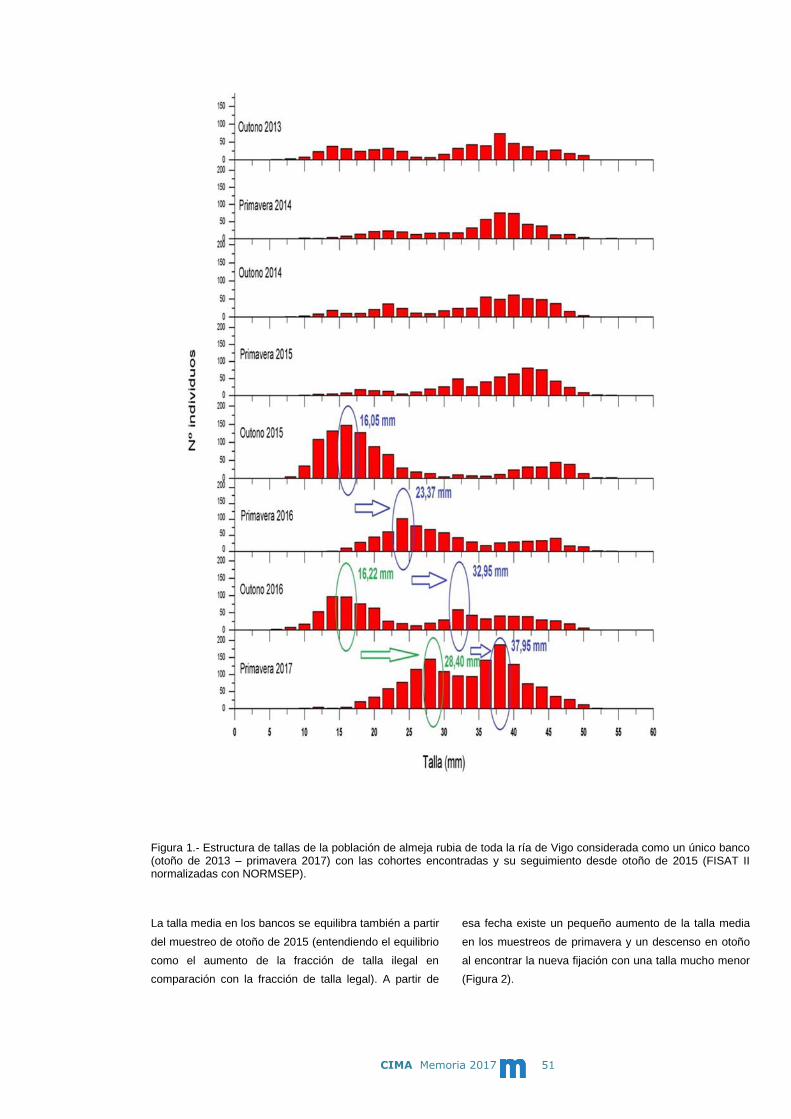
CIMA Memoria 2017 51
Figura 1.- Estructura de tallas de la población de almeja rubia de toda la ría de Vigo considerada como un único banco (otoño de 2013 – primavera 2017) con las cohortes encontradas y su seguimiento desde otoño de 2015 (FISAT II normalizadas con NORMSEP).
La talla media en los bancos se equilibra también a partir
del muestreo de otoño de 2015 (entendiendo el equilibrio
como el aumento de la fracción de talla ilegal en
comparación con la fracción de talla legal). A partir de
esa fecha existe un pequeño aumento de la talla media
en los muestreos de primavera y un descenso en otoño
al encontrar la nueva fijación con una talla mucho menor
(Figura 2).
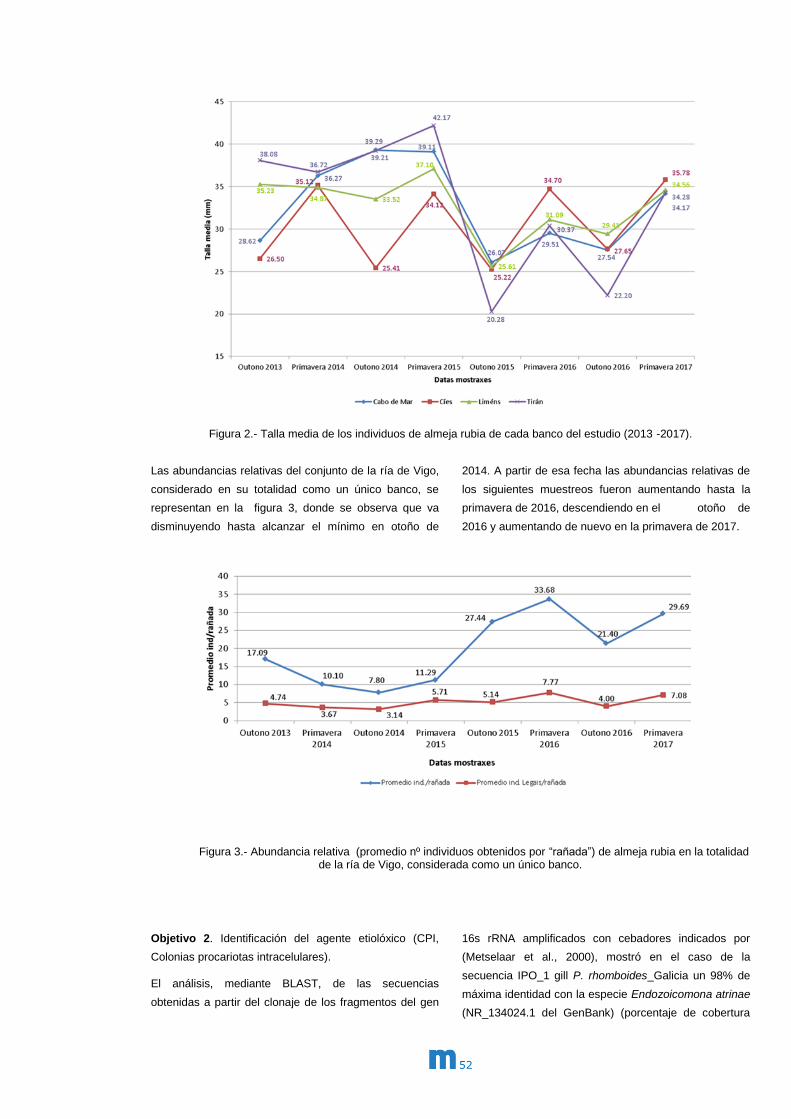
52
Figura 2.- Talla media de los individuos de almeja rubia de cada banco del estudio (2013 -2017).
Las abundancias relativas del conjunto de la ría de Vigo,
considerado en su totalidad como un único banco, se
representan en la figura 3, donde se observa que va
disminuyendo hasta alcanzar el mínimo en otoño de
2014. A partir de esa fecha las abundancias relativas de
los siguientes muestreos fueron aumentando hasta la
primavera de 2016, descendiendo en el otoño de
2016 y aumentando de nuevo en la primavera de 2017.
Figura 3.- Abundancia relativa (promedio nº individuos obtenidos por “rañada”) de almeja rubia en la totalidad de la ría de Vigo, considerada como un único banco.
Objetivo 2. Identificación del agente etiolóxico (CPI,
Colonias procariotas intracelulares).
El análisis, mediante BLAST, de las secuencias
obtenidas a partir del clonaje de los fragmentos del gen
16s rRNA amplificados con cebadores indicados por
(Metselaar et al., 2000), mostró en el caso de la
secuencia IPO_1 gill P. rhomboides_Galicia un 98% de
máxima identidad con la especie Endozoicomona atrinae
(NR_134024.1 del GenBank) (porcentaje de cobertura

CIMA Memoria 2017 53
del 100%) y la secuencia IPO_2 gill P.
rhomboides_Galicia un 97% de máxima identidad con
distintas secuencias de Candidatus Endonucleobacter
bathymodioli encontradas en el GenBank. Estas
bacterias pertenecen al grupo Gammaproteobacteria
subgrupo Proteobacteria, y el Candidatus
Endonucleobacter bathymodioli también está incluído en
la familia Endozoicomonaceae
(http://www.uniprot.org/taxonomy). Datos similares fueron
observados por Cano et al. (2018) para un organismo
procariota intracelular de la glándula digestiva y
branquia de la vieira (Pecten maximus) en Lyme Bay
(Sudeste de Inglaterra), caracterizando ese organismo
como del tipo-Endozoicomona (KX780138 do GenBank).
Objetivo 3. Observación al microscopio óptico de las
preparaciones histológicas recogidas en la ría de Vigo.
En los bancos de Liméns, Cíes y Tirán de la ría de Vigo
se observan prevalencias muy altas (en torno al 100%)
de CPI1-Br en todas las fechas. Las intensidades medias
de infección (Imi) (barras rojas) fueron moderadas (más
de 100 col./10 c.) en Liméns 2014 y 2015, y en Cíes en
2012, siendo el resto bajas (menos de 100 col./10 c.) o
muy bajas (menos de 50 col./10 c.). En lo que respecta a
las CPI2-Br, las prevalencias fueron variables (línea azul)
y las Imi muy bajas en los distintos bancos.
En el banco de Cabo de Mar (ría de Vigo), donde se
registró el episodio de mortandad en el año 2010, las
prevalencias de las CPI1-Br igualmente fueron muy altas
durante todo el período de estudio. La intensidad media
(Imi) fue muy alta durante el evento de mortandad en el
año 2010 (más de 300 col./10 c.) y alta (más de 200 col.
/10 c.) a lo largo de 2011. Se observó un descenso
progresivo hasta alcanzar en la actualidade valores muy
bajos (muy inferiores a 50 col./10 c.). En lo que respecta
a las prevalencias de CPI2-Br (línea azul), éstas fueron
muy altas igualmente en el año 2010 y 2011,
observándose un descenso progresivo hasta la
actualidad. La Imi detectada en el 2010 (mortandad) fue
baja aunque registró el recuento más alto de todos los
bancos analizados (23 col./10 c.) y a partir de 2011 llega
a alcanzar valores muy bajos.
Objetivo 4. Estudio epidemiológico centrándose en los
agentes patógenos que fueron identificados como
posibles responsables de causar efectos deletéreos en la
almeja rubia.
En lo tocante a los organismos procariotas, hay que
resaltar que no se comentarán en este apartado, ya que
fueron evaluadas al detalle en el objetivo anterior. De los
demás procariotas, en lo que respecta a los quistes
bacterianos en branquia, en la ría de Vigo destacaron los
niveles elevados de prevalencias en los bancos de Tirán
(zona interna) y en Cabo de Mar (zona media sur),
mientras que los bancos de Liméns (zona media norte) e
Islas Cíes (zona externa) los niveles fueron inferiores. En
el banco de Cabío (ría de Arousa) se observa una
variabilidad en las prevalencias (17% al 100%). Para
evaluar estadísticamente estos resultados, se realizó un
análisis de las prevalencias globales por pares,
obteniéndose que los bancos de Cabo de Mar y Tirán
tienen las prevalencias significativamente más altas de
los bancos analizados, seguidos de Cabío con valores
significativamente mayores que Liméns y éste, a su vez,
tiene valores significativamente mayores que las Islas
Cíes, que alcanza niveles significativamente más bajos.
Las colonias procariotas intracelulares (CPI) observadas
en la glándula digestiva alcanzaron prevalencias
variables, bajas en general en todos los bancos. El
análisis de las prevalencias globales por pares mostró
que Cabo de Mar y Liméns presentaron, los valores
significativamente mayores, seguidos de Islas Cíes, Tirán
y Cabío. Del grupo de los protozoos, destacaron las
gregarinas del género Nematopsis, que se detectaron en
todas las muestras, con prevalencias altas. Aún así, las
intensidades fueron bajas en general, con casos
puntuales de intensidades más altas, y nunca
coincidentes con los episodios de mortandad. Además
no se observó que causaran daños en los tejidos.
Un protozoo del género Perkinsus fue detectado en
ambas rías, con prevalencias bajas en general y sin
mostrar estados en fase de multiplicación. Plasmodios,
de un supuesto haplosporidio, fueron observados en las
dos rías con prevalencias variables, así como coccidios
en el riñón, con prevalencias bajas en general.
Del grupo de los metazoos, lo más destacable son las
larvas de trematodos en fase de esporoquiste con
cercarias que en el caso de intensidades altas se
observó invasión de la gónada, produciendo castración.
Conclusiones:
1. Los episodios de mortandad que se dieron en
bancos de almeja rubia de la ría de Vigo, en 2010, se
debieron a un bloom de las colonias de organismos
procariotas intracelulares que se demuestra que son
simbiontes habituales en la branquia de esta especie de
almeja. Estos procariotas parecen ser

54
ganmaproteobacterias de la familia
Endozoicomonaceae, que incluye especies simbiontes
de un amplio abanico de organismos marinos. La causa
de ese importante incremento de la intensidad de
infección no fue identificada.
2. Estos episodios de mortandad supusieron una
caída muy relevante en la producción de ese año y de
los posteriores, lo que queda reflejado en la producción
de esta especie en Galicia. La intensidad media de
infección por las colonias parásitas (CPI1-Br) en el banco
de Cabo de Mar recupera valores normales a principios
de 2012 y ya en otoño de 2013 se registra en este banco
una estructura de tallas equilibrada. Es a partir del otoño
de 2015 cuando en toda la ría se observa un incremento
de la abundancia relativa de la población de almeja
rubia, coincidiendo con intensidades de infección de las
CPI1 bajas en todos los bancos analizados. Los
desequilibrios de la estructura de tallas entre 2013 y
antes del otoño de 2015 pueden tener como causa otros
factores distintos de las infecciones por las CPI1-Br, ya
que no en todos los casos se corresponden valores
bajos de las intensidades de infección con recuperación
del equilibrio.
3. Si no aparecen nuevos episodios de
mortandad, y siguiendo el patrón oscilante de la
extracción de almeja rubia a lo largo de los años, los
datos de este estudio apuntan una mejoría en la
producción de la almeja rubia en la ría de Vigo en un
futuro inmediato.

CIMA Memoria 2017 55
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Estudio de la eliminación de virus entéricos patógenos
en ostra mediante procesos de reinstalación y
depuración.
INICIO: octubre, 2016
FINALIZACIÓN: octubre, 2018
ENTIDAD FINANCIADORA:
Consellería do Mar. Xunta de Galicia.
Código: CIMA 16/08
INVESTIGADORA PRINCIPAL:
López Gómez, C.
INVESTIGADORES/AS:
CIMA: Febrero Mayor, F.M.
INTECMAR: Álavares Álvarez, C.; Darriba Couñago,
S.
USC: López Romalde, J.
APOYO A LA INVESTIGACIÓN:
CIMA: Meléndez Ramos, M.I.
INTECMAR: Rodríguez Ruibal, L.
BECARIO:
CIMA: Grande Ocampo, O.
RESUMEN
El proyecto tiene como objetivos, por una parte, el
establecimiento de un plan integrado de vigilancia que
reduzca de forma significativa los efectos del riesgo de
contaminación microbiológica y viral durante la
producción primaria de las ostras comerciales producidas
en Galicia, enfocando el esfuerzo en el estudio de la
efectividad de la depuración en la eliminación de estos
virus.
Por otra parte, el proyecto pretende establecer la
capacidad infectiva de estos virus después del proceso
de depuración. Este parámetro nos acercará a
establecer el riesgo real de estos patógenos, ya que no
todas las partículas virales detectadas por las técnicas
de cuantificación actuales son infectivas.
El plan de trabajo establecido fue el siguiente:
1) Selección de las zonas a estudio
Se escogieron como zonas representativas de las zonas
de producción dos zonas que dispongan de parques de
cultivo de bateas y de bancos de infaunales, y que
dispongan de las dos especies comerciales de ostra,
Ostrea edulis y Crasssotrea gigas.
Ostrea edulis: la zona elegida fue el polígono Cambados
D (ría de Arousa) (Figura 1).
Crasssotrea gigas: Se eligió la ría de O Barqueiro por
ser la zona donde hay abundancia de esta especie de
ostra.
Como zona de reinstalación se escogió la ría de Aldán
(Figura 2).
2) Diseño del plan de muestreo
Se diseñaron 10 experiencias de
reinstalación/depuración con cada especie de ostra. La
frecuencia de muestreo durante la reinstalación quedó
fijada con el siguiente régimen: días 0 (antes de llevar a
la zona de reinstalación), 15, 22 y 30. El día 30,
dependiendo de los niveles de contaminación, las ostras
se trasladan a la depuradora experimental en el CIBUS
(Universidad de Santiago de Compostela) para su
depuración y se realizan análisis los días 32 (48h de
depuración) y 37 (1 semana de depuración).
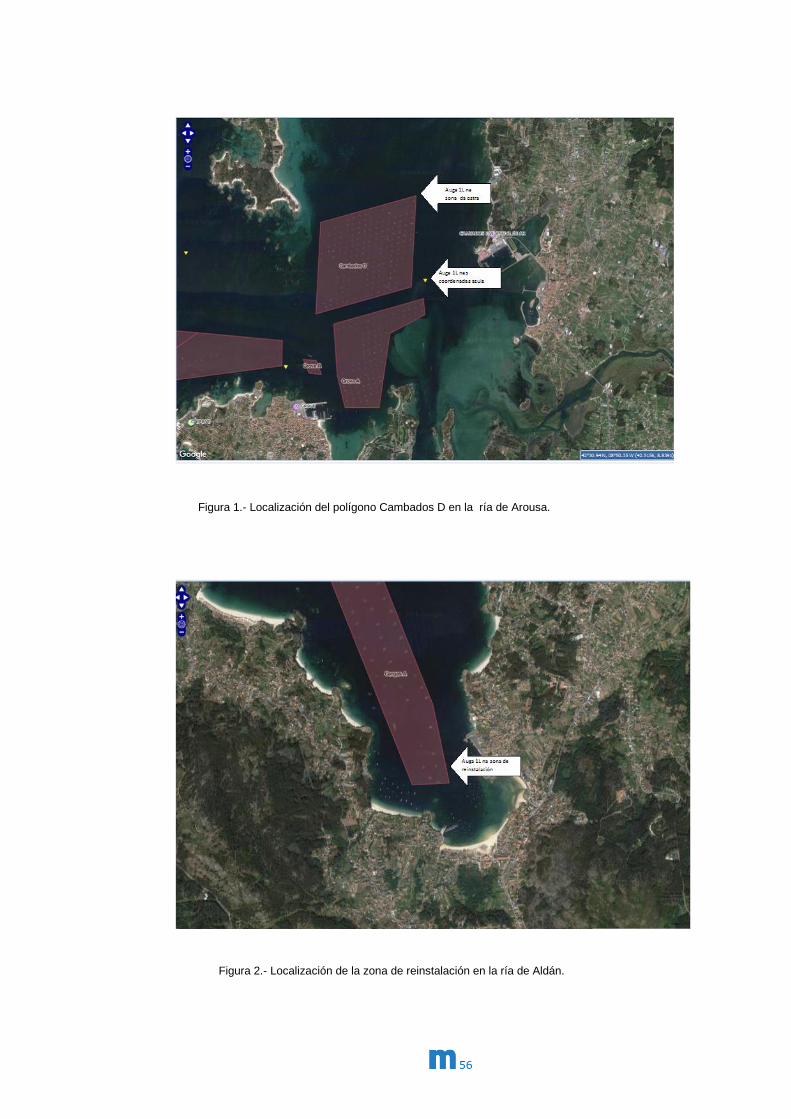
56
Figura 1.- Localización del polígono Cambados D en la ría de Arousa.
Figura 2.- Localización de la zona de reinstalación en la ría de Aldán.

CIMA Memoria 2017 57
Se están recogiendo de modo paralelo dos muestras de
agua en la zona de origen de la ostra, en un punto
próximo al emisario de la EDAR más influyente en la
zona y en el punto de recogida de la muestra, así como
una muestra de agua en la zona de reinstalación.
3) Transporte y recogida de las muestras
La reinstalación y recogida de las muestras, coordinada
por el CETMAR, está siendo realizada por personal
cualificado, así como por personal contratado
específicamente para este trabajo.
El traslado del producto se realizará siguiendo el método
Coleman de refrigeración según se realiza en la Unidad
de Microbiología y Virología del Intecmar. Las muestras
de agua se recogerán en botes estériles y se
conservarán también en refrigeración hasta el momento
del ensayo.
4) Realización de ensayos
En el año 2016 se realizaron dos experiencias, la última
finalizada en 2017, año en el que se realizaron otras dos
experiencias con ostra plana y una con ostra japonesa.
Se realizaron además dos experiencias de
bioacumulación en el mes de junio de 2017, con ambas
especies debido a la baja carga viral ambiental detectada
en las experiencias realizadas hasta la fecha y en
sustitución de la 5ª experiencia de ostra plana que fue
anulada.
A) Carga bacteriana de E. coli / 100 g en el molusco
Cada fecha de muestreo se lleva al INTECMAR una
parte de la muestra para el cálculo de carga bacteriana
mediante la técnica del número más probable (NMP).
Los resultados obtenidos hasta el momento son
coherentes con los históricos tanto de la zona de
producción como de la zona de reinstalación, y no
indican eventos de contaminación extraordinarios en
dichas zonas.
B) Presencia de virus entéricos (Norovirus y hepatitis A
HAV)
Para la detección de virus entéricos se emplea la técnica
de reacción en cadena de la polimerasa en tiempo real
(RT-PCR) con sondas TaqMan siguiendo las
recomendaciones de la norma ISO/TS 15216-1:2013.
Estos análisis se están realizando en paralelo en los
laboratorios de la Universidad de Santiago de
Compostela y del INTECMAR, como prueba de ensayo
interlaboratorio.
Hasta el momento se detectaron en ostra plana en las
fechas de febrero de 2017 Noroviru GI y GII con 40 y 711
copias/g respectivamente y en abril de 2017 Norovirus
GII con 18 copias/g; la procedencia de ambas muestras
es una zona de producción de origen, el polígono
Cambados D. En ostra japonesa, en la experiencia de
noviembre se detectan los dos virus Norovirus GI y GII
en la muestra inicial de origen y en la muestra depurada
en reinstalación después de 21 días; se detectó
Norovirus GII en todas las muestras de esta experiencia.
También se detectó HAV, a nivel inferior al límite de
cuantificación, en la muestra de agua recogida en el
polígono de origen, Cambados D.
Cabe destacar lo atípico de la meteorología durante los
meses de otoño y comienzo de invierno con pocas
lluvias lo que podría ser un factor importante para la no
presencia de los virus en las muestras de ostra y de
agua.
C) Estudio histológico e inmunohistoquímico
Así mismo, se recogieron muestras para llevar a cabo el
estudio histológico e inmunohistoquímico mediante
técnicas clásicas en tejidos de ostra en origen, después
de la reinstalación y después de la depuración, con el fin
de determinar la presencia de partículas virales en los
tejidos. La negatividad de las muestras por otros
métodos hizo innecesario el análisis de las muestras
aunque estas fueron procesadas para el estudio
histológico.

58

CIMA Memoria 2017 59
PROYECTO DE INVESTIGACIÓN EQUIPO
TÍTULO:
Poblaciones bacterianas en criadero de bivalvos.
Procesos de patogénesis
INICIO: junio, 2015
FINALIZACIÓN: diciembre, 2017
ENTIDAD FINANCIADORA:
Ministerio de Economía y Competitividad
INVESTIGADOR PRINCIPAL:
USC: Barja Pérez, J.L.
INVESTIGADORES/AS:
CIMA: Montes Pérez, J.; Novoa Vázquez, S.; Ojea
Martínez, J.
USC: Prado Plana, S.; Dubert Pérez, J.
APOYO A LA INVESTIGACIÓN:
CIMA: Bouzamayor Yáñez, M.V.; Brianes Beiras, M.J.;
Cotarelo Jardón, M.J.; Meléndez Ramos, I.; Páez Pérez,
F.J.; Penas Pampín; E.; Rúa Santervás, P.; Ruíz Misioné,
M.
USC: Núñez García, S.
BECARIOS/AS:
Castilla del Río, S.; Matesanz Salgado, D.; Ulla Refojo, L
RESUMEN
El proyecto “Poblaciones bacterianas en criaderos de
bivalvos: Procesos de patogénesis” avanzó en su
desarrollo durante esta anualidad, obteniendo además
una prórroga para terminar tareas previstas durante el
año 2018.
Dado el objetivo general de avanzar en el conocimiento
de los procesos de infección bacteriana limitantes de la
producción en criaderos, y el interés en obtener
información realista y alcanzar resultados prácticos, una
parte importante de los trabajos se llevó a cabo en
colaboración directa entre la USC y el CIMA, en el
criadero del Centro de Cultivos Mariños de Ribadeo.
Para obtener nuevos conocimientos de la dinámica de
los patógenos bacterianos en el criadero, se estudiaron
las interacciones de los patógenos larvarios con los
cultivos de fitoplancton de diferentes especies usados
como alimento. Bacterias del género Vibrio constituyen
una importante amenaza para la producción de bivalvos
en criadero, contando el grupo de trabajo con una
colección de patógenos demostrados e identificados
como miembros de distintas especies. Por otro lado, en
fases anteriores quedó establecido que el fitoplancton es
una de las principales fuentes de bacterias en los
cultivos en criadero, siendo los vibrios componentes
ocasionales de su microbiota, con una abundancia y
diversidad influidas por factores como temperatura o
requerimientos de producción. El paso siguiente fue
estudiar específicamente la supervivencia de vibrios
patógenos en los cultivos de fitoplancton de la
instalación. En una primera aproximación, se ensayó en
cultivos a pequeña escala de las siete especies de
microalgas que forman parte de la dieta habitual del
criadero. El diseño permitió un chequeo con un enfoque
práctico para evaluar simultáneamente en la planta la
capacidad de supervivencia de distintos patógenos. Esta
quedó demostrada, pero también la capacidad de
algunas de las especies de microalgas ensayadas de
inhibir in vivo a estas bacterias. La demostración de esta
actividad in vivo constituye un resultado novedoso y abre
una interesante vía de estudio. Como resultados más
destacables, Tetraselmis suecica fue la especie de
microalga con un mayor potencial inhibidor, mientras
Vibrio ostreicida fue el patógeno más sensible a distintas
microalgas.
Los prometedores resultados pusieron de manifiesto la
necesidad de realizar ensayos específicos con las
diferentes especies de microalgas, en condiciones de
cultivo activo y a concentraciones que favorezcan la

60
competencia fitoplancton-vibrio, evaluando también el
posible efecto de la bacteria sobre el crecimiento
microalgal o una potencial patogenicidad. Se diseñaron
por tanto complejos experimentos in vivo en condiciones
de cultivo en la instalación. Los resultados obtenidos
hasta el momento permiten establecer que los vibrios
patógenos larvarios no lo son para las dos especies de
fitoplancton ensayadas, T. suecica e Isochrysis galbana.
En lo que respecta al efecto de las microalgas sobre los
patógenos, fue especialmente relevante la confirmación
de la capacidad de inhibición de algunos de ellos, no
sólo de T. suecica, como ya se observara, sino también
de I. galbana. Así, queda demostrado el riesgo de
transmisión de patógenos larvarios en cultivos de
microalgas que no se ven afectados por su presencia,
pero también el potencial del fitoplancton para el control
de la proliferación de vibrios en la instalación.
Otro aspecto que mencionar de los trabajos de esta
anualidad es el estudio de la formación de biopelículas
bacterianas por los vibrios patógenos en las superficies
de los tanques de cultivo, como forma de persistencia en
la instalación. Estas estructuras pueden hacer a los
microorganismos que las integran resistentes a
tratamientos como los antibióticos, la lejía o la
desecación, habituales como métodos de limpieza y
desinfección. Se abordó por ello un objetivo ambicioso
como es el estudio de la capacidad in vivo de los
diferentes patógenos larvarios para formar estas
biopelículas. Este trabajo implica una gran complejidad y
requiere de un cuidado extremo por la manipulación de
los patógenos en la instalación. Al mismo tiempo, es una
oportunidad de obtener información realista, en las
condiciones ambientales del criadero. Durante esta
anualidad se puso a punto todo el procedimiento,
ensayándose diferentes métodos para evaluar la
formación de las biopelículas. Los resultados obtenidos
muestran la utilidad de la microscopía electrónica para la
visualización, una vez ajustado el protocolo más
apropiado para las condiciones particulares de los
experimentos. Los ensayos realizados hasta el momento
parecen confirmar la capacidad de los patógenos, en
mayor o menor medida, de formación de biopelículas.
Hay que citar también los estudios de la microbiota
asociada a las fases iniciales del desarrollo de las
puestas y su relación con el éxito/fracaso del cultivo
larvario. Este punto trata de vincular los distintos
compartimentos del criadero y su importancia en la
microbiota de los cultivos. Supondría un avance en el
conocimiento de la dinámica de los patógenos en la
instalación y de los mecanismos de infección, en un
momento en el que tienen influencia los reproductores, el
agua, el alimento y los tanques de cultivo.
Además de los trabajos eminentemente prácticos que
vertebran la línea principal de actuación del proyecto, en
paralelo se continúa con los estudios genómicos de los
patógenos y los análisis bioinformáticos que permitan
identificar genes que puedan tener relación con la
patogenicidad.
El proyecto está demostrando la importancia de los
trabajos realizados in vivo para alcanzar resultados que
ayuden a una mejora real de los cultivos de bivalvos en
criadero. También está poniendo de manifiesto la
complejidad de estos trabajos, sólo posibles con la
implicación total del grupo investigador y de todo el
personal. En resumen, confirma la necesidad de una
orientación práctica para aplicar los conocimientos
científicos en beneficio de la producción.

CIMA Memoria 2017 61
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Epidemiología de la enfermedad causada por Marteilia
cochillia en las poblaciones de berberecho
Cerastoderma edule de Galicia.
INICIO: marzo, 2015
FINALIZACIÓN: diciembre, 2017
ENTIDAD FINANCIADORA:
Consellería do Mar. Xunta de Galicia.
Código: PGIDIT-CIMA 15/02
INVESTIGADORA PRINCIPAL:
Carballal Durán, M.J.
INVESTIGADORES/AS:
CIMA: Cao Hermida, A.; Iglesias Estepa, D.; No Couto,
E.; Villalba García, A.
IIM: González González, A.; Pascual del Hierro, S.
CETMAR: Abollo Rodríguez, E.
INTECMAR: Darriba Couñago, S.; Montero Vila, P.
Cofradía de Pescadores “San Antonio” de Cambados:
Mariño Balsa, J.C.
Delegación de Carril de la Consellería do Mar:
Fernández Barreiro, J.
APOYO A LA INVESTIGACIÓN:
Brianes Beiras, M.J.; García Alves, L.; González
Fontela, A.I.; Martínez Verde, G.; Meléndez Ramos, I.;
Penas Pampín, E.; Rúa Santervas, P.
BECARIA:
Fernández González, J.
RESUMEN
La marteiliosis es una enfermedad infecciosa originada
por parásitos del género Marteilia que afectan a
moluscos bivalvos de gran interés económico para la
acuicultura europea. La marteiliosis se detectó por
primera vez en Cerastoderma edule en 1975 en Francia
(Comps et al., 1975). En España se observó en 2008 en
berberechos del Delta del Ebro, con un 40% de
prevalencia, asociada a un episodio de elevada
mortalidad (Carrasco et al., 2011). En Galicia se detectó
por primera vez en febrero de 2012 en berberechos C.
edule de Lombos do Ulla asociada a una mortalidad
masiva del hospedador. En los meses siguientes, la
enfermedad se observó en otros bancos de berberechos
de la ría de Arousa causando intensas mortalidades. El
parásito que causa la marteiliosis en C. edule de Galicia
fue identificado como M. cochillia que es la misma
especie que afecta a los berberechos del Delta del Ebro
(Carrasco et al., 2013; Villalba et al., 2014).
En los años 2013-2014 desarrollamos la Acción de
Investigación “Estudio da marteliose que afecta a os
berberechos Cerastoderma edule de Galicia” (PGIDIT-
CIMA 13/03). En este proyecto de investigación se llevó
a cabo un programa de vigilancia de la enfermedad en
las rías limítrofes a la ya afectada, lo que permitió la
detección del parásito en la ría de Pontevedra en 2013 y
en bancos de la ría de Vigo en 2014. En esos años se
registró un nuevo brote de M. cochillia que afectó a todos
los bancos de berberecho de la ría de Arousa, aunque
con diferente intensidad.
En marzo de 2015 se inició este nuevo proyecto cuyo
objetivo general fue la adquisición de conocimientos
sobre la enfermedad causada por el parásito Marteilia
cochillia en las poblaciones de Cerastoderma edule de
Galicia, para facilitar la toma de decisiones en la gestión
del recurso y, si es el caso, el desarrollo de estrategias
de lucha que conduzcan a la mitigación de sus efectos.
Este estudio incluyó aspectos epidemiológicos y
ambientales y el conocimiento del ciclo de vida del
parásito. El proyecto desarrolló los siguientes objetivos
específicos.
Objetivo 1. Desarrollar un programa activo de vigilancia
de la enfermedad en poblaciones de berberecho aún no
afectadas, con el fin de conocer el estatus sanitario de
las poblaciones.

62
Se recogieron muestras de berberechos C. edule de
cada población seleccionada en dos ocasiones:
primavera y otoño de los años 2015 y 2016. Los bancos
seleccionados son los que figuran en la red de muestreo
del programa oficial de control patológico de moluscos
bivalvos, que desarrolla la Unidad de Patología del
INTECMAR. Debido a las mortalidades causadas por la
marteiliosis, en el muestreo de primavera de 2015 no fue
posible obtener un número suficiente de individuos de C.
edule en algunos bancos y, como alternativa, se
analizaron berberechos de la especie C. glaucum. Al
inicio del proyecto, la marteiliosis afectaba a las
poblaciones de berberecho de la ría de Arousa, ría de
Pontevedra y ría Vigo. Ante el riesgo de propagación de
la enfermedad a la ría de Noia, que es la de mayor
producción de berberecho de Galicia, se realizó un
programa de vigilancia intensivo en esta ría, en el que se
hicieron muestreos con periodicidad mensual el primer
año del proyecto, y después bimestral. Para todos estos
estudios, se recogieron 30 berberechos de cada banco y
se realizaron cortes histológicos de los principales
tejidos de cada individuo. Las secciones histológicas se
analizaron al microscopio óptico para determinar si
estaban o no afectados por el parásito. Los resultados de
este estudio mostraron que en los berberechos C. edule
de las rías altas (O Barqueiro, Espasante, Barrallobre,
Miño, Anllóns, Camariñas), los de la ría de Muros-Noia y
los de Baiona no se observaba M. cochillia. Por el
contrario, los berberechos de la ría de Arousa (A Pobra,
Cabo de Cruz, Lombos do Ulla, Cambados), Pontevedra
(Combarro) y Vigo (Moaña, Redondela) si estaban
parasitados. La especie C. glaucum no estaba afectada
por esta parasitosis a pesar de convivir con berberechos
C. edule intensamente parasitados por la marteiliosis.
Objetivo 2. Estudio de la dinámica temporal de la
enfermedad y evaluación de la influencia de variables
ambientales en el desarrollo de la misma.
Durante los años 2015-2017 se recogieron
mensualmente muestras de berberecho en dos
importantes bancos de la ría de Arousa: Lombos do Ulla,
banco mayoritariamente submareal situado en la zona
más interna de la ría y muy influenciado por el río Ulla, y
O Sarrido, banco intermareal situado en el interior de la
ensenada de O Grove en la desembocadura del río
Umia. El tamaño de muestra fue de 30 berberechos y de
cada individuo se realizaron cortes histológicos que
fueron analizados al microscopio óptico con el fin de
diagnosticar la presencia de Marteilia cochillia. Se
calculó la prevalencia mensual del parásito en cada
banco como el porcentaje de berberechos afectados por
M. cochillia del total de analizados. Se evaluó, asimismo,
la mortalidad contabilizando, cada mes, el número de
berberechos supervivientes del total de individuos que
previamente se habían dispuesto en cajas de control que
se situaban enterradas en el substrato, en el caso del
banco intermareal de O Sarrido, y en una estructura
dispuesta sobre el fondo, que se recuperaba con ayuda
de una embarcación dotada de grúa, en el caso del
banco submareal de Lombos do Ulla.
En Lombos do Ulla se detectó en octubre un nuevo brote
de M. cochillia en la cohorte de berberechos de 2015 con
una prevalencia inicial del 80%. Este brote siguió un
modelo de desarrollo rápido al igual que los brotes de
años anteriores, de manera que la prevalencia y
mortalidad aumentaron progresivamente hasta provocar
la extinción de la cohorte de berberecho reclutada en
2015 en abril de 2016. En julio de 2016 se observa
fijación de la nueva cohorte que se infecta en octubre
con una prevalencia inicial del 3%. El desarrollo de este
nuevo brote sigue un modelo similar al del 2015, y en
abril 2016 se produce la desaparición de esta cohorte.
En junio de 2017 se fijan los nuevos reclutas con una
prevalencia inicial del 43%. A lo largo del tiempo, la
prevalencia de este nuevo brote desciende y la
mortalidad mensual también desciende de acuerdo con
la progresión de la enfermedad. No se observa una
influencia de la temperatura ni de la salinidad en el
desarrollo de la enfermedad (Fig.1).

CIMA Memoria 2017 63
En O Sarrido, la enfermedad no se detectó en la cohorte
de berberechos de 2015 hasta febrero de 2016, con una
prevalencia inicial del 37,9%. Se recogieron muestras de
individuos de esta cohorte hasta junio de 2016, momento
en la que quedó prácticamente agotada. En julio de 2016
se detecta fijación de berberecho y los primeros casos
de infección por M. cochillia se observaron en diciembre
con una prevalencia del 3,4%. El desarrollo de este brote
es más lento que el del año 2015 y se pudieron recoger
individuos de esta cohorte durante todo el año. En julio
de 2017 se fijan los reclutas de la nueva cohorte que en
agosto ya está infectada. La progresión de la
enfermedad es similar en estas cohortes de 2016 y 2017,
incrementándose la prevalencia hasta alcanzar un valor
máximo del 75,9% en noviembre. No se observa una
influencia de la temperatura ni de la salinidad en la
progresión de la enfermedad (Fig.2).
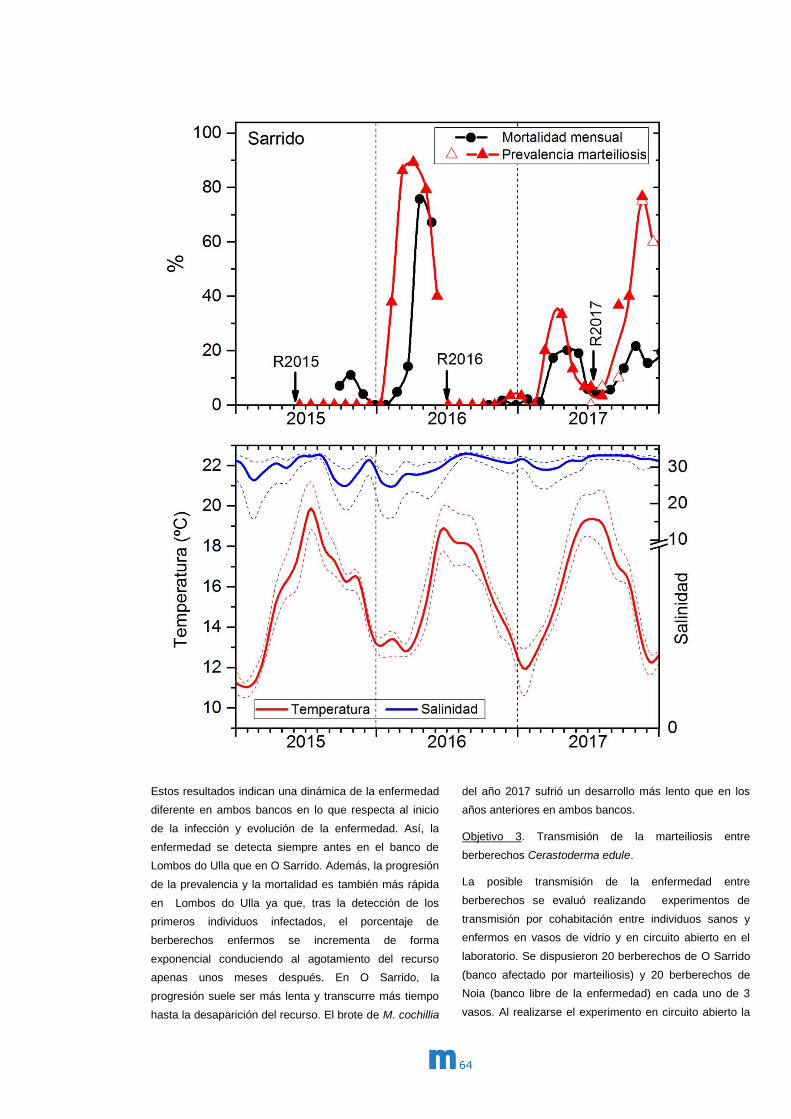
64
Estos resultados indican una dinámica de la enfermedad
diferente en ambos bancos en lo que respecta al inicio
de la infección y evolución de la enfermedad. Así, la
enfermedad se detecta siempre antes en el banco de
Lombos do Ulla que en O Sarrido. Además, la progresión
de la prevalencia y la mortalidad es también más rápida
en Lombos do Ulla ya que, tras la detección de los
primeros individuos infectados, el porcentaje de
berberechos enfermos se incrementa de forma
exponencial conduciendo al agotamiento del recurso
apenas unos meses después. En O Sarrido, la
progresión suele ser más lenta y transcurre más tiempo
hasta la desaparición del recurso. El brote de M. cochillia
del año 2017 sufrió un desarrollo más lento que en los
años anteriores en ambos bancos.
Objetivo 3. Transmisión de la marteiliosis entre
berberechos Cerastoderma edule.
La posible transmisión de la enfermedad entre
berberechos se evaluó realizando experimentos de
transmisión por cohabitación entre individuos sanos y
enfermos en vasos de vidrio y en circuito abierto en el
laboratorio. Se dispusieron 20 berberechos de O Sarrido
(banco afectado por marteiliosis) y 20 berberechos de
Noia (banco libre de la enfermedad) en cada uno de 3
vasos. Al realizarse el experimento en circuito abierto la

CIMA Memoria 2017 65
enfermedad podía transmitirse a través del agua, y por
ello se dispusieron 40 berberechos de Noia en cada uno
de 3 vasos que se usaron como control. Los berberechos
que se morían se eliminaban, y se hacían cortes
histológicos para evaluar la presencia del parásito. El
experimento finalizó a los 71 días de su inicio, y la
enfermedad no se transmitió entre berberechos. En los
berberechos de los vasos control tampoco se observó
marteiliosis. Estos resultados confirman los de otro
experimento de transmisión en tanques que realizamos
previamente, y señalan hacia la existencia de un ciclo de
vida complejo en M. cochillia con varios hospedadores
intermediarios al igual que sucede en M. refringens y M.
sydneyi.
Objetivo 4. Identificación de potenciales hospedadores
intermediarios en el ciclo biológico de Marteilia cochillia.
Se realizaron arrastres pelágicos en 3 transeptos de la
ría de Arousa durante el periodo comprendido entre
noviembre 2015 a marzo 2017. Los transeptos
correspondían a los bancos marisqueros de O Sarrido,
Lombos do Ulla y Corón. Se empleó una red de Bongo
de dos mangas con malla de 200 µm de luz para obtener
muestras de zooplancton en la columna de agua. En
cada transepto, la muestra procedente de una de las
mangas de la red de Bongo fue conservada en alcohol
de 96% para extracción de ADN, y la procedente de la
otra manga fue fijada en 10% de formaldehído en agua
de mar para el estudio histológico. Se equipó la red de
Bongo con un medidor de flujo de corriente para estimar
abundancias (Nº individuos/m3) de la comunidad
mesozooplanctónica a partir del volumen de agua de mar
filtrado en cada transepto. Para la identificación de
especies se pipeteó un volumen concreto en una cámara
de recuento de zooplancton. Cuando resultó posible
cada individuo fue identificado hasta nivel de especie. En
el mes de febrero se pudo observar una clara
disminución de la abundancia de individuos en todas las
zonas de estudio. Por el contrario, en los meses de abril
y mayo de 2016 se observó un incremento bastante
elevado de la abundancia de individuos respecto al resto
de los meses. Este resultado es el esperado por entrada
de la primavera, época de reproducción de la mayoría de
las especies. En el periodo de análisis, comprendido
desde noviembre de 2015 hasta marzo de 2017, se
identificaron un total de 43 taxones. Los taxones
identificados se reagruparon en dos categorías,
holoplancton (organismos que durante todo su ciclo vital
forman parte de la comunidad planctónica como
copépodos, apendicularias, cladóceros, medusas,
radiolarios,…) y meroplancton (que está constituido por
seres que están en esta comunidad sólo durante una
parte de su vida, como pueden ser fases larvarias de
moluscos, crustáceos o peces). Del análisis de la
composición taxonómica de la comunidad
mesozooplanctónica en las tres zonas de estudio se
puede destacar que el grupo de copépodos es
principalmente el dominante, salvo excepciones en las
que el holoplancton está constituido mayoritariamente
por apendicularias o cladóceros, o en las que el
meroplancton es el dominante debido a la elevada
abundancia de larvas. Los copépodos calanoides Acartia
clausi, Acartia discaudata y Paracartia grani fueron
dominantes a lo largo del periodo de estudio.
Las especies de mesozooplancton separadas en cada
muestreo fueron analizadas para determinar la presencia
de Marteilia cochillia mediante pruebas moleculares. Se
hicieron pools de cada especie para cada una de las 3
áreas de estudio. Se realizó la extracción de ADN de
cada pool y la presencia del parásito en la muestra se
determinó mediante nested PCR y ensayo de restricción
(Lopez-Flores y col., 2004; Carrasco y col., 2012). Los
resultados mostraron que el mayor número de especies
PCR positivas a M. cochillia se corresponde con el
holoplancton aunque se registraron algunos positivos en
el meroplancton. Dentro del holoplancton, las especies
PCR positivas fueron principalmente copépodos
calanoides (orden Calanoida) aunque también se
registran positivos en copépodos del orden Ciclopoida y
en las apendicularias. Dentro del meroplancton, se
registró un valor PCR positivo en larvas de decápodos,
en larvas de equinodermos y en nauplius de cirrípedos.
Es de señalar que los positivos detectados mediante
PCR indican la presencia de un agente infeccioso, pero
la técnica no verifica la existencia de una infección, ya
que la presencia del parásito podría deberse a la
ingestión del mismo entre otras causas. Así, para
identificar hospedadores intermediarios es necesario
realizar ensayos de hibridación in situ (ISH) para
verificar la identidad del parásito y confirmar que el
hospedador intermediario está infectado. En el caso de
M. cochillia se están realizando pruebas de ISH para
confirmar que estos resultados positivos detectados en el
mesozooplancton se corresponden con verdaderas
parasitosis.

66
Referencias
Carrasco, N., Roque, A., Andree, K. B., Rodgers, C.,
Lacuesta, B., Furones, M. D. 2011. A Marteilia parasite
and digestive epithelial virosis lesions observed during a
common edible cockle Cerastoderma edule mortality
event in the Spanish Mediterranean coast. Aquaculture
321: 197-202.
Carrasco, N., Hine, M., Durfort, M., Andree, K. B.,
Malchus, N., Lacuesta, B., González, M., Roque, A.,
Rodgers, C., Furones, M. D. 2013. Marteilia cochillia sp.
nov., a new Marteilia species affecting the edible cockle
Cerastoderma edule in European waters. Aquaculture,
412-413: 223-230.
Comps, M., Grizel, H., Tigé,G., Duthoit, J. L. 1975.
Parasites nouveaux de la glande digestive des
mollusques marins Mytilus edulis L. et Cardium edule.
Comp. Rendus Acad. Sci.paris Sér. D 281: 179-181.
Villalba A., Iglesias D., Ramilo A., Darriba S., Parada J.
M., No E., Abollo E., Molares, J., Carballal M. J. 2014.
Cockle Cerastoderma edule fishery collapse in the Ría
de Arousa (Galicia NW Spain) associated with the
protistan parasite Marteilia cochillia. Diseases of Aquatic
Organisms, 109: 55-80.

CIMA Memoria 2017 67
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Determinación de la posible resistencia a la marteiliosis
de una cohorte de berberecho Cerastoderma edule
detectada en el banco de O Ariño (Vilanova de Arousa).
INICIO: julio, 2017
FINALIZACIÓN: diciembre, 2018
ENTIDAD FINANCIADORA:
Consellería do Mar. Xunta de Galicia
Código: CIMA 17/06
INVESTIGADOR PRINCIPAL:
Iglesias Estepa, D.
INVESTIGADORES/AS:
CIMA: Carballal Durán, M.J.; Fuentes González, J.M.
Cofradía de Pescadores “La Pastoriza” de Vilanova de
Arousa: Abella Rey, E.
APOYO A LA INVESTIGACIÓN:
Martínez Verde, G.; Penas Pampín, E.; Brianes Beiras,
M.J.
RESUMEN
El berberecho Cerastoderma edule fue tradicionalmente
la principal especie explotada del marisqueo en Galicia
en lo que se refiere a volumen de capturas. En el año
2012 se detectó, por primera vez en Galicia, el parásito
Marteilia cochillia infectando berberechos de la ría de
Arousa y causando una mortalidad sin precedentes que
originó el colapso de su explotación en esta ría. En los
años 2013 y 2014 la marteiliosis se extendió a las rías de
Pontevedra y Vigo originando mortalidades elevadas de
berberecho en las mismas y una caída dramática en la
producción de esta especie en Galicia. Desde la
aparición de esta enfermedad, en las rías afectadas se
han detectado brotes anuales de marteiliosis que
habitualmente causan la extinción de cada nueva
cohorte reclutada e impiden la recuperación de la
explotación marisquera.
En la navidad del 2016 se constató que en la lonja de
Vilanova de Arousa se estaban comercializando
berberechos de gran tamaño (ejemplares de hasta 37
mm) que conseguían un precio de primera venta muy
elevado (precios máximos de hasta 18 €/kg). Mientras
tanto, en el resto de lonjas de la ría de Arousa apenas
existía berberecho C. edule de tamaño legal en las
subastas. Indagando en el origen de estos ejemplares de
berberecho de gran tamaño, se confirmó que procedían
del banco del Ariño y, atendiendo a su tamaño, muy
probablemente correspondían a una cohorte reclutada
en el año 2015. En este supuesto se trataría de
berberechos supervivientes al brote de marteiliosis que a
principios del año 2016 asoló la ría de Arousa causando
mortalidades masivas del berberecho fijado en 2015.
En la presente acción de investigación se proponía
analizar la situación de esta población de berberecho
para tratar de dilucidar sí (1) el banco del Ariño es una
zona refugio libre de marteiliosis dentro de la ría de
Arousa; (2) si el berberecho en esta zona presenta una
tasa de crecimiento mayor que el resto de zonas de la ría
de Arousa, lo que explicaría su elevado tamaño aún
tratándose de la cohorte reclutada en el año en curso; o
sí, (3) efectivamente, el berberecho C. edule de esta
zona presenta características particulares que la hacen
resistente a la marteiliosis. De tratarse de este último
supuesto los individuos de este banco constituirían una
verdadera oportunidad para el desarrollo de estrategias
de lucha contra la marteiliosis basados en programas de
selección genética.
Con el fin de contrastar las hipótesis anteriores se inició
a finales de junio de 2017, coincidiendo con la aparición
de los primeros reclutas de la cohorte de 2017, un
seguimiento del estado sanitario del berberecho de este
banco. Para ello se recogieron mensualmente 30
individuos de C. edule de O Ariño que fueron procesados
mediante histología y analizados al microscopio óptico
con el fin de diagnosticar si estaban infectados por el
parásito M. cochillia. Asimismo, desde septiembre de
2017, se realizaron trimestralmente muestreos de
evaluación de C. edule, con el fin de analizar la evolución
de la densidad y de la estructura de tallas de esta
especie y estimar diferentes parámetros de la dinámica
poblacional como crecimiento y mortalidad. Para ello se
recogieron dos muestras de 0,05 m2 de superficie,
empleando calicatas cilíndricas enterradas hasta unos 20
cm de profundidad, en cada una de las estaciones de
muestreo de una red de 8 distribuidas regularmente a lo
largo de todo el banco. Las muestras se lavaron y
tamizaron en el campo a través de una malla metálica de
5 mm de luz, se recogieron en bolsas plásticas
etiquetadas y se llevaron a los laboratorios del CIMA,

68
donde se contabilizaron y midieron los berberechos
presentes en cada muestra. A mayores, a comienzos de
diciembre de 2017 se dispusieron en este banco tres
cajas plásticas de control de mortalidad enterradas en el
sustrato. En cada una de ellas se depositaron 30
berberechos y las cajas fueron cubiertas con una malla
plástica de 5 mm de luz para evitar la pérdida y
depredación de estos individuos control. Las cajas se
revisaron mensualmente contabilizando el número de
berberechos supervivientes, con el fin de estimar la tasa
de mortalidad mensual. Se repusieron los berberechos
muertos recogiendo individuos del banco de manera que
de nuevo hubiera 30 individuos de control en cada una
de las cajas. Fijados a estas cajas de control se
dispusieron sondas de registro continuo de temperatura
y salinidad, con el fin de monitorizar estas variables
ambientales.
Aunque esta acción de investigación no finaliza hasta
diciembre de 2018, con los resultados de 2017 ya se
pueden establecer ciertas conclusiones con respecto a
las hipótesis de partida. Así, tal y como se muestra en la
figura 1A, se detectó un brote de marteiliosis afectando a
la cohorte de berberecho C. edule del banco del Ariño
reclutada en 2017. En julio, apenas un mes después de
la detección en el banco de los primeros reclutas, ya el
21 % de los individuos analizados estaban infectados por
M. cochillia. La prevalencia (porcentaje de individuos de
la población afectados por la enfermedad) alcanzó
valores máximos en septiembre, con un 79 % de los
berberechos infectados, y a partir de ahí disminuyó
paulatinamente hasta valores en torno al 24 % en
diciembre de 2017. La caída de los valores de
prevalencia pueden explicarse por la muerte de los
individuos enfermos, tal y como se puede deducir de los
elevados valores de mortalidad acumulada entre
septiembre y diciembre (Fig. 1A), calculados a partir de
la disminución de la densidad de berberecho en el banco
(Figs. 1B y 2). Se constata que los berberechos del
banco del Ariño están infectados por M. cochillia y, por lo
tanto, se descarta la hipótesis de que este banco
constituya una zona refugio libre de marteiliosis dentro
de la ría de Arousa.
La fuerte caída en la densidad de población entre finales
de septiembre y comienzos de diciembre, en un
momento en que las variables ambientales (salinidad y
temperatura) estuvieron dentro del rango de tolerancia
para el berberecho (Fig. 1C), y en el que prácticamente
no había habido explotación comercial del recurso,
sugieren la existencia de fuertes mortalidades asociadas
a la marteiliosis. Parece pues que los berberechos de O
Ariño tampoco presentan características particulares que
los hagan resistentes a la marteiliosis, si bien este
término tiene que ser evaluado en profundidad a lo largo
del año 2018 a través de la estima de la mortalidad
natural en cajas de control.
Llama la atención, sin embargo, el elevado crecimiento
del berberecho en este banco, ya que partiendo de un
stock de reclutas fijados en el banco a lo largo del verano
de 2017, en diciembre ya existía una densidad de
berberecho comercial en el banco de 45 individuos/m2. A
partir del análisis de la progresión de la moda de esta
cohorte se estima un crecimiento en este período de 2,4
mm/mes (Fig. 2), valores muy elevados que hacen que el
recurso alcance tamaño comercial, y por lo tanto resulte
explotable, tan solo entre 4 y 6 meses tras la fijación de
los juveniles en el sustrato. A falta de una verificación de
este modelo a lo largo de 2018, todo apunta a que en
este banco el elevado crecimiento del berberecho
permite su explotación comercial en el año en curso
antes de que se dispare la mortalidad debida a la
marteiliosis y se produzca la extinción de la cohorte.
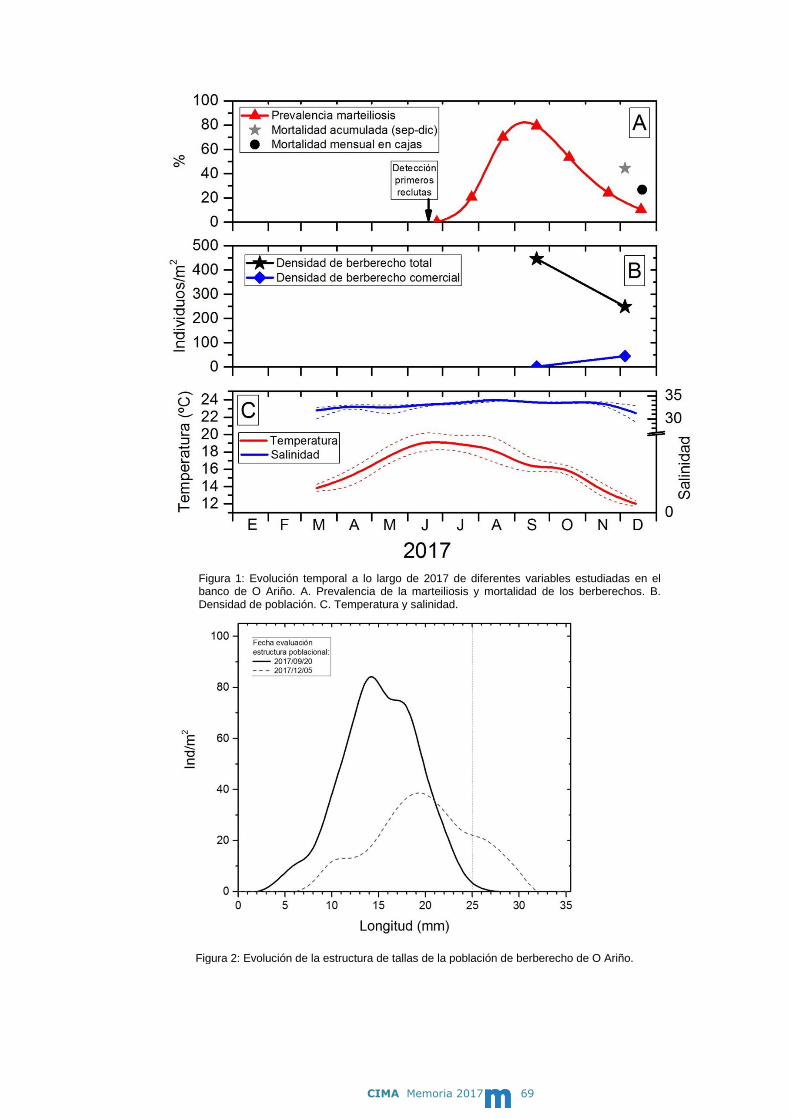
CIMA Memoria 2017 69
Figura 1: Evolución temporal a lo largo de 2017 de diferentes variables estudiadas en el banco de O Ariño. A. Prevalencia de la marteiliosis y mortalidad de los berberechos. B. Densidad de población. C. Temperatura y salinidad.
Figura 2: Evolución de la estructura de tallas de la población de berberecho de O Ariño.

70

CIMA Memoria 2017 71
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Evaluación del incremento de supervivencia en el
engorde de ostra rizada Crassostrea gigas por el uso de
semilla descendiente de supervivientes a la exposición
prolongada al virus herpes de ostreidos tipo 1.
Identificación de marcadores moleculares de resistencia
para su aplicación en un programa de selección de
estirpes resistentes al virus.
INICIO: marzo, 2016
FINALIZACIÓN: diciembre, 2018
ENTIDAD FINANCIADORA:
Consellería do Mar. Xunta de Galicia.
Código: PGIDIT-CIMA 16/02
INVESTIGADOR PRINCIPAL:
Villalba García, A.
INVESTIGADORES/AS:
Cao Hermida, M.A; Fuentes González, J.M.
APOYO A LA INVESTIGACIÓN:
Brianes Beiras, M.J.; Giráldez Rivero, R.; González
Fontela, A.I.; Gregorio Chenlo, M.V.; Meléndez Ramos,
M.I.; Penas Pampín, E.; Rúa Santervas, P.
BECARIOS/AS:
Alonso Costas, J.B.; Campañó Piñeiro, M.S.; Pena
Thomas, G.; Viturro García, R.
RESUMEN
Introducción
El engorde de la ostra rizada, Crassostrea gigas, en la
ría de Arousa se ve afectado desde hace al menos 7
años por una mortalidad muy alta de la semilla, causada
por la infección con la microvariante del virus herpes 1
de ostreidos (OsHV1-µvar). Se trata de una enfermedad
detectada inicialmente en Francia, en el verano del 2008,
que se ha expandido a otros países europeos así como a
Australia, Nueva Zelanda y Corea del Sur. Nuestro grupo
se ha implicado en el estudio de esta enfermedad, en
particular en la búsqueda de procedimientos que
permitan aumentar la supervivencia de la ostra rizada en
el engorde en Galicia. Otros autores han demostrado
que la temperatura del agua y la edad y el tamaño de la
ostra influyen en la dinámica de la enfermedad; así, por
debajo de 16 ºC no hay mortalidad relevante y por
encima de 24 ºC la mortalidad se reduce
significativamente; la susceptibilidad de la ostra a la
infección y mortalidad se reduce notablemente en los
individuos mayores, con peso superior a 20 g. En un
proyecto previo evaluamos la eficacia de procedimientos
alternativos de engorde cuyo diseño se había basado en
la información sobre la influencia de la temperatura y la
edad y tamaño de las ostras en la mortalidad pero los
resultados no fueron satisfactorios pues la supervivencia
fue baja. Teniendo en cuenta que en Francia se habían
obtenido resultados muy esperanzadores en la lucha
contra la infección por OsHV1-µvar mediante programas
de mejora genética para aumentar la resistencia a la
enfermedad, basados en utilizar como reproductores
ostras supervivientes en zonas afectadas por la
enfermedad, repitiéndose el proceso en generaciones
sucesivas, decidimos desarrollar un nuevo proyecto de
investigación para evaluar si, también en la ría de
Arousa, la utilización de supervivientes de una zona
afectada como reproductores conduciría a un aumento
de la supervivencia de sus descendientes durante el
engorde y, en tal caso, sentar las bases para desarrollar
un programa de selección genética. Además, se planteó
la búsqueda de marcadores moleculares de resistencia a
la enfermedad que sirvan de apoyo a un ulterior
programa de mejora genética, mediante dos enfoques,
uno proteómico, desarrollado por nuestro grupo, y otro
genómico, a desarrollar por el equipo de la Dra. María
Prado Álvarez, investigadora del Instituto de
Investigaciones Marinas (CSIC, Vigo).
Material y métodos
La primera etapa del proyecto implicó la producción de
varias familias de semilla de ostra rizada de dos orígenes
diferentes, (1) una población nunca afectada por el virus
y (2) supervivientes tras la exposición al virus en un área
afectada. Un banco naturalizado de ostra rizada en la ría
de O Barqueiro (Mariña de Lugo) se utilizó como fuente
de reproductores adultos de una población nunca
afectada, lo que se ha ido confirmando por muestreos y
análisis sucesivos desde 2012 y ausencia de mortalidad
anormal. Como “supervivientes”, se utilizaron ostras de
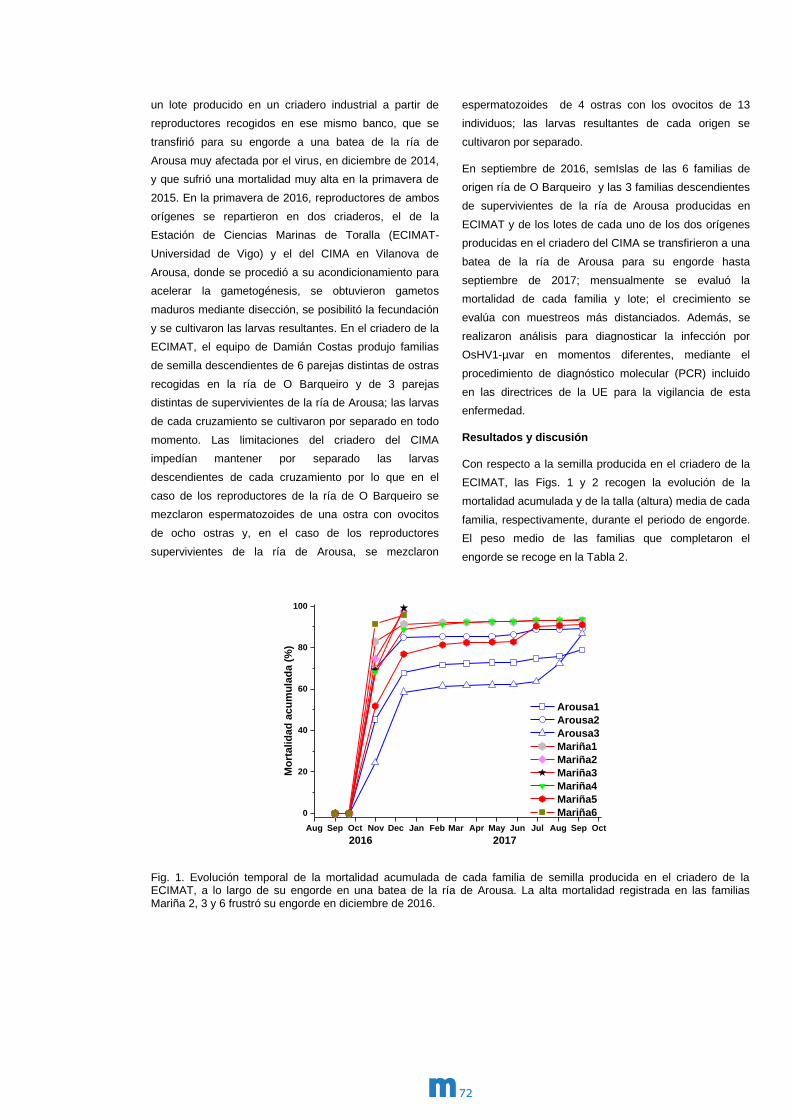
72
un lote producido en un criadero industrial a partir de
reproductores recogidos en ese mismo banco, que se
transfirió para su engorde a una batea de la ría de
Arousa muy afectada por el virus, en diciembre de 2014,
y que sufrió una mortalidad muy alta en la primavera de
2015. En la primavera de 2016, reproductores de ambos
orígenes se repartieron en dos criaderos, el de la
Estación de Ciencias Marinas de Toralla (ECIMAT-
Universidad de Vigo) y el del CIMA en Vilanova de
Arousa, donde se procedió a su acondicionamiento para
acelerar la gametogénesis, se obtuvieron gametos
maduros mediante disección, se posibilitó la fecundación
y se cultivaron las larvas resultantes. En el criadero de la
ECIMAT, el equipo de Damián Costas produjo familias
de semilla descendientes de 6 parejas distintas de ostras
recogidas en la ría de O Barqueiro y de 3 parejas
distintas de supervivientes de la ría de Arousa; las larvas
de cada cruzamiento se cultivaron por separado en todo
momento. Las limitaciones del criadero del CIMA
impedían mantener por separado las larvas
descendientes de cada cruzamiento por lo que en el
caso de los reproductores de la ría de O Barqueiro se
mezclaron espermatozoides de una ostra con ovocitos
de ocho ostras y, en el caso de los reproductores
supervivientes de la ría de Arousa, se mezclaron
espermatozoides de 4 ostras con los ovocitos de 13
individuos; las larvas resultantes de cada origen se
cultivaron por separado.
En septiembre de 2016, semIslas de las 6 familias de
origen ría de O Barqueiro y las 3 familias descendientes
de supervivientes de la ría de Arousa producidas en
ECIMAT y de los lotes de cada uno de los dos orígenes
producidas en el criadero del CIMA se transfirieron a una
batea de la ría de Arousa para su engorde hasta
septiembre de 2017; mensualmente se evaluó la
mortalidad de cada familia y lote; el crecimiento se
evalúa con muestreos más distanciados. Además, se
realizaron análisis para diagnosticar la infección por
OsHV1-µvar en momentos diferentes, mediante el
procedimiento de diagnóstico molecular (PCR) incluido
en las directrices de la UE para la vigilancia de esta
enfermedad.
Resultados y discusión
Con respecto a la semilla producida en el criadero de la
ECIMAT, las Figs. 1 y 2 recogen la evolución de la
mortalidad acumulada y de la talla (altura) media de cada
familia, respectivamente, durante el periodo de engorde.
El peso medio de las familias que completaron el
engorde se recoge en la Tabla 2.
Aug Sep Oct Nov Dec Jan Feb Mar Apr May Jun Jul Aug Sep Oct
0
20
40
60
80
100
Mo
rta
lid
ad
ac
um
ula
da
(%
)
2016 2017
Arousa1
Arousa2
Arousa3
Mariña1
Mariña2
Mariña3
Mariña4
Mariña5
Mariña6
Fig. 1. Evolución temporal de la mortalidad acumulada de cada familia de semilla producida en el criadero de la ECIMAT, a lo largo de su engorde en una batea de la ría de Arousa. La alta mortalidad registrada en las familias Mariña 2, 3 y 6 frustró su engorde en diciembre de 2016.
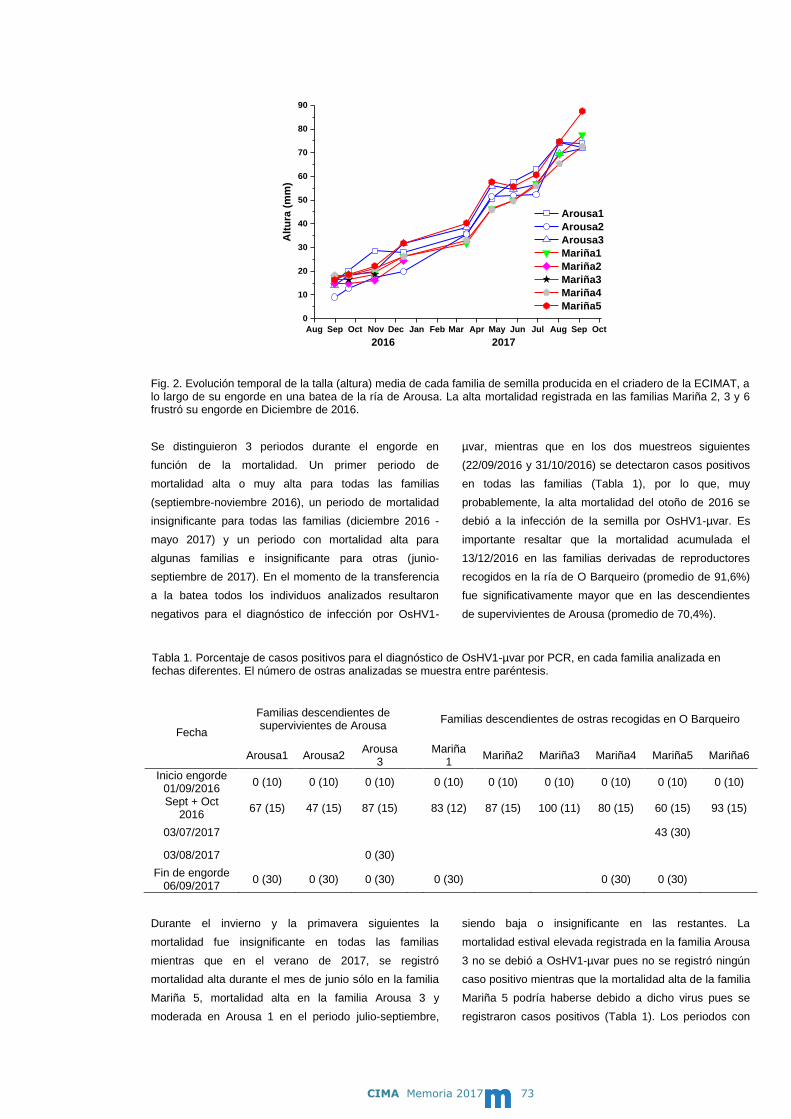
CIMA Memoria 2017 73
Aug Sep Oct Nov Dec Jan Feb Mar Apr May Jun Jul Aug Sep Oct
0
10
20
30
40
50
60
70
80
90
Alt
ura
(m
m)
2016 2017
Arousa1
Arousa2
Arousa3
Mariña1
Mariña2
Mariña3
Mariña4
Mariña5
Fig. 2. Evolución temporal de la talla (altura) media de cada familia de semilla producida en el criadero de la ECIMAT, a lo largo de su engorde en una batea de la ría de Arousa. La alta mortalidad registrada en las familias Mariña 2, 3 y 6 frustró su engorde en Diciembre de 2016.
Se distinguieron 3 periodos durante el engorde en
función de la mortalidad. Un primer periodo de
mortalidad alta o muy alta para todas las familias
(septiembre-noviembre 2016), un periodo de mortalidad
insignificante para todas las familias (diciembre 2016 -
mayo 2017) y un periodo con mortalidad alta para
algunas familias e insignificante para otras (junio-
septiembre de 2017). En el momento de la transferencia
a la batea todos los individuos analizados resultaron
negativos para el diagnóstico de infección por OsHV1-
µvar, mientras que en los dos muestreos siguientes
(22/09/2016 y 31/10/2016) se detectaron casos positivos
en todas las familias (Tabla 1), por lo que, muy
probablemente, la alta mortalidad del otoño de 2016 se
debió a la infección de la semilla por OsHV1-µvar. Es
importante resaltar que la mortalidad acumulada el
13/12/2016 en las familias derivadas de reproductores
recogidos en la ría de O Barqueiro (promedio de 91,6%)
fue significativamente mayor que en las descendientes
de supervivientes de Arousa (promedio de 70,4%).
Tabla 1. Porcentaje de casos positivos para el diagnóstico de OsHV1-µvar por PCR, en cada familia analizada en fechas diferentes. El número de ostras analizadas se muestra entre paréntesis.
Fecha
Familias descendientes de supervivientes de Arousa
Familias descendientes de ostras recogidas en O Barqueiro
Arousa1 Arousa2 Arousa
3
Mariña1
Mariña2 Mariña3 Mariña4 Mariña5 Mariña6
Inicio engorde 01/09/2016
0 (10) 0 (10) 0 (10) 0 (10) 0 (10) 0 (10) 0 (10) 0 (10) 0 (10)
Sept + Oct 2016
67 (15) 47 (15) 87 (15) 83 (12) 87 (15) 100 (11) 80 (15) 60 (15) 93 (15)
03/07/2017 43 (30)
03/08/2017 0 (30)
Fin de engorde 06/09/2017
0 (30) 0 (30) 0 (30) 0 (30) 0 (30) 0 (30)
Durante el invierno y la primavera siguientes la
mortalidad fue insignificante en todas las familias
mientras que en el verano de 2017, se registró
mortalidad alta durante el mes de junio sólo en la familia
Mariña 5, mortalidad alta en la familia Arousa 3 y
moderada en Arousa 1 en el periodo julio-septiembre,
siendo baja o insignificante en las restantes. La
mortalidad estival elevada registrada en la familia Arousa
3 no se debió a OsHV1-µvar pues no se registró ningún
caso positivo mientras que la mortalidad alta de la familia
Mariña 5 podría haberse debido a dicho virus pues se
registraron casos positivos (Tabla 1). Los periodos con
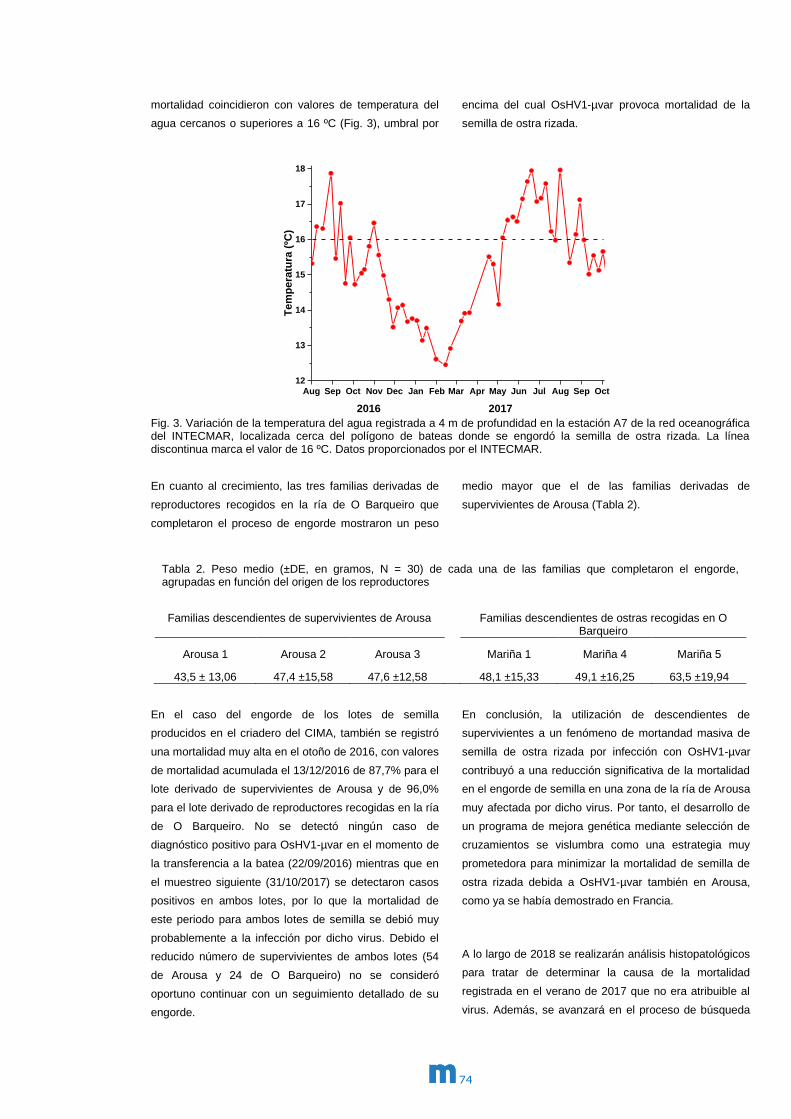
74
mortalidad coincidieron con valores de temperatura del
agua cercanos o superiores a 16 ºC (Fig. 3), umbral por
encima del cual OsHV1-µvar provoca mortalidad de la
semilla de ostra rizada.
Aug Sep Oct Nov Dec Jan Feb Mar Apr May Jun Jul Aug Sep Oct
12
13
14
15
16
17
18
Tem
pera
tura
(ºC
)
2016 2017 Fig. 3. Variación de la temperatura del agua registrada a 4 m de profundidad en la estación A7 de la red oceanográfica del INTECMAR, localizada cerca del polígono de bateas donde se engordó la semilla de ostra rizada. La línea discontinua marca el valor de 16 ºC. Datos proporcionados por el INTECMAR.
En cuanto al crecimiento, las tres familias derivadas de
reproductores recogidos en la ría de O Barqueiro que
completaron el proceso de engorde mostraron un peso
medio mayor que el de las familias derivadas de
supervivientes de Arousa (Tabla 2).
En el caso del engorde de los lotes de semilla
producidos en el criadero del CIMA, también se registró
una mortalidad muy alta en el otoño de 2016, con valores
de mortalidad acumulada el 13/12/2016 de 87,7% para el
lote derivado de supervivientes de Arousa y de 96,0%
para el lote derivado de reproductores recogidas en la ría
de O Barqueiro. No se detectó ningún caso de
diagnóstico positivo para OsHV1-µvar en el momento de
la transferencia a la batea (22/09/2016) mientras que en
el muestreo siguiente (31/10/2017) se detectaron casos
positivos en ambos lotes, por lo que la mortalidad de
este periodo para ambos lotes de semilla se debió muy
probablemente a la infección por dicho virus. Debido el
reducido número de supervivientes de ambos lotes (54
de Arousa y 24 de O Barqueiro) no se consideró
oportuno continuar con un seguimiento detallado de su
engorde.
En conclusión, la utilización de descendientes de
supervivientes a un fenómeno de mortandad masiva de
semilla de ostra rizada por infección con OsHV1-µvar
contribuyó a una reducción significativa de la mortalidad
en el engorde de semilla en una zona de la ría de Arousa
muy afectada por dicho virus. Por tanto, el desarrollo de
un programa de mejora genética mediante selección de
cruzamientos se vislumbra como una estrategia muy
prometedora para minimizar la mortalidad de semilla de
ostra rizada debida a OsHV1-µvar también en Arousa,
como ya se había demostrado en Francia.
A lo largo de 2018 se realizarán análisis histopatológicos
para tratar de determinar la causa de la mortalidad
registrada en el verano de 2017 que no era atribuible al
virus. Además, se avanzará en el proceso de búsqueda
Tabla 2. Peso medio (±DE, en gramos, N = 30) de cada una de las familias que completaron el engorde, agrupadas en función del origen de los reproductores
Familias descendientes de supervivientes de Arousa Familias descendientes de ostras recogidas en O Barqueiro
Arousa 1 Arousa 2 Arousa 3
Mariña 1 Mariña 4 Mariña 5
43,5 ± 13,06 47,4 ±15,58 47,6 ±12,58
48,1 ±15,33 49,1 ±16,25 63,5 ±19,94

CIMA Memoria 2017 75
de genes y proteínas de la ostra rizada candidatos a ser
considerados como marcadores de resistencia a la
infección por OsHV1-µvar, para lo que se compararán el
proteoma y el transcriptoma de individuos de cada
familia recogidos en el momento de la transferencia a
batea con los de individuos de cada familia
supervivientes recogidos en diciembre de 2016, tras el
episodio de mortandad masiva de semilla.

76
PROYECTO DE INVESTIGACIÓN EQUIPO
TÍTULO:
Co-operation for restoring cockle shellfisheries and its
ecosystem-services in the Atlantic Area (AA).
ACRÓNIMO: COCKLES
INICIO: octubre, 2017
FINALIZACIÓN: septiembre, 2020
ENTIDAD FINANCIADORA:
Unión Europea, Programa INTERREG Atlantic Area
(EAPA_458/2016)
COORDINADOR DEL PROYECTO:
CETMAR: Fernández Otero, R.
INVESTIGADOR PRINCIPAL EN EL CIMA:
Villalba García, A.
INVESTIGADORES/AS:
Cao Hermida, M.A; Carballal Durán, M.J.; Iglesias
Estepa, D.
APOYO A LA INVESTIGACIÓN:
Brianes Beiras, M.J.; González Fontela, A.I.; Martínez
Verde, G.; Meléndez Ramos, M.I.; Penas Pampín, E.;
Rúa Santervas, P.
BECARIOS/AS:
Fernández González, J.; Pena Thomas, G; Viturro
García, R.
RESUMEN
Los berberechos proporcionan beneficios ambientales,
socioeconómicos y culturales significativos a las
comunidades costeras de los países implicados en el
proyecto (España, Portugal, Francia, Irlanda y Reino
Unido) pero están amenazados por enfermedades y una
gestión incorrecta. El propósito de este proyecto es
recuperar y aumentar la producción de berberecho así
como los servicios que este molusco proporciona en las
regiones del Espacio Atlántico de la UE, a través de los
objetivos siguientes: (1) Evaluar la salud, diversidad e
interrelaciones de las poblaciones de berberecho a lo
largo del Espacio Atlántico caracterizando la dinámica de
poblaciones; diversidad genética y transporte larvario;
amenazas por enfermedades, contaminación, especies
invasoras y cambio climático. (2) Cuantificar los
beneficios económicos, sociales y culturales de los
servicios ecosistémicos que aportan los berberechos
(marisqueo, acuicultura, biodiversidad, alimento para
aves, turismo, servicios sociales) realizando encuestas,
entrevistas y análisis socioeconómicos. (3) Proveer de
técnicas de gestión nuevas desarrollando procedimientos
y tecnologías de recuperación de bancos de berberecho,
cultivo en criadero, programas de mejora genética para
producir estirpes resistentes a enfermedades y de
crecimiento rápido y conservación de la estructura y
diversidad genética. (4) Proporcionar orientación y
asesoramiento sobre buenas prácticas a productores,
administraciones, agencias ambientales y ONGs
evaluando y poniendo en común los procedimientos
mejores a lo largo del Espacio Atlántico y optimizando la
gestión mediante enseñanzas mutuas. Todo ello
redundará en la mejora de la producción de berberecho,
una industria robusta, viable y sostenible, con patentes
beneficios sociales y de biodiversidad.
Grupos de tareas del proyecto:
1. Coordinación. El objetivo es facilitar un desarrollo
fluido de las actividades, gestionar la coordinación global
del equipo de personas y los recursos implicados y
garantizar una implementación eficiente y responsable y
la emisión en tiempo y forma de los informes pertinentes.
2. Comunicación. Se pretende promover un diálogo
abierto entre todos los estamentos afectados: científicos,
mariscadores, ONGs y otros implicados en el uso de
berberechos y sensibilizar a todos sobre la pluralidad y
valor de los servicios y beneficios aportados por los
berberechos. Como instrumentos se usará una página
web, actos públicos, talleres de trabajo y una conferencia
final en la que se presentarán y discutirán los logros del
proyecto. Su diseminación continuará a través de la web,
artículos científicos y de divulgación, reuniones técnicas

CIMA Memoria 2017 77
y congresos y un taller de trabajo específico transcurrido
un año desde la conclusión del proyecto.
3. Capitalización. Acciones orientadas a asegurar que los
resultados continuarán generando impacto más allá de la
duración del proyecto, de forma que todos a quienes
atañe (productores, administraciones, ONGs,
comunidades costeras) incorporen los resultados y
productos del proyecto y los utilicen en sus ámbitos
respectivos, con capacitación y pericia optimizadas, junto
con la creación de redes de cooperación a largo plazo.
4. Evaluación de la situación de partida: caracterización
de las poblaciones, capacidad reproductiva y estructura
genética. Evaluación exhaustiva e integradora de la
situación de las poblaciones de berberecho en el
Espacio Atlántico, incluyendo distribución geográfica,
abundancia, dinámica de poblaciones e influencia de
factores geográficos, con vistas a predecir tendencias
futuras en escenarios diferentes. Se incluirá la
evaluación de la capacidad reproductiva y de la
estructura y diversidad genéticas.
5. Identificación de las amenazas principales. Análisis de
las enfermedades que padecen las poblaciones de
berberecho en cada país, evaluación de riesgos y
determinación de procedimientos específicos de lucha.
Caracterización del sistema inmunitario del berberecho y
de su microbiota como vías del berberecho para superar
condiciones de estrés, con énfasis en patógenos y
cambio climático. Evaluación de la amenaza por
especies invasoras.
6. Modelado de flujos: transporte larvario y cascadas
tróficas. Desarrollo de modelos biofísicos y ecológicos
integrados que simulen las conexiones entre bancos de
berberecho vía dispersión larvaria. Desarrollo de
modelos ecológicos que, estimando el flujo de
materia/energía entre ecosistemas a través de
organismos filtradores (berberechos y otros), permitan
predecir la capacidad de carga y los efectos del
berberecho sobre producción primaria y cascadas
tróficas. El uso de estos modelos predictivos mejorará la
gestión de los bancos de berberecho.
7. Procedimientos de cultivo de berberecho y mejora
genética para producir estirpes resistentes a la
marteiliosis. Desarrollo de procedimientos de cultivo:
producción de semilla en criadero y engorde en el medio
natural. Para producir estirpes de berberecho resistentes
a la marteiliosis se estudiarán las bases moleculares de
la respuesta del berberecho a esta enfermedad y se
buscarán marcadores moleculares de resistencia. Se
diseñará un protocolo de selección genética para
resistencia a la marteiliosis. Los protocolos de cultivo y
de selección genética y toda la información sobre
demografía y estructura genética de las poblaciones de
berberecho y su dispersión larvaria se integrarán para
diseñar procedimientos para recuperar bancos de
berberecho agotados o en declive.
8. Cuantificación de los beneficios socioeconómicos y
para la biodiversidad proporcionados por el berberecho.
Análisis exhaustivo de los beneficios que proporcionan
los berberechos para mantener la salud del ecosistema
así como de los socioeconómicos. Con los resultados del
análisis se creará una herramienta para cuantificar todos
estos beneficios que será muy útil para la gestión de los
bancos.
ENTIDAD PARTICIPANTE PAÍS/REGIÓN TIPO DE
PARTICIPACIÓN
Centro Tecnológico del Mar, Fundación CETMAR España/Galicia Socio coordinador
Consellería do Mar, Xunta de Galicia (Centro de Investigacións Mariñas)
España/Galicia Socio
Universidade de Santiago de Compostela (ACUIGEN - Dpto. de Genética)
España/Galicia Socio
Université de Bordeaux Francia/Aquitaine Socio
Groupe d'Etude des Milieux Estuariens et Littoraux
Francia/ Hautes-de-France Socio
Université de Caen Normandie Francia/Basse Normandie Socio

78
University College Cork Irlanda/Southern and Eastern Socio
Bangor University Reino Unido/ West Wales and The Valleys
Socio
NERC Centre for Ecology and Hydrology, Bangor Reino Unido/ West Wales and The Valleys
Socio
Cyfoeth Naturiol Cymru Socio
Groupe d'Etude des Milieux Estuariens et Littoraux
Francia/ Hautes-de-France Socio
Instituto Português do Mar e da Atmosfera Portugal/Lisboa Socio
Universidade de Aveiro Portugal/Centro Socio
Faculdade de Ciências da Universidade de Lisboa
Portugal/Lisboa Socio
Confraría de Pescadores San Antonio de Cambados
España/Galicia Asociado
Secretaría Xeral de Calidade e Avaliación Ambiental-Xunta de Galicia
España/Galicia Asociado
Instituto Tecnolóxico para o Control do Medio Mariño de Galicia
España/Galicia Asociado
Institut Français de Recherche pour l'Exploitation de la Mer
Francia/Nouvelle Aquitaine Asociado
Fédération des Sociétés pour l’Etude, la Protection, et l’Aménagement de la Nature dans le Sud-Ouest
Francia/Nouvelle Aquitaine Asociado
Centre National de la Recherche Scientifique Délégation Aquitaine
Francia/Nouvelle Aquitaine Asociado
Marine Institute Ireland Irlanda/Border, Midland and Western
Asociado
Formosa, Cooperativa de Viveiristas da Ria Formosa, C.R.L.
Portugal/Algarve Asociado
Instituto da Conservação da Natureza e das Florestas
Portugal/Lisboa Asociado

CIMA Memoria 2017 79
7 PROCESOS OCEANOGRÁFICOS COSTEROS

80
RESUMEN
Esta acción es la continuación de la acción DEMETOX.
El control de la toxicidad de tipo paralizante (PSP) y
lipofílico se lleva a cabo en la actualidad por medio de
bioensayo con ratón y LC-MS/MS, respectivamente. Con
todo, es inminente la adopción de un método de
cromatografía líquida de alta eficacia (HPLC), con
oxidación pre-columna y detección de fluorescencia,
como referencia en la UE para la determinación de la
toxicidad PSP, substituyendo al bioensayo de ratón, lo
cual hace necesaria su adopción en el sistema de control
de la Comunidad Autónoma de Galicia.
Ambas metodologías analíticas (HPLC y LC-MS/MS)
presentan problemas cuando se implementan en
sistemas de control con una gran carga de trabajo, como
es el caso del Intecmar.
Por otra parte, existe unha creciente preocupación por
dos grupos de toxinas emergentes en Europa –el de la
tetrodotoxina y el de la palytoxina- que son de elevada
potencia y que, bien por la expansión de su área de
distribución, bien por su posible introducción en
productos importados, están aumentando su riesgo en
los países de la UE. A pesar de esta preocupación no
existen aún métodos oficiales, ni en algunos casos
materiales de referencia, para llevar a cabo la
cuantificación de estas toxinas, por lo cual resulta
importante contribuír a su desarrollo de forma que los
que finalmente se adopten puedan estar bien adaptados
a la dinámica de un sistema de control como el de
Galicia.
Se continuó desarrollando la metodología cromatográfica
para los análisis de PSP con oxidación pre-columna. La
mejor columna identificada en DEMETOX (Waters
Cortecs T3) resultó soportar un número reducido de
inyecciones (aprox. 300) por lo que fue substituida por
una Atlantis T3 de 3µm y 75mm de longitud, que dio una
buena resolución con cromatogramas de una duración
inferior a 3.5 min, y una vida más larga (más de 900
inyecciones). Se comprobó que las determinaciones
obtenidas por este método coinciden bien con las
obtenidas por el método oficial (con cromatogramas de
14 min).
Se evaluó la posibilidad de substituír el fraccionamiento
de la mezcla de toxinas con SPE por otro con
cromatografía. Se comprobaron diversas fases y
formatos sin resultados satisfactorios en algunos casos.
En los casos en los que el fraccionamiento fue
satisfactorio, tanto el tiempo como el coste por muestra
fueron similares a los del SPE, pero dado que la
separación cromatográfica requeire controles
adicionales, se consideró mejor continuar con la
utilización de SPE.
Se procedió a la purificación por medio de columna de
cromatografía de exclusión molecular Sephadex LH-20
de extractos de Ostreopsis como primer paso para la
obtención de soluciones purificadas de ovatoxinas para
ser usadas como materiales de referencia. Igualmente se
purificaron extractos de Prorocentrum belizeanum, para
purificar diol-ésteres de ácido okadaico y otros derivados
más complejos.
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Mejora analítica de la detección de toxinas en
moluscos bivalvos para maximizar el flujo de trabajo
en sistemas de control.
ACRÓNIMO: MADeTox
INICIO: 2017
FINALIZACIÓN: 2019
ENTIDAD FINANCIADORA:
Consellería do Mar. Xunta de Galicia
Código: CIMA 17/01
INVESTIGADOR PRINCIPAL:
Blanco Pérez, J.C.
INVESTIGADORES/AS:
INTECMAR: Arévalo Bartolomé, F.; Moroño Mariño, A.;
Correa Jorge, J.
APOYO A LA INVESTIGACIÓN:
CIMA: Campaña Ferro, E.; Mariño Cadarso, C.; Martín
Sánchez, H.
INTECMAR: Muñiz Romero, S.

CIMA Memoria 2017 81
8 RECURSOS MARINOS

82
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Evaluación y seguimiento de las Reservas Marinas de
Interés Pesquero de Galicia.
ACRÓNIMO: ESCAMPE 2
INICIO: 2014
FINALIZACIÓN: 2017
ENTIDAD FINANCIADORA:
Consellería do Mar. Xunta de Galicia.
Código: PGIDIT-CIMA 14/05
INVESTIGADOR PRINCIPAL:
Cuiñas Olmedo, P.
INVESTIGADOR:
Febrero Mayor, F.
RESUMEN
Las Reservas Marinas de Interés Pesquero (RMIP) son
figuras administrativas creadas para asegurar la
conservación, la protección y la gestión sostenible de
espacios naturales en los que se lleva a cabo la
explotación pesquera y marisquera de recursos naturales
marinos.
En la actualidad en la Comunidad Autónoma de Galicia
existen dos Reservas Marinas de Interés Pesquero
(RMIP): “Os Miñarzos”, creada en el año 2007 y “Ría de
Cedeira” creada en el año 2009. Cada una de estas
RMIP cuenta con un “Órgano de Gestión Seguimiento y
Control” (Oxesco) constituido por representantes de la
Administración Pesquera de la Xunta de Galicia y
representantes de las Organizaciones Sectoriales de
pesca.
Durante el período de vigencia de esta acción en el año
2017, se continuó dando apoyo científico-técnico a los
Órgaños de Gestión Seguimiento y Control de las RMIPs
de “Os Miñarzos” y “Ría de Cedeira”.
Dentro de la actividad referente a la selección de
indicadores útiles para la evaluación de la evolución de
la actividad pesquera, no fue posible contar con datos
específicos referidos estrictamente a la actividad llevada
a cabo dentro de las reservas. Esto fue debido a la
demora en la instalación de los sistemas de seguimiento
de buques (AIS-VMS), que está previsto que sea
obligatorio para todas las embarcaciones que faenen en
las RMIPs.
Debido a lo anterior, se optó por utilizar los datos del
sistema de registro del programa informático
denominado “Gestión de Cofradías” de la Xunta de
Galicia, del que se obtuvieron datos diarios, del volumen
de capturas (kg), precio de venta en lonja (€) y artes
despachadas para cada embarcación censada en las
RMIPs.
Estos datos no discriminan las capturas hechas dentro y
fuera de la RMIP, por lo que la información corresponde
a las capturas totales de la flota censada en cada RMIP.
La zona de trabajo de estas flotas incluye la RMIP y
otros bancos próximos, por lo que consideramos que los
resultados reflejan el estado de los recursos de una zona
donde se encuentra incluida la reserva.
Como indicadores se contemplaron los siguientes:
- Esfuerzo pesquero: presenta la evolución
del número de embarcaciones/día explotando los
recursos mediante las distintas artes
En las Figuras 1 y 2 se muestra el esfuerzo realizado,
(suma de jornadas totales realizadas por todas las
embarcaciones) con las artes que más se utilizan.
En el caso de la flota de Lira son la nasa de pulpo, los
miños, y las vetas y los recursos específicos (solénidos,
erizo y percebe).
Se puede apreciar que durante el período estudiado se
mantuvo un nivel estable de actividad para todas ellas, lo
que puede indicar que las especies objetivo mantuvieron
poblaciones adecuadas para una explotación rentable.
La nasa de pulpo fue el arte más utilizada con gran
diferencia (con una media anual de 2.646 jornadas
acumuladas, representa el 43% de la totalidad de las
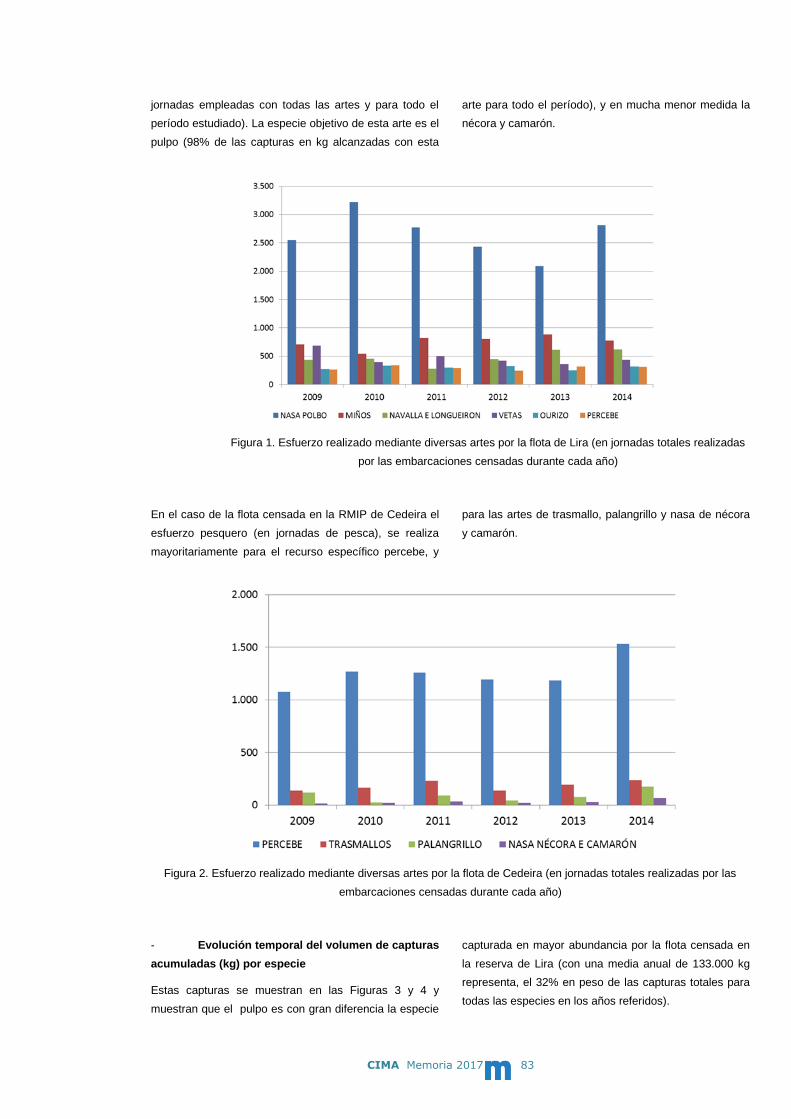
CIMA Memoria 2017 83
jornadas empleadas con todas las artes y para todo el
período estudiado). La especie objetivo de esta arte es el
pulpo (98% de las capturas en kg alcanzadas con esta
arte para todo el período), y en mucha menor medida la
nécora y camarón.
Figura 1. Esfuerzo realizado mediante diversas artes por la flota de Lira (en jornadas totales realizadas
por las embarcaciones censadas durante cada año)
En el caso de la flota censada en la RMIP de Cedeira el
esfuerzo pesquero (en jornadas de pesca), se realiza
mayoritariamente para el recurso específico percebe, y
para las artes de trasmallo, palangrillo y nasa de nécora
y camarón.
Figura 2. Esfuerzo realizado mediante diversas artes por la flota de Cedeira (en jornadas totales realizadas por las
embarcaciones censadas durante cada año)
- Evolución temporal del volumen de capturas
acumuladas (kg) por especie
Estas capturas se muestran en las Figuras 3 y 4 y
muestran que el pulpo es con gran diferencia la especie
capturada en mayor abundancia por la flota censada en
la reserva de Lira (con una media anual de 133.000 kg
representa, el 32% en peso de las capturas totales para
todas las especies en los años referidos).
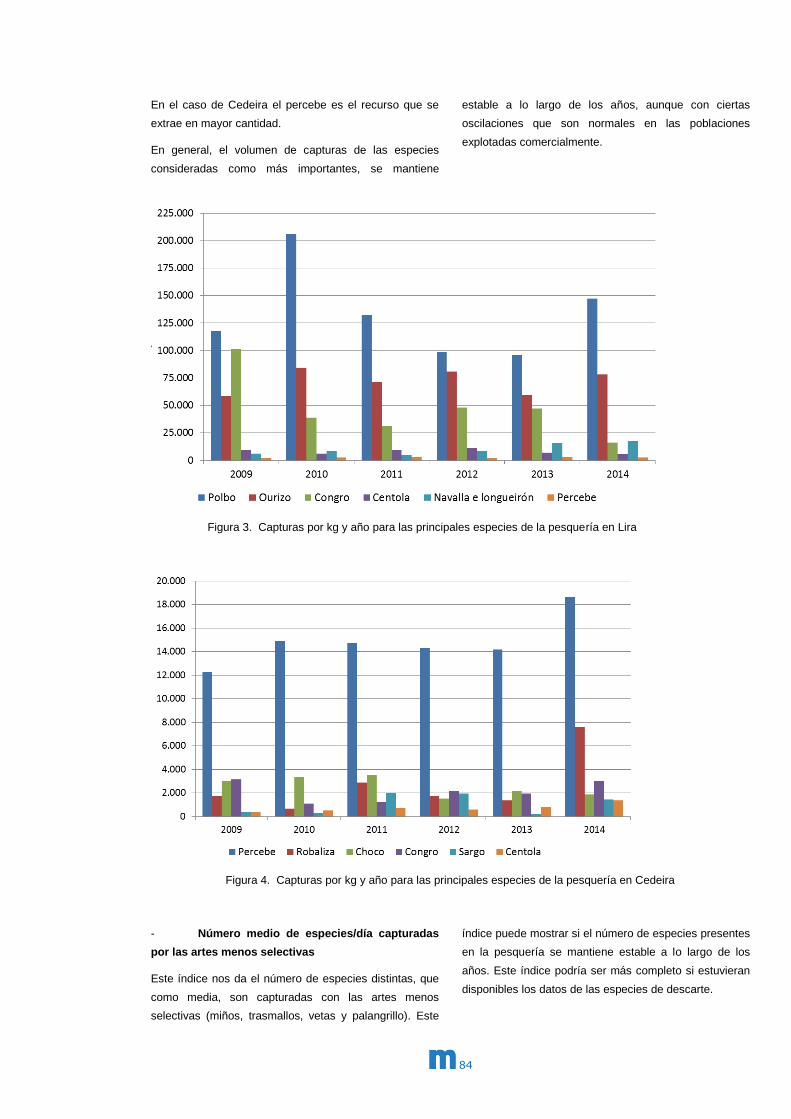
84
En el caso de Cedeira el percebe es el recurso que se
extrae en mayor cantidad.
En general, el volumen de capturas de las especies
consideradas como más importantes, se mantiene
estable a lo largo de los años, aunque con ciertas
oscilaciones que son normales en las poblaciones
explotadas comercialmente.
Figura 3. Capturas por kg y año para las principales especies de la pesquería en Lira
Figura 4. Capturas por kg y año para las principales especies de la pesquería en Cedeira
- Número medio de especies/día capturadas
por las artes menos selectivas
Este índice nos da el número de especies distintas, que
como media, son capturadas con las artes menos
selectivas (miños, trasmallos, vetas y palangrillo). Este
índice puede mostrar si el número de especies presentes
en la pesquería se mantiene estable a lo largo de los
años. Este índice podría ser más completo si estuvieran
disponibles los datos de las especies de descarte.
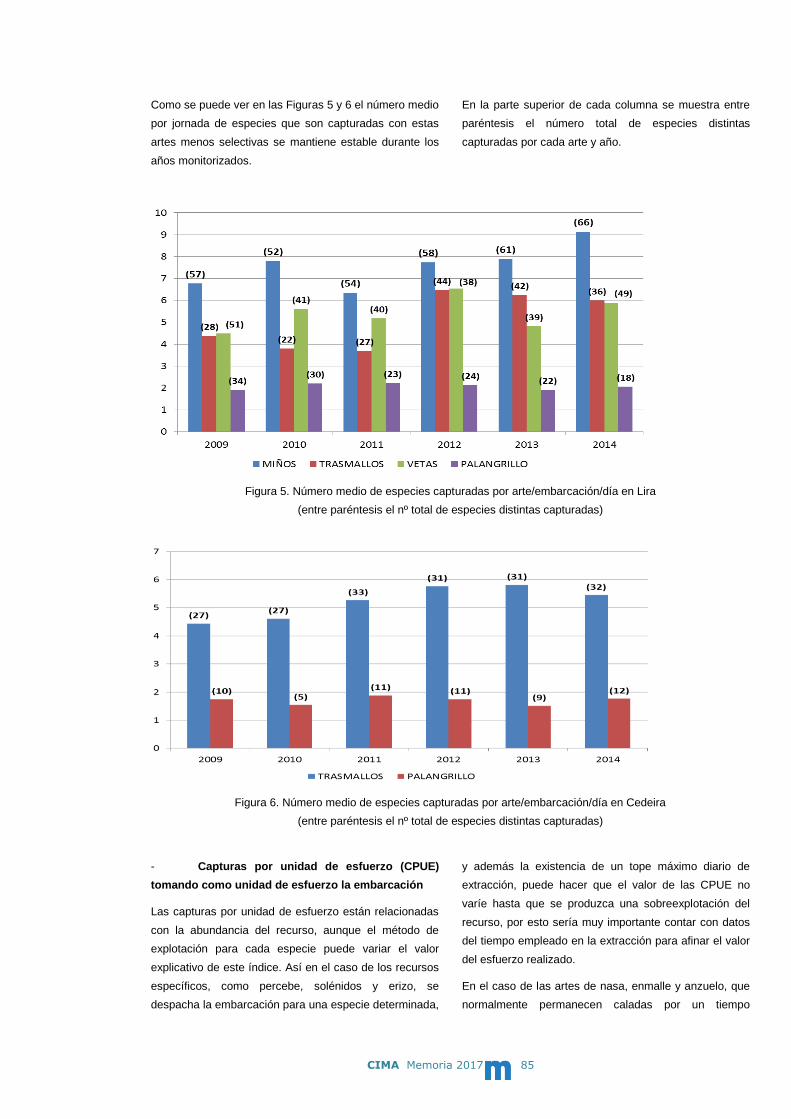
CIMA Memoria 2017 85
Como se puede ver en las Figuras 5 y 6 el número medio
por jornada de especies que son capturadas con estas
artes menos selectivas se mantiene estable durante los
años monitorizados.
En la parte superior de cada columna se muestra entre
paréntesis el número total de especies distintas
capturadas por cada arte y año.
Figura 5. Número medio de especies capturadas por arte/embarcación/día en Lira
(entre paréntesis el nº total de especies distintas capturadas)
Figura 6. Número medio de especies capturadas por arte/embarcación/día en Cedeira
(entre paréntesis el nº total de especies distintas capturadas)
- Capturas por unidad de esfuerzo (CPUE)
tomando como unidad de esfuerzo la embarcación
Las capturas por unidad de esfuerzo están relacionadas
con la abundancia del recurso, aunque el método de
explotación para cada especie puede variar el valor
explicativo de este índice. Así en el caso de los recursos
específicos, como percebe, solénidos y erizo, se
despacha la embarcación para una especie determinada,
y además la existencia de un tope máximo diario de
extracción, puede hacer que el valor de las CPUE no
varíe hasta que se produzca una sobreexplotación del
recurso, por esto sería muy importante contar con datos
del tiempo empleado en la extracción para afinar el valor
del esfuerzo realizado.
En el caso de las artes de nasa, enmalle y anzuelo, que
normalmente permanecen caladas por un tiempo
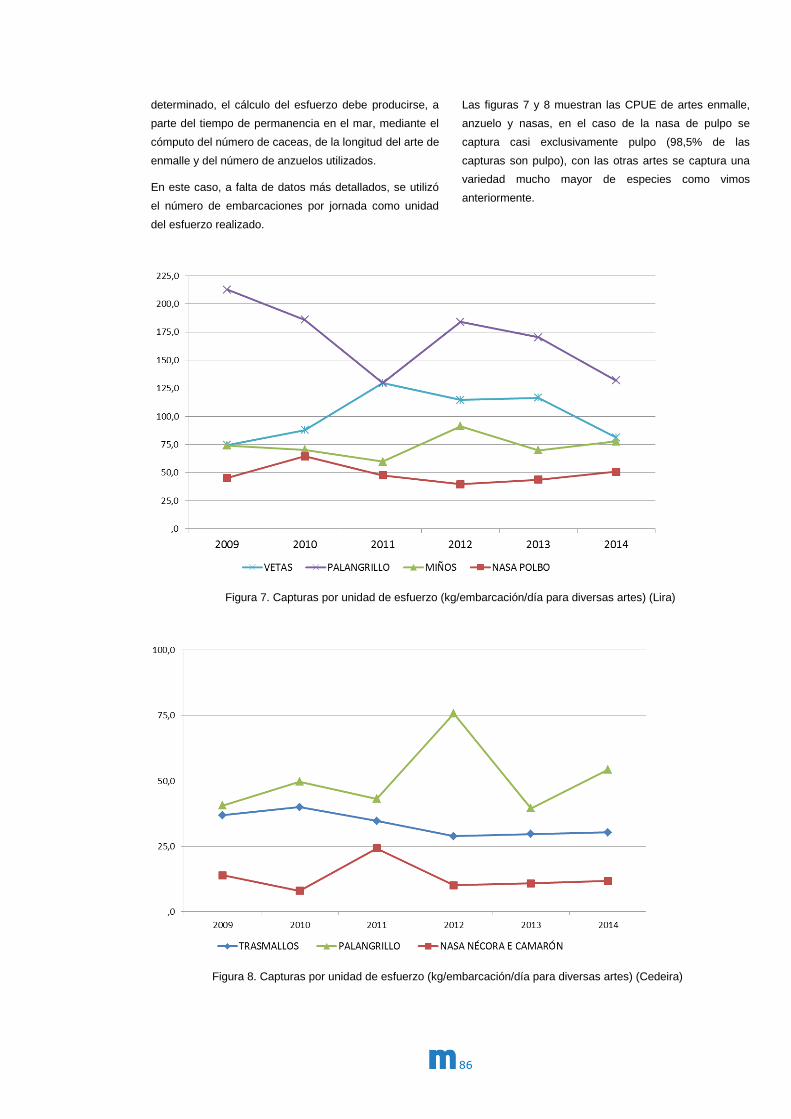
86
determinado, el cálculo del esfuerzo debe producirse, a
parte del tiempo de permanencia en el mar, mediante el
cómputo del número de caceas, de la longitud del arte de
enmalle y del número de anzuelos utilizados.
En este caso, a falta de datos más detallados, se utilizó
el número de embarcaciones por jornada como unidad
del esfuerzo realizado.
Las figuras 7 y 8 muestran las CPUE de artes enmalle,
anzuelo y nasas, en el caso de la nasa de pulpo se
captura casi exclusivamente pulpo (98,5% de las
capturas son pulpo), con las otras artes se captura una
variedad mucho mayor de especies como vimos
anteriormente.
Figura 7. Capturas por unidad de esfuerzo (kg/embarcación/día para diversas artes) (Lira)
Figura 8. Capturas por unidad de esfuerzo (kg/embarcación/día para diversas artes) (Cedeira)

CIMA Memoria 2017 87
Para finalizar es necesario reiterar que los datos
utilizados corresponden a zonas de trabajo donde
operan las flotas censadas en las RMIPs “Os Miñarzos”,
y “Ría de Cedeira” que incluyen las zonas de pesca
adyacentes a las reservas, en algunos casos bastante
apartadas. Por lo que la extrapolación de estos datos a
la zona de reserva debe considerarse como una
interpretación de que los recursos marinos de la zona de
la reserva comparten el estado general de una zona más
amplia, sobre todo para el caso de los recursos
pesqueros con más capacidad de movimiento
(demersales y pelágicos), para los recursos marinos
sésiles, las variaciones espaciales pueden ser mucho
más acusadas.

88

CIMA Memoria 2017 89
RESUMEN
El objetivo de esta acción es determinar las causas
naturales que están afectando a los bancos de Donax
sp. y los efectos que sobre ellos tiene la actividad
humana de diferentes orígenes (industrial, de la
agricultura, minería, construcción, urbanización o
deforestación entre otros). Las causas naturales pueden
estar vinculadas a la naturaleza de los sedimentos, a la
topografía de las playas y a sus cambios por efectos
ambientales y antropológicos, a diferentes toxicidades, a
cambios por causas hidrológicas o climáticas o a
diversas patologías que ya afectan a los individuos en
una talla pequeña. De la misma forma que se pretende
abordar el problema conociendo sus causas, se propone
comenzar a estudiar la posible recuperación futura de los
bancos, haciendo pequeños ensayos piloto de siembras
con semilla obtenida en criadero.
En el banco de Vilarrube se siguieron haciendo
muestreos trimestrales, es decir, en cada estación del
año, desde enero a diciembre de 2017, en los que se
recogieron, en los puntos marcados en toda la extensión
del banco, muestras de los individuos para ver
densidades y tamaño, muestras de sedimento para ver
sus características. Todo esto en cada punto para
después poder relacionarlos entre ellos. También se
estudia el perfil de la playa, la topografía. Además se
continúa con el control de las patologías y de su
reproducción.
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Determinación de las causas de la
disminución de los bancos de coquina.
Condicionantes naturales y
antropogénicos.
INICIO: junio, 2016
FINALIZACIÓN: diciembre, 2018
ENTIDAD FINANCIADORA:
Consellería do Mar. Xunta de Galicia.
Código: CIMA 16/06
COORDINADORA:
Martínez Patiño, D.
INVESTIGADORES/AS:
CIMA: Febrero Mayor, F.; Montes Pérez, J.; Nóvoa Vázquez,
S.; Ojea Martínez, J.
UNI VIGO: Nombela Castaño, M.A.
CSIC: Álvarez Salgado, X.A.; Otero Villar, J.
Xefatura Territorial de Celeiro. Consellería do Mar: Cerdeira
Arias, D.; Teijeiro Golpe, O.
Xefatura Territorial de Vigo. Consellería do Mar: Rodríguez
González, L.M.
Xefatura Comarcal de Ferrol. Consellería do Mar: Brezmes
Comesaña, C.; Liñán Gil, I.M.
USC: Barja Pérez, J.L.; Dubert, J.; Olveira, G.; Prado Plana,
S.
ASISTENCIAS TÉCNICAS:
Cofradía de O Vicedo: Gómez Brandariz, D.
Cofradía de O Barqueiro: del Río Rocha, G.
Cofradía de Cedeira: Quintana Couto, A.
APOYO A LA INVESTIGACIÓN:
CIMA: Bouzamayor Yáñez, V.; Cotarelo Jardón, M.J.;
Meléndez Ramos, I.; Páez Pérez, J.; Penas Pampín, E.; Ruíz
Misioné, M.
Xefatura Territorial de Celeiro. Consellería do Mar: Martín
Salas, F.
BECARIOS/AS:
Castilla del Río, S.; Granjel Piñeiro, E.; Matesanz Salgado, D.
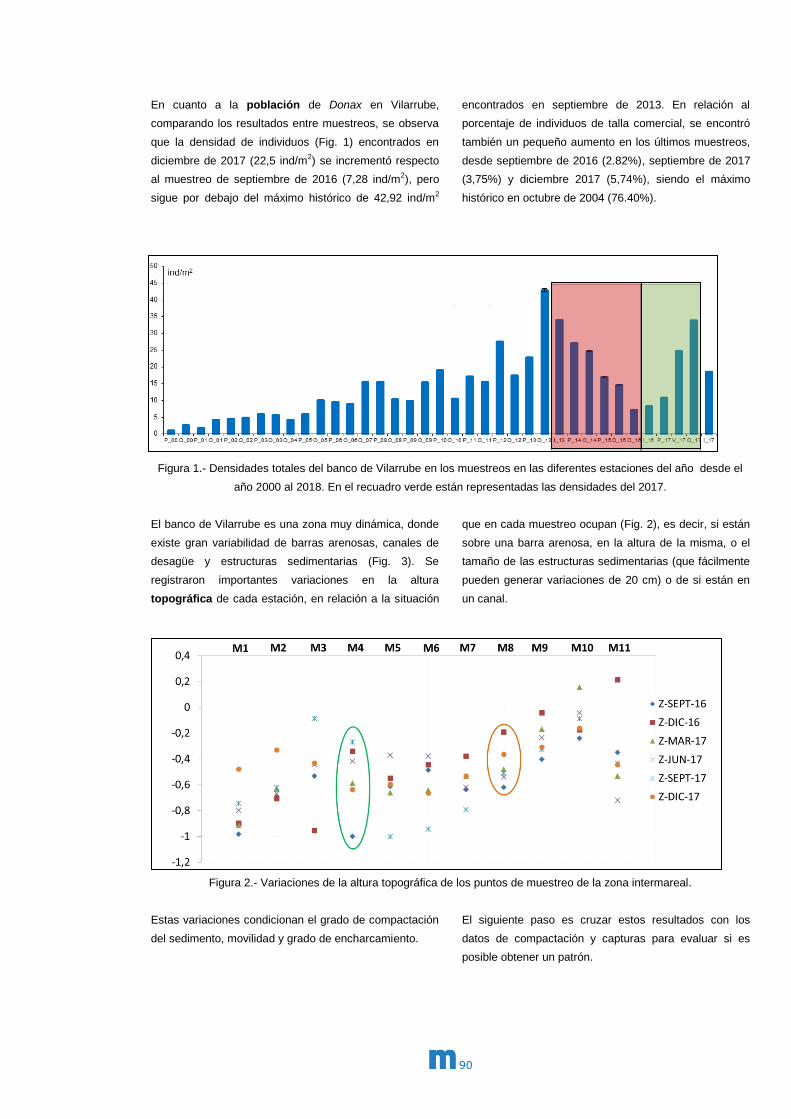
90
En cuanto a la población de Donax en Vilarrube,
comparando los resultados entre muestreos, se observa
que la densidad de individuos (Fig. 1) encontrados en
diciembre de 2017 (22,5 ind/m2) se incrementó respecto
al muestreo de septiembre de 2016 (7,28 ind/m2), pero
sigue por debajo del máximo histórico de 42,92 ind/m2
encontrados en septiembre de 2013. En relación al
porcentaje de individuos de talla comercial, se encontró
también un pequeño aumento en los últimos muestreos,
desde septiembre de 2016 (2.82%), septiembre de 2017
(3,75%) y diciembre 2017 (5,74%), siendo el máximo
histórico en octubre de 2004 (76.40%).
Figura 1.- Densidades totales del banco de Vilarrube en los muestreos en las diferentes estaciones del año desde el
año 2000 al 2018. En el recuadro verde están representadas las densidades del 2017.
El banco de Vilarrube es una zona muy dinámica, donde
existe gran variabilidad de barras arenosas, canales de
desagüe y estructuras sedimentarias (Fig. 3). Se
registraron importantes variaciones en la altura
topográfica de cada estación, en relación a la situación
que en cada muestreo ocupan (Fig. 2), es decir, si están
sobre una barra arenosa, en la altura de la misma, o el
tamaño de las estructuras sedimentarias (que fácilmente
pueden generar variaciones de 20 cm) o de si están en
un canal.
Figura 2.- Variaciones de la altura topográfica de los puntos de muestreo de la zona intermareal.
Estas variaciones condicionan el grado de compactación
del sedimento, movilidad y grado de encharcamiento.
El siguiente paso es cruzar estos resultados con los
datos de compactación y capturas para evaluar si es
posible obtener un patrón.
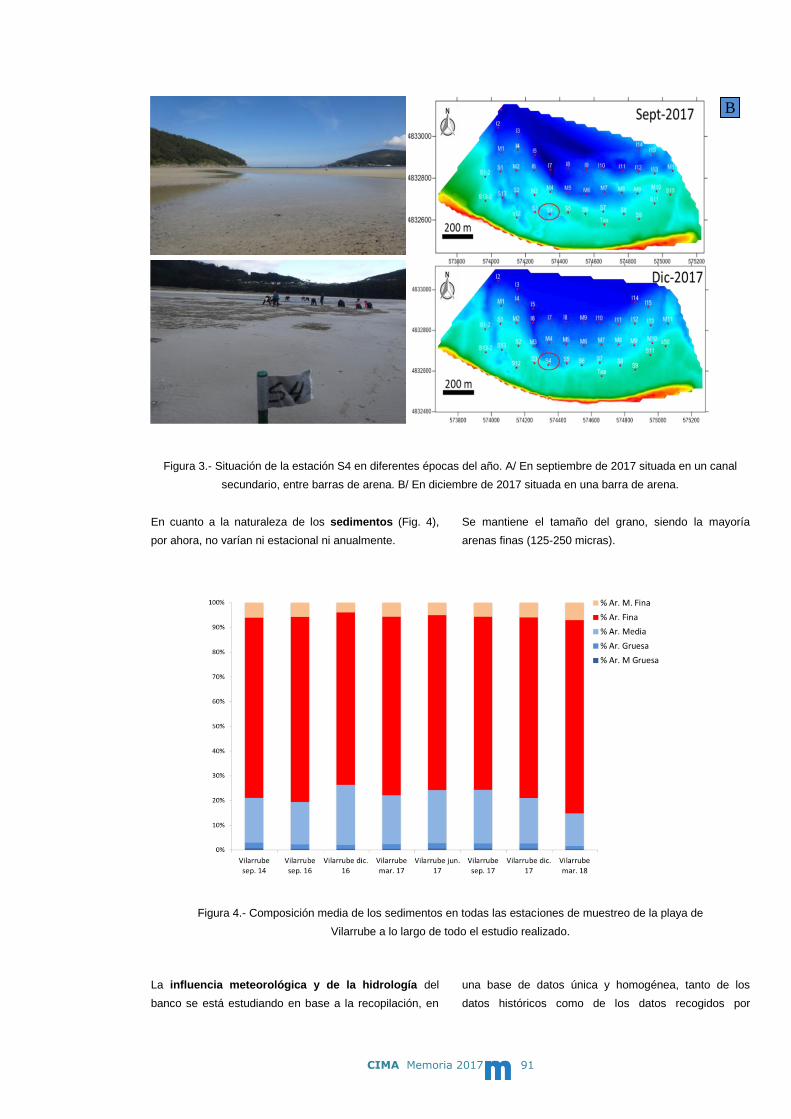
CIMA Memoria 2017 91
Figura 3.- Situación de la estación S4 en diferentes épocas del año. A/ En septiembre de 2017 situada en un canal
secundario, entre barras de arena. B/ En diciembre de 2017 situada en una barra de arena.
En cuanto a la naturaleza de los sedimentos (Fig. 4),
por ahora, no varían ni estacional ni anualmente.
Se mantiene el tamaño del grano, siendo la mayoría
arenas finas (125-250 micras).
Figura 4.- Composición media de los sedimentos en todas las estaciones de muestreo de la playa de
Vilarrube a lo largo de todo el estudio realizado.
La influencia meteorológica y de la hidrología del
banco se está estudiando en base a la recopilación, en
una base de datos única y homogénea, tanto de los
datos históricos como de los datos recogidos por
A
B

92
nosotros de densidades y estructura de tamaños de las
poblaciones de coquina. También de los datos históricos
y actuales de las extracciones y de las condiciones
ambientales (meteorológicas, hidrológicas e
hidrográficas) que les afectan.
Hasta ahora, de los resultados obtenidos del
cruzamiento de estos datos, se puede observar que hay
mucha variación pero, parece ser que: las menores
densidades están en la zona central del banco; las tallas
más grandes están en el medio; la ría de Cedeira tiene
una mayor influencia oceánica que continental= A la ría
le afectan más los afloramientos oceánicos que los
aportes de los ríos.
Para el estudio de la patología, en el año 2017 se
realizaron un total de 5 muestreos en playa América y de
4 muestreos en Vilarrube. El total de ejemplares
procesados fue de 131 en playa América (entre 15-20
mm), y de 91 en Vilarrube (más de 25 mm). También se
procesaron 9 ejemplares de A Lanzada y 49 ejemplares
de Corrubedo procedentes de muestreos puntuales. En
ambos casos la talla era muy variable.
Hasta el momento no se han detectado parasitologías ni
alteraciones patológicas, que pudieran considerarse
como preocupantes para los bancos de coquina.
Tampoco, en la actualidad se han detectado infecciones
por herpesvirus.
En el estudio realizado en este año sobre su ciclo
reproductivo (Fig. 5), se vio que los resultados eran
coincidentes con otros anteriores de la población de
Vilarrube.
Figura 5.- Representación de los estadíos del ciclo reproductivo de la coquina en las tallas (10-15; 15-20; 20-25; 25-30; 30-35;>35) desde marzo a septiembre de 2017. E0: Reposo sexual; E1: Inicio del desarrollo gametogénico; E2:
Gametogénesis avanzada; E3: Madurez y puesta; E4: Postpuesta.
Los individuos capturados en los muestreos del banco de
Vilarrube del año 2017 fueron estabulados en el CIMA de
Ribadeo obteniendo desoves desde febrero hasta junio.
A partir del cultivo larvario de dos de estos desoves (de
mayo y junio) se consiguió semilla de coquina. Se hizo
una siembra de esta semilla marcada con hojas de
poliéster pegadas con pegamento de cianocrilato, en la
playa de Abrela (ría de Viveiro).
Se realizaron muestreos de las poblaciones de los
distintos bancos naturales del Cantábrico Gallego: la
playa de Vilela y el Lombo das Navallas (ría de O
Barqueiro); las playas de Abrela y de Area (ría de
Viveiro) y la playa de Altar (ría de Foz). Los últimos
muestreos efectuados en las playas este pasado otoño a
lo largo del litoral cantábrico gallego, muestran datos
bajos de densidad (ind/m2) y poca presencia de adultos
(% comercial), como se aprecian en la tabla I. Aún así,
se constata una ligera recuperación de los bancos, con
un aumento relativo de la densidad y del porcentaje
comercial de las poblaciones de coquina.
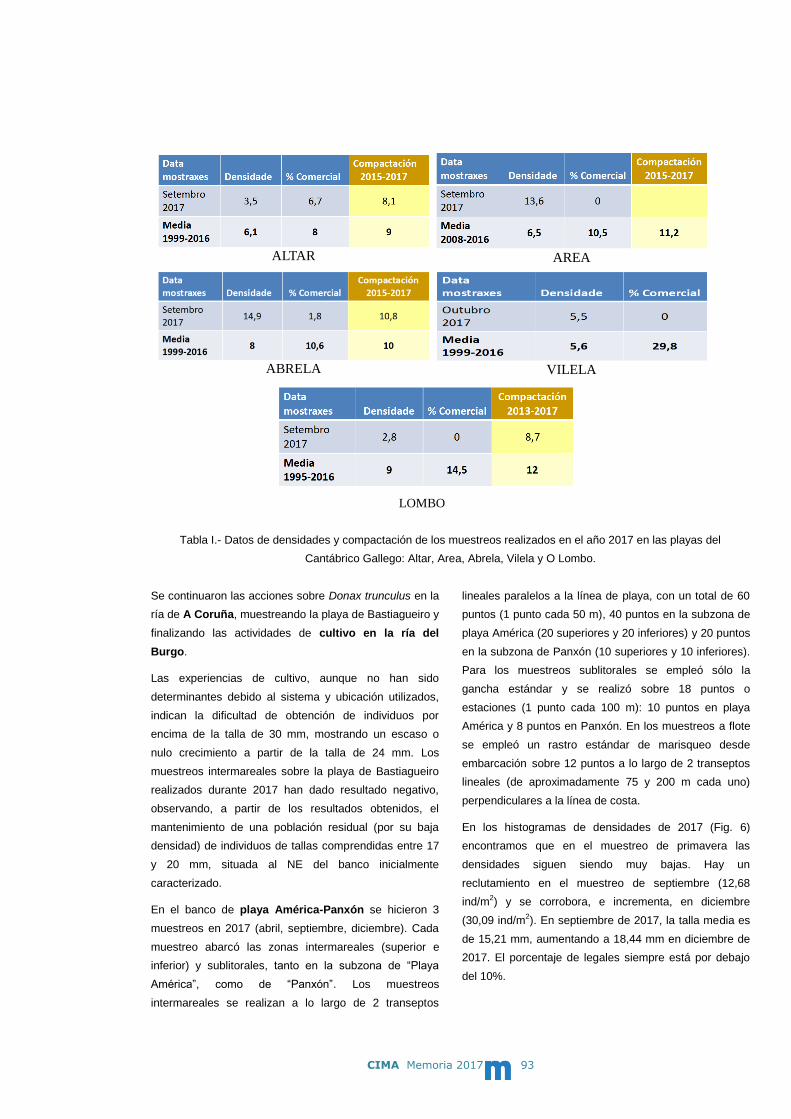
CIMA Memoria 2017 93
Tabla I.- Datos de densidades y compactación de los muestreos realizados en el año 2017 en las playas del
Cantábrico Gallego: Altar, Area, Abrela, Vilela y O Lombo.
Se continuaron las acciones sobre Donax trunculus en la
ría de A Coruña, muestreando la playa de Bastiagueiro y
finalizando las actividades de cultivo en la ría del
Burgo.
Las experiencias de cultivo, aunque no han sido
determinantes debido al sistema y ubicación utilizados,
indican la dificultad de obtención de individuos por
encima de la talla de 30 mm, mostrando un escaso o
nulo crecimiento a partir de la talla de 24 mm. Los
muestreos intermareales sobre la playa de Bastiagueiro
realizados durante 2017 han dado resultado negativo,
observando, a partir de los resultados obtenidos, el
mantenimiento de una población residual (por su baja
densidad) de individuos de tallas comprendidas entre 17
y 20 mm, situada al NE del banco inicialmente
caracterizado.
En el banco de playa América-Panxón se hicieron 3
muestreos en 2017 (abril, septiembre, diciembre). Cada
muestreo abarcó las zonas intermareales (superior e
inferior) y sublitorales, tanto en la subzona de “Playa
América”, como de “Panxón”. Los muestreos
intermareales se realizan a lo largo de 2 transeptos
lineales paralelos a la línea de playa, con un total de 60
puntos (1 punto cada 50 m), 40 puntos en la subzona de
playa América (20 superiores y 20 inferiores) y 20 puntos
en la subzona de Panxón (10 superiores y 10 inferiores).
Para los muestreos sublitorales se empleó sólo la
gancha estándar y se realizó sobre 18 puntos o
estaciones (1 punto cada 100 m): 10 puntos en playa
América y 8 puntos en Panxón. En los muestreos a flote
se empleó un rastro estándar de marisqueo desde
embarcación sobre 12 puntos a lo largo de 2 transeptos
lineales (de aproximadamente 75 y 200 m cada uno)
perpendiculares a la línea de costa.
En los histogramas de densidades de 2017 (Fig. 6)
encontramos que en el muestreo de primavera las
densidades siguen siendo muy bajas. Hay un
reclutamiento en el muestreo de septiembre (12,68
ind/m2) y se corrobora, e incrementa, en diciembre
(30,09 ind/m2). En septiembre de 2017, la talla media es
de 15,21 mm, aumentando a 18,44 mm en diciembre de
2017. El porcentaje de legales siempre está por debajo
del 10%.
LOMBO
ALTAR AREA
ABRELA VILELA

94
Figura 6.- Densidades (ind/m2) de la población de playa América a lo largo de los muestreos realizados en el año 2017.
La población de coquina del banco de playa América-
Panxón se encuentra en un estado de supervivencia,
donde observamos reclutamientos no continuados (cada
3 años: 2014, 2017). Hasta el momento, todo parece
indicar que, una vez realizados los desoves los
individuos progenitores desaparecen. Se observó en
este año de estudio, un aumento del número de
individuos desde el mes de abril, que se incrementa en el
mes de diciembre. Desde el comienzo de este estudio
(2013), estos son los datos más favorables hasta el
momento.
Se realizaron muestreos intermareales en otras dos
playas. En la playa A Ladeira (Corrubedo) el 21 de
diciembre de 2017 sobre 12 puntos, con una separación
aproximada de 50 m y en la playa de A Lanzada el 18
de diciembre de 2017 sobre 11 puntos, con una
separación aproximada de 50 m, empleando en ambos
casos una gancha.
Los resultados obtenidos con estos muestreos se
muestran en la siguiente tabla:
A LANZADA A LADEIRA (CORRUBEDO)
Talla Media (mm) 21,71 Talla Media (mm) 25,84
% Comerciales (>35mm) 15,63% % Comerciales (>35mm) 20,00%

RESUMEN
En Galicia coexisten dos especies de berberecho:
Cerastoderma edule y Cerastoderma glaucum. Las
poblaciones naturales de C. edule están
tradicionalmente afectadas por episodios recurrentes de
mortalidad (Villalba et al., 2001), de los cuales siempre
se recuperaron con rapidez. Sin embargo, a partir del
año 2012, gran parte de las poblaciones de C. edule de
las rías gallegas están siendo diezmadas por el parásito
Marteilia cochillia, (Villalba et al., 2014), sin que las
poblaciones de C. glaucum se vean afectadas. La lucha
contra este tipo de enfermedades en el medio natural
usando agentes terapéuticos es compleja. Una forma
diferente de lucha es mediante programas de selección,
bien a través de la obtención de estirpes tolerantes a la
enfermedad (da Silva et al., 2005; Fuentes y Villalba,
2007) o bien mediante la producción de variedades de
rápido crecimiento que alcancen la talla comercial antes
de la aparición de los episodios de Marteiliosis. En una
anterior acción de investigación (PGIDIT-CIMA 14/03;
Puesta a punto de las técnicas de cultivo en mini-
criadero de las especies del género Cerastoderma de
Galicia) demostramos que es posible obtener, mediante
cultivo suspendido en batea, individuos de berberecho,
Cerastoderma edule, de talla comercial antes de la
aparición de los graves brotes de mortalidad
ocasionados por el parásito Marteilia cochillia.
El objetivo principal de esta nueva acción de
investigación es realizar un proceso de escalado y
probar una estrategia semi-industrial de engorde en
batea de semilla de berberecho producida en criadero.
Esta prueba se está llevando a cabo en colaboración
con la Cofradía de Pescadores “San Antonio” de
Cambados y los resultados no son todavía definitivos.
Otro de los objetivos de esta acción de investigación es
la optimización de las técnicas de maduración artificial
en criadero (acondicionamiento) mediante la realización
de un estudio comparado del proceso de maduración
entre un sistema en abierto (FTS) y un sistema en
recirculación (RAS). Para ello acondicionamos en
criadero dos lotes de berberechos procedentes de zonas
libres de Marteilia, un lote de la desembocadura del río
Anllóns (CE-RA) y otro de la ría de Noia (CE-N). En
ambos casos utilizamos dos sistemas alternativos: un
sistema en circuito abierto (FTS, Imagen 1) y un sistema
en recirculación (RAS, Imagen 2), este último conectado
a una unidad con filtro biológico (Imagen 3). En ambos
casos se suministró agua de mar filtrada por 1 micra,
esterilizada por UVA y calentada progresivamente hasta
alcanzar los 16ºC. Los individuos acondicionados se
alimentaron, mediante una bomba dosificadora, con una
mezcla de seis especies diferentes de microalgas. El
acondicionamiento del lote CE-RA se inició el 1 de
diciembre de 2016 y el del lote CE-N el 3 de febrero de
2017.
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Engorde en batea de semilla de berberecho producida en
criadero. Mejora de la metodología de cultivo.
INICIO: octubre, 2016
FINALIZACIÓN: diciembre, 2018
ENTIDADE FINANCIADORA:
Consellería do Mar. Xunta de Galicia
Código: CIMA 16/10
INVESTIGADOR PRINCIPAL:
Fuentes González, J. M.
INVESTIGADORES/AS:
Cao Hermida, M.A.; Iglesias Estepa, D.; Mariño Balsa,
J. C.
APOYO A LA INVESTIGACIÓN:
Fernández Abuín, I.; Giráldez Rivero, R.; Gregorio
Chenlo, M.V.; Solla Vicente, M.C.

Imagen 1. Cubetas en el sistema en circuito abierto (FTS)
Imagen 2. Cubetas en el sistema en recirculación (RAS)

CIMA Memoria 2017 97
Imagen 3. Unidad de filtración biológica conectada al sistema RAS
La mortalidad de los individuos acondicionados fue muy
diferente en función del sistema de acondicionamiento
utilizado (Figura 1). En FTS la mortalidad final fue muy
alta, del 100% en el lote CE-RA y del 96% en el lote CE-
N, transcurridas 8 y 4 semanas de acondicionamiento,
respectivamente. Por el contario, la mortalidad final en
RAS fue mucho menor (59% para el lote CE-RA y del
76% para el lote CE-N), pero el acondicionamiento pudo
prolongarse durante más tiempo (15 y 8 semanas,
respectivamente).
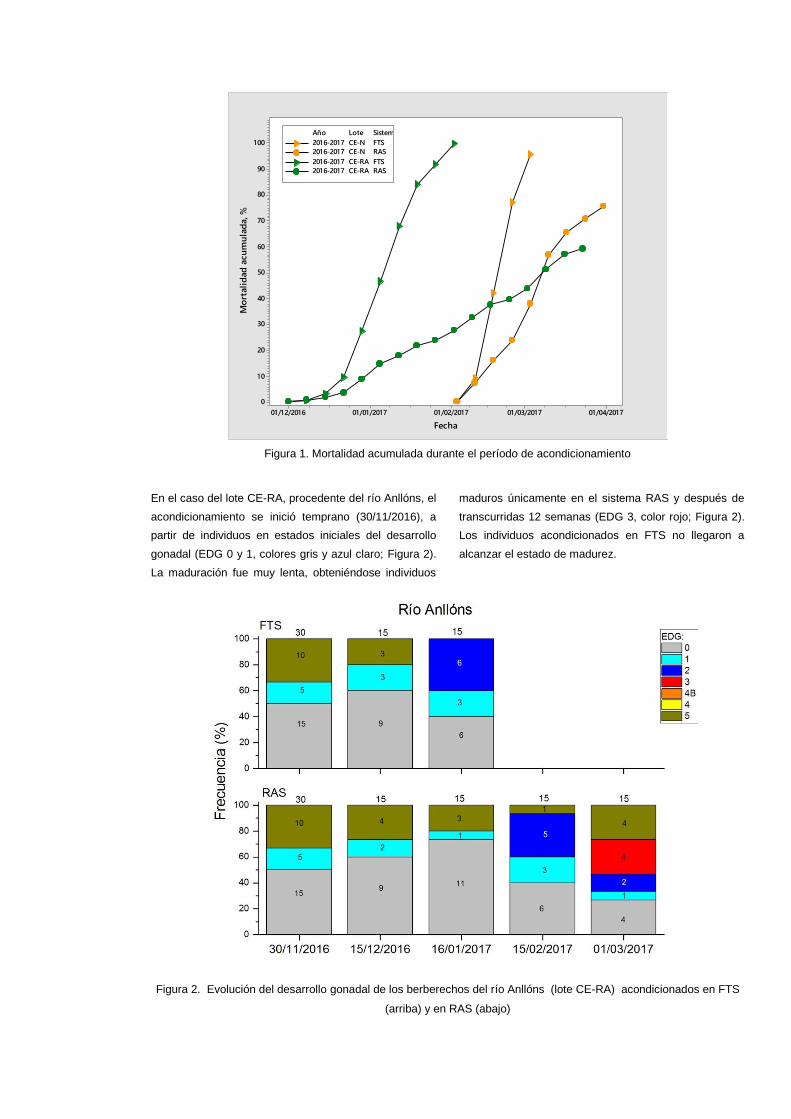
Figura 1. Mortalidad acumulada durante el período de acondicionamiento
En el caso del lote CE-RA, procedente del río Anllóns, el
acondicionamiento se inició temprano (30/11/2016), a
partir de individuos en estados iniciales del desarrollo
gonadal (EDG 0 y 1, colores gris y azul claro; Figura 2).
La maduración fue muy lenta, obteniéndose individuos
maduros únicamente en el sistema RAS y después de
transcurridas 12 semanas (EDG 3, color rojo; Figura 2).
Los individuos acondicionados en FTS no llegaron a
alcanzar el estado de madurez.
Figura 2. Evolución del desarrollo gonadal de los berberechos del río Anllóns (lote CE-RA) acondicionados en FTS
(arriba) y en RAS (abajo)
01/04/201701/03/201701/02/201701/01/201701/12/2016
100
90
80
70
60
50
40
30
20
10
0
Fecha
Mo
rtali
dad
acu
mu
lad
a, %
2016-2017 CE-N FTS
2016-2017 CE-N RAS
2016-2017 CE-RA FTS
2016-2017 CE-RA RAS
Año Lote Sistem
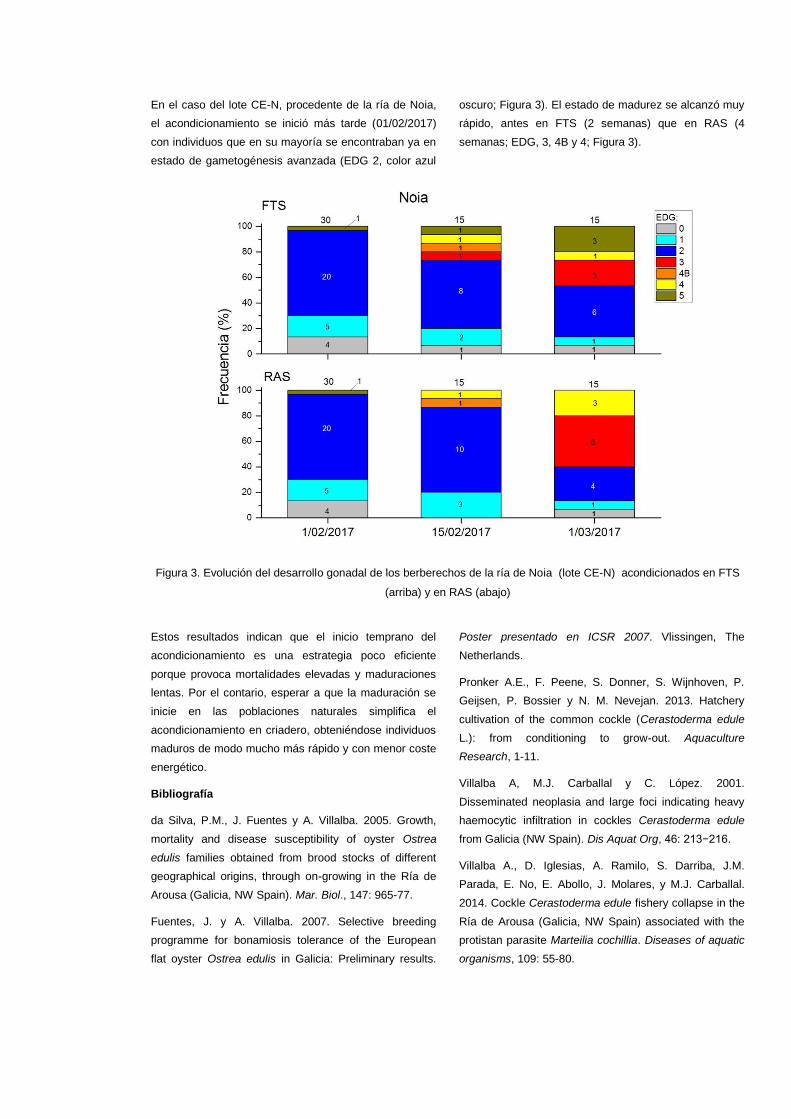
En el caso del lote CE-N, procedente de la ría de Noia,
el acondicionamiento se inició más tarde (01/02/2017)
con individuos que en su mayoría se encontraban ya en
estado de gametogénesis avanzada (EDG 2, color azul
oscuro; Figura 3). El estado de madurez se alcanzó muy
rápido, antes en FTS (2 semanas) que en RAS (4
semanas; EDG, 3, 4B y 4; Figura 3).
Figura 3. Evolución del desarrollo gonadal de los berberechos de la ría de Noia (lote CE-N) acondicionados en FTS
(arriba) y en RAS (abajo)
Estos resultados indican que el inicio temprano del
acondicionamiento es una estrategia poco eficiente
porque provoca mortalidades elevadas y maduraciones
lentas. Por el contario, esperar a que la maduración se
inicie en las poblaciones naturales simplifica el
acondicionamiento en criadero, obteniéndose individuos
maduros de modo mucho más rápido y con menor coste
energético.
Bibliografía
da Silva, P.M., J. Fuentes y A. Villalba. 2005. Growth,
mortality and disease susceptibility of oyster Ostrea
edulis families obtained from brood stocks of different
geographical origins, through on-growing in the Ría de
Arousa (Galicia, NW Spain). Mar. Biol., 147: 965-77.
Fuentes, J. y A. Villalba. 2007. Selective breeding
programme for bonamiosis tolerance of the European
flat oyster Ostrea edulis in Galicia: Preliminary results.
Poster presentado en ICSR 2007. Vlissingen, The
Netherlands.
Pronker A.E., F. Peene, S. Donner, S. Wijnhoven, P.
Geijsen, P. Bossier y N. M. Nevejan. 2013. Hatchery
cultivation of the common cockle (Cerastoderma edule
L.): from conditioning to grow-out. Aquaculture
Research, 1-11.
Villalba A, M.J. Carballal y C. López. 2001.
Disseminated neoplasia and large foci indicating heavy
haemocytic infiltration in cockles Cerastoderma edule
from Galicia (NW Spain). Dis Aquat Org, 46: 213−216.
Villalba A., D. Iglesias, A. Ramilo, S. Darriba, J.M.
Parada, E. No, E. Abollo, J. Molares, y M.J. Carballal.
2014. Cockle Cerastoderma edule fishery collapse in the
Ría de Arousa (Galicia, NW Spain) associated with the
protistan parasite Marteilia cochillia. Diseases of aquatic
organisms, 109: 55-80.

100
RESUMEN
La primera tarea realizada fue la adquisición de material
y montaje de las series de acuarios necesarios para las
pruebas experimentales previstas (Fig.1).
Figura 1.- Acuarios en funcionamiento en laboratorio del CIMA.
Los trabajos de investigación realizados, de acuerdo a
los objetivos enumerados en la acción, fueron:
- Muestreos en el medio natural: ensenada y
litoral de O Grove, litoral de Cambados, bateas
cercanas y zona de O Galiñeiro, y zona interna
de la ría de Vigo, con nasas, rastro de vieira y
buzos de navaja para posible localización de
Rapana venosa, cangrejo azul (Callinectes
sapidus) y otras especies exóticas.
- Muestreos en zona rocosa de la costa lucense
de Stramonita haemastoma.
- Experiencias de acuarios de tasa alimentaria a
diferentes temperaturas (CIMA-Vilaxoán) y
seguimiento de ciclo reproductivo de
Stramonita haemastoma (CIMA–Ribadeo).
- Reuniones con responsables del sector para
informar de la acción y solicitar su colaboración
(Consello Regulador do Mexillón y Cofradías).
ACCIÓN DE INVESTIGACIÓN EQUIPO
TÍTULO:
Estudio de la presencia de Rapana venosa en la ría
de Arousa, expansión de Stramonita hemastoma en
la costa lucense y litoral de Galicia y Callinectes
sapidus, en base a la evolución de su distribución
actual, potencial reproductivo, crecimiento y
capacidad alimentaria. Análisis de riesgos.
INICIO: 2017
FINALIZACIÓN: 2019
ENTIDADE FINANCIADORA:
Consellería do Mar. Xunta de Galicia
Código: CIMA 17/03
INVESTIGADOR PRINCIPAL:
De Coo Martín, A.
INVESTIGADORES/AS:
Fernández Álvarez, A.; García Fernández, A.

CIMA Memoria 2017 101
Figura 2.- Larvas recién eclosionadas de ooteca de Stramonita
Resultados provisionales alcanzados:
- No se encontraron, por el momento ejemplares de
Rapana ni de Cangrejo azul.
- Se está elaborando un mapa de la expansión de
Stramonita en la costa lucense.
- Se detectó una expansión de Crepipatella dilatata en
los fondos de batea y se detectaron los primeros
indicios de que esta especie está comenzando a
instalarse en los mejillones cultivados en cuerda.
- Se inició la comprobación de la posible presencia de
Caprella mutica en bancos de navaja y longueirón en la
Ensenada de O Grove. Pendiente de confirmación
directa.
- Las diferencias de tasas alimentarias con la
temperatura son muy notorias (Fig.3).
- Se sigue la puesta de Stramonita en el CIMA de
Ribadeo.
- Para las reuniones con el sector se elaboró un póster
de diverso material (Fig.4) con muy buena receptividad.
Figura 3.- Consumo a diferente temperatura de Stramonita en acuarios
0
5
10
15
20
25
30
35
% V
ian
da
/d
ia
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Probas
Acuario 1-Tª-18ºc Acuario 2-Tª-15ºC Acuario 3-Tª-12ºC

102
Figura 4.- Material informativo
Asímismo, se realizaron los muestreos en las playas de
Vilaxoán próximas al CIMA para continuar el
seguimiento de las poblaciones de longueirón vello y
navaja que se está realizando de forma ininterrumpida
desde hace años (Fig.5).
Figura 5.- Diferentes clases de edad de población de longueirón vello y C1+ de navaja.

CIMA Memoria 2017 103
9 ARTÍCULOS EN REVISTAS SCI
AFONSO, C., CARDOSO, C., FREIRE M. SILVA, I.E. LINARES, F., VILLANUEVA,
J.L.R., VALENTE, L.M.P., BANDARRA, N.M. The impact of alternative dietary lipids
on the in vitro bioaccessibility of sole fillets for human consumption. Aquaculture, 474:
66-74.
AMARO, R., BOUZA, C., GÓMEZ, B., CASTRO, J., SAN MIGUEL, E., VILLALBA, A.,
LOIS, S., OUTEIRO, A., ONDINA, P. Identification of novel gender-associated
mitochondrial haplotypes in Margaritifera margaritifera (Linnaeus, 1758). Zoological Journal of the Linnean Society,
179:738-750.
BLANCO, J., ARÉVALO, F., MOROÑO, A., CORREA, J., MUÑÍZ, S., MARIÑO, C., ET AL. Presence of azaspiracids in
bivalve molluscs from Northern Spain. Toxicon, 137: 135–143.
BLANCO, J., MARTÍN, H., & MARIÑO, C. Reduction of diarrhetic shellfish poisoning (DSP) toxins accumulation in
cultured mussels by means of rope clustering and hydrodynamic barriers. Aquaculture, 479, 120–124.
CARBALLAL, M.J., IGLESIAS, D., DARRIBA, S., CAO, A., MARIÑO, J.C., RAMILO, A., NO, E., VILLALBA, A.
Parasites and pathological conditions of the lagoon cockle Cerastoderma glaucum from Galicia (NW Spain) and its
resistance to Marteilia cochillia. Diseases of Aquatic Organisms, 122:137-152.
CARNEGIE, R.B., DUNGAN, C.F., VILLALBA, A., REECE, K.S. Honored life member Eugene M. Burreson March 25,
2009. Journal of Shellfish Research 36:1-2.
DÍAZ, S., VILLALBA, A., CARBALLAL, M.J. Assessment of transmissibility of the disseminated neoplasia affecting
cockles Cerastoderma edule in Galicia (NW Spain). Bulletin of the European Association of Fish Pathologists, 37:5-15.
HASANUZZAMAN, A.F.M., GÓMEZ-TATO, A., ALVAREZ DIOS, J.A., HARRISON, P.W., CAO, A., FERNÁNDEZ-
BOO, S., VILLALBA, A., PARDO, B.G., MARTÍNEZ, P. Transcriptomic profile of Manila clam (Ruditapes philippinarum)
haemocytes in response to Perkinsus olseni infection. Aquaculture, 467: 170-181.
HASANUZZAMAN, A.F.M., RUBIOLO, J.A., ROBLEDO, D., GÓMEZ-TATO, A., FERNÁNDEZ-BOO, S., CAO, A.,
VILLALBA, A., PARDO, B.G., MARTÍNEZ, P. Gene expression analysis of Ruditapes philippinarum haemocytes after
experimental Perkinsus olseni zoospore challenge and infection in the wild. Fish and Shellfish Immunology. En prensa.
MENDES, M.C. DE Q., NUNES, J.M.C., MENEZES, M., FRAGA, S., RODRÍGUEZ, F., VÁZQUEZ, J.A., ET AL. Toxin
production, growth kinetics and molecular characterization of Ostreopsis cf. ovata isolated from Todos os Santos Bay,
tropical southwestern Atlantic. Toxicon, 13: 18–30.
NANTÓN, A., ARIAS-PÉREZ, A., FREIRE, R., FERNÁNDEZ-PÉREZ, J., NÓVOA, S., MÉNDEZ, J. Microsatellite
variation in Donax trunculus from the Iberian Peninsula, with particular attention to Galician estuaries (NW Spain).
Estuarine, Coastal and Shelf Science, 197: 27-34.
PAZOS, A.J., VENTOSO, P., MARTÍNEZ-ESCAURIAZA, R., PEREZ-PARALLÉ, M.L., BLANCO, J., TRIVIÑO, J.C.,
ET AL. Transcriptional response after exposure to domoic acid-producing Pseudo-nitzschia in the digestive gland of
the mussel Mytilus galloprovincialis. Toxicon, 140: 60–71.

104
RAMILO, A., VILLALBA, A., ABOLLO, E., CARBALLAL, M.J. A Minchinia mercenariae-like parasite infects cockles
Cerastoderma edule in Galicia (NW Spain). Journal of Fish Diseases, 41:41-48.
STENTIFORD, G.D., RAMILO, A., ABOLLO, E., KERR, R., BATEMAN, K.S., FEIST, S.W., BASS, D., VILLALBA, A.
Hyperspora aquatica n.gn., n.sp. (Microsporidia), hyperparasitic in Marteilia cochillia (Paramyxida) is closely related to
crustacean-infecting microsporidian taxa. Parasitology, 144:186-199.
TILLMANN, U., JAEN, D., FERNANDEZ, L., GOTTSCHLING, M., WITT, M., BLANCO, J., ET AL. Amphidoma
languida (Amphidomatacea, Dinophyceae) with a novel azaspiracid toxin profile identified as the cause of molluscan
contamination at the Atlantic coast of southern Spain. Harmful Algae, 6.

CIMA Memoria 2017 105
10 ARTÍCULOS EN REVISTAS NO SCI
CARBALLAL DURÁN, M.J., IGLESIAS ESTEPA, D. Parásitos y
enfermedades de Nassarius reticulatus y su rol como transmisores de
enfermedades a los bivalvos comerciales. Libro de Actas del XVI Congreso
Nacional de Acuicultura, págs. 336-337.
VÁZQUEZ-LUIS, M., ÁLVAREZ, E., BARRAJÓN, A., GARCÍA-MARCH, J.R.,
GRAU, A., HENDRIKS, I.E., JIMÉNEZ, S., KERSTING, D., MORENO, D.,
PÉREZ, M., RUIZ, J.M., SÁNCHEZ, J., VILLALBA, A., DEUDERO, S. Pinna
nobilis: a mass mortality event in western Mediterranean Sea. Frontiers in
Marine Science, 4:220.

106
11 LIBROS
El ctra Galicia pasado, presente y futuro.
El cultivo de la ostra rizada en Galicia – Pasado, presente y futuro
Editores: ABOLLO, E., VILLALBA, A., SIEIRO, P., MAROTO, J.
Publicado por: Centro Tecnológico del Mar-FUNDACIÓN CETMAR; Centro de
Investigacións Mariñas, Consellería do Mar, Xunta de Galicia.
ISBN: 978-84-697-3753-8
IGLESIAS, D., GÓMEZ, J.L. Problemática asociada a la introducción de Crassostrea gigas y
análisis de la situación en Galicia. Pp. 9-38.
GÓMEZ, J.L, RODRÍGUEZ, L.M., MONTES, J., IGLESIAS, D. Cultivo y explotación de la ostra rizada Crassostrea
gigas en Galicia. Pp. 41-67.
VILLALBA, A., CAO, A., RAMILO, A., ABOLLO, E. El reto de la alta mortalidad en el engorde de la ostra rizada
Crassostrea gigas: infección por la microvariante del virus herpes 1 de ostreidos. Pp. 90-120.
Oysters and clams: cultivation, habitat, threats and ecological impact.
Editor: J.L. Romalde.
Publicado por: Nova Sci. Publ. NY.
BARJA, J.L., DUBERT, J., MONTES, J., OLVEIRA, J.G., PRADO, S. & MARTÍNEZ-PATIÑO, D. Microbiological
approaches to understand the threats for the wedge-shell clam (Donax trunculus) in natural beds. Pp. 67-77.
LOUZÁN, A., NÓVOA, S., OJEA, J., MARTÍNEZ PATIÑO, D. Culture of the wedge-shell clam Donax trunculus: new
developments. Pp. 15-28.

CIMA Memoria 2017 107
2 PONENCIAS Y PANELES
Ponencias
18th International Conference of the European Association of Fish Pathologists
“Diseases of Fish and Shellfish”. Belfast (Reino Unido).
Autores/as: IGLESIAS, D., VILLALBA, A., NO E., DARRIBA, S., CAO, A., MARIÑO,
J.C., FERNÁNDEZ, J., CARBALLAL, M.J.
Título: Epidemiological patterns of marteiliosis affecting the common cockle
Cerastoderma edule in Galicia (NW Spain).
Autores/as: VILLALBA, A., CAO, A.
Título: Assessing procedures to reduce mortality of hatchery-produced Pacific oyster
spat due to OsHV1-µvar during growing out in Galicia (NW Spain).
Evolution 2017 (Portland, Oregon, Estados Unidos)
Autores/as: METZGER, M., VILLALBA, A., CARBALLAL, M., IGLESIAS, D., SHERRY, J., REINISCH, C., MUTTRAY,
A., BALDWIN, S., GOFF, S.
Título: Contagious Clam Cancer: Evolution at the intersection of cancer and infectious disease.
2nd International Symposium on Advances in Mussel Research, Sète (Francia).
Autores/as: BERNARD, I., ALLAIN, G., CHARLES, M., VILLALBA, A., BURIOLI, E., HOUSSIN, M., DEYDIER, Y.
Título: Investigation on blue mussel high mortality in France: a phenomenon with high spatio-temporal variability but
no conclusive cause.
Presentación del libro “El cultivo de la ostra rizada en Galicia”, Vigo.
Autores/as: VILLALBA, A.
Título: Producción de la ostra rizada en Galicia y su salubridad.
Jornada de presentación de la Red Gallega de Investigación y Transferencia en Acuicultura INMUNOGENOM,
Santiago de Compostela.
Autores/as: VILLALBA, A.
Título: Transmisión horizontal de cáncer en berberechos.
Autores/as: VILLALBA, A.
Título: Grupo de Patología de Moluscos del CIMA (GPM-CIMA).

108
Aquaculture Europe 2017. Dubrovnik (Croacia)
Autores/as: PAPADAKI, M., PELETEIRO, J.B., ALVAREZ-BLÁZQUEZ, B., RODRÍGUEZ, J.L., LINARES, F., VILAR,
A., PÉREZ RIAL, E., LLUCH, N., FAKRIADIS, I., MYLONAS, C.
Título: Description of the endocrine reproductive cycle of the wreckfish Polyprion americanus in captivity.
Autores/as: ÁLVAREZ-BLÁZQUEZ, B., RODRÍGUEZ, J.L., VILAR, A., MYLONAS, C., PAPANDROULAKIS, N.,
PÉREZ RIAL, E., LLUCH, N., PAZOS, G., LINARES, F.
Título: Progress in the wreckfish intensive culture. New candidate species for aquaculture.
Paneles
7th congress of european microbiologists (FEMS). Valencia
Autores/as: RODRÍGUEZ, S., BALBOA, S., OLIVEIRA, G., MONTES, J., MORENO, D., BARRAJÓN, A., BARJA, J.
Título: First report of mass mortalities in natural population of Pinna nobilis. A microbial perspective.
XVIII Conference da EAFP 2017 (Belfast, Irlanda del Norte)
Autores/as: LOPEZ, C. Y DARRIBA, S.
Titulo: Survey of the Rickettsiales-like colonies associated with a mortality event in Tapes virgineus in Galicia (NW
Spain).
18th International Conference of the European Association of Fish Pathologists “Diseases of Fish and
Shellfish” Belfast (Reino Unido).
Autores/as: DARRIBA, S., IGLESIAS, D., VILLALBA, A., CARBALLAL, M.J.
Título: Infection with a haplosporidan-like protist detected in the main production area of Cerastoderma edule in
Galicia (NW Spain).
XVI Congreso Nacional de Acuicultura (Zaragoza, España)
VILLALBA GARCÍA, A.
Miembro del Comité Científico.
Autores/as: CARBALLALL, M.J., IGLESIAS, D.
Título: Parásitos y enfermedades de Nassarius reticulatus y su rol como trasmisores de enfermedades a los bivalvos
comerciales.
Autores/as: COSTA, D.P., NÓVOA, S., OJEA, J., MARTÍNEZ, D.
Título: Desarrollo larvario en criadero de almeja bicuda Polititapes aureus (Gmelin, 1791).
Autores/as: NÓVOA, S., PÉREZ-PARALLÉ, M.L., OJEA, J., SÁNCHEZ, J.L., MARTÍNEZ, D.
Título: Inducción de la fijación de larvas de almeja japonesa Ruditapes philippinarum (Adams & Reeve, 1850)
utilizando diferentes compuestos neuroactivos.
Autores/as: PRADO, S., NÓVOA, S., MARTÍNEZ-PATIÑO, D., BARJA, J.L.
Título: Interacción microalgas-bacterias en criadero de bivalvos: Supervivencia en el fitoplancton de vibrios patógenos
de larvas.
21st International Pectinid Workshop. Portland, Maine (Estados Unidos)
Autores/as: VENTOSO, P., MARTÍNEZ-ESCAURIAZA, R., SÁNCHEZ, J.L., PÉREZ-PARALLÉ, M.L., BLANCO, J.,
TRIVIÑO, J.C., PAZOS, A.J.
Título: RNA-seq analysis of differential gene expression in response to domoic acid-producing Pseudo-nitzschia in the
digestive gland of Aequipecten opercularis (L.)

CIMA Memoria 2017 109
Aquaculture Europe 17. Dubrovnik (Croacia)
Autores/as: MARTÍNEZ-ESCAURIAZA, R., VENTOSO, P., SÁNCHEZ, J.L., PÉREZ-PARALLÉ, M.L., BLANCO, J.,
TRIVIÑO, J.C. AND PAZOS, A.J.
Título: Differentially expressed genes in the digestive gland of Aequipecten opercularis (L.) in response to domoic acid
accumulation.
Autores/as: RODRIGUEZ, J.L., ALVAREZ-BLÁZQUEZ, B., PÉREZ, E., MARTÍNEZ, J.M., MÉNDEZ, J.C., ACUÑA, I.,
LINARES, F.
Título: Evaluation of wreckfish (Polyprion americanus) growth in Galicia (Spain).
11th International Conference on Molluscan Shellfish Safety. Galway, Ireland
Autores/as: BLANCO, J., MARTÍN, H., MARIÑO, C.
Título: Evaluation of chromatographic columns for the analysis of PSP toxins after pre-column oxidation.
Autores/as: ALVAREZ, G., GODOY, M., ARAYA, M., GANUZA, I., PINO, R., RENGEL, J., URIBE, E., BLANCO, J.
Título: Effects of Harmful Algal Bloom of Alexandrium catenella in the mass mortality of the surf clam Mesodesma
donacium in Chiloé Island, Chile.
Autores/as: ÁLVAREZ, G., RENGEL, J., URIBE, E., BLANCO, J.
Título: Anatomical distribution of cadmium in Argopecten purpuratus: effects of evisceration on industrial processes to
reduce the metal concentration in scallops used for human consumption.
XX Foro de Recursos Marinos y Acuicultura de las Rías Gallegas
Autores/as: NÓVOA, S., OJEA, J., CASTILLA, S., GRANJEL, E., MARTÍNEZ, D.
Título: Estado reproductivo de una población de coquina Donax trunculus (Linnaeus, 1758) de la playa de Vilarrube
(Galicia).
Autores/as: CATOIRA, J.L., SEOANE, P., MARTÍNEZ, D., OJEA. J.
Título: Relaciones biométricas y crecimiento de coquina (Bivalvia: Donacidae) en sistemas de cultivo en el medio
natural

110
13 EDICIÓN DE PUBLICACIONES
VILLALBA GARCÍA, A.
Diseases of Aquatic Organisms.
Journal of Shellfish Research.

CIMA Memoria 2017 111
14 INFORMES
Título: Informe del año 2016 correspondiente a las “Experiencias para el
cultivo del besugo realizadas en las Jaulas Experimentales” de las que es
titular el Centro de Investigacións Mariñas (Consellería do Mar) ubicadas en
Bueu.
Autor/a: Linares Cuerpo, F.
Solicitante: Subdirectora Xeral de Acuicultura. Consellería do Mar.
Título: Informe y exposición de resultados correspondientes a las actividades del WP12. “Nutrición de la cherna
(Polyprion americanus) para la reunión anual de coordinación del proyecto.
Autor/a: Linares Cuerpo, F.
Solicitante: Coordinador WP Nutrición Proyecto Diversify
Título: Informe de resultados para el 3ndReport (período junio 2016-noviembre 2017) del Proyecto Diversify de las
actividades relacionadas con el desarrollo de la “Nutrición de la cherna ((Polyprion americanus) WP.12)”.
Autor/a: Linares Cuerpo, F.
Solicitante: Coordinador WP Nutrición Proyecto Diversify

112
15 REUNIONES
REUNIÓN: Jornada de presentación de la Red Gallega de Investigación y
transferencia en Acuicultura: INMUNOGENOM.
ASISTENTES: Gabín Sánchez, C., Villalba García, A., Cao Hermida, A.
REUNIÓN: Reunión del Comité de Organización para la puesta en marcha de
la RED IMMUNOGEOM.
ASISTENTES: Cao Hermida, C., Villalba García, A.
REUNIÓN: II Reunión del Comité de Organización de la RED
IMMUNOGEOM.
ASISTENTES: Cao Hermida, C., Villalba García, A.
REUNIÓN: Reunión del Grupo de Trabajo Técnico de Acuicultura de la PTEPA para actualizar el Plan Estratégico de
Innovación y Desarrollo Tecnológico de la Pesca y la Acuicultura.
ASISTENTES: Villalba García, A.
REUNIÓN: Reunión de coordinación del MAPAMA, Comunidades Autónomas del Mediterráneo y centros de
investigación para consensuar medidas de protección de la nacra.
ASISTENTES: Villalba García, A.
REUNIÓN: Reunión de coordinación y puesta en marcha del proyecto de investigación “Co-Operation for Restoring
Cockle Shellfiseries and its Ecosystem Services in the Atlantic Area (COCKLES)”.
ASISTENTES: Villalba García, A.
REUNIÓN: Reuniones de planificación y coordinación de actividades con los participantes de Galicia en el proyecto
“Exploring the biological and socio-economic potential of new/emerging candidate fish species for the expansión of the
European aquaculture industry”. “Diversify”.
ASISTENTES: Linares Cuerpo, F.
REUNIÓN: Reunión anual de coordinación del proyecto “Exploring the biological and socio-economic potential of
new/emerging candidate fish species for the expansión of the European aquaculture industry”. “Diversify”.
ASISTENTES: Linares Cuerpo, F.
REUNIÓN: Órgano de Gestión Seguimento y Control (Oxesco) de la Reserva Marina de interés pesquero ría de
Cedeira.
ASISTENTES: Cuiñas Olmedo, P., Febrero Mayor, F.
REUNIÓN: Órgano de Gestión Seguimento y Control (Oxesco) de la Reserva Marina de interés pesquero Os
Miñarzos.
ASISTENTES: Cuiñas Olmedo, P., Febrero Mayor, F.
CIMA Memoria 2017 113
16 OTROS
16.1. ASESORAMIENTO. OTROS
CARBALLAL DURÁN, M.J.
Participación en una prueba de inter-comparación basada en 50 preparaciones histológicas para detectar patógenos
de declaración obligatoria para la UE, organizada por el Laboratorio Nacional de Referencia.
LINARES CUERPO, F.
Convenio de colaboración entre la Xunta de Galicia a través de la Consellería do Mar y la Asociación de Acuicultores
“Illa do Santo” de Bueu (Pontevedra), para el cultivo de peces en jaulas.
VILLALBA GARCÍA, A.
Asesor en el proceso de evaluación para el nombramiento de la Dra. Kimberly L. Reece como Chancellor Professor
en el Virginia Institute of Marine Science (VIMS), College of William and Mary.
VILLALBA GARCÍA, A., IGLESIAS ESTEPA, D., CAO HERMIDA, A., CARBALLAL DURÁN, M.J.
Miembros del “International Common Cockle Genome Consortium” para la secuenciación y anotación del genoma del
berberecho Cerastoderma edule, financiado a través del proyecto SCUBA CANCER (ERC Starting Grant, 716290),
cuyo investigador principal es José Manuel Castro Tubío (Universidad de Santiago de Compostela), quien también
preside el consorcio.
Asesores del proyecto “Projet d’échantillonnage pour la détection de la néoplasie hémocytaire dans les moules de
Bretagne nord”, cuyo investigador principal es Ismaël Bernard (Eureka Modélisation, Francia), que se desarrolla en
Francia con el objetivo global de identificar las causas de la mortalidad masiva de mejillón cultivado en el Norte de
Francia.

114
16.2. REVISIÓN DE ARTÍCULOS EN PUBLICACIONES CIENTÍFICAS
BLANCO PÉREZ, J.
Marine Drugs
Nº de artículos: 1 (2 rev)
Aquatic Toxicology
Nº de artículos: 4
Chemosphere
Nº de artículos: 1 (4 rev)
Hydrobiología
Nº de artículos: 1 (2 rev)
Water
Nº de artículos: 1
Marine Environmental Research
Nº de artículos: 1
Journal of Oceanology and Limnology
Nº de artículos: 1
VILLALBA GARCÍA, A.
Diseases of Aquatic Organisms
Journal of Invertebrate Pathology

CIMA Memoria 2017 115
17 TESIS DOCTORALES
17.1. DIRECCION/TUTORIZACIÓN DE TESIS
TEMA: Estudio proteómico del sistema inmune de la ostra plana europea (Ostrea edulis) y su interacción con
parásitos Bonamia spp.
DOCTORANDO/A: Rodríguez de la Ballina, N.
TITULACIÓN: Licenciada en Biología
TUTOR/A Y DIRECTOR/A DE LA TESIS: Villalba García, A. (tutor y codirector), Cao Hermida, A. (codirectora),
Hermida Prieto, M. (codirector).
TEMA: Consideraciones biológicas, ecológicas y productivas relativas al cultivo de la ostra rizada Crassostrea gigas
(Thunberg) en las rías gallegas.
DOCTORANDO/A: Rodríguez González, L.
TITULACIÓN: Licenciado en Biología
TUTOR/A Y DIRECTOR/A DE LA TESIS: Montes Pérez, J., Iglesias Estepa, D.
TEMA: Optimización del cultivo intensivo de la almeja fina Ruditapes decussatus en criadero.
DOCTORANDO/A: Aranda Burgos, J.A.
TITULACIÓN: Licenciado en Biología
TUTOR/A Y DIRECTOR/A DE LA TESIS: Martínez Patiño, D. (tutora y directora); da Costa González, F. (codirector).
17.2 PARTICIPACIÓN EN TRIBUNALES DE TESIS
VILLALBA GARCÍA, A.
Título: Caracterización del antígeno reconocido por el anticuerpo monoclonal M22.8 y su posible implicación en la
biomineralización de bivalvos.
Doctorando/a: Calvo Iglesias, J.
Universidad: Universidad de Vigo

116
18 BECARIOSAS FP 2º GRADO
TUTOR/A: Cao Hermida, M.A.
BECARIO/A: Viturro García, R.
ESPECIALIDAD: Técnico Superior en Análisis y Control de Calidad
TUTOR/A: Cao Hermida, M.A.
BECARIO/A: Pena Thomas, G.
ESPECIALIDAD: Técnico Superior en Análisis y Control de Calidad
TUTOR/A: Carballal Durán, M.J.
BECARIO/A: Fernández González, J.
ESPECIALIDAD: Técnico Superior Anatomía, Patología y Citología
TUTOR/A: Linares Cuerpo, F.
BECARIO/A: Araujo Sendín, A.
ESPECIALIDAD: Técnico Superior en Producción Acuícola
TUTOR/A: Linares Cuerpo, F.
BECARIO/A: Fuentes Villar, N.
ESPECIALIDAD: Técnico Superior en Producción Acuícola
TUTOR/A: López Gómez, M.C.
BECARIO/A: Grande Ocampo, O.
ESPECIALIDAD: Técnico Superior Anatomía Patológica y Técnico Superior en Acuicultura
TUTOR/A: Nóvoa Vázquez, S.
BECARIO/A: Castilla del Río, S.
ESPECIALIDAD: Técnica Superior en Producción Acuícola
TUTOR/A: Ojea Martínez, J.
BECARIO/A: Matesanz Salgado, D. (hasta 16/05/18)
ESPECIALIDAD: Técnico Superior en Producción Acuícola
TUTOR/A: Ojea Martínez, J.
BECARIO/A: Granjel Piñeiro, Eduardo (desde 01/07/17)
ESPECIALIDAD: Técnico Superior en Producción Acuícola

CIMA Memoria 2017 117
19 CURSOS
19.1. CURSOS IMPARTIDOS
Máster Interuniversitario de Acuicultura (UDC/USC/UVIGO)
BLANCO PÉREZ, J. (compartido con Ana Gago (UVIGO))
“Mareas Tóxicas”.
MARTÍNEZ PATIÑO, D., NÓVOA VÁZQUEZ, S., OJEA MARTÍNEZ, J.
Impartición de prácticas sobre cultivo de moluscos bivalvos.
MARTÍNEZ PATIÑO, D., NÓVOA VÁZQUEZ, S.
Tutorización de Trabajo Fin de Máster.
VILLALBA GARCÍA, A.
“Enfermedades en invertebrados”:
Tema 4: Enfermedades neoplásicas.
Tema 5: Estrategias de lucha frente a enfermedades en invertebrados.
Sesión de prácticas.
Curso EGAP “Espectrometría de masas, el ajuste perfecto a las técnicas híbridas. Aplicación a muestras
marinas y ambientales”.
BLANCO PÉREZ, J.
Aplicación de la técnica LC-MS-QQQ al análisis de las biotoxinas marinas.
11th International Postgraduate Course" "Histology and Histopathology of Aquatic Animals" de la Universidad
del País Vasco (Estación Marina de Plentzia), correspondiente al programa del "European Master of Science
In Marine Environment and Resources.
VILLALBA GARCÍA, A.
“General Histopathology of molluscs”.
“Neoplastic lesions in molluscs”.
Practical session 2: Microscopical observations on general molluscan histopathology.
Practical session 3: Microscopical observations on general molluscan histopathology (cont.) and identification of
neoplastic lesions.
“Líneas actuales en investigación en Biología Animal”, correspondiente al Máster Universitario en
Investigación en Ciencias, Especialidad en Biología Animal, de la Universidad de Alcalá.
VILLALBA GARCÍA, A.
Epidemias de cáncer en moluscos bivalvos.

118
Ciclo de Seminarios del Instituto de Investigaciones Agroambientales y de Economía del Agua (INAGEA, Islas
Baleares).
VILLALBA GARCÍA, A.
Aplicaciones biotecnológicas en la lucha contra enfermedades de moluscos.
International Postgraduate Course “Research in Marine Environment and Resources” dentro del “Erasmus
Mundus European MSc in Marine Environment and Resources awarded by the MER Consortium
(Universidades del País Vasco, Southampton, Burdeos y Lieja).
VILLALBA GARCÍA, A.
Marine Molluscs as Emerging Animal Model for Human Cancer?” y participación en la mesa redonda “Marine Animals
as Alternative Research Models in Biomedicine.
19.2. ASISTENCIA A CURSOS
Escola Galega de Administración Pública. EGAP. Santiago de Compostela
BLANCO PÉREZ, J.
Estadística avanzada con software libre R.
Lengua inglesa. Nivel avanzado 1 (C1).
IGLESIAS ESTEPA, D.
Aplicaciones informáticas de bases de datos relacionais. (LIBREOFFICE BASE).
Aplicaciones informáticas de hojas de cálculo (LIBREOFFICE CALC).
Aplicaciones informáticas de presentaciones gráficas de información (LIBREOFFICE IMPRESS).
Aplicaciones informáticas de tratamiento de textos (LIBREOFFICE WRITER).
Manejo de técnicas y dispositivos de orientación básica.
Estadística avanzada con software libre R.
LINARES CUERPO, F.
Lengua inglesa. Nivel avanzado 1 (C1).

CIMA Memoria 2017 119
20 ESTANCIAS
20.1. ESTUDIANTES EN PRÁCTICAS
Alumno/a: Varela Chorén, T.
Procedencia: Facultad de Biología, Ciencias Ambientales y Química.
Universidad de Alcalá
Duración: 824 horas
Tutor/a: Cao Hermida, M.A.; Viéitez Martín, J.M.
Alumno/a: Castedo Moreira, M.I.
Procedencia: IES Lucus Augusti. Lugo
Duración: 410 horas
Tutor/a: Martínez Patiño, D.
Alumno/a: Pousada Linares, I.
Procedencia: Facultad de Ciencias. Universidad de A Coruña
Duración: 265 horas
Tutor/a: Ojea Martínez, J.
Alumno/a: Pilré da Costa, D.
Procedencia: Instituto Politécnico de Leiria. Portugal
Duración: 1.620 horas
Tutor/a: Martínez Patiño, D.
Alumno/a: Escrivá Perales, J.
Procedencia: Universidad Politécnica de Valencia
Duración: 30 días
Tutor/a: Martínez Patiño, D.
20.2. TFM: MÁSTER INTERUNIVERSITARIO GALLEGO DE ACUICULTURA
Alumno/a: Rodrigues Fonseca Baptista Verde, M.F.
Tutores/as: Martínez Patiño, D.
Alumno/a: Fernández Pena, P.
Tutores/as: Nóvoa Vázquez, S.

120
21 GASTOS
I.- Distribución del presupuesto por capítulos
Capítulo
Cantidad (Euros)
Capítulo I: Gastos de personal 2.271.553,49 €
Capítulo II: Gastos corrientes en bienes y servicios 182.000,00 €
Capítulo IV: Transferencias corrientes 82.200,00 €
Capítulo VI: Inversiones reales 1.218.265,87 €
Capítulo VII: Transferencias de capital
Total 3.754.019,36 €

CIMA Memoria 2017 121
22 FUENTES DE FINANCIACIÓN
ORIGEN DE LOS FONDOS
CANTIDAD (€)
Propia: Dirección Xeral de Desenvolvemento Pesqueiro. Consellería do
Mar. Xunta de Galicia
a) Acciones de Investigación que desarrolla el CIMA
b) Convenios con entidades externas
3.735.753,69 €
I+D de la Xunta de Galicia
Administración General del Estado
Unión Europea (INTERREG, VI Programa Marco,...) 37.804,62 €
Total 3.773.558,31 €

122
m





![Practice For May: Cell Ultrastructure [114 marks]blogs.4j.lane.edu/.../2018/02/Cell-Ultrastructure-Test-1.pdfPractice For May: Cell Ultrastructure [114 marks]1. Which structure found](https://cdn.vdocuments.us/doc/165x107/5eda4db5b3745412b5711d9c/practice-for-may-cell-ultrastructure-114-marksblogs4jlaneedu201802cell-ultrastructure-test-1pdf.jpg)

