de ciencias biologicas y' de la salud - …148.206.53.84/tesiuami/uam2657.pdf · division de...
TRANSCRIPT

' U . A. M. IZTAPALAPA BIBLIOTECA I
UNIVERSIDAD AUTONOMA METROPOLITANA
UNIDAD IZTAP,.\l..?PA
DIVISION DE CIENCIAS BIOLOGICAS Y' DE LA SALUD
INFORW FINAL DEL SERVICIO SOCIAL
DETERMINACION DE LOS NIVELES DE RADICALES; LIBRES DURANTE LA MADURACION DE MANGO VARIEDAD 'KEIIT".
PRESENTADO POR:
José Antonio González Rodriguez.,
LICENCIATURA EN BIOLOGIA EXPERIMENTAL
ASESOR: Dra. Laura J. Pérez Flores. Profesor Titular "C" , T. Completo.
Lab. de Bioquímica y Fisiología Vegetal. Depto. de Ciencias de la Salud.
DICIEMBRE, 1995

INTRODUCCIQN
La senescencia~ representa los procesos degenerativos controlados de manera endógena (al interior del organismo) que conducen a la muerte. Este concepto de la senescencia permite abordar este proceso no colno ¡a mera descripción de un estado fisiológico sino a partir del estudio de sus mecanismos de regulación y de los factores que participan en ellos. Actuaimente la idea de !a regulacibn de la senescencia por genes específicos es aceptada por la mayoría de los investigadores en el campo. For otro lado, el envejecimiento comprende una amplia gama de procesos degenerativos pasivos o no regulados, inducidos primordialmente por factores exógenos (ambientales) (Ponce de León et al., 1992).
En este sentida, Harman (1956) planteó una teoría que proporre que el envejecimiento es una consecuencia del clafio irreparable camado por ia acurnulaci6n de radicales libres en los organismos.
Un radical libre puede ser definido como un átorno o grupo de iintorraos con un efectrón desapareado, el cual dota al &tomo o grupo de itornos de ia capacidad de remover o donar un electr6n a una molécula vecina. Debido a su alta reactividad los radicales libres pueden atacar a sus rnol&;ulas vecinas y dar por resultado un daño al organismo. Entre los radicales más nocivos están los hidroxilos (OH.-) p los superóxidos (O,.-), adem6s de algunos de origen orgimico! corno los pravenieates de ¡as semiquinonas y de ios Bcidos grasos poliinsaturados.
Los organismos vivos poseen mecanismos para contrarrestar los efectos de estos radicales libres ya sea previniendo su forrrlación, acelerando su descomposicibn o secuestrándolos para disminuir sus niveles. En dichos mecanismos participan agentes antioxidantes tales corno e 1 &cid0 ascbrbico, los tocoferoles (como la vitamina E), las citocininas y algunas enzinas como la superbxido dkrnutasa (SOD) y la peroxidasa (Leshem, f98-1).
Las condiciones del medio con el que interactúan los orgar?isrnos pueden influir tanto sobre la produccibn de radicales libres corno sobre ios mecanismos de defensa.
Actualmente se sugiere que el componente yeni??:ics del programa de senescencia consta de las capacidades antioxidantes y reparadores del organismo las que por interacciCln con factores del ambierste determinan fa duracibrt de la vida (Psnce de Le6n ef al., 1992), Sohal (1989) señala que la hipótesis de i4armat-i 10
explica c6mo la acurjluiaciijn de radicales libres. que en la rnayaría de las c6llelas son subproductos de la utilízacibn del oxígeno, pueden, a! pro'vocar un darlo al azar: dar lugar a un patr6n reproducible de cambios que sucede3 de m8ne:1% arláloga durante 21 envejecimiento en todos los inclivich!os de ~ n a poblaciSy: especifica.
En un estudio realizado en Musca domestica (Sohai, 1989) se er*tcn:-!!r6 que la tasa be consumo de oxígeno de estos organismos mantiene una relación directa csn la terrbperatura del ambiente, m!icntras qtie la daracibtb de su v;dii
guarda m a relación inversa con respecto a la temperatura; si bien este resultado

no es inesperado sí sorprende el hecho de qtie el C O R S , U ~ ~ total de energía durante la vida ("potencial rnetabdico") se haya mantenido relativamente canstante.
En el nivel molecular, estos resultados significasian que el "reloj" para el programa genetico de la senescencia registra más que, el paso del tiempo. el paso de eventos metabólicos (potencial rnetabdico).
Hay evidencias que tienden a mostrar que la longevidad en animales es función, mrirs que del paso del tiempo, de la tasa de utiiisac:ión de energía o tasa metabdica ya que existe una cantidad finita de gasto total de energia durante la vida de un organismo (Ponce de León et al., 1992).
Sohal (1987) sugiere que el mecanismo por el cuial la tasa metabrjlica ejerce influencia en la tasa de envejecimiento puede involucrar a los radicales libres e hidroperóxidos como mediadores . El mismo autor proporre en 1989 una hip6tesis denominada "hipótesis del estrés oxidativo del envejecimiento" que contiene parcialmente a la teoría de Harrnan pues mantiene la relacjSn del incremento de radicales libres asociada al envejecimiento pero incluye el concepto del aumento de un estrés oxidativo con la edad, el cual s'e manifiesta porque el estado redox se vuelve progresivamente rn&s oxidante o nienas reductor durante
x' e! procese, de envejecirniento. El estrés oxidativo se determina, entre rjtros factores, por la disminución en las relaciones NADPH/NADP", glutlatiin redusidolglutatión oxidado y por un aumento en los niveles de W,O,. Los cambios
disminucion de los mecanismos antioxidantes de defensa como por un amento en la generación de radicales libres y de H,O,. La hipótesis de Sohal atribuye al estres oxidativo un función fisiológica en la inducción y mantenimiento de la diferenciaciór~ ceiuiar. En este sentido, se propone con base en Ias evidencias experimentales que los radicales libres de oxígeno establecen un estado redox 6pt;mo para inducir y mantener el estado celular diferenciada
Si la senescencla se contempla como una continuación del proceso de dewrollo dentro del marco del proceso ontogen&ico, el estrés oxidativo proveeria el vincuio entre estas dos fases de la vida.
El incremento en el estrks oxidativo es predecihle drlrante fases especificas de !a ontogenia y puede ser provocado por estírncdos experimentales por lo que se puede suponer q ~ e ocurre en respuesta a sefiales genómicas especificas.
SI bien existen resultados que apoyan la hip6tesis del estrés oxidativo, q ~ e d a a ú n por dilucidar 6) mecanismo por el que se transduce la seAal de dicho eswis err la 'rnducci6n de genes especifieos en las cktulas, entre los cuales pc3:ian encontrarse Dos genes responsables de la senescencia ( P Q ~ c ~ de he6n et ai., '1 992),
En !as membranas biolbgicas los radicales libres reacciona1 cot7 los ilpidos de las membranas produciendo la lipoperoxidación, Aparecen reacciones de ini:iacibn, propagacirjn y terminacibn. Se ha reportado que las plantas sornetidas a diferentes tipos de estr6s biótico y abidtico, aumentan los niveles de Ir;soperoxrdaciOn dands lugar finalmente a la perdida cfe la integridad celular, sL;~r:ue al pxecer el proceso de lipopesoxidacidn es un evento ~metabólico rrarrnal
\..
i
?.
Is en el estado redox del estrhs oxidativo pueden expiiearse tanto por una

que se encuentra tanto en plantas estresadas como en no estresadas (Purvis & Shewfelt, 1993).
MADURACION DE FRIJTOS.
La maduración es la fase final del crecimiento y desarrollo del fruto, en la que se producen una serie de cambios generalmente coordinados , que conducen a la madurez de consumo y posteriormente a la senescerlcia .
La maduración de muchos frutos se caracteriza por un incremento de la actividad respiratoria (pico climatérico) y un aumento en la síntesis de etileno, a estos frutos se les denomina climatéricos. Los frutos de otras especies maduran sin mostrar ninguna de estas características (frutos no climatéricos), y frutos de otras especies tienen un comportamiento intermedio. La maduración del fruto suele coincidir con cambios en su apariencia, textura y sabor. Los cambios en la apariencia involucran cambios en el color de la piel y la pulpa debidos al remplazo de clorofilas por antocianinas y carotenoides. Las alteraciones en la firmeza de los frutos conllevan el adelgazamiento de la pared celular, la disolución de la lámina media, la pérdida de turgencia y la degradación de los productos de reserva, originando constituyentes semilíquidos. Duranse la maduración del fruto existe una producción de aromas provenientes de una mezcla compleja de compuestos volátiles que interaccionan con la producción de otros constituyentes, especialmente carbohidratos, ácidos orgánicos y compuestos fenólicos. El aroma es característico de cada especie y variedad (Azcón-Bieto & Talón, M, 1993).
El mango (Mangifera indica L) es un fruto climatérico. Durante su maduración el mango sufre cambios en la expresión genética que son disparados por factores endógenos como el etileno y por factores ambientales como la temperatura (Pérez-Flores et a/. , 1993).
El incremento en la tasa respiratoria se debe a la actividad rnitacondrial. Las mitocondrias vegetales presentan dos vías respiratorias: la vía de los citocromos y la via alternativa o vía resistente a cianuro (Pérez-Flores et al., 1993). Algunos reportes sugieren que esta última vía se encuentra presente e incluso se incrementa durante la maduración de algunos frutos (Romani, 1991). En estudios realizados en mango se observó un aumento de la contribución de la vía alternativa durante la maduración de éste (Kumar et al., 1998).
La vía alternativa se desvía de la vía citocrómica a nivel de la poza de ubiquinona y pasa los electrones de la ubiquinona seducida directamente a la oxidasa alternativa que a su vez los transfiere ai oxígeno molecular, produciendo agua como producto reducido (McIntosh, 1994). No se genera ningún gradiente protón-motriz durante el flujo de electrones entre la ubiquinona reducida y la oxidasa alternativa. Por lo tanto, toda la energia libre producida durante el flujo de electrones desde la ubiquinona hasta la vía alternativa se pierde C O ~ Q calor y es inutilizable para la sintesis de ATP (Moore et al., 1978).
Se ha reportado que la proteína oxidasa alternativa , puede encontrarse en dos formas: una dimérica unida a través de puentes disulfuro y otra monomérica que corresponde aproximadamente a un peso.molecular de 36kD. La actividad de la vía alternativa se incrementa al reducir el puente diSlJlfUrQ y pasar a la forma
4

monomerica. Con base en estos resultados se sugiere que la transicibn entre estas dos formas dependiente del estado redox proporcionaría un mecanismo para reguiar fa actividad de esta proteina (Umbach & Siedow, 1993). Un segundo mecanismo regulatorio reportado recientemente para la vía alternativa involucra la estimulaci6n de su actividad por adición de ácidos orgánicos (por ejemplo, piruvato). Se propone que el piruvato actlSa como un efector alostérica de la sxidasa alternativa (Millar et al., 1993).
No se tiene una idea clara en cuanto a la función' metabólica de la vía alternativa excepto su efecto termogénico en las aráceas. E% estas plantas la vía alternativa produce calor en las flores e inflorescencias, este calor generado volatiiiza compuestos odoríferos que atraen a los insectos; polinizadores (P&rez.- Flores et al., 1993).
Existen reportes de distintas condiciones que inducen la expresión de la vía aiternativa entre las cuales se incluyen: las bajas temperaturas, el envejecimiento inducida por heridas, el ataque por patbgenos, el aurnel?io de la concentración de sustratos respiratorios (por ejemplo, carbohidratos solubfesj, la adicibn de etileno, fa maduración y en algunos casos el itnuemento en los niveles de acido salicílico (Siedow & Umbach, 1995).
Algunos investigadores han propuesto que fa via alternativa se utiliza para regular un sobreflujo de electrones cuando la via de los citocromos se satura o se vz limitada por un aumento er; sustratos respiratorios o por la presencia de inhibidores de la vía citocrórnica (McIntosh, 1994). En este sentido, en un estudio realizado en radículas de maíz (Leprince et al., 1992) se cncontr6 que el aumento en la actividad de la vía alternativa correlaciona con ei aumento en el contenido de monosacáridos (sustratos respiratorios). Estudios recientes, donde se obtuvieron piantas transgénicas de Nicotiana tabacum, corroboran la hipotksis antes mencionada. En dichos experimentas se Iogr6 que las plantas transgknicas ?resentasan altos niveles de la oxidasa alterna debido a la intraduccidn de LIE
ransgen con sentido de Aoxl , el gen nuclear que cedlfica para la oxidasa alterna. Esta se utilizó para investigar los mecanismos regdadores de la actividdd de Is cxidasa alterna. Por otra parte, se obtuvieron plantas transg6nicas deficientes en ia proteina oxidasa alternativa por insercción antisentido del get? AOXI. Los -esuitados muestran que bajo condiciones en las cuales el rnetaboíismo :-=;spiratcrio del carbono se ve restringido por !a capacidad del transpork d'2 & X % ~ Q ~ E S mitocondrial, ocurre una activación de la oxidasa akma~iva por pcdw -sductcr mnitoeondrial y por piruvato lo cual actGa previniendlo que el rnelabolismo
-: ' , 1 $@S'.. f"zr otro lado, ¡as conceutracicnes d e 14DP y F'f :m-. ; x f.z::[:;:?s tmis
!-?portar?tes en la regulacibn de la respiracibn n-iitororrdsi:,:1. Nis~eies rc:dt,lr;idos de ADP c un gran incremento en la relacirin APP/AD" res~k3 on el ferlcis~~er~o bief, -;r?nocic'o de conlrol respiratorio por adenilato. C'us~da e! .4DP o el PI se eacuentran en concentraciones tirnit:xias. el gradient@ srot6n-rnchiz se incrw-nenta J' ia veioc!dad del acop!amiento del transporte c.i-r? rz?~ec:rcms a la vía cr txr j rn ica 4 -2. res-. $7-
c '~spcwf~' ~ . J C !a via aiternativa centrcwve a la sobrevrvencia de /as4 ~ ~ a l ~ t a s
*-. -CA - a $ carbono se redirija hacia, por ejemplo, vias iermntativas ('~./anledmgh~, et
.." (Sheodorou & ?laxti>;,, '1 993). En x t e sentidc, otras invasligalcjorEs

crecidas, durante largos periodos, en suelos deficientes en fosfato inórganico. La operación de la vía alternativa cuando la actividad de la vía citocrómica se suprime debido a un bajo nivel de Pi puede permitir el funcionamiento del ciclo de Krebs al igual que limitar la producción de ATP y por lo tanto ayudar en la sobrevivencia ternpara! de la planta en esas condiciones desfavorables (Rychter & Mikuiska, 1990; Rychter et a¡. , 1992).
Uno de los sitios de producción de radicales libres en las células vegetales esta a nivel de la ubiquinona presente en las mitocondrias, la cual en su estado sernirreducido es tin radicai libre que puede reaccionar directamente con el oxigeno molecular generando el radical superóxido. Recientemente se ha propuesto que una funci6n importante de la vía alternativa podria ser la de captar electrones de la ubiquinona reduciendo asÍ los niveles de la ubiquinona reducida y por tanto la posibilidad de generación de radicales libres en las plantas, reduciendo en gran medida el daño por lipoperoxidación de membranas y otros dar'los mediados por radicales libres (Purvis & Shewfelt, 1993).
La medicihn de los niveles de radicaies libres se puede efectuar de manera directa, es decir, determinando la presencia de radicales libres directamente de la muestra por medio de un método espectroscópicc:, denominado resonancia rmma;-,:nética del electr6n (EQR). Una segunda forma de determinar los nivetes de radicales libres en muestras biológicas es utilizando técnicas bioquímicas que nos permiten ~r3nocer el indice de lipoperoxidación de membranas, esta es manera indirecta de determinar los radicales libres que interactuaron con los lipidos de membrana.
La resonancia paramagnktica del electrbn, o resonancia de1 sph de! electrón (ESR), es una tecnica que permite el estudio de materiales paramagnéticos, los cuafes se caracterizan por su capacidad de magnetizarse en presencia de un campo magnetico en proporción a la magnitud de ktstc, dicha propiedad se asocia can las sustancias que poseen en sus rnol&culas electrones desapareados (Cammack, 1993). Ejemplos de tales cornpuestos son los radicales libres. La tecnica de EPR se basa e;) la absorcicjn de radiaci6n c?e microondas por un electrón no apareado expuesto a un campo magnetico intenso, generando así una respuesta característica propia, pur otro lado, se reporta que la sefial de radicales libres se observa a 3300 Gauss(Skoog, 2987).
El rnktodo para determinar la lipaperoxidacibn se h a m en /a reacci6n reportada por Buege 8 Aust en 1978. En el cual el rnalo;ldlaldehido (MDA), forrnado durante el ronspimiertto de los ácidos grasos poliinsafuradrss, sirve C W ~ L
un indice conveniente para determinar la extensi6n de fa reaccijn tle peroxidacizjn. El malonclialdehido ha sido ijenlificad;, COIYIO el producto de /a peroxidacibn de lipidos que reacciona, corno ya se mer!cior-r5, con el ác. tiobarbitDrico dando una especie coiorida qae absorbe a 535nm.
El siguiente proyecto de servicio social se encarnin6 a almrdar el estudio d e los niveles de radicales libres durante la maduración del mango (~Mangifera indk-2 var. kent), para 10 c ~ ~ a l se determinó la lipoperoxidacibn de n-ierrrbranas cop1 el fin de tener un indicador de senescencia q w adern& se cxrreiacione c m otros estudios C G ~ Q [o son: ios cambies en la estructl:ra celular y is actividacl rje ' 5 alternaliva durante la rrraduracibn del fruto.

OBJETIVO GENERAL: Contribuir al conocimiento del metabolismo celular durante la maduración del mango 'Kent' mediante el 4análisis de la acurrrulacibn de radicales libres como indicadores de senescencia y de los cambios celulares y ultraestructurales.
OBJETIVOS PARTICULARES:
1. Determinas tos niveles de radicales libres (lipoperoxidacibn y/o EPR) durante la maduracidn del mango 'Kent'.
2. Determinar Iss niveles de radicales libres ( ~ ~ ~ ~ p e r ~ ~ x i d a ~ i ~ n ylo EPR) durante ia rnaduraci6n del mango variedad 'Haden'.
3. Correlacionar Cos niveles de radicales fibres con la actividad de la via alternativa durante la rnaduracibn del mango 'Kent' (la rnedici6n de la actividad de la via alternativa forma parte de otro proyecto).
4. Elabosacibn del informe final.

DISEÑO EXPERIMENTAL En este proyecto de servicio social se trabajó con mangos de la variedad
'Kent' calidad exportación cosecha 1994 provenientes de Tecuala, Nayarit (empresa ARIVANIA). Los frutos se cosecharon en estado de madurez sazón (madurez fisiológica, la cual presentó las siguientes características: color verde olivo, llenado de los hombros, 7OBrix y 1.3% de ácido cítrico). Los mangos se transportaron, el mismo día, en trailer refrigerado a 13OC desde la empacadora Arivania en Tecuala, Nayarit hasta las instalaciones de l a UAM-lztapalapa. una vez que llegaron los mangos a la UAM, se seleccionaron 33 mangos al azar, se iimpiaron, se pesaron y se numeraron. Los frutos se mantuvieron durante el experimento en un cuarto a temperatura ambiente (25' -+ 2) y con una humedad relativa de 80-90%, midiendose esta última con un higrómetro.
Dos días después de la cosecha se inició el experimento, finalizando hasta el día doce . Diariamente se obtuvieron muestras de tres mangos para su posterior análisis.
También se analizaron mangos de la variedad 'Haden' cosecha 1995 provenientes de Sinaloa, los cuales se sometieron a las' mismas condiciones que los mangos anteriores. El mismo día que se cosecharon se inició con el tratamiento
METODOLOGIA.
De cada mango se obtienen aproximadamente I O gramos de mesocarpio (puipa). El tejido se coloca en empaques de papel alurninio grueso previamente etiquetados. Los empaques se congelan inmediatamente en nitrógeno líquido para prevenir la formación de radicales libres por exposición al oxígeno del aire y para detener los procesos metabólicos del fruto. De cada empaque se toma el tejido necesario para los ensayos y el resto se guarda en un ultracongelador. Las muestras se muelen perfectamente en un mortero con nitrógeno líquido obteniendose una harina de la cual se pesan las cantidades requeridas tanto para la determinacion de EPR como para el ensayo de lipoperoxidación. Se recomienda obtener al mismo tiempo las muestras para EPR y para lipoperoxidación, para evitar la exposicíbn frecuente del tejido al aire.
DETERMINACION DE LA LlPOPERQXlDACIOPJ.
REACTIVOS:
stock TCA-TBA-HCI: Acido tricloroacético 15% plv Acido tiobarbitúrico 0.5 % p/v Acido clorhídrico 0.25 N

Esta solución puede ser ligeramente calentada para ayudar en la disolución de4 hcIdo tiobarbitúrico. NOTA: El stock TCA-TBA-HCI se prepara en el momento de la determinación.
-__. PROCEDIMIENTO; En un vaso de precipitados pequeiío se colocan aproximadamente 0.29 de
la harina de mango y se disuelve con 1.2 ml de agua desionizada, dicha solución tendr6 de 0.1 a 2.0 mg de proteína de membrana aproximadamente. Se toma una aticuota de I m l de la solución para la determinacibn de la lipopel-oxidacicjn (ensayo del $cid0 tiobarbitcrico) y otra alícuota de 25p.l para la deterrninacicjn de proteha (Lowry).
Para GI ensayo del ácido tiobarbitúrico se combina Irni de muestra con 2nd del stock de 'Y'CA-TBA-HCI en un tubo de ensaye, se agita perfectamente en un vortex. Dicha solución se calienta en bario María por 25 minutos. Despuks de enfriarse, la soluci6r: se distribuye en tubos eppendori y se cerrtrifuga a 3000 g (aprox. 3500 rpm) durante 10 minutos en urra microcentrifLlga Shelton Scientific modelo VSMC-13 con el fin de eliminar el preciprlado formado durante la reaccih, el sobrenadante se recupera y la soluci6n colorida se lee a 535nrn en un espectrofotómetro de doble haz Perkin-Elmer modelo Lamda 2 contra un blanco que contenga todos los reactivos excepto la muestra problema. La concentración de rnaiordialdehído (MDA) de la muestra p~ede ser calculada usando la siguiente fdrmula' CMDA=A,&; donde, CMDA es la eorrcentracibn de nralondialdehído en nrnoles por m l , A,,, es ia absorbancia a 535nm y a es e l coeficiente de extinción molar el c ~ a l corresponde a 1.56~185 Molar-' Centirnetro-1 . La concentracibn final dei malondialdehtds se expresa en nmoies de MOA por ing de mrsltei1-m o er3 nmoies de h4DA por gramo de peso fresco de tejido.

ACTIVIDADES RlEALIZADAS.
Las actividades realizadas durante el servicio social se presentan a continuación en forma mensual.
MES UNO. Acondicionamiento del lugar donde se colocarán los mangos para el experimento. Espesar la llegada de los mangos desde Tecuaia, Nayarit hasta las instalaciones de la UAM. Una vez llegados los mangos se limpian, numeran y pesan. Preparación de reactivos. Obtención diaria de lasl muestras.
MES DOS. Preparación de reactivos para la determinacibn de fa lipsperoxidacicita y para la deterrninacibn de proteína. lrnplementacion de la metodología para la determinación de lipoperoxidación. Liofilizado.
MES TRES. Determinación de los nivetes de lipoperoxid$acion. Determinación de proteína. Liofilizado. Investigación bibliográfica.
ME§ CUATRO. Determinacibn de los niveles de lipoperoxidación Determinación de proteína. Liofilizado. Investigación bibliográfica.
MES CINCO. Determinaci6n de los niveles de lipoperoxidación Determinacibn de proteína. Investigación bibliográfica.
MES SEIS. Reaiización de los cálculos con los datos obt.enidos. Investigación bibliogriifica.
ME§ SIETE. Andisis de Dos resultados. Elaboracibn del informe final.

OBJETIVOS Y METAS ALCANZADAS.
Los siguientes objetivos fueron alcanzados:
-Se contribuy6 al conocimiento del metabolismo celular durante la maduracídn de2 mango variedad 'Kent' mediante el análisis de la acumulacibn de radicales libres como indicadores de senescencia.
--Se determinaron los niveles de radicales libres (Lipoperoxidaci6n) derrante la anaduraciór~ del mango 'Kent'.
-§e determinaron los niveles de radicales libres (lipoparoxidacitm) durante la maduraci6n del mango 'Haden'.
-Se correlacionaron las niveles de radicales libres con la actividad de la .via alternativa durante la maduraci6n de9 mango 'Kent'.
-Se eiaborb el informe de servicio social.

RESULTADOS Y CONCLUSIONES.
La figura 1 muestra la curva de respiración del rnango 'Kent' con el fin de poder establecer las fases preclimatéricas, climatéricas y postclimatéricas durante la maduración del mango 'Kent'. Se grafica la respiración del mango en mg de COJkg-hr contra días de postcosecha a temperatura ambiente. En la gráfica observamos un aumento en la respiración a partir del día cero de postcosecha hasta alcanzar su máximo en el día seis (pico climatérico) disminuyendo de manera gradual para después mantenerse de modo constante hasta el Artavo. día de almacenamiento. Se presentó un patrón respiratorio climatérico típico que coincide con reportes anteriores (Laksminarayana, 1975, 1977; Burg & Burg, 1962) para la variedad 'Kent'.
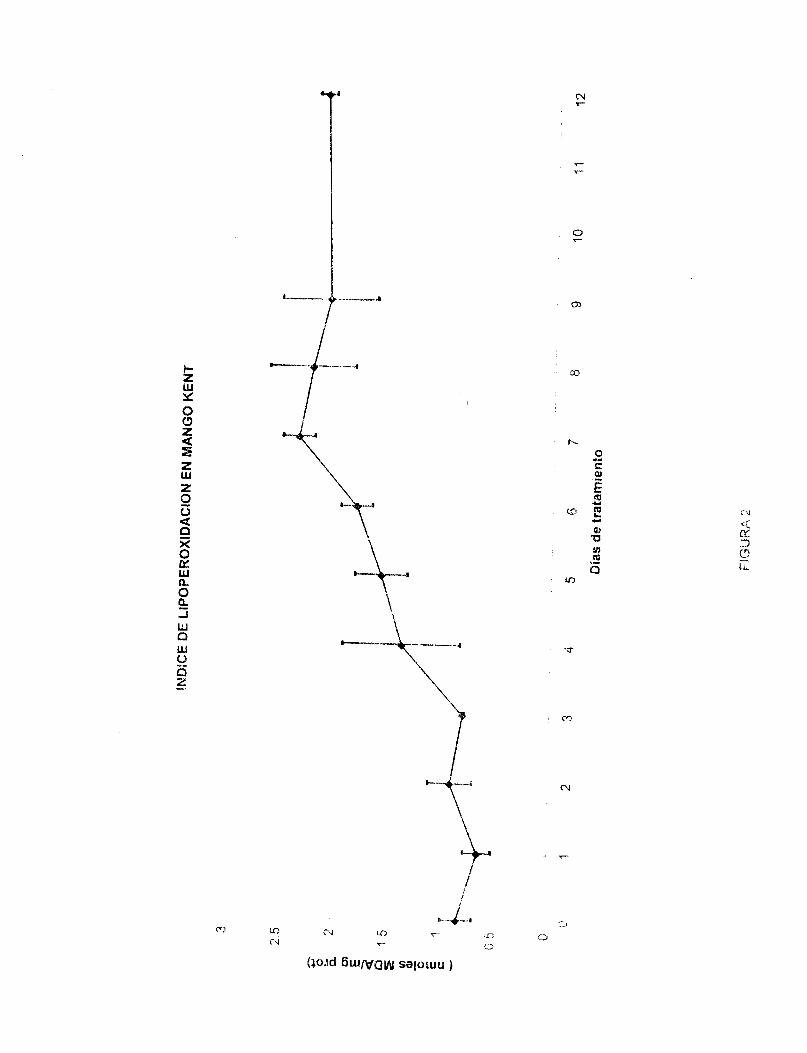
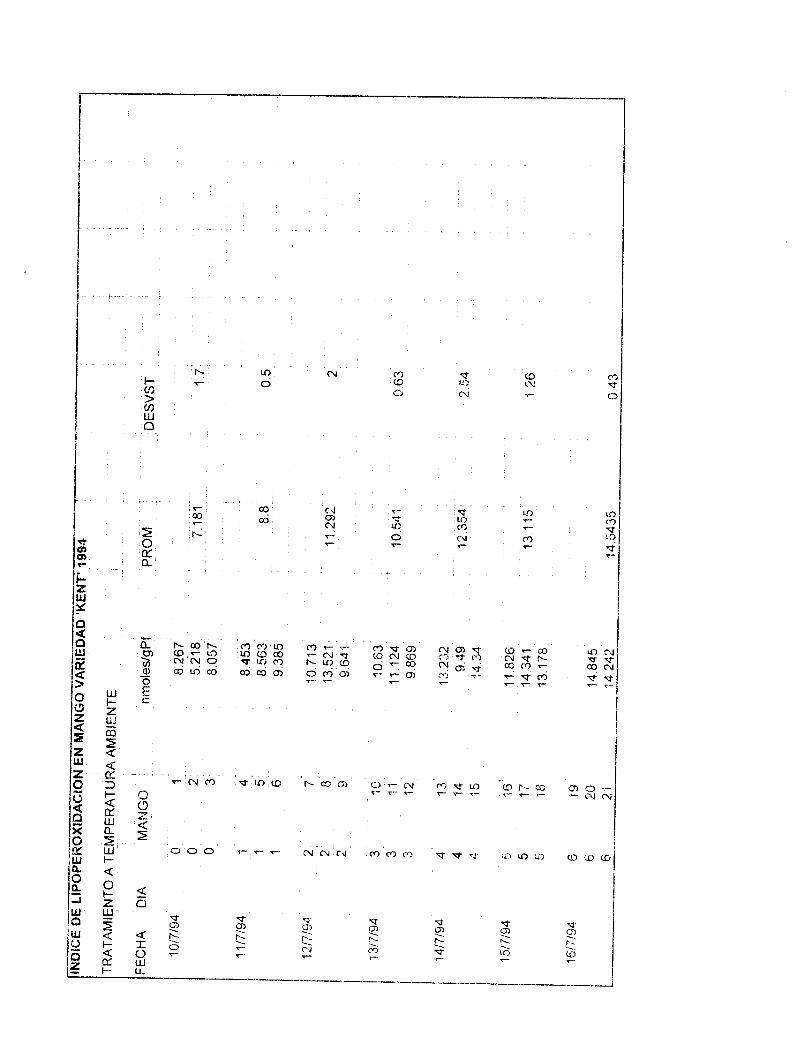
Los datos de la figura 2 muestran los niveles de lipoperoxidación del mango 'Kent' durante su maduración a temperatura ambiente,. Se grafican nmsles de matonaldehído (MDA) vs días de tratamiento. Como SE? observa en la figura el índice de lipoperoxidación aumenta de manera rápida a partir del cuarto día (dia 6 de postcosecha) alcanzando su máximo el día siete de tratamiento (2.378 nmoles MDNmg proteina) para después mantenerse constante hasta el doceavo día. Es interesante notar que en los días donde ocurre la fase climatérica, el índice de lipoperoxidación se incrementa rapidamente para llegar a su máximo durante la fase postclimatérica (senescencia). El mismo comportamiento en cuanto al incremento de la lipoperoxidación se observa en mangos de la variedad 'Haden'.
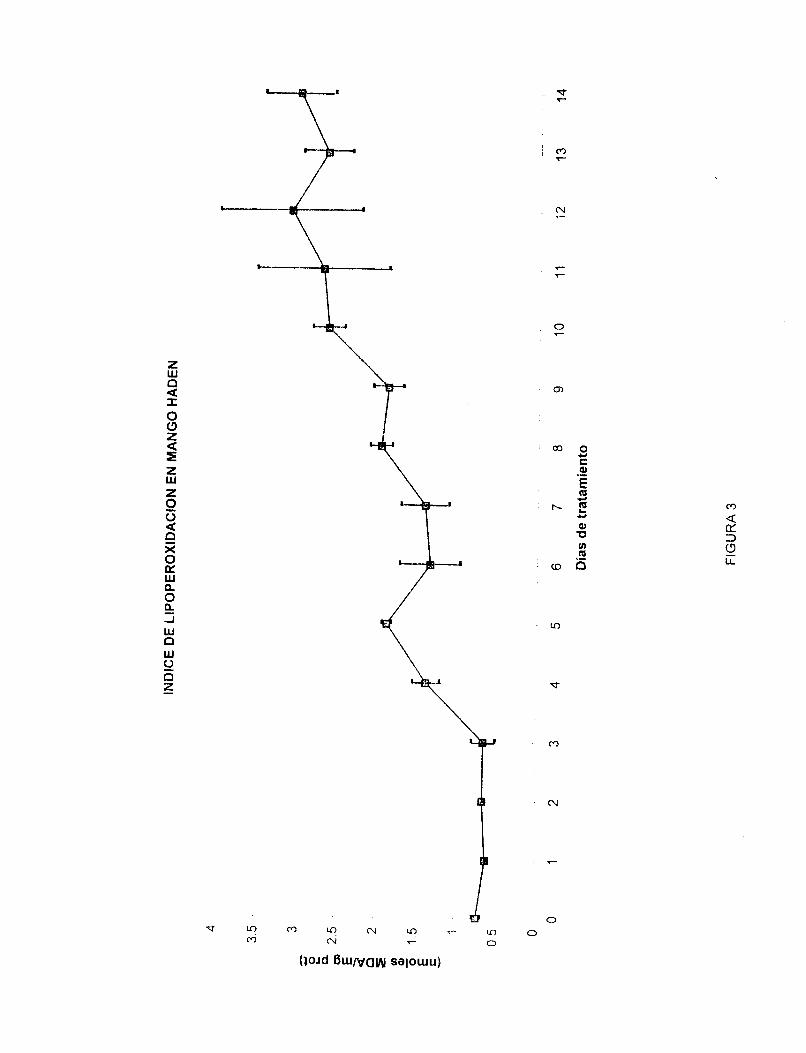
La figura 3 muestra el índice de lipoperoxidación del mango 'Haden' durante su maduración a temperatura ambiente. Se grafican nrrloles de MDA contra días de tratamiento. En la figura se observa el aumento en el índice de lipoperoxidación a partir del tercer día postcosecha para después ir aumentando paulatinamente hasta alcanzar un máximo entre el décimo y el doceavo día postcosecha (2.6298 nmoles/mg prot. y 3.1034 nmoles/mg prot., respectivamente) manteniéndose constante hasta el día catorce.
La determinación de la lipoperoxidación se hizo de acuerdo a la tkcnica descrita por Buege ¿? Aust (1 978). Sin embargo, Du & Bramlage (1 992) reportan una técnica modificada para la determinacibn de lipoperoxidación en tejidos vegetales ricos en amQcares, ya que en algunos frutos los azúcares (por ejemplo, sacarosa) incrementan las lecturas a una absorbancia de 535 nm en el espectrofotometro, por lo cual se hizo la determinación de una curva estándar de sacarosa con el fin de observar si ocurria alguna interfelrencia con las lecturas de hpoperoxidación tanto para el mango 'Kent' como para el mango 'Haden' (figura 4) Los datos de la figura 4 muestran que las lecturas de las absorbancias par2 sacarosa son muy bajas, no interferiendo por tanto con las lecturas de fipoperoxidación.

La tabla 1 muestra la participación de la vía alternativa durante la maduración del mango 'Kent' (Díaz de León et a/, 1995). Como se observa en la tabla la participación de la vía alternativa aumenta mielntras que la de la via cttocr6rnica disminuye durante la maduración. La máxima participación ocurre al quinto dia de postcosecha (séptimo día de tratamiento) justo cuando ia actividad respiratoria del mango se vio incrementada. Esto hace pensar que la via alterncdiva participa de manera importante cuando la actividad respiratoria aumenta antes del pico climatérico.
La participación máxima de la via alternativa (día 5 de postcosecha) ocurre cuando los niveles de lipoperoxidación comienzan a incrementarse rapidamente, pero durante la disminución de la participación de la vía alternativa la lipoperoxidacibn sigue aumentando y llega a su rrláxirno para despues estabilizarse. En relación a esto último, se requieren estudios posteriores para establecer ¡a relacibn entre la produccibn de radicales libres y los sistemas enzimáticos responsables de su atrapamiento asi conno de los niveles de antioxidantes durante la maduraci6n del mango 'Kent'. Estos resultados sor? consistentes con estudios realizados durante la germinacibn de maiz (Leprince. 6994) y otros tejidos vegetales sometidos a estrés (Purvis 8, Shewfelt, 1993).

c11 O c
eu &5
o CJ
N T”
O r
c b i n
P o w a
T”
z w
w n w
f? z o I.
7 I,
c."
"" I_ "".,-,, \
t" ,-
o r-
o
LD
z w
J
n w
w o n z
'c1 o,
In m ' ( D ii
d
m

3 O +
I “ + d v- F d
lh m
c! d CD

m O o d
a Y-
"
a3

l" " " "1
. .
c co co I"
. .
M U
9 O
(D
2 o
5) u U) -3-
o
f . ,
o \-.
1 . .
. . .
?u t- z w a 5
O 0 0
o !"

N r- w - o
. . . N E?
:¿? 'N
. .
. .
r- co 10 v-
hj
. .
. .
. . I .
. .
: / ,
2 . . in v-
d T.-
+
i3 c c r c
b ' l 'c-
F ' P J O'
a 0 0
..
m m ( 9 N (u N' li-I in ii?

io r- ,- Y
. .
1



I
c t w u O u z a I 2 w Z o
w
\ 1
O O

RECOMENDACIONES.
-Realizar la determinacián de radicales libres por EPR., que no se realizh en este proyecto debido a que el aparato de EPW de CE3 se encontraba en reparación.
-Deteaminar los niveles de antioxidantes (vitimina C, vitamina E, &c.) durante la rnaduracibn del mango.
-Determinar los niveles de enzirnas involucradas en el metabolismo de radicales libres (peroxidasa, la superóxido dismutasa, etc.) ciurante la rnaduracibn de! mango.
-Correlacionar los cambios de las estructuras celulares con los niveles de radicales libres durante el process de maduración del rnlango.

BIBLIOGRAFIA.
Azcón-Bieto, J. & Talón, M. (1993). Fisiología y Bioquímica Vegetal. la edición. Edit. lnteramericana Mcgraw-Hill. España. pp 463-464. Buege, J.A. & Aust, S.D. (1978). Microsomal lipid Peroxidation. In: Methods in Enzimology, vol. 52. Fleisher, S. & Packer, L. (Eds). Alcademic Press. New York.
Burg, S.P. & Burg, E.A. (1962). Role of ethylene in fruit ripening. Plant Physiol. 37:
Cammack,R.(1993). Electron Paramagnetic Resonance Spectroscopy of Metalloproteins. In: Methods in Molecular Biology vol. 17. Jones, C. et al (Eds). Human Press. New Jersey. pp 327-344. Díaz de León, F., Sánchez, G., Contreras, C., Kerbel, C., González, J.A., Ponce de León, L. & Pérez, L. (1995). Regulation of alternative oxidase activity by pyruvate in mango mitochondria. Memorias del VI1 Congreso Nacional de Bioquímica y Biología Molecular de Plantas. pp 121. Harman, D.(1956). Journal of Geront. 1 1 :298-300. Kurnar, S., Patil, B.C. & Sinha, S.K.(1990). Cyanide resistant respiration involved temperature rise in ripening mangoes. Biochem. Biophys. Res. Commun. 168:818- 822. Laksminarayana, S. (1 975). Relation of time harvest on respiration chemical constituyens and storage life of mangoes. Proc. Fla. State Hort Soc. 88:477-481. Laksminarayana, S. et al (1 977). Effect of refrigerate temperatures on the incidence of chilling injury and ripening quality of mango fruit. Proc. Fla. State Hort. soc. 90: Leprince, O. vander Wed, A., Deltour, R. & Lambers H.(1992). Respiratory pathways in germinating maize radicles correlated with desiccation tolerance and soluble sugars. Phisiol. Plant. 85:581-588. Leprince, O., Atherton, N.M.,Deltour, R. & Hendry, G.F.(1994). The involvement
of respiration in free radical processes during loss of desiccation tolerance in geminating Zea mays L.: An electron paramagnetic: resonance study. Plant
Leshem Y.Y.(1981). 3xy free radicals atld plant senescence.what's New Plant
McIntosh, L. (1994) Molecular Biology of the Alternative Oxidase . Plant. Physiol.
Millar, H.A., Wiskich, J.T., Whelan, J., Day, U.A. (1993) Organic acid activation of the alternative oxidase of plant mitochondrra. FEBS Lett. 392:259-262. Moore A.L. Bonner W.D. & Rich, P.R. (1978). The determination of the proton- motive force during cyanide-insensitive respiration in plant mitochondria. Arch. Biochem. Biophys. 186:298-306.
pp 302-31 O.
179-1 89.
Physiol. 104: 1333-1 339..
Physioi . 1211-3.
105:781-786.

Pérez-Flores, L., Ponse de León, L., Bósquez-Molinla, E. & Pelayo-Saldívar, C. (1993) La vía alternativa en el metaboDismo respiratorio de plantas superiores. Ciencia 44:41-52. Pome de león, L., Pérez-Flores, L., Pelays-Saidívar', C. & Bósquer-Molina, E. (1993) El proceso de senescencia. Ciencia 45121-1 26. Purvis, A.C. 8 Shewfelt, R.L. (1993). Does the alternative pathway ameliorate chilling injury in sensitive plant tissues?. Physiol. Plant. 88:712-718. Romani, R.J.(1991) A perpective on postharvest physiology and biochemistry. HortsScience 26: 1257-1 260. Rychter A.N. & Mikulska M.( 1990) The relationship between phosphate status and Cyanide-resistant respiration in bean roots. Physiol plant. 79:663-667. Rychter A.N., Chauveau M., Bomsef, J.L. & Lance, C. (1992) The effect of phospbate deficiency on mitochondrial activity and adenylate levels in bean roots. Physiol Plant. 84:80-86. Siedow, J.N. & Umbach, A.L. (1995). Plant mitochondrial electron transfer and molecular biology. Plant Cell 7:821-831. Skoos, DA. (1987). Anidisis Instrumental. 2a edic. Edit. Interarnericana. Mexico.
Soha!, R,S. (1987) The free radical theory of aging: a Critique. Review of Biological Research in Aging. Rothstein (Ed). New York, pp 385-415. Sohal, R.S. (1989) Adv. in Miochem. 221-34. Theodorou, M.E. & Plaxton, W.C. (1993). Metabolic adaptations of plant respiration to nutritional phosphate deprivation. Plant Physiol. 1 O1 :339-344. Umbach, A.L. & Siedow J.N. (1993) Covalent and nocovalent dimers of the cyanide-resistant alternative oxidase protein in higher plant mitochondria and their relationship to enzime activity. Plant Physiol. 103:845-$54. Vanlerberghe, G.C., Day, DA. , 'eagiskich, J.T., Vanlerberghe, A.E. & Mclntosh, L(1995) Alternative oxidase activity in tobacco leaf mitochondria: Dependence on tricarboxilic acid cycle-mediated redox regulation and pyruvate activation. Plant Physiol. 1 Q9:353-361.
pp 350-391.