chondrogenic potential of skeletal cell populations: selective growth of chondrocytes and their...
TRANSCRIPT

Chondrogenic Potential of Skeletal Cell Populations:Selective Growth of Chondrocytes and Their Morphogenesisand Development In VitroLOUIS C. GERSTENFELD,1* CYRIL D. TOMA,2 JONATHAN L. SCHAFFER,3 AND WILLIAM J. LANDIS4
1Musculoskeletal Research Laboratory, Boston University Medical Center, Boston, Massachusetts 021182Department of Orthopedic Surgery, University of Vienna Medical School, Vienna, Austria3Department of Orthopedic Surgery, Brigham and Woman’s Hospital and Harvard Medical School, Boston, Massachusetts 021154Department of Orthopedic Surgery, Children’s Hospital and Harvard Medical School, Boston, Massachusetts 02115
KEY WORDS type X collagen; hypertrophic cartilage; bone formation
ABSTRACT Most vertebrate embryonic and post-embryonic skeletal tissue formation occursthrough the endochondral process in which cartilage serves a transitory role as the anlage for thebone structure. The differentiation of chondrocytes during this process in vivo is characterized byprogressive morphological changes associated with the hypertrophy of these cells and is defined bybiochemical changes that result in the mineralization of the extracellular matrix. The mechanisms,which, like those in vivo, promote both chondrogenesis in presumptive skeletal cell populations andendochondral progression of chondrogenic cells, may be examined in vitro. The work presented heredescribes mechanisms by which cells within presumptive skeletal cell populations become restrictedto a chondrogenic lineage as studied within cell populations derived from 12-day-old chicken embryocalvarial tissue. It is found that a major factor associated with selection of chondrogenic cells is theelimination of growth within serum-containing medium. Chondrogenesis within these cell popula-tions appears to be the result of permissive conditions which select for chondrogenic proliferationover osteogenic cell proliferation. Data suggest that chondrocyte cultures produce autocrine factorsthat promote their own survival or proliferation. The conditions for promoting cell growth,hypertrophy, and extracellular matrix mineralization of embryonic chicken chondrocytes in vitroinclude ascorbic acid supplementation and the presence of an organic phosphate source. Thedifferentiation of hypertrophic chondrocytes in vitro is associated with a 10–15-fold increase inalkaline phosphatase enzyme activity and deposition of mineral within the extracellular matrix.Temporal studies of the biochemical changes coincident with development of hypertrophy in vitrodemonstrate that proteoglycan synthesis decreases 4-fold whereas type X collagen synthesisincreases 10-fold within the same period. Ultrastructural examination reveals cellular andextracellular morphology similar to that of hypertrophic cells in vivo with chondrocytes embedded ina well formed extracellular matrix of randomly distributed collagen fibrils and proteoglycan.Mineral deposition is seen in the interterritorial regions of the matrix between the cells and isapatitic in nature. These characteristics of chondrogenic growth and development are very similarin vivo and in vitro and they suggest that studies of chondrogenesis in vitro may provide a valuablemodel for the process in vivo. Microsc. Res. Tech. 43:156–173, 1998. r 1998 Wiley-Liss, Inc.
INTRODUCTIONMuch of skeletal formation during vertebrate embryo-
genesis and postnatal growth occurs by a processreferred to as the endochondral sequence. This is adevelopmentally regulated series of stages that pro-ceeds in a highly coordinated temporal and spatialmanner, in which there is sequential recruitment anddifferentiation of cells that form cartilage, vascular, andbone tissues. During the initial phases of this process,mesenchymal stem cells are recruited, and they con-dense and differentiate into chondrocytes to produce aninitially unmineralized and avascular cartilage modelof a developing bone. Subsequently, the chondrocyteswithin this model further differentiate and their elabo-rated matrix mineralizes to form calcified cartilage inspecific areas. The mineralization proceeds and contin-
ues in the presence of vascularization of the tissue. Thisis initiated by means of blood vessels accompanied byundifferentiated mesenchymal cells entering cartilageregions containing hypertrophic chondrocytes. Chondro-cytes within lacunae localized in these mineralizedareas die and perivascular cells adjacent to cores ofcalcified cartilage secrete new matrix that results inbone deposition. The process is completed when me-taphyseal remodeling occurs as the mineralized extra-cellular matrix of both bone and cartilage is ultimately
Contract grant sponsor: NIH; Contract grant numbers: HD 22400, AR41452,N01-HD-2–3144; Contract grant sponsor: Max Kade Foundation.
*Correspondence to: Dr. Louis C. Gerstenfeld, Musculoskeletal ResearchLaboratory, Boston University Medical Center, 715 Albany St., Housman 2,Boston, MA 02118.
Received 16 July 1998; accepted in revised form 19 July 1998
MICROSCOPY RESEARCH AND TECHNIQUE 43:156–173 (1998)
r 1998 WILEY-LISS, INC.

resorbed and replaced with bone alone (Farnum andWilsman, 1987; Ham and Cormack, 1979; Holtrop,1972; Hunziker et al., 1984; Shapiro et al., 1977).
In normal vertebrate development in vivo, an endo-chondral phase seldom occurs during the formation ofthe calvaria and the facial bones of the cranial skeleton.These tissues are generated by direct induction througha process known as membranous bone formation. Onthe other hand, cartilage tissue is present in someinstances, including that of articular cartilage surfacesthat form on the maxillary processes of the jaw by directdifferentiation of ‘‘secondary’’ cartilage within the peri-osteum (Thorogood, 1979). In addition, specific experi-mental manipulation in vivo such as that causingcalcium deficiency by growing chick embryos in ashell-less environment has been shown to give rise tocartilage formation within the calvaria (Jacenko andTuan, 1986). Such changes to either cartilage or boneduring vertebrate development would not be unantici-pated considering that cell populations derived fromembryonic calvaria from several species exhibit thepotential to differentiate into one of several multiplecell lineages including those of osteoblasts (Aronow etal., 1990; Bellows and Aubin, 1989; Gerstenfeld et al.,1987), adipocytes (Bellows et al., 1994; Beresford et al.,1992; Kodama et al., 1982; Yamaguchi and Kahn, 1991),myoblasts (Grigoriadis et al., 1988; Yamaguchi andKahn, 1991), and chondrocytes (Asahina et al., 1993;Bellows et al., 1989; Rifas et al., 1982; Toma et al.,1997). These results indicate that, although not nor-mally observed, there is a subset of cells within the cellpopulations of the cranial bones that retains chondro-genic potential and can express a cartilage phenotypein vitro. The selection of cells having a cartilage pheno-type in cultured calvarial cell populations has beenshown to occur as a result of a variety of experimentalconditions including treatment of the cultures withosteogenic protein (OP-1) (Asahina et al., 1993) orcorticosteroids (dexamethasone) (Grigoriadis et al.,1988); growth of the cells on gelatin bone matrix(Terashima and Urist, 1975); or growth of the cells inthe absence of serum (Cole et al., 1992; Rifas et al.,1982; Toma et al., 1997). These observations suggestthat the epigenetic factors that regulate normal lineageprogression of skeletal cells and presumably direct thechanges so noted might be both identified and studiedin vitro.
Studies from several groups also indicate that bothgenetic factors (Gerstenfeld et al., 1989) and environ-mental factors (Eavey et al., 1988) alter chondrocytephenotype and promote cell hypertrophy. With respectto the latter, chondrocyte hypertrophy can be stimu-lated in vitro by culturing cells in a three-dimensional(collagen or agarose) gel, selecting an appropriate cellplating density (Solursh and Meier, 1974), or supple-menting the cells with appropriate nutrient and vita-min D3 metabolites (Gerstenfeld et al., 1990) or ascorbicacid (Gerstenfeld and Landis, 1991; Leboy et al., 1989).The identification of the required growth conditions hasled to research studies that have examined the pheno-typic developmental progression of chondrocyte hyper-trophy and morphogenesis in vitro.
The present report examines the mechanism(s) andcontributory role of developmental versus environmen-tal factors in the selection of cells to differentiate into
the chondroblastic lineage and to undergo furtherhypertrophic differentiation. For this purpose, 12-day-old embryonic chicken calvaria and vertebrae havebeen used as tissue sources for presumptive skeletalcells and committed growth chondrocytes, respectively.The results detail underlying mechanisms of cell typeselection and limitation for the development of chondro-genic and osteogenic lineages, define phenotypic changesthat accompany hypertrophic differentiation, and de-scribe chondrocyte and matrix morphogenesis duringthe hypertrophic progression of events.
MATERIALS AND METHODSCell Culture
Skeletal cell populations were isolated by three se-quential trypsin-collagenase treatments of 12-day-oldchicken calvaria as previously described (Gerstenfeld etal., 1996). Only cells released from the third consecu-tive digest were used for experimental determinations.Cells were suspended in minimal essential medium(MEM) supplemented with 10% fetal bovine serum(FBS) (Sigma Chemical Co., St. Louis, MO) and platedat 1.25 x 105 cells/60 mm diameter and 5 x 105 cells/100mm diameter culture dishes. After 24 hours, cellswere washed once in phosphate buffered saline(PBS) with a subsequent change of medium to eitherMEM supplemented with 10% NU-Serum IV CultureSupplement (Collaborative Biomedical Products, Bed-ford, MA) (Wong and Tuan, 1993) or 10% FBS. Cultureswere maintained in MEM/10% FBS or MEM/10% NU-Serum for 2–3 weeks with medium changes every 2days until cells reached confluence. At that time, themedium was changed to BGJb (Fitton-Jacksonmodification) supplemented with 10% FBS or 10%NU-Serum. After another 2 days, the medium wassupplemented with 10 mM b-glycerophosphate (bGPO4)and 2 days later this medium was changed into ‘‘miner-alizing medium,’’ containing 10% FBS or 10% NU-Serum, 10 mM bGPO4, and 12.5 µg/ml ascorbic acid.The time at which the cultures were introduced to themineralizing medium was denoted as day 0. Thereafter,the medium was changed every 3 days until the experi-ments were terminated. All time points are referencedto day 0.
In studies focusing on growth chondrocyte matura-tion and hypertrophic development, committed chondro-cytes were prepared from vertebral bodies of 12-day-oldembryonic chickens. In these experiments, the chondro-cyte cultures were grown under conditions that werepermissive for growth chondrocyte differentiation aspreviously described (Gerstenfeld and Landis, 1991). Inother experiments, committed chondrocytes were pre-pared from the cephalic halves of 17-day embryonicchicken sterna and these cultures were grown underconditions identical to those used for the vertebralchondrocytes. In other investigations, conditioned me-dium supplementation experiments were carried outusing medium from the sternal chondrocyte cultures inthe following manner: Cephalic chondrocytes were se-lected and grown for at least 2 weeks in culture and,beginning at the end of the second week, medium wascollected (this was termed ‘‘conditioned medium’’). Col-lected fresh from the sternal chondrocyte culturesthrough the duration of the experiments, this condi-
157CHONDROGENIC POTENTIAL OF SKELETAL CELLS

tioned medium was mixed at a ratio of either one orthree parts to either three or one part, respectively, ofDulbecco’s modified eagle’s medium (DME) (SigmaChemical Co.) containing 10% FBS, 10 mM bGPO4, and12.5 µg/ml ascorbic acid. Calvarial cells from 12- or17-day-old embryonic chickens were then grown for 2weeks in 1:3 or 3:1 DME:conditioned supplement. Thismedium was replenished with conditioned mediumevery 2 days over the time period.
Bromodeoxyuridine LabelingBrdU labeling was as previously described (Gersten-
feld et al., 1996). Six days after their initiation, cellcultures prepared from 12-day-old embryonic chickencalvaria were pulsed for 3 hours with BrdU (SigmaChemical Co.) at a final concentration of 20 mM. At theend of the pulse period, cell layers were fixed for 30minutes at 4°C with 4% formaldehyde in 0.1 M phos-phate buffer, pH 7.2. Cell layers were then reacted for60 minutes with anti-BrdU antibody supplementedwith nuclease (Amersham Inc., Arlington Heights, IL),diluted 1:10 in PBS buffer containing 0.05% bovineserum albumin (BSA) (w/v) and 0.5% Tween (v/v).Secondary antibody reactions followed with anti-mouseFTIC-conjugated IgG.
Isolation and Analysis of mRNATotal RNA and DNA were isolated using Tri-Re-
agentTM, (Molecular Research Center, Cincinnati, OH)according to the manufacturer’s instructions. RNA wasresolved on 1% agarose gels containing 2.2 M formalde-hyde (Thomas, 1980) and 5 µg of total RNA based onOD260 were loaded per gel lane. Equal loading of RNAwas verified by ethidium bromide staining of the gelbefore blotting onto Biotrans nylon membranes (ICNBiomedical Inc., Aurora, OH). Chicken cDNAs used forthese studies were pro a1(I) collagen (Lehrach et al.,1979), pro a1(II) collagen (Young et al., 1984), osteocal-cin (Neugebauer et al., 1995), osteopontin (Moore et al.,1991), and aggrecan (Sai et al., 1986). Type X collagenwas detected using a 790 bp sequence beginning at thesplice junction of exon 3 of a chicken genomic type Xcollagen clone (gcCOLX; L.C. Gerstenfeld, unpublisheddata). This sequence corresponds to the sequence pub-lished by LuValle et al. (1988).
32P-radiolabeled cDNA probes were synthesized byrandom priming and hybridization was carried out at65°C in 50 mM PIPES, 100 mM NaCl, 50 mM Naphosphate, pH 7.0, 1 mM EDTA buffer containing 5%SDS (w/v) and 60 mg/ml single stranded salmon spermDNA. Hybridization was carried out for 18 hours in arotating hybridization oven (Robins Scientific, Sun-nyvale, CA). Autoradiograms were quantitated usingan LKB Ultrascan II laser scanning densitometer (LKB,Broma, Sweden) and values were normalized to 18Sribosomal RNA obtained by hybridization of each blot toa conserved nucleotide sequence probe of the 18Sribosomal subunit (Ambion Corp., Austin, TX).
Analysis of Collagen and Proteoglycan SynthesisCollagen protein synthesis was analyzed by pulse
labeling with 3H-proline. Cultures were labeled for 24hours using 5 ml of 50 µCi/ml 3H-proline (110 mCi/
mmol) (Amersham Inc.) after 6 days of growth inmineralizing medium. The total cell layer-associatedproteins and secreted medium proteins were processedin the presence of proteolytic inhibitors as describedpreviously (Gerstenfeld et al., 1984). Fifty thousandcpm of medium-labeled proteins and 100,000 cpm of celllayer-associated proteins were applied per lane for eachgel. Collagens were resolved by electrophoresis on5–10% continuous gradient SDS-polyacrylamide gels(Laemmli, 1970) and fluorography was carried out(Bonner and Laskey, 1974). Quantitation of fluoro-graphs was performed on an LKB Ultrascan II laserscanning densitometer (LKB, Broma, Sweden).
For proteoglycan analysis, cell cultures were grownin 33 mm tissue culture wells. At day 6 after the changeto complete medium, replicates (n 5 6) were labeledwith 50 µCi/ml Na2[35S]O4 (21 mCi/mmol) in 1.5 ml for24 hours. Proteoglycans were precipitated overnightwith 1.3% potassium acetate (v/v) in 95% ethanol.Samples were collected by centrifugation at 16,000g ina microcentrifuge for 30 minutes, resuspended, andprecipitated a second time. Incorporation was deter-mined by filtration of the precipitate onto glass filtersfollowed by scintillation counting (Gerstenfeld et al.,1990).
Statistical AnalysisBiochemical assays were performed with a minimum
of 6 replicates and were repeated with at least 2–3separate preparations of cells. All values were normal-ized to DNA content. Data were tabulated as themean 6 standard deviation (or standard error of themean where appropriate) with a minimum sample sizeof 6. Statistical analysis consisted of ANOVA, Student’st-test of significance, and the Tukey HSD test usingVersion 6.11 of SAS for Windows software package (SASInstitute Inc., Cary, NC). Values of p , 0.05 wereconsidered significant. Mean ratios for cell count datawere compared using an independent samples Stu-dent’s t-test (Bland and Altman, 1995). For all calcula-tions, two-tailed values of p , 0.05 were consideredstatistically significant.
Histological and Ultrastructural Analysisof Chondrocyte Cultures
Alkaline phosphatase expression was examined byhistochemical staining. Duplicate cultures were fixedwith 2% paraformaldehyde in 0.1 M cacodylate bufferat pH 7.4 for 10 minutes, rinsed with fresh buffer, andstained for alkaline phosphatase activity using naphtolAS-MX phosphate and Fast Red TR salts (Sigma Chemi-cal Co.). Matrix calcification was determined by hema-toxylin/eosin and von Kossa (phosphate) staining. Forultrastructural analysis, samples (cells and matrix)were fixed and embedded directly in culture platesusing either aqueous (glutaraldehyde-OsO4) or anhy-drous (ethylene glycol) techniques (Landis et al., 1977).Specimens were sectioned (,80 nm) with diamondknives on a Reichert Ultracut S ultramicrotome (Leica,Deerfield, IL) and floated briefly on an aqueous oranhydrous (ethylene glycol) trough fluid (Landis et al.,1977). Some sections were stained with 8% uranylacetate in absolute ethanol and with Sato’s lead.
158 L.C. GERSTENFELD ET AL.

Ultrastructural features of such sections were exam-ined by conventional transmission microscopy at 60 or80 kV in either a Philips EM 300 (Philips Electronic
Instruments, Inc., Mahwah, NJ) or a JEOL 100C(JEOL USA, Peabody, MA) electron microscope. Min-eral in unstained sections was studied by selected area
Fig. 1. Selection of chondrogenesis by growth of presumptive skeletalcell populations in serum-substituted medium. Left (top to bottom):Histochemical analysis of 12-day-old embryonic chicken calvarial cellcultures grown 6 days in medium containing either 10% fetal bovineserum (FBS) or 10% serum substitute (serum subt., SS). Cell layers werestained for either alkaline phosphatase (Apase) activity or proteoglycansbased on alcian blue reactivity. Representative staining characteris-tics of culture dish regions are denoted. Right (top): Bar graph ofalcian blue staining nodules in cultures grown in SS (solid bars) or10% FBS (open bars). The number of alcian blue staining nodules wascounted in 6 dishes and compared in 2 separate experiments. Error
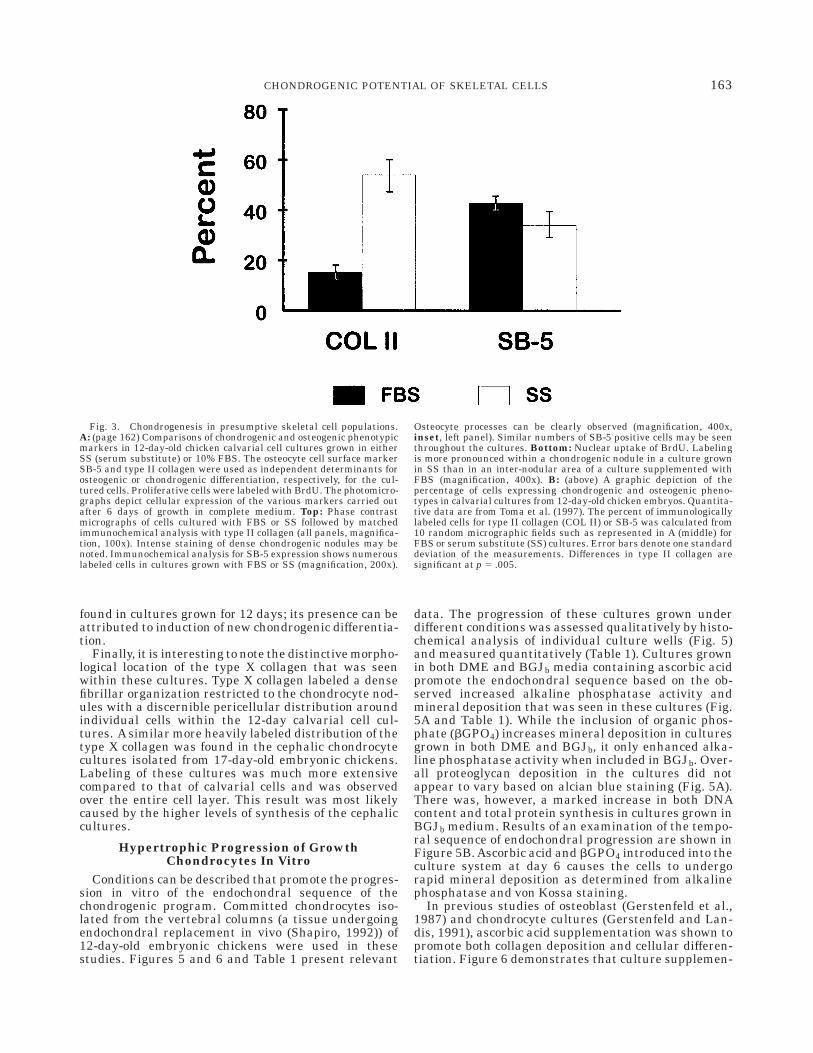
bars denote one standard deviation of these data. Right (middle):Analysis of proteoglycan synthesis in cultures grown 12 days in SS(solid bars) or 10% FBS (open bars). Cells were labeled on day 6 with50 µCi/ml Na2[35S]O4 (21 mCi/mmol) in 1.5 ml for 24 hours and counted asnoted in Materials and Methods. Error bars denote one standard deviation(n 5 10) for two separate experiments. Right (bottom): Analysis ofalkaline phosphatase enzyme activity (by p-nitrophenol, p-NP conver-sion) of cultures grown in SS (solid bars) or 10% FBS (open bars). Enzymeactivity was measured on day 6. The results in all panels were determinedfrom 12 wells in 2 separate experiments (exp 1 and exp 2). Error barsdenote one standard deviation. Left panel magnification, 100x.
159CHONDROGENIC POTENTIAL OF SKELETAL CELLS

electron diffraction at 80 or 100 kV in the Philipsinstrument and by electron probe microanalysis at 25kV in a STEM-modified JSM-50A scanning elec-tron microscope (JEOL USA) (Landis and Glimcher,1978, 1982). All microscopes were equipped with aliquid nitrogen anticontaminator to limit specimenartifacts.
RESULTSChondrogenic Selection Within Presumptive
Skeletal Cell Populations Isolated From12-Day-Old Embryonic Chicken Calvaria
The growth of 12-day-old embryonic chicken calvarialcell populations in medium in which FBS was replacedwith a serum substitute has been shown to promotechondrogenesis (Toma et al., 1997). As Figure 1 shows,histological staining of whole cultures with alcian blueidentified the presence of chondrogenic cell nodulesthroughout the cell layers in such serum-substitutedsamples. The number of alcian blue staining nodulescould be counted and compared between cultures grownin the absence or presence of medium containing FBS(Fig. 1, top right). These results demonstrate that thetotal number of chondrogenic nodules increased approxi-mately 5–10-fold in the cultures grown in serum substi-tute. Chondrogenesis was biochemically determined by
35SO4 labeling of sulfated proteoglycans (Fig. 1, middleright), and these results independently confirm an,8–20-fold increase in the chondrogenic phenotype ofthe serum-substituted cultures.
Differences in alkaline phosphatase enzyme activitywere assessed in the two sets of cultures. At early timepoints, histochemical staining for alkaline phosphatasewas seen only in the cells localized between the chondro-cyte nodules in the cultures containing serum substi-tute (Fig. 1, middle left). In contrast, alkaline phospha-tase staining was intense throughout the entire celllayer of the cultures containing FBS (Fig. 1, bottomleft). Such histochemical results suggest that the en-zyme activity would be much higher in the culturesgrown in FBS. However, biochemical measurement oftotal enzyme activity was considerably higher in cul-tures grown in serum-substituted medium after 12days (Fig. 1, bottom right). Histochemical staining after12 days was also seen throughout the cell layers ofcultures grown in either FBS or serum substitute (datanot shown). Thus, since both hypertrophic chondrocytesand osteoblasts express alkaline phosphatase, this as-say by itself cannot discriminate between osteogeniccells and hypertrophic chondrocytes. Moreover, as thechondrogenic cells progress toward hypertrophy, they
Fig. 2. Comparison of chondrogenic and osteogenic expression in12-day-old embryonic calvarial cultures grown under conditions per-missive for chondrogenesis. A: RNAs were isolated from cells after 12days of culture growth in mineralizing medium supplemented witheither SS (serum substitute) or 10% FBS. Northern blot analysis wasthen carried out for bone and cartilage mRNAs, including aggrecan(AG), osteopontin (OPN), osteocalcin (OC), and a1(I), a1(II), and a1(X)collagen chains. The efficiency and uniformity of blot transfers are
demonstrated by hybridization profiles to an 18S ribosomal probe andstaining with ethidium bromide (EtBr). B: Collagen protein synthesisin cultures grown for 12 days in complete medium supplemented witheither FBS or SS. The electrophoretic positions of 3H-proline-labeleda1(I 1 II), a2(I), pro a1(X), and a1(X) collagen chains isolated fromeither cell layers or medium are indicated. The horizontal linemarking to the left of some blots indicates the position of 27S mRNAused as a molecular weight indicator.
160 L.C. GERSTENFELD ET AL.

will express very high levels of alkaline phosphatase(see below).
The expression and hypertrophic progression of thechondrogenic phenotype were also examined by grow-ing the 12-day-old embryonic chicken calvarial culturesunder conditions that both permitted chondrogenesisand promoted mineralization. In these studies, a num-ber of genes that are specific for either the chondrogenicor osteogenic cells were examined (Fig. 2A). A definitiveincrease in aggrecan and type II and X collagen geneexpression (chondrogenesis markers) was seen in cul-tures grown in serum-substituted medium. However,as was previously demonstrated (Toma et al., 1997), theexpression of type X collagen was observed only afterlonger periods of growth in serum-substituted mediumthat had been specifically supplemented with ascorbicacid and bGPO4 (see below). While chondrogenic cellswere selectively increased when grown in serum substi-tute, there was only a slight diminishment in theosteogenic phenotype of the same cultures as deter-mined from osteopontin and osteocalcin gene expres-sion (osteogenesis markers), demonstrating that thecultures maintained their overall osteogenic differentia-tion.
The relationship between levels of mRNA expressionand new collagen synthesis was assessed in the cul-tures by pulse labeling with 3H-proline after 12 days ofgrowth in mineralizing medium (Fig. 2B). Type I colla-gen synthesis was detected in cultures grown in bothserum-substituted and FBS-containing medium as evi-denced by the detection of the a2(I) collagen chain. Inboth cultures a2(I) was detected as a major secretoryproduct based on its prevalence in the medium. Anincreased type II collagen expression was indicated by agreatly increased a1 to a2 ratio seen in serum-substituted cultures. In comparing the cultures grownunder the different conditions, proline labeled bands atthe position of pro a1(X) and its processed form weredetected only in the cultures grown in serum-substi-tuted medium. Consistent with previous analysis (Ger-stenfeld and Landis, 1991), type X collagen was shownto increase preferentially in its accumulation in the celllayers of the mineralizing culture.
The mechanisms by which various growth conditionspromote the selection of specific cell lineages within apopulation of presumptive skeletal cells may be exam-ined by determining the total number of cells within thepopulation that express osteogenic or chondrogeniclineage markers (Gerstenfeld et al., 1996; Toma et al.,1997). In the current study, this was carried out byimmunolabeling the cell populations grown under thedifferent culture conditions with markers selective forchondrogenic (type II collagen) and osteogenic (osteo-cyte SB-5) cells (Fig. 3). Analysis shows dense nodulesof birefringent rounded cells cultured with serum sub-stitute selectively labeled for type II collagen but fewsuch chondrogenic cells in cultures grown under nor-mal conditions in the absence of serum substitute (Fig.3A, first and second rows of panels). In contrast, withincell populations grown under either culture condition,the terminal osteogenic marker SB-5 was present oncells throughout both cultures (Fig. 3A, third row ofpanels).
In order to determine if there was selective prolifera-tion of chondrogenic cells compared to osteogenic cells,proliferative cells were labeled by BrdU incorporation
(Fig. 3A, bottom panels). There was selective incorpora-tion of BrdU within the chondrogenic nodules, while incomparison few cells were labeled in the internodularregions composed primarily of osteogenic cells. Theseresults show that the chondrogenic cells were moreproliferative than the osteogenic cells.
The percentage of cells that expressed either osteo-genic or chondrogenic markers was determined fromcell counts taken from random microscopic fields atidentical magnifications and these results are summa-rized in Figure 3B. Under control growth conditions inmedium supplemented with FBS, ,15% of the cells inthe embryonic cultures expressed a chondrogenic pheno-type while about ,40% of the cells in these cultureswere positive for the expression of the osteogenic SB-5marker. When these same cultures were grown inmedium containing serum substitute, there was a,3.5-fold increase in the cells that expressed the chon-drogenic phenotype such that ,50% of the cells werenow positive. This increase in the number of chondro-genic cells was associated with a slight decrease in thenumber of cells that expressed the osteocyte specificmarker to ,35% of the cell population. This decreasewas not statistically significant compared to the cellnumbers determined from FBS cultures. Thus, growthof these cells in serum substitute did not change theoverall percentage of cells that expressed an osteogenicphenotype based on their immunoreactivity for SB-5.
The results presented in Figure 3 suggest that chon-drocytes may produce autocrine factors that are selec-tive for their own survival or further proliferation.Preliminary studies carried out to test this hypothesisare depicted in Figure 4.
In these experiments, committed growth chondro-cytes derived from the cephalic portions of 17-day-oldembryonic chicken sterna were grown in culture andtheir medium was collected. This conditioned mediumwas added as a supplement to the medium of 12-day-oldchicken calvarial cells grown in FBS. The 12-day-oldembryonic calvarial cell populations were allowed togrow in the presence of different ratios of conditioned:FBS medium and the chondrogenic phenotype of thecultures was subsequently examined for expression oftype X collagen within the cell layers. Figure 4 shows acomparison of the type X collagen labeling seen in thesecultures supplemented in ratios of 1:3 or 3:1 andwithout conditioned medium. Increased labeling inten-sity and numbers of observable type X collagen-reactivechondrocyte nodules are found, dependent on the ratioof conditioned medium that was added. Immunostainedcephalic chondrocyte cultures grown under conditionspromoting chondrocyte hypertrophy were used as apositive control. Calvarial cell cultures from 17-day-oldembryonic chickens were used as negative controlssince previous studies demonstrated such cell popula-tions lacked the ability to undergo chondrogenesis inresponse to growth in serum-substituted medium (Tomaet al., 1997). This latter control was also used to addressthe question of whether there was adsorption of thetype X collagen to the osteoblast cell layers because oftype X presence in the conditioned medium isolatedfrom the cephalic chondrocyte cultures. No immunofluo-rescent labeling with type X collagen antibody wasapparent for the 17-day-old embryonic chicken cal-varial cell populations supplemented with a 3:1 ratio ofchondrocyte-conditioned medium. Type X collagen was
161CHONDROGENIC POTENTIAL OF SKELETAL CELLS

Fig. 3 A.
found in cultures grown for 12 days; its presence can beattributed to induction of new chondrogenic differentia-tion.
Finally, it is interesting to note the distinctive morpho-logical location of the type X collagen that was seenwithin these cultures. Type X collagen labeled a densefibrillar organization restricted to the chondrocyte nod-ules with a discernible pericellular distribution aroundindividual cells within the 12-day calvarial cell cul-tures. A similar more heavily labeled distribution of thetype X collagen was found in the cephalic chondrocytecultures isolated from 17-day-old embryonic chickens.Labeling of these cultures was much more extensivecompared to that of calvarial cells and was observedover the entire cell layer. This result was most likelycaused by the higher levels of synthesis of the cephaliccultures.
Hypertrophic Progression of GrowthChondrocytes In Vitro
Conditions can be described that promote the progres-sion in vitro of the endochondral sequence of thechondrogenic program. Committed chondrocytes iso-lated from the vertebral columns (a tissue undergoingendochondral replacement in vivo (Shapiro, 1992)) of12-day-old embryonic chickens were used in thesestudies. Figures 5 and 6 and Table 1 present relevant
data. The progression of these cultures grown underdifferent conditions was assessed qualitatively by histo-chemical analysis of individual culture wells (Fig. 5)and measured quantitatively (Table 1). Cultures grownin both DME and BGJb media containing ascorbic acidpromote the endochondral sequence based on the ob-served increased alkaline phosphatase activity andmineral deposition that was seen in these cultures (Fig.5A and Table 1). While the inclusion of organic phos-phate (bGPO4) increases mineral deposition in culturesgrown in both DME and BGJb, it only enhanced alka-line phosphatase activity when included in BGJb. Over-all proteoglycan deposition in the cultures did notappear to vary based on alcian blue staining (Fig. 5A).There was, however, a marked increase in both DNAcontent and total protein synthesis in cultures grown inBGJb medium. Results of an examination of the tempo-ral sequence of endochondral progression are shown inFigure 5B. Ascorbic acid and bGPO4 introduced into theculture system at day 6 causes the cells to undergorapid mineral deposition as determined from alkalinephosphatase and von Kossa staining.
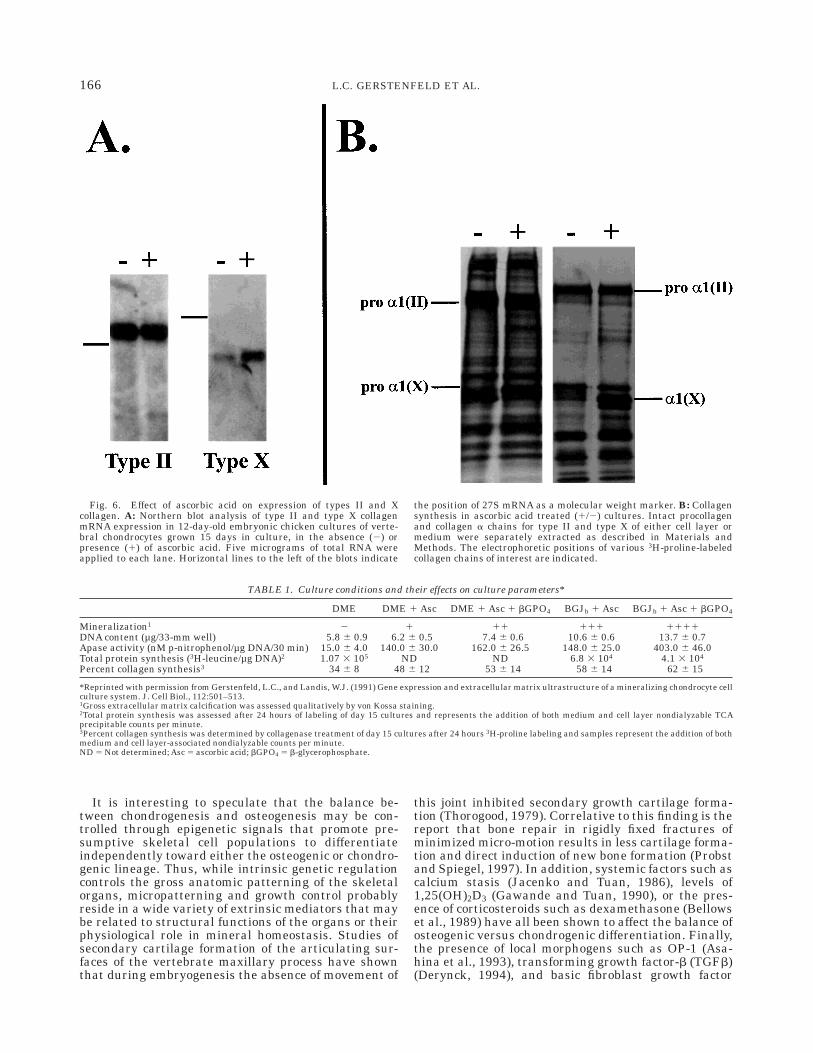
In previous studies of osteoblast (Gerstenfeld et al.,1987) and chondrocyte cultures (Gerstenfeld and Lan-dis, 1991), ascorbic acid supplementation was shown topromote both collagen deposition and cellular differen-tiation. Figure 6 demonstrates that culture supplemen-
Fig. 3. Chondrogenesis in presumptive skeletal cell populations.A: (page 162) Comparisons of chondrogenic and osteogenic phenotypicmarkers in 12-day-old chicken calvarial cell cultures grown in eitherSS (serum substitute) or 10% FBS. The osteocyte cell surface markerSB-5 and type II collagen were used as independent determinants forosteogenic or chondrogenic differentiation, respectively, for the cul-tured cells. Proliferative cells were labeled with BrdU. The photomicro-graphs depict cellular expression of the various markers carried outafter 6 days of growth in complete medium. Top: Phase contrastmicrographs of cells cultured with FBS or SS followed by matchedimmunochemical analysis with type II collagen (all panels, magnifica-tion, 100x). Intense staining of dense chondrogenic nodules may benoted. Immunochemical analysis for SB-5 expression shows numerouslabeled cells in cultures grown with FBS or SS (magnification, 200x).
Osteocyte processes can be clearly observed (magnification, 400x,inset, left panel). Similar numbers of SB-5 positive cells may be seenthroughout the cultures. Bottom: Nuclear uptake of BrdU. Labelingis more pronounced within a chondrogenic nodule in a culture grownin SS than in an inter-nodular area of a culture supplemented withFBS (magnification, 400x). B: (above) A graphic depiction of thepercentage of cells expressing chondrogenic and osteogenic pheno-types in calvarial cultures from 12-day-old chicken embryos. Quantita-tive data are from Toma et al. (1997). The percent of immunologicallylabeled cells for type II collagen (COL II) or SB-5 was calculated from10 random micrographic fields such as represented in A (middle) forFBS or serum substitute (SS) cultures. Error bars denote one standarddeviation of the measurements. Differences in type II collagen aresignificant at p 5 .005.
163CHONDROGENIC POTENTIAL OF SKELETAL CELLS

tation with ascorbic acid promotes chondrocyte hyper-trophic differentiation as demonstrated by the inductionof high levels of type X collagen mRNA expression andprotein synthesis. Figure 7 summarizes the overalltemporal relationships between proteoglycan and colla-gen synthesis to mineral deposition and relates these toaggrecan and types II and X collagen mRNA expres-sion. Here, it is apparent that proteoglycan synthesisand message levels fall rapidly after a few days ofculture time while collagen content increases and thendrops and mineral continuously increases. The rise andfall in collagen synthesis is mirrored by a precedingtype II collagen message expression. Type X expressiongradually rises throughout the culture period.
The morphogenesis of the chondrocytes and theirmatrices under culture growth conditions permissivefor endochondral progression is shown in Figures8–11. Corresponding to the changes in gene expressionand biochemistry described above, there are changes in
the structure of the chondrocytes and their extracellu-lar regions that likewise reflect a progression throughthe endochondral sequence. The temporal developmentof the cultures morphologically shows increasing num-bers of cells having rounded shapes and residencewithin lacunae (Fig. 8). Changes in cell size, shape,distribution, and cytoplasmic content in the culturesare apparent (Figs. 8–11) with larger chondrocytesbeing associated with not only more mature cultures(grown for longer periods) but also the relatively deeperregions of a culture dish where mineralization is becom-ing prominent (Figs. 8, 10). Representative calvarialchondrocytes from 12-day-old embryonic chickens andgrown for 17–18 days in culture are generally markedby typical intracellular organelles, including a singlenucleus, extensive endoplasmic reticulum, vesicles, andvacuoles (Fig. 9). Many chondrocytes, particularly thelarger ones, are observed within a highly stainingpericellular region characterized by the thin fibrils of
Fig. 4. Autocrine selection of chondrogenic growth of 12-day-oldembryonic calvarial cell populations. Type X collagen matrix deposi-tion in 12-day-old embryonic chicken calvarial cultures grown in thepresence of conditioned medium from hypertrophic chondrocytesderived from 17-day-old embryonic chicken cephalic sterna. Theidentity and distribution of type X collagen extracellular matrixaccumulation within chondrogenic nodules were determined by immu-nolocalization. Top: Calvarial cell cultures grown for 12 days in
complete medium containing 10% FBS without conditioned medium(control) or with a 1:3 or 3:1 ratio of conditioned medium from cephalicsternal chondrocyte cultures. Bottom: Negative and positive controls,respectively, of 17-day-old chicken embryo calvarial cell populationstreated with a 3:1 ratio of chondrocyte-conditioned medium (left) andcephalic sternal chondrocyte cultures after 12 days of growth inmineralizing medium. All photomicrographs: magnification, 400x.
164 L.C. GERSTENFELD ET AL.

type II collagen and small condensations suggestive ofproteoglycans (aggrecan) (Fig. 11). Mineral in the formof dense, irregularly shaped deposits is exclusivelyextracellular in location and is found overlying thecollagen (Fig. 10), frequently following individual fibrilsand in clear association with them (Fig. 11). There aremany cytoplasmic processes that invade the extracellu-lar regions of the cultures (Fig. 11) and these appear toterminate in small vesicles (Fig. 11). Consistent associa-tion of such vesicles with mineral is equivocal. Withtime, the cultures mineralize more completely, and thenumerous mineral deposits can be identified structur-ally as apatitic on the basis of their electron diffractioncharacter (data not presented; see Gerstenfeld andLandis, 1991).
DISCUSSIONThe data presented here focus on the growth condi-
tions in vitro that either induce chondrogenesis frompresumptive or uncommitted cells or promote chondro-cyte maturation and terminal differentiation toward ahypertrophic phenotype. The underlying mechanism(s)
by which mesenchymal stem cells are restricted to theosteogenic vs. the chondrogenic lineage was examinedin skeletal cell populations isolated from embryonicchicken calvaria. The use of cell populations fromcalvaria is particularly informative since this tissueundergoes direct bone induction in the absence of anendochondral stage. Thus, the appearance of chondro-cytes in these cell populations arises through theinduction and differentiation of the mesenchymal stemcells to chondrocytes. The data presented and reviewedhere indicate that the developmental restriction ofskeletal cells of the calvaria was not a result of positiveselection for osteogenic differentiation but a negativeselection against the progressive growth of chondro-genic cells in the absence of either a permissive orinductive environment. This conclusion is based on thefindings that the 12-day-old embryonic calvarial cellpopulations selectively proliferated to chondrogeniccells but were little changed in their percentage ofosteogenic cells when they were induced to undergochondrogenesis.
Fig. 5. Histochemical analysis of proteoglycan, alkaline phospha-tase, and mineral deposition in cultures of vertebral chondrocytesgrown under different conditions. A: All cultures were grown for 15days in BGJb or DME with or without b-glycerophosphate (b-GPO4).On day 15, separate 33 mm wells were analyzed using histochemicalstaining to assess proteoglycan deposition (alcian blue), mineral
deposition (von Kossa silver nitrate), or alkaline phosphatase (Apase,fast red). B: Time course over 20 days of mineral accumulation andalkaline phosphatase activity in 33 mm culture wells containingvertebral chondrocytes grown in BGJb medium supplemented withb-GPO4. Images are of the full 33 mm wells.
165CHONDROGENIC POTENTIAL OF SKELETAL CELLS
It is interesting to speculate that the balance be-tween chondrogenesis and osteogenesis may be con-trolled through epigenetic signals that promote pre-sumptive skeletal cell populations to differentiateindependently toward either the osteogenic or chondro-genic lineage. Thus, while intrinsic genetic regulationcontrols the gross anatomic patterning of the skeletalorgans, micropatterning and growth control probablyreside in a wide variety of extrinsic mediators that maybe related to structural functions of the organs or theirphysiological role in mineral homeostasis. Studies ofsecondary cartilage formation of the articulating sur-faces of the vertebrate maxillary process have shownthat during embryogenesis the absence of movement of
this joint inhibited secondary growth cartilage forma-tion (Thorogood, 1979). Correlative to this finding is thereport that bone repair in rigidly fixed fractures ofminimized micro-motion results in less cartilage forma-tion and direct induction of new bone formation (Probstand Spiegel, 1997). In addition, systemic factors such ascalcium stasis (Jacenko and Tuan, 1986), levels of1,25(OH)2D3 (Gawande and Tuan, 1990), or the pres-ence of corticosteroids such as dexamethasone (Bellowset al., 1989) have all been shown to affect the balance ofosteogenic versus chondrogenic differentiation. Finally,the presence of local morphogens such as OP-1 (Asa-hina et al., 1993), transforming growth factor-b (TGFb)(Derynck, 1994), and basic fibroblast growth factor
Fig. 6. Effect of ascorbic acid on expression of types II and Xcollagen. A: Northern blot analysis of type II and type X collagenmRNA expression in 12-day-old embryonic chicken cultures of verte-bral chondrocytes grown 15 days in culture, in the absence (2) orpresence (1) of ascorbic acid. Five micrograms of total RNA wereapplied to each lane. Horizontal lines to the left of the blots indicate
the position of 27S mRNA as a molecular weight marker. B: Collagensynthesis in ascorbic acid treated (1/2) cultures. Intact procollagenand collagen a chains for type II and type X of either cell layer ormedium were separately extracted as described in Materials andMethods. The electrophoretic positions of various 3H-proline-labeledcollagen chains of interest are indicated.
TABLE 1. Culture conditions and their effects on culture parameters*
DME DME 1 Asc DME 1 Asc 1 bGPO4 BGJb 1 Asc BGJb 1 Asc 1 bGPO4
Mineralization1 2 1 11 111 1111DNA content (µg/33-mm well) 5.8 6 0.9 6.2 6 0.5 7.4 6 0.6 10.6 6 0.6 13.7 6 0.7Apase activity (nM p-nitrophenol/µg DNA/30 min) 15.0 6 4.0 140.0 6 30.0 162.0 6 26.5 148.0 6 25.0 403.0 6 46.0Total protein synthesis (3H-leucine/µg DNA)2 1.07 3 105 ND ND 6.8 3 104 4.1 3 104
Percent collagen synthesis3 34 6 8 48 6 12 53 6 14 58 6 14 62 6 15
*Reprinted with permission from Gerstenfeld, L.C., and Landis, W.J. (1991) Gene expression and extracellular matrix ultrastructure of a mineralizing chondrocyte cellculture system. J. Cell Biol., 112:501–513.1Gross extracellular matrix calcification was assessed qualitatively by von Kossa staining.2Total protein synthesis was assessed after 24 hours of labeling of day 15 cultures and represents the addition of both medium and cell layer nondialyzable TCAprecipitable counts per minute.3Percent collagen synthesis was determined by collagenase treatment of day 15 cultures after 24 hours 3H-proline labeling and samples represent the addition of bothmedium and cell layer-associated nondialyzable counts per minute.ND 5 Not determined; Asc 5 ascorbic acid; bGPO4 5 b-glycerophosphate.
166 L.C. GERSTENFELD ET AL.

Fig. 7. Comparison of chondrocyte min-eral deposition, collagen synthesis, proteogly-can synthesis, and gene expression over a20-day growth period in vitro. Top: Totalcollagen synthesis (l), proteoglycan synthe-sis (●), and mineral deposition (o) were deter-mined from triplicate 33 mm culture dishes ofduplicate experiments with 12-day-old embry-onic chicken vertebral chondrocytes. Bot-tom: Comparison of type II collagen (l), typeX collagen (o), and aggrecan (●) mRNA geneexpression over 21 days. The left axis of theplot pertains to quantitative levels of aggre-can (Ag) and the right axis to quantitativelevels of either type II or type X collagen.Error bars denote total range of experimentalvariation.
167CHONDROGENIC POTENTIAL OF SKELETAL CELLS

(FGF) (Rousseau et al., 1994) are also suggested toinfluence the balance between chondrogenesis and os-teogenesis for presumptive skeletal cell populations.
The apparent local control in the balance of chondro-genic and osteogenic differentiation has led to theconcept that these processes are coupled in some man-ner (Gerstenfeld and Shapiro, 1996). For example,promotion of chondrogenesis by an absence of serum ingrowth medium is consistent with studies showing thateither embryonic calvarial cell cultures or undifferenti-ated limb bud cells in serum-free conditions becomechondrogenic rather than osteogenic (Cole et al., 1992;Rifas et al., 1982; Toma et al., 1997; Wong and Tuan,1995). Unlike many other cell types, chondrocytesappear to be specifically adapted to grow in serum-freeconditions in vitro (Ballock and Reddi, 1994; Bruckneret al., 1989; Quarto et al., 1992). These data might alsosuggest that serum contains factors that either inhibitchondrogenesis or promote selective growth of osteo-genic cells.
Chondrocytes themselves may produce factors thatpromote their own differentiation or growth. This con-clusion is supported by studies that have shown that
chondrogenesis can occur when cells are grown at highplating densities or in micromass cultures, even in thepresence of serum supplementation (Osdoby andCaplan, 1979). Other studies demonstrate that chondro-cytes cultured at high densities produce autocrinefactors that enable them to survive at low cell densitiesin the absence of serum (Ishizaki et al., 1994), and it isinteresting to speculate that growth in serum substi-tute reported here and elsewhere (Toma et al., 1997;Wong and Tuan, 1995) may have either promotedchondrogenesis or maintained critical numbers of chon-drogenic cells that allow their survival and expansionin culture. Further work reports that calvarial cellpopulations isolated from 14-day avian embryos andgrown in vitro produce factors that inhibit limb budmesenchymal cells from undergoing chondrogenesis(Wong and Tuan, 1995). Moreover, the limb bud mesen-chymal cells produce factors that stimulate selectedcalvarial cell populations to undergo chondrogenesis(Wong and Tuan, 1995). The results in the presentstudy provide yet additional evidence that chondro-cytes produce factors that promote chondrogenic differ-entiation. All the above results lead to the conclusionthat, under normal growth conditions in vivo, osteo-genic differentiation predominates as a result of regu-lated selection against progressive chondrogenic differ-entiation, which itself is probably under the controlprimarily of autocrine and paracrine factors producedby the local precursor cell population.
Considerable data has accumulated pertaining to theculture conditions that promote growth chondrocytedifferentiation. From the research summarized hereand that of several other groups (Gerstenfeld andLandis 1991; Leboy et al., 1989; Sandell and Daniel,1988), one of the most critical factors leading to mainte-nance of the chondrocyte phenotype, progressive cul-ture growth, and hypertrophic differentiation of thesecells is the inclusion of ascorbic acid in the growthmedia. The role of ascorbic acid as an essential cofactorin collagen synthesis is well documented (Lyons andSchwartz, 1984), and the biochemical data presentedhere show that ascorbic acid increased both total colla-gen synthesis and matrix accumulation. Ultrastruc-tural analysis further demonstrates that over the pe-riod of chondrocyte growth in vitro, these cellselaborated an extensive extracellular matrix that re-tained morphological features comparable to those seenin vivo, including increasing collagen content withtime. Recent studies of both chondrocytes and osteo-blast growth in vitro describe the crucial role theextracellular matrix (ECM) plays in stimulating cellu-lar differentiation (Franceschi et al., 1994; Gerstenfeldet al., 1988; Gerstenfeld and Landis, 1991). Thesereports and the present data suggest that the differen-tiation of specialized connective tissue cells not only iscontrolled by extrinsic soluble regulators but also isdependent on some form of feedback regulatory processrelated to the assembly and deposition of their matri-ces. While the exact mechanism(s) by which the ECMmay exert its effects on cellular differentiation in thismanner are incompletely understood, considerable in-formation has accumulated to support the concept thatboth the maintenance of normal cell morphology andspecific receptor interactions with the ECM are quite
Fig. 8. A light micrograph of a 12-day-old embryonic chickencalvarial chondrocyte culture grown for 18 days in mineralizingmedium. The culture has been sectioned to show its full thicknessprofile. The tissue was fixed in glutaraldehyde and osmium tetroxideand stained with toluidine blue. The interface between the culturedish and the cultured cells and matrix is marked by the relativelystraight line along the bottom of the micrograph. The culture consistsof numerous cells, each having a generally round shape and residing insingle lacunae. The size, shape, and disposition of the cells are similarto those features of growth plate chondrocytes observed in situ. Theextracellular matrices vary in their intensity of staining as do some ofthe cells. Pericellular staining marks the lacunae of certain cells(arrowheads) and, in the deeper portions of the culture, nearer theculture dish bottom, there is deposition of considerable mineral(arrows). Magnification, 1,200x; bar 5 10 µm.
168 L.C. GERSTENFELD ET AL.

important in promoting normal skeletal cell differentia-tion (Franceschi, 1992).
While the data presented here define growth condi-tions that promote growth chondrocyte differentiation,they also summarize several fundamental features thatcharacterize synthetic changes associated with hyper-trophic cartilage maturation and matrix formation.One of the most striking is an inverse relationshipbetween the levels of collagen and proteoglycan synthe-sis in comparison to mineral deposition. Thus, increas-ing collagen synthesis may be correlated with mineralaccumulation while proteoglycan synthesis levels de-crease. Such a relationship is consistent with earlierwork (Gerstenfeld and Landis, 1991) and with thesuggestions that proteoglycans inhibit mineral nucle-ation (Buckwalter et al., 1987) while the development ofa collagenous matrix is a prerequisite to mineral deposi-tion (Glimcher, 1992). It is also apparent that thesynthetic levels in collagen and proteoglycan observedhere are primarily controlled at a pre-translationallevel as reflected by comparisons in collagen and aggre-can mRNA. Further, while overall collagen synthesis isincreasing, it is largely a result of increasing levels oftype X collagen expression. The reciprocal relationshipfound here in the expression of types II and X collagenis comparable to the reported temporal patterns of
expression for these genes observed during the endo-chondral differentiation of appendicular growth carti-lage in vivo (Schmid and Linsenmayer, 1985).
Both the cells and extracellular matrices within thecultures described here develop and retain a number ofultrastructural features that have their close correlateswith the biochemical results presented above as well asthe structural characteristics of chondrocytes and ma-trix in vivo. These include, for example, the presence ofhigh levels of alkaline phosphatase as mineralizationoccurs and progresses in culture. Microscopically, alka-line phosphatase and other enzymes may be identifiedby specific immunoreactivity of culture sections orinferred principally through the observation of electrondense condensates appearing within extracellularvesicles. In this paper, the many such dense vesicles arepresent in the extracellular regions of calvarial chondro-cytes as cultures of these cells grow and mineralize.Their detection follows the progressive changes inalkaline phosphatase measured with nitrophenol. Asecond correlate is reflected in the overall chondrocyteculture thickness that is elaborated and increases withtime. The changes observed can be attributed to asignificant degree of hydration of the culture matrixthrough water retention mediated by proteoglycans.Evidence for these macromolecules is clear in micro-
Fig. 9. An electron micrograph of a calvarial chondrocyte culturegrown for 17 days in mineralizing medium. The region contains anumber of cells typically having a single nucleus and nucleolus as wellas cytoplasmic organelles marked by numerous vesicles and vacuolesand extensive endoplasmic reticulum. The cells reside within lacuna(L) defined by relatively electron transparent pericellular spaces (S).
The extracellular matrix beyond the pericellular regions of the tissueis characterized principally by collagen fibrils (C). Mineral (M) ap-pears as electron dense, small punctate and larger irregularly shapedextracellular deposits. This specimen is torn (T) in places as a result ofsectioning through the mineral. Magnification, 2,850x; bar 5 5.0 µm.
169CHONDROGENIC POTENTIAL OF SKELETAL CELLS

graphs taken from regions spanning wide portions ofthe culture volume, a result that is consistent with theknown presence, location, and distribution of proteogly-cans in vivo. In addition, the ultrastructural studieshere reveal numerous type II collagen fibrils compris-ing the culture matrices and whose appearance corre-sponds to their biochemical detection in the sameculture samples as well as their documented presencein vivo. Perhaps the most important observation madein this work is that these culture systems will mineral-ize and they do so in a reproducible manner. As such,these systems are distinct from others that mineralizeunpredictably or not at all or elaborate mineral that isabnormal. In the present cultures, the mineral isapatitic, having a chemical nature identical by electrondiffraction means to that found in vivo. The mineral isalso discretely deposited and is associated principallywith the many type II collagen fibrils comprising thematrices. In these aspects, the chondrocyte culturesagain follow the manner of mineralization that has
been reported extensively, based on many studies ofgrowth cartilage in vivo.
In summary, the data reviewed here indicate thatstudies in vitro have led to considerable understandingof mechanisms that regulate chondrocyte differentia-tion, morphogenesis, extracellular matrix formation,and mineralization. These data suggest that chondro-genesis occurs in three stages: the differentiation ofchondrogenic cells within a population of presumptiveskeletal cells; the maintenance and expansion of thesechondrocytes; and chondrocyte terminal maturation.The progressive differentiation of chondrocytes in vitrodemonstrates that the cells develop in a manner compa-rable to that found in vivo. Chondrocyte maturation ischaracterized by an increasing expression of type Xcollagen and alkaline phosphatase and a decreasingexpression of type II collagen and aggrecan. Thesechanges are accompanied by correlative morphogeneticmodulations including chondrocyte progression to ahypertrophic phenotype and the development of a ma-
Fig. 10. Microscopic view of the hypertrophic state in vitro. Thelower portions of a culture nearer to the culture dish bottom may bemarked by calvarial cells (C) undergoing hypertrophy and havingsparse cytoplasmic elements. The extracellular matrix at these loca-
tions contains many mineral deposits (M) overlying or otherwise inapparent association with collagen fibrils of narrow diameter. Thismicrograph is taken from a culture grown for 17 days. Magnification,4,400x; bar 5 5.0 µm.
170 L.C. GERSTENFELD ET AL.

trix characterized ultimately by extensive apatitic min-eral deposition corresponding to that observed in vivo.
ACKNOWLEDGMENTSThis investigation was supported in part by NIH
grant HD22400 (L.C.G.), NIH AR41452 (W.J.L.), andthe Max Kade Foundation (C.D.T.). The authors ex-press thanks to Ms. Karen J. Hodgens (Children’sHospital, Boston, MA) for superb technical assistanceand manuscript preparation and to Drs. Tom Linsen-mayer (Tufts University School of Medicine, Boston,MA), Scott Bruder (Osiris Corp., Baltimore, MD), andArnold Caplan (Case Western Reserve University, Cleve-land, OH) for generously providing type X collagen andosteoblast lineage-specific antibodies, respectively,which were used in these studies. The authors are alsoindebted to Mr. Christopher George (Boston UniversityMedical Center) for assistance with the computer gener-
ated figures. Other antibodies were obtained from theDevelopmental Studies Hybridoma Bank, which ismaintained by the Department of Pharmacology andMolecular Sciences at Johns Hopkins University Schoolof Medicine, Baltimore, MD, and the Department ofBiology at the University of Iowa, Des Moines, IA,under contract NO1-HD-2–3144 from the NationalInstitutes of Health.
REFERENCESAronow, M.A., Gerstenfeld, L.C., Owen, T.A., Tassinari, M.S., Stein,
G.S., and Lian, J.B. (1990) Factors that promote progressive devel-opment of the osteoblast phenotype in cultured fetal rat calvariacells. J. Cell. Physiol., 143:213–221.
Asahina, I., Sampath, T.K., Nishimura, I., and Hauschka, P.V. (1993)Human osteogenic protein-1 induces both chondroblastic and osteo-blastic differentiation of osteoprogenitor cells derived from newbornrat calvaria. J. Cell Biol., 123:921–933.
Ballock, R.T., and Reddi, A.H. (1994) Thyroxine is the serum factor
Fig. 11. Electron micrograph of the chondrocyte-matrix interface.An enlargement of a region from a 17-day-old cell culture showscytoplasmic organelles, including mitochondria (arrows), the Golgiapparatus (G), dense vacuoles (DV), endoplasmic reticulum (ER), andother elements. The cell appears to be secreting numbers of thincollagen fibrils adjacent to the cell envelope and its infoldings. Thenarrow diameter and faint periodicity (arrowheads) of the fibrils areconsistent with structural features of type II collagen. Cell processes
(P) sectioned into a variety of profiles, possible vesicles (V) containingalkaline phosphatase, highly dense condensations of putative proteo-glycans (PG), and mineral deposits (M) may be observed in theextracellular matrix. The mineral is located over the collagen networkin the matrix and some deposits follow individual collagen fibrils(double arrowheads). The cellular and matrix characteristics illus-trated here are similar to those described for cartilage in vivo.Magnification, 25,600x; bar 5 1.0 µm.
171CHONDROGENIC POTENTIAL OF SKELETAL CELLS

that regulates morphogenesis of columnar cartilage from isolatedchondrocytes in chemically defined medium. J. Cell Biol., 126:1311–1318.
Bellows, C.G., and Aubin, J.E. (1989) Determination of numbers ofosteoprogenitors present in isolated fetal rat calvaria cells in vitro.Dev. Biol., 133:8–13.
Bellows, C.G., Heersche, J.N.M., and Aubin, J.E. (1989) Effects ofdexamethasone on expression and maintenance of cartilage inserum-containing cultures of calvaria cells. Cell Tissue Res., 256:145–151.
Bellows, C.G., Wang, Y.H., Heersche, J.N.M., and Aubin, J.E. (1994)1,25-dihydroxyvitamin D3 stimulates adipocyte differentiation incultures of fetal rat calvaria cells: Comparison with the effects ofdexamethasone. Endocrinology, 134:2221–2229.
Beresford, J.N., Bennett, J.H., Devlin, C., Leboy, P.S., and Owen, M.E.(1992) Evidence for an inverse relationship between the differentia-tion of adipocytic and osteogenic cells in rat marrow stromal cellcultures. J. Cell Sci., 102:341–351.
Bland, J.M., and Altman, D.G. (1995) Multiple significance tests: TheBonferroni method. Br. Med. J., 310:170.
Bonner, W.M., and Laskey, R.A. (1974) A film detection method fortritium-labeled proteins and nucleic acids in polyacrylamide gels.Eur. J. Biochem., 46:83–88.
Bruckner, P., Horler, I., Mendler, M., Houze, Y., Winterhalter, K.H.,Eich-Bender, S.G., and Spycher, M.A. (1989) Induction and preven-tion of chondrocyte hypertrophy in culture. J. Cell Biol., 109:2537–2545.
Buckwalter, J.A., Rosenberg, L.C., and Ungar, R. (1987) Changes inproteoglycan aggregates during cartilage mineralization. Calcif.Tissue Int., 41:228–236.
Cole, A.A., Luchene, L.J., Linsenmayer, T.F., and Schmid, T.M. (1992)The influence of bone and marrow on cartilage hypertrophy anddegradation during 30-day serum-free culture of the embryonicchick tibia. Dev. Dyn., 193:277–285.
Derynck, R. (1994) Transforming growth factor-b. In: Cytokine Hand-book. 2nd ed. A.W. Thomson, ed. Academic Press, London and SanDiego, pp. 319–342.
Eavey, R.D., Schmid, T.M., and Linsenmayer, T.F. (1988) Intrinsic andextrinsic controls of the hypertrophic program of chondrocytes in theavian columella. Dev. Biol., 126:57–62.
Farnum, C.E., and Wilsman, N.J. (1987) Morphologic stages of theterminal hypertrophic chondrocyte of growth plate cartilage. Anat.Rec., 219:221–232.
Franceschi, R.T. (1992) The role of ascorbic acid in mesenchymaldifferentiation. Nutr. Rev., 50:65–70.
Franceschi, R.T., Iyer, B.S., and Cui, Y. (1994) Effects of ascorbic acidon collagen matrix formation and osteoblast differentiation inmurine MC3T3-E1 cells. J. Bone Miner. Res., 9:843–854.
Gawande, S.R., and Tuan, R.S. (1990) Action of 1,25(OH)2 vitamin D3on calcium metabolism and skeletal development of the developingchick embryo. Trans. Ortho. Res. Soc. 15:401.
Gerstenfeld, L.C. and Shapiro, F.D. (1996) Expression of bone-specificgenes by hypertrophic chondrocytes: Implications of the complexfunctions of the hypertrophic chondrocyte during endochondral bonedevelopment. J. Cell. Biochem., 62:1–9.
Gerstenfeld, L.C., Crawford, D.R., Boedtker, H., and Doty, P. (1984)Expression of type I and III collagen genes during differentiation ofembryonic chicken myoblasts in culture. Mol. Cell. Biol., 4:1483–1492.
Gerstenfeld, L.C., Chipman, S.D., Glowacki, J., and Lian, J.B. (1987)Expression of differentiated function by mineralizing cultures ofchicken osteoblasts. Dev. Biol., 122:49–60.
Gerstenfeld, L.C., Chipman, S.D., Kelly, C.M., Hodgens, K.J., Lee,D.D., and Landis, W.J. (1988) Collagen expression, ultrastructuralassembly, and mineralization in cultures of chicken embryo osteo-blasts. J. Cell Biol., 106:979–989.
Gerstenfeld, L.C., Finer, M.H., and Boedtker, H. (1989) Quantitativeanalysis of collagen expression in embryonic chick chondrocyteshaving different developmental fates. J. Biol. Chem., 264:5112–5120.
Gerstenfeld, L.C., Kelley, C.M., Von Deck, M., and Lian, J.B. (1990)Effect of 1,25-dihydroxyvitamin D3 on induction of chondrocytematuration in culture: Extracellular matrix gene expression andmorphology. Endocrinology, 126:1599–1609.
Gerstenfeld, L.C., Zurakowski, D., Schaffer, J.L., Nichols, D.P., Toma,C.D., Broess, M., Bruder, S.P., and Caplan, A.I. (1996) Variablehormone responsiveness of osteoblast populations isolated at differ-ent stages of embryogenesis and its relationship to the osteogeniclineage. Endocrinology, 137:3957–3968.
Glimcher, M.J. (1992) The nature of the mineral component of boneand the mechanism of calcification. In: Disorders of Bone and
Mineral Metabolism. Coe, F.L. and Favus, M.J., eds. Raven Press,Ltd., New York, pp. 265–286.
Grigoriadis, A.E., Heersche, J.N.M., and Aubin, J.E. (1988) Differentia-tion of muscle, fat, cartilage, and bone from progenitor cells presentin a bone-derived clonal cell population: Effect of dexamethasone. J.Cell Biol., 106:2139–2151.
Ham, A.W., and Cormack, D.H. (1979) Histology. 8th ed. J.B. Lippin-cott Company, Philadelphia and Toronto, pp. 377–462.
Holtrop, M.E. (1972) The ultrastructure of the epiphyseal plate. II:The hypertrophic chondrocyte. Calcif. Tissue Res., 9:140–151.
Hunziker, E.B., Herrmann, W., Schenk, R.K., Mueller, M., and Moor,H. (1984) Cartilage ultrastructure after high pressure freezing,freeze substitution, and low temperature embedding. I: Chondro-cyte ultrastructure—implications for the theories of mineralizationand vascular invasion. J. Cell Biol., 98:267–276.
Ishizaki, Y., Burne, J.F., and Raff, M.C. (1994) Autocrine signalsenable chondrocytes to survive in culture. J. Cell Biol., 126:1069–1077.
Jacenko O., and Tuan, R.S. (1986) Calcium deficiency induces expres-sion of cartilage-like phenotype in chick embryonic calvaria. Dev.Biol., 115:215–232.
Kodama, H.A., Amagai, Y., Koyama, H., and Kasai, S. (1982) Hor-monal responsiveness of a preadipose cell line derived from newbornmouse calvaria. J. Cell. Physiol., 112:83–88.
Laemmli, U.K. (1970) Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature, 227:680–685.
Landis, W.J., and Glimcher, M.J. (1978) Electron diffraction andelectron probe microanalysis of the mineral phase of bone tissueprepared by anhydrous techniques. J. Ultrastruct. Res., 63:188–223.
Landis, W.J., and Glimcher, M.J. (1982) Electron optical and analyti-cal observations of rat growth plate cartilage prepared by ultracryo-microtomy: The failure to detect a mineral phase in matrix vesiclesand the identification of heterodispersed particles as the initial solidphase of calcium phosphate deposited in the extracellular matrix. J.Ultrastruct. Res., 78:227–268.
Landis, W.J., Paine, M.C., and Glimcher, M.J. (1977) Electron micro-scopic observations of bone tissue prepared anhydrously in organicsolvents. J. Ultrastruct. Res., 59:1–30.
Leboy, P.S., Vaias, L., Uschmann, B., Golub, E., Adams, S.L., andPacifici, M. (1989) Ascorbic acid induces alkaline phosphatase, typeX collagen, and calcium deposition in cultured chick chondrocytes. J.Biol. Chem., 264:17281–17286.
Lehrach, H., Frischauf, A.M., Hanahan, D., Wozney, J., Fuller, F., andBoedtker, H. (1979) Construction and characterization of pro a1collagen complementary deoxyribonucleic acid clones. Biochem.,18:3146–3152.
LuValle, P., Ninomiya, Y., Rosenblum, N.D., and Olsen, B.R. (1988)The type X collagen gene. Intron sequences split the 5’-untranslatedregion and separate the coding regions for the non-collagenousamino-terminal and triple-helical domains. J. Biol. Chem., 263:18378–18385.
Lyons, B.L., and Schwarz, R.I. (1984) Ascorbate stimulation of PATcells causes an increase in transcription rates and a decrease indegradation rates of procollagen mRNA. NucleicAcids Res., 12:2569–2579.
Moore, M.A., Gotoh, Y., Rafidi, K., and Gerstenfeld, L.C. (1991)Characterization of a cDNA for chicken osteopontin: Expressionduring bone development, osteoblast differentiation, and tissuedistribution. Biochemistry, 30:2501–2508.
Neugebauer, B.M., Moore, M.A., Broess, M., Gerstenfeld, L.C., andHauschka, P.V. (1995) Characterization of structural sequences inthe chicken osteocalcin gene: Expression of osteocalcin by maturingosteoblasts and by hypertrophic chondrocytes in vitro. J. BoneMiner. Res., 10:157–163.
Osdoby, P., and Caplan, A.I. (1979) Osteogenesis in cultures of limbmesenchymal cells. Dev. Biol., 73:84–102.
Probst, A., and Spiegel, H.U. (1997) Cellular mechanisms of bonerepair. J. Invest. Surg., 10:77–86.
Quarto, R., Campanile, G., Cancedda, R., and Dozin, B. (1992) Thyroidhormone, insulin, and glucocorticoids are sufficient to supportchondrocyte differentiation to hypertrophy: A serum-free analysis.J. Cell Biol., 119:989–995.
Rifas, L., Uitto, J., Memoli, V.A., Kuettner, K.E., Henry, R.W., andPeck, W.A. (1982) Selective emergence of differentiated chondro-cytes during serum-free culture of cells derived from fetal ratcalvaria. J. Cell Biol., 92:493–504.
Rousseau, F., Bonaventure, J., Legeai-Mallet, L., Pelet, A., Rozet, J.M.,Maroteaux, P., Le Merrer, M., and Munnich, A. (1994) Mutations inthe gene encoding fibroblast growth factor receptor-3 in achondropla-sia. Nature, 371:252–254.
172 L.C. GERSTENFELD ET AL.

Sai, S., Tanaka, T., Kosher, R.A., and Tanzer, M.L. (1986) Cloning andsequence analysis of a partial cDNA for chicken cartilage proteogly-can core protein. Proc. Natl. Acad. Sci., U.S.A., 83:5081–5085.
Sandell, L.J., and Daniel, J.C. (1988) Effects of ascorbic acid oncollagen mRNA levels in short term chondrocyte cultures. Connect.Tissue Res., 17:11–22.
Schmid T.M., and Linsenmayer, T.F. (1985) Developmental acquisitionof type X collagen in the embryonic chick tibiotarsus. Dev. Biol.,107:373–381.
Shapiro, F. (1992) Vertebral development of the chick embryo duringdays 3–19 of incubation. J. Morphol., 213:317–333.
Shapiro, F., Holtrop, M.E., and Glimcher, M.J. (1977) Organizationand cellular biology of the perichondrial ossification groove ofRanvier: A morphological study in rabbits. J. Bone Joint Surg.,59A:703–723.
Solursh, M., and Meier, S. (1974) Effects of cell density on theexpression of differentiation by chick embryo chondrocytes. J. Exp.Zool., 187:311–322.
Terashima, Y., and Urist, M.R. (1975) Differentiation of cartilage fromcalvarial bone under the influence of bone matrix gelatin in vitro.Clin. Orthop. Rel. Res., 113:168–177.
Thomas, PS. (1980) Hybridization of denatured RNA and small DNA
fragments transferred to nitrocellulose. Proc. Natl. Acad. Sci.,U.S.A., 77:5201–5205.
Thorogood, P. (1979) In vitro studies on skeletogenic potential ofmembrane bone periosteal cells. J. Embryol. Exp. Morphol., 54:185–207.
Toma, C., Schaffer, J., Meazzini, M.C., Zurakowski, D., Nah, H.D., andGerstenfeld, L.C. (1997) Developmental restriction of embryonicmesenchymal stem cells as characterized by their in vitro potentialfor differentiation. J. Bone Miner. Res., 12:2024–2039.
Wong, M., and Tuan, R.S. (1993) Nuserum, a synthetic serum replace-ment, supports chondrogenesis of embryonic chick limb bud mesen-chymal cells in micromass culture. In Vitro Cell. Dev. Biol., Animal,29A:917–922.
Wong, M., and Tuan, R.S. (1995) Interactive cellular modulation ofchondrogenic differentiation in vitro by subpopulations of chickembryonic calvarial cells. Dev. Biol., 167:130–147.
Yamaguchi, A., and Kahn, A.J. (1991) Clonal osteogenic cell linesexpress myogenic and adipocytic developmental potential. Calcif.Tissue Int., 49:221–225.
Young, M.F., Vogeli, G., Nunez, A.M., Fernandez, M.P., Sullivan, M.,and Sobel, M.E. (1984) Isolation of cDNA and genomic DNA clonesencoding type II collagen. Nucleic Acids Res., 12:4207–4228.
173CHONDROGENIC POTENTIAL OF SKELETAL CELLS