biomimetic materials in tissue engineering
TRANSCRIPT

ISSN:1369 7021 © Elsevier Ltd 2010JAN–FEB 2010 | VOLUME 13 | NUMBER 1-214
Biomimetic materials in tissue engineering
Cells receive numerous signals from their immediate
microenvironment, the extracellular matrix (ECM)1. Within a
biomechanical context provided by this elastic milieu2, cells
adhere by receptor-mediated interactions with ECM components
such as fibronectin and laminin (among many others, as reviewed
elsewhere), mediated by specialized adhesion receptors such as
integrins and others3. These receptors transmit stress from the
ECM, through the membrane (the receptors are transmembrane
proteins), to the cytoskeleton within the cell in a dynamic and
concerted manner4. The adhesion receptors do much more
than transmit stress, however; in particular within clusters
of adhesion receptors in the membrane, biochemical signal
transduction takes place through kinase activation and other
mechanisms3,4. In addition to adhesion proteins, the ECM
also sequesters and presents a number of morphoregulatory
molecules including growth factors, which control processes of
cell division, differentiation, and multicellular morphogenesis2,5.
The growth factors bind ECM components such as heparan sulfate
proteoglycans6 and fibronectin7, among others, and reside there
until released by enzymatic processes or dissociation. These
Biomaterial matrices are being developed that mimic the key characteristics of the extracellular matrix, including presenting adhesion sites and displaying growth factors in the context of a viscoelastic hydrogel. This review focuses on two classes of materials: those that are derived from naturally occurring molecules and those that recapitulate key motifs of biomolecules within biologically active synthetic materials. For biologically derived materials, methods are being sought to gain molecular-level control over biological characteristics and biomechanics. For synthetic, biomimetic materials, chemical schemes are being developed to enable in situ cross-linking and protease-dependent degradation and release of incorporated growth factors. These materials will open new doors to biosurgical therapeutics in tissue engineering and regenerative medicine.
Jennifer Patterson1,+, Mikaël M. Martino1,+, and Jeffrey A. Hubbell1,2*1Institute of Bioengineering, Ecole Polytechnique Fédérale de Lausanne (EPFL), Lausanne, Switzerland 2Institute of Chemical Sciences and Engineering, Ecole Polytechnique Fédérale de Lausanne (EPFL), Lausanne, Switzerland
*E-mail [email protected]+These authors contributed equally.
MT131-2p14_22.indd 14 01/02/2010 10:35:43

Biomimetic materials in tissue engineering REVIEW
JAN–FEB 2010 | VOLUME 13 | NUMBER 1-2 15
factors, when released and sometimes also when matrix-bound8,
bind to cell-surface receptors and trigger signaling, principally
through kinase activation. Thus, the ECM serves as a reservoir of
signaling molecules, both adhesion molecules and growth factors,
to instruct cell decision processes.
These characteristics of the ECM guide the design of biomaterials
for use in tissue engineering and regenerative medicine, with the
overall goal of mimicking these key features: the presentation of
adhesion molecules and the sequestration, display and release of
growth factors. These features may be accompanied by aspects of
formation and removal of the biomaterial ECM analogs. Namely, the
materials should be formable in situ, in the presence of cells without
damaging them and biomolecules without altering them. Further,
cells should be able to exert their enzymatic processes to remodel
and eventually replace the matrices, principally through matrix
metalloproteinases (MMPs)9, as they do with the natural ECM.
Thus, we organize this review from the perspective of biomimicry.
Having above begun, by very briefly outlining some of the biological
goals of biomimetic material design in tissue engineering, below we
proceed to review some of the biologically-derived materials that form
our design basis for biomimicry, following which we move to chemical
approaches that are being employed to actually mimic these materials.
Thus we proceed from biological, through biosynthetic, to completely
synthetic material systems.
Biologically derived and biologically produced materials CollagenCollagen type I, the most abundant protein in mammals, has a triple-
helical structure made of three polypeptide chains containing repeating
Gly-X-Y triplets in which the X and Y positions are frequently occupied
by proline and 4-hydroxyproline, respectively10, the latter of which
is very important for intermolecular hydrogen bonding. Collagen can
be readily purified from animal tissues, such as skin and tendon, and
from discarded human tissues, such as placenta. This ECM protein
can be reconstituted into a fibrillar matrix (Fig. 1a) by increasing the
Fig. 1 Scanning electron micrographs of biologically-derived and biologically-produced materials. (a) Fibrillar structure of collagen type I hydrogel. Reprinted with adaptation with permission from94. © 2007 Mary Ann Liebert, Inc. (b) Fibrin network. Reprinted with adaptation with permission from94. © 2007 Mary Ann Liebert, Inc. (c) Cross-linked adipic acid dihydrazide-modified hyaluronic acid hydrogel. Reprinted with adaptation with permission from95. © 2005 Elsevier. (d) Self assembling peptide RADA16-II hydrogel. Reprinted with adaptation with permission from96. © 2008 Wiley Periodicals, Inc. Scale bar = 500 nm.
(b)(a)
(c) (d)
MT131-2p14_22.indd 15 01/02/2010 10:35:46

REVIEW Biomimetic materials in tissue engineering
JAN–FEB 2010 | VOLUME 13 | NUMBER 1-216
pH and temperature of a precursor solution. Widely used, collagen
type I is frequently treated with proteases to remove small nonhelical
telopeptides that are present at the ends of the triple-helical domain
and that contribute to most of the protein’s cross-species immunogenic
character. Nevertheless, for clinical application of this material,
concerns of immunogenicity and disease transmission remain11. To
avoid these risks, methods for the recombinant expression of collagen
have been developed in several eukaryotic expression systems12, and
recombinant human collagen types I and III are commercially available.
Because in vivo application of collagen gels is limited by a deficiency
in mechanical strength, several procedures have been described in
order to form collagen matrices that possess sufficient mechanical
proprieties to at least partially resist cell-induced contraction. For
example, chemical glycation can adjust, in a rather controllable manner,
the elastic character of collagen gels13. Collagen hydrogels can be
enzymatically cross-linked14 or compressed to relatively high density15.
Heat and chemical treatments have also been developed to produce
cross-linked collagen sponges for bone and cartilage repair16, although
these materials become so dense that they are no longer really
hydrogels.
Interestingly, the dipole moment of fibrillar collagen gives rise to
the possibility to obtain permanent microscopic and even macroscopic
alignment of fibrillar collagen matrices. For instance, collagen fibril
alignment under a strong magnetic field has been demonstrated to
impart special characteristics for inducing directed cell migration, such as
neurites which grow preferentially in the direction of fibril alignment17.
Fig. 2 Improvement of biologically-derived and biologically-produced materials with biomolecules. (a) Fibrin matrices can be functionalized with recombinant ECM fragments or growth factors. (i-v) The factor XIIIa enzymatic substrate sequence NQEQVSPL is fused to bioactive peptides or recombinant proteins, allowing these molecules to be covalently incorporated during the fibrin’s natural polymerization process via factor XIIIa-induced transglutamination31,97. The molecules are engineered in a manner that depends on their biological function and local cell-induced proteolysis. (ii) For example, growth factor release by cell-demand can be accelerated by incorporating a protease sensitive sequence after the factor XIIIa enzymatic substrate sequence69. (iii) Because ECM peptides or fragments are mostly active when they are bound to the matrix, the molecule is designed without a protease sensitive sequence (PHSRN and RGD are the critical sequences from fibronectin for engagement of integrins)33. (iv) Since molecules such as heparin naturally bind certain growth factors, systems combining this interaction and transglutamination have been developed. For example, a heparin-binding peptide can be fused to the factor XIIIa enzymatic substrate and cross-linked into the matrix with affinity-bound growth factor. A complex between the three molecules is formed, and the growth factor release depends mostly on its affinity for heparin. (v) Fibrin matrices can also be functionalized noncovalently with growth factors fused to a fibrin binding sequence such as Gly-Pro-Arg-Pro (GPRP)29,30. (b) Hyaluronic acid derivatives can be functionalized with biomolecules such as protease sensitive sequences or ECM fragments. For example, a recombinant ECM fragment engineered with a single free cysteine is first reacted with divinylsulfone for further cross-linking into thiol modified hyaluronic acid hydrogel by Michael-type addition41. (c) Similarly to fibrin matrix, collagen matrix can be functionalized with a growth factor fused to a collagen binding sequence derived, for example, from fibronectin, von Willebrand factor (vWF) or collagenase. The growth factor release depends on the binding sequence affinity for collagen19-22. (d) Hydrogels based on self assembling peptides can be functionalized with biomolecules such as protease sensitive sequences or ECM fragments fused to the self assembling building block peptide (to both or to one extremity), as long as it does not disturb hydrogel self-assembly51-53,57.
(b) Hyaluronic acid
(a) Fibrin
(c) Collagen
(d) Self-assembling peptides
MT131-2p14_22.indd 16 01/02/2010 10:35:52

Biomimetic materials in tissue engineering REVIEW
JAN–FEB 2010 | VOLUME 13 | NUMBER 1-2 17
While collagens bind several cellular receptors that modulate cells’
behavior18, collagen gel bioactivity can be increased with growth
factors that have been engineered to bind the matrix in a noncovalent
manner. Collagen binding sequences derived from collagenase, von
Willebrand factor or fibronectin have been recombinantly fused
to growth factors in order to delay their release from collagen
scaffolds19-22 (Fig. 2c). For example, injection of collagen binding
vascular endothelial growth factor (VEGF) is able to improve cardiac
function after an acute myocardial infarction23.
FibrinClinically available from autologous sources and from cryoprecipitated
pooled human blood plasma, fibrin is a specialized protein network that
is formed principally in spontaneous tissue repair. The fibrin matrix
forms spontaneously by polymerization of fibrinogen, a circulating
glycoprotein homodimer of a heterotrimer, in the presence of thrombin
protease. Thrombin cleaves the so-called fibrinopeptides on fibrinogen
that prevent physicochemical self-assembly or polymerization of the
molecule. The resulting network is chemically cross-linked by the
blood transglutaminase factor XIIIa24, and its complex fibril structure
(Fig. 1b) and cross-linked character depend upon the details of its
formation25-27. Although fibrin is not an ECM in the usual sense,
since cells in the local environment do not produce it, the material is
nevertheless a crucial member of the body’s repertoire of matrices and
serves the role of a provisional matrix, being remodeled and replaced
with ECM molecules.
In contrast to fibrillar collagen matrices where cell migration
proceeds both through mechanisms that are dependent and
independent of proteolytic degradation, cell migration in fibrin is
almost exclusively dependent upon cell-associated proteolytic activity
(e.g., from plasmin and MMPs24). This distinction in cellular behavior
from fibrillar collagen probably results from the smaller mesh size of
the fibrin matrices and the stronger fibril–fibril interactions, owing to
the nature of network formation and covalent stabilization.
Several proteins are naturally incorporated into fibrin matrix
during coagulation, such as fibronectin and alpha-2-plasmin inhibitor,
which are covalently cross-linked into the matrix by factor XIIIa24.
Other biomolecules, such as fibroblast growth factor-2 (FGF-2) bind
noncovalently to fibrin and are able to provide growth factor-specific
bioactivity, such as potentiating of endothelial cell proliferation,
even when bound28. Nevertheless, fibrin matrices remain naturally
poorly active for most cell types, leading to their functionalization
with ECM peptides, recombinant ECM protein domains or growth
factors. Similarly to collagen matrices as described above, engineered
growth factors or ECM fragments can be incorporated into fibrin
non-covalently by fusing a fibrin-binding domain to the recombinant
protein29. For example, the native knob:pocket interactions during
fibrin assembly can be exploited to delay the release of recombinant
proteins30. However, to have even better incorporation and controlled
release, biomolecules can be covalently incorporated. In a powerful
approach, the factor XIIIa enzymatic substrate sequence of alpha-2-
plasmin inhibitor, Asn-Gln-Glu-Gln-Val-Ser-Pro-Leu (NQEQVSPL), can
be fused to bioactive peptides or recombinant proteins, allowing these
molecules to be covalently incorporated during the fibrin’s natural
polymerization process via factor XIIIa-induced cross-linking31. For
example, VEGF, fibronectin fragments and parathyroid hormone 1-34
(PTH1-34) have been engineered and covalently incorporated to be
released in a manner that depends on local cell-induced proteolysis32-
34 (Fig. 2a). In the case of the PTH1-34 fusion, the hormone is active
only when released: modification of its N-terminus with the factor XIIIa
substrate fusion inhibits the activity of the hormone and this activity
is regained after cell-induced proteolysis between the fusion and the
PTH1-34 domain. This engineered peptide in fibrin is currently in human
clinical evaluation for local bone repair.
GlycosaminoglycansThe structural proteins of the ECM are augmented in their
biomechanical and biochemical functions by long unbranched
polysaccharides, the glycosaminoglycans. In most cases, these are
components of proteoglycans of the ECM, except in the case of
hyaluronic acid (also called hyaluronan), which is not covalently
attached to a protein core and is entangled within the extracellular
space. These strongly anionic polymers absorb water, which provides
compressive strength to the ECM, while the glycosaminoglycans also
directly affect tissue organization via interactions with cell-surface
receptors35.
Hyaluronic acid can be isolated from animal tissue, such as
the rooster comb, and can be biotechnologically produced using
Streptococcus bacterium. Because the material absorbs enormous
amounts of water at equilibrium, it forms a hydrogel that is non-
fibrillar (Fig. 1c), and owing to entanglement associated with its high
molecular weight, up to several million Daltons, the gel dissolves only
very slowly.
Several chemical hyaluronic acid derivatives have been prepared
and by controlling the functional group (e.g., pendant hydrophobic
groups), the type of covalent bond (e.g., stable or hydrolytically-
sensitive), and the cross-linking density, it is possible to create a wide
range of physically diverse materials. A number of modifications of the
carboxyl and hydroxyl groups of hyaluronic acid have been developed,
to crosslink the material into an elastic gel that resists dissolution or
rendering the polymer controllably more hydrophobic and thus less
soluble36.
Hyaluronic acid hydrogels have been used for various applications,
including keratinocyte transfer for dermal wound healing and chondrocyte
transplantation for cartilage repair37,38. While hyaluronic acid interacts
with at least three cell surface receptors (CD44, RHAMM, and ICAM-1)39,
its biological activity can be significantly augmented by the incorporation
of other functional biomolecules. For example, hyaluronic acid gels have
MT131-2p14_22.indd 17 01/02/2010 10:35:54

REVIEW Biomimetic materials in tissue engineering
JAN–FEB 2010 | VOLUME 13 | NUMBER 1-218
been functionalized with peptides or protein fragments derived from
fibronectin to improve fibroblast proliferation and wound healing40,41
(Fig. 2b). Similarly, cellular infiltration into the hydrogel can be
improved by using MMP-sensitive cross-linkers, since hyaluronic acid gel
degradation is principally due to hyaluronidase42 whereas cell migration is
driven principally by the activities of various MMPs.
Self-assembling polypeptides Inspired by the understanding of protein self-assembly, important
progress has been made using supramolecular self-assembly of
biomolecules to form nanofibrillar matrices in situ. These approaches
use non-covalent intermolecular interactions to fabricate higher order
structures by self-assembly of oligomeric peptide, nucleotide and non-
biological amphiphilic building blocks43.
Whereas many of these systems require nonphysiologic conditions
for self-assembly, several can gel at cell-tolerable conditions. Zhang
and coworkers developed a class of nanofibrillar gels with very high
water content (> 99%) cross-linked by assembly of self-complementary
amphiphilic peptides in physiological medium44 (Fig. 1d). These ionic
self-complementary peptides are characterized by a periodic repetition
of alternating ionic hydrophilic and hydrophobic amino acids. Upon
exposure to aqueous solutions with neutral pH, they form stable
β-strand and β-sheet structures, partitioning the side-chains to two sides,
one polar and the other nonpolar. They self-assemble to form nanofibers
with the nonpolar residues inside and positively and negatively charged
residues forming complementary ionic interactions, like a checkerboard.
A number of similar scaffolds have been reported and customized to
deliver growth factors and cells43,45. For example, gels of RAD self-
assembling peptide have been used to encapsulate several growth factors
to accelerate dermal wound healing and myocardial repair46-48.
Self-assembled peptide hydrogels are also being developed as
a tool for three-dimensional cell culture. For example, cell types
such as chondrocytes49 and neural stem cells50 have been cultured
within these scaffolds. However, although these gels biomechanically
organize cells in a three-dimensional fashion, they show no specific
cell interaction, because they are not equipped with any specific
biofunctional ligands. To engineer receptor-mediated biospecificity,
active and functional motifs from the ECM have been employed to
significantly enhance their interactions with cells and tissues. Stupp and
coworkers have designed self-assembling oligomeric-amphiphiles that
allow incorporation of specific biomolecular signals without inhibiting
the self-assembling properties and nanofiber formation. Using
scaffolds presenting the laminin-derived peptide Ile-Lys-Val-Ala-Val,
encapsulated neural progenitor cells were observed to differentiate into
neurons51. Following this trend, a variety of self-assembling peptides
with different functional motifs such as cell adhesion sites or protease
sensitive sequences have been produced50,52 (Fig. 2d).
Another class of polypeptides that forms hydrogels is derived
from the Val-Pro-Gly-X-Gly pentapeptide repeat (where X is a guest
amino acid other than proline) found in human tropoelastin. Called
elastin-like-polypeptides (ELPs), the chains are soluble in aqueous
solution, but as the solution temperature is raised, ELPs become
insoluble and aggregate at a critical temperature, termed the inverse
transition temperature53. By changing the composition and chain
length of guest residue(s), the transition temperature can be precisely
tuned between 0-100°C for specific applications such as recombinant
protein purification54 or cell culture55. For example, ELPs have been
demonstrated to promote the synthesis and retention of cartilaginous
matrix from encapsulated chondrocytes and adult stem cells when
cultured in vitro56. Similarly to self-assembling peptides, ELPs have also
been modified with ECM ligands derived from fibronectin, in order to
promote better cell attachment57.
Synthetic materials to mimic biological functionalityThe biological materials described above serve as a point of departure
for biomimicry, the use of synthetic materials to recapitulate salient
materials features of natural ECM molecules, such as in situ cross-
linking, presentation of adhesion ligands, binding of growth factors,
and susceptibility to cell-derived proteases. Synthetic analogs can
offer several advantages. The chemistries used for matrix formation
and functionalization are becoming increasingly straightforward and
easy to control. Many reactions can be performed under gentle, often
physiological, conditions that allow incorporation of cells or biological
molecules with little loss of viability or function, respectively. The use
of entirely synthetic materials eliminates the purification problems
that can occur with naturally derived materials as well as reduces
the potential for an immune response or pathogen transmission.
Finally, polymeric-based hydrogels, which represent a large class of
synthetic materials used for this application, mimic the highly hydrated
viscoelastic properties of the natural ECM, allow for transport by
diffusion and interstitial flow, and can present soluble, affinity-bound,
or covalently bound biological factors.
Synthetic cell-responsive hydrogels for use in tissue engineering
can be formed from a number of hydrophilic synthetic polymers or
polysaccharides, including poly(ethylene glycol) (PEG), poly(vinyl
alcohol) (PVA), poly(hydroxyethyl methacrylate) (poly(HEMA)),
alginate derivatives, and others, and have been reviewed extensively
in the literature58-61. Here, we focus on the mechanisms of hydrogel
formation, the engineering of biological functionality into hydrogel
constructs, and the presentation of multiple signals in a temporally
dynamic and/or spatially patterned manner.
Mechanisms of hydrogel formationPolymers can be covalently cross-linked into hydrogel networks by
several mechanisms, which can be broadly grouped into chain-growth
polymerizations, such as photopolymerization, and step-growth
polymerizations, such as Michael-addition reactions. Examples of
MT131-2p14_22.indd 18 01/02/2010 10:35:54

Biomimetic materials in tissue engineering REVIEW
JAN–FEB 2010 | VOLUME 13 | NUMBER 1-2 19
photopolymerization include photo-initiated reactions between PEG
diacrylate (PEG-DA) molecules62 (Fig 3a) or between thiol-acrylates63
or thiol-enes64. Lutolf and Hubbell developed vinyl sulfone modified
multi-arm PEG macromers (PEG-VS) to allow a Michael addition
reaction between the acrylated PEG and a thiol group (Fig. 3b), which
is most often presented as a free cysteine on a peptide or protein65.
In this system, the PEG-VS is first functionalized with mono-cysteine
species, such as cell adhesion ligands, growth factor binding ligands,
or even growth factors themselves. The functionalized PEG-VS is then
cross-linked into a biodegradable gel network using peptides that
contain a protease-sensitive substrate sequence flanked by cysteine-
containing domains. Michael addition reactions are not limited to
acrylates and also occur between maleimide and thiol groups (Fig. 3c),
as has been shown for the crosslinking of a p-maleimidophenyl-
modified dextran66.
Recently, sequential copper-free click chemistry (Fig. 3d) has
been applied for the formation of hydrogels and their subsequent
patterning with biomolecules67. Specifically, an azide can react
with an alkyne to form a triazole using a di-fluorinated cyclooctyne
(DIFO3) moiety during hydrogel formation. Orthogonal thiol-ene
photocoupling, such as with peptides containing the photoreactive
allyl ester Fmoc-Lys(alloc)-OH, can then be used to pattern
biomolecules. Anseth’s group has used this click chemistry scheme
to prepare PEG hydrogels with MMP-cleavable sequences and
patterned incorporation of Arg-Gly-Asp (RGD) peptides to provide
localized cell attachment67. In addition to Michael addition reactions
and click chemistry, step-wise polymerizations can also be achieved
through enzyme-catalyzed crosslinking of peptide-functionalized
materials. Sperinde and Griffith first used transglutaminase to cross-
link multifunctional glutaminyl-PEG with polypeptides containing
alternating lysine and phenylalanine residues68. Alternatively, a
mixture of multi-arm PEG molecules, each conjugated with one of
two different counter-reactive peptide substrates for factor XIIIa,
can be crosslinked in the presence of this transglutaminase69 (Fig
3e). Similar to the approach used to covalently functionalize fibrin
matrices, biofunctional peptides tagged with one of these factor XIIIa
substrates can also be incorporated into PEG hydrogels in a simple “one
pot” reaction. Increasingly sophisticated means have been developed
to provide triggered activation of transglutaminase to initiate the
crosslinking process in situ, both for peptide-grafted polymers70 and
for alginates or fibrinogen71. In these systems, thermally triggered
release of calcium from phospholipid vesicles was used to activate the
Fig. 3 (a) Cyclic RGD-modified PEG-DA macromer for photopolymerization, after62 (single letter amino acid nomenclature). (b) Michael addition macromers for reaction between a vinyl sulfone group and a thiol, after79. (i) Multi-arm PEG vinyl sulfone (PEG-VS). (ii) Peptide crosslinker containing free thiols from cysteine residues. (c) Michael addition macromers for reaction between maleimide and thiol groups, after66. (i) p-maleimidophenyl isocyanate (PMPI) conjugated dextran. (ii) Peptide crosslinker containing free thiols from cysteine residues. (d) Functionalized macromers for click chemistry reactions, after67. (i) PEG tetra-azide. (ii) Bis(DIFO3) di-functionalized peptide crosslinker. (e) Peptide functionalized macromers for transglutaminase catalyzed crosslinking, after69. Reactive functionality is either FKGG or NQEQVSPL peptide tag (single letter amino acid nomenclature). (f) Silk-inspired multiblock copolymers that self-assemble, after72. “(g) Photolabile cross-linking macromer for the formation of photodegradable hydrogels, after93.
(b)(a) (c)
(g)
(e)(d) (f)
i i
i
ii ii
ii
MT131-2p14_22.indd 19 01/02/2010 10:35:54
REVIEW Biomimetic materials in tissue engineering
JAN–FEB 2010 | VOLUME 13 | NUMBER 1-220
factor XIII (which is calcium ion-dependent) and initiate rapid hydrogel
formation.
In addition to the covalent crosslinking mechanisms described
above, hydrogels can also be formed by physical or ionic interactions
between molecules. This behavior is observed in the self-assembly of
peptide amphiphiles into fibrillar β-sheet structures, as described above,
and during the complexation of polymers or polysaccharides with ions.
Taking inspiration from nature, hybrid hydrogels have been developed by
replacing the amorphous peptide domain of N. calvipes silk with PEG72.
Poly(Ala) self-assembling domains, as in the silk, have been combined
with short-chain PEG segments (Fig. 3f) to form self-assembling hydrogels
with good mechanical properties and retention of the natural β-sheet
structure72. Complexation via the divalent cation calcium leads to
hydrogel formation for alginate or modified alginate polysaccharides73-
75. Alginate is composed of D-mannuronic acid and L-guluronic acid in
a multiblock copolymer form, the guluronic acid units of which allow
for crosslinking by the formation of an “egg-box” conformation upon
the complexation with calcium76. Altering the bulk composition or
distribution of mannuronic and guluronic acid units as well as the overall
molecular weight of the alginate affects the final hydrogel properties75,77.
Engineering biological functionality in synthetic hydrogelsPeptide-conjugated polymers provide a facile means for the
presentation of ECM-derived biomolecular signals. Extensive work has
distilled key elements of proteins, such as cell adhesion sites, protein
binding sites, or protease substrate sites, down to short peptide
sequences that display similar functionality. These peptides can be
conjugated to polymer networks using the chemistries described above,
or incorporated using other strategies that do not interfere with the
reactive functional groups involved in network formation62, and have
been shown to impart biological activity to otherwise inert materials.
The RGD peptide sequence mentioned above, found in several ECM
proteins and known to bind the integrin family of cell-surface adhesion
receptors78, has frequently been used as a cell adhesion ligand and has
been shown to influence 3D fibroblast migration in functionalized PEG
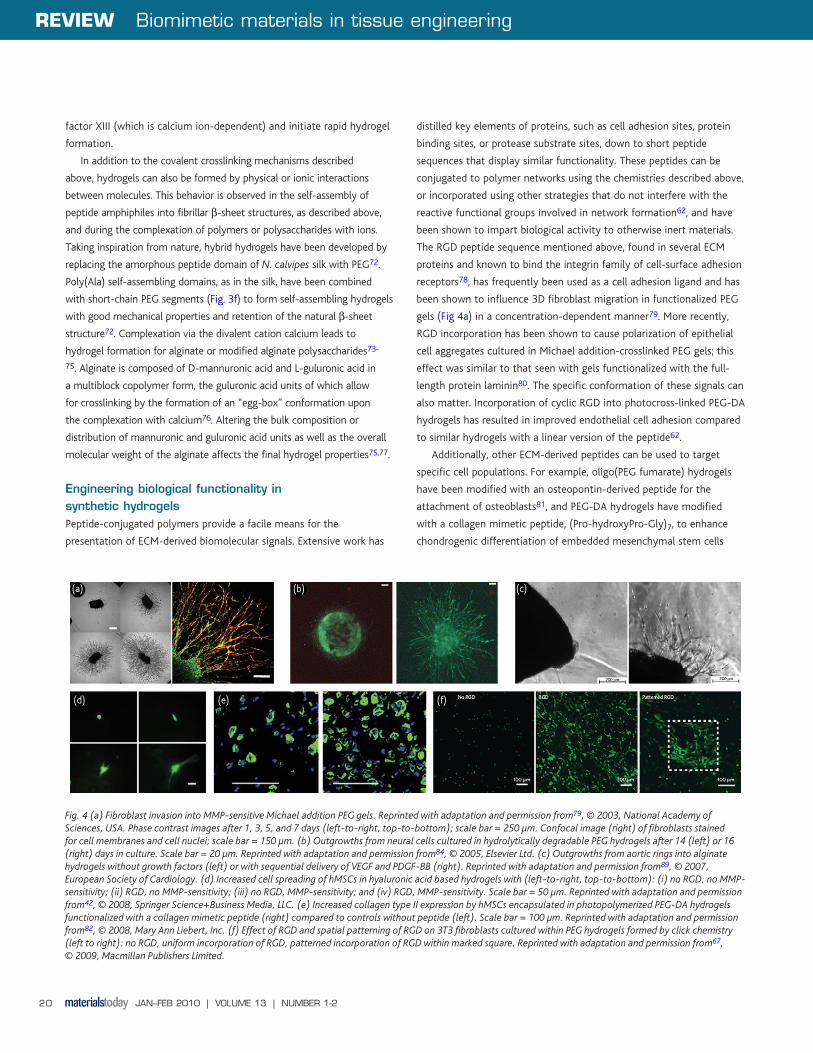
gels (Fig 4a) in a concentration-dependent manner79. More recently,
RGD incorporation has been shown to cause polarization of epithelial
cell aggregates cultured in Michael addition-crosslinked PEG gels; this
effect was similar to that seen with gels functionalized with the full-
length protein laminin80. The specific conformation of these signals can
also matter. Incorporation of cyclic RGD into photocross-linked PEG-DA
hydrogels has resulted in improved endothelial cell adhesion compared
to similar hydrogels with a linear version of the peptide62.
Additionally, other ECM-derived peptides can be used to target
specific cell populations. For example, oligo(PEG fumarate) hydrogels
have been modified with an osteopontin-derived peptide for the
attachment of osteoblasts81, and PEG-DA hydrogels have modified
with a collagen mimetic peptide, (Pro-hydroxyPro-Gly)7, to enhance
chondrogenic differentiation of embedded mesenchymal stem cells
Fig. 4 (a) Fibroblast invasion into MMP-sensitive Michael addition PEG gels. Reprinted with adaptation and permission from79, © 2003, National Academy of Sciences, USA. Phase contrast images after 1, 3, 5, and 7 days (left-to-right, top-to-bottom); scale bar = 250 μm. Confocal image (right) of fibroblasts stained for cell membranes and cell nuclei; scale bar = 150 μm. (b) Outgrowths from neural cells cultured in hydrolytically degradable PEG hydrogels after 14 (left) or 16 (right) days in culture. Scale bar = 20 μm. Reprinted with adaptation and permission from84, © 2005, Elsevier Ltd. (c) Outgrowths from aortic rings into alginate hydrogels without growth factors (left) or with sequential delivery of VEGF and PDGF-BB (right). Reprinted with adaptation and permission from89, © 2007, European Society of Cardiology. (d) Increased cell spreading of hMSCs in hyaluronic acid based hydrogels with (left-to-right, top-to-bottom): (i) no RGD, no MMP-sensitivity; (ii) RGD, no MMP-sensitivity; (iii) no RGD, MMP-sensitivity; and (iv) RGD, MMP-sensitivity. Scale bar = 50 μm. Reprinted with adaptation and permission from42, © 2008, Springer Science+Business Media, LLC. (e) Increased collagen type II expression by hMSCs encapsulated in photopolymerized PEG-DA hydrogels functionalized with a collagen mimetic peptide (right) compared to controls without peptide (left). Scale bar = 100 μm. Reprinted with adaptation and permission from82, © 2008, Mary Ann Liebert, Inc. (f) Effect of RGD and spatial patterning of RGD on 3T3 fibroblasts cultured within PEG hydrogels formed by click chemistry (left to right): no RGD, uniform incorporation of RGD, patterned incorporation of RGD within marked square. Reprinted with adaptation and permission from67, © 2009, Macmillan Publishers Limited.
(b)(a) (c)
(e)(d) (f)
MT131-2p14_22.indd 20 01/02/2010 10:35:55

Biomimetic materials in tissue engineering REVIEW
JAN–FEB 2010 | VOLUME 13 | NUMBER 1-2 21
(MSCs) (Fig. 4e) and collagen retention within the gels82. Further,
these cell adhesive materials have been used in vivo and shown to have
improved efficacy compared to non-biologically active materials. As
one example, this has been demonstrated in a rat myocardial infarct
model treated with alginate hydrogels where the gels modified with
RGD had the greatest angiogenic response83.
Another key parameter for the design of cell responsive synthetic
materials is their ability to degrade. In tissue engineering, the ultimate
goal is to have the implanted materials completely remodeled and
replaced by living tissue. Whether cells are incorporated into the
matrix before implantation or the scaffold simply provides signals
and physical support for host cells to invade, it is important that the
cells can have space to secrete their own ECM to eventually replace
the initial support provided by the hydrogel. Nonspecific hydrogel
degradation can be implemented by the incorporation of hydrolytically
degradable segments, such as poly(glycolic acid) (PGA) or poly(lactic
acid) (PLA) domains, within the hydrogel cross-links84 (Fig. 4b) or by
using hydrolytically degradable polymers, such as partially oxidized
alginate75, as the backbone of the matrix.
In vivo, ECM molecules are enzymatically, rather than hydrolytically,
degraded, with matrix degradation coupled to cell migration through
the expression of cell-associated or cell-secreted proteases. Thus, most
biologically derived hydrogels are naturally degradable, such as the
hyaluronic acid and fibrin hydrogels described above. Cell-mediated
control of degradation can be engineered into synthetic hydrogels
by the incorporation of protease substrate sequences42,52,66,79
(Fig. 4a, d). In these examples, peptide sequences derived from the
protease cleavage site in type I collagen85 or from combinatorial library
screening86 have been utilized to render covalently cross-linked42,66,79
or self-assembled52 hydrogels enzymatically degradable. Going one
step further, exogenous control of degradation can be achieved by
the incorporation of enzyme labile units into hydrogels. Degradation
of photocross-linked PEG gels containing caprolactone units within
the linkers occurred only in the presence of lipase, and the timing and
duration of lipase exposure affected the deposition and organization of
type II collagen by encapsulated chondrocytes in vitro87.
In addition to functionalities that allow cells to interact with the
hydrogel scaffolds, either by binding to or degrading the matrix,
truly biofunctionalized scaffolds will also provide signals necessary to
influence cell behavior. Thus, many groups seek to provide controlled
release of morphogens by physical entrapment, affinity-binding,
or covalent binding within hydrogels. The current state of the art
recognizes that delivery of a single growth factor may be insufficient to
recapitulate natural processes, and thus attention has turned to novel
delivery approaches that allow the release of multiple growth factors
at different doses and with different release rates88. In the area of
angiogenesis, where different growth factors stimulate vessel growth
and maturation, sequential delivery from hydrogel scaffolds has been
achieved by several different strategies. Affinity binding by alginate-
sulfate incorporated into alginate hydrogels resulted in different
delivery rates for bound VEGF, platelet-derived growth factor (PDGF)-
BB, and transforming growth factor (TGF)-β1 and led to increased
vessel density and maturity in scaffolds implanted subcutaneously74.
Improvements in cardiac function and neo-vessel maturity were also
seen with alginate hydrogels sequentially delivering VEGF and PDGF-BB
that were injected in the border zone of induced myocardial infarcts in
rats89, and sprouting was observed from aortic rings embedded in these
gels (Fig. 4c).
Spatially patterned or temporally dynamic hydrogels for presentation of multiple signalsIncreasingly complex hydrogel-based systems allow for presentation of
multiple signals, with the possibilities of spatial patterns of molecules
and/or temporal variation in exposure or release. Drawing from
microfluidics and microfabrication technologies, it is becoming possible
to engineer hydrogels down to the microscale90. Modular approaches
have been used to create in vitro engineered tissues, as seen with
the assembly of cell-embedded collagen microgels91, and processing
conditions such as tissue printing92 can be used to control the
architecture and spatial composition of cell-laden polymeric scaffolds.
The sequential click reactions described above67 also allow for localized
photopatterning of biological molecules after hydrogel formation
(Fig. 4f).
Using stimulus-sensitive linkers, protecting groups, or other
exposing mechanisms, it is possible to create temporally dynamic in
addition to spatially patterned matrices. One recent example comes
from Anseth’s group where gel properties can be manipulated in situ
by photodegradable segments (a nitrobenzyl ether-derived moiety)
incorporated within PEG hydrogels (Fig. 3g)93. The hydrogel itself
is cleaved, resulting in reduced local network cross-link density by
release of PEG; changes in stiffness, water content, and diffusivity;
and eventually complete erosion by release of poly(acrylate) chains.
To allow space for cell spreading or ECM production, the hydrogels
can be eroded to produce channels, leading to directed cell migration
or cell-cell connectivity, and three-dimensional features such as
interconnected channels can be produced by two-photon methodology.
Further, local modification of the chemical environment can be
achieved by the incorporation of photolabile tethered biologically
active functionalities, which was demonstrated to lead to enhanced
chondrogenic differentiation of encapsulated human MSCs upon
removal of RGD93.
SummaryIn the sections above, we have illustrated some of the goals and
challenges of creating biofunctionality in synthetic materials:
the biological materials from which the design concepts arise,
approaches to use those biological materials directly without and with
modification, and schemes by which to mimic the key features of the
MT131-2p14_22.indd 21 01/02/2010 10:35:57

REVIEW Biomimetic materials in tissue engineering
JAN–FEB 2010 | VOLUME 13 | NUMBER 1-222
natural ECM in completely synthetic implementations. Some of these
approaches are being evaluated in clinical trials and more are making
their way through preclinical investigation toward clinical studies.
These materials will enable new biosurgical approaches to tissue
engineering and regenerative medicine for repairing and replacing
damaged or diseased tissue, such as broken bones, acute and chronic
dermal wounds, damaged articular cartilage, and infarcted cardiac
muscle.
REFERENCES
1. Kleinman, H. K., et al., Curr Opin Biotechnol (2003) 14 (5), 526.
2. Discher, D. E., et al., Science (2009) 324, 1673.
3. Berrier, A. L., and Yamada, K. M., J Cell Physiol (2007) 213, 565.
4. Hinz, B., J Biomech (2009) 43 (1), 146.
5. Schultz, G. S., and Wysocki, A., Wound Repair Regen (2009) 17, 153.
6. Lindahl, U., and Li, J. P., Int Rev Cell Molec Biol (2009) 276, 105.
7. Wijeleath, E. S., et al., Circ Res (2006) 2006, 853.
8. Makarenkova, H. P., et al., Sci Signal (2009) 2, ra55.
9. Page-McCaw, A., et al., Nat Rev Mol Cell Biol (2007) 8, 221.
10. Kadler, K. E., et al., J Cell Sci (2007) 120 (Pt 12), 1955.
11. Lynn, A. K., et al., J Biomed Mater Res B (2004) 71 (2), 343.
12. Ruggiero, F., and Koch, M., Methods (2008) 45 (1), 75.
13. Girton, T. S., et al., J Biomech Eng (2000) 122 (3), 216.
14. Orban, J. M., et al., J Biomed Mater Res (2004) 68A, 756.
15. Abou Neel, E. A., et al., Soft Matter (2006) 2, 986.
16. Glowacki, J., and Mizuno, S., Biopolymers (2008) 89 (5), 338.
17. Ceballos, D., et al., Exp Neurol (1999) 158 (2), 290.
18. Leitinger, B., and Hohenester, E., Matrix Biol (2007) 26 (3), 146.
19. Nishi, N., et al., Proc Natl Acad Sci U S A (1998) 95 (12), 7018.
20. Andrades, J. A., et al., Exp Cell Res (1999) 250 (2), 485.
21. Ishikawa, T., et al., J Biochem (2001) 129 (4), 627.
22. Lin, H., et al., Biomaterials (2006) 27 (33), 5708.
23. Zhang, J., et al., Circulation (2009) 119 (13), 1776.
24. Mosesson, M. W., J Thromb Haemost (2005) 3 (8), 1894.
25. Lorand, L., and Graham, R. M., Nat Rev Mol Cell Biol (2003) 4 (2), 140.
26. Standeven, K. F., et al., Blood (2007) 110 (3), 902.
27. Weisel, J. W., Biophys Chem (2004) 112 (2-3), 267.
28. Sahni, A., et al., J Biol Chem (1998) 273 (13), 7554.
29. Zhao, W., et al., Tissue Eng Part A (2009) 15 (5), 991.
30. Soon, A. S., et al., Biomaterials (2009), .
31. Schense, J. C., and Hubbell, J. A., Bioconjug Chem (1999) 10 (1), 75.
32. Ehrbar, M., et al., J Control Release (2005) 101 (1-3), 93.
33. Martino, M. M., et al., Biomaterials (2009) 30 (6), 1089.
34. Arrighi, I., et al., Biomaterials (2009) 30, 1763.
35. Toole, B. P., Nat Rev Cancer (2004) 4 (7), 528.
36. Prestwich, G. D., and Kuo, J. W., Curr Pharm Biotechnol (2008) 9 (4), 242.
37. Price, R. D., et al., J Plast Reconstr Aesthet Surg (2007) 60 (10), 1110.
38. Tognana, E., et al., Cells Tiss Org (2007) 186 (2), 97.
39. Turley, E. A., et al., J Biol Chem (2002) 277 (7), 4589.
40. Park, Y. D., et al., Biomaterials (2003) 24 (6), 893.
41. Ghosh, K., et al., Tissue Eng (2006) 12 (3), 601.
42. Kim, J., et al., J Mater Sci: Mater Med (2008) 19, 3311.
43. Branco, M. C., and Schneider, J. P., Acta Biomaterialia (2009) 5 (3), 817.
44. Zhang, S., et al., Biomaterials (1995) 16 (18), 1385.
45. Gelain, F., et al., Macromol Biosci (2007) 7 (5), 544.
46. Schneider, A., et al., PLoS One (2008) 3 (1), e1410.
47. Segers, V. F., et al., Circulation (2007) 116 (15), 1683.
48. Hsieh, P. C., et al., Circulation (2006) 114 (7), 637.
49. Kisiday, J., et al., Proc Natl Acad Sci U S A (2002) 99 (15), 9996.
50. Gelain, F., et al., PLoS One (2006) 1, e119.
51. Silva, G. A., et al., Science (2004) 303 (5662), 1352.
52. Chau, Y., et al., Biomaterials (2008) 29, 1713.
53. Chilkoti, A., et al., Curr Opin Chem Biol (2006) 10 (6), 652.
54. Meyer, D. E., and Chilkoti, A., Nat Biotechnol (1999) 17 (11), 1112.
55. Betre, H., et al., Biomacromolecules (2002) 3 (5), 910.
56. Betre, H., et al., Biomaterials (2006) 27 (1), 91.
57. Liu, J. C., et al., Biomacromolecules (2004) 5 (2), 497.
58. Lutolf, M. P., and Hubbell, J. A., Nat Biotechnol (2005) 23 (1), 47.
59. Tibbitt, M. W., and Anseth, K. S., Biotechnol Bioeng (2009) 103 (4), 655.
60. Lin, C.-C., and Anseth, K. S., Pharm Res (2009) 26 (3), 631.
61. Jia, X., and Kiick, K. L., Macromolec Biosci (2009) 9, 140.
62. Zhu, J., et al., Bioconjugate Chem (2009) 20, 333.
63. Salinas, C. N., and Anseth, K. S., Macromolecules (2008) 41 (16), 6019.
64. Khire, V. S., et al., J Polym Sci Part A Polym Chem (2006) 44 (24), 7027.
65. Lutolf, M. P., and Hubbell, J. A., Biomacromolecules (2003) 4, 713.
66. Lévesque, S. G., and Shoichet, M. S., Bioconjugate Chem (2007) 18, 874.
67. DeForest, C. A., et al., Nat Mater (2009) 8, 659.
68. Sperinde, J. J., and Griffith, L. G., Macromolecules (1997) 30, 5255.
69. Ehrbar, M., et al., Biomaterials (2007) 28, 3856.
70. Sanborn, T. J., et al., Biomaterials (2002) 23, 2703.
71. Westhaus, E., and Messersmith, P. B., Biomaterials (2001) 22, 453.
72. Rathore, O., and Sogah, D. Y., J Am Chem Soc (2001) 123, 5231.
73. Hori, Y., et al., Acta Biomaterialia (2009) 5, 969.
74. Freeman, I., and Cohen, S., Biomaterials (2009) 30, 2122.
75. Boontheekul, T., et al., Biomaterials (2005) 26 (15), 2455.
76. Sikorski, P., et al., Biomacromolecules (2007) 8 (7), 2098.
77. Kong, H. J., et al., Biomacromolecules (2004) 5 (5), 358.
78. Ruoslahti, E., Annu Rev Cell Dev Biol (1996) 12, 697.
79. Lutolf, M. P., et al., Proc Nat Acad Sci USA (2003) 100 (9), 5413.
80. Chung, I.-M., et al., Biomaterials (2008) 29, 2637.
81. Shin, H., et al., Biomaterials (2004) 25, 895.
82. Lee, H. J., et al., Tissue Engineering: Part A (2008) 14 (11), 1843.
83. Yu, J., et al., Biomaterials (2009) 30, 751.
84. Mahoney, M. J., and Anseth, K. S., Biomaterials (2006) 27, 2265.
85. Nagase, H., and Fields, G. B., Biopolymers (Peptide Sci) (1996) 40, 399.
86. Turk, B. E., et al., Nat Biotechnol (2001) 19 (7), 661.
87. Rice, M. A., and Anseth, K. S., Tissue Eng (2007) 13 (4), 683.
88. Richardson, T. P., et al., Nat Biotechnol (2001) 19, 1029.
89. Hao, X., et al., Cardiovasc Res (2007) 75, 178.
90. Khademhosseini, A., and Langer, R., Biomaterials (2007) 28, 5087.
91. McGuigan, A. P., and Sefton, M. V., Proc Nat Acad Sci USA (2006) 103 (31), 11461.
92. Fedorovich, N. E., et al., Biomacromolecules (2009) 10, 1689.
93. Kloxin, A. M., et al., Science (2009) 324, 59.
94. Stegemann, J. P., et al., Tissue Eng (2007) 13 (11), 2601.
95. Hou, S., et al., J Neurosci Methods (2005) 148 (1), 60.
96. Sieminski, A. L., et al., J Biomed Mater Res A (2008) 87 (2), 494.
97. Zisch, A. H., et al., J Control Release (2001) 72 (1-3), 101.
MT131-2p14_22.indd 22 01/02/2010 10:35:57