` requirement for nuclear calcium signaling in drosophila ... · ` requirement for nuclear calcium...
TRANSCRIPT

(274), ra33. [DOI: 10.1126/scisignal.2003598] 6Science SignalingMartina Bujard, Christoph M. Schuster and Hilmar Bading (7 May 2013) Jan-Marek Weislogel, C. Peter Bengtson, Michaela K. Müller, Jan N. Hörtzsch,Memory
Long-TermDrosophilaRequirement for Nuclear Calcium Signaling in `
This information is current as of 8 May 2013. The following resources related to this article are available online at http://stke.sciencemag.org.
Article Tools http://stke.sciencemag.org/cgi/content/full/sigtrans;6/274/ra33
Visit the online version of this article to access the personalization and article tools:
MaterialsSupplemental
http://stke.sciencemag.org/cgi/content/full/sigtrans;6/274/ra33/DC1 "Supplementary Materials"
Related Content
http://stke.sciencemag.org/cgi/content/abstract/sigtrans;2006/356/re11 http://stke.sciencemag.org/cgi/content/abstract/sigtrans;3/112/ra18 http://stke.sciencemag.org/cgi/content/abstract/sigtrans;5/237/pe34 http://stke.sciencemag.org/cgi/content/abstract/sigtrans;6/274/pc12
's sites:ScienceThe editors suggest related resources on
References http://stke.sciencemag.org/cgi/content/full/sigtrans;6/274/ra33#BIBL
1 article(s) hosted by HighWire Press; see: cited byThis article has been
http://stke.sciencemag.org/cgi/content/full/sigtrans;6/274/ra33#otherarticlesThis article cites 86 articles, 27 of which can be accessed for free:
Glossary http://stke.sciencemag.org/glossary/
Look up definitions for abbreviations and terms found in this article:
Permissions http://www.sciencemag.org/about/permissions.dtl
Obtain information about reproducing this article:
the American Association for the Advancement of Science; all rights reserved. byAssociation for the Advancement of Science, 1200 New York Avenue, NW, Washington, DC 20005. Copyright 2008
(ISSN 1937-9145) is published weekly, except the last week in December, by the AmericanScience Signaling
on May 8, 2013
stke.sciencemag.org
Dow
nloaded from

R E S E A R C H A R T I C L E
N E U R O S C I E N C E
Requirement for Nuclear Calcium Signaling inDrosophila Long-Term MemoryJan-Marek Weislogel,* C. Peter Bengtson, Michaela K. Müller, Jan N. Hörtzsch,Martina Bujard, Christoph M. Schuster, Hilmar Bading†
Dow
nload
Calcium is used throughout evolution as an intracellular signal transducer. In the mammalian central ner-vous system, calcium mediates the dialogue between the synapse and the nucleus that is required fortranscription-dependent persistent neuronal adaptations. A role for nuclear calcium signaling in similar pro-cesses in the invertebrate brain has yet to be investigated. Here, we show by in vivo calcium imaging ofadult brain neurons of the fruit fly Drosophila melanogaster, that electrical foot shocks used in olfactoryavoidance conditioning evoked transient increases in cytosolic and nuclear calcium concentrations in neu-rons. These calcium signals were detected in Kenyon cells of the flies’mushroom bodies, which are sites oflearning and memory related to smell. Acute blockade of nuclear calcium signaling during conditioningselectively and reversibly abolished the formation of long-term olfactory avoidance memory, whereasshort-term, middle-term, or anesthesia-resistant olfactory memory remained unaffected. Thus, nuclear cal-cium signaling is required in flies for the progression of memories from labile to transcription-dependentlong-lasting forms. These results identify nuclear calcium as an evolutionarily conserved signal needed inboth invertebrate and vertebrate brains for transcription-dependent memory consolidation.
ed fr
on May 8, 2013 stke.sciencem
ag.orgom
INTRODUCTION
One aspect common to most forms of persistent neuronal adaptations, in-cluding memory formation, addiction, and acquired neuroprotection (aprocess through which synaptic activity enhances the ability of neuronsto tolerate harmful conditions), is the requirement for a dialogue betweenthe synapse and the nucleus; this communication pathway is signal-inducedand controls specific genomic responses that are critical for functionalchanges to be long-lasting (1–7). One possible mechanism for achievingspecificity in the cellular responses is to dedicate specific signaling path-ways to particular cellular adaptations. Alternatively, if a diverse set of adap-tations has a common requirement, universal signaling mechanisms mayhave evolved that initiate appropriate genomic responses whenever adjust-ments need to be implemented long-term.
Studies of the mammalian nervous system suggest that calcium fulfillssuch a role as a universal signal transducer. Calcium acts locally near thesite of calcium signal generation, for example, at the synapse (8, 9), but itcan also propagate information toward and into the cell nucleus to couplesynaptic activity to genomic responses (10–14). Indeed, in mice, nuclearcalcium and one of its main targets, calcium/calmodulin-dependent pro-tein kinase IV (Ca2+/CaMKIV), have emerged as key players in the con-solidation and maintenance of memory, addictive behavior, developmentof chronic pain, and neuronal survival (14–22). For example, in hippocam-pal neurons, the establishment of a neuroprotective transcriptional profile,consisting of a defined set of prosurvival and antiapoptotic genes that pro-tect against a variety of otherwise toxic insults, requires the activation ofsynaptic N-methyl-D-aspartate (NMDA) receptors, nuclear calcium signal-ing, and CaMKIV activation of a transcriptional response mediated bythe complex of adenosine 3′,5′-monophosphate (cAMP) response element–binding protein (CREB) and CREB-binding protein (CBP), collectivelyreferred to as CREB/CBP (14, 22–24). Thus, a broad spectrum of per-
Department of Neurobiology, Interdisciplinary Centre for Neurosciences(IZN), University of Heidelberg, INF 364, 69120 Heidelberg, Germany.*Present address: A*STAR Neuroscience Research Partnership, 61 BiopolisDrive, #04-12 Proteos, Singapore 138673, Singapore.†Corresponding author. E-mail: [email protected]
sistent neuronal adaptations involves nuclear calcium signaling as a com-mon mediator.
To assess whether the role of nuclear calcium signaling in controllinglong-lasting neuronal adaptations is evolutionarily conserved and, hence,may also be used in transcription-dependent processes in invertebrate brains,we focused on olfactory memory formation in the fruit fly Drosophilamelanogaster. Olfactory learning in flies involves several brain structures,including the mushroom bodies that are considered the primary associa-tion center of olfactory and aversive or appetitive stimuli (3, 25–30) (seefig. S1A). Odors are detected in the antennae and maxillary palp by ol-factory receptor neurons, which project to odor-specific synapse clusterscalled glomeruli located in the antennal lobe. Projection neurons carry theoutput from glomeruli to higher centers of learning and integration, such asthe mushroom bodies and the lateral horn. In the mushroom body calyces,projection neuron axons synapse with mushroom body Kenyon cells, withsomata dorsal to the calyx and with axons that form prominent stalk-likestructures termed lobes. Kenyon cells have bifurcating axons that projectto either a and b lobes (collectively the a/b lobes) or to a′ and b′ lobes orto g lobes. Flies can be efficiently conditioned to avoid a certain odor if itis presented together with an aversive stimulus, such as an electric footshock (31–33), known to activate dopaminergic neurons that form multipleclusters throughout the brain and project to regions including the mush-room body calyces and lobes (34, 35). The exact nature of the conditioningprotocol determines the type of the resulting memories, which are classifiedas short-term memory (STM), middle-term memory (MTM), anesthesia-resistant memory (ARM), and long-term memory (LTM) (3, 4). Of thesememory forms, only LTM is dependent on gene transcription (36) andwould, therefore, be expected to require an intact communication betweenthe synapse and the nucleus. Several genes and signaling molecules areinvolved in LTM in D. melanogaster, which include calcium-permeableNMDA-type glutamate receptors (3, 37–39), Ca2+/CaMKs (40–42), compo-nents of the cAMP-signaling pathway (31, 43, 44), and the transcriptionalregulator CREB (42, 45). Imaging experiments revealed that LTM trainingenhances calcium responses to odors in subpopulations of mushroom bodyneurons (42, 46–48) and other brain regions (49) or cell types (50), suggesting
www.SCIENCESIGNALING.org 7 May 2013 Vol 6 Issue 274 ra33 1
R E S E A R C H A R T I C L E
that calcium may be an important signal during LTM formation. Here, weused recombinant calcium sensors in in vivo imaging experiments and in-hibitors of nuclear Ca2+/CaM signaling expressed in adultD. melanogasterin a temporally and spatially restricted manner to assess the potential role ofnuclear calcium in olfactory avoidance memory. We provide evidence for akey role of nuclear calcium in memory consolidation in flies.
RESULTS
Electrical foot shocks evoke transient increases innuclear calcium in mushroom body neuronsTo investigate the potential role of nuclear calcium signaling in olfactoryavoidance conditioning of adult D. melanogaster, we assessed whether the
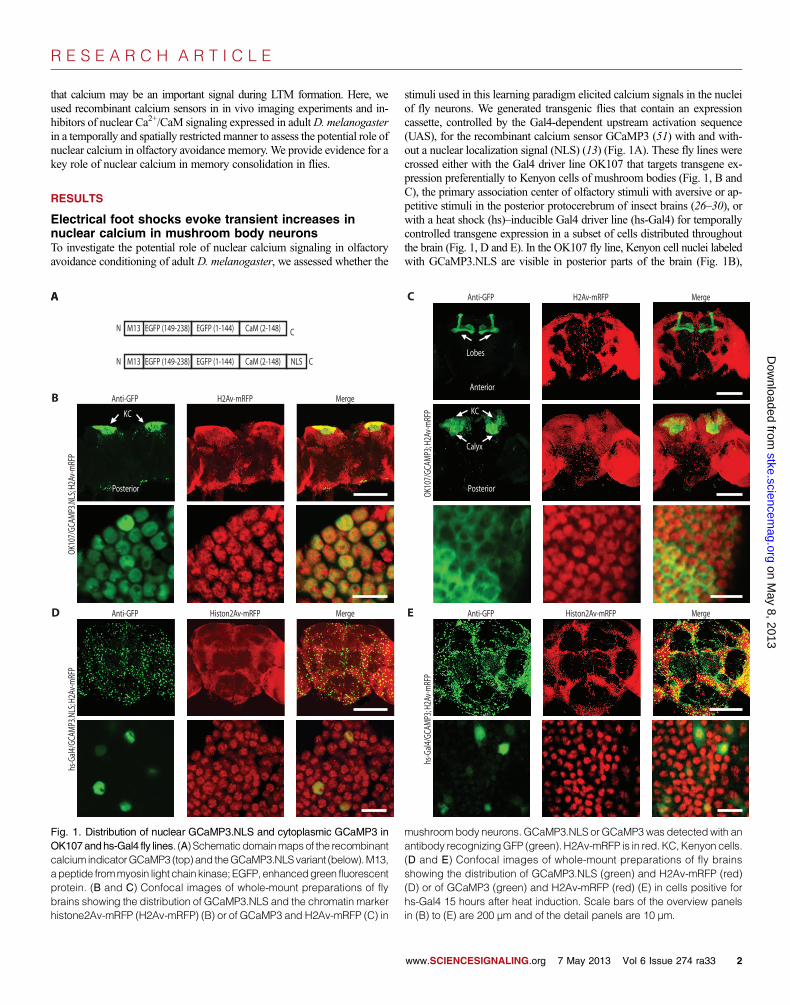
stimuli used in this learning paradigm elicited calcium signals in the nucleiof fly neurons. We generated transgenic flies that contain an expressioncassette, controlled by the Gal4-dependent upstream activation sequence(UAS), for the recombinant calcium sensor GCaMP3 (51) with and with-out a nuclear localization signal (NLS) (13) (Fig. 1A). These fly lines werecrossed either with the Gal4 driver line OK107 that targets transgene ex-pression preferentially to Kenyon cells of mushroom bodies (Fig. 1, B andC), the primary association center of olfactory stimuli with aversive or ap-petitive stimuli in the posterior protocerebrum of insect brains (26–30), orwith a heat shock (hs)–inducible Gal4 driver line (hs-Gal4) for temporallycontrolled transgene expression in a subset of cells distributed throughoutthe brain (Fig. 1, D and E). In the OK107 fly line, Kenyon cell nuclei labeledwith GCaMP3.NLS are visible in posterior parts of the brain (Fig. 1B),
on May 8, 2013
stke.sciencemag.org
Dow
nloaded from
AA
B
OK10
7/GC
AMP3
.NLS
; H2A
v-m
RFP
C
C
N EGFP (149-238)
N EGFP (149-238)
EGFP (1-144)
EGFP (1-144)
CaM (2-148)
CaM (2-148) NLSM13
M13
KC
Anti-GFP
Posterior
Anti-GFPD
hs-G
al4/G
CAM
P3.N
LS; H
2Av-
mRF
P
H2Av-mRFP
Histon2Av-mRFP
Merge
Merge
Merge
Merge
C Anti-GFP
Anterior
Posterior
KC
Calyx
Lobes
OK10
7/GC
AMP3
; H2A
v-m
RFP
Anti-GFPE
hs-G
al4/G
CAM
P3; H
2Av-
mRF
P
H2Av-mRFP
Histon2Av-mRFP
Fig. 1. Distribution of nuclear GCaMP3.NLS and cytoplasmic GCaMP3 in mushroombody neurons. GCaMP3.NLS or GCaMP3 was detected with an
OK107andhs-Gal4 fly lines. (A) Schematicdomainmapsof the recombinantcalcium indicatorGCaMP3 (top) and theGCaMP3.NLSvariant (below).M13,apeptide frommyosin light chain kinase; EGFP, enhancedgreen fluorescentprotein. (B and C) Confocal images of whole-mount preparations of flybrains showing the distribution of GCaMP3.NLS and the chromatin markerhistone2Av-mRFP (H2Av-mRFP) (B) or of GCaMP3 and H2Av-mRFP (C) inantibody recognizingGFP (green). H2Av-mRFP is in red. KC,Kenyon cells.(D and E) Confocal images of whole-mount preparations of fly brainsshowing the distribution of GCaMP3.NLS (green) and H2Av-mRFP (red)(D) or of GCaMP3 (green) and H2Av-mRFP (red) (E) in cells positive forhs-Gal4 15 hours after heat induction. Scale bars of the overview panelsin (B) to (E) are 200 µm and of the detail panels are 10 µm.
www.SCIENCESIGNALING.org 7 May 2013 Vol 6 Issue 274 ra33 2

R E S E A R C H A R T I C L E
on May 8, 2013
stke.sciencemag.org
Dow
nloaded from
whereas GCaMP3 was detected in cell bodies, dendrites, and axons ofKenyon cells that form the calyces, peduncles, and the characteristic ver-tical and horizontal lobes of the mushroom bodies (Fig. 1C and fig. S1A).
We confirmed that GCaMP3.NLS was limited to the nucleus by confocalanalysis of its colocalization with the nuclear chromatin marker histone2Av-mRFP (52) (Fig. 1, B and D). In OK107/GCaMP3.NLS;H2Av-mRFPflies, colocalization analysis with an automatic threshold approach (53)revealed a 93.5% overlap between the two fluorescent signals that repre-sented ~98% of the total GCaMP3.NLS fluorescence intensity (n = 403nuclei). These data indicated that the amount of extranuclear GCaMP3.NLS is less than 2%. Localization of GCaMP3 and GCaMP3.NLS outsidethe nucleus was also directly assessed in OK107 flies by quantifying co-localization with tubulin, a cytoplasmic protein. This revealed cytoplasmicand nuclear localization of GCaMP3 (fig. S1B) and an almost completenuclear localization of GCaMP3.NLS in the mushroom body Kenyoncells of adult flies (fig. S1C). We observed a similarly strict nuclear local-ization of GCaMP3.NLS, when expressed with the OK6 line that targetstransgene expression in motor neuron cell bodies of the ventral cord (54)(fig. S1D) or when the transgene was expressed in muscle cells of 3rd instarlarvae with the muscle-specific promoter MHC (55) (fig. S1E). We alsoquantified the colocalization of GCaMP3.NLS with cytoplasmic markersin cultured cells: cultured mouse hippocampal neurons (fig. S1F) and theinsect cell line High Five (fig. S1G). Quantitative analyses revealed that~5% of GCaMP3.NLS was extranuclear in vivo and in the cultured cells(fig. S1H).
For in vivo calcium imaging, we mounted the flies such that they couldbe exposed to the conditioning stimuli, a controlled stream of odor [3-octanol(OCT) or 4-methyl-cyclohexanol (MCH)] as the conditioned stimulus(CS) and a pulsed electric foot shock as the unconditioned stimulus (fig.S2). In flies expressing GCaMP3.NLS in mushroom body neurons(OK107/GCaMP3.NLS), foot shocks elicited calcium transients in thecell body region of Kenyon cells (Fig. 2A), which may reflect the fact thatthe mushroom body calyx receives a direct shock input from the smalldopaminergic cell cluster PPL2ab (34, 35). The calyx also receives directodor inputs from antennal lobe projection neurons (35); however, they appearinsufficient to generate detectable increases in calcium in nuclei of mushroombody neurons (Fig. 2B). In flies expressing GCaMP3 (OK107/GCaMP3),robust fluorescence changes were observed after electrical and odor stimu-lations in the horizontal lobes of the mushroom bodies and in the calyxand Kenyon cell body region of the mushroom bodies (Fig. 2, C and D).These shock- and odor-induced cytoplasmic calcium transients most likelyresulted from direct inputs onto Kenyon cell axons and dendrites from an-tennal lobe projection neurons that relay olfactory information to the Kenyoncells and from dopaminergic neurons that present information aboutaversive stimuli to the mushroom bodies (3, 35, 56, 57). These results showedthat the shock, but not the odor input, to the mushroom bodies resulted ina cue-dependent generation of nuclear calcium transients in Kenyon cells.
We also investigated the responses to odor or foot shock when the cal-cium sensors were expressed in subsets of cells distributed throughout thebrain using the hs-Gal4 driver line. Foot shocks, but not odors, elicitedrobust calcium responses with both nuclear (Fig. 2, E and F) and cyto-plasmic (Fig. 2, G and H) indicators. We observed nuclear calcium tran-sients in dorsomedial subsuperficial brain areas evoked by foot shocks(Fig. 2E), but not by odor stimulation (Fig. 2F). The lack of odor responsesin this brain region can be explained by the fact that insect olfactorysystems show conserved major projections from the antennal lobes tothe mushroom bodies and lateral horn but not to dorsomedial brain re-gions (58, 59). Thus, foot shocks induced nuclear calcium transients notonly in mushroom body neurons but also in other brain areas that may par-ticipate in relaying or processing shock information. Both shock and odor
evoked cytoplasmic responses in subsuperficial dorsolateral brain regions(Fig. 2, G and H). The cytoplasmic signals induced by odor stimulationswere significantly smaller than those observed using the mushroom bodydriver line (Fig. 2, D, H, and I), suggesting only a partial overlap with theodor pathway to Kenyon cells.
Expression of nuclear calcium blockers during olfactoryavoidance conditioning blocks LTMWe investigated the functional importance of nuclear calcium signalingduring olfactory avoidance conditioning in D. melanogaster using trans-genic fly lines that enabled Gal4-mediated expression of one of the twomyc-tagged nuclear Ca2+/CaM signaling blockers, CaMBP4 (14, 22, 60) or2xM13.NLS, or an inactive, scrambled version of 2xM13.NLS (S2mM13.NLS)as control (Fig. 3A). CaMBP4 and 2xM13.NLS are both nuclear proteinsthat contain four or two copies, respectively, of M13, which is a CaM-bindingpeptide. CaMBP4 is derived from myosin light chain kinase of rabbit skel-etal muscle (60), and 2xM13.NLS is derived from myosin light chain ki-nase of chicken smooth muscle (61). CaMBP4 does not buffer calciumbut instead binds to and inactivates nuclear CaM complexes; it selectivelyblocks gene expression regulated by nuclear calcium signaling in culturedmouse hippocampal neurons and the long-term implementation of differ-ent forms of neuroadaptations in mice (14, 17, 18, 20, 22–24). To confirmthe effectiveness of 2xM13.NLS in the same system in which CaMBP4was characterized, we tested the effect of expression of 2xM13.NLS orS2mM13.NLS in cultured mouse hippocampal neurons. Similar to the ef-fects of CaMBP4 expression in these cells (14), we observed that both pro-teins localized to the nuclei of expressing neurons (fig. S3A) and thatexpressing 2xM13.NLS, but not S2mM13.NLS, attenuated the action po-tential bursting-induced increase in the abundance of the immediate earlytranscription factors c-Fos and activating transcription factor 3 (ATF3) (fig.S3, B and C). In adult flies, immunoblot analysis of brain extracts ofhs-Gal4–driven genotypes revealed that heat shock–induced production ofCaMBP4, 2xM13.NLS, or S2mM13.NLS occurred between 4 and24 hours after heat shock and reached a peak in abundance between12 and 18 hours after heat shock (Fig. 3, B and C). All three proteinsshowed a widespread distribution of expressing cells in fly brains (Fig. 3D).We observed an exclusive nuclear localization of CaMBP4 with the hs-Gal4driver line and the mushroom body driver line OK107 (Fig. 3E) that areexpressed in neurons projecting to the a, a′, b, b′, and g lobes of the mush-room bodies. Similarly, both 2xM13.NLS and S2mM13.NLS localized tothe cell nucleus in adult flies (fig. S4).
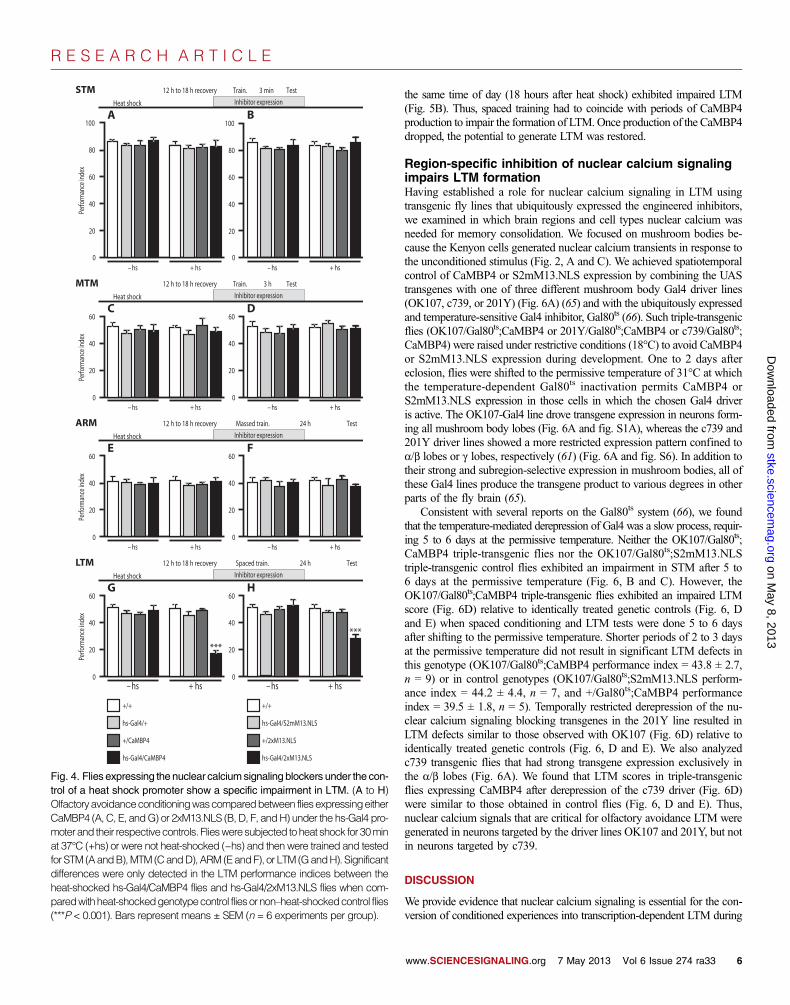
To assess whether nuclear calcium signaling plays a role in any of thevarious forms of olfactory avoidance memory in flies, we expressed either ofthe nuclear calcium signal blockers, CaMBP4 or 2xM13.NLS, or the mutantcontrol S2mM13.NLS using hs-Gal4. We used the heat shock expression sys-tem because it offers a tight temporal control of transgene expression (Fig. 3, Band C) in ubiquitously distributed random subsets of cells (Figs. 1, D and E,and 3D), which include neurons essential for olfactory learning and mem-ory in flies (37, 38, 45, 62, 63).We conditioned flieswhen transgene expres-sion reached its peak (12 to 18 hours after heat shock) during the respectiveconditioning trials (see schematic drawings in Fig. 4).We found no significantimpairment in any of the tested transcription-independent forms of learning,STM,MTM, orARM, in flies expressing any of the transgenes (Fig. 4, A to F).However, LTMwas significantly impaired by the expression of either of thetwo nuclear calcium signaling blockers, CaMBP4 or 2xM13.NLS (Fig. 4, Gand H). In contrast, all other genotype control flies [flies with only hs-Gal4(hs-Gal4/+), flieswith onlyCaMBP4 (+/CaMBP4), flieswith hs-Gal4 and thescrambled control (hs-Gal4/S2mM13.NLS), flies with only 2xM13.NLS(+/2xM13.NLS)], exposed or not exposed to heat shock, had LTMperform-ance indices thatwere indistinguishable from thoseofwild-typecontrol flies (+/+)
www.SCIENCESIGNALING.org 7 May 2013 Vol 6 Issue 274 ra33 3

R E S E A R C H A R T I C L E
on May 8, 2013
stke.sciencemag.org
Dow
nloaded from
Baseline
G hs-Gal4/GCaMP3
H
∆F
hs-Gal4/GCaMP3
Baseline Odor
Foot shock
Odor
∆F
∆F
High
Low
CA
20 s
20%
∆F
∆F
B OK107/GCaMP3.NLS
5 s
10%
Foot shock
Baseline Odor
Baseline
∆F
OK107/GCaMP3
c/Kc
hl p
D OK107/GCaMP3
Baseline Foot shock
Baseline Odor
∆F
OdorOdor
I
20
40
60
0
80
8*
7*
11
9**
16**
10*
4 5*
Shock Odor
OK107
Shock Odor
hs-Gal4
GCaMP3
GCaMP3.NLS
Peak response amplitude
E
∆F
hs-Gal4/GCaMP3.NLS
F
∆F
hs-Gal4/GCaMP3.NLS
Baseline Foot shock
Baseline Odor
Odor
OK107/GCaMP3.NLS
Foot shock Foot shockFoot shockFoot shock
∆F
∆F
High
Low
% ∆F
/F0
Fig. 2. In vivo imaging of nuclear calcium signals in Drosophila. (A to H) Each panel showsrepresentative examples of quantitative image analysis of GCaMP3.NLS or GCaMP3 fluorescencechanges in vivo in OK107 flies (A to D) and hs-Gal4 flies (E to H) in response to the application ofelectric foot shocks (A, C, E, and G) or odor (B, D, F, and H). Grayscale image panels showoverview images of fly brains (scale bars, 30 µm). Color images (scale bars, 10 µm) showcolor-coded (see color bar) fluorescence intensity during baseline and stimulation (foot shock orodor) and their difference (DF) from the boxed region shown in the corresponding overview image.Pairs of imageshavebeenscaled identically, but image thresholdingand some imagingparametersvary from fly to fly. The quantitative analysis shown at the bottom of each panel came from threeregions of interest in the color images. hl, horizontal lobes; p, peduncle; c/Kc, calyx and Kenyon cellbodies. (I) Summary statistics showing the mean (± SEM) amplitude of responses to odor and footshock (*P < 0.05, **P < 0.001, one-sample t test). N values are indicated above each bar.
www.SCIENCESIGNALING.org 7 May 2013 Vol 6 Issue 274 ra33 4

R E S E A R C H A R T I C L E
on May 8, 2013
stke.sciencemag.org
Dow
nloaded from
(Fig. 4, G and H). The failure to form LTM in animals expressing eithernuclear calcium signaling blocker was not due to an impairment in percep-tion, propagation, or association of the sensory stimuli delivered in theconditioning trials, nor was it due to an impaired recall of stored information,because the performance indices of all other transcription-independentmemory forms (STM, MTM, and ARM) were comparable to those of wild-type control flies and genotype control flies exposed or not exposed to heatshock (Fig. 4, A to F).
In contrast, when we disrupted the cAMP signaling pathway in parallelexperiments by heat shock–mediated ubiquitous overexpression of dunce(64), which encodes a phosphodiesterase that degrades cAMP, we found,as expected, significant deficits in STM (fig. S5), indicating that the ex-perimental protocol applied in our experiments can detect failures in STMperformance. These findings demonstrated that the nuclear calcium sig-naling blockers selectively impaired the formation of LTM.
Impairment of LTM formation by inhibition of nuclearcalcium signaling is reversibleTo rule out that inhibition of nuclear calcium signaling in the fly brain byCaMBP4 was due to permanent damage that would compromise neuronalfunction and nonspecifically impair the formation of LTM or its recall, weexpressed CaMBP4 in a temporally restricted manner with the heat shockpromoter and performed LTM conditioning either during or after the CaMBP4expression. CaMBP4 was readily detected 18 hours after its inductionwith heat shock but was undetectable after an additional 24 hours (thatis, 42 hours after heat shock) (Fig. 5A). We found that spaced training done42 hours after heat shock resulted in LTM scores in hs-Gal4/CaMBP4 fliesthat were similar to those obtained in hs-Gal4/CaMBP4 flies that had notbeen subjected to heat shock (Fig. 5B). In contrast, hs-Gal4/CaMBP4 fliesthat were subjected to heat shock in parallel experiments but receivedspaced training during the period when CaMBP4 was abundant and at
E
OK10
7/
H2Av
-mRF
P; Ca
MBP
4
H2Av-mRFP MergeAnti-myc
hs-G
al4/
H2Av
-mRF
P; Ca
MBP
4
A
S2mM13.NLS (control)
2xM13.NLS
N M13 M13 CNLS myc
CaMBP4
N M13 M13 CmycM13 M13
N mM13 mM13 NLS myc C
hs-Gal4/2xM13.NLSB24 h12 h8 h4 h0 h 36 h18 h15 h After hs
Anti-Gal4
Anti-myc
Anti-tubulin
C hs-Gal4/CaMBP4 hs-Gal4/2xM13.NLShs-Gal4/S2mM13.NLS
5
15
20
0
10
24 h12 h8 h4 h 18 h15 h0 h
hs-G
al4/S
2mM
13.N
LShs
-Gal4
/2xM
13.N
LS
D Anti-myc MergeHoechst
hs-G
al4/C
aMBP
4
).u.a( ecnadnuba nietorP
36 h
Fig. 3. Temporal analysis of the production of the nuclear calciumsignaling blockers. (A) Schematic representation of the domains of thetransgenic proteins used to interfere with nuclear calcium signaling.CaMBP4 contains four cryptic NLS in the N-terminal regions of theM13 domains, which are not shown and are sufficient for its nuclear lo-calization (60). The expression of all constructs is under the control ofthe UAS site. (B) Immunoblot analysis of protein extracts prepared fromhs-Gal4/2xM13.NLS fly brains at various time points (0 to 36 hours) afterheat shock (hs). 2xM13.NLS was detected with the antibody against themyc epitope. Heat shock induction is shown by the production of theGal4 protein. Tubulin is shown as a loading control. (C) Quantitativeanalysis of the protein abundance profile of the nuclear calcium sig-
naling blockers and control from fly brains at various time points afterheat shock (n = 3 each). a.u., arbitrary units. (D) Confocal images ofwhole-mount preparations of fly brains expressing the indicated trans-genes. Preparations were labeled with Hoechst (nuclear chromatin inblue) and the myc antibody (anti-myc) for immunofluorescence (red).hs-Gal4 flies were fixed 15 hours after heat shock induction. Scale bars,75 µm. (E) Confocal images of whole-mount preparations of fly brainshowing the distribution of CaMBP4 (green) and H2Av-mRFP (red) inheat shock–positive cells 15 hours after heat induction or OK107-positiveneurons. The recombinant nuclear calcium signaling blocker CaMBP4 isimmunolabeled with antibody recognizing the myc tag (anti-myc). Scalebars, 5 µm.
www.SCIENCESIGNALING.org 7 May 2013 Vol 6 Issue 274 ra33 5
R E S E A R C H A R T I C L E
on May 8, 2013
stke.sciencemag.org
Dow
nloaded from
the same time of day (18 hours after heat shock) exhibited impaired LTM(Fig. 5B). Thus, spaced training had to coincide with periods of CaMBP4production to impair the formation of LTM.Once production of the CaMBP4dropped, the potential to generate LTM was restored.
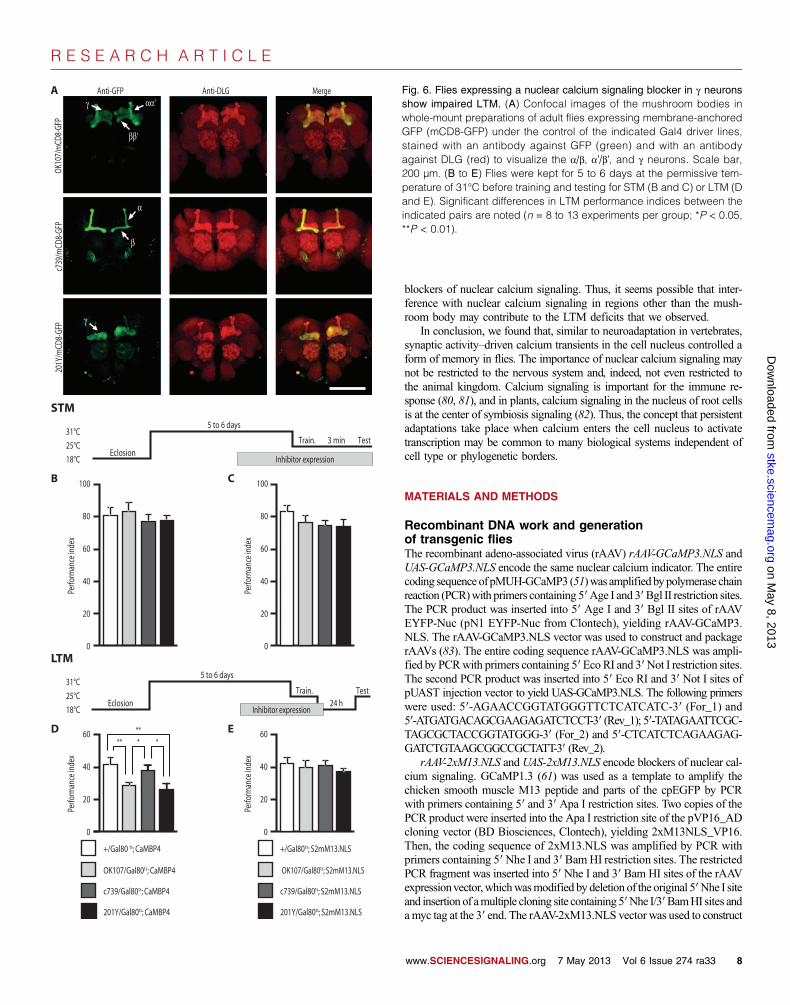
Region-specific inhibition of nuclear calcium signalingimpairs LTM formationHaving established a role for nuclear calcium signaling in LTM usingtransgenic fly lines that ubiquitously expressed the engineered inhibitors,we examined in which brain regions and cell types nuclear calcium wasneeded for memory consolidation. We focused on mushroom bodies be-cause the Kenyon cells generated nuclear calcium transients in response tothe unconditioned stimulus (Fig. 2, A and C). We achieved spatiotemporalcontrol of CaMBP4 or S2mM13.NLS expression by combining the UAStransgenes with one of three different mushroom body Gal4 driver lines(OK107, c739, or 201Y) (Fig. 6A) (65) and with the ubiquitously expressedand temperature-sensitive Gal4 inhibitor, Gal80ts (66). Such triple-transgenicflies (OK107/Gal80ts;CaMBP4 or 201Y/Gal80ts;CaMBP4 or c739/Gal80ts;CaMBP4) were raised under restrictive conditions (18°C) to avoid CaMBP4or S2mM13.NLS expression during development. One to 2 days aftereclosion, flies were shifted to the permissive temperature of 31°C at whichthe temperature-dependent Gal80ts inactivation permits CaMBP4 orS2mM13.NLS expression in those cells in which the chosen Gal4 driveris active. The OK107-Gal4 line drove transgene expression in neurons form-ing all mushroom body lobes (Fig. 6A and fig. S1A), whereas the c739 and201Y driver lines showed a more restricted expression pattern confined toa/b lobes or g lobes, respectively (61) (Fig. 6A and fig. S6). In addition totheir strong and subregion-selective expression in mushroom bodies, all ofthese Gal4 lines produce the transgene product to various degrees in otherparts of the fly brain (65).
Consistent with several reports on the Gal80ts system (66), we foundthat the temperature-mediated derepression of Gal4 was a slow process, requir-ing 5 to 6 days at the permissive temperature. Neither the OK107/Gal80ts;CaMBP4 triple-transgenic flies nor the OK107/Gal80ts;S2mM13.NLStriple-transgenic control flies exhibited an impairment in STM after 5 to6 days at the permissive temperature (Fig. 6, B and C). However, theOK107/Gal80ts;CaMBP4 triple-transgenic flies exhibited an impaired LTMscore (Fig. 6D) relative to identically treated genetic controls (Fig. 6, Dand E) when spaced conditioning and LTM tests were done 5 to 6 daysafter shifting to the permissive temperature. Shorter periods of 2 to 3 daysat the permissive temperature did not result in significant LTM defects inthis genotype (OK107/Gal80ts;CaMBP4 performance index = 43.8 ± 2.7,n = 9) or in control genotypes (OK107/Gal80ts;S2mM13.NLS perform-ance index = 44.2 ± 4.4, n = 7, and +/Gal80ts;CaMBP4 performanceindex = 39.5 ± 1.8, n = 5). Temporally restricted derepression of the nu-clear calcium signaling blocking transgenes in the 201Y line resulted inLTM defects similar to those observed with OK107 (Fig. 6D) relative toidentically treated genetic controls (Fig. 6, D and E). We also analyzedc739 transgenic flies that had strong transgene expression exclusively inthe a/b lobes (Fig. 6A). We found that LTM scores in triple-transgenicflies expressing CaMBP4 after derepression of the c739 driver (Fig. 6D)were similar to those obtained in control flies (Fig. 6, D and E). Thus,nuclear calcium signals that are critical for olfactory avoidance LTM weregenerated in neurons targeted by the driver lines OK107 and 201Y, but notin neurons targeted by c739.
DISCUSSION
We provide evidence that nuclear calcium signaling is essential for the con-version of conditioned experiences into transcription-dependent LTM during
Test
Heat shock
12 h to 18 h recovery 3 minTrain.
Inhibitor expression
60
80
40
20
+ hs
100
0
A100
80
60
40
20
B
– hs + hs0
– hs
MTM Test
Heat shock
12 h to 18 h recovery 3 hTrain.
+ hs
Inhibitor expression
20
40
60
0– hs + hs
20
40
60
0
DC
– hs
ARMHeat shock
12 h to 18 h recovery 24 h TestMassed train.
– hs + hs
20
40
60
0
EInhibitor expression
– hs + hs
F
20
40
60
0
LTMHeat shock
12 h to 18 h recovery 24 h TestSpaced train.
– hs + hs
20
40
60
0
GInhibitor expression
– hs + hs
H
20
40
60
0
***
***
hs-Gal4/2xM13.NLS
hs-Gal4/S2mM13.NLS
+/+
+/2xM13.NLS
+/+
+/CaMBP4
hs-Gal4/CaMBP4
hs-Gal4/+
STMPe
rform
ance
inde
xPe
rform
ance
inde
xPe
rform
ance
inde
xPe
rform
ance
inde
x
Fig. 4. Flies expressing the nuclear calciumsignaling blockers under the con-trol of a heat shock promoter show a specific impairment in LTM. (A to H)
Olfactoryavoidanceconditioningwascomparedbetween fliesexpressingeitherCaMBP4 (A, C, E, andG) or 2xM13.NLS (B, D, F, andH) under the hs-Gal4 pro-moterand their respectivecontrols. Fliesweresubjected toheat shock for30minat 37°C (+hs) or were not heat-shocked (−hs) and then were trained and testedfor STM (AandB),MTM (CandD),ARM (EandF), or LTM (GandH). Significantdifferences were only detected in the LTM performance indices between theheat-shocked hs-Gal4/CaMBP4 flies and hs-Gal4/2xM13.NLS flies when com-paredwithheat-shockedgenotypecontrol fliesornon–heat-shockedcontrol flies(***P < 0.001). Bars represent means ± SEM (n = 6 experiments per group).www.SCIENCESIGNALING.org 7 May 2013 Vol 6 Issue 274 ra33 6

R E S E A R C H A R T I C L E
on May 8, 2013
stke.sciencemag.org
Dow
nloaded from
spaced olfactory avoidance training of adult D. melanogaster. The signalingpathway activated by shock-induced calcium transients in the cell nucleuswas, however, dispensable for the formation of short-lasting, transcription-independent forms of memory. Thus, intact nuclear calcium signaling wasneeded for the progression of these olfactory memories from short-lived tolong-lasting and transcription-dependent forms. Transient repression ofnuclear calcium signalingwith the nuclear calcium signaling inhibitor CaMBP4did not induce permanent damage to fly neurons, but rather acutely and tran-siently impaired the formation of olfactory avoidance LTM.
Together with the fact that the mammalian nervous system requires nu-clear calcium signaling to establish a broad spectrum of transcription-dependent adaptations (1, 67), it emerges that nuclear calcium acts as auniversal and evolutionarily conserved signal that initiates appropriate ge-nomic responses needed for the implementation of stable adjustments.
In mammalian neurons, nuclear calcium acts primarily through nuclearCaMKs and controls the activity of CREB/CBP (12, 68–71) and methyl-CpGbinding protein 2 (MeCP2) (72). The nuclear calcium–regulated gene poolin mouse hippocampal neurons contains nearly 200 genes, many of whichare known or putative CREB/CBP targets and play roles in acquired neuro-protection, memory consolidation, and the development of chronic inflamma-tory pain (14, 18, 20, 22, 24, 73). The activation of synaptic NMDA receptorsis important for the initiation of the nuclear Ca2+/CaMK-CREB/CBP sig-naling module after synaptic stimulation (1, 11). In D. melanogaster, a simi-lar pathwaymay link nuclear calcium signaling to LTM formation. SuppressionofNMDA receptor, CaMKII, orCREB function inmushroombodya/b or gneurons blocks the transcription-dependent formation of time- and cell-specific LTM traces encoding this memory (36, 39, 42, 46, 62), which isconsistent with activation of NMDA receptors in Kenyon cells generating acalcium signal that, at least in mushroom body g neurons, may be trans-mitted into the nuclei to activate a potential nuclear isoform of CaMKII(41). CREB is a target of CaMKs and has a well-documented role in theformation of LTM in flies (42, 45, 46, 62). Thus, it seems possible thatan evolutionarily conserved nuclear Ca2+/CaMK-CREB signaling modulemay be primarily responsible for the induction of a learning-associated geneprogram in both vertebrate and invertebrate neurons.
With spatially restricted interference and imaging methods in fly brains,distinct memory traces have been identified in subregions of the flies’mush-
room body and other brain regions, suggesting that different memory formsand memory phases are encoded in different brain structures. These includeshort-lived memory traces located in the antennal lobe (74) and the g neu-rons of the mushroom bodies (75), and middle-term and slightly delayedmemory traces in dorsal paired medial (DPM) neurons (76) and a′/b′mushroom body neurons (77). LTM traces have been mapped to the a/band the g branches of the mushroom bodies (42, 46, 48, 75), although othercells, such as the two dorsal-anterior-lateral (DAL) neurons (78), two pairsof PPL1 dopaminergic neurons (49), cer-expressing glial cells surroundingthe Kenyon cell bodies (79), or NMDA receptor–expressing neurons in theellipsoid body of the central complex (38), are also required for memoryconsolidation.
In the a/b and g neurons of mushroom bodies, we identified a dichot-omy in the requirements of these cells for cAMP signaling and nuclearcalcium. For LTM formation, a/b neurons (targeted by c739) required in-tact cAMP signaling (75), but not nuclear calcium signaling. Conversely,in g neurons of the mushroom bodies (targeted by 201Y), nuclear calciumwas critical for the formation of LTM, whereas cAMP signaling has beenreported as dispensable (75). The a/b neurons may contribute to LTMthrough a mechanism that involves activation and nuclear translocationof cAMP-dependent protein kinase A and subsequent stimulation ofCREB-dependent transcription (42, 46). In contrast, g neurons may usenuclear calcium signaling, possibly to activate CaMKs, to control a poten-tially distinct transcriptional program. Differences in the genomic responsesmay be one reason why LTM traces that consist of an enhancement ofthe stimulus-associated calcium influx are detectable 9 to 24 hours afterspaced training in a/b neurons (48, 77) and 18 to 48 hours after training ing neurons (42).
Together, it seems that distinct subsets of mushroom body neurons thathave been previously implicated in LTM use distinct different signalingcascades to control the appropriate LTM-associated genomic responses.Although mushroom body g neurons may represent key cells in nuclearcalcium–dependent memory consolidation, it should be noted that theGal4 driver lines used in this study can lead to expression of the transgeneoutside the mushroom body (65). For example, using the hs-Gal4 driverline, we observed shock-evoked nuclear calcium transients in regionsoutside the mushroom bodies and deficits in LTM upon expression of
18 h after heat-shock 42 h after heat-shock A
Anti-
DLG
No heat-shock
Anti-
myc
B
25°C
37°C
18°C Inhibitor expression
Train.18 h
Test18 h
Train.42 h
42 h
hs
0 h 66 h
Test42 h
– hs
40
20
60
0
Perfo
rman
ce in
dex
**
+ hs
hs-Gal4/CaMBP4 (train 18 h, –hs)
hs-Gal4/CaMBP4 (train 18 h, +hs)
hs-Gal4/CaMBP4 (train 42 h, +hs)
+ hs
Fig. 5. The block in LTMmediated by inhibition of nuclear calcium signaling isreversible. (A) Confocal images show the nuclear calcium signaling blocker
body against discs large protein (DLG) to show its major neuropil structures(red). (B) Flies were trained for LTM 18 hours (when CaMBP4 was actively
CaMBP4 in whole-mount brain preparations from flies without heat shock or18 or 42 hours after heat shock. CaMBP4 was detected with an antibodyagainst themyc epitope (green). Fly brainswere counterstainedwith an anti-
producedandabundant)and42hours (whenCaMBP4wasno longerproduced)after heat shock induction of CaMBP4 expression. LTM was tested in allgroups 24 hours after training (**P < 0.01, n = 8 experiments per group).
www.SCIENCESIGNALING.org 7 May 2013 Vol 6 Issue 274 ra33 7
25°C
31°C
STM
18°C
B
60
80
40
20
100
0
25°C
31°C
18°C
LTM
EclosionTest
5 to 6 days
24 hTrain.
Inhibitor expression
20
40
60
0
D****
**
+/Gal80 ts; CaMBP4
OK107/Gal80ts; CaMBP4
c739/Gal80ts; CaMBP4
201Y/ Gal80ts; CaMBP4
Test
Inhibitor expressionEclosion
5 to 6 days
3 minTrain.
Perfo
rman
ce in
dex
60
20
40
0
E
+/Gal80ts; S2mM13.NLS
OK107/Gal80ts; S2mM13.NLS
c739/Gal80ts; S2mM13.NLS
201Y/ Gal80ts; S2mM13.NLS
C 100
60
80
40
20
0
γ
OK10
7/m
CD8-
GFP
α
β
ββ’
201Y
/mCD
8-GF
Pc7
39/m
CD8-
GFP
Perfo
rman
ce in
dex
Perfo
rman
ce in
dex
Perfo
rman
ce in
dex
R E S E A R C H A R T I C L E
stkD
ownloaded from
blockers of nuclear calcium signaling. Thus, it seems possible that inter-ference with nuclear calcium signaling in regions other than the mush-room body may contribute to the LTM deficits that we observed.
In conclusion, we found that, similar to neuroadaptation in vertebrates,synaptic activity–driven calcium transients in the cell nucleus controlled aform of memory in flies. The importance of nuclear calcium signaling maynot be restricted to the nervous system and, indeed, not even restricted tothe animal kingdom. Calcium signaling is important for the immune re-sponse (80, 81), and in plants, calcium signaling in the nucleus of root cellsis at the center of symbiosis signaling (82). Thus, the concept that persistentadaptations take place when calcium enters the cell nucleus to activatetranscription may be common to many biological systems independent ofcell type or phylogenetic borders.
on May 8, 2013
e.sciencemag.org
MATERIALS AND METHODS
Recombinant DNA work and generationof transgenic fliesThe recombinant adeno-associated virus (rAAV) rAAV-GCaMP3.NLS andUAS-GCaMP3.NLS encode the same nuclear calcium indicator. The entirecoding sequenceof pMUH-GCaMP3 (51)was amplifiedbypolymerase chainreaction (PCR)with primers containing 5′Age I and 3′Bgl II restriction sites.The PCR product was inserted into 5′ Age I and 3′ Bgl II sites of rAAVEYFP-Nuc (pN1 EYFP-Nuc from Clontech), yielding rAAV-GCaMP3.NLS. The rAAV-GCaMP3.NLS vector was used to construct and packagerAAVs (83). The entire coding sequence rAAV-GCaMP3.NLS was ampli-fied by PCRwith primers containing 5′EcoRI and 3′Not I restriction sites.The second PCR product was inserted into 5′ Eco RI and 3′ Not I sites ofpUAST injection vector to yield UAS-GCaMP3.NLS. The following primerswere used: 5′-AGAACCGGTATGGGTTCTCATCATC-3′ (For_1) and5′-ATGATGACAGCGAAGAGATCTCCT-3′ (Rev_1); 5′-TATAGAATTCGC-TAGCGCTACCGGTATGGG-3′ (For_2) and 5′-CTCATCTCAGAAGAG-GATCTGTAAGCGGCCGCTATT-3′ (Rev_2).
rAAV-2xM13.NLS and UAS-2xM13.NLS encode blockers of nuclear cal-cium signaling. GCaMP1.3 (61) was used as a template to amplify thechicken smooth muscle M13 peptide and parts of the cpEGFP by PCRwith primers containing 5′ and 3′ Apa I restriction sites. Two copies of thePCR product were inserted into the Apa I restriction site of the pVP16_ADcloning vector (BD Biosciences, Clontech), yielding 2xM13NLS_VP16.Then, the coding sequence of 2xM13.NLS was amplified by PCR withprimers containing 5′Nhe I and 3′ BamHI restriction sites. The restrictedPCR fragment was inserted into 5′ Nhe I and 3′ Bam HI sites of the rAAVexpressionvector,whichwasmodified by deletion of the original 5′Nhe I siteand insertion of amultiple cloning site containing5′Nhe I/3′BamHI sites andamyc tag at the 3′ end. The rAAV-2xM13.NLS vector was used to construct
Fig. 6. Flies expressing a nuclear calcium signaling blocker in g neuronsshow impaired LTM. (A) Confocal images of the mushroom bodies inwhole-mount preparations of adult flies expressing membrane-anchoredGFP (mCD8-GFP) under the control of the indicated Gal4 driver lines,stained with an antibody against GFP (green) and with an antibodyagainst DLG (red) to visualize the a/b, a′/b′, and g neurons. Scale bar,200 µm. (B to E) Flies were kept for 5 to 6 days at the permissive tem-perature of 31°C before training and testing for STM (B and C) or LTM (Dand E). Significant differences in LTM performance indices between theindicated pairs are noted (n = 8 to 13 experiments per group; *P < 0.05,**P < 0.01).
Anti-DLG Aγ αα’
Anti-GFP Merge
www.SCIENCESIGNALING.org 7 May 2013 Vol 6 Issue 274 ra33 8

R E S E A R C H A R T I C L E
on May 8, 2013
stke.sciencemag.org
Dow
nloaded from
and package rAAVs (83). Finally, 2xM13.NLSwas subcloned into 5′Bgl IIand3′Not I sites of the pUASTinjectionvector toyieldUAS-2xM13.NLS. Thefollowing primers were used: 5′-TGGGCCCAGTCGACTCAT-3′ (For_1)and 5′-TAAGGGCCCTCGATGTTG-3′ (Rev_1); 5′-TAAGCTAGC-GCCGCCACCATGGGCCC-3′ (For_2) and 5′-TAAGGATCCGTCCAGATC-GAAATCGTC-3′ (Rev_2); 5′-AGATCTAAGCTAGCGCCGCCATTT-3′(For_3) and5′-ATTGCGGCCGCTTACAGATCCTCTTCTGAG-3′ (Rev_3).
rAAV-S2mM13.NLS and UAS-S2mM13.NLS each encode a scrambledcontrol for the blocker of nuclear calcium signaling. The two copies of theM13 peptide of rAAV-2xM13.NLS were replaced by two copies of a scram-bled M13 version (S2mM13). 5′ Nhe I, 3′ Bam HI, and 3′ Xho I restrictionsites were added to the ends of the synthesized peptide sequence (GeneART).The scrambled peptide was inserted in-frame into 5′ Nhe I and 3′ Xho I sitesof rAAV-2xM13.NLS, yielding rAAV-S2mM13.NLS. Finally, S2mM13.NLSwas subcloned into 5′ Bgl II and 3′Not I sites of the pUAST injection vector.The following primers were used: 5′-AGCGGATCCACCATGGGCCC-3′(For_1) and 5′-ATTGCGGCCGCTTACAGATCCTCTTCTGAG-3′(Rev_1).
UAS-CaMBP4 encodes a blocker of nuclear calcium signaling. TheCaMBP4 construct (60) was amplified by PCR with primers containing5′ Nhe I and 3′ Bam HI restriction sites. CaMBP4 consists of four copiesof the rabbit skeletal muscle M13 peptide, each of which contains anN-terminal cryptic NLS (60). The PCR product was inserted into 5′ Nhe Iand 3′ Bam HI sites of modified rAAVexpression vector, yielding rAAV-CaMBP4, and then subcloned into 5′Bgl II and 3′Not I sites of the pUASTinjection vector to yield UAS-CaMBP4. The following primers were used:5′-AATTGGATCCCCCATGGGACCC-3′ (For_1) and 5′-ATTGCGG-CCGCTTACAGATCCTCTTCTGAG-3′ (Rev_1).
Fly stocksWild-type 2202U and hs-Gal4 (P26) flies were obtained from Y. Zhong,histone2Av-mRFP and histone2Av-GFP flies were obtained from J. Grosshans,and UAS-dunce flies were obtained from H. L. Atwood. UAS-GCaMP3was obtained from L. L. Looger. The OK107, 201Y, and c739 driver linesand the UAS-mCD8-GFP line (84) were obtained from the BloomingtonStock Center. All flies used in the behavior experiments were outcrossedto the genetic background of the wild-type flies by crossing heterozygousvirgins with 2202U males for six generations. Wild-type controls in all ex-periments were indicated as +/+. Embryo injections to generate the trans-genic flies (UAS-2xM13.NLS, UAS-S2mM13.NLS, UAS-CaMBP4, andUAS-GCaMP3.NLS) were done by Best Gene.
Heat shock regimenFlies were cultured in standard medium [agar (8 g/liter), yeast (18 g/liter),soy flour (10 g/liter), yellow cornmeal (80 g/liter), light corn syrup (22 g/liter),malt extract (80 g/liter), and propionic acid (6.25 ml/liter)] at 60 to 75%relative humidity and at 18°C to prevent leaky expression. For heat shockinduction, flies were collected within 1 to 2 days after eclosion, placed infresh food vials containing a strip of Whatman filter paper to absorb extrahumidity, and kept at 18°C. Twelve to 18 hours before training, the vialswere then submerged in a 37°C water bath until the bottom of the foamstopper (inside the vials) was below the surface of the water, thereby en-suring that the flies could not escape the heat shock. After the vials re-mained submerged for 30 min, the vials were transferred to the climatechamber room (25°C and 75% relative humidity). Training began imme-diately after the incubation period.
Immunoblotting and immunohistochemistryFor immunoblot analysis, either 10 heads of adult flies per sample werehomogenized directly in 40 µl of preheated 1× Laemmli or hippocampal
neurons were lysed by adding 200 ml of preheated 1× Laemmli. Addition-ally, samples were boiled for 5 min to denature all proteins. The super-natants were centrifuged at 14,000 rpm in a microfuge at room temperaturefor 5 min, loaded on 12% SDS-polyacrylamide gels, and run at 35 mA/gelfor 90 min. Proteins were transferred directly onto a nitrocellulose mem-brane with a pore size of 0.45 µm (Schleicher & Schuell) in a wet transferchamber for 1.5 hours at a constant potential of 20 V. As a control, themembrane was counterstained with Ponceau red (Serva) for 5 min. Afterincubation with blocking solution [5% milk powder in PBST (1× phosphate-buffered saline + 0.01% Tween 20)] for 1 hour, primary antibodies [mouseantibody against Myc (SC-40, Santa Cruz Biotechnology), mouse anti-body against tubulin (Sigma), rabbit antibody against Gal4 DNA bindingdomain (Santa Cruz Biotechnology), rabbit antibody against CREB (NewEngland Biolabs), rabbit antibody against pCREB (Upstate), rabbit anti-body against ATF3 (C-19, Santa Cruz Biotechnology), or rabbit antibodyagainst c-Fos (SC-52, Santa CruzBiotechnology)] were added, and themem-brane was incubated overnight at 4°C. On the following day, the membranewaswashedwith PBST. Secondary antibodies [goat antibody againstmousehorseradish peroxidase (HRP) or goat antibody against rabbit HRP (Dianova)]were then applied for 30minand thenwashedoffwithPBST.Finally, themem-brane was subjected to an enhanced chemiluminescence detection system(ECL, Amersham) and afterward exposed to Kodak Hyperfilm for the desiredtime. Films were developed in a chemiluminescence film developer (M35M-Omat Processor, Kodak).
Two- to 3-day-old flies were used for whole-mount brain preparation(85). Briefly, fly brains were dissected in ice-cold Schneider’s DrosophilaMedium (Gibco Invitrogen) and fixed in 4% paraformaldehyde/PBS, 4%sucrose solution overnight at 4°C. The brains were washed in PAT (PBScontaining 1% bovine serum albumin, 0.5% Triton X-100, and 0.05%Na-azide) at room temperature. After incubation with 3% normal goat serumfor 1 hour, primary antibodies [mouse antibody against myc (SC-40, SantaCruz Biotechnology), rabbit antibody against GFP (Molecular Probes),rabbit antibody against GFP conjugated to Alexa 488 (Invitrogen), rabbitantibody against DLG, or mouse antibody against a-tubulin] were addedand incubated overnight at 4°C. On the following day, the brains were washedagain in PAT, and secondary antibodies [goat antibody against rabbit con-jugated to Cy3 (Dianova), goat antibody against mouse conjugated toAlexa 488 (Molecular Probes), or goat antibody against mouse conjugatedto Alexa 568 (Invitrogen)] were added and incubated overnight at 4°C. Thebrains were then washed again in PAT for 24 hours before being counter-stained and mounted in Vectashield containing 4′,6-diamidino-2-phenylindole(1.5 µg/ml). The brains were imaged using either a Leica SP2 confocalmicroscope with an HCX PL APO CS 40× 1.25 oil UV objective (LeicaMicrosystems GmbH) or a Zeiss LSM 700 confocal microscope with a40×1.25oil or 63×1.32oil objective.Confocal z-stackswere processedusingImageJ and Adobe Photoshop software. Colocalization was calculated asthe number of pixels above threshold for both channels (GCaMP3.NLSand H2Av-mRFP) divided by the number of pixels above threshold forGCaMP3.NLS, where threshold was determined automatically accordingto the method of Costes et al. (53). Colocalization (GCaMP3.NLS andtubulin) was calculated with the same method.
Recombinant protein expression in hippocampalneurons and insect cell linesThe procedure used to isolate and culture hippocampal neurons hasbeen described previously (24, 86). Briefly, hippocampal neurons fromnewborn C57Bl6J mice (Charles River) were cultured in Neurobasalmedium (Invitrogen) containing 1% rat serum, B27 (Invitrogen), and pen-icillin and streptomycin (Sigma). The vectors used to construct and packagerAAVs have been described previously (22). Neurons were infected with
www.SCIENCESIGNALING.org 7 May 2013 Vol 6 Issue 274 ra33 9

R E S E A R C H A R T I C L E
on May 8, 2013
stke.sciencemag.org
Dow
nloaded from
rAAV-GCaMP3.NLS, rAAV 2xM13.NLS, or rAAV S2mM13.NLS after4 days in vitro. A total volume of 2.5 ml containing about 1 × 1011 genomicvirus particles was added directly to each 35-mm culture dish.
The coding sequences of rAAV-GCaMP3.NLS and rAAV-Btg2-mCherry.NES from recombinant adeno-associated viral mammalian expression vectorswere PCR-subcloned and inserted into a pIE vector (Novagen) containing anIE1 insect cell–specific promoter. The resulting pIE-GCaMP3.NLS andpIE-Btg2-mCherry.NES were verified by sequencing and transfected intoHI5 insect cells by standard calcium phosphate precipitation–based trans-fection techniques according to the manufacturer’s recommendations.
Behavior studiesOlfactory associative learning was measured by training 2- to 3-day-old flieswith a Pavlovian conditioning procedure (33). Groups of about 60 to 100flies were placed in the training chamber with air flow through the chamberat 750 ml/min. The flies were exposed to this novel environment for 90 s.Flies received one-session training, during which they were exposed for 60 ssequentially to the first odor, which was OCTor MCH paired with electricshocks [conditioned stimulus (CS+)], and then to the second odor, MCHor OCT without the electric shocks [unconditioned stimulus (CS−)]. Theelectric shocks were delivered at 60 V in 1.5-s pulses, repeated every 5 s for1 min. Conditioned odor avoidance was tested immediately after training.During testing, flies were given 2 min to choose between the CS+ and CS−in each arm of a T-maze before being trapped, anesthetized, and counted.From these counts, a performance index was calculated as the difference ofthe number of flies avoiding the CS+ minus that avoiding the CS− dividedby the total number of flies and multiplied by 100. Thus, an equal numberof flies in each arm (representing no learning) yielded a performance indexof 0, and an avoidance of the CS+ by all flies yielded a performance indexof 100. The pure odors were diluted 1.5:1000 (OCT) and 1:1000 (MCH) inmineral oil. To eliminate naïve odor bias, experiments were performed in acounterbalanced design where both the allocation of OCT and MCH to theCS+ and the CS− and the order of presentation of the CS+ and the CS−during training were varied.
STM (3-min memory) was evaluated after one-session training fol-lowed immediately by the testing. MTM (3-hour memory) was evaluatedafter one-session training followed by a 3-hour interval at 25°C and 75%relative humidity before testing. Two forms of 24-hour memory were eval-uated after either spaced (LTM) or massed training (ARM) (36). Spacedtraining as an LTM paradigm consisted of 10 cycles of one-session train-ing, with a 15-min interval between each session. Massed training as anARM paradigm consisted of 10 cycles of one-session training, where eachsession immediately follows the previous one. For both spaced and massedtraining, flieswere stored at 18°Cand60 to 75% relative humidity for 24 hoursbefore training and were maintained at 25°C and 75% relative humidity for24 hours between training and testing. Details of each procedure are shownas schematics in the relevant figure panels.
In vivo calcium imagingFlies were anesthetized with CO2, their wings were removed, and theireyes and thorax were glued with three-component dental cement (ProtempII, 3M ESPE Dental Products) to a thin plastic coverslip covered with thinpolyethylene foil. A hole was cut in the foil and head cuticle in a droplet ofEste’s Ringers solution (130 mM NaCl, 5 mM KCl, 2 mM MgCl2, 2 mMCaCl2, 5 mM Hepes, 36 mM sucrose). Trachea and fatty tissue were re-moved to reveal the underlying brain. The preparation was then mountedonto a wide-field upright microscope (BX51WI, Olympus) equipped witha 20× objective (XLUMPLFL20xW, numerical aperture 0.95, Olympus)and an electron-multiplying charge-coupled device camera (Andor iXonDV885, BFi OPTiLAS) connected through a software interface (Cell^R,
w
Olympus) to a Xenon fluorescent excitation source and filter wheel (MT-20,Olympus). GCaMP3.NLS was imaged with 470/40-nm excitation and525/50-nm emission filters (AHFAnalysentechnik). GCaMP3 showed someexponential decay in emission fluorescence at the start of imaging, but re-covered fluorescence after several seconds in the dark, indicating that thefluorescence decrease results from photoisomerization (reversible bleach-ing) but not from permanent bleaching. For this reason, all experimentswere performed at a constant exposure (15 to 30 ms with 2 × 2 binning)and imaging rate (2 Hz), and stimulations were commenced after baselineintensities had stabilized. All images were corrected for background fluo-rescence using a measurement from the same image in a region devoid ofdetectable recombinant fluorescent protein. Quantitative data using GCaMP3are presented for each region of interest as DF/F0 = (F − F0)/F0, whereF represents the background-subtracted emission fluorescence intensity ofGCaMP3 and F0 represents the baseline F measured before each stimu-lation series. In vivo imaging was analyzed with Fiji where necessary,using the StackReg plugin to realign images to reduce artifacts due to res-piratory movement. Electrical shocks (10 to 100 µA for 1.5 s, repeatedevery 5 s for 1 min) generated by an isolated pulse stimulator (AM SystemsModel 2100, Science Products GmbH) in constant current mode were de-livered to the fly’s feet through a copper grid brought into contact with thefeet of the fly with a micromanipulator (Narishige NMN-25, ScienceProducts GmbH).
Data analysisIn all conditioning experiments, the performance indices were subjected toa one-way analysis of variance (ANOVA) followed by post hoc analysiswith Tukey’s test. Statistical significance is indicated by asterisks. All datawere presented as means ± SEM. Statistical analysis of other data is de-scribed in the figure legends.
SUPPLEMENTARY MATERIALSwww.sciencesignaling.org/cgi/content/full/6/274/ra33/DC1Fig. S1. Confocal analysis of the nuclear localization of GCaMP3.NLS in vivo and incultured cells.Fig. S2. Schematic diagram of the in vivo calcium imaging setup.Fig. S3. Characterization of nuclear Ca2+/CaM inhibitor rAAV-2xM13.NLS and thescrambled control rAAV-S2mM13.NLS in cultured mouse hippocampal neurons.Fig. S4. Confocal images of fly brains expressing S2mM13.NLS and 2xM13.NLS.Fig. S5. Flies overexpressing the cAMP phosphodiesterase–encoding gene (dunce)under the control of a heat shock promoter show impaired STM.Fig. S6. Confocal images of fly brains expressing CaMBP4 and H2Av-GFP in the gneurons of the mushroom bodies.
REFERENCES AND NOTES1. G. E. Hardingham, H. Bading, Synaptic versus extrasynaptic NMDA receptor
signalling: Implications for neurodegenerative disorders. Nat. Rev. Neurosci. 11,682–696 (2010).
2. R. Menzel, U. Muller, Learning and memory in honeybees: From behavior to neuralsubstrates. Annu. Rev. Neurosci. 19, 379–404 (1996).
3. R. L. Davis, Olfactory memory formation in Drosophila: From molecular to systemsneuroscience. Annu. Rev. Neurosci. 28, 275–302 (2005).
4. M. Heisenberg, Mushroom body memoir: From maps to models. Nat. Rev. Neurosci.4, 266–275 (2003).
5. P. L. Greer, M. E. Greenberg, From synapse to nucleus: Calcium-dependent genetranscription in the control of synapse development and function. Neuron 59,846–860 (2008).
6. E. R. Kandel, The molecular biology of memory storage: A dialogue between genesand synapses. Science 294, 1030–1038 (2001).
7. A. M. Hagenston, H. Bading, Calcium signaling in synapse-to-nucleus communication.Cold Spring Harb. Perspect. Biol. 3, a004564 (2011).
8. G. J. Augustine, F. Santamaria, K. Tanaka, Local calcium signaling in neurons.Neuron 40, 331–346 (2003).
ww.SCIENCESIGNALING.org 7 May 2013 Vol 6 Issue 274 ra33 10

R E S E A R C H A R T I C L E
on May 8, 2013
stke.sciencemag.org
Dow
nloaded from
9. T. R. Soderling, CaM-kinases: Modulators of synaptic plasticity. Curr. Opin. Neurobiol.10, 375–380 (2000).
10. C. R. Raymond, S. J. Redman, Spatial segregation of neuronal calcium signalsencodes different forms of LTP in rat hippocampus. J. Physiol. 570, 97–111(2006).
11. G. E. Hardingham, F. J. Arnold, H. Bading, Nuclear calcium signaling controls CREB-mediated gene expression triggered by synaptic activity. Nat. Neurosci. 4, 261–267(2001).
12. G. E. Hardingham, S. Chawla, C. M. Johnson, H. Bading, Distinct functions of nuclearand cytoplasmic calcium in the control of gene expression. Nature 385, 260–265(1997).
13. C. P. Bengtson, H. E. Freitag, J. M. Weislogel, H. Bading, Nuclear calcium sensorsreveal that repetition of trains of synaptic stimuli boosts nuclear calcium signaling inCA1 pyramidal neurons. Biophys. J. 99, 4066–4077 (2010).
14. S. J. Zhang, M. Zou, L. Lu, D. Lau, D. A. Ditzel, C. Delucinge-Vivier, Y. Aso, P. Descombes,H. Bading, Nuclear calcium signaling controls expression of a large gene pool: Identificationof a gene program for acquired neuroprotection induced by synaptic activity. PLoSGenet. 5,e1000604 (2009).
15. N. Ho, J. A. Liauw, F. Blaeser, F. Wei, S. Hanissian, L. M. Muglia, D. F. Wozniak,A. Nardi, K. L. Arvin, D. M. Holtzman, D. J. Linden, M. Zhuo, L. J. Muglia, T. A. Chatila,Impaired synaptic plasticity and cAMP response element-binding protein activation inCa2+/calmodulin-dependent protein kinase type IV/Gr-deficient mice. J. Neurosci. 20,6459–6472 (2000).
16. H. Kang, L. D. Sun, C. M. Atkins, T. R. Soderling, M. A. Wilson, S. Tonegawa, Animportant role of neural activity-dependent CaMKIV signaling in the consolidation oflong-term memory. Cell 106, 771–783 (2001).
17. K. Limbäck-Stokin, E. Korzus, R. Nagaoka-Yasuda, M. Mayford, Nuclear calcium/calmodulinregulates memory consolidation. J. Neurosci. 24, 10858–10867 (2004).
18. D. Mauceri, H. E. Freitag, A. M. Oliveira, C. P. Bengtson, H. Bading, Nuclear calcium-VEGFD signaling controls maintenance of dendrite arborization necessary formemory formation. Neuron 71, 117–130 (2011).
19. T. J. Ribar, R. M. Rodriguiz, L. Khiroug, W. C. Wetsel, G. J. Augustine, A. R. Means,Cerebellar defects in Ca2+/calmodulin kinase IV-deficient mice. J. Neurosci. 20,RC107 (2000).
20. M. Simonetti, A. M. Hagenston, D. Vardeh, H. E. Freitag, D. Mauceri, J. Lu, V. P. Satagopam,R. Schneider, M. Costigan, H. Bading, R. Kuner, Nuclear calcium signaling in spinal neuronsdrives a genomic program required for persistent inflammatory pain. Neuron 77, 43–57(2013).
21. F. Wei, C. S. Qiu, J. Liauw, D. A. Robinson, N. Ho, T. Chatila, M. Zhuo, Calciumcalmodulin-dependent protein kinase IV is required for fear memory. Nat. Neurosci.5, 573–579 (2002).
22. S. J. Zhang, M. N. Steijaert, D. Lau, G. Schütz, C. Delucinge-Vivier, P. Descombes,H. Bading, Decoding NMDA receptor signaling: Identification of genomic programsspecifying neuronal survival and death. Neuron 53, 549–562 (2007).
23. S. Papadia, P. Stevenson, N. R. Hardingham, H. Bading, G. E. Hardingham, NuclearCa2+ and the cAMP response element-binding protein family mediate a late phase ofactivity-dependent neuroprotection. J. Neurosci. 25, 4279–4287 (2005).
24. S. J. Zhang, B. Buchthal, D. Lau, S. Hayer, O. Dick, M. Schwaninger, R. Veltkamp,M. Zou, U. Weiss, H. Bading, A signaling cascade of nuclear calcium-CREB-ATF3activated by synaptic NMDA receptors defines a gene repression module that protectsagainst extrasynaptic NMDA receptor-induced neuronal cell death and ischemicbrain damage. J. Neurosci. 31, 4978–4990 (2011).
25. R. L. Davis, Mushroom bodies and Drosophila learning. Neuron 11, 1–14 (1993).26. J. S. de Belle, M. Heisenberg, Associative odor learning in Drosophila abolished by
chemical ablation of mushroom bodies. Science 263, 692–695 (1994).27. M. Heisenberg, A. Borst, S. Wagner, D. Byers, Drosophila mushroom body mutants
are deficient in olfactory learning. J. Neurogenet. 2, 1–30 (1985).28. S. E. McGuire, P. T. Le, R. L. Davis, The role of Drosophila mushroom body signaling
in olfactory memory. Science 293, 1330–1333 (2001).29. A. Pascual, T. Préat, Localization of long-term memory within the Drosophila mushroom
body. Science 294, 1115–1117 (2001).30. T. Zars, M. Fischer, R. Schulz, M. Heisenberg, Localization of a short-term memory in
Drosophila. Science 288, 672–675 (2000).31. Y. Dudai, Y. N. Jan, D. Byers, W. G. Quinn, S. Benzer, dunce, a mutant of Drosophila
deficient in learning. Proc. Natl. Acad. Sci. U.S.A. 73, 1684–1688 (1976).32. W. G. Quinn, W. A. Harris, S. Benzer, Conditioned behavior in Drosophila melanogaster.
Proc. Natl. Acad. Sci. U.S.A. 71, 708–712 (1974).33. T. Tully, W. G. Quinn, Classical conditioning and retention in normal and mutant
Drosophila melanogaster. J. Comp. Physiol. A 157, 263–277 (1985).34. Z. Mao, R. L. Davis, Eight different types of dopaminergic neurons innervate the
Drosophila mushroom body neuropil: Anatomical and physiological heterogeneity.Front. Neural Circuits 3, 5 (2009).
35. T. Riemensperger, T. Völler, P. Stock, E. Buchner, A. Fiala, Punishment prediction bydopaminergic neurons in Drosophila. Curr. Biol. 15, 1953–1960 (2005).
w
36. T. Tully, T. Preat, S. C. Boynton, M. Del Vecchio, Genetic dissection of consolidatedmemory in Drosophila. Cell 79, 35–47 (1994).
37. S. Xia, T. Miyashita, T. F. Fu, W. Y. Lin, C. L. Wu, L. Pyzocha, I. R. Lin, M. Saitoe,T. Tully, A. S. Chiang, NMDA receptors mediate olfactory learning and memory inDrosophila. Curr. Biol. 15, 603–615 (2005).
38. C. L. Wu, S. Xia, T. F. Fu, H. Wang, Y. H. Chen, D. Leong, A. S. Chiang, T. Tully,Specific requirement of NMDA receptors for long-term memory consolidation inDrosophila ellipsoid body. Nat. Neurosci. 10, 1578–1586 (2007).
39. T. Miyashita, Y. Oda, J. Horiuchi, J. C. Yin, T. Morimoto, M. Saitoe, Mg2+ block ofDrosophila NMDA receptors is required for long-term memory formation andCREB-dependent gene expression. Neuron 74, 887–898 (2012).
40. S. I. Ashraf, A. L. McLoon, S. M. Sclarsic, S. Kunes, Synaptic protein synthesisassociated with memory is regulated by the RISC pathway in Drosophila. Cell 124,191–205 (2006).
41. L. C. Griffith, L. M. Verselis, K. M. Aitken, C. P. Kyriacou, W. Danho, R. J. Greenspan,Inhibition of calcium/calmodulin-dependent protein kinase in Drosophila disruptsbehavioral plasticity. Neuron 10, 501–509 (1993).
42. D. B. Akalal, D. Yu, R. L. Davis, A late-phase, long-term memory trace forms in theg neurons of Drosophila mushroom bodies after olfactory classical conditioning.J. Neurosci. 30, 16699–16708 (2010).
43. M. S. Livingstone, P. P. Sziber, W. G. Quinn, Loss of calcium/calmodulin respon-siveness in adenylate cyclase of rutabaga, a Drosophila learning mutant. Cell 37,205–215 (1984).
44. E. M. Skoulakis, D. Kalderon, R. L. Davis, Preferential expression in mushroombodies of the catalytic subunit of protein kinase A and its role in learning and memory.Neuron 11, 197–208 (1993).
45. J. C. Yin, J. S. Wallach, M. Del Vecchio, E. L. Wilder, H. Zhou, W. G. Quinn, T. Tully,Induction of a dominant negative CREB transgene specifically blocks long-termmemory in Drosophila. Cell 79, 49–58 (1994).
46. D. Yu, D. B. Akalal, R. L. Davis, Drosophila a/b mushroom body neurons form abranch-specific, long-term cellular memory trace after spaced olfactory conditioning.Neuron 52, 845–855 (2006).
47. R. L. Davis, Traces of Drosophila memory. Neuron 70, 8–19 (2011).48. D. B. Akalal, D. Yu, R. L. Davis, The long-term memory trace formed in the Drosophila
a/b mushroom body neurons is abolished in long-term memory mutants. J. Neurosci.31, 5643–5647 (2011).
49. P. Y. Plaçais, S. Trannoy, G. Isabel, Y. Aso, I. Siwanowicz, G. Belliart-Guérin, P. Vernier,S. Birman, H. Tanimoto, T. Preat, Slow oscillations in two pairs of dopaminergic neuronsgate long-term memory formation in Drosophila. Nat. Neurosci. 15, 592–599 (2012).
50. C. Liu, P. Y. Plaçais, N. Yamagata, B. D. Pfeiffer, Y. Aso, A. B. Friedrich, I. Siwanowicz,G. M. Rubin, T. Preat, H. Tanimoto, A subset of dopamine neurons signals reward forodour memory in Drosophila. Nature 488, 512–516 (2012).
51. L. Tian, S. A. Hires, T. Mao, D. Huber, M. E. Chiappe, S. H. Chalasani, L. Petreanu,J. Akerboom, S. A. McKinney, E. R. Schreiter, C. I. Bargmann, V. Jayaraman, K. Svoboda,L. L. Looger, Imaging neural activity in worms, flies and mice with improved GCaMPcalcium indicators. Nat. Methods 6, 875–881 (2009).
52. T. Kanesaki, C. M. Edwards, U. S. Schwarz, J. Grosshans, Dynamic ordering ofnuclei in syncytial embryos: A quantitative analysis of the role of cytoskeletal networks.Integr. Biol. 3, 1112–1119 (2011).
53. S. V. Costes, D. Daelemans, E. H. Cho, Z. Dobbin, G. Pavlakis, S. Lockett, Automaticand quantitative measurement of protein-protein colocalization in live cells. Biophys. J.86, 3993–4003 (2004).
54. H. Aberle, A. P. Haghighi, R. D. Fetter, B. D. McCabe, T. R. Magalhães, C. S. Goodman,wishful thinking encodes a BMP type II receptor that regulates synaptic growth inDrosophila. Neuron 33, 545–558 (2002).
55. C. M. Schuster, G. W. Davis, R. D. Fetter, C. S. Goodman, Genetic dissection ofstructural and functional components of synaptic plasticity. I. Fasciclin II controlssynaptic stabilization and growth. Neuron 17, 641–654 (1996).
56. M. Schwaerzel, M. Monastirioti, H. Scholz, F. Friggi-Grelin, S. Birman, M. Heisenberg,Dopamine and octopamine differentiate between aversive and appetitive olfactorymemories in Drosophila. J. Neurosci. 23, 10495–10502 (2003).
57. A. Claridge-Chang, R. D. Roorda, E. Vrontou, L. Sjulson, H. Li, J. Hirsh, G. Miesenböck,Writing memories with light-addressable reinforcement circuitry. Cell 139, 405–415(2009).
58. C. G. Galizia, W. Rossler, Parallel olfactory systems in insects: Anatomy and function.Annu. Rev. Entomol. 55, 399–420 (2010).
59. J. P. Martin, A. Beyerlein, A. M. Dacks, C. E. Reisenman, J. A. Riffell, H. Lei,J. G. Hildebrand, The neurobiology of insect olfaction: Sensory processing in acomparative context. Prog. Neurobiol. 95, 427–447 (2011).
60. J. Wang, B. Campos, G. A. Jamieson Jr., M. A. Kaetzel, J. R. Dedman, Functionalelimination of calmodulin within the nucleus by targeted expression of an inhibitorpeptide. J. Biol. Chem. 270, 30245–30248 (1995).
61. J. Nakai, M. Ohkura, K. Imoto, A high signal-to-noise Ca2+ probe composed of asingle green fluorescent protein. Nat. Biotechnol. 19, 137–141 (2001).
ww.SCIENCESIGNALING.org 7 May 2013 Vol 6 Issue 274 ra33 11

R E S E A R C H A R T I C L E
on May
stke.sciencemag.org
Dow
nloaded from
62. B. Perazzona, G. Isabel, T. Preat, R. L. Davis, The role of cAMP responseelement-binding protein in Drosophila long-term memory. J. Neurosci. 24, 8823–8828(2004).
63. M. J. Krashes, S. Waddell, Rapid consolidation to a radish and protein synthesis-dependent long-term memory after single-session appetitive olfactory conditioningin Drosophila. J. Neurosci. 28, 3103–3113 (2008).
64. U. S. Cheung, A. J. Shayan, G. L. Boulianne, H. L. Atwood, Drosophila larval neuro-muscular junction’s responses to reduction of cAMP in the nervous system. J. Neurobiol.40, 1–13 (1999).
65. Y. Aso, K. Grübel, S. Busch, A. B. Friedrich, I. Siwanowicz, H. Tanimoto, The mush-room body of adult Drosophila characterized by GAL4 drivers. J. Neurogenet. 23,156–172 (2009).
66. S. E. McGuire, P. T. Le, A. J. Osborn, K. Matsumoto, R. L. Davis, Spatiotemporalrescue of memory dysfunction in Drosophila. Science 302, 1765–1768 (2003).
67. C. P. Bengtson, H. Bading, Nuclear calcium signaling. Adv. Exp. Med. Biol. 970,377–405 (2012).
68. E. E. Corcoran, A. R. Means, Defining Ca2+/calmodulin-dependent protein ki-nase cascades in transcriptional regulation. J. Biol. Chem. 276, 2975–2978(2001).
69. G. E. Hardingham, S. Chawla, F. H. Cruzalegui, H. Bading, Control of recruitment andtranscription-activating function of CBP determines gene regulation by NMDA receptorsand L-type calcium channels. Neuron 22, 789–798 (1999).
70. S. Chawla, G. E. Hardingham, D. R. Quinn, H. Bading, CBP: A signal-regulated tran-scriptional coactivator controlled by nuclear calcium and CaM kinase IV. Science 281,1505–1509 (1998).
71. S. Impey, A. L. Fong, Y. Wang, J. R. Cardinaux, D. M. Fass, K. Obrietan, G. A. Wayman,D. R. Storm, T. R. Soderling, R. H. Goodman, Phosphorylation of CBP mediatestranscriptional activation by neural activity and CaM kinase IV. Neuron 34, 235–244(2002).
72. B. Buchthal, D. Lau, U. Weiss, J. M. Weislogel, H. Bading, Nuclear calcium signalingcontrols methyl-CpG-binding protein 2 (MeCP2) phosphorylation on serine 421following synaptic activity. J. Biol. Chem. 287, 30967–30974 (2012).
73. A. M. Oliveira, T. J. Hemstedt, H. Bading, Rescue of aging-associated decline inDnmt3a2 expression restores cognitive abilities. Nat. Neurosci. 15, 1111–1113(2012).
74. D. Yu, A. Ponomarev, R. L. Davis, Altered representation of the spatial code for odorsafter olfactory classical conditioning; memory trace formation by synaptic recruitment.Neuron 42, 437–449 (2004).
75. A. L. Blum, W. Li, M. Cressy, J. Dubnau, Short- and long-term memory inDrosophila require cAMP signaling in distinct neuron types. Curr. Biol. 19, 1341–1350(2009).
76. D. Yu, A. C. Keene, A. Srivatsan, S. Waddell, R. L. Davis, Drosophila DPM neuronsform a delayed and branch-specific memory trace after olfactory classical conditioning.Cell 123, 945–957 (2005).
77. Y. Wang, A. Mamiya, A. S. Chiang, Y. Zhong, Imaging of an early memory trace in theDrosophila mushroom body. J. Neurosci. 28, 4368–4376 (2008).
w
78. C. C. Chen, J. K. Wu, H. W. Lin, T. P. Pai, T. F. Fu, C. L. Wu, T. Tully, A. S. Chiang,Visualizing long-term memory formation in two neurons of the Drosophila brain. Science335, 678–685 (2012).
79. D. Comas, F. Petit, T. Preat, Drosophila long-term memory formation involves regulationof cathepsin activity. Nature 430, 460–463 (2004).
80. R. S. Lewis, Calcium signaling mechanisms in T lymphocytes. Annu. Rev. Immunol.19, 497–521 (2001).
81. M. Oh-hora, A. Rao, Calcium signaling in lymphocytes. Curr. Opin. Immunol. 20,250–258 (2008).
82. G. E. Oldroyd, J. A. Downie, Nuclear calcium changes at the core of symbiosissignalling. Curr. Opin. Plant Biol. 9, 351–357 (2006).
83. M. Klugmann, C. W. Symes, C. B. Leichtlein, B. K. Klaussner, J. Dunning, D. Fong,D. Young, M. J. During, AAV-mediated hippocampal expression of short and longHomer 1 proteins differentially affect cognition and seizure activity in adult rats.Mol. Cell. Neurosci. 28, 347–360 (2005).
84. T. Lee, L. Luo, Mosaic analysis with a repressible cell marker for studies of genefunction in neuronal morphogenesis. Neuron 22, 451–461 (1999).
85. A. Fiala, T. Spall, In vivo calcium imaging of brain activity in Drosophila by transgeniccameleon expression. Sci. STKE 2003, PL6 (2003).
86. H. Bading, M. E. Greenberg, Stimulation of protein tyrosine phosphorylation by NMDAreceptor activation. Science 253, 912–914 (1991).
Acknowledgments: We are grateful to D. Lau for the construction of the expressionvectors pIE-GCaMP3.NLS and pIE-Btg2-mCherry.NES. We thank A. Fiala for helpfuldiscussions, I. Bünzli-Ehret for culture preparations, R. Geiger and M. Knirr for supportwith the transgenic fly lines, C. Schäfer for technical support with fly dissections,M. Müller for assistance in conditioning trials, and M. Müller for HI5 cell lines. We thankY. Zhong, J. Grosshans, H. L. Atwood, and L. L. Looger for providing fly lines. Funding:This work was supported by the Alexander von Humboldt Foundation (Wolfgang PaulPrize to H.B.), an ERC Advanced Grant (to H.B.), and the Sonderforschungsbereich(SFB) 636 of the Deutsche Forschungsgemeinschaft (to C.M.S.). C.M.S. and H.B. aremembers of the Excellence Cluster CellNetworks at Heidelberg University. Authorcontributions: H.B. and C.M.S. conceived the study; J.-M.W., C.P.B., C.M.S., and H.B.designed the experiments; J.-M.W., C.P.B., M.K.M., and J.N.H. performed the experiments;J.-M.W. designed and cloned the plasmids for transgenic flies; J.-M.W. and M.B. characterizedprotein expression; J.-M.W., C.P.B., J.N.H., C.M.S., and H.B. analyzed and interpreted thedata; J.-M.W., C.P.B., C.M.S., and H.B. wrote the paper. Competing interests: The authorsdeclare that they have no competing interests.
Submitted 7 September 2012Accepted 9 April 2013Final Publication 7 May 201310.1126/scisignal.2003598Citation: J.-M. Weislogel, C. P. Bengtson, M. K. Müller, J. N. Hörtzsch, M. Bujard,C. M. Schuster, H. Bading, Requirement for nuclear calcium signaling in Drosophilalong-term memory. Sci. Signal. 6, ra33 (2013).
8,ww.SCIENCESIGNALING.org 7 May 2013 Vol 6 Issue 274 ra33 12
2013
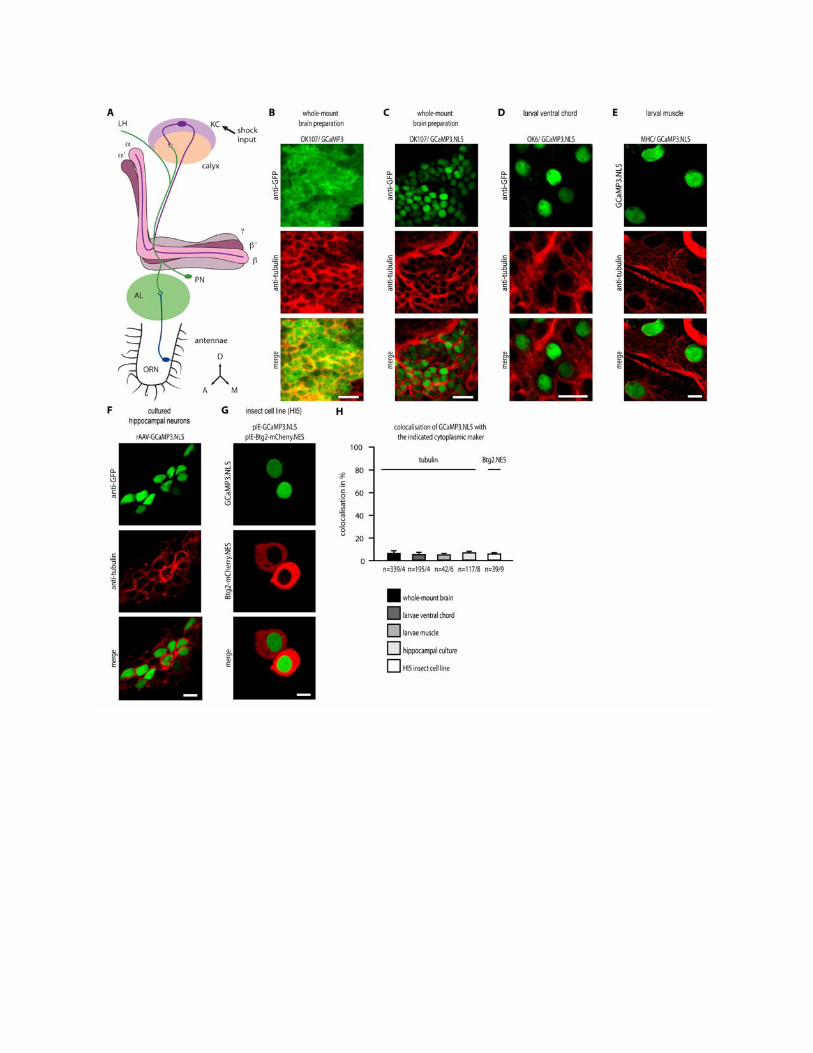

Fig. S1. Confocal analysis of the nuclear localization of GCaMP3.NLS in vivo and in cultured cells. (A) Simplified schematic representation focusing on the Drosophila mushroom bodies (purple, gray, peach) and their olfactory inputs. See introduction for details. Abbreviations: Olfactory receptor neuron (ORN), projection neuron (PN), antennal lobe (AL), mushroom body Kenyon cell (KC) and lobes (, ’, ’, ), lateral horn (LH), anterior (A), dorsal (D), medial (M). (B, C) Confocal images of whole-mount preparations of fly brains expressing GCaMP3 (OK107/ GCaMP3) or GCaMP3.NLS (OK107/ GCaMP3.NLS) in mushroom body neurons. The upper row shows immunolabeling with an antibody against GFP (green) of the recombinant calcium indicators GCaMP3 (B) visible in the cytoplasm and the nucleus and GCaMP3.NLS (C) visible exclusively in the nucleus. Flies were counterstained with an antibody against the cytoplasmic marker tubulin (red). Merged images in the lower panels show colocalized signal in yellow. The scale bar is 10 µm. (D) Confocal images of motor neurons in the 3rd instar larval ventral chord expressing GCaMP3.NLS (OK6/ GCaMP3.NLS) and immunolabeled with an antibody against GFP (green) and an antibody against tubulin (red). Merged imaged shows the lack of colocalization. The scale bar is 10 µm. (E) Confocal images of muscle cells in the 3rd instar larvae expressing GCaMP3.NLS (MHC/ GCaMP3.NLS) detected with an antibody against GFP (green) and tubulin (red). Merged imaged shows the lack of colocalization. The scale bar is 10 µm. (F) Confocal images of cultured hippocampal neurons from mouse infected with AAV-GCaMP3.NLS. GCaMP3.NLS staining is green (detected with by immunolabeling with an antibody against GFP) and tubulin is red. Merged imaged shows the lack of colocalization. The scale bar is 10 µm. (G) Confocal images of the insect cell line HI5 cotransfected with pIE-GCaMP3.NLS (green) and pIE-Btg2-mCherry.NES (red). Merged imaged shows the lack of colocalization. The scale bar is 10 µm. (H) Quantitative analysis of the percentage of the GCaMP3.NLS fluorescence signal colocalizing with the cytoplasmic markers tubulin or Btg2.NES in each of the experiments shown in panels C to G. Costes technique for automatic thresholding was used. N values below each bar indicate the number of cells / the number of flies or larvae or coverslips analyzed.

Fig. S2. Schematic diagram of the in vivo calcium imaging setup. In vivo calcium imaging was performed with a wide field fluorescence microscope through an opening in the head cuticle of flies whose heads were glued to a coverslip and whose feet were in contact with a copper grid that delivered the electric shocks. Foot shocks consisted of repeated electric pulses (13 × 1.5 sec 10 µA pulses at 0.2 Hz). The fly’s antennae received a constant stream of air passed through a vial containing either mineral oil alone or 10% OCT or MCH in mineral oil.
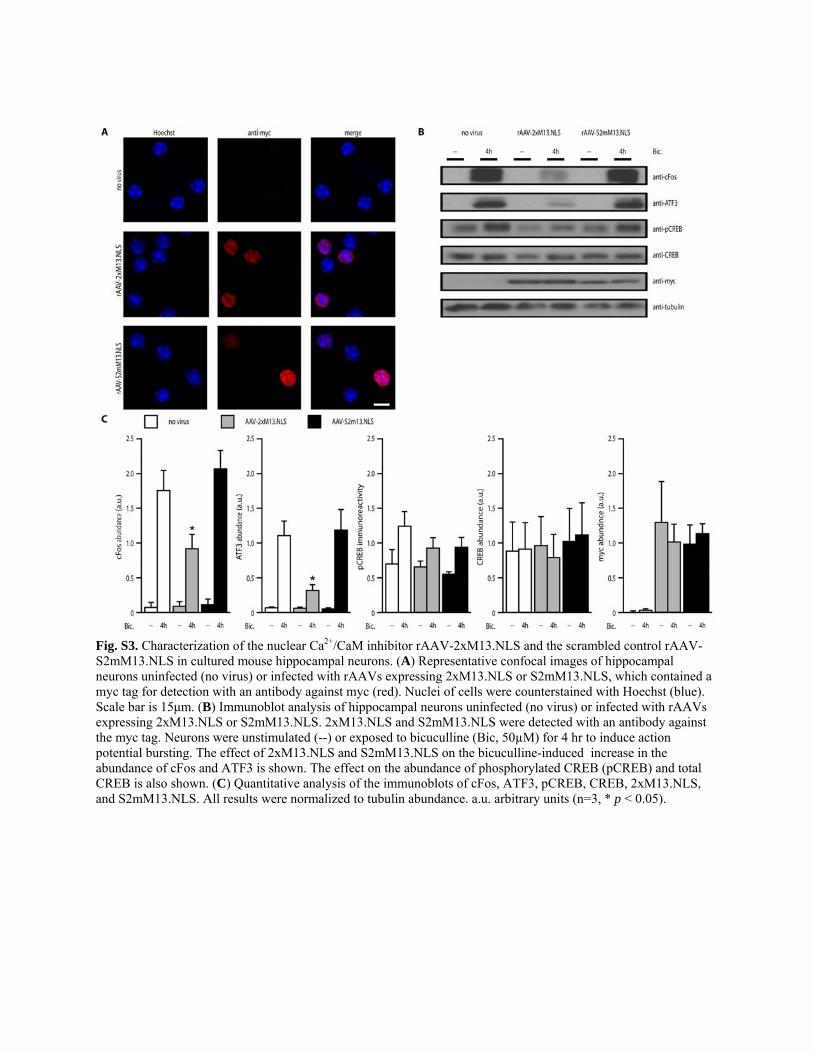
Fig. S3. Characterization of the nuclear Ca2+/CaM inhibitor rAAV-2xM13.NLS and the scrambled control rAAV-S2mM13.NLS in cultured mouse hippocampal neurons. (A) Representative confocal images of hippocampal neurons uninfected (no virus) or infected with rAAVs expressing 2xM13.NLS or S2mM13.NLS, which contained a myc tag for detection with an antibody against myc (red). Nuclei of cells were counterstained with Hoechst (blue). Scale bar is 15μm. (B) Immunoblot analysis of hippocampal neurons uninfected (no virus) or infected with rAAVs expressing 2xM13.NLS or S2mM13.NLS. 2xM13.NLS and S2mM13.NLS were detected with an antibody against the myc tag. Neurons were unstimulated (--) or exposed to bicuculline (Bic, 50µM) for 4 hr to induce action potential bursting. The effect of 2xM13.NLS and S2mM13.NLS on the bicuculline-induced increase in the abundance of cFos and ATF3 is shown. The effect on the abundance of phosphorylated CREB (pCREB) and total CREB is also shown. (C) Quantitative analysis of the immunoblots of cFos, ATF3, pCREB, CREB, 2xM13.NLS, and S2mM13.NLS. All results were normalized to tubulin abundance. a.u. arbitrary units (n=3, * p < 0.05).

Fig. S4. Confocal images of fly brains expressing S2mM13.NLS and 2xM13.NLS. Images show anti-myc (red) and nuclear chromatin labeled with Hoechst (blue). Non-linear scaling of Hoechst images was necessary to visualize entire nuclei. The scale bars are 7.5µm.
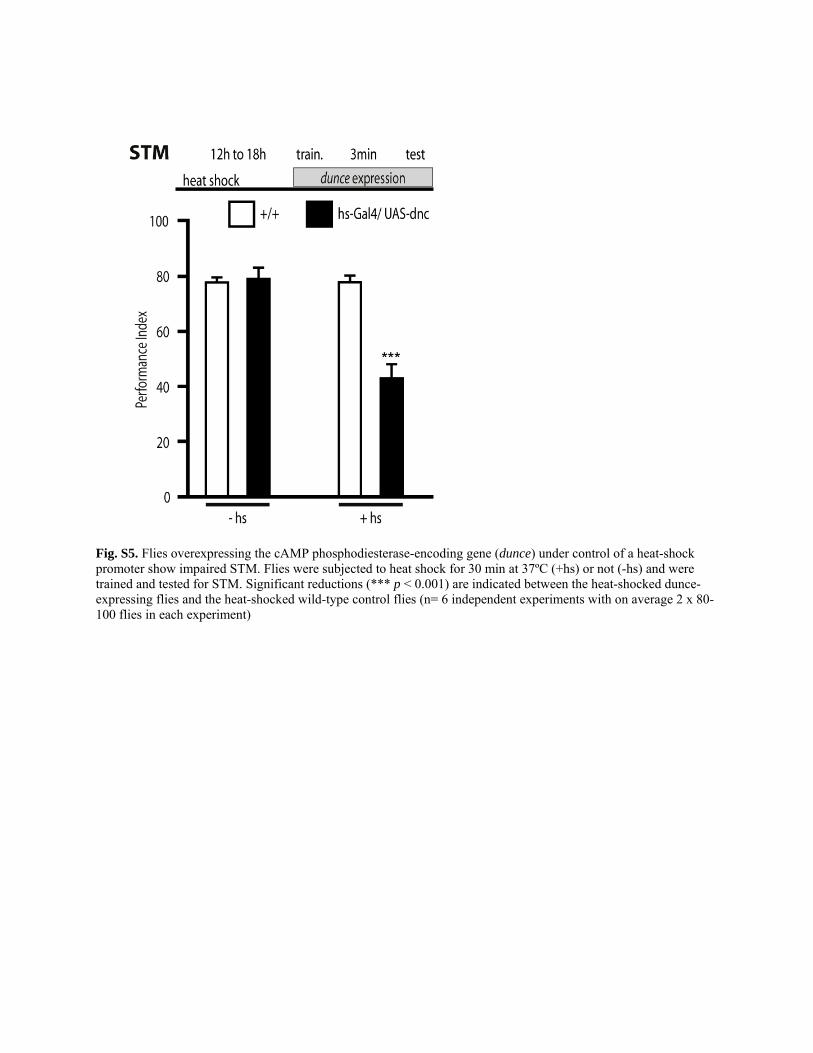
Fig. S5. Flies overexpressing the cAMP phosphodiesterase-encoding gene (dunce) under control of a heat-shock promoter show impaired STM. Flies were subjected to heat shock for 30 min at 37ºC (+hs) or not (-hs) and were trained and tested for STM. Significant reductions (*** p < 0.001) are indicated between the heat-shocked dunce-expressing flies and the heat-shocked wild-type control flies (n= 6 independent experiments with on average 2 x 80-100 flies in each experiment)

Fig. S6. Confocal images of fly brains expressing CaMBP4, H2Av-GFP in the -neurons of the mushroom bodies. The images show 201Y; CaMBP4/ H2Av-GFP fly brains expressing histone2Av-GFP (H2Av-GFP) under control of histon2Av promoter immunolabeled with anti-GFP (green) and either CaMBP4 immunolabeled with anti-myc (red, A) or the cytoplasmic marker tubulin immunolabeled with anti-tubulin (red, B). Scale bars are 10µm.