voltage-gated ca 2+ channels. history fatt & katz (1953) discovered an exception to na aps: –...
TRANSCRIPT

Voltage-Gated Ca2+ Channels

History• Fatt & Katz (1953) discovered an exception to Na APs:– Na substitution made APs stronger!– TEA antagonism spread AP even more (why?)– procaine enhanced excitability!– thus, crustacean muscle doesn’t use Na+ for APs
• “Ca2+ spiking”• ECa even more positive than ENa
• Fatt & Ginsborg– AP requires Ca2+, Sr2+, or Ba2+
• Hagiwara & Naka (1964)– lowering [Ca]in (free) increases response in barnacle muscle– must be <0.1 mM for all-or-nothing response

[Ca]-sensitive Channels?
• some Ca channels are inactivated by [Ca]in > 0.1-1.0 mM• [Ca]in-sensitive K channels also exist, activated by ↑[Ca]in
• both mechanisms (decrease [Ca]in)would serve to generate the barnacle muscle cell experimental results
• No TTX-sensitive Na currents in arthropod muscle fibers, only in axons
• Hagiwara & Naka (1964) also measured 45Ca fluxes, to nail it definitively– 2-6 pmol of Ca2+ per mF of membrane C (only 0.5 needed for
cation to depolarize by 100 mV

Ubiquity of Ca Channels
• permeant divalent cation competition• divalent transition metal blockage (Ni2+, Cd2+, Co2+)• variety of subtypes, via characterization based on
– activation ranges– ion selectivity– sensitivity to blockage– kinetics
• functions– secretion– sarcomeric shortening (contraction)– exclusively for APs in myocytes of arthropods, nematodes,
tunicates, molluscs, and in vert. SmM, vert. CardM

Revelation Delayed
• Ca channels don’t occur on clampable membranes in high density with easy accessibility– nerve terminals, dendrites, infoldings of myocytes– nerve soma, confounded by other currents, e.g. K
• suction pipettes were developed for voltage clamp (precursor of gigaseal [109W] whole cell patch)
• gastropod ganglion cells were used• blockage of INa with TTX or substitution (externally)• blockage of IK with Cs+ or TEA substitution
(internally)

ICa• inward current• can be masked
by INa
• smaller magnitude (SA-normalized): ~40-50 mA/cm2 or 100x less than INa in vert. SkM, axons• slower than INa
why this shape?
Early Nomenclature• High-Voltage Activated (HVA)
– often lack rapid inactivation– require much larger depolarizations to activate than
• Low-Voltage Activated (LVA) Ca channels– usually have rapid Vm-dependent inactivation (measure peaks
for I-V curves)– activate in response to smaller depols
• Recall, classical interaction betweenVm and g• Ca channels have 2 major roles:
– electrical excitation– regulation
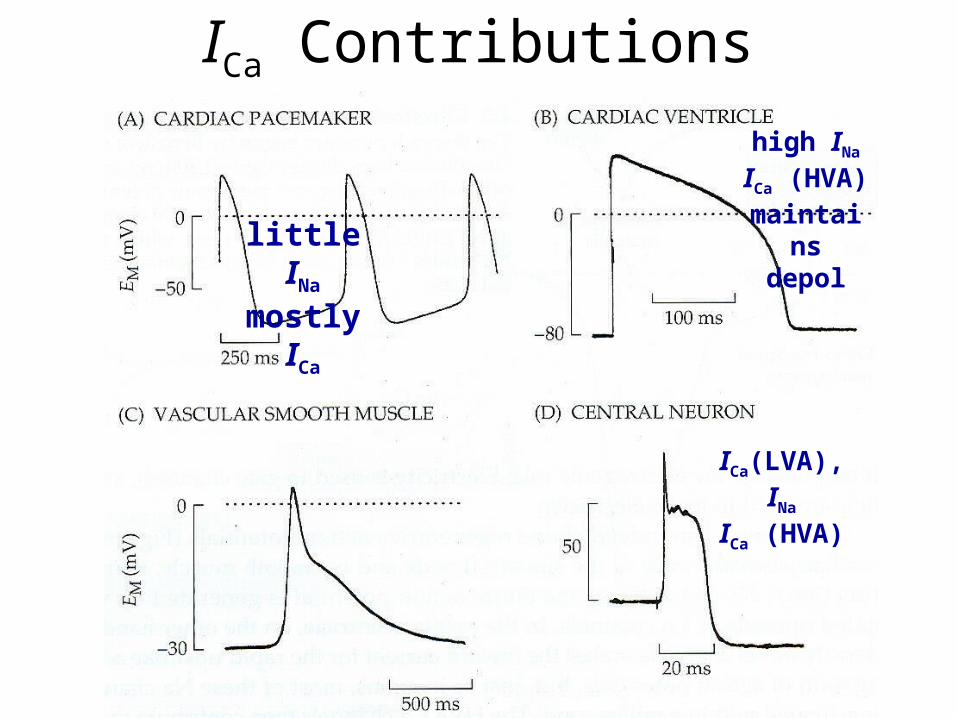
ICa Contributions
little INa
mostly ICa
high INa
ICa (HVA) maintains
depol
ICa(LVA), INa
ICa (HVA)

Ventricular Myocytes• Na+ influx depolarizes cell• Na+ Ch inactivation is not
accompanied by repolarization• plateau potential (~0 mV) is
maintained– K+ chs remain/close– Ca2+ chs open
• Ca2+ chs (Vm-gated) open slowly and remain open long (L-type)
• ICa2+ balances IK+
• repolarization occurs when gK+, gCa2+ return to original state
typical
long-lasting
atypical

Pacemaker Potentials• spontaneous, rhythmical
de/repolarizations• most prominent in SA node
Mechanisms:1. ↓PK+ due to previous AP2. opening of chs at polarized levels
(If): mostly Na+
3. T-type Ca2+ chs open
APs via L-type Ca2+ chs, repolarized after delay by IK+ chs
SA node captures other PMPs
typicalbut slow
Na+

Ca: Excitation & Messenger• neither the nervous,
skeletal muscle, cardiac muscle, nor endocrine systems are a purely electrical systems. They are electro-chemical/mechanical systems.• in addition to its role in
excitation, Ca is an important mediator in both mechanical force generation and cellular physiological responses
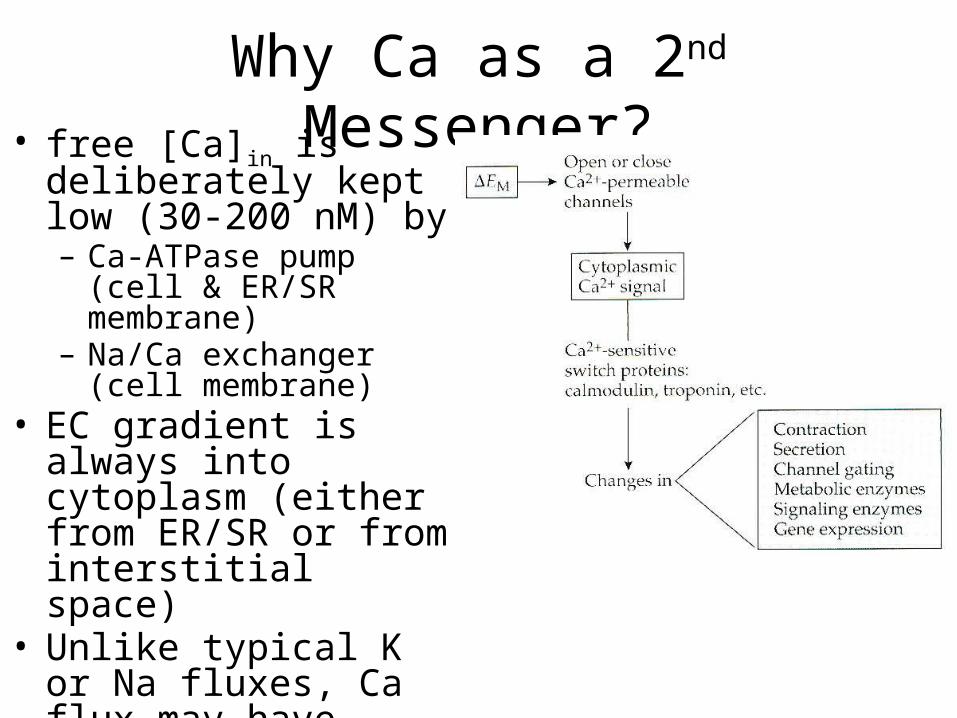
Why Ca as a 2nd Messenger?• free [Ca]in is deliberately
kept low (30-200 nM) by– Ca-ATPase pump (cell &
ER/SR membrane)– Na/Ca exchanger (cell
membrane)• EC gradient is always into
cytoplasm (either from ER/SR or from interstitial space)
• Unlike typical K or Na fluxes, Ca flux may have dramatic effect on [Ca], with huge effects on cellular processes

Biophysical Interests
• Contraction (muscle)• Secretion (neurotransmission, NMJ,
endocrine)• Gating (Ca, K channels)

Muscles• source of Ca is often SR (cardiac/neurons are more
complex, depend on both external and internal sources)• free cytosolic Ca binds to calmodulin, troponin, and other
molecules• this may result in activation (and inactivation) of classes
of enzymes, including kinases, that result in short-term and long-term physiological changes.
• in muscles, Ca binding uncovers myosin binding site on actin, and ATPase portion of actin-myosin complex is free to produce displacement (shortening), the physical underpinning of contraction
• Ca-calmodulin acts on cytoskeletal complexes to influence motility, migration

Synaptic Transmission• AP travels down axon to terminal• Depolarization opens Vm-gated Ca2+ channels (N-type)• Ca2+ flows into terminals, initiates exocytosis of vesicles
(i.e., vesicle fusion with presyn memb)• Prior to this, SNARE proteins interact in vesicle memb,
presynapt memb, and terminal cytoplasm• Once fusion occurs, vesicle contents (neurotrans, cotrans) is
released into synaptic cleft and diffuses to postsyn memb• Ions flow in as a result of NT binding to postsyn receptors.
Opening (1st mess) or shutting (2nd mess).• delay between arrival of AP & D in postsyn Vm ~0.2 ms

Synaptic Transmission1. AP invades axon terminals2. Vm-dep Ca2+ Chs open
3. [Ca2+]in rises
4. synaptic vesicles fuse to plasma memb
5. neurotransmitter released into synaptic cleft
6. NT binds to postsynaptic Rs, opening them
7. Cations or anions flow inThus, electrochemical
communication

Synaptic/NMJ Potentials• are inhibited by
Mg2+, which blocks Ca channels
• Squid giant synapse allowed for Ca imaging, voltage clamping, and injection of Ca directly.
• Can now image and patch onto vertebrate CNS synapses

• presynaptic [Ca] changes are rapid in onset
• buffered slowly• cumulative• Ca must be
pumped actively• at Ca chs near
synaptic vesicles: >100X greater rise and faster onset and decay
• unknown sensor molecules: several vesicle-associated proteins bind Ca

[Ca]in and Gating
• several Ca, K (0.1-0.9 mM), and Cl (0.2-0.8 mM) channels are sensitive to [Ca]in
• so are some non-specific cation channels (1-6 mM)
• NMDA channels conduct Ca2+ (large) and Na+ (smaller)
• all have binding sites for Ca2+ or calmodulin on intracellular surfaces

Why this shape?
Why this effect?
Note: this is spatial average!
Local [Ca]?
Why this effect?
What’s this about?KDR + K(Ca)
What’s the implication here?
why biggesteffect here?
IK

Bottom Line• Many [Ca]in-dependent processes are Vm-
dependent, since many Ca channels are Vm-dependent and Ca will enter from the extracellular space.
• These processes may or may not be Vm-sensitive irrespective of [Ca]in

Electrical Characterization
• Based on Vm-dependence & kinetics– HVA: large depol for act, slow kinetics, no rapid inact– LVA: small depol for act, faster kinetics, rapid Vm-dep
inact
How would you distinguish between them, experimentally, if they were both present in the
same cell?

Separation of HVA & LVA…by Vhold during
voltage-clamp

L- vs. T-type Nomenclature• based on unitary
conductance measurements
• L = long lasting, larger conductances (HVA)
• T= transient, tiny conductances (LVA)
• note: driving force on Ba2+ is less at more depolarized potentials, so currents are even more disproportional

Ca Channel Antagonists• clinical uses:– L-type blockers (cardiac and SmM) such as 1,4-dihydropyridines
(fig. 4.13):• D-600, nifedipine, nitrendipine, diltiazem: 20nM-50mM ½ blocking
– lipid-soluble drugs can block other channel types at high concentrations
• DHPs have complex effects– may ↑ (agonist; BAY K 8644) or ↓ (antagonist; nif., nit.) ICa
– same compound may serve as both, because they contain a mixture of 2 optical enantiomers (isomers)
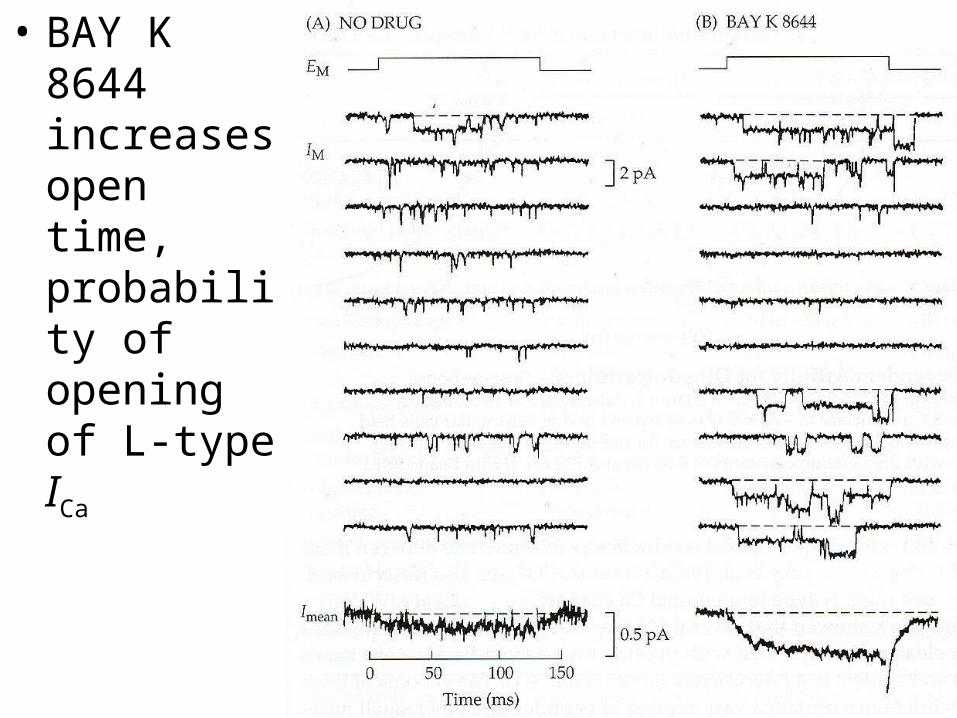
• BAY K 8644 increases open time, probability of opening of L-type ICa

Vm-dependent Antagonism• binding of blockers is Vm-dependent: this is rule
Implication?• ~2000x more nitrendipine is needed to block L-type
channels at -80 mV than at -15 mV (see KD values)
inactivated“deinactivated”

Another Type of Ca Channel• “HVAish” channels have been found on neurons (N-type):
dominant form at presynaptic terminals• resistant to DHP antagonism• intermediate conductances, between T- and L-type• Subtypes can be distinguished by peptide toxins from
Pacific cone snails (Conus) and spiders (e.g., Agelenopsis aperta). Predatory invertebrate venoms have been purified and are commercially available, making channel ID easier.
• Subtypes: N (w-conotoxin GVIA sensitive), P/Q (w-Aga IVA; Purkinje cells), R (resistant), etc. (See Table 4.1 for a summary)

Genetics• transverse tubules in SkM are the first and richest known
source of L-type Ca channels• DHP served as a purification label• short AA seqs from a1 subunit were used to obtain a full-
length cDNA clone– 4 repeats of 6TM segments (including S4 and P regions) in each
repeat
• aux proteins: a2dbg
• cDNA library screens provided at least 10 mammalian variants
• Also, alternative splicing (small exons portions included or excluded), proteolytic cleavage of C-terminal tail, etc.

N

Permeation & Ionic Block• divalent cations strontium, barium, calcium (alkali
earth metals) all permeate Ca channels• Ba2+ yields the largest currents, and blocks K
channels too.• Vmcontrols r and
driving force
• [Ca]out = 104 to105 x [Ca]in, thus, outward ICa is miniscule, even at Vm more positive than ECa (e.g., +200mV). This is due to lack of ions.

‘Open-Channel’I-V for Ca
• model based on diffusion theory
• matches observation
• even though PK/PCa = 1/1000, IK rivals ICa
• this is due to relative scarcity of Ca2+, availability of K+ ions
[K]o = 2 mM[K]i = 100 mM[Ca]o = 2 mM[Ca]i = 100 nMECa = +124 mVreversal potential = observationequilibrium potential = calculated thermodynamicsErev ≠ Eequ! Why not?

More Outrageous Claims!• Even though Ca ions move down their EC gradient,
their movement differs from free diffusion:– ICa does not increase linearly with [Ca]o
– ICa is a saturating function of [Ca]o
– hence, maximum velocity for passing ions, “binding site” theory, and single file conduction: upper limit
• Ca channels become highly permeable to monovalent cations in the absence of divalent ones
• When Ca is present, at least one ion is “bound” within the channel at all times, preventing monovalent ions from entering

Inactivation
• LVAs (e.g., T-type) inactivate rapidly, and with Vm-dependence, while HVAs do not (at least not so quickly)• recall that [Ca]in below 100 nM is needed for maximal
responses, as if high [Ca]in blocks Ca channels• Brehm & Eckert (1978) and Tillotson (1979) remembered this
and suggested that free intracellular Ca blocks the channels somehow!• Thus, HVA function would be self-limiting, inactivating channels
until local [Ca]in is reduced via pumping
• In this scheme, inactivation is [Ca]in-dependent, and only indirectly Vm-dependent.

HVA (L-type) Inactivation• EGTA is a Ca chelator, and
is used as a buffer to keep [Ca]in low (fixed)• Ba2+ serves as a good
current carrier, but poor “inactivator”• little inactivation at very
large depolarizations, near ECa, where there is little ICa

2-Pulse Experiments• extent of inactivation ~
extent of Ca influx during pre-pulse• inactivation is maximal where
ICa is peak, not at peak depolarization
How does this compare to Na inactivation?
Note: this is not true for all Ca channels, but is true for HVA
pre-pulse:ICa = IBa
test pulse:ICa ≠ IBa

Molecular Mechanisms
HVA [Ca]in-dependent inactivation may be mediated by calmodulin molecules tethered to cytoplasmic regions (particularly, C-term) of the channel, bound with high affinity– inactivation disappears with mutation (deletion or
alteration) of calmodulin binding region– inactivation disappears if calmodulin is altered so
as not to bind Ca2+ (x4)

Activation: Delayed and Vm-dep
• rise of peak opening probability can be modeled by a Boltzmann relation with 3-5 gating charges moving across the membrane to control activation
• delay can be modeled by 2nd, 3rd, 5th, or 6th-order function of a probability particle (cf., m3 for Na, n4 for K)
• T-type act/inact has tm and th 20-50X longer than those for Na at same temp
• Closing (deactivation) begins immediately upon repolarization, with single or double exponential time course (tails), sometimes reflecting multiple types of ICa (T-LVA and L-HVA)

Single Channel Kinetics
• stochastic, as usualC ↔ C ↔ O
• opening viewed as 2-step process (e.g., m2), both steps Vm-dep• L-type channels can show sudden changes in gating
kinetics• typical sweeps: many sub-ms openings in 200 ms sweeps• other sweeps: longer channel openings, with k-2 100X slower
than typical sweeps• hence, “mode 0” (no opening) vs. “mode 1” vs. “mode 2”,
switching between the modes controlled at molecular level, and agonists stabilize mode 2, antagonists mode 0/1. ???
k1
k-1
k2
k-2

Review• permeating ions compete for binding sites within
the pore• Ca channels types differ in Vm-dep, inact, etc.• single channel gating described by state models
and chemical kinetics, which may change• drug-receptor sites change gating/mode, in Vm-dep
manner• Ca chs in all excitable cells, 2 roles:
– maintain inward current during prolonged depolarizations (no brisk inactivation)
– serve as ONLY LINK between depolarization and non-electrical activity

• axons: only important at terminals, ????• dendrites: electrical boosting, summation,
spreading of synaptic currents (t, l)• secretory/endocrine glands: slow inact allows for
prolonged depol and release• CardM & SmM: important electrical and
transducing roles during prolonged depol/contraction• Ca messenger (activity surrogate) affecting
replenishment, growth, and gene transcription•more to come!