viral vaccines. introduction the development of antiviral vaccines is highly dependent upon the...
TRANSCRIPT

Viral VaccinesViral Vaccines

IntroductionIntroduction
• The development of antiviral vaccines is highly dependent upon the infection cycle of The development of antiviral vaccines is highly dependent upon the infection cycle of the viral pathogen itself.the viral pathogen itself.
• Viruses that have an extracellular, viremic stage in the infection cycle are accessible to Viruses that have an extracellular, viremic stage in the infection cycle are accessible to soluble humoral components – are susceptible to antibodies.soluble humoral components – are susceptible to antibodies.
• Viruses that are highly cell associated, either through their ability to transport directly Viruses that are highly cell associated, either through their ability to transport directly from one infected cell to an adjacent susceptible host cell, or have a cell-associated from one infected cell to an adjacent susceptible host cell, or have a cell-associated viremic stage, are not accessible to humoral components – require direct recognition of viremic stage, are not accessible to humoral components – require direct recognition of infected cells – implicates T cell responses.infected cells – implicates T cell responses.
• The basis for successful vaccination is the generation of a long-term protective The basis for successful vaccination is the generation of a long-term protective acquired immune response – requires a knowledge of how immunological memory acquired immune response – requires a knowledge of how immunological memory develops and how this can be best implemented.develops and how this can be best implemented.
• There are many different methods that can be used to vaccinate susceptible individuals There are many different methods that can be used to vaccinate susceptible individuals – live attenuated viruses, inactivated or killed viruses, virus-like particles, plasmid DNA, – live attenuated viruses, inactivated or killed viruses, virus-like particles, plasmid DNA, individual viral components, recombinant virus, pseudovirus, replicon – each approach individual viral components, recombinant virus, pseudovirus, replicon – each approach has its advantages and disadvantages.has its advantages and disadvantages.
• The use of non-replicating immunogens as vaccines often requires the use of The use of non-replicating immunogens as vaccines often requires the use of adjuvants to optimize the subsequent immune response.adjuvants to optimize the subsequent immune response.
• Immunization can still result in immune escape variants.Immunization can still result in immune escape variants.

HistoryHistory
• Although early in history the basis of disease was not known, the presence of a life-Although early in history the basis of disease was not known, the presence of a life-long immunity to disease was understood as early as the 4long immunity to disease was understood as early as the 4 thth century. century.
• The first documentation of “immunization” was the process of variolation – the removal The first documentation of “immunization” was the process of variolation – the removal of pus from smallpox lesions and the subsequent scratching of an uninfected person in of pus from smallpox lesions and the subsequent scratching of an uninfected person in the 10the 10thth century in India – variolation did not always work, for reasons unknown. century in India – variolation did not always work, for reasons unknown.
• In 1796, Edward Jenner observed that milk maids exposed to cowpox (vaccinia virus) In 1796, Edward Jenner observed that milk maids exposed to cowpox (vaccinia virus) did not acquire smallpox – he predicted that deliberately infecting an individual with did not acquire smallpox – he predicted that deliberately infecting an individual with vaccinia would protect against smallpox (variola virus) – Sarah Nelmes donated fluid vaccinia would protect against smallpox (variola virus) – Sarah Nelmes donated fluid from her cowpox-infected hands, which was inoculated into James Phipps – produced from her cowpox-infected hands, which was inoculated into James Phipps – produced a lesion similar to cowpox – later challenged James Phipps with fluid from a smallpox a lesion similar to cowpox – later challenged James Phipps with fluid from a smallpox lesion, but no subsequent smallpox developed – this was the first recorded incidence of lesion, but no subsequent smallpox developed – this was the first recorded incidence of “vaccination”.“vaccination”.
• Jenner would be imprisoned for this type of experiment today, but the James Phipps Jenner would be imprisoned for this type of experiment today, but the James Phipps vaccination led to the development of the smallpox vaccine and the eradication of vaccination led to the development of the smallpox vaccine and the eradication of naturally occurring infections worldwide.naturally occurring infections worldwide.

Induction of the B Cell Response andInduction of the B Cell Response andB Cell MemoryB Cell Memory

Primary and Secondary AntibodyPrimary and Secondary AntibodyResponsesResponses

Virus-specific T Cell Responses ~Virus-specific T Cell Responses ~CD4 and CD8 T CellsCD4 and CD8 T Cells
Antiviral CD8Antiviral CD8++ and CD4 and CD4++ T-cell responses. T-cell responses. The three phases of the T-cell immune response (expansion, contraction and memory) are The three phases of the T-cell immune response (expansion, contraction and memory) are indicated. Antigen-specific T cells clonally expand during the first phase in the presence of antigen. Soon after the virus is cleared, the indicated. Antigen-specific T cells clonally expand during the first phase in the presence of antigen. Soon after the virus is cleared, the contraction phase ensues and the number of antigen-specific T cells decreases due to apoptosis. After the contraction phase, the number of contraction phase ensues and the number of antigen-specific T cells decreases due to apoptosis. After the contraction phase, the number of virus-specific T cells stabilizes and can be maintained for great lengths of time (the memory phase). Note that, typically, the magnitude of the virus-specific T cells stabilizes and can be maintained for great lengths of time (the memory phase). Note that, typically, the magnitude of the CD4CD4++ T-cell response is lower than that of the CD8 T-cell response is lower than that of the CD8++ T-cell response, and the contraction phase can be less pronounced than that of CD8 T-cell response, and the contraction phase can be less pronounced than that of CD8++ T T cells. The number of memory CD4cells. The number of memory CD4++ T cells might decline slowly over time. T cells might decline slowly over time.
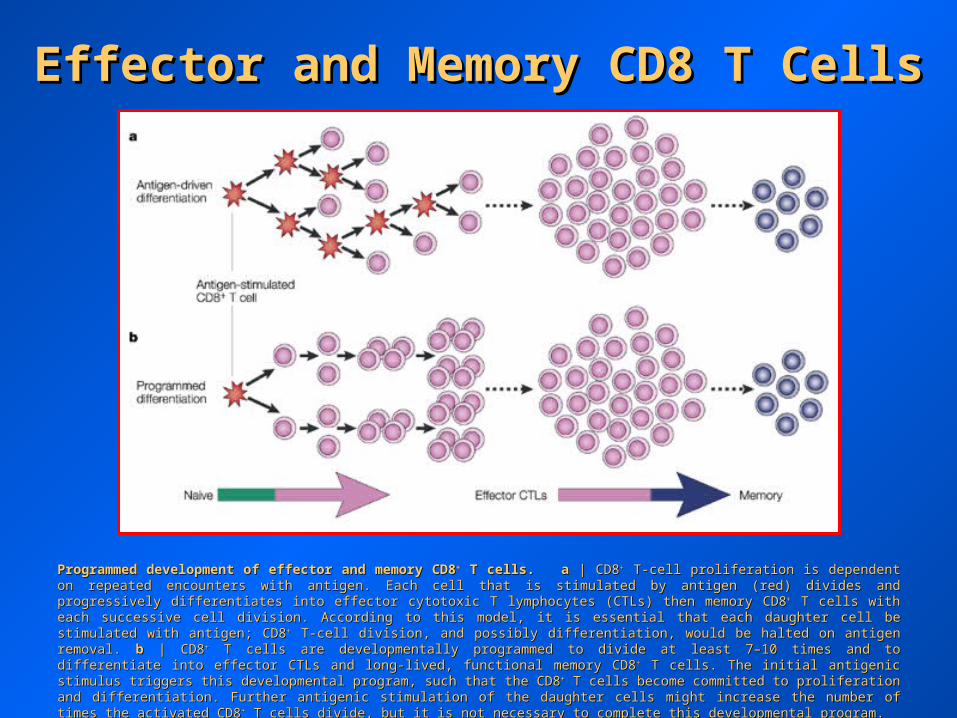
Effector and Memory CD8 T CellsEffector and Memory CD8 T Cells
Programmed development of effector and memory CD8Programmed development of effector and memory CD8++ T cells. T cells. aa | CD8 | CD8++ T-cell proliferation is dependent on repeated encounters with T-cell proliferation is dependent on repeated encounters with antigen. Each cell that is stimulated by antigen (red) divides and progressively differentiates into effector cytotoxic T lymphocytes (CTLs) then antigen. Each cell that is stimulated by antigen (red) divides and progressively differentiates into effector cytotoxic T lymphocytes (CTLs) then memory CD8memory CD8++ T cells with each successive cell division. According to this model, it is essential that each daughter cell be stimulated with T cells with each successive cell division. According to this model, it is essential that each daughter cell be stimulated with antigen; CD8antigen; CD8++ T-cell division, and possibly differentiation, would be halted on antigen removal. T-cell division, and possibly differentiation, would be halted on antigen removal. bb | CD8 | CD8++ T cells are developmentally T cells are developmentally programmed to divide at least 7–10 times and to differentiate into effector CTLs and long-lived, functional memory CD8programmed to divide at least 7–10 times and to differentiate into effector CTLs and long-lived, functional memory CD8++ T cells. The initial T cells. The initial antigenic stimulus triggers this developmental program, such that the CD8antigenic stimulus triggers this developmental program, such that the CD8++ T cells become committed to proliferation and differentiation. Further T cells become committed to proliferation and differentiation. Further antigenic stimulation of the daughter cells might increase the number of times the activated CD8antigenic stimulation of the daughter cells might increase the number of times the activated CD8++ T cells divide, but it is not necessary to T cells divide, but it is not necessary to complete this developmental program. complete this developmental program.

Subsets of memory T cells. Subsets of memory T cells. Several subsets of CD4Several subsets of CD4++ and CD8 and CD8++ T cells based on differential expression of CD45, CD62L and CCR7 have T cells based on differential expression of CD45, CD62L and CCR7 have been characterized from human PBMC populations. The relative frequencies of CD4been characterized from human PBMC populations. The relative frequencies of CD4++ and CD8 and CD8++ T cells expressing such markers from PBMC T cells expressing such markers from PBMC populations of either healthy individuals or those exposed to chronic viral infection are shown. CCR7populations of either healthy individuals or those exposed to chronic viral infection are shown. CCR7++ memory cells are T memory cells are TCMCM; CCR7; CCR7--memory cells memory cells
are Tare TEMEM. In normal individuals, there are very few CD4. In normal individuals, there are very few CD4++CD45RACD45RA++CCR7CCR7-- cells compared with CD8 cells compared with CD8++ T cells. 2°, secondary. T cells. 2°, secondary.
Subsets of Memory T CellsSubsets of Memory T Cells
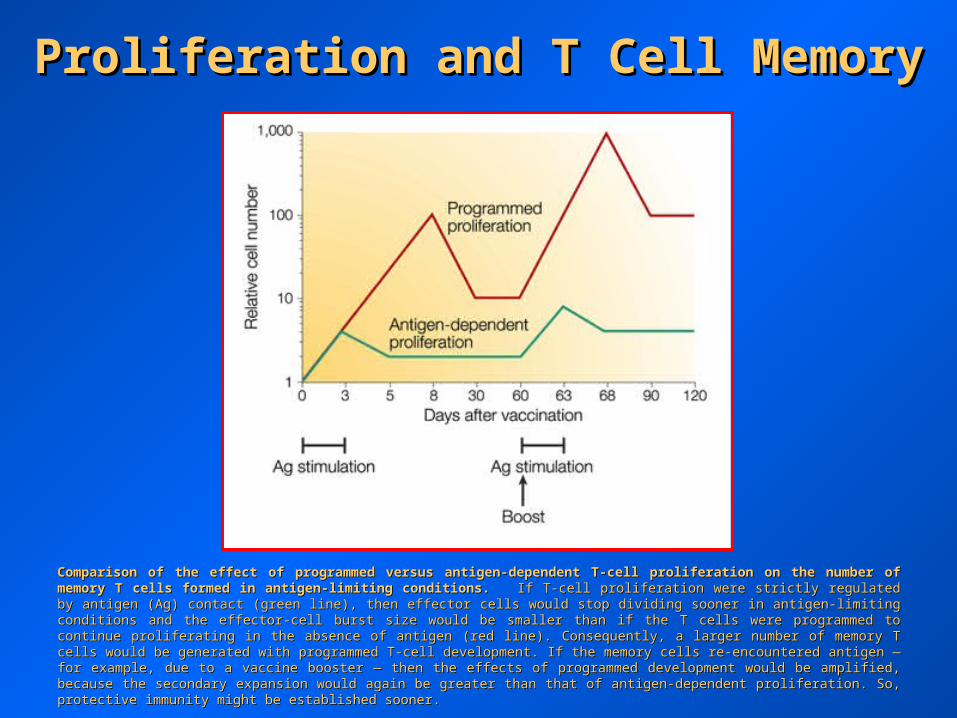
Comparison of the effect of programmed versus antigen-dependent T-cell proliferation on the number of memory T cells formed in Comparison of the effect of programmed versus antigen-dependent T-cell proliferation on the number of memory T cells formed in antigen-limiting conditions.antigen-limiting conditions. If T-cell proliferation were strictly regulated by antigen (Ag) contact (green line), then effector cells would stop If T-cell proliferation were strictly regulated by antigen (Ag) contact (green line), then effector cells would stop dividing sooner in antigen-limiting conditions and the effector-cell burst size would be smaller than if the T cells were programmed to continue dividing sooner in antigen-limiting conditions and the effector-cell burst size would be smaller than if the T cells were programmed to continue proliferating in the absence of antigen (red line). Consequently, a larger number of memory T cells would be generated with programmed T-cell proliferating in the absence of antigen (red line). Consequently, a larger number of memory T cells would be generated with programmed T-cell development. If the memory cells re-encountered antigen — for example, due to a vaccine booster — then the effects of programmed development. If the memory cells re-encountered antigen — for example, due to a vaccine booster — then the effects of programmed development would be amplified, because the secondary expansion would again be greater than that of antigen-dependent proliferation. So, development would be amplified, because the secondary expansion would again be greater than that of antigen-dependent proliferation. So, protective immunity might be established sooner.protective immunity might be established sooner.
Proliferation and T Cell MemoryProliferation and T Cell Memory
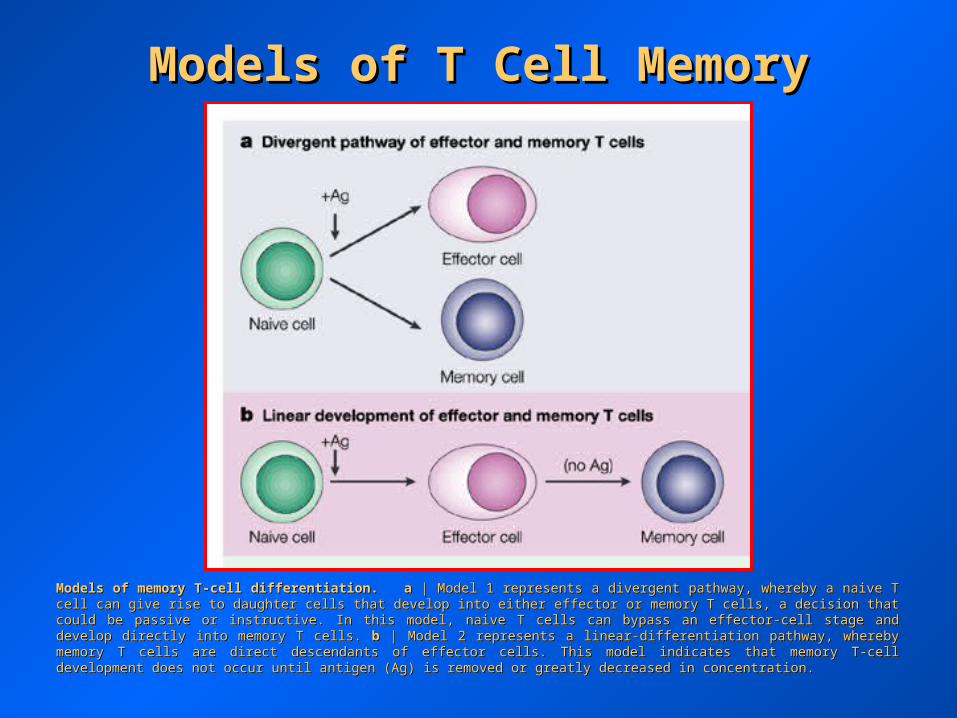
Models of memory T-cell differentiation.Models of memory T-cell differentiation. aa | Model 1 represents a divergent pathway, whereby a naive T cell can give rise to daughter cells | Model 1 represents a divergent pathway, whereby a naive T cell can give rise to daughter cells that develop into either effector or memory T cells, a decision that could be passive or instructive. In this model, naive T cells can bypass an that develop into either effector or memory T cells, a decision that could be passive or instructive. In this model, naive T cells can bypass an effector-cell stage and develop directly into memory T cells. effector-cell stage and develop directly into memory T cells. bb | Model 2 represents a linear-differentiation pathway, whereby memory T cells are | Model 2 represents a linear-differentiation pathway, whereby memory T cells are direct descendants of effector cells. This model indicates that memory T-cell development does not occur until antigen (Ag) is removed or direct descendants of effector cells. This model indicates that memory T-cell development does not occur until antigen (Ag) is removed or greatly decreased in concentration.greatly decreased in concentration.
Models of T Cell MemoryModels of T Cell Memory
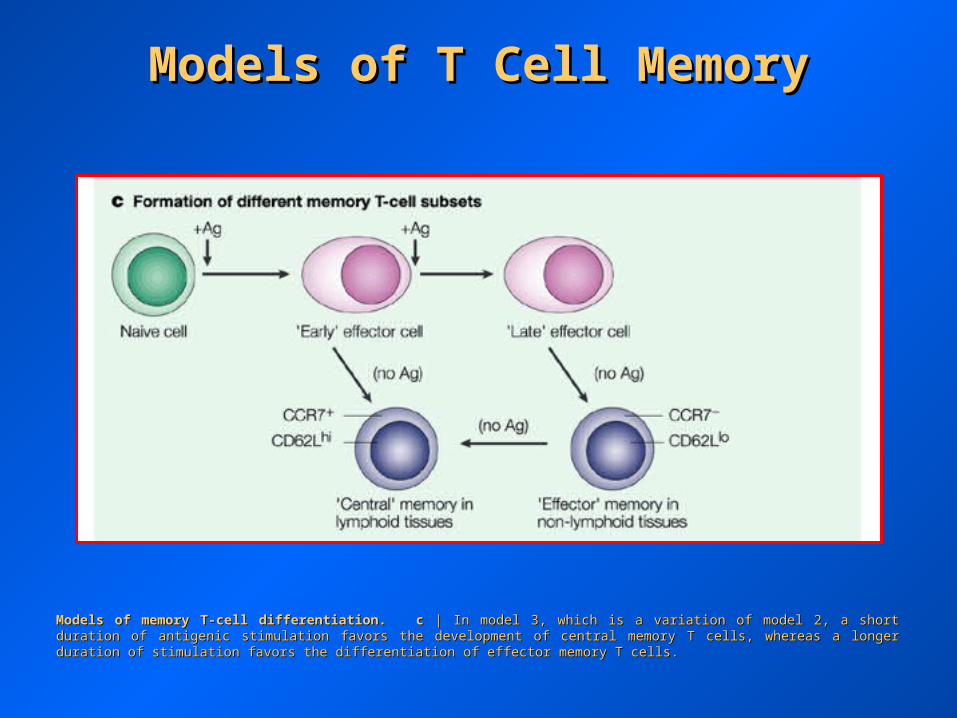
Models of memory T-cell differentiation.Models of memory T-cell differentiation. cc | In model 3, which is a variation of model 2, a short duration of antigenic stimulation favors the | In model 3, which is a variation of model 2, a short duration of antigenic stimulation favors the development of central memory T cells, whereas a longer duration of stimulation favors the differentiation of effector memory T cells.development of central memory T cells, whereas a longer duration of stimulation favors the differentiation of effector memory T cells.
Models of T Cell MemoryModels of T Cell Memory

Models of memory T-cell differentiation.Models of memory T-cell differentiation. dd | Model 4 represents the decreasing-potential hypothesis, which suggests that effector T-cell | Model 4 represents the decreasing-potential hypothesis, which suggests that effector T-cell functions steadily decrease as a consequence of persisting antigen (as observed in chronic infections). In addition, accumulative encounters functions steadily decrease as a consequence of persisting antigen (as observed in chronic infections). In addition, accumulative encounters with antigen lead to increased susceptibility of effector cells to apoptosis, and reduced numbers of memory T cells are formed. As suggested in with antigen lead to increased susceptibility of effector cells to apoptosis, and reduced numbers of memory T cells are formed. As suggested in model 2, the development of memory T cells occurs following antigen clearance. It is not known whether dysfunctional effector T cells can give model 2, the development of memory T cells occurs following antigen clearance. It is not known whether dysfunctional effector T cells can give rise to functional memory T cells, but this model suggests that T cells might regain function over time following the removal of antigen.rise to functional memory T cells, but this model suggests that T cells might regain function over time following the removal of antigen.
Models of T Cell MemoryModels of T Cell Memory

Modulation of the three phases of T-cell responses and the quantitative Modulation of the three phases of T-cell responses and the quantitative effects on memory T-cell formation.effects on memory T-cell formation. Alteration of any phase of a T-cell Alteration of any phase of a T-cell response — expansion, contraction or memory — can affect the number of response — expansion, contraction or memory — can affect the number of memory T cells that is formed and whether protective immunity is memory T cells that is formed and whether protective immunity is established. The dashed line denotes the minimum number of antigen-established. The dashed line denotes the minimum number of antigen-specific memory T cells required to confer protection against disease. If all specific memory T cells required to confer protection against disease. If all three phases occur optimally, then the number of memory T cells formed is three phases occur optimally, then the number of memory T cells formed is above the threshold for protective immunity (red line), but if above the threshold for protective immunity (red line), but if aa | priming and | priming and expansion are suboptimal; expansion are suboptimal; bb | cell death is increased; or | cell death is increased; or cc | memory T cells | memory T cells are not maintained, then the number of memory T cells that is formed will be are not maintained, then the number of memory T cells that is formed will be insufficient for long-term immunological protection (green line). Pink shading insufficient for long-term immunological protection (green line). Pink shading indicates the suboptimal phase of the response. Therapeutic intervention at indicates the suboptimal phase of the response. Therapeutic intervention at any of these three steps might lead to greater numbers of memory T cells any of these three steps might lead to greater numbers of memory T cells and protective immunity.and protective immunity.
Impacts on T Cell MemoryImpacts on T Cell Memory

Vaccine DesignsVaccine DesignsApproachApproach PurposePurpose
Vectored vaccinesVectored vaccines To employ an attenuated virus or bacterium To employ an attenuated virus or bacterium with limited replication potential to carry a with limited replication potential to carry a gene or genes for antigens from a pathogen gene or genes for antigens from a pathogen into the bodyinto the body
Nucleic acid vaccinesNucleic acid vaccines To use a plasmid containing genes coding for To use a plasmid containing genes coding for one or more antigens from a pathogen so that one or more antigens from a pathogen so that the body itself becomes a factory for the the body itself becomes a factory for the antigen(s) in question; there are variations on antigen(s) in question; there are variations on this themethis theme
Peptide vaccinesPeptide vaccines To construct a polymer out of a number of To construct a polymer out of a number of peptides, frequently T cell epitopes, thereby peptides, frequently T cell epitopes, thereby creating an immunogencreating an immunogen
Mucosal vaccinesMucosal vaccines To administer an antigen not by injection but To administer an antigen not by injection but through a mucosal surface (e.g., orally or through a mucosal surface (e.g., orally or intranasally), so as to engage the mucosal intranasally), so as to engage the mucosal immune system in protectionimmune system in protection
Transdermal vaccinesTransdermal vaccines To administer the antigen through the skinTo administer the antigen through the skin
Edible vaccinesEdible vaccines To genetically engineer a plant so that it To genetically engineer a plant so that it comes to contain antigens in a form that is comes to contain antigens in a form that is immunogenic when eatenimmunogenic when eaten
Prime-boost strategiesPrime-boost strategies To administer two separate versions of a To administer two separate versions of a vaccine sequentially, typically a DNA vaccine vaccine sequentially, typically a DNA vaccine followed by a vectored vaccine, or either of followed by a vectored vaccine, or either of these followed by a protein vaccinethese followed by a protein vaccine

Optimization of Vaccines ~Optimization of Vaccines ~AdjuvantsAdjuvants
• The nature of vaccine design may result in suboptimal generation of immune The nature of vaccine design may result in suboptimal generation of immune responses due to low immunogenicity, or suboptimal presentation to APCs – this can responses due to low immunogenicity, or suboptimal presentation to APCs – this can be ameliorated by the use of adjuvants or by designing vaccines that specifically target be ameliorated by the use of adjuvants or by designing vaccines that specifically target APCs. APCs.
• Uptake of antigens by APCs is vital; hence, rendering antigens polymeric or particulate Uptake of antigens by APCs is vital; hence, rendering antigens polymeric or particulate helps.helps.
• Activation of APCs is essential; hence, agents that engage TLR work well.Activation of APCs is essential; hence, agents that engage TLR work well.
• Depot effects and delayed absorption can enhance immunity; hence, emulsions and Depot effects and delayed absorption can enhance immunity; hence, emulsions and biodegradable microparticles underlie many adjuvant approaches.biodegradable microparticles underlie many adjuvant approaches.
• Many cytokines increase immune responses.Many cytokines increase immune responses.
• Molecular targeting of antigens to APCs through conjugation of ligands can enhance Molecular targeting of antigens to APCs through conjugation of ligands can enhance immune responses.immune responses.
• Targeting to dendritic cells (DC) specifically is likely to be important for antiviral Targeting to dendritic cells (DC) specifically is likely to be important for antiviral vaccine development.vaccine development.
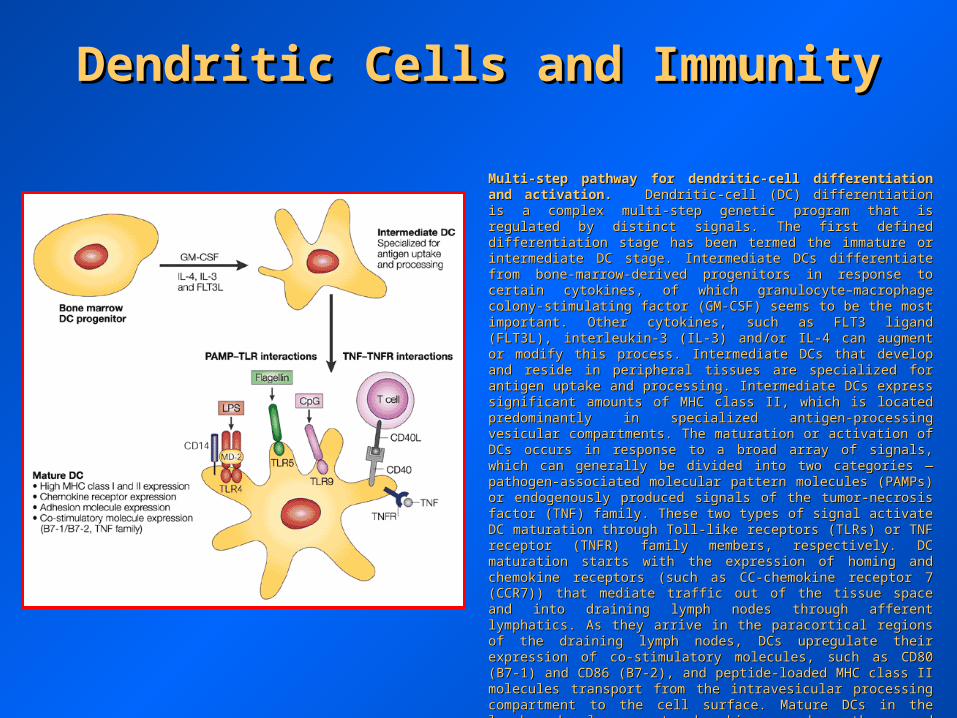
Multi-step pathway for dendritic-cell differentiation and activation.Multi-step pathway for dendritic-cell differentiation and activation. Dendritic-cell (DC) differentiation is a complex multi-step genetic program Dendritic-cell (DC) differentiation is a complex multi-step genetic program that is regulated by distinct signals. The first defined differentiation stage that is regulated by distinct signals. The first defined differentiation stage has been termed the immature or intermediate DC stage. Intermediate DCs has been termed the immature or intermediate DC stage. Intermediate DCs differentiate from bone-marrow-derived progenitors in response to certain differentiate from bone-marrow-derived progenitors in response to certain cytokines, of which granulocyte–macrophage colony-stimulating factor (GM-cytokines, of which granulocyte–macrophage colony-stimulating factor (GM-CSF) seems to be the most important. Other cytokines, such as FLT3 CSF) seems to be the most important. Other cytokines, such as FLT3 ligand (FLT3L), interleukin-3 (IL-3) and/or IL-4 can augment or modify this ligand (FLT3L), interleukin-3 (IL-3) and/or IL-4 can augment or modify this process. Intermediate DCs that develop and reside in peripheral tissues are process. Intermediate DCs that develop and reside in peripheral tissues are specialized for antigen uptake and processing. Intermediate DCs express specialized for antigen uptake and processing. Intermediate DCs express significant amounts of MHC class II, which is located predominantly in significant amounts of MHC class II, which is located predominantly in specialized antigen-processing vesicular compartments. The maturation or specialized antigen-processing vesicular compartments. The maturation or activation of DCs occurs in response to a broad array of signals, which can activation of DCs occurs in response to a broad array of signals, which can generally be divided into two categories — pathogen-associated molecular generally be divided into two categories — pathogen-associated molecular pattern molecules (PAMPs) or endogenously produced signals of the pattern molecules (PAMPs) or endogenously produced signals of the tumor-necrosis factor (TNF) family. These two types of signal activate DC tumor-necrosis factor (TNF) family. These two types of signal activate DC maturation through Toll-like receptors (TLRs) or TNF receptor (TNFR) maturation through Toll-like receptors (TLRs) or TNF receptor (TNFR) family members, respectively. DC maturation starts with the expression of family members, respectively. DC maturation starts with the expression of homing and chemokine receptors (such as CC-chemokine receptor 7 homing and chemokine receptors (such as CC-chemokine receptor 7 (CCR7)) that mediate traffic out of the tissue space and into draining lymph (CCR7)) that mediate traffic out of the tissue space and into draining lymph nodes through afferent lymphatics. As they arrive in the paracortical regions nodes through afferent lymphatics. As they arrive in the paracortical regions of the draining lymph nodes, DCs upregulate their expression of co-of the draining lymph nodes, DCs upregulate their expression of co-stimulatory molecules, such as CD80 (B7-1) and CD86 (B7-2), and peptide-stimulatory molecules, such as CD80 (B7-1) and CD86 (B7-2), and peptide-loaded MHC class II molecules transport from the intravesicular processing loaded MHC class II molecules transport from the intravesicular processing compartment to the cell surface. Mature DCs in the lymph node also compartment to the cell surface. Mature DCs in the lymph node also secrete chemokines, such as thymus and activation-regulated chemokine secrete chemokines, such as thymus and activation-regulated chemokine (TARC; CC-chemokine17), which attract naive T cells. Each of these DC (TARC; CC-chemokine17), which attract naive T cells. Each of these DC maturation and activation steps is potentially amenable to regulation by maturation and activation steps is potentially amenable to regulation by engineered immunotherapy approaches. LPS, lipopolysaccharide; CD40L, engineered immunotherapy approaches. LPS, lipopolysaccharide; CD40L, CD40 ligand.CD40 ligand.
Dendritic Cells and ImmunityDendritic Cells and Immunity

Approaches to enhance antigen targeting to antigen-presenting cells for improved immunization.Approaches to enhance antigen targeting to antigen-presenting cells for improved immunization. Several approaches are being Several approaches are being actively explored to enhance the processing and presentation of antigens by crucial antigen-presenting cells (APCs), such as dendritic cells. actively explored to enhance the processing and presentation of antigens by crucial antigen-presenting cells (APCs), such as dendritic cells. Recombinant viruses that directly infect APCs can be engineered to express antigen together with various MHC-targeting signals that will Recombinant viruses that directly infect APCs can be engineered to express antigen together with various MHC-targeting signals that will enhance antigen transport into the MHC class I and/or II processing pathways. Targeting to the MHC class I processing pathway can be enhance antigen transport into the MHC class I and/or II processing pathways. Targeting to the MHC class I processing pathway can be enhanced by linking antigen-encoding genes to genes that encode heat-shock proteins (HSPs), VP22 (an important coat protein of Marek's enhanced by linking antigen-encoding genes to genes that encode heat-shock proteins (HSPs), VP22 (an important coat protein of Marek's disease virus) or PET (disease virus) or PET (PseudomonasPseudomonas exotoxin translocation subunit). Targeting to the MHC class II pathway can be enhanced by linking the exotoxin translocation subunit). Targeting to the MHC class II pathway can be enhanced by linking the antigen to the lysosome-associated membrane protein 1 (LAMP1) or invariant chain (Ii; a molecule that is involved in MHC class II formation) antigen to the lysosome-associated membrane protein 1 (LAMP1) or invariant chain (Ii; a molecule that is involved in MHC class II formation) targeting signals. Additionally, genes that encode co-stimulatory molecules can be engineered into recombinant vectors, thereby endowing targeting signals. Additionally, genes that encode co-stimulatory molecules can be engineered into recombinant vectors, thereby endowing infected cells with super-physiological levels of co-stimulatory ligands. Similar modifications have been engineered into DNA vaccines. infected cells with super-physiological levels of co-stimulatory ligands. Similar modifications have been engineered into DNA vaccines. Engineered protein vaccines are being constructed that link antigens to ligands for receptors on the surface of APCs (such as DEC-205 and the Engineered protein vaccines are being constructed that link antigens to ligands for receptors on the surface of APCs (such as DEC-205 and the granulocyte–macrophage colony-stimulating factor (GM-CSF) receptor), which can effectively mediate endocytosis and targeting into MHC granulocyte–macrophage colony-stimulating factor (GM-CSF) receptor), which can effectively mediate endocytosis and targeting into MHC processing pathways. DEC-205 is also known as lymphocyte antigen 75 (LY75). processing pathways. DEC-205 is also known as lymphocyte antigen 75 (LY75).
Targeting Antigen Presenting CellsTargeting Antigen Presenting Cells

Vaccine FormulationsVaccine Formulations• Unlike vaccines that induce antibodies, T cell-inducing vaccines pose particular problems. The table below gives an Unlike vaccines that induce antibodies, T cell-inducing vaccines pose particular problems. The table below gives an
overview of the types of vaccines currently under development to specifically generate an antiviral T cell response.overview of the types of vaccines currently under development to specifically generate an antiviral T cell response.

Priming entityPriming entity Boosting entityBoosting entity
Viral vectorViral vector Recombinant proteinRecombinant protein
Naked DNANaked DNA Recombinant proteinRecombinant protein
Naked DNANaked DNA Viral vectorViral vector
Viral vectorViral vector Naked DNA*Naked DNA*
Viral vectorViral vector Different viral vectorDifferent viral vector
Some Examples of “Prime-Some Examples of “Prime-boost” Strategiesboost” Strategies
** Not very effective Not very effective
• Optimization of vaccination can be achieved by different combinations of priming and boosting – the table below Optimization of vaccination can be achieved by different combinations of priming and boosting – the table below gives an overview of some of the approaches that can be used.gives an overview of some of the approaches that can be used.
• The use of different “prime-boost” strategies can be used to overcome the possible reduction of the efficacy of the The use of different “prime-boost” strategies can be used to overcome the possible reduction of the efficacy of the boost phase due to a pre-existing neutralizing antibody response generated during priming – this existing antibody boost phase due to a pre-existing neutralizing antibody response generated during priming – this existing antibody response may remove the boosting phase before further activation takes place if the same entity is used in both response may remove the boosting phase before further activation takes place if the same entity is used in both phases.phases.

• Antiviral antibodies can impact viral infection in multiple ways.Antiviral antibodies can impact viral infection in multiple ways.
The antiviral activities of antibodies.The antiviral activities of antibodies. aa | Activities against free virus (an enveloped virus is shown). Neutralizing antibodies probably act | Activities against free virus (an enveloped virus is shown). Neutralizing antibodies probably act primarily by binding to the envelope protein (Env) at the surface of the virus and blocking infection (neutralization). They can also trigger effector primarily by binding to the envelope protein (Env) at the surface of the virus and blocking infection (neutralization). They can also trigger effector systems that can lead to viral clearance, as discussed in the text. systems that can lead to viral clearance, as discussed in the text. bb | Activities against infected cells. These activities can be mediated by both | Activities against infected cells. These activities can be mediated by both neutralizing and non-neutralizing antibodies. Neutralizing antibodies bind to the same proteins on infected cells as on free virus. Non-neutralizing neutralizing and non-neutralizing antibodies. Neutralizing antibodies bind to the same proteins on infected cells as on free virus. Non-neutralizing antibodies bind to viral proteins that are expressed on infected cells but not, to a significant degree, on free virus particles. Examples include antibodies bind to viral proteins that are expressed on infected cells but not, to a significant degree, on free virus particles. Examples include altered forms of Env protein and certain non-structural (NS) proteins, such as NS1 of dengue virus. The binding of neutralizing and/or non-altered forms of Env protein and certain non-structural (NS) proteins, such as NS1 of dengue virus. The binding of neutralizing and/or non-neutralizing antibodies to infected cells can lead to clearance of such cells or the inhibition of virus propagation as shown. neutralizing antibodies to infected cells can lead to clearance of such cells or the inhibition of virus propagation as shown.
Targets for Antiviral AntibodiesTargets for Antiviral Antibodies
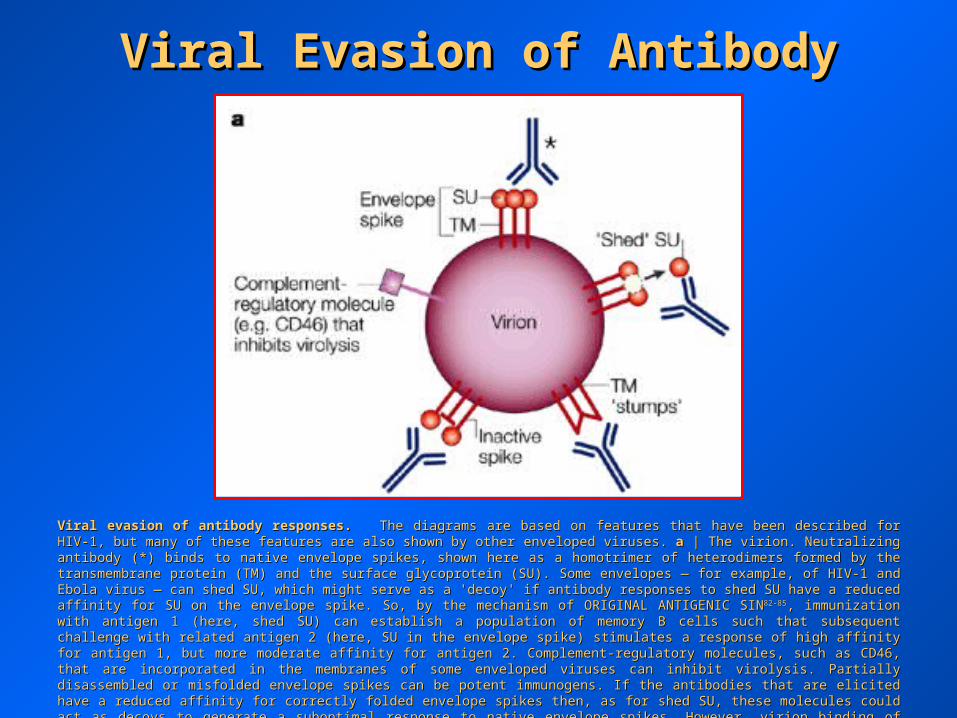
Viral Evasion of AntibodyViral Evasion of Antibody
Viral evasion of antibody responses.Viral evasion of antibody responses. The diagrams are based on features that have been described for HIV-1, but many of these features The diagrams are based on features that have been described for HIV-1, but many of these features are also shown by other enveloped viruses. are also shown by other enveloped viruses. aa | The virion. Neutralizing antibody (*) binds to native envelope spikes, shown here as a | The virion. Neutralizing antibody (*) binds to native envelope spikes, shown here as a homotrimer of heterodimers formed by the transmembrane protein (TM) and the surface glycoprotein (SU). Some envelopes — for example, of homotrimer of heterodimers formed by the transmembrane protein (TM) and the surface glycoprotein (SU). Some envelopes — for example, of HIV-1 and Ebola virus — can shed SU, which might serve as a 'decoy' if antibody responses to shed SU have a reduced affinity for SU on the HIV-1 and Ebola virus — can shed SU, which might serve as a 'decoy' if antibody responses to shed SU have a reduced affinity for SU on the envelope spike. So, by the mechanism of ORIGINAL ANTIGENIC SINenvelope spike. So, by the mechanism of ORIGINAL ANTIGENIC SIN82-8582-85, immunization with antigen 1 (here, shed SU) can establish a , immunization with antigen 1 (here, shed SU) can establish a population of memory B cells such that subsequent challenge with related antigen 2 (here, SU in the envelope spike) stimulates a response of population of memory B cells such that subsequent challenge with related antigen 2 (here, SU in the envelope spike) stimulates a response of high affinity for antigen 1, but more moderate affinity for antigen 2. Complement-regulatory molecules, such as CD46, that are incorporated in high affinity for antigen 1, but more moderate affinity for antigen 2. Complement-regulatory molecules, such as CD46, that are incorporated in the membranes of some enveloped viruses can inhibit virolysis. Partially disassembled or misfolded envelope spikes can be potent the membranes of some enveloped viruses can inhibit virolysis. Partially disassembled or misfolded envelope spikes can be potent immunogens. If the antibodies that are elicited have a reduced affinity for correctly folded envelope spikes then, as for shed SU, these immunogens. If the antibodies that are elicited have a reduced affinity for correctly folded envelope spikes then, as for shed SU, these molecules could act as decoys to generate a suboptimal response to native envelope spikes. However, virion binding of these non-neutralizing molecules could act as decoys to generate a suboptimal response to native envelope spikes. However, virion binding of these non-neutralizing antibodies could also, in principle, facilitate complement-mediated virolysis and phagocytosis.antibodies could also, in principle, facilitate complement-mediated virolysis and phagocytosis.
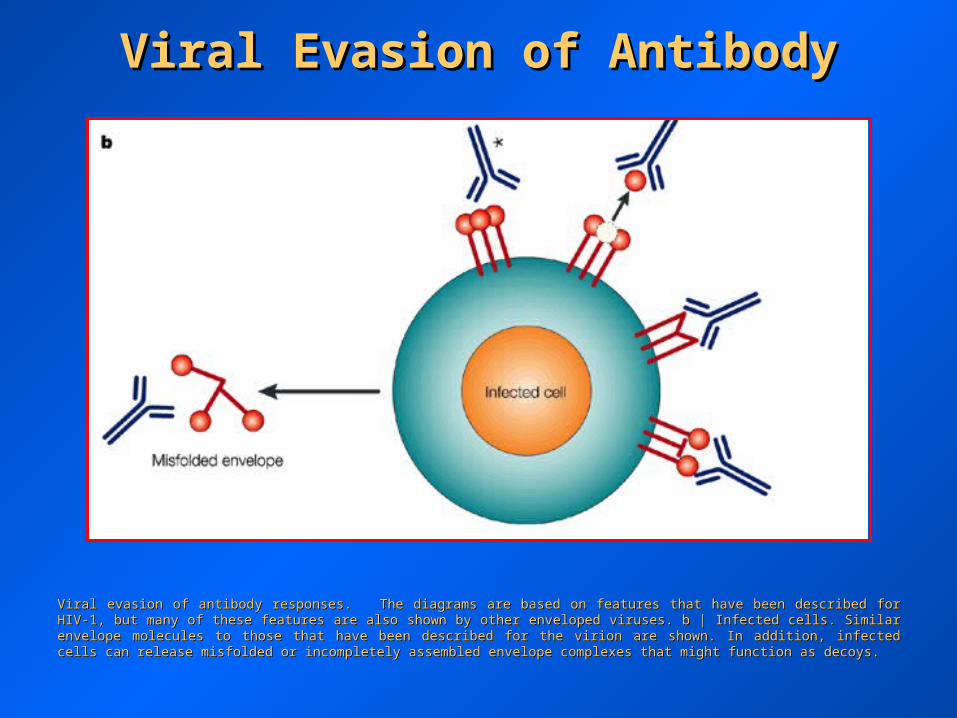
Viral Evasion of AntibodyViral Evasion of Antibody
Viral evasion of antibody responses. The diagrams are based on features that have been described for HIV-1, but many of these features are Viral evasion of antibody responses. The diagrams are based on features that have been described for HIV-1, but many of these features are also shown by other enveloped viruses. b | Infected cells. Similar envelope molecules to those that have been described for the virion are also shown by other enveloped viruses. b | Infected cells. Similar envelope molecules to those that have been described for the virion are shown. In addition, infected cells can release misfolded or incompletely assembled envelope complexes that might function as decoys.shown. In addition, infected cells can release misfolded or incompletely assembled envelope complexes that might function as decoys.

Viral Evasion of AntibodyViral Evasion of Antibody
Viral evasion of antibody responses.Viral evasion of antibody responses. The diagrams are based on features that have been described for HIV-1, but many of these features The diagrams are based on features that have been described for HIV-1, but many of these features are also shown by other enveloped viruses. are also shown by other enveloped viruses. cc | Surface glycoprotein. An immunodominant, but variable, loop favors the induction of strain- | Surface glycoprotein. An immunodominant, but variable, loop favors the induction of strain-specific rather than strain-cross-neutralizing antibodies, as for HIV-1 and influenza virus. Carbohydrate chains and oligomer formation conceal specific rather than strain-cross-neutralizing antibodies, as for HIV-1 and influenza virus. Carbohydrate chains and oligomer formation conceal large parts of the protein surface, which reduces antigenicity and immunogenicity. A recessed receptor site might also reduce antigenicity and large parts of the protein surface, which reduces antigenicity and immunogenicity. A recessed receptor site might also reduce antigenicity and immunogenicity. immunogenicity.

Structure of HIV-1.Structure of HIV-1. aa | Schematic of HIV-1 showing the envelope | Schematic of HIV-1 showing the envelope glycoproteins that are targets for neutralizing antibodies and the structural, as glycoproteins that are targets for neutralizing antibodies and the structural, as well as enzymatic, proteins that are targets for T cells. The longest three well as enzymatic, proteins that are targets for T cells. The longest three reading frames of the virus transcribe the Gag, Env and Pol polyproteins. The reading frames of the virus transcribe the Gag, Env and Pol polyproteins. The Gag polyprotein is processed into MA, CA, NC and p7, which make up the inner Gag polyprotein is processed into MA, CA, NC and p7, which make up the inner core of the viral particle. Glycoprotein 120 (gp120; SU) and gp41 (TM) are core of the viral particle. Glycoprotein 120 (gp120; SU) and gp41 (TM) are derived from the envelope (Env) polyprotein and are the outer membrane derived from the envelope (Env) polyprotein and are the outer membrane proteins of the virus. Processing of the polymerase (Pol) polyprotein yields the proteins of the virus. Processing of the polymerase (Pol) polyprotein yields the enzymes protease (PR), reverse transcriptase (RT) and integrase (IN), which enzymes protease (PR), reverse transcriptase (RT) and integrase (IN), which are encapsulated in the core of the inner particle. The accessory proteins Vif, are encapsulated in the core of the inner particle. The accessory proteins Vif, Vpr and Nef are encoded by three other reading frames in the virus. The Vpr and Nef are encoded by three other reading frames in the virus. The surface envelope glycoprotein is glycosylated at 25–30 sites, and contains surface envelope glycoprotein is glycosylated at 25–30 sites, and contains three variable loops that mask receptor-binding sites. Adapted, with permission, three variable loops that mask receptor-binding sites. Adapted, with permission, from Ref. 115. © (1998) Academic Press. from Ref. 115. © (1998) Academic Press. bb | Schematic depicting | Schematic depicting conformational changes of the gp120 subunit of Env that expose the conserved conformational changes of the gp120 subunit of Env that expose the conserved CD4- and co-receptor-binding sites. The '3' symbolizes the 3-fold axis, from CD4- and co-receptor-binding sites. The '3' symbolizes the 3-fold axis, from which gp41 interacts with gp120 to generate a functional trimer. The left-hand which gp41 interacts with gp120 to generate a functional trimer. The left-hand schematic depicts the form of gp120 on the surface of a virion. The CD4-schematic depicts the form of gp120 on the surface of a virion. The CD4-binding site is partially occluded by variable loops 1 and 2 (V1/V2) — shown in binding site is partially occluded by variable loops 1 and 2 (V1/V2) — shown in orange. The middle form represents a conformational change that is depicted orange. The middle form represents a conformational change that is depicted as an inner–outer domain shift, with the purple shape denoting the formation of as an inner–outer domain shift, with the purple shape denoting the formation of the CD4-binding cavity. The right-hand schematic depicts a third conformational the CD4-binding cavity. The right-hand schematic depicts a third conformational form, in which the gp120 'bridging sheet' and sequences in the third variable form, in which the gp120 'bridging sheet' and sequences in the third variable loop (shown in green) bind to the chemokine co-receptor. loop (shown in green) bind to the chemokine co-receptor.
Strategies for HIV VaccinesStrategies for HIV Vaccines

Strategies for HIV VaccinesStrategies for HIV Vaccines
Two strategies for HIV DNA vaccine inserts.Two strategies for HIV DNA vaccine inserts. aa | Depiction of the nine open reading frames — Gag, Env, Pol, Tat, Rev, Vpu, Vif, Vpr and Nef | Depiction of the nine open reading frames — Gag, Env, Pol, Tat, Rev, Vpu, Vif, Vpr and Nef —in the HIV-1 genome. —in the HIV-1 genome. bb | Example of the expression of several proteins from a single DNA by HIV regulatory protein (Rev)-dependent | Example of the expression of several proteins from a single DNA by HIV regulatory protein (Rev)-dependent subgenomic splicing. The construct is the HIV-1 DNA vaccine that we are advancing into human trials. 'subgenomic splicing. The construct is the HIV-1 DNA vaccine that we are advancing into human trials. 'XX' denotes a safety mutation that ' denotes a safety mutation that inactivates reverse transcriptase or the zinc fingers for packaging viral RNA. inactivates reverse transcriptase or the zinc fingers for packaging viral RNA. cc | Example of the expression of a codon-optimized fusion protein. | Example of the expression of a codon-optimized fusion protein. This DNA vaccine expresses the largest number of sequences among current codon-optimized vaccines, and was constructed by deleting the This DNA vaccine expresses the largest number of sequences among current codon-optimized vaccines, and was constructed by deleting the frameshift between the genes that encode group-specific antigen (Gag) and the viral enzymes protease (PR), reverse transcriptase (RT) and frameshift between the genes that encode group-specific antigen (Gag) and the viral enzymes protease (PR), reverse transcriptase (RT) and integrase (IN). The integrase (IN). The PRPR gene includes an inactivating mutation. CA, p24 capsid protein; Env, envelope protein; LTR, long terminal repeat; MA, gene includes an inactivating mutation. CA, p24 capsid protein; Env, envelope protein; LTR, long terminal repeat; MA, p17 matrix protein; NC, p7 nucleocapsid protein; RT, p66/p51 reverse transcriptase; SU, gp120 surface envelope protein; TM, gp41 p17 matrix protein; NC, p7 nucleocapsid protein; RT, p66/p51 reverse transcriptase; SU, gp120 surface envelope protein; TM, gp41 transmembrane envelope protein. vif, vpr, vpu and nef are viral accessory proteins; tat and rev are viral regulatory proteins. transmembrane envelope protein. vif, vpr, vpu and nef are viral accessory proteins; tat and rev are viral regulatory proteins.

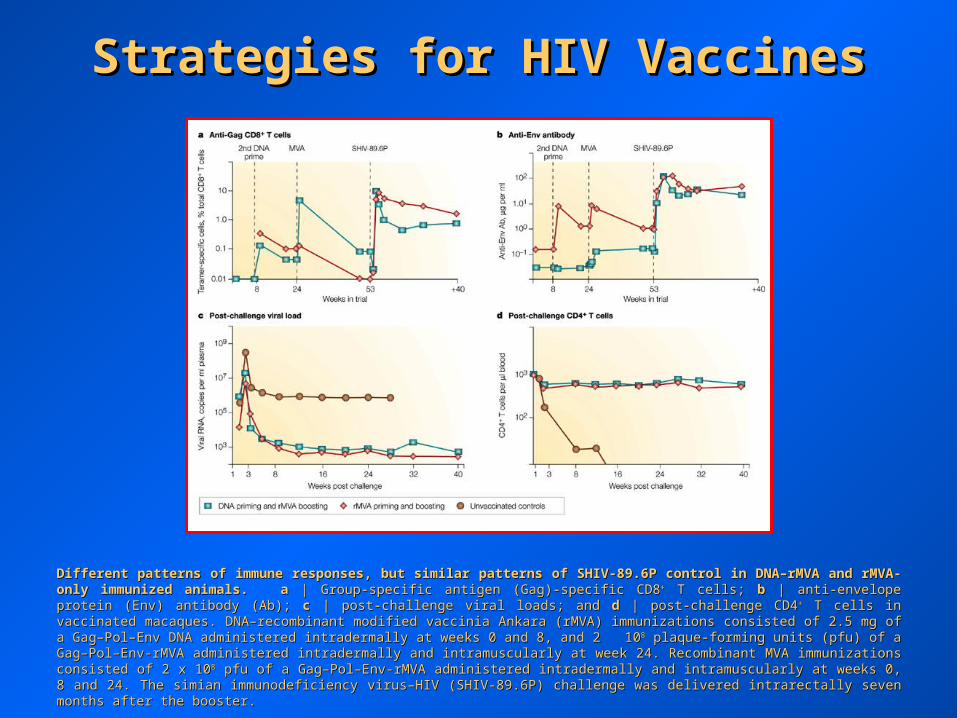
Strategies for HIV VaccinesStrategies for HIV Vaccines
Different patterns of immune responses, but similar patterns of SHIV-89.6P control in DNA–rMVA and rMVA-only immunized animals.Different patterns of immune responses, but similar patterns of SHIV-89.6P control in DNA–rMVA and rMVA-only immunized animals. aa | Group-specific antigen (Gag)-specific CD8 | Group-specific antigen (Gag)-specific CD8++ T cells; T cells; bb | anti-envelope protein (Env) antibody (Ab); | anti-envelope protein (Env) antibody (Ab); cc | post-challenge viral loads; and | post-challenge viral loads; and dd | post- | post-challenge CD4challenge CD4++ T cells in vaccinated macaques. DNA–recombinant modified vaccinia Ankara (rMVA) immunizations consisted of 2.5 mg of a T cells in vaccinated macaques. DNA–recombinant modified vaccinia Ankara (rMVA) immunizations consisted of 2.5 mg of a Gag–Pol–Env DNA administered intradermally at weeks 0 and 8, and 2 10Gag–Pol–Env DNA administered intradermally at weeks 0 and 8, and 2 1088 plaque-forming units (pfu) of a Gag–Pol–Env-rMVA administered plaque-forming units (pfu) of a Gag–Pol–Env-rMVA administered intradermally and intramuscularly at week 24. Recombinant MVA immunizations consisted of 2 x 10intradermally and intramuscularly at week 24. Recombinant MVA immunizations consisted of 2 x 1088 pfu of a Gag–Pol–Env-rMVA administered pfu of a Gag–Pol–Env-rMVA administered intradermally and intramuscularly at weeks 0, 8 and 24. The simian immunodeficiency virus–HIV (SHIV-89.6P) challenge was delivered intradermally and intramuscularly at weeks 0, 8 and 24. The simian immunodeficiency virus–HIV (SHIV-89.6P) challenge was delivered intrarectally seven months after the booster.intrarectally seven months after the booster.


HIV/SIV CD8 T Cell-InducingHIV/SIV CD8 T Cell-InducingVaccine EfficacyVaccine Efficacy

T Cell-based Vaccine EfficacyT Cell-based Vaccine Efficacy