vergne al, aubin t, martin s, mathevon n, 2012. acoustic communication in crocodilians
TRANSCRIPT

ORIGINAL PAPER
Acoustic communication in crocodilians: information encodingand species specificity of juvenile calls
Amelie L. Vergne • Thierry Aubin •
Samuel Martin • Nicolas Mathevon
Received: 21 November 2011 / Revised: 2 July 2012 / Accepted: 2 July 2012 / Published online: 21 July 2012
� Springer-Verlag 2012
Abstract In the Crocodylia order, all species are known
for their ability to produce sounds in several communica-
tion contexts. Though recent experimental studies have
brought evidence of the important biological role of young
crocodilian calls, especially at hatching time, the juvenile
vocal repertoire still needs to be clarified in order to
describe thoroughly the crocodilian acoustic communica-
tion channel. The goal of this study is to investigate the
acoustic features (structure and information coding) in the
contact call of juveniles from three different species (Nile
crocodile Crocodylus niloticus, Black caiman, Melanosu-
chus niger and Spectacled caiman, Caiman crocodilus).
We have shown that even though substantial structural
differences exist between the calls of different species, they
do not seem relevant for crocodilians. Indeed, juveniles and
adults from the species studied use a similar and non-
species-specific way of encoding information, which relies
on frequency modulation parameters. Interestingly, using
conditioning experiments, we demonstrated that this tol-
erance in responses to signals of different acoustic struc-
tures was unlikely to be related to a lack of discriminatory
abilities. This result reinforced the idea that crocodilians
have developed adaptations to use sounds efficiently for
communication needs.
Keywords Crocodiles � Caimans �Acoustic communication � Species recognition �Information coding � Conditioning experiments
Introduction
In crocodiles, acoustic signalling is used for social inter-
actions, particularly between adults during courtship and
territorial defence (Garrick et al. 1982), and within family
groups where survival of the young depends on maternal
care (Campbell 1973; Herzog and Burghardt 1977; Vergne
and Mathevon 2008; Senter 2008; Vergne et al. 2011; for a
review of acoustic communication in crocodilians, see
Vergne et al. 2009). Juvenile crocodilians produce various
sounds that have been classified into three main functional
categories. First, ‘‘hatching calls’’ (with the sub-categories,
pre-hatching, hatching and post-hatching) that are uttered
by embryos and newborns to solicit the mother to open the
nest and have been shown to fine-tune hatching synchrony
among siblings (Vergne and Mathevon 2008); second,
‘‘contact calls’’ produced by juveniles from hatching to
several days or weeks old. These mainly support cohesion
between juveniles (Vergne et al. 2011). And the third is
‘‘distress calls’’ that induce parental (mother) protection
(for an experimental approach of the biological roles of
contact and distress calls, see Vergne et al. 2011). Finally,
juveniles may emit ‘‘threat and disturbance’’ calls when
threatened (Britton 2001; Vergne et al. 2009). Although the
acoustic structure and the biological roles of these juvenile
signals have recently started to be studied (Britton 2001;
A. L. Vergne � N. Mathevon (&)
Equipe de Neuro-Ethologie Sensorielle, CNPS, Universite de
Lyon—Saint-Etienne, CNRS UMR 8195, Saint-Etienne, France
e-mail: [email protected]
A. L. Vergne � T. Aubin � N. Mathevon
Centre de Neurosciences Paris-Sud, Centre National de la
Recherche Scientifique, UMR 8195, Paris, France
T. Aubin
Equipe ‘Communications acoustiques’, CNPS, Universite Paris
XI, CNRS UMR 8195, Orsay, France
S. Martin
La Ferme aux Crocodiles, Pierrelatte, France
123
Anim Cogn (2012) 15:1095–1109
DOI 10.1007/s10071-012-0533-7

Vergne and Mathevon 2008; Vergne et al. 2007, 2009,
2011), we still do not know the specific acoustic features
that make these sounds relevant to crocodilians.
Species specificity is one of the most consistent char-
acteristic of animal vocalizations, especially in the context
of reproduction (Dooling et al. 1992). This is related to the
fact that most sounds are directed towards conspecifics
whose reactions should be appropriate to the content of the
message. The species specificity of vocalizations is of
primary importance when two close-related species live in
sympatry and when risks of hybridization or mis-directed
parental care can occur. Among the 23 species of crocod-
ilians spread over the subtropical and tropical regions of
the New and Old World (Trutnau and Sommerlad 2006),
sympatry is rare and most interactions are likely to be intra-
specific (though there are some exceptions, for example,
Spectacled caiman Caiman crocodilus and Black caiman
Melanosuchus niger in South America). Due to genetic
drift, differences in the acoustic structure of vocalizations
may also arise in allopatry. Except Britton (2001) who
conducted a preliminary structural analysis of juvenile calls
from different species, there is no published investigation
on the species specificity of juvenile calls.
In the present study, we focused on juvenile contact
calls. These vocalizations, used by the group to gather,
seem to be mostly directed towards other juveniles (Vergne
et al. 2011). By investigating the acoustic structure of
contact calls from different species, our goal was to dis-
cover whether calls were species-specific or whether they
share a common rule for encoding information. First, we
compared the acoustic structure of contact calls from three
different species (one species belonging to the family
Crocodylidae: Nile crocodile Crocodylus niloticus; two
species from the family Alligatoridae: Black caiman,
Melanosuchus niger and Spectacled caiman, Caiman
crocodilus), and we looked for the presence of species-
specific information in these recorded calls using playback
experiments. Then, modified signals were broadcast to find
the acoustic features that trigger the receiver’s behavioural
response. Finally, conditioning experiments helped to
determine the details of some aspects of crocodile auditory
discriminatory abilities.
Experiment 1: species specificity of juvenile calls
Methods
Sound analysis
We analysed calls from juveniles of 3 different species
(Nile Crocodile, Spectacled Caiman and Black Caiman; 25
calls per species, from 5 individuals/species). Animals were
recorded either at the laboratory (Nile crocodiles and
Spectacled caimans, provided by the zoo ‘‘La Ferme aux
Crocodiles’’, Pierrelatte, France), or in the field (Black
caimans, studied along the Rupununi river, Guyana, South
America). All recorded calls were spontaneously emitted in
contexts where juveniles were gathered as a group, without
any visible disturbance. They thus can be considered as
typical juvenile ‘‘contact calls’’ (Vergne et al. 2009, 2011).
These acoustic signals are ‘‘complex’’ sounds, that is,
composed of a fundamental frequency and a harmonic
series, modulated in frequency and amplitude (Fig. 1).
The Nile crocodiles recorded were 4–6 days old. They
hatched in our laboratory, and vocalizations were recorded
at the time the crocodiles were released into a ‘‘home tank’’
where they could meet other siblings for the first time.
Spectacled caimans were older individuals (2–3 weeks old).
They hatched at the zoo ‘‘La Ferme aux Crocodiles’’ (Pi-
errelatte, France) and were brought to the laboratory
2 weeks later. Their vocalizations were recorded when we
released them together in their own home tank. The Black
caimans’ contact calls were recorded during a field trip to
Guyana (South America). Individuals were about 1 week
old. Contact calls were emitted spontaneously by juveniles
gathered on the riverbank. All recordings were performed at
a distance of 30–40 cm from the animals with an omnidi-
rectional microphone SENNHEISER MD42 connected to a
Marantz PMD670 tape recorder.
We conducted a sound analysis to describe the calls’
acoustic structure. For the Nile crocodile contact calls, the
analysis had already been performed and published else-
where (Vergne et al. 2009). For comparison purposes, we
used the same method to analyse the contact calls of the two
other species. Briefly, 7 acoustic variables were measured in
both temporal and frequency domains using Avisoft-SAS
Lab-Pro (http://www.avisoft.com/) and Praat (http://www.
praat.org) (Fig. 2). The total duration of the call (DT) was
measured from the oscillogram (Fig. 2b). We performed a
spectrographic analysis (window size: 1,024, sampling
frequency: 48 kHz, overlap 90 %) and measured two vari-
ables related to the fundamental frequency: the maximal
frequency value (F0max, Hz) and the ending frequency
value (Fend, Hz; Fig. 2a). To describe the frequency
modulation, we calculated the slope of the first temporal
quartile of the call (Slope 1) and the slope of the last three
temporal quartiles of the call (Slope 2; Fig. 2a). To describe
the spectral energy distribution, we measured two param-
eters on a spectrum of the entire call: the bandpass (Band-
pass, Hz) and the frequency at the maximum amplitude
(PicF, Hz) (Fig. 2c).
Statistical tests were conducted using the Statistica
package (version 6, Statsoft France). First, a one-way
MANOVA based on the 7 measured acoustic variables was
performed to compare the calls between the 3 species. This
1096 Anim Cogn (2012) 15:1095–1109
123

analysis was followed by a one-way ANOVA on each
acoustic variable and a post hoc Fisher’s LSD procedure at a
95 % confidence level to identify statistically homogeneous
groups. A cross-validated discriminant function analysis
using the 7 measured acoustic variables, preceded by a
principal component analysis, provided a classification
procedure that assigned each call to its appropriate species
(correct assignment) or to one of the two others (incorrect
assignment).
Playback experiments
Five young Nile crocodiles (age: 1 month old, sex
unknown) were tested. These individuals were kept
together in a home tank (dimensions: 120 9 180 9 50 cm;
water temperature: 30 �C; air temperature: 28–30 �C;appropriate luminance to this tropical species was pro-
grammed 12 h day/12 h night). During experimental peri-
ods, the tested individual was isolated from the others in a
test tank whose characteristics—particularly temperature,
luminance and feeding conditions—were the same as for
the home tank. Both tanks had a shelter at one end and the
crocodiles hid in it most of the time. The test tank was
located in an acoustically isolated room. To limit the
possible stress on the crocodiles due to carrying them from
on tank to another, we always moved the tested individual
in the test tank on the morning before the playback so that
there were several hours of non-disturbance before the
Fig. 1 Spectrograms (top) and oscillograms (bottom) of juvenile contact calls (a: Nile crocodile; b: spectacled caiman; c: black caiman)
Fig. 2 Acoustic parameters
used for the analysis of juvenile
calls. a Spectrogram.
b Oscillogram. c Average
frequency spectrum. DT (s):
total duration, F0max (Hz):
maximal frequency of the
fundamental, Fend (Hz): final
frequency of the fundamental,
slopes 1 and 2 (Hz/s): frequency
modulation slopes, PicF (Hz):
frequency at the maximum
power amplitude of the
spectrum
Anim Cogn (2012) 15:1095–1109 1097
123

tests. All playback tests were conducted during the night
(from 8 p.m. to 2 a.m.)—a preliminary study of the croc-
odile circadian behavioural activity having shown a pref-
erential nocturnal activity. A low intensity red neon light
was placed above the aquarium to allow the monitoring of
behaviours at night.
To test the species specificity of calls, each of the 5
individuals was successively challenged with Nile crocodile
calls, black caiman calls, spectacled caiman calls and a
control noise (white noise band-pass filtered between 100
and 5,000 Hz; duration = 0.15 s; emitted at 50 dBSPL;
built with Syntana software, Aubin 1994). The spectrum
bandwidth, duration and intensity of the control noise were
chosen to mimic those of a juvenile natural contact call. A
loudspeaker was suspended at one end of the test tank (at
the opposite side from the shelter). Bubble wrap covered the
tank’s inner walls in order to minimize reverberation of
acoustic waves. The loudspeaker was connected to a com-
puter located outside the experimental room to minimize
disturbance for the isolated crocodile. This computer was
programmed to control the emission of acoustic stimuli.
Emission level conformed to the natural intensity of calls
(51 ± 4 dBSPL at 1 metre from the loudspeaker, measured
with a sound level meter SL-4001, Digital Instruments).
Every 2 h starting at 8 p.m., the tested young Nile crocodile
was exposed to a 90-second playback of an experimental
acoustic sequence. Each sequence was composed of 5
identical series of experimental signals; interval between
series = 12 s; 1 series = 4 repetitions of a given experi-
mental signal with a natural emission rhythm, approxi-
mately one call every 2 s; total duration of one series = 8 s.
During each experimental night, each tested individual was
challenged with 1 sequence of each species’ calls and 1
sequence of control noise. The order of sequences was
changed for each individual. Pseudo-replication within
sequences of a given species’ calls was avoided by building
sequences using different calls from different individuals.
Analysis of behavioural responses
All behavioural and vocal responses were recorded. We
were able to follow precisely the crocodile’s movements
with two webcams (IP DCS-900) suspended above the two
opposite sides of the tank to get a complete view. Go1984
software (http://www.go1984.com/) was programmed to
activate the webcams and record the crocodile’s activity
during sequences of 15 min (5 min before the start of the
playback sequence, during the 90 s of playback and 10 min
after). Any vocal activity was recorded during the same
duration via a microphone (Labtec desk microphone).
Two parameters were chosen to measure the responses to
playbacks: (1) Latency (t), the time between the first call
played back and the first observable reaction (head or body
movement) of the tested individual. A 4-level scale was
defined: 0 if t[ 60 s or no reaction, 1 if 41 B t B 60 s, 2 if
21 B t B 40 s, and 3 if t B 20 s; (2) animal displacements
were also quantified according to a 4-levels scale: 0 was
scored if the animal did not move at all during the 90 s of
playback, 1 if we observed a head and/or body orientation
towards the loudspeaker but no displacement, 2 if the indi-
vidual moved towards the loudspeaker but stayed at more
than 20 cm from it, and 3 if the individual came at less than
20 cm from the loudspeaker. Note that we only took into
consideration experiments when the crocodile was at the
opposite side of the speaker before the start of a test (that
was the case 99 % of the time). Also, we did not need
negative scores to assess displacements because once a
tested animal moved towards the speaker, it never moved
away from it during our recordings. These two scores (head
or bodymovement and displacements) were then summed to
obtain a general score representing the behavioural response
to playback, varying from 0 to 6, with 0 corresponding to the
weakest behavioural response and 6 to the strongest.
Using the behavioural response scores, we first per-
formed a Friedman two-way repeated measures analysis of
variance by ranks to detect differences in Nile crocodiles’
responses throughout the different playback tests. Second,
we did Wilcoxon tests to compare responses to the Nile
crocodile calls with responses to the two other signals.
Statistical tests were conducted using Statistica package.
Results
Acoustic differences between the contact calls of the three
species
Although the acoustic structure of calls of the three species
show strong similarities, as shown by spectrographic rep-
resentations (Fig. 1), the analysis sheds light on significant
differences between the measured parameters from one
species to another (Table 1, MANOVA, F(14,132) = 21.2,
P\ 0.001). The one-way ANOVAs and the Fisher’s LSD
tests reveal that all measured acoustic parameters except the
slopes of the frequency modulation allow differentiating a
Nile crocodile contact call from a Black or a spectacled
caiman’s call (ANOVAs: df = 2,72; P\ 0.001 for each of
the seven acoustic variables, ‘‘DT’’: F = 178, ‘‘F0max’’:
F = 21.2, ‘‘Fend’’: F = 9.29, ‘‘Bandpass’’: F = 31.2,
‘‘Slope 1’’: F = 7.65, ‘‘Slope 2’’: F = 8.30, ‘‘PicF’’:
F = 22.9; homogeneous groups: see Table 2). Nile croco-
dile calls are higher pitched (F0max, Fend, Bandpass),
possess more energy towards high frequencies (PicF) and
have a longer duration (DT) than calls of the other tested
species. Differences were also measured between contact
calls of Spectacled and Black caimans. For instance, the
maximum of the fundamental frequency (F0max) and the
1098 Anim Cogn (2012) 15:1095–1109
123

frequency bandpass (Bandpass) are higher pitched in
Spectacled caiman calls than in Black caimans’ ones. The
frequency modulation slopes (Slope 1 and 2) also differ-
entiate Spectacled caiman contact calls from the two others,
the frequency modulation of the calls from this species
being more pronounced (Table 2).
In summary, though the basic acoustic structure of
crocodilian calls (a complex sound with multiple harmonic,
modulated in frequency and amplitude) appears similar
between different species, differences exist and are sub-
stantial enough to assign each call in its proper category
with confidence. A cross-validated discriminant function
analysis correctly classified 96 % of the Nile crocodile
contact calls, 88 % of the Black Caiman contact calls and
80 % of the Spectacled caiman calls (Fig. 3).
Behavioural response to contact calls
The Friedman analysis found significant differences
between Nile crocodiles’ responses to the broadcast signals
(P\ 0.02, Fig. 4): four individuals out of the five tested
did not react with the noise stimulus; conversely, the three
types of crocodilians’ calls elicited strong behavioural
responses from all juvenile Nile crocodiles. There was no
significant difference between responses to Nile crocodile,
Spectacled caiman and Black caiman calls.
Because acoustic analysis had shown significant struc-
tural differences between the calls of the three species, one
hypothesis that could explain why Nile crocodiles indi-
viduals respond similarly to calls of their own species and
to heterospecific calls is that all these signals encode the
same information. Investigation of the acoustic parameters
that are responsible for the crocodiles’ behavioural
responses may shed some light on this matter.
Experiment 2: identification of calls’ salient acoustic
parameters
Methods
From playback experiments 1, it appeared that Nile croc-
odile juveniles react similarly to calls of their own species
and heterospecific calls. However, they differentiate croc-
odilian calls from noise, meaning that calls convey some
‘‘crocodilian’’ information. In order to identify the relevant
parameters of calls (in the frequency and/or temporal
domains), we ran playback experiments using acoustic
lures (modified signals). Each acoustic parameter was
tested independently. The original sounds used to prepare
experimental signals were selected from the bank of
recorded contact calls previously described.
Adult crocodiles (especially females) are also known to
react to juvenile calls (Vergne and Mathevon 2008). Thus,
we decided to also test adult crocodilians in order to see
whether the coding rules might change with age.
Experiments on juveniles
Twenty-two naıve individuals (6 Spectacled caimans—
2 months old, 16 Nile crocodiles—1 month old, sex
unknown) were available for our experiments. To avoid
habituation due to multiple testing, we performed playback
experiments on two different sets of individuals:
The first experimental set was composed of 6 Spectacled
caimans and 6 Nile crocodiles. We tested them individually
with a series of 7 experimental signals (Fig. 5). Each
individual was tested with natural and modified signals
from its own species. To avoid pseudo-replication, we used
a different series of experimental signals for each indi-
vidual—each series being built from a different natural
contact call (NAT). Spectacled caiman natural calls came
from 2- to 3-week-old individuals while Nile crocodile
natural calls came from 4- to 6-days old individuals (see
details in Experiment 1 above). We worked with synthetic
copies of calls so that we could modify all parameters of
the signal. Modified versions of natural contact calls were
created with Avisoft-SAS Lab-Pro software. The six fol-
lowing signals were built from a NAT signal (Fig. 5): (1) a
synthetic copy of the natural call (Scontrol; control signal),
(2) a signal without amplitude modulation (noAM; the
frequency modulation of the NAT was retained), (3) a
signal without frequency modulation (FM1; the amplitude
modulation as well as the energy distribution between the
different harmonics was identical to the NAT), (4) a signal
without any harmonic structure (1H, only the first
Table 1 Mean ± SD of acoustic variables from contact calls of three species of crocodilians—Nile crocodiles, Spectacled caimans and Black
caimans (5 individuals, 5 calls/individual)
DT(s) F0max (Hz) Fend (Hz) Bandpass (Hz) Slope 1 (Hz/s) Slope 2 (Hz/s) PicF (Hz)
Nile crocodiles (N = 25) 0.195 ± 0.029 506 ± 116 211 ± 61 3,925 ± 760 -2,225 ± 1,536 -1,291 ± 429 721 ± 222
Black caimans (N = 25) 0.085 ± 0.014 319 ± 65 169 ± 46 2,573 ± 193 -2,784 ± 1,607 -1,437 ± 595 651 ± 242
Spectacled caimans (N = 25) 0.09 ± 0.024 379 ± 122 149 ± 49 3,289 ± 694 -4,005 ± 1,784 -2,091 ± 1,048 599 ± 123
DT (s): total duration, F0max (Hz): maximal frequency of the fundamental, Fend (Hz): final frequency of the fundamental, slopes 1 and 2 (Hz/s):
frequency modulation slopes, PicF (Hz): frequency at the maximum power amplitude of the spectrum
Anim Cogn (2012) 15:1095–1109 1099
123

harmonic, which is also the one of highest energy, was
retained), (5) a temporally reversed natural call (Srev; a
temporally mirror of the frequency modulation slope of the
original call), and (6) a temporally reversed version of 1H
(1Hrev).
The second experimental set was composed of 10 naıve
Nile crocodiles (2 weeks old). Besides the control NAT
signal, they were challenged with 6 synthetic signals
(Fig. 5): (1) a synthetic copy of the natural call (synthetic
control signal, Scontrol); (2) a signal with a frequency
modulation slope reduced by one-third compared to the
natural slope (SLOPE1): in the Nile crocodile SLO-
PE1 = 1,017 ± 384 Hz s-1, in the Black caiman: SLO-
PE1 = 1,182 ± 417 Hz s-1 and in the Spectacled caiman:
SLOPE1 = 1,713 ± 557 Hz s-1; (3) a signal with a fre-
quency modulation slope reduced by a half compared to the
natural slope (SLOPE2): in the Nile crocodile SLO-
PE2 = 763 ± 288 Hz s-1, in the Black caiman:
SLOPE2 = 887 ± 313 Hz s-1 and in the Spectacled cai-
man: SLOPE2 = 1,284 ± 418 Hz s-1; (4) a signal with-
out any frequency modulation or harmonics series (FM2);
(5) a signal with a modified energy distribution across the
spectrum (ENER, with 80 % of energy between 2,500 and
5,000 Hz instead of between 200 and 2,500 Hz as in the
control signal); (6) a noise having the same duration, fre-
quency bandpass and amplitude modulation as the control
signal (NOISE). The first four and the NOISE experimental
signals were built using the Avisoft-SAS Lab-Pro software.
The ENER signal was built using PRAAT software.
The experimental procedures and the assessment of
behavioural responses were identical to those described in
the experiments testing the species specificity of calls
(Experiment 1; air and water temperatures were 28–30 and
30 �C, respectively). All statistical tests were conducted
using Statistica package. We performed a Friedman two-
way repeated measures analysis of variance by ranks to
detect differences in crocodilian responses between the
different playback tests, and Wilcoxon two-tailed tests for
comparisons of behavioural responses to the Natural call
with responses to other signals tested.
Experiments on adults
Adult experiments were performed at the zoo ‘‘La Ferme
aux Crocodiles’’ (hosting 350 adult Nile crocodiles in a
8,000 m2 tropical greenhouse; 80 % of the crocodiles are
mature females of 3–8 years old; the air and water tem-
peratures are 28–30 and 30 �C, respectively). Although it
was possible to approach animals conveniently, it was
impossible to test them individually. We thus played back
experimental signals to clusters of 7–24 adults and moni-
tored the response of all individuals together. Five clusters
were carefully chosen within the greenhouse to avoid
possible interferences between them: they were located as
far away as possible from each other (more than 30 m
apart). To limit the beam size of propagated sounds, we
used a directional loudspeaker (Audax) and only the tested
cluster was situated in front of the loudspeaker (the closest
Table 2 Results of Fisher LSD tests to identify statistically homo-
geneous groups of calls on the basis of 7 measured acoustic
parameters
Acoustic parameter Homogeneous groups
F0max (Hz) (Nile) (Black) (Spec)
Bandpass (Hz) (Nile) (Black) (Spec)
Slope 1 (Hz/s) (Nile, Black) (Spec)
Slope 2 (Hz/s) (Nile, Black) (Spec)
DT (s) (Nile) (Black, Spec)
Fend (Hz) (Nile) (Black, Spec)
PicF (Hz) (Nile) (Black, Spec)
‘‘Nile’’: Nile crocodile calls; ‘‘Black’’: black caimans calls; ‘‘Spec’’:
spectacled caiman calls. Calls within the same brackets cannot be
distinguished on the basis of the parameter considered
Fig. 3 Classification of crocodilian calls by discriminant function
analysis. The DFA correctly classified 96 % of the Nile crocodile
contact calls, 88 % of the Black Caiman contact calls and 80 % of the
spectacled caiman calls
Fig. 4 Behavioural reaction of Nile crocodile juveniles to contact
calls from different species and to white noise (each of the 5 tested
individuals is represented by a different colour) (colour figure online)
1100 Anim Cogn (2012) 15:1095–1109
123

individual of the cluster was at least 10 m from the loud-
speaker). The amplitude of emitted sounds was of
61 ± 4 dBSPL at 1 m from the loudspeaker. The maximum
range of propagated signals from the loudspeaker was
approximately 20 m (at this distance, the intensity level
was below the background noise). Given the minimum
distance between clusters (30 m), we assumed that indi-
viduals within a cluster were not able to hear the signals
emitted during another cluster’s test. The experiments took
place in April, a month when most adults are sexually
active. Females had already started to lay their eggs; males
were defending their harems and thus tended to stay around
the same area in the greenhouse (±10 m). In addition, all
the experiments only lasted 2 days, so it is likely that the
composition of clusters remained stable during this time
and that no animal was tested twice.
To avoid habituation, each cluster was challenged once,
with only one experimental signal. Due to the limited
number of clusters that could be tested independently
(N = 5), we thus only used 5 experimental signals chosen
from those used for Nile crocodile juvenile tests: (1) a
natural contact call (control signal, NAT, number of adult
crocodiles within the tested cluster n = 24), (2) a signal
without amplitude modulation (noAM, n = 7), (3) a signal
without any harmonic structure (1H, n = 20), (4) a
signal without frequency modulation (FM1, n = 7), (5) a
signal without any frequency modulation nor harmonic
series (FM2, n = 20).
Behavioural responses were assessed according to a
4-level scale: 0 was scored if the playback did not provoke
any observable response during the 90 s of playback, 1 if
there was a head and/or body orientation towards the
loudspeaker but no displacement, 2 if the individual moved
towards the loudspeaker but stayed at more than 5 m from
it, and 3 if it came at less than 5 m from the loudspeaker.
The behaviour of each animal within a cluster was assessed
independently, allowing calculation of the proportion of
females with scores of 0, 1, 2 and 3 (expressed in % to the
total number of observed females) for each tested cluster
and thus each experimental signal. Due to the limited
number of tests, no statistical tests were performed on these
adult data and only raw results are presented.
Results
The frequency modulation is a key parameter
for both juveniles and adults
Playback experiments run on juvenile Spectacled caimans
and Nile crocodiles showed that the shape of the frequency
modulation seems to be a biologically relevant call
parameter: the responses to FM1, 1Hrev and Srev were
significantly weaker than the responses to the synthetic
copy of the natural call (Scontrol) for both species (Fig. 6a;
Wilcoxon tests, in Spectacled caimans, N = 6, T = 0.0,
P = 0.028; N = 6, T = 1, P = 0.046; N = 6, T = 0.0,
Fig. 5 Experimental signals used for playback experiments (NAT:
natural contact call; Scontrol: synthetic copy of the natural
call = control signal; noAM: signal without amplitude modulation;
FM1: signal without frequency modulation; 1H: signal without any
harmonic structure; Srev: temporally reversed natural call; 1Hrev:
temporally reversed 1H signal; SLOPE1: signal with a frequency
modulation slope one-third reduced compared to the natural slope;
SLOPE2: signal with a frequency modulation slope half reduced
compared to the natural slope; FM2: signal without any frequency
modulation nor harmonics series; ENER: signal with a modified
energy distribution among the spectrum; NOISE: noise having the
same duration, frequency bandpass and amplitude modulation as the
control signal)
Anim Cogn (2012) 15:1095–1109 1101
123
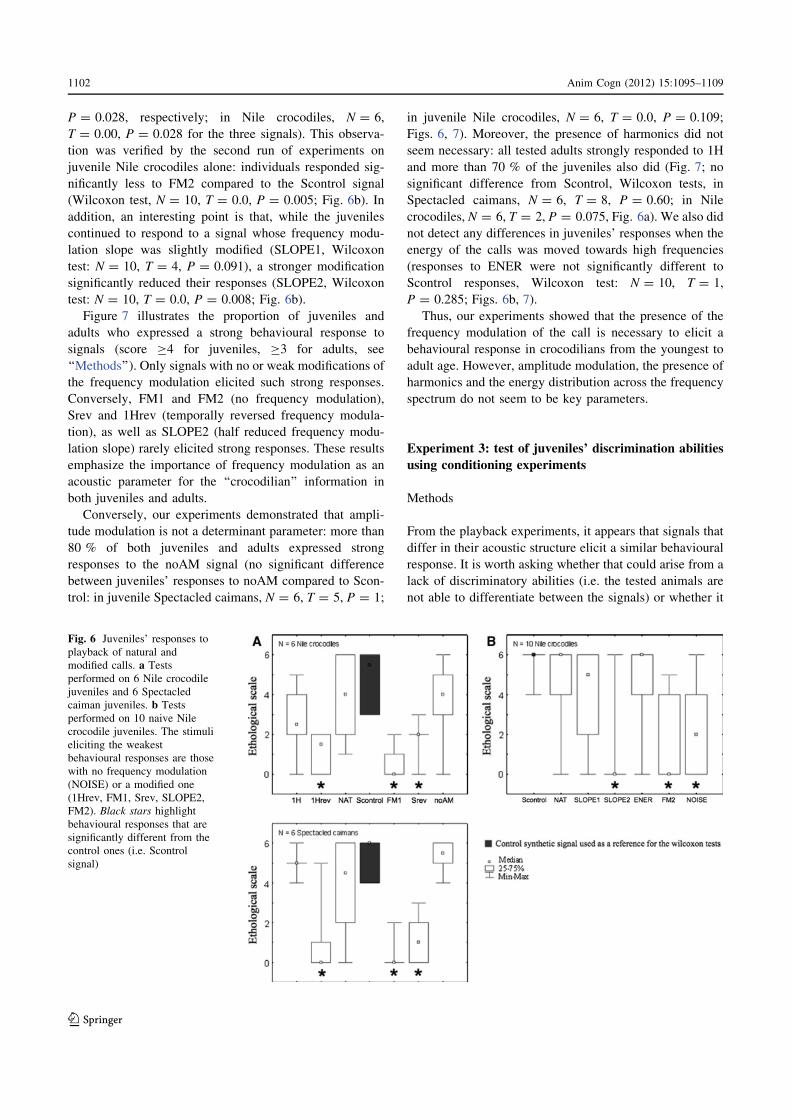
P = 0.028, respectively; in Nile crocodiles, N = 6,
T = 0.00, P = 0.028 for the three signals). This observa-
tion was verified by the second run of experiments on
juvenile Nile crocodiles alone: individuals responded sig-
nificantly less to FM2 compared to the Scontrol signal
(Wilcoxon test, N = 10, T = 0.0, P = 0.005; Fig. 6b). In
addition, an interesting point is that, while the juveniles
continued to respond to a signal whose frequency modu-
lation slope was slightly modified (SLOPE1, Wilcoxon
test: N = 10, T = 4, P = 0.091), a stronger modification
significantly reduced their responses (SLOPE2, Wilcoxon
test: N = 10, T = 0.0, P = 0.008; Fig. 6b).
Figure 7 illustrates the proportion of juveniles and
adults who expressed a strong behavioural response to
signals (score C4 for juveniles, C3 for adults, see
‘‘Methods’’). Only signals with no or weak modifications of
the frequency modulation elicited such strong responses.
Conversely, FM1 and FM2 (no frequency modulation),
Srev and 1Hrev (temporally reversed frequency modula-
tion), as well as SLOPE2 (half reduced frequency modu-
lation slope) rarely elicited strong responses. These results
emphasize the importance of frequency modulation as an
acoustic parameter for the ‘‘crocodilian’’ information in
both juveniles and adults.
Conversely, our experiments demonstrated that ampli-
tude modulation is not a determinant parameter: more than
80 % of both juveniles and adults expressed strong
responses to the noAM signal (no significant difference
between juveniles’ responses to noAM compared to Scon-
trol: in juvenile Spectacled caimans, N = 6, T = 5, P = 1;
in juvenile Nile crocodiles, N = 6, T = 0.0, P = 0.109;
Figs. 6, 7). Moreover, the presence of harmonics did not
seem necessary: all tested adults strongly responded to 1H
and more than 70 % of the juveniles also did (Fig. 7; no
significant difference from Scontrol, Wilcoxon tests, in
Spectacled caimans, N = 6, T = 8, P = 0.60; in Nile
crocodiles, N = 6, T = 2, P = 0.075, Fig. 6a). We also did
not detect any differences in juveniles’ responses when the
energy of the calls was moved towards high frequencies
(responses to ENER were not significantly different to
Scontrol responses, Wilcoxon test: N = 10, T = 1,
P = 0.285; Figs. 6b, 7).
Thus, our experiments showed that the presence of the
frequency modulation of the call is necessary to elicit a
behavioural response in crocodilians from the youngest to
adult age. However, amplitude modulation, the presence of
harmonics and the energy distribution across the frequency
spectrum do not seem to be key parameters.
Experiment 3: test of juveniles’ discrimination abilities
using conditioning experiments
Methods
From the playback experiments, it appears that signals that
differ in their acoustic structure elicit a similar behavioural
response. It is worth asking whether that could arise from a
lack of discriminatory abilities (i.e. the tested animals are
not able to differentiate between the signals) or whether it
Fig. 6 Juveniles’ responses to
playback of natural and
modified calls. a Tests
performed on 6 Nile crocodile
juveniles and 6 Spectacled
caiman juveniles. b Tests
performed on 10 naive Nile
crocodile juveniles. The stimuli
eliciting the weakest
behavioural responses are those
with no frequency modulation
(NOISE) or a modified one
(1Hrev, FM1, Srev, SLOPE2,
FM2). Black stars highlightbehavioural responses that are
significantly different from the
control ones (i.e. Scontrol
signal)
1102 Anim Cogn (2012) 15:1095–1109
123

could be the result of a true behavioural choice (i.e. the
tested animals perceive the difference between calls of
different acoustic structures but do not show any differ-
ential responses to them). This situation (similar responses
to structurally different calls) occurred during our playback
experiments on juvenile Nile Crocodiles when, for
instance, we compared the behavioural responses to the
control synthetic Scontrol signals and to the ENER signals
(Scontrol signals = synthetic copies of natural calls;
ENER = signals with a modified energy distribution
among the spectrum, that is, 80 % of energy between 2,500
and 5,000 Hz instead of between 200 and 2,500 Hz in the
Scontrol signals). With the setup of experiment 2, behav-
ioural responses elicited by Scontrol and ENER signals
were not different (see ‘‘Results’’ of experiment 2 above);
however, this result does not prove that the tested animals
are not able to distinguish between both the two. It is
possible that both signals have similar biological relevance.
Conditioning experiments are a way to test whether this
undifferentiated response is linked to a something other
than discriminatory ability. Although conditioning experi-
ments have already been used in several situations with
crocodiles (Davidson 1966; Williams 1968; Burghardt
1977, to our best knowledge, this method had never been
used on crocodiles to test their discrimination abilities with
acoustic signals. To test whether young crocodiles can
learn to respond to a sound stimulus, we first started with a
simple GO/NOGO task. Second, we used a GO/NOGO
procedure to test the crocodile’s ability to discriminate
between Scontrol and ENER signals.
Animals and experimental conditions
Three naıve young Nile crocodiles (1 month old) partici-
pated in the experiments. The home tank in which the
individuals were kept had the same characteristic as in the
previous experiments. Before the beginning of condition-
ing experiments, we stopped feeding the young crocodiles
in their home tank. Feeding was used as a reinforcer during
the conditioning tasks and thus occurred only during
experimental periods when the individual was isolated
from the two others in the test tank. As the crocodiles were
used to being fed once every 3 days before the experi-
ments, we maintained this feeding rhythm and tested each
of the three crocodiles every 3 days. All experiments
occurred during the night between 8 p.m. and 5 a.m. (water
temperature: 30 �C; air temperature: 28–30 �C). All trialswere video-recorded. The tested individual was isolated in
the test tank the morning before the test. We used 4
‘‘speaking’’ feeders (CAT Gato) located in the four corners
of the test tank (Fig. 8a). The feeders were put on bricks at
about 5 cm above the water level, not only to protect them
from getting wet but also so that it would require a physical
effort for the crocodile to access them and check for the
presence of food. Each speaking feeder was composed of
four compartments of exactly the same size (Fig. 8b). The
opening of each compartment was electronically pro-
grammable. It took 15 s for each compartment to open
completely, and during that time, a soft and regular
mechanical noise could be heard. Once the compartment
was open, we could choose to programme the feeder to
play 20 s of a sound sequence or to remain silent. Each
feeder possessed an integrated microphone and a loud-
speaker (Fig. 8b).
Experimental signals and conditioning protocol
The first conditioning protocol (pre-training) aimed at
testing whether it was possible to condition a crocodile to
respond to a feeder opening associated with a sound
Fig. 7 Proportion of
responding individuals to
experimental signals (score C 4
for juveniles, C3 for adults).
The behavioural scores were
established according to a scale
(see text). Only few individuals
showed a strong behavioural
response to the signals with no
or disrupted frequency
modulation (Srev, 1Hrev, Noise,
SLOPE2, FM1 and FM2).
n = total number of tested
individuals (Nil = juvenile Nile
crocodiles; Spec = juvenile
Spectacled caimans;
Ad = adult Nile crocodile)
Anim Cogn (2012) 15:1095–1109 1103
123

stimulus. On the morning before any nocturnal test and
before we brought the crocodile in the experimental room,
the test tank was organized with two ‘‘food’’ feeders filled
with food (the four compartments of each feeder were each
filled with a piece of food, for example, fish, frog’s legs,
shrimps, chicken, meat…) and programmed to play 20 s of
a pre-recorded sound sequence (3 repetitions of the same
series; one series = 7 successive Scontrol signals from
different individuals; interval between Scontrol sig-
nals = 1 s; interval between series = 1.5 s). The pre-
recorded sound sequences differed between the 2 ‘‘food’’
feeders (different Scontrol signals), and we also made sure
not to use the same sequences from one night to another for
the same individual. The two other ‘‘no-food’’ feeders were
soaked with the smell of food (the same as the one placed
in the other feeders) but contained no food, and their
compartment opening was not programmed to be followed
by sounds. The position of the 4 feeders in each corner of
the tank was chosen at random and changed for every test.
We programmed the opening times in alternation between
the feeders so that every 30 min one opening occurred
from a given feeder. The order of openings between feeders
was randomly determined and changed for every test. The
experiment was run during 45 consecutive nights. Each of
the three individuals was tested every three nights (16
openings per night 9 15 nights/individual = 240 learning
trials/individual).
The second conditioning protocol (training) involved the
same three pre-trained crocodiles. There was no rest night
between the two protocols. The presence of food was still
associated with the Scontrol signal. However, we replaced
the silent signal associated with no food by a sound
sequence built with the ENER signal previously used
during the coding-decoding tests. The pre-recorded sound
sequences were different for each feeder (new Scontrol and
ENER signals), and we also made sure not to use the same
sequences from one night to another one on the same
individual. As in the first experiment, the two ‘‘no-food’’
feeders were soaked with the smell of the same kinds of
food as those placed in the other feeders, and the position
of the 4 feeders in each corner of the tank was chosen at
random and changed every day.
Measurements and analysis of behavioural responses
As for experiments 1 and 2, all conditioning tests were
controlled by a computer located outside the experimental
room. The audio and video recordings were set up to start
5 min before each feeder’s opening and to stay on for a
total of 15 min. We quantified the responses of the croc-
odiles using the video recordings by blindly assessing the
following 3 parameters (blind evaluation was achieved by
analyzing the behavioural responses without knowing what
was the sound stimuli that triggered them): (1) first, reac-
tion (head or body movement) of the tested individual
following the beginning of the feeder’s opening. We
expressed the number of positive responses (i.e. the event
‘‘crocodile reaction’’ occurred) as a percentage of the total
number of tests per category (i.e. no-food feeder or food
feeder). Every night, a crocodile was exposed to 16 fee-
der’s openings per category. In addition, we measured the
reaction latency T0 (i.e. the time in seconds between the
beginning of a feeder opening and the animal’s first reac-
tion). A 3-level scale was set up with a score of 3 assigned
when T0 B 10 s, 2 when 30 s B T0 B 11 s, 1 when
60 s B T0 B 31 s or 0 when T0[ 60 s (or no response).
(2) approach: the crocodile displacements towards the
feeder. A displacement was considered as an approach if
the crocodile came within 5 cm of the active feeder. As for
the reaction parameter, we expressed the number of
approaches as a percentage of the total number of tests per
feeder’s category. We also measured the time to approach
the feeder T1 (i.e. the time in seconds from the first reac-
tion to come within the 5 cm around a feeder). We used the
same scale as for the reaction parameter with 3 if
T0 B 10 s, 2 if 30 s B T0 B 11 s, 1 if 60 s B T0 B 31 s,
0 if T0[ 60 s. (3) climbing behaviour: once the crocodile
had reached the feeder, it had to decide whether or not to
Fig. 8 Test tank and speaking
feeders used during
conditioning experiments.
a Position of the 4 speaking
feeders at each corner of the test
tank. b Detail of a speaking
feeder. It is composed of 4
compartments whose openings
are programmable. The
integrated microphone was used
to record a 20 s sound sequence
which could be played via the
integrated loudspeaker
1104 Anim Cogn (2012) 15:1095–1109
123

climb on the feeder’s platform to look for the presence of
food. Note that every time a crocodile climbed onto a
feeder, it ate the food if it was available. We expressed the
number of climbing behaviours as a percentage of the total
number of tests per feeder category. We also measured the
climbing time T3 (i.e. the time in seconds from the end of
the approach to the feeder to come up onto the feeder to
check for the presence of food). The same scale as for the
two previous parameters was used.
For data analysis and statistical tests, we grouped each
three consecutive days of test (i.e. from day 1 to day 3
together, day 4 to day 6…until day 13 to day 15). For each
of these groups, we used Wilcoxon tests to compare
crocodiles’ responses to ‘‘food feeders’’ (associated to the
Scontrol signal) with their responses to ‘‘no-food feeders’’
(associated with Silence in the pre-training protocol or
ENER signals in the training protocol).
Results
Discrimination between calls with different energy
distribution over the frequency spectrum
The pre-training tests showed that it was possible to teach
the crocodiles to respond to a sound stimulation versus
silence. Results of the pre-training conditioning protocol
are shown Fig. 9. For the reaction parameter (Fig. 9a),
statistical tests show significant differences between
crocodiles’ reaction to ‘‘Scontrol feeders’’ compared to
‘‘Silent feeders’’ from the first day of test until the last:
crocodiles reactions were stronger in response to Scontrol
feeders. However, during the whole experiment, the tested
individuals were highly motivated and we have observed a
response in more than 80 % of all tests for both silent and
Scontrol feeders triggers. Times to react did not improve
with time or depend on the type of feeder (graphs not
shown). Conversely, the approach parameter (Fig. 9b)
shows that the crocodiles quickly learnt to associate the
presence of food with the Scontrol signal. This learning
was apparent in a significant decrease in approach behav-
iours towards the silent feeders. Until day 9, we still
observed a high percentage of errors (crocodiles approa-
ched both types of feeders in more than 80 % of the time:
no significant differences between responses to Scontrol
compared to Silence, D7–D9: P = 0.14, z = 2.38). How-
ever, from day 10, the number of approaches in response to
silent feeders decreased significantly (from 68 % of
approaches from day 10 to 12 to 55 % during the last
3 days of tests) while responses to Scontrol feeders remain
constant and close to 100 %. The speed of approaching the
active feeder did not change with time or depend on the
type of feeder. The climbing parameter was even more
discriminatory. Crocodiles climbed significantly less on
silent feeders compared to Scontrol feeders (Fig. 9c).
Indeed, while crocodiles climbed on Scontrol feeders more
than 90 % of the time, we observed a decrease from above
Fig. 9 Results of the pre-training tests: crocodiles learnt to associate
the presence of food with the Scontrol signal compared to silence.
a Reaction parameter: crocodiles answered during all the tests period
([80 % of reaction for both conditions) but analyses showed a
significantly stronger response to the Scontrol feeders compared to the
Silence feeders from the first days of experiment. b Approach
parameter: from day 10 the number of approaches in response to
Silence feeders decreased significantly (from 83 % of approaches at
day 7–9 to 55 % at day 13–15 of tests) while the number of
approaches to Scontrol feeders remained constant and close to 100 %.
c Climbing parameter: crocodiles climbed significantly less on
Silence feeders compared to Scontrol feeders and got better with
time (while crocodiles climbed on Scontrol feeders in more than 90 %
of the time, we observed a decrease from more than 60 % of climbing
behaviours at days 1–3 to less than 20 % at days 13–15 of tests in
response to Silence feeders). Results for Wilcoxon tests are presented
on each chart, *P\ 0.05, **P\ 0.02, ***P\ 0.01
Anim Cogn (2012) 15:1095–1109 1105
123

60 % of climbing behaviours the first 3 days to below
20 % the last 3 days of tests in response to silent feeders.
Also, times to climb did not change with time or depend on
the type of feeder. It is thus likely that crocodiles learnt
quickly to associate the presence of food with the Scontrol
sound sequence.
Second and interestingly, the training protocol showed
that crocodiles are able to discriminate between Scontrol
and ENER signals (Fig. 10). For the reaction parameter,
there is no significant difference between crocodiles’
responses to ‘‘Scontrol feeders’’ compared to ‘‘ENER
feeders’’ from the beginning to the end of our conditioning
tests (Fig. 10a). Furthermore, reaction times comparisons
still did not bring any information about the learning pro-
cess. The reaction parameter was thus not the most sensi-
tive for detecting learning. The crocodiles also seemed to
approach Scontrol feeders as much as ENER feeders (no
significant differences, from the beginning to the end;
Fig. 10b). Thus, the ‘‘approach’’ parameter did not help to
demonstrate any associative learning nor did the time to
make these displacements towards the two types of feeders.
However, the climbing parameter did show that crocodiles
learnt to differentiate between Scontrol and ENER signals
(Fig. 10c). From day 7, the number of climbing behaviours
in response to ENER feeders decreased significantly
compared to those in response to Scontrol feeders (between
70 and 92 % of climbing behaviours in response to Scon-
trol feeders during the 15 days versus a decrease from
85 % the first 3 days to about 50 % on the last 3 days of
tests in response to ENER feeders). Hence, crocodiles
learnt that the presence of food was associated with
Scontrol and not ENER and are thus able to differentiate
between these acoustic signals.
From a methodological point of view, note that playing
the sound after the feeder was opened gave us another clue
as to whether the crocodiles were able to learn that the
discrimination was based on sounds. The opening of the
feeder was relatively silent but crocodiles did react to it.
However, after a while we could see that the crocodiles
merely reacted to the sound and stayed motionless during
the feeder operation. This process confirmed that croco-
diles were attending to the broadcast sounds and not to the
opening of the feeders.
General discussion
The aim of this study was to investigate the acoustic
structure and information coding in the contact calls of
juvenile crocodilians. On the basis of acoustic analyses and
playback experiments, the following points have been
demonstrated:
First, in spite of common acoustic structure between
the contact calls of juvenile Nile crocodiles, Spectacled
caimans and Black caimans, most of the measured acoustic
parameters showed significant inter-specific differences
and it is straightforward to classify calls according to their
species origin using a multivariate analysis. Although calls
of all crocodilian species share the same overall acoustic
features, previous studies had already shown inter-specific
Fig. 10 Results of the training test: crocodiles were able to learn
discriminating between Scontrol and ENER signals. a Reaction
parameter: crocodiles did not express any significant differences in
their responses to Scontrol versus ENER feeders. They reacted to both
feeders in more than 80 % of the tests. b Approach parameter: they
approached both feeders and did not seem to learn with time not to
approach ENER feeders. c Climbing parameter: conversely, from day
7, crocodiles started climbing significantly less on ENER feeders
compared to Scontrol ones (between 70 and 92 % of climbing
behaviours in response to Scontrol feeders during the 15 days versus a
decrease from 85 % at days 1–3 to about 50 % at days 13–15 in
response to ENER feeders). Results for Wilcoxon tests are presented
on each chart, *P\ 0.05, **P\ 0.02, ***P\ 0.01
1106 Anim Cogn (2012) 15:1095–1109
123

variations (reviewed in Vergne et al. 2009). However, these
inter-specific differences may be biased in the present
study due to the relative age heterogeneity of our animals:
we showed in a previous study with newborn Nile croco-
diles’ distress calls that acoustic variables related to call
duration and to the fundamental frequency vary with the
individual’s age (Vergne et al. 2007). Specifically, the
youngest/smallest individuals produce the highest-pitched
calls. This may explain why the contact calls of the Nile
crocodile we studied possessed more energy towards high
frequencies and a broader frequency bandwidth than those
of the spectacled or black caimans. Nevertheless, the slope
of the frequency modulation has been found to remain
stable with the age in Nile crocodile’s distress calls (Ver-
gne et al. 2007), and in the present study, this parameter
was one of those that distinguished most strongly between
the contact calls of the different species. Hence, the vari-
ability in age between the animals recorded is unlikely to
explain all the observed differences and it is not unrea-
sonable to assume that inter-specific differences in the fine
structure of juvenile calls do exist.
Second, playback experiments showed that these inter-
specific structural differences between calls seemed not to
be relevant to the animals. We observed no significant
behavioural differences in juvenile Nile crocodile’s
responses to calls of their own species compared to calls of
other species. We have to be cautious with this result
because of the small sample size and unknown power of
the experimental system: in a more subtle paradigm and a
fortiori in the wild, discrimination between species calls
remains conceivable. Nevertheless, this result supports a
previously established hypothesis about a possible inter-
specific repertoire of crocodilian calls with behavioural
responses not restricted to the conspecific calls (Britton
2001; Campbell 1973) and is in accordance with several
preliminary experiments we have made in the wild. That
gives us several reasons to think that species-specific rec-
ognition based on juvenile calls is extremely weak or
nonexistent in crocodiles. The species specificity of juve-
nile calls may be irrelevant information for these animals.
One main point is that effective sympatry is rare among
crocodiles. The Nile crocodile lives in Africa while Black
and Spectacled caimans are American species. This geo-
graphical distribution might explain why the Nile croco-
diles have not developed the ability to discriminate their
calls from those of other species. The Black and the
Spectacled caimans are sympatric but do not usually meet
in the field, their preferential habitat being slightly different
(and observations have been made of Black caimans
chasing the other species, P. Taylor pers.com.). Based on
our own observations in the field, it is likely that acoustic
exchanges between juveniles and between adults and
juveniles occur only within family groups (i.e. a female and
her young; for example, Black caiman family clusters stay
away from other individuals, Vergne et al. 2011).
Third, our study shed light on the key acoustic param-
eters responsible for the biological relevance of the juve-
niles’ contact calls and yielded information on the process
of coding and decoding crocodilian information by juve-
niles and adults. The importance of the slope of the fre-
quency modulation and the tolerance of tested individuals
to slight modifications of this parameter is in accordance
with the inter-specific responses we observed during
Experiments 1, as FM slopes differed slightly between
species. An information encoding process using frequency
modulation is widespread in animals using acoustic to
communicate (Becker 1982). A coding based on a slow
frequency modulation has the advantage of being robust,
particularly in the face of propagation, because modulation
characteristics are only slightly damaged during transmis-
sion over long ranges (Wiley and Richards 1982). Also, as
the crocodilian contact call’s frequency modulation
extends over a large frequency band, its characteristics are
useful for improving the localization of the sound source
(Aubin and Jouventin 2002b). In addition, playback
experiments using modified signals showed that amplitude
modulation does not play a major role in inducing a
behavioural response. It is known that this parameter is
quickly modified during signal transmission throughout the
environment (Aubin et al. 2000; Mathevon and Dabelsteen
2002; Wiley and Richards 1982). Previous propagation
experiments using crocodilian calls have confirmed this
result (Vergne et al. pers. obs.). In birds (Jouventin et al.
1999) and mammals (Charrier et al. 2002), amplitude
modulation is never a parameter encoding specific or
individual identity. Nevertheless, this parameter can play a
crucial role during sound localization as has been demon-
strated for instance in the barn owl Tyto alba (Konishi
1973; Shalter and Schleidt 1977) and in the King Penguin
Aptenodytes patagonicus (Aubin and Jouventin 2002b).
Further experiments would be necessary to determine
whether this is also the case in crocodilians. Playback
experiments also showed that the entire frequency spec-
trum is not necessary to induce a behavioural response.
One unique harmonic signal seems to be enough to main-
tain the biological effectiveness of the signal. Experiments
with birds and mammals have shown that tolerance
towards such a modification is extremely variable from
one species to another. For instance in Adelie Penguin
Pygoscelis adeliae, parent-offspring recognition needs the
entire frequency spectrum while the same type of recog-
nition in Subantarctic Fur Seal Arctocephalus tropicalis or
in King Penguins is effective with a reduced number of
harmonics (Aubin and Jouventin 2002a; Charrier et al.
2002; Searby et al. 2004). Young crocodilians seem to be
quite tolerant towards modifications of this parameter. Just
Anim Cogn (2012) 15:1095–1109 1107
123

as for amplitude modulation, it does not mean that this
parameter could not contribute to the communication
process. In particular, the fact that harmonic series extend
over a large spectral bandwith is also likely to considerably
improve the localization of the emitter (Wiley and Richards
1982). We have to remember that calls are often emitted in
a context when the emitter is soliciting parental care or in
order to gather siblings: being easily localized by
receiver(s) thus makes sense. In the field, it is known that
adult crocodiles can localize juvenile calls accurately (Hunt
and Watanabe 1982; Passek and Gillingham 1999). How-
ever, research on sound localization in crocodilians is still
in its infancy. Only one study (Carr et al. 2009) has
investigated neurophysiological processes involved in
localization tasks. Future experiments are necessary to test
whether sound energy distribution across a wide spectrum
effectively reinforces the reliability of localization of the
emitter in crocodiles. Finally, we observed that shifting the
signal’s energy towards high frequencies (ENER signals)
did not seem to modify the level of response of the animals.
In crocodilians, the youngest and smallest individuals are
the ones who produce the highest-pitched sounds with
more energy towards high frequencies, especially in
stressful conditions (emission of high-pitched distress calls,
Vergne et al. 2007). Playback experiments would be nec-
essary to discover whether age information or stress level
could be carried in the calls and could trigger different
behavioural responses on the receiver’s side. However,
testing crocodilians’ abilities to discriminate between high-
pitched calls and natural calls via conditioning experiments
was a first step and further investigations are now needed.
Indeed, fourth and finally, the conditioning experiments
showed that crocodilians are able to learn an associative
task based on acoustic stimuli. In our experiment, indi-
viduals learnt to discriminate between a control contact call
and a modified call with more energy towards high fre-
quencies. Lack of discriminatory abilities thus cannot
explain why crocodilians did not express different behav-
ioural responses during playback experiments using these
two signals. It is likely that we observed the results of true
behavioural choices by the individuals tested. However,
this conditioning experiment remains quite preliminary and
requires further investigations. For instance, if tested ani-
mals could discriminate between both control and modified
calls, we must wonder why they would approach both
loudspeakers equally. We of course took great care to give
the same odour to all feeders, and thus, a crocodile that has
approached a feeder was unlikely to get olfactory cues to
tell whether there was food available. Also, from a com-
parison between climbing behaviours during the pre-
training and training experiments (Figs. 9c, 10c), it appears
that there were far more errors (i.e. animals climbing on the
wrong no-food feeder) when no-food feeders were
associated with the modified sound stimulus than with
silent no-food feeders. If crocodiles had cues by which they
could identify feeders with food, they should have suc-
ceeded equally in the two tests. So, a reasonable hypothesis
is that crocodiles had some difficulty in distinguishing
between the two acoustic stimuli whereas they had of
course no difficulty in distinguishing between the control
sound and silence during the pre-training conditioning
experiment. In spite of possible unknown biases in the
experimental setup, and although field experiments are
irreplaceable, this first investigation using conditioning to
assess discrimination between acoustic signals enhances
the interest of testing crocodiles in the laboratory.
This study has given new insights on acoustic com-
munication in crocodilians. In addition to previous work
that has demonstrated the biological roles of sounds in
crocodilians, especially in the context of parent-offspring
interactions (Vergne et al. 2009, 2011), here, by studying
the features (acoustic structure and information coding) of
a crocodile sound, we have found evidence that crocodiles
calls are true communication signals. A crocodile sound
carries information in the form of a code decipherable by
crocodilian receivers and which includes several acoustic
characteristics likely to make acoustic communication in
crocodilians particularly effective. Crocodiles appear to
be tolerant of substantial modifications of the acoustic
structure of juvenile calls. A previous study has shown
that adults are also tolerant to variations of adult signals
(Wang et al. 2009). Taken together, these results under-
line that information coding is resistant to signal degra-
dation. Crocodilians together with birds are the modern
representatives of the Archosauria phylum that includes
the extinct dinosaurs and pterosaurs (Hopson 1975;
Walker 1972). Most species of these modern archosaurs
provide parental care in terms of food provisioning (birds)
and protection (birds and crocodiles), and sound signals
play a major role in these parent-offspring interactions.
Birds and crocodiles share several important traits
regarding acoustic communication, and this supports the
hypothesis that one of the shared behavioural features of
past and present Archosaurs is the use of acoustic com-
munication, especially in the context of parental care and
sibling interactions.
Acknowledgments We thank Luc Fougeirol and the staff of ‘‘La
Ferme aux Crocodiles’’, Pierrelatte, France, for their great help. We
thank Peter Taylor and his staff for help with logistics during the field
study in Guyana; and Toshao Isaac Rogers for permission to conduct
work at Yupukari. This study was funded by the Institut universitaire
de France (NM), the Centre National de la Recherche Scientifique
(TA) and the Ministere de l’Education Nationale et de la Recherche
(AV). Experiments were performed under the authorization no
42–218-0901–38 SV 09, (ENES Lab, Direction Departementale des
Services Veterinaires de la Loire) and were in agreement with the
French legislation regarding experiments on animals.
1108 Anim Cogn (2012) 15:1095–1109
123

References
Aubin T (1994) Syntana: a software for the synthesis and analysis of
animal sounds. Bioacoustics 6:80–81
Aubin T, Jouventin P (2002a) How to identify vocally a kin in a
crowd? The penguin model. Adv Stud Anim Behav 31:243–277
Aubin T, Jouventin P (2002b) Localisation of an acoustic signal in a
noisy environment: the display call of the king penguin
Aptenodytes patagonicus. J Exp Biol 205:3793–3798
Aubin T, Jouventin P, Hildebrand C (2000) Penguins use the two-
voice theory to recognise each other. Proc R Soc Lond B 267:
1081–1087
Becker PH (1982) The coding of species-specific characteristics in
bird sounds. In: Kroodsma DE, Miller EH (eds) Acoustic
communication in birds, vol 1. Academic Press, New York
Britton ARC (2001) Review and classification of call types of juvenile
crocodilians and factors affecting distress calls. In: Grigg GC,
Seebacher F, Franklin CE (eds) Crocodilian biology and evolu-
tion. Surrey Beatty & Sons, Chipping Norton, pp 364–377
Burghardt GM (1977) Learning processes in reptiles. In: Gans C
(ed) Biology of the reptilia, vol 7. Academic Press, New York,
pp 555–681
Campbell HW (1973) Observations on the acoustic behavior behav-
iour of crocodilians. Zoologica 58:1–11
Carr CE, Soares D, Smolders J, Simon JZ (2009) Detection of interaural
time differences in the alligator. J Neurosc 29:7978–7982
Charrier I, Mathevon N, Jouventin P (2002) How does a fur seal
mother recognize the voice of her pup? An experimental study of
Arctocephalus tropicalis. J Exp Biol 205:603–612
Davidson RS (1966) Operant stimulus control applied tomaze behavior
behaviour: heat escape conditioning and discrimination reversal in
Alligator mississipiensis. J Exp Anal Behav 9:671–676
Dooling RJ, Brown SD, Klump GM, Okanoya K (1992) Auditory
perception of conspecific and heterospecific vocalizations in
birds: evidence for special processes. J Comp Psychol 106:20–28
Garrick LD, Lang JW, Herzog HA (1982) Social signals of adult
American alligators. Bul Am Mus Nat Hist 160:53–192
Herzog HA, Burghardt GM (1977) Vocalization in juvenile crocod-
ilians. Z Tierpsychol 44:294–303
Hopson JA (1975) The evolution of cranial display features in
hadrosaurian dinosaurs. Paleobiology 1:21–43
Hunt R, Watanabe M (1982) Observations on maternal behavior of
the American alligator (Alligator mississipiensis). J Herpetol
15:235–239
Jouventin P, Aubin T, Lengagne T (1999) Finding a parent in a king
penguin colony: the acoustic system of individual recognition.
Anim Behav 57:1175–1183
Konishi M (1973) Locatable and nonlocatable acoustic signals for
barn owls. Am Nat 107:775–785
Mathevon N, Dabelsteen T (2002) Why do songbirds sing intensively
at dawn? A test of the acoustic transmission hypothesis. Acta
Ethol 4:65–72
Passek KM, Gillingham JC (1999) Absence of kin discrimination in
hatching American alligators, Alligator mississipiensis. Copeia3:831–835
Searby A, Jouventin P, Aubin T (2004) Acoustic recognition in
macaroni penguins: an original signature system. Anim Behav
67:615–625
Senter P (2008)Homology and antiquity of stereotyped communicatory
behavior behaviours of crocodilians. J Herpetol 42:354–360
Shalter M, Schleidt W (1977) The ability of barn owls Tyto alba to
discriminate and localize avian alarm calls. Ibis 119:22–27
Trutnau L, Sommerlad R (2006) Crocodilians. Their natural history
and captive husbandry. Brahm, A S, Frankfurt
Vergne AL, Mathevon N (2008) Crocodile egg sounds signal hatching
time. Cur Biol 18:12
Vergne AL, Avril A, Martin S, Mathevon N (2007) Parent-offspring
communication in the Nile crocodile Crocodylus niloticus: donewborns’ calls show an individual signature? Naturwissens-
chaften 94:49–54
Vergne AL, Mathevon N, Pritz MB (2009) Acoustic communication
in crocodilians: from behaviour to brain. Biol Rev 84:391–411
Vergne AL, Aubin T, Taylor P, Mathevon N (2011) Acoustic signals
of baby black caimans. Zoology 114:313–320
Walker AD (1972) New light on the origin of birds and crocodiles.
Nature 237:257–263
Wang X, Wang D, Wu X, Wang C, Wang R, Xia T (2009) Response
specificity to advertisement vocalization in the Chinese alligator
(Alligator sinensis). Ethology 115:832–839
Wiley RH, Richards DG (1982) The coding of species-specific
characteristics in bird sounds. In: Kroodsma DE, Miller EH (eds)
Acoustic Communication in Birds. Academic Press, New York,
pp 214–244
Williams JT (1968) Reversal-learning in the spectacled caiman. Am J
Psychol 81:258–261
Anim Cogn (2012) 15:1095–1109 1109
123