top articles supplement - future medicine
TRANSCRIPT

Transgenerational Epigenetic Inheritance TOP ARTICLES SUPPLEMENT
CONTENTSEDITORIAL:: Epigenetics: where environment, society and genetics meet Epigenomics Vol. 6 Issue 1
REVIEW: Transgenerational epigenetic inheritance: adaption through the germline epigenome? Epigenomics Vol. 7 Issue 5
PERSPECTIVE: Transgenerational epigenetic inheritance: an open dicussion Epigenomics Vol. 7 Issue 5

1ISSN 1750-1911Epigenomics (2014) 6(1), 1–4
Keywords: environment • epigenetics • noncoding RNA • socioeconomic status • transgenerational epigenetic inheritance
Understanding how environment, social fac-tors and genetics combine to affect patterns of health is an urgent priority. Social factors that affect health patterns include race and socioeconomic status (SES), while biological factors include sex, genetics and epigenetics. The consequence of biology and environ-ment can be observed in life expectancy data. At 25 years old, life expectancy for white men is 4.4 years greater than for Afri-can–American men. Females follow a similar pattern. Income also affects life expectancy with high income earners outliving their low income counterparts, and white males and females outliving African–American males and females by 2–4 years at every income level [1]. Environment also affects perina-tal mortality. US-born African–American women experience greater rates of perinatal mortality than their foreign-born counter-parts [2]. Similar patterns can be observed with multiple health conditions, including cancer, depression, diabetes and heart disease highlighting the importance of this issue [1]. What is less clear is the mechanism by which these disparities are caused. One way in which biology and environment can both leave their fingerprint is through epigenetic changes.
Epigenetics refers to the regulation of gene expression through modifications to DNA or DNA-associated proteins. Epigenetic regula-tion includes histone modifications, DNA methylation and nonprotein coding RNA. DNA methylation has been a particular focus of environment-mediated changes in health. Methylation changes have been associated
with environmental chemical exposure [3], race, gender [4] and income level [5]. How-ever, epigenetic gene regulation is bigger than methylation alone.
Human and animal studies have begun to investigate environmental effects on epi-genetics. While these publications are increas-ing, some factors need to be considered, these include tissue specificity, epigenetic specific-ity and transgenerational studies.
Tissue specificityTissue-specific epigenetic profiles are well accepted, and so it must be noted that blood cells also have cell-specific profiles. Circulat-ing blood is a convenient source of material for epigenetic testing. It is minimally invasive to collect and can be collected multiple times over a period of time. However, develop-ment and environment changes the cellular makeup of blood.
Cellular heterogeneity may confound epi-genetic results, since epigenetics differs from one blood cell to another [6,7]. For example, neutrophils predominate blood of a new-born, but, by 2–4 years, lymphocytes are the majority population [8]. Furthermore, cord blood also varies in its cellular make up compared to peripheral blood [9]. These two factors can confound the comparison of methylation profiles from a newborn to an adult, even in the same individual. The con-stituents of the blood can also be changed if the individual is experiencing inflammation, an infection or high amounts of stress [10–12]. All of these factors can be influenced by an individual’s SES. These differences highlight
Epigenetics: where environment, society and genetics meet
Amber V MajnikAuthor for correspondence:
Department of Pediatrics Medical
College of Wisconsin, Milwaukee,
WI, USA
Tel.: +1 414 955 2374
Robert H LaneDepartment of Pediatrics Medical
College of Wisconsin, Milwaukee,
WI, USA
“The goal is to use epigenetics to anticipate health in the individual and, more importantly, the population. Before this can be done, several
challenges must be faced.”
part of
Editorial
10.2217/EPI.13.83 © 2014 Future Medicine Ltd
Editorial

2 Epigenomics (2014) 11(1)
how developmental and environmental changes can alter epigenetic results.
The importance of different epigenetic patterns between specific cell types and whole blood will vary depending on disease or condition. Subtle differences are likely more important for multifactorial diseases, like obesity, where the environment, genetics and social factors all contribute to pathology. Furthermore, while methylation patterns may appear similar between cell types, changes in histone code or noncoding RNA may differ.
Epigenetic specificityWe believe social and environmental factors rely on epigenetics to effect biologic change. The study of how the environment affects phenotype is environmental epigenetics. A cell has many epigenetic tools to modify its gene expression in response to the environment. However, to understand environmental epigenetics, we must look beyond gene expression and protein coding regions and pay attention to regulatory and noncod-ing regions. This includes studying methylation and histone modifications, as well as noncoding RNA.
The majority of SES studies give a limited view of the epigenetic landscape. The majority of these stud-ies investigate SES and changes in global methylation. Some studies have taken an extra step to investigate methylation in disease relevant genes. However, to fully appreciate the epigenetic impact of the environ-ment, analysis must go beyond methylation. Animal models allow for a more detailed investigation of the epigenetic profile. Pena et al. analyzed DNA methyla-tion and histone modifications of the estrogen recep-tor alpha gene in the brains of dams and pups in a maternal care model [13]. These types of tissue specific, detailed epigenetic studies provide a more rigorous demonstration of epigenetic consequences.
While methylation is the best studied epigenetic mark, followed by histone modifications, the composi-tion of our DNA is telling us that RNA may be much more important. The discovery that <3% of the human genome encodes protein-coding genes was surprising to many people. But perhaps what is more astounding is that scientists have now found that approximately 76% of the human DNA is transcribed into some sort of RNA [14]. Much of the 76% is thought to have a regulatory role, including a role in epigenetics.
The role for noncoding RNA in epigenetics is just beginning to be understood; however, a number of studies indicate that RNA could be a powerful player in transgenerational epigenetics. The role for RNA as a mechanism of transfer of transgenerational phenotype come from experiments of microinjections of oliogri-bonucleotides into egg and sperm cells [15]. Given the
established effect of environment on epigenetics, envi-ronmental factors such as those effecting SES could disrupt this process. While the role for noncoding RNA is still being investigated, many studies have begun to investigate how the environment can change the life course of multiple generations and the role of epigenetics in this process.
Transgenerational epigenetic inheritanceOne of the most challenging and intriguing aspects of epigenetics is the transmission of modifications to sub-sequent generations. Transgenerational studies indicate that epigenetic modifications are not wiped clean for each generation, as was originally thought [16,17]. This, transgenerational epigenetic inheritance, coupled with the environmental influences of epigenetics indicates that the environment of one generation can modify the phenotype of the following generations. This far-reaching epigenetic effect indicates the importance of understanding environmental influences, such as SES, on epigenetics.
The life course approach determines how the social and environmental context of one generation can affect another. For example, a maternal grandmother’s exposure to neighborhood poverty during pregnancy predicts the risk for infant low birth weight and sub-sequent mortality among African–American women, independent of the mother’s status [18]. A detailed understanding of transgenerational epigenetics is nec-essary to understand this complex inheritance.
Stress is a core component of socioeconomic disad-vantage. Both prenatal as well as postnatal stress has been shown to effect long-term health. Epigenetic changes in offspring exposed to maternal stress have been noted in multiple species. In Drosophila, heat stress induces phosphorylation of the transcription fac-tor dATF-2 [19]. This leads to a loss of heterochroma-tin structures at several region of the genome, which is transmitted to the next generation. In mice, the early life stress of chronic unpredictable maternal separation leads to changes in behavior and alters the methylation profile in the promoter of candidate genes in the germ-line and brain of affected males, as well as in the germ-line of the offspring [20]. The licking and grooming behavior of rats has also been shown to be associated with epigenetic changes and inherited [13].
Several additional aspects of SES have been modeled in studies of transgenerational epigenetic inheritance;
“The role for noncoding RNA in epigenetics is just beginning to be understood; however, a number of studies indicate that RNA could
be a powerful player in transgenerational epigenetics.”
future science group
Editorial Majnik & Lane

www.futuremedicine.com 3
one such aspect is prenatal diet. Areas of socioeconomic disadvantage are often part of food deserts, or areas with limited access to healthy affordable food [21]. This can lead to high-fat, low-nutrient diets and contribute to SES health disparity. This type of diet has negative consequences for pregnant women, as well as subse-quent generations. Animal studies have shown that maternal exposure to high-fat diet results in increased body size and reduced sensitivity to insulin through two generations via both the maternal and paternal lineages [22]. This effect continues into the third generation via the paternal lineage. This transgenerational epigenetic inheritance of diet is associated with imprinted gene patterns and changes in methylation.
Transgenerational epigenetic studies have also begun to investigate the social and racial disparity in toxicant burden. Economically disadvantaged areas have an increased risk of environmental toxin exposure [23–25]. These toxins include endocrine disruptors, heavy met-als, tobacco smoke and air pollution, among others. Similar to diet studies, exposure of gestating females to environmental toxins leads to increased disease for multiple generations [3]. Also associated with the toxin derived phenotype are changes in epigenetics. Several studies have found changes in differential methylation regions, which are proposed to act as biomarkers of exposure [3].
It is clear that numerous environmental factors can contribute to an individual’s epigenetic inheritance of health. The next step forward in advancing our under-standing should be to integrate multiple environmen-tal influences. This will not only create a better life course model, but the increased complexity will help to discern the causality of health disparities.
Epigenetics exists at the intersection between genet-ics and the environment. The goal is to use epigenetics to anticipate health in the individual and, more impor-tantly, the population. Before this can be done, several challenges must be faced. These include investigating the nonprotein coding region of the genome, studying common diseases where the environment likely contrib-utes to the gradation of severity, and developing new, complex models that include multiple environmental influences and have transgenerational impacts.
Financial & competing interests disclosureThe authors have no relevant affiliations or financial involve-
ment with any organization or entity with a financial inter-
est in or financial conflict with the subject matter or mate-
rials discussed in the manuscript. This includes employment,
consultancies, honoraria, stock ownership or options, expert
testimony, grants or patents received or pending, or royalties.
No writing assistance was utilized in the production of this
manuscript.
References1 Williams DR, Mohammed SA, Leavell J, Collins C. Race,
socioeconomic status, and health: complexities, ongoing challenges, and research opportunities. Ann. NY Acad. Sci. 1186, 69–101 (2010).
2 Collins JW Jr, Soskolne GR, Rankin KM, Bennett AC. Differing first year mortality rates of term births to white, African-American, and Mexican-American US-born and foreign-born mothers. Matern. Child Health J. 17(10), 1776–1783 (2012).
3 Manikkam M, Tracey R, Guerrero-Bosagna C, Skinner MK. Plastics derived endocrine disruptors (BPA, DEHP and DBP) induce epigenetic transgenerational inheritance of obesity, reproductive disease and sperm epimutations. PLoS ONE 8(1), e55387 (2013).
4 Zhang FF, Cardarelli R, Carroll J et al. Significant differences in global genomic DNA methylation by gender and race/ethnicity in peripheral blood. Epigenetics 6(5), 623–629 (2011).
5 Tehranifar P, Wu HC, Fan X et al. Early life socioeconomic factors and genomic DNA methylation in mid-life. Epigenetics 8(1), 23–27 (2013).
6 Wu HC, Wang Q, Delgado-Cruzata L, Santella RM, Terry MB. Genomic methylation changes over time in peripheral blood mononuclear cell DNA: differences by assay type and baseline values. Cancer Epidemiol. Biomarkers Prevent. 21(8), 1314–1318 (2012).
7 Adalsteinsson BT, Gudnason H, Aspelund T et al. Heterogeneity in white blood cells has potential to confound DNA methylation measurements. PloS ONE 7(10), e46705 (2012).
8 Orkin SH,Nathan DG, Ginsburg D, Look AT, Fisher DE, Lux SE,Nathan and Oski’s Hematology of Infancy and Childhood. Saunders Elsevier, PA, USA (2009).
9 Lopez MC, Palmer BE, Lawrence DA. Phenotypic differences between cord blood and adult peripheral blood. Cytometry B Clin. Cytom. 76(1), 37–46 (2009).
10 Landmann RM, Muller FB, Perini C, Wesp M, Erne P, Buhler FR. Changes of immunoregulatory cells induced by psychological and physical stress: relationship to plasma catecholamines. Clin. Exp. Immunol. 58(1), 127–135 (1984).
11 Williams DM, Johnson NW. Alterations in peripheral blood leucocyte distribution in response to local inflammatory stimuli in the rat. J. Pathol. 118(3), 129–141 (1976).
12 Bonneau RH, Sheridan JF, Feng NG, Glaser R. Stress-induced effects on cell-mediated innate and adaptive memory components of the murine immune response to herpes simplex virus infection. Brain Behav. Immun. 5(3), 274–295 (1991).
13 Pena CJ, Neugut YD, Champagne FA. Developmental timing of the effects of maternal care on gene expression and epigenetic regulation of hormone receptor levels in female rats. Endocrinology 154(11), 4340–4351 (2013).
14 Consortium TEP. An integrated encyclopedia of DNA elements in the human genome. Nature 489(7414), 57–74 (2012).
future science group
Epigenetics: where environment, society & genetics meet Editorial

4 Epigenomics (2014) 11(1)
15 Kiani J, Grandjean V, Liebers R et al. RNA–mediated epigenetic heredity requires the cytosine methyltransferase Dnmt2. PLoS Genet. 9(5), e1003498 (2013).
16 Lane N, Dean W, Erhardt S et al. Resistance of IAPs to methylation reprogramming may provide a mechanism for epigenetic inheritance in the mouse. Genesis 35(2), 88–93 (2003).
17 Guerrero-Bosagna C, Skinner MK. Environmentally induced epigenetic transgenerational inheritance of phenotype and disease. Mol. Cell. Endocrinol. 354(1–2), 3–8 (2012).
18 Collins JW Jr, David RJ, Rankin KM, Desireddi JR. Transgenerational effect of neighborhood poverty on low birth weight among African Americans in Cook County, Illinois. Am. J. Epidemiol. 169(6), 712–717 (2009).
19 Seong K-H, Li D, Shimizu H, Nakamura R, Ishii S. Inheritance of stress-induced, ATF-2-dependent epigenetic change. Cell 145(7), 1049–1061 (2011).
20 Franklin TB, Russig H, Weiss IC et al. Epigenetic transmission of the impact of early stress across generations. Biol. Psychiatry 68(5), 408–415 (2010).
21 Beaulac J, Kristjansson E, Cummins S. A systematic review of food deserts, 1966–2007. Prevent. Chronic Dis. 6(3), A105 (2009).
22 Dunn GA, Bale TL. Maternal high-fat diet effects on third-generation female body size via the paternal lineage. Endocrinology 152(6), 2228–2236 (2011).
23 Hajat A, Diez-Roux AV, Adar SD et al. Air pollution and individual and neighborhood socioeconomic status: evidence from the multi-ethnic study of atherosclerosis (MESA). Environ. Health Perspect. 121(11–12), 1325–1333 (2013).
24 Tyrrell J, Melzer D, Henley W, Galloway TS, Osborne NJ. Associations between socioeconomic status and environmental toxicant concentrations in adults in the USA: NHANES 2001-2010. Environ. Int. 59, 328–335 (2013).
25 Vrijheid M, Martinez D, Aguilera I et al. Socioeconomic status and exposure to multiple environmental pollutants during pregnancy: evidence for environmental inequity? J. Epidemiol. Commun. Health 66(2), 106–113 (2012).
future science group
Editorial Majnik & Lane

829Epigenomics (2015) 7(5), 829–846 ISSN 1750-1911
part of
Review
10.2217/epi.15.36 © 2015 Future Medicine Ltd
Epigenomics
Review 2015/07/287
5
846
2015
Epigenetic modifications direct the way DNA is packaged into the nucleus, making genes more or less accessible to transcriptional machinery and influencing genomic stability. Environmental factors have the potential to alter the epigenome, allowing genes that are silenced to be activated and vice versa. This ultimately influences disease susceptibility and health in an individual. Furthermore, altered chromatin states can be transmitted to subsequent generations, thus epigenetic modifications may provide evolutionary mechanisms that impact on adaptation to changed environments. However, the mechanisms involved in establishing and maintaining these epigenetic modifications during development remain unclear. This review discusses current evidence for transgenerational epigenetic inheritance, confounding issues associated with its study, and the biological relevance of altered epigenetic states for subsequent generations.
Keywords: epigenetic inheritance • epigenetics • evolution • germline • transgenerational epigenetic inheritance
Multicellular organisms are composed of hundreds of highly specialized cell types that are ultimately derived from a single cell, the zygote, which forms at fertilization. The diverse range of cell types found in each individual is accomplished through speci-fication and differentiation of cells during embryonic and fetal development. Changes in cell specialization are achieved through complex interactions involving cell signaling, transcriptional control and establishment of mitotically heritable epigenetic modifi-cations, which regulate global expression patterns and restrict cells to their defined lineages. Epigenetic modifications include DNA methylation and histone modifica-tions, which establish specialized chromatin states that restrict or permit gene expression. Although epigenetic modifications provide long-term cell lineage information, they are also dynamic and can respond to influences such as developmental cues or environmen-tal factors. This flexibility permits changes in cell differentiation and function during
embryonic development, while retaining lin-eage information and guiding organization of the body plan.
After fertilization, the zygote has the capacity to make all cell types in the body, a quality known as totipotency. As the embryo develops, cells lose totipotency and become restricted to specialized lineages. However, to ensure reproductive capacity, sexually reproducing species must maintain a popu-lation of germ cells that have the capability of regaining totipotency after fertilization. Thus, germ cells facilitate transmission of the genome and epigenome between parent and offspring in a manner that supports embryo-genesis. In mammals, this is achieved in part through the ability of germ cells to remove existing epigenetic information and estab-lish an epigenetic program that is compatible with totipotency.
Epigenetic reprogramming includes the widespread removal and re-establishment of epigenetic information, and occurs exten-sively in germ cells and preimplantation
Transgenerational epigenetic inheritance: adaptation through the germline epigenome?
Lexie Prokopuk1,2, Patrick S Western‡,1,2 & Jessica M Stringer*,‡,1,2
1Centre for Genetic Diseases, Hudson
Institute of Medical Research, 27–31
Wright Street, Clayton, Victoria 3168,
Australia 2Molecular & Translational Science,
Monash University, Clayton, Victoria 3168,
Australia
*Author for correspondence:
Tel.: +61 3 9594 7009
[email protected] ‡Authors contributed equally
For reprint orders, please contact: [email protected]
830 Epigenomics (2015) 7(5)
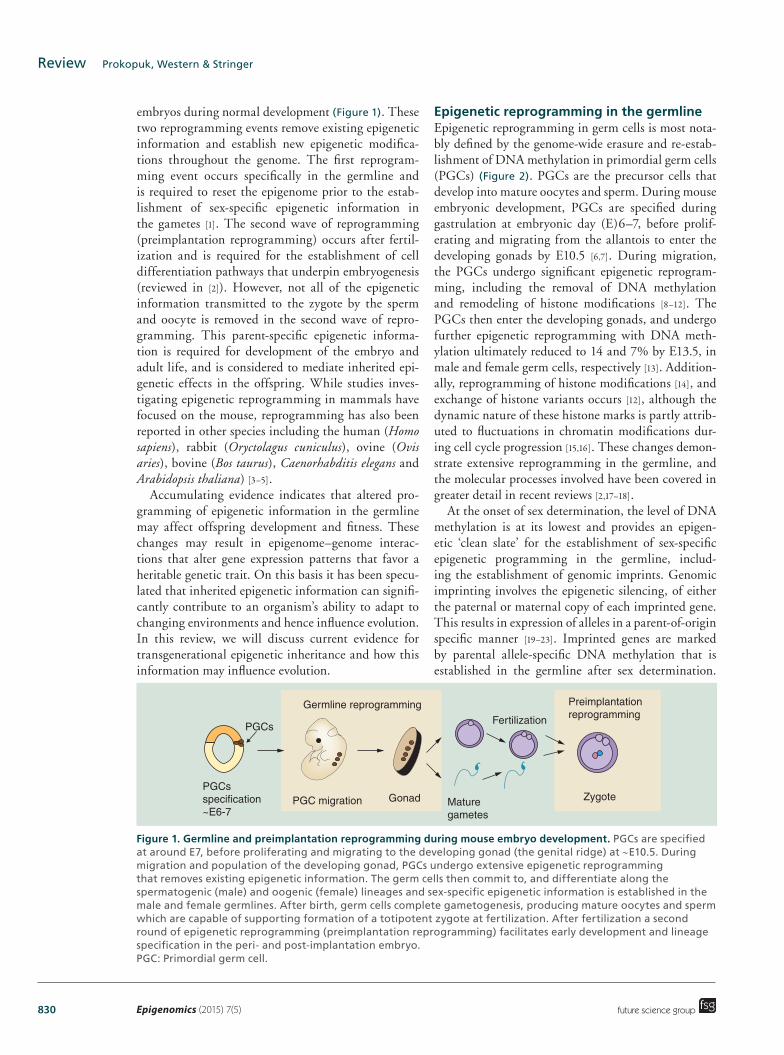
Figure 1. Germline and preimplantation reprogramming during mouse embryo development. PGCs are specified at around E7, before proliferating and migrating to the developing gonad (the genital ridge) at ∼E10.5. During migration and population of the developing gonad, PGCs undergo extensive epigenetic reprogramming that removes existing epigenetic information. The germ cells then commit to, and differentiate along the spermatogenic (male) and oogenic (female) lineages and sex-specific epigenetic information is established in the male and female germlines. After birth, germ cells complete gametogenesis, producing mature oocytes and sperm which are capable of supporting formation of a totipotent zygote at fertilization. After fertilization a second round of epigenetic reprogramming (preimplantation reprogramming) facilitates early development and lineage specification in the peri- and post-implantation embryo. PGC: Primordial germ cell.
Germline reprogramming
PGCs
PGCsspecification~E6-7
Preimplantationreprogramming
PGC migration Gonad Maturegametes
Fertilization
Zygote
future science group
Review Prokopuk, Western & Stringer
embryos during normal development (Figure 1). These two reprogramming events remove existing epigenetic information and establish new epigenetic modifica-tions throughout the genome. The first reprogram-ming event occurs specifically in the germline and is required to reset the epigenome prior to the estab-lishment of sex-specific epigenetic information in the gametes [1]. The second wave of reprogramming (preimplantation reprogramming) occurs after fertil-ization and is required for the establishment of cell differentiation pathways that underpin embryogenesis (reviewed in [2]). However, not all of the epigenetic information transmitted to the zygote by the sperm and oocyte is removed in the second wave of repro-gramming. This parent-specific epigenetic informa-tion is required for development of the embryo and adult life, and is considered to mediate inherited epi-genetic effects in the offspring. While studies inves-tigating epigenetic reprogramming in mammals have focused on the mouse, reprogramming has also been reported in other species including the human (Homo sapiens), rabbit (Oryctolagus cuniculus), ovine (Ovis aries), bovine (Bos taurus), Caenorhabditis elegans and Arabidopsis thaliana) [3–5].
Accumulating evidence indicates that altered pro-gramming of epigenetic information in the germline may affect offspring development and fitness. These changes may result in epigenome–genome interac-tions that alter gene expression patterns that favor a heritable genetic trait. On this basis it has been specu-lated that inherited epigenetic information can signifi-cantly contribute to an organism’s ability to adapt to changing environments and hence influence evolution. In this review, we will discuss current evidence for transgenerational epigenetic inheritance and how this information may influence evolution.
Epigenetic reprogramming in the germlineEpigenetic reprogramming in germ cells is most nota-bly defined by the genome-wide erasure and re-estab-lishment of DNA methylation in primordial germ cells (PGCs) (Figure 2). PGCs are the precursor cells that develop into mature oocytes and sperm. During mouse embryonic development, PGCs are specified during gastrulation at embryonic day (E)6–7, before prolif-erating and migrating from the allantois to enter the developing gonads by E10.5 [6,7]. During migration, the PGCs undergo significant epigenetic reprogram-ming, including the removal of DNA methylation and remodeling of histone modifications [8–12]. The PGCs then enter the developing gonads, and undergo further epigenetic reprogramming with DNA meth-ylation ultimately reduced to 14 and 7% by E13.5, in male and female germ cells, respectively [13]. Addition-ally, reprogramming of histone modifications [14], and exchange of histone variants occurs [12], although the dynamic nature of these histone marks is partly attrib-uted to fluctuations in chromatin modifications dur-ing cell cycle progression [15,16]. These changes demon-strate extensive reprogramming in the germline, and the molecular processes involved have been covered in greater detail in recent reviews [2,17–18].
At the onset of sex determination, the level of DNA methylation is at its lowest and provides an epigen-etic ‘clean slate’ for the establishment of sex-specific epigenetic programming in the germline, includ-ing the establishment of genomic imprints. Genomic imprinting involves the epigenetic silencing, of either the paternal or maternal copy of each imprinted gene. This results in expression of alleles in a parent-of-origin specific manner [19–23]. Imprinted genes are marked by parental allele-specific DNA methylation that is established in the germline after sex determination.
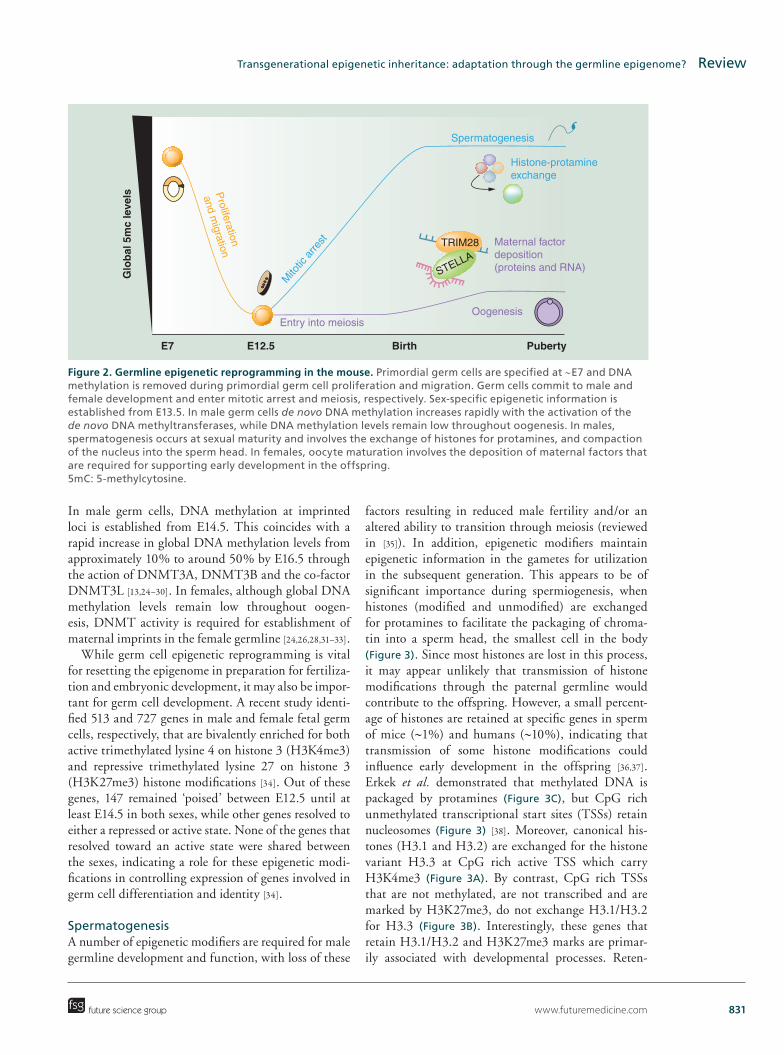
www.futuremedicine.com 831
Figure 2. Germline epigenetic reprogramming in the mouse. Primordial germ cells are specified at ∼E7 and DNA methylation is removed during primordial germ cell proliferation and migration. Germ cells commit to male and female development and enter mitotic arrest and meiosis, respectively. Sex-specific epigenetic information is established from E13.5. In male germ cells de novo DNA methylation increases rapidly with the activation of the de novo DNA methyltransferases, while DNA methylation levels remain low throughout oogenesis. In males, spermatogenesis occurs at sexual maturity and involves the exchange of histones for protamines, and compaction of the nucleus into the sperm head. In females, oocyte maturation involves the deposition of maternal factors that are required for supporting early development in the offspring. 5mC: 5-methylcytosine.
Glo
bal
5m
c le
vels
E7 E12.5 Birth Puberty
OogenesisEntry into meiosis
Proliferation
and migration
Mito
tic a
rrest
Spermatogenesis
Histone-protamineexchange
Maternal factordeposition(proteins and RNA)
TRIM28
STELLA
future science group
Transgenerational epigenetic inheritance: adaptation through the germline epigenome? Review
In male germ cells, DNA methylation at imprinted loci is established from E14.5. This coincides with a rapid increase in global DNA methylation levels from approximately 10% to around 50% by E16.5 through the action of DNMT3A, DNMT3B and the co-factor DNMT3L [13,24–30]. In females, although global DNA methylation levels remain low throughout oogen-esis, DNMT activity is required for establishment of maternal imprints in the female germline [24,26,28,31–33].
While germ cell epigenetic reprogramming is vital for resetting the epigenome in preparation for fertiliza-tion and embryonic development, it may also be impor-tant for germ cell development. A recent study identi-fied 513 and 727 genes in male and female fetal germ cells, respectively, that are bivalently enriched for both active trimethylated lysine 4 on histone 3 (H3K4me3) and repressive trimethylated lysine 27 on histone 3 (H3K27me3) histone modifications [34]. Out of these genes, 147 remained ‘poised’ between E12.5 until at least E14.5 in both sexes, while other genes resolved to either a repressed or active state. None of the genes that resolved toward an active state were shared between the sexes, indicating a role for these epigenetic modi-fications in controlling expression of genes involved in germ cell differentiation and identity [34].
SpermatogenesisA number of epigenetic modifiers are required for male germline development and function, with loss of these
factors resulting in reduced male fertility and/or an altered ability to transition through meiosis (reviewed in [35]). In addition, epigenetic modifiers maintain epigenetic information in the gametes for utilization in the subsequent generation. This appears to be of significant importance during spermiogenesis, when histones (modified and unmodified) are exchanged for protamines to facilitate the packaging of chroma-tin into a sperm head, the smallest cell in the body (Figure 3). Since most histones are lost in this process, it may appear unlikely that transmission of histone modifications through the paternal germline would contribute to the offspring. However, a small percent-age of histones are retained at specific genes in sperm of mice (∼1%) and humans (∼10%), indicating that transmission of some histone modifications could influence early development in the offspring [36,37]. Erkek et al. demonstrated that methylated DNA is packaged by protamines (Figure 3C), but CpG rich unmethylated transcriptional start sites (TSSs) retain nucleosomes (Figure 3) [38]. Moreover, canonical his-tones (H3.1 and H3.2) are exchanged for the histone variant H3.3 at CpG rich active TSS which carry H3K4me3 (Figure 3A). By contrast, CpG rich TSSs that are not methylated, are not transcribed and are marked by H3K27me3, do not exchange H3.1/H3.2 for H3.3 (Figure 3B). Interestingly, these genes that retain H3.1/H3.2 and H3K27me3 marks are primar-ily associated with developmental processes. Reten-
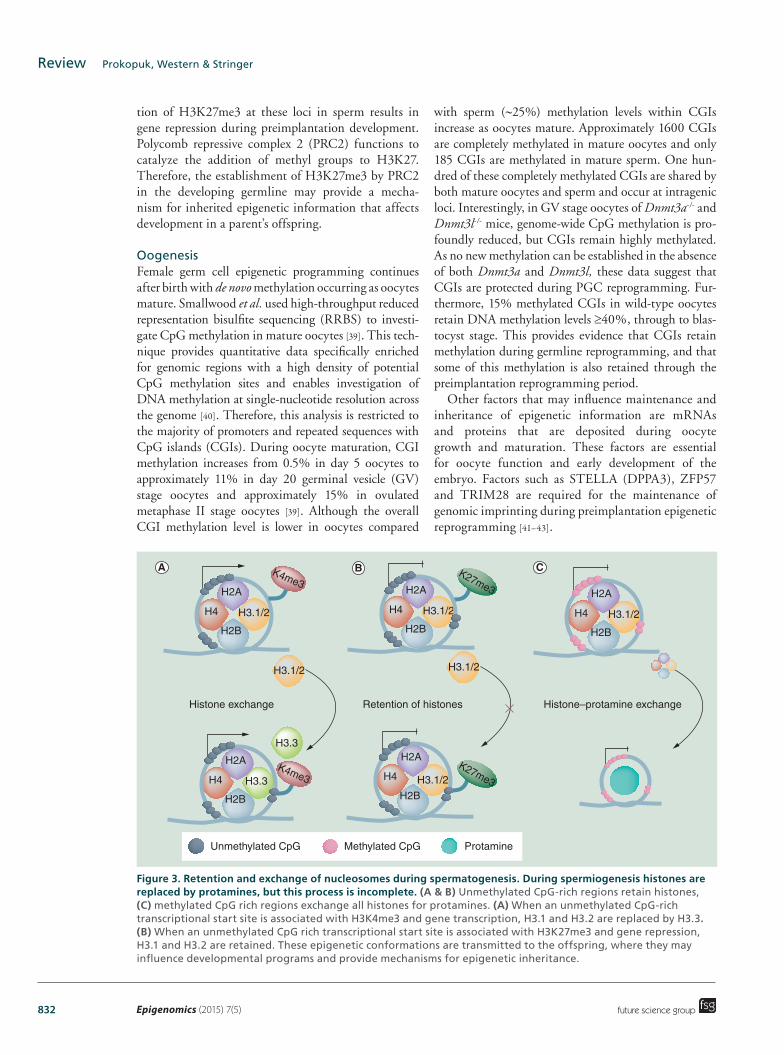
832 Epigenomics (2015) 7(5)
Figure 3. Retention and exchange of nucleosomes during spermatogenesis. During spermiogenesis histones are replaced by protamines, but this process is incomplete. (A & B) Unmethylated CpG-rich regions retain histones, (C) methylated CpG rich regions exchange all histones for protamines. (A) When an unmethylated CpG-rich transcriptional start site is associated with H3K4me3 and gene transcription, H3.1 and H3.2 are replaced by H3.3. (B) When an unmethylated CpG rich transcriptional start site is associated with H3K27me3 and gene repression, H3.1 and H3.2 are retained. These epigenetic conformations are transmitted to the offspring, where they may influence developmental programs and provide mechanisms for epigenetic inheritance.
H2A
H4
H2B
H3.1/2
H3.1/2
H3.3
H3.3
A B C
H2A
H4
H2B
Histone exchange Retention of histones Histone–protamine exchange
Unmethylated CpG Methylated CpG Protamine
K4me3
K4me3
H3.1/2
H3.1/2
H2A
H4
H2B
H2A
H4
H2B
H3.1/2
H2A
H4
H2B
H3.1/2
K27me3
K27me3
future science group
Review Prokopuk, Western & Stringer
tion of H3K27me3 at these loci in sperm results in gene repression during preimplantation development. Polycomb repressive complex 2 (PRC2) functions to catalyze the addition of methyl groups to H3K27. Therefore, the establishment of H3K27me3 by PRC2 in the developing germline may provide a mecha-nism for inherited epigenetic information that affects development in a parent’s offspring.
OogenesisFemale germ cell epigenetic programming continues after birth with de novo methylation occurring as oocytes mature. Smallwood et al. used high-throughput reduced representation bisulfite sequencing (RRBS) to investi-gate CpG methylation in mature oocytes [39]. This tech-nique provides quantitative data specifically enriched for genomic regions with a high density of potential CpG methylation sites and enables investigation of DNA methylation at single-nucleotide resolution across the genome [40]. Therefore, this analysis is restricted to the majority of promoters and repeated sequences with CpG islands (CGIs). During oocyte maturation, CGI methylation increases from 0.5% in day 5 oocytes to approximately 11% in day 20 germinal vesicle (GV) stage oocytes and approximately 15% in ovulated metaphase II stage oocytes [39]. Although the overall CGI methylation level is lower in oocytes compared
with sperm (∼25%) methylation levels within CGIs increase as oocytes mature. Approximately 1600 CGIs are completely methylated in mature oocytes and only 185 CGIs are methylated in mature sperm. One hun-dred of these completely methylated CGIs are shared by both mature oocytes and sperm and occur at intragenic loci. Interestingly, in GV stage oocytes of Dnmt3a-/- and Dnmt3l-/- mice, genome-wide CpG methylation is pro-foundly reduced, but CGIs remain highly methylated. As no new methylation can be established in the absence of both Dnmt3a and Dnmt3l, these data suggest that CGIs are protected during PGC reprogramming. Fur-thermore, 15% methylated CGIs in wild-type oocytes retain DNA methylation levels ≥40%, through to blas-tocyst stage. This provides evidence that CGIs retain methylation during germline reprogramming, and that some of this methylation is also retained through the preimplantation reprogramming period.
Other factors that may influence maintenance and inheritance of epigenetic information are mRNAs and proteins that are deposited during oocyte growth and maturation. These factors are essential for oocyte function and early development of the embryo. Factors such as STELLA (DPPA3), ZFP57 and TRIM28 are required for the maintenance of genomic imprinting during preimplantation epigenetic reprogramming [41–43].

www.futuremedicine.com 833future science group
Transgenerational epigenetic inheritance: adaptation through the germline epigenome? Review
Preimplantation reprogrammingPreimplantation epigenetic reprogramming was first reported in the mouse as DNA methylation levels were observed to be lower in the preimplantation embryos, compared with the levels in mature sperm or oocytes and those found after implantation [44,45]. More recent studies have determined the extent and timing of this reprogramming event in a variety of species [3,4]. In the mouse the paternal genome releases protamines and is re-packaged by maternal nucleosomal histones to form the paternal pronucleus 1 h postfertilization [46]. By approximately 7–8 h postfertilization, the paternal genome appears to lose a substantial amount of DNA methylation, whereas low levels are retained in the maternal genome until E2.5 [47–50]. DNA methylation levels in the blastocyst inner cell mass (ICM) are simi-lar to those observed in the maternal pronucleus, after which DNA methylation rapidly increases as cells of the ICM undergo differentiation and begin to establish somatic lineages of the embryo.
A study by Li et al. challenged the paradigm of active global demethylation during preimplantation reprogramming using immunolocalization in preim-plantation mouse embryos [51]. In this study, trypsin-digestion resulted in detection of DNA methylation in one-cell and two-cell stage embryos, indicating that DNA demethylation may be less pronounced at these stages than previously thought. In other species such as Xenopus laevis (African clawed frogs) [52] and bovine [53], very little DNA demethylation occurs in zygotes, while there is no reduction in DNA methylation in Danio rerio (zebra fish) and rabbit zygotes [53,54]. With the caveat that analytical differences may exist, these stud-ies suggest that preimplantation reprogramming may not occur to the same extent in all species.
Recently, Smith and colleagues used RRBS to provide a more detailed analysis of CGI methylation throughout preimplantation development in mice [55]. The authors identified dynamic changes in DNA methylation during two developmentally important stages (Figure 4). Upon fertilization CGIs in the pater-nal genome are rapidly demethylated. This is followed by a more gradual demethylation, during cleavage, such that the level of DNA methylation is at its low-est in the ICM, before increasing postimplantation. These global DNA methylation dynamics were also identified in human embryos [56]. These genome-wide epigenetic data reveal epigenetic processes through which developmental programs are likely to be set for regulating differentiation of somatic lineages during postimplantation development.
The process of demethylation in both the PGCs and the zygote is thought to involve both passive, and active mechanisms involving TET enzymes (Figure 5) [57].
TET enzymes catalyze the conversion of 5-methyl-cytosine (5mC) to 5-hydroxymethylcytosine (5hmC) and depletion of TET1 results in decreased levels of 5hmC and the retention of 5mC levels (Figure 5A) [58]. In the mature oocyte and zygote, Tet3 is present at high levels, before declining after the first zygotic divi-sion. This coincides with the majority of hydroxylation of 5mC to 5hmC in the paternal genome, which results in loss of DNA methylation after DNA replication [57]. Critically, during preimplantation epigenetic repro-gramming some epigenetic modifications, including those involved in the regulation of imprinted genes, escape demethylation and maintain their methylated state into adulthood [55,59]. Maternal factors such as STELLA and the components of the TRIM28 complex are thought to be vital for maintaining this epigenetic information during reprogramming [60,61].
Levels of the maternal factor STELLA are high in the early embryo and are required for preimplanta-tion development [41]. STELLA binds to H3K9me2, which is found at maternally imprinted loci and some paternally imprinted loci, and protects imprinted genes from epigenetic reprogramming (Figure 5B) [60,62]. Similarly another maternal factor, TRIM28, is a scaffolding protein that recruits epigenetic machin-ery involved in gene silencing, such as SETDB1 and UHRF1, which methylate H3K9me3 and recruit DNMT1 and DNMT1o, respectively, maintaining methylation within imprinted differentially methyl-ated regions (DMRs) (Figure 5C) [43,63–67]. ZFP57 is a KRAB zinc finger protein, which recognizes and binds KRAB domains, and recruits TRIM28 (Figure 5C). KRAB domains are located within the majority of imprinting DMRs as well as other regions within the genome [42]. Deletion of either Trim28 or Zfp57 results in hypomethylation of some, but not all imprinted DMRs [42,68]. Therefore, other factors and complexes may be involved in maintaining epigenetic information during epigenetic reprogramming in preimplantation embryos.
Evidence of epigenetic inheritanceEpigenetic inheritance refers to the germline transmis-sion of epigenetic information to the somatic cells of the resulting offspring. To achieve this, the epigenetic information transmitted by the sperm and/or oocyte must escape preimplantation reprogramming and be maintained during fetal, and later development. There is clear evidence that epigenetic information is inherited from parents to offspring (intergenerational inheritance), with the strongest demonstration pro-vided by imprinted genes. However, whether epigen-etic information is transmitted over multiple genera-tions without reprogramming in the germline is less
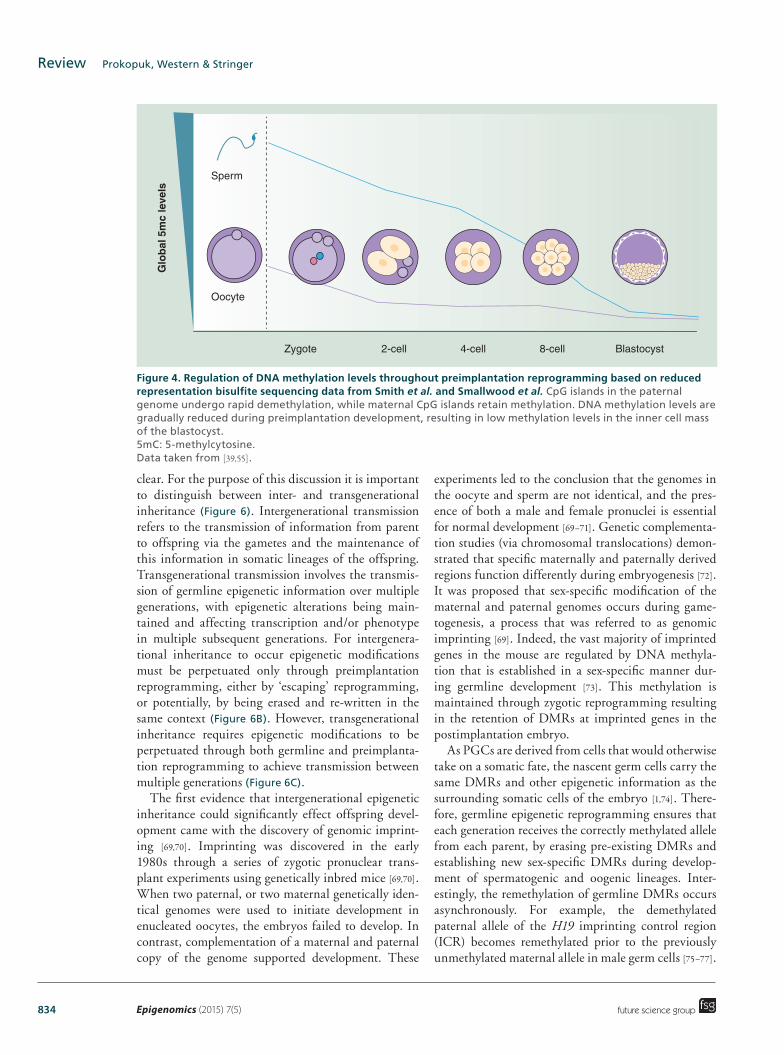
834 Epigenomics (2015) 7(5)
Figure 4. Regulation of DNA methylation levels throughout preimplantation reprogramming based on reduced representation bisulfite sequencing data from Smith et al. and Smallwood et al. CpG islands in the paternal genome undergo rapid demethylation, while maternal CpG islands retain methylation. DNA methylation levels are gradually reduced during preimplantation development, resulting in low methylation levels in the inner cell mass of the blastocyst. 5mC: 5-methylcytosine. Data taken from [39,55].
Glo
bal
5m
c le
vels
Sperm
Oocyte
Zygote 2-cell 4-cell 8-cell Blastocyst
future science group
Review Prokopuk, Western & Stringer
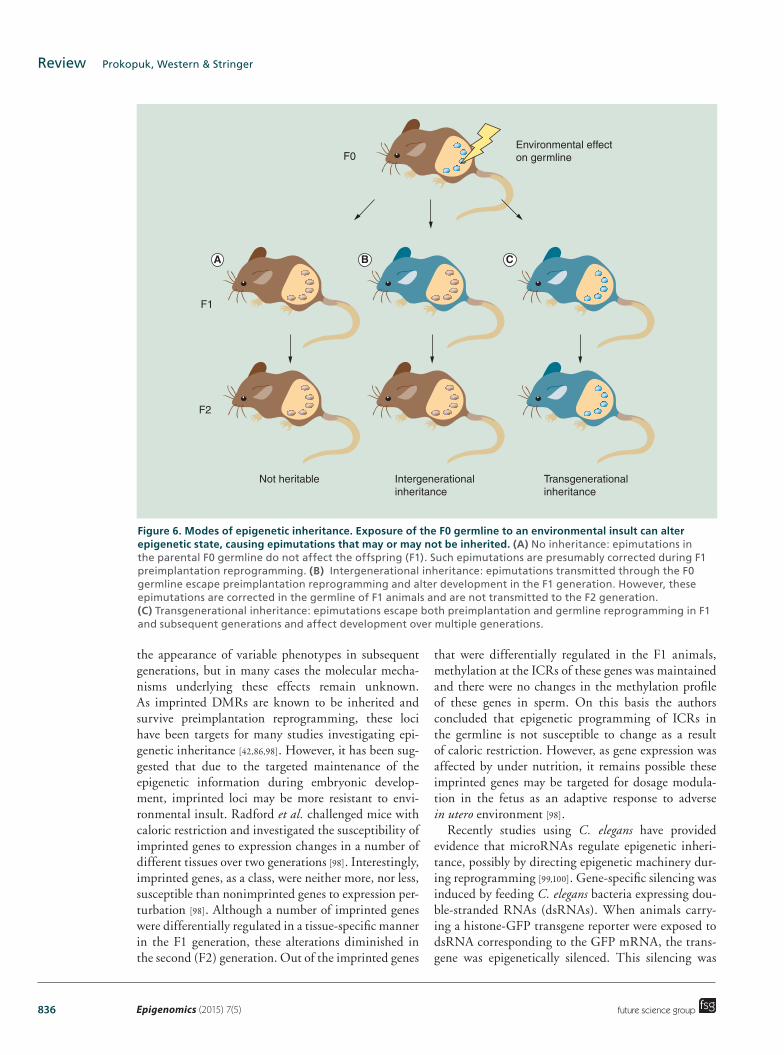
clear. For the purpose of this discussion it is important to distinguish between inter- and transgenerational inheritance (Figure 6). Intergenerational transmission refers to the transmission of information from parent to offspring via the gametes and the maintenance of this information in somatic lineages of the offspring. Transgenerational transmission involves the transmis-sion of germline epigenetic information over multiple generations, with epigenetic alterations being main-tained and affecting transcription and/or phenotype in multiple subsequent generations. For intergenera-tional inheritance to occur epigenetic modifications must be perpetuated only through preimplantation reprogramming, either by ‘escaping’ reprogramming, or potentially, by being erased and re-written in the same context (Figure 6B). However, transgenerational inheritance requires epigenetic modifications to be perpetuated through both germline and preimplanta-tion reprogramming to achieve transmission between multiple generations (Figure 6C).
The first evidence that intergenerational epigenetic inheritance could significantly effect offspring devel-opment came with the discovery of genomic imprint-ing [69,70]. Imprinting was discovered in the early 1980s through a series of zygotic pronuclear trans-plant experiments using genetically inbred mice [69,70]. When two paternal, or two maternal genetically iden-tical genomes were used to initiate development in enucleated oocytes, the embryos failed to develop. In contrast, complementation of a maternal and paternal copy of the genome supported development. These
experiments led to the conclusion that the genomes in the oocyte and sperm are not identical, and the pres-ence of both a male and female pronuclei is essential for normal development [69–71]. Genetic complementa-tion studies (via chromosomal translocations) demon-strated that specific maternally and paternally derived regions function differently during embryogenesis [72]. It was proposed that sex-specific modification of the maternal and paternal genomes occurs during game-togenesis, a process that was referred to as genomic imprinting [69]. Indeed, the vast majority of imprinted genes in the mouse are regulated by DNA methyla-tion that is established in a sex-specific manner dur-ing germline development [73]. This methylation is maintained through zygotic reprogramming resulting in the retention of DMRs at imprinted genes in the postimplantation embryo.
As PGCs are derived from cells that would otherwise take on a somatic fate, the nascent germ cells carry the same DMRs and other epigenetic information as the surrounding somatic cells of the embryo [1,74]. There-fore, germline epigenetic reprogramming ensures that each generation receives the correctly methylated allele from each parent, by erasing pre-existing DMRs and establishing new sex-specific DMRs during develop-ment of spermatogenic and oogenic lineages. Inter-estingly, the remethylation of germline DMRs occurs asynchronously. For example, the demethylated paternal allele of the H19 imprinting control region (ICR) becomes remethylated prior to the previously unmethylated maternal allele in male germ cells [75–77].
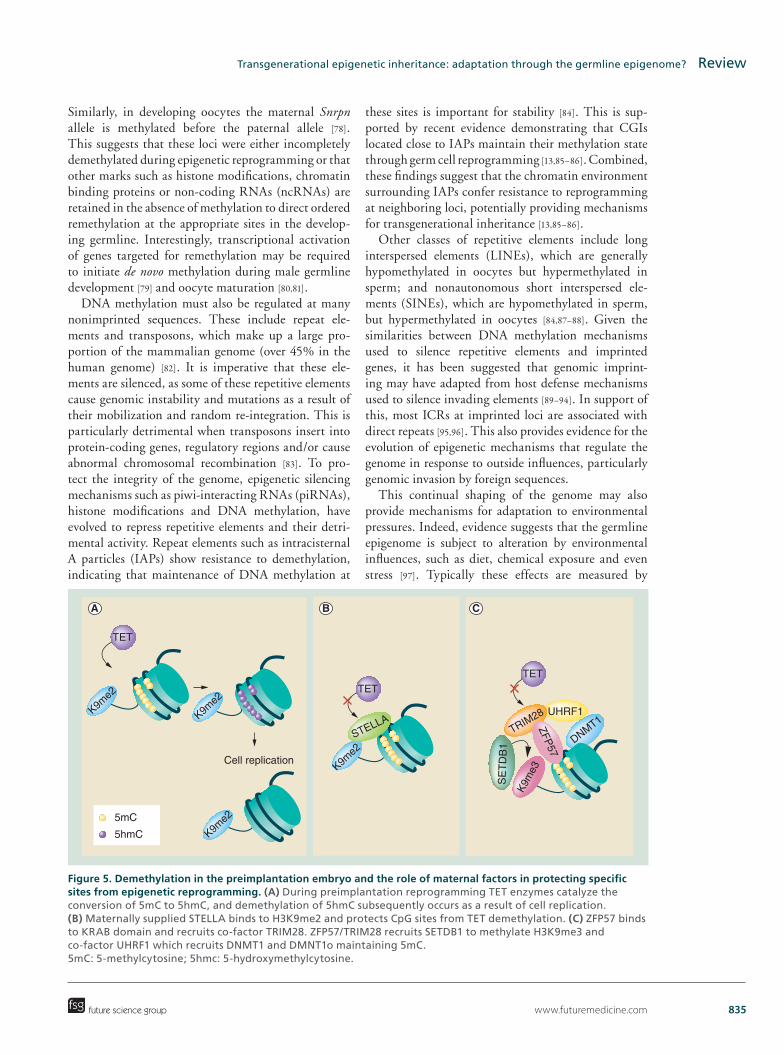
www.futuremedicine.com 835
Figure 5. Demethylation in the preimplantation embryo and the role of maternal factors in protecting specific sites from epigenetic reprogramming. (A) During preimplantation reprogramming TET enzymes catalyze the conversion of 5mC to 5hmC, and demethylation of 5hmC subsequently occurs as a result of cell replication. (B) Maternally supplied STELLA binds to H3K9me2 and protects CpG sites from TET demethylation. (C) ZFP57 binds to KRAB domain and recruits co-factor TRIM28. ZFP57/TRIM28 recruits SETDB1 to methylate H3K9me3 and co-factor UHRF1 which recruits DNMT1 and DMNT1o maintaining 5mC. 5mC: 5-methylcytosine; 5hmc: 5-hydroxymethylcytosine.
DNMT1
A B C
K9me2
5mC
Cell replication
5hmC K9me2
K9me2
TETTET
TET
SE
TD
B1
K9m
e3
K9me2
STELLAUHRF1
TRIM28
ZFP57
future science group
Transgenerational epigenetic inheritance: adaptation through the germline epigenome? Review
Similarly, in developing oocytes the maternal Snrpn allele is methylated before the paternal allele [78]. This suggests that these loci were either incompletely demethylated during epigenetic reprogramming or that other marks such as histone modifications, chromatin binding proteins or non-coding RNAs (ncRNAs) are retained in the absence of methylation to direct ordered remethylation at the appropriate sites in the develop-ing germline. Interestingly, transcriptional activation of genes targeted for remethylation may be required to initiate de novo methylation during male germline development [79] and oocyte maturation [80,81].
DNA methylation must also be regulated at many nonimprinted sequences. These include repeat ele-ments and transposons, which make up a large pro-portion of the mammalian genome (over 45% in the human genome) [82]. It is imperative that these ele-ments are silenced, as some of these repetitive elements cause genomic instability and mutations as a result of their mobilization and random re-integration. This is particularly detrimental when transposons insert into protein-coding genes, regulatory regions and/or cause abnormal chromosomal recombination [83]. To pro-tect the integrity of the genome, epigenetic silencing mechanisms such as piwi-interacting RNAs (piRNAs), histone modifications and DNA methylation, have evolved to repress repetitive elements and their detri-mental activity. Repeat elements such as intracisternal A particles (IAPs) show resistance to demethylation, indicating that maintenance of DNA methylation at
these sites is important for stability [84]. This is sup-ported by recent evidence demonstrating that CGIs located close to IAPs maintain their methylation state through germ cell reprogramming [13,85–86]. Combined, these findings suggest that the chromatin environment surrounding IAPs confer resistance to reprogramming at neighboring loci, potentially providing mechanisms for transgenerational inheritance [13,85–86].
Other classes of repetitive elements include long interspersed elements (LINEs), which are generally hypomethylated in oocytes but hypermethylated in sperm; and nonautonomous short interspersed ele-ments (SINEs), which are hypomethylated in sperm, but hypermethylated in oocytes [84,87–88]. Given the similarities between DNA methylation mechanisms used to silence repetitive elements and imprinted genes, it has been suggested that genomic imprint-ing may have adapted from host defense mechanisms used to silence invading elements [89–94]. In support of this, most ICRs at imprinted loci are associated with direct repeats [95,96]. This also provides evidence for the evolution of epigenetic mechanisms that regulate the genome in response to outside influences, particularly genomic invasion by foreign sequences.
This continual shaping of the genome may also provide mechanisms for adaptation to environmental pressures. Indeed, evidence suggests that the germline epigenome is subject to alteration by environmental influences, such as diet, chemical exposure and even stress [97]. Typically these effects are measured by
836 Epigenomics (2015) 7(5)
Figure 6. Modes of epigenetic inheritance. Exposure of the F0 germline to an environmental insult can alter epigenetic state, causing epimutations that may or may not be inherited. (A) No inheritance: epimutations in the parental F0 germline do not affect the offspring (F1). Such epimutations are presumably corrected during F1 preimplantation reprogramming. (B) Intergenerational inheritance: epimutations transmitted through the F0 germline escape preimplantation reprogramming and alter development in the F1 generation. However, these epimutations are corrected in the germline of F1 animals and are not transmitted to the F2 generation. (C) Transgenerational inheritance: epimutations escape both preimplantation and germline reprogramming in F1 and subsequent generations and affect development over multiple generations.
A B C
Environmental effecton germlineF0
F1
F2
Not heritable Intergenerational inheritance
Transgenerationalinheritance
future science group
Review Prokopuk, Western & Stringer
the appearance of variable phenotypes in subsequent generations, but in many cases the molecular mecha-nisms underlying these effects remain unknown. As imprinted DMRs are known to be inherited and survive preimplantation reprogramming, these loci have been targets for many studies investigating epi-genetic inheritance [42,86,98]. However, it has been sug-gested that due to the targeted maintenance of the epigenetic information during embryonic develop-ment, imprinted loci may be more resistant to envi-ronmental insult. Radford et al. challenged mice with caloric restriction and investigated the susceptibility of imprinted genes to expression changes in a number of different tissues over two generations [98]. Interestingly, imprinted genes, as a class, were neither more, nor less, susceptible than nonimprinted genes to expression per-turbation [98]. Although a number of imprinted genes were differentially regulated in a tissue-specific manner in the F1 generation, these alterations diminished in the second (F2) generation. Out of the imprinted genes
that were differentially regulated in the F1 animals, methylation at the ICRs of these genes was maintained and there were no changes in the methylation profile of these genes in sperm. On this basis the authors concluded that epigenetic programming of ICRs in the germline is not susceptible to change as a result of caloric restriction. However, as gene expression was affected by under nutrition, it remains possible these imprinted genes may be targeted for dosage modula-tion in the fetus as an adaptive response to adverse in utero environment [98].
Recently studies using C. elegans have provided evidence that microRNAs regulate epigenetic inheri-tance, possibly by directing epigenetic machinery dur-ing reprogramming [99,100]. Gene-specific silencing was induced by feeding C. elegans bacteria expressing dou-ble-stranded RNAs (dsRNAs). When animals carry-ing a histone-GFP transgene reporter were exposed to dsRNA corresponding to the GFP mRNA, the trans-gene was epigenetically silenced. This silencing was

www.futuremedicine.com 837future science group
Transgenerational epigenetic inheritance: adaptation through the germline epigenome? Review
maintained after removal of the dsRNA and was inher-ited in over 60% of individuals for at least four gen-erations, suggesting that this silencing was maintained through epigenetic programming in the germline [99]. The PIWI argonaute proteins, and their associated piRNAs are distinct from small-interfering RNAs (siRNAs) as they have a specific and evolutionarily conserved role in transposon silencing in the germline in many invertebrate [101] and vertebrate [102] species (reviewed in [103]). It has been suggested that piR-NAs have the ability to initiate an epigenetic memory of ‘self ’ and ‘nonself ’ RNA [100]. When ‘nonself ’ for-eign RNA is detected, targeted silencing occurs [104]. While a piRNA trigger initially induced the targeted silencing, both small RNA factors and chromatin regulators were required to complete and maintain multigenerational transgene silencing, including the putative H3K9me3 methyltransferases, HPL-2 and SET-25/32 [99]. Although the piRNA and siRNA sys-tems vary between C. elegans and vertebrate species, mechanisms regulating piRNAs and histone modifi-cations are highly conserved and may produce similar downstream silencing.
Confounding issues in the study of epigenetic inheritanceTracing the inheritance of DNA is relatively straight-forward as mutations are rare and it is easy to track the heritage of specific mutations through each gen-eration by DNA sequencing. While the majority of epigenetic information, including DNA methylation, is mitotically stable and important for cellular iden-tity, some aspects are more plastic and can be altered in response to different environmental stimuli. This enables cells to change, and potentially provides a mechanism for individuals to adapt to altered environ-ments. For transgenerational epigenetic inheritance to occur, altered epigenetic states need to be resistant to reprogramming and maintained in both germline and somatic lineages (Figure 6C). Therefore, to identify and trace specific epigenetic mutations across genera-tions, the epigenome in both the germline and affected somatic cells must be examined in each generation. In addition, to test the effect of a specific environmen-tal factor on the epigenome in a founding individual, the environment that each subsequent generation is exposed to must be free of the stimulus of interest, and other confounding factors. For example, to examine the effect of maternal diet, it is necessary to distinguish between the effect a mother’s diet has on the germline of the fetus, and the effect on the somatic cells of the offspring that may induce confounding factors in the following generation. These confounding factors may in turn, affect outcomes in the following generation.
Similarly, a single stimulus may cause a cascade of gene expression changes that are relayed through networks of genes to result in the observed phenotype. Clearly, in some cases the indirect nature of these mechanisms can make it difficult to identify the causative modifica-tion that initiated the gene expression cascade. These considerations provide profound challenges for iden-tifying phenotypic changes that can be definitively linked to transgenerational inheritance over multiple generations. For these reasons, some studies identi-fying transgenerational epigenetic inheritance may be confounded by influences of the parent in each subsequent generation [105].
Maternal transmissionParents can influence their offspring in numerous ways, including mechanisms mediated directly through uter-ine environment, or more indirectly through mecha-nisms such as behavioral cues. For example, studies have suggested that maternal care in mice is affected by stress, with stressed mothers more likely to produce stressed offspring [106,107]. However, what was origi-nally thought of as a case of transgenerational epigen-etic inheritance was subsequently attributed to altera-tions in the hypothalamic–pituitary–adrenal (HPA) axis in the offspring [107]. Francis et al. carried out a series of experiments in rats, which involved cross-fostering pups from stressed mothers to control moth-ers [106]. Naturally occurring variations in maternal care, such as licking and grooming (LG) behavior were assessed to observe the effect on offspring stress [106]. They showed that female offspring of high LG moth-ers showed significantly higher LG behavior (toward their own pups) in comparison to the offspring of low LG mothers. However, cross fostering pups from low LG mothers to high LG mothers resulted in those pups having higher LG behavior in motherhood, which is presumably a behavior learned from the high LG foster mothers or due to factors and stress hormones absorbed during lactation. Although this model provides inter-esting insights into the influence of maternal behavior on offspring behavior, it does not provide evidence of transgenerational epigenetic inheritance.
During pregnancy a mother, her embryo and the embryo’s germ cells may all be exposed to the same environmental influences (via the placenta). This expo-sure may alter the epigenome in the germ cells of the embryo, and therefore, individuals derived from the embryo’s germline (i.e., the mother’s grandchildren). In mice, the methylation-sensitive metastable agouti viable yellow (Avy) allele can influence coat color of genetically identical offspring (Figure 7). Mothers fed a diet rich in methyl donors produce a reduced percent-age of yellow-coated pups [108]. The increased avail-

838 Epigenomics (2015) 7(5)
Figure 7. Epigenetic control and inheritance at the Avy allele. Coat color variation ranging from 100% yellow, mottled (∼50% yellow), to agouti (0% yellow) in isogenic mice carrying the methylation-sensitive metastable Avy allele. Increased methylation at the long terminal repeat at the agouti locus correlates with loss of transcription at the Avy allele and a consequent increase in agouti coat color. LTR: Long terminal repeat.
Percentage methylation of LTR at agouti locus
future science group
Review Prokopuk, Western & Stringer
ability of methyl donors shifts the coat color toward pseudoagouti due to higher DNA methylation and epi-genetic silencing at the Avy allele [109]. In experiments by Cropley et al. pregnant females were fed a methyl donor rich diet, whereas their offspring were not and the coat color of F2 animals also shifted significantly toward pseudoagouti, compared with controls [110]. As the F1 generation was only exposed to a methyl donor diet in utero, this indicates that the diet altered Avy pro-gramming in the germ cells of the F1 animals (i.e., the founding germline for the F2 generation). However, the phenotypic effect was heritable for only one gen-eration [111]. Therefore, it is likely that the phenotype was lost through reprogramming in the germ cells of the F2 animals (founders of the F3 generation) [112]. Importantly, this indicates that the effect of feeding methyl donors was maintained only as an intergenera-tional effect (i.e., only in the offspring derived from the affected PGCs), not as a transgenerational effect.
The Avy allele is controlled by an IAP inserted upstream of the agouti allele. Interestingly, the spec-trum of phenotypes in offspring from maternally inher-ited Avy skews toward the maternal phenotype with a higher proportion of pseudoagouti pups produced by Avy females. However, when the Avy allele is inherited from the father, a higher percentage of pseudoagouti offspring are produced regardless of the father’s coat color [108,113]. One possible explanation for this is that IAPs are more stringently targeted for silencing by the PIWI pathway in the male germline (reviewed in [114]), leading to greater silencing of the IAP-linked Avy locus. The observed maternal inheritance of the Avy allele was initially presumed to be due to the maternal environ-ment that the pups were exposed to in utero. Yellow Avy mice are obese and hyperinsulinemic, whereas pseu-doagouti Avy mice are lean and healthy. These pheno-typic differences provide a confounding environmental
effect that may alter methylation at the Avy allele in the offspring. However, the results from embryo-transfer experiments and specific breeding schemes indicate that the presence of the IAP in the Avy allele is more likely to retain epigenetic marks when inherited from the oocyte rather than through the sperm [113]. Inter-estingly, Blewitt et al. demonstrated that the Avy allele is completely demethylated in the blastocyst before implantation suggesting that DNA methylation may not be the inherited epigenetic mark in this case [115]. Although it is possible that DNA methylation leaves a footprint for remethylation after erasure in the epiblast, the observations of Blewitt et al. indicate that epigen-etic effects observed using the Avy may be controlled by other epigenetic mechanisms.
To avoid the effects of maternal environment, embryo transfer methods have been utilized to study transgen-erational epigenetic effects. Padmanabhan et al. found that a hypomorphic mutation in the mouse Mtrr gene resulted in intrauterine growth restriction, develop-mental delay and congenital malformations (neural tube, heart and placental defects) [116]. The MTRR enzyme is necessary for the metabolism of folate, a methyl-rich B group vitamin required for successful pregnancy and development of the fetus. Wild-type F2 and F3 offspring showed abnormal developmental phe-notypes consistent with their maternal Mtrr heterozy-gous grandparents. Global DNA methylation was sig-nificantly reduced in maternal Mtrr deficient mice, and their F2 and F3 wild-type offspring. To demonstrate that these effects were independent of maternal uterine environment embryo transfers were performed. Wild-type preimplantation embryos derived from pedigrees with one heterozygous maternal grandparent were col-lected at E3.25 [116], prior to DNA remethylation dur-ing preimplantation epigenetic reprogramming. These embryos were transferred to pseudo-pregnant wild-type females and allowed to develop until E10.5. Growth defects (intrauterine growth restriction and develop-mental delay) were not observed in the transferred lit-ters. However, congenital malformations (neural tube, heart and placental defects) persisted in animals up to the F3 generation, indicating that the effects were due to transgenerational epigenetic inheritance, although the mechanisms were not identified.
Paternal transmissionRather than tracing epigenetic effects through the maternal line, it can be simpler to trace epigenetic transmission via the paternal germline. Paternal trans-mission of environmentally induced phenotypes pro-vides a robust model for studying epigenetic inheri-tance, as the male contribution to the next generation is mediated only through the sperm, without contribu-

www.futuremedicine.com 839future science group
Transgenerational epigenetic inheritance: adaptation through the germline epigenome? Review
tions from the uterine environment. Females not only have the potential to influence the next generation via the oocyte, but also prenatally via the placenta and postnatally via lactation and maternal care. A recent study of paternal epigenetic inheritance demonstrated that mice exposed to early life trauma sired progeny with reduced fear and insulin hypersensitivity [117]. Deep sequencing revealed that several microRNAs were upregulated in the sperm of the traumatized mice compared with control mice. Reduced fear and insulin hypersensitivity were observed in the F1, F2 and F3 animals compared with control animals. Changes in miRNA expression were observed in blood serum and brain (hippocampus), but not in the sperm of the F2 mice. Injection of the mutant sperm RNAs into wild-type oocytes resulted in offspring with similar behav-ioral and metabolic abnormalities as those observed in traumatized males [117]. This study provides novel evidence that an RNA-dependent mechanism affects paternal transmission of environmentally triggered traits in mammals.
The rapid global rise in diabetes, obesity and car-diovascular disease suggests that nongenetic factors significantly contribute to disease risk. This has driven debate over the past decade around the possibility that environment can influence disease inheritance through epigenetic mechanisms [118]. While the underlying mechanisms are yet to be determined, environmentally induced inter- and trans-generational effects have been reported [119–126]. Metabolic effects may be modulated through epigenetic mechanisms induced by high-fat diet. Dunn et al. exposed female mice to high-fat diet before, throughout and after pregnancy [119]. First- and second-generation offspring of both maternal and paternal lineages showed reduced insulin sensitivity and increased body length. In the third generation only increased body length was detected and this phe-notype was paternally, but not maternally, transmitted to female offspring. While these phenotypes were ini-tially transmitted through both lineages, supporting a role for epigenetic inheritance, the mechanisms of this inheritance remain unclear. Furthermore, DNA meth-ylation in the brain (arcuate nucleus) of second-gener-ation offspring was reduced at the promoter region of Ghsr, which correlated with an increase in transcription and a larger body size. However, it was not determined whether this difference in DNA methylation origi-nated in the germline of the first-generation animals. Therefore, although these data provide evidence that glucose homeostasis and body length are modulated by maternal diet/caloric availability in utero, the role for germline epigenetic inheritance is less clear [127]. It remains possible that the in utero environment affects programming of the fetal germ cells, resulting in a phe-
notype in the second-generation, but further studies are required to determine whether this is the case.
Male mice exposed to folate-deficient diets (FD) in utero and throughout life, sire offspring with mul-tiple musculoskeletal defects and placental abnor-malities [128]. Methylated DNA immunoprecipitation promoter array (me-DIP-chip) of FD male sperm com-pared with control sperm identified DMRs associated with functions in the central nervous system and mus-cle tissue. In the placentas of FD sired offspring (F2), only 2 out of approximately 400 differentially expressed loci were correlated to DMRs in the FD sperm. How-ever, no differences in DNA methylation were observed at these two loci in the F2 placenta. Therefore, the dif-ferences in DNA methylation observed in the sperm are unlikely to have directly caused the observed phe-notypes. In a similar study, Radford et al. exposed pregnant females to 50% caloric restriction during late gestation, coinciding with the time of epigenetic pro-gramming in developing male germ cells [126]. Caloric restriction decreased DNA methylation at 111 loci, and increased methylation at a further 55 loci in the sperm of F1 male mice. When F1 males (control and restricted calorie groups) were mated to control females and fed a normal calorie diet, differential methylation was lost in the F2 tissues examined (brain and liver of E16.5 embryos). However, differences in gene expression asso-ciated with glucose tolerance and metabolism persisted in the absence of differential DNA methylation. In both of these studies [126,128] it is speculated that gene expres-sion differences might be perpetuated by mechanisms other than DNA methylation, such as histone modi-fications. Interestingly, global H3K4 and K9 mono-methylation and H3K9 trimethylation were reduced in the FD sperm compared with controls [128], indicating that histone methylation levels may be sensitive to folate availability. However, it should also be noted that folate deficiency has been linked to increased DNA damage and therefore genetic mutations could contribute in these models [129]. Indeed, a marker of DNA damage (γ-H2AX) was detected at higher levels in FD sper-matocytes compared with controls. This damage was not observed in mature sperm [128]. Therefore, although it is possible that the observed phenotypes were due to alterations in the sperm epigenome, it is also plausible that aberrant DNA repair mechanisms may also con-tribute. Therefore, it would be of particular interest to identify specific regions within the genome that are par-ticularly sensitive to folate-driven DNA damage and to determine if these genomic regions are associated with the observed phenotypes. These studies [126,128] high-light the potential for confounding effects that can influence the interpretation of inherited phenotypes, even when using a paternal inheritance model.

840 Epigenomics (2015) 7(5) future science group
Review Prokopuk, Western & Stringer
Epigenetic inheritance may also occur in an indirect, trans-acting manner. Disrupting epigenetic modifiers can result in multiple errors during epigenetic program-ming in the germline that may subsequently alter pre-implantation reprogramming in the next generation. Using yellow (Avy/a) mothers, Chong et al. examined the coat colors of wild-type offspring sired by agouti (a/a) males that were either heterozygous or wild-type for a point mutation in either Dnmt1 or Smarca5; epigenetic modifiers that alter DNA methylation and histone orga-nization, respectively [130]. Mutations in either of these genes caused a shift in the penetrance at the Avy allele, such that wild-type animals from heterozygous fathers were more likely to be yellow than offspring sired from a wild-type father. Significantly this work indicates that mutating epigenetic modifiers in the paternal germ-line can affect gene expression in the next generation through nongenetic information carried by the sperm to the offspring. Moreover, the Avy allele was inherited from the mother, indicating that the effects resulting from the germline epimutations in the male caused a trans-acting effect on gene expression in the offspring. As this type of trans-acting effect is presumably indi-rect, Chong et al. speculate that RNA or chromatin modifying proteins in the sperm may be responsible for altering expression of maternal alleles in the off-spring [130]. Indeed, recent studies using C. elegans have associated small RNAs and histone modifications to multigenerational epigenetic inheritance [99–100,131–132]. These studies provide important insights into the rela-tionship between germline epigenetic patterning and mechanisms facilitating the transmission of epigenetic information from one generation to the next.
Adaptive evolution & natural selectionIt is possible that with each generation the epigenetic state can change to benefit survival of the offspring, however due to confounding influences in most stud-ies, the significance of epigenetic mechanisms in inheri-tance remain to be clarified [133,134]. Although epigen-etic modifications have mostly been studied in the context of development and disease, they may also have a role in evolutionary processes. Adaptive evolution, first described by Charles Darwin and Alfred Wallace, involves the gradual process whereby environmental pressure results in the selection of phenotypes within a population that benefit reproductive fitness (survival of the fittest). Over 70 years ago this Darwinian view of evolution was combined with the theory of Mendelian genetics to produce the modern synthesis model for evo-lution [135]. Under this model, transmission of germline genes is equated to variations in DNA sequence and importantly is not affected by the developmental his-tory of the individual. Selection occurs at the individ-
ual level with minimal contribution of group selection. This results in gradual evolution through the selection of individuals with more advantageous phenotypes. It is now clear that soft inheritance [136], which refers to nongenetic inheritance, such as epigenetic modifi-cations, niche construction and possibly learning and cultural influences, provides a potentially important influence on evolution. For example, mother–offspring interactions in utero or postnatally affect the develop-ment of an individual. These interactions could alter both survival and reproductive fitness. If these interac-tions alter heritable epigenetic marks or are genetically linked, these factors could influence evolution. Herita-ble epialleles are common in plants, fungi, invertebrates and vertebrates and have been proposed to be particu-larly important for producing variation and promoting adaptation in the absence of genetic variation [133]. Epi-genetic information in the germline, such as genomic imprinting, may also initiate reproductive isolation with variations resulting in incompatibilities within hybrid individuals [137]. Therefore, the modern synthe-sis view of heredity and evolution has been expanded to incorporate these soft inheritance mechanisms [138–142].
Epigenetic processes may have evolved to calibrate organisms to stochastic environments. However, it has also been speculated that extreme adverse environmen-tal influences can promote programming defects in the germline that impair the individual to adapt to environ-mental stresses [143]. This may be particularly relevant for human societies that have altered diets contain-ing high-sugar and fat, and a greater exposure to toxic chemicals, which may alter epigenetic processes in the germline. A study by Li et al. used a mouse model with natural-onset maternal obesity and diabetes to demon-strate that induced epigenetic changes have an adaptive function [144]. Mice carrying the active Avy allele are pre-disposed to becoming obese and developing type-2 dia-betes [145]. When a/a offspring from obese yellow Avy/a mothers were compared with isogenic a/a offspring from lean pseudoagouti Avy/a mothers after being fed con-trol and high-fat diets, offspring of yellow obese mice showed a latent defect in glucose and lipid metabolism when fed a high-fat diet. Remarkably, this phenotype can be evaded by avoiding high-fat diets. This subop-timal metabolic response is likely the result of damage induced by an abnormal intrauterine environment. Moreover, the molecular changes that occur due to the in utero exposure do not directly indicate disrupted metabolic pathways, but rather developmental ontolo-gies [144]. Therefore, it is possible that programming that occurred in utero facilitated survival of the devel-oping embryo, at the expense of increased disease risk later in life. Although, adult function may be negatively affected, if a mutation promotes early survival and repro-

www.futuremedicine.com 841future science group
Transgenerational epigenetic inheritance: adaptation through the germline epigenome? Review
ductive success, then this adaptation, albeit outwardly suboptimal, will disseminate through the population and influence the evolution of the species [146].
Genomic imprinting provides another example of what might appear as ‘suboptimal’ epigenetic inheri-tance. For imprinting to have evolved, the selective advantage of monoallelic expression must have out-weighed the cost of increased mortality due to the increased rate of exposing recessive deleterious alleles. Genomic imprinting affects both the growth and func-tion of organs involved in nutrient supply (e.g., pla-centa), and thereby modulates fetal nutrient supply and growth [147]. Epigenetic regulation of these factors is vital for both embryonic and maternal survival during pregnancy and lactation [19–21,23,125]. This may provide a balance between nutritional supply that ensures off-spring success, and sufficient nutrient restriction that prevents a mother from being disadvantaged to the point of compromised survival or future reproductive success [148–151]. Genomic imprinting involves com-plex and strictly regulated mechanisms of epigenetic programming that is essential for development. It is unlikely that such a mechanism of epigenetic inheri-tance has evolved without evolutionary relevance, and is likely an active mechanism that promotes survival
of resulting offspring and therefore the species. Fur-ther developing our understanding of the mechanisms involved in epigenetic programming and mainte-nance of epigenetic modifications in the germline will enhance our understanding of inherited epigenetic information and its possible role in evolution.
Future perspectiveWhile evidence for transgenerational epigenetic inheri-tance is strengthening, it remains challenging to isolate its specific effects on offspring from confounding factors. New technologies including genome-wide epigenetic studies are beginning to improve our understanding of epigenetic programming in the germline and during pre-implantation development. Expansion of this work will provide a more complete scaffold for understanding epi-genetic programming and the transmission of epigenetic information between parent and offspring. These new technologies are now being complemented with experi-mental systems that directly test the function of epi-genetic modifiers in the germline by using models such as C. elegans and mice. These models will identify how changes in epigenetic modifications in the germline affect development of the offspring and the epigenetic modifiers that mediate these effects. This is essential to understand
Executive summary
Epigenetic reprogramming in the germline• Germline epigenetic reprogramming involves the genome-wide erasure and re-establishment of DNA
methylation in germ cells.• This mechanism is vital for the establishment of sex-specific epigenetic information and correct development
in the next generation.• Histone marks are dynamic during germ cell epigenetic reprogramming, with some histone modifications
remaining in a ‘poised’ state.Preimplantation reprogramming• Preimplantation reprogramming is essential for embryonic development, allowing cells to proliferate and
differentiate into all cell types in the developing embryo.• Maternal factors such as STELLA and TRIM28 protect imprinted genes from epigenetic reprogramming in
preimplantation embryos.Evidence of epigenetic inheritance• Epigenetic modifications need to ‘escape’ both germline and preimplantation epigenetic reprogramming to
be inherited in a transgenerational manner.• Mechanisms maintaining retrotransposon silencing may have been adapted to enable epigenetic inheritance.Confounding issues of the study of epigenetic inheritance• Transgenerational epigenetic inheritance is difficult to study, due to limitations in isolating cause and effect,
such as separating germline epigenetic changes from confounding in utero effects in the developing fetus.Adaptive evolution & natural selection• Epigenetic inheritance may provide evolutionary mechanisms that facilitate adaptation of an organism to
adverse environments.Future perspective• Further study of how imprinted genes and IAPs escape reprogramming will advance our understanding of
how loci are resistant to epigenetic reprogramming.• Genome-wide analyses of histone modifications throughout early development (germline and
preimplantation reprogramming) will provide a greater understanding about epigenetic reprogramming mechanisms.

842 Epigenomics (2015) 7(5) future science group
Review Prokopuk, Western & Stringer
the molecular mechanisms underlying intergenerational and transgenerational epigenetic inheritance and will have wide-ranging implications for human health.
AcknowledgementsThe authors would like to thank K Hogg for manuscript review
and comments.
Financial & competing interests disclosureThis work was supported by funding from the Monash Univer-
sity Faculty of Medicine, Nursing and Health Sciences funding
granted to PS Western and the Victorian Government’s Op-
erational Infrastructure Support Program. L Prokopuk is sup-
ported by an Australian Postgraduate Award. PS Western and
JM Stringer are supported by National Health and Medical Re-
search Grants 1043939 and 1051223 awarded to PS Western.
The authors have no other relevant affiliations or financial in-
volvement with any organization or entity with a financial inter-
est in or financial conflict with the subject matter or materials
discussed in the manuscript apart from those disclosed.
No writing assistance was utilized in the production of this
manuscript.
ReferencesPapers of special note have been highlighted as: • of interest; •• of considerable interest
1 Hajkova P, Erhardt S, Lane N et al. Epigenetic reprogramming in mouse primordial germ cells. Mech. Dev. 117(1–2), 15–23 (2002).
• Demonstratesepigeneticreprogramminginprimordialgermcells(PGCs).
2 Saitou M, Kagiwada S, Kurimoto K. Epigenetic reprogramming in mouse pre-implantation development and primordial germ cells. Development 139(1), 15–31 (2012).
3 Morgan HD, Santos F, Green K, Dean W, Reik W. Epigenetic reprogramming in mammals. Hum. Mol. Genet. 14(Suppl. 1), R47–R58 (2005).
4 Feng S, Jacobsen SE, Reik W. Epigenetic reprogramming in plant and animal development. Science 330(6004), 622–627 (2010).
5 Kelly WG. Transgenerational epigenetics in the germline cycle of Caenorhabditis elegans. Epigenetics Chromatin 7(1), 6 (2014).
6 Mintz B, Russell ES. Gene-induced embryological modifications of primordial germ cells in the mouse. J. Exp. Zool. 134(2), 207–237 (1957).
7 Tam PPL, Snow MHL. Proliferation and migration of primordial germ-cells during compensatory growth in mouse embryos. J. Embryol. Exp. Morph. 64, 133–147 (1981).
8 Tachibana M, Ueda J, Fukuda M et al. Histone methyltransferases G9a and GLP form heteromeric complexes and are both crucial for methylation of euchromatin at H3-K9. Genes Dev. 19(7), 815–826 (2005).
9 Yabuta Y, Kurimoto K, Ohinata Y, Seki Y, Saitou M. Gene expression dynamics during germline specification in mice identified by quantitative single-cell gene expression profiling. Biol. Repro. 75(5), 705–716 (2006).
10 Seki Y, Yamaji M, Yabuta Y et al. Cellular dynamics associated with the genome-wide epigenetic reprogramming in migrating primordial germ cells in mice. Development 134(14), 2627–2638 (2007).
11 Seki Y, Hayashi K, Itoh K, Mizugaki M, Saitou M, Matsui Y. Extensive and orderly reprogramming of genome-wide chromatin modifications associated with specification and early development of germ cells in mice. Dev. Biol. 278(2), 440–458 (2005).
12 Hajkova P, Ancelin K, Waldmann T et al. Chromatin dynamics during epigenetic reprogramming in the mouse germ line. Nature 452(7189), 877–881 (2008).
13 Seisenberger S, Andrews S, Krueger F et al. The dynamics of genome-wide DNA methylation reprogramming in mouse primordial germ cells. Mol. Cell 48(6), 849–862(2012).
•• UsedwholegenomebisulfitesequencingandRNA-seqtoidentifydynamicsofDNAmethylationreprogramminginmousePGCs.
14 Ancelin K, Lange UC, Hajkova P et al. Blimp1 associates with Prmt5 and directs histone arginine methylation in mouse germ cells. Nat. Cell. Biol. 8(6), 623–630 (2006).
15 Kagiwada S, Kurimoto K, Hirota T, Yamaji M, Saitou M. Replication-coupled passive DNA demethylation for the erasure of genome imprints in mice. EMBO J. 32(3), 340–353 (2013).
16 Aoto T, Saitoh N, Sakamoto Y, Watanabe S, Nakao M. Polycomb group protein-associated chromatin is reproduced in post-mitotic G1 phase and is required for S phase progression. J. Biol. Chem. 283(27), 18905–18915 (2008).
17 Hackett JA, Zylicz JJ, Surani MA. Parallel mechanisms of epigenetic reprogramming in the germline. Trends Genet. 28(4), 164–174 (2012).
18 Stringer J, Barrand S, Western P. Fine-tuning evolution: germ-line epigenetics and inheritance. Reproduction 146(1), R37–R48 (2013).
19 Stringer JM, Pask AJ, Shaw G, Renfree MB. Post-natal imprinting: evidence from marsupials. Heredity 113(2), 145–155 (2014).
20 Constancia M, Kelsey G, Reik W. Resourceful imprinting. Nature 432(7013), 53–57 (2004).
21 Li Y, Sasaki H. Genomic imprinting in mammals: its life cycle, molecular mechanisms and reprogramming. Cell Res. 21(3), 466–473 (2011).
22 Bartolomei MS, Ferguson-Smith AC. Mammalian genomic imprinting. Cold Spring Harb. Perspect. Biol. 3(7), pii:a002592 (2011).
23 Charalambous M, Da Rocha ST, Ferguson-Smith AC. Genomic imprinting, growth control and the allocation of nutritional resources: consequences for postnatal life. Curr. Opin. Endocrinol. Diabetes Obes. 14(1), 3–12 (2007).
24 Kaneda M, Okano M, Hata K et al. Essential role for de novo DNA methyltransferase Dnmt3a in paternal and maternal imprinting. Nature 429(6994), 900–903 (2004).

www.futuremedicine.com 843future science group
Transgenerational epigenetic inheritance: adaptation through the germline epigenome? Review
25 Kato Y, Kaneda M, Hata K et al. Role of the Dnmt3 family in de novo methylation of imprinted and repetitive sequences during male germ cell development in the mouse. Hum. Mol. Genet. 16(19), 2272–2280 (2007).
26 La Salle S, Mertineit C, Taketo T, Moens PB, Bestor TH, Trasler JM. Windows for sex-specific methylation marked by DNA methyltransferase expression profiles in mouse germ cells. Dev. Biol. 268(2), 403–415 (2004).
27 Sakai Y, Suetake I, Shinozaki F, Yamashina S, Tajima S. Co-expression of de novo DNA methyltransferases Dnmt3a2 and Dnmt3L in gonocytes of mouse embryos. Gene Expr. Patterns 5(2), 231–237 (2004).
28 Shovlin TC, Bourc’his D, La Salle S et al. Sex-specific promoters regulate Dnmt3L expression in mouse germ cells. Hum. Reprod. 22(2), 457–467 (2007).
29 Webster KE, O’bryan MK, Fletcher S et al. Meiotic and epigenetic defects in Dnmt3L-knockout mouse spermatogenesis. Proc. Natl Acad. Sci. USA 102(11), 4068–4073 (2005).
30 Western PS, Van Den Bergen JA, Miles DC, Sinclair AH. Male germ cell differentiation involves complex repression of the regulatory network controlling pluripotency. FASEB J. 24(8), 3026–3035 (2010).
31 Bourc’his D, Xu GL, Lin CS, Bollman B, Bestor TH. Dnmt3L and the establishment of maternal genomic imprints. Science 294(5551), 2536–2539 (2001).
32 Hata K, Okano M, Lei H, Li E. Dnmt3L cooperates with the Dnmt3 family of de novo DNA methyltransferases to establish maternal imprints in mice. Development 129(8), 1983–1993 (2002).
33 Lucifero D, La Salle S, Bourc’his D, Martel J, Bestor TH, Trasler JM. Coordinate regulation of DNA methyltransferase expression during oogenesis. BMC Dev. Biol. 7, 36 (2007).
34 Lesch BJ, Dokshin GA, Young RA, Mccarrey JR, Page DC. A set of genes critical to development is epigenetically poised in mouse germ cells from fetal stages through completion of meiosis. Proc. Natl Acad. Sci. USA 110(40), 16061–16066 (2013).
•• PerformedChIP-seqandRNA-seqanalysisonmaleandfemalegermcellsduringembryogenesisandidentifiedasetofdevelopmentallycriticalepigeneticallypoisedgenes.
35 Kota SK, Feil R. Epigenetic transitions in germ cell development and meiosis. Dev. Cell 19(5), 675–686 (2010).
36 Hammoud SS, Nix DA, Zhang H, Purwar J, Carrell DT, Cairns BR. Distinctive chromatin in human sperm packages genes for embryo development. Nature 460(7254), 473–478 (2009).
37 Brykczynska U, Hisano M, Erkek S et al. Repressive and active histone methylation mark distinct promoters in human and mouse spermatozoa. Nat. Struct. Mol. Biol. 17(6), 679–687 (2010).
38 Erkek S, Hisano M, Liang CY et al. Molecular determinants of nucleosome retention at CpG-rich sequences in mouse spermatozoa. Nat. Struct. Mol. Biol. 20(7), 868–875 (2013).
•• Analyzedthelocationofnucleosomeretentioninmousespermatozoa,supportingthehypothesisfornucleosomalepigeneticinheritance.
39 Smallwood SA, Tomizawa S, Krueger F et al. Dynamic CpG island methylation landscape in oocytes and preimplantation embryos. Nat. Genet. 43(8), 811–814 (2011).
•• High-throughputbisulfiteanalysisofmouseoocytes,spermandpreimplantationembryos.
40 Chatterjee A, Rodger EJ, Stockwell PA, Weeks RJ, Morison IM. Technical considerations for reduced representation bisulfite sequencing with multiplexed libraries. J. Biomed. Biotechnol. 2012, 741542 (2012).
41 Payer B, Saitou M, Barton SC et al. Stella is a maternal effect gene required for normal early development in mice. Curr. Biol. 13(23), 2110–2117 (2003).
42 Li X, Ito M, Zhou F et al. A maternal-zygotic effect gene, Zfp57, maintains both maternal and paternal imprints. Dev. Cell 15(4), 547–557 (2008).
43 Messerschmidt DM, De Vries W, Ito M, Solter D, Ferguson-Smith A, Knowles BB. Trim28 is required for epigenetic stability during mouse oocyte to embryo transition. Science 335(6075), 1499–1502 (2012).
44 Monk M, Boubelik M, Lehnert S. Temporal and regional changes in DNA methylation in the embryonic, extraembryonic and germ cell lineages during mouse embryo development. Development 99(3), 371–382 (1987).
45 Howlett SK, Reik W. Methylation levels of maternal and paternal genomes during preimplantation development. Development 113(1), 119–127 (1991).
46 Van Der Heijden GW, Dieker JW, Derijck AA et al. Asymmetry in histone H3 variants and lysine methylation between paternal and maternal chromatin of the early mouse zygote. Mech. Dev. 122(9), 1008–1022 (2005).
47 Mayer W, Niveleau A, Walter J, Fundele R, Haaf T. Demethylation of the zygotic paternal genome. Nature 403(6769), 501–502 (2000).
48 Oswald J, Engemann S, Lane N et al. Active demethylation of the paternal genome in the mouse zygote. Curr. Biol. 10(8), 475–478 (2000).
49 Santos F, Hendrich B, Reik W, Dean W. Dynamic reprogramming of DNA methylation in the early mouse embryo. Dev. Biol. 241(1), 172–182 (2002).
50 Wossidlo M, Nakamura T, Lepikhov K et al. 5-hydroxymethylcytosine in the mammalian zygote is linked with epigenetic reprogramming. Nat. Commun. 2, 241 (2011).
51 Li Y, O’Neill C. Persistence of cytosine methylation of DNA following fertilisation in the mouse. PLoS ONE 7(1), e30687 (2012).
52 Stancheva I, Meehan RR. Transient depletion of xDnmt1 leads to premature gene activation in Xenopus embryos. Genes Dev. 14(3), 313–327 (2000).
53 Beaujean N, Hartshorne G, Cavilla J et al. Non-conservation of mammalian preimplantation methylation dynamics. Curr. Biol. 14(7), R266–R267 (2004).
54 Macleod D, Clark VH, Bird A. Absence of genome-wide changes in DNA methylation during development of the zebrafish. Nat. Genet. 23(2), 139–140 (1999).

844 Epigenomics (2015) 7(5) future science group
Review Prokopuk, Western & Stringer
55 Smith ZD, Chan MM, Mikkelsen TS et al. A unique regulatory phase of DNA methylation in the early mammalian embryo. Nature 484(7394), 339–344 (2012).
•• High-throughputDNAmethylationanalysisofmousegametesandfromzygotetopostimplantation.
56 Smith ZD, Chan MM, Humm KC et al. DNA methylation dynamics of the human preimplantation embryo. Nature 511(7511), 611–615 (2014).
57 Inoue A, Zhang Y. Replication-dependent loss of 5-hydroxymethylcytosine in mouse preimplantation embryos. Science 334(6053), 194 (2011).
58 Tahiliani M, Koh KP, Shen Y et al. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science 324(5929), 930–935 (2009).
59 Borgel J, Guibert S, Li Y et al. Targets and dynamics of promoter DNA methylation during early mouse development. Nat. Genet. 42(12), 1093–1100 (2010).
60 Nakamura T, Liu YJ, Nakashima H et al. PGC7 binds histone H3K9me2 to protect against conversion of 5mC to 5hmC in early embryos. Nature 486(7403), 415–419 (2012).
• DemonstratedthatSTELLAbindsH3K9me2toprotectCpGsitesagainstTETdemethylation.
61 Messerschmidt DM. Should I stay or should I go: protection and maintenance of DNA methylation at imprinted genes. Epigenetics 7(9), 969–975 (2012).
62 Nakamura T, Arai Y, Umehara H et al. PGC7/Stella protects against DNA demethylation in early embryogenesis. Nat. Cell Biol. 9(1), 64–71 (2007).
63 Ratnam S, Mertineit C, Ding F et al. Dynamics of Dnmt1 methyltransferase expression and intracellular localization during oogenesis and preimplantation development. Dev. Biol. 245(2), 304–314 (2002).
64 Quenneville S, Verde G, Corsinotti A et al. In embryonic stem cells, ZFP57/KAP1 recognize a methylated hexanucleotide to affect chromatin and DNA methylation of imprinting control regions. Mol. Cell 44(3), 361–372 (2011).
65 Zuo X, Sheng J, Lau HT et al. Zinc finger protein ZFP57 requires its co-factor to recruit DNA methyltransferases and maintains DNA methylation imprint in embryonic stem cells via its transcriptional repression domain. J. Biol. Chem. 287(3), 2107–2118 (2012).
66 Kurihara Y, Kawamura Y, Uchijima Y et al. Maintenance of genomic methylation patterns during preimplantation development requires the somatic form of DNA methyltransferase 1. Dev. Biol. 313(1), 335–346 (2008).
67 Cirio MC, Ratnam S, Ding F, Reinhart B, Navara C, Chaillet JR. Preimplantation expression of the somatic form of Dnmt1 suggests a role in the inheritance of genomic imprints. BMC Dev. Biol. 8, 9 (2008).
68 Mackay DJ, Callaway JL, Marks SM et al. Hypomethylation of multiple imprinted loci in individuals with transient neonatal diabetes is associated with mutations in ZFP57. Nat. Genet. 40(8), 949–951 (2008).
69 Surani MA, Barton SC, Norris ML. Development of reconstituted mouse eggs suggests imprinting of the genome during gametogenesis. Nature 308(5959), 548–550 (1984).
70 McGrath J, Solter D. Completion of mouse embryogenesis requires both the maternal and paternal genomes. Cell 37(1), 179–183 (1984).
71 Mann JR, Lovell-Badge RH. Inviability of parthenogenones is determined by pronuclei, not egg cytoplasm. Nature 310(5972), 66–67 (1984).
72 Searle AG, Beechey CV. Noncomplementation phenomena and their bearing on nondisjunctional effects. Basic Life Sci. 36, 363–376 (1985).
73 Li E, Beard C, Jaenisch R. Role for DNA methylation in genomic imprinting. Nature 366(6453), 362–365 (1993).
74 Saitou M, Payer B, Lange UC, Erhardt S, Barton SC, Surani MA. Specification of germ cell fate in mice. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 358(1436), 1363–1370 (2003).
75 Davis TL, Trasler JM, Moss SB, Yang GJ, Bartolomei MS. Acquisition of the H19 methylation imprint occurs differentially on the parental alleles during spermatogenesis. Genomics 58(1), 18–28 (1999).
76 Davis TL, Yang GJ, McCarrey JR, Bartolomei MS. The H19 methylation imprint is erased and re-established differentially on the parental alleles during male germ cell development. Hum. Mol. Genet. 9(19), 2885–2894 (2000).
77 Li JY, Lees-Murdock DJ, Xu GL, Walsh CP. Timing of establishment of paternal methylation imprints in the mouse. Genomics 84(6), 952–960 (2004).
78 Lucifero D, Mann MR, Bartolomei MS, Trasler JM. Gene-specific timing and epigenetic memory in oocyte imprinting. Hum. Mol. Genet. 13(8), 839–849 (2004).
79 Henckel A, Chebli K, Kota SK, Arnaud P, Feil R. Transcription and histone methylation changes correlate with imprint acquisition in male germ cells. EMBO J. 31(3), 606–615 (2012).
80 Chotalia M, Smallwood SA, Ruf N et al. Transcription is required for establishment of germline methylation marks at imprinted genes. Genes Dev. 23(1), 105–117 (2009).
81 Smith EY, Futtner CR, Chamberlain SJ, Johnstone KA, Resnick JL. Transcription is required to establish maternal imprinting at the Prader-Willi syndrome and Angelman syndrome locus. PLoS Genet. 7(12), e1002422 (2011).
82 Lander ES, Linton LM, Birren B et al. Initial sequencing and analysis of the human genome. Nature 409(6822), 860–921 (2001).
83 Slotkin RK, Martienssen R. Transposable elements and the epigenetic regulation of the genome. Nat. Rev. Genet. 8(4), 272–285 (2007).
84 Lane N, Dean W, Erhardt S et al. Resistance of IAPs to methylation reprogramming may provide a mechanism for epigenetic inheritance in the mouse. Genesis 35(2), 88–93 (2003).
85 Hackett JA, Sengupta R, Zylicz JJ et al. Germline DNA demethylation dynamics and imprint erasure through 5-hydroxymethylcytosine. Science 339(6118), 448–452 (2013).
86 Guibert S, Forne T, Weber M. Global profiling of DNA methylation erasure in mouse primordial germ cells. Genome Res. 22(4), 633–641 (2012).

www.futuremedicine.com 845future science group
Transgenerational epigenetic inheritance: adaptation through the germline epigenome? Review
87 Peaston AE, Evsikov AV, Graber JH et al. Retrotransposons regulate host genes in mouse oocytes and preimplantation embryos. Dev. Cell 7(4), 597–606 (2004).
88 Rubin CM, Vandevoort CA, Teplitz RL, Schmid CW. Alu repeated DNAs are differentially methylated in primate germ cells. Nucleic Acids Res. 22(23), 5121–5127 (1994).
89 Sapienza C. Sex-linked dosage-sensitive modifiers as imprinting genes. Development 108(Suppl.), 107–113 (1990).
90 Barlow DP. Methylation and imprinting: from host defense to gene regulation? Science 260(5106), 309–310 (1993).
91 McDonald JF. Transposable elements: possible catalysts of organismic evolution. Trends Ecol. Evol. 10(3), 123–126 (1995).
92 Surani MA. Imprinting and the initiation of gene silencing in the germ line. Cell 93(3), 309–312 (1998).
93 Matzke MA, Mette MF, Matzke AJM. Transgene silencing by the host genome defense: implications for the evolution of epigenetic control mechanisms in plants and vertebrates. Plant Mol. Biol. 43(2), 401–415 (2000).
94 Mcdonald JF, Matzke MA, Matzke AJ. Host defenses to transposable elements and the evolution of genomic imprinting. Cytogenet. Genome Res. 110(1–4), 242–249 (2005).
95 Constancia M, Pickard B, Kelsey G, Reik W. Imprinting mechanisms. Genome Res. 8(9), 881–900 (1998).
96 Reik W, Walter J. Genomic imprinting: parental influence on the genome. Nat. Rev. Genet. 2(1), 21–32 (2001).
97 Daxinger L, Whitelaw E. Understanding transgenerational epigenetic inheritance via the gametes in mammals. Nat. Rev. Genet. 13(3), 153–162 (2012).
98 Radford EJ, Isganaitis E, Jimenez-Chillaron J et al. An unbiased assessment of the role of imprinted genes in an intergenerational model of developmental programming. PLoS Genet. 8(4), 41–53 (2012).
99 Ashe A, Sapetschnig A, Weick EM et al. piRNAs can trigger a multigenerational epigenetic memory in the germline of C. elegans. Cell 150(1), 88–99 (2012).
100 Shirayama M, Seth M, Lee HC et al. piRNAs initiate an epigenetic memory of nonself RNA in the C. elegans germline. Cell 150(1), 65–77 (2012).
101 Bagijn MP, Goldstein LD, Sapetschnig A et al. Function, targets, and evolution of Caenorhabditis elegans piRNAs. Science 337(6094), 574–578 (2012).
102 Lim SL, Tsend-Ayush E, Kortschak RD et al. Conservation and expression of PIWI-interacting RNA pathway genes in male and female adult gonad of amniotes. Biol. Reprod. 89(6), 136 (2013).
103 Malone CD, Hannon GJ. Small RNAs as guardians of the genome. Cell 136(4), 656–668 (2009).
104 Lee HC, Gu W, Shirayama M, Youngman E, Conte D Jr., Mello CC. C. elegans piRNAs mediate the genome-wide surveillance of germline transcripts. Cell 150(1), 78–87 (2012).
105 Grossniklaus U, Kelly B, Ferguson-Smith AC, Pembrey M, Lindquist S. Transgenerational epigenetic inheritance: how important is it? Nat. Rev. Genet. 14(3), 228–235 (2013).
106 Francis D, Diorio J, Liu D, Meaney MJ. Nongenomic transmission across generations of maternal behavior and stress responses in the rat. Science 286(5442), 1155–1158 (1999).
107 Francis DD, Champagne FA, Liu D, Meaney MJ. Maternal care, gene expression, and the development of individual differences in stress reactivity. Ann. NY Acad. Sci. 896, 66–84 (1999).
108 Wolff GL, Kodell RL, Moore SR, Cooney CA. Maternal epigenetics and methyl supplements affect agouti gene expression in Avy/a mice. FASEB J. 12(11), 949–957 (1998).
109 Waterland RA, Jirtle RL. Transposable elements: targets for early nutritional effects on epigenetic gene regulation. Mol. Cell. Biol. 23(15), 5293–5300 (2003).
110 Cropley JE, Suter CM, Beckman KB, Martin DI. Germ-line epigenetic modification of the murine A vy allele by nutritional supplementation. Proc. Natl Acad. Sci. USA 103(46), 17308–17312 (2006).
111 Cropley JE, Dang TH, Martin DI, Suter CM. The penetrance of an epigenetic trait in mice is progressively yet reversibly increased by selection and environment. Proc. Biol. Sci. 279(1737), 2347–2353 (2012).
112 Waterland RA, Travisano M, Tahiliani KG. Diet-induced hypermethylation at agouti viable yellow is not inherited transgenerationally through the female. FASEB J. 21(12), 3380–3385 (2007).
113 Morgan HD, Sutherland HG, Martin DI, Whitelaw E. Epigenetic inheritance at the agouti locus in the mouse. Nat. Genet. 23(3), 314–318 (1999).
114 Zamudio N, Bourc’his D. Transposable elements in the mammalian germline: a comfortable niche or a deadly trap? Heredity 105(1), 92–104 (2010).
115 Blewitt ME, Vickaryous NK, Paldi A, Koseki H, Whitelaw E. Dynamic reprogramming of DNA methylation at an epigenetically sensitive allele in mice. PLoS Genet. 2(4), e49 (2006).
116 Padmanabhan N, Jia D, Geary-Joo C et al. Mutation in folate metabolism causes epigenetic instability and transgenerational effects on development. Cell 155(1), 81–93 (2013).
•• Showedtransgenerationalepigeneticeffectsusingembryotransferexperiments.
117 Gapp K, Jawaid A, Sarkies P et al. Implication of sperm RNAs in transgenerational inheritance of the effects of early trauma in mice. Nat. Neurosci. 17(5), 667–669 (2014).
118 Schübeler D. Function and information content of DNA methylation. Nature 517(7534), 321–326 (2015).
119 Dunn GA, Bale TL. Maternal high-fat diet promotes body length increases and insulin insensitivity in second-generation mice. Endocrinology 150(11), 4999–5009 (2009).
120 Carone BR, Fauquier L, Habib N et al. Paternally induced transgenerational environmental reprogramming of metabolic gene expression in mammals. Cell 143(7), 1084–1096 (2010).
121 Ng SF, Lin RC, Laybutt DR, Barres R, Owens JA, Morris MJ. Chronic high-fat diet in fathers programs beta-cell dysfunction in female rat offspring. Nature 467(7318), 963–966 (2010).

846 Epigenomics (2015) 7(5) future science group
Review Prokopuk, Western & Stringer
122 Pembrey ME. Male-line transgenerational responses in humans. Hum. Fertil. (Camb) 13(4), 268–271 (2010).
123 Jimenez-Chillaron JC, Isganaitis E, Charalambous M et al. Intergenerational transmission of glucose intolerance and obesity by in utero undernutrition in mice. Diabetes 58(2), 460–468 (2009).
124 Wei Y, Yang C-R, Wei Y-P et al. Paternally induced transgenerational inheritance of susceptibility to diabetes in mammals. Proc. Natl Acad. Sci. USA 111(5), 1873–1878 (2014).
125 Guerrero-Bosagna C, Settles M, Lucker B, Skinner MK. Epigenetic transgenerational actions of vinclozolin on promoter regions of the sperm epigenome. PLoS ONE 5(9), e13100 (2010).
126 Radford EJ, Ito M, Shi H et al. In utero effects. In utero undernourishment perturbs the adult sperm methylome and intergenerational metabolism. Science 345(6198), 1255903 (2014).
127 Dunn GA, Bale TL. Maternal high-fat diet effects on third-generation female body size via the paternal lineage. Endocrinology 152(6), 2228–2236 (2011).
128 Lambrot R, Xu C, Saint-Phar S et al. Low paternal dietary folate alters the mouse sperm epigenome and is associated with negative pregnancy outcomes. Nat. Commun. 4, 2889 (2013).
129 Blount BC, Mack MM, Wehr CM et al. Folate deficiency causes uracil misincorporation into human DNA and chromosome breakage: implications for cancer and neuronal damage. Proc. Natl Acad. Sci. USA 94(7), 3290–3295 (1997).
130 Chong S, Vickaryous N, Ashe A et al. Modifiers of epigenetic reprogramming show paternal effects in the mouse. Nat. Genet. 39(5), 614–622 (2007).
131 Gu SG, Pak J, Guang S, Maniar JM, Kennedy S, Fire A. Amplification of siRNA in Caenorhabditis elegans generates a transgenerational sequence-targeted histone H3 lysine 9 methylation footprint. Nat. Genet. 44(2), 157–164 (2012).
132 Buckley BA, Burkhart KB, Gu SG et al. A nuclear Argonaute promotes multigenerational epigenetic inheritance and germline immortality. Nature 489 (7416) 447 –451 (2012).
133 Youngson NA, Whitelaw E. Transgenerational epigenetic effects. Annu. Rev. Genomics Hum. Genet. 9, 233–257 (2008).
134 Daxinger L, Whitelaw E. Transgenerational epigenetic inheritance: more questions than answers. Genome Res. 20(12), 1623–1628 (2010).
135 Huxley J. Evolution. The Modern Synthesis. Allen & Unwin, London (1942).
136 Mayr E. The Growth of Biological Thought: Diversity, Evolution, and Inheritance. Harvard University Press, USA (1982).
137 Wolf JB, Oakey RJ, Feil R. Imprinted gene expression in hybrids: perturbed mechanisms and evolutionary implications. Heredity 113(2), 167–175 (2014).
138 Jablonka E, Lamb MJ. Epigenetic Inheritance and Evolution: The Lamarckian Dimension. Oxford University Press, UK, p360 (1995).
139 Jablonka E, Lamb MJ. The evolution of information in the major transitions. J. Theor. Biol. 239(2), 236–246 (2006).
140 Jablonka E, Lamb MJ. Soft inheritance: challenging the modern synthesis. Genet. Mol. Biol. 31(2), 389–395 (2008).
• SummarizestheassumptionsoftheModernSynthesismodelforevolutionanddiscussestheneedforexpansiontoincorporatesoftinheritance.
141 Bossdorf O, Richards CL, Pigliucci M. Epigenetics for ecologists. Ecol. Lett. 11(2), 106–115 (2008).
142 Pigliucci M, Müller GB. Evolution, the Extended Synthesis. MIT Press, Cambridge, USA (2010).
143 Suter CM, Boffelli D, Martin DI. A role for epigenetic inheritance in modern evolutionary theory? A comment in response to Dickins and Rahman. Proc. Biol. Sci. 280(1771), 20130903; 20131820 (2013).
144 Li CC, Young PE, Maloney CA et al. Maternal obesity and diabetes induces latent metabolic defects and widespread epigenetic changes in isogenic mice. Epigenetics 8(6), 602–611 (2013).
145 Duhl DM, Vrieling H, Miller KA, Wolff GL, Barsh GS. Neomorphic agouti mutations in obese yellow mice. Nat. Genet. 8(1), 59–65 (1994).
146 Bateson P, Gluckman P. Plasticity and robustness in development and evolution. Int. J. Epidemiol. 41(1), 219–223 (2012).
147 Reik W, Constancia M, Fowden A et al. Regulation of supply and demand for maternal nutrients in mammals by imprinted genes. J. Physiol. 547(Pt 1), 35–44 (2003).
148 Haig D, Westoby M. Parent-specific gene expression and the triploid endosperm. Am. Nat. 134(1), 147–155 (1989).
149 Haig D. Genomic imprinting and kinship: how good is the evidence? Annu. Rev. Genet. 38(1), 553–585 (2004).
150 Wolf JB, Hager R. A maternal–offspring coadaptation theory for the evolution of genomic imprinting. PLoS Biol. 4(12), e380 (2006).
151 Keverne EB, Curley JP. Epigenetics, brain evolution and behaviour. Front. Neuroendocrinol. 29(3), 398–412 (2008).

781Epigenomics (2015) 7(5), 781–790 ISSN 1750-1911
part of
Perspective
10.2217/epi.15.46 © 2015 Future Medicine Ltd
Epigenomics
Perspective 2015/07/287
5
2015
Much controversy surrounds the idea of transgenerational epigenetics. Recent papers argue that epigenetic marks acquired through experience are passed to offspring, but as in much of the field of epigenetics, there is lack of precision in the definitions and perhaps too much eagerness to translate animal research to humans. Here, we review operational definitions of transgenerational inheritance and the processes of epigenetic programing during early development. Subsequently, based on this background, we critically examine some recent findings of studies investigating transgenerational inheritance. Finally, we discuss possible mechanisms that may explain transgenerational inheritance, including transmission of an epigenetic blueprint, which may predispose offspring to specific epigenetic patterning. Taken together, we conclude that presently, the evidence suggesting that acquired epigenetic marks are passed to the subsequent generation remains limited.
Keywords: animal models • early development • histone modifications • human • intergenerational • methylation • transgenerational
Conrad Waddington first used the term epi-genetics over half a century ago and defined it as ‘the branch of biology which studies the causal interactions between genes and their products which bring the phenotype into being.’ A more modern definition of epigenetics is: heritable chemical modifica-tions to DNA capable of influencing tran-scriptional activity with no direct alteration to the DNA sequence itself. It should be noted that heritable in this definition refers to ‘mitotic stability’ [1] or, in other words, to the fact that epigenetic information is passed to, and maintained, in the daughter cell upon mitotic division. Certainly, this type of epi-genetic heritability is of relevance to cancer research [2]. However, the concept of epigen-etic heritability has broadened over the years. Studies have progressively suggested that not only might epigenetic marks be inherited through mitotic processes, but also through meiosis, a concept akin to genetic inheri-tance. That is, similar to the way we inher-ited genetic traits from our parents through
germline transmission; epigenetic marks might also be propagated forward through the germline to offspring. This notion was fuelled by both research studies in plants and longitudinal epidemiological studies in humans, which have suggested that the effects of environmental influences were observed in subsequent generations. Stud-ies in the plant Arabidopsis thaliana showed that alterations in DNA methylation can be transmitted through several generations [3]. Famously in humans, epidemiological stud-ies examining the Dutch famine found that those individuals who were exposed to the famine while in utero, had increased sus-ceptibility to diseases in adulthood such as, diabetes, cardiovascular disease and obesity, to name a few [4]. These findings begged the question of lastingness, expressly, whether these predisposing factors, which seemingly stemmed from in utero exposure to famine, could be transmitted to the next generation. Though there is no direct molecular link to these major health concerns, second-
Transgenerational epigenetic inheritance: an open discussion
Corina Nagy1 & Gustavo Turecki*,1,2
1McGill Group for Suicide Studies,
Douglas Hospital University Institute,
6875 Lasalle boul, Montreal, QC, Canada 2Department of Psychiatry, McGill
University, Montreal, QC, Canada
*Author for correspondence:
For reprint orders, please contact: [email protected]

782 Epigenomics (2015) 7(5) future science group
Perspective Nagy & Turecki
generation offspring was found to demonstrate the same lower birth weights, as seen in first-generation affected individuals [4].
In contrast to the mitotic stability seen in somatic cells, transgenerational epigenetics requires germline transmission of acquired epigenetic information across generations. In cases of male exposure, the second generation may be sufficient to assess transgenera-tional effects. This is because exposure will affect the F0 male as well as his germline (F1), therefore only starting with the F2 generation can the effects of trans-generational epigenetic inheritance be considered. In females, however, studies require a third generation to test whether any of these DNA modifications are truly transmitted from one generation to the next. This is because one needs to rule out in utero exposure of the second generation to an environmental effect associ-ated with epigenetic changes. As an illustration, expos-ing a gestating female, her fetus and thus the fetal germline to a given environment, produces an F1 gen-eration with an associated epigenetic change reflecting direct environmental effects through in utero exposure (Figure 1). Prior to the F2 generation in males and the F3 generation in female, the inheritance detected in the offspring can only be considered intergenerational, in other words, parental effect due to direct expo-sure. This is important information to consider when reviewing some of the recent publications in this field.
Early developmental mechanisms: epigenetic resetEarly epigenetic reprogramming has been extensively studied in rodents. Though the same processes of early developmental epigenetic erasure occur in humans, the definitive characteristics and timeline of these events have still to be established [5]. Work in mice has demonstrated that maternal and paternal cells not only undergo a global erasure of DNA methyla-tion but the chromatin also undergoes active remod-eling. Global erasure occurs at two points. First, as the primordial germ cells (PGC) or gamete precursors develop and become part of the embryo, the epigenetic landscape is cleared to allow for cellular totipotency and the development of future generations through PGCs. This stage is followed by gametogenesis when the genomes undergo de novo methylation, a process that occurs later in the maternal genome. The second wave of demethylation occurs after fertilization as the gametes fuse to form the zygote (Figure 2). Here, however, imprinted genes escape the second round of demethylation (Figure 2, dotted lines), to carry for-ward parent-specific monoallelic expression in somatic cells. If epigenetic marks were to be maintained across generations, it is presumably at this point that they
could be carried forward. In fact, some loci, includ-ing repeated elements such as retrotransposons, do not undergo reprogramming during this phase [6,7]. Interestingly, Molaro and colleagues [8] found a strik-ing difference in methylation of retrotransposon sub-families when examining the sperm of humans and chimps. Furthermore, when comparing these sperm methylomes to embryonic stem cells, which they con-sider a mature germ cell having the product of the two reprogramming events in mammal, they found distinct characteristic, suggesting that sperm and somatic cells have different features, which deter-mine DNA methylation patterns in each cell type. Furthermore, approximately 100 nonimprinted, non-repetitive genes have been identified as maintaining promoter methylation levels throughout a range of developmental stages, from mature gametes to blas-tocysts [9]. Nonmammalian organisms on the other hand, appear to not undergo whole genome demeth-ylation upon fertilization [10]. In a comprehensive study using zebrafish, Jiang and colleagues [10] were able to demonstrate that no global erasure occurs at fertilization and by the midblastula stage the embryo’s methylome is almost identical to the of sperm, while the oocyte’s methylome contribution lessens by the 16-cell state. This suggests that the early reprogram-ming mechanisms are different between mammalian and non-mammalian organisms.
As mentioned above, both the mammalian mater-nal and paternal genomes undergo demethylation and they also experience active chromatin remodeling. This is particularly evident for the paternal genome. The process of spermatogenesis in humans requires that anywhere between 85 and 96% of histones be replaced by protamines, resulting in a tenfold compaction of the DNA [12,13]. It is believed that this process protects the paternal genome from physical and chemical dam-age. Protamines, like histones, carry post-translational modifications such as phosphorylation. The role of these modifications remains unknown, although they are believed to act similarly to histone modifications altering the availability of the DNA sequencing to the cellular transcriptional machinery [12]. Thus, the majority of paternally derived histone marks are lost during this process while the histones not replaced by protamines are thought to belong to genes expressed early in development [13]. Interestingly, the repressive mark, histone H3 lysine 27 trimethylation, is retained on some genes in the human and mouse spermatozoa. This is arguably another possible avenue for inheri-tance; conversely, it could be a methodological arti-fact, and simply be the result of a histone mark being re-established so quickly that its period of absence is not detected. Gradually, studies are demonstrating
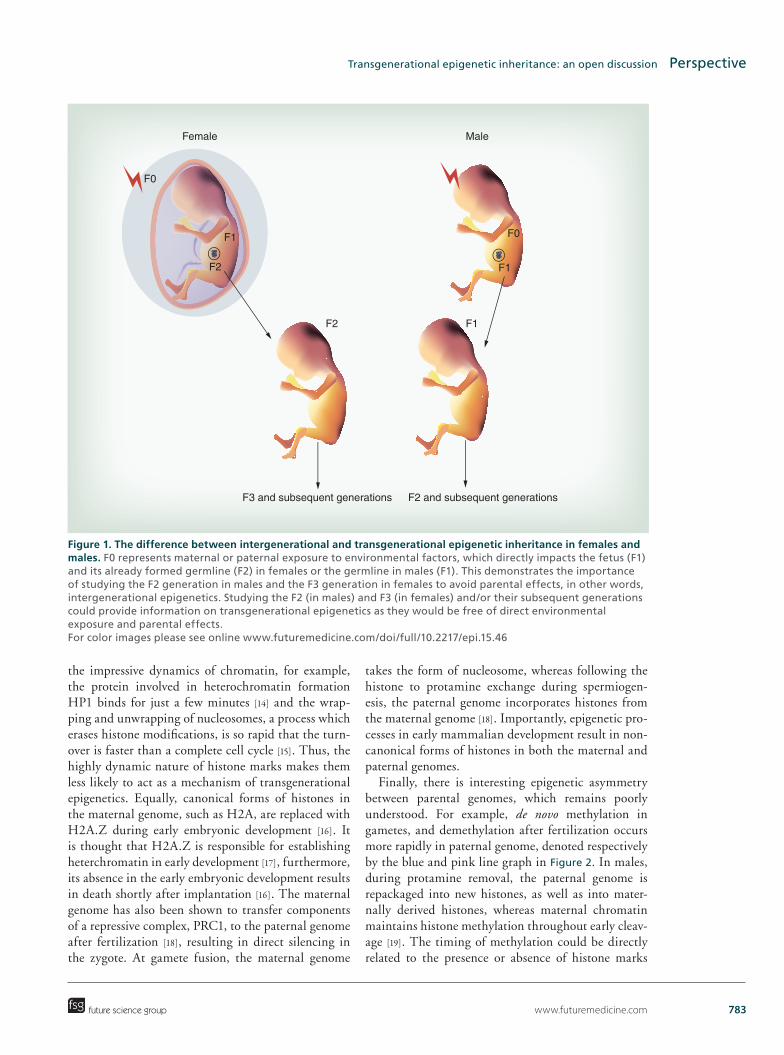
www.futuremedicine.com 783
Figure 1. The difference between intergenerational and transgenerational epigenetic inheritance in females and males. F0 represents maternal or paternal exposure to environmental factors, which directly impacts the fetus (F1) and its already formed germline (F2) in females or the germline in males (F1). This demonstrates the importance of studying the F2 generation in males and the F3 generation in females to avoid parental effects, in other words, intergenerational epigenetics. Studying the F2 (in males) and F3 (in females) and/or their subsequent generations could provide information on transgenerational epigenetics as they would be free of direct environmental exposure and parental effects. For color images please see online www.futuremedicine.com/doi/full/10.2217/epi.15.46
Female Male
F1
F1
F1
F0
F0
F2 and subsequent generationsF3 and subsequent generations
F2
F2
future science group
Transgenerational epigenetic inheritance: an open discussion Perspective
the impressive dynamics of chromatin, for example, the protein involved in heterochromatin formation HP1 binds for just a few minutes [14] and the wrap-ping and unwrapping of nucleosomes, a process which erases histone modifications, is so rapid that the turn-over is faster than a complete cell cycle [15]. Thus, the highly dynamic nature of histone marks makes them less likely to act as a mechanism of transgenerational epigenetics. Equally, canonical forms of histones in the maternal genome, such as H2A, are replaced with H2A.Z during early embryonic development [16]. It is thought that H2A.Z is responsible for establishing heterchromatin in early development [17], furthermore, its absence in the early embryonic development results in death shortly after implantation [16]. The maternal genome has also been shown to transfer components of a repressive complex, PRC1, to the paternal genome after fertilization [18], resulting in direct silencing in the zygote. At gamete fusion, the maternal genome
takes the form of nucleosome, whereas following the histone to protamine exchange during spermiogen-esis, the paternal genome incorporates histones from the maternal genome [18]. Importantly, epigenetic pro-cesses in early mammalian development result in non-canonical forms of histones in both the maternal and paternal genomes.
Finally, there is interesting epigenetic asymmetry between parental genomes, which remains poorly understood. For example, de novo methylation in gametes, and demethylation after fertilization occurs more rapidly in paternal genome, denoted respectively by the blue and pink line graph in Figure 2. In males, during protamine removal, the paternal genome is repackaged into new histones, as well as into mater-nally derived histones, whereas maternal chromatin maintains histone methylation throughout early cleav-age [19]. The timing of methylation could be directly related to the presence or absence of histone marks
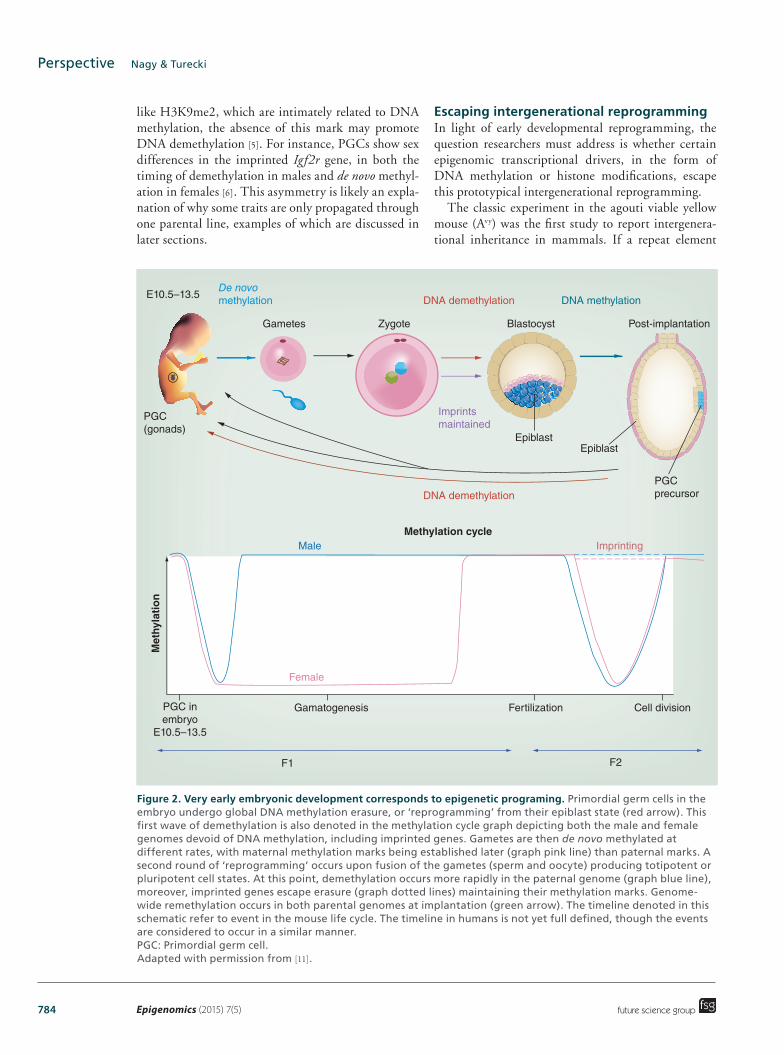
784 Epigenomics (2015) 7(5) future science group
Perspective Nagy & Turecki
like H3K9me2, which are intimately related to DNA methylation, the absence of this mark may promote DNA demethylation [5]. For instance, PGCs show sex differences in the imprinted Igf2r gene, in both the timing of demethylation in males and de novo methyl-ation in females [6]. This asymmetry is likely an expla-nation of why some traits are only propagated through one parental line, examples of which are discussed in later sections.
Escaping intergenerational reprogrammingIn light of early developmental reprogramming, the question researchers must address is whether certain epigenomic transcriptional drivers, in the form of DNA methylation or histone modifications, escape this prototypical intergenerational reprogramming.
The classic experiment in the agouti viable yellow mouse (Avy) was the first study to report intergenera-tional inheritance in mammals. If a repeat element
Figure 2. Very early embryonic development corresponds to epigenetic programing. Primordial germ cells in the embryo undergo global DNA methylation erasure, or ‘reprogramming’ from their epiblast state (red arrow). This first wave of demethylation is also denoted in the methylation cycle graph depicting both the male and female genomes devoid of DNA methylation, including imprinted genes. Gametes are then de novo methylated at different rates, with maternal methylation marks being established later (graph pink line) than paternal marks. A second round of ‘reprogramming’ occurs upon fusion of the gametes (sperm and oocyte) producing totipotent or pluripotent cell states. At this point, demethylation occurs more rapidly in the paternal genome (graph blue line), moreover, imprinted genes escape erasure (graph dotted lines) maintaining their methylation marks. Genome-wide remethylation occurs in both parental genomes at implantation (green arrow). The timeline denoted in this schematic refer to event in the mouse life cycle. The timeline in humans is not yet full defined, though the events are considered to occur in a similar manner. PGC: Primordial germ cell. Adapted with permission from [11].
Gamatogenesis Fertilization Cell division
Methylation cycle
Met
hyla
tio
n
PGC inembryo
E10.5–13.5
Female
Male Imprinting
F1 F2
Gametes
De novomethylation
Zygote
DNA demethylation
DNA demethylation
DNA methylation
Post-implantation
PGC precursor
EpiblastEpiblast
Blastocyst
Imprints maintained
PGC (gonads)
E10.5–13.5

www.futuremedicine.com 785future science group
Transgenerational epigenetic inheritance: an open discussion Perspective
called an intracisternal A particle (IAP), of which there are thousand of copies throughout the mouse genome, inserts itself upstream of the agouti locus, phenotypic variations can arise including altered coat color and obesity [20]. DNA methylation at the promoter region of this IAP, which in turn regulates the agouti gene, is inversely related to its transcriptional activity [20]. As indicated above, repeat elements may escape repro-gramming, which appears to be the case for this epial-lele that is passed along the maternal but not paternal germline [20]. Interesting as this may be, this is not the result of a past environmental factor being propa-gated forward via epigenetic inheritance. Though the insertion of the IAP at the agouti locus may not be entirely random [21], it does not result from the effects of environmental factors. However, once the IAP is inserted and susceptible to methylation changes, envi-ronment can play a role. This has been demonstrated by altering the food source of pregnant dams. Diets high in methyl groups such as folic acid or betaine can effectively ‘turn off ’ the Avy locus by methylating the promoter region of the IAP [22,23]. Though it is tempt-ing to look at this as transgenerational inheritance, it is rather the result of direct environmental exposure on the germline, and is thus the result of parental influ-ence, more appropriately termed, intergenerational inheritance.
Early life environment is also known to alter meth-ylation states as classically demonstrated through rat maternal behavior paradigms [24]. Caution must be taken when conducting transgenerational experi-ments to control for both in utero and early life expe-riences to avoid confounding early-life environmental effects with transgenerational inheritance. To avoid these potential confounding effects, both in utero and in early life, studies in rodent models have focused on paternal lineage as males are not involved in the in utero environment and are less likely to be involved in the early-life postnatal environment. An interesting example of this was found using outbred rats exposed to the fungicide, vinclozolin in utero. As a result, males exhibited diminished fertility over three to four gen-erations of offspring, which was found to be transmit-ted through the male germline [25]. Interestingly, these effects were not observed with another strain of rats. It is possible that this discrepancy results from method-ological differences between studies but it also raises the possibility that interaction with genetic variation might be responsible for the observed effect [26]. If this is the case, it represents an example of secondary epi-mutations, whereby DNA sequence variants have an effect on epigenetic marks (in cis or trans) [27]. Indeed a number of studies have demonstrated that genetic-based alterations can be responsible for the inheri-
tance of an epigenetic state [28–32]. In other words, in some cases, transmission of epigenetic marks can be secondary to inherited DNA sequence mutations.
Epigenetic inheritance: acquiring knowledgeA number of recent studies have implied that epi-genetic alteration induced by environmental factors affect the behavior of subsequent generations. Vas-soler and colleagues [33] found decreased cocaine self-administration in the first generation offspring of cocaine exposed male rats. The behavioral outcome suggests an almost protective effect of fathers consum-ing cocaine, which is not consistent with the clinical literature [34,35]. Molecularly, this is associated with increased Bdnf levels as well as increased H3 acetyla-tion, an epigenetic mechanism potentially responsible for this change. The authors of this intriguing study correctly suggested that more generations are required to definitively conclude that cocaine exposure results in transgenerational epigenetic alterations [36]. Cer-tainly, cocaine itself can alter chromatin states [37], but it is possible that cocaine directly influenced the gam-etes of the parental generation, and thus the F1 prog-eny. In a similar vein, a recent study in Caenorhabditis elegans showed paternal transmission to the embryo of the repressive mark H3K27me3, which is under the control of the Polycomb repression complex 2 [38]. Interestingly, histones containing this mark are not replaced by protamines during spermatogenesis in mammals [13]. This very interesting finding demon-strates mechanistic properties for the propagation of epigenetic marks through generations, but as pointed out previously, there is a need to demonstrate trans-generational inheritance beyond the first generation of progeny.
A study examining early stress in mice addressed the question of whether transgenerational inheritance could be explained by the effect of parental environ-ment on the gametes or through the transmission of acquired epigenetic marks to the offspring, by observ-ing the transmission of depressive-like behaviors up to the third generation [39,40]. The behavioral traits cose-gregated with altered DNA methylation in the male germ line. The same genes were tested for methylation levels in both the F2 sperm and the F2 female brains. The methylation patterns in these cell types were simi-lar, though certainly not identical. However, of note was a discrepancy in behavioral phenotype, where the F1 and F3 males showed depressive-like behaviors but not the F2 males. The depressive-like phenotype was only seen in the F2 females and the molecular changes reported in this study were correlational, as there was no experiment in this study showing a causal link

786 Epigenomics (2015) 7(5) future science group
Perspective Nagy & Turecki
between methylation changes and phenotype. Further-more, the exposure of females to stressed males, even for a short period, could have had subtle effects on the female behaviors and hormonal expression, which in turn may have affected maternal care. In vitro fertiliza-tion (IVF) of the sperm directly into control females has been used to avoid these potential confounding effects from fathers. When IVF was used in social defeat paradigms, which is an animal model of depres-sion, the depressive-like phenotype that seemed to be propagated forward under normal mating conditions was largely absent [40]. Together, these results suggest that broad behavioral conditioning can be inherited. Although there is a possibility that these may cosegre-gate with specific epigenetic profiles, additional work is necessary to definitively make such conclusions.
Another study using the same early stress paradigm in mice showed depressive-like behaviors associated with an upregulation of several miRNA in sperm which appeared to affect serum and hippocampal miRNAs levels in the subsequent generation [41]. Because small RNAs are highly present in sperm, they have become candidate vectors for conveying trans-generational inheritance [42]. However, in this study, although the behavioral phenotype was detected in the F3 generation, the miRNA levels of five candi-date miRNAs in F2 sperm were unchanged and the alterations seen in these candidate miRNAs from F2 were no longer detectable in F3. The authors suggest that the initial change in miRNA levels resulting from early stress may have been transferred to other epigen-etic marks, but give no evidence of this effect. Another study from the same group set out to demonstrate a causal mechanism for the inherited stress responses in the offspring of stressed sires [43]. In this study, the authors were able to pharmacologically induce the same behavioral traits seen in the stressed offspring, with corresponding alteration to gene expression levels resulting from histone methyl- and acetyl-transferase inhibitors, suggesting a causal role for mineral corti-coid receptors in the altered stress response of pater-nally stressed offspring. Here, however, the epigenetic changes seen in the male germline differed from what was found in the adult hippocampus of the offspring, specifically methylation changes where seen in the sperm whereas altered histone modifications were seen in the brains. Collectively, it appears that whatever environmentally induced epigenetic alteration is tak-ing place in one generation may be propagated for-ward, but as noted above, although possible, it is still premature to conclusively suggest a clear association between one acquired epigenetic mark and transgener-ational behavioral phenotypes. An enticing possibility is that epigenetic changes transmitted across genera-
tions may act in a probabilistic manner. In other words, these changes are more or less likely to occur in broad genomic regions, resulting in altered behavior without producing a consistent molecular phenotype. A recent paper demonstrating that paternal sugar consumption in fruit flies influences the metabolic properties of the F1 generation, leading to an obesity phenotype, sup-ports this concept. Here, the authors demonstrate that a network of genes is required for proper intergenera-tional metabolic reprogramming involving a number of changes to chromatin structure. No specific sites were identified but rather a pattern or signature of gene dysfunction was identified as conferring susceptibility to obesity [44].
Epigenetic inheritance: from rodents to humansThe vast majority of studies examining transgenera-tional inheritance have been conducted in animals other than humans. Though mice and rats provide convenient models for human disorders and behav-ioral traits, there are different physiological and bio-logical processes between humans and rodents. For instance, a recent paper exploring the methylation landscape of early human embryos showed features distinctive to humans, specifically levels of methyla-tion and timing of genome-wide demethylation [45], while another study suggests the symmetry of epigen-etic reprogramming cycles may differ between spe-cies [6]. Moreover, many transcriptionally relevant epigenetic marks show considerable sequence diver-gence between mice and humans [46]. Furthermore, information about epigenetic erasure during early development comes primarily from mouse mod-els, and it is possible to be different in humans, but remains, as yet, untested. Transgenerational effects have been observed in humans through longitudinal studies [47–49], whereas gene–environment interplay is suspected as a mediating factor in health outcome. Some researchers have attempted to approach epigen-etic inheritance using in a systems biology approach, whereby acquired behavioural and physiological changes, are considered through the framework of a “two hit” model. This approach recognizes both, the biology established in a previous generation and the organism’s present environment, as involved in the development of these changes [50]. In a study explor-ing vinclozolin-lineage rats presumed to be carry-ing epigenetic alterations from previous generations, the authors identified gene network and behavioural alterations associated with the lineage specific rats when subjected to a stress paradigm [51]. Though this study does not answer the question of whether or not these genetic and behavioural changes would have

www.futuremedicine.com 787future science group
Transgenerational epigenetic inheritance: an open discussion Perspective
occurred in the absence of the induced environmental stress (context) nor does it confirm the presence of epigenetically altered states, it nonetheless represents the attractive notion of dynamic interactions between context dependence and “predisposing factors”, in this case, environment and inherited epigenetic state.Intuitively, most people can recognize that their envi-ronment plays a role in health and behavior, but we still lack conclusive evidence to indicate that such transgenerational effects are explained by acquired epigenetic mechanisms inherited from one generation to the next.
ConclusionThe studies reviewed above suggest that there is some evidence that certain epigenetic marks escape erasure in early development. There is also evidence that cer-tain acquired behavioral phenotypes are transmitted through subsequent generations. Although promising, the evidence suggesting that acquired epigenetic marks cosegregate with acquired behavioral phenotypes remains inconclusive and open to a number of potential alternative interpretations, including methodological explanations.
One should be particularly cautious when interpret-ing the results of studies testing transgenerational epi-genetic inheritance in animals to the lay media or gen-eral public. Though likely not the intention, as pointed out previously [52], animal studies have been vulgarized and commonly anthropomorphized into a public mes-sage that overemphasizes the molecular impact of the environment, including parents and grandparents onto their children.
The field of epigenetics is fascinating and holds great potential in medicine, both to uncover dis-ease biomarkers and therapeutic interventions. The results suggesting that acquired epigenetic factors may be transmitted through generations and explain acquired phenotypic traits are promising and intrigu-ing, but nevertheless, studies published thus far have limitations and prevent us from making definitive conclusions.
Future perspectiveAn important missing link in the study of transgen-erational epigenetic inheritance is a mechanism by which gene-regulatory information is transferred from somatic cells to germs cells. Efforts to uncover
Executive Summary
Early developmental mechanisms: epigenetic reset• Epigenetic reprogramming occurs at two points during early development, after fertilization and in primordial
germ cells.• Some genomic features, such as retrotransposons, as well as certain histone marks can carry forward
epigenetic marks in spite of these erasure mechanisms, supporting the possibility that epigenetic marks can be maintained across generations.
Escaping intergenerational reprogramming• Mice and rat models have been used to show that certain epigenetic marks can escape intergenerational
reprogramming, affecting the phenotype of future generations.• Effects resulting from parental influences result in intergeneration inheritance, which is not to be confused
with transgenerational inheritance (effects that survive across generations in the absence of direct exposure).• Secondary epimutations represent another factor that can influence possible transgenerational inheritance. In
these cases, epigenetic marks are secondary to inherited DNA sequence mutations.Epigenetic inheritance: acquiring knowledge• A number of studies including recent articles providing mechanistic insight show that epigenetic marks can be
inherited intergenerationally.• Data for transgenerational inheritance on the other hand show certain inconsistencies, which may be
suggestive of probabilistic epigenetic changes capable of influencing phenotypic outcome in subsequent generations.
Epigenetic inheritance: from rodents to humans• It is difficult to provide concrete evidence that transgenerational epigenetics occurs in humans owing to the
lack of studies in humans and the physiological and biological differences between humans and rodents.Conclusion• There is evidence that epigenetic inheritance occurs intergenerationally by escaping erasure in early
development.• However, evidence is lacking for the co-segregation of acquired epigenetic marks with acquired behavioral
traits transgenerationally.• The field is very promising but more studies are required in order to provide definitive conclusions on the
topic of transgenerational inheritance, particularly in humans.

788 Epigenomics (2015) 7(5) future science group
Perspective Nagy & Turecki
these mechanisms have been made in nonmamma-lian animals. For example, a number of studies in C. elegans have implicated small RNAs in the process of inherited epigenetic marks [53–55]. It was recently demonstrated in that dsRNA act as mobile elements that mediating intertissue transfer of regulatory infor-mation by entering the cytosol via a dsRNA-selective importer [56]. Equally, it has been speculated, notably by a paper discussed here [41] and others [57,58], that small noncoding RNAs, like miRNA, mediate soma to germline transfer of regulatory information in mammals, however, experimental evidence in mam-mals is lacking. Tackling this question in mammals and particularly in humans is an important next step in the research of transgenerational epigenetics.
Moving forward in the field of transgenerational epigenetics requires more precision in experimental design. First, studies should investigate both acquired
phenotypic traits and acquired epigenetic marks, and their co-segregation, through three generations at least. In addition to correlational evidence, studies should also investigate causative links between the molecular changes investigated and the phenotypes. Finally, although more difficult to conduct, studies are also necessary in humans.
Financial & competing interests disclosureThe authors have no relevant affiliations or financial in-
volvement with any organization or entity with a financial
interest in or financial conflict with the subject matter or
materials discussed in the manuscript. This includes employ-
ment, consultancies, honoraria, stock ownership or options,
expert testimony, grants or patents received or pending, or
royalties.
No writing assistance was utilized in the production of
this manuscript.
ReferencesPapers of special note have been highlighted as: • of interest; •• of considerable interest
1 Skinner MK. Environmental epigenetic transgenerational inheritance and somatic epigenetic mitotic stability. Epigenetics 6(7), 838–842 (2011).
2 Dawson MA, Kouzarides T. Cancer epigenetics: from mechanism to therapy. Cell 150(1), 12–27 (2012).
3 Johannes F, Porcher E, Teixeira FK et al. Assessing the impact of transgenerational epigenetic variation on complex traits. PLoS Genet. 5(6), e1000530 (2009).
4 Painter RC, Osmond C, Gluckman P, Hanson M, Phillips DI, Roseboom TJ. Transgenerational effects of prenatal exposure to the dutch famine on neonatal adiposity and health in later life. BJOG 115(10), 1243–1249 (2008).
5 Leitch HG, Tang WW, Surani MA. Primordial germ-cell development and epigenetic reprogramming in mammals. Curr. Top. Dev. Biol. 104, 149–187 (2013).
6 Hyldig SM, Croxall N, Contreras DA, Thomsen PD, Alberio R. Epigenetic reprogramming in the porcine germ line. BMC Dev. Biol. 11, 11 (2011).
7 Law JA, Jacobsen SE. Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat. Rev. Genet. 11(3), 204–220 (2010).
8 Molaro A, Hodges E, Fang F et al. Sperm methylation profiles reveal features of epigenetic inheritance and evolution in primates. Cell 146(6), 1029–1041 (2011).
9 Borgel J, Guibert S, Li Y et al. Targets and dynamics of promoter DNA methylation during early mouse development. Nat. Genet. 42(12), 1093–1100 (2010).
10 Jiang L, Zhang J, Wang JJ et al. Sperm, but not oocyte, DNA methylome is inherited by zebrafish early embryos. Cell 153(4), 773–784 (2013).
• Inthisstudy,theauthorsdemonstratethatthetimingandinheritanceofepigeneticmarksinzebrafisharedrivenbythemalegermline.
11 Fresard L, Morisson M, Brun JM et al. Epigenetics and phenotypic variability: some interesting insights from birds. Genet. Sel. Evol. 45, 16 (2013).
12 Brunner AM, Nanni P, Mansuy IM. Epigenetic marking of sperm by post-translational modification of histones and protamines. Epigenetics Chromatin 7(1), 2 (2014).
13 Brykczynska U, Hisano M, Erkek S et al. Repressive and active histone methylation mark distinct promoters in human and mouse spermatozoa. Nat. Struct. Mol. Biol. 17(6), 679–687 (2010).
14 Festenstein R, Pagakis SN, Hiragami K et al. Modulation of heterochromatin protein 1 dynamics in primary mammalian cells. Science 299(5607), 719–721 (2003).
15 Deal RB, Henikoff S. Histone variants and modifications in plant gene regulation. Curr. Opin. Plant Biol. 14(2), 116–122 (2011).
16 Faast R, Thonglairoam V, Schulz TC et al. Histone variant h2a.Z is required for early mammalian development. Curr. Biol. 11(15), 1183–1187 (2001).
17 Banaszynski LA, Allis CD, Lewis PW. Histone variants in metazoan development. Dev. Cell 19(5), 662–674 (2010).
18 Puschendorf M, Terranova R, Boutsma E et al. Prc1 and suv39h specify parental asymmetry at constitutive heterochromatin in early mouse embryos. Nat. Genet. 40(4), 411–420 (2008).
19 Zhou LQ, Dean J. Reprogramming the genome to totipotency in mouse embryos. Trends Cell. Biol. 25(2), 82–91 (2014).
20 Morgan HD, Sutherland HG, Martin DI, Whitelaw E. Epigenetic inheritance at the agouti locus in the mouse. Nat. Genet. 23(3), 314–318 (1999).
21 Baillie JK, Barnett MW, Upton KR et al. Somatic retrotransposition alters the genetic landscape of the human brain. Nature 479(7374), 534–537 (2011).
22 Waterland RA, Jirtle RL. Transposable elements: targets for early nutritional effects on epigenetic gene regulation. Mol. Cell. Biol. 23(15), 5293–5300 (2003).

www.futuremedicine.com 789future science group
Transgenerational epigenetic inheritance: an open discussion Perspective
• Evidencesthattransposableelementscanescapeearlydevelopmentalreprogramming.
23 Wolff GL, Kodell RL, Moore SR, Cooney CA. Maternal epigenetics and methyl supplements affect agouti gene expression in avy/a mice. FASEB J. 12(11), 949–957 (1998).
24 Weaver IC, Cervoni N, Champagne FA et al. Epigenetic programming by maternal behavior. Nat. Neurosci. 7(8), 847–854 (2004).
• Demonstrateshowenvironmentcanhavearelativelylong-lastingeffectonbehaviorthroughepigeneticalterations.
25 Anway MD, Cupp AS, Uzumcu M, Skinner MK. Epigenetic transgenerational actions of endocrine disruptors and male fertility. Science 308(5727), 1466–1469 (2005).
26 Schneider S, Kaufmann W, Buesen R, Van Ravenzwaay B. Vinclozolin – the lack of a transgenerational effect after oral maternal exposure during organogenesis. Reprod. Toxicol. 25(3), 352–360 (2008).
27 Oey H, Whitelaw E. On the meaning of the word ‘epimutation’. Trends Genet. 30(12), 519–520 (2014).
28 Bell JT, Pai AA, Pickrell JK et al. DNA methylation patterns associate with genetic and gene expression variation in hapmap cell lines. Genome Biol. 12(1), R10 (2011).
29 Gertz J, Varley KE, Reddy TE et al. Analysis of DNA methylation in a three-generation family reveals widespread genetic influence on epigenetic regulation. PLoS Genet. 7(8), e1002228 (2011).
30 Gutierrez-Arcelus M, Ongen H, Lappalainen T et al. Tissue-specific effects of genetic and epigenetic variation on gene regulation and splicing. PLoS Genet. 11(1), e1004958 (2015).
31 Zhang D, Cheng L, Badner JA et al. Genetic control of individual differences in gene-specific methylation in human brain. Am. J. Hum. Genet. 86(3), 411–419 (2010).
32 Cini G, Carnevali I, Quaia M et al. Concomitant mutation and epimutation of the mlh1 gene in a lynch syndrome family. Carcinogenesis 36(4), 452–458 (2015).
33 Vassoler FM, White SL, Schmidt HD, Sadri-Vakili G, Pierce RC. Epigenetic inheritance of a cocaine-resistance phenotype. Nat. Neurosci. 16(1), 42–47 (2013).
34 Johnson JL, Leff M. Children of substance abusers: overview of research findings. Pediatrics 103(5 Pt. 2), 1085–1099 (1999).
35 Delaney-Black V, Chiodo LM, Hannigan JH et al. Prenatal and postnatal cocaine exposure predict teen cocaine use. Neurotoxicol. Teratol. 33(1), 110–119 (2011).
36 Vassoler FM, Sadri-Vakili G. Mechanisms of transgenerational inheritance of addictive-like behaviors. Neuroscience 264, 198–206 (2014).
37 Maze I, Feng J, Wilkinson MB, Sun H, Shen L, Nestler EJ. Cocaine dynamically regulates heterochromatin and repetitive element unsilencing in nucleus accumbens. Proc. Natl Acad. Sci. USA 108(7), 3035–3040 (2011).
38 Gaydos LJ, Wang W, Strome S. Gene repression. H3K27me and PRC2 transmit a memory of repression across generations and during development. Science 345(6203), 1515–1518 (2014).
39 Franklin TB, Russig H, Weiss IC et al. Epigenetic transmission of the impact of early stress across generations. Biol. Psychiatry 68(5), 408–415 (2010).
40 Dietz DM, Laplant Q, Watts EL et al. Paternal transmission of stress-induced pathologies. Biol. Psychiatry 70(5), 408–414 (2011).
41 Gapp K, Jawaid A, Sarkies P et al. Implication of sperm rnas in transgenerational inheritance of the effects of early trauma in mice. Nat. Neurosci. 17(5), 667–669 (2014).
42 Kawano M, Kawaji H, Grandjean V, Kiani J, Rassoulzadegan M. Novel small noncoding rnas in mouse spermatozoa, zygotes and early embryos. PLoS ONE 7(9), e44542 (2012).
43 Gapp K, Soldado-Magraner S, Alvarez-Sanchez M et al. Early life stress in fathers improves behavioural flexibility in their offspring. Nat. Commun. 5, 5466 (2014).
•• Providesevidenceinmammalsthatcertainhistonemarkscanbemantainedthroughdevelopmentalreprogramming.
44 Ost A, Lempradl A, Casas E et al. Paternal diet defines offspring chromatin state and intergenerational obesity. Cell 159(6), 1352–1364 (2014).
45 Guo H, Zhu P, Yan L et al. The DNA methylation landscape of human early embryos. Nature 511(7511), 606–610 (2014).
46 Yue F, Cheng Y, Breschi A et al. A comparative encyclopedia of DNA elements in the mouse genome. Nature 515(7527), 355–364 (2014).
47 Bygren LO, Kaati G, Edvinsson S. Longevity determined by paternal ancestors’ nutrition during their slow growth period. Acta Biotheor. 49(1), 53–59 (2001).
48 Kaati G, Bygren LO, Edvinsson S. Cardiovascular and diabetes mortality determined by nutrition during parents’ and grandparents’ slow growth period. Eur. J. Hum. Genet. 10(11), 682–688 (2002).
49 Pembrey ME, Bygren LO, Kaati G et al. Sex-specific, male-line transgenerational responses in humans. Eur. J. Hum. Genet. 14(2), 159–166 (2006).
50 Crews D, Gore AC. Transgenerational epigenetics – current controversiesand debates. In: Transgenerational Epigenetics. Tollefsbol T (Ed.). Elsevier, MA, USA, 371–387 (2014).
51 Crews D, Gillette R, Scarpino SV, Manikkam M, Savenkova MI, Skinner MK. Epigenetic transgenerational inheritance of altered stress responses. Proc. Natl Acad. Sci. USA. 109(23), 9143–9148 (2012).
52 Juengst ET, Fishman JR, McGowan ML, Settersten RA Jr. Serving epigenetics before its time. Trends Genet. 30(10), 427–429 (2014).
53 Buckley BA, Burkhart KB, Gu SG et al. A nuclear argonaute promotes multigenerational epigenetic inheritance and germline immortality. Nature 489(7416), 447–451 (2012).
54 Ashe A, Sapetschnig A, Weick EM et al. Pirnas can trigger a multigenerational epigenetic memory in the germline of C. elegans. Cell 150(1), 88–99 (2012).
55 Burton NO, Burkhart KB, Kennedy S. Nuclear rnai maintains heritable gene silencing in Caenorhabditis elegans. Proc. Natl Acad. Sci. USA 108(49), 19683–19688 (2011).
56 Devanapally S, Ravikumar S, Jose AM. Double-stranded RNA made in C. elegans neurons can enter the germline and

790 Epigenomics (2015) 7(5) future science group
Perspective Nagy & Turecki
cause transgenerational gene silencing. Proc. Natl Acad. Sci. USA 112(7), 2133–2138 (2015).
57 Sharma A. Bioinformatic analysis revealing association of exosomal mrnas and proteins in epigenetic inheritance. J. Theor. Biol. 357, 143–149 (2014).
58 Sharma A. Novel transcriptome data analysis implicates circulating microRNAs in epigenetic inheritance in mammals. Gene 538(2), 366–372 (2014).