thesis section: restorative neurology
DESCRIPTION
Thesis section: Restorative neurologyhttp://yassermetwally.comhttp://yassermetwally.netTRANSCRIPT

i
Restorative neurologyEssay
In NeuropsychiatrySubmitted for partial fulfillment of Master Degree
BySamy moussa Seliem
M.B.B.CH
Supervisors of
Prof. Mohammed Yasser Metwally
Professor of Neuropsychiatry
Faculty of Medicine-Ain Shams University
www.yassermetwally.com
Prof. Naglaa Mohamed Elkhayat
Professor of Neuropsychiatry
Faculty of Medicine-Ain Shams University
Dr. Haitham Hamdy salem
Lecturer of Neuropsychiatry
Faculty of Medicine-Ain Shams University
Faculty of Medicine
Ain Shams University
2011

i
CCoonntteennttss
Subject
1. Introduction and aim of the work
2. Stem cell
3. Stem cell therapy in Parkinson disease
4. Stem cell therapy in in stroke
5. Stem cell therapy in demyelinating disease
6. Stem cell therapy in in amyotrophic lateral sclerosis
7. Stem cell therapy in muscular dystrophy
9. Stem cell therapy for Alzheimer's Disease
10.Stem cell therapy in degenerative diseases in children
11.Stem cell therapy in retinal degeneration
12.Stem cell therapy in spinal cord injury
13.Stem cell therapy in peripheral nerve injury
8. Stem cell therapy Huntington chorea
References
Summary

i
Introduction
Stem cells are unspecialized cells in the human body that are
capable of becoming specialized cells, each with new
specialized cell functions. The best example of a stem cell is the
bone marrow stem cell that is unspecialized and able to
specialize into blood cells, such as white blood cells and red
blood cells, and these new cell types have special functions,
such as being able to produce antibodies, act as scavengers to
combat infection and transport gases. Thus one cell type stems
from the other and hence the term “stem cell.” Basically, a stem
cell remains uncommitted until it receives a signal to develop
into a specialized cell. Stem cells have the remarkable properties
of developing into a variety of cell types in the human body.
They serve as a repair system by being able to divide without
limit to replenish other cells. When stem cell divides, each new
cell has the potential to either remain as a stem cell or become
another cell type with new special functions, such as blood cells,
brain cells, etc. (Bongso and Lee, 2005).
Stem cells also known as progenitor cells which are cells
that have not undergone differentiation to acquire specific
structure or role. They have the potential to self-renew, divide
and differentiate into specialized cell types. They are also,
sometimes, termed ‘pluripotent’ or ‘undifferentiated’ cells

ii
because they can differentiate and develop into various cell lines
(Metwally, 2009).
Scientists and researchers are interested in stem cells for
several reasons. Although stem cells do not serve any one
function, many have the capacity to serve any function after
they are instructed to specialize. Every cell in the body, for
example, is derived from first few stem cells formed in the early
stages of embryological development. Therefore, stem cells
extracted from embryos can be induced to become any desired
cell type. This property makes stem cells powerful enough to
regenerate damaged tissue under the right conditions (Crosta,
2010).
Perhaps, the most important reason that stem cell
development is so appealing to neurologists can be found in the
statement “The adult human brain, in contrast to other organs
such as skin and liver, lacked the capacity for self repair and
regeneration” (Lin et al., 2007).
The types of stem cells include: Bone marrow-derived
mesenchymal stem cells (BMSCs), embryonic stem cells
(ESCs), Adult (somatic) stem cells, and neural stem cells
(NSCs). BMSCs also termed bone marrow stromal cells are
another example of a somatic stem cell being studied for its
therapeutic potential in the central nervous system (CNS) and in
other tissue (Abdallah and Kassem, 2008).

iii
BMSCS generate neurotransmitter-responsive cells with
electro-physiological properties similar to neurons (Diana and
Gabriel, 2008).
ESCs are pluripotent cells isolated from the inner cell mass
of day 5-8 blastocyte with indefinite self-renewal capabilities as
well as of the ability to differentiate into all cell types derived
from the three embryonic germ layers. The primary therapeutic
goal of ESCs research is cell replacement therapy (Aoki et al.,
2007).
Adult (somatic) stem cells: it has a capacity to differentiate
into tissue-specific types and represent a potential source of
autologus cells for transplantation therapy that eliminate
immunological complications associated with allogenic donor
cells as well as bypass ethical concern associated with ESCs,
All types are generally characterized by their potency, or
potential to differentiate into different cell types (such as skin,
muscle, bone, etc) (Lin et al., 2007).
Scientists discovered ways to obtain or derive stem cells
from early mouse embryos more than 20 years ago. Many years
of detailed study of biology of mouse stem cells led to the
discovery, in 1998, of how to isolate stem cells from human
embryos and grow the cells in the laboratory. These are called
human embryonic stem cells. The embryos used in these studies
were created for infertility purposes through in vitro fertilization

iv
procedures and when they were no longer needed for that
purpose, they were donated for research with the informed
consent of the donor (Ordrico et al., 2001).
The concept that the adult mammalian CNS contains NSCs
was first discovered from evidence of neuronal turnover in the
olfactory bulb and hippocampus in the adult organism cells with
more restricted neural differentiation capabilities committed to
specific subpopulation lineage, have been generated from
human ESCs or directly isolated from neurogenic regions of
fetal and adult CNS, such as the subventricular zone, which
provides neuroblasts to replenish inhibitory interneurons in the
olfactory bulb (Lin et al., 2007).
Stem cell differentiation must be turned on, given direction,
and turned off as needed in order to properly supply the basic
building blocks of tissues in different organ systems. This
requirement for precise regulation applies to an even greater
degree to the differentiation of neuronal progenitor cells,
because effective neural function depends on establishing
precise linkage and interactions between different individual
neurons and classes of neurons (Metwally, 2009).
Most tissue repair events in mammals are dedifferentiation
independent events brought about by the activation of pre-
existing stem cells or progenitor cells. By definition, a
progenitor cell lies in between a stem cell and a terminally
differentiated cell (Crosta, 2010).

v
With the therapeutic application of NSCs for
neurorestoration in mind, a clearer picture is emerging. Both in
normal neurodevelopment and stem cell biology, the precursor
cells display preprogrammed behavior modified by cues from
the local environment. The fundamental assumption is that
differentiation and predictable behavior of NSCs can be
achieved if the appropriate cocktail of soluble/diffusible or
contact-mediated signals is present. In addition, several
corollary considerations are quickly evident. For example, can
we use NSCs from different sources in an equivalent fashion?
The answer to this important question requires that we
understand the developmental potential of all the types of NSCs
(Marquez et al., 2005).
Medical researchers believe that stem cell therapy has the
potential to dramatically change the treatment of human disease.
A number of adult stem cell therapies already exist, particularly
bone marrow transplants that are used to treat leukemia. In the
future, medical researchers anticipate being able to use
technologies derived from stem cell research to treat a wider
variety of diseases including cancer, Parkinson's disease, spinal
cord injuries, Amyotrophic lateral sclerosis, multiple sclerosis,
and muscle damage, amongst a number of other impairments
and conditions (Goldman and Windrem, 2006).

vi
Aim of the work:
The aim of this work is to study and summarize recent
progress in stem cell therapies aimed at neurodegenerative
disorder and illustrate how some of aforementioned methods
and strategies are being utilized to formulate clinically viable
treatments.

1
Stem cells
Definition: A stem cell is a cell that has the ability to divide
(self replicate) for indefinite periods, often throughout the life of
the organism. Under the right conditions, or given the right
signals, stem cells can give rise (differentiate) to the many
different cell types that make up the organism. That is, stem
cells have the potential to develop into mature cells that have
characteristic shapes and specialized functions, such as heart
cells, skin cells, or nerve cells (Charron et al., 2009).
The word “stem” actually originated from old botanical
monographs from the same terminology as the stems of plants,
where stem cells were demonstrated in the apical root and shoot
meristems that were responsible for the regenerative
competence of plants. Hence also the use of word “stem” in
“meristem” (Kiessling and Anderson, 2003).
Historical overview of stem cell therapy:The stem cell is the origin of life. As stated first by the great
pathologist (Rudolph Virchow), “All cells come from cells”.
The fertilized egg is formed from fusion of the haploid progeny
of germinal stem cells. The fertilized egg is totipotent; from it
forms all the tissues of the developing embryo. During
development of the embryo, germinal stem cells are formed,
which persist in adult to allow the cycle of life to continue. In

2
the adult, tissue is renewed by proliferation of specialized stem
cells, which divide to form one cell that remains a stem cell and
another cell that begins the process of differentiation to
specialized function of a mature cell type, normal tissue renewal
is accomplished by the differentiating progeny of stem cells, the
so-called transit amplifying cells. For example, blood cells are
mature cells derived from hematopoietic stem cells in the bone
marrow; the lining cells of the gastrointestinal tract are formed
from transit amplifying cells, progeny of stem cell in the base of
intestinal glands (Crosta, 2010).
Nineteenth century pathologists first hypothesized the
presence of stem cells in the adult as “embryonal rests” to
explain the cellular origin of cancer and the studies indicate that
the most cancers arise from stem cells or their immediate
progeny, the transit-amplifying cells. Cancer results from an
imbalance between the rate at which cells are produced and the
rate at which they terminally differentiate or die. Understanding
how to control the proliferation and differentiation of stem cells
and their progeny is not only the key to controlling and treating
cancer, but also to cell replacement and gene therapy for many
metabolic, degenerative, and immunological diseases (Virchow,
1985).
Stem cell properties:
Stem cells have a capacity for self-renewal giving rise to
more stem cells, and the ability to differentiate into tissues of

3
various lineages under appropriate conditions. They may be
totipotent, pluripotent or multipotent, depending on type. Only
the embryo is totipotent. Embryonic stem cells (ESCs) are
pluripotent, as they are capable of differentiating into many
tissue types, whereas differentiation of adult stem cells is
generally restricted to the tissue in which they reside, as with
hepatocytes in the liver, and haemopoietic stem cells in blood
(figure 1) (Bongso and Lee, 2005).
Figure (1): Stem cell self-renewal and differentiation (Bongso and
Lee, 2005).

4
A) Stem cell self renewal:
The defining feature of a true stem cell is the capacity for
self-renewal. Self renewal occurs when a cell that has been
activated to divide does so asymmetrically. The result produces
one cell that is exactly like the mother cell and one cell that
takes on biological functions that are different from those of the
mother cell. Without self-renewal, each activation event would
result in the progressive loss of the originating stem cell
population (Andeson et al., 2001).
B) The stem cell life cycle:
Stem cell activation is generally followed by a clonal
expansion of the daughter cell that is produced. This is
associated with a series of biological processes that include
proliferation, migration, differentiation, and at some point cell
death. Regulation of these downstream events determines the
net effect that, each stem cell activation has on new tissue
formation (Song et al., 2007).
C) Stem cell plasticity:
The term plasticity means that a stem cell from one adult
tissue can generate the differentiated cell types of another tissue.
At this time, there is no formally accepted name for this
phenomenon in the scientific literature. It is variously referred
to as “plastisity” “unorthodox differentition” or
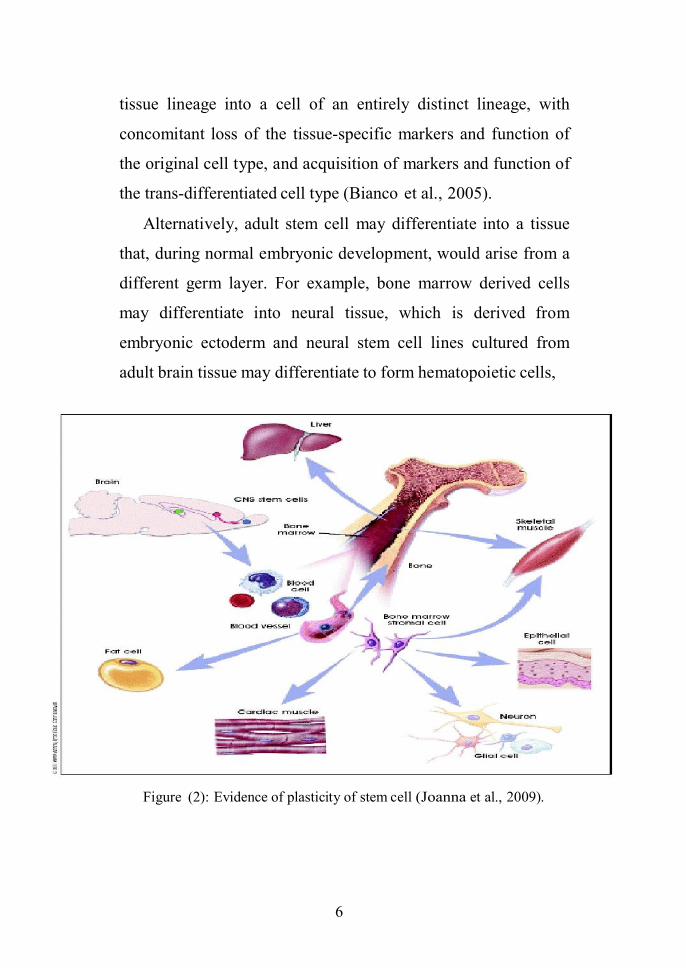
“transdifferentiation” (figure 2) (Joanna et al., 2009).

5
To show that the adult stem cells can generate other cell
types requires them to be tracked in their new environment,
whether it is in vitro or in vivo. In general, this has been
accomplished by obtaining the stem cells from a mouse that has
been genetically engineered to express a molecular tag in all its
cells. It is then necessary to show that the labeled adult stem
cells have adopted key structural and biochemical
characteristics of the new tissue they are claimed to have
generated (Gussoni et al., 2002).
Also it is necessary to demonstrate that the cells can
integrate into their new tissue environment, survive in the tissue,
and function like the mature cells may assume the characteristic
of cells that have developed from the same primary germ layer
or a different germ layer, for example, much plasticity
experiments involve stem cells derived from bone marrow,
which is a mesodermal derivative. The bone marrow stem cells
may then differentiate into another mesodermally derived tissue
such as skeletal muscle, cardiac muscle or liver (Kocher et al.,
2001).
Stem cell lineage differentiation and commitment is
conventionally viewed as progressively
downstream, unidirectional and irreversible. The
notion of unidirectional tissue-lineage commitment of stem cells
is being challenged by evidence of plasticity, or lineage
conversion, in adult stem cells. Mechanisms allowing for
such plasticity include trans- differentiation
which describes the conversion of a cell of one
6
tissue lineage into a cell of an entirely distinct lineage, with
concomitant loss of the tissue-specific markers and function of
the original cell type, and acquisition of markers and function of
the trans-differentiated cell type (Bianco et al., 2005).
Alternatively, adult stem cell may differentiate into a tissue
that, during normal embryonic development, would arise from a
different germ layer. For example, bone marrow derived cells
may differentiate into neural tissue, which is derived from
embryonic ectoderm and neural stem cell lines cultured from
adult brain tissue may differentiate to form hematopoietic cells,
Figure (2): Evidence of plasticity of stem cell (Joanna et al., 2009).

7
or even give rise to many different cell types in embryo. In
both cases cited above, the cells would be deemed to show
plasticity, but in the case of bone-marrow stem cells generating
brain cells, the finding is less predictable (Song et al., 2007).
Alternative mechanisms for explaining apparent stem cell
plasticity involve cell-cell fusion between a stem cell and a
tissue specific cell, the existence of multiple stem cell
populations in one pool of cells, and the ability of the stem cells
to differentiate to a more primitive, less specialized cell lineage,
and then re-differentiate down another lineage (Bongso and
Lee, 2005).
The differentiation potential of stem cells:Many of the terms used to define stem cells depend on the
behavior of the cells in the intact organism (in vivo), under
specific laboratory conditions (in vitro), or after transplantation
in vivo, often to a tissue that is different from the one from
which the stem cells were derived (Joanna et al., 2009).
So they are three classes of stem cells exist: totipotent,
pluripotent multipotent and unipotent.
1) Totipotent:
Totipotency is the ability of a cell to divide and produce all
of the undifferentiated cells within an organism, from the Latin
word totus, meaning entire; For example, the fertilized egg is
said to be totipotent, because it has the potential to generate all
the cells and tissues that make up an embryo and that support its
development in uterus. After fertilization, the cell begins to

8
divide and produce other totipotent cells; these totipotent cells
begin to specialize within a few days after fertilization. The
totipotent cells specialize into pluripotent cells, which they
develop into the tissues of the developing body. Pluripotent
cells can further divide and specialize into multipotent cells,
which produce cells of a particular function (Svendsen and
Ebert, 2008).
Adult mammals, including humans, consist of more than 200
kinds of cells. These include nerve cells (neurons), muscle cells
(myocytes), skin (epithelial) cells, blood cells (erythrocytes,
monocytes, lymphocytes, etc.), bone cells (osteocytes) and
cartilage cells (chondrocytes). Other cells, which are essential
for embryonic development but are not incorporated into the
body of the embryo, include the extraembryonic tissues,
placenta, and umbilical cord. All of these cells are generated
from a single, totipotent cell, the zygote or fertilized egg
(Joanna et al., 2009).
2) Pluripotent:
Pluripotent stem cells can give rise to any type of cell in the
body except those needed to develop a fetus or adult because
they lack the potential to support the extraembryonic tissue
(e.g., the placenta). Most scientist use the term pluripotent to
describe stem cells that can give rise to cells derived from all
three embryonic germ layers (endoderm, mesoderm, and

9
ectoderm). These three germ layers are the embryonic source of
all cells of the body (figure 3) (Svendsen and Ebert, 2008).
Figure (3): Pluripotent stem cells (Svendsen and Ebert, 2008).
The term “pluri” is derived from the Latin word plures,
means several or many. Thus, pluripotent cells have the
potential to give rise to any type of cell, a property observed in
the natural course of embryonic development and under certain
laboratory conditions. Pluripotent stem cells are isolated from
embryos that are only several days old; cells from these stem

10
cell lines can be cultured in the lab and grown without limit
(Sonja et al., 2006).
3) Multipotent:
Multipotent cells, in contrast, can only give rise to a small
number of cell types and they can produce only cells of a
closely related family cell. As haematopiotic stem cells that
differentiate to red blood cells, white blood cells and platelets.
A hematopoietic cell, or a blood stem cell, can develop into
several types of blood cells but cannot develop into liver cells or
other types of cells; the differentiation of the cell is limited in
scope. A multipotent blood cell can produce red and white
blood cells (figure 4) (Svendsen and Ebert, 2008).
Figure (4): Multipotent stem cell (Svendsen and Ebert, 2008).

11
4) Unipotent:
Unipotent stem cells, a term that is usually applied to a cell
in adult organisms, means that the cells in question are capable
of differentiating along only one lineage. The term “uni” is
derived from the Latin word unus, which means one. Also, it
may be that the adult stem cells in many differentiated,
undamaged tissues are typically unipotent and give rise to just
one cell type under normal conditions. This process would
allow for a steady state of self renewal for the tissue. However,
if the tissue becomes damaged and the replacement of multiple
cell types is required, pluripotent stem cells may become
activated to repair the damage (Avasthe et al., 2008).
F igu re (5): Differentiation of human stem cells (Bongso and Lee,
2005).

12
Classification of stem cells according to their sources:
Stem cells can be classified into four broad types based on
their origin, stem cells from embryos; stem cells from the fetus;
stem cells from umbilical cord; and stem cells from the adult.
Each of these can be grouped into subtypes (Andeson et al.,
2001).
1) Embryonic stem cells:In mammals; the fertilized oocyte, zygote, 2-cells, 4-cells, 8-
cells and morula resulting from cleavage of the early embryo
are examples of totipotent cells (ability to form a complete
organism) (figure 6) (Avasthe et al., 2008).

13
Figure (6): Development and differentiation of human tissues (Avasthe
et al., 2008).
The inner cell mass (ICM) of the 5 to 6 days old human
blastocyte is the source of pluripotent embryonic stem cells
(HESCs) and consisting of 50–150 cells (figure 7) (Bongso and
Lee, 2005).

14
Figure (7): Human blastocyst
showing inner cell mass and
trophectoderm (Bongso and Lee,
2005).

15
Figure (8): How human embryonic stem cells are derived? (Bongso
and Lee, 2005).
Characteristics of human embryonic stem cells:
They can maintain undifferentiated phenotype and these
cells are able to renew themselves continuously through many
passages leading to the claim that they are immortal, also these

16
cells are pluripotent, meaning that they are able to create all
three germ layers of the developing embryo and thus they can
develop into each of the more than 200 cell types of the adult
body (figure 9) (Junying et al., 2006).
Figure (9): Characteristics of embryonic stem cells (Junying et al.,
2006).

17
Nearly all research to date has taken place using mouse
embryonic stem cells (MES) or Human embryonic stem cells
(HESCs). Both have the essential stem cell characteristics, yet
they require very different environments in order to maintain an
undifferentiated state. Mouse ES cells are grown on a layer of
gelatin and require the presence of Leukemia Inhibitory Factor
(LIF) (Bongso and Lee, 2005).
HESCs are grown on a feeder layer of mouse embryonic
fibroblasts (MEFs) and require the presence of basic Fibroblast
Growth Factor (bFGF or FGF-2). Without optimal culture
conditions or genetic manipulation, embryonic stem cells will
rapidly differentiate (Avasthe et al., 2008).
Identification of the human embryonic stem cells:
Laboratories that grow human embryonic stem cell lines use
several kinds of tests to identify the human embryonic stem
cells.
These tests include:
1- Growing and sub-culturing the stem cells for many
months. This ensures that the cells are capable of long
term self-renewal. Scientists inspect the cultures through
a microscope to see that the cells look healthy and
remain undifferentiated (Lawrence et al., 2006).
2- Using specific techniques to determine the presence of
surface markers that are found only on undifferentiated
cells. Another important test is for the presence of a

18
protein called oct-4, which undifferentiated cells
typically make. Oct-4 is a transcription factor, meaning
that it helps turn genes on and off at the right time, which
is an important part of the processes of cell
differentiation and embryonic development.
3- Examining the chromosomes under a microscope. This is
a method to assess whether the chromosomes are
damaged or if the number of chromosomes has changed.
It does not detect genetic mutations in the cells.
4- Determining whether the cells can be subculture after
freezing, thawing, replanting (junying et al., 2006).
Differentiation of human embryonic stem cells:
In order to start differentiation, the HESCs must be removed
from the feeder layer and the cell replated and will form
embryoid bodies (Ebs), spherical aggregates in which the
HESCs undergo mixed spontaneous differentiation toward
lineages of all three dermal layers. Another protocol of
differentiation directly without formation of embryoid bodies
stage have resulted in more controlled differentiation and better
yield of the required cells (figure 10) (Joanna et al., 2009).

19
Figure (10): Fluorescent markers can be used to identify stem cells hiddenamong ordinary adult cells. Here, human embryonic stem cells arerecognized by the marker proteins they express (green) (Joanna et al.,2009).
Ethical considerations:The promise of stem cell therapy has ignited public dispute
on the ethics of using aborted embryos for medical purposes.
Individual attitudes are usually influenced by religious and
liberal views but also by concerns that the practice of embryonic
tissue transplantation will increase the pressure to perform
abortions and create a black market in which pregnancy and
aborted tissues will be sold to the highest bidder. The regulated
banking of stem cell lines may solve some of the ethical issues.
As in other cases in which medical and scientific advances
found society without the means to deal with their ethical, legal,
and social consequences, it is important to discuss these issues
in public, with the active participation of the medical and
scientific community (Christopher, 2008).

20
2) Fetal stem cells:The identification of human fetal stem cells has raised the
possibility of using autologus cells for in utero treatments. The
human fetal stem cells population extracted from fetal blood
contains adherent cells that divide in culture for 20 to 40
passages and can differentiate into mesenchymal lineages
including bone and cartilage, but also have the ability to form
oligodendrocytes and hematopoiotic cells. These cells, which
can be found circulating only during the first trimester, are
similar to hematopoiotic populations in fetal liver and bone
marrow (Avasthe et al., 2008).
3) Umbilical cord stem cells:
These are cells harvested from the cord blood. Cord blood is
rich in the stem cells and after appropriate human leukocyte
antigen [HLA] matching may be used to treat a variety of
conditions. Characteristics of these cells are identical to adult
stem cells except that they are not derived from adults and that
their concentration is far more in umbilical blood as compared
to adults. The use of umbilical cord stem cells in orthopedics is
still in a nascent stage and most studies currently focus on the
use of the stem cell (Crosta, 2010).
4) Adult stem cell:It is an undifferentiated cell that is found in a differentiated
tissue, it can renew itself and become specialized to yield all the

21
specialized cell types of the tissue from which it originated.
Adult stem cells, like all stem cells, share at least two
characteristics. First, they can make identical copies of
themselves for long period of time; this ability to proliferate is
referred to as long term self renewal. Second, they can give rise
to mature cell types that have characteristic morphologies
(shapes) and specialized functions (Charron et al., 2009).
Typically, stem cells generate an intermediate cell type or
types before they achieve their fully differentiated state. The
intermediate cell is called a precursor cells in fetal or adult
tissues are partially differentiated cells that divide and give rise
to differentiated cells. Such cells are usually regarded as
“committed” to differentiating along a particular cellular
development pathway, although this characteristic may not be as
definitive as once thought (Bianco et al., 2005).
Adult stem cells are rare. Their primary functions are to
maintain the steady state functioning of a cell, called
(homeostasis) and with limitation to replace cells that die due to
injury or disease. For example, only an estimated 1 in 10,000 to
15,000 cells in the bone marrow is a hematopoietic (blood-
forming) stem cell (HSC). Furthermore, adult stem cells are
dispersed in tissues throughout the nature of animal and behave
very differently, depending on their local environment. For
example, HSCs are constantly being generated in the bone
marrow where they differentiate into mature types of blood

22
cells. Indeed, the primary role of HSCs is to replace blood cells
(Abdallah and Kassem 2008).
Unlike embryonic stem cells, which are defined by their
origin (the inner cell mass of the blastocyte), adult stem cells
share no such definitive means of characterization. In fact, no
one knows the origin of adult stem cells in any mature tissue.
Some have proposed that stem cells are somehow set aside
during fetal development and restrained from differentiating.
Definition of adult stem cells vary in the scientific literature
range from a simple description of the cells to a rigorous set of
experimental criteria that must be met before characterizing a
particular cell as an adult stem cell. Most of the information
about adult stem cells comes from studies of mice. The list of
adult tissues reported to contain stem cells is growing and
includes bone marrow, peripheral blood, brain, spinal cord,
dental pulp, blood vessels, skeletal muscle, epithelia of skin and
digestive system, cornea, retina, liver, and pancreas
(Christopher, 2008).
Ideally, adult stem cells should also be clonogenic. In other
words, a single adult stem cell should be able to generate a line
of genetically identical cells, which then gives rise to all the
appropriate, differentiated cell types of the tissue in which it
resides. Again, this property is difficult to demonstrate in vivo;
in practice, scientists show either that a stem cell is clonogenic
in vitro, or that a purified population of candidate stem cells can
repopulate the tissue (Avasthe et al., 2008).

23
Sources of adult stem cells:(I) Bone Marrow-Derived Stem/Progenitor Cells:
Adult bone marrow-derived stem cells are presently the cell
types most widely used in stem cell therapy. A heterogeneous
subset there of, termed autologous bone marrow-derived
mononuclear cells (ABMMNCs), comprises the following types
of stem cells, (Mesenchymal stem cells, Hematopoietic stem
cells and Endothelial progenitor cells), that have potential
therapeutic uses (figure 11) (Svendsen and Ebert, 2008).
Figure (11): Some of the known sources of adult stem cells (Svendsen andEbert, 2008).

24
(a) Mesenchymal stem cells (MSCs):
MSCs are a proper stem cell which can be greatly and
efficiently expanded in culture and can differentiate to several
specific mesenchymal cell lineages. Mesenchymal (Stromal)
stem cells (MSCs) are found in various niches of adult tissue.
MSCs are rare in bone marrow (<0.01% of nucleated cells, by
some estimates) and 10 times less abundant than hematopoietic
progenitor cells but MSCs can be readily grown in culture.
However, more recently, other sources of MSCs have been
described including placenta, adipose tissue, cord blood and
liver (junying et al., 2006).
The human Mesenchymal stem cells (HMSCs) from bone
marrow can be cloned and expanded in vitro more than 1
million-fold and retain the ability to differentiate to several
mesenchymal lineages. Researchers have not yet found
conditions that allow continuous, indefinite HMSC growth, yet
it is possible to produce billions of MSCs in vitro for cellular
therapy from a modest bone marrow aspirate drawn through the
skin. MSCs need to be expanded ex vivo because they
apparently are very contact inhibited, and there is little evidence
of in vivo expansion as MSCs labeled with membrane dyes, that
would be diluted and undetected from dividing cells after about
3 divisions, are found months later even in repairing tissue
(Sottile et al., 2002).

25
Advantages of Mesenchymal Stem Cells:
Ease of isolation, high expansion potential, genetic stability,
reproducible characteristics in widely dispersed laboratories,
compatibility with tissue engineering principles and potential to
enhance repair in many vital tissues. There they may be the
current preferred stem cells model for cellular therapeutic
development (Diana and Gabriel, 2008).
Biology of mesenchymal stem cells (MSCs):
The anatomical locations of phenotype of MSCs have no yet
been well defined in vivo. Some have used expression of Stro-1
and VCAM-1 to analyses putative MSC in vivo in human. A
general consensus among researchers in the field is that MSC
can be successfully defined based on staining with surface
markers such as CD44, CD90, CD73, CD105 and CD166.
However, none of these antigens are unique to MSC. Using
markers such as Stro-1 (human) and Sca-1 (mouse), several
reports indicate that MSC reside adjacent to endothelium in the
bone marrow and possibly other tissues (Zannettino et al.,
2007).
(b) Hematopoietic stem cells (HSCs):
HSCs are presented in umbilical cord blood with a frequency
of just under one in 1 million mononuclear cells (one in 3
million MNCs) or mobilized peripheral blood (one in 6 million
MNCs). They are capable of unlimited cell proliferation in bone
marrow and must undergo at least 20 to 23 divisions on their

26
way to produce mature blood cells, even assuming no cell death
along the way (Emerson et al., 2008).
Biology of heamtopiotic stem cells:
Much effort has been focused on discovering cell surface
markers that can identify those cells that have true functional
stem cell properties. Perhaps clinically most familiar is CD34, a
glycoprotein present on the cell surface of stem and progenitor
cells which is used to enrich stem cells mobilization and
collection for HSCs, but even within the CD34+ population,
only a small percentage are HSCs (Emerson et al., 2008).
For decade scientists and hematologists have struggled with
the difficulty that HSCs cannot be purified based on
phenotypical characteristics and perhaps more importantly,
cannot be expanded and cloned ex vivo. Recent evidence has
emerged suggesting that HSCs can be expanded ex vivo. But
there is still no evidence to support the idea of clonality. For
these reasons HSCs are not ideally suited for in vitro
experiments designed to test plasticity. In this regard HSCs
differ dramatically from MSCs in bone marrow and neural stem
cells (NSCs) in the central nervous system, both of which can
be clonally derived and tested for multiple differentiation
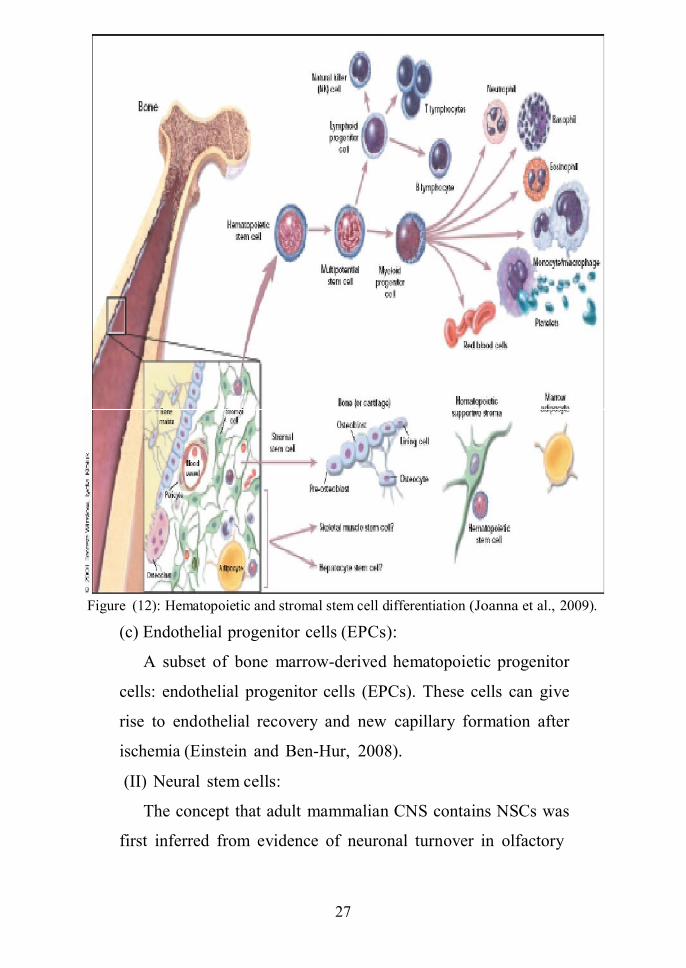
pathways (figure 12) (Joanna et., 2009).
27
Figure (12): Hematopoietic and stromal stem cell differentiation (Joanna et al., 2009).
(c) Endothelial progenitor cells (EPCs):
A subset of bone marrow-derived hematopoietic progenitor
cells: endothelial progenitor cells (EPCs). These cells can give
rise to endothelial recovery and new capillary formation after
ischemia (Einstein and Ben-Hur, 2008).
(II) Neural stem cells:
The concept that adult mammalian CNS contains NSCs was
first inferred from evidence of neuronal turnover in olfactory

28
bulb and hippocampus in the adult organism. The multipotency
of NSCs was demonstrated in vitro in 1990 by their ability to
differentiate into neurons, astrocytes, and oligodendrocytes as
well as various forms of neural precursors. In addition, in vivo
delivery of these cells to animal models of neurodegenerative
diseases was associated with varying degrees of functional
recovery. Currently, there is no set of markers or protein
expression profiles that precisely define and fully characterize
undifferentiated NSCs. Neural stem cells (NSCs) and neural
precursor cells (NPCs) can be isolated from the developing or
adult CNS and can be safely expanded in chemically defined
culture media for an extended (Song et al., 2007).
(a) Adult neural precursor cells (NPCs):
New neurons are derived in adulthood from a population of
adult NPCs, which are primarily found in the subependymal
layer of the ventricular zone and the dentate gyrus of the
hippocampus, although they are also probably found in other
sites. However, the behavior of the neural precursor cells
(NPCs) found in all these sites is different, and may relate as
much to the environment in which they find themselves as to
their intrinsic properties, eg; nigral NPCs appear to only
differentiate into astrocytes in situ or when grafted to the adult
nigra, but when they are cultured in vitro or transplanted into the
hippocampus they can form neurons (Gronthos et al., 2003).

29
Properties of neural stem cells:
(1) Immunosupressive effect of NSCs:
Although NSCs may exert their therapeutic effects by
directly replacing missing cells, transplantation rarely results in
significant numbers of transplant-derived terminally
differentiated neurons. The beneficial effect of NSCs in disease
models may be attributable to alternative biologic properties.
The first indication of an anti-inflammatory effect of NPCs
came from transplantation experiments in rats with experimental
autoimmune encephalomyelitis (EAE). It was shown
transplantation of NPCs reduced brain inflammation and clinical
disease severity, it was suggested that the benefit of NPC
transplantation was mediated by an anti-inflammatory effect
(Raisman and Li, 2007).
The exact mechanisms by which transplanted NPCs
attenuate brain inflammation are unclear. Some suggests an
immunomodulatory effect by which NPCs promote apoptosis of
type 1 T-helper cells, shifting the inflammatory process in the
brain toward a more favorable climate of dominant type 2 T-
helper cells. Alternatively, a nonspecific bystander
immunosuppressive effect of NPCs on T-cell activation and
proliferation has been suggested. The suppressive effect of
NPCs on T cells was accompanied by a significant suppression
of pro-inflammatory cytokines. This nonspecific anti-
inflammatory mechanism may be of major importance in the
application of transplantation therapy in immune-mediated

30
diseases because it can protect the host CNS and graft from
additional immune attacks (Einstein and Ben-Hur, 2008).
(2) Neuroprotictive effects of transplanted NSCs:
Neuroprotective effect was observed in other non
autoimmune experimental disease models. Neural stem cells
rescued dopaminergic neurons of the mesostriatal system in a
Parkinson disease (PD) model in rodents. These findings led to
the concept that NSCs are endowed with inherent mechanisms
for rescuing dysfunctional neurons. This effect was found to be
important in other neurologic diseases. Neural stem cells seeded
on a synthetic biodegradable scaffold and grafted into the hemi-
sectioned adult rat spinal cord induced significant improvement
in animal movement by reduction of necrosis in the surrounding
parenchyma and by prevention of inflammation, glial scar
formation, and extensive secondary cell loss (Einstein and Ben-
Hur, 2008).
(3) Neurotrophic effects of transplanted NSCs:
After sectioning of the adult spinal cord, NSC
transplantation induced a permissive environment for axonal
regeneration. Similarly, in a model of retinal degeneration, NPC
transplantation promoted neural growth in the optic nerve. In
both cases, this effect was mediated by induction of matrix
metalloproteinases that degrade the impeding extracellular
matrix and cell surface molecules, enabling axons to extend
through the glial scar. Transplantation of olfactory-ensheathing
cells into the sectioned spinal cord also promoted axonal

31
regeneration in long fiber tracts, with a return of lost function.
This was explained by the creation of proper realignment,
enabling axonal growth through a permissive tract. In addition,
the cells increased axonal sprouting, remyelination, and
vascularization of the injured spinal cord (Raisman and Li,
2007).
Isolation of human NSCs:
To date, they are primary isolated and propagated in vitro as
cells that form free-floating neurospheres when cultured in
serum-free medium on non adherent surfaces in the presence of
mitogenic factors such as basic FGF or FGF-2 and epidermal
growth factors, although there have also been reports of
monolayer cultures (McBride et al., 2004).
(III) Pancreatic stem/progenitor cells:
There is strong evidence that new pancreatic islets can
derive from progenitor cells present within the ducts and islets,
in a process called “neogenesis”. Furthermore, when these
pseudo-islets were transplanted into non-obese diabetic (NOD)
mice, diabetes reversal was observed. Candidate pancreatic
stem/progenitor cells have also been described within acini, but
contamination with endocrine and ductal cells in cultures could
not be excluded in these experiments (Limmbert et al., 2008).
The isolation of a distinct stem/progenitor cell within the
endocrine pancreas depends on the identification of a specific
progenitor marker. The exciting observation that nestin positive
islets cells display endocrine differentiating capacity led to the

32
hypothesis that this intracytoplasmic filament protein might
correspond to a pancreatic stem/progenitor cell marker. More
recently, in two important studies a population of cells in the
developing and adult mouse pancreas was identified, which
under differentiation conditions, released insulin in a glucose-
dependant manner. After differentiation, these cells expressed
specific developmental pancreatic endocrine genes (e.g. Ngn3,
Pax-4, Pax-6 and PDX-1) and contamination with mature beta
cells was ruled out (Limbert et al., 2008).
While mature beta cell replication appears to be major
physiological beta-cell regenerative process, identification of
pancreatic cells with progenitor features might open an
important and promising strategy for cell replacement and
regeneration therapy. Anyhow, to be clinically relevant, in vitro
proliferation of progenitor cells from human pancreas must
produce large amounts of cells, in order to allow cells isolated
from one single donor to be sufficient to treat a given diabetic
patient. It would be even better to have one single donor for
several diabetics. For these reasons, acinar isolated
stem/progenitor cells might be of interest, considering that
exocrine tissue constitutes 90% of pancreatic tissue and is
discarded during islet isolation (Kushner et al., 2005).
(IV) Other sites:
First identified in human bone marrow, a population of
mesenchymal progenitor/stem cells (MSC) with well
characterized immunophenotype and distinct from
33
hematopoietic stem cells, was shown to possess a high
proliferation rate and great plasticity. Under specific culture
conditions these cells differentiate into mesenchymal tissues,
such as bone, cartilage, muscle, tendon, adipose and stroma, as
well as neuronectodermal tissues (Limbert et al., 2008).
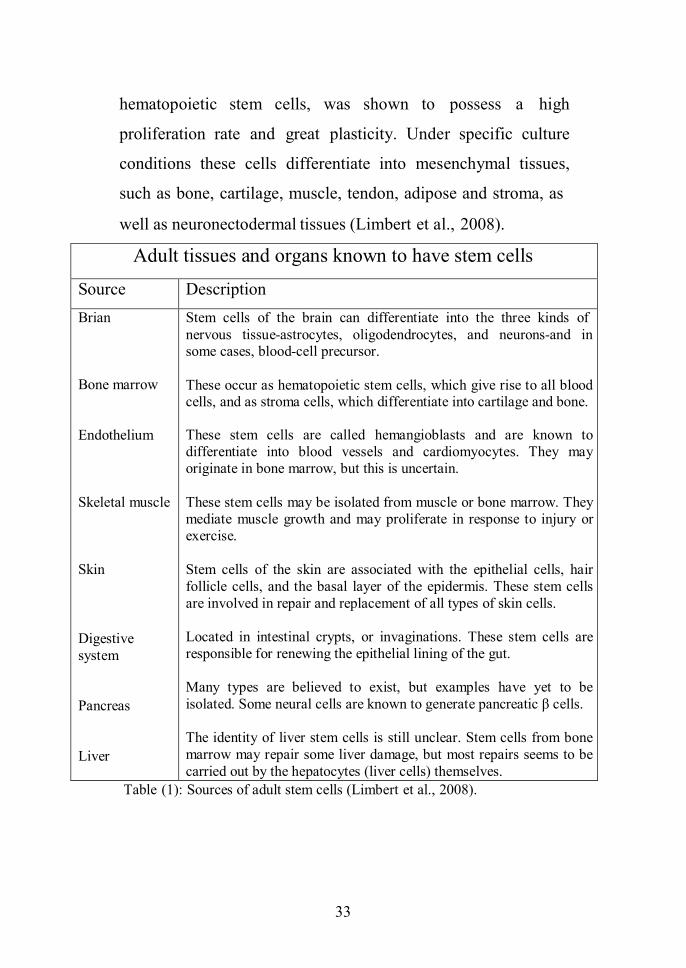
Adult tissues and organs known to have stem cells
Source DescriptionBrian
Bone marrow
Endothelium
Skeletal muscle
Skin
Digestivesystem
Pancreas
Liver
Stem cells of the brain can differentiate into the three kinds ofnervous tissue-astrocytes, oligodendrocytes, and neurons-and insome cases, blood-cell precursor.
These occur as hematopoietic stem cells, which give rise to all bloodcells, and as stroma cells, which differentiate into cartilage and bone.
These stem cells are called hemangioblasts and are known todifferentiate into blood vessels and cardiomyocytes. They mayoriginate in bone marrow, but this is uncertain.
These stem cells may be isolated from muscle or bone marrow. Theymediate muscle growth and may proliferate in response to injury orexercise.
Stem cells of the skin are associated with the epithelial cells, hairfollicle cells, and the basal layer of the epidermis. These stem cellsare involved in repair and replacement of all types of skin cells.
Located in intestinal crypts, or invaginations. These stem cells areresponsible for renewing the epithelial lining of the gut.
Many types are believed to exist, but examples have yet to beisolated. Some neural cells are known to generate pancreatic β cells.
The identity of liver stem cells is still unclear. Stem cells from bonemarrow may repair some liver damage, but most repairs seems to becarried out by the hepatocytes (liver cells) themselves.
Table (1): Sources of adult stem cells (Limbert et al., 2008).

34
Identification of the adult stem cells:
The scientists often use one or more of the following three
methods to identify and test adult stem cells:
1- Labeling the cells in a living tissue with molecular markers
and then determining the specialized cell types they
generate. Then
2- Removing the cells from living animals, labeling them in
cell culture, and transplanting them back into another animal
to determine whether the cells repopulate their tissue of
origin. Then
3- Isolating the cells, growing them in cell culture, and
manipulating them, often by adding growth factors or
introducing new genes, to determine what differentiated
cells types they can become (Raisman and Li, 2007).
The similarities and differences between embryonic and adult
stem cells:
The adult and embryonic stem cells differ in the number and
types of differentiated cells types they can become. Embryonic
stem cells can become all cell types of the body because are
pluripotent. Adult stem cells are generally limited to
differentiating into different cell types of their tissue of origin.
However, some evidence suggests that adult stem cell plasticity
may exist; increasing the number of cell types a given adult
stem cell can become (figure 13). Large numbers of embryonic
stem cells can be relatively easily grown in culture, while adult
stem cells are rare in mature tissues and methods for expanding

35
their numbers in cell culture have not yet been worked out. This
is an important distinction, as a large number of cells are needed
for stem cell replacement therapies (Limbert et al., 2008).
A potential advantage of using stem cells from an adult is
that the patient’s own cells could be expanded in culture and
then reintroduced into the patient. The use of patient’s own
adult stem cells would means that the cell would not be rejected
by the immune system. This represents a significant advantage
as immune rejection is a difficult problem that can only be
circumvented with immunosuppressive drugs. Embryonic stem
cells from a donor introduced into a patient could cause
transplant rejection, however, whether the recipient would reject
donor embryonic stem cells has not been determined in human
experiments (Sonja et al., 2006).
Figure (13): Sources of stem cells (Limbert et al., 2008).

36
Types of Stem Cell transplantation:
Stem cell transplantation can be classified according to the
genetic relation between the donor and recipient into 4 classes:
1- Autograft: In which the donor and recipient is the same
individual.
2- Isograft or syngenic graft: In which the donor and
recipient are genetically identical (e.g., monozygotic
twins).
3- Allograft or homograft: In which the donor and recipient
are genetically unrelated but belong to the same species.
4- Xenograft or heterograft: In which the donor and
recipient belong to different species (David, 2009).
Application of stem cells:
1) Basic science application:
Stem cells are ideally suited to allow for the study of complex
processes that direct early unspecialized cells to differentiate
and develop into the more than two hundred cell types in the
human body (Bianco et al., 2005).
2) Medical research applications:
Stem cell studies may allow researchers to follow the processes
by which diseases and impairments caused by genetic
abnormalities first manifest themselves biochemical or
structurally in cells and tissues. Using stem cells to produce
large numbers of genetically uniform cultures of organ tissues
for example, liver, muscle, or neural would allow controlled
comparison of the effects of drugs or chemical on these tissues.

37
Alternatively, testing drugs against stem cell tissues varying
genetic makeup could allow tissue specific stem cell may
provide a constant in vitro source of such cellular material
(Bianco et al., 2005).
The site of stem cell implantation:
The transplantation can be described as orthotropic or
heterotropic:
1- Neurologic transplantation: Refers to donor tissue
implantation in the anatomically correct position in the
recipient.
2- Heterotropic transplantation: Refers to the relocation of
the implant in the recipient at a site different from the
normal anatomy (David, 2009).
Route of stem cell delivery:
Reports have indicated that after stereotactic
intraparenchymal, intracerebro-ventricular, intravenous and
intraarterial transplantation, stem cells can home to sites of
injury in the CNS and induce functional recovery. Of these
various transplantation techniques, those that depend on
intravascular delivery of stem cells for stroke are particularly
attractive.
Intravascular delivery:
In addition to its minimal invasive nature, intravascular
delivery may allow stem cells to have a superior interaction
with injured tissue. A comparative study revealed that direct
intracerebral transplantations resulted in the largest number of

38
cells at the lesion site, followed by intracerebro-ventricular and
intravenous transplantations (Guzman et al., 2008).
However, researchers in that study only assessed the
absolute number of cells in the perilesional area and took no
account of whether these cells were therapeutically distributing
to all injured areas of brain parenchyma on a microscopic level.
Many believe that intravascular delivery of stem cells may lead
to a wider distribution of cells around the lesion as compared
with focal perilesional transplants, thereby leading to superior
stem cell–injured tissue interactions (Xiao et al., 2007).
Mechanism of wide distribution:
The cells travel in the blood stream and follow a chemo
attractant gradient generated by inflammation in the injured
brain. Unfortunately, intravenously delivered cells pass through
the systemic and pulmonary circulation systems and home to
other organs as well, which significantly reduces cell homing to
the injured brain. Intravenous injection of human MSCs into
rats 24 hours after stroke showed that only 4% of the cells
entered the brain, the number of cells entering the brain
increased over time and peaked at Day 21 post-stroke. At Day
56, 60% of these surviving cells differentiated into glia, and
20% into neurons. Despite the fact that the number of cells
entering the brain was limited, functional recovery was
enhanced by intravenous delivery (Pluchino et al., 2005).
39
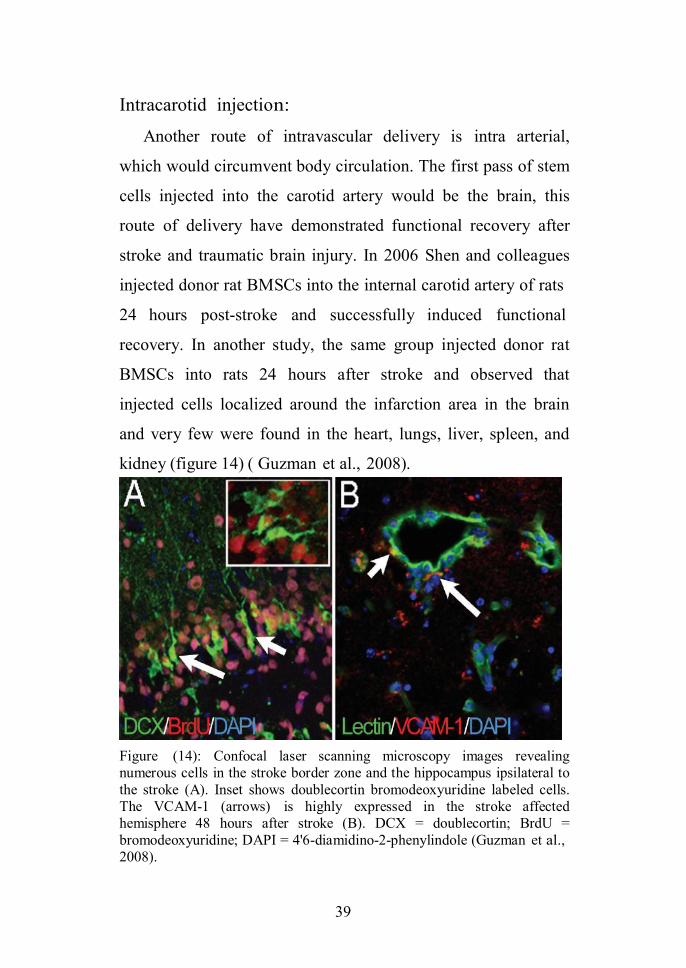
Intracarotid injection:
Another route of intravascular delivery is intra arterial,
which would circumvent body circulation. The first pass of stem
cells injected into the carotid artery would be the brain, this
route of delivery have demonstrated functional recovery after
stroke and traumatic brain injury. In 2006 Shen and colleagues
injected donor rat BMSCs into the internal carotid artery of rats
24 hours post-stroke and successfully induced functional
recovery. In another study, the same group injected donor rat
BMSCs into rats 24 hours after stroke and observed that
injected cells localized around the infarction area in the brain
and very few were found in the heart, lungs, liver, spleen, and
kidney (figure 14) ( Guzman et al., 2008).
Figure (14): Confocal laser scanning microscopy images revealingnumerous cells in the stroke border zone and the hippocampus ipsilateral tothe stroke (A). Inset shows doublecortin bromodeoxyuridine labeled cells.The VCAM-1 (arrows) is highly expressed in the stroke affectedhemisphere 48 hours after stroke (B). DCX = doublecortin; BrdU =bromodeoxyuridine; DAPI = 4'6-diamidino-2-phenylindole (Guzman et al.,2008).

40
The debate over the best delivery route is further
complicated by the fact that there is still a great deal of
controversy concerning the mechanism by which stem cells lead
to enhanced functional recovery in patients who have
experienced stroke. The 2 most discussed mechanisms are as
follows: 1) cellular replacement, by way of the functional
integration of stem cells; and 2) secretion of neurotrophic and
angiogenic factors. If the mechanism of recovery is cellular
replacement, then transendothelial migration is necessary and
the methods that allow the highest concentrations of stem cells
in the injured brain areas ought to be pursued; however, there is
significant evidence that stem cells may provide their benefits
by secreting various neuroprotective factors (Guzman et al.,
2008).
In summary, the best route of human stem cell delivery has
not been determined, but the intravascular route is particularly
attractive because of its ease of administration, minimal
invasiveness, and potential for widespread cell distribution
together with widespread secretion of neuroprotective,
proangiogenic, and immunomodulatory factors. Intuitively, the
intraarterial route of delivery seems better than the intravenous,
given that injected cells first pass the target organ that is, the
brain prior to being redistributed in the systemic circulation
(Pluchino et al., 2005).

41
Timing of transplantation
Undoubtedly the fate and function of transplanted cells will
depend on any or all of these alterations, and the optimal time of
transplantation is unknown. The timing of transplantation
depends mainly on the goal of treatment, for example,
neuroprotection, which should happen early after the insult, or
neuroregeneration/cell replacement, which can be done once a
lesion has stabilized. We can envision a future in which we will
rely on multimodal stem cell treatment, depending on a
combination of early and late administrations of different cell
types (Guzman et al., 2008).
Early intravascular cell delivery
In animal models with a neuro inflammatory component
such as stroke, traumatic brain injury, spinal cord injury, and
multiple sclerosis, therapeutic somatic stem cells (for example,
BMSCs, umbilical cord blood stem cells, MSCs, and NPCs)
target inflamed CNS areas where they persist for months and
promote recovery through neuroprotective mechanisms. It is
thought that the process of transendothelial migration of somatic
stem cells may be regulated in a manner similar to that of
inflammatory cells. As early as 30 minutes after stroke, the
infiltration of leukocytes, both polymorphonuclear leukocytes
and monocytes/macrophages, can be observed (Goldman and
Windrem, 2006).
Chemoattraction, adhesion, and transendothelial migration
of inflammatory cells is regulated by specific inflammatory

42
mediators, which have been identified in experimental and
human stroke. The temporal expression profile of adhesion
molecules, cytokines, and chemokines after stroke has been well
described. Vascular cell adhesion molecule–1 (VCAM-1) has
been shown to reach a peak level 24 hours after experimental
stroke. At the bedside, soluble VCAM-1 concentration in
plasma is increased in patients with acute stroke. Intercellular
adhesion molecule–1 levels have been elevated as early as 4
hours after stroke with sustained levels for up to 1 week (Zhang
and Lodish, 2005).
Monocyte chemo-attractant protein–1, a key chemoattractant
factor for the recruitment of circulating peripheral cells to the
stroke area and an important factor for stem cell migration, is
upregulated 3 days after stroke and then returns to baseline after
1 week. Similarly, stromal-derived factor–1(SDF-1) is known to
be a potent chemoattractant for inflammatory as well as stem
cells (including BMSCs and NSCs) and is expressed early after
stroke. Anatomically, adhesion molecule upregulation as well as
chemokine expression has been shown to be highest in the
stroke-affected penumbral region. Blocking the different
pathways of chemoattraction and cell adhesion in stroke-
affected rodents reduced the number of infiltrating
inflammatory cells (Belmadani et al., 2006).
In mice lacking intercellular adhesion molecule–1 (ICAM-
1), a significant reduction in inflammatory cellular infiltrate and
a reduction in lesion size were noted. Treatment with anti–

43
ICAM-1 antibodies was a successful neuroprotective means of
reducing lesion size and apoptosis in experimental stroke.
However, a clinical trial exploring the feasibility of using an
ICAM-1 blocking antibody failed to demonstrate any beneficial
effects in the patients. There is some evidence that
intravascularly administered stem cells undergo the same
process as inflammatory cells, including chemoattraction,
adhesion, and transendo-thelial migration after stroke,
potentially making this route an ideal way of cell delivery (Hill
et al., 2004).
Late intraparenchymal cell transplantation
In contrast to the acute intravascular cell treatment, the
intraparenchymal approach has been hindered by poor outcomes
if the stem cells are transplanted too early after stroke.
Excitotoxicity, oxidative stress, and inflammation post ischemia
make the ischemic brain a hostile environment for
intracerebrally transplanted cells. In fact, we have found a
negative correlation between graft survival and inflammation.
Additionally, we demonstrated that human NSCs transplanted
too close or into the stroke area have very limited survival at
days after stroke (Belmadani et al., 2006).
Transplanting cells 3 weeks after stroke, when there is a
significant decrease in inflammation, led to greater graft
survival than transplanting 5–7 days after stroke. Taken
together, early intravascular cell therapy might benefit from the
processes tied to post-stroke inflammation but might be

44
detrimental to cells directly transplanted intra-parenchymally.
Therefore, intra-parenchymal cell replacement therapy might be
useful as a second line or delayed stem cell treatment strategy
(Grabowski, 2010).

45
Stem cell therapy in Parkinsonism
Parkinson’s disease (PD) otherwise known as ‘’paralysis
agitans’’ or ‘’shaking palsy’’ was classically described by James
Parkinson in 1817. His description of “Involuntary tremulous
motion with lessened muscular power, in parts not in action and
even when supported, with a propensity to bend the trunk
forward and to pass from a walking to a running pace, the
senses and intellect being uninjured” has stood the test of time.
PD is also defined as a debilitating neurodegenerative disorder
of insidious onset in middle or late age characterized by the
selective loss of nigrostriatal dopaminergic neurons and loss of
dopamine in the striatum (Abayomi, 2002).
Parkinson’s disease is second only to Alzheimer’s disease
with a prevalence of 1 in 10,000. Although it is uncommon in
people under age 40 years, the incidence of PD greatly increases
with age, affecting approximately 1% of individuals older than
60 years (Lane et al., 2008).
Pathology:
The basic pathology is cell degeneration and loss of
pigmented neurons in the pars compacta of the substantia nigra
and locus ceruleus with atrophy and glial scarring. The
degenerated pigmented neurons contain Lewy bodies which are
intracytoplasmic eosinophilic hyaline inclusions composed of
protein filaments (ubiquitin & synuclein), and do not have the
electronic microscropic appearance of any known viral or

46
infective agent (fig. 15). Lewy bodies are characteristic of
Parkinson’s disease except in post-encephalitic Parkinson’s and
parkengene mutants. However, they could be found in 4% of
brain without parkinsonian features and these are likely cases of
subclinical Parkinson’s as 80% of the zona compacta cells must
degenerate before clinical symptoms become apparent
(Abayomi, 2002).
The pars compacta contains 450,000 dopaminergic neurons.
With the loss of dopaminergic neurons at those sites, there is
deficiency of dopamine in the basal ganglia, chiefly the striatum
(caudate nucleus and putamen). Furthermore, the enzymes
required for dopamine synthesis, DOPA decarboxylase and the
rate limiting enzyme tyrosine hydroxylase are reduced. In
addition, there is deficiency of neurotropic factors such as glial
and brain derived neurotrophic factors. However, neurons in the
striatum with dopamine receptors remain intact and are
responsible for the therapeutic effects of levodopa. In the
Parkinsonism unresponsive to levodopa, striatal neurons are
degenerated. Genetic and environmental factors are important in
the mechanism of neuronal deaths due to neuronal necrosis or
apoptosis. In neuronal necrosis there is disintegration of cell and
organelles and subsequent removal by phagocytic and
inflammatory response with increased cellular permeability. In
apoptosis on the other hand, there is rapid programmed cell
death in response to a toxic stimuli. There is chromatin
condensation, DNA fragmentation and cell shrinkage, with

47
relative sparing of organelles without inflammatory changes or
increased cellular permeability (Golbe, 2003).
Among the factors that have been implicated in neuronal
degeneration in Parkinson’s disease are mitochondrial
dysfunction, oxidative stress, and the actions of excitotoxins,
deficient neurotrophic support and immune mechanisms. HLA-
DR positive reactive microglial cells and cytokines such as
interleukin 1 (IL-1) and tumor necrosis factor-a play significant
role in the pathogenesis of Parkinson’s disease. Oxidative stress
with excess reactive oxygen species and free radical damage
involving one or more unpaired electrons react with nucleic
acids, proteins and lipids, this metabolic derangement results in
generation of toxic byproducts and increased oxidative stress
with resultant cellular damage (figure 15) (Lane et al., 2008).

48
Figure (15): Neuronal Pathways that degenerate in Parkinson's disease.Signals that control body movements travel along neurons that project fromthe substantia nigra to the caudate nucleus and putamen (collectively calledthe striatum) (Lane et al., 2008).
Cell replacement therapy:
The idea of growing dopamine cells in the laboratory to treat
Parkinson’s is the most recent step in the long history of cell or
tissue transplantation to reverse this devastating disease. The
concept was, and still is, straightforward: implant cells into the
brain that can replace the lost dopamine releasing neurons.
Although conceptually straight forward, this is not an easy task.
Fully developed and differentiated dopamine neurons do not
survive transplantation, so direct transplantation of fully
developed brain tissue from cadavers, for example, is not an

49
option. Moreover, full functional recovery depends on more
than cell survival and dopamine release; transplanted cells must
also make appropriate connections with their normal target
neurons in the striatum (Lindvall and Kokaia, 2010).
Under the basic principle of restoring dopamine producing
neurons via neural grafts, extensive studies have been done to
bring this to fruition. One of the first attempts at using cell
transplantation in humans was tried in the 1980s. This surgical
approach involved the transplantation of dopamine producing
cells found in the adrenal glands, which sit atop the kidneys in
the abdomen dramatic improvement in Parkinson’s patients by
transplanting dopamine producing chromaffin cells from several
patients’ own adrenal glands to the nigrostriatal area of their
brains; it showed dramatic improvement in Parkinson’s patients.
Another strategy was previously attempted in the 1970s, in
which cells derived from fetal tissue from the mouse substantia
nigra was transplanted into the adult rat eye and found to
develop into mature dopamine neurons (Panchision, 2006).
The functional integration of dopamine neuron grafts prove
the efficacy of the cell replacement principle, but in reality, this
clinical outcome is extremely inconsistent with respect to the
percentage of cells that survive the grafting procedure and the
amount of dopamine produced by the new neurons. In fact,
average functional improvement of patients in the experiments
only rises about 20%. Across the board, subjects achieve
functioning levels less than or equal to that of patients

50
undergoing deep brain stimulation, which carries a lower
morbidity risk (Lane et al., 2008).
Cell transplantation:
Transplantation of primary ventral mesencephalic tissue into
the striatum aims to restore brain circuitry and function lost as a
result of PD. The main objective of primary tissue
transplantation has been to provide proof of principle that
grafted dopaminergic neurons can i) survive and restore
regulated dopamine release, ii) integrate with the host brain to
reinstate frontal cortical connections and activation, and iii) lead
to measurable clinical benefits together with improved quality
of life. Preclinical work in animal models of PD has shown that
grafted dopaminergic neurons, extracted from the developing
ventral mesencephalon (VM) can survive, reinnervate the
lesioned striatum, and improve motor function (Winkler et al
2005).
Over the past two decades, a series of open label clinical
trials have provided convincing evidence to show that human
embryonic nigral neurons taken at a stage of development when
they are committed to a dopaminergic phenotype can survive,
integrate and function over a long time in the human brain.
There is good evidence of graft survival, with grafted neurons
developing afferent and efferent projections with the host
neurons. Long term survival of dopaminergic grafts is possible
up to 10 years after transplantation, and there have been no

51
reported cases of overt immunorejection even after several years
of withdrawal from immuno-suppression (Olanow et al 2003).
Evidence from Positron Emission Tomography (PET)
scanning has revealed significant increases in activation in the
areas reinnervated by the grafted cells, and longitudinal clinical
assessments indicate significant functional recovery for motor
control, in some cases for more than 10 years, in the most
successful cases, patients have either reduced dependency for or
completely withdrawn from L-dopa treatment. Post mortem
studies similarly show good survival of transplanted neurons
and well integrated grafts (figure 16) (Winkler et al., 2005).
Figure (16): Dopamine Neuron Transplantation: PET images from aParkinson’s patient before and after fetal tissue transplantation, the imagetaken before surgery (left) shows uptake of a radioactive form of dopamine(red) only in the caudate nucleus, indicating that dopamine neurons havedegenerated. Twelve months after surgery, an image from the same patient(right) reveals increased dopamine function, especially in the putamen(Winkler et al., 2005).

52
The precise mechanism responsible for these dyskinesia
remains unknown but it does not appear to be related to graft
overgrowth resulting in excessive dopamine release. One
possibility surrounds the quality of dissected tissue. Successful
trials have used either freshly dissected tissue or tissue that has
been stored in culture for only a few days. One of the trials
reporting cases of severe dyskinesias used tissue stored in
culture for up to four weeks and it may be that holding tissue in
this way reduces its dopaminergic composition (Olanow et al.,
2003).
A further issue concerns the identification of dense
hyperdopaminergic areas within the graft of some patients with
graft induced dyskinesias. This may have caused uneven striatal
innervation and excessive dopamine release into non
reinnervated areas. It is also possible that variable side effects of
graft induced dyskinesias are related to patient selection.
Greater functional improvement is associated with younger
patients, and in patients with less advanced disease. This is most
likely because the neuropathology is relatively confined to the
nigrostriatal pathway and may have better trophic support
compared to patients with more advanced disease (Winkler et
al., 2005).
Stem cells:
Stem cells could provide one such source and would
overcome the issue of limited availability of fresh primary fetal
cells. A wide range of stem cells are being investigated as

53
potential sources of dopaminergic neurons for transplantation,
stem cells can be obtained from various sources (Morizane et
al., 2009).
The majority of research thus far with respect to the
formation of dopaminergic neurons for the treatment of PD is in
embryonic stem cells and neural stem cells. Dopaminergic
neurons are more easily obtained from neural stem cells in the
developing ventral mesencephalon (VM) than other parts of the
developing central nervous system but the number of
dopaminergic cells produced is still very low. Despite genetic
manipulation and the addition of various growth and
differentiation factors, generating large numbers of
dopaminergic cells from this cell type has had mixed results
(Panchision, 2006).
However, greater success has been achieved with the more
complex ES cells. Derived from blastocysts donated following
in vitro fertilization these cells are truly pluripotent. Promising
data have been obtained with dopaminergic neurons derived
from mouse ES cells, significantly improving motor function in
a rat model of PD. However, directing the differentiation of
human ES cells has proved complex and while 50% of cells
spontaneously differentiate into neurons upon leukemia
inhibitory factor (LIF) withdrawal, few are dopaminergic. Thus,
there is the need to develop protocols to ‘direct’ differentiation.
The most successful published protocols describe multiple
culture stages in which different transcription and growth

54
factors are added at controlled time points (Goldman and
Windrem, 2006).
However, despite good yield of dopaminergic neurons in
vitro, clinically relevant long term survival and behavioral
recovery in animal models rivaling that of primary tissue has yet
to be convincingly demonstrated. Neuronal stem cells unlike
embryonic stem cells, which are only derived from the
embryonic blastocyst, neural stem cells can be found both in
embryonic neural tissue and also in specific neurogenic regions
of the adult brain. If the in vivo survival of neural stem cells can
be improved they hold the potential to provide autologous
transplantation as patients provide the cells for their own
recovery (Morizane et al., 2009).
Interestingly, stem cells may not just be useful as dopamine
factories in the striatum. Some studies in both rodent and
primate models have shown significant behavioral recovery
following transplantation with neural stem cells. In addition to
the generation of a small population of dopaminergic neurons
other cells within the graft were found to be releasing growth
factors which are purportedto exert neuro-protective or
neuroregenerative influences. While more evidence needs to be
accumulated on the longevity of this effect, it broadens the
potential of neural stem cells from simple dopamine
replacement to preserving and enhancing remaining
dopaminergic neurons (Svendsen and Langston, 2004).

55
Stem cell based approaches could be used to provide
therapeutic benefits in two ways: first, by implanting stem cells
modified to release growth factors, which would protect existing
neurons and/or neurons derived from other stem cell treatments;
and second, by transplanting stem cell derived DA neuron
precursors/neuroblasts into the putamen, where they would
generate new neurons to ameliorate disease-induced motor
impairments (figure 17) (Lindvall and Kokaia, 2010).
Figure (17): Stem cell based therapies for PD. PD leads to the progressivedeath of DA neurons in the substantia nigra and decreased DA innervationof the striatum, primarily the putamen (Lindvall and Kokaia, 2010).
Neurogenesis:
As mentioned above endogenous stem cells are present in
specific regions of the brain. While the occurrence of
neurogenesis in the striatum and substantia nigra is debated, one

56
indisputable neurogenic region is the subventricular zone (SVZ)
lying adjacent to the striatum. The cells in the region are an
assortment of stem and progenitor cells that have the potential to
be mobilized and induced to differentiate by the presence of
growth factors or other small molecules. In the normal condition
75%–99% or the cells differentiate into granular GABAergic
neurons, with the rest forming periglomular neurons expressing
either tyrosine hydroxylase or GABA. The control of
proliferation and mobilisation of these cells may be
dopaminergic as both MPTP and 6-OHDA mediated dopamine
depletion reportedly decreases proliferation in this zone (Zhao
et al., 2008).
An additional source of endogenous source of new
dopaminergic neurons may be described presence of tyrosine
hydroxylase positive cell bodies in the striatum, which increase
in quantity with dopaminergic denervation. As yet there are no
imminent therapeutic strategies heading towards the clinic that
manipulate these endogenous systems but their potential is
waiting to be harnessed. Therapeutic strategies to increase
striatal dopamine could involve recruiting newly produced
neurons in the SVZ and encouraging them to migrate into the
striatum and differentiate into dopaminergic neurones or to
stimulate cells resident in the striatum. In order for this to be
achieved understanding more about these two processes of
neurogenesis and phenotypic switching in the striatum is
necessary, determining the intrinsic or extrinsic factors

57
responsible may provide an alternative set of mechanisms that
could be utilized to treat PD (Lane et al., 2008).
Graft standardization:
Grafting methods for the past 20 years have differed in
everything from procurement process to tissue composition to
implantation technique. To add even more variability, multiple
donors are needed to create a graft large enough to carry some
promise of efficacy. This, undoubtedly, plays an important role
in determining survival, growth, and integration of the
transplant (Morizane et al., 2009).
As stem cells can theoretically provide an endless source of
quality consistent neurons, standardization of the transplant
tissue will enhance the reliability of the procedure and its
results. One promising study has shown that implantation of
undifferentiated human neural stemcells (hNSCs) in
Parkinsonian primate brains can restore functionality.
Furthermore, the repair process not only reestablishes the gross
anatomical structure of the organ but does so with appropriate
proportions of neuron types. Guided by signals of the
microenvironment of the damaged brain, uncommitted hNSCs
are induced to differentiate into dopaminergic neurons, as well
as other cells that mediate neuroprotection (Lane et al., 2008).
Patient standardization:
With standardization of transplant material, patients must
likewise be evaluated for variables in their presentation of the
disease. Specifically, the distribution of the damaged neurons

58
should be taken into account before and after graft implantation.
In earlier studies, patient selections overlooked the preoperative
magnitude of the lesions, making it difficult to evaluate the
extent of the graft incorporation. Similarly, it was also unknown
whether continued postoperative degeneration of non grafted
regions would affect clinical response. Conversely, patients with
little or no postoperative damage showed the best functional
outcome. Because the decline of dopaminergic cells in areas
outside the nigrostriatal region seems to arise as PD progresses
to a more severe state, it may be that implantation during earlier
stages will exhibit a higher rate of success. This is mirrored in
the survival of transplanted tissue, which survives and integrates
better in younger patients. The reasons why this occurs have yet
to be fully determined, but it is known that neural growth factors
are expressed more in younger brains (Morizane et al., 2009).

59
Stem cell therapy in stroke
Stroke is defined as an abrupt focal loss of brain function
resulting from interference with the blood supply to part of the
central nervous system. It is one of the major causes of death
and disability among the adult population in the world. In spite
of the extensive research in the field of stroke biology, there is
little effective treatment for a completed stroke. Most strokes
fall into two main categories: ischemic (80%) or hemorrhagic
(20%) (Caplan, 2011).
The biology of cellular transplantation:
The transplantation of human neuronal cells is an approach
to reducing the functional deficits caused by CNS disease or
injury. Several investigators have evaluated the effects of
transplanted fetal tissue, rat striatum, or cellular implants into
small animal stroke models for the most part, clinical trial
designs using primary human fetal tissue into patients with
neurodegenerative diseases have lessened. The widespread
clinical use of primary human tissue is likely to be limited due
to the ethical and technical difficulties in obtaining large
quantities of fetal neurons at the same time; much effort has
been devoted to developing alternate sources of human neurons
for use in transplantation (Kondziolka and Wechsler, 2008).
When transplanted, these neuronal cells survived, extended
processes, expressed neurotransmitters, formed functional
synapses, and integrated with the host. During the retinoic acid

60
induction process, significant changes were seen in the neuron
precursor cells that resulted in the loss of neuroepithelial
markers and the appearance of neuronal markers. The final cell
product was a ≥ 95% pure population of human neuronal cells
that appeared virtually indistin-guishable from terminally
differentiated, post mitotic neurons. The cells were capable of
differentiation to express a variety of neuronal markers
characteristic of mature neurons, including all 3 neurofilament
proteins (L, M, and H); microtubule associated protein 2, the
somatic/dendritic protein, the axonal protein. Thus, the neuronal
phenotype made these cells a promising candidate for
replacement in patients with CNS disorders, as a virtually
unlimited supply of pure, post mitotic, and differentiated human
neuronal cells (Lindvall and Kokaia, 2010).
The concept of restoring function after a stroke by
transplanting human neuronal cells into the brain was conceived
in the mid 1990s. Research conducted in a rat model of transient
focal cerebral ischemia demonstrated that transplantation of
fetal tissue restored both cognitive and motor functions.
Ischemic stroke leads to the death of multiple neuronal types
and astrocytes, oligodendrocytes, and endothelial cells in the
cortex and subcortical regions. Stem cell based therapy could be
used to restore damaged neural circuitry by transplanting stem
cell derived neuron precursors/neuroblasts. Also, compounds
could be infused that would promote neurogenesis from
endogenous SVZ stem/progenitor cells, or stem cells could be

61
injected systemically for neuroprotection and modulation of
inflammation (Kondziolka and Wechsler, 2008).
Behavioral testing was conducted using a passive avoidance
learning and retention task and a motor asymmetry measure.
Animals that received transplants of neurons and treatment with
cyclosporine showed amelioration of ischemia induced
behavioral deficits throughout the 6 month observation period.
They demonstrated complete recovery in the passive avoidance
test, as well as normalization of motor function in the elevated
body swing test. In comparison, control groups receiving
transplants of rat fetal cerebellar cells, medium alone, or
cyclosporine failed to show significant behavioral improvement.
Subsequent studies have shown that these cells released glial
derived neurotrophic factor after transplantation into ischemic
rats (figure 18) (Lindvall and Kokaia, 2010).
Figure (18): Stem cell based therapies for stroke (Lindvall and Kokaia,2010).

62
Embryonic stem cells:
Animal models have demonstrated that ESCs, when
transplanted into adult hosts, differentiate and develop into cells
and tissues and thus may be applicable for treating a variety of
conditions, including Parkinson’s disease, multiple sclerosis,
spinal injuries, stroke, and cancer. Transplanted ESCs are
exposed to immune reactions similar to those acting on organ
transplants; hence, immunosuppression of the recipient is
generally required. It is possible, however, to obtain ESCs that
are genetically identical to the patient’s own cells using
therapeutic cloning techniques (Kalluri and Dempsey, 2008).
Several studies showed that these cells are able to migrate in
response to damage, using MRI, that ESCs that were implanted
into the healthy hemisphere of rat brains 2 weeks after focal
cerebral ischemia (FCI), migrated along the corpus callosum to
the ventricular walls, and populated en masse at the border zone
of the damaged brain tissue (i.e., the hemisphere opposite to the
implantation sites). Another study showed that undifferentiated
ESCs xenotransplanted into the rat brain at the hemisphere
opposite to the ischemic injury migrated along the corpus
callosum toward the damaged tissue and differentiated into
neurons at the border zone of the lesion. In the homologous
mouse brain, the same murine ESCs did not migrate, but
produced highly malignant teratocarcinomas at the site of
implantation, independent of whether they were
predifferentiated in vitro to NPCs. These results imply that

63
ESCs might migrate to the damaged site. However, the
production of teratocarcinoma raises concerns about the safety
of ESC transplantation in patients with stroke (Battler and Leor,
2006).
Adult neural stem cells:
During the last century, the dogma existed that the adult
CNS was incapable of generating new neurons (neurogenesis).
Over the past decades, convincing evidence emerged that
neurogenesis in the adult CNS is a continuous physiological
process. Neurogenesis is present in two regions: the
subventricular zone (SVZ) and the subgranular zone of the
dentate gyrus (figure 19) (Kalluri and Dempsey, 2008).
Figure (19): Schematic drawings showing ischemia induced damage in thecortex and neural stem cell proliferation in the subventricular zone anddentate gyrus; these hypothetical sections of brain illustrate the ischemictissue and the neurogenic regions. A: Drawing showing the subventricularzone of the lateral ventricles. B: Drawing showing the dentate gyrus in thehippocampus. Note the migration of cells from the subventricular zonetoward the infarcted area. Red dots represent proliferating and/or migratingneural stem and/or progenitor cells (Kalluri and Dempsey, 2008).

64
Additionally, other studies also indicated the existence of
NSCs in other regions of the CNS, namely the striatum, spinal
cord and neocortex. SVZ and dentate gyrus derived NSCs are
characterized by long term, self renewal capacity and
multipotency. Adult SVZ and dentate gyrus derived NSCs
persist throughout the life span of mammals including humans.
It is important to note that neurogenesis occurs in a
physiological mode or is exogenously modulated by external
signals or pathophysiological processes. External global
stimulants such as enriched environment, physical activity and
stress or application of defined molecules such as fibroblast
growth factor-2, vascular endothelial growth factor (VEGF),
brain derived neurotrophic factor (BDNF) and erythropoietin
differentially modulate adult neurogenesis. Finally, CNS disease
conditions such as seizures and traumatic brain injury
represented by respective animal models induce neurogenesis
(Panchision, 2006).
Role of neural stem cells:
Proliferation:
Neural stem cell proliferation involves the sequential
activation of several cell cycle dependent enzymes and proteins
to initiate either symmetrical (2 stem cells) or asymmetrical cell
division (1 stem and 1 progenitor cell). Erroneous activation of
cell cycle enzymes in differentiated cells like neurons, however,
can lead to either apoptotic death of the neurons or to the
formation of cancer cells. A plethora of growth factors are

65
expressed by the ischemic tissue, which may be responsible for
the ischemia induced neurogenesis. The increase in ischemia
induced neurogenesis could therefore be due to the upregulation
of growth factor content or their receptor expression (Hass et
al., 2005).
In addition, not all growth factors are stimulatory in
function. Although a variety of growth and trophic factors are
upregulated following ischemic injury, some are stimulators of
neural progenitor proliferation, whereas others block the self
renewal of cells. It is important to note that ischemia induced
migration of neuroblasts has been shown to be due to the
expression of monocyte chemoattractant protein (MCP-1),
which can attract the progenitor cells away from the neurogenic
niche, blocking their proliferation. Likewise, inhibition of
growth factor activity by its binding proteins may also interfere
with proliferation of cells. Interestingly, insulin like growth
factor–I (IGF-I) can stimulate the proliferation of progenitor
cells only in the presence of mitogens like fibroblast growth
factor–2 (FGF-2) and promotes differentiation following
mitogen (FGF-2) withdrawal (figure 20) (Kalluri and Dempsey,
2008).
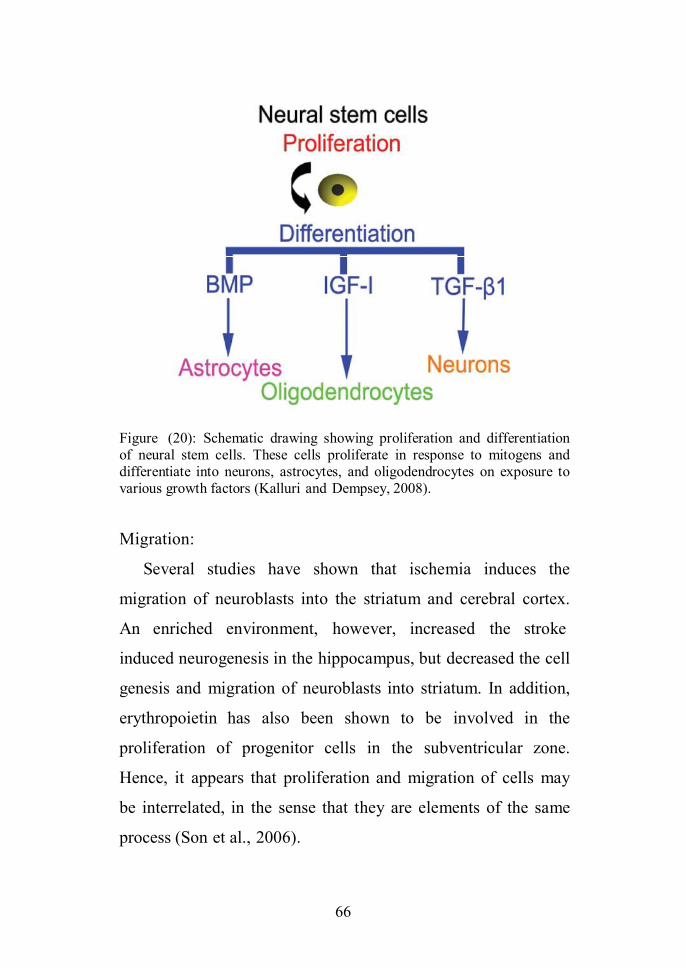
66
Figure (20): Schematic drawing showing proliferation and differentiationof neural stem cells. These cells proliferate in response to mitogens anddifferentiate into neurons, astrocytes, and oligodendrocytes on exposure tovarious growth factors (Kalluri and Dempsey, 2008).
Migration:
Several studies have shown that ischemia induces the
migration of neuroblasts into the striatum and cerebral cortex.
An enriched environment, however, increased the stroke
induced neurogenesis in the hippocampus, but decreased the cell
genesis and migration of neuroblasts into striatum. In addition,
erythropoietin has also been shown to be involved in the
proliferation of progenitor cells in the subventricular zone.
Hence, it appears that proliferation and migration of cells may
be interrelated, in the sense that they are elements of the same
process (Son et al., 2006).

67
It is thought that chemokines proteins released at the site of
injury by the inflammatory cells induce migration of the cells.
The expression of chemokine receptors on neural stem cells is
important for an efficient response to chemokines. The
expression of chemokine receptors is regulated by retinoic acid,
and that animals fed on a diet lacking retinoic acid have
decreased numbers of double cortin positive cells. Retinoic acid
has been shown to induce the differentiation of neurons in vitro.
Hence, it appears that lack of retinoic acid may have decreased
the neuronal differentiation and/ or the expression of chemokine
receptors, which results in decreased numbers of neuroblasts
(immature neuronal cells expressing double cortin) migrating
out of the neurogenic niche (figure 21) (Kondziolka and
Wechsler, 2008).

68
Figure (21): Schematic drawing showing ischemia induced neurogenesis.Ischemia increases the expression of IGF-I, FGF- 2, TGFb1, and MCP-1.The IGF-I and FGF-2 enhance the proliferation of cells, whereas MCP-1increases the migration of cells away from neurogenic regions toward theischemic tissue. The stem cells leaving the neurogenic regions exit the cellcycle, during which time TGFb1 promotes the differentiation of stem cellsinto neuroblasts (migrating immature neurons). Alternately, if proliferatingcells in the neurogenic region are exposed to TGFb1, it will inhibit theprocess, thus slowing down the proliferation of cells (Kondziolka andWechsler, 2008).
Differentiation:
The process of generating specialized cells from neural stem
cells is called differentiation. Neural stem cells can be
differentiated into neurons, astrocytes, and oligodendrocytes;
this involves interplay between intrinsic cellular programs and
extrinsic cues like growth factors provided by the surrounding
environment. Although neural progenitor cells proliferate in
response to ischemia, several immature neurons (double cortin
positive cells) were shown to be migrating toward striatum and
cortex (Battler and Leor, 2006).

69
Although IGF-I induces the proliferation and differentiation
of progenitor cells after ischemia, most of the cells that are
migrating toward the injury site are neuroblasts (immature
neurons) but not oligodendrocytes. Although speculative, a
neuronal differentiation factor like transforming growth factor
b1 (TGFb1), which is upregulated after ischemia, may be
responsible for the formation of neuroblasts during the post
ischemic neurogenesis. The differential response of IGF-I
toward proliferation and differentiation was shown to be
regulated by a mitogen activated protein kinase pathway.
Because matrix metalloproteinase (MMPs) play an important
role in both extracellular matrix (ECM) digestion and IGF
regulation, it is crucial to understand their direct (ECM
digestion) and indirect (IGF metabolism) effects on progenitor
cell differentiation. Hence it is possible that after stroke, cells
proliferate in response to mitogens and growth factors, some of
which exit the cell cycle due to the chemokine mediated
migration out of the neurogenic area, and differentiate into
neuroblasts in response to differentiation promoting factors such
as TGFb1. Thus, the final outcome depends on the spatial and
temporal expression of these factors following ischemia (Son et
al., 2006).
Potential mechanism involved in stem cell mediated
recovery after stroke:
Initial transplantation studies were focused on the potential
of NSCs to replace lost circuitry. Transplanted neural progenitor

70
cell (NPCs) in a rat model of global ischemia have been
reported to express synaptic proteins post transplantation.
However, only limited evidence demonstrates that transplanted
cells are able to sustain CNS repair through massive cell
replacement, especially to the extent that might be required after
stroke. Regardless of the characteristics of the experimental
disease, functional recovery achieved through NPC
transplantation does not correlate with absolute numbers of
transplant derived, newly generated, and terminally
differentiated neural cells (Emerson et al., 2008).
Stem cell induced neuroprotection:
Transplanted stem cells can provide neuroprotection and
reduce host cell death in the post stroke brain. Most authors
have reported functional recovery and a reduction in lesion size
when cells are transplanted within the first 24-48 hours after
stroke. The short timeframe in which NPCs affect recovery
cannot be explained by the regeneration of new neurons and
synapses, suggesting an important role for neuroprotection in
enhancing recovery. In fact, NPCs are known to exert direct
neuroprotection through the neutralization of free radicals,
inflammatory cytokines, excitotoxins, lipases, peroxidases, and
other toxic metabolites that are released following an ischemic
event (Ourednik et al., 2005).
The neuroprotective effects of transplanted NPCs are usually
accompanied by increased in vivo bioavailability of main
neurotrophins such as nerve growth factor, brain derived

71
neurotrophic factor (BDNF), ciliary neurotrophic factor,
vascular endothelial growth factor (VEGF), fibroblast growth
factor, and glial derived neurotrophic factor (GDNF).
Alternatively, one could argue that the cells did adhere to the
endothelium in the affected brain area and exerted their effect
through the secretion of GDNF but did not eventually engraft. A
different aspect of neuroprotection would be an effect on
endogenous neurogenesis after stroke. Endogenous
neurogenesis is increased after a stroke. Its exact function has
yet to be determined, but it may signify a natural repair
mechanism of the brain, which could be enhanced by
transplanted cells (Guzman et al., 2008).
Stem cell induced angiogenesis:
Transplanted NPCs can also enhance endogenous
angiogenesis. Increased vascularization in the penumbra within
a few days after stroke is associated with spontaneous functional
recovery. As early as 3 days after ischemic injury, new blood
vessels are observed in the stroke affected penumbra, and
growth continues to increase for at least 21 days. Transplanted
cell induced blood vessel formation has been reported with
BMSCs, NSCs, and cells from human cord blood or peripheral
blood. The ability of transplanted cells to increase endogenous
levels of angiogenic factors and chemoattractant factors (for
example, SDF-1) induces the proliferation of existing vascular
endothelial cells (angiogenesis) and mobilization and homing of

72
endogenous endothelial progenitors (vasculogenesis) (Shyu et
al., 2006).
The direct injection of recombinant stromal derived factor–1
(SDF-1) into the stroke affected rat brain resulted in the
increased recruitment of BMSCs and increased vascular density
in the ischemic brain. A similar effect was achieved by
treatment of stroke affected rats with granulocyte colony
stimulating factor (Ourednik et al., 2005).
Stem cell induced immunomodulation:
In addition to neuroprotection and enhancement of
angiogenesis, transplanted NPCscan also decrease post
ischemic inflammatory damage. A landmark study highlights
the remarkable ability of transplanted NPCs to promote
neuroprotection through an immunomodulatory strategy, once
within inflamed CNS areas, systemically injected NPCs persist
around the perivascular space where reactive astrocytes,
inflamed endothelial cells, and bloodborne infiltrating T cells
coreside. Adult NPCs can promote neuroprotection by releasing
antiinflammatory chemokines and by expressing
immunomodulatory molecules (Pluchino et al., 2005).
In animal models of stroke, decreased infiltration of
mononuclear cells has been demonstrated at the lesion borders
of ischemic areas in the CNS where NPCs accumulate. Working
under the paradigm of bidirectional communication between
transplanted cells and the inflammatory ischemic
microenvironment, one group has attempted to combine NPC

73
based and immunotherapy based approaches in spinal cord
injury. In 2006 Ziv and colleagues described a two stage process
of repair. In the first stage, myelin specific T cells induce
transplanted NPC migration to the injury site and also activate
microglia to adopt a neuroprotective phenotype; in the second
stage, NPC interaction with T cells and microglia leads to better
tissue preservation, increased endogenous neurogenesis, and
improved functional recovery.
Although basic differences between the post ischemic brain
and the post injury spinal cord might preclude the translation of
this approach to stroke, the common themes of a limited
therapeutic window to promote a more neurogenic
microenvironment beckon a closer look. Indeed, studies by
these same authors as well as others have revealed how
modulation and/or suppression of inflammation after stroke can
promote neuroprotection and neurogenesis. This fascinating
cross talk between neural and immune cells reveals additional
therapeutic opportunities for attenuating post ischemic
inflammatory damage and providing a
more hospitable environment for the
persistence of both endogenous and transplanted NPCs (Shyu et
al., 2006).
Stem cell induced neuroplasticity:
The ability of cell based therapies to enhance injury induced
plasticity has been demonstrated with other somatic stem cell
types. Human cord blood cells in the ischemic cortex increase
nerve fiber sprouting from the contralateral to the ischemic

74
hemisphere. Neural progenitor cells were first reported to alter
plasticity in models of epilepsy. Transplanted NPCs have also
been shown to scavenge inhibitory molecules that limit the
reorganization of the injured brain. We have found that NPCs
possess the potential to alter synaptogenesis by secreting
thrombospondins, which are a family of large extracellular
matrix proteins. Thrombospondins 1 and 2, secreted by
immature astrocytes, are necessary and sufficient for the
development of ultrastructurally normal synapses (Xiao et al.,
2007).
Possible ways to monitor cell treatment:
The clinical application of cell transplantation therapy for
stroke requires a means of noninvasive monitoring these cells in
patients. Several imaging modalities, including MRI,
bioluminescence imaging, PET, and in vivo fluorescence
microscopy, have been used to track stem cells in vivo. Of these
methods, however, only MRI and PET are practicable for
clinical use. MRI is a high spatial resolution method for tracking
cells in real time. Cells are preloaded with molecules that
substantially alter proton resonance, such as superparamagnetic
iron oxide (SPIO) and ultra SPIO (USPIO) particles. In vivo,
SPIO labeled cells appear as hypointense areas in tissues,
because of a susceptibility artifact most pronounced in iron
sensitive T2- and T2*-weighted images. With high resolution
MR imaging, even single mammalian cells have been depicted
after SPIO labeling. Several studies have shown the ability of

75
MR imaging to longitudinally track transplanted iron labeled
cells in different animal models, including stroke. Using clinical
grade human fetal derived NSCs (Guzman et al., 2008).
Future imaging modalities may involve a combination of the
high spatial resolution and excellent anatomical detail of MRI
with the high sensitivity of PET. An alternative direction for
stem cell imaging is the development of MR reporter genes
(Hass et al., 2005).
One great advantage of this technique is that no external
contrast agent needs to be administered for imaging. However,
the application of MR reporter genes for in vivo studies has
been extremely limited, mainly because of the low sensitivity of
the technique. In addition to tracking the transplanted stem cells
after stroke, surveillance of the microenvironment and
monitoring the therapeutic effect of the stem cells will play a
major role in the assessment of clinical efficacy. Several
preclinical studies have demonstrated the possibility of
monitoring the inflammatory reaction after stroke by using
USPIO particles (Kelly et al., 2004).

76
Stem Cell Therapy forDemyelinating Diseases
Multiple sclerosis (MS) is an autoimmune inflammatory
demyelinating disease of the central nervous system (CNS) that
is a leading cause of disability in young adults. This disease is
notorious for making a patient totally bed bound, not only
resulting in bed sores but also inviting all kinds of infections to
the disabled, depressed patient. Although intensive therapy has a
key role in managing this complex disease, even in this era of
advancement, the slowing accumulation of neurological damage
in MS has remained mostly irreversible. Along with therapies
that improve blood circulation, mitochondrial energy
production, and anti-inflammatory, antimicrobial,and
antioxidant factors, there is increased optimism among scientists
about slowing and perhaps reversing the disease with a
comprehensive program that includes stem cell therapies (Olek,
2010).
Precursor cells in the adult CNS are capable of regenerating
oligoden-drocytes and myelin:
In different experimental models of focal demyelination, it
has been shown that endogenous cells in the adult rodent CNS
have the potential for regenerating oligodendrocytes and myelin.
Several studies have investigated the nature of the cells that
remyelinate the adult CNS after induction of demyelinated
lesions. The lack of spontaneous remyelination in

77
experimentally chemical demyelinated lesions that were X-
irradiated to kill proliferating cells, suggests that cell division is
an absolute prerequisite for myelin regeneration. Although
differentiated oligodendrocytes may survive within such lesions,
they are unable to rebuild myelin sheaths (Charron et al.,
2009).
Myelin regeneration fails in MS:
Attempts to regenerate myelin can be also recognized
pathologically in brains of MS patients by the existence of
shadow plaques, which are partially remyelinated lesions. The
process of spontaneous functional remyelination is often
incomplete and limited, leading to permanent axonal loss and
fixed neurologic disability. Failure of remyelination could stem
either from insufficiency of endogenous remyelinating cells or
from lack of environmental support for this process (johnson,
2011).
Data from experimental models of demyelination and from
human brain tissue indeed suggest that several factors may have
a role in limiting myelin regeneration in the adult brain and its
subsequent failure. In experimental focal demyelination, it has
been shown that only a subpopulation of local progenitor cells
react to injury and generate new oligodendrocytes and myelin.
Although the existence of progenitor cells was demonstrable in
acute and chronic MS lesions, they did not exhibit reactive
increase in cell number as compared with normal white matter
(Battler and Leor, 2006).

78
This suggests that the response of the progenitor cell
population to the demyelinating process in the human brain is
deficient. It has also been suggested that repeated demyelinating
episodes in chronic and relapsing MS causes depletion in the
endogenous pool of progenitor cell transplantation for diseases
of myelin cells. Although progenitor cells decrease in number
after experimental focal demyelination, this was not observed in
pathological specimens of chronic MS lesions (Charles and
Alfredo, 2005).
In some patients, there was progressive loss of
oligodendrocytes and myelin without reactive remyelination,
whereas in others, who exhibited strong T cell and macrophage
activity, there was robust remyelination, indicating the
important role of tissue support to the remyelinating response.
Cell migration seems to be another limiting factor in myelin
regeneration. It has been shown that only progenitor cells that
reside at the margins of experimental lesions migrate into the
lesion core and remyelinate it, whereas long distance migration
of progenitor cells does not occur in the brain parenchyma
(Crosta, 2010).
Both environmental factorsand basic properties of
endogenous adult progenitor cells limit the degree of
spontaneous remyelination. The apparent linkage between the
acute inflammatory phase and myelin regeneration and the
necessity to remyelinate before axonal damage occurs, may
define a narrow time window when remyelination is feasible.

79
Although this time window may be too narrow for adequate
endogenous progenitor cell mobilization, it may also determine
the window of opportunity for therapeutic cell transplantation
(Olek, 2010).
Rationale for cell transplantation into the CNS to repair
myelin:
Basically, there are two therapeutic approaches to induce
remyelination in MS and both have shown promising results in
experimental animals:
1. Promotion of endogenous remyelination by growth factors
therapy.In acquired demyelinating diseases, such as MS, growth
factor therapy has proven effective in enhancing the endogenous
brain’s capacity for repair. Insulin like growth factor-1 (IGF-1)
and glial growth factor-2 (GGF2) are neurotrophic factors that
promote survival and proliferation in the oligodendrocyte
lineage. Treatment with these factors was beneficial clinically
and pathologically in animals with EAE.
2. Transplantation of myelin forming cells is a mode of
delivering the entire “cell factory” that manufactures myelin.
This approach may be advantageous over other modes of gene
therapy, in which targeting the gene to specific cells and tissues
and controlling its degree of expression may be problematic or
even detrimental (johnson, 2011).
Experimental transplantation has been performed in several
animal models of myelin disease. Transplanted myelin forming
cells remyelinated focal lesions in the optic nerve and spinal

80
cord and restored normal conduction properties, indicating fully
functional regenerated myelin (Crosta, 2010).
Remyelination by various myelin forming cells:
The traditional primary condition for any cell type to be a
candidate for therapeutic cell transplantation in myelin disorders
has been its ability to remyelinate and to restore nerve function.
Neural transplantation for remyelination has been studied for
more than 25 years, following early work by several
researchers. Various cell populations were shown to myelinate
efficiently after transplantation into experimental animals
(Einestien and Ben-Hurr, 2008).
The oligodendrocyte lineage:
Studies designed to identify the oligodendrocyte lineage
cells that had best myelinating potential showed that when
focally injected to chemically induce demyelinated lesions,
postmitotic oligodendrocytes had poor remyelinating capacity,
whereas OPCs showed greater mitotic, migratory, and
regenerative properties. Interestingly, transplanted OPCs were
found to be more efficient in remyelination than endogenous
OPCs (Charles and Alfredo, 2005).
Schwann cells:
Remyelination of CNS axons has been achieved by other
myelin forming cells as well. Schwann cells, the peripheral
myelin forming cells, have excellent myelinating properties in
the CNS. They produce thick and compact myelin after
transplantation into the CNS and can restore normal conduction

81
velocity in the dorsal columns of the spinal cord, indicating
functional recovery. The main advantage of using these
peripheral nervous system myelin forming cells is that Schwann
cells can be isolated from a sural nerve biopsy of patients
affected with a myelin disorder of the CNS, cultured and
expanded in vitro under appropriate conditions, cryopreserved,
and finally serve for autologous transplantation into
demyelinated CNS areas. Another potential advantage of
transplanted autologous Schwann cells is that they might escape
the autoimmune attack in MS that is directed against central
myelin antigens (Battler and Leor, 2006).
Olfactory nerve ensheathing cells:
Olfactory nerve ensheathing cells (OECs) display properties
of both astrocytes and Schwann cells. These cells are unique in
that they continue to develop in the olfactory epithelium
throughout life from which they migrate to the olfactory bulb.
Studies have shown that although these cells do not normally
make myelin, they are able to do so when transplanted to areas
of demyelination in the brain or spinal cord. These cells have a
capacity to grow in vitro and to remyelinate large axons with a
Schwann cell like pattern of myelin and improve conduction
properties after transplantation into the demyelinated adult rat
CNS. These cells seem also to promote axonal growth and
secrete neurotrophic molecules (Emerson et al., 2008).
Thus, the relative availability of these cells, their apparent
myelinating properties, and their trophic effect on axonal

82
growth make them another promising candidate for autologous
therapeutic transplantation. However, it is not yet clear whether
OECs can be expanded in sufficient amounts for human
transplantation and whether they will need to be isolated only
from the olfactory bulb, located intracranially, or also more
easily from the olfactory mucosa, situated at the back of the
nose outside the cranium (Einestien and Ben-Hurr, 2008).
Neural stem cells:
The main advantage of stem cells is that they are non
transformed precursors that are potentially able to self renew
indefinitely, allowing their expansion in large quantities. NSCs
can be expanded in vitro, maintain their capacity for self
renewal, and generate a progeny of the three neural cell lineages
(Charron et al., 2009).
NSCs retain their functional plasticity after in vitro
passaging and after several freezing thawing cycles and they can
still be modulated in vitro by exposure to different growth
factors. These uncommitted NSCs can integrate and repair the
damaged CNS and thus might represent a renewable source of
cells that can be used for transplantation procedures. Stem cells
can adapt their lineage fate and function according to
environmental needs (Crosta, 2010).
Transplanted NSCs of various origins improved the clinical
outcome in experimental models of stroke, spinal cord trauma,
and proved to have good myelinating properties as well.
Intraventricularly transplanted clonal NSCs in the newborn shi

83
mouse disseminated in widespread brain areas and participated
in myelination. Some recipient animals showed a decrease in
their symptomatic tremor. In adult animals with traumatic spinal
cord injury, rat NSCs migrated in the spinal cord and
differentiated into myelinating oligodendrocytes (Charles and
Alfredo, 2005).
Recently, it was also shown that intraventricular, as well as
intravenous, transplantation of stem cells into mice with chronic
EAE, resulted in clinical improvement. This was correlated by
graft derived and endogenous remyelination and by reduction in
axonal pathology. These data suggest that cell transplantation in
de- and dysmyelinated human disease may have therapeutic
potential for functional restoration. In the process of neural
development, there is continuous functional and lineage
potential specification of the stem cell, before entering the
neuronal, oligodendroglial, or astroglial lineage (Olek, 2010).
Although early NSCs in the developing cortical plate expand
very rapidly, they are not migratory cells. In the SVZ of the
CNS reside stem cells that generate mainly glia progeny. Donor
derived myelin was observed after SVZ precursor cells were
propagated in culture with epidermal growth factor (EGF) and
transplanted into the spinal cords of md rat and sh pup and to
the retinas of young mice (Battler and Leor, 2006).
Embryonic stem cells
ES cell lines can actually be established from virtually all
mammals and can be banked and propagated in vitro almost

84
indefinitely, with maintenance of a normal karyotype and
totipotency, as was shown by the culturing of mouse ES cell
lines in the presence of leukemia inhibitory factor. Mouse ES
cells can be induced to differentiate in vitro into neurons
(David, 2009).
The sequential use of growth factors, such as fibroblast
growth factor-2 (FGF2), EGF, and platelet derived growth
factor (PDGF), in a program that mimics embryonic
development, has been successful to derive glial precursors
from mouse ES cells. The myelinogenic potential of mouse ES-
derived OPCs, that were expanded in vitro, was demonstrated in
the embryonic md rat brains, when these cells extensively
myelinated the brain and spinal cord. When transplanted in a
rodent model of chemically induced demyelination and in spinal
cords of shi mice, mouse ES-derived progenitor’s cells were
also able to differentiate into glial cells and remyelinate
demyelinated axons in vivo (Goldman and Windrem, 2006).
Since the isolation of human ES cells, it has been possible to
generate an endless source of transplantable human ES-derived
neural precursors. Transplanted human ES-derived neural
progenitors into newborn mice differentiated into all three
neural lineages, including oligodendrocytes. The most important
potential hazard in stem cell (and especially ES cell)
transplantation is the risk of tumor formation. By definition, ES
cells are capable of forming teratomas after transplantation.
These teratomas contain cells of the three embryonic layers, i.e.,

85
ectoderm, mesoderm, and endoderm. When undifferentiated
mouse ES cells were transplanted into the brain, they could
generate brain cells, but at the expense of teratoma formation
(Svendsen and Ebert, 2008).
Clearly, the commitment to a restricted neural lineage should
be complete before transplantation in order to eliminate this
problem. Indeed, transplantation of mouse and human ES-
derived neural precursors, obtained after multiple in vitro
passages and exposure to various growth factors, did not result
in teratoma formation. Another problem in stem cell
transplantation is the possibility of graft rejection. In most
studies of xeno-transplantation of human cells into rodents, graft
rejection did not pose a significant problem. However, there is
no long term systematic follow up of the survival of these cells
in the brain (Einestien and Ben-Hurr, 2008).
Bone marrow stromal cells:
A central issue in the field of stem cell biology is the
suggestion that plasticity of stem cells is marked to the degree
of promiscuity, where stem cells of one tissue may generate
cells of other tissues. It has been shown that adult mouse and
human bone marrow stromal cells (BMSCs) can differentiate in
vitro into other cell types, including muscle, skin, liver, lung,
and neural cells. In humans affected by hematologic
malignancies, peripherally injected BMSCs enter the brain and
produce new neurons and microglia. Recent reports suggest that
these cells could contribute to the generation of new neural cells

86
in the adult brain by a direct conversion of transplanted BMSCs
into neurons or oligodendroglial cells, referred to as
transdifferentiation and/or assimilation of transplanted cells or
their progeny into existing neurons and formation of
heterokaryons, referred to as cell fusion (Crosta, 2010).
In rats with a demyelinated lesion of the spinal cord,
intravenous or brain injection of isolated mononuclear BMSCs
resulted in varying degrees of remyelination. In addition, bone
marrow derived stromal cells from transgenic green fluorescent
protein (GFP) mice that were injected directly into the
demyelinated spinal cord of immunosuppressed rats produced
myelin and improved axonal conduction velocity. These
observations stress the notion that it may not be mandatory to
introduce glial committed cells for remyelination in vivo, as the
developing and acutely demyelinated CNS may instruct other
cells to differentiate into the required lineage. Accordingly,
BMSCs might be useful as a therapeutic tool for brain repair by
autologous transplantation (David, 2009).
Problematic issues in cell transplantation for demyelinating
diseases:
Most experimental data on cell transplantation for
remyelination has been obtained from genetic dysmyelinating
models, where transplanted cells integrate into the normal
developmental program of the CNS, or in models of acquired
focal demyelination. However, the chronic and multifocal

87
nature of MS raises several crucial issues that need to be
considered in order to bring the therapeutic cell transplantation
approach closer to clinical reality (Battler and Leor, 2006).
Route of cell delivery:
Another key issue for cell transplantation in MS is the route
of cell delivery. Because MS is a multifocal disease, it is
impossible to introduce regenerating cells into all foci of
disease. Moreover, it is often difficult to determine which of the
multiple foci observed in the brain by magnetic resonance
imaging (MRI) is most important clinically. Also, current
neuroimaging techniques do not identify the specific
pathological pattern of the lesion, and whether it is amenable for
effective remyelination (Svendsen and Ebert, 2008).
Therefore, it is necessary to contemplate the optimal route of
cell delivery that will promote efficient targeted migration of
transplanted cells into multiple lesions for repair. Because most
white matter tracts that are involved in MS are in close
proximity to ventricular and spinal subarachnoid spaces, then
intraventricular and intrathecal transplantation may serve as an
efficient route of delivering remyelinatingcells. After
intracerebro-ventricular injection, transplanted cells may
disseminate throughout the neuroaxis without a separating
barrier from the CNS white matter. Intraventricular
transplantation of OPCs and stem cells led to widespread
myelination in the genetic dysmyelinating models of the shi
mouse and the md rat (Goldman and Windrem, 2006).

88
Migration of transplanted cells in the brain:
A crucial feature of transplanted cells is their ability to
migrate into inflamed brain areas, integrate, and differentiate.
The regenerative potential of transplanted cells is dependent on
their ability to arrive to the active inflammatory demyelinated
lesions. Cell migration is a major limiting factor in
remyelination. In the lesioned CNS, spontaneous remyelination
is a local event because of the limited migration of endogenous
remyelinating cells. Whereas transplanted multipotential NSCs
migrate and integrate in the embryonic and newborn rodent
CNS and adopt cellular identity according to local and temporal
cues, the normal adult brain does not permit large distance
migration and does not support transplanted neural cell survival.
Transplanted precursors were found to possess superior
migratory capabilities (figure 22) (Battler and Leor, 2006).
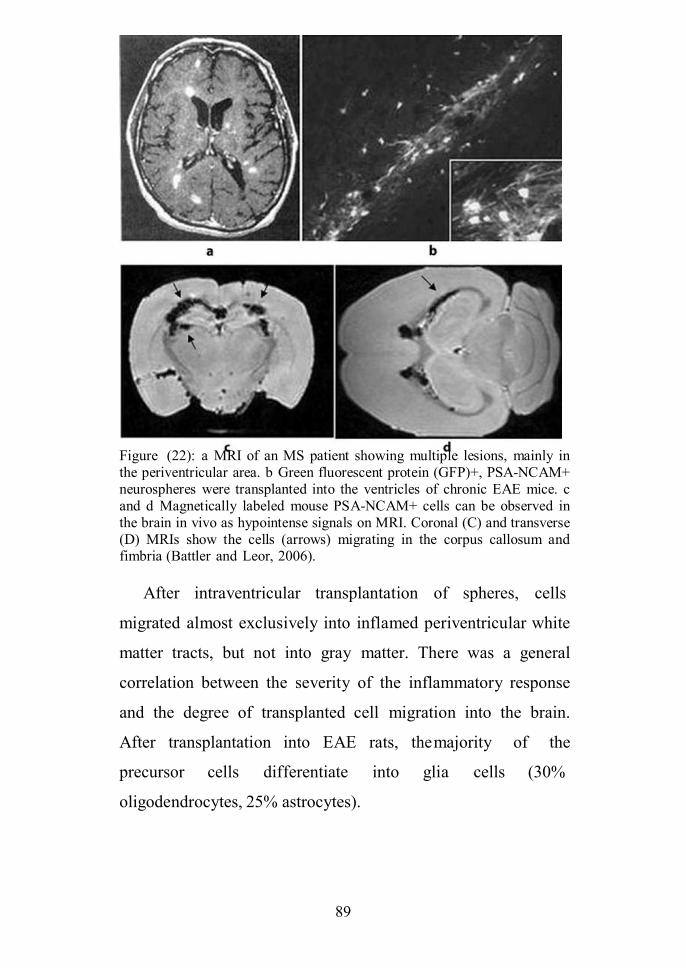
89
Figure (22): a MRI of an MS patient showing multiple lesions, mainly inthe periventricular area. b Green fluorescent protein (GFP)+, PSA-NCAM+neurospheres were transplanted into the ventricles of chronic EAE mice. cand d Magnetically labeled mouse PSA-NCAM+ cells can be observed inthe brain in vivo as hypointense signals on MRI. Coronal (C) and transverse(D) MRIs show the cells (arrows) migrating in the corpus callosum andfimbria (Battler and Leor, 2006).
After intraventricular transplantation of spheres, cells
migrated almost exclusively into inflamed periventricular white
matter tracts, but not into gray matter. There was a general
correlation between the severity of the inflammatory response
and the degree of transplanted cell migration into the brain.
After transplantation into EAE rats, themajority of the
precursor cells differentiate into glia cells (30%
oligodendrocytes, 25% astrocytes).

90
Because MS is a multifocal relapsing disease, these findings
exemplified the potential use of an intraventricular
transplantation as a route of cell delivery in MS, bringing cells
close to white matter tracts and enabling their inflammation-
induced targeted migration. These findings suggest a linkage
between parenchymal inflammation and setting regenerative
mechanisms in motion. As such, the inflammatory process that
develops in disease foci during clinical relapses of
demyelinating diseases may serve to attract remyelinating cells.
Thus, the brain inflammatory process may have a dual,
contrasting action in inflicting brain injury and recruiting the
regenerative process simultaneously (Olek, 2010).
This stresses the notion that combination of cell
transplantation and immunomodulation for MS in the future will
need to be developed as nonreciprocally antagonistic modes of
treatments. To this end, it is important to dissect the pro
regenerative components in the inflammatory process and target
the immunomodulatory treatment without inhibiting
regenerative processes. Tracking transplanted cells can be
performed non-invasively to develop successful clinical (stem)
cell based therapies, it will be important to develop methods that
can assess the fate and distribution of cells non-invasively. It is
obvious that traditional histopathological methods for cell
detection used in animal studies, which requires the removal of
tissue, cannot be applied to patients in most cases (Charles and
Alfredo, 2005).

Stem Cell Therapy for Amyotrophic
Lateral Sclerosis
Amyotrophic lateral sclerosis (ALS), first described by
Charcot in the nineteenth century is a relentlessly progressive
neurodegenerative disorder of motor neurons in cerebral cortex,
brain stem and spinal cord that causes muscle weakness,
disability, and eventually death, with a median survival of three
to five years. ALS is also known by the eponym "Lou Gehrig's
Disease," after the famous baseball player who was affected
with the disease. Motor neuron disease (MND) is the preferred
term in the United Kingdom, but in the United States ALS and
MND are sometimes used interchangeably (Maragakis et al.,
2010).
Cell transplant based neuroprotection:
A variety of different cell types have been examined with
the intent of modulating the multifactorial cascade of events
contributing to the pathological interplay between and within
microglia, astrocytes, and neurons at the local neuronal
microenvironment. These attempts to affect neuronal survival
have predominantly incorporated cell types known to directly or
indirectly serve a physiological support role, including olfactory
ensheathing cells, Sertoli cells, ESCs, and neural precursor cells
at varying levels of lineage restriction and differentiation.
Finally, the modification of local immunological function has

been alternately attempted through the use of both bone marrow
stromal cells and umbilical cord blood (Crosta, 2010).
Physiological support of cell engraftment:
Both olfactory ensheathing cells and Sertoli cells have been
exploited in mutant superoxide dismutase 1 (mSOD1) ALS
animal models because of their known in vivo physiological
support roles. Olfactory ensheathing cells provide trophic
support and guidance to advancing axons of the primary
olfactory receptor neurons of the nasal mucosa as they traverse
the cribriform plate before synaptic connection with projection
neurons of the olfactory bulb. In vitro studies have indicated
OEC production of mRNA encoding nerve growth factor,
BDNF, GDNF, and CNTF, whereas alternate studies have
corroborated the production of corresponding neurotrophic
proteins (Svendsen and Ebert, 2008).
Approaches to glial cell replacement:
The supplementation or replacement of endogenous astroglia
has been attempted with a variety of different cell types of
varied lineage restriction and differentiation potential from
embryonic, fetal, or adult tissues. The first successful attempts
to promote indefinite ESC survival in an in vitro culture utilized
murine cells supplemented with media containing a basal feeder
layer of inactivated mouse embryonic fibroblasts either with or
without leukemia inhibitory factor (LIF) supplementation.
Removal of LIF or the basal feeder layer, when present, resulted
in the initial differentiation of these cells, as evidenced by

embryoid body (EB) formation. Subsequent discoveries have
demonstrated that LIF application does not prevent EB
formation, as does the use of a basal feeder layer, when applied
to hESCs (Lindvall and Kokaia, 2010).
Cell transplant based neurorestoration:
The restoration of lost neurological function is a goal
relevant to the repair of damage due to a wide variety of causes,
including neurodegenera-tive conditions such as ALS.
However, this process requires successful negotiation of
multiple complex issues, including ability for transplanted
neuronal precursor cells to survive transplantation, terminally
differentiate, send peripheral axonal projections, generate
synaptic connections at the neuromuscular junction, and result
in functional incorporation (Charron et al., 2009).
Approaches to Motor Neuron Production:
Achieving the appropriate culture conditions required to
reproducibly generate cell types of a neuronal or neuronal
subtype lineage has required the application of detailed
knowledge of the microenvironment encountered by ESCs
residing within the developing neural tube. In the developing
embryo, growth promoting chemicals called “morphogens”
provide directional cues along both dorsoventral and
rostrocaudal planes, guiding neuronal growth. Therefore,
identification of the appropriate morphogens followed by their
subsequent application at empirically derived concentrations
and time points represents a means to recapitulate the in vivo

conditions promoting terminal differentiation into cell types of
defined lineages. The application of retinoic acid as a
caudalizing factor and sonic hedgehog as a ventralizing factor to
age appropriate embryonic cells can be used to preferentially
differentiate this pluripotent cell type into a terminally
differentiated motor neuron. Enhancement of neurite outgrowth
has been extensively studied in vitro and is affected by a variety
of substances such as cAMP analogs, fatty acid derivatives,
phosphodiesterase inhibitors, and Rho kinase inhibitors (Jodi et
al., 2004).
Role of Stem Cell Therapies in ALS:
Stem cells could help patients with ALS in several ways.
Ideally, they could be induced to differentiate into lower motor
neurons in order to replace those neurons that die because of
ALS. Perhaps stem cells could rescue dying motor neurons by
reconnecting these neurons to partly denervated muscle before it
has died completely. Better yet, they could be induced to
differentiate into upper motor neurons in the cortex and connect
to the lower motor neurons (Metwally, 2009).
Stem cells also play a supportive role in maintaining the
viability of or extending the function of surviving motor
neurons. The stem cells could be induced to differentiate into
supporting cells, glia, or interneurons that might produce factors
that would support motor neurons, or perhaps the stem cells
themselves might produce such factors (Svendsen and
Langston, 2004).

95
Stem Cell Therapy for Muscular
Dystrophy
The muscular dystrophies are an inherited group of
progressive myopathic disorders resulting from defects in a
number of genes required for normal muscle function. The
Duchenne and Becker muscular dystrophies are caused by
mutations of the dystrophin gene and are therefore named
dystrophinopathies. Weakness is the principal symptom as
muscle fiber degeneration is the primary pathologic process.
Muscular dystrophies, include X-linked recessive as in
Duchenne MD, autosomal recessive as in limb-girdle MD type
2, or autosomal dominant as in Facioscapulohumeral MD,
myotonic dystrophy and limb girdle MD type 1 (Darras, 2010).
The dystrophinopathies have varying clinical characteristics:
• Duchenne muscular dystrophy (DMD) is associated with themost severe clinical symptoms
• Becker muscular dystrophy (BMD) has a similar presentationto DMD, but a relatively milder clinical course
• An intermediate group of patients, known as "outliers," maybe classified clinically as having either mild DMD or severeBMD (Gussoni et al., 2002).

96
Cell therapy:
The concept of a cell based therapy to promote muscle
regeneration, in particular in recessive types of MD, originated
with the observation of the intrinsic ability of myofibers to fuse
to each other. Thus, functional correction could be obtained by
generating hybrid muscle fibers, where the donor nuclei provide
the missing gene product. Based on this premise, several
investigators have assessed the ability of adult myoblasts to treat
DMD, which is characterized by the lack of dystrophin (David,
2009).
Murine models for muscular dystrophy include the naturally
occurring mdx mice. The mdx mice are dystrophin deficient due
to a stop codon point mutation in exon 23 of the dystrophin
gene. Although initial results in mdx mice were encouraging,
early clinical trials failed due especially to the poor survival and
limited migratory ability of injected myoblasts (Gabellini et al.,
2006).
The lesson learned from these early studies is that a more
primitive cell population endowed with self renewal and
differential potential would be preferable for therapeutic
applications. Accordingly, a number of studies involving
different sources of stem cells, including muscle satellite cells,
vessel derived stem cells (mesoangioblasts) and embryonic stem
cells have provided better outcomes with wider distribution of
transplanted cells, higherlevels of engraftment, and
importantly, improvement in the contractile properties of

97
muscles from transplanted dystrophic mice (Cerletti et al.,
2008).
Because all these studies were performed in recessive forms
of MD, it is unknown to what extent dominant forms of MD
could benefit from stem cell based therapy. Since it takes fewer
nuclei to add back a missing product than it takes to dilute out a
deleterious product, the bar is expected to be higher for effective
treatment of a dominant disease, however this will depend on
the specific disease mechanism (Sampaolesi et al., 2006).
In experimental studies, engraftment and ameliorate the
phenotype in a mouse model would produced for a dominant
neuromuscular disorder. For these studies, we have chosen mice
over expressing FRG1, which have been proposed as a mouse
model for Facioscapulohumeral dystrophy (FSHD). Transgenic
mice over expressing FRG1 (medium and high expressers)
develop a myopathy and phenotypic abnormalities that are
typical of MD, including abnormal spinal curvature, skeletal
muscle atrophy, increased variability in fiber size, fiber
necrosis, centrally located nuclei and fibrosis (D’Antona et al.,
2007).
Although there is some controversy in the field as to the
relationship of this pathophysiology to human FSHD, for the
purposes of this study, the salient feature is that it is dominantly
inherited. A total of 24 FRG1-medium mice at 12 weeks of age
were used for these experiments. All mice were treated with an
immunosuppressive agent (tacrolimus; Sigma) to prevent

98
rejection of nonisogenic cells at a daily dose of 5mg/kg
intraperitoneally (IP) from the day before cell injection until the
time of euthanasia (30 days) (David, 2009).
EBs were subjected to Pax3 induction (doxycycline was
added to the EB culture medium) from day 2 to day 5 of EB
differentiation, at which point early myogenic progenitors were
purified by FACS using antibodies to PDGFαR and Flk-1.
PDGFαR+Flk-1- cells were expanded in the same medium
containing doxycycline for 7-10 days and then transplanted into
FRG1 transgenic mice In the first series of experiments, female
FRG1 mice were transplanted with Pax3-induced ES derived
myogenic progenitors through intramuscular (i.m.; n=4) and
intravenous (i.v.; n=4) injections. 24 hours before
transplantation, 15 μl of cardiotoxin (10μM, Sigma) was
injected into the tibialis anterior (TA) muscle of each mouse
(both legs) to induce muscle injury (Sampaolesi et al., 2006).
In the intramuscular group, mice received 1 × 106 cells by
direct intramuscular injection into the left tibialis anterior (TA)
muscle whereas the right TA muscle received only PBS
(control). In the systemic i.v. group, mice received 5 × 105 cells
through tail vein injection while the control group (n=4)
received PBS. After 30 days, muscles were harvested and
analyzed by immunofluorescence to assess engraftment by the
presence of GFP (donor cells) and MHC, a marker of terminal
muscle differentiation. In a second set of experiments, female
and male FRG1 mice were transplanted with Pax3-induced ES-

99
derived myogenic progenitors through intramuscular injection
(a total of 6 mice in each group), as described above, and
muscle function was assessed 30 days following the
transplantation. Muscle cryosections were also evaluated for
engraftment (Deasy et al., 2007).
Muscles were frozen in isopentane cooled in liquid nitrogen.
Tissue cryosections (6-8 mm) were fixed with cold acetone for
5 min., permeabilized with 0.5% Triton X-100 (Sigma) for 20
min., blocked with 10% goat serum (or 3% BSA) for 1 hr, and
then incubated with primary antibodies: anti-MHC (1:20
overnight 4°C) and chicken anti-GFP (1:500, 1hr RT; Abcam).
For secondary staining, appropriate secondary alexa fluor
antibodies were used (Invitrogen). As observed, whereas PBS-
injected FRG1 mice were negative for GFP, transplantation of
Pax3-induced ES-derived myogenic progenitors, independent of
the route of administration, resulted in substantial engraftment
as evidenced by the presence of GFP+MHC+ myofibers. In the
case of intramuscular injection, transplanted cells migrated and
engrafted well around the site of injection. As observed
previously, these cells were able to home to the muscle
following their systemic injection. TA muscles from these mice
contained about 14.5 ± 1.07 % of GFP+ myofibers (D’Antona
et al., 2007).
The measurement of muscle contractile properties was
performed as previously described. Interestingly we observed
gender differences in regard to the contractile ability of TA

100
muscles from FGR1 mice. Whereas male FRG1 mice showed
significant reduction in their contractility ability when compared
to the B/6 control group, female mice demonstrated no
functional deficit. Accordingly, cell transplantation had no
effect in this latter group, while the transplantation of Pax3-
induced ES-derived myogenic progenitors in male FRG1
resulted in improvement of maximal isometric tetanic and
specific force. It is important to note that engraftment levels
were similar among female and male groups. CSA and weight
were not affected by cell transplantation. According to our
previous results, while systemic delivery of ES-derived
myogenic progenitors into mdx mice reduced these parameters
close to wild-type mice, intramuscular transplantation did not
affect CSA and weight, as observed here. However it is
important to note that mdx and FRG1 mice present quite distinct
phenotypes. While mdx mice are hypertrophic due to the
multiple rounds of degeneration and regeneration, FRG1 mice
are atrophic (David, 2009).
Gender differences in skeletal muscle have been
demonstrated with energy metabolism, hormonal status, and
muscle fiber types. For instance male skeletal muscles generally
contain more fast fibers than female muscles. Conversely,
during repeated contractions, female muscles are generally more
fatigue resistant and recover faster. It has been shown in FRG1
mice that fast fibers are preferentially involved in the pathology
(D’Antona et al., 2007).

101
Accordingly, staining for slow and fast isoforms of MHC in
male and female FRG1 mice showed gender specific
differences. Female FRG1 mice contained significantly more
slow myofibers than males, as evidenced by NADH tetrazolium
reductase staining, while immunohistochemistry for a fast
isoform of MHC revealed a lower frequency of fast myofibers.
This may explain, at least in part, the results observed here.
Consistently, female FRG1 mice presented much less fibrosis
than their male counterparts, as evidenced by Mason’s
Trichrome staining, which corroborates their milder phenotype.
It is important to note that cell transplantation did not affect
fibrosis or the ratio of fast and slow myofibers in male or female
FRG1 mice. Moreover, it has recently been pointed out that
gender also affects stem cell mediated skeletal muscle
regeneration, with female muscle derived stem cells (MDSCs)
having higher potential for muscle regeneration than their male
counterparts. Although these studies were performed in mdx
mice through transplantation experiments, one could
hypothesize that resident MDSCs in female FRG1 mice are
better able to respond to the damage caused by overexpression
of FRG1 (Deasy et al., 2007).

102
Stem Cell Therapy for Huntington's
Disease
Huntington disease (HD) is an inherited progressive
neurodegenerative disorder characterized by choreiform
movements, psychiatric problems, and dementia. The disease is
named after the American doctor George Huntington, who first
described the condition in detail in 1872. It is caused by a
trinucleotide (CAG) expansion in the Huntington gene on
chromosome 4p and inherited in an autosomal dominant pattern.
The pathophysiology of HD is not fully understood, although it
is thought to be related to toxicity of the mutant huntingtin
protein (Sutton and Suchowersky 2010).
Current work in HD cell therapy is broadly classified into 1
of 3 aims:
1) To harness the ability of the brain to self repair through the
upregulation of endogenous stem cells/neurogenesis;
2) To replace dead and/or dying neurons through fetal or stem
cell transplantation; and
3) To protect neurons vulnerable to disease progression through
the administration of neuroprotective trophic factors via cell or
viral delivery.
Clinical trials of fetal derived cell transplantation in the
striatum have been performed since the 1990s in patients with

103
HD. However, a more cogent understanding of the pathological
processes that give rise to HD and the behavior of both
endogenous and exogenous cells in the diseased brain will
contribute to not only additional or improved therapeutic
strategies, but also a better context through which the success of
therapeutic manipulations can be analyzed and understood
(figure 23) (Claire et al., 2008).
Figure (23): Drawing illustrating therapies in HD. Sites of celldegeneration (caudate and putamen) in HD appear in red, and ongoing adultneurogenesis in the human brain appears in green. Target of fetal derivedcell transplantation into the diseased striatum is represented by the syringe(Claire et al., 2008).

104
Restoring or manipulating neurogenesis as endogenous cell
therapy in HD:
One focus of cell therapy has been to understand the deficits
in endogenous stem cell regulation in HD, primarily in the adult
hippocampal, dentate gyrus (DG) and SVZ, with the aim that
restoring or upregulating endogenous neurogenesis may
partially ameliorate disease symptoms and repair the damaged
brain. Neurogenesis occurs in the hippocampus and SVZ of
adult mammals, including humans (figure 24) (Johann et al.,
2007).
Figure (24): Drawing depicting sites of neurogenesis in the rodent brain. A:Newborn hippocampal neurons (blue) are born in the subgranular zone ofthe hippocampus and migrate into the granule cell layer of the DG as theymature and integrate. B: Neurons (blue) originate in the SVZ lining thelateral ventricles and migrate via the rostral migratory stream (RMS) to theolfactory bulb. CA1 = cornu ammonis 1 of the hippocampus proper; CA3 =cornu ammonis 3 of the hippocampus proper; GCL = granule cell layer; GL= glomerular layer (Johann et al., 2007).

105
Neural stem cells in the SVZ olfactory bulb and DG can
differentiate into all lineages of the adult CNS including
neurons. Furthermore, endogenous NSCs outside the 2 standard
neurogenic regions have been shown, in some instances, to have
the capacity to differentiate into neurons and integrate
appropriately in regions undergoing cell death (Chen et al.,
2004).
Neurodegeneration or injury can upregulate proliferation and
promote the migration of newborn cells to the site of damage.
However, it is not clear if this increase in cell genesis is
neuroprotective and/or linked to functional recovery, if the
increase is of a magnitude to be of therapeutic value, or if this
upregulation in proliferation represents pathological network
reorgani-zation (Johann et al., 2007).
Neural stem cells in the SVZ and hippocampal subgranular
zone develop into electrically mature and fully integrated
neurons. In the subgranular zone ofthe hippocampus,
progenitor cells become neurons predominantly and
functionally integrate into the DG. Neural precursor cells
originating in the SVZ of the lateral ventricle migrate
tangentially through the rostral migratory stream, guided by
astrocytes to the olfactory bulb where they become fully
differentiated and electrically competent. Neurogenesis has
been implicated in learning and memory and may be necessary
for the functional integrity of the circuits into which new cells

106
generate. The rate of neurogenesis in adult mammals is
dynamically regulated by a variety of physiological and
pathological factors (Jin et al., 2005).
The microenvironment plays a key role in maintaining the
neurogenic potential of the adult brain; it also plays a role in the
fate restriction of endogenous NSCs in neurogenic regions.
Local conditions of the micro-environment, including factors
released by astrocytes, play a crucial role in neurogenesis.
Dysregulation of this environment in the degenerating brain can
result in a non permissive environment for neurogenesis
(Svendsen and Ebert, 2008).
Despite the expression of mutant the huntingtin gene (htt),
the proliferation, longevity, differentiation, and survival of
NPCs derived from the hippocampus and SVZ of HD transgenic
mice are similar to those of wild type derived NPCs in vitro,
suggesting that abnormalities in neuro-genesis in vivo are due to
the microenvironment in which the NPC resides and not the
intrinsic properties of the precursor itself. Although there is no
change in basal SVZ NPC proliferation or maturation in a
rodent model of HD, the absence of increased proliferation of
SVZ NPCs in response to quinolinic acid indicates deficits in
the SVZ microenvironments (Phillips et al., 2005).
Although little work has been done on the effect of HD on
adult human hippocampal neurogenesis, the SVZ in adult
humans with HD has been well characterized, increased cell
proliferation in, and the thickness of, the SVZ correlate with

107
pathological grade and CAG repeat length. It is unclear whether
this increase in proliferation is a protective response by the CNS
to replace or protect damaged cells in the striatum or whether
the upregulation in neurogenesis contributes to pathology in the
striatum, Most of the cells born in the diseased SVZ exhibit
phenotypes of glial cells (progenitors or mature glial fibrillary
acidic protein positive astrocytes) with fewer neurons.
Furthermore, this effect is region specific there is increased
proliferation in the central and ventral regions of the SVZ,
whereas striatal atrophy occurs mostly in the dorsal region
(Battler and Leor, 2006).
The birth site of new neurons in proximity to the
degenerating striatum as well as the upregulation of
neurogenesis in this region in response to damage in HD raises
the possibility that the brain can at least partially compensate for
neurodegeneration. Indeed, in a rat lesion model of HD, SVZ
progenitor cells have been shown to migrate from the site of
their inception toward the lesion. A better understanding of the
mechanisms underlying adult neurogenesis within the context of
HD would contribute to our ability to harness the therapeutic
potential of these cells (Svendsen and Ebert, 2008).
Strikingly, the beneficial impact on cognitive and motor
behavioral measures through the restoration of adult
neurogenesis in rodent models of HD has revealed the
therapeutic potential of endogenous NPCs. The induction of
neurogenesis through trophic factors or antidepressant treatment

108
has alleviated motor and cognitive deficits in rodent models of
HD. In a mouse model of HD, treatment with fibroblast growth
factor–2, a neuroprotective growth factor, increased cell
proliferation by 150% in the SVZ (compared with 30% in wild
type littermate controls) and was accompanied by an increase in
immature doublecortin positive neurons and a decrease in
aggregations formed by mutant huntingtin protein. These cells
migrated to the striatum (the primary site of pathology) and
formed projections. Improved motor performance on a test for
motor coordination (rotarod) and extended lifespan were also
observed (Jin et al., 2005).
Cognitive and hippocampal neurogenic deficits in HD
transgenic mice can be rescued (that is, return to normal or near
normal levels of behavior) by chronic administration of the
antidepressant fluoxetine without affecting motor deficits
characteristic of HD and thus suggesting that in addition to
striatal pathology, secondary abnormalities in adult
hippocampal neuro-genesis play a role in the affective
symptoms of HD. This finding is in line with the dependency of
behavioral outcomes on hippocampal neurogenesis following
antidepressant treatment. Finally, the forced induction of
neostriatal neurogenesis also slows disease progression and
alleviates motor and cognitive impairments in a mouse model of
HD By correcting abnormalitiesin adult hippocampal
neurogenesis or manipulating endogenous neurogenic regions to
contribute to the repair of degenerating neuronal populations, it

109
is theoretically possible to nonsurgically treat both the
underlying pathologies and the cognitive and motor symptoms
of HD (Chen et al., 2004).
Transplantation of fetal derived cells as exogenous cell
therapy in HD:
One of the most studied areas of HD cell therapy is the
transplantation of fetal derived cells into the brain of a
symptomatic patient with HD. Extensive work on the safety and
efficacy of the transplantation of fetal striatal allografts into a
diseased adult brain has been completed in both rodent and
primate models, and several clinical trials have been performed
since the 1990s. Although full recovery has not been observed
following fetal allografts in the striatum of humans with HD,
some suggestions of delayed disease progression indicate
positive functional outcomes. The contribution of these grafts to
functional recovery is enhanced by the fact that implanted cells,
lacking the disease causing gene, do not themselves appear
vulnerable to neurodegenerative processes, an effect that is also
seen in transplants in patients with PD (Bachoud-Lévi et al.,
2006).
All clinical trials to date have been focused on the
transplantation of fetal derived cells into the diseased striatum.
Because HD pathophysiology involves several brain regions,
including nonstriatal sites, cell implantation into the striatum
alone may limit the ability of grafts to address or reverse HD
symptoms. Whereas grafted regions are relatively spared from

110
neurodegeneration, surrounding brain regions continue to
degenerate, which may limit the functional outcome of
transplantation. Furthermore, the timing of transplantation may
be crucial to the functional benefits of the graft; the majority of
patients involved in clinical transplantation trials have had mild
to moderate HD, and the impact of the disease stage on
transplant efficacy remains to be measured (Claire et al., 2008).
Much evidence has accumulated about the efficacy of fetal
derived cell transplantations in animal models of HD. All
striatal cell types from the allo and xenotransplantation of fetal
or embryonic derived striatal tissue have been shown to survive,
grow, and establish functional afferent and efferent connections
to host tissue and display appropriate electrophysiolo-gical
properties in both rodent and primate models of HD. authors
have demonstrated that behavioral motor deficits are improved
by the transplantation of striatal tissue into the degenerate
striatum of rodents and primates. Both pre and postoperative
motor training in animal models of HD have also had a
significant impact on functional recovery, presumably by
enhancing the plasticity of grafts and host circuitry (Svendsen
and Ebert, 2008).
In primates, striatal allografts and xenografts survive,
differentiate into mature dopamine and cyclic adenosine
monophosphate regulated phosphoprotein positive striatal cells,
and receive dopaminergic innervation. These changes are
correlated with improvements in both motor performance,

111
including recovery in a test of skilled motor performance, and
cognitive function. Cellular and behavioral improvements
following fetal derived striatal cell transplantation are thought to
occur through circuit reconstruction, the normalization of
neurotransmitter release, and/or the production of trophic
factors (Claire et al., 2008).
In this respect, van Dellen and colleagues have demonstrated
the potential therapeutic benefit of targeting the host cortex as a
site of transplantation. Wild type fetal cortical cells were
transplanted into the anterior cingulate cortex of neonatal HD
transgenic mice. This therapy delayed the onset of an overt
motor deficit but did not enhance motor coordination. This
study highlights the potential benefit of targeting the widespread
pathophysiology of HD, not just neurodegeneration in the
striatal regions, as it may allow preservation of striatal function
through the delivery of neuroprotective trophic factors, such as
brain derived neurotrophic factor (BDNF), from spared
nonstriatal regions that project to the striatum (Bachoud-Lévi et
al., 2006).
Clinical trials involving the transplantation of fetal striatal
tissue in humans were begun in the late 1990s. As in animal
models of HD, most transplanted tissue was derived from the
human fetal striatum (whole or partial ganglionic eminence) and
transplanted into the host striatum. The amount and cellular
composition of the transplanted cells must be regulated, as
transplants with, 30% striatal content are ineffective, and the

112
selective dissection of striatal primordial tissue (that is, the
lateral region of the lateral ganglionic eminence) has proven
disappointing from a functional perspective. As in PD, the
safety of allografts of fetal striatal neural tissue has been
demonstrated in HD. Similar to its successful use in cell therapy
for PD, hibernation medium used to store harvested cells for up
to 8 days has allowed the collection of adequate amounts of
tissue and the flexibility to prepare tissue at sites and times other
than the site and time of transplantation surgery (figure 24)
(Claire et al., 2008).
Figure (25): Flair MRI obtained in a patient with HD who had received agraft of fetal derived striatal tissue into the caudate and putamen, showingthe patient’s status 3 (A), 8 (B), 13 (C), and 32 months (D) aftertransplantation. The origin of the apparent persistent signal along the impacttract is unknown but may represent surviving donor tissue or host glialreaction (Claire et al., 2008).
Graft survival has been reported in several clinical trials.
MRI has been successfully used to locate and monitor the
growth of grafted tissue, and metabolic assessment has also

113
been effective in evaluating the efficacy of transplantation.
Tissue at the site of grafting has been shown on MRI to increase
in growth by 6 months post transplantation, without overgrowth.
Furthermore, inferences regarding the health of the graft, based
on MR spectroscopy and PET, suggest that graft sites are
populated by adult neurons that are metabolically active. For
example, the maturation of transplanted cells led to the
stabilization of brain striatal and cortical metabolism in 3 of 5
patients receiving bilateral fetal striatal grafts in an open label
study. In the remaining 2 patients, striatal and cortical hypo
metabolism progressed (Johann et al., 2007).
However, reductions in metabolic activity, including a
significant loss of dopamine receptor binding, have been
documented in patients 2 years following intrastriatal
transplantation of fetal derived cells. A 7% decline in striatal
metabolism was observed on fluorine-18 labeled
fluorodeoxyglucose PET over a 6 year period in patients who
had received functional benefit from the transplants; although
not significantly different, this decline was lower than the
average for patients with HD who did not receive implants. On
average, dopamine receptor (D2) binding declines by 5-15% in
control patients with HD as measured on PET. In general,
increases in hypometabolism in patients with HD who have
received transplants are consistent with those in control patients
with HD, although 1 patient showing clinical improvement after

114
transplantation also showed improvement on striatal raclopride
PET scans (Claire et al., 2008).
In addition, postoperative FLAIR imaging showed a
persistent signal that could represent grafted tissue, host reaction
(gliosis), or both. Postmortem histological evidence for the
survival and integration of fetal derived cell striatal
transplantation has also been reported in 3 patients who died
between 18 and 79 months after the procedure. Fetal lateral
ganglionic eminence was bilaterally grafted into striata with
HD. Autopsy results revealed that grafts were clearly
demarcated and grew to 5-10% of the normal human caudate
putamen tissue volume after 18 months in 1 patient. Grafted
cells displayed morphological features characteristic of both the
developing and the mature striatum and were innervated by
appropriate host target regions, although the presence of
efferents was very limited in the tissue from patients surviving
6 years after transplantation, despite minimal gliosis within the
grafted tissue (Keene et al., 2007).
Notably, grafted fetal striatal cells survive, develop, and
integrate into host tissue without being affected by the disease
process itself; that is, no mutant huntingtin aggregates were
observed within the transplanted region, and the graft showed
no signs of immunological rejection. The sparing of grafted
tissue from disease progression has also been reported in other
transplant cases, including in patients with PD. The success of
allografts of fetal derived striatal tissue in patients with HD

115
hinges on motor and cognitive improvements following
transplantation (Chen et al., 2004).
Improvements on the Unified Huntington’s Disease Rating
Scale motor score have been reported as early as 1 month after
unilateral grafting of fetal derived striatal tissue and have
stabilized between 6 months and 2 years after transplantation.
Patients assessed up to 6 years after bilateral transplantation
have demonstrated that immediate improvements in motor
scores (between 6 months and 2 years) may not persist long
term. However, the rate of motor and cognitive decline may be
reduced in some patients who have undergone transplantation.
Of those patients exhibiting initial functional and metabolic
benefits from transplantation in the clinical trial, clinical
improvement reached a plateau after 2 years and declined
variably from 4 to 6 years. Dystonia progressed more rapidly
and consistently than other motor symptoms, but transplantation
of fetal derived striatal tissue appeared to alleviate choreatic
symptoms (Bachoud-Lévi et al., 2006).
Cognitive performance was also stabilized on non timed
tests in these patients. The remaining 2 patients who showed
neither metabolic nor functional improvement at the initial 6
month follow up continued to decline over the 6 year course of
study at a rate similar to that seen in patients with HD who had
not received transplants (Rosser et al., 2002).
Clinical trials have included patients in earlier stages of the
disease, which reduces the risks of this complication.

116
Complications from immunosuppressive drug treatment
following transplantation have been generally predictable with
largely reversible side effects and thus have not been a major
safety concern. However, the issues of whether
immunosuppressive therapy is needed for intracerebral
transplantation and the optimal duration of immunosuppressive
treatment have not been resolved (Claire et al., 2008).
The use of fetal derived striatal cells for transplantation has
shown that grafts into the degenerative adult brain are safe and
at least partially efficacious in terms of some immediate
functional response. However, this approach seems to be less
viable over the long term. In addition to the ethical and practical
considerations regarding the source of cells for human fetus
derived allografts and the limited repair potential of striatal only
grafts, many groups have turned to alternative strategies for
obtaining cells for therapy in HD, the most notable of which has
been the utilization of stem cells (Keene et al., 2007).
Stem Cell Based Cell Therapy in HD
Stem cell therapy in the adult brain with HD has received
increasing attention for its potential to mitigate pathological
neurodegeneration by replacing lost neuronal and/ornon
neuronal populations, by triggering endogenous repair
mechanisms, or by protecting existing cells primarily through
trophic support. Stem cell transplantation is a promising field in
cell therapy for HD as stem cells are relatively easy to obtain
compared with primary fetal tissue and have the potential to be

117
manipulated to eliminate possible problems of host rejection.
Neural stem cells can be isolated from the fetal, neonatal, and
adult brain and propagated in culture (McBride et al., 2004).
Grafted NSCs can develop both morphological and electrical
properties of mature neurons and integrate into host circuitry.
Transplanted immature neurons or embryo derived NPCs have
been shown to migrate toward the site of degeneration,
differentiate, and form synaptic connections in several models
of neurodegeneration. Transplanted ESCs can differentiate into
neurons in the HD affected striatum and appear to migrate to
nearby cortical regions where they have been found to express
markers of immature neurons. One of the first challenges to
stem cell therapy in HD is to determine which source of stem
cells is most efficacious, and many sources have been
examined. In addition to human ESCs, stem cells derived from
mesenchyme in adults have been investigated as a readily
available source of stem cells in HD (Song et al., 2007).
Whereas few neurons have been formed from mesenchyme
derived stem cell grafts, transplantation of these grafts has
elicited some behavioral recovery. Similarly,
xenotransplantation of adult peripheral precursor cells
originating from porcine Sertoli cells of the testes has shown the
rescue of locomotor impairments in an HD lesion model in rats,
probably acting through neuroprotection given the effectiveness
after only a short time (McBride et al., 2004).

118
Neural stem cell treatments have resulted in either full or
partial functional recovery in tests of lesion induced
impairment. Transplanted human and mouse derived NPCs in
an excitotoxic HD lesion model not only have survived in the
striatal site of implantation, but also have migrated and stained
for markers of immature neurons in cortical regions. Following
transplantation into a mouse model of HD, murine ESC derived
NPCs, genetically modified to promote neuronal differentiation,
formed GABAergic neurons with appropriate outgrowth. The
grafts were also functionally beneficial given that the animals
showed improvement in rotational behavior. Human NSCs
appear to behave similarly to murine derived NSCs in rodent
models of HD. Intravenously transplanted human NSCs migrate
to the striatum, reduce striatal atrophy, and contribute to
functional improvement in a rodent lesion model of HD (Song
et al., 2007).
The differentiation of stem cells or treatment with growth
factors in vitro prior to implantation may facilitate fate
determination while mitigating the risk of tumor formation
posed by stem cells. Additionally, stem cell therapy has the
potential to alleviate cognitive deficits in degenerative disease,
such as memory impairment, through the replacement of
degenerating cells and the restoration or preservation of proper
network function (Claire et al., 2008).
Although the gold standard in stem cell treatment has been
the production of neurons, the differentiation of transplanted

119
cells into non neuronal lineages, such as astrocytes, may serve
neuroprotective or regenerative roles in degenerative
environments. Additionally, there is at least corollary evidence
that diseased glial cells contribute to HD pathophysiology.
Endogenous or transplant derived nonneuronal cells must be
included in any consideration of the efficacy of stem cell
therapy in HD. In neurogenic regions, astrocytes interact with
NSCs and NPCs to promote proliferation and fate determination
and may stimulate the maturation and integration of newly
formed neurons (Song et al., 2007).
The contribution by non neuronal cells may be particularly
relevant in regions affected by HD in which it is hypothesized
that abnormalities in the microenvironment, rather than neuron
intrinsic properties, primarily contribute to the degenerative
environment. Taken together; results from initial stem cell
therapy investigations have indicated that stem cells are viable
cell sources for transplantation in animal models of HD (Claire
et al., 2008).

120
Stem cell therapy for Alzheimer's
Disease
One hundred years ago, 1906, the German psychiatrist Alois
Alzheimer presented the case of a 54 years old woman who
suffered from progressive dementia. He described the clinical
and pathological symptoms for the first time, now known as
Alzheimer’s disease (AD) (Ermini, 2006).
AD is a neurodegenerative disorder characterized by
progressive dementia with memory loss as the major clinical
manifestation. Impairment of short term memory is often
prominent, but remote memory loss also appears to be affected
over time. Other important features of AD are an altered
behavior including paranoia, delusions, depression, impairments
of attention, perception, reasoning and comportment, and a
progressive decline in language function (Grabowski, 2010).
The main neuropathologic findings in the AD brain are
amyloid plaques which are multicellular lesions containing
extracellular deposits of amyloid β-protein (Aβ) consisting of
amyloid fibrils (7-10 nm) intermixed with non fibrillar forms of
the Aβ peptide. Neuronal and synaptic loss, neurofibrillary
tangles, and deposits of β amyloid protein in senile plaques
involve the basal forebrain cholinergic system, amygdala,
hippocampus, and cortical areas. The plaques are surrounded by
dystrophic neuritis, another form of Aß deposition appears as

121
diffuse or pre amyloid plaques where Aβ occurs in a non-
fibrillar, less dense and amorphous form in the neuropil,
cerebral amyloid angiopathy (CAA) is detected in 30 % of the
general population, compared to 80% in AD patients (Ermini,
2006).
Stem cell based therapies for AD:
The situation for neuronal replacement aiming at functional
restoration in AD is extremely complex because the stem cells
would have to be pre differentiated in vitro to many different
types of neuroblasts for subsequent implantation in a large
number of brain areas. Since acetylcholinesterase inhibitors,
which enhance cholinergic function, induce some temporary
improvement in AD patients, the cognitive decline could
hypothetically be improved by transplantation of stem cell
derived basal forebrain cholinergic neurons (figure 26) (Lindvall
and Kokaia, 2010).

122
Figure (26): Stem cell based therapies for AD. AD leads to neuronal loss inthe basal forebrain cholinergic system, amygdala, hippocampus, andcortical areas of the brain; formation of neurofibrillary tangles; and amyloidprotein accumulation in senile plaques. Stem cell based therapy could beused to prevent progression of the disease by transplanting stem cellsmodified to release growth factors (Lindvall and Kokaia, 2010).
Experimental evidence indicates that it should be possible to
generate such cells from stem cells. However, to give long
lasting symptomatic benefit, a cholinergic cell replacement
approach would require intact target cells, host neurons that the
new cholinergic neurons can act on, and they are probably
damaged in AD. Stem cell based cell replacement strategies are
very far from clinical application in AD. The disease symptoms
in AD could partly be due to impaired formation of new
hippocampal neurons from endogenous NSCs in the subgranular
zone of the dentate gyrus, which is believed to contribute to
mood regulation, learning, and memory. Mouse models of AD

123
have provided equivocal data, with studies demonstrating both
increased and decreased hippocampal neurogenesis, one
important factor being disease severity, with a compensatory
increase in progenitor proliferation in the early stages and
decreased proliferation and survival with advanced pathology
(Zhao et al., 2008).
Formation of immature hippocampal neurons has been
reported to increase in senile AD patients, while in another
study, neurogenesis was not altered in pre senile cases. Later
studies have indicated deficient maturation of new neurons in
AD brains. As a result, approaches to enhance neurogenesis
and/or maturation could be considered potential stem cell based
therapies for AD. Clearance of brain β-amyloid has been
proposed to be of value in halting disease progression in AD.
Active β-amyloid vaccination in young AD mice, using as
antigen a sequence of the β-amyloid peptide, decreased β-
amyloid burden and increased hippocampal neurogenesis.
Moreover, passive β-amyloid immunotherapy with an antibody
specific for aggregated β-amyloid restored neurogenesis and
morphological maturation of new hippocampal neurons in aged
transgenic mice with β-amyloid related impairments of
neurogenesis (Charles and Alfredo, 2005).
Stem cell based gene therapy could deliver factors
modifying the course of AD and may be advantageous because
of the capacity of stem cells to migrate and reach large areas of
the brain. Preclinical studies that provide a rationale for this

124
approach include one demonstrating that basal forebrain grafts
of fibroblasts producing nerve growth factor (NGF), which
counteracts cholinergic neuronal death, stimulate cell function
and improve memory in animal models of AD (Zhao et al.,
2008).
Stem cells could also be engineered to carry other genes,
such as that encoding BDNF, which has substantial
neuroprotective effects in AD models. Transplantation of
fibroblasts producing the β-amyloid degradingprotease
neprilysin have been shown to reduce plaque burden in AD
mice, although this may not lead to reduction in the number of
pathogenic β-amyloid oligomers or prevent cognitive deficits
(Lindvall and Kokaia (2010).

125
Stem Cell Therapy for Cerebral palsy
Cerebral palsy (CP) consists of a heterogeneous group of
non progressive clinical syndromes that are characterized by
motor and postural dysfunction, it is caused by abnormal
development or damage in one or more parts of the brain that
control muscle tone and motor activity. The resulting
impairments first appear early in life, usually in infancy or early
childhood. Infants with cerebral palsy are usually slow to reach
developmental milestones such as rolling over, sitting, crawling,
and walking. Although the disorder itself is not progressive, the
appearance of neuropathologic lesions and their clinical
expression may change over time as the brain matures (Miller,
2010).
Stem cell implantation:
In clinical study, more than 88 cases are already treated by
stem cells with clinical improvement in around 70 % cases.
Cells of highest purity, viability and integrity from world quality
laboratory were used. As it has been proved that more
transplantation cycles gives better and better results. Any
perinatal injury to infant brain or hypoxic damage to cerebral
cortex is mainly responsible for cerebral palsy, few factors,
which needed to be thought over. When we are thinking about
treatment of cerebral palsy:

126
1. Bone marrow transplantation is autologous, so there is no fear
of rejection or mismatch or need for immunosuppressant
required in other transplantation surgeries.
2. No confirmed documentary evidence that autologous stem
cells are dangerous (except embryonic stem cells). In
combination with BMSCT, We also use cord stem cells which
we are using do not have any surface antigen, so they have no
immune or rejection from body. Cord stem cells also have much
better regeneration potential. Combination therapy compliments
each other as bone marrow stem cell produces much favorable
environment for cord stem cell to act.
3. Good results are documented in otherwise dead end patients.
Other treatment modalities are only supportive and empirical.
4. Many of cases have shown definite clinical improvement till
to date (Battler and Leor, 2006).
Stem cell therapy is used to induce regeneration in various
neurological disorders, including cerebral palsy. During the
treatment authors give 6 doses of stem cells for transplantation
cycles, 3 autologous bone marrow related and 3 cord
mesenchymal cells in high doses. As it has been proved that
more transplantation cycles gives better and better results.
Treatment lasts for 17 days and Patient will need admission for
6 days and 11 days gap in between (Cohen et al., 2009).

127
Treatment results:
Follow up statistics from 50 cerebral palsy patients showthat close to 67% experienced improvements after stem celltherapy. The type of improvements reported include: decreasedspasticity; better coordination; improved motor function,improved posture stability; better cognition resulting incommunication improvements; gaining the ability to sit, standor even walk unassisted. Improved speech & I.Q. was observedin 56% of patients. 43% reported a decrease or even absence ofepileptic seizures following treatment. About 20% showedimproved cognition (Battler and Leor 2006).

128
Batten DiseaseBatten disease is a rare but fatal neurodegenerative disease
that affects children and for which no treatment exists, other
than possible future stem cell research treatments. The disease is
named after the British physician Frederick Batten, who first
identified and described the condition in 1903, it is also known
as Spielmeyer Vogt Sjogren Batten disease. Batten disease
manifests itself in children somewhere between the ages of 4
and 10 years. Early signs might include vision impairment, poor
circulation, hyperventilation, reduced communicative ability,
and behavioral changes. As a neurodegenerative disease, Batten
disease causes the patient to progressively lose motor skills,
communication skills, and brain functions (Svendsen and Ebert,
2008).
The results are distressing not just to the patients concerned
but also to their careers. Batten disease is a genetically inherited
disease that leads to a mutation in the patient that prevents the
elimination of toxins from the brain. It also leads to a build up
of lipofuscins in the body, and these combinations of proteins
and fats are the symptoms by which the disease is detected.
Since the 1990s, a team of scientists have discovered certain
elements of the X chromosome, resulting in the discovery of the
genetic causes of Batten disease, among other conditions. In the
years since, six genes have been discovered that are associated
with the onset of the disease, although it has not yet been

129
determined what functions most of these genes possess (Hough
et al., 2010).
However, the determination of the cause of the disease has
led to an indication of how treatments could be created. These
treatments are based on injecting genetic material directly into
the affected area, the brain of an affected child. In the last few
years, several teams have been working with injecting fetal stem
cell material into the brains of children with Batten disease, and
there have been reports of positive outcomes, although it is too
soon to determine whether a permanent cure is possible or
whether the treatment is temporary in effect. Nevertheless,
patients have responded well to the treatment, and certain motor
skills and communication skills have been returned to them.
Parents who are able to talk to their children after years of them
being unable to speak find their quality of life to be greatly
improved, irrespective of whether that improvement will be
sustained permanently (Svendsen and Ebert, 2008).
The treatment is a proprietary preparation of human central
nervous system stem cells. To avoid patients rejecting the
foreign cells, the immune system must be suppressed to some
extent, and it is in this area that most problems are anticipated to
arise. However, tests show that this problem has been
successfully negotiated within the trials conducted (Miller,
2010).

130
A further stream of research has been sparked by the
detection of mature onset Batten disease in a breed of dogs.
Testing shows that perhaps 5% of these dogs may suffer from
the disease. Dog owners and their associations have been
working in partnership with patients and their careers in sharing
information and jointly sponsoring and supporting research. It is
hoped that new forms of treatment might, in due course, arise
from this collaboration (Hough et al., 2010).
The source of most of the genetic material used to treat
Batten disease is aborted fetuses, which are the main source of
the stem cells required. However, fetal cells are currently the
only known source for the stem cells required, and the treatment
relieves terrible suffering. In the future, alternative sources of
stem cells might be identified, including, for example, children
who have died from natural causes or as the victims of road
traffic or other accidents. The use of organs from such sources is
much more widely accepted in most societies. A further ethical
issue concerns the use of children in medical experimentation.
Although only children suffer from Batten disease, there is still
the need to consider whether they are able to give informed
consent to a new and unproven form of treatment, especially
given the progressive damage to their brains. The basic principle
that applies is that consent is provided on behalf of the children
by their parents or other legally appointed guardian (Svendsen
and Ebert, 2008).

131
Stem Cell Therapy for AutismAutism spectrum disorders (ASD) are a group of
biologically based neurodevelopmental disorders characterized
by impairments in three major domains: socialization,
communication, and behavior. These disorders include autistic
disorder (classic autism, sometimes called early infantile autism,
childhood autism, or Kanner's autism), Rett disorder, childhood
disintegrative disorder, pervasive developmental disorder not
otherwise specified (PDD-NOS), and Asperger disorder (also
known as Asperger syndrome) (Augustyn et al., 2011).
Hypoperfusion of brain in autism:
Children with autism have been consistently shown to have
impaired, or subnormal CNS circulation, as well as resulting
hypoxia. Defects include basal hypoperfusion, and decreased
perfusion in response to stimuli that under normal circumstances
upregulates perfusion. In numerous studies the areas affected by
hypoperfusion seem to correlate with regions of the brain that
are responsible for functionalities that are abnormal in autism.
For example, specific temporal lobe areas associated with face
recognition, social interaction, and language comprehension,
have been demonstrated to be hypoperfusedn in autistic but not
control children (Augustyn et al., 2011).
Immune deregulation in autism:
The fundamental interplay between the nervous system and
the immune system cannot be understated. Philosophically, the

132
characteristics of self/non self recognition, specificity, and
memory are only shared by the immune system and the nervous
system. Physically, every immune organ is innervated and bi-
directional communication between neural and immune system
cells has been established in numerous physiological systems. In
autism, several immunological abnormalities have been detected
both in the peripheral and the central nervous systems.
Astrocytes surround various portions of the cerebral
endothelium and play a critical role in regulating perfusion, and
blood brain barrier function (Vargas et al., 2005).
Physiologically, astrocytes play an important protective role
against infection, generating inflammatory cytokines. Through
secretion of various chemokines, astrocytes play an important
role in shaping adaptive immune responses in the CNS.
Astrocyte secretion of a soluble neurotoxic substance has been
demonstrated to be involved in disease progression. Astrocyte
hyperactivation has been demonstrated in this disease by
imaging, as well as autopsy studies. In multiple sclerosis,
astrocytes play a key role in maintaining autoreactive responses
and pathological plaque formation (Ichim et al., 2007).
Treatment of hypoperfusion defect by umbilical cord blood
CD34+ stem cell administration:
Therapeutic angiogenesis, the induction of new blood
vessels from preexisting arteries for overcoming ischemia, has
been experimentally demonstrated in peripheral artery disease,
myocardial ischemia, and stroke. Angiogenesis is induced

133
through the formation of collateral vessels and has been
observed in hypoperfused tissues. This process is believed to be
coordinated by the oxygen sensing transcription factor hypoxia
inducible factor-1 (HIF-1). During conditions of low oxygen
tension, various components of the transcription factor dimerize
and coordinately translocate into the nucleus causing
upregulation of numerous cytokines and proteins associated
with angiogenesis (Ichim et al., 2007).
The potency of tissue ischemia stimulating angiogenesis is
seen in patients after myocardial infarction in which bone
marrow angiogenic stem cells mobilize into systemic circulating
in response to ischemia induced chemotactic factors. The
association between neural angiogenesis and neurogenesis after
brain damage is not only temporally linked but also connected
by common mediators, for example, SDF-1 secreted in response
to hypoxia also induces migration of neural progenitors.
Angiogenic factors such as vascular endothelial growth factor
(VEGF) and angiopoietin have been implicated in post ischemia
neurogenesis (Ishibashi et al., 2009).
Methods to enhance angiogenesis and as a result
neurogenesis are numerous and have utilized approaches that
upregulate endogenous production of reparative factors, as well
as administration of exogenous agents. For example,
administration of exogenous cytokines such as fibroblast growth
factor-2 (FGF-2), erythropoietin, and G-CSF, has been

134
performed clinically to accelerate healing with varying degrees
of success (Kalluri and Dempsey, 2008).
A promising method of increasing angiogenesis in situations
of ischemia is administration of cells with potential to produce
angiogenic factors and the capacity to differentiate into
endothelial cells themselves. Accordingly, the use of CD34+
stem cells has been previously proposed as an alternative to
growth factor administration. Therapeutic administration of
bone marrow derived CD34+ cells has produced promising
results in the treatment of end stage myocardial ischemia, as
well as a type of advanced peripheral artery disease called
critical limb ischemia. Additionally, autologous peripheral
blood CD34+ cells have also been used clinically with induction
of therapeutic angiogenesis (Augustyn et al., 2011).
Cord blood has been used successfully for stimulation of
angiogenesis in various models of ischemia. In one report, the
CD34+, CD11b+ fraction, which is approximately less than half
of the CD34+ fraction of cord blood, was demonstrated to
possess the ability to differentiate into endothelial cells. In
another report, VEGF-R3+, CD34+ cells demonstrated the
ability to differentiate into endothelial cells and were able to be
expanded 40-fold expansion. The concentration of this potential
endothelial progenitor fraction in cord blood CD34+ cells is
approximately ten fold higher as compared to bone marrow

135
CD34+ cells (1.9% +/- 0.8% compared to 0.2% +/- 0.1%)
(Vargas et al., 2005).
Given the potency of cord blood CD34+ cells to induce
angiogenesis in areas of cerebral hypoperfusion, we propose
that this cell type may be particularly useful for the treatment of
autism, in which ischemia is milder than stroke induced
ischemia, and as a result the level of angiogenesis needed is
theoretically lower. However at face value, several
considerations have to be dealt with. Firstly, cord blood contains
a relatively low number of CD34+ cells for clinical use.
Secondly, very few patients have access to autologous cord
blood; therefore allogeneic cord blood CD34+ cells are needed
if this therapy is to be made available for widespread use. There
is a belief that allogeneic cord blood cells can not be used
without immune suppression to avoid host versus graft
destruction of the cells (Ichim et al., 2007).
Safety concerns regarding allogeneic CD34+ cells are
divided into fears of graft versus host reactions, as well as host
versus graft. The authors of the current paper have published a
detailed rationale for why administration of cord blood cells is
feasible in absence of immune suppression. Essentially, GVHD
occurs in the context of lymphopenia caused by bone marrow
ablation. Administration of cord blood has been reported in over
500 patients without a single one suffering GVHD if no immune
suppression was used. Although host versus graft may
conceptually cause immune mediated clearing of cord blood

136
cells, efficacy of cord blood cells in absence of immune
suppression has also been reported. Accordingly, we believe
that systemic administration of expanded cord blood derived
CD34+ cells may be a potent tool for generation of neo
angiogenesis in the autistic brain (Vargas et al., 2005).
Immune modulation by mesenchymal stem cells:
The treatment of immune deregulation in autism is expected
to not only cause amelioration of intestinal and systemic
symptomology, but also to profoundly influence neurological
function. Reports exist of temporary neurological improvement
by decreasing intestinal inflammation through either antibiotic
administration or dietary changes. Although, some anti
inflammatory treatments have yielded beneficial effects, no
clinical agent has been developed that can profoundly suppress
inflammation at the level of the fundamental immune
abnormality. We believe mesenchymal stem cell administration
may be used for this purpose. This cell type, in allogeneic form,
is currently in Phase III clinical studies for Crohn's disease and
Phase II results have demonstrated profound improvement
(Svendsen and Ebert, 2008).
An important characteristic of MSCs is their ability to
constitutively secrete immune inhibitory factors such as IL-10
and TGF-b while maintaining ability to present antigens to T
cells. This is believed to further allow inhibition of immunity in
an antigen specific manner, as well as to allow the use of such
cells in an allogeneic fashion without fear of immune mediated

137
rejection. Antigen specific immune suppression is believed to
allow these cells to shut off autoimmune processes (Keilhoff et
al., 2006).
Further understanding of the immune inhibitory effects of
MSCs comes from the fact that during T cell activation, two
general signals are required for the T cell in order to mount a
productive immune response, the first signal is recognition of
antigen, and the second is recognition of costimulatory or
coinhibitory signals. MSCs present antigens to T cells but
provide a coinhibitory signal instead of a co stimulatory signal,
thus specifically inhibiting T cells that recognize them, and
other cells expressing similar antigens. Supporting this concept,
it was demonstrated in a murine model that MSCs
transplantation leads to permanentdonor specific
immunotolerance in allogeneic hosts and results in long-term
allogeneic skin graft acceptance (Augustyn et al., 2011).
Other studies have shown that MSCs are inherently immuno-
suppressive through production of PGE-2, interleukin-10 and
expression of the tryptophan catabolizing enzyme indoleamine
2, 3, dioxygenase as well as Galectin-1. These stem cells also
have the ability to non specifically modulate the immune
response through the suppression of dendritic cell maturation
and antigen presenting abilities. Immune suppressive activity is
not dependent on prolonged culture of MSCs since functional
induction of allogeneic T cell apoptosis was also demonstrated

138
using freshly isolated, irradiated MSCs (Svendsen and Ebert,
2008).
Others have also demonstrated that MSCs have the ability to
preferentially induce expansion of antigen specific T regulatory
cells with the CD4+ CD25+ phenotype. Supporting the potential
clinical utility of such cells, it was previously demonstrated that
administration of MSCs inhibits antigen specificT cell
responses in the murine model of multiple sclerosis,
experimental autoimmune encephalomyelitis, leading to
prevention and/or regression of pathology. Safety of infusing
MSCs was illustrated in studies administering 1–2.2 × 106
cells/kg in order to enhance engraftment of autologous bone
marrow cell. No adverse events were associated with infusion,
although level of engraftment remained to be analyzed in
randomized trials. The ability of MSCs on one hand to suppress
pathological immune responses but on the other hand to
stimulate hematopoiesis leads to the possibility that these cells
may also be useful for treatment of the defect in T cell numbers
associated with autism (Ichim et al., 2007).

139
Stem cell therapy for Retinal
Degeneration
In the back of the eye exists a thin layer of neural cells that
convert external light into neural signals, known as the retina; In
comparison with other CNS targets, the retina offers some
unique advantages that make it considerably more favorable for
the development of transplantation and stem cell therapies. The
neural sensory retina is a unique structure of the CNS, This
consists of many neuronal types, but the rods are mainly
responsible for detecting low levels of light and the cones can
detect different wave lengths (color) (Diana and Gabriel,
2008).
Other types of neuron connect back to the brain (retinal
ganglion cells) and transmit signals from the rods and cones into
the processing region of the cortex, and finally the visual cortex
where the information is converted into an image. Other
important layers in the eye include the pigment epithelial layer,
which is a pigmented single layer of cells that supports rods and
cones through taking up waste material generated during the
process of detecting light. Loss of function in any of these cells
can result in diminished or lost sight. Retinal degeneration can
occur early or late in life (Svendsen and Ebert, 2008).
The most common cause of blindness in people older than
their 60s is macular degeneration, the degeneration of

140
photoreceptors and epithelial layers. The macula is the part of
the retina that handles detailed vision. Macular degeneration
arises when the retinal pigment epithelial cells, the layer of cells
under the retina, become dysfunctional. This failure leads to
degeneration of the macula, the center of the retina, and
diminishing central vision. The patient slowly loses the ability
to read, recognize faces, and see fine details. There are two
forms of macular degeneration. Dry degeneration comes from
the atrophy of the retinal pigment epithelial layer, which
generates a loss of rod and cone function and, eventually,
blindness. Wet macular degeneration arises from abnormal
growth of blood vessels, which produces a leakage of blood and
protein below the macula, producing irreversible damage to the
photoreceptors (Aoki et al., 2007).
Dry degeneration occurs in 90 % of the age-related cases.
Diabetic retinopathy, which damages blood vessels, is another
leading cause of vision loss in the world, and retinitis
pigmentosa is a hereditary genetic disease that affects the rods
and cones. Many researchers are working on retinal stem cells
of one type or another. It is clear that during development there
are retinal stem cells that can be isolated and grown in culture
for many passages. These seem to lose their ability to make rods
and cones very quickly, however, embryonic stem cells have
been used as a source of retinal neural stem cells, these cells are
more plastic and can give rise to many retinal cell types,
although the exact cocktails of factors needed to produce retinal

141
cells is the subject of intense investigation right now (Diana
and Gabriel, 2008).
The goal is to be able to generate large numbers of retinal
cells that will be useful for both potential transplant therapies
and drug screening, the adult cilliary margin of the human and
mouse eye has been shown to harbor adult retinal stem cells that
appear to grow in culture fairly easily. With regards for treating
eye diseases with stem cells there have been no trials to date
using either fetal or embryonic tissue, although there may be
some plans to do this in the near future. Interestingly, there have
been many pre clinical studies where various types of stem cell
have been injected into the eye and shown to have functional
effects, which of these eventually make it to the clinic remains
speculative at this time (Ofri and Narfström, 2007).
There have been 2 general approaches to the transplantation
of stem cells or stem cell derived cells to rescue the
degenerating retina:
1) Photoreceptor survival promoted by restoring the supportive
functions of the retinal pigment epithelial cells (RPECs) through
subretinal grafts of stem cell derived RPE like cells.
2) Directly replacing lost photoreceptors with transplanted stem
cells and retinal precursor cells coaxed to differentiate and
integrate into the outer nuclear layer of the degenerating retina
(Charles and Alfredo, 2005).
In contrast to therapies for Huntington disease and ALS,
which have already entered clinical trials, and Parkinson

142
disease, for which graft viability and host integration have been
investigated and verified, the potential of these cell based
therapies for retinal degenerative disorders is still being
determined in animal models. A study of subretinal injections of
human ESC derived RPECs rat model of retinal degeneration
(caused by inhibition of RPEC phagocytosis of photoreceptor
outer segments due to mutation in the receptor tyrosine kinase
gene) reported rescue of visual response and acuity assessed
using electroretinography, optomotor acuity thresholds, and
luminance threshold recordings in the superior colliculus (Lund
et al., 2007).
However, there is no direct evidence that the observed
functional improvements were due to rescue of RPEC
phagocytosis rather than a general neuroprotective effect
brought on by the transplanted cells. Indeed, a variety of cell
types have been shown to provide temporary rescuing effects
through secretion of neurotrophic factors when transplanted into
the retina, and the human ESC derived RPECs have been shown
to produce pigment endothelium derived factor, a trophic factor
with protective and morphogenetic effects on photoreceptors.
Furthermore, the grafted cells were observed to aggregate at the
injection site instead of integrating into the host retina with poor
long term cell retention that parallels the decline in rescue
effects at 4 weeks after transplantation (Svendsen and Ebert,
2008).

143
Attempting to rescue the degenerating retina by cellular
replacement of lost photoreceptor neurons presents its own
unique set of challenges. In cases of severe retinal degeneration,
the retina may have already undergone substantial postreceptor
remodeling (in the early stages of these diseases the retina
beyond the photoreceptor layer remains relatively anatomically
and functionally intact). Therefore, grafted stem cells would
need to differentiate into functional photoreceptor neurons and
establish the intricate functional connections with bipolar and
horizontal cells in order to restore the neural retinal circuitry.
Subretinal injections of photoreceptor precursor cells (isolated
from the retina during rod photoreceptor genesis) into mouse
models of retinal degeneration demonstrated cell integration
into the retina, expression of the synapse protein bassoon,
formation of synaptic contacts with bipolar cells identified by
immunostaining with phosphokinase C, positive rod
photoreceptor differentiation based on immunostaining for
proteins involved in the phototransduction pathway (phosducin
and rhodopsin), and improved visual responses assessed with
extracellular field potentials of the retinal ganglion cell layer
and papillary reflex to light stimulation (MacLaren et al.,
2006).
Furthermore, advances in therapeutic cloning could lead to
successful generation of stem cells from adult somatic cells to
facilitate transplantation of autologous cells for treatment of
patients with retinal degeneration. In addition to treating

144
pathological entities that primarily involve the death of
photoreceptors in the outer retina, stem cell therapies are also
being investigated for treating degeneration of the inner retina
specifically, diseases that cause the degeneration of the optic
nerve and retinal ganglion cell (RGCs) (for example, glaucoma,
Leber hereditary optic neuropathy, and ischemic optic
neuropathy) for which there are currently no treatments
available to improve vision once vision loss has occurred.
Regeneration of the optic nerve after injury or disease is
difficult for a number of reasons (Ofri and Narfström 2007).
Optic nerve degeneration is usually accompanied by RGC
apoptosis, whether it is due to elevated intraocular pressure, as
in glaucoma, or excitotoxicity, as in retinal ischemia and
reperfusion injury. Furthermore, as is the case with most CNS
neurons, RGCs in the adult mammalian retina do not have the
ability to reinitiate axonal growth after injury on their own
despite having this capacity during development. Finally,
similar to other areas of the CNS, reactive gliosis and
upregulation of factors such as myelin-associated protein in the
local environment following injury inhibit regrowth of axons
that need to reestablish synaptic connections with the lateral
geniculate nucleus. Overcoming these challenges by using stem
cell therapies would likely require a combination of cell
transplantation and neuroprotective strategies (Lund et al.,
2007).

145
One major advantage of the inner retina and optic nerve is
that molecularly the mechanisms that ultimately result in RGC
apoptosis and optic nerve loss are the same classic mechanisms
described for neurodegenerative events in other parts of the
CNS. As such, both basic and clinical stem cell therapy
investigations that target the inner retina and optic nerve might
provide results that translate to or, at least guide, further
research into similar therapies for other neurodegenerative
disorders. In a different and intriguing line of research
investigators are attempting to use ESCs to essentially grow
eyelike structures rather than replace a lost cell type or provide a
neuroprotective environment (Aoki et al., 2007).
Most of these neurons in N-methyl.D-aspartate treated retina
went on to express ganglion cell specific markers. However,
functional assessment in a more physiologically relevant in vivo
model remains to be done, and the clinical impact of this
approach, if any, remains to be determined. Finally, the adult
retina offers another unique potential source of transplantable
cells (MacLaren et al., 2006).

146
Stem cell therapy for spinal cord
injury
Spinal cord injury is defined as damage to the spinal cord
that results in partial or complete loss of function, whether
temporary or permanent. The three most common causes of
spinal cord injury are direct trauma, compression by disk
herniation, and damage caused by occluded spinal arteries.
Although the range of symptoms that result from a spinal cord
injury is largely determined by the size and location of the
lesion, most patients will at least experience chronic pain
accompanied by lifelong heart and lung complications. Current
therapies focus on physical rehabilitation and counseling to deal
with the emotional frustration of disability, but nothing can be
done to regenerate the spinal cord. As such, research for
treatments targets four main concepts: limiting the damage,
neuroreconstru-ction, stimulating regrowth of neurons, and
retraining neural circuits to restore body functions. Stem cells
have shown great promise and potential for restoration of the
damaged spine (Eftekharpour et al., 2008).
Recent advances in cell replacement therapy for SCI:
Cell death is inevitable after SCI. The adult spinal cord
harbors endogenous stem/progenitor cells, collectively referred
to as NPCs that might be responsible for normal turnover of the

147
cells. However, the proliferative activity of endogenous NPCs is
too limited to support significant self repair after SCI. Thus,
various cellular transplantation strategies have been adopted in
models of SCI. Cell replacement approaches in the setting of
SCI can be used to achieve 2 broad goals: 1) regeneration,
which seeks to replace lost or damaged neurons and induce
axonal regeneration or plasticity; and 2) repair, which seeks to
replace supportive cells such as oligodendrocytes in order to
induce remyelination and prevent progressive myelin loss (Ziv
et al., 2006).
Spinal cord injury leads to interruption of ascending and
descending axonal pathways, loss of neurons and glial cells,
inflammation, and demyelination. Stem cell based therapies
could be used to treat individuals with spinal cord injury in
several ways. First, transplanting stem cell derived spinal
neuroblasts could lead to the replacement of damaged or dead
motor and other neurons. Second, transplanting stem cell
derived OPCs could promote remyelination. Last, transplanting
stem cells modified to release different factors could counteract
detrimental inflammation (figure 27) (Lindvall and Kokaia,
2010).

148
Figure (27): Stem cell based therapies for spinal cord injury (Lindvall andKokaia, 2010).
Human pluripotent stem cells:
The derivation of HESCs in 1998 with their pluripotential
ability to produce all cell types of the human body has sparked
significant interest in their use as therapeutic tools. Considering
the complexity of the nervous tissue and the importance of non
neuronal elements, the pluripotentiality of HESCs makes them
an optimal candidate that can potentially differentiate into
different cell types for the reformation of cellular networks.
Using an excitotoxicity brain injury models, it has been shown
that transplanted hESCs express immunological labeling for
vascular endothelial cells, glia, and different neuronal subtypes
with morphologically normal synapses (Eftekharpour et al.,
2008).
The generation of autologous pluripotent stem cells from
skin cells has been reported. By using viral gene transfer into
somatic cells of adult humans, pluripotent cells can be generated

149
that have similar immunological characteristics to the donor.
These pluripotent cells are similar to ESCs in that they retain the
developmental potential to differentiate into any cell type. This
discovery will obviate the ethical issues surrounding the source
of ESCs and eliminate the need for immunological matching
between the host and donor. However, several issues including
the potential mutation of these cells after integration of the
carrier viral gene into the host DNA must be resolved before
these cells can be used in clinical trials (Lindvall and Kokaia,
2010).
Fetal stem cells:
Fetal tissue grafts were tested in human patients with CNS
disorders and Parkinson disease as early as the 1970s. Some
reports have suggested that fetal derived NPCs can differentiate
into neurons and oligodendrocytes after transplantation into a
contusion SCI model with some improved behavioral outcome.
The transplantation of olfactory ensheathing cell (OECs) from
fetal sources, has gained considerable media attention; however,
the limited scientific data from this study severely limit
enthusiasm for this procedure as a therapeutic option in patients
(Raisman and Li, 2007).
Banking of umbilical cord blood cells, another type of fetal
stem cells, for the cure of immunological and hematological
disorders as well as a potential source for stem cell based
therapeutic approaches, has expanded rapidly. The
multipotentiality of these cells to produce the main cell types of

150
nervous tissue has been recently reported. These cells have also
been used in experimental SCI models and were shown to
improve functional recovery due to neurotrophic function or
increased myelination. However, further work must be done to
fully characterize the extent of functional recovery induced by
umbilical cord blood cells and to further delineate their
mechanisms of action (Lindvall and Kokaia, 2010).
Adult derived stem and progenitor cells:
Adult derived stem/progenitor cells have been identified in
many tissues including those from skin, eye, pancreas, brain,
and spinal cord. These somatic cells have an extensive capacity
for self renewal and the multipotentiality to produce all the
major cell types of their specialized tissue of origin. However,
experimental evidence has suggested the possibility of
transdifferentiation of these cells into other cell types, when
transplanted ectopically into other tissues such as the spinal
cord. Although this topic is the subject of active research, in this
review we focus on selected reports that have shown some
beneficial outcome when adult cells were transplanted into an
SCI model (Yiu and He, 2006).
Schwann cells and OECs:
The PNS myelinating Schwann cells would seem to be an
obvious substitute when considering a cell based remyelination
strategy. Interestingly, infiltration of endogenous Schwann cells
at the site of injury and their contribution to spontaneous
myelination after SCI is well documented, although the impact

151
of such schwannosis which is associated with increased CSPG
formation and may have deleterious effects on regeneration is
unclear and could even promote neuropathic pain. Schwann
cells can be easily harvested from the patient’s peripheral
nerves, and a relatively short time is required to produce
clinically suitable material for autologous transplantation (Rishi
et al., 2009).
After PNS injury, Schwann cells provide a growth
promoting environment for regenerating axons through
secretion of neurotrophic factors1 and extracellular matrix
elements and ultimately myelinate the axons. Several groups
have studied transplantation of Schwann cells after SCI and
have shown successful integration of these cells in the host
tissue with some regeneration of sensory axons and myelination
of these axons associated with modest improvement of hind
limb function. A poor survival rate has been reported for these
cells when transplanted into the epicenter cavity, although the
endogenous Schwann cells will fill up the cavity (Raisman and
Li, 2007).
The therapeutic effects of Schwann cells have been shown to
be enhanced when these cells are transplanted with their
phenotypically similar cells, OECs. These are peripheral glial
cells that support the growth of newly generated axons from the
olfactory neurons to the olfactory bulb but do not myelinate the
olfactory axons in their native environment. However, some
authors believe that OECs are not only involved in axonal

152
regeneration but also in remyelination after SCI or in a
demyelinating disease. In contrast, others have demonstrated a
lack of myelination in axons associated with OECs transplanted
after SCI. The controversial myelinating ability of OECs is
affected by several experimental variables including the age of
the donor and the isolation methods used (Yiu and He, 2006).
Mesenchymal stem cells:
The ability of these cells to differentiate into neural cells is
controversial; therefore, it has been suggested that the functional
improvement might be effected through neuroprotective
pathways. The in vivo expression of neural tissue markers
nestin and GFAP in MSCs or their neuron like morphological
characteristics under certain tissueculture conditions as
evidence of the transdifferentiation ability of MSCs to neural
tissue cells. Nonetheless, nestin and GFAP are not specific
markers for nervous tissue and can be found in mesodermal
tissues such as muscle and cartilage (Ziv et al., 2006).
The reported neuron like morphological characterristics of
MSCs after tissue culture manipulation can best be described as
tissue culture artifact and has not been supported by functional
properties associated with normal neurons such as
electrophysiological assessment. The possibility of MSC
transdifferentiation into neuronal progeny remains to be
investigated. Alternatively the functional benefits of MSC
transplantation in CNS injuries can be explained by their ability
to provide the host tissue with growth factors or modulate the

153
host immune system. These cells can be used in combinatorial
therapies as tools for growth factor delivery. Transplantation of
bone marrow MSCs, engineered to deliver neurotrophins, can
induce axonal growth through the glial scar in a model of
chronic SCI in rats; however, it seems that these approaches do
not result in functional recovery after chronic SCI, indicating
the lack of appropriate synapse formation by the regenerated
axons with the host tissue (Lindvall and Kokaia, 2010).
Adult derived NPCs:
Adult NPCs contribute to neurogenesis and gliogenesis in
some regions of the CNS in various species including humans.
These tissue specific somatic stem cells are located in the
periventricular regions and in white matter tissue throughout the
brain and spinal cord and can differentiate into all neural cell
types under defined conditions. Using selective in vitro
expansion and cell sorting based on antigenic properties and
molecular techniques, these cells can be directly derived from
the adult tissue; however, so far no specific marker has been
identified for human adult NPCs (Eftekharpour et al., 2008).
Recently, using a combined therapeutic strategy, we have
shown the beneficial effects of adult NPC transplantation in a
clinically relevant model of SCI in rats. This compressive model
consists of a modified aneurysm clip that delivers a calibrated
closing force. The 23-g clip used in our studies produces a
moderately severe SCI with central cavitation and loss of 80%
of axons in the spinal cord white matter, demyelination of the

154
surviving axons in the residual subpial rim, and spastic
paraparesis. Therefore, this model is of translational relevance
to the majority of patients with chronic SCI (Lindvall and
Kokaia, 2010).
The adult NPCs were harvested from the subventricular zone
of adult mice that express the yellow fluorescence protein (YFP)
gene, allowing for easy tracking of transplanted cells. Cells
were clonally driven in vitro in the presence of growth factors.
These free floating colonies (called neurospheres) were
composed of 1% neural stem cells and 99% progenitor cells.
The dissociated adult NPCs transplanted into the areas rostral
and caudal to the injury epicenter in a delayed fashion at 2 or 8
weeks post injury to represent 2 clinically relevant phases in
SCI pathophysiology, the subacute and chronic stages,
respectively. We adopted a novel post transplantation delivery
of epidermal growth factor, basic fibroblast growth factor, and
platelet derived growth factor AA approach aimed at:
1) Promoting the survival of the grafted adult NPCs.
2) In vivo differentiation of undifferentiated NPCs towards
an oligodendroglial lineage (Rishi et al., 2009).
Transplantation of adult NPCs into the subacutely
injured spinal cord:
When transplantation was delayed to 2 weeks after injury, a
substantial survival rate of 40% of the transplanted cells was
observed (figure 28).

155
Figure (28): Photomicrographs demonstrating that transplanted adult NPCssuccessfully survive, migrate, and integrate with the injured spinal cord.Confocal images from a longitudinal section of injured spinal cord takenfrom the dorsal spinal cord of an injured rat that received cell transplants 2weeks post injury. The low magnification image (A) shows a substantialsurvival of YFP-NPCs within the injured spinal cord at 8 weeks posttransplantation. Grafted YFP-NPCs (green) were dispersed along therostrocaudal axis of the spinal cord approximately 5 mm from theimplantation sites (asterisks). The YFP-NPCs also migrated to thecontralateral side of the spinal cord to a lesser extent. Double labeling withthe neuronal marker bIII tubulin (Tuj1) showed that YFP-NPCs residepredominantly in the white matter (WM) area. GM = gray matter(Eftekharpour et al., 2008).
The multipotent capability of the subventricularly derived
adult NPCs to differentiate into the 3 main cell types of the
nervous tissue has been shown in our experiments as well as in
previous reports in vitro, but the transplanted NPCs were mainly
driven towards a glial fate. The lack of neuronal differentiation

156
by engrafted NPCs in the adult spinal cord has been reported
repeatedly by different groups of authors (Ziv et al., 2006).
The tendency of NPCs toward glial differentiation after
transplantation may also reflect the intrinsic properties of the
spinal cord that favor glial differentiation. This may be
attributed to potential inhibitory or lack of promoting factors for
neurogenesis by NPCs in different CNS tissues and needs to be
taken into consideration when cell therapy strategies are
designed for neuronal replacement. Because loss of
oligodendrocytes and myelin is a major contributor to white
matter damage after trauma, it is rather intriguing to speculate
that the propensity of adult NPCs to form oligodendrocytes after
transplantation into the adult injured spinal cord may reflect the
influence of the host tissue to replenish a particular cell type to
maximize repair. However, whether demyelinated axons exert
such effects on NPC integration and differentiation remains to
be investigated (Eftekharpour et al., 2008).
Myelin is critical to the precise molecular organization of
axons. The nodal distribution of Na+ channels, paranodal
preference of contactin associated protein, and juxtaparanodal
adherence of K+ channels is closely related to myelin and is
disrupted following demyelination (Sasaki et al., 2007).
However, the potential contribution of endogenous
myelinating cells in these models cannot be ruled out unless
mutant dysmyelinated models such as neonatal myelin deficient
rats or shiverer mice are used. Beneficial structural and

157
functional effects of neural stem cells in genetic mutant models
indicate that long term molecular abnormalities can be
successfully reversed in the adult dysmyelinated CNS axons.
Although neuroanatomical assessment of myelination is a
common approach determining the success of cell therapy
approaches, direct physiological assessment to investigate the
functionality of the new myelin has been overlooked in
literature. Using in vivo spinal cord evoked potential recordings,
we observed that adult NPC induced myelination promoted
enhanced spinal cord axonal conduction in the shiverer mice, as
evidenced by increased amplitude, reduced latency, and
enhanced estimated conduction velocity.Although
remyelination has been shown to be involved in adult NPC
mediated recovery after SCI, we cannot exclude other potential
protective/trophic effects of adult NPCs. It is possible that the
transplanted adult NPCs may increase axonal sprouting and
plasticity that could be responsible for some of the observed
locomotor recovery (Rishi et al., 2009).
Functional locomotor recovery following subacute
transplantation of adult NPCs has been also reported by other
investigators. However, recent reports have noted an increase in
pain sensation, known as allodynia, which was observed when
NPCs chiefly differentiated into astrocytes. Allodynia was
diminished after genetic modification of the naive adult NPCs to
enhance their differentiation towards oligodendrocyte lineage.
Of note, there are no observed changes in pain sensation in

158
experiments, which can be explained by low rates of astrocytic
differentiation by the transplanted adult NPCs in our
transplantation protocol. However, these reports suggest that the
therapeutic evaluation of any intervention for the repair of SCI
should include detailed sensory and motor assessments
(Lindvall and Kokaia, 2010).
Transplantation of NPCs into chronic spinal cord lesions:
Chronic SCI continues to pose a major challenge for
patients, physicians and translational scientists. To date, limited
success has been achieved in applying regenerative technologies
to the complex pathobiology of chronic SCI. This may be
attributed to the environmental restrictions present in
chronically injured spinal cord including glial scarring and
syrinx formation. In our experience with adult NPC therapy for
the repair of SCI, we have learned that the chronically injured
spinal cord environment is not as hospitable as the subacutely
injured tissue for the transplanted cells. Grafted adult NPCs
mostly failed to survive after transplantation into the chronically
injured spinal cord (Chen et al., 2004).
These results suggest the presence of inhibitory components
or the lack of promoters such as growth factors in the
environment of chronically injured spinal cord. Considering the
complexity of chronic SCI pathobiology, it is important to adopt
a multifactorial approach to overcome the environmental
restrictions present in the chronically injured spinal cord. Such

159
strategies will need to overcome the inhibitory effects for the
glial scar, endogenous inhibitory molecules such as Nogo and
Rho, and the presence of post traumatic cyst (Sasaki et al.,
2007).

160
Stem cell therapy for Peripheral nerves
Injuries
Peripheral nerves Injuries are common and debilitating,
affecting 2.8% of trauma patients and resulting in considerable
long term disability. Although peripheral nerves do exhibit the
potential to regenerate axons and re innervate the end organ,
outcome following severe nerve injury, even after repair,
remains relatively poor. This is likely because of the extensive
injury zone that prevents axon outgrowth. Even if outgrowth
does occur, a relatively slow growth rate of regeneration results
in prolonged denervation of the distal nerve (Metwally, 2009).
Whereas denervated Schwann cells (SCs) are key players in
the early regenerative success of peripheral nerves, protracted
loss of axonal contact renders Schwann cells unreceptive for
axonal regeneration. The assumption has been that peripheral
nerve injuries recover, given the observation of spontaneous
axonal regeneration following insult. While this capacity for
regeneration is higher than that of the central nervous system,
complete recovery is fairly infrequent, misdirected, or
associated with debilitating neuropathic pain. In fact,
satisfactory results only tend to occur following relatively minor
injuries, such as neurapraxia or axonotmesis (Keilhoff et al.,
2006).

161
Nerve transection is associated with notoriously poor
outgrowth compared with other injuries, particularly when the
distance between injury and target is long. Poor outcome from
peripheral nerve injury is especially evident when repair is
performed after a temporal delay, occurring frequently in
clinical practice. Due to the nature of most nerve injuries where
the nerve is left in physical continuity, the propensity for
spontaneous recovery is not immediately known. As such,
surgical repair is significantly delayed in a great number of
cases. Even patients undergoing immediate nerve repair are
subject to a lengthy denervation of the distal nerve as a result of
the low rate of regeneration (~ 1 mm/day in humans) and the
long regeneration distances required to reach the end organ
(Metwally, 2009).
Elongation of regenerating axons is initially supported by
resident Schwann cells that undergo a phenotypic change from
myelinating to growth supportive following initial denervation.
Proliferating Schwann cells are a rich source of neurotrophins,
cell adhesion molecules, and cytokines that support axonal
regeneration and recruit further cells into the injury site. Unless
axonal contact is reestablished in a timely fashion, however, this
growth supportive environment is not maintained (Cui et al.,
2008).
Denervated Schwann cells progressively lose their ability to
express regeneration assisting genes and in effect become
"turned off." As the capacity of the denervated distal nerve to

162
support axonal regeneration is highly dependent on proliferating
Schwann cells within the basal lamina tubes that guide
elongating axons to their denervated target, this loss of vitality
and functionality in distal Schwann cells directly translates to
poor muscle reinnervation outcomes. Although placement of
interposed autologous nerve grafts offers a cell rich material
through which axons can regenerate, their use is not ideal
because of donor site morbidity, lack of donor tissue
availability, and nonspecific regeneration. New advances in
tissue engineering have introduced synthetic nerve guide
conduits that are capable of bridging small defects in peripheral
nerves (up to ~ 3 cm in humans), but their relatively inert
microenvironment reduces their value for larger or more chronic
injuries (Bellamkonda, 2006).
It appears that combined approaches with cells or trophic
factors within synthetic tubes may extend their functionality.
Indeed, delivery of SCs in a variety of repair paradigms has
been successful in promoting regeneration and remyelination of
the injured spinal cord and peripheral nerve. However, human
SCs must be derived from invasive nerve biopsies in sufficient
numbers for regeneration and are only available after a lengthy
expansion time in vitro. Therefore, several groups have turned
their attention to identifying more accessible sources of SCs like
cells for transplant therapies (Metwally, 2009).

163
Considerations for optimizing stem cell therapy for
peripheral nerve repair:
Number and method of stem cell delivery:
Although often not reported, the number of cells delivered to
nerve injuries in animal models varies considerably between
studies. While some have used as few as 4 × 103 cells, others
have transplanted 2 × 107 cells, but there has often been little
explanation for the selection of cell numbers in these studies. It
is admittedly difficult to compare the number of cells delivered
in widely different repair paradigms, but it is fair to state that
there are likely an ideal number of cells that should be
determined for each cell type or repair scenario. Just as too few
cells may not translate to a therapeutic effect, delivery of too
many cells may also have detrimental results. This was
exemplified beautifully by a study using transplanted Schwann
cells delivered in 10 mm nerve gap (Cui et al., 2008).
Optimization strategies should take the number of delivered
cells into account. Similarly, the way in which cells are
delivered to the injury site has varied between studies, ranging
from direct microinjection, suspension within artificial tubes,
and seeding within devitalized muscle or nerve grafts. Although
the choice of stem cell delivery method may depend on the type
and extent of nerve injury in question, it may be optimized by
providing transplanted cells an environment that will favor their
survival and integration, such as within structured fibers or
biomatrices (Keilhoff et al., 2006).

164
Differentiation state of delivered stem cells:
Part of the appeal of using precursor or stem cells for
supplementing peripheral nerve repair is their capacity for self
renewal, such that it is possible to deliver large numbers of
dividing cells to the injury site. By delivering stem cells into the
injured nerve in a naive state, this proliferative capacity is
maintained, and it is expected that cells will be prompted by the
microenvironment to differentiate into the required cell type. In
vitro studies have demonstrated that neural stem/progenitor cells
in coculture with cells from the nervous system will take on a
phenotype similar to their partner tissue’s origin: dorsal root
ganglion cultures will induce a peripheral neuron/Schwann
cell/smooth muscle phenotype, and a cerebellar feeder layer will
induce differentiation into CNS neurons (Brannvall et al.,
2008).
Nevertheless, incidence of differentiation from naive
precursor cells within the peripheral nerve is rather low in many
cases. Choosing to pre-differentiate stem cells toward a desired
phenotype prior to delivery into the repair site may be an
effective strategy to ensure a more precise and complete
therapeutic effect. It may be that cells at later developmental
stages (vs embryonic stem cells, for example) possess more
mature intrinsic molecular programs to direct them to their
target destination (Bellamkonda, 2006).

165
Because it is well known that mature SCs survive
denervation events by secreting autocrine factors such as insulin
like growth factor, neurotrophin-3, and plant derived growth
factor BB, might an appropriately differentiated stem cell also
possess similar machinery for self preservation and thus be an
ideal candidate for supplementing the injured peripheral nerve?
We have found that SKPCs, when injected as naive sphere
forming cells, do differentiate into GFAP positive SCs in
response to cues found in the local environment of the injured
peripheral nerve. However, long term survival and maintenance
of SCs markers is greatly improved by pre-differentiating the
cells to SCs phenotype prior to delivery. On the other hand,
others have reported that allowing stem cells to differentiate
before delivery accelerates post transplant cell death, perhaps
owing to increased expression of major histocompatibility
complex antigens or reduced proliferation rates (Svendsen and
Ebert, 2008).
In addition to survival of stem cells, their effect on
surrounding tissues may be modified based on their level of
differentiation prior to transplantation. For example, when naive
adult neural stem cells were injected into a lesioned spinal cord,
the resulting aberrant sprouting resulted in profound allodynia.
If gliogenesis in these cells was suppressed by prior treatment
with neurogenin-2, there was an overall greater functional
improvement (Brannvall et al., 2008).

166
Improving survival of transplanted stem cells:
Whether due to technical challenges or oversight, it is an
unfortunate reality that survival of stem cells delivered to nerve
injury sites is reported only infrequently. When quantified,
precursor cells have shown between 0.5 and 38% survival,
depending on evaluation time point and cell type. When naive
SKPCs delivered into an acutely injured nerve, survival after 2
weeks was ~ 10.5%, whereas when delivered into a nerve that
had been previously chronically denervated, the number of
detected skin derived precursor cells (SKPCs) decreased to
5.8%. Seeing that 78% of the surviving stem cells in the chronic
model had differentiated into GFAP-positive Schwann cells, we
next used pre-differentiated (Schwann cell like) SKPCs and
found that we could increase survival to ≥ 8% (Pan et al.,
2007).
Without quantification of survival in stem cell
transplantation experiments, it is difficult to determine whether
they are being retained long enough and in enough numbers to
confer a sufficient benefit to regeneration. The danger of
exogenous cell therapy is of course cell death caused by
immune system attack. Although some authors have reported
considerable phagocytosis of transplanted stem cells, this may
be due to species/strain mismatching of donor and recipient, as
many others have not observed this trend. In fact, we have
observed a highly interesting pattern of surviving transplanted

167
SKPCs that are spatially separate from phagocytic ED-1
positive macrophages (Keilhoff et al., 2006).
If a minimum survival time of stem cells is indeed required
to observe a therapeutic effect, strategies should be devised to
increase the amount of time cells remain in grafted regions.
Survival and effectiveness of transplanted cells can be improved
by ex vivo genetic manipulation or concomitant delivery of
protective agents or trophic factors. Pan and colleagues found
that administration of granulocyte colony stimulating factor to
animals receiving transplants of amniotic fluid mesenchymal
stem cells not only improved survival of transplanted cells but
also augmented nerve regeneration over that of a primarily cell
based approach. Additionally, differences in the material in
which stem cells are delivered have demonstrated varying
capacities to support long term cell survival. Finally,
immunosuppressive regimens, especially in the light of
interspecific transplants may protect stem cells from premature
clearance from the nerve injury site (figure 29) (Metwally,
2009).

168
Figure (29): Confocal image. The SKPC Schwann cells are not
immediately cleared by host immune system. Eight weeks following
injection into an initially decellularized (by repetitive freeze-thawing) nerve
graft bridging a 12-mm defect created in the rodent sciatic nerve, SKPC
Schwann cells (red), and ED-1-positive macrophages (green) are spatially
segregated within the longitudinal extent of the nerve graft. The finding that
there is very little colabeling of SKPC–Schwann cells with ED-1 positive
macrophages suggests that transplanted cells are not phagocytosed in any
large quantity within the host nerve, Original magnification ×400
(Metwally, 2009).
Influence of final stem cell phenotype on regenerative
success:
As with survival, differentiation of stem cells within the
injury site has demonstrated a mixed correlation to therapeutic
effect. Some studies have demonstrated a need for
differentiation to glial phenotype to observe adequate
regeneration of neural tissue, and others have shown
improvement with little to no differentiation of stem cells at the

169
assessment end point. Furthermore, the glial differentiation of
transplanted stem cells within the injured peripheral nerve has
tended to vary between studies, the difference in repair
paradigms (devitalized muscle graft versus crush injured nerve)
may explain the disparity in the ability of these cells to
differentiate, outlining the need for careful consideration of the
method of delivering stem cells to the injury site (Keilhoff et al.,
2006).
In the cases in which adequate regeneration and
improvement of outcomes occurs without SCs differentia-tion
of transplanted precursors, it may be that the cells are
supporting axonal growth by additional mechanisms such as the
production of cytokines or harnessing the inflammatory
response. Although C17.2 neural stem cells show little
differentiation into SCs phenotype in the chronically denervated
peripheral nerve, their secretion of various matrix
metalloproteinases, capable of breaking down growth inhibiting
chondroitin sulfate proteoglycans, likely underlies their ability
to elicit superior regeneration (Pan et al., 2007).
Careful examination of ultimate cell fate with correlation to
functional outcome is strongly recommended for future
precursor transplant studies and will be required to fully answer
this question for each cell type and repair strategy. It may be
that, at least for some precursor cell types, there is a minimum
level of differentiation to S100β/MBP/GFAP-positive SCs that
is required for acceptable regeneration outcomes. If this is the

170
case, effectiveness of precursor transplantation could be
improved using technology that exists to directly alter the
regenerative microenvironment by continuous delivery of
neuregulins, forskolin, or other differentiation-promoting factors
(Svendsen and Ebert, 2008).
Methods for tracking fate of transplanted stem cells:
Stem cell transplantation for peripheral nerve repair should
give careful thought on strategies to track the fate of
transplanted cells over time. There is often little importance
placed on prelabeling cells prior to delivery into the injured
nerve, and as such authors cannot comment on the mechanism
of any advantage conferred by cell therapy. Others have used
labeling techniques that are not sufficiently robust or long
lasting to be detected at the study end points. Chemical markers
such as bisbenzimide and PKH26 have been used to label SCs
delivered to peripheral nerve injuries, but their usefulness is
limited to the short term and may in fact affect the viability and
phenotype of transplanted cells (Bellamkonda, 2006).
Genetic labeling with either fluorescent protein such as GFP
is increasingly popular and appears to be a relatively long
lasting method that is not deleterious to transduced cells. We
have used the lipophilic carbocyanine derivative Cell T racker
CM-DiI (Molecular Probes) to reliably label SKPCs within a
variety of nerve injury models with no dilution or loss of signal
for ≥ 10 weeks following transplantation. These dyes have the

171
advantage of being technically simple to use, rapid, and resistant
to leakage to nearby cells. Emerging technologies such as
quantum dots offer an exciting alternative to traditional cell
labeling methods. These nanoparticles are available in a wide
range of photostable colors and are resistant to chemical and
metabolic degradation, making them ideal for use in long-term
fate tracking of transplanted stem cells (Brannvall et al., 2008).

172
Summary
Stem cells are emerging as one of the most exciting new
areas of neuroscience, not only in terms of revealing insights
into normal development, but also as a therapeutic agent for a
range of neurological diseases. In both of these aspects, they
will affect neurological practice by providing insights into
mechanisms of disease as well as curative cell therapies.
Cell therapy is in fact, a type of organ transplant which has
also been referred to as "xenotransplant therapy", "cellular
suspensions", "glandular therapy" or "fresh cell therapy". The
procedure involves the injection of either whole fetal xenogenic
(animal) cells (e.g. from sheep, cows, pigs, and sharks) or cell
extracts from human tissue. The latter is known as autologous
cell therapy if the cells are extracted from and transplanted back
into the same patient. Several different types of cells can be
administrated simultaneously as embryonic, fetal (derived from
fetal tissues), umbilical cord and adult (somatic) stem cells
including haemopoietic, mesenchymal, hepatic, neuronal,
epidermal and pancreatic stem cells.
The types of cells that are administered correspond in some
way with the organ or tissue in the patient that is failing. No one
knows exactly how cell therapy works, but it was claimed that
the injected cells travel to the similar organ from which they
were taken to revitalize and stimulate that organ's function and

173
regenerate its cellular structure. In other words, the cells are not
species specific, but only organ specific.
Replacing the worn out or injured cells by functional cells to
restore the normal function of the tissues or organs is the
underlying principle of CRT, otherwise also called as
regenerative or restorative medicine. It is hoped that the organs
or tissues treated by this approach can perform their normal
function more efficiently than the ones treated by conventional
therapies like transplantation and pharmocological therapy.
Cell therapy has been used successfully to rebuild damaged
cartilage in joints, repair spinal cord injuries, strengthen a
weakened immune system, treat autoimmune diseases, Acquired
Immunodeficiency Syndrome (AIDS) and help patients with
neurological disorders such as Parkinson's disease (PD), stroke
and Alzheimer's disease and further uses have shown positive
results in the treatment of a wide range of chronic conditions
such as arteriosclerosis, congenital defects, and, myocardial
infarction, PD , diabetes which are due to the progressive (acute
/ chronic) loss of functional cells due to disease or injury to the
cells or ageing.
Cell therapy for PD has significantly advanced in recent
years. Open label clinical trials have provided proof of principle
that transplantation of fetal DA neurons can improve patients’
neurological symptoms. Stem cell based approaches could be
used to provide therapeutic benefits by implanting modified
stem cells to release growth factors which protect existing

174
neurons and transplanting stem cell derived DA neuron
precursors into the putamen.
Compared with neurodegenerative disease, stroke possesses
special conditions that impact the potential success of
transplantation to enhance neurological recovery including; the
anatomy, time of the stroke, the vascular supply, site of
implantation and type of patients enrolled in clinical trials.
Another disease of interest is demyelinating disease such as
multiple sclerosis which presents particular and serious
problems to those attempting to develop cell-based therapies:
the occurrence of innumerable lesions scattered throughout the
CNS, axon loss, astrocytosis, and a continuing inflammatory
process, to name but a few. Nevertheless, the limited and
relatively focused nature of damage to oligodendrocytes and
myelin, at least in early disease, the large body of available
knowledge concerning the biology of oligodendrocytes, and the
success of experimental myelin repair, have allowed cautious
optimism that therapies may be possible.
Amyotrophic lateral sclerosis is characterized by progressive
dysfunction and degeneration of motor neurons in cerebral
cortex, brain stem and spinal cord. To have long-term value,
stem cell therapy must restore function of both upper and lower
motor neurons. Successful replacement of cortical motor
neurons requires not only re-establishment of spinal cord
connections but also precise functional integration of the new
neurons into cortical circuitries.

175
Also, there have been several different experimental
approaches to cell therapy for Muscular dystrophies which are a
heterogeneous group of inherited disorders clinically manifested
by progressive muscle weakness and wasting and, in some
cases, death. Although much of the molecular genetics of
muscular dystrophies have been elucidated, there is still no
effective cure for any of them.
Huntington’s disease is an autosomal dominant
neurodegenerative disorder associated with progressive cell loss
and atrophy predominantly in the striatum and neocortex, Cell
therapy in Huntington's disease aims at restoring brain function
by replacing these neurons.
Alzheimer’s disease is a neurodegenerative disorder
characterized by progressive dementia with memory loss and
altered behavior including paranoia, delusions, depression, and
impairments of attention, perception, reasoning and
comportment, Stem cell-based therapy could be used to prevent
progression of the disease by transplanting stem cells modified
to release growth factors. Alternatively, compounds and/or
antibodies could be infused to restore impaired hippocampal
neurogenesis.
Cerebral palsy consists of a heterogeneous group of non
progressive clinical syndromes that are characterized by motor
and postural dysfunction, Stem cell therapy is used to induce
neuronal regeneration, The type of improvements reported
include: decreased spasticity; better coordination; improved

176
motor function, improved posture stability; better cognition
resulting in communication improvements.
Autism spectrum disorders are a group of biologically based
neurodevelopmental disorders characterized by impairments in
three major domains: socialization, communication, and
behavior, stem cells induce angiogenesis and neurogenesis in
areas of cerebral hypoperfusion, their ability to constitutively
secrete immune inhibitory factors leads to Immune modulation,
(in autism, several immunological abnormalities have been
detected both in the peripheral and the central nervous systems).
Neural retinal degeneration is a common cause of blindness
in people older than 60 years, stem cell transplantation rescue
the degenerating retina by supportive functions of the retinal
pigment epithelial cells and directly replacing lost
photoreceptors and retinal precursor cells.
Spinal cord injury leads to interruption of ascending and
descending axonal pathways, loss of neurons and glial cells,
inflammation, and demyelination. Stem cell-based therapies
could be used to treat individuals with spinal cord injury in
several ways. First, transplanting stem cell-derived spinal
neuroblasts could lead to the replacement of damaged or dead
motor and other neurons. Second, transplanting stem cell-
derived OPCs could promote remyelination. Last, transplanting
stem cells modified to release different factors could counteract
detrimental inflammation.

177
Peripheral nerves Injuries are common and debilitating,
affecting 2.8% of trauma patients and resulting in considerable
long term disability, stem cells delivery in a variety of repair
paradigms has been successful in promoting regeneration and
remyelination of the injured peripheral nerve.
Despite the fact that there is much basic work left to do and
many fundamental questions still to be answered, researchers
are hopeful that effective repair for once hopeless clinical
problems may eventually be achieved. Whether through
developing replacement cells or activating the body's own stem
cells in vivo, research on the use of stem cells for
neurodegenerative disorders is a rapidly advancing and
promising field.

178
References
Abayomi Sh: Parkinson’s disease - A review. Aferican Journal of Neurologicl.
Science 2002; 21: 30-37.
Abdallah BM and Kassem M: Human mesenchymal stem cells from basic
biology to clinical applications. Gene Therap 2008; 15: 109-116.
Andeson DJ, Gage FH and Weissman IL: Can stem cells cross lineage
boundaries? Nat. Med 2001; 7: 393-95.
Aoki H, Hara A, Niwa M, Motohashi T and Kunisada T: An in vitro mouse
model for retinal ganglion cell replacement therapy using eye-like
structures differentiated from ES cells. Exp. Eye Res 2007; 84: 868–75.
Augustyn M, Patterson MC and Bridgemohan C: Diagnosis of autism
spectrum disorders. Exp. Brain Res 2011; 103: 355–71.
Avasthe S, Srivastava R, Singh A and Srivastava M: Stem cell past, present
and future, Internet Journal of medical update 2008; 3: 22-30.
Bachoud-Lévi AC, Gaura V, Brugières P, Lefaucheur JP and Boissé MF:
Effect of fetal neural transplants in patients with Huntington’s disease 6
years after surgery: a long-term follow up study. Lancet Neurol 2006; 5:
303–9.
Battler A and Leor J: Stem cell and Gene-Based Therapy; frontiers in
regenerative medicine, Section 2 Neuro 2006: 123-32.
Bellamkonda RV: Peripheral nerve regeneration: an opinion on channels,
scaffolds and anisotropy. Biomaterials 2006; 27: 3515–18.

179
Belmadani A, Tran PB, Ren D and Miller RJ: Chemokines regulate the
migration of neural progenitors to sites of neuro inflammation. J
Neurosci 2009; 26: 3182–91.
Bianco P, Riminucci M, Gronthos S and Robey PG: Bone marrow stromal
stem cells: Nature, biology and potential applications. Stem Cell 2005;
19: 180-92.
Bongso A, Lee EH: Stem cells: their definition, classification and sources. In:
Bongso A, Lee EH, eds. Stem cells: from bench to bed side. Singapore:
World Scientific Publishing 2005; 1: 79-85.
Brannvall K, Corell M and Forsberg N: Environmental cues from CNS, PNS,
and ENS cells regulate CNS progenitor differentiation. Neuroreport
2008; 19: 1283–89.
Caplan LR: Clinical diagnosis of stroke subtypes. Neuroscience 2011; 106:
201-16.
Charles YL and Alfredo QH: Stem cell opportunities for neurosurgery.
Neurosurg Focus 2005; Vol 19: 289-304.
Charron D, Boissel CS and Al-Daccak RJ: A Challenge of Stem Cell
Therapy. J. Medical Sci. 2009; 2: 130-38.
Chen J, Magavi S and Macklis J: Neurogenesis of corticospinal motor
neurons extending spinal projections in adult mice. Proc. Nat Acad Sci
U S A 2004; 101: 16357-62.
Christopher T S: Stem cells; new frontiers of ethics, law, and policy.
Neurosurg Focus 2008; 24 (3&4): E23.
Claire D, Clelland B, Roger A and Colin W: Cell therapy in Huntington
disease Neurosurg Focus 2008; 24 (3&4): E8, 1-12.

180
Cohen M, Lahat E and Bistritzer T: Evidence based review of bone strength
in children and youth with cerebral palsy. J Child Neurol 2009; 24: 959.
Crosta P: What are stem cells? Information section for Medical News Today.
The National Institutes of Health resource for stem cell research,
Wikipedia 2010; 147: 869-78.
Cui L, Jun J, Ling W, Xin Z and Joy S: Transplantation of Embryonic Stem
Cells Improves Nerve Repair and Functional Recovery After Severe
Sciatic Nerve Axotomy in Rats. Stem Cells 2008; 26: 1356 -65.
D’Antona G, Brocca L, Pansarasa O, Rinaldi C and Tupler R: Structural and
functional alterations of muscle fibres in the novel mouse model of
facioscapulohumeral muscular dystrophy. J Physiol 2007; 584: 997–
1009.
Darras BT: Treatment of Duchenne and Becker muscular dystrophy. Nat Med
2010; 8: 963–70.
David D: The present and future of stem cell therapy, J. Vincent’s Sci. 2009; 3:
148-52.
Deasy BM, Lu A, Sun B, Urish KL and Cao B: A role for cell sex in stem cell
mediated skeletal muscle regeneration: female cells have higher muscle
regeneration efficiency. J Cell Biol 2007; 177: 73–86.
Diana YU and Gabriel AS: Stem cell sources and therapeutic approaches for
central nervous system and neural retinal disorders. Neurosurg Focus
2008; 24 (3&4): E10: 1-14.
Eftekharpour E, Abdolrezaee SK and Fehlings MG: Current status of
experimental cell replacement approaches to spinal cord injury,
Neurosurg. Focus 2008; 24: 1-13.

181
Emerson C, Silva GV and Fernandes MR: Regenerative Medicine,
Application in Cardiovascular Diseases, Hematopoietic Stem Cell
Transplantation 2008; 3: 713-14.
Ermini FV: Neural stem cell biology and neurogenisis in mouse models of
aging and Alzheimer disease, Philosophisch Naturwissenschaftlichen
Fakultät der Universität Basel 2006; 2: 14-18.
Golbe L: Genetics of Parkinson’s disease. In: Ellenberg JH, Koller WC,
Langston KW. Eds. Etiology of Parkinson’s disease. New York: Marcel
Dekker 2003; 19: 115-40.
Goldman S and Windrem M: "Cell replacement therapy in neurological
disease". Philos Trans R Soc Lond B Biol Sci 2006; 361: 1463–75.
Grabowski TJ: Clinical manifestations and diagnosis of Alzheimer disease.
Arch Neurol 2010; 62: 860-64.
Gronthos S, Hay S, Shi S, Graves S and Simmons P: Molecular and cellular
characterization of highly purified stromal stem cells derived from
human bone marrow. J Cell Sci 2003; 116: 1827- 35.
Gussoni E, Buzney EA, Khan MK, Flint AF and Mulligan RC: Dystrophin
expression in the mdx. Mouse restored by stem cell transplantation.
Nature 2002; 401: 390-94.
Guzman R, Choi R, Gera A, Andres R, and Steinberg G: Intravascular cell
replacement therapy for stroke Neurosurg Focus 2008; 24 (3&4): E14.
Hass S, Weidner N and Winkler J: Adult stem cell therapy in stroke. J
Neurosci 2005; 3: 59-65.
Hill WD, Hess DC, Martin-Studdard A, Carothers JJ and Hale D: SDF-1
(CXCL12) is upregulated in the ischemic penumbra following stroke:

182
association with bone marrow cell homing to injury. J Neuropathol Exp.
Neurol 2004; 63: 84–96.
Hough JP, Boyd RN and Keating JL: Early Diagnosis and Therapy in
cerebral palsy. Pediatrics 2010; 125: 670-75.
Ichim TE, Solano F, Glenn E, Morales F and Smith L: Stem Cell Therapy
for Autism. Journal of Translational Medicine 2007; 5: 173-79.
Ishibashi S, Sakaguchi M, Kuroiwa T, Yamasaki M and Shizuko I: Human
neural stem/progenitor cells, expanded in long term neurosphere culture,
promote functional recovery after focal ischemia in Mongolian gerbils. J
Neurosci Res 2009; 78: 215–23.
Jin K, LaFevre-Bernt M, Sun Y, Chen S and Crippen D: FGF-2 promotes
neurogenesis and neuroprotection and prolongs survival in a transgenic
mouse model of Huntington’s disease. Proc Nat Acad Sci U S A 2005;
102: 18189–94.
Joanna E, Emanuela B and Dianes K: Plasticity of bone marrow-derived
stem cells. Stem cells 2009; 22: 487-500.
Jodi LM, Soshana PB, Er-Yun CH: Human Neural Stem Cell Transplants
Improve Motor Function in a Rat Model of Huntington’s Disease, The
Journal of Comparative Neurology 2004; 475: 211–19.
Johann V, Schiefer J, Sass C, Mey J and Brook G: Time of transplantation
and cell preparation determine neural stem cell survival in a mouse
model of Huntington’s disease. Exp. Brain Res 2007; 177: 458–70.
Johnson D: Advancements in Stem Cell Research and Multiple Sclerosis
Treatments; Nat. Biotechnol 2011; 21:1200-1207.
Junying Yu and James A and Thomson: Embryonic stem (ES) cells .Science
20 Jan 2006; Vol. 311. No. 5759, p. 335.

183
Kalluri H and Dempsey R: Growth factors, stem cells, and stroke. Neurosurg
Focus 2008; 24 (3&4): E13
Keene CD, Sonnen JA, Swanson PD, Kopyov O and Leverenz JB: Neural
transplantation in Huntington disease: longterm grafts in two patients.
Neurology 2007; 68: 2093–98.
Keilhoff G, Stang F and Goihl A: Transdifferentiated mesenchymal stem cells
as alternative therapy in supporting nerve regeneration and myelination.
Cell Mol Neurobiol 2006; 26:1235–52.
Kelly S, Bliss TM, Shah AK, Sun GH and Foo WC: Transplanted human fetal
neural stem cells survive, migrate, and differentiate in ischemic rat
cerebral cortex. Proc. Nat Acad Sci U S A 2004; 101: 11839–44.
Kissling AA and Anderson SC: Human embryonic stem cells. Dev. Boil 2003;
277: 271-78.
Kocher A, Schuster M, Wang J, Homma S and Itescu S: Neovascu1arisation
of ischemic myocardium by human bone marrow derived angioblasts
prevents cardiomyocyte apoptosis, reduces remodeling and improves
cardiac function. Nat. Med 2001; 7: 430-36.
Kondziolka D and Wechsler L: Stroke repair with cell transplantation:
neuronal cells, neuroprogenitor cells, and stem cells. Neurosurg Focus
2008; 24 (3&4): E12.
Kushner JA, Ciemerych MA, Sicinska E, Wartschow LM and Long SY:
Cyclins D2 and Dl are essential for postnatal pancreatic beta-cell
growth, Mol. Cell Biol 2005; 25: 3752-62.
Lane EL, Handley OJ, Rosser AE and Dunnett SB: Potential cellular and
regenerative approaches for the treatment of Parkinson’s disease.
Neuropsychiatric Disease and Treatment 2008; 4(5) 835–45.

184
Lawrence G, Douglas J, Francois C, John F and Lynn A: Identification of
Candidate Regulators of Embryonic Stem Cell Differentiation, Journal
of Molecular and Cellular Proteomics 2006; 5: 57-67.
Limbert CA, Jakob FB and Seufert JA: Diabetes research and clinical
practice, Beta-cell replacement and regeneration, Strategies of cell-
based therapy for type I diabetes mellitus 2008; 4: 389-99.
Lin Y, Weisdorf DJ, Solovey A and Hebbel RP: Origins of circulating
endothelial cells and endothelial outgrowth from blood. J. Clin. Invest
2007; 105: 71-89.
Lindvall O and Kokaia Z: Stem cells in human neurodegenerative disorders -
time for clinical translation? The Journal of Clinical Investigation
http://www.jci.org 2010; 120: 29-40.
Lund RD, Wang S, Lu B, Girman S, Holmes T and Sauvé Y: Cells isolated
from umbilical cord tissue rescue photoreceptors and visual functions in
a rodent model of retinal disease. Stem Cells 2007; 25: 602–11.
MacLaren RE, Pearson RA, MacNeil A, Douglas RH and Salt TE: Retinal
repair by transplantation of photoreceptor precursors. Nature 2006; 444:
203–207.
Maragakis N, Galvez-Jimenez N, Shefner J and Targoff I: Epidemiology and
pathogenesis of amyotrophic lateral sclerosis. Journal of neurology
neurosurgery and psychiatry 2010; 74: 553-57.
Marquez YD, Wang MY and Liu CY: Cellular signaling in neural stem cells:
implications for restorative neurosurgery. Neurosurg Focus 2005; 19
(3): E2: 1-4.

185
McBride JL, Chen EY, Jakel RJ, Siegel I, and Svendsen CN: Human neural
stem cell transplants improve motor function in a rat model of
Huntington disease. J Comp Neurol 2004; 475: 211-19.
Metwally Y: Stem Cell therapy of Amyotrophic Lateral Sclerosis 2009 @
http://yasser metwally.net.
Metwally Y: Use of Stem Cells for Peripheral Nerve Repair 2009@
http://yasser metwally.net.
Miller G: Diagnosis of cerebral palsy. Neuroscience 2010; 106: 201-16.
Morizane A, Li J and Brundin P: From bench to bed: the potential of stem
cells for the treatment of Parkinson’s disease. Cell Tissue Res 2009;
333: 323–36.
Ofri R and Narfström K: Light at the end of the tunnel? Advances in the
understanding and treatment of glaucoma and inherited retinal
degeneration. Vet J 2007; 174: 10–22.
Olanow CW, Goetz CG and Kordower JH: A double-blind controlled trial of
bilateral fetal nigral transplantation in Parkinson’s disease. Ann Neurol
2003; 54: 403–14.
Olek M J: Treatment of progressive multiple sclerosis in adults. Nat Neurosci
2010; 7: 269–77.
Ourednik J, Ourednik V, Lynch WP, Schachner M and Snyder EY: Neural
stem cells display an inherent mechanism for rescuing dysfunctional
neurons. Nat Biotechnol 2005; 20: 1103–10.
Pan HC, Cheng FC and Chen CJ: Post injury regeneration in rat sciatic nerve
facilitated by neurotrophic factors secreted by amniotic fluid
mesenchymal stem cells. J Clin Neurosci 2007; 14: 1089–98.

186
Panchision D: Regenerative Medicine, repairing of nervous system with stem
cells Neuroscience 2006; 3: 35-43.
Phillips W, Morton AJ and Barker RA: Abnormalities of neurogenesis in the
R6/2 mouse model of Huntington’s disease are attributable to the in vivo
microenvironment. J Neurosci 2005; 25: 11564–76.
Pluchino S, Zanotti L, Rossi B, Ottoboni L and Salani G: Neurosphere-
derived multipotent precursors promote neuroprotection by an
immunomodulatory mechanism. Nature 2005; 436: 266–271.
Raisman G and Li Y: Repair of neural pathways by olfactory ensheathing
cells. Nat Rev Neurosci 2007; 8(4): 312-19.
Rishi S, Andres H, Ronald H, Andre G and Martin O: Stem Cell–Based
Therapies for Spinal Cord Injury, J Spinal Cord Med. Apr 2009; 32(2):
105–14.
Rosser AE, Barker RA, Harrower T, Watts C and Ho AK: Unilateral
transplantation of human primary fetal tissue in four patients with
Huntington’s disease. J Neurol Neurosurg Psychiatry 2002; 73: 678–85.
Sampaolesi M, Blot S, Mognol P, Perani L and Cossu G.: Mesoangioblast
stem cells ameliorate muscle function in dystrophic dogs. Nature 2006;
444: 574–79.
Sasaki M, Li B, Lankford KL, Radtke C and Kocsis JD: Remyelination of the
injured spinal cord. Prog Brain Res 2007; 161: 419–33.
Shyu WC, Lin SZ, Chiang MF, Su CY and Li H: Intracerebral peripheral
blood stem cell (CD34+) implantation induces neuroplasticity by
enhancing beta1 integrin-mediated angiogenesis in chronic stroke rats. J
Neurosci 2006; 26: 3444–53.

187
Son BR, Marquez-Curtis LA, Kucia M, Turner AR and Ratajczak MZ:
Migration of bone marrow and cord blood mesenchymal stem cells in
vitro is regulated by stromal derived factor-1-cXCR4 and hepatocyte
growth factor-c-met axes and involves matrix metalloproteinases. Stem
Cells 2006; 24: 1254–64.
Song J, Lee ST, Kang W, Park JE and Lee SE: Human embryonic stem cell
derived neural precursor transplants attenuate apomorphine-induced
rotational behavior in rats with unilateral quinolinic acid lesions.
Neurosci Lett 2007; 423: 58–61.
SonJa K, Nicolas Z, Pierre V, Luc S and Gottfried D: Similarities and
differences between human Adult and Embryonic Stem Cells, American
Journal of Reproductive Immunology 2006; (55)3: 169-180.
Sottile V, Halleux C, Bassilana F, Keller H and Seuwen K: Stem cell
characteristics of human trabecular bone- derived cells. Bone 2002; 30:
699-704.
Sutton B and Suchowersky O: Huntington disease: Clinical features and
diagnosis. Neurosci Biobehav Rev 2010; 21: 289–93.
Svendsen CN, Ebert AD: Encyclopedia of stem cell research /QU 13 E565
2008; 366-90.
Svendsen CN and Langston JW: Stem cells for Parkinson disease and ALS:
replacement or protection? Nature Med 2004; 10: 224-25. Abstract.
Vargas DL, Nascimbene C, Krishnan C, Zimmerman AW, Pardo CA:
Neuroglial activation and neuroinflammation in the brain of patients
with autism. Ann Neurol 2005; 57: 67-81.
Virchow A: History of Stem Cells, J. Pathol. Anat 1985; 407: 379-85.

188
Winkler C, Kirik D and Bjorklund A: Cell transplantation in Parkinson’s
disease: how can we make it work? Trends Neurosci 2005; 28: 86–92.
Xiao J, Nan Z, Motooka Y, Low WC: Transplantation of a novel cell line
population of umbilical cord blood stem cells ameliorates neurological
deficits associated with ischemic brain injury. Stem Cells Dev 2007; 14:
722–33.
Yiu G and He Z: Glial inhibition of CNS axon regeneration. Nat Rev Neurosci
2006; 7: 617-27.
Zannettino A, Paton S, Arthur A, Khor F, and Gimble J: Multi-potential
human adipose-derived stromal stem cells exhibit a perivascular
phenotype in Vitro and in vivo. J. Cell Physiol 2007; 210: 85-90.
Zhang C and Lodish H: Murine hematopoietic stem cells change their surface
phenotype during ex vivo expansion. Blood 2005; 105: 4314-20.
Ziv Y, Avidan H, Pluchino S, Martino G and Schwartz M: Synergy between
immune cells and adult neural stem/progenitor cells promotes functional
recovery from spinal cord injury. Proc Nat Acad Sci U S A 2006; 103:
13174–79.