the ulk3 kinase is critical for convergent control of cancer …€¦ · · 2017-09-12by csl and...
TRANSCRIPT

Cell Reports, Volume 20
Supplemental Information
The ULK3 Kinase Is Critical for Convergent Control
of Cancer-Associated Fibroblast Activation
by CSL and GLI
Sandro Goruppi, Maria-Giuseppina Procopio, Seunghee Jo, Andrea Clocchiatti, VictorNeel, and G. Paolo Dotto




1
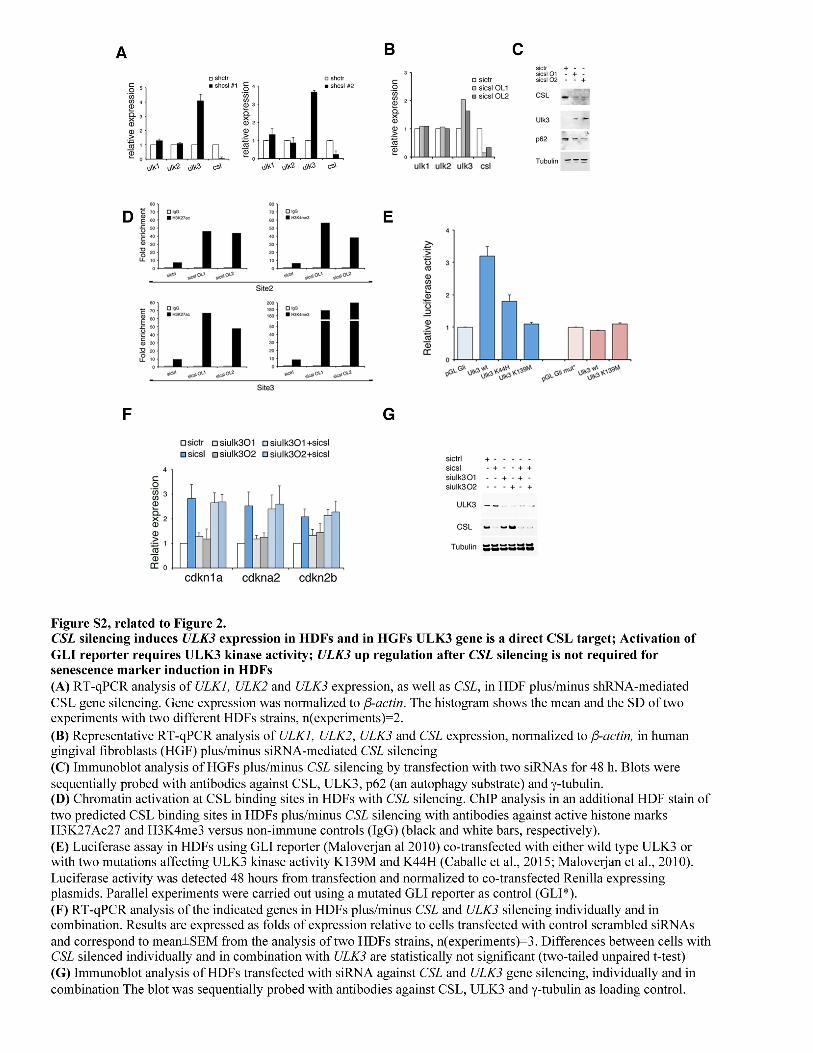
SUPPLEMENTAL EXPERIMENTAL PROCEDURES Cell Manipulations Human dermal fibroblasts were prepared from discarded skin samples of abdominoplasty patients at Massachusetts General Hospital (Boston, Massachusetts, USA) as in (Procopio et al., 2015). Conditions for culturing cells, viral infection, siRNA-mediated gene silencing, RT–qPCR and ChIP were previously reported (Brooks et al., 2014; Hu et al., 2012). For derivation of CAFs, surgically excised discarded skin SCCs samples, or non-affected skin samples, were cut into 1–2 mm pieces after removal of fat excess, followed by incubation in 2 ml of PBS containing 0.25 mg ml−1 of Liberase TL (Roche) for 40 min at 37 °C with gentle shaking. After equal volume addition of fetal bovine serum to stop the enzymatic digestion, the dissociated tissue was passed through a 10 ml syringe attached to a 70µm sieve. Cells from the flow through were centrifuged, washed three times with DMEM 10% fetal bovine serum and seeded in a 10 cm tissue culture dishes. Adherent cells were expanded for characterization as with HDFs. In some cases we derived simultaneously matched normal HDFs from the same patient using discarded non-diseased area. Skin SCC13 cells were originally reported in (Restivo et al., 2011). For in co-culture and in vivo approaches, SCC13 were infected with a high titer EGFP-expressing lentivirus (Tiscornia et al., 2003) and expanded. HDFs and CAFs strains stably infected with a doxycycline-inducible lentiviral vector for Myc-tagged CSL in parallel with empty vector control. CAFs were infected and treated for 5 days plus/minus doxycycline (500 ng ml−1) (Procopio et al., 2015) before RT-PCR analysis. RNAi experiments were carried out with SilencerTM oligos from Ambion (for CSL, ULK3, atg5, atg7, becn1 and GLI2) and Quiaperfect reagents (Qiagen) or by using shRNA-targeting vectors from Sigma (for CSL, atg5 and ULK3), as previously described (Hu et al., 2012; Procopio et al., 2015). Nucleotide sequences used for RNA-interference are given in Tables S1 and S2. For co-culture experiments HDFs were first infected with shulk3 or shctr viruses and selected with puromycin for 3 days. After expansion, the HDFs were re-infected with shCSL or shctr and selected with G418 for one week. The resulting cells were co-cultured in an insert with 4x103 EGFP-SCC13 cells for 4 days, in duplicate. Each day, four immunofluorescence pictures were taken and used to determine the cell number. After 4 days the SCC13 cultures were used for RNA analysis by RT-qPCR. ULK3 overexpression was obtained by infecting HDFs with an ULK3-V5 tagged virus (Broad Institute) and selecting with 10µg/ml blasticidin for the indicated times. HDFs were infected with atg5 silencing lentivirus and selected for 3 days with puromycin. After 7 days cells were infected with a high titer lentivirus expressing a ULK3-V5 tagged protein and harvested after additional 3 days without selection. Spontaneously immortalized MEFs Gli wild type (+/+) and with combined gene deletion of gli1 and gli 2 (-/-)(Lipinski et al., 2008) were infected with shulk3 or shctr viruses and selected with puromycin for 3 days. After 2 weeks of expansion cells were analyzed by immunofluorescence and for gene expression, then further expanded and used for the in vivo studies. Human gingival fibroblasts were from Sciencell, MEF from knockout mice for GLI1/2 have been characterized(Lipinski et al., 2008). All HDFs, HGFs, MEFs, CAFs and HEK293 cellular strains used were routinely checked for the absence of mycoplasma. Immunofluorescence, autophagy studies and cell assays Western blots and immunofluorescences were performed as in (Kong et al., 2010; Procopio et al., 2015) using anti LC3B, CSL, p62, p-Beclin S15, GLI2, PKM2, LDHA and β-actin antibodies (Cell Signaling); ULK3, TNC, Postn and TOM20 (Santa Cruz); γ-tubulin, SMA and p62 were from Sigma. For immunofluorescence cells were seeded on coverslips, fixed in 3% PFA and processed as in(Kong et al., 2010). Autophagosome evaluation studies were performed essentially as in (Kong et al., 2010). Two different strains of HDFs were infected with pHcRed1-LC3 encoding for the far-red fluorescent protein (Clontech) fused to LC3 (Kong et al., 2010). After silencing endogenous CSL with siRNAs for 48 h, the percentage of cells having Red-LC3 translocation into dots (a minimum of 200 cells/sample, in four experiments) was counted in the Red-LC3 expressing HDF cells, fixed with 3.7% paraformaldehyde (Sigma) in PBS containing 1µg/ml DAPI -for nuclear DNA visualization. Cells with only nuclear Red-LC3 were considered negatives while cells with more than five dots were considered as positive(Klionsky et al., 2016; Kong et al., 2010). Cells with only cytosolic dots were considered positive (if more than five). To monitor autophagosomal flux, 400nM Bafilomycin A1 (Calbiochem) was added to the cells for 4h as lysosome inhibitor; 10µmM KU0063794 was used as positive control of autophagy activation. Luciferase assays were performed as in (Kong et al., 2010) co-transfecting in 6 well culture dishes HDFs with 8xGLI BS reporter (Addgene) with either ULK3 wild type or point mutations described in (Caballe et al., 2015). Renilla luciferase expressing vector was added to normalize transfection efficiency and each experiment was carried out in triplicate wells. The assays were analyzed after 48h using a commercial kit (Dual assay luciferase system Promega).

2
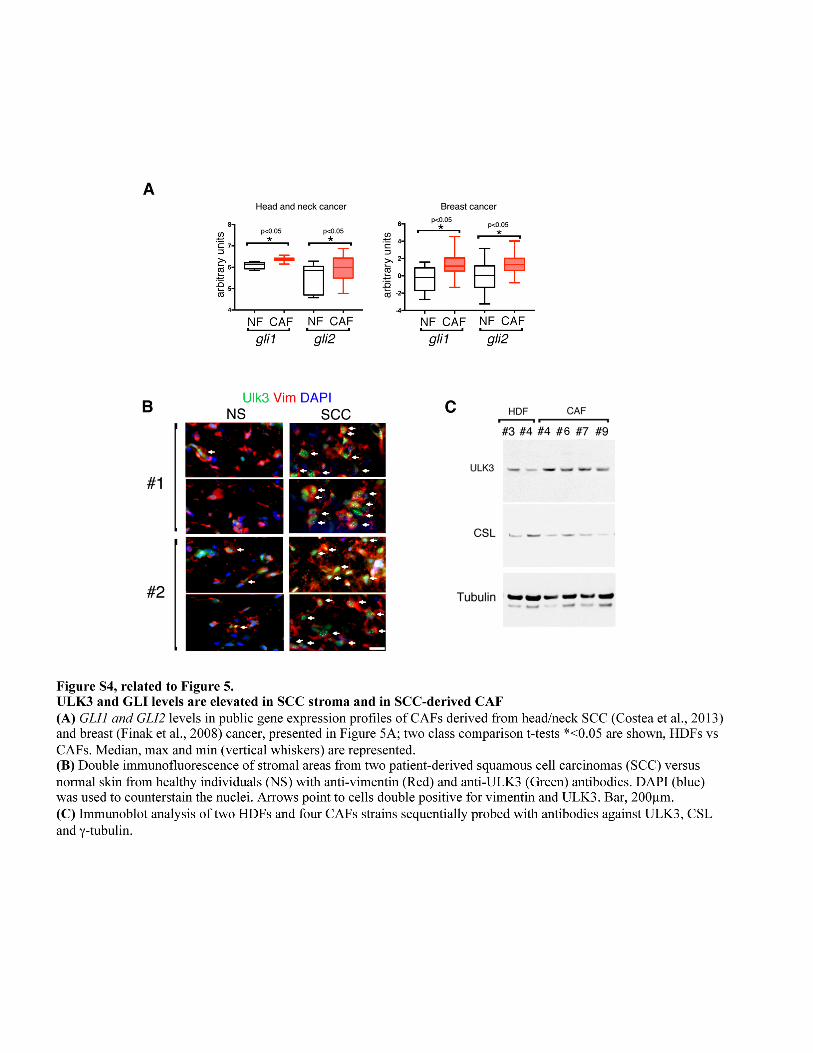
Colorimetric determination of lactate was carried out using an EnzyChrom L-lactate assay kit as indicated by the manufacturer (BioAssay Systems). The cell culture supernatant of several HDFs strains with CSL silencing or overexpressing ULK3-V5 was collected after three days of culture. The concentration of lactate was determined monitoring the reduction of MTT by NADH produced by the oxidation of lactate by the lactate dehydrogenase (LDH) in the medium. The absorbance (565nm) of the reactions was measured at time 0 and after 30-60 min, and the values were normalized for the total amount of cellular proteins in each experiment. Each sample reaction was carried out in triplicate and a standard curve of lactate was used to determine the lactate concentration. For mitochondrial labeling with MitoTracker Red FM (Thermofisher), HDFs plus/minus CSL silencing for three days were pulse labeled with 100nM of dye, then fixed with 3% PFA in PBS and analyzed using confocal microscopy (Zeiss Observer Z1) using a Zen Pro 2.3 software. DAPI was used to counterstain the nuclei. Protein interaction and immunoprecipitations Proximity ligation assay (Soderberg et al., 2006) was performed using Duolink PLA kit (Sigma) according to manufacturer’s protocol. Briefly, cells were fixed with 4% formaldehyde and permeabilized in 0.1% Triton X-100. After blocking with PLA blocking solution, HDFs were incubated with primary antibody solution containing ULK3 and GLI2 antibodies (both from Santa Cruz). After washing with PLA wash buffer, cells were incubated with PLA probes, anti-rabbit PLUS, anti-mouse MINUS, then washed, ligated, amplified by rolling circle amplification. Images were obtained with a Nikon Eclipse Ti confocal microscope. Gene expression and Chromatin immunoprecitation Chromatin immunoprecitation (ChIP) assays with antibodies against CSL versus non-immune IgG controls (both Cell Signaling), and against rabbit anti H3K27Ac, H3K4me3 and IgG controls (all from Abcam) were carried out as previously described (Brooks et al., 2014; Hu et al., 2012), followed by determination of binding enrichment for the indicated sites within human ULK3 promoter by qPCR; CSL binding sites in ULK3 regulatory region were predicted using Matinspector (Genomatix). Oligonucleotides for qPCR were designed using Primer 3 (http://bioinfo.ut.ee/primer3-0.4.0/). Primer sequences are in Tables S3 and S4; mouse sequences as in (Hu et al., 2012). Public data sets of gene expression in the tumor stroma compared to normal skin fibroblasts were the following: Head and neck cancer GSE38517; prostate cancer GSE34312; breast cancer GSE9014. Human samples and LCM experiments Normal human skin samples and samples of squamous cell carcinoma (SCC) were obtained at the Department of Dermatology, Massachusetts General Hospital, as discarded parts not needed for diagnosis. All samples were processed as approved by the Institutional Review Board. H&E and immunohistochemistry analysis was performed using anti ULK3 antibodies (Santa Cruz) and anti Vimentin antibodies (Abcam) as in (Procopio et al., 2015). Matched normal skin and SCC paraffin samples used for LCM and RT-PCR were provided by the Department of Dermatology, University of Tubingen with institutional review board approvals and informed consent. LCM was made using an Arcturus XT micro-dissection system (Applied Biosystems) as before (Hu et al., 2012; Procopio et al., 2015). Gene expression was normalized to 36β4, and the absence of contaminating leucocytes in the selected areas confirmed with CD45 oligos in RT-qPCR assays. Animal studies Experiments with gli 1/2 -/- MEFs: Mouse ear injections with the MEFs were carried out in 8 to 10-week-old female SKID (CB17sc-m) mice (Taconic), as in (Procopio et al., 2015). EGFP expressing SCC13 cells (1 × 105) were admixed with equal numbers of MEFs (either gli +/+ or gli1 and gli2 -/-) with shRNA-mediated silencing of Csl, or a control. Cells were injected 5µl per site using a 33-gauge micro syringe (Hamilton). Six mice were used with gli +/+ and six mice were injected with gli1 and 2 -/- MEFs. Starting the day after injection, the mice ears were imaged using a fluorescent stereomicroscope (Leica MZ-FLIII), every three days for 21 days. The mice were sacrificed after 24 days and images of the ears taken using bright field and fluorescence stereomicroscopy. For the ear injection and the imaging the mice were injected intra peritoneal with xylazine-ketamine solution in PBS. The mice were housed as a group and their health was monitored daily by the MGH housing facility. The mice developed tumors as follow: 4/6 for shctr and 5/6 for shcsl MEFs gli +/+; 5/6 for shctr and 6/6 for shcsl MEFs gli1 and gli2 -/-. All the tumors obtained were analyzed (data in Figure 1I and 1J) or only matched ear pairs (data in Figure S1D). No lesion/pair was excluded from the specific statistical analysis. No blinding of the personnel. No adverse event found and no modification to the experimental protocol was made. Experiment with patient-derived CAFs: Mouse ear injections of cells were carried out in 8 to 10-week-old female SKID (CB17sc-m) mice (Taconic). EGFP expressing SCC13 cells (1 × 105) were admixed with equal numbers of CAFs (strain#1 or strain#3) with shRNA-mediated silencing of ULK3, or a control. Cells were injected (5µl per site) and the following day the efficient injection was confirmed using a fluorescent stereomicroscope (Leica MZ-FLIII). A total of eight mice in two experiments were injected with CAF1 strain and six mice were injected with CAF3 strain. For the ear injection and the confirmation of efficient injection the next day the mice were injected intra peritoneal with avertin solution. The mice were housed as a group and their health was monitored daily by the MGH housing facility and every three days by lab personnel. Mice were sacrificed after 3 weeks and images of the ears taken using bright field and

3
fluorescence stereomicroscopy. The mice developed tumors as follow: 4/8 for shctr and 6/8 for CAF1 strain; 5/6 for shctr and 4/6 for CAF3 strain. All the tumors obtained were analyzed (data in Figure 6A) or only matched ear pairs (data in FigureS6B). A total of three experiments were made. No lesion/pair was excluded from the specific statistical analysis. No blinding of the personnel. No adverse event found and no modification to the experimental was protocol made. All the animal studies were approved by the MGH/Partners institutional animal care and use committee. Immunohistochemistry of the tumors was performed as in (Brooks et al., 2014; Hu et al., 2012; Procopio et al., 2015) and quantification of ki67 and p63 protein levels was made using the watershed algorithm (http://imagej.nih.gov/ij/plugins/watershed.html) and ImageJ, NIH. Quantification of all other tissue immunocytochemistry staining was performed using ImageJ. Antibodies against p63 and Tenascin C were from Santa Cruz; Pan cytokeratin, ki67 and periostin from Abcam; anti smooth muscle actin from Sigma and anti CD31/PECAM from BD-Pharmingen. All were used as indicated by manufacturer. A detailed list of all the antibodies and the condition(s) used is in TABLE S5. Statistical analysis Data are presented as mean± SEM, mean ± SD, or as ratio among treated and controls, as indicated in the Figure legends. For gene expression and functional testing assays, statistical significance of differences between experimental groups and controls was assessed by two-tailed unpaired or paired t-test, and one sample t-test, as indicated in the figure legends. A value for p < 0.05 was considered as statistically significant. For each experiment, two to three separate HDF strains were used in the independent experiments. For ear injection/tumorigenicity assays, we minimize the individual animal variability issue through parallel ear injections of the same animals with control versus experimental combinations of cells. No statistical method was used to predetermine sample size in animal experiments. No exclusion criteria were adopted for studies and sample collection. No randomization was used and the researchers were not blinded. Brooks, Y.S., Ostano, P., Jo, S.H., Dai, J., Getsios, S., Dziunycz, P., Hofbauer, G.F., Cerveny, K., Chiorino, G., Lefort, K., et al. (2014). Multifactorial ERbeta and NOTCH1 control of squamous differentiation and cancer. J Clin Invest 124, 2260-2276. Caballe, A., Wenzel, D.M., Agromayor, M., Alam, S.L., Skalicky, J.J., Kloc, M., Carlton, J.G., Labrador, L., Sundquist, W.I., and Martin-Serrano, J. (2015). ULK3 regulates cytokinetic abscission by phosphorylating ESCRT-III proteins. Elife 4, e06547. Costea, D.E., Hills, A., Osman, A.H., Thurlow, J., Kalna, G., Huang, X., Pena Murillo, C., Parajuli, H., Suliman, S., Kulasekara, K.K., et al. (2013). Identification of two distinct carcinoma-associated fibroblast subtypes with differential tumor-promoting abilities in oral squamous cell carcinoma. Cancer Res 73, 3888-3901. Finak, G., Bertos, N., Pepin, F., Sadekova, S., Souleimanova, M., Zhao, H., Chen, H., Omeroglu, G., Meterissian, S., Omeroglu, A., et al. (2008). Stromal gene expression predicts clinical outcome in breast cancer. Nat Med 14, 518-527. Hara, T., Nakamura, K., Matsui, M., Yamamoto, A., Nakahara, Y., Suzuki-Migishima, R., Yokoyama, M., Mishima, K., Saito, I., Okano, H., et al. (2006). Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature 441, 885-889. Hu, B., Castillo, E., Harewood, L., Ostano, P., Reymond, A., Dummer, R., Raffoul, W., Hoetzenecker, W., Hofbauer, G.F., and Dotto, G.P. (2012). Multifocal epithelial tumors and field cancerization from loss of mesenchymal CSL signaling. Cell 149, 1207-1220. Klionsky, D.J., Abdelmohsen, K., Abe, A., Abedin, M.J., Abeliovich, H., Acevedo Arozena, A., Adachi, H., Adams, C.M., Adams, P.D., Adeli, K., et al. (2016). Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy 12, 1-222. Kong, D.K., Georgescu, S.P., Cano, C., Aronovitz, M.J., Iovanna, J.L., Patten, R.D., Kyriakis, J.M., and Goruppi, S. (2010). Deficiency of the transcriptional regulator p8 results in increased autophagy and apoptosis, and causes impaired heart function. Mol Biol Cell 21, 1335-1349. Lipinski, R.J., Bijlsma, M.F., Gipp, J.J., Podhaizer, D.J., and Bushman, W. (2008). Establishment and characterization of immortalized Gli-null mouse embryonic fibroblast cell lines. BMC Cell Biology 9, 49-49. Maloverjan, A., Piirsoo, M., Michelson, P., Kogerman, P., and Osterlund, T. (2010). Identification of a novel serine/threonine kinase ULK3 as a positive regulator of Hedgehog pathway. Exp Cell Res 316, 627-637. Procopio, M.G., Laszlo, C., Al Labban, D., Kim, D.E., Bordignon, P., Jo, S.H., Goruppi, S., Menietti, E., Ostano, P., Ala, U., et al. (2015). Combined CSL and p53 downregulation promotes cancer-associated fibroblast activation. Nat Cell Biol 17, 1193-1204.

4
Restivo, G., Nguyen, B.C., Dziunycz, P., Ristorcelli, E., Ryan, R.J., Ozuysal, O.Y., Di Piazza, M., Radtke, F., Dixon, M.J., Hofbauer, G.F., et al. (2011). IRF6 is a mediator of Notch pro-differentiation and tumour suppressive function in keratinocytes. EMBO J 30, 4571-4585. Soderberg, O., Gullberg, M., Jarvius, M., Ridderstrale, K., Leuchowius, K.J., Jarvius, J., Wester, K., Hydbring, P., Bahram, F., Larsson, L.G., et al. (2006). Direct observation of individual endogenous protein complexes in situ by proximity ligation. Nature methods 3, 995-1000. Tasdemir, E., Maiuri, M.C., Galluzzi, L., Vitale, I., Djavaheri-Mergny, M., D'Amelio, M., Criollo, A., Morselli, E., Zhu, C., Harper, F., et al. (2008). Regulation of autophagy by cytoplasmic p53. Nat Cell Biol 10, 676-687. Tiscornia, G., Singer, O., Ikawa, M., and Verma, I.M. (2003). A general method for gene knockdown in mice by using lentiviral vectors expressing small interfering RNA. Proc Natl Acad Sci U S A 100, 1844-1848.




TABLE1-identifiers for the SilencerTM oligonucleotides (Ambion and Thermofisher) used for RNA-
interference experiments
gene name #cat #id
control 4390846 NA
ulk3 O1 4392420 24888
ulk3 O2 4392420 24886
gli2 O1 4392420 5817
gli2 O2 4392420 5818
csl O1 4392420 7252
csl O2 4392420 7253
atg5 6345S 6345
atg7 AM16708A 135754
becn1 AM16708A 137196

TABLE 2-identifiers for the shRNA sequences (Sigma) used for RNA-interference experiments
human
gene targeted
#id Sequence
ulk3 #1
CCGGGCAGACTTTGGTTTCGCACAACTCGAGTTGTGCGAAACCAAAGTCTGCTTTTTG
ulk3 #2
CCGGACTCGTGAAGTGGTAGCCATACTCGAGTATGGCTACCACTTCACGAGTTTTTTG
csl #1 CCGGGCTGGAATACAAGTTGAACAACTCGAGTTGTTCAACTTGTATTCCAGCTTTTT
csl #2 CCGGCCCTAACGAATCAAACACAAACTCGAGTTTGTGTTTGATTCGTTAGGGTTTTT
mouse
gene targeted
csl #1 CCGGCCCTGTGCGTTTATTGGAATACTCGAGTATTCCAATAAACGCACAGGGTTTTTG
atg5 #1 CCGGGCCAAGTATCTGTCTATGATACTCGAGTATCATAGACAGATACTTGGCTTTTTG

TABLE3-Sequence of the oligonucleotides used for PCR experiments
gene Sequence
ulk1 TCGAGTTCTCCCGCAAGG
CGTCTGAGACTTGGCGAGGT
ulk2 TGGGTCCTCCCAACTATCTACAAGT
CGAGATGTTGTGTGGCACCAA
ulk3 TGAAGGAGCAGGTCAAGATGAGG
GCTACGAACAGATTCCGACAGTCC
ulk3_N2 AAGGAGCAGGTCAAGATGAG
GTGCAAGAGCTACGAACAGA
il6 CCTTCCAAAGATGGCTGAAA
GCTCTGGCTTGTTCCTCACT
ptgs2 GTTTTGACATGGGTGGGAAC
CCCTCAGACAGCAAAGCCTA
p15 GCGCGCGATCCAGGTCATGATG
CAGCACCACCAGCGTGTC
p16 CTGCCCAACGCACCGAATAGTTAC
CACCACCAGCGTGTCCAGGAAG
p21 GATTAGCAGCGGAACAAGGA
CAACTACTCCCAGCCCCATA
tnc TAACAGCATCACCCTGGAAT
TCCTTGTCTTCCTTCACAGC
acta-2 AGCGCAAATACTCTGTCTGG
AGGCATAATTCCACAGGACA
beta-actin GTTGTCGACGACGAGCG
GCACAGAGCCTCGCCTT
csl CAAAAGTTGCACAGAAGTCATA
TGCTGCATTTCTTGGTCAC
gli1 GTGCAAGTCAAGCCAGAACA
GGGGGTAATGGGAAAAGAGA
gli2 CAGTGTGCATGCCTGGTATC
TGGAATTTGGAACTGGCTTC
gli2_N2 CCTCATTAAGGCCAAGGTCA
TGATGCAGTTCCTTTATTATCAGG
postn CAAAACTGAAGGACCCACAC
TATTTCCACAGGCACTCCAT
mmp3 CAATTTCATGAGCAGCAACG

AGGGATTAATGGAGATGCCC il-8 TCTGGCAACCCTAGTCTGCT GCTTCCACATGTCCTCACAA il-1 AGCCTCAAGATGAAGGCAAA GATGCCTGGTCACACTCAGA becn1 AGGTTGAGAAAGGCGAGACA GCTTTTGTCCACTGCTCCTC atg5 GCAAGCCAGACAGGAAAAAG GACCTTCAGTGGTCCGGTAA mki67 CCCCACCTCAGAGAGTTTTG GGGCTTGCAGAGCATTTATC

TABLE4-Sequence of the oligonucleotides used for ChIP studies of Ulk3 promoter
Name Sequence
OL1 SE CCTCCTGAAAGCTGATGGAT
OL1 AS ACCAGCCGTATGCTCAAAGA
OL2 SE TCGTGCCAGTAGCTCTCCTT
OL2 AS GCGCAGAGGCGTCTAGAGAT
OL3 SE CGGGGAGACTCTGAGAGGTTA
OL3 AS AAGACCTAAGCGTGGCAGTC
OL4 SE CCGTGCTCTCAGTCTTCCTC
OL4 AS AAAAACATCTCGTGGCAAGG
OL5 SE CCCTCAGCAGGGACTGATAA
OL5 AS TCCACCCTTAAAACCAGCAC
OL6 SE GTGGAGCCACCCAGTGCT
OL6 AS CTTCATCCTCACCGAGCG

TABLE5- List of antibodies used
Target Species Company and catalog # Assay and dilution used
CSL rabbit Cell Signaling #5313 WB (1:1000), IF(1:50), ChIP, IP
H3K27ac3 rabbit Abcam #4729 ChIP
H3K4me3 rabbit Abcam #8580 ChIP
LC3B rabbit Cell Signaling #3868 WB(1:1000), IF(1:50)
SQSTSM/p62 mouse Santa Cruz #28359 PLA(1:100)
P-Beclin1 rabbit Cell Signaling #13825 WB(1:500)
Gli2 rabbit Cell Signaling #2585 WB(1:500)
β-actin rabbit Cell Signaling #13E5 WB(1:2000)
γ-tubulin mouse Sigma #GTU-88 WB(1:2000)
p63 rabbit Santa Cruz #8343 IF(1:100)
ki67 rabbit Abcam #15580 IF(1:200)
Vimentin mouse Abcam #20346 IF(1:200)
Ulk3 rabbit Santa Cruz #137897 WB(1:500), IF(1:100); PLA(1:100)
Smooth muscle actin mouse Sigma #1A4 WB(1:2000), IF(1:100)
Tenascin-C mouse Santa Cruz #25328 IF(1:100)
CD31/PECAM rat BD/Pharmingen #550274 IF(1:100)
Pan cytokeratin mouse Abcam #7753 IF(1:200)
SQSTSM/p62 rabbit Sigma# P0067 WB(1:5000); PLA (1:200); IP (1:1000)
Periostin rabbit Abcam #92460 IF(1:100)
GFP rabbit Santa Cruz #8334 WB(1:11000)
Gli2 mouse Santa Cruz #271786 WB(1:500); PLA(1:100)
Atg5 rabbit Epitomics #3167-1 WB(1:500)
P-Gli2 rabbit Xing et al, Cell 2014 WB(1:500)
V5 Tag rabbit Sigma#1306079 WB(1:1000); IP(1:200)
Gli2 rabbit Santa Cruz#28674 WB(1:1000)
CD68 rat Abcam#53444 IF(1:100)
Vimentin rabbit Abcam#45939 IF(1:100)
TOM20 mouse Santa Cruz#17764 IF(1:100); WB (1:500)
PKM2 rabbit Cell Signaling #9198S IF(1:100); WB(1:1000)
LDHA rabbit Cell Signaling #3582S IF(1:100); WB(1:1000)
Ulk3 mouse MyBiosource#MBS9200567 PLA (1:50); IHC (1:100)