the role of different modes of interactions among
TRANSCRIPT

Faculty of Environmental Sciences
The role of different modes of interactions among neighbouring plants in driving population dynamics Dissertation for awarding the academic degree Doctor rerum naturalium ( Dr. rer. nat. ) Submitted by M.Sc. Yue Lin Supervisor: Mrs. Prof. Dr. Uta Berger Dresden University of Technology Mr. Prof. Dr. Volker Grimm Helmholtz Centre for Environmental Research – UFZ University of Postdam
Dresden, 22.01.2013

II
Explanation of the doctoral candidate This is to certify that this copy is fully congruent with the original copy of the dissertation with the topic: „ The role of different modes of interactions among neighbouring plants in driving population dynamics “
Dresden, 22.01.2013
……………………………………….….
Place, Date
Lin, Yue
……………………………………….….
Signature (surname, first name)

III
To my doctoral advisors
Prof. Dr. Uta Berger
and Prof. Dr. Volker Grimm
whose wisdoms furnish the best of sounding boards,
whose reliance and optimisms dispel all my misgivings,
whose enthusiasms and professionalisms are contagious,
who welcomed me from China to Germany with open arms,
and who are my mentors forever.
To my dear and loving wife
Qian-Ru Ji
who has made numerous sacrifices to support my studies.

IV

V
Table of contents
Summary ....................................................................................................... VII
Zusammenfassung ......................................................................................... IX
Chapter 1: General introduction ......................................................................1
Chapter 2: Plant competition and metabolic scaling theory ......................... 26
Chapter 3: Mechanisms mediating plant mass-density relationships:
the role of biomass allocation, above- and belowground
competition ............................................................................... 57
Chapter 4: Exploring the interplay between modes of positive and
negative plant interactions ........................................................ 93
Chapter 5: Metabolic scaling theory predicts near ecological equivalence
in plants ................................................................................... 129
Chapter 6: General discussion .................................................................... 139
Acknowledgments ...................................................................................... 151

VI

VII
Summary
The general aim of my dissertation was to investigate the role of plant interactions in
driving population dynamics. Both theoretical and empirical approaches were employed.
All my studies were conducted on the basis of metabolic scaling theory (MST), because
the complex, spatially and temporally varying structures and dynamics of ecological
systems are considered to be largely consequences of biological metabolism. However,
MST did not consider the important role of plant interactions and was found to be invalid
in some environmental conditions. Integrating the effects of plant interactions and
environmental conditions into MST may be essential for reconciling MST with observed
variations in nature. Such integration will improve the development of theory, and will
help us to understand the relationship between individual level process and system level
dynamics.
As a first step, I derived a general ontogenetic growth model for plants which is
based on energy conservation and physiological processes of individual plant. Taking the
mechanistic growth model as basis, I developed three individual-based models (IBMs) to
investigate different topics related to plant population dynamics:
1. I investigated the role of different modes of competition in altering the prediction
of MST on plant self-thinning trajectories. A spatially-explicit individual-based
zone-of-influence (ZOI) model was developed to investigate the hypothesis that MST
may be compatible with the observed variation in plant self-thinning trajectories if
different modes of competition and different resource availabilities are considered. The
simulation results supported my hypothesis that (i) symmetric competition (e.g.
belowground competition) will lead to significantly shallower self-thinning trajectories
than asymmetric competition as predicted by MST; and (ii) individual-level metabolic
processes can predict population-level patterns when surviving plants are barely affected
by local competition, which is more likely to be in the case of asymmetric competition.
2. Recent studies implied that not only plant interactions but also the plastic biomass
allocation to roots or shoots of plants may affect mass-density relationship. To investigate
the relative roles of competition and plastic biomass allocation in altering the
mass-density relationship of plant population, a two-layer ZOI model was used which

VIII
considers allometric biomass allocation to shoots or roots and represents both above- and
belowground competition simultaneously via independent ZOIs. In addition, I also
performed greenhouse experiment to evaluate the model predictions. Both theoretical
model and experiment demonstrated that: plants are able to adjust their biomass
allocation in response to environmental factors, and such adaptive behaviours of
individual plants, however, can alter the relative importance of above- or belowground
competition, thereby affecting plant mass-density relationships at the population level.
Invalid predictions of MST are likely to occur where competition occurs belowground
(symmetric) rather than aboveground (asymmetric).
3. I introduced the new concept of modes of facilitation, i.e. symmetric versus
asymmetric facilitation, and developed an individual-based model to explore how the
interplay between different modes of competition and facilitation changes spatial pattern
formation in plant populations. The study shows that facilitation by itself can play an
important role in promoting plant aggregation independent of other ecological factors (e.g.
seed dispersal, recruitment, and environmental heterogeneity).
In the last part of my study, I went from population level to community level and
explored the possibility of combining MST and unified neutral theory of biodiversity
(UNT). The analysis of extensive data confirms that most plant populations examined are
nearly neutral in the sense of demographic trade-offs, which can mostly be explained by a
simple allometric scaling rule based on MST. This demographic equivalence regarding
birth-death trade-offs between different species and functional groups is consistent with
the assumptions of neutral theory but allows functional differences between species. My
initial study reconciles the debate about whether niche or neutral mechanisms structure
natural communities: the real question should be when and why one of these factors
dominates.
A synthesis of existing theories will strengthen future ecology in theory and
application. All the studies presented in my dissertation showed that the approaches of
individual-based and pattern-oriented modelling are promising to achieve the synthesis.

IX
Zusammenfassung
Das primäre Ziel meiner Dissertation umfasste die Erforschung der Rolle von
Wechselwirkungen zwischen Pflanzen für die Populationsdynamik. Dazu wurden sowohl
theoretische als auch empirische Ansätze angewendet. Alle meine Untersuchungen
wurden auf Basis der Metabolic Scaling Theory (MST) durchgeführt, da angenommen
wird, dass die komplexen räumlich und zeitlich variierenden Strukturen ökologischer
Systeme weitgehend die Konsequenzen von biologischem Metabolismus sind. Allerdings
berücksichtigt die MST nicht die wichtige Rolle der Wechselwirkungen zwischen
Pflanzen und wurde für einige Umweltbedingungen als unzutreffend befunden. Die
Integration dieser Wechselwirkungen und Umweltbedingungen in die MST hingegen ist
essenziell, um die MST mit den beobachteten Abweichungen in der Natur abzustimmen.
Diese Integration wird die Entwicklung von Theorien verbessern und trägt zum
Verständnis der Zusammenhänge zwischen Prozessen auf Individual- und Systemebene
bei.
In einem ersten Schritt leitete ich ein generelles ontogenetisches Wachstumsmodell für
Pflanzen ab, welches auf Energieersparnis und physiologischen Prozessen der
individuellen Pflanzen basiert. Auf dieser Grundlage entwickelte ich drei
individuenbasierte Modelle (IBM), um verschiedene Fragestellungen in Bezug auf die
Populationsdynamik der Pflanzen zu analysieren.
In Kapitel 2 untersuchte ich den Einfluss verschiedener Arten von Konkurrenz auf die
Prognosen der MST in Bezug auf Trajektorien der Selbstausdünnung. Ein räumlich
explizites, individuenbasiertes zone-of-influence (ZOI) Modell wurde entwickelt, um die
Hypothese zu eruieren, dass die MST kompatibel zu den Abweichungen in
Selbstausdünungstrajektorien ist, wenn verschiedene Arten der Konkurrenz und
unterschiedliche Ressourcenverfügbarkeiten in Betracht gezogen werden. Die Ergebnisse
der Simulationen bestätigen meine Hypothese, dass (i) symmetrische Konkurrenz (z.B.:
unterirdisch – Wurzeln) zu signifikant flacheren Selbstausdünnungstrajektorien führt als
bei asymmetrischer Konkurrenz durch die MST vorhergesagt wird; und (ii) metabolische
Prozesse auf Individualebene Muster auf Systemebene vorhersagen können, wenn die
überlebenden Pflanzen kaum durch lokale Konkurrenz beeinflusst werden, was bei
asymmetrischer Konkurrenz meist der Fall ist.
Aktuelle Studien unterstellen, dass nicht nur die Interaktion zwischen Pflanzen, sondern
auch die Biomasseallokation zu Wurzeln oder Trieben der Pflanzen das Masse-Dichte
Verhältnis beeinflussen kann. In Kapitel 3 untersuchte ich die Rolle der Konkurrenz und
Biomasseallokation im Hinblick auf Veränderung des Masse-Dichte Verhältnisses der
Pflanzenpopulation. Dazu wurde ein ZOI Modell mit zwei Ebenen entwickelt, welches
die allometrische Biomasseallokation zu Wurzeln oder Trieben berücksichtigt und sowohl

X
die unter- wie überirdische Konkurrenz durch zwei unabhängige ZOI simultan darstellt.
Zusätzlich führte ich ein Gewächshausexperiment durch, um die Modellprognosen zu
evaluieren. Sowohl das theoretische Modell und das Experiment zeigen, dass Pflanzen in
der Lage sind, ihre Biomasseallokation je nach Umweltfaktoren anzupassen. Diese
individuellen Anpassungsstrategien jedoch verändern die relative Bedeutung der ober-
wie unterirdischen Konkurrenz und damit das Masse-Dichte Verhältnis auf
Populationsebene. Falsche Vorhersagen der MST treten mit höherer Wahrscheinlichkeit
auf, wenn unterirdische Konkurrenz (symmetrisch) vorliegt.
In Kapitel 4 führte ich ein neues Konzept verschiedener Arten der Förderung ein, z.B.:
symmetrische versus asymmetrische Förderung und entwickelte ein individuenbasiertes
Modell um zu ergründen, wie das Zusammenspiel zwischen verschiedenen Arten der
Konkurrenz und Förderung die räumlichen Muster in Pflanzenpopulationen verändert.
Diese Studie zeigt, dass die Förderung allein die Aggregation von Pflanzen maßgeblich
begünstigen kann, unabhängig von anderen ökologischen Faktoren (z.B.:
Samenausbreitung, Mortalität, Heterogenität).
In Kapitel 5 betrachte ich die Ebene der Pflanzengesellschaften und prüfe die
Möglichkeit, die MST mit der Unified Neutral Theory of Biodiversity (UNT) zu
kombinieren. Die Analyse umfangreicher Daten bestätigt, dass die meisten untersuchten
Pflanzenpopulationen nahezu neutral im Sinne des demographischen Trade-off
(Geburt/Sterberate) sind, was durch eine simple Skalierung basierend auf der MST erklärt
werden kann. Dieses demographische Gleichgewicht von Geburt und Mortalität zwischen
verschiedenen Arten und funktionalen Gruppen steht im Einklang mit der Annahme der
Neutral Theory, gestattet jedoch funktionale Abweichungen für manche Arten. Meine
Untersuchung bringt die Debatte darüber, ob Nischen oder neutrale Mechanismen
natürliche Pflanzengesellschaften strukturieren, in Einklang: die eigentlich Frage sollte
lauten, wann und warum einer dieser Faktoren dominiert.
Das abschließende Kapitel 6 beinhaltet eine generelle Diskussion meiner
Untersuchungen und Fragestellungen. Die Synthese bestehender Theorien wird die
Ökologie in Theorie und Anwendung zukünftig stärken. Alle in meiner Dissertation
durchgeführten Untersuchungen zeigten, dass der Ansatz der individuenbasierten und
musterorientierten Modellierung vielversprechend scheint, diese Synthese zu erreichen.

Chapter 1
General introduction
In the mid-19th century, the word ‘ecology’ (Ökologie) was first coined by Ernst
Haeckel in Morphology of Organisms (1866), and was originally defined as the
study of the relationship of organisms with their environment. The environment
of an organism is comprised of all the external factors that could influence it,
including physical and chemical factors (abiotic) or/and other organisms
(biotic). Despite how precise ecology is defined to date (Table 1.1), it is no
doubt that the interactions with those abiotic and biotic factors are fundamental
on the elaboration of ecological theories.
However, some new ecological theories such as the metabolic scaling
theory (MST) attempts at scaling up from the individuals to the ecosystem,
which is based on simple physical rules but overlooks (consciously or
unconsciously) the important role of such interactions. Although claimed to be
a ‘universal’ theory, the validity and universality of MST are still under debate
because some empirical observations in plant populations and communities
are inconsistent with the predictions of MST.
The major goal of my dissertation is to investigate the effects of different
modes of interactions in driving plant population dynamics. All my studies are
under a context of MST, because I am attempted to integrate the effects of
plant interactions and environmental conditions into metabolic scaling theory,

- 2 - Chapter 1
in order to reconcile metabolic scaling theory with observed variations in
nature.
In the following, I will first introduce the field of plant interactions and
then to the overall approach chosen in this thesis, individual-based and
pattern-oriented modelling. Third, I will discuss two ‘unifying’ theories for which
plant interactions play a key role: the Metabolic Scaling Theory and the Unified
Neutral Theory. Finally, I will outline the structure and content of my thesis.
Table 1.1 Textbook definitions of Ecology (after Scheiner and Willig 2008)
Definition Source
The study of the structure and function of nature Odum (1971)
The scientific study of the relationships between organisms
and their environments
McNaughton and
Wolf (1973)
The study of the natural environment, particularly the
interrelationships between organisms and their surroundings
Ricklefs (1979)
The study of animals and plants in relation to their habits and
habitats
Colinvaux (1986)
The study of the relationship between organisms and their
physical and biological environments
Ehrlich and
Roughgarden (1987)
The study of interactions between organisms and between
organisms and their environments
Stiling (1992)
The study of the relationships, distribution, and abundance of
organisms, or groups of organisms, in an environment
Dodson et al. (1998)
The scientific study of the interactions that determine the
distribution and abundance of organisms
Krebs (2001)
The scientific study of the interactions between organisms
and their environment
Begon et al. (2006)
The study of the relationships between living organisms and
their environments, the interactions of organisms with one
another, and the patterns and causes of the abundance and
distribution of organisms in nature
Gurevitch et al.
(2006)

Chapter 1 - 3 -
1.1 Ecology and interactions
1.1.1 Plant competition
In natural situations, the limitation of resources is the crucial issue that links
organisms with their environment. Resource limitation can be caused by both
abiotic and biotic factors. Competition, which per definition is about limiting
resources, is considered perhaps the most important but not the only form of
biotic interactions, which occurs naturally among living organisms that coexist
in same environment. The importance of crop-weed competition was expound
in the ancient Chinese encyclopedia of agriculture The Valuable Techniques
for The Welfare of People (齊民要術, Qí Mín Yào Shù), which had been
completed between A.D. 533 and 544. Although farmers (and agronomists)
were already aware of the importance of competition in managing their
agro-ecosystems long before, the first academic report about competition was
published in the 14th century (Grace and Tilman 1990). Since Darwin (1859),
competition has been established as the conceptual basis for his concept of
‘struggle for life’. Henceforth, most evolutionary biologists view competition as
the main driving force of ‘natural selection’ and, in turn, of evolution. In plant
ecology, the competition is one of the most important topics: it is believed to be
the main force in driving plant phenotype plasticity, life history evolution,
population dynamics, and community assembly (Tilman 1988, Grace and
Tilman 1990).
However, as Grace and Tilman (1990) emphasized in Perspectives on
Plant Competition, although a great deal of research was focused on the topics
of competition for a long time, it is still not well understood. Even the definition
of competition is not consistent. Many ecologists attempted to give an accurate
and unified definition of competition (Milne 1961, Harper 1977, Grime 1979,
Tilman 1988, Begon et al. 2005), but so far, none of them has been generally
accepted. This terminological confusion indicates the complexity of
competition. Begon et al. (2005) defined competition in a very precise way:

- 4 - Chapter 1
“competition is an interaction between individuals, brought about by a shared
requirement for a resource, and leading to a reduction in the survivorship,
growth and/or reproduction of at least some of the competing individuals
concerned”. I advocate using this definition because it gives both the cause
and the consequence of competition, especially when applying in plant
population ecology.
In plant ecology, competition is usually categorized as asymmetric- or
symmetric competition according to the competitive effects (Weiner 1990,
Schwinning and Weiner 1998, Freckleton and Watkinson 2001, Berger et al.
2008). Asymmetric competition is an unequal or disproportional division of
resources amongst competing plants which can be either inter- or intraspecific.
Therefore, competition may be asymmetric in the sense that some species or
individuals have a competitive advantage over others in taking a
disproportionately large amount of resource. The mechanisms determining the
degree of inter- and intraspecific asymmetry are similar: they are
size-dependent and, related to the nature of the limiting resources (Schwinning
and Weiner 1998, Freckleton and Watkinson 2001). For example, taller plants
will have a disproportionate advantage over smaller individuals when
competing for light, because the limiting resource, light, is ‘pre-emptable’. This
can leads to a growth depression of the latter, which has also been referred to
as ‘dominance and suppression’ and ‘one-sided competition’ (Weiner 1990,
Schwinning and Weiner 1998, Stoll et al. 2002, Berger et al. 2008). In contrast,
when the limiting resources are not ‘pre-emptable’ such as water and nutrient,
plants will share resources equally or proportionally to their size (Schwinning
and Weiner 1998, Berger et al. 2008). It is therefore assumed that competition
for light (aboveground competition) is more size asymmetric whereas
competition for water and nutrient (belowground competition) is more size
symmetric (Schwinning and Weiner 1998, Stoll et al. 2002, Berger et al. 2008).
The particular mode of interactions seems to vary depending on
environmental conditions, and whether the interactions occur above- or

Chapter 1 - 5 -
belowground. The latter strongly depends on the type of the shared resources
and their availability. However, due to logistic and technical difficulties, so far
only few studies exist where belowground competition was explored (Morris
2003, Deng et al. 2006, Berger et al. 2008). Most of the empirical experiments
as well as theoretical models only consider light competition as the crucial
process (but see Casper et al. 2003, May et al. 2009, Schiffers et al. 2011). In
heterogeneous or stressful environments, including intra- and interspecific
competition, the relative effect and importance of above- versus belowground
interactions and how they control the structure and function of populations and
communities are still poorly understood (Berger et al. 2008).
In view of its complexity, ecologists have intensely debated the
relevance of different aspects of competition to particular theories. There is
one debate on the prevalence and importance of competition along
environmental gradient which is commonly mentioned as two opposite theories
of competition, and is usually associated with two of its main protagonists,
Grime and Tilman (for a summary, see e.g. Goldberg et al. 1999, Brooker et al.
2005). One view point suggested that plant competition is more dominant
within plant systems in productive and mild environments, but the role of
competition played within plant systems will decreases when productivity
decreases and environmental gradient increases (Grime 1979, Huston 1979,
Keddy 1989). The opposite view point is that competition is predominant within
plant systems irrespective of system productivity or environmental stress,
while the mechanisms by which plants compete can vary, namely, plants
compete strongly for light or space in productive and mild environments, while
in unproductive and harsh environments plants compete mainly for water or
soil nutrients (Newman 1973, Tilman 1982, 1987, 1988, Grubb 1985). Both
points of view have been supported by empirical evidences, and were
considered to be irreconcilable at their fundamental assumptions.
Grace (1991) pointed out that this debate was primarily aggravated by a
failure of distinguishing between two components of competition, the

- 6 - Chapter 1
‘importance’ and ‘intensity’ of competition. The differences between those two
concepts are explicitly (Welden and Slauson 1986): The intensity of
competition is the absolute impact, which relates directly to the physiological
aspects of individuals, but indirectly and conditionally to their fitness, as well as
the dynamics of populations and the structure of communities. The importance
of competition is also not necessarily correlated with the intensities of other
ecological processes. In contrast, the importance of competition is its impact
relative to the environmental factors that influence individual success, which
relates indirectly to their physiological aspects but directly to the ecology and
fitness of individuals. The importance of competition is necessarily relative to
the importance of other ecological processes. Furthermore, intensity refers to
the impact of present competition on individuals, whereas importance refers to
the products and consequences of past competition on individuals and
possibly the systems.
Although it has been argued that Tilman’s theory concerns intensity
whereas Grime’s theory more emphasize importance (see e.g. Grace 1991),
the concept of importance is still frequently neglected. Such widespread
confusion between intensity and importance also arise within studies of plant
positive interactions (Brooker et al. 2005, Brooker and Kikvidze 2008), and
becomes a barrier to our understandings about the role of interactions may
play in plant populations and communities.
1.1.2 Plant facilitation
Over the last decade or so, the role of plant positive interactions has received
increasing attention and is now widely recognized in both empirical and
theoretical ecology (Bertness and Callaway 1994, Brooker et al. 2008,
Bronstein 2009, Maestre et al. 2009). In plant ecology, positive interactions are
usually referred to as facilitation, which I define here as: the interaction
between individual plants via moderation of biotic and abiotic stress,

Chapter 1 - 7 -
enrichment of resource or increased access to resource, which leads to an
increase in the survivorship, growth and/or reproduction of at least some of the
interacting individuals involved. This definition of facilitation is complementary
to the definition of competition given by Begon et al. (2005; see above).
Facilitation seems to be particularly important under harsh
environmental conditions, i.e. abiotic stress. The ‘stress gradient hypothesis’
(SGH) proposes that competition and facilitation may act simultaneously, but
the relative importance of facilitation and competition will vary inversely along
gradients of physical stress. Under high stress conditions, facilitation should be
dominant over competition in shaping of community structures (Bertness and
Callaway 1994, Brooker et al. 2008, Maestre et al. 2009). The SGH has been
supported by many studies: the interplay between facilitation and competition
can drive population dynamics (Chu et al. 2008, 2009, 2010, McIntire and
Fajardo 2011), community structure (Gross 2008, Xiao et al. 2009), community
diversity (Cavieres and Badano 2009), ecosystem functions (Callaway et al.
2002, Kikvidze et al. 2005), and evolutionary consequences (Bronstein 2009,
McIntire and Fajardo 2011). However, there are also studies which do not
support SGH predictions, as facilitative effects have not been detected in some
extreme stress situations (Tielbörger and Kadmon 2000, Maestre et al. 2005,
2009). The mechanistic explanations for SGH as well as facilitation are not
well presented. This indicates that the conceptual framework underlying the
SGH might need further refinement (Maestre et al. 2009). And in fact, whereas
numerous studies exist that explore the consequences of different modes of
competition, i.e. symmetric versus asymmetric competition (Schwinning and
Weiner 1998, Weiner et al. 2001, Stoll and Bergius 2005, Berger et al. 2008),
so far different modes of facilitation have not been well explored. Inconsistent
definitions of facilitation and the lack of differentiation between the impacts of
plant-plant interactions on beneficiary and benefactor individuals have recently
been identified as important gaps in current research (Brooker et al. 2008,
Bronstein 2009, Brooker and Callaway 2009, Pakeman et al. 2009). Refining

- 8 - Chapter 1
and clarifying the concept of facilitation is crucial for understanding how
facilitation arises, persists and evolves.
1.1.3 The interplay between negative and positive interactions
Theoretically, there are three fundamental forms of interaction between
two individuals (species): neutral (0), positive (+) and negative (−). According
to different definitions, modes of facilitation can be mutualistic (+/+),
commensal (+/0) or even antagonistic (+/−) among plants (Brooker et al. 2008,
Bronstein 2009). If competition and facilitation occur at the same time, there
are six possible combinations: −/−, 0/0, +/+, −/0, +/0 and −/+. Considering the
facts that when individuals are together or apart show different modes of
interaction, the possible combinations could increase to 10 (Table 1.2).
However, an approach which incorporates the existing definitions of both
competition and facilitation as special cases might be more useful. Thus, it is
efficient to completely transfer, albeit conversely, the concept of modes of
competition to the new concept of modes of facilitation (Table 1.2). This new
conceptual model has three advantages: first, it is analogous, and therefore
directly comparable to the widely used and important concept of symmetric
and asymmetric interaction (Schwinning and Weiner 1998, Weiner et al. 2001,
Stoll and Bergius 2005, Berger et al. 2008); second, it offers a quantitative and
operational means of evaluating both competitive and facilitative impacts; and
last, it can help us to integrate the facilitation with competition theories
(Callaway et al. 2002, Berger et al. 2008, Brooker et al. 2008, Bronstein 2009,
Maestre et al. 2009).

Chapter 1 - 9 -
Table 1.2 Modes of interactions among two individuals (or species) (A and B).
Defined according to the effect of this interaction for each organism (positive +,
negative −, or no effect 0). A: asymmetric; S: symmetric; C: competition; F: facilitation.
Forms of interaction Together Apart Possible combinations of
the modes of interaction A B A B
Neutralism 0 0 0 0 Neutrality or SC + SF
Competition − − 0 0 SC
Amensalism 0 − 0 0 AC
Parasitism + − − 0 AC + AF
Commensalism + 0 − 0 AF
Mutualism + + − − SF
Unnamed + + 0 − SF or AF
Proto-cooperation + + 0 0 SF
Unnamed + − 0 0 AC + AF
Unnamed + 0 0 0 AF

- 10 - Chapter 1
1.2 Ecology, individual-based models and pattern-oriented
modelling
A main goal of ecology as a science is to understand the interactions of
organisms among themselves and their environment. Individual-based models
(IBMs, or also known as agent-based models ABMs in social science) are thus
a natural tool for ecology because IBMs are capable of taking into account
local interactions, individual variability, adaptive behaviour, environmental
heterogeneous, abiotic stress, disturbance and other ecological factors
(DeAngelis and Gross 1992, Grimm 1999, Grimm and Railsback 2005, Grimm
et al. 2005, Berger et al. 2008).
A model is a simplified version of the real world only dealing with a
limited number of factors which are considered most relevant. Ecological
models attempt to capture the essences of ecological system for addressing
specific questions about the given system (Grimm and Railsback 2005). The
systems of interest in ecology are usually made up of myriad interacting
organisms. Organisms are so different from each other, interacting with others
and physical environment in their unique ways. Grimm and Railsback (2005)
argued that ecology and biology are still lacking in strong mathematical tools to
understand and predict such individual-based complex systems.
With the development of computer science, bottom-up simulation
modelling approaches have been developed a lot in ecology. Using bottom-up
approaches, ecologists compile relevant information at a lower level of the
system, formulate and implement rules about individual’s behaviour in a
computer simulation, and then observe and understand the emergence of
properties related to particular questions at a higher level of integration (Grimm
et al. 2005). In particular, individual-based modelling follows the bottom-up
framework and helped in understanding emergent properties of complex
systems such as population dynamics or community assembly out of the
ecological traits, behaviours and interactions of individual organisms

Chapter 1 - 11 -
(DeAngelis and Gross 1992, Breckling et al. 2005, Grimm and Railsback
2005).
In plant ecology, individual-based approaches are widely used for
modelling plant interactions (Czárán 1997, Berger et al. 2008). In a recent
review, Berger et al. (2008) comprehensively compared different approaches.
Based on Berger et al.’s classification, there are three important approaches
which I think are most advisable: cellular automaton (CA, or the so called
grid-based models), zone-of-influence (ZOI), and field-of-neighbourhood (FON)
(Berger and Hildenbrandt 2000, 2003, Berger et al. 2004). These approaches,
from simple to more sophisticated, have their advantages and limitations and
should be applied to different research questions accordingly (see Berger et al.
2008 for details). However, existing modelling approaches for investigating
plant interactions show three major gaps. New approaches are needed that: (i)
help understanding the mechanisms of plants’ development and response to
their local environment, (ii) consider plants’ adaptations to changing
environmental conditions, and (iii) simultaneously consider positive and
negative interactions among neighbouring plants as well as above- and
belowground interactions.
Modelers greatly rely on the principle of parsimony in developing
models, known as the ‘Occam’s razor’. However, the Occam‘s razor should be
use appropriately in individual-based modelling, because the relationship
between a model’s complexity and payoff is no longer monotonically negative
(as assumed for classical models), but really a hump-shape (Fig 1.1). John
Holland (1995) had formulated this as “Model building is the art of selecting
those aspects of a process that are relevant to the question being asked”. His
interpretation not only highlights the principle of parsimony, but also highlights
that the question being asked is the element of the scientific problem that
should be referenced to determine the components of a model.

- 12 - Chapter 1
Fig. 1.1 A model’s payoff (in terms of how much we can learn from it) versus its
complexity (after Grimm and Railsback 2005). In contrast to analytical models of
classical theoretical ecology (dashed line), where payoff is high only for very simple
models and then declines continuously with complexity, IBMs and other bottom-up
simulation models (humped curve) have a “Medawar zone” at intermediate complexity
where payoff is maximized.
IBMs should be neither too simple nor too complex if they are to be
useful (Grimm and Railsback 2005). To make the bottom-up modelling and
individual-based modelling more rigorous and comprehensive, a strategy
named pattern oriented modelling (POM) was developed (Grimm et al. 1996,
2005, Wiegand et al. 2003, Grimm and Railsback 2005). Because patterns are
the observations of any kind showing specialized, sustained, repeated and
nonrandom characteristics of the system, which contain information on the
mechanisms how they emerge from lower level and the essential structures
and processes, therefore could be used to guide model design (Grimm et al.
1996, 2005, Wiegand et al. 2003, Grimm and Railsback 2005).
The protocol of POM includes four steps (Wiegand et al. 2003, Grimm
and Railsback 2005, Piou 2007): (1) “aggregation of individual based biological
information” for model construction, (2) “determination of parameter values”, (3)

Chapter 1 - 13 -
“systematic comparison between the observed pattern and the simulated
pattern produced by the model” for model evaluation, and (4) “secondary
predictions”. The steps (1) and (2) ensure that properties or patterns at higher
levels emerge from lower-level information incorporated in IBMs. The steps (3)
and (4) leading to the increase in reliability of the IBMs. In this sense, POM is
useful not just for individual-based modelling, but for any kind of modelling. In
addition, POM can also be used to test and refine theory, and can help to
advance a synthesis of diverse theories across scales (Fig 1.2).
Fig. 1.2 Synthesis of theories based on pattern oriented modelling approach.
Synthesis of Theories
Hypothesis
IBMs design
Observed Patterns
Deduction Induction

- 14 - Chapter 1
1.3 Are there universal laws or unified theories in ecology?
More than a decade ago, John Lawton (1999) wrote that, “Parts of science,
areas of physics for instance, have deep universal laws, and ecology is deeply
envious because it does not”. This kind of discontent is perhaps driven by a
fact that use of the laws and models in ecology are usually very local and
highly contingent, and the system studied in ecology are too complex and
contingent to apply the general laws and models.
However, this pessimism regarding a theoretical foundation of ecology
represents a misplaced ‘physical envy’ (Cohen 1971). It is misplaced on at
least two aspects: first, in ecology, the building blocks of systems are living
organisms not particles (Grimm and Railsback 2005). Open a bottle of perfume
in a closed room, the liquid of perfume will become a smelly cloud and fills up
the room eventually, which is state by the second law of thermodynamics.
However, organisms try their best to avoid this smelly cloud of equilibrium,
they can reproduce, interacting with others and environment, seeking fitness,
acquiring resources to maintain internal order and build complexity by
exporting entropy. Erwin Schrödinger (1944) also pondered these questions
and described the negative entropy among organisms: “The essential thing in
metabolism is that the organism succeeds in freeing itself from all the entropy it
cannot help producing while alive”. It is metabolism that makes living
organisms to be unique creatures. Second, the laws of physics are all ‘ceteris
paribus’ laws assumed in ideal vacuums, but the actual governance of, for
instance, particle movement is much more complicated than these general
laws would imply (Cartwright 1983, Simberloff 2004). Therefore, ecology
cannot and should not just try and copy physics.
Over the last decade, several new theories emerged which address the
core principles of ecology, such as Metabolic scaling theory (Brown et al.
2004), Neutral theory in ecology (Hubbell 2001), Ecological stoichiometry
(Sterner and Elser 2002), Maximum entropy of ecology (Harte 2011), Dynamic

Chapter 1 - 15 -
energy budget theory (Kooijman 2009), etc., all having their advantages and
shortages but claiming to be the ‘general’, ‘universal’ or ‘unified’ theory. There
are two striking theories (Gaston and Chown 2005), which attract me most
among all those theories. The first one is Metabolic Scaling Theory (MST)
(also named as Metabolic theory of ecology, Brown et al. 2004), which is an
attempt to link physiological processes of individual organisms with
macroecology. Based on a fractal model of circulatory networks, such as the
vascular system in animals and plants, MST predicts how metabolic rate
increases with body size and temperature following a quarter-power scaling
law. This simple scaling law has been observed from intracellular levels to
individual levels of mammals, covering 27 orders of magnitude (West and
Brown 2005), this even more than the span between earth and whole galaxy
(18 orders of magnitude). Consequently, a large numbers of studies have
explored the ecological consequences of scaling relationships between
organism body size and developmental time, reproduction, population energy
use, abundance, population demographic rate, community structure and
species diversity. Many of these studies were published in high-profile journals
which reflect the promise and potential significance of such a general theory.
However, like any other claim for a ‘universal’ theory, MST has also
generated controversy. For instance, MST predicts a ‘-4/3 universal scaling
law’ for plant mean mass-density relationships (Enquist et al. 1998), but
empirical observations are more variable. Empirical evidences from harsh
areas or from plantation experiments with low resource levels often deviate
from these predictions and show significantly less negative exponents, i.e.
shallower slopes of the self-thinning trajectory or log mass-log density
relationship, respectively (Morris 2003, Deng et al. 2006, Liu et al. 2006).
Consequently, both the validity and universality of scaling exponents and MST
are still unclear and require further analyses of the underlying mechanisms
(Coomes 2006, Deng et al. 2006, Coomes et al. 2011).

- 16 - Chapter 1
A core assumption of MST is that the processes internal to individuals
determine mass-density relationships, whereas interactions among individuals
are largely ignored. Therefore, population dynamic and community assembly
in MST are just a spatial packing process (Reynolds and Ford 2005). An
alternative view is that internal mechanism may play an important role and set
limits on mass-density relationships, but that ecological interactions can be
more important in determining the relationships in the field. Thus, variation in
the ecological conditions and the interactions among individuals can explain
the observed variation in scaling exponents. Specifically, it has been argued
that competition among plants will change mass-density relationships from
those predicted by MST (Coomes et al. 2011).
The second significant theory recently development in ecology is the
Unified Neutral Theory of biodiversity and biogeography (UNT) (Hubbell 2001).
UNT is based on a seemingly unrealistic assumption of ecological equivalence,
that all individuals are functionally equivalent on a per capita basis, i.e. with
respect to their birth, death, dispersal and speciation. Even so, it successfully
explains many patterns about the relative species abundance across different
communities (Hubbell 2001, Chave 2004, Gaston and Chown 2005). UNT
emphasizes ecological drift, dispersal and speciation as the main drivers of
community assembly, thus the abundance of individuals of all species should
be a ‘martingale’ in communities. It severely bucks the basis of ecology by
leaving niche differences and, hence, selection out.
Not surprisingly, criticism in this theory has been predominantly directed
at the fundamental assumption of neutrality. Hubbell (Hubbell 2001, 2005,
2006, 2008) argue that even though the equivalence assumption is not
apposite, the neutral model heavily relies on the principle of parsimony and
predicts real patterns in nature, therefore should not be misjudged. However,
in a POM view, the neutral model is located apparently on left side of the
‘Medawar zone’. It is too simple hence fails in reality and justification that
species are apparently different in varied aspects, and some details on the

Chapter 1 - 17 -
assumption are needed. Indeed, the neutral model stimulated ecologists to
revise the equivalence assumption. Some researchers relaxed the strict
neutrality assumption by considering fitness equivalence via ecological
trade-offs (K. Lin et al. 2009). Their modified neutral models clearly showed
that birth-death trade-offs can lead to a similar result as strict neutral model did.
However, the mechanisms underlying this demographic trade-off are still
unknown.
As Simon Levin (2000) laid out, “the most important challenge for
ecologists remains to understand the linkages between what is going on at the
level of the physiology and behaviour of individual organisms and emergent
properties such as the productivity and resiliency of ecosystems”. Both MST
and UNT or similar theories that are based on simple physical rules attempt to
scale up from the individual to the system. Predictions made by those theories
sometimes become invalid in nature because macroscopic features of the
system can emerge from the large numbers of interacting individual organisms.
Individual-based ecology, with IBM and POM (Grimm and Railsback 2005,
Grimm et al. 2005), I believe, can help us to ultimately achieve a synthesis of
theories in ecology which focus on different processes, organization levels, or
scales.

- 18 - Chapter 1
1.4 Objectives and content of the dissertation
My major goal in this dissertation is to investigate the effects of different modes
of interactions in driving plant population dynamics. Specifically, I explored the
relative role of asymmetric- versus symmetric competition, above- versus
belowground competition, and negative versus positive interactions in shaping
plant population along the environmental gradients by using spatially-explicit
individual-based models. The pattern-oriented modelling approach is applied
in my individual-based modelling studies. Some of the model predictions are
tested with greenhouse experiments and field studies. Although my studies in
this dissertation are mostly on mono-population dynamics, the models and
theories presented are general, so that in principle they can be extended to
communities. All my studies are under a context of metabolic scaling theory
(MST), because I am attempted to integrate the effects of plant interactions
and environmental conditions into metabolic scaling theory, in order to
reconcile metabolic scaling theory with observed variations in nature.
First of all, I derived a general ontogenetic growth model for vascular
plants which is based on energy conservation and physiological process of
individual plant (see Appendix A in Chapter 2). This new mechanistic model is
thus different from the phenomenological growth models which are generally
used in individual-based models of plant interactions. Some extensions and
predictions based on the new growth model are tested by empirical data. My
individual-based simulation models are developed on the basis of this general
growth model.
In Chapter 2, I present a generic spatially-explicit individual-based
model, which I named as pi (plant interaction) model. The pi model implements
different modes of competition (from completely asymmetric to completely
symmetric) among individual plants via their overlapping zone-of-influence
(ZOI). I used the ZOI approach because it is easy to implement and to
combine with metabolic scaling theory. The pi model was used here to

Chapter 1 - 19 -
investigate the hypothesis that MST may be compatible with the observed
variation in plant self-thinning trajectories if different modes of competition and
different resource availabilities are considered. Specifically, two hypotheses
were investigated by using a one-layer ZOI model: (1) if surviving plants are
not highly affected by local interactions, then individual-level metabolic
processes can predict population-level mass-density relationships; and (2)
size-symmetric competition (e.g. belowground competition) will lead to
shallower self-thinning trajectories.
Recent studies on plant competition and MST imply that not only plant
interactions (Coomes et al. 2011, Rüger and Condit 2012) but also the plastic
biomass allocation to roots or shoots of plant can affect mass-density
relationship (Morris 2003, Deng et al. 2006, Zhang et al. 2011). However, the
relative roles of those two factors, competition and plastic biomass allocation,
are still unknown. In Chapter 3, both a theoretical model and an experiment
are used to answer this question. Since the one-layer pi model is too simple,
though, to explore the relative role of plasticity of biomass allocation and
below- versus aboveground competition, I developed a new two-layer model
which represents both above- and belowground competition simultaneously
via independent ZOIs. In the tow-layer pi model, plant growth and biomass
allocation are represented by the growth function based on MST (Enquist 2002,
Niklas 2005, Lin et al. 2012), which tries to mechanistically capture the plastic
responses of plants to changing environmental conditions. In addition, a
greenhouse experiment with tree seedlings is employed to evaluate the
behaviour of the simulated plants related to the allocation patterns and to
validate our model predictions regarding the mass-density relationship.
Specifically, I focus on the following research questions: (1) how does plant
phenotype plasticity (root/shoot biomass allocation in heterogeneous
environments) affect emergent patterns observed at the system level (e.g.,
variation of plant mass-density relationship)? (2) What are the resulting effects

- 20 - Chapter 1
of above- and belowground competition modified by plants’ morphological
adaptations to environmental severity on population dynamics?
In Chapter 4, the effects of both negative and positive plant interactions
on population spatial dynamics are investigated. I introduce a new concept of
symmetric vs. asymmetric facilitation and present the modified pi model. This
new model is able to simultaneously implements different modes of both
facilitation and competition among individual plants. Because different modes
of facilitation are considered as a continuum related to the environment, I
explored this concept within the context of the stress gradient hypothesis
(Bertness and Callaway 1994, Brooker et al. 2008). Specifically, I address the
following questions at both the plant population and individual levels: (1) How
does the interplay of different modes of competition and facilitation change
spatial pattern formation during self-thinning in conspecific cohorts that initially
have a random or aggregated distribution; and (2) How do combinations of
modes of competition and facilitation alter the intensity of local plant
interactions along a stress gradient?
In Chapter 5, I attempt to combine neutral theory and metabolic scaling
theory. Demographic equivalence regarding birth-death trade-offs between
different species is consistent with the assumption of neutral theory but allows
differences between species as suggested by niche theory (Lin et al. 2009).
Based on MST, I deduced an allometric scaling rule which can explain
observed demographic trade-offs. In this preliminary study, I tested the validity
of deduced demographic trade-off by a broad array of plant species. The
ultimately aim of my study is the synthesis of existing theories to strengthen
future ecology in theory and application.
The concluding Chapter 6 offers the general discussion and outlook
related to my study topics. An opinionated review about the development of
theories in ecology is also presented.

Chapter 1 - 21 -
References
Begon, M., C. R. Townsend, and J. L. Harper. 2005. Ecology: From Individuals to
Ecosystems. John Wiley & Sons.
Berger, U., and H. Hildenbrandt. 2000. A new approach to spatially explicit modelling of
forest dynamics: spacing, ageing and neighbourhood competition of mangrove trees.
Ecological Modelling 132:287–302.
Berger, U., and H. Hildenbrandt. 2003. The strength of competition among individual trees
and the biomass-density trajectories of the cohort. Plant Ecology 167:89–96.
Berger, U., H. Hildenbrandt, and V. Grimm. 2004. Age‐related decline in forest
production: modelling the effects of growth limitation, neighbourhood competition and
self‐thinning. Journal of Ecology 92:846–853.
Berger, U., C. Piou, K. Schiffers, and V. Grimm. 2008. Competition among plants:
concepts, individual-based modelling approaches, and a proposal for a future
research strategy. Perspectives in Plant Ecology, Evolution and Systematics
9:121–135.
Bertness, M. D., and R. Callaway. 1994. Positive interactions in communities. Trends in
Ecology & Evolution 9:191–193.
Breckling, B., F. Müller, H. Reuter, F. Hölker, and O. Fränzle. 2005. Emergent properties
in individual-based ecological models—introducing case studies in an ecosystem
research context. Ecological Modelling 186:376–388.
Bronstein, J. L. 2009. The evolution of facilitation and mutualism. Journal of Ecology
97:1160–1170.
Brooker, R., Z. Kikvidze, F. I. Pugnaire, R. M. Callaway, P. Choler, C. J. Lortie, and R.
Michalet. 2005. The importance of importance. Oikos 109:63–70.
Brooker, R. W., and R. M. Callaway. 2009. Facilitation in the conceptual melting pot.
Journal of Ecology 97:1117–1120.
Brooker, R. W., and Z. Kikvidze. 2008. Importance: an overlooked concept in plant
interaction research. Journal of Ecology 96:703–708.
Brooker, R. W., F. T. Maestre, R. M. Callaway, C. L. Lortie, L. A. Cavieres, G. Kunstler, P.
Liancourt, K. Tielb rger, J. M. . Travis, and F. Anthelme. 2008. Facilitation in plant
communities: the past, the present, and the future. Journal of Ecology 96:18–34.
Brown, J. H., J. F. Gillooly, A. P. Allen, V. M. Savage, and G. B. West. 2004. Toward a
metabolic theory of ecology. Ecology 85:1771–1789.
Callaway, R. M., R. W. Brooker, P. Choler, Z. Kikvidze, C. J. Lortie, R. Michalet, L. Paolini,
F. I. Pugnaire, B. Newingham, and E. T. Aschehoug. 2002. Positive interactions
among alpine plants increase with stress. Nature 417:844–848.
Cartwright, N. 1983. How the laws of physics lie. Cambridge University Press.
Casper, B. B., H. J. Schenk, and R. B. Jackson. 2003. Defining a plant’s belowground
zone of influence. Ecology 84:2313–2321.
Cavieres, L. A., and E. I. Badano. 2009. Do facilitative interactions increase species
richness at the entire community level? Journal of Ecology 97:1181–1191.
Chave, J. 2004. Neutral theory and community ecology. Ecology Letters 7:241–253.

- 22 - Chapter 1
Chu, C. J., F. T. Maestre, S. Xiao, J. Weiner, Y. S. Wang, Z. H. Duan, and G. Wang. 2008.
Balance between facilitation and resource competition determines biomass–density
relationships in plant populations. Ecology Letters 11:1189–1197.
Chu, C. J., J. Weiner, F. T. Maestre, Y. S. Wang, C. Morris, S. Xiao, J. L. Yuan, G. Z. Du,
and G. Wang. 2010. Effects of positive interactions, size symmetry of competition and
abiotic stress on self-thinning in simulated plant populations. Annals of Botany
106:647–652.
Chu, C. J., J. Weiner, F. T. Maestre, S. Xiao, Y. S. Wang, Q. Li, J. L. Yuan, L. Q. Zhao, Z.
W. Ren, and G. Wang. 2009. Positive interactions can increase size inequality in
plant populations. Journal of Ecology 97:1401–1407.
Cohen, J. E. 1971. Mathematics as metaphor. Science 172:674–675.
Coomes, D. A. 2006. Challenges to the generality of WBE theory. Trends in ecology &
evolution 21:593–596.
Coomes, D. A., E. R. Lines, and R. B. Allen. 2011. Moving on from Metabolic Scaling
Theory: hierarchical models of tree growth and asymmetric competition for light.
Journal of Ecology 99:748–756.
Czárán, T. 1997. Spatiotemporal models of population and community dynamics.
Springer.
DeAngelis, D. L., and L. J. Gross. 1992. Individual-based models and approaches in
ecology: populations, communities and ecosystems. Chapman & Hall.
Deng, J. M., G. X. Wang, E. C. Morris, X. P. Wei, D. X. Li, B. Chen, C. M. Zhao, J. Liu, and
Y. Wang. 2006. Plant mass–density relationship along a moisture gradient in
north-west China. Journal of Ecology 94:953–958.
Enquist, B. J. 2002. Universal scaling in tree and vascular plant allometry: toward a
general quantitative theory linking plant form and function from cells to ecosystems.
Tree Physiology 22:1045–1056.
Enquist, B. J., J. H. Brown, and G. B. West. 1998. Allometric scaling of plant energetics
and population density. Nature 395:163–165.
Freckleton, R. P., and A. R. Watkinson. 2001. Asymmetric competition between plant
species. Functional Ecology 15:615–623.
Gaston, K. J., and S. L. Chown. 2005. Neutrality and the niche. Functional Ecology
19:1–6.
Goldberg, D. E., T. Rajaniemi, J. Gurevitch, and A. Stewart-Oaten. 1999. Empirical
approaches to quantifying interaction intensity: competition and facilitation along
productivity gradients. Ecology 80:1118–1131.
Grace, J. B. 1991. A clarification of the debate between Grime and Tilman. Functional
Ecology 5:583–587.
Grace, J. B., and D. Tilman. 1990. Perspectives on plant competition. Blackburn Press.
Grime, J. P. 1979. Plant strategies and vegetation processes. John Wiley & Sons.
Grimm, V. 1999. Ten years of individual-based modelling in ecology: what have we
learned and what could we learn in the future? Ecological modelling 115:129–148.
Grimm, V., K. Frank, F. Jeltsch, R. Brandl, J. Uchmański, and C. Wissel. 1996.
Pattern-oriented modelling in population ecology. Science of the Total Environment
183:151–166.

Chapter 1 - 23 -
Grimm, V., and V. G. & S. F. Railsback. 2005. Individual-Based Modeling and Ecology.
Princeton University Press.
Grimm, V., E. Revilla, U. Berger, F. Jeltsch, W. M. Mooij, S. F. Railsback, H. H. Thulke, J.
Weiner, T. Wiegand, and D. L. DeAngelis. 2005. Pattern-oriented modeling of
agent-based complex systems: lessons from ecology. Science 310:987–991.
Grubb, P. J. 1985. Plant populations and vegetation in relation to habitat, disturbance and
competition: problems of generalization. In The population structure of vegetation, pp.
595-621
Harper, J. L. 1977. Population biology of plants. Academic Press.
Harte, J. 2011. Maximum Entropy and Ecology: A Theory of Abundance, Distribution, and
Energetics. Oxford University Press, USA.
Holland, J. H. 1995. Hidden Order: How Adaptation Builds Complexity. Basic Books.
Hubbell, S. P. 2001. The unified neutral theory of biodiversity and biogeography.
Princeton University Press.
Hubbell, S. P. 2005. Neutral theory in community ecology and the hypothesis of functional
equivalence. Functional ecology 19:166–172.
Hubbell, S. P. 2006. Neutral theory and the evolution of ecological equivalence. Ecology
87:1387–1398.
Hubbell, S. P. 2008. Approaching ecological complexity from the perspective of symmetric
neutral theory. Tropical forest community ecology. Wiley-Blackwell, Chichester,
UK:143–159.
Huston, M. 1979. A general hypothesis of species diversity. American naturalist:81–101.
Keddy, P. A. 1989. Effects of competition from shrubs on herbaceous wetland plants: a
4-year field experiment. Canadian Journal of Botany 67:708–716.
Kikvidze, Z., F. I. Pugnaire, R. W. Brooker, P. Choler, C. J. Lortie, R. Michalet, and R. M.
Callaway. 2005. Linking patterns and processes in alpine plant communities: a global
study. Ecology 86:1395–1400.
Kooijman, B. 2009. Dynamic Energy Budget Theory for Metabolic Organisation, 3rd
edition. Cambridge University Press.
Lawton, J. H. 1999. Are there general laws in ecology? Oikos 84:177–192.
Levin, S. A. 2000. Fragile Dominion: Complexity and the Commons. Basic Books.
Lin, K., D. Y. Zhang, and F. He. 2009. Demographic trade-offs in a neutral model explain
death-rate-abundance-rank relationship. Ecology 90:31–38.
Lin, Y., U. Berger, V. Grimm, and Q.-R. Ji. 2012. Differences between symmetric and
asymmetric facilitation matter: exploring the interplay between modes of positive and
negative plant interactions. Journal of Ecology 100:1482–1491.
Liu, J., L. Wei, C. M. Wang, G. X. Wangl, and X. P. Wei. 2006. Effect of Water Deficit on
Self‐thinning Line in Spring Wheat (Triticum aestivum L.) Populations. Journal of
Integrative Plant Biology 48:415–419.
Maestre, F. T., R. M. Callaway, F. Valladares, and C. J. Lortie. 2009. Refining the
stress-gradient hypothesis for competition and facilitation in plant communities.
Journal of Ecology 97:199–205.

- 24 - Chapter 1
Maestre, F. T., F. Valladares, and J. F. Reynolds. 2005. Is the change of plant–plant
interactions with abiotic stress predictable? A meta‐analysis of field results in arid
environments. Journal of Ecology 93:748–757.
May, F., V. Grimm, and F. Jeltsch. 2009. Reversed effects of grazing on plant diversity:
the role of belo-ground competition and size symmetry. Oikos 118:1830–1843.
McIntire, E. J., and A. Fajardo. 2011. Facilitation within Species: A Possible Origin of
Group-Selected Superorganisms. The American naturalist 178:88–97.
Milne, A. 1961. Definition of competition among animals. Pages 40–61 Symposia of the
Society for Experimental Biology. Cambridge University Press.
Morris, E. C. 2003. How does fertility of the substrate affect intraspecific competition?
Evidence and synthesis from self-thinning. Ecological Research 18:287–305.
Newman, E. I. 1973. Competition and diversity in herbaceous vegetation. Nature 244:310.
Niklas, K. J. 2005. Modelling below-and above-ground biomass for non-woody and woody
plants. Annals of Botany 95:315–320.
Pakeman, R. J., F. I. Pugnaire, R. Michalet, C. J. Lortie, K. Schiffers, F. T. Maestre, and J.
M. . Travis. 2009. Is the cask of facilitation ready for bottling? A symposium on the
connectivity and future directions of positive plant interactions. Biology Letters
5:577–579.
Piou, C. 2007. Patterns and individual-based modeling of spatial competition within two
main components of Neotropical mangrove ecosystems. PhD Thesis, University of
Bremen.
Reynolds, J. H., and E. D. Ford. 2005. Improving competition representation in theoretical
models of self‐thinning: a critical review. Journal of Ecology 93:362–372.
Rüger, N., and R. Condit. 2012. Testing metabolic theory with models of tree growth that
include light competition. Functional Ecology 26:759–765.
Schiffers, K., K. Tielbörger, B. Tietjen, and F. Jeltsch. 2011. Root plasticity buffers
competition among plants: theory meets experimental data. Ecology 92:610–620.
Schrödinger, E. 1944. What is life?: The physical aspect of the living cell. Cambridge
University press.
Schwinning, S., and J. Weiner. 1998. Mechanisms determining the degree of size
asymmetry in competition among plants. Oecologia 113:447–455.
Simberloff, D. 2004. Community ecology: Is it time to move on? The American Naturalist
163:787–799.
Sterner, R. W., and R. W. S. J. J. Elser. 2002. Ecological Stoichiometry: The Biology of
Elements from Molecules to the Biosphere. Princeton University Press.
Stoll, P., J. Weiner, H. Muller-Landau, E. Müller, and T. Hara. 2002. Size symmetry of
competition alters biomass–density relationships. Proceedings of the Royal Society of
London. Series B: Biological Sciences 269:2191–2194.
Tielbörger, K., and R. Kadmon. 2000. Temporal environmental variation tips the balance
between facilitation and interference in desert plants. Ecology 81:1544–1553.
Tilman, D. 1982. Resource Competition and Community Structure. Princeton University
Press.
Tilman, D. 1987. On the meaning of competition and the mechanisms of competitive
superiority. Functional Ecology 1:304–315.

Chapter 1 - 25 -
Tilman, D. 1988. Plant strategies and the dynamics and structure of plant communities.
Princeton University Press.
Weiner, J. 1990. Asymmetric competition in plant populations. Trends in Ecology &
Evolution 5:360–364.
Welden, C. W., and W. L. Slauson. 1986. The intensity of competition versus its
importance: an overlooked distinction and some implications. Quarterly Review of
Biology:23–44.
West, G. B., and J. H. Brown. 2005. The origin of allometric scaling laws in biology from
genomes to ecosystems: towards a quantitative unifying theory of biological structure
and organization. Journal of Experimental Biology 208:1575–1592.
Wiegand, T., F. Jeltsch, I. Hanski, and V. Grimm. 2003. Using pattern-oriented modeling
for revealing hidden information: a key for reconciling ecological theory and
application. Oikos 100:209–222.
Zhang, W.P., X. Jia, Y.Y. Bai, and G.X. Wang. 2011. The difference between above- and
below-ground self-thinning lines in forest communities. Ecological Research
26:819–825.

Chapter 2
Plant competition and metabolic scaling theory *
Abstract
Metabolic scaling theory (MST) is an attempt to link physiological processes of
individual organisms with macroecology. It predicts a power law relationship
with an exponent of -4/3 between mean individual biomass and density during
density-dependent mortality (self-thinning). Empirical tests have produced
variable results, and the validity of MST is intensely debated. MST focuses on
organisms' internal physiological mechanisms but we hypothesize that
ecological interactions may be more important in determining plant
mass-density relationships induced by density. We employ an individual-based
model of plant stand development that includes three elements: a model of
individual plant growth based on MST, different modes of local competition
(size-symmetric vs. -asymmetric), and different resource levels. Our model is
consistent with the observed variation in the slopes of self-thinning trajectories.
Slopes were significantly shallower than -4/3 if competition was
size-symmetric. We conclude that when the size of survivors is influenced by
strong ecological interactions, these can override predictions of MST, whereas
* A slightly revised version of this chapter has been submitted to PLoS ONE (status:
minor revision) as Yue Lin1, 2
, Uta Berger1, Volker Grimm
2, 3, Franka Huth
4, and Jacob
Weiner5: Plant interactions alter the predictions of metabolic scaling theory
1 Institute of Forest Growth and Computer Science, Dresden University of Technology,
P.O. 1117, 01735 Tharandt, Germany; 2 Helmholtz Centre for Environmental Research –
UFZ, Department of Ecological Modelling, 04318 Leipzig, Germany; 3 Institute for
Biochemistry and Biology, University of Potsdam, Maulbeerallee 2, 14469 Potsdam,
Germany; 4 Institute of Silviculture and Forest Protection, Dresden University of
Technology, 01735 Tharandt, Germany; 5 Department of Plant and Environmental
Sciences, University of Copenhagen, DK-1958 Frederiksberg, Denmark

Chapter 2 - 27 -
when surviving plants are less affected by interactions, individual-level
metabolic processes can scale up to the population level. MST, like
thermodynamics or biomechanics, sets limits within which organisms can live
and function, but there may be stronger limits determined by ecological
interactions. In such cases MST will not be predictive.
Keywords: mass-density relationships, plant competition, self-thinning,
size-symmetric competition, zone-of-Influence model
2.1 Introduction
Metabolic Scaling Theory (MST) offers a quantitative framework for linking
physiological processes of individual organisms with higher-level dynamics of
populations and communities. It predicts that an individual’s metabolic rate, B,
scales with body mass, m, as m3/4 (West et al. 1999). For plants, it is assumed
that B is proportional to their rate of resource use, Q, and increases with body
mass, m, as BQm3/4 (Enquist et al. 1998). When the rate of resource supply,
R, per unit area is held constant, the relationship between maximum
population density, N, and mean body mass is predicted to be mN-4/3. Thus, if
mass-density relationships during self-thinning reflect MST, the relation
between m and N is predicted to be a power law with a mass–density scaling
exponent of –4/3.
While some empirical observations seem to be consistent with this
prediction (Enquist et al. 1998, Enquist 2002), data from self-thinning
populations are more variable (Morris 2003, Deng et al. 2006, Dai et al. 2009,
Zhang et al. 2011). Especially the data from arid regions or areas with low
resource levels often deviate from the predictions of MST and show
significantly shallower trajectories, i.e. less negative exponents (Morris 2003,
Deng et al. 2006). While some researchers assume that the mass–density
scaling exponent is universal but disagree about the correct value, others
argue that there is real biological variation in the exponent, thus questioning
the generality of MST (Deng et al. 2006, Coomes and Allen 2007, Coomes et
al. 2011, Rüger and Condit 2012).

- 28 - Chapter 2
A core assumption of MST is that processes internal to individuals
determine mass-density relationships. An alternative view is that internal
mechanism may play an important role and set limits on mass-density
relationships, but that ecological interactions can be more important in
determining the relationships in the field. Thus, variation in the ecological
conditions can explain the observed variation in scaling exponents.
Specifically, it has been argued that competition among plants will change
mass-density relationships from those predicted by MST (Morris 2003,
Coomes et al. 2011). The well-documented plasticity of plant form in response
to competition (Weiner and Thomas 1992, Dai et al. 2009) suggests that
competitive interactions could affect mass-density relationships.
Many empirical studies on plant mass-density relationships have based on
data where competition for light dominates (Enquist et al. 1998, Enquist and
Niklas 2001, Deng et al. 2006), but in areas where belowground resources
such as nutrients and water are more limiting canopies can remain unclosed.
In such areas belowground competition may affect growth and mortality much
more than aboveground competition (Morris 2003, Deng et al. 2006, Allen et al.
2008).
Below- and aboveground competition are qualitatively different.
Aboveground, the limiting resource, light, is directional and therefore
“pre-emptable”, i.e. taller plants will have a disproportionate advantage over
smaller individuals when competing for light, which has also been referred to
as "size-asymmetric competition", “dominance and suppression” or “one-sided
competition” (Schwinning and Weiner 1998, Stoll et al. 2002, Berger et al.
2008). In contrast, belowground resources such as water and nutrients are not
generally pre-emptable so that competing plants tend to share belowground
resources in proportion to their sizes. There is much evidence that
aboveground competition tends to be size-asymmetric, while belowground
competition is more size-symmetric (Schwinning and Weiner 1998, Stoll et al.
2002, Berger et al. 2008). This could influence mass-density relationships.
There is evidence to support this claim. For example, for the desert shrub
Larrea tridentata, the individuals’ allometric growth and root-shoot biomass
allocation patterns are consistent with MST, but the log mass - log density
relationship is shallower than predicted by MST with a substantial variation

Chapter 2 - 29 -
(Allen et al. 2008). This suggests that belowground competition, which is more
size-symmetric, may leads to shallower self-thinning trajectories. Results from
an individual-based Zone-of-Influence plant population model indicate that the
size-symmetry or asymmetry of competition will affect self-thinning trajectories
(Stoll et al. 2002, Chu et al. 2010). These studies used a phenomenological
model for individual plant growth (Weiner et al. 2001) that does not
accommodate the physical and biological principles of MST. And indeed, the
range of slopes produced by Chu et al.'s model (Chu et al. 2010), from -0.820
to 1.609, is larger than the range observed in the field. For example, in 1266
plots within six biomes and 17 forest types across China, the estimated log
mass - log density slopes ranged from -1.103 to -1.441 (Li et al. 2006).
We hypothesize that MST may be compatible with the observed variation in
self-thinning trajectories if different modes of competition and different
resource availabilities are considered. We investigate two hypotheses: 1,
size-symmetric competition (e.g. belowground competition) will lead to
shallower self-thinning trajectories. 2, Individual-level metabolic processes can
predict population-level mass-density relationships if surviving plants are not
highly affected by local interactions.
To investigate our hypothesis, we modify a widely used individual-based
Zone-of-Influence model of individual growth and competition, in which
competition can be size-symmetric or -asymmetric (Weiner et al. 2001). To
make our model compatible with the assumptions of MST, we use an
individual growth model and allometric relationships derived from MST
(Appendix A, Lin et al. 2012).
2.2 Methods
2.2.1 The model
The individual plant growth model used here is consistent with MST (see
Appendix A for details, Lin et al. 2012), which was based on an energy
conservation equation (Enquist and Niklas 2001, West et al. 2001, Hou et al.
2008). It takes into consideration three basic processes that require energy:
maintenance of biomass, ion transport and biosynthesis (Lambers et al. 2008).

- 30 - Chapter 2
Using empirical measurements and theoretical assumptions, MST predicts
quantitative relationships among these processes (Enquist 2002, Enquist et al.
2009), and we use these as the basis of our individual growth model for plants:
dm/dt = am3/4 – bm = am3/4 [1 – ( m / M0 )1/4] (2-1)
where m is the plant's total biomass and a and b species-specific constants
(Appendix A). Our derivation of this model is similar to the derivation of growth
models for animals (West et al. 2001, Hou et al. 2008). The value of M0 = (a/b)4
is the asymptotic maximum body mass of plant (calculated for dm/dt = 0),
which depends on species-specific traits and is determined by the systematic
variation of the in vivo metabolic rate within different taxa (West et al. 2001).
The gain term (am3/4) in equation (2-1) dominates early in plant growth, and
has some empirical support (Brown et al. 2004, Enquist et al. 2009). Equation
(2-1) is similar to the “von Bertalanffy growth model”, but its derivation here is
based on physical and biological principles of MST (West et al. 2001, Hou et al.
2008).
In our spatially explicit, individual-based model [17], plants are modelled as
circles growing in 2-dimensional space (Weiner et al. 2001). The area of the
circle, A, represents the resources available to the plant, and this area is
allometrically related to the plant’s body mass, m, as m3/4 = c0A (Enquist and
Niklas 2001), where c0 is a normalization constant. Plants compete for
resources in areas in which they overlap, and the mode of competition is
reflected in the rules for dividing up the overlapping areas. Resource
competition is incorporated by using a dimensionless competition index, fp,
which value is determined by the overlap with neighbors (can be reduced from
1 to 0). With these assumptions, equation (2-1) becomes:
dm/dt = fp fr am3/4 – bm = fp fr cA[1 – ( m / M )1/4] (2-2)
where M = (fpfr)4M0 represents maximum achievable biomass under resource
limitation and competition, and where c = ac0 is the initial growth rates in units
of mass per area and time interval. We represent resource limitation with a
dimensionless efficiency factor, fr, as different levels of resource availability.
For simplicity, we use a linear form here, i.e. fr = 1 – RL, where RL indicates
the level of resource limitation, and ranges from 0 (no resource limitation) to 1

Chapter 2 - 31 -
(maximum resource limitation; Table A1). The mode of resource-mediated
competition among plants can be defined anywhere along a continuum from
completely size-asymmetric competition (all the contested resources are
obtained by largest plants) to completely symmetric competition (resources in
areas of overlap are divided equally among all overlapping individuals,
independent of their relative sizes) (Schwinning and Weiner 1998). To
represent the different modes of competition explicitly, we define the effect of
competition, fp, as
,1
1
( ) /io
j
pn
p no o k nk p
jj
mf A A A
m
(2-3)
This index refers to the fraction of resources available in the ZOI which the
plant i could obtain after a loss of potential resources due to areas overlapped
by nj individuals of sizes mj (Schwinning and Weiner 1998). Ano is the area not
overlapping with any neighbors, and Ao,k indicates the area overlapped by
neighbors. Parameter p determines the mode of competition, ranging from
complete symmetry (p = 0) to complete asymmetry (p = ∞).
In MST, individuals’ mortality rate is assumed to be proportional to their
mass-specific metabolism (Brown et al. 2004). Based on this, we assume that
individuals die if their actual growth rate (realistic metabolic rate) falls below a
threshold fraction of their basal metabolic rate (scaled by current biomass, i.e.
2% of m3/4). Therefore, individual plants may die due to metabolic inactivation
driven by resource limitation, competition, senescence (when m approaches M)
or combinations thereof. The model follows ODD protocol (Overview, Design
concepts, Details) for describing individual- and agent-based models (Grimm
et al. 2006, 2010) (Appendix A) and implemented in NetLogo 3.1.4 (Wilensky
1999).
2.2.2 Simulations and Analysis
In our simulations, we investigated 4 resource limitation levels (RL equal to 0,
0.1, 0.5 and 0.9), 4 modes of competition (p=∞: completely asymmetric; p=10:
highly size-asymmetric; p=1: perfectly size-symmetric; p=0: completely
symmetric) and one initial density (8100 individuals per total area). We also
investigated other initial densities and the results were very similar to those

- 32 - Chapter 2
presented below. Simulations for the resulting 16 scenarios were repeated five
times using different random initializations.
We used the relative interaction index RII (Armas et al. 2004) to evaluate
the effects of local competition on shaping plant mass-density relationship:
RII = (mx-mnc) / (mx+mnc) (2-4)
where mx and mnc are the performance (mean biomass) of surviving plants at
the same resource level with and without local competition (i.e., isolated
plants), respectively. Values of RII from -1 to 1 indicate the intensity of
interactions as competition (from -1 to 0), neutral interaction (equal to 0) and
facilitation (from 0 to 1). To estimate mnc, we use the growth equation (2-2)
without the competitive factor fp.
For linear fits of the self-thinning trajectories obtained with our model, we
selected data points on the basis of mortality (Westoby 1984): After
density-dependent mortality starts, data points with surviving plants no less
than 10% of the initial density (not less than 800 surviving plants here) and
with the relative mortality larger than a threshold (the mean value of relative
mortality at each time step through self-thinning process) were selected to fit
the self-thinning trajectories. The thinning trajectories (log-log transformed
data of mean biomass vs. density of survivors) were fitted by reduced major
axis (type II model) regression, which assumes error in both variables and is
widely used to investigate mass-density relationships. All statistical analyses
were conducted using R 2.11.1.
2.3 Results
Variation in the mode of competition, the level of resource limitation and their
interaction produced significant variation in the self-thinning trajectory (Fig. 2.1,
Table S1). The mode of competition had a greater effect on the slope of
self-thinning trajectories than did the level of resource limitation. For given
resource limitation, RL, symmetric competition made self-thinning trajectories
significantly shallower (ninety-five percent confidence intervals for the four

Chapter 2 - 33 -
modes of competition did not overlap), but within same mode of competition
the level of resource limitation did not change slopes much (Fig. 2.2).
In scenarios with more symmetric competition, the relative interaction index
RII is close to -1 and thus the effect of competition on surviving individuals is
quite strong (Fig. 2.3). In contrast, in scenarios with more asymmetric
competition, surviving plants are less affected by interactions with other plants
(RII close to 0). The growth curves of plants also showed same results (Fig.
2.4). Both resource limitation and asymmetric competition lowered the position
of self-thinning trajectories: less biomass can be accumulated at a given
density under resource limitation or with more asymmetric competition (Fig.
2.1). Resource limitation decreased intercepts within the same mode of
competition (Fig. 2.2), which means that the maximum biomass of plants is
smaller in harsh conditions.
Fig. 2.1 Self-thinning trajectories for different levels of resource limitation and modes
of competition. RL indicates the level of resource limitation (from 0 to 1 indicating no
limitation to extreme limitation), p indicates the modes of competition (∞: completely
asymmetric; 10: highly size-asymmetric; 1: perfectly size-symmetric; 0: completely
symmetric). For comparison, the solid lines in figures have same intercepts and with
slopes equal to-4/3.

- 34 - Chapter 2
Fig. 2.2 Slopes and intercepts of self-thinning trajectories of mean individual biomass
versus survivor density under different levels of resource limitation and modes of
competition. RL indicates the level of resource limitation (from 0 to 1 indicating no
limitation to extreme limitation), p indicates the modes of competition (∞: completely
asymmetric; 10: highly size-asymmetric; 1: perfectly size-symmetric; 0: completely
symmetric). Bars indicate 95% confidence intervals.
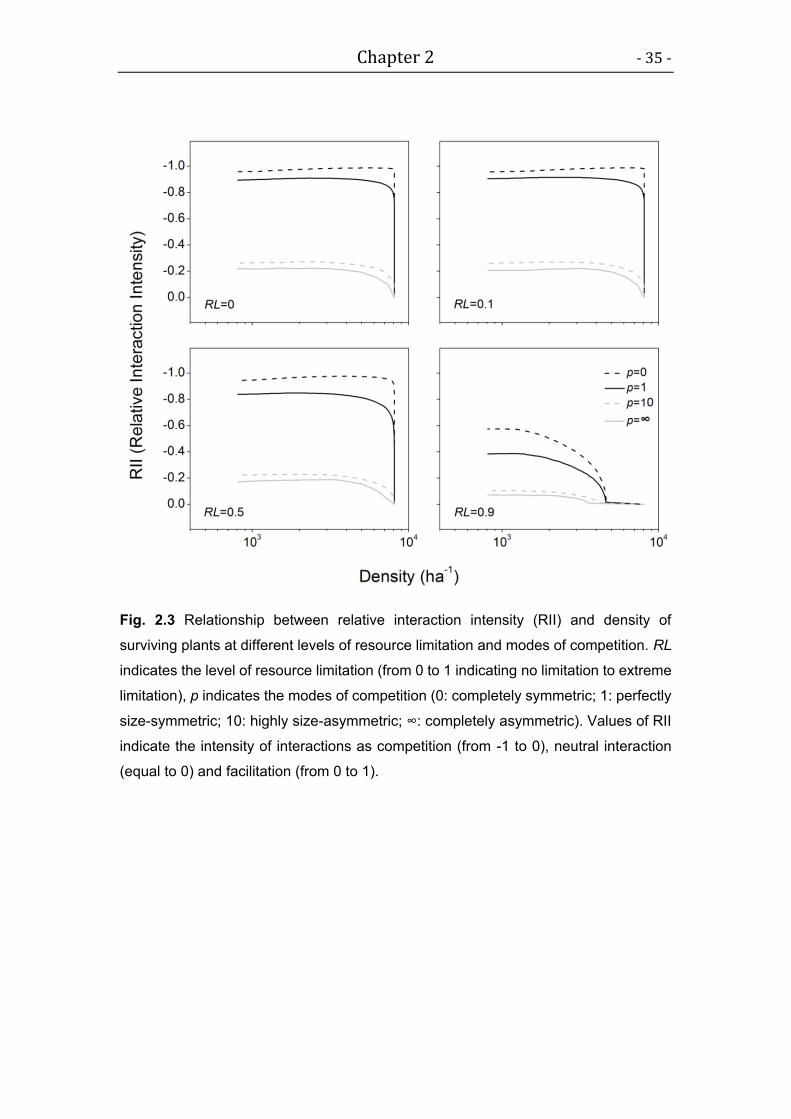
Chapter 2 - 35 -
Fig. 2.3 Relationship between relative interaction intensity (RII) and density of
surviving plants at different levels of resource limitation and modes of competition. RL
indicates the level of resource limitation (from 0 to 1 indicating no limitation to extreme
limitation), p indicates the modes of competition (0: completely symmetric; 1: perfectly
size-symmetric; 10: highly size-asymmetric; ∞: completely asymmetric). Values of RII
indicate the intensity of interactions as competition (from -1 to 0), neutral interaction
(equal to 0) and facilitation (from 0 to 1).

- 36 - Chapter 2
Fig. 2.4 Growth curves of surviving plants at different levels of resource limitation and
modes of competition. RL indicates the level of resource limitation (from 0 to 1
indicating no limitation to extreme limitation), p indicates the modes of competition (0:
completely symmetric; 1: perfectly size-symmetric; 10: highly size-asymmetric; ∞:
completely asymmetric; NC: isolated growth without competition). Values of RII
indicate the intensity of interactions as competition (from -1 to 0), neutral interaction
(equal to 0) and facilitation (from 0 to 1).

Chapter 2 - 37 -
2.4 Discussion
Our results and those of previous studies (Stoll et al. 2002, Chu et al. 2010)
are consistent with our first hypothesis that symmetric competition can lead to
shallower self-thinning trajectories than asymmetric competition. This
suggests that deviations from the slope predicted by MST are likely to occur
when competition below ground is stronger than above ground because the
former is size-symmetric (Weiner et al. 1997, Schwinning and Weiner 1998).
Indeed, several empirical studies show that slopes of self-thinning trajectories
are significantly flatter under severe water stress (Deng et al. 2006) and low
nutrient levels (Morris 2003), conditions in which competition below ground is
thought to be more important than above ground (Schwinning and Weiner
1998, Deng et al. 2006, Berger et al. 2008). Furthermore, boreal coniferous
forests tend to have steeper slopes than deciduous broadleaved forests
because the canopy of conifers is denser (with lower light transmittance)
suggesting that asymmetric competition for light is more intense (Westoby
1984, Stoll et al. 2002, Li et al. 2006).
Why does symmetric competition lead to flatter trajectories in our model
even though it makes surviving plants larger on average at a given density (Fig.
2.1)? With symmetric competition, the growth of all individuals is significantly
reduced but the onset of mortality is delayed (Weiner et al. 2001 p. 20). Plants
can survive and grow even at relatively high densities, meaning that more
biomass can be maintained at a given density when competition is
size-symmetric (Stoll et al. 2002).
When competition is highly size asymmetric, surviving plants are less
affected by their neighbors and thus individual-level metabolic processes of
MST predict plant mass-density relationship at the population level: the slope
of the self-thinning trajectories are close to -4/3. This is consistent with our
second hypothesis that MST's predictions of population-level mass-density
relationships are successful when surviving plants are barely affected by local
interactions. In stands of Nothofagus solandri (mountain beech), taller trees
are relatively unhindered by competition for light and show the scaling of
diameter growth which is consist with prediction of MST, whereas small trees
affected by asymmetric competition do not follow the growth trajectory of

- 38 - Chapter 2
prediction (Coomes and Allen 2007, Coomes et al. 2011). In a tropical
rainforest, the trees under high-light conditions, i.e. being barely affected by
neighboring trees, their diameter growth was not significantly different from the
predictions of MST (Rüger and Condit 2012), which we also find in our model
(Fig. S1) and support our second hypothesis.
In contrast to our findings, Coomes and coworkers concluded that
deviations from predictions of MST in forests are caused by size-asymmetric
competition (Coomes and Allen 2007, Coomes et al. 2011). On closer
inspection, our results are not inconsistent with those of Coomes and
coworkers’. Coomes's studies focused on diameter growth of individuals,
including small suppressed individuals that are experiencing size-asymmetric
competition for light, whereas we focus here on the mass-density relationship
of populations during self-thinning, i.e. on plants surviving competition. When
competition is highly size-asymmetric, total biomass is primarily due to the
largest individuals, which are not highly affected by neighbors. If one looks,
however, at smaller individuals suffering from asymmetric competition before
they die, they will be highly affected by their larger neighbors, and this will be
more important for their growth and density than the internal relationships that
form the basis for MST. It would be worthwhile to analyze individual-level
diameter growth in our model to compare the patterns with those from
empirical studies (Coomes and Allen 2007, Coomes et al. 2011, Rüger and
Condit 2012) to see if the conclusions are consistent. Nevertheless, we agree
with Coomes and coworkers on the central point: interactions among
individuals can overrule the predictions of MST.
Both size-asymmetric competition and resource limitation lowered
self-thinning trajectories (Fig. 2.2). Resource limitation reduces the growth of
individual plants, leading to smaller individuals. Size-asymmetric competition
results in faster mortality. The reduction of biomass due to mortality is not
immediately compensated by the growth of survivors, so there is less total
biomass at a given density.
Using a similar model, Chu et al. (2010) found the same effect of the mode
of competition (size symmetric vs. asymmetric) on the slope of self-thinning
trajectories. The most important difference between Chu et al.'s model and
ours is that we used an individual growth model that is derived from MST,

Chapter 2 - 39 -
whereas they used the phenomenological growth equation of Weiner et al.
(2001). Chu et al. (2010) focused on the effects of mode of competition,
resource levels, and facilitation on self-thinning per se, so they do not refer to
MST or focus on their choice of their individual growth model. This may be why
the range of scaling exponent predicted with our model (-1.083 to -1.486) is
closer to the observed range of exponents (-1.103 to -1.441 for 1266 plots of
six biomes and 17 forest types across China) (Li et al. 2006) than models
using phenomenological growth functions (-0.8204 to -1.6095) (Chu et al.
2010). This suggests that the consideration of both neighborhood interactions
and constraints provided by MST are necessary to explain the
biomass-density relationships observed in the field.
Our results point to the scope and limits of MST at the population and
community level. MST applies to individual organisms, not always and
necessarily to populations or ecosystems. In some cases, for example where
resources are not limiting and competition is highly size-asymmetric, the
mass-density scaling exponent predicted by MST matches observations very
well. This is because individual acquisition of resources and accumulation of
biomass is driven primarily by what the individual itself does rather than by
interactions with other individuals (Fig. 2.3). On the other hand, when
individual behaviour is determined more by interactions with their neighbors
rather than processes that are the bases of MST, the population-level
behaviour will deviate from the predictions of MST.
Acknowledgements
We thank Sven Wagner, Angelika Körner, Christine Lemke, Cyril Piou, Antje
Karge, and Heinz Wolf for their kind help. James Brown and Jianming Deng
gave constructive comments on an earlier version of the manuscript. We also
thank two anonymous reviewers for their constructive comments on the
manuscript. The first author is supported by Helmholtz Impulse and
Networking Fund through Helmholtz Interdisciplinary Graduate School for
Environmental Research (HIGRADE).

- 40 - Chapter 2
References
Allen, A. P., W. T. Pockman, C. Restrepo, and B. T. Milne. 2008. Allometry, growth and
population regulation of the desert shrub Larrea tridentata. Functional Ecology
22:197–204.
Armas, C., R. Ordiales, and F. I. Pugnaire. 2004. Measuring plant interactions: a new
comparative index. Ecology 85:2682–2686.
Berger, U., C. Piou, K. Schiffers, and V. Grimm. 2008. Competition among plants:
concepts, individual-based modelling approaches, and a proposal for a future
research strategy. Perspectives in Plant Ecology, Evolution and Systematics
9:121–135.
Brown, J. H., J. F. Gillooly, A. P. Allen, V. M. Savage, and G. B. West. 2004. Toward a
metabolic theory of ecology. Ecology 85:1771–1789.
Chu, C. J., J. Weiner, F. T. Maestre, Y. S. Wang, C. Morris, S. Xiao, J. L. Yuan, G. Z. Du,
and G. Wang. 2010. Effects of positive interactions, size symmetry of competition
and abiotic stress on self-thinning in simulated plant populations. Annals of
Botany.
Coomes, D. A., and R. B. Allen. 2007. Effects of size, competition and altitude on tree
growth. Journal of Ecology 95:1084–1097.
Coomes, D. A., E. R. Lines, and R. B. Allen. 2011. Moving on from Metabolic Scaling
Theory: hierarchical models of tree growth and asymmetric competition for light.
Journal of Ecology 99:748–756.
Dai, X., X. Jia, W. Zhang, Y. Bai, J. Zhang, Y. Wang, and G. Wang. 2009. Plant
height–crown radius and canopy coverage–density relationships determine
above-ground biomass–density relationship in stressful environments. Biology
Letters 5:571–573. doi: 10.1098/rsbl.2009.0228.
Deng, J. M., G. E. N. . Wang, E. C. Morris, X. P. Wei, D. X. I. . Li, B. A. O. . Chen, C. M.
Zhao, J. Liu, and Y. Wang. 2006. Plant mass–density relationship along a
moisture gradient in north‐west China. Journal of Ecology 94:953–958.
Enquist, B. J. 2002. Universal scaling in tree and vascular plant allometry: toward a
general quantitative theory linking plant form and function from cells to
ecosystems. Tree Physiology 22:1045.
Enquist, B. J., J. H. Brown, and G. B. West. 1998. Allometric scaling of plant energetics
and population density. Nature 395:163–165.
Enquist, B. J., and K. J. Niklas. 2001. Invariant scaling relations across tree-dominated
communities. Nature 410:655–660.
Enquist, B. J., G. B. West, and J. H. Brown. 2009. Extensions and evaluations of a
general quantitative theory of forest structure and dynamics. Proceedings of the
National Academy of Sciences 106:7046.
Grimm, V., U. Berger, F. Bastiansen, S. Eliassen, V. Ginot, J. Giske, J. Goss-Custard, T.
Grand, S. K. Heinz, and G. Huse. 2006. A standard protocol for describing
individual-based and agent-based models. Ecological Modelling 198:115–126.
Grimm, V., U. Berger, D. L. DeAngelis, J. G. Polhill, J. Giske, and S. F. Railsback. 2010.
The ODD protocol: a review and first update. Ecological Modelling.
Hou, C., W. Zuo, M. E. Moses, W. H. Woodruff, J. H. Brown, and G. B. West. 2008.
Energy uptake and allocation during ontogeny. science 322:736.
Lambers, H., Chapin, F.S. & Pons, T.L. (2008). Plant physiological ecology. Springer
Verlag. pp. 134-143

Chapter 2 - 41 -
Li, H. T., X. G. Han, and J. G. Wu. 2006. Variant scaling relationship for mass-density
across tree-dominated communities. Journal of Integrative Plant Biology
48:268–277.
Lin, Y., U. Berger, V. Grimm, and Q.-R. Ji. 2012. Differences between symmetric and
asymmetric facilitation matter: exploring the interplay between modes of positive
and negative plant interactions. Journal of Ecology 100:1482–1491.
Morris, E. C. 2003. How does fertility of the substrate affect intraspecific competition?
Evidence and synthesis from self-thinning. Ecological Research 18:287–305.
Rüger, N., and R. Condit. 2012. Testing metabolic theory with models of tree growth that
include light competition. Functional Ecology 26:759–765.
Schwinning, S., and J. Weiner. 1998. Mechanisms determining the degree of size
asymmetry in competition among plants. Oecologia 113:447–455.
Stoll, P., J. Weiner, H. Muller-Landau, E. Müller, and T. Hara. 2002. Size symmetry of
competition alters biomass–density relationships. Proceedings of the Royal
Society of London. Series B: Biological Sciences 269:2191.
Weiner, J., P. Stoll, H. Muller-Landau, and A. Jasentuliyana. 2001. The effects of density,
spatial pattern, and competitive symmetry on size variation in simulated plant
populations. The American Naturalist 158:438–450.
Weiner, J., and S. C. Thomas. 1992. Competition and allometry in three species of annual
plants. Ecology 21:648–656.
Weiner, J., D. B. Wright, and S. Castro. 1997. Symmetry of below-ground competition
between Kochia scoparia individuals. Oikos 79:85–91.
West, G. B., J. H. Brown, and B. J. Enquist. 1999. The fourth dimension of life: fractal
geometry and allometric scaling of organisms. Science 284:1677.
West, G. B., J. H. Brown, and B. J. Enquist. 2001. A general model for ontogenetic growth.
Nature 413:628–631.
Westoby, M. 1984. The self-thinning rule. Advances in ecological research 14:167–226.
Wilensky, U. 1999. Netlogo. http://ccl.northwestern. edu/netlogo/. Center for Connected
Learning and Computer-Based Modeling, Northwestern University, Evanston, IL.
Zhang, W.P., X. Jia, Y.Y. Bai, and G.X. Wang. 2011. The difference between above- and
below-ground self-thinning lines in forest communities. Ecological Research
26:819–825.

- 42 - Chapter 2
Chapter 2 – Appendix A
A general ontogenetic growth model for plants
The essential basis for modelling the ontogeny of individual plants starts with
an energy conservation equation (Enquist and Niklas 2001; West et al. 2001;
Hou et al. 2008):
Bp = Br + Bs = Br + Esdm/dt (2-A1)
where Bp is defined as total energy intake rate (i.e. gross photosynthetic rate).
A fraction of this assimilated energy is consumed by respiration, Br, the
remainder is stored as reserves and Bs is used for synthesizing new tissues
(and for reproducing) (Figure A1). Es is the metabolic energy stored in one unit
of biomass and dm/dt is the change in biomass (m) per unit time (t).
Figure A1 Assimilated energy partition of plants during ontogenetic growth.
The rate of energy consumed by respiration, Br, depends on three major
processes that require energy (Figure A1): maintenance of biomass (Bmaint),
ion transport (Btran) and biosynthesis (Bsyn), which can be summarized as
(Lambers et al. 2008):
Br = Bmaint + Btran + Bsyn = ∑Bmmliving + Btran + Ecdm/dt (2-A2)
Whole-plant photosynthetic
rate (Bp)
Energy consumed by
respiration (Br)
Rate of energy for maintenance
(Bmaint)
Rate of energy for photosynthetic
tissue (BL=BmmL)
Rate of energy for conducting tissue
(BC=BmmC)
Rate of energy for ion transport
(Btran)
Rate of energy for biosyntheses (Bsyn=Emdm/dt)
Rate of energy stored
(Bs=Esdm/dt)

Chapter 2 - 43 -
where Bm denotes average mass-specific maintenance metabolic rate, mliving
stands for the biomass of living tissues, and Ec designates the energy required
to synthesize a unit of biomass.
Typical average biological parameters of plant cells (tissue) are taken as a
fundamental unit here, and possible differences between tissues are ignored
(West et al. 2001). Note that the terms Bs=Esdm/dt in Eqn (2-A1) and
Bsyn=Ecdm/dt in Eqn (2-A2) are quite different: S stands for the rate of
cumulative energy content of new biomass, whereas Bsyn refers to the
metabolic energy expended on biosynthesis which is dissipated as heat
instead of obtained as stored biomass (Hou et al. 2008). Combining Eqns
(2-A1) and (2-A2), we get
Bp = Bmaint + Btran + Bsyn + Bs = ∑Bmmliving + Btran + E0dm/dt (2-A3)
where E0 = Ec+Es, is constant for a given taxon and stands for the sum of
energy stored in a unit of biomass plus the energy used to synthesize this
biomass, i.e. the synthesis costs of a unit of biomass.
Equation (2-A3) is quite general, but Bmaint may vary between woody and
non-woody plants, as woody plants contain nonliving tissues (e.g. heartwood
in stem and root) which do not need energy for their maintenance (Enquist et
al. 2009). Also large trees with a large amount of heartwood, contain much
less living tissues (mliving) in comparison to the total biomass (m). We assume
that during ontogeny, woody plants mainly expend energy for maintaining their
photosynthetic tissues (leaves), mL, and conducting tissues (standing
sapwood of stem and root), mC, and suppose
Bmaint = ∑Bmmliving = BmmL + BmmC = BL + BC (2-A4)
for woody plants, where BL and BC specify the metabolic rate for maintaining
photosynthetic and conducting tissues, respectively (Figure A1). Combining
Eqns (2-A3) and (2-A4) we can get the energy conservation equation for
woody plants:
Bp = BmmL + BmmC + Btran + E0 dm/dt (2-A5)
Based on empirical measurements and theoretical assumptions linking
biomass and metabolism, MST (West et al. 1999; Enquist 2002; Price et al.
2007; Enquist et al. 2009; West et al. 2009) predicts that whole-plant, or gross,
photosynthesis rate, Bp, and ion transport metabolic rate Btran allometrically
scale with the total biomass of a plant, m, as BpBtranmθ, where θ ≡ 1/(2α+β)
and α and β representing the geometry and biomechanics of the vascular
network. Their values may vary across different taxa (Price et al. 2007).
Although the Eqn (2-A5) can be easily recast by using empirical values of
α and β, we use α = 1/2 and β = 1/3 as common and idealized cases here
(Price et al. 2007; West et al. 2009), so that θ = 3/4. Models based on these

- 44 - Chapter 2
scaling relationships predict that the standing leaf biomass, mL, scales with
respect to total biomass as mL ∝ m~3/4 across woody plants which was
confirmed by empirical data (Sack et al. 2002; Niklas 2005).
The relationship between standing sapwood biomass (mC) and total
biomass m is largely unknown, therefore here we assume that the tissue- or
species-specific wood density of conducting tissues, dC, is constant for a given
plant. Its total volume of conducting tissues, vC, can be formulated as vCASh,
where AS is the mean cross-sectional area of sapwood and h is the height of
plant. Because ASm3/4 and hm1/4 (Enquist 2002; Savage et al. 2010), we
therefore derive the allometric relationship mC=dCvCAShm3/4m1/4m for
woody plants. Substituting the allometric relationship on biomass for all related
terms in Eqn (2-A5) gives
B0m3/4 = BmaLm
3/4 + BmaSm + atranm3/4 + E0 dm/dt (2-A6)
where Bp=B0m3/4 reflects the total energy intake rate (i.e. gross photosynthetic
rate) under optimal situation, B0 is constant for a given taxon (West et al. 1999),
aL, aS and atran are normalization constants. Eqn (2-A6) can therefore be
rewritten as
dm/dt = a1m3/4 – b1m
= a1m3/4 [ 1 – ( m / M1)
1/4 ] (2-A7)
with a1 = (B0 – BmaL– atran)/E0 and b1 = BmaS/E0. The value M1 = (a1/b1)4 is
asymptotic maximum body size of the woody plant (calculated for dm/dt = 0),
which depends on species-specific traits and is determined by the systematic
variation of the in vivo metabolic rate within different taxa (West et al. 2001).
The gain term (a1m3/4) in Eqn (2-A7) dominates while plants grow to a
moderate size, which has been shown to be a good quantitative description of
plant growth (Niklas and Enquist 2001; Enquist et al. 2009).
Across non-woody plants which lack secondary tissues (or juveniles of
woody plant which have not accumulated much secondary tissue), the total
biomass of living tissues (as leaves, mL, stem, mS, and roots, mR is
approximately equal to the whole plant mass, mliving=mL+mS+mR≈m (Enquist et
al. 2007). Combining Eqn (2-A3) with those scaling relationship leads to
B0m3/4 = Bmm + atranm + E0 dm/dt (2-A8)
for non-woody plants.
Taking the parameters in Eqn (2-A8) in the same sense as before, Eqn
(2-A8) can be re-expressed as
dm/dt = a2m3/4 – b2m = a2m
3/4 [ 1 – ( m / M2)1/4 ] (2-A9)
with a2 = B0/E0, b2 = (Bm+atran)/E, and M2 = (a2/b2)4, which is the asymptotic
maximum body size of a non-woody plant.

Chapter 2 - 45 -
Eqns (2-A7) and (2-A9) have same form, and we use a general form of the
growth function for both woody and non-woody plants:
dm/dt = am3/4 – bm = am3/4 [ 1 – ( m / M0)1/4 ] (2-A10)
where a is a general constant and M0 is the generally asymptotic maximum
body size of plant.
Equation (2-A10) has similar form as the “von Bertalanffy growth function”
(von Bertalanffy 1941, 1957) and other phenomenological logistic functions
used for describing plant growth (Hunt 1982; Weiner et al. 2001; Stoll et al.
2002; Stoll and Bergius 2005; Chu et al. 2008; Chu et al. 2010). However, the
ontogenetic functions derived here are based on more fundamental principles,
in which all parameters determining plant growth are directly linked to physical
and biological processes.
Plant growth under abiotic stress
Because ‘stress’ is not a precise concept, the characteristics of abiotic stress
factors are different and can be resource-independent or dependent (Maestre
et al. 2009). We assume simply that abiotic stress factors act in two ways:
restricting the energy intake rate or burdening the maintenance of plant or
even concurrently, this is presumably always true for plants (Lambers et al.
2008). Since the plant growth rate is negatively and linearly related to the
degree of abiotic stress (Chu et al. 2008, 2010), incorporating abiotic stress in
Eqn (2-A10), we have
dm/dt = am3/4 – bm – Sam3/4 = (1 – S)am3/4 [ 1 – ( m / Ms)1/4 ] (2-A11)
where S is a dimensionless efficiency factor that indicates the level of stress
ranges from 0 (no stress) to 1 (extreme stress), Sam3/4 is the energy restricted
or burdened by abiotic stress which is proportional to total energy intake rate
and increasing with the degree of stress level, and Ms=(1–S)4M0 is the
maximum achievable biomass of plant under stress. This also implies that a
plant’s final size is usually smaller than its asymptotic maximum size (optimal
body size, M0) under the environmental stress and/or resource competition,
but can increase by the beneficial effects of neighbour plants via the
amelioration of habitat.

- 46 - Chapter 2
Extensions and predictions based on the general growth model of plant
A general stem diameter growth model of plants
Sometimes, to get the whole biomass of plants is difficulty, especially when we
are going to measure trees. It is therefore flexible to use stem diameter (D)
instead of biomass (m). Since D∝m3/8, which is equivalent to f(D)=dD/dm∝
m-5/8, then dD/dt=f(D)(dm/dt) combine with Eqs. (2-A7), (2-A9) and (2-A10), we
have
dD/dt = adD1/3 – bdD
4/3 = adD1/3 [ 1 – (D / Dmax) ] (2-A12)
where ad and bd are constants, Dmax =ad /bd is the asymptotic maximum stem
diameter.
The sigmoid growth curve of plants
A general sigmoid growth model for plants can be obtained from integrating
Equation (2-A10):
1/4
1/4
1/4 1/4
/40
0 0
4
1/4 /40
0
1 1
or 1 [1 ( ) ]
at M
at M
mme
M M
mam e
b M
(2-A13)
where m0 is the initial biomass of the plant (biomass at birth, t=0).
In the expression related to stem diameter,
1/3d max
1/3 1/3
/30
max max
1 1
a t DDDe
D D (2-A14)
where D0 is the initial diameter of the plant (t=0).
Currently, we are not able to combine the data sets that are precise
enough and also cover a broad plant species to test our model. However,
based on a recent publishes work (Deng et al. 2012), we tested our model with
three non-woody plants, the mechanistic model are proved to be reliable
(Figure A2). How well of our model in capturing the fundamental features of
growth in woody and non-woody plants are remains to be seen.

Chapter 2 - 47 -
Figure A2 Growth curves for wheat, flax, and Arabidopsis. The curves are the
predictions of the general growth model (Equation. 2-E2). Growth data are estimated
from Deng et al. (2012).
A general model for respiratory metabolism of plant
Overall, the respiratory metabolism of plant can be generally summarized in
equation (2-A2). For woody plant, because of the growth rate dm/dt ≈ acm3/4,
where ac is a constant, hence the respiration rate of woody plants is:
Br = BmaLm3/4 + BmaSm + atranm
3/4 + Ec dm/dt = arm3/4 + brm (2-A15)
where ar= BmaL+atran+ Ecac and br=BmaS.
Equation (2-A15) indicates a mixed scaling relationship of rate of energy
consumed by respiration for woody and non-woody plants, i.e. the theoretical
scaling exponent between whole respiration rate of plant and its whole
biomass, Br vs. m, can range from 0.75 to 1 which depends on the values of ar
and br, and is related to the productive biomass partitioning among different
compartments (e.g. leaf, branch, stem, root, and seed reproduction). However,
for woody plants with much of secondary tissues, the scaling should approach
0.75, but for non-woody plants or juveniles of woody plant which haven’t
accumulated much of secondary tissues, the scaling exponent between
respiratory metabolism and biomass will approach to 1. These variations were
perceived in empirical researches (Reich et al. 2006; Enquist et al. 2007;Mori
et al. 2010; Cheng et al. 2010).

- 48 - Chapter 2
Scaling relationship between reproductive biomass and mean body mass of
seed plants
Based on MST and equation (2-A10), the plant reproductive biomass should
proportional to its metabolic rate, and then we predict a simple scaling
relationship between reproductive biomass (mrep.) and plant body mass (m) as
mrep. m 3/4 (2-A16)
using a broad data set (Niklas and Enquist 2004) of seed plants, we proved the
validity of this prediction (Figure A3).
Figure A3 The scaling relationship between mean plant mass (m) and mean
reproduction mass (mrep.) across different species of of seed plants. The solid line
shows the fitted Reduced Major Axis (RMA, type II model) regression of equation (95%
C.I. of the exponent 0.70–0.77).

Chapter 2 - 49 -
ODD protocol – model description of one-layer pi model
The following model description follows the ODD protocol (Overview, Design
concepts, Details) for describing individual- and agent-based models (Grimm
et al. 2006; Grimm et al. 2010),
Purpose
The aim of this model is to evaluate the multiple effects of the mode of
competition and resource limitation on regulating plant population dynamics,
specifically on self-thinning trajectories and density-dependent mortality. In
particular, we investigate whether interactions on individual plant level can
alter the slope and intercept of the self-thinning line. The model does not
represent specific species, but generic ones.
Entities, state variables, and scales
The entities in the model are plants and square habitat units, or patches (Table
A1). Plants are characterized by the following state variables: initial growth
rate, initial biomass, maximum biomass (asymptotic biomass), current
biomass and their position, i.e. coordinates of the stem. Each individual plant
has its own circular zone-of-influence (ZOI). The ZOI stands for the physical
space occupied by a plant, and represents the energy and resources
potentially available to this plant, which is allometrically related to its body
mass. Neighboring plants only compete for the resources when their ZOIs are
overlapping.
In order to make the spatial calculations of resource competition easier,
ZOIs are projected onto a grid of patches. To avoid edge effects, we use a
torus world with a size of 200 × 200 patches (Grimm & Railsback 2005). Each
patch represents 0.25 m2 or 0.25 cm2 for woody- and non-woody plants,
respectively. The state of each patch is characterized by its resource
availability. We use a homogeneous environment here as all patches have the
same, and constant, degree of resource limitation. One time step in the model
represents approximately one year for woody plants and one day for
non-woody plants.

- 50 - Chapter 2
Table A1. State variables and initialization in the individual-based model. Actual
values are drawn from the given intervals to introduce a certain degree of
heterogeneity among individuals.
Variable Description Initial Value [unit]
(woody/non-woody)
Plants
c Initial growth rate 1 ± 0.1 [kg m-2 time step-1] /
[mg cm-2 time step-1]
m0 Initial body mass 2 ± 0.2 [kg] / [mg]
M Maximal biomass 2×106 ± 2×105 [kg] / [mg]
m Current biomass [kg] / [mg]
A Zone of influence [m-2] / [cm-2]
Patches
RL Level of resource limitation [0, 1]
Initialization
Mortality Threshold of death 2% of m3/4
Density Number of plants 8100 ha-1 / m-2
Process overview and scheduling
After initialization, all individual plants with a given density are randomly
distributed in the world. The processes of resource competition, growth and
mortality of each plant are fulfilled within each time step. In each step,
individual plants first sense the resource qualities of patches within their ZOIs,
the area (radius) of an individual plant’s ZOI is determined by its current
biomass. When their ZOIs are overlapping, individuals compete within the
overlapping area. Thus, the overlapping area reflecting resources is divided
according to the competition mode. Considering the outcome of the
competition process, all individual plants grow according to the growth function.
Plants with growth rates falling below a threshold die and are removed
immediately. The state variables of the plants are synchronously updated
within the subroutines, i.e. changes to state variables are updated only after all
individuals have been processed (Grimm and Railsback 2005).
Design concepts
Basic principles: From “Metabolic Scaling Theory”, we derived a general
ontogenetic growth model for individual plants. We combine this model, via the
ZOI approach, with the effects of different modes of competition and resource
limitation.
Emergence: All features observed at the population level, e.g. mass-density
relationship or self-thinning trajectories (i.e. size distribution and spatial
distribution, respectively), emerge from the interaction of individual plants with
their neighbors and the resource level of their abiotic environment.

Chapter 2 - 51 -
Interaction: Individual plants interact via competition for resources in the
overlapping area of their ZOIs.
Stochasticity: Initial growth rate, initial biomass, maximum biomass and initial
position of plants are randomly taken from the intervals given in Table 1. This
introduces a certain level of heterogeneity among individual characteristics to
take into account that real plants are never exactly identical.
Observation: Population size, biomass of each plant, and mean biomass of all
living plants are the main observations.
Initialization
Initially, individual plants are randomly distributed according to the chosen
initial density. Resources are spatially and temporally constant. Each plant has
an initial biomass (m0), maximal biomass (M) and initial growth rate (c1 or c2)
drawn from truncated normal distributions with average and intervals given in
Table 1.
Input
After initialization, the model does not include any external inputs, i.e. the
abiotic environment is constant.
Submodels
Plant growth
In our individual-based model the plant’s ZOI stands for the physical space
occupied by a plant and represents the energy and resources potentially
available to this plant. This space is allometrically related to the plant’s body
mass, m, as c0A=m3/4 (Enquist & Niklas 2001), where c0 is a normalization
constant. Accordingly, based on equation (2-A1) we have:
dm/dt = cA[ 1 – ( m / M0 )1/4 ] (2-M1)
where c = ac0 is initial growth rates in units of mass per area and time interval.
For simplicity, we choose 1 ± 0.1 in our model. We also simulate the model
with different c values. As expected, the results from different values were
qualitatively similar (the slopes didn’t change).
Resource competition and limitation
Resource limitation and competition usually cause a reduction of resource
availability for plants. We therefore represent resource limitation via a
dimensionless efficiency factor or index, fR, for different levels of resource
availability. Resource competition is incorporated by using a dimensionless
competition factor or index, fp, leading to
dm/dt = fRfpcA[ 1 – ( m / M )1/4 ] (2-M2)

- 52 - Chapter 2
where M = (fRfp)4M0, the maximum body size with resource limitation and
competition.
The efficiency factor fR, can take different forms depending on the
characteristics and level of the limiting resource. For simplification, we use a
linear form here, i.e. fR = 1–RL, where RL indicates the level of resource
limitation, with its value ranging from 0 (no resource limitation) to 1 (maximum
resource limitation; Table A1).
As for competition, the mode of resource-mediated competition among
plants can be located somewhere along a continuum between completely
asymmetric competition (largest plants obtain all the contested resources) and
completely symmetric competition (resource uptake is equal for all plants,
independent of their relative sizes; Schwinning & Weiner 1998). To represent
different modes of competition explicitly, we describe the competitive factor fp
as
,1
1
( ) /
o
j
pn i
p no o k nk p
jj
mf A A A
m (2-M3)
This factor thus refers to the percentage of realized resource plant i could
uptake from the amount of resources available (the entire resource that plant
could potentially occupy) among nj competitors with sizes mj (Schwinning &
Weiner 1998). Ano is the area not overlapping with neighbors, Ao,k denotes the
area overlapping with neighbors. Parameter p determines the mode of
competition, ranging from complete symmetry (p = 0) to complete asymmetry
(p approaching infinity; for details and examples see Figure A4).
Figure A4. An example of calculating the competitive index (equation 2-M3)
with different modes of resource competition in an individual-based model as a

Chapter 2 - 53 -
way of dividing plants’ ZOI (zone-of-influence). Three plants with sizes m1, m2
and m3 are competing in this example. For plant 1, its ZOI (A) was divided into
four parts: Ano, the area not overlapping with the other two plants; Ao,1, the area
overlapping with plant 2; Ao,2, the area overlapping with plants 2 and 3; Ao,3,
the area overlapping with plant 3.
Then the actual area that plant 1 can take from Ao,1 is
1 1,1 ,12
1 21
p p
o o p pp
jj
m mA A
m mm
For Ao,2,
1 1,2 ,23
1 2 31
p p
o o p p pp
jj
m mA A
m m mm
And for Ao,3,
1 1,3 ,32
1 31
p p
o o p pp
jj
m mA A
m mm
Therefore, the competitive index for plant 1 is:
Where 3/4
1 0/A m c
In total, equation (2-M2) clearly shows how a plant’s growth rate is jointly
determined by resource availability, fR, and competition, fp. This also implies
that a plant’s final size is usually smaller than its asymptotic maximum size (M)
during resource limitation and local competition.
Mortality
An individual’s mortality rate is proportional to its mass-specific metabolism
(Brown et al. 2004). Based on this, we assume that individuals die if their
actual growth rate (dm/dt) falls below a threshold of their current scaled body
mass, i.e. 2% of m3/4. Therefore, individual plants may die due to metabolic
inactivation driven by resource limitation, competition, senescence (when m
approaches M) or combinations thereof. This provides a more realistic
representation of relevant ecological process than in previous models (Stoll et
al. 2002; Chu et al. 2010).

- 54 - Chapter 2
References
von Bertalanffy, L. 1941. Stoffwechseltypen und Wachstumstypen. Untersuchungen über
die Gesetzlichkeit des Wachstums VII, Biologisches Zentralblatt, 61, 511–532.
von Bertalanffy, L. 1957. Quantitative laws in metabolism and growth. Quarterly Review of
Biology 32:217–231.
Brown, J.H., Gillooly, J.F., Allen, A.P., Savage, V.M. & West, G.B. (2004). Toward a
metabolic theory of ecology. Ecology, 85, 1771–1789.
Cheng, D.L., T. Li, Q.L. Zhong, and G.X. Wang. 2010. Scaling relationship between tree respiration rates and biomass. Biology Letters 6:715 –717.
Chu, C. J., F. T. Maestre, S. Xiao, J. Weiner, Y. S. Wang, Z. H. Duan, and G. Wang. 2008. Balance between facilitation and resource competition determines biomass–density relationships in plant populations. Ecology Letters 11:1189–1197.
Chu, C.J., Weiner, J., Maestre, F.T., Wang, Y.S., Morris, C., Xiao, S., et al. (2010). Effects
of positive interactions, size symmetry of competition and abiotic stress on
self-thinning in simulated plant populations. Annals of Botany, 106, 647–652.
Deng, J., J. Ran, Z. Wang, Z. Fan, G. Wang, M. Ji, J. Liu, Y. Wang, J. Liu, and J. H. Brown. 2012. Models and tests of optimal density and maximal yield for crop plants. Proceedings of the National Academy of Sciences 109:15823–15828.
Enquist, B. J. 2002. Universal scaling in tree and vascular plant allometry: toward a general quantitative theory linking plant form and function from cells to ecosystems. Tree Physiology 22:1045.
Enquist, B. J., A. P. Allen, J. H. Brown, J. F. Gillooly, A. J. Kerkhoff, K. J. Niklas, C. A. Price, and G. B. West. 2007. Biological scaling: Does the exception prove the rule? Nature 445:E9–E10.
Enquist, B.J. & Niklas, K.J. (2001). Invariant scaling relations across tree-dominated
communities. Nature, 410, 655–660.
Enquist, B. J., G. B. West, and J. H. Brown. 2009. Extensions and evaluations of a general quantitative theory of forest structure and dynamics. Proceedings of the National Academy of Sciences 106:7046–7052.
Grimm, V., Berger, U., Bastiansen, F., Eliassen, S., Ginot, V., Giske, J., et al. (2006). A
standard protocol for describing individual-based and agent-based models.
Ecological Modelling, 198, 115–126.
Grimm, V., Berger, U., DeAngelis, D.L., Polhill, J.G., Giske, J. & Railsback, S.F. (2010).
The ODD protocol: A review and first update. Ecological Modelling, 211,
2760–2768.
Grimm, V. & Railsback, S.F. (2005). Individual-based modeling and ecology. Princeton
University Press.
Hou, C., W. Zuo, M. E. Moses, W. H. Woodruff, J. H. Brown, and G. B. West. 2008. Energy uptake and allocation during ontogeny. Science 322:736.
Hunt, R. 1982. Plant growth curves: the functional approach to plant growth analysis. Arnold.
Lambers, H., F. S. Chapin, and T. L. Pons. 2008. Plant physiological ecology. Springer Verlag.
Maestre, F. T., R. M. Callaway, F. Valladares, and C. J. Lortie. 2009. Refining the stress-gradient hypothesis for competition and facilitation in plant communities. Journal of Ecology 97:199–205.
Mori, S., K. Yamaji, A. Ishida, S. G. Prokushkin, O. V. Masyagina, A. Hagihara, A. T. M. R. Hoque, R. Suwa, A. Osawa, T. Nishizono, T. Ueda, M. Kinjo, T. Miyagi, T. Kajimoto, T. Koike, Y. Matsuura, T. Toma, O. A. Zyryanova, A. P. Abaimov, Y. Awaya, M. G. Araki, T. Kawasaki, Y. Chiba, and M. Umari. 2010. Mixed-power scaling of whole-plant respiration from seedlings to giant trees. Proceedings of the National Academy of Sciences of the United States of America 107:1447-1451

Chapter 2 - 55 -
Niklas, K. J. 2005. Modelling below-and above-ground biomass for non-woody and woody plants. Annals of Botany 95:315.
Niklas, K. J., and B. J. Enquist. 2001. Invariant scaling relationships for interspecific plant biomass production rates and body size. Proceedings of the National Academy of Sciences of the United States of America 98:2922.
Price, C. A., B. J. Enquist, and V. M. Savage. 2007. A general model for allometric covariation in botanical form and function. Proceedings of the National Academy of Sciences 104:13204.
Reich, P. B., M. G. Tjoelker, J.-L. Machado, and J. Oleksyn. 2006. Universal scaling of respiratory metabolism, size and nitrogen in plants. Nature 439:457–461.
Sack, L., T. Maranon, P. J. Grubb, B. J. Enquist, and K. J. Niklas. 2002. Global allocation rules for patterns of biomass partitioning. Science 296:1923.
Savage, V. M., L. P. Bentley, B. J. Enquist, J. S. Sperry, D. D. Smith, P. B. Reich, and E. I. von Allmen. 2010. Hydraulic trade-offs and space filling enable better predictions of vascular structure and function in plants. Proceedings of the National Academy of Sciences 107:22722.
Schwinning, S. & Weiner, J. (1998). Mechanisms determining the degree of size
asymmetry in competition among plants. Oecologia, 113, 447–455.
Stoll, P. & Bergius, E. (2005). Pattern and process: competition causes regular spacing of
individuals within plant populations. Journal of Ecology, 93, 395–403.
Stoll, P., J. Weiner, H. Muller-Landau, E. Müller, and T. Hara. 2002. Size symmetry of competition alters biomass–density relationships. Proceedings of the Royal Society of London. Series B: Biological Sciences 269:2191.
Weiner, J., P. Stoll, H. Muller-Landau, and A. Jasentuliyana. 2001. The effects of density, spatial pattern, and competitive symmetry on size variation in simulated plant populations. American Naturalist 158:438–450.
West, G. B., J. H. Brown, and B. J. Enquist. 2001. A general model for ontogenetic growth. Nature 413:628–631.
West, G. B., B. J. Enquist, and J. H. Brown. 2009. A general quantitative theory of forest structure and dynamics. Proceedings of the National Academy of Sciences 106:7040–7045.

- 56 - Chapter 2
Slopes and intercepts of self-thinning trajectories produced by the model
Table S1. Slope and intercept (log-log transformed) of self-thinning trajectories for
simulated plants under different levels of resource limitation and modes of
competition*.
RL p
Slope Intercept
R2 Mean 95% C.I. Mean 95% C.I.
0 ∞ -1.478 -1.525 -1.429 6.258 6.096 6.424 0.994
0 10 -1.342 -1.361 -1.322 5.848 5.808 5.898 0.999
0 1 -1.140 -1.156 -1.125 5.714 5.664 5.769 0.997
0 0 -1.083 -1.085 -1.082 6.208 6.204 6.212 0.999
0.1 ∞ -1.478 -1.526 -1.427 6.217 6.074 6.377 0.994
0.1 10 -1.354 -1.374 -1.333 5.848 5.778 5.918 0.998
0.1 1 -1.144 -1.158 -1.130 5.697 5.650 5.744 0.997
0.1 0 -1.094 -1.098 -1.091 6.185 6.180 6.190 0.999
0.5 ∞ -1.480 -1.516 -1.445 6.176 6.060 6.297 0.995
0.5 10 -1.385 -1.402 -1.365 5.805 5.736 5.875 0.997
0.5 1 -1.188 -1.198 -1.179 5.554 5.519 5.585 0.998
0.5 0 -1.124 -1.126 -1.122 5.978 5.972 5.984 0.999
0.9 ∞ -1.486 -1.499 -1.472 5.918 5.873 5.963 0.996
0.9 10 -1.395 -1.401 -1.389 5.637 5.619 5.657 0.999
0.9 1 -1.201 -1.209 -1.194 5.370 5.345 5.395 0.998
0.9 0 -1.130 -1.139 -1.122 5.474 5.446 5.502 0.997
*RL indicates the level of resource limitation (0–1), p indicates the modes of
competition (with 0: completely symmetric; 1: perfectly size-symmetric; 10: highly
size-asymmetric; ∞: completely asymmetric). C.I. is confidence intervals.

Chapter 3
Mechanisms mediating plant mass-density
relationships: the role of biomass allocation,
above- and belowground competition *
Abstract
Metabolic scaling theory (MST) predicts a ‘universal scaling law’ for plant
mass-density relationships, but empirical observations are more variable.
Possible explanations of this deviation include plasticity in biomass allocation
between the above- and belowground compartment and the mode of
competition, which can be asymmetric or symmetric. Although complex
interactions of these factors are reasonable, the majority of modelling and
empirical studies have been focusing on mono-factorial explanations so far.
We present a generic individual-based model, which allows exploring MST
predictions in realistic settings by representing plasticity of biomass allocation
and different modes of competition in the above- and belowground
compartment simultaneously. To evaluate the behaviour of the simulated
plants related to the allocation patterns and to validate model predictions
* A slightly revised version of this chapter has been submitted to Ecology (status: under
review) as Yue Lin1, 2
, Franka Huth3, Uta Berger
1, and Volker Grimm
2, 4: Belowground
competition and biomass allocation explain deviations from the plant mass-density
relationship predicted by metabolic scaling theory
1 Institute of Forest Growth and Computer Science, Dresden University of Technology, P.O.
1117, 01735 Tharandt, Germany; 2 Helmholtz Centre for Environmental Research – UFZ,
Department of Ecological Modelling, 04318 Leipzig, Germany; 3 Institute of Silviculture
and Forest Protection, Dresden University of Technology, 01735 Tharandt, Germany; 4
Institute for Biochemistry and Biology, University of Potsdam, Maulbeerallee 2, 14469
Potsdam, Germany

- 58 - Chapter 3
regarding the mass-density relationship, we conducted greenhouse
experiments with tree seedlings. The simulated model well captured empirical
patterns both at plant individual and population level. Without belowground
resource limitation, aboveground processes dominated and the slopes of
mass-density relationships followed the predictions of MST. In contrast,
resource limitation resulted to an increased allocation of biomass to
belowground parts of the plants. The subsequent dominance of symmetric
belowground competition led to significantly shallower slopes of the
mass-density relationship. We conclude that changes in biomass allocation
patterns control the constraints at different levels of biological organization and
explain the deviations from the mass-density relationship predicted by MST.
Taking into account the plasticity of biomass allocation and its linkage to the
visible aboveground competition and the invisible belowground competition is
critical for fully representing many forests and eco-regions, in particular for
correctly predicting the response of carbon storage and sequestration to
changing environmental conditions.
Keywords: plasticity, biomass allocation, optimization theory, symmetric
competition, asymmetric competition, ontogenetic growth of plant,
individual-based model, allometric exponent, power law
3.1 Introduction
The relationship between plant performance and density is one of the central
topics in plant ecology (Stoll et al. 2002, Deng et al. 2006, Chu et al. 2010,
Weiner and Freckleton 2010). It is widely accepted that resource competition
among plants is the main force in determining plant mass-density relationships
induced by density-dependent mortality. In self-thinning stands of plants, the
general relationship between density (N) and mean biomass (My) of surviving
plants can be described by a power function, My=kNγ, where k and γ are
referred to as the thinning coefficient and exponent, respectively. Based on
Euclidean geometry, it was proposed that the value of γ is approximately equal
to -3/2, which is well-known as the “-3/2 power rule” of self-thinning (Yoda et al.

Chapter 3 - 59 -
1963). More recently, based on a fractal model metabolic scaling theory (MST)
predicted a ‘universal scaling law’ with γ equal to -4/3 (Enquist et al. 1998,
West et al. 1999, Enquist and Niklas 2001, Enquist 2002, Brown et al. 2004,
Savage et al. 2010).
The exponent predicted by MST is generally confirmed in studies
addressing aboveground biomass across different types of ecological
communities in environments without major resource limitation or physical
stress (Deng et al. 2006). However, findings from arid areas or from plantation
experiments with low resource levels often deviate from these predictions and
show significantly less negative exponents, i.e. shallower slopes of the
self-thinning trajectory or log mass-log density relationship, respectively
(Morris 2003, Deng et al. 2006, Liu et al. 2006). Consequently, both the validity
and universality of scaling exponent and MST are still unclear and require
further analyses of the underlying mechanisms (Coomes 2006, Deng et al.
2006, Coomes et al. 2011).
On sites where belowground resources such as nutrients and water are
limited, plants tend to allocate more biomass to their belowground parts and
develop extended root systems for acquiring limited resources while canopies
remain unclosed (Deng et al. 2006). In such situations belowground
competition is believed to affect plant growth and mortality more than
aboveground competition (Deng et al. 2006, Berger et al. 2008). Nevertheless,
only few studies have explored the role of belowground process for plant
mass-density relationships (Morris 2003, Deng et al. 2006, Berger et al. 2008).
This is because observations of the root systems are hampered by logistic
difficulties (Cahill and Casper 2000, Casper et al. 2003, Berger et al. 2008,
Rewald and Leuschner 2009, Schiffers et al. 2011).
Deng et al. (2006) found that the mass-density relationship of natural
vegetation was altered by changing root versus shoot biomass allocation along
a precipitation gradient. The root:shoot ratio (RSR) of biomass increased with
aridity but for the belowground compartment the slope of the plant
mass-density trajectory was still consistent with MST predictions. In contrast,
the slopes of the aboveground compartment became less negative. This study
suggested that plant mass-density relationships are altered by the
well-documented plasticity of biomass allocation in response to resource

- 60 - Chapter 3
limitation (McConnaughay and Coleman 1999, Weiner 2004, Deng et al. 2006,
Berger et al. 2008, May et al. 2009, Schiffers et al. 2011).
Apart from this, the mode of competition (asymmetry vs. symmetry) has
also been discussed to affect plant mass-density relationships (Schwinning
and Weiner 1998, Stoll et al. 2002, Coomes and Allen 2009, Berger et al. 2008,
Chu et al. 2010, Coomes et al. 2011). Aboveground competition for light is
considered to be asymmetric, whereas belowground competition is considered
to be rather symmetric (Schwinning and Weiner 1998). In various studies with
belowground recourse limitation, belowground competition was shown to alter
the plant mass-density relationship, i.e. the exponents became less negative
than predicted by MST (Morris 2003, Deng et al. 2006, Chu et al. 2010, Lin et
al., in revision). Nevertheless, there also seem to be cases where belowground
competition can be asymmetric (Fransen et al. 2001, Rajaniemi 2003, Rewald
and Leuschner 2009).
Although plant ecologist agree that the biomass allocation patterns and the
relative importance of above- and belowground competition change along
environmental gradients (Tilman 1988, Deng et al. 2006, Berger et al. 2008), it
remains unknown which factor or which combination of them dominates the
alteration of the mass-density relationship in plant populations and
communities.
In a previous one-layer individual-based model (IBM), which uses the zone
of influence (ZOI) approach (Wyszomirski 1983, Wyszomirski et al. 1999,
Weiner et al. 2001, May et al. 2009) but does not distinguish the above- and
belowground compartments, we found that symmetric competition can alter
the predictions of MST (Lin et al., in revision). This model is too simple, though,
to explore the role of resource allocation and below- versus aboveground
competition. Therefore, based on recent modelling studies (May et al. 2009,
Lin et al., 2012, Lin et al., in revision), we developed a new two-layer IBM
which represents both above- and belowground competition simultaneously
via independent ZOIs. In the presented model, plant growth and biomass
allocation are represented by the growth function based on MST (Enquist 2002,
Niklas 2005, Lin et al., 2012), which tries to mechanistically capture the plastic
responses of plants to changing environmental conditions.
To validate our new model, a greenhouse experiment was also employed.

Chapter 3 - 61 -
We determined root and shoot biomass allocation and relative mass-density
relationships of birch seedlings grown under severe root competition at
different resource levels. The combination of modelling and experiment
enabled us to identify the relative role of two key factors (above- vs.
belowground biomass allocation and asymmetric vs. symmetric competition)
and effects on plant mass-density relationships.
3.2 Methods
3.2.1. Individual-based model
To explicitly assess the interplay between biomass allocation patterns and the
different modes of subsequent shoot competition and root competition, we
extended an established one-layer zone-of-influence (ZOI) model (Lin et al.
2012, Lin et al. in revision) to a two-layer model, which describes the above-
and belowground part of a single plant by separate ZOIs.
A detailed description of the whole model, which follows the ODD protocol
(Overview, Design concepts, Details) for describing individual-based models
(Grimm et al. 2006, 2010), is provided in the supplementary material
(Appendix A) as well as the model implementation in NetLogo 4.1.3 (Wilensky
1999). Here, we only describe the core elements of the model.
The fundamental function representing the ontogenetic growth of an
individual plant is based on MST and does not differ from the one-layer model
(Lin et al. 2012):
∆m/∆t = am3/4 – bm = am3/4[ 1 – ( m / M0)1/4 ] (3-1)
where m is the current biomass of whole plant, a and b the species-specific
constants, and M0=(a/b)4 refers to the asymptotic maximum body size of a
plant (calculated for ∆m/∆t =0).
The general description of the mode of operation of the zone-of-influence
(ZOI; Weiner et al. 2001) is the same above- and belowground: the particular
circular ZOI with area A stands for the physical space occupied by a plant,
which also represents the potential energy and resources that are available to

- 62 - Chapter 3
this plant and is proportional to the plant’s metabolic rate (B). Each ZOI is
allometrically related to the respective biomass, m, as m3/4=c0A (Enquist and
Niklas 2001), where c0 is a normalization constant. Resource limitation and
competition usually cause a reduction of resource availability for plants. We
therefore represent resource limitation via a dimensionless efficiency index, fR,
for different levels of resource availability. Resource competition is
incorporated by using a dimensionless competition index, fp. Equation (3-1)
becomes:
∆m/∆t = fRfpcA[ 1 – ( m / M)1/4 ] (3-2)
where c=ac0, is the initial growth rate and M=(fRfp)4M0 the maximum body size
with resource limitation and competition. The efficiency index, fR, can take
different forms depending on the characteristics and level of the limiting
resource. For simplification, we use a linear form, fR = 1–RL, where RL
indicates the level of resource limitation, with its value ranging from 0 (no
resource limitation) to 1 (maximum resource limitation). As for competition, the
modes of resource-mediated competition among plants can be located along a
continuum between completely asymmetric competition (largest plants obtain
all the contested resources) and completely symmetric competition (resource
uptake is equal for all plants, independent of their relative sizes; Schwinning
and Weiner 1998). Different modes of competition are incorporated in our
model by using a competition index fp as
,1
1
( ) /
o
j
pn i
p no o knk p
jj
mf A A A
m (3-3)
This index represents the fraction of resources available in the ZOI which plant
i could obtain after the loss of potential resources on the areas overlapped by
neighbours of sizes mj (Schwinning and Weiner 1998). Ano is the area not
overlapping with neighbours, Ao,k are the no areas overlapping with nj different
neighbours. Parameter p adjusts the mode of competition (Schwinning and
Weiner 1998). Here, we consider two theoretically important p values reflecting
two modes of competition: allometric asymmetry (p=10, larger individuals get a
disproportional share of overlapped areas) and size symmetry (p=1, the

Chapter 3 - 63 -
overlapped areas are divided among all overlapping individuals proportional to
their sizes; for detail and an example see Figure A2).
In the one-layer version of our ZOI model (Chapter 2, Lin et al. in revision),
equation (3-2) lumps above- and belowground competition together. In the
study presented, equation (3-2) is applied to both ZOIs representing the
above- and belowground physical space a plant occupies and on which it has
potential access to the resources light, water and nutrients, respectively
(Figure A3). Through this modification, we take into account that the relative
importance of shoot versus root competition and the size of the corresponding
ZOIs can depend on environmental factors (Casper et al. 2003, Deng et al.
2006, O’Brien et al. 2007, May et al. 2009).
We assumed that (i) under optimal conditions without resource limitation
and competition, the abilities of above- and belowground resource uptake are
balanced, with relationships between metabolic rate, B, and biomass, m, being
B=cshootmshoot3/4=crootmroot
3/4 (Niklas 2005, Cheng and Niklas 2007; see
Appendix A), where cshoot and croot are normalization constants (to simplify, we
assume cshoot=croot=1), and “shoot” and “root” refer to the above- and
belowground compartment, respectively; (ii) the plant’s above- and
belowground ZOIs are proportional to the plant metabolic rate, B, and
allometrically related to the plant’s shoot and root biomass (Enquist and Niklas
2001, May et al. 2009), Aa=camshoot3/4 and Ab=cbmroot
3/4, where ca and cb are
normalization constants (to simplify, we use ca=cb=1); (iii) growth of the entire
plant is limited by the compartment with smaller resource uptake rate (May et
al. 2009). Thus, equation (3-2) is first independently applied for both the
above- and belowground compartment, then growth of the entire plant is set to
that of the more limited compartment, multiplied by two to account for the case
where below- and aboveground growth are the same (May et a. 2009):
1/4
, ,
1/4
, ,
2 2 [1 ( / ) ],
2 2 [1 ( / ) ], (3-4)
,
R a p a a a a
R b p b b b b
AGR f f c A m M AGR BGR
mBGR f f c A m M AGR BGR
tAGR BGR AGR BGR
where ∆AGR and ∆BGR are above- and belowground growth rate and the
subscripts a and b indicate the above- and belowground compartment,

- 64 - Chapter 3
respectively. Then, allocation of the gained biomass to the two compartments
is assumed to be plastic (McConnaughay and Coleman 1999, Weiner 2004,
Berger et al. 2008), with more biomass being allocated to the compartment
with the smaller, and thus more limiting, growth rate. Adopting optimization
theory (Johnson 1985, McConnaughay and Coleman 1999, May et al. 2009)
and metabolic scaling theory (Niklas 2005), we assumed partitioning of
biomass growth between shoot and root to be:
3/4
3/4 3/4
3/4
3/4 3/4
(3-5a)
(3-5b)
shoot
root
m m BGR
t t AGR BGR
m m AGR
t t AGR BGR
An allometric form (3/4) of resource allocation was used here as metabolic
balance; we also tested the original exponent of 1 (Johnson 1985, May et al.
2009), which did not change our general findings.
An individual’s mortality rate is proportional to its mass-specific
metabolism, i.e. current total metabolic rate divided by body mass (Brown et al.
2004). Based on this, we assume that individuals die if their actual growth rate
(actual metabolic rate, ∆m/∆t) falls below a threshold fraction of their basal
metabolic rate (scaled by current body mass, i.e. 3% of m3/4). Therefore,
individual plants may die due to metabolic inactivation driven by above- or/and
belowground resource limitation, competition, senescence (when m
approaches M), or combinations thereof. In total, Equations (3-4), (3-5a) and
(3-5b) represent how growth, biomass allocation and mortality of a plant are
jointly determined by above- and belowground resource levels and local
competition. This provides a mechanistic and quantitative basis for linking the
energetic metabolism and growth of plants to local interactions and population
dynamics under different environmental conditions, i.e. to identify the
compartment which mostly influences the plant growth at individual level and
mass-density relationship at population level. The design of our model also
allows us to explicitly ascribe the mortality of individual plants to above- or/and
belowground processes in simulations.

Chapter 3 - 65 -
3.2.2. Scenarios
In our simulation experiments, we tested high initial densities (104 = 10, 000
individuals per total area), two modes of aboveground (shoot) competition (CAA:
allometric asymmetry, pa = 10; CSS: size symmetry, pa = 1), and two modes of
belowground (root) competition (CAA: allometric asymmetry, pb = 10; CSS: size
symmetry, pb = 1) at three resource-limitation levels for the belowground
compartment (RLb equal to 0, 0.4 and 0.8 representing no, medium and strong
limitation of belowground resources respectively). Simulations for the resulting
nine scenarios were repeated five times, using different random initializations.
We also tested low initial densities (103.5 ≈ 3,163 individuals per total area) and
other combinations of competition parameters, which lead to similar
conclusions as we presented here (Table S1).
3.2.3. Greenhouse experiment
To test the predictions of our model, we carried out a greenhouse experiment
at the research station of Hetzdorf, Dresden University of Technology,
Germany. A common tree species of central Europe, Betula pendula Roth
(silver birch), was chosen because it is fast-growing. The germination
percentage of birch in our controlled pre-test under climate chamber conditions
was 45% (ISTA 1993). Seeds of B. pendula were sown in the pots at two initial
densities (calculated as 0.09g and 0.27g per pot), then two initial seedling
densities with approximately 250 and 750 individuals per pot have been
established, which correspond to 10000 and 33000 plants m-2. Each pot was
filled with a natural mixed spruce forest humus substrate. The plastic pot size
was 20×16.3 cm at top and 17×13.5 cm at bottom with 5 cm in depth.
Consequently, root competition is expected to be very intensive in such
shallow pots (Wilson 1988).
On 11 April 2010, seeds were sown into each pot and covered by a thin
layer of sand against drought damages. We set two nutrient levels, as fertilized
and non-fertilized groups. For the fertilized group, sustained-release fertilizer
was placed uniformly over the ground substance surface in each pot (4.6 g per
pot). In total, we used 40 pots, for two densities, two nutrient levels and ten
replicates. All pots were arranged on benches in the greenhouse, and were

- 66 - Chapter 3
randomly rearranged once per week to avoid possible effects of environmental
heterogeneity. On 28 July 2010, we terminated the planting experiment and
harvested all plants. Self-thinning had largely reduced density of seedlings in
all harvested pots. In order to avoid edge effects, we set up a 7×7 cm subplot
in the center of each pot for measurement. Shoot and root dry biomass were
measured for each individual plant after oven-drying to constant weights at
60°C. Two pots in the fertilization group were lost (one pot in each initial
density), thus 38 pots were evaluated.
3.2.4. Statistical analysis
The mass-density relationship (log-log transformed data of mean biomass vs.
live plant density) has the form log My = log k + γ log N, where My (mean
biomass of either total plants, shoots, or roots) and N (plant density) are the
variables plotted on the ordinate and abscissa, respectively. The constant log k
is the intercept and γ the slope of the regression line (the allometric or scaling
exponent). We used the data of simulated population with high initial density
(i.e. 10,000 individuals per total area) for analysis (the thinning trajectories of
low initial densities were identical to high initial density in simulations; Fig. S1).
Data points were selected a posteriori for linear fits in our simulation
experiment (Weller 1987, Chu et al. 2010): for each scenario, after
density-dependent mortality starts, the data points with surviving plants
between 7,000 and 500 during self-thinning were selected, which excluded the
data points that may cause a non-linearity effect on linear regression. Data
points were fitted by standard major axis regression (SMA, type II model),
which concerns the presence of error in both variables and is widely used to
investigate mass-density relationships (Warton et al. 2006). All statistical
analyses were conducted using R 2.11.1 (R Development Core Team, 2010).
3.3 Results
3.3.1 Individual-based model
Plants allocated more biomass to roots for acquiring belowground resources
(RLb) when these resources became limited. As expected, root : shoot ratio

Chapter 3 - 67 -
(RSR) was significantly higher with belowground resource limitation (ANOVA,
F = 11.48, P < 0.001; Fig. 3.1a). Consequently, belowground competition
among plants became more intense leading to increased mortality (Fig. 3.1b).
This suggests that the relative importance of above- and belowground
competition for the population dynamics varies inversely along a gradient of
belowground resource limitation.
Also, the plasticity of biomass allocation contributes to variable
mass-density patterns (Fig. S1; Table S1). When the mode of competition is
entirely asymmetric (CAA AA , for both above and below ground) and without
resource limitation (RLb=0), the slopes of log mass-log density relationships for
mean total, above- and belowground biomass are all consistent with the
prediction of MST (Fig. 3.2, Table S1). With increasing belowground resource
limitation, the slopes of log mass-log density relationships for belowground
biomass still supports MST but the slopes for aboveground part are
significantly shallower (95% CI do not include -4/3; Fig. 3.2, Table S1).
However, at all resource levels, entirely symmetric competition (CSS SS ) results in
slopes of the log mass-log density relationships significantly shallower than
predicted by MST (95% CI do not include -4/3; Fig. 3.2, Table S1), for both
above- and belowground biomass. This result indicates that the mode of
competition contributed more to altering the mass-density relationship than
biomass allocation did.
In the combination CAA SS without resource limitation (RLb=0, Fig. 3.1b),
aboveground competition had a greater effect on population mortality than
belowground competition because there was no resource limitation. The
allometric exponents were very close to those of the combination CAA AA (95% CI
overlapped; Fig. 3.2, Table S1) and also compatible with prediction of MST (95%
CI include -4/3), but significantly differ from CSS SS (95% CI did not overlap; Fig.
3.2, Table S1). When belowground resources were limited (RLb=0.4), plant
allocated more biomass to belowground parts; the aboveground mass-density
relationship of CAA SS was significantly different from the prediction of MST (95%
CI did not include -4/3) and CAA AA (95% CI did not overlap; Fig. 3.2, Table S1).
However, exponents of belowground showed little variation and were still
consistent with MST (also CAA AA) reflecting that mortality was mainly induced by
asymmetric aboveground competition (Fig. 3.1b).

- 68 - Chapter 3
With further increased belowground resource limitation (RLb=0.8), the
mass-density relationships of CAA SS for mean total, above- and belowground
biomass were very similar to the case of CSS SS (Fig. 3.2, Table S1) and thus
significantly different from the predictions of MST and CAA AA (Fig. 3.2, Table S1).
In this scenario, symmetric belowground competition was the main factor
driving mortality (Fig. 3.1b). This result again demonstrated that the
mass-density relationships are more affected by the mode of competition
independent of which compartment is dominant in terms of growth limitation.
Fig. 3.1 The (a) root:shoot ratio (mean ± 1 SD) and (b) percentage of mortality
induced by above- or belowground competition in simulated plant populations at
different levels of belowground resource limitation, RLb, ranging from 0 (no resource
limitation) to 1 (maximum resource limitation). C with superscript and subscript
indicated the mode of competition for above- and belowground part correspondingly
(AA: allometric asymmetry; SS: size symmetry). Bars in the same group that share
the same letter do not differ significantly (P > 0.05, Holm-Sidak test).

Chapter 3 - 69 -
Fig. 3.2 Slopes of mass-density relationships between the mean total, above- and
belowground biomass and densities in simulated plant populations (log-log
transformed; as estimated by standard major axis; all regressions were significant at
P < 0.0001). RLb defined the level of belowground resource limitation, with its value
ranging from 0 (no resource limitation) to 1 (maximum resource limitation). C with
superscript and subscript indicated the mode of competition for above- and
belowground part correspondingly (AA: allometric asymmetry; SS: size symmetry).
The dotted lines indicate the value of exponent (-4/3) predicted by MST.
3.3.2 Greenhouse experiment
Our greenhouse experiment confirmed the results of the simulation
experiments. For non-fertilized plants the mean RSR of biomass was
significantly higher than for fertilized plants (ANOVA, F = 9.09, P < 0.001; Fig.
3.3). The mass-density relationship varied between fertilization and
non-fertilization treatment (Table S2, Fig. 3.4a). As for fertilized plants, the
slopes of total, above- and belowground log mass-log density relationship
were not significantly different from the prediction of MST (95% CI included
-4/3; Table S2). However, the slopes became shallower in the group of
non-fertilized plants (Fig. 3.4b) and differed significantly from the prediction of
MST (95% CI do not include -4/3; Table S2). The overall pattern of differences
in allometric exponents between fertilization and non-fertilization treatment
(Fig. 3.4b) was similar to the pattern of C AA SS changing along the
resource-limited gradient explored in the simulation experiments (Fig. 3.2)

- 70 - Chapter 3
confirming the dominance change from aboveground (asymmetric) to
belowground (symmetric) competition. Therefore, the importance of
belowground competition as driving force in plant population and communities
is predicted to increase along gradients of decreasing belowground resource
availability, whereas the importance of aboveground competition is predicted
to decrease.
Fig. 3.3 The root:shoot ratio (mean ± 1 SE) of Betula pendula seedlings in
greenhouse experiment with different nutrient treatments and initial densities (L: low
density, H: high density, L+H: combined data). Bars that share the same letter do not
differ significantly (P > 0.05, Holm-Sidak test).

Chapter 3 - 71 -
Fig. 3.4 The mass-density relationships of Betula pendula seedlings in greenhouse
experiment with different nutrient treatments. (a) Relationships between mean total,
above- and belowground biomass and densities (log-log transformed). (b) Slopes of
mass-density relationships between the mean total, above- and belowground
biomass and densities (log-log transformed, as estimated by standard major axis; all
regressions were significant at P < 0.001, 95% CI of the slopes indicated that the
exponents of non-fertilization treatment were statistically different from −4/3).
3.4 Discussion
We presented a two-layer individual-based model that is rooted in metabolic
scaling theory (MST). The purpose of the model was to test whether a generic
model that explicitly represents biomass allocation and different modes of

- 72 - Chapter 3
competition for both above- and belowground compartments would be able to
capture observed variations and patterns in plant mass-density relationships.
In addition to simulation experiments, we conducted a greenhouse experiment
to verify the model’s predictions.
The main finding was that changes in the allometry of plants, induced by
their plasticity of biomass allocation in response to environmental factors, can
alter the relative importance of above- or belowground competition on driving
density-dependent mortality, which affects plant mass-density relationships.
Root competition, which is assumed to be more symmetric, can have a strong
effect on altering the allometric exponent, i.e. flattening plant log mass-log
density relationships (also self-thinning trajectories). Deviations from the
mass-density scaling exponent predicted by MST are thus likely to occur if
competition occurs belowground (symmetric) rather than aboveground
(asymmetric). Consequently, models or theories explaining mass-density
relationships and self-thinning which are solely based on aboveground parts
are unlikely to be appropriate for the cases where belowground processes
predominate (Deng et al. 2006, Berger et al. 2008).
3.4.1. The advantages of using a new mechanistic growth model
It was important that we used a mechanistic growth model derived from MST
rather than a phenomenological growth model, as usually has been done so
far for analyzing biomass-density relationships (e.g., Weiner et al. 2001, Stoll
et al. 2002, Chu et al. 2010). First, our generic growth model is based on the
energy budget of the individuals during growth. It thus captures the salient
features of energy acquisition and allocation to maintain and replace the
existing tissue and the production of new tissue, and relates ontogenetic
growth to metabolic energy at the cellular level (West et al. 2001). This
certainly is an advantage by itself because models driven by “first principles”
are more flexible and usually capture more adaptive responses of individuals
to their environment than phenomenological models, which are statistically
fitted to existing data (Grimm and Railsback 2005, Martin et al. 2012). Second,
in contrast to previous models (Wyszomirski et al. 1999, Weiner et al. 2001,
Stoll et al. 2002, Chu et al. 2010), our model is able to simultaneously explain

Chapter 3 - 73 -
the observed variation in both root-shoot allocation patterns and mass-density
relationships.
It is worth noting that an association of the scaling exponents with root :
shoot ratios (RSR) predicted by our model matched the observed values of
natural vegetations along a precipitation gradient in China very well. Deng et al.
(2006) found that with drought severity increasing, the mean RSR of plant dry
biomass increased approximately from 1 to 2 (RSR±SD: 0.92±0.31, 1.92±1.42
and 2.18±1.61) and the scaling exponent of plant total biomass-density
relationship changed from -1.33 to -1.27 correspondingly (-1.33, -1.28 and
-1.27). In our model, we found very similar associations between RSR and
scaling exponent (Table S1; e.g., C AA SS : RLb=0, RSR±SD=1.09±0.03,
Slope=-1.36; RLb=0.4, RSR±SD=1.66±0.16, Slope=-1.29), which are
statistically identical to the empirical results (P > 0.05, Holm-Sidak test).
3.4.2. Above- and belowground competition
Why do the predictions of MST on plant mass-density relationship often turn
out to be correct? The principal assumption of energy equivalence in MST
implies that light is the limiting factor and competition is a canopy-packing
process among plants (Enquist et al. 1998, Reynolds and Ford 2005, Deng et
al. 2006). If then in benign environments only aboveground biomass is
measured, as in most empirical studies, it is to be expected that we obtain the
slope of -4/3 (scaling exponent) as shown in our model.
Our results imply the importance of adaptive behaviours of plants for
system-level properties like biomass-density relationship (Grimm and
Railsback 2005, Grimm et al. 2005). With “adaptive behaviour” we refer to
morphological and physiological plasticity that allows plants to adapt to
changing biotic and abiotic environmental conditions (Weiner 2004, Berger et
al. 2008, Schiffers et al. 2011). It is recognized that features of vegetation can
change in drought or oligotrophic environments (Deng et al. 2006). Based on
optimization theory, plants compete primarily for limited belowground
resources in those environments, which can cause an increase of RSR as well
as root space, but a decrease in cover where plant canopies are not closed
(Deng et al. 2006). In analogy to Liebig’s law of limiting factors, belowground

- 74 - Chapter 3
competition can overtake aboveground competition and becomes dominant in
these situations. Consequently, adaptive behaviour not only alters the way
plants grow but also their mode of competition.
Empirical experiments show that slopes of log mass-log density
relationship are manifestly shallower under severe water stress (Deng et al.
2006, Liu et al. 2006) and low nutrient levels (reviewed in Morris 2003). Our
model and greenhouse experiments confirm these results. As demonstrated,
the mode of competition played a more important role for the plant
mass-density relationship.
3.4.3. Asymmetric vs. symmetric competition
With symmetric competition, resource acquisition is more evenly distributed
among interacting plants, so that competitive suppression and the
corresponding onset of mortality are delayed (Stoll et al. 2002). Consequently,
even at high densities plants can still survive and grow (Hautier et al. 2009,
Lamb et al. 2009). This leads to the shallower self-thinning lines as has been
proven in several studies (Chu et al. 2010, Lin et al. in revision).
Accordingly, one should also expect that slopes become shallower if
aboveground competition is less asymmetric. This was indeed observed for
low light conditions (Lonsdale and Watkinson 1982, Westoby 1984, Dunn and
Sharitz 1990), where plants often show flexible shoot morphology and grow
taller instead of wider, and competition was considered more symmetric
(Schwinning and Weiner 1998, Stoll et al. 2002).
However, some researchers proposed that size-asymmetric (light)
competition is more important for explaining the deviations predicted by MST
in forests (Coomes and Allen 2009; Coomes et al. 2011). At first glance, this
seems to be in contrast to our findings here, but this is not the case because
those studies focused on individual’s diameter growth. Asymmetric competition
from larger neighbours can highly affect the growth of smaller individuals
before they die. Thus, for these individuals, asymmetric competition is more
important than the relationships predicted by MST. In contrast, we here
focused on the biomass and density of surviving plants.
It is noteworthy to mention that it also has been surmised that

Chapter 3 - 75 -
belowground competition might be rather asymmetric in heterogeneous
resource conditions (Schwinning and Weiner 1998, Fransen et al. 2001,
Rajaniemi 2003, Rewald and Leuschner 2009), though the experimental
evidence for asymmetric belowground competition is scarce (Rajaniemi 2003,
von Wettberg and Weiner 2003, Rewald and Leuschner 2009). Our model and
greenhouse experiment, nevertheless, confirm the majority of empirical
experiments which suggest that belowground competition is more symmetric.
We conclude that an appropriate evaluation of the mode of competition should
not only consider environmental heterogeneity and species-specific ecological
traits of plants (Schwinning and Weiner 1998, Rajaniemi 2003, Pretzsch 2006,
Rewald and Leuschner 2009), but also the interaction between above- and
belowground competition (Cahill 2002).
Conclusion
It seems that Liebig’s law even applies, in a generalized sense, to the debate
about whether or not MST is a universally valid and relevant theory. Deviation
in observed plant mass-density relationships indicates that MST is less general
than originally thought. Our findings confirm that MST by itself applies to
individual organisms, but not necessarily to populations or ecosystems (Lin et
al. in revision). System-level features of ecological systems may be
constrained by individual metabolism, which is taken into account in MST, or
by ecological factors beyond the individual, i.e. the type of limiting resource
and the mode of competition among con-specifics. The overall claim that MST
provides a universal and mechanistic basis for quantitatively linking the
energetic metabolism of individuals to ecological community dynamics is thus
neither completely right nor wrong: the real question is when and which factors
dominate in a given situation.
In addition to previous studies, which do not address the linkage between
aboveground and belowground competition explicitly, the most important
conclusion of our study is that changes in biomass allocation patterns can
modify ecological mechanisms and subsequently the constraints which MST
sets. The observed variation in mass-density relationships represents changes
in the dominance of the most limiting factor in a given ecological context. This

- 76 - Chapter 3
has important implications, e.g., for using allometry-based simulation models
to predict carbon storage and sequestration in plant systems. Taking into
account not only the visible aboveground competition, but also the invisible
belowground competition, is thus critical for many forests and eco-regions.
Acknowledgements
We thank Angelika Körner, Christine Lemke, Antje Karge, Heinz Wolf and
Sven Wagner for their kind help. We are particularly grateful to Jacob Weiner
who showed us once again the beauty and seriousness science. The first
author was supported by Helmholtz Impulse and Networking Fund through
Helmholtz Interdisciplinary Graduate School for Environmental Research
(HIGRADE).
References
Berger, U., C. Piou, K. Schiffers, and V. Grimm. 2008. Competition among plants: concepts, individual-based modelling approaches, and a proposal for a future research strategy. Perspectives in Plant Ecology, Evolution and Systematics 9:121–135.
Brown, J. H., J. F. Gillooly, A. P. Allen, V. M. Savage, and G. B. West. 2004. Toward a metabolic theory of ecology. Ecology 85:1771–1789.
Cahill J. F., Jr. and B. B. Casper. 2000. Investigating the relationship between neighbor root biomass and belowground competition: field evidence for symmetric competition belowground. Oikos 90:311–320.
Cahill J. F., Jr. 2002. Interactions between root and shoot competition vary among species. Oikos 99:101–112.
Casper, B. B., H. J. Schenk, and R. B. Jackson. 2003. Defining a plant’s belowground zone of influence. Ecology 84:2313–2321.
Cheng, D. L., and K. J. Niklas. 2007. Above- and belowground biomass relationships across 1534 forested communities. Annals of botany 99:95–102.
Chu, C. J., J. Weiner, F. T. Maestre, Y. S. Wang, C. Morris, S. Xiao, J. L. Yuan, G. Z. Du, and G. Wang. 2010. Effects of positive interactions, size symmetry of competition and abiotic stress on self-thinning in simulated plant populations. Annals of botany 106:647–652.
Coomes, D. A. 2006. Challenges to the generality of WBE theory. Trends in ecology & evolution 21:593–596.
Coomes, D. A., and R. B. Allen. 2009. Testing the metabolic scaling theory of tree growth. Journal of Ecology 97:1369–1373.
Coomes, D. A., E. R. Lines, and R. B. Allen. 2011. Moving on from Metabolic Scaling Theory: hierarchical models of tree growth and asymmetric competition for light. Journal of Ecology 99:748–756.
Deng, J. M., G. X. Wang, E. C. Morris, X. P. Wei, D. X. Li, B. M. Chen, C. M. Zhao, J. Liu, and Y. Wang. 2006. Plant mass–density relationship along a moisture gradient in north‐west China. Journal of Ecology 94:953–958.
Dunn, C. P., and R. R. Sharitz. 1990. The relationship of light and plant geometry to self‐thinning of an aquatic annual herb, Murdannia keisak (Commelinaceae). New Phytologist 115:559–565.
Enquist, B. J. 2002. Universal scaling in tree and vascular plant allometry: toward a

Chapter 3 - 77 -
general quantitative theory linking plant form and function from cells to ecosystems. Tree physiology 22:1045–1064.
Enquist, B. J., J. H. Brown, and G. B. West. 1998. Allometric scaling of plant energetics and population density. Nature 395:163–165.
Enquist, B. J., and K. J. Niklas. 2001. Invariant scaling relations across tree-dominated communities. Nature 410:655–660.
Enquist, B. J., G. B. West, and J. H. Brown. 2009. Extensions and evaluations of a general quantitative theory of forest structure and dynamics. Proceedings of the National Academy of Sciences, USA 106:7046–7051.
Fransen, B., H. de Kroon, and F. Berendse. 2001. Soil nutrient heterogeneity alters competition between two perennial grass species. Ecology 82:2534–2546.
Grimm, V., U. Berger, F. Bastiansen, S. Eliassen, V. Ginot, J. Giske, J. Goss-Custard, T. Grand, S. K. Heinz, and G. Huse. 2006. A standard protocol for describing individual-based and agent-based models. Ecological Modelling 198:115–126.
Grimm, V., U. Berger, D. L. DeAngelis, J. G. Polhill, J. Giske, and S. F. Railsback. 2010. The ODD protocol: A review and first update. Ecological Modelling 221:2760–2768.
Grimm, V., and S. F. Railsback. 2005. Individual-based modelling and ecology. Princeton University Press, USA
Grimm, V., E. Revilla, U. Berger, F. Jeltsch, W. M. Mooij, S. F. Railsback, H. H. Thulke, J. Weiner, T. Wiegand, and D. L. DeAngelis. 2005. Pattern-oriented modelling of agent-based complex systems: lessons from ecology. Science 310:987–991.
Hautier, Y., P. A. Niklaus, and A. Hector. 2009. Competition for light causes plant biodiversity loss after eutrophication. Science 324:636–638.
ISTA. 1993. International rules for seed testing. Seed Science and Technology 21:1–288. Johnson, I. R. 1985. A model of the partitioning of growth between the shoots and roots of
vegetative plants. Annals of Botany 55:421–431. Lamb, E. G., S. W. Kembel, and J. F. Cahill Jr. 2009. Shoot, but not root, competition
reduces community diversity in experimental mesocosms. Journal of Ecology 97:155–163.
Lin, Y., Berger, U., Grimm, V., and Q. R. Ji. 2012. Differences between symmetric and asymmetric facilitation matter: exploring the interplay between different modes of positive and negative plant interactions. Journal of Ecology DOI: 10.1111/j.1365-2745.2012.02019.x
Liu, J., L. Wei, C. M. Wang, G. X., and X. P. Wei. 2006. Effect of Water Deficit on Self‐thinning Line in Spring Wheat (Triticum aestivum L.) Populations. Journal of Integrative Plant Biology 48:415–419.
Lonsdale, W. M., and A. R. Watkinson. 1982. Light and self-thinning. New Phytologist:431–445.
Martin, B. T., E. I. Zimmer, V. Grimm, and T. Jager. 2012. Dynamic Energy Budget theory meets individual-based modelling: a generic and accessible implementation. Methods in Ecology and Evolution 3:445–449.
May, F., V. Grimm, and F. Jeltsch. 2009. Reversed effects of grazing on plant diversity: the role of below‐ground competition and size symmetry. Oikos 118:1830–1843.
McConnaughay, K. D. M., and J. S. Coleman. 1999. Biomass allocation in plants: ontogeny or optimality? A test along three resource gradients. Ecology 80:2581–2593.
Morris, E. C. 2003. How does fertility of the substrate affect intraspecific competition? Evidence and synthesis from self‐thinning. Ecological Research 18:287–305.
Niklas, K. J. 2005. Modelling below-and aboveground biomass for non-woody and woody plants. Annals of Botany 95:315–321.
O’Brien, E. E., J. S. Brown, and J. D. Moll. 2007. Roots in space: a spatially explicit model for belowground competition in plants. Proceedings of the Royal Society B: Biological Sciences 274:929–935.
R Development Core Team (2010) R: a language and environment for statistical computing. R Foundation for Statistical Computing, URL http://www.R-project.org. Last accessed 17 February 2012.

- 78 - Chapter 3
Pretzsch, H. 2006. Species-specific allometric scaling under self-thinning: evidence from long-term plots in forest stands. Oecologia 146:572–583.
Rajaniemi, T. K. 2003. Evidence for size asymmetry of belowground competition. Basic and Applied Ecology 4:239–247.
Rewald, B., and C. Leuschner. 2009. Does root competition asymmetry increase with water availability? Plant Ecology & Diversity 2:255–264.
Reynolds, J. H., and E. D. Ford. 2005. Improving competition representation in theoretical models of self‐thinning: a critical review. Journal of Ecology 93:362–372.
Savage, V. M., L. P. Bentley, B. J. Enquist, J. S. Sperry, D. D. Smith, P. B. Reich, and E. I. von Allmen. 2010. Hydraulic trade-offs and space filling enable better predictions of vascular structure and function in plants. Proceedings of the National Academy of Sciences, USA 107:22722–22727.
Schiffers, K., K. Tielbörger, B. Tietjen, and F. Jeltsch. 2011. Root plasticity buffers competition among plants: theory meets experimental data. Ecology 92:610–620.
Schwinning, S., and J. Weiner. 1998. Mechanisms determining the degree of size asymmetry in competition among plants. Oecologia 113:447–455.
Stoll, P., J. Weiner, H. Muller-Landau, E. Müller, and T. Hara. 2002. Size symmetry of competition alters biomass–density relationships. Proceedings of the Royal Society of London. Series B: Biological Sciences 269:2191–2195.
Tilman, D. 1988. Plant strategies and the dynamics and structure of plant communities. Princeton University Press, USA
Warton, D. I., I. J. Wright, D. S. Falster, and M. Westoby. 2006. Bivariate line‐fitting methods for allometry. Biological Reviews 81:259–291.
Weiner, J. 2004. Allocation, plasticity and allometry in plants. Perspectives in Plant Ecology, Evolution and Systematics 6:207–215.
Weiner, J., and R. P. Freckleton. 2010. Constant final yield. Annual Review of Ecology, Evolution, and Systematics 41:173–192.
Weiner, J., P. Stoll, H. Muller‐Landau, and A. Jasentuliyana. 2001. The effects of density, spatial pattern, and competitive symmetry on size variation in simulated plant populations. The American Naturalist 158:438–450.
Weller, D. E. 1987. Self-thinning exponent correlated with allometric measures of plant geometry. Ecology 68:813–821.
West, G. B., J. H. Brown, and B. J. Enquist. 1999. A general model for the structure, function, and allometry of plant vascular systems. Nature 400:664–667.
West, G. B., J. H. Brown, and B. J. Enquist. 2001. A general model for ontogenetic growth. Nature 413:628–631.
Westoby, M. 1984. The self-thinning rule. Advances in Ecological Research 14:167–225. von Wettberg, E. J., and J. Weiner. 2003. Larger Triticum aestivum plants do not preempt
nutrient-rich patches in a glasshouse experiment. Plant Ecology 169:85–92. Wilensky, U. (1999). NetLogo. Center for Connected Learning and Computer-Based
Modelling, Northwestern University. Available at: http://ccl.northwestern.edu/netlogo/. Last accessed 17 February 2012.
Wilson, J. B. 1988. Shoot competition and root competition. Journal of Applied Ecology 25:279–296.
Wyszomirski, T. 1983. Simulation model of the growth of competing individuals of a plant population. Ekologia Polska 31.
Wyszomirski, T., I. Wyszomirska, and I. Jarzyna. 1999. Simple mechanisms of size distribution dynamics in crowded and uncrowded virtual monocultures. Ecological Modelling 115:253–273.
Yoda, T. Kira, H. Ogawa, and K. Hozumi. 1963. Self-thinning in overcrowded pure stands under cultivated and natural conditions (Intraspecific competition among higher plants XI). Journal of Biology, Osaka City University 14:107–129.

Chapter 3 - 79 -
Chapter 3 – Appendix A
ODD protocol – model description of two-layer pi model
The following model description follows the ODD protocol (Overview, Design
concepts, Details) for describing individual- and agent-based models (Grimm
et al. 2006; Grimm et al. 2010),
1. Purpose
The aim of this model is to evaluate the multiple effects of the mode of
competition (above- and belowground part) and resource limitation on
regulating plant population dynamics, specifically on mass-density
relationships (self-thinning trajectories) and density-dependent mortality. In
particular, we test whether interactions on individual plant level can alter the
slope and intercept of the log mass-log density relationship under different
environmental conditions. The model does not represent specific species, but
generic ones.
2. Entities, state variables, and scales
The entities in the model are plants and square habitat units, or patches (Table
A1). Plants are characterized by the following state variables: initial growth rate,
initial biomass, maximum biomass (asymptotic biomass), current biomass
(both shoot and root) and their position, i.e. coordinates of the stem. Each
individual plant has its own circular zone-of-influence (ZOI) for both above- and
belowground compartment. The pair of ZOIs stand for the physical space
occupied by a plant’s shoot and root respectively, and represents the energy
and resources potentially available to this plant for above- and belowground
part, which ZOIs are allometrically related to its shoot and root mass separately.
Neighbouring plants only compete for the resources when their above- or
belowground ZOIs are overlapping.
In order to make the spatial calculations of resource competition easier,
ZOIs are projected onto a grid of patches. To avoid edge effects, we use a
torus world with a size of 200 × 200 patches (Grimm & Railsback 2005), and
each patch represents 0.25 cm2 in reality. The state of each patch is
characterized by its resource availability. We use a homogeneous environment
here as all patches have the same, and constant, degree of resource limitation
for both above- and belowground part. One time step in the model represents
approximately one week for simulated plants.
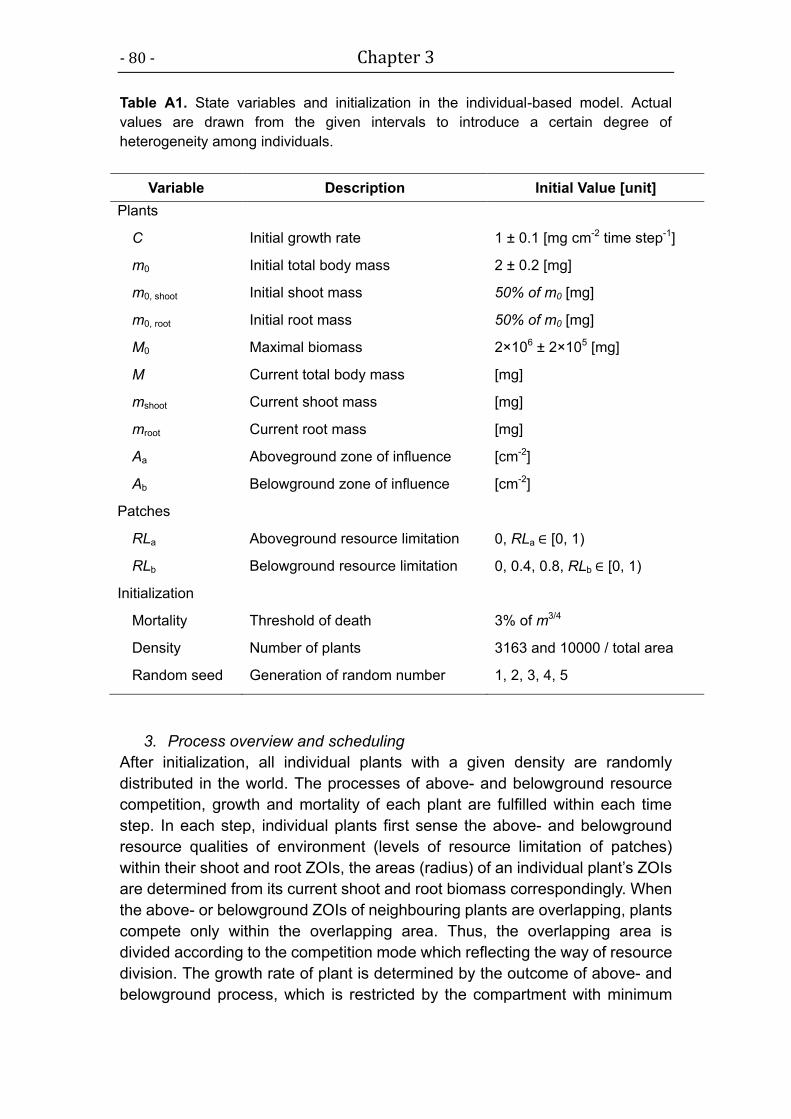
- 80 - Chapter 3
Table A1. State variables and initialization in the individual-based model. Actual
values are drawn from the given intervals to introduce a certain degree of
heterogeneity among individuals.
Variable Description Initial Value [unit]
Plants
C Initial growth rate 1 ± 0.1 [mg cm-2 time step-1]
m0 Initial total body mass 2 ± 0.2 [mg]
m0, shoot Initial shoot mass 50% of m0 [mg]
m0, root Initial root mass 50% of m0 [mg]
M0 Maximal biomass 2×106 ± 2×105 [mg]
M Current total body mass [mg]
mshoot Current shoot mass [mg]
mroot Current root mass [mg]
Aa Aboveground zone of influence [cm-2]
Ab Belowground zone of influence [cm-2]
Patches
RLa Aboveground resource limitation 0, RLa ∈ [0, 1)
RLb Belowground resource limitation 0, 0.4, 0.8, RLb ∈ [0, 1)
Initialization
Mortality Threshold of death 3% of m3/4
Density Number of plants 3163 and 10000 / total area
Random seed Generation of random number 1, 2, 3, 4, 5
3. Process overview and scheduling
After initialization, all individual plants with a given density are randomly
distributed in the world. The processes of above- and belowground resource
competition, growth and mortality of each plant are fulfilled within each time
step. In each step, individual plants first sense the above- and belowground
resource qualities of environment (levels of resource limitation of patches)
within their shoot and root ZOIs, the areas (radius) of an individual plant’s ZOIs
are determined from its current shoot and root biomass correspondingly. When
the above- or belowground ZOIs of neighbouring plants are overlapping, plants
compete only within the overlapping area. Thus, the overlapping area is
divided according to the competition mode which reflecting the way of resource
division. The growth rate of plant is determined by the outcome of above- and
belowground process, which is restricted by the compartment with minimum

Chapter 3 - 81 -
resource uptake rate according to growth function. The synthesized biomass is
allocated to shoot and root optimally which follows the rule of functional
balanced growth (Niklas 2005; May et al. 2009). Plants with growth rates
falling below a threshold die and are removed immediately. The state variables
of the plants are synchronously updated within the subroutines, i.e. changes to
state variables are updated only after all individuals have been processed
(Grimm & Railsback 2005).
4. Design concepts
Basic principles: From “Metabolic Scaling Theory”, we derived a general
ontogenetic growth model for individual plants. We combine this model, via the
ZOI approach, with the effects of different modes of competition for both
above- and belowground compartment and resource limitation.
Emergence: All features observed at the population level, e.g. mass-density
relationship or self-thinning trajectories, size distribution and spatial distribution,
emerge from the interaction of individual plants with their neighbours and the
resource level of their abiotic environment.
Interaction: Individual plants interact via shoot and root competition for
resources in the overlapping area of their ZOIs.
Stochasticity: Initial growth rate, initial biomass (for shoot and root
respectively), maximum biomass and initial position of plants are randomly
taken from the intervals given in Table 1. This introduces a certain level of
heterogeneity among individual characteristics to take into account that real
plants are never exactly identical.
Observation: Population size, shoot and root biomass of each plant, and mean
biomass of all living plants are the main observations.
5. Initialization
Initially, individual plants are randomly distributed according to the chosen
initial density. Resources are spatially and temporally constant. Each plant has
an initial biomass (m0), initial shoot and root biomass (m0, shoot and m0, root),
maximal biomass (M) and initial growth rate (c) drawn from truncated normal
distributions with average and intervals given in Table 1.
6. Input
After initialization, the model does not include any external inputs, i.e. the
abiotic environment is constant.
7. Submodels
One layer model – Plant growth, resource limitation and competition
In our individual-based model the plant’s ZOI, A, stands for the physical space
occupied by the plant and represents the energy and resources potentially
available to this plant. This space is allometrically related to the plant’s body
mass, m, as c0A=m3/4 (Enquist & Niklas 2001), where c0 is a normalization

- 82 - Chapter 3
constant. Since plant growth in our simulation is discrete, therefore equation
(3-1) can be rewritten as
∆m/∆t = cA[ 1 – ( m / M0)1/4 ] (A1)
where c=ac0, is the initial growth rates in units of mass per area and time
interval. For simplicity, we choose 1 ± 0.1 in our model.
Resource limitation and competition usually cause a reduction of resource
availability for plants. We therefore represent resource limitation via a
dimensionless efficiency factor or index, fR, for different levels of resource
availability. Resource competition is incorporated by using a dimensionless
competition factor or index, fp, leading to
∆m/∆t = fRfpcA[ 1 – ( m / M)1/4 ] (A2)
where M=(fRfp)4M0 is the maximum body size with resource limitation and
competition.
The efficiency factor fR, can take different forms depending on the
characteristics and level of the limiting resource. For simplification, we use a
linear form here, i.e. fR = 1–RL, where RL indicates the level of resource
limitation, with its value ranging from 0 (no resource limitation) to 1 (maximum
resource limitation).
As for competition, the modes of resource-mediated competition among
plants can be located somewhere along a continuum between completely
asymmetric competition (largest plants obtain all the contested resources) and
completely symmetric competition (resource uptake is equal for all plants,
independent of their relative sizes; Schwinning & Weiner 1998). To represent
different modes of competition explicitly, we describe the competitive index fp
as
,1
1
( ) /o
j
pn i
p no o knk p
jj
mf A A A
m
(A3)
This factor thus refers to the fraction of resources available in the area
which plant i could obtain after a loss of potential resources due to areas
overlapped by neighbours of sizes mj (Schwinning and Weiner 1998). Ano is
the area not overlapping with neighbours, Ao,k denotes the no areas
overlapping with nj different neighbours. Parameter p determines the mode of
competition, ranging from complete symmetry (p = 0) to complete asymmetry
(p approaching infinity; for details and examples see Figure A2).

Chapter 3 - 83 -
Figure A2. An example of calculating the interactive indexes (Eqns A3) with
different modes of competition and facilitation in an individual-based model as
a way of dividing plants’ ZOI (zone-of-influence). Three plants with sizes m1,
m2 and m3 are interacting in this example. For plant 1, its ZOI (A) was divided
into four parts: Ano, the area not overlapping with the other two plants; Ao,1, the
area overlapping with plant 2; Ao,2, the area overlapping with plants 2 and 3;
Ao,3, the area overlapping with plant 3. Then the actual area that plant 1 can
take from Ao,1 is
1 1,1 ,12
1 21
p p
o o p pp
jj
m mA A
m mm
For Ao,2,
1 1,2 ,23
1 2 31
p p
o o p p pp
jj
m mA A
m m mm
And for Ao,3,
1 1,3 ,32
1 31
p p
o o p pp
jj
m mA A
m mm
Therefore, the competitive index for plant 1 is:
Where 3/4
1 0/A m c
Two layer model – Shoot versus root competition, biomass allocation and
mortality
Because competition among plants can occurs at both above- and
belowground simultaneously, the relative importance of shoot versus root
competition and their ZOI can varies depending on the environmental factors
(Deng et al. 2006, May et al. 2009). Therefore, the one layer model however
cannot properly represent this property. We developed the two layer

- 84 - Chapter 3
individual-based model to represent the plant’s shoot and root competition. In
our two layer model, a plant has two ZOIs stand for the above- and
belowground physical space occupied by the plant, which ZOIs represent the
corresponding levels of energy and resources (e.g. light, water and nutrient)
potentially available to this plant.
We assume that (i) under optimal conditions without resource limitation
and competition, the abilities of above- and belowground resource uptake are
balanced, with relationships between metabolic rate, B, and biomass, m, being
B=cshootmshoot3/4=crootmroot
3/4 (Niklas 2005, Cheng and Niklas 2007), where cshoot
and croot are normalization constants (to simplify, we assume cshoot=croot=1),
and “shoot” and “root” refer to the above- and belowground compartment,
respectively; (ii) the plant’s above- and belowground ZOIs are proportional to
the plant metabolic rate, B, and then allometrically related to the plant’s shoot
and root biomass (Enquist and Niklas 2001, May et al. 2009), Aa=camshoot3/4
and Ab=cbmroot3/4, where ca and cb are normalization constants (to simplify, we
use ca=cb=1); (iii) growth of the entire plant is limited by the compartment with
smaller resource uptake rate (May et al. 2009). In a view of comparability with
one-layer model, the real growth rate of whole plant in two-layer model has
been doubled. Accordingly, equation (A2) can be applied to above- and
belowground compartment, and then we get the whole plant growth rate as
1/4
, ,
1/4
, ,
2 2 [1 ( / ) ],
2 2 [1 ( / ) ], (A4)
,
R a p a a a a
R b p b b b b
AGR f f c A m M AGR BGR
mBGR f f c A m M AGR BGR
tAGR BGR AGR BGR
where ∆AGR and ∆BGR are above- and belowground determined growth rate.
The factor of resource availability (fR), competitive index (fp) and ZOI (A) were
applied to both two layers independently, with the corresponding subscript a
and b indicate the above- and belowground compartment respectively (see
Figure A3).
Adjustability of root/shoot allocation as the morphological plasticity allows
plants to adapt to changing biotic and abiotic environmental conditions (Berger
et al. 2008). We adopt the optimal allocation theory, functional balance growth
hypothesis and metabolic scaling theory to quantify the partition of growth
between the shoots and roots (Weiner 2004; Niklas 2005; May et al. 2009):
3/4
3/4 3/4
3/4
3/4 3/4
(A5a)
(A5b)
shoot
root
m m BGR
t t AGR BGR
m m AGR
t t AGR BGR
which means plant allocates more biomass to the compartment that is most

Chapter 3 - 85 -
limiting growth for increasing its uptake of resource. An allometric form (3/4) of resources allocation was used here as metabolic balance, we also tested the original allocation of linearity which only lead to a small difference on root/shoot ratio but do not change our general findings.
An individual’s mortality rate is proportional to its mass-specific
metabolism (as current total metabolic rate divide by body mass; Brown et al.
2004). Based on this, we assume that individuals die if their actual growth rate
(∆m/∆t, represent actual metabolic rate) falls below a threshold of their basal
metabolic rate (allometrically scaled with body mass), i.e. 3% of m3/4.
Therefore, individual plants may die due to metabolic inactivation driven by
above- or/and belowground resource limitation, competition, senescence
(when m approaches M) or combinations thereof. This provides a more
realistic representation of relevant ecological process than in previous models
(Stoll et al. 2002; Chu et al. 2009, 2010). In addition, we are able to ascribe the
mortality of individual plants to above- or belowground process explicitly.
In total, equations (A4), (A5a) and (A5b) clearly showed how a plant’s
growth, biomass allocation and mortality are jointly determined by above- and
belowground resource level and local competition.
Figure A3. A visual illustration of the two-layer zone-of-influence (ZOI) approach
including both above- and belowground competition. The above- and belowground
ZOIs (green: aboveground, gray: belowground) are allometrically related to the plant
shoot and root biomass, respectively. Plants only compete for the resources in
overlapped areas with their neighbours which can occur independently at above- and
belowground compartment (arrows indicate the overlapped area of aboveground
competition).

- 86 - Chapter 3
References
Berger, U., Piou, C., Schiffers, K. & Grimm, V. (2008). Competition among plants: Concepts, individual-based modelling approaches, and a proposal for a future research strategy. Perspect. Plant Ecol., 9, 121-135.
Brown, J.H., Gillooly, J.F., Allen, A.P., Savage, V.M. & West, G.B. (2004). Toward a metabolic theory of ecology. Ecology, 85, 1771-1789.
Chu, C.J., Weiner, J., Maestre, FT, Xiao, S., Wang, Y.-S., Li, Q., Yuan, J.L., Zhao, L.Q., Ren, Z.W. and Wang, G.(2009), Positive interactions can increase size inequality in plant populations. J. Ecol., 97, 1401–1407
Chu, C.J., Weiner, J., Maestre, F.T., Wang, Y.S., Morris, C., Xiao, S. et al. (2010). Effects of positive interactions, size symmetry of competition and abiotic stress on self-thinning in simulated plant populations. Ann. Bot., 106, 647-652.
Deng, J.M., Wang, G.X., Morris, E.C., Wei, X.P., Li, D.X., Chen, B.M. et al. (2006). Plant mass-density relationship along a moisture gradient in north-west China. J. Ecol., 94, 953–958.
Enquist, B.J. (2002). Universal scaling in tree and vascular plant allometry: toward a general quantitative theory linking plant form and function from cells to ecosystems. Tree Physiol., 22, 1045-1064.
Enquist, B.J. & Niklas, K.J. (2001). Invariant scaling relations across tree-dominated communities. Nature, 410, 655-660.
Enquist, B.J., West, G.B. & Brown, J.H. (2009). Extensions and evaluations of a general quantitative theory of forest structure and dynamics. Proc. Natl Acad. Sci. USA., 106, 7046-7051.
Grimm, V., Berger, U., Bastiansen, F., Eliassen, S., Ginot, V., Giske, J. et al. (2006). A standard protocol for describing individual-based and agent-based models. Ecol. Model., 198, 115-126.
Grimm, V., Berger, U., DeAngelis, D.L., Polhill, J.G., Giske, J. & Railsback, S.F. (2010). The ODD protocol: A review and first update. Ecol. Model.. 221, 2760-2768.
Grimm, V. & Railsback, S.F. (2005). Individual-based modeling and ecology. Princeton University Press, Princeton and Oxford.
Hou, C., Zuo, W., Moses, M.E., Woodruff, W.H., Brown, J.H. & West, G.B. (2008). Energy uptake and allocation during ontogeny. Science, 322, 736-739.
Lambers, H., Chapin, F.S. & Pons, T.L. (2008). Plant physiological ecology. 2nd edition. Springer Verlag, New York, pp. 134–143.
Maestre, F.T., Callaway, R.M., Valladares, F. & Lortie, C.J. (2009). Refining the stress-gradient hypothesis for competition and facilitation in plant communities. J. Ecol., 97, 199–205.
May, F., Grimm, V. & Jeltsch, F. (2009). Reversed effects of grazing on plant diversity: the role of below-ground competition and size symmetry. Oikos, 118, 1830–1843.
Molofsky, J. & Bever, J.D. (2002). A novel theory to explain species diversity in landscapes: positive frequency dependence and habitat suitability. Proc. R. Soc. Lond., B, Biol. Sci., 269, 2389–2393.
Molofsky, J., Bever, J.D. & Antonovics, J. (2001). Coexistence under positive frequency dependence. Proc. R. Soc. Lond., B, Biol. Sci., 2001, 268–277.
Niklas, K.J. (2005). Modelling below-and above-ground biomass for non-woody and woody plants. Ann. Bot., 95, 315-321.
Niklas, K.J. & Enquist, B.J. (2001). Invariant scaling relationships for interspecific plant biomass production rates and body size. Proc. Natl Acad. Sci. USA., 98, 2922-2927.
Price, C.A., Enquist, B.J. & Savage, V.M. (2007). A general model for allometric covariation in botanical form and function. Proc. Natl Acad. Sci. USA., 104, 13204-13209.
Sack, L., Maranon, T., Grubb, P.J., Enquist, B.J. & Niklas, K.J. (2002). Global allocation rules for patterns of biomass partitioning. Science, 296, 1923a.
Schwinning, S. & Weiner, J. (1998). Mechanisms determining the degree of size asymmetry in competition among plants. Oecologia, 113, 447-455.

Chapter 3 - 87 -
Stoll, P., Weiner, J., Muller-Landau, H., Müller, E. & Hara, T. (2002). Size symmetry of competition alters biomass–density relationships. Proc. R. Soc. Lond. B: Biol. Sci., 269, 2191-2195.
Travis, J.M.J., Brooker, R.W., Clark, E.J. & Dytham, C. (2006). The distribution of positive and negative species interactions across environmental gradients on a dual-lattice model. J. Theor. Biol., 241, 896–902.
Weiner, J., Stoll, P., Muller-Landau, H. & Jasentuliyana, A. (2001). The effects of density, spatial pattern, and competitive symmetry on size variation in simulated plant populations. The American Naturalist, 158, 438-450.
Weiner, J. (2004). Allocation, plasticity and allometry in plants. Perspectives in Plant Ecology, Evolution and Systematics, 6, 207-215.
West, G.B., Brown, J.H. & Enquist, B.J. (1999). A general model for the structure and allometry of plant vascular systems. Nature, 400, 664-667.
West, G.B., Brown, J.H. & Enquist, B.J. (2001). A general model for ontogenetic growth. Nature, 413, 628-631.
West, G.B., Enquist, B.J. & Brown, J.H. (2009). A general quantitative theory of forest structure and dynamics. Proc. Natl Acad. Sci., 106, 7040-7045.
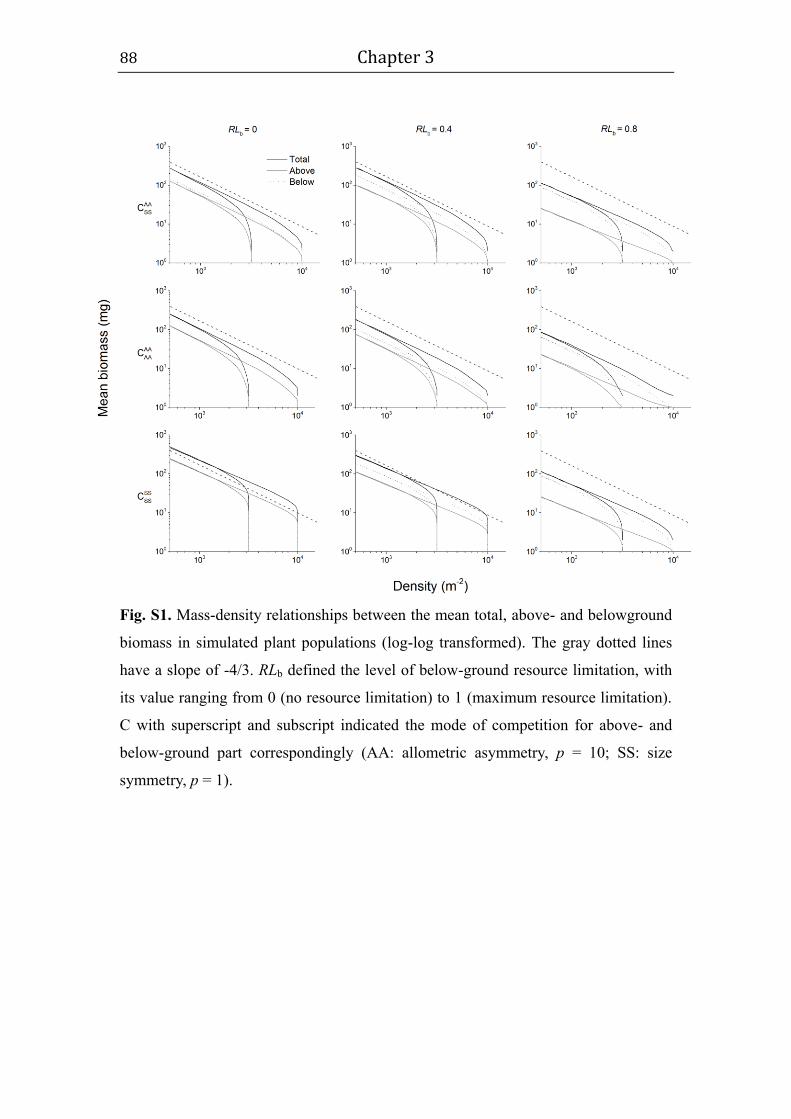
88 Chapter 3
Fig. S1. Mass-density relationships between the mean total, above- and belowground
biomass in simulated plant populations (log-log transformed). The gray dotted lines
have a slope of -4/3. RLb defined the level of below-ground resource limitation, with
its value ranging from 0 (no resource limitation) to 1 (maximum resource limitation).
C with superscript and subscript indicated the mode of competition for above- and
below-ground part correspondingly (AA: allometric asymmetry, p = 10; SS: size
symmetry, p = 1).

Chapter 3 89
Table S1. Slopes and intercepts of mass-density relationships between the mean total,
above- and belowground biomass in simulated plant populations (log-log transformed;
as estimated by standard major axis regression). RLb defined the level of belowground
resource limitation, with its value ranging from 0 (no resource limitation) to 1
(maximum resource limitation). CA: complete asymmetry, p = ∞; AA: allometric
asymmetry, p = 10; SS: size symmetry, p = 1; CS: complete symmetry, p = 0.
Resource limitation Mode of Competition Mean total biomass
RLb Aboveground Belowground Slope 95% CI Intercept 95% CI R2
0 CA CA -1.450 -1.509 -1.399 6.423 6.243 6.616 0.990
0 CA AA -1.475 -1.537 -1.416 6.502 6.290 6.707 0.988
0 CA SS -1.417 -1.461 -1.377 6.333 6.196 6.479 0.993
0 CA CS -1.439 -1.482 -1.401 6.420 6.289 6.563 0.993
0 AA CA -1.475 -1.538 -1.413 6.502 6.288 6.710 0.988
0 AA AA -1.351 -1.391 -1.316 6.109 5.987 6.242 0.996
0 AA SS -1.359 -1.394 -1.329 6.174 6.068 6.286 0.996
0 AA CS -1.401 -1.426 -1.379 6.348 6.273 6.430 0.998
0 SS CA -1.417 -1.462 -1.376 6.333 6.195 6.480 0.993
0 SS AA -1.359 -1.394 -1.329 6.174 6.068 6.286 0.996
0 SS SS -1.100 -1.110 -1.092 5.647 5.618 5.678 0.999
0 SS CS -1.351 -1.365 -1.342 6.764 6.728 6.805 0.994
0 CS CA -1.439 -1.482 -1.401 6.420 6.289 6.563 0.993
0 CS AA -1.401 -1.425 -1.380 6.348 6.273 6.429 0.998
0 CS SS -1.351 -1.363 -1.340 6.764 6.727 6.803 0.994
0 CS CS -1.060 -1.060 -1.059 5.868 5.866 5.870 1.000
0.4 CA CA -1.319 -1.344 -1.298 5.845 5.776 5.921 0.997
0.4 CA AA -1.319 -1.345 -1.294 5.849 5.771 5.929 0.996
0.4 CA SS -1.391 -1.421 -1.363 6.261 6.170 6.352 0.992
0.4 CA CS -1.425 -1.448 -1.403 6.405 6.337 6.474 0.994
0.4 AA CA -1.315 -1.340 -1.293 5.834 5.762 5.911 0.997
0.4 AA AA -1.288 -1.309 -1.269 5.758 5.697 5.822 0.998
0.4 AA SS -1.292 -1.313 -1.273 5.980 5.921 6.044 0.996
0.4 AA CS -1.311 -1.345 -1.279 6.286 6.190 6.387 0.957
0.4 SS CA -1.325 -1.352 -1.301 5.884 5.810 5.970 0.996
0.4 SS AA -1.281 -1.298 -1.267 5.754 5.707 5.805 0.999
0.4 SS SS -1.096 -1.101 -1.093 5.428 5.416 5.441 1.000
0.4 SS CS -1.017 -1.017 -1.016 5.391 5.390 5.393 1.000
0.4 CS CA -1.333 -1.361 -1.308 5.919 5.841 6.005 0.996
0.4 CS AA -1.285 -1.303 -1.268 5.775 5.722 5.831 0.998
0.4 CS SS -1.102 -1.108 -1.097 5.483 5.466 5.501 0.999
0.4 CS CS -1.021 -1.021 -1.020 5.398 5.397 5.400 1.000
0.8 CA CA -1.286 -1.303 -1.271 5.436 5.387 5.489 0.996
0.8 CA AA -1.263 -1.274 -1.251 5.373 5.336 5.409 0.998
0.8 CA SS -1.143 -1.151 -1.135 5.155 5.131 5.179 0.999
0.8 CA CS -1.038 -1.041 -1.035 4.889 4.880 4.898 1.000
0.8 AA CA -1.285 -1.301 -1.270 5.433 5.387 5.484 0.996

90 Chapter 3
0.8 AA AA -1.256 -1.267 -1.244 5.350 5.318 5.387 0.998
0.8 AA SS -1.132 -1.137 -1.127 5.118 5.102 5.134 0.999
0.8 AA CS -1.034 -1.035 -1.032 4.874 4.870 4.878 1.000
0.8 SS CA -1.284 -1.301 -1.269 5.434 5.385 5.482 0.996
0.8 SS AA -1.257 -1.258 -1.237 5.329 5.297 5.361 0.998
0.8 SS SS -1.127 -1.132 -1.123 5.106 5.092 5.121 0.999
0.8 SS CS -1.033 -1.034 -1.032 4.870 4.867 4.873 1.000
0.8 CS CA -1.285 -1.302 -1.269 5.436 5.387 5.487 0.996
0.8 CS AA -1.248 -1.259 -1.238 5.331 5.300 5.365 0.998
0.8 CS SS -1.122 -1.127 -1.118 5.092 5.079 5.106 0.999
0.8 CS CS -1.033 -1.034 -1.032 4.871 4.867 4.874 1.000
Resource limitation Mode of Competition Mean aboveground biomass
RLb Aboveground Belowground Slope 95% CI Intercept 95% CI R2
0 CA CA -1.450 -1.507 -1.396 6.122 5.932 6.310 0.990
0 CA AA -1.477 -1.543 -1.417 6.204 5.991 6.423 0.988
0 CA SS -1.387 -1.432 -1.348 5.907 5.775 6.054 0.993
0 CA CS -1.392 -1.432 -1.353 5.922 5.790 6.056 0.993
0 AA CA -1.474 -1.533 -1.414 6.198 5.989 6.395 0.988
0 AA AA -1.351 -1.392 -1.315 5.807 5.685 5.943 0.996
0 AA SS -1.333 -1.367 -1.303 5.758 5.658 5.870 0.996
0 AA CS -1.359 -1.382 -1.338 5.869 5.800 5.944 0.997
0 SS CA -1.443 -1.491 -1.401 6.145 5.999 6.299 0.993
0 SS AA -1.384 -1.420 -1.350 5.977 5.861 6.091 0.996
0 SS SS -1.100 -1.110 -1.109 5.346 5.315 5.377 0.999
0 SS CS -1.308 -1.321 -1.299 6.282 6.244 6.321 0.995
0 CS CA -1.471 -1.516 -1.430 6.293 6.154 6.436 0.993
0 CS AA -1.438 -1.464 -1.414 6.203 6.124 6.290 0.997
0 CS SS -1.390 -1.404 -1.378 6.623 6.583 6.667 0.993
0 CS CS -1.060 -1.060 -1.059 5.567 5.565 5.569 1.000
0.4 CA CA -1.300 -1.324 -1.280 5.414 5.349 5.486 0.997
0.4 CA AA -1.299 -1.322 -1.278 5.409 5.343 5.478 0.997
0.4 CA SS -1.314 -1.340 -1.290 5.586 5.510 5.667 0.994
0.4 CA CS -1.318 -1.339 -1.298 5.616 5.557 5.681 0.995
0.4 AA CA -1.294 -1.316 -1.274 5.396 5.333 5.462 0.998
0.4 AA AA -1.271 -1.289 -1.253 5.327 5.273 5.383 0.999
0.4 AA SS -1.227 -1.242 -1.212 5.340 5.292 5.389 0.997
0.4 AA CS -1.138 -1.170 -1.107 5.232 5.137 5.330 0.950
0.4 SS CA -1.313 -1.338 -1.291 5.496 5.423 5.573 0.997
0.4 SS AA -1.272 -1.287 -1.258 5.368 5.325 5.416 0.999
0.4 SS SS -1.070 -1.073 -1.067 4.929 4.920 4.938 1.000
0.4 SS CS -0.888 -0.889 -0.886 4.434 4.431 4.438 1.000
0.4 CS CA -1.333 -1.359 -1.309 5.583 5.506 5.665 0.996
0.4 CS AA -1.286 -1.303 -1.270 5.436 5.385 5.488 0.998
0.4 CS SS -1.100 -1.105 -1.095 5.089 5.073 5.107 1.000
0.4 CS CS -0.899 -0.900 -0.897 4.450 4.444 4.456 0.999
0.8 CA CA -1.152 -1.155 -1.149 4.477 4.467 4.488 1.000
0.8 CA AA -1.138 -1.114 -1.136 4.438 4.432 4.444 1.000
0.8 CA SS -1.020 -1.022 -1.018 4.138 4.132 4.144 1.000
Chapter 3 91
0.8 CA CS -0.911 -0.917 -0.905 3.792 3.773 3.811 0.996
0.8 AA CA -1.149 -1.152 -1.146 4.469 4.459 4.479 1.000
0.8 AA AA -1.134 -1.136 -1.133 4.428 4.422 4.434 1.000
0.8 AA SS -1.021 -1.024 -1.018 4.144 4.135 4.152 1.000
0.8 AA CS -0.921 -0.927 -0.915 3.821 3.802 3.840 0.996
0.8 SS CA -1.147 -1.151 -1.143 4.469 4.457 4.481 1.000
0.8 SS AA -1.129 -1.131 -1.128 4.418 4.413 4.424 1.000
0.8 SS SS -1.013 -1.015 -1.010 4.125 4.116 4.133 1.000
0.8 SS CS -0.917 -0.923 -0.911 3.808 3.788 3.828 0.996
0.8 CS CA -1.147 -1.152 -1.143 4.473 4.461 4.487 1.000
0.8 CS AA -1.129 -1.131 -1.127 4.420 4.414 4.426 1.000
0.8 CS SS -1.011 -1.013 -1.009 4.121 4.114 4.129 1.000
0.8 CS CS -0.918 -0.924 -0.911 3.808 3.789 3.830 0.996
Resource limitation Mode of Competition Mean belowground biomass
RLb Aboveground Belowground Slope 95% CI Intercept 95% CI R2
0 CA CA -1.450 -1.508 -1.395 6.122 5.933 6.317 0.990
0 CA AA -1.474 -1.534 -1.415 6.198 5.986 6.399 0.988
0 CA SS -1.443 -1.491 -1.401 6.145 5.999 6.299 0.993
0 CA CS -1.471 -1.514 -1.429 6.293 6.154 6.436 0.993
0 AA CA -1.477 -1.541 -1.419 6.204 5.999 6.421 0.988
0 AA AA -1.351 -1.389 -1.314 5.807 5.681 5.936 0.996
0 AA SS -1.384 -1.419 -1.350 5.977 5.862 6.092 0.996
0 AA CS -1.438 -1.464 -1.414 6.203 6.124 6.289 0.997
0 SS CA -1.387 -1.432 -1.348 5.907 5.775 6.054 0.993
0 SS AA -1.333 -1.367 -1.303 5.758 5.658 5.870 0.996
0 SS SS -1.100 -1.109 -1.092 5.346 5.316 5.375 0.999
0 SS CS -1.390 -1.405 -1.378 6.623 6.581 6.666 0.993
0 CS CA -1.392 -1.432 -1.352 5.922 5.791 6.058 0.993
0 CS AA -1.359 -1.380 -1.339 5.869 5.790 5.945 0.997
0 CS SS -1.308 -1.319 -1.297 6.282 6.246 6.320 0.995
0 CS CS -1.060 -1.060 -1.059 5.567 5.565 5.569 1.000
0.4 CA CA -1.333 -1.361 -1.308 5.650 5.571 5.737 0.996
0.4 CA AA -1.334 -1.364 -1.308 5.660 5.576 5.752 0.996
0.4 CA SS -1.440 -1.473 -1.407 6.213 6.110 6.315 0.992
0.4 CA CS -1.487 -1.514 -1.462 6.411 6.334 6.494 0.993
0.4 AA CA -1.331 -1.359 -1.306 5.643 5.564 5.728 0.996
0.4 AA AA -1.312 -1.335 -1.289 5.563 5.489 5.633 0.998
0.4 AA SS -1.332 -1.355 -1.312 5.908 5.840 5.978 0.995
0.4 AA CS -1.395 -1.430 -1.364 6.387 6.291 6.491 0.958
0.4 SS CA -1.334 -1.361 -1.309 5.657 5.575 5.470 0.996
0.4 SS AA -1.289 -1.307 -1.274 5.525 5.478 5.579 0.998
0.4 SS SS -1.114 -1.119 -1.108 5.271 5.255 5.288 1.000
0.4 SS CS -1.072 -1.072 -1.071 5.418 5.415 5.421 1.000
0.4 CS CA -1.332 -1.360 -1.308 5.650 5.572 5.735 0.996
0.4 CS AA -1.284 -1.302 -1.269 5.510 5.462 5.563 0.998
0.4 CS SS -1.104 -1.110 -1.099 5.258 5.242 5.277 0.999
0.4 CS CS -1.070 -1.071 -1.069 5.414 5.411 5.417 1.000
0.8 CA CA -1.340 -1.365 -1.320 5.485 5.416 5.559 0.993
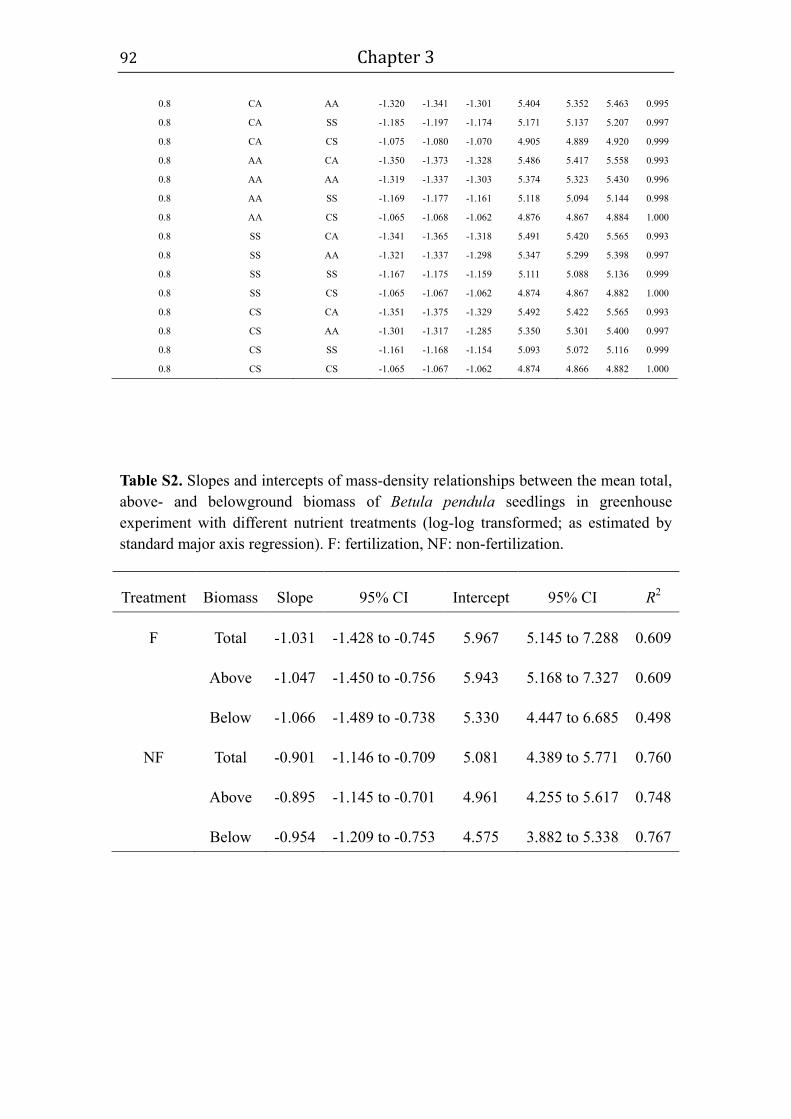
92 Chapter 3
Table S2. Slopes and intercepts of mass-density relationships between the mean total,
above- and belowground biomass of Betula pendula seedlings in greenhouse
experiment with different nutrient treatments (log-log transformed; as estimated by
standard major axis regression). F: fertilization, NF: non-fertilization.
0.8 CA AA -1.320 -1.341 -1.301 5.404 5.352 5.463 0.995
0.8 CA SS -1.185 -1.197 -1.174 5.171 5.137 5.207 0.997
0.8 CA CS -1.075 -1.080 -1.070 4.905 4.889 4.920 0.999
0.8 AA CA -1.350 -1.373 -1.328 5.486 5.417 5.558 0.993
0.8 AA AA -1.319 -1.337 -1.303 5.374 5.323 5.430 0.996
0.8 AA SS -1.169 -1.177 -1.161 5.118 5.094 5.144 0.998
0.8 AA CS -1.065 -1.068 -1.062 4.876 4.867 4.884 1.000
0.8 SS CA -1.341 -1.365 -1.318 5.491 5.420 5.565 0.993
0.8 SS AA -1.321 -1.337 -1.298 5.347 5.299 5.398 0.997
0.8 SS SS -1.167 -1.175 -1.159 5.111 5.088 5.136 0.999
0.8 SS CS -1.065 -1.067 -1.062 4.874 4.867 4.882 1.000
0.8 CS CA -1.351 -1.375 -1.329 5.492 5.422 5.565 0.993
0.8 CS AA -1.301 -1.317 -1.285 5.350 5.301 5.400 0.997
0.8 CS SS -1.161 -1.168 -1.154 5.093 5.072 5.116 0.999
0.8 CS CS -1.065 -1.067 -1.062 4.874 4.866 4.882 1.000
Treatment Biomass Slope 95% CI Intercept 95% CI R2
F Total -1.031 -1.428 to -0.745 5.967 5.145 to 7.288 0.609
Above -1.047 -1.450 to -0.756 5.943 5.168 to 7.327 0.609
Below -1.066 -1.489 to -0.738 5.330 4.447 to 6.685 0.498
NF Total -0.901 -1.146 to -0.709 5.081 4.389 to 5.771 0.760
Above -0.895 -1.145 to -0.701 4.961 4.255 to 5.617 0.748
Below -0.954 -1.209 to -0.753 4.575 3.882 to 5.338 0.767

Chapter 4
Exploring the interplay between modes of positive
and negative plant interactions *
Abstract
Facilitation (positive interaction) has received increasing attention in plant
ecology over the last decade. Just as for competition, distinguishing different
modes of facilitation (mutualistic, commensal or even antagonistic) may be
crucial. We therefore introduce the new concept of symmetric vs. asymmetric
facilitation and present a generic individual-based zone-of-influence model.
The model simultaneously implements different modes of both facilitation and
competition among individual plants via their overlapping zone of influence.
Because we consider facilitation modes as a continuum related to
environmental context, we integrated this concept with the stress gradient
hypothesis (SGH) by exploring differences in spatial pattern formation in
self-thinning plants along a stress gradient in our model. The interplay among
* A slightly revised version of this chapter has been published as Yue Lin1, 2
, Uta Berger1,
Volker Grimm2, 3
, and Qian-Ru Ji4 (2012) Differences between symmetric and asymmetric
facilitation matter: exploring the interplay between modes of positive and negative plant
interactions. Journal of Ecology 100: 1482–1491.
1 Institute of Forest Growth and Computer Science, Dresden University of Technology, P.O.
1117, 01735 Tharandt, Germany; 2 Helmholtz Centre for Environmental Research – UFZ,
Department of Ecological Modelling, 04318 Leipzig, Germany; 3 Institute for Biochemistry
and Biology, University of Potsdam, Maulbeerallee 2, 14469 Potsdam, Germany; 4
Institute of Hydrobiology, Dresden University of Technology, Zellescher Weg 40, 01062
Dresden, Germany

- 94 - Chapter 4
modes of interaction creates distinctly varied spatial patterns along stress
gradients. When competition was symmetric, symmetric facilitation (mutualism)
consistently led to plant aggregation along stress gradients. However,
asymmetric facilitation (commensalism) produces plant aggregation only under
more benign conditions but tends to intensify local competition and spatial
segregation when conditions are harsh. When competition was completely
asymmetric, different modes of facilitation contributed little to spatial
aggregation. Symmetric facilitation significantly increased survival at the
severe end of the stress gradient, which supports the claim of the SGH that
facilitation should have generally positive net effects on plants under high
stress levels. Asymmetric facilitation, however, was found to increase survival
only under intermediate stress conditions, which contradicts the current
predictions of the SGH. Our modelling study demonstrates that the interplay
between modes of facilitation and competition affects different aspects of plant
populations and communities, implying context-dependent outcomes and
consequences. The explicit consideration of the modes and mechanisms of
interactions (both facilitation and competition) and the nature of stress factors
will help to extend the framework of the SGH and foster research on facilitation
in plant ecology.
Keywords: asymmetry, competition, metabolic scaling theory, plant-plant
interaction, plant population and community dynamics, self-thinning, spatial
pattern, stress gradient hypothesis, symmetry

Chapter 4 - 95 -
4.1 Introduction
The role of positive interactions in driving population and community dynamics
has received significant attention and is now widely recognized in both
empirical and theoretical ecology (Bertness & Callaway 1994; Brooker et al.
2008; Bronstein 2009; Maestre et al. 2009; Fajardo & McIntire 2011; McIntire &
Fajardo 2011). In plant ecology, positive interactions are usually referred to as
facilitation, which has been defined as the beneficial effects of neighbours via
the amelioration of habitat (Bertness & Callaway 1994; Bronstein 2009); e.g.
via moderation of stress, enrichment of nutrients, or increased access to
nutrients. Facilitation has been shown particularly important when considering
the performance of plants under stressful environmental conditions. ‘Stress’ is
not a precise concept (Maestre et al. 2009), and can be biotic and abiotic
(Bronstein 2009). The best-understood examples of plant facilitation were
mostly carried out under abiotic stress conditions (Bronstein 2009). Moreover,
the characteristics of abiotic stress factors are also different and can be
resource-independent (e.g. wind, frost and salinity) or resource-dependent
(e.g. water, nutrient and light; Maestre et al. 2009).
The “stress gradient hypothesis” (SGH) proposes that competition and
facilitation may act simultaneously, but the relative importance of facilitation
and competition will vary inversely along gradients of abiotic stress (Bertness
& Callaway 1994). Under high stress conditions, facilitation should be
dominant over competition in affecting community structures (Bertness &
Callaway 1994; Brooker et al. 2008; Maestre et al. 2009). The SGH was
originally formulated at the interspecific level, but recent studies revealed that
SGH is also valid at the intraspecific level (Chu et al. 2008, 2009; Eränen &
Kozlov 2008; McIntire & Fajardo 2011). The interplay between facilitation and
competition can thus drive intraspecific population dynamics (Chu et al. 2008,
2009, 2010; Jia et al. 2011; McIntire & Fajardo 2011), community structure

- 96 - Chapter 4
(Gross 2008; Xiao et al. 2009), community diversity (Cavieres & Badano 2009),
and ecosystem functions (Callaway et al. 2002; Kikvidze et al. 2005); and can
even have evolutionary consequences (Bronstein 2009; McIntire & Fajardo
2011).
However, there are also studies which do not support SGH predictions, as
facilitative effects have not been detected under some extreme stress
conditions (Tielbörger & Kadmon 2000; Maestre et al. 2005, 2009). This
indicates that the conceptual framework underlying the SGH might need
further refinement (Maestre et al. 2009). Moreover, whereas numerous studies
have explored the consequences of different modes of competition, i.e.
symmetric versus asymmetric competition (Schwinning & Weiner 1998;
Weiner et al. 2001; Stoll & Bergius 2005; Berger et al. 2008), different modes
of facilitation have not yet been explored. Inconsistent definitions of facilitation
and the lack of differentiation between the impacts of plant-plant interactions
on beneficiary and benefactor individuals have recently been identified as
important gaps in current research (Brooker et al. 2008; Bronstein 2009;
Brooker & Callaway 2009; Pakeman et al. 2009). Refining and clarifying the
concept of facilitation is crucial for understanding how facilitation arises,
persists and evolves, and then could help extend the general SGH framework
and improve plant ecology research in general (Brooker et al. 2008; Bronstein
2009; Maestre et al. 2009).
According to different definitions, modes of facilitation can be mutualistic
(+/+) or commensal (+/0) amongst plants (Brooker et al. 2008; Bronstein 2009).
However, a more continuous approach to facilitative interactions might be
more accurate and useful, as has been the case for the corresponding
continuous approach to exploring competitive interactions (Schwinning &
Weiner 1998). We therefore suggest using a new concept of modes of
facilitation: any facilitation among plants, no matter whether inter- or
intra-specific, can be placed along a continuum ranging from completely
symmetric facilitation (interacting plants receive the same amount of benefit

Chapter 4 - 97 -
from each other, irrespective of their species or sizes) to completely
asymmetric facilitation (Vellend 2008; the beneficiary plant receives all benefits
but there are no positive effects on the benefactor; Table 4.1). Thus,
mutualistic cases are expected to be at the symmetric end of the facilitation
continuum, and commensal cases at the asymmetric end (Fig. S1).
Table 4.1 Definition and description of facilitation modes. The index q determines the
mode of facilitation among plants (see equation 5-5 and Appendix A).
Mode of
facilitation Effect and definition
Index
value
Expected prevalent
nature of stress factor
Complete
symmetry
All plants receive the same
amount of benefit from each
other, irrespective of their
species or sizes
q = 0
Symmetry:
temperature, moisture,
nutrient, salinity,
pollution, wind
desiccation, altitude etc.
Partial
symmetry
Benefit increases with
benefactor’s size, but less
than proportionally
0 < q < 1
Proportional
symmetry
Benefit is proportional to
benefactor’s size (equal gain
per unit size)
q = 1
Partial
asymmetry
Benefit increases with
benefactor’s size
super-linearly
q > 1 Asymmetry:
light, UV-radiation,
transpiration, tide, water,
nutrient fixation,
pollution, wind,
herbivory, etc.
Complete
asymmetry
The beneficiary plant
receives all benefits, with no
advantage to the benefactor
plants
q = ∞
This new conceptual model (with its terminology) has two main advantages:
it is analogous, and therefore directly comparable to the widely used and
important concept of symmetric and asymmetric competition (Schwinning &

- 98 - Chapter 4
Weiner 1998; Weiner et al. 2001; Stoll & Bergius 2005; Berger et al. 2008); it
also offers a quantitative and operational means of evaluating facilitative
impacts.
The analogy between different modes of competition and facilitation is
evident. In reality, competition and facilitation often interact: clusters of cohorts
facilitate each other against cold or wind desiccation (symmetric facilitation),
but they may also compete for nutrients and water (symmetric competition;
Fajardo & McIntire 2011). Adult nurse plants facilitate the growth and survival
of small plants of their own or other species (asymmetric facilitation), which
may lead to asymmetric light competition if the crown of an adult plant is very
dense or the small plants are not “stress-tolerant” (Reinhart et al. 2006;
Maestre et al. 2009). In general, modes of interaction depend on both the
ecological traits of the interacting plants and the nature of the stress factors
themselves (Maestre et al. 2009).
Competition usually leads to the spatial segregation of plants, implying that
distributions are more regular than aggregated (Stoll & Bergius 2005; Perry et
al. 2006). However, spatial aggregation is ubiquitous amongst varied plant
systems, especially in harsh environments (Bertness & Callaway 1994; Haase
2001; Perry et al. 2006). Because SGH predicts that facilitation should
dominate in such harsh environments, facilitation is believed to be an
important factor explaining plant aggregation in addition to other ecological
factors (e.g. topography, resource availability and dispersal) (Bertness &
Callaway 1994; Haase 2001; Perry et al. 2006). It has indeed been shown that
facilitation tends to maintain the aggregation of seedling cohorts and
established plants (Bertness & Callaway 1994; Fajardo & McIntire 2011; Jia et
al. 2011; McIntire & Fajardo 2011), but we do not yet know whether this is
generally true for different modes of facilitation and at different stress levels.
Consequently, we do not know how different modes of competition modify
the effects of facilitation on plants and structure their populations and
communities in different environmental contexts (Brooker et al. 2008). To

Chapter 4 - 99 -
address this issue, we implemented different modes of facilitation and
competition in a generic individual-based model based on the
zone-of-influence (ZOI) approach of Weiner et al. (2001). Plant growth and
density-dependent mortality are described according to a growth model as
deriving from “metabolic scaling theory” (MST; Brown et al. 2004; Savage et al.
2010) to provide mechanistic representation of plant response to stress. To
simplify, we restrict ourselves to intraspecific plant interactions. Specifically,
we addressed the following questions at both the plant population and
individual levels: (1) How does the interplay of different modes of competition
and facilitation change spatial pattern formation during self-thinning in
conspecific cohorts that initially have a random or aggregated distribution; and
(2) How do combinations of modes of competition and facilitation alter the
intensity of local plant interactions along a stress gradient?
4.2 Methods
4.2.1 The model
Metabolic scaling theory (MST) predicts quantitative relationships amongst
metabolic processes using empirical measurements and theoretical
assumptions (West et al. 2001; Enquist 2002; Enquist et al. 2009; Savage et al.
2010). We adopted these relationships as the basis of our individual growth
model for plants (see Chapter 2). The model is derived from an energy
conservation equation (Enquist & Niklas 2001; West et al. 2001; Hou et al.
2008) and takes into account respiration and three basic energy-demanding
processes: biomass maintenance, ion transport and biosynthesis (Lambers et
al. 2008). It provides a mechanistic and quantitative basis for linking the
energy used in metabolism of plants under abiotic stress to local interactions
and population dynamics. The growth model is:

- 100 - Chapter 4
dm/dt = am3/4 – bm = am3/4 [ 1 – ( m / M0)1/4 ] (4-1)
where m is total plant biomass, a and b are species-specific constants
(Chapter 2) determined by systematic variation of the in vivo metabolic rate of
different taxa (West et al. 2001), and M0 = (a/b)4 is the asymptotic maximum
size of a plant (calculated for dm/dt = 0). The term am3/4 in equation (4-1)
dominates during early plant growth and provides a good quantitative
description of plant growth (Enquist et al. 2009).
Because stress can be resource-independent or resource-dependent
(Maestre et al. 2009), we assume abiotic stress factors act in one or both of
two ways: by restricting the energy intake rate and burdening biomass
maintenance. This assumption provides a mechanistic basis for representing
the effects of stress on plant performance (Lambers et al. 2008). Accordingly,
the growth model with stress is:
dm/dt = (1 – S) am3/4 [ 1 – ( m / Ms)1/4 ] (4-2)
where S is a dimensionless efficiency factor that indicates the level of stress,
ranging from 0 (no stress) to 1 (extreme stress). Ms = (1–S)4M0 is the
maximum plant biomass achievable under stress.
Our individual-based model (IBM; Grimm & Railsback 2005) is described in
detail in the Supporting Information, following the ODD protocol (Overview,
Design concepts, Details) for describing individual-based models (Grimm et al.
2006; Grimm et al. 2010). In the following, we describe the model’s main
elements. In our IBM, a plant’s circular ZOI (Weiner et al. 2001), A, is the
physical space in which a plant acquires resources and represents the energy
and resources potentially available to the plant. This space is allometrically
related to plant biomass, m, as c0A = m3/4 (Enquist & Niklas 2001), where c0 is
a normalization constant. We represent plant interaction by calculating the
overlapping areas among the plants’ ZOIs (Weiner et al. 2001; Chu et al. 2008,
2009, 2010). Competition and facilitation under abiotic stress are incorporated

Chapter 4 - 101 -
by using dimensionless factors or indexes, fp and fq, respectively. With these
assumptions, equation (4-2) becomes:
dm/dt = fpfqcA[ 1 – ( m / M)1/4 ] (4-3)
where fp is the index of competition, fq refers to the abiotic stress (S) modified
by facilitation (see below), c = ac0 is the initial growth rate in units of biomass
per area and time, and M = (fpfq)4M0 is the maximum biomass achievable
under stress in the presence of competition and facilitation. Modes of
competition among plants can be defined along a continuum from completely
asymmetric competition (largest plants obtain all contested resources) to
completely symmetric competition (resources in areas of overlap are divided
equally among all overlapping individuals, irrespective of their relative sizes;
Schwinning & Weiner 1998). To represent the different modes of competition,
we define the index of competition, fp, as
,1
1
( ) /o
j
pn i i
p no o knk p
j jj
v mf A A A
v m
(4-4)
This index refers to the fraction of resources available in a given area to a plant
i after the loss of potential resources from areas overlapping neighbours with
biomass mj (Schwinning & Weiner 1998). Ano is the area with no overlap from
neighbours, and Ao,k denotes the no areas overlapping nj different neighbours.
The number of overlapping areas, no, can vary due to the position and number
of neighbours (see Appendix A). Parameter p indicates the mode of
competition, ranging from complete symmetry (p = 0) to complete asymmetry
(p approaching infinity). In this article we restrict ourselves to intraspecific
competition and facilitation, and assume, therefore, that the species-specific
weighting constants of competition vi and vj equal 1.
Similarly, we define the index of facilitation modifying abiotic stress, fq, as

- 102 - Chapter 4
,1
1
1 11
(1 ) 1o
j
q qn
i ifo knk q
j jj
S Sf
wmAA
w m
(4-5)
This term is based on SGH, which reflects the facilitative effect of relieving
stress, and is consistent with earlier models (Chu et al. 2008, 2009, 2010). Our
definition includes earlier indices of facilitation as a special case, all of which
only represent symmetric facilitation (Brooker et al. 2008; Chu et al. 2008,
2009, 2010; Jia et al. 2011; see Appendix A). Here, Af indicates the benefits
gained by a plant from all interactive neighbours under abiotic stress (S), which
is calculated as the sum of the areas overlapping ZOIs of neighbour plants.
Index q determines the mode of facilitation among plants, ranging from
complete symmetry (q = 0, algorithmic equivalent to the form used in Chu et al.
2008, 2009) to complete asymmetry (q approaching infinity; Table 1). When
there is no facilitation (Af = 0), equation (4-5) becomes 1–S, which reflects the
effect of abiotic stress. As for competition, the species-specific weighting
constants of facilitation, wi and wj, equal 1 when considering intraspecific
interactions, as we do here.
Equation (4-3) describes how plants grow under local competition, abiotic
stress and facilitation. Equations (4-1), (4-2) and (4-3) are similar to the von
Bertalanffy growth function and other phenomenological growth functions
(Weiner et al. 2001; Chu et al. 2008, 2009, 2010; Jia et al. 2011). However,
these equations are derived from first principles and their parameters and are
directly linked to physical and biological processes.
Because individual plant mortality is proportional to mass-specific metabolic
rate (Brown et al. 2004), we assume that individuals die if their actual growth
rate (realistic metabolic rate) falls below a threshold fraction of their basal
metabolic rate (scaled by current biomass, i.e. 5% of m3/4). Therefore,
individual plants may die due to metabolic inactivation caused by
environmental stress, local competition, senescence (when m approaches M)

Chapter 4 - 103 -
or combinations thereof.
Our model was implemented in NetLogo 4.1.3 (Wilensky 1999). The source
code is available in the online supporting information of the publication
(http://onlinelibrary.wiley.com/doi/10.1111/j.1365-2745.2012.02019.x/suppinfo).
4.2.2 Scenarios and analysis
We investigated 2 modes of facilitation (q = 0: completely symmetric and q = ∞:
completely asymmetric) and 2 modes of competition (p = 0: completely
symmetric and p = ∞: completely asymmetric), at 3 stress levels (S = 0.1, 0.5
and 0.9). Two initial conditions were used: 300 initial plants distributed over
space either with aggregation or randomly (Fig. 4.1). For both aggregated and
random initial location scenarios, all simulations (42 scenarios in total) began
with exactly the same plant locations (i.e. using the same random number
seed), so that differences in results can be ascribed entirely to the interplay
among modes of competition and facilitation at different stress levels. (In
simulations not reported here, we used other initial densities, initial locations,
and interaction combinations to confirm that our general conclusions were not
artefacts of initial conditions.)
Ripley’s K function is widely used to analyse the spatial point pattern of
plants (Ripley 1981; Stoll & Bergius 2005; Perry et al. 2006). Here, we
employed the variance-stabilizing K function, the so-called Ripley’s L function,
to evaluate spatial pattern dynamics. The L function, L(r), characterizes the
point pattern at certain scales (r), with an expected value of zero under the null
hypothesis of complete spatial randomness (CSR). We carried out 499 Monte
Carlo simulations for each scenario to determine the 95% confidence
envelopes of the L function for CSR. Observed L(r) values out of the envelopes
indicate significant aggregation or regularity. Spatial point pattern data were
collected at 6 densities (300, 250, 200, 150, 100 and 50 plants) during the
self-thinning process. All statistical analyses were accomplished using R
2.11.1 (R Development Core Team, 2010).
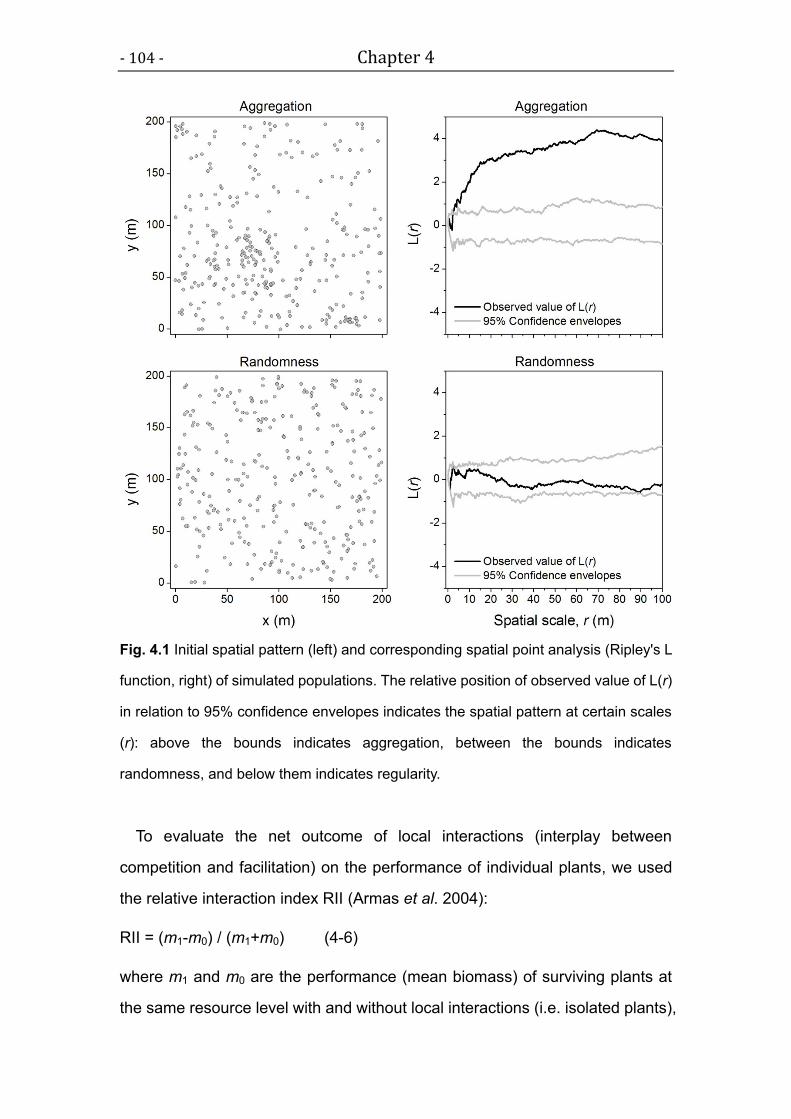
- 104 - Chapter 4
Fig. 4.1 Initial spatial pattern (left) and corresponding spatial point analysis (Ripley's L
function, right) of simulated populations. The relative position of observed value of L(r)
in relation to 95% confidence envelopes indicates the spatial pattern at certain scales
(r): above the bounds indicates aggregation, between the bounds indicates
randomness, and below them indicates regularity.
To evaluate the net outcome of local interactions (interplay between
competition and facilitation) on the performance of individual plants, we used
the relative interaction index RII (Armas et al. 2004):
RII = (m1-m0) / (m1+m0) (4-6)
where m1 and m0 are the performance (mean biomass) of surviving plants at
the same resource level with and without local interactions (i.e. isolated plants),

Chapter 4 - 105 -
respectively. Values of RII from -1 to 1 indicate the net outcome of interactions
as negative (from -1 to 0), neutral (equal to 0) and positive (from 0 to 1). To
estimate m0, we using equation (4-2) for plant growth in all scenarios.
4.3 Results
The interplay between competition and facilitation strongly influenced spatial
pattern formation in the plant population along the stress gradient. Different
modes of competition and facilitation also led to distinct spatial patterns. With
aggregated initial locations (Fig. 4.1), facilitation was vital for maintaining
aggregated patterns if competition was completely symmetric (CCS, p = 0) (Fig.
4.2). Without facilitation (NF), aggregation was maintained only until the
number of surviving plants decreased to 200 (Fig. 4.2a−c). With symmetric
facilitation (FCS, q = 0), plant aggregation patterns can be maintained (Fig.
4.2d−f) even at quite low density (100 plants), depending on the level of stress.
Aggregated patterns are particularly robust at high stress levels, which is
consistent with the predictions of SGH.
In contrast, with asymmetric facilitation (FCA, q = ∞), aggregating (Figs 4.2g
and 4.2h) can be maintained only at mild or intermediate stress levels (S = 0.1
and 0.5). Under harsh conditions (S = 0.9), aggregation disappears early in the
self-thinning process (Fig. 4.2i), a result that deviates from predictions of SGH.
However, modes of facilitation had little effect on maintaining aggregation
when competition was completely asymmetric (CCA, p = ∞; Fig. S2).
The importance of facilitation for creating aggregation became more obvious
when the initial pattern was random (Fig. 4.3). Under completely symmetric
competition without facilitation and under benign condition, there was some
slight spatial aggregation (values of L(r) were very close to the upper boundary
of the 95% confidence envelopes defining lack of aggregation) at very small
scales (Fig. 4.3a), but this aggregation instantly disappeared when stress

- 106 - Chapter 4
increased (Fig 4.3b and 4.3c). In contrast, more pronounced aggregation
patterns emerged in the presence of facilitation: symmetric facilitation
consistently forced aggregation under harsh conditions (Fig. 4.3d−f);
asymmetric facilitation led to aggregation only when stress was milder (Fig.
4.3g−i). Nevertheless, asymmetric competition can largely override the effects
of facilitation on spatial pattern formation at all stress levels, leading to
non-aggregative (even regular) spatial distributions (Fig. S3). This is in
agreement with empirical findings (Stoll & Bergius 2005).
Fig. 4.2 Spatial dynamics (black, aggregation; grey, randomness; white, regularity)
during self-thinning, with aggregated initial locations and completely symmetric
competition (CCS, p = 0) in the absence (NF) or presence of facilitation (FCS,
completely symmetric facilitation, q = 0; FCA, completely asymmetric facilitation, q = ∞)
at different levels of abiotic stress (S).
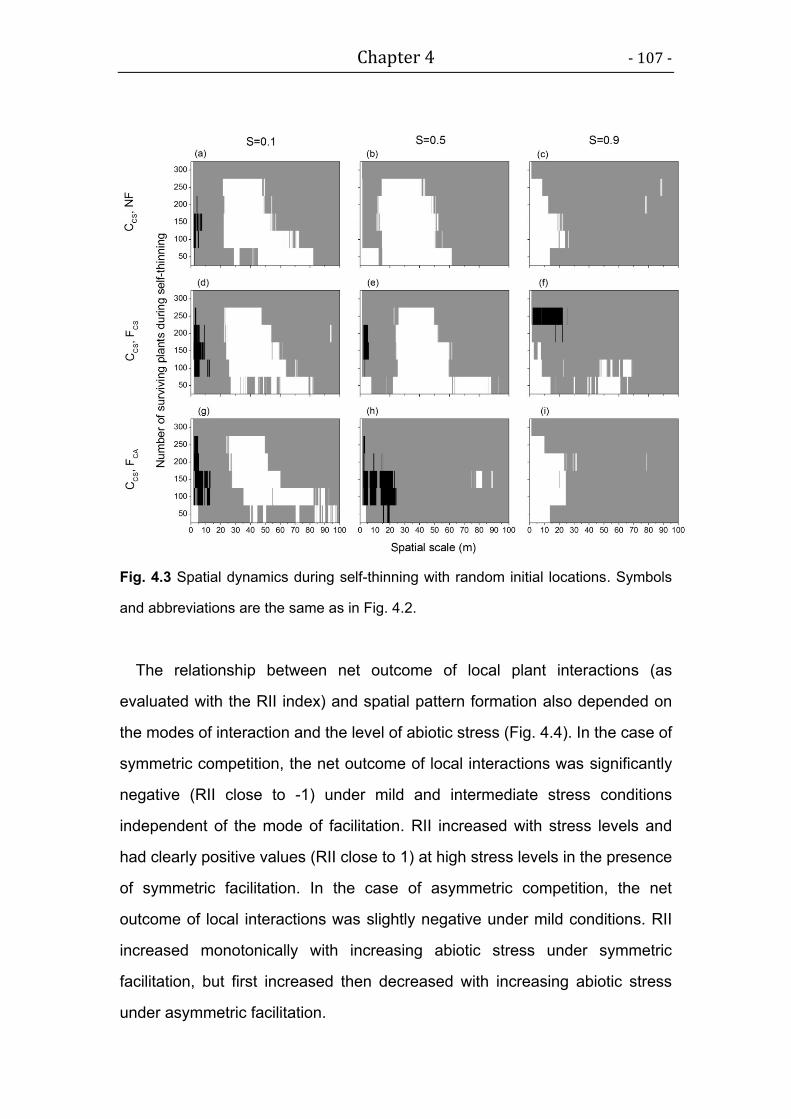
Chapter 4 - 107 -
Fig. 4.3 Spatial dynamics during self-thinning with random initial locations. Symbols
and abbreviations are the same as in Fig. 4.2.
The relationship between net outcome of local plant interactions (as
evaluated with the RII index) and spatial pattern formation also depended on
the modes of interaction and the level of abiotic stress (Fig. 4.4). In the case of
symmetric competition, the net outcome of local interactions was significantly
negative (RII close to -1) under mild and intermediate stress conditions
independent of the mode of facilitation. RII increased with stress levels and
had clearly positive values (RII close to 1) at high stress levels in the presence
of symmetric facilitation. In the case of asymmetric competition, the net
outcome of local interactions was slightly negative under mild conditions. RII
increased monotonically with increasing abiotic stress under symmetric
facilitation, but first increased then decreased with increasing abiotic stress
under asymmetric facilitation.

- 108 - Chapter 4
Fig. 4.4 Relative interaction intensity (RII) during self-thinning under different modes
of competition (CCS, completely symmetric competition, p = 0; CCA, completely
asymmetric competition, p = ∞) in the absence (NF) or presence of facilitation (FCS,
completely symmetric facilitation, q = 0; FCA, completely asymmetric facilitation, q = ∞)
at different levels of abiotic stress (S), and with aggregated initial locations.
Facilitation sometimes resulted in spatial aggregation (Figs 4.2 and 4.3)
even though the net outcome (RII) of local interaction was negative. This
spatial patterning also depended on the mode of competition. Such results
indicate that even though facilitation was responsible for resulting plant
aggregation, the net outcome of local interaction was not always positive under
harsh conditions. In other words, modes of interaction can have different
effects on different population characteristics, i.e. here individual growth (RII)
and spatial pattern.

Chapter 4 - 109 -
Symmetric facilitation not only mitigated abiotic stress but also delayed the
onset of mortality in the case of symmetric competition, at all stress levels
(Duncan’s test, P < 0.05). Symmetric facilitation delayed the self-thinning
process, as indicated by the increased survival over time (Fig. 4.5).
Asymmetric facilitation had the same effect only at intermediate stress levels,
not at harsh conditions (Duncan’s test, P > 0.05). However, in the case of
asymmetric competition, the effects of facilitation were relatively weak.
Fig. 4.5 Temporal dynamics of density-dependent mortality under different modes of
competition (CCS, complete symmetric competition, p = 0; CCA, complete asymmetric
competition, p = ∞) in the absence (NF) or presence of facilitation (FCS, completely
symmetric facilitation, q = 0; FCA, completely asymmetric facilitation, q = ∞) at different
levels of abiotic stress (S), and for both initial location scenarios.

- 110 - Chapter 4
4.4 Discussion
We have introduced the new concept of modes of facilitation, i.e. symmetric
versus asymmetric facilitation, and have used an individual-based model to
explore how these facilitation modes affect plant populations differently.
Specifically, we explored how the interplay between different modes of
competition and facilitation changes spatial pattern formation during
self-thinning in populations that start with either random or aggregated
distributions, and how combinations of competition and facilitation modes alter
the intensity of local plant interactions along a stress gradient.
Our main finding was that the spatial aggregation of plants can be attributed
to different modes of facilitation if competition is symmetric rather than
asymmetric, whereas non-aggregative (regular) patterns indicate strong
asymmetric competition driving density-dependent mortality (Stoll & Bergius
2005). Facilitation by itself can play an important role in promoting plant
aggregation independent of other ecological factors ignored in our scenarios
(e.g. seed dispersal, recruitment, and environmental heterogeneity); moreover,
different modes of facilitation led to different spatial and temporal patterns.
In our simulations of harsh conditions, plants had highest biomass
accumulation and survival under the combination of symmetric competition
and facilitation. This result emerged from growing in association with
neighbours due to facilitation and the increased importance of facilitation at
higher stress levels as predicted by SGH. McIntire & Fajardo (2011) found that
spatial aggregation and symmetric interactions in Nothofagus pumilio are
essential for the species’ success under harsh conditions. Seedlings growing
in clustered cohorts facilitate each other and have higher survival rates than
isolated individuals under stress of wind desiccation. With seedlings growing in
clusters, natural grafts (physiological and physical merging of stem, branch or
root) occur in later growth stages, and multi–stemmed trees survive better than
single-stemmed trees. The mode of facilitation and competition among grafted

Chapter 4 - 111 -
plants is considered to be symmetric because the transport of resources and
assimilates is bidirectional (Silvertown & Charlesworth 2001; Lambers et al.
2008). The combination of symmetric facilitation and competition in these tree
clusters decreases the impact of stress and the onset of competition among
them (Fajardo & McIntire 2010; Tarroux & DesRochers 2011). Therefore,
mortality is lower within clusters and leads to aggregation, an outcome we also
found in our model.
Because we based our model of facilitation on SGH, a monotonic increase
in the importance of facilitation with increasing stress should be expected due
to the assumptions underlying equation (4-5). Surprisingly, this effect was
outweighed by asymmetric facilitation. Our study indicated that with symmetric
competition, asymmetric facilitation first promotes plant aggregation under mild
and intermediate stress conditions, but then brings about spatial
disaggregation at the more stressful end of the gradient (Fig. 4.2g−i).
Moreover, asymmetric facilitation was found to delay the self-thinning process
(indicated by increased survival over time) only under intermediate stress
conditions (Fig. 4.5). This is because under very stressful conditions, plants
are very sensitive to physical stress and local competition. Competition will
thus aggravate the mortality of those individuals that were disadvantaged by
asymmetric facilitation. Asymmetric facilitation can thus promote plant survival
and aggregation only if the environment is less harsh, as we observed in our
model.
Our results regarding asymmetric facilitation are consistent with recent
empirical findings that the SGH is not fully supported by observations (Maestre
et al. 2009). Reduced positive effects on plant survival have been observed in
arid areas at high stress levels (Tielbörger & Kadmon 2000; Maestre et al.
2005; Maestre et al. 2009). A switch from negative to positive and back to
negative effects on plant survival was found in a semi-arid steppe along a
gradient of decreasing rainfall (Maestre & Cortina 2004). Similarly, Tielbörger &
Kadmon (2000) found that the effect of desert shrubs on abundance and

- 112 - Chapter 4
reproductive success of understory annuals shifted from positive to neutral and
to negative with decreasing annual rainfall.
Our findings suggest a mechanism explaining such exceptions to the SGH’s
prediction of monotonic increasing net positive effect with stress: the nature of
abiotic stress factors is also important (Maestre et al. 2009). Abiotic stress
factors not only alter the mode of competition (Schwinning & Weiner 1998;
Berger et al. 2008), but also the mode of facilitation (Maestre et al. 2009). It is
therefore important to ask how symmetric and asymmetric facilitation are
interrelated with different kinds of stress factors. In the light of the new concept
presented here, it should be expected that if the effect of stress is
symmetrically mitigated by other individuals, symmetric facilitation should
usually be prevalent (detected via plant aggregation or positive RII in our
simulation). Using an individual-based model, Chu et al. (2009) found that
symmetric facilitation (mutualism) among plants can increase plant biomass
and size inequality in conspecific populations. Their results are consistent with
empirical findings for an annual species (Elymus nutans) in alpine meadows
(although they worked with clonal ramets, which probably confounded
physiological integration with positive interactions among individuals; Fajardo
& McIntire 2011). The mode of facilitation in their experiment was probably
symmetric because frost is the most important stress factor in their research
area (Chu et al. 2009): all individuals endure the same degree of low
temperature stress, which is not asymmetrically mitigated by other individuals.
In contrast, when the stress factor is “pre-mitigable” and/or directional (e.g.
higher ultraviolet radiation due to ozonosphere depletion or direct damage
caused by wind), facilitation should be more asymmetric (Fig. S1). For
example, in a conspecific plant population under strong ultraviolet radiation,
taller plants (benefactor) will suffer most from radiation stress but their crowns
can reduce stress in the understory microenvironment. As a result, smaller
plants (beneficiary) can receive disproportionately more benefit from their
neighbouring taller plants for maintenance and growth, and therefore survive

Chapter 4 - 113 -
longer. Asymmetric facilitation should thus reduce size inequality in plant
populations, which is in contrast to symmetric facilitation that can increase size
inequality (Chu et al. 2009). This effect of asymmetric facilitation is observed in
our model and consistent with empirical studies (Zhang et al. 2012). To the
best of our knowledge, effects of different modes of facilitation have not been
addressed systematically in empirical experiments yet, so we suggest this
topic for future research.
To answer our second research question about the effect of different
combinations of competition and facilitation on local interactions, we employed
the index RII to measure the direction and magnitude of local interactions
along a stress gradient. We found a monotonically increasing strength of
facilitation relative to competition for symmetric facilitation but not for
asymmetric facilitation. Negative interaction was dominant under mild and less
harsh conditions.
Our results indicate that even though facilitation can lead to plant
aggregation at the population level, the net outcome and intensity of local
interactions at the individual plant level is not necessarily positive. In particular,
we found that for some scenarios (e.g. CCS, FCA, S = 0.5), competition is the
dominant process when assessing plant growth (the net outcome is negative
as indicated by RII; Fig. 4.4), whereas the corresponding spatial aggregation
and greater survival probability indicate a dominance of facilitation (Figs 4.2,
4.3 and 4.5). In other words, the intensity of interaction (as the net outcome
between competition and facilitation) is insufficient to express the relative
importance of competition and facilitation on structuring plant systems
(Brooker et al. 2005). Furthermore, our findings imply that facilitation may be
more common than generally believed, but its important role in structuring
populations and communities can be hidden simply because it is hard to detect.
Thus, establishing whether competition or facilitation is the dominant process
in plant populations or communities can be difficult, because competition and
facilitation act simultaneously (Callaway 2007) but can affect different aspects

- 114 - Chapter 4
of plant populations or communities.
Nevertheless, the importance of facilitation has been clearly detected in arid,
alpine and arctic habitats, which are assumed highly sensitive to global change
(Callaway et al. 2002; Brooker et al. 2008). It is therefore crucial to better
understand both the mode of facilitation and nature of stress factors, because
the facilitative effect from key species is essential for system diversity and
stability in such conditions (Brooker et al. 2008; Vellend 2008). Although we
focused here on intraspecific interactions, our approach can easily be used to
analyse communities. Our conclusions may thus also be relevant for plant
communities.
To conclude, our study is the first to quantitatively define different modes of
facilitation, and it is also the first attempt to integrate different modes of
facilitation with different modes of competition into the SGH. We showed that
facilitation can have an important influence on population structure. Moreover,
different modes of facilitation and competition can affect different aspects of
plant populations and communities, implying context-dependent outcomes and
consequences. Explicit consideration of modes and mechanisms of interaction
(both facilitation and competition) and the nature of stress factors may help us
extend the SGH framework and foster research on facilitation in ecology
(Brooker et al. 2008; Maestre et al. 2009).
Acknowledgements
We thank Anne Fetsch, Jacob Weiner, Franka Huth, Rui-Chang Zhang and
Ming Yue for their kind help. We thank Steve Railsback for language editing.
We thank three anonymous reviewers and the handling editor for valuable
comments and suggestions. The first author was supported by Helmholtz
Impulse and Networking Fund through Helmholtz Interdisciplinary Graduate
School for Environmental Research (HIGRADE).

Chapter 4 - 115 -
References
Armas, C., Ordiales, R. & Pugnaire, F.I. (2004). Measuring plant interactions: a new
comparative index. Ecology, 85, 2682-2686.
Berger, U., Piou, C., Schiffers, K. & Grimm, V. (2008). Competition among plants:
concepts, individual-based modelling approaches, and a proposal for a future
research strategy. Perspectives in Plant Ecology, Evolution and Systematics, 9,
121-135.
Bertness, M.D. & Callaway, R. (1994). Positive interactions in communities. Trends in
Ecology & Evolution, 9, 191-193.
Bronstein, J.L. (2009). The evolution of facilitation and mutualism. Journal of Ecology, 97,
1160-1170.
Brooker, R.W. & Callaway, R.M. (2009). Facilitation in the conceptual melting pot. Journal
of Ecology, 97, 1117-1120.
Brooker, R., Kikvidze, Z., Pugnaire, F.I., Callaway, R.M., Choler, P., Lortie, C.J. &
Michalet, R. (2005) The importance of importance. Oikos, 109, 63-70.
Brooker, R.W., Maestre, F.T., Callaway, R.M., Lortie, C.L., Cavieres, L.A., Kunstler, G., et
al. (2008). Facilitation in plant communities: the past, the present, and the future.
Journal of Ecology, 96, 18-34.
Brown, J.H., Gillooly, J.F., Allen, A.P., Savage, V.M. & West, G.B. (2004). Toward a
metabolic theory of ecology. Ecology, 85, 1771-1789.
Callaway, R.M. (2007) Positive Interactions and Interdependence in Plant Communities.
Springer, Dordrecht.
Callaway, R.M., Brooker, R.W., Choler, P., Kikvidze, Z., Lortie, C.J., Michalet, R., et al.
(2002). Positive interactions among alpine plants increase with stress. Nature,
417, 844-848.
Cavieres, L.A. & Badano, E.I. (2009). Do facilitative interactions increase species richness
at the entire community level? Journal of Ecology, 97, 1181-1191.
Chu, C.J., Maestre, F.T., Xiao, S., Weiner, J., Wang, Y.S., Duan, Z.H., et al. (2008).
Balance between facilitation and resource competition determines
biomass–density relationships in plant populations. Ecology Letters, 11,
1189-1197.
Chu, C.J., Weiner, J., Maestre, F.T., Xiao, S., Wang, Y.S., Li, Q., et al. (2009). Positive
interactions can increase size inequality in plant populations. Journal of Ecology,
97, 1401-1407.
Chu, C.J., Weiner, J., Maestre, F.T., Wang, Y.S., Morris, C., Xiao, S., et al. (2010). Effects
of positive interactions, size symmetry of competition and abiotic stress on
self-thinning in simulated plant populations. Annals of Botany, 106, 647-652.

- 116 - Chapter 4
Enquist, B.J. (2002). Universal scaling in tree and vascular plant allometry: toward a
general quantitative theory linking plant form and function from cells to
ecosystems. Tree Physiology, 22, 1045-1064.
Enquist, B.J. & Niklas, K.J. (2001). Invariant scaling relations across tree-dominated
communities. Nature, 410, 655-660.
Enquist, B.J., West, G.B. & Brown, J.H. (2009). Extensions and evaluations of a general
quantitative theory of forest structure and dynamics. Proceedings of the National
Academy of Sciences, 106, 7046.
Eränen, J.K. & Kozlov, M.V. (2008). Increasing intraspecific facilitation in exposed
environments: consistent results from mountain birch populations in two subarctic
stress gradients. Oikos, 117, 1569–1577.
Fajardo, A. & McIntire, E.J. (2010). Merged trees in second-growth, fire-origin forests in
Patagonia, Chile: positive spatial association patterns and their ecological
implications. American journal of botany, 97, 1424-1430.
Fajardo, A. & McIntire, E.J. (2011). Under strong niche overlap conspecifics do not
compete but help each other to survive: facilitation at the intraspecific level.
Journal of Ecology, 99, 642-650.
Graham, B.F. & Bormann, F.H. (1966). Natural root grafts. The Botanical Review, 32,
255-292.
Grimm, V., Berger, U., Bastiansen, F., Eliassen, S., Ginot, V., Giske, J., et al. (2006). A
standard protocol for describing individual-based and agent-based models.
Ecological modelling, 198, 115-126.
Grimm, V., Berger, U., DeAngelis, D.L., Polhill, J.G., Giske, J. & Railsback, S.F. (2010).
The ODD protocol: A review and first update. Ecological modelling, 221,
2760-2768.
Grimm, V. & Railsback, S.F. (2005). Individual-based modeling and ecology. Princeton
University Press, Princeton and Oxford.
Haase, P. (2001). Can isotropy vs. anisotropy in the spatial association of plant species
reveal physical vs. biotic facilitation? Journal of Vegetation Science, 12, 127-136.
Hou, C., Zuo, W., Moses, M.E., Woodruff, W.H., Brown, J.H. & West, G.B. (2008). Energy
uptake and allocation during ontogeny. Science, 322, 736.
Jia, X., Dai, X.F., Shen, Z.X., Zhang, J.Y. & Wang, G.X. (2011). Facilitation can maintain
clustered spatial pattern of plant populations during density-dependent mortality:
insights from a zone-of-influence model. Oikos, 120, 472-480.
Kikvidze, Z., Pugnaire, F.I., Brooker, R.W., Choler, P., Lortie, C.J., Michalet, R., et al.
(2005). Linking patterns and processes in alpine plant communities: a global study.
Ecology, 86, 1395-1400.
Lambers, H., Chapin, F.S. & Pons, T.L. (2008). Plant physiological ecology. 2nd edition.
Springer Verlag, New York.

Chapter 4 - 117 -
Maestre, F.T., Callaway, R.M., Valladares, F. & Lortie, C.J. (2009). Refining the
stress-gradient hypothesis for competition and facilitation in plant communities.
Journal of Ecology, 97, 199-205.
Maestre, F.T. & Cortina, J. (2004) Do positive interactions increase with abiotic stress? A
test from a semi-arid steppe. Proceedings of the Royal Society of London, Series
B, 271, S331-S333.
Maestre, F.T., Valladares, F. & Reynolds, J.F. (2005). Is the change of plant–plant
interactions with abiotic stress predictable? A meta-analysis of field results in arid
environments. Journal of Ecology, 93, 748-757.
McIntire, E.J. & Fajardo, A. (2011). Facilitation within Species: A Possible Origin of
Group-Selected Superorganisms. The American Naturalist, 178, 88-97.
Pakeman, R.J., Pugnaire, F.I., Michalet, R., Lortie, C.J., Schiffers, K., Maestre, F.T., et al.
(2009). Is the cask of facilitation ready for bottling? A symposium on the
connectivity and future directions of positive plant interactions. Biology Letters, 5,
577-579.
Perry, G.L., Miller, B.P. & Enright, N.J. (2006). A comparison of methods for the statistical
analysis of spatial point patterns in plant ecology. Plant Ecology, 187, 59-82.
R Development Core Team (2010) R: a language and environment for statistical
computing. R Foundation for Statistical Computing, URL http://www.R-project.org.
Last accessed 17 February 2012.
Reinhart, K.O., Maestre, F.T. & Callaway, R.M. (2006) Facilitation and inhibition of
seedlings of an invasive tree (Acer platanoides) by different tree species in a
mountain ecosystem. Biological Invasions, 8, 231–240.
Ripley, B.D. (1981) Spatial Statistics. John Wiley and Sons, New York.
Savage, V.M., Bentley, L.P., Enquist, B.J., Sperry, J.S., Smith, D.D., Reich, P.B., et al.
(2010). Hydraulic trade-offs and space filling enable better predictions of vascular
structure and function in plants. Proceedings of the National Academy of Sciences,
107, 22722.
Schwinning, S. & Weiner, J. (1998). Mechanisms determining the degree of size
asymmetry in competition among plants. Oecologia, 113, 447-455.
Silvertown, J.W. & Charlesworth, D. (2001) Introduction to plant population biology.
Blackwell Publishing Limited, Oxford, UK.
Stoll, P. & Bergius, E. (2005). Pattern and process: competition causes regular spacing of
individuals within plant populations. Journal of Ecology, 93, 395-403.
Sthultz C.M, Gehring C.A, & Whitham T.G. 2007. Shifts from competition to facilitation
between a foundation tree and a pioneer shrub across spatial and temporal scales
in a semi-arid woodland. New Phytologist, 173, 135-145.
Tarroux, E. & DesRochers, A. (2011). Effect of natural root grafting on growth response of
jack pine (Pinus banksiana; Pinaceae). American Journal of botany, 98, 967-974.

- 118 - Chapter 4
Tielbörger, K. & Kadmon, R. (2000). Temporal environmental variation tips the balance
between facilitation and interference in desert plants. Ecology, 81, 1544-1553.
Vellend, M. (2008). Effects of diversity on diversity: consequences of competition and
facilitation. Oikos, 117,1075-1085.
Weiner, J., Stoll, P., Muller-Landau, H. & Jasentuliyana, A. (2001). The effects of density,
spatial pattern, and competitive symmetry on size variation in simulated plant
populations. The American Naturalist, 158, 438-450.
West, G.B., Brown, J.H. & Enquist, B.J. (2001). A general model for ontogenetic growth.
Nature, 413, 628-631.
Wilensky, U. (1999). NetLogo. Center for Connected Learning and Computer-Based
Modeling, Northwestern University. Available at:
http://ccl.northwestern.edu/netlogo/. Last accessed 17 February 2012.
Zhang, R.C., Lin, Y., Yue, M., Li, Q., Zhang, X.F., Liu, X., Chi, H., Chai, Y.F. & Wang, M.
(2012) Effects of Ultraviolet-B Irradiance on Intraspecific Competition and
Facilitation of Plants: Self-Thinning, Size Inequality, and Phenotypic Plasticity.
PLoS ONE, 7, e50822.

Chapter 4 - 119 -
Chapter 4 – Appendix A
ODD protocol – model description of pi model with competition
and facilitation
The following model description follows the ODD protocol (Overview, Design
concepts, Details) for describing individual- and agent-based models (Grimm
et al. 2006; Grimm et al. 2010),
Purpose
The aim of this model is to evaluate the multiple effects of the modes of
competition and facilitation under abiotic stress on regulating plant population
dynamics, specifically on spatial pattern formation induced by
density-dependent mortality. In particular, we test whether different modes of
facilitations at individual plant level can result in the spatial aggregation of the
population. The model does not represent specific species, but generic ones.
Entities, state variables, and scales
The entities in the model are plants and square habitat units, or patches (Table
A1). Plants are characterized by the following state variables: initial growth rate,
initial biomass, maximum biomass (asymptotic biomass), current biomass and
their position, i.e. coordinates of the stem. Each individual plant has its own
circular zone-of-influence (ZOI). The ZOI stands for the physical space
occupied by a plant, and represents the energy and resources potentially
available to this plant, which is allometrically related to its body mass.
Neighbouring plants only compete for the resources when their ZOIs are
overlapping.
In order to make the spatial calculations of resource competition easier,
ZOIs are projected onto a grid of patches. To avoid edge effects, we use a
torus world with a size of 200 × 200 patches (Grimm & Railsback 2005). Each
patch represents 1 m2 or 1 cm2 for woody- and non-woody plants, respectively.
The state of each patch is characterized by its resource availability. We use a
homogeneous environment here as all patches have the same, and constant,
degree of abiotic stress. One time step in the model represents approximately
one year for woody plants and one day for non-woody plants.

- 120 - Chapter 4
Table A1. State variables and initialization in the individual-based model. Actual
values are drawn from the given intervals to introduce a certain degree of
heterogeneity among individuals.
Variable Description Initial Value [unit]
(woody/non-woody)
Plants
c Initial growth rate 1 ± 0.1 [kg m-2 time step-1] /
[mg cm-2 time step-1]
m0 Initial body mass 2 ± 0.2 [kg] / [mg]
M0 Maximal biomass 2×106 ± 2×105 [kg] / [mg]
m Current biomass [kg] / [mg]
A Zone of influence [m-2] / [cm-2]
Patches
S Abiotic stress level [0, 1]
Initialization
Mortality Threshold of death 5% of m3/4
Density Number of plants 300 [ha-1] / [m-2]
Random seed To generate random number 123456789
(Aggregation) Number of cohort 6 (50 individuals / cohort)
(Aggregation) Cluster scale (diameter) 50% of the world size
Process overview and scheduling
After initialization, all individual plants with a given density are randomly
distributed in the world. The processes of local competition, facilitation, growth
and mortality of each plant are fulfilled within each time step. In each step,
individual plants first sense the environment qualities of patches within their
ZOIs, the area (radius) of an individual plant’s ZOI is determined by its current
biomass. When their ZOIs are overlapping, individuals compete and facilitate
within the overlapping area. Thus, the overlapping area reflecting resources is
divided according to the mode of competition. At the same time, under abiotic
stress in the presence of facilitation, the overlapping area reflecting the
ameliorated habitat by neighbours is also divided according to the mode of
facilitation. Considering the outcome of the interaction process, all individual
plants grow according to the growth function. Plants with growth rates falling
below a threshold die and are removed immediately. The state variables of the

Chapter 4 - 121 -
plants are synchronously updated within the subroutines, i.e. changes to state
variables are updated only after all individuals have been processed (Grimm &
Railsback 2005).
Design concepts
Basic principles: From “Metabolic Scaling Theory”, we derived a general
ontogenetic growth model for individual plants. We combine this model, via the
ZOI approach, with the effects of different modes of competition and facilitation
under abiotic stress.
Emergence: All features observed at the population level, e.g. mass-density
relationship or self-thinning trajectories (i.e. size distribution and spatial
distribution, respectively), population size inequality and spatial pattern, are
emerged from local interactions among plants under abiotic stress of their
environment.
Interaction: Individual plants interact via competition for resources and
facilitation from neighbours in the overlapping area of their ZOIs.
Stochasticity: Initial growth rate, initial biomass, maximum biomass and
initial position of plants are randomly taken from the intervals given in Table 1.
This introduces a certain level of heterogeneity among individual
characteristics to take into account that real plants are never exactly identical.
Observation: Spatial point patterns of plants, population size, biomass of
each plant, and mean biomass of all living plants are the main observations.
Initialization
If the initial spatial pattern is aggregation, we randomly choose several patches
(determined by the number of cohorts) as centers and to transplant groups of
individual plants (initial density / number of cohorts) among the centers within a
certain cluster diameter. In the case of randomness, individual plants are
randomly distributed according to the chosen initial density. Resources and
abiotic stress are spatially and temporally constant. Each plant has an initial
biomass (m0), maximal biomass (M0) and initial growth rate (c) drawn from
truncated normal distributions with average and intervals given in Table A1.
Input
After initialization, the model does not include any external inputs, i.e. the
abiotic environment is constant.
Submodels
Plant growth

- 122 - Chapter 4
In our individual-based model the plant’s ZOI stands for the physical space
occupied by a plant and represents the energy and resources potentially
available to this plant. This space is allometrically related to the plant’s body
mass, m, as c0A=m3/4 (Enquist & Niklas 2001), where c0 is a normalization
constant. Accordingly, Eqn (4-1) can be rewritten as
dm/dt = cA [ 1 – ( m / M0)1/4 ] (A1)
and with abiotic stress, it becomes
dm/dt = (1 – S) cA [ 1 – ( m / Ms)1/4 ] (A2)
where c = ac0, is the initial growth rates in units of mass per area and time
interval. For simplicity, we choose c = 1 ± 0.1 in our model. In addition, we
simulate the model with different c values. As expected, the results from
different values were qualitatively similar (consist with our findings).
Competition and facilitation under abiotic stress
Competition and facilitation are incorporated by using dimensionless factors or
indexes, fp and fq respectively. With the above assumptions, equation (A2)
becomes:
dm/dt = fpfqcA[ 1 – ( m / M)1/4 ] (A3)
where M = (fpfq)4M0 is the maximum achievable biomass under stress with
competition and facilitation.
As for competition, the modes of resource-mediated competition among
plants can be located somewhere along a continuum between completely
asymmetric competition (largest plants obtain all the contested resources) and
completely symmetric competition (resource uptake is equal for all plants,
independent of their relative sizes; Schwinning & Weiner 1998). To represent
different modes of competition explicitly, we describe the competitive index fp
as
,1
1
( ) /o
j
pn i i
p no o knk p
j jj
v mf A A A
v m
(A4)
This factor thus refers to the fraction of resources available in the area
which plant i could obtain after a loss of potential resources due to areas
overlapped by neighbours of sizes mj (Schwinning and Weiner 1998). Ano is
the area not overlapping with neighbours, Ao,k denotes the no areas
overlapping with nj different neighbours. Parameter p determines the mode of

Chapter 4 - 123 -
competition, ranging from complete symmetry (p=0) to complete asymmetry (p
approaching infinity; for details and examples see Figure A2). In this research,
we restrict ourselves to intraspecific facilitation and competition, and assume
therefore the species-specific weighting constant of competition vi and vj equal
1 here, as conspecific case.
Simultaneously, assuming the effect of facilitation is additive (Molofsky
2001; Molofsky & Bever 2002; Chu et al. 2008, 2009), we define the effect of
different modes of facilitation, fq, as
,1
1
1 11
(1 ) 1o
j
q qn
i ifo knk q
j jj
S Sf
wmAA
w m
(A5)
This factor is based on SGH, which reflects the facilitative effect of
relieving stress and is consist with other model (Chu et al. 2008, 2009, 2010;
Jia et al. 2011; for details and examples see Figure A2). To keep things simple,
we follow the assumption of Chu et al. (2008, 2009) that the ZOI, A, is the
same for competition and facilitation (they could be different). Where Af refers
to the benefit gained by plant from all interactive neighbours, and is calculated
as the sum of the areas (ZOIs) overlapped with neighbour plants. The index q
determines the mode of facilitation among plants, ranging from complete
symmetry (q = 0, algorithmic equivalent to the form used in Chu et al. 2008;
2009) to complete asymmetry (q approaching infinity; see Table 1 for the
complete form and definitions). Since we investigate the monopopulation here,
to simplify, we assume the species-specific weighting constant w equals 1 here
as the conspecific case, so the facilitation is size dependent. When there is no
facilitation (Af =0), equation (A5) becomes 1–S, which reflects the effect of
abiotic stress.
Figure A2. An example of calculating the interactive indexes (equations A4
and A5) with different modes of competition and facilitation by dividing plants’

- 124 - Chapter 4
ZOI (zone-of-influence). Three plants with sizes m1, m2 and m3 are interacting
in this example. For plant 1, its ZOI (A) was divided into four parts: Ano, the
area not overlapping with the other two plants; Ao,1, the area overlapping with
plant 2; Ao,2, the area overlapping with plants 2 and 3; Ao,3, the area
overlapping with plant 3.
Then the actual area that plant 1 can take from Ao,1 is
1 1,1 ,12
1 21
p p
o o p pp
jj
m mA A
m mm
For Ao,2,
1 1,2 ,23
1 2 31
p p
o o p p pp
jj
m mA A
m m mm
And for Ao,3,
1 1,3 ,32
1 31
p p
o o p pp
jj
m mA A
m mm
therefore, the competitive index for plant 1 is:
where 3/4
1 0/A m c . For plant 1, the benefit received from neighbours is
and the facilitative index for plant 1 under abiotic stress is:
In total, equation (A3) clearly shows how a plant’s growth rate is jointly
determined by abiotic stress, S, competition, fp, and facilitation fq. This also
implies that a plant’s final size is usually smaller than its asymptotic maximum
size (M0) during environmental stress and local competition, but can increase
by the beneficial effects of neighbour plants via the amelioration of habitat.
Mortality
An individual’s mortality rate is proportional to its mass-specific metabolism (as
current total metabolic rate divide by body mass; Brown et al. 2004). Based on

Chapter 4 - 125 -
this, we assume that individuals die if their actual growth rate (dm/dt, realistic
metabolic rate) falls below a threshold fraction of their basal metabolic rate
(allometrically scaled with body mass), i.e. 5% of m3/4. Therefore, individual
plants may die due to metabolic inactivation driven by abiotic stress,
competition, senescence (when m approaches M) or combinations thereof.
This provides a more realistic representation of relevant ecological process
than in previous models (Stoll et al. 2002; Chu et al. 2009, 2010; Jia et al.
2011).
References
Brown, J.H., Gillooly, J.F., Allen, A.P., Savage, V.M. & West, G.B. (2004). Toward a
metabolic theory of ecology. Ecology, 85, 1771-1789.
Chu, C.J., Maestre, F.T., Xiao, S., Weiner, J., Wang, Y.S., Duan, Z.H. et al. (2008).
Balance between facilitation and resource competition determines
biomass–density relationships in plant populations. Ecol. Lett., 11, 1189-1197.
Chu, C.J., Weiner, J., Maestre, FT, Xiao, S., Wang, Y.S., Li, Q., Yuan, J.L., Zhao, L.Q.,
Ren, Z.W. and Wang, G.(2009), Positive interactions can increase size inequality
in plant populations. J. Ecol., 97, 1401–1407
Chu, C.J., Weiner, J., Maestre, F.T., Wang, Y.S., Morris, C., Xiao, S. et al. (2010). Effects
of positive interactions, size symmetry of competition and abiotic stress on
self-thinning in simulated plant populations. Ann. Bot., 106, 647-652.
Enquist, B.J. (2002). Universal scaling in tree and vascular plant allometry: toward a
general quantitative theory linking plant form and function from cells to
ecosystems. Tree Physiol., 22, 1045-1064.
Enquist, B.J. & Niklas, K.J. (2001). Invariant scaling relations across tree-dominated
communities. Nature, 410, 655-660.
Enquist, B.J., West, G.B. & Brown, J.H. (2009). Extensions and evaluations of a general
quantitative theory of forest structure and dynamics. Proc. Natl Acad. Sci. USA.,
106, 7046-7051.
Grimm, V., Berger, U., Bastiansen, F., Eliassen, S., Ginot, V., Giske, J. et al. (2006). A
standard protocol for describing individual-based and agent-based models. Ecol.
Model., 198, 115-126.
Grimm, V., Berger, U., DeAngelis, D.L., Polhill, J.G., Giske, J. & Railsback, S.F. (2010).
The ODD protocol: A review and first update. Ecol. Model., 221, 2760-2768.
Grimm, V. & Railsback, S.F. (2005). Individual-based modeling and ecology. Princeton
University Press, Princeton and Oxford.
Hou, C., Zuo, W., Moses, M.E., Woodruff, W.H., Brown, J.H. & West, G.B. (2008). Energy
uptake and allocation during ontogeny. Science, 322, 736-739.
Jia, X., Dai, X.F., Shen, Z.X., Zhang, J.Y. & Wang, G.X. (2011) Facilitation can maintain
clustered spatial pattern of plant populations during density-dependent mortality:
insights from a zone-of-influence model. Oikos, 120, 472-480.
Lambers, H., Chapin, F.S. & Pons, T.L. (2008). Plant physiological ecology. 2nd edition.

- 126 - Chapter 4
Springer Verlag, New York, pp. 134-143.
Maestre, F.T., Callaway, R.M., Valladares, F. & Lortie, C.J. (2009). Refining the
stress-gradient hypothesis for competition and facilitation in plant communities. J.
Ecol., 97, 199–205.
Molofsky, J. & Bever, J.D. (2002). A novel theory to explain species diversity in landscapes:
positive frequency dependence and habitat suitability. Proc. R. Soc. Lond., B, Biol.
Sci., 269, 2389–2393.
Molofsky, J., Bever, J.D. & Antonovics, J. (2001). Coexistence under positive frequency
dependence. Proc. R. Soc. Lond., B, Biol. Sci., 2001, 268–277.
Niklas, K.J. (2005). Modelling below-and above-ground biomass for non-woody and
woody plants. Ann. Bot., 95, 315-321.
Niklas, K.J. & Enquist, B.J. (2001). Invariant scaling relationships for interspecific plant
biomass production rates and body size. Proc. Natl Acad. Sci. USA., 98,
2922-2927.
Price, C.A., Enquist, B.J. & Savage, V.M. (2007). A general model for allometric
covariation in botanical form and function. Proc. Natl Acad. Sci. USA., 104,
13204-13209.
Sack, L., Maranon, T., Grubb, P.J., Enquist, B.J. & Niklas, K.J. (2002). Global allocation
rules for patterns of biomass partitioning. Science, 296, 1923a.
Schwinning, S. & Weiner, J. (1998). Mechanisms determining the degree of size
asymmetry in competition among plants. Oecologia, 113, 447-455.
Stoll, P., Weiner, J., Muller-Landau, H., Müller, E. & Hara, T. (2002). Size symmetry of
competition alters biomass–density relationships. Proc. R. Soc. Lond. B: Biol. Sci.,
269, 2191-2195.
Travis, J.M.J., Brooker, R.W., Clark, E.J. & Dytham, C. (2006). The distribution of positive
and negative species interactions across environmental gradients on a dual-lattice
model. J. Theor. Biol., 241, 896–902.
West, G.B., Brown, J.H. & Enquist, B.J. (1999). A general model for the structure and
allometry of plant vascular systems. Nature, 400, 664-667.
West, G.B., Brown, J.H. & Enquist, B.J. (2001). A general model for ontogenetic growth.
Nature, 413, 628-631.
West, G.B., Enquist, B.J. & Brown, J.H. (2009). A general quantitative theory of forest
structure and dynamics. Proc. Natl Acad. Sci., 106, 7040-7045.

Chapter 4 - 127 -
Fig. S1. Conceptual model of the occurrence of mutualism versus commensalism in a
continuum of symmetric versus asymmetric facilitation.
Fig. S2. Spatial dynamics (black, aggregation; grey, randomness; white, regularity)
during self-thinning with aggregated initial locations and completely asymmetric
competition (CCA, p = ∞) in the absence (NF) or presence of facilitation (FCS,
completely symmetric facilitation, q = 0; FCA, completely asymmetric facilitation, q = ∞)
at different levels of abiotic stress (S).
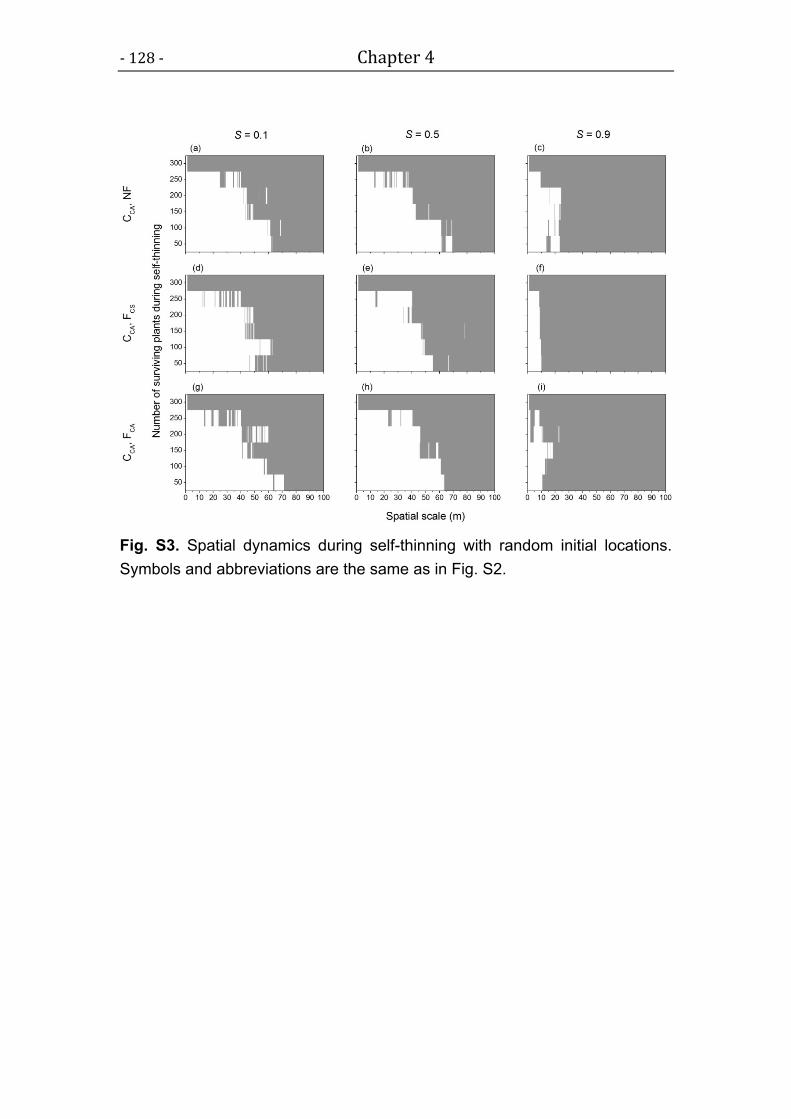
- 128 - Chapter 4
Fig. S3. Spatial dynamics during self-thinning with random initial locations.
Symbols and abbreviations are the same as in Fig. S2.

Chapter 5
Metabolic scaling theory predicts near ecological
equivalence in plants *
Abstract
Both niche and neutral theory aim to explain biodiversity patterns but seem
incompatible: whilst niche theory is based on the differences between species,
neutral models assume there are no such differences. Ecological equivalence
mediated by trade-offs however, is consistent with neutral theory whilst
allowing for ecological differences as suggested by niche theory. Here we show
with data from 375 plant species that metabolic scaling theory predicts such
trade-offs and thus supports ecological equivalence, at least as a near
approximation. This highlights a potentially important role for metabolic scaling
and points the way to a possible reconciliation between niche and neutral
theory.
Keywords: allometric scaling, neutral theory, ecological equivalence,
functional equivalence, demography, trade-offs, birth rate, death rate
* Manuscript in preparation
Yue Lin, Uta Berger, Volker Grimm, Qian-Ru Ji, James Rosindell, Jens Kattge, Helge
Bruelheide, and Richard Sibly

- 130 - Chapter 5
5.1 Introduction
Ecological difference among species (niche difference) is the conceptual basis
has been established since Darwin for explaining species coexistence and
diversity. By contrast, neutral theory (Hubbell 2001) provides models that
assume all species are ecologically identical. The idea received enormous
attention due to its accurate predictions despite its surprising simplicity, and
controversial assumption of ecological equivalence (Chave 2004; Rosindell et
al. 2012) which seems to be violated by many observable species-specific
characteristics.
One important aspect of neutral theory, however, has not yet been
sufficiently appreciated: ecological differences do not necessarily lead to
differences in net fitness (Chave 2004; Allouche & Kadmon 2009; Lin, Zhang,
& He 2009). Ecological equivalence regarding trade-offs between birth and
death rates among different species is still consistent with the neutral
assumption but allows for functional differences as suggested by niche theory
(Hubbell 2001; Chave 2004; Allouche & Kadmon 2009; Lin et al. 2009). It has
been shown in the simulation models that these birth-death trade-offs can be
incorporated into neutral theory without impairing its ability to predict
biodiversity patterns (Chave 2004; Lin et al. 2009; Rosindell et al. 2012).
Although predictions of neutral model have been tested extensively
(Hubbell 2001, 2005, 2006, 2008; Chave 2004; Allouche & Kadmon 2009; Lin
et al. 2009; Rosindell, Hubbell, & Etienne 2011), the cornerstone of neutral
theory which is the hypothesis of ecological equivalence has rarely been tested
(Hubbell 2008). Such ecological equivalence amongst individuals (species),
namely the demographic trade-offs, can be attributed to the intrinsic
explanations as neutral assumption assumed and/or the extrinsic explanations
as niche differences suggested. The intrinsic explanations are determined
essentially by the metabolism of organisms, whereas the extrinsic

Chapter 5 - 131 -
explanations are mainly related to external factors of environment such as
competition, resource, disease, predation or disaster (McCoy & Gillooly 2008).
In this study, we attempt to give a mechanistic, generic explanation for
these observed birth-death trade-offs, and thus provide a new and compelling
explanation for the success of neutral theory in a non-neutral world. To test our
predictions, we compiled data of demographic rates of 375 plant species which
include different functional groups and cover 11 orders of magnitude in plant
size ranging from mosses to trees.
The explanation we seek is provided by building on a base given by the
metabolic scaling theory (MST). This theory of ecology predicts quantitative
relationships between various functional traits of organisms and their body size
based on their metabolic rates (Brown et al. 2004; Price et al. 2010).
Extensions of MST yield that birth and death rates, b and d, are strongly
governed by the mass-specific metabolic rate (B), and thus should show the
body size (M) and temperature dependence (Brown et al. 2004; McCoy &
Gillooly 2008; Price et al. 2010) as:
b ∝ d ∝ B ∝ M -1/4e-E/kT (5-1)
where e-E/kT is the Boltzmann-Arrhenius factor which describes the exponential
increase in biochemical reaction rates with body temperature (Brown et al.
2004; Marbà et al. 2007; McCoy & Gillooly 2008; Price et al. 2010), E the
average activation energy of rate-limiting biochemical metabolic reactions (≈
0.32 eV for plants), k the Boltzmann’s constant (= 8.62×10-5 eV K-1), and T
the absolute temperature (Kelvin).
Although such scaling relationships have partly been statistically validated
before (Brown et al. 2004; Marbà, Duarte, & Agustí 2007; Price et al. 2010), so
far they were not linked to neutral theory. A reasonable estimation of overall
fitness (R0) can be measured as the ratio of birth rate over death rate which

- 132 - Chapter 5
gives the lifetime reproductive success or intrinsic rate of increase (Hubbell
2001; Chave 2004; Niklas 2007; Allouche & Kadmon 2009) leading to
R0 = b / d ∝ M -1/4 e-E/kT / M -1/4 e-E/kT ∝ M 0 (5-2)
Thus, fitness is predicted to be invariant with respect to body size, temperature,
as well as the metabolic rate across different functional groups of plant species.
It is almost self-explanatory that birth-death trade-offs is strongly governed by
the metabolic constrains of organisms, in other words, the intrinsic
mechanisms are expected to be the primary explanations. If this is true, the
overall fitness should be a constant among individuals and species, and the
ecological equivalence is to be expected as R0 = 1, which is also the parameter
setting of neutral models (Hubbell 2001; Chave 2004; Allouche & Kadmon
2009).
5.2 Materials and Methods
Temperature dependence of metabolism. Coexisting plant species in a
community can be assumed to experience the same temperature, so the
Boltzmann-Arrhenius factor can be omitted among species that coexisting in a
certain community. Moreover, the data set of birth and death rates analyzed in
this study also shows temperature independence (Student’s t-test, P = 0.43 and
P = 0.50 respectively), which is consistent with previous studies suggesting that
metabolic rates of plant are less temperature dependent than those of animals
(Marbà et al. 2007; Niklas 2007; Allouche & Kadmon 2009). Such findings
might also due to either statistical ‘‘noise’’ or the narrow band of temperature in
the data set (Marbà et al. 2007; Niklas 2007).
Data Compilation. Because the predictions of MST are base on a fractal-like
architecture of transporting network, so we compiled data on individual
average body masses, natural birth and death rates from a broad array of
higher plant species (excluding phytoplankton, algae, and macro-algae which

Chapter 5 - 133 -
do not have fractal-like architectures). Our data set includes mosses,
seaweeds, ferns, grasses, herbs, succulents, shrubs, lianas, mangroves as
well as other tree species. Each taxonomic group is represented by a diversity
of species and with plant body sizes span a very broad range (7.0×10-3 to
1.1×107g). All these data were primarily compiled from published data sets and
some are gathered from large census plots.
Measure of birth rate. We calculated the birth rates (b, in year -1) from initial
number of individuals (N0) and the number of new-born individuals (Nb) during
the time past between census interval (t, in year) as:
0 0ln ( ) ln ( )bN N Nb
t
(5-3)
Measure of death rate. The death rates (d, in year -1) are calculated as:
0ln ( ) ln ( )N Sd
t
(5-4)
where S indicates the number of survivors in the later census.
Estimate of tree mass. Tree mass (M, dry biomass in grams) that cannot
directly get was estimated from DBH (diameter at breast height in centimeters)
by using an allometric equation:
M = 78·DBH 2.5 (5-5)
this equation was fitted for a compilation of independent data set which
enclosing a very broad range of size for tree flora (Marbà et al. 2007).
Data Analysis. Because we are intend to answer different biological questions,
both Model I and II regressions are applied in our work by following the
suggestions about model selection (Smith 2009). To test the relationship
between body size (M) and the birth rates, death rates, and overall fitness (R0),
the allometric equations are fitted to log-transformed data by using ordinary
least squares regression (OLS, Model I regression). This is because we are
interested about how demographic rates change with plant body size. We use

- 134 - Chapter 5
the reduced major axis regression (RMA, Model II regression) for
log-transformed data fitting between birth and death rates. Because we
attempt to ascertain the actuality of ecological equivalence (b = d), meaning
that the bivariate relationship is symmetric and regardless of which variable is
dependent and which is independent.
5.3 Results and Discussion
Our analysis of data confirms that most plant populations examined here
are ecologically equivalent. Death rate, d, is proportional to the birth rate, b, as
d = 1.104 b 1.001 (Fig. 5.1 A). This does not differ statistically from the
equivalence in vital rates (b = d) assumed by neutral model. Furthermore,
observed distribution of the intrinsic rate of increase (R0 = b / d) is unimodal
with a mean of 1 (Fig. 5.1 B), implying a validity of nearly ecological
equivalence (Hubbell 2008).
In agreement with theoretical predictions, the data shows that species
balance high birth rates with high death rates according to mass-specific
metabolic rates and scale as -1/4 power of body mass (Fig. 5.2, A and B). The
overall fitness, R0, scales as M 0.006, and does not differ statistically from the
predicted mass invariance (M 0) (Fig. 5.2 C). Such results imply that ecological
equivalence as demographic trade-offs among different plant species are
largely constrained by intrinsic mechanisms of individual metabolic rate,
despite the fact that ecologically extrinsic factors exist in nature.
Nevertheless, the results also point to substantial variation (Fig. 5.1A) that
cannot be explained by metabolic constraints alone. Such variation might be
viewed as an indicator of non-neutral process among species from particular
communities, suggesting that both intrinsic and extrinsic constraints may
contribute to population and community dynamics. Therefore, it is more
relevant to test the relative contributions of intrinsic and extrinsic factors in
different populations and communities.

Chapter 5 - 135 -
Fig. 5.1 (A) The relationship between birth rates and death rates of plants (375
species). Red line shows the fitted reduced major axis regression (P < 0.0001, 95%
confidence intervals, exponent: 0.950 to 1.056, constant: 0.966 to 1.271). (B)
Distribution of the intrinsic rate of increase (overall fitness, calculated as the ratio
between birth rate and mortality rate, R0 = b / d) of plant species.

- 136 - Chapter 5
Fig. 5.2 (A) The relationship between plant body size and birth rates. Black
line shows the fitted ordinary least squares regression (OLS, Model I
regression; P < 0.0001, 95% confidence intervals, exponent: -0.315 to -0.232,
constant: 0.237 to 0.372). (B) The relationship between plant body size and

Chapter 5 - 137 -
death rates. Black line shows the fitted OLS regression (P < 0.0001, 95%
confidence intervals, exponent: -0.273 to -0.241, constant: 0.408 to 0.545). (C)
The relationship between plant body size and overall fitness (intrinsic rate of
increase, calculated as the ratio between birth rate and death rate, R0 = b / d).
Black line shows the fitted OLS regression (P < 0.707, 95% confidence
intervals, exponent: -0. 024 to 0.036, constant: 0.697 to 0.964).
Overall, our study indicates that neutrality as assumed by neutral theory
might be the consequence of scaling relationships which emerges from
constraints, trade-offs, and evolved characteristics of metabolism and
physiology. Meanwhile, MST also predicts different functional traits across
different functional groups of species (Brown et al. 2004; Price et al. 2010).
Thus, niches and neutrality are not excluding each other but might in fact have
the same mechanistic root. The current debate about whether niche or neutral
mechanisms structure natural communities might miss the point. The real
question should be when and why one of these factors dominates. Our
initiative work may provide some insights regarding the relative importance of
niche vs. neutral based process on community assembly and species
diversity.
References
Allouche, O. & Kadmon, R. (2009) A general framework for neutral models of
community dynamics. Ecology letters, 12, 1287–1297.
Brown, J.H., Gillooly, J.F., Allen, A.P., Savage, V.M. & West, G.B. (2004)
Toward a metabolic theory of ecology. Ecology, 85, 1771–1789.
Chave, J. (2004) Neutral theory and community ecology. Ecology Letters, 7,
241–253.
Hubbell, S.P. (2001) The Unified Neutral Theory of Biodiversity and
Biogeography. Princeton University Press.
Hubbell, S.P. (2005) Neutral theory in community ecology and the hypothesis
of functional equivalence. Functional ecology, 19, 166–172.
Hubbell, S.P. (2006) Neutral theory and the evolution of ecological
equivalence. Ecology, 87, 1387–1398.

- 138 - Chapter 5
Hubbell, S.P. (2008) Approaching ecological complexity from the perspective
of symmetric neutral theory. Tropical forest community ecology.
Wiley-Blackwell, Chichester, UK, 143–159.
Lin, K., Zhang, D.Y. & He, F. (2009) Demographic trade-offs in a neutral model
explain death-rate-abundance-rank relationship. Ecology, 90, 31–38.
Marbà, N., Duarte, C.M. & Agustí, S. (2007) Allometric scaling of plant life
history. Proceedings of the National Academy of Sciences, 104, 15777.
McCoy, M.W. & Gillooly, J.F. (2008) Predicting natural mortality rates of plants
and animals. Ecology letters, 11, 710–716.
Niklas, K.J. (2007) Sizing up life and death. Proceedings of the National
Academy of Sciences, 104, 15589.
Price, C.A., Gilooly, J.F., Allen, A.P., Weitz, J.S. & Niklas, K.J. (2010) The
metabolic theory of ecology: prospects and challenges for plant biology.
New Phytologist, 188, 696–710.
Rosindell, J., Hubbell, S.P. & Etienne, R.S. (2011) The Unified Neutral Theory
of Biodiversity and Biogeography at Age Ten. Trends in Ecology &
Evolution.
Rosindell, J., Hubbell, S.P., He, F., Harmon, L.J. & Etienne, R.S. (2012) The
case for ecological neutral theory. Trends in Ecology & Evolution, 27,
203–208.
Smith, R.J. (2009) Use and misuse of the reduced major axis for line‐fitting.
American journal of physical anthropology, 140, 476–486.

Chapter 6
General discussion
The general aim of my dissertation was to investigate the role of plant
interactions in driving population dynamics. Both theoretical and empirical
approaches were employed, such as individual-based models (IBMs) and
greenhouse experiment. All my studies are conducted on the basis of
metabolic scaling theory (MST, Brown et al. 2004, Price et al. 2010), because
the complex, spatially and temporally varying structures and dynamics of
ecological systems are considered to be largely consequences of biological
metabolism (Brown et al. 2004). On the other hand, MST didn’t consider the
important role of plant interactions and was found to be invalid in some
environmental conditions. Integrating the effects of plant interactions and
environmental conditions into MST may be essential for reconciling MST with
observed variations in nature. Such integration will improve the development
of theory, and will help us to understand the relationship between individual
level process and system level dynamics.
In the following, I will discuss what we in general learned from the work
presented in this thesis. First, I will discuss the pros and cons of mechanistic
and phenomenological models, in particular whether it paid to develop and use
an ontogenetic growth model. Then I will turn to plant competition and how
modelling it can be integrated with MST. Next, facilitation in combination with
competition will be discussed as a framework for exploring interactions of
plants with each other and their abiotic environment. The final two sections will
be devoted to synthesis: first, of plant competition, MST, and Neutral Theory,
and finally of the approach to addressing ecological questions used in this
thesis, Individual-Based Ecology.

- 140 - Chapter 6
Mechanistic models versus Phenomenological models
In the beginning of this dissertation, we deduced a mechanistic growth model
for plant. It is important that we use a mechanistic growth model derived from
MST rather than a phenomenological growth model, as usually has been done
so far in other plant IBMs (e.g., Weiner et al. 2001, Stoll et al. 2002, Stoll and
Bergius 2005, Chu et al. 2008, 2009, 2010, May et al. 2009). First, our
mechanistic growth model is based on the energy budget of the individuals
during growth which captures the key features of metabolic process of
organisms (West et al. 2001, Hou et al. 2008). Such mechanistic models
driven by ‘first principles’ are more flexible and are capable of capturing
adaptive responses of individuals to their environment than phenomenological
models, which are statistically fitted to existing data and forego any attempt in
explaining the underlying mechanisms (Hilborn and Mangel 1997, Grimm and
Railsback 2005, Martin et al. 2012). Second, the mechanistic models can be
easily extended to predict further patterns by deduction (e.g., the reproduction
biomass of individual plant, in Appendix of Chapter 1), whereas
phenomenological models are usually induced by patterns that were observed.
Mechanistic models are more likely to work correctly when extrapolating
beyond the current states than empirical models. Third, ecological or
mathematical artifacts are easier to be prevented if using mechanistic models
deduced from physical or physiological rules. For instance, in many IBMs plant
growth is assumed to be negatively and linearly related to resource
competition and environmental stress (Weiner et al. 2001, Stoll et al. 2002,
Stoll and Bergius 2005, Chu et al. 2008, 2009, 2010, May et al. 2009, Jia et al.
2011), with maximum body size kept constant. This phenomenological
assumption is both mathematical and ecological incorrect, because it implies
that plants can reach their optimal size regardless environmental conditions.
Fourth, in contrast to previous models used in plant IBMs (Wyszomirski 1983,
Wyszomirski et al. 1999, Weiner et al. 2001, May et al. 2009), our model is not

Chapter 6 - 141 -
only able to explain the observed variation in both root-shoot allocation
patterns, but also simultaneously describe the mass-density relationships (in
Chapter 3).
Yet, the merits of phenomenological/statistical models should not be
underestimated, especially when the aim is only to describe (or predict) rather
than to understand the mechanisms. Both the mechanistic models and
phenomenological models are useful, it is more relevant to ask how to choose
a model properly, in particular in IBMs. The POMIC (pattern-oriented modeling
information criterion, Piou et al. 2009) offers a rigorous statistical approach of
model selection which is in a context of POM (pattern-oriented modeling) and
is more efficient and specialized for IBMs. Nevertheless, in IBMs, with all other
things being equal, mechanistic models are arguably more powerful because
they can explain underlying processes driving observed patterns. Meanwhile,
the phenomenological models can also be used to stimulate and improve our
understandings on the mechanisms underlying observed patterns. A case in
point is how the ‘Kleiber’s law’ urge MST to come out (Brown et al. 2004).
Plant competition
In Chapter 2, we investigated the role of different modes of competition in
altering the prediction of MST on plant self-thinning trajectories. Our
spatially-explicit individual-based zone-of-influence (ZOI) model was
developed to investigate the hypothesis that MST may be compatible with the
observed variation in plant self-thinning trajectories if different modes of
competition and different resource availabilities are considered. Our one-layer
ZOI model supported our hypothesis that (i) size-symmetric competition (e.g.
belowground competition) will lead to significantly shallower self-thinning
trajectories than size-asymmetric competition as predicted by MST; and (ii)
individual-level metabolic processes can predict population-level patterns
when surviving plants are barely affected by local competition, which is more
likely to be in the case of asymmetric competition. In contrast to our findings,

- 142 - Chapter 6
some researchers proposed that asymmetric competition is more important to
explain the deviations predicted by MST (Coomes and Allen 2009, Coomes et
al. 2011). However, this debate again is due to a failure of distinguishing
between the ‘importance’ and ‘intensity’ of competition (Welden and Slauson
1986): we respect to the competitive effect on population level (mass-density
relationship) which is about the importance of competition, whereas they focus
on competitive response of single tree (individual tree growth) which is more
related to the intensity of competition.
In Chapter 3, we go further and add a bit more complexity by considering
the phenotypic plasticity and adaptive behaviour of plants in changing
environments. A two-layer ZOI model was developed which considers
allometric biomass allocation to shoots or roots and represents both above-
and belowground competition simultaneously via independent ZOIs. In
addition, we also performed greenhouse experiment to evaluate the model
predictions. Both our theoretical model and experiment demonstrated that:
plants are able to adjust their biomass allocation in response to environmental
factors, and such adaptive behaviours of individual plants, however, can alter
the relative importance of above- or belowground competition, thereby
affecting plant mass-density relationships at population level. Root competition,
which is assumed to be more symmetric, can strongly affect the growth of
surviving plants therefore alter the mass-density relationships. Invalid
predictions of MST are likely to occur where competition occurs belowground
(symmetric) rather than aboveground (asymmetric). Consequently, models or
theories which only consider aboveground part are not appropriate for the
cases where belowground processes predominate (Deng et al. 2006, Berger
et al. 2008). In addition to previous studies which do not consider the linkage
between above- and belowground competition, our study implies that adaptive
behaviours of plants such as allometry can modify ecological mechanisms and
subsequently the constraints set by MST. This has important implications for
many forests and eco-regions, e.g., for using allometry-based models to

Chapter 6 - 143 -
predict carbon storage and sequestration.
Plant competitions and metabolic scaling theory
Chapter 2 and 3 jointly showed that, MST, like principles from energetics and
biomechanics, sets limits on the behaviour of individuals and therefore of
populations and communities. In some ecological situations these limits will
dominate, and MST will predict higher-level behaviour. In many cases however,
other constraints will determine the patterns observed. Our most important
conclusion is that the behaviour of populations and communities may be
dominated by internal physiological mechanisms addressed by MST or by
ecological factors beyond the individual level, such as the type of resource
limitation and the mechanisms of competition among individuals. In the latter
cases MST will not be predictive, although nor will it be violated. The claim that
MST provides a ‘universal law’ for quantitatively linking the energetic
metabolism of individuals to ecological system dynamics is thus neither
completely right nor wrong: the real question is when and which factors
dominate in a given situation. However, our studies show that cross-level tests
are needed to explore and develop individual-level theories. Martin et al.
(2012), who used an individual-based population model of Daphnia based on
Dynamic Energy Budget theory (Kooijman 2009), arrived at the same
conclusion. Without doubts, the consideration of plant interactions is critical for
understanding variation in observed patterns.
Plant competition and facilitation
In Chapter 4, we introduced the new concept of modes of facilitation, i.e.
symmetric versus asymmetric facilitation, and developed an individual-based
model to explore how the interplay between different modes of competition and
facilitation changes spatial pattern formation in plant populations, and how
combinations of competition and facilitation modes alter the intensity of local

- 144 - Chapter 6
plant interactions along a stress gradient. Our study shows that facilitation by
itself can play an important role in promoting plant aggregation independent of
other ecological factors (e.g. seed dispersal, recruitment, and environmental
heterogeneity). Such finding identifies the origin of an important mechanism
underlying self-organization: the process of local facilitation among plants that
set against the background of overall control by environmental constrains.
Our results also indicate that the indices widely used to estimate intensity
of local interactions is too limited to represent the relative importance of
different modes of interactions in structuring plant system dynamics. Different
modes of facilitation and competition can affect different aspects of plant
populations and communities, leading to variously spatial and temporal
patterns, and implying context-dependent outcomes and consequences. For
instance, we find that the net outcome of local interactions among plants can
show a dominance of competition when using the index of relative interaction
intensity, in spite of the fact that local facilitation plays an important role in
structuring spatial aggregation. Although the intensity of interaction as the net
outcome is measurable, but the relative importance of competition and
facilitation in structuring system dynamics is much more complicated. To clarify
some debates in current ecological researches, it is necessary to distinguish
the differences between intensity and importance of different modes of
competition and facilitation, and to develop a comprehensive approach rather
than only use some indices. Moreover, a moderate mixture of positive and
negative interactions is demonstrated to stabilize population dynamics (Mougi
and Kondoh 2012). The diversity of species and interaction types may be the
essential element of biodiversity that maintains ecological communities, and
may hold the key to understanding population and community dynamics
(Grimm and Railsback 2005, Mougi and Kondoh 2012).
Towards a synthesis of metabolic scaling, niche, and neutral theory
In Chapter 5, we went from population level to community level and explored

Chapter 6 - 145 -
the possibility of combining the MST and unified neutral theory of biodiversity
(UNT, Hubbell 2001). Studies addressing UNT have largely focused on testing
the predictions of neutral models, whereas the fundamental assumption of
ecological equivalence has rarely been tested with empirical studies. Our
analysis of extensive data confirms that most plant populations examined are
nearly neutral in a sense of demographic trade-offs, which can mostly be
explained by a simple allometric scaling rule that based on MST. This
demographic equivalence regarding birth-death trade-offs between different
species and functional groups is consistent with the assumption of neutral
theory but allows functional differences between species (K. Lin et al. 2009).
Both niche and neutral mechanisms may be essential processes in community
assembly. In agreement with this, a new theoretical framework proposed four
basic processes in a community: natural selection (niche mechanisms),
ecological drift (neutral mechanisms), dispersal limitation and speciation. But
the relative importance of these four processes varies among communities
(Vellend 2008).
Current debate about whether niche or neutral mechanisms structure
natural communities apparently missing the point. Our initial study provides
some insights into this debate: the real question should be when and why one
of these factors dominates. Because UNT emphasizes neutral mechanisms
(ecological drift) as the main drivers of community assembly, for this reason
the demographic dynamics of all species in a certain community should be a
martingale. However, our results also show the variation of demographic
trade-offs that cannot be explained by metabolic constraints alone. Such
variation may be viewed as an indicator of non-neutral mechanisms (e.g.,
interactions and natural selection) among species in the communities, and the
goodness of fit (the coefficient of determination) may be viewed as the
contribution of neutral mechanisms. The empirical tests about the relative
importance of different mechanisms in natural communities are urgently
needed.

- 146 - Chapter 6
MST or similar bio-energetic theories (e.g., Dynamic energy budget
theory, Kooijman 2009) have the potential to unify existing biodiversity theories,
which are focused on abundances and species number, and link them to
questions regarding ecosystem functioning, e.g. productivity, plasticity, stability,
respiration, carbon flux etc. Such a synthesis of bio-energetic and
demographic theories is needed to better understand the relationship between
diversity and function of ecosystems and to unify future research in biodiversity
theory and management.
The Individual-Based Ecology
Ecology as a developing science is full of fresh ideas, but most of the
influential ideas still have not been mathematically well developed (Lawton
1999, Ghilarov 2003, Simberloff 2004, Grimm and Railsback 2005, Scheiner
and Willig 2008, 2011, Odenbaugh 2011a). Elaborate theories of ecology
which reflect ‘physics envy’ often omit the differences among organisms and
their traits, which I so-called ‘the assumption of spherical trees in a vacuum’.
However, since organisms are not just atoms such ‘physics envy’ theories are
of little use if we are going to understand the emergent properties of complex
systems such as population dynamics or community assembly out of
ecological traits, behaviours and interactions of individual organisms
(DeAngelis and Gross 1992, Breckling et al. 2005, Grimm and Railsback 2005).
The legendary ecologist MacArthur claimed “the best ecologists had blurry
vision so they could see the big patterns without being overly distracted by the
contradictory details” (Odenbaugh 2010, 2011b). Yet, an approach that can
‘zoom in and zoom out’ would be more appropriate if we are going to
understand the ecologically complex systems from individual levels (Grimm
and Railsback 2005). Accounting for omitted details often leads to more
refined theories. For instance, the van der Waals equation was derived as a
more precise version of the ideal gas law by incorporating the facts that
molecules have a nonzero volume and that they’re attracted to each other, for

Chapter 6 - 147 -
which Johannes van der Waals received the Nobel Prize for Physics in 1910
(Fox 2011). As in all such quests, it is important to keep in mind Einstein’s
dictum ‘seek simplicity, and distrust it’. Nevertheless, the main challenge in
ecology is to understand complexity and how it emerges from the adaptive
traits of individuals (Levin 2000, Grimm and Railsback 2005) which is the
major goal of individual-based ecology. The approaches of individual-based
and pattern-oriented modelling are promising to achieve the synthesis.
References
Berger, U., C. Piou, K. Schiffers, and V. Grimm. 2008. Competition among plants:
concepts, individual-based modelling approaches, and a proposal for a future
research strategy. Perspectives in Plant Ecology, Evolution and Systematics
9:121–135.
Breckling, B., F. Müller, H. Reuter, F. Hölker, and O. Fränzle. 2005. Emergent properties
in individual-based ecological models—introducing case studies in an ecosystem
research context. Ecological Modelling 186:376–388.
Brown, J. H., J. F. Gillooly, A. P. Allen, V. M. Savage, and G. B. West. 2004. Toward a
metabolic theory of ecology. Ecology 85:1771–1789.
Chu, C. J., F. T. Maestre, S. Xiao, J. Weiner, Y. S. Wang, Z. H. Duan, and G. Wang. 2008.
Balance between facilitation and resource competition determines
biomass–density relationships in plant populations. Ecology Letters
11:1189–1197.
Chu, C. J., J. Weiner, F. T. Maestre, Y. S. Wang, C. Morris, S. Xiao, J. L. Yuan, G. Z. Du,
and G. Wang. 2010. Effects of positive interactions, size symmetry of competition
and abiotic stress on self-thinning in simulated plant populations. Annals of Botany
106:647–652.
Chu, C. J., J. Weiner, F. T. Maestre, S. Xiao, Y. S. Wang, Q. Li, J. L. Yuan, L. Q. Zhao, Z.
W. Ren, and G. Wang. 2009. Positive interactions can increase size inequality in
plant populations. Journal of Ecology 97:1401–1407.
Coomes, D. A., and R. B. Allen. 2009. Testing the metabolic scaling theory of tree growth.
Journal of Ecology 97:1369–1373.
Coomes, D. A., E. R. Lines, and R. B. Allen. 2011. Moving on from Metabolic Scaling
Theory: hierarchical models of tree growth and asymmetric competition for light.
Journal of Ecology 99:748–756.
DeAngelis, D. L., and L. J. Gross. 1992. Individual-based models and approaches in
ecology: populations, communities and ecosystems. Chapman & Hall.
Deng, J. M., G. X. Wang, E. C. Morris, X. P. Wei, D. X. Li, B. Chen, C. M. Zhao, J. Liu, and

- 148 - Chapter 6
Y. Wang. 2006. Plant mass–density relationship along a moisture gradient in
north-west China. Journal of Ecology 94:953–958.
Fox, J. 2011. Why macroecology needs ‘microecology’: revisiting an Oikos classic.
http://oikosjournal.wordpress.com/2011/05/03/why-macroecology-needs-microec
ology-revisiting-an-oikos-classic/
Ghilarov, A. M. 2003. The changing place of theory in 20th century ecology: from
universal laws to array of methodologies. Oikos 92:357–362.
Grimm, V., and S. F. Railsback. 2005. Individual-Based Modeling and Ecology. Princeton
University Press.
Hilborn, R., and R. H. M. Mangel. 1997. The Ecological Detective: Confronting Models
with Data. Princeton University Press.
Hou, C., W. Zuo, M. E. Moses, W. H. Woodruff, J. H. Brown, and G. B. West. 2008.
Energy uptake and allocation during ontogeny. Science 322:736–739.
Hubbell, S. P. 2001. The unified neutral theory of biodiversity and biogeography.
Princeton University Press.
Jia, X., X. F. Dai, Z. X. Shen, J. Y. Zhang, and G. X. Wang. 2011. Facilitation can maintain
clustered spatial pattern of plant populations during density-dependent mortality:
insights from a zone-of-influence model. Oikos 120:472–480.
Kooijman, B. 2009. Dynamic Energy Budget Theory for Metabolic Organisation, 3rd
edition. Cambridge University Press.
Lawton, J. H. 1999. Are there general laws in ecology? Oikos:177–192.
Levin, S. A. 2000. Fragile Dominion: Complexity and the Commons. Basic Books.
Lin, K., D. Y. Zhang, and F. He. 2009. Demographic trade-offs in a neutral model explain
death-rate-abundance-rank relationship. Ecology 90:31–38.
Martin, B. T., E. I. Zimmer, V. Grimm, and T. Jager. 2012. Dynamic Energy Budget theory
meets individual-based modelling: a generic and accessible implementation.
Methods in Ecology and Evolution 3:445–449.
May, F., V. Grimm, and F. Jeltsch. 2009. Reversed effects of grazing on plant diversity:
the role of below‐ground competition and size symmetry. Oikos 118:1830–1843.
Mougi, A., and M. Kondoh. 2012. Diversity of Interaction Types and Ecological
Community Stability. Science 337:349–351.
Odenbaugh, J. 2010. Robert H. MacArthur.
Odenbaugh, J. 2011a. A General, Unifying Theory of Ecology? The Theory of Ecology:51.
Odenbaugh, J. 2011b. Philosophical themes in the work of Robert H. MacArthur.
Philosophy of Ecology 11:109.
Piou, C., U. Berger, and V. Grimm. (2009) Proposing an information criterion for
individual-based models developed in a pattern-oriented framework. Ecological
Modelling 220: 1957-1967.
Price, C. A., J. F. Gilooly, A. P. Allen, J. S. Weitz, and K. J. Niklas. 2010. The metabolic
theory of ecology: prospects and challenges for plant biology. New Phytologist
188:696–710.
Scheiner, S. M., and M. R. Willig. 2008. A general theory of ecology. Theoretical Ecology
1:21–28.
Scheiner, S. M., and M. R. Willig. 2011. The Theory of Ecology. University of Chicago

Chapter 6 - 149 -
Press.
Simberloff, D. 2004. Community ecology: Is it time to move on? The American Naturalist
163:787–799.
Stoll, P., and E. Bergius. 2005. Pattern and process: competition causes regular spacing
of individuals within plant populations. Journal of Ecology 93:395–403.
Stoll, P., J. Weiner, H. Muller-Landau, E. Müller, and T. Hara. 2002. Size symmetry of
competition alters biomass–density relationships. Proceedings of the Royal
Society of London. Series B: Biological Sciences 269:2191–2195.
Vellend M (2010) Conceptual synthesis in community ecology. Quarterly Review of
Biology, 85, 183–206.
Weiner, J., P. Stoll, H. Muller‐Landau, and A. Jasentuliyana. 2001. The effects of density,
spatial pattern, and competitive symmetry on size variation in simulated plant
populations. The American Naturalist 158:438–450.
Welden, C. W., and W. L. Slauson. 1986. The intensity of competition versus its
importance: an overlooked distinction and some implications. Quarterly Review of
Biology:23–44.
West, G. B., J. H. Brown, and B. J. Enquist. 2001. A general model for ontogenetic growth.
Nature 413:628–631.
Wyszomirski, T. 1983. Simulation model of the growth of competing individuals of a plant
population. Ekologia Polska 31.
Wyszomirski, T., I. Wyszomirska, and I. Jarzyna. 1999. Simple mechanisms of size
distribution dynamics in crowded and uncrowded virtual monocultures. Ecological
Modelling 115:253–273.


Acknowledgments - 151 -
Acknowledgments
I have met many wonderful people during my PhD studies, I am really grateful to all
of them!
First of all, I would like to thank my advisor Uta Berger for all her generous help.
Uta gave me the freedom, trust and support to bring in my own ideas into this research
topic. But her opportune advices can always remind me the dreamer to put my feet on
the ground. I appreciate that she opened the door of mangroves to me and brought me
to an interesting field.
Many thanks go to my co-advisor Volker Grimm, who just has the magic to make
me be full of enthusiasm in my studies. When I was not convinced of my work, he
was the one who believed the merits of it even more than me. His ‘Scientific writing
course’ is the best course I have ever had.
I cannot express myself in words how thankful I am to Uta and Volker. I greatly
profited from their experience in science and life. The ‘facilitations’ I have received
from them are ‘completely asymmetric’. Without their supervision, I would never
have the opportunity to arrive the present thesis. As I said before, they are my mentors
forever.
There are a lot of people who helped me for some of my studies. I would like to thank
wholeheartedly: Franka Huth for her kind help on the greenhouse experiments, it
was so pleasure working with her. Jacob Weiner who is a co-author of one of my
manuscripts, his insightful comment not only improved the manuscript but also my
knowledge and experience of doing science. Juliane Vogt and Peter Frohberg, we
are “the best team in mangrove”. Prof. Dr. Steven Railsback for his help on my paper.
Prof. Dr. C. Edward Proffitt, Prof. Dr. Donna Devlin, and Prof. Dr. Candy Feller
for their kindly help during our research stays in Florida. Dr. Ulf Mehlig and Dr.
Moirah Menezes for their warmly help on our work in Brazil. Prof. Dr. James
Brown, Prof. Dr. Brian Enquist, Prof. Dr. Chen Hou, and Dr. Jian-Ming Deng for
their contributive suggests to my studies. Prof. Dr. Richard Sibly and Dr. James
Rosindell, Dr. Jens Kattge, and Prof. Dr. Helge Bruelheide for their kind help and
discussions on one of my manuscript. Dr. Stephen Wolfram, the conversation with
him inspired me to think the relationship between physics and ecology.
I wish to thank everyone in the Institut für Waldwachstum und Forstliche Informatik
for a friendly working atmosphere. Many thanks go to all my colleagues for their
moral support in daily PhD life, especially, Martin Schubert, Ronny Peters,
Andreas Tharang, Uwe Grüters, Mariana Vidal, Soledad Luna, Aor Pranchai,
Ursula Behr and Gisela Metzig. I am also grateful to my “doctoral bother”
Muhammad Ali Imron who is now in Indonesia.

- 152 - Acknowledgments
I would like also to thank Prof. Dr. Florian Jeltsch for accepting to evaluate my
dissertation, and Prof. Dr. Sven Wagner and Prof. Dr. Michael Müller to be on my
defense committee. I am also grateful for the help of Mrs. Bärbel Lommatzsch.
I could not have completed this doctoral dissertation without the support of
HIGRADE (Helmholtz Impulse and Networking Fund through Helmholtz
Interdisciplinary Graduate School for Environmental Research) fellowship. Many
thanks to my colleagues in UFZ.
At last, my love goes to my wife, Qian-Ru Ji for all her support and love during this
PhD thesis, for cheering me up and for her always so proud of me.
感谢我的母亲和父亲,没有你们的理解和支持,就没有我现在的一切。
感谢我的岳父和岳母,把茜茹交给了我,让我们可以自由自在的生活。
你们,是我们永远的港湾,我们取得的一切成绩也同样属于你们。