the revolution that wasn't: a new interpretation of the origin of
TRANSCRIPT

Sally McBreartyDepartment of Anthropology,University of Connecticut,Storrs, Connecticut 06269,U.S.A. E-mail:[email protected]
Alison S. BrooksDepartment of Anthropology,George WashingtonUniversity, Washington,DC 20052, U.S.A. E-mail:[email protected]
Received 3 June 1999Revision received 16 June2000 and accepted 26 July2000
Keywords: Origin of Homosapiens, modern behavior,Middle Stone Age, Africanarchaeology, MiddlePleistocene.
The revolution that wasn’t: a newinterpretation of the origin of modernhuman behavior
Proponents of the model known as the ‘‘human revolution’’ claimthat modern human behaviors arose suddenly, and nearly simul-taneously, throughout the Old World ca. 40–50 ka. This fundamentalbehavioral shift is purported to signal a cognitive advance, a possiblereorganization of the brain, and the origin of language. Because theearliest modern human fossils, Homo sapiens sensu stricto, are found inAfrica and the adjacent region of the Levant at >100 ka, the ‘‘humanrevolution’’ model creates a time lag between the appearance ofanatomical modernity and perceived behavioral modernity, andcreates the impression that the earliest modern Africans were behav-iorally primitive. This view of events stems from a profound Euro-centric bias and a failure to appreciate the depth and breadth of theAfrican archaeological record. In fact, many of the components ofthe ‘‘human revolution’’ claimed to appear at 40–50 ka are found inthe African Middle Stone Age tens of thousands of years earlier.These features include blade and microlithic technology, bone tools,increased geographic range, specialized hunting, the use of aquaticresources, long distance trade, systematic processing and use ofpigment, and art and decoration. These items do not occur suddenlytogether as predicted by the ‘‘human revolution’’ model, but at sitesthat are widely separated in space and time. This suggests a gradualassembling of the package of modern human behaviors in Africa, andits later export to other regions of the Old World. The African Middleand early Late Pleistocene hominid fossil record is fairly continuousand in it can be recognized a number of probably distinct species thatprovide plausible ancestors for H. sapiens. The appearance of MiddleStone Age technology and the first signs of modern behavior coincidewith the appearance of fossils that have been attributed to H. helmei,suggesting the behavior of H. helmei is distinct from that of earlierhominid species and quite similar to that of modern people. If onanatomical and behavioral grounds H. helmei is sunk into H. sapiens,the origin of our species is linked with the appearance of MiddleStone Age technology at 250–300 ka.
� 2000 Academic Press
Journal of Human Evolution (2000) 39, 453–563doi:10.1006/jhev.2000.0435Available online at http://www.idealibrary.com on
Introduction and background
The human revolution in EuropeFor at least the past 15 years, most recon-structions of later human evolutionaryhistory have featured a relatively brief anddramatic shift known as the ‘‘human revol-ution’’ (Binford, 1985, 1989; Mellars &Stringer, 1989; Klein, 1989a, 1994, 1995,
0047–2484/00/110453+111$35.00/0
2000; Diamond, 1992; Mellars, 1995,1996; Nobel & Davidson, 1991; Tattersall,1995; Bar-Yosef, 1998). The ‘‘humanrevolution’’ model proposes a dramaticalteration in human behavior at the MiddlePaleolithic to Upper Paleolithic transition atabout 40 ka. This behavioral breakthroughis thought by some to correspond toincreased cognitive sophistication, the
� 2000 Academic Press

454 . . .
manipulation of symbols, and the origin oflanguage (e.g., White, 1982; Mellars &Stringer, 1989; Diamond, 1992; Byers,1994; Mithen, 1994, 1996; Klein, 1995; butsee Kay et al., 1998). We believe that themodel of the ‘‘human revolution’’ is fatallyflawed. Modern humans and modernhuman behaviors arose first in Africa, andwe examine the African record to reveal adifferent picture of the nature of events.
The concept of a ‘‘human revolution’’ andthe periodization of Stone Age prehistoryhave their roots in the nineteeth-centuryprobings of the Western European archaeo-logical record. The first paleolithic classifi-catory schemes were based on the WesternEuropean large mammal succession (Lartet& Christy, 1865–1875; Lyell, 1868), andthese authors emphasized the wide techno-logical gulf separating the l’age du renne(Upper Paleolithic) from the earlier phases(Lartet & Christy, 1865–1875:25). By the1920s the concept of an Upper Paleolithicdistinguished by the appearance of engrav-ing, sculpture, painting, beads, and workedbone tools had become current. A tripartitedivision into Lower, Middle and UpperPaleolithic based upon stone tool technology(De Mortillet, 1900; Obermeier, 1924;Burkitt, 1921, 1928, 1933; Kendrick, 1925;Menghin, 1931) echoed the three-age sys-tems of Thomsen (1837) and Worsaae(1849) that partitioned the total prehistoricrecord into ages of Stone, Bronze, and Iron.
The Lower, Middle and Upper Paleolithicdivisions of the western European recordhave continued to dominate discourse in thefield, despite problems in the application ofthese divisions to sequences in Eastern andSouthern Europe (Morselli, 1926:292).None of these temporal divisions wasintended as an evolutionary scheme, butrather they were thought to reflect repeatedinvasions by outsiders with new ideas.Perhaps not surprisingly, a picture ofEurope conquered by invaders with superiortechnology had little appeal in the light of
two great European wars, and the trendin archaeology in the second half of thetwentieth century has been the study oflocal sequences and the application ofmodels of cultural evolution (Otte & Keeley,1990).
In terms of developments in world pre-history, however, Western Europe is aremote cul de sac with a somewhat anom-alous prehistoric record. We argue here thatmodels derived from the unique record ofEuropean prehistory do not explain eventsin Africa where the origin of modern peopleactually occurred. In the Holocene, westernEurope experienced a series of incursionsfrom the less peripheral portions of the OldWorld. Each arrival of a wave of invadersand alien technology induced a fairlysudden, rapid cultural turnover. Thesedisruptive episodes are reflected in theEuropean archaeological record as dis-continuities that punctuate industrialperiods of relatively long duration. Theyhave been sometimes described as ‘‘revol-utions,’’ such as the ‘‘neolithic revolution’’of Childe (1936, 1942).
Recent paleoclimatic data and refinedchronologies have supported the early sug-gestion of Howell (1951) that regions ofPleistocene Europe were repeatedly isolatedby ice and mountain barriers, so that itshominid populations were periodicallyreduced or even eliminated (Howell, 1952;Gamble, 1986, 1994; Jochim, 1987;Hublin, 1998a). Moreover, it has becomeincreasingly clear that the Neanderthalswere replaced by modern humans in Europewithin too short a period for the former tohave evolved into the latter (Mellars,1998a,b, 1999, Bocquet-Appel & Demars,2000). Thus, the ‘‘revolutionary’’ nature ofthe European Upper Paleolithic is mostprobably due to discontinuity in thearchaeological record rather than to the sortof rapid cultural, cognitive, and/or biologicaltransformation that has been argued byproponents of the ‘‘human revolution.’’

455 ’
The earliest modern Europeans wereAfricans
Who were the earliest modern Europeans? Itis becoming increasingly difficult to denythat they were Africans. Although the‘‘mitochondrial Eve’’ hypothesis, firstarticulated by Cann et al. (1987), has beenrevised in light of criticism (Templeton,1992; Hedges et al., 1992; Ayala, 1995), andpopulation size and structure have effects onthe distribution of genetic characters thatwere not taken into account in early recon-structions (Harpending et al., 1993, 1998;Sherry et al., 1994; Relethford, 1995;Relethford & Harpending, 1995), geneticdata either directly support or are consistentwith an African origin for modern humans(Wainscoat et al., 1986; Cann, 1988;Stringer & Andrews, 1988; Vigilant et al.,1991; Stoneking, 1993; Stoneking et al.,1993; Relethford & Harpending, 1994;Ayala, 1995; Nei, 1995; Goldstein, 1995;Tishkoff et al., 1996; Ruvolo, 1996, 1997;Irish, 1998; Pfeiffer, 1998; Zietkiweicz et al.,1997; Pritchard et al., 1999; Quintana-Murci, 1999; Relethford & Jorde, 1999;Tishkoff et al., 2000; see Relethford, 1998and Jorde et al., 1998 for recent reviews).As Howell (1994:306) observes, ‘‘Thephylogenetic roots of modern humans aredemonstrably in the Middle Pleistocene.The distribution of those antecedentpopulations appear to lie outside of westernand eastern Eurasia, and more probablycentered broadly on Africa.’’1
The fossil evidence for an African originfor modern humans is robust. It is clear thatmodern humans (H. sapiens sensu stricto)were certainly present in Africa by 130 ka(Day & Stringer, 1982; Deacon, 1989), andperhaps as early as 190 ka if specimenssuch as Singa are considered modern(McDermott et al., 1996; Stringer, 1996).Modern humans do not appear in Europe orCentral Asia before ca. 40 ka; earliest datesfor the Levant range between ca. 80 ka and120 ka (Day, 1969, 1972; Day & Springer,1982, 1991; Stringer, 1989, 1992;McBrearty, 1990b; Stringer et al., 1989;Brauer, 1984a,b, 1989; Stringer & Andrews,1988; Valladas et al., 1988; Grun &Stringer, 1991; Miller et al., 1991; Foley &Lahr, 1992; Mercier et al., 1993; Deacon,1993b; Brooks et al., 1993a,b; Stringer,1993a; Schwarcz, 1994; Straus, 1994;Bar-Yosef, 1994, 1995a, 1998; but seeHowells, 1989). Recent evidence suggeststhat modern humans were present inAustralia as early as 62 ka (Stringer, 1999;Thorne et al., 1999).
Although some, notably Brauer (1984a,b,1989), favor a scenario involving some inter-breeding among Neanderthal and modernhuman populations, the successful extrac-tion and analysis of fragmentary mito-chondrial DNA (mtDNA) from both theNeanderthal type fossil (Krings et al., 1997,1999) and additional material from thenorthern Caucasus (Ovchinnikov et al.,2000) appears to remove the Neanderthalsfrom modern human ancestry. Body propor-tions of early European H. sapiens fossilssuggest a tropical adaptation and support anAfrican origin (Holliday & Trinkaus, 1991;Ruff, 1994; Pearson, 1997, 2000; Holliday,1997, 1998, 2000). A single migration orpopulation bottleneck was originally envis-aged in the ‘‘African Eve hypothesis’’ (Cannet al., 1987), but a succession of populationdispersals, subsequent isolation induced byclimatic events and local adaptation maybetter account for the complexity of the
1. The Middle to Late Pleistocene boundary is thebeginning of the last interglacial, at approximately130 ka; the base of the Middle Pleistocene is the shiftfrom reversed to normal magnetic polarity at theMatuyama–Brunhes boundary, dated to about 780 ka(Butzer & Isaac, 1975; Imbrie & Imbrie, 1980; Bergeret al., 1984; Martinson et al., 1987; Shackleton et al.,1990; Deino & Potts, 1990; Cande & Kent, 1992;Baksi et al., 1992; Tauxe et al., 1992). Further evidencemay confirm recent suggestions (Schneider et al., 1992;Singer & Pringle, 1996; Hou et al., 2000) that the ageof this geomagnetic polarity reversal be revised toca. 790 ka.

456 . . .
fossil record and the genetic composition ofpresent human populations (Howells, 1976,1989, 1993; Boaz et al., 1982; Foley & Lahr,1992; Lahr & Foley, 1994, 1998; Ambrose,1998b).
It can be deduced from the archaeologicalevidence that on a continent-wide scale theAfrican record differs markedly from thatof Europe in its degree of population con-tinuity. While parts of Africa, such as theSahara or the interior of the Cape Provinceof South Africa, do appear to have experi-enced interruptions in human settle-ment during glacial maxima (Deacon &Thackeray, 1984; Williams, 1984; Butzer,1988b; Brooks & Robertshaw, 1990;Mitchell, 1990), climatic reconstructionssuggest that the contiguous expanse ofsteppe, savanna and woodland biomes avail-able for human occupation, especially in thetropical regions of the continent, was alwayssubstantially larger than the comparableregions in Europe. Perhaps as a result,hominid populations in Africa, while prob-ably widely dispersed, appear to havebeen consistently larger (Relethford &Harpending, 1995; Jorde et al., 1998;Relethford & Jorde, 1999; Tishkoff et al.,2000).
Revolution or evolution? The Africandata
How might the archaeological signature ofcontinuous evolutionary change be expectedto differ from that of abrupt replacement? Ifthe entire human species experienced a sim-ultaneous, punctuated, genetically encodedevent, such as the development of moderncapacities for language (Klein, 1995;Diamond, 1992), one would expect thetransition to modern human behavior to beabrupt, in Africa as well as in Europe andAsia. On the other hand, if aspects ofmodern human culture in Africa were devel-oped by hominids using existing cognitivecapabilities and transmitted by cultural
rather than by genetic processes, the mostlikely scenario would be an accretionaryprocess, a gradual accumulation of modernbehaviors in the African archaeologicalrecord (cf. Allsworth-Jones, 1993). Thischange need not be unidirectional or con-fined to a single location. Rather, we mightexpect innovative behaviors to appear atdifferent times and in different regions, anddue to low population densities we mightexpect the transmission of new ideas to besporadic.
As early as the 1920s it was clear that theAfrican archaeological record could not beaccommodated within the European Paleo-lithic model. A separate scheme of Earlier,Middle and Later Stone Ages (ESA, MSA,and LSA) was devised for Stone Age Africa(Goodwin & van Riet Lowe, 1929) toemphasize its distinctiveness from theLower, Middle, and Upper Paleolithic ofEurope. The ESA, MSA and LSA were firstdefined on technological grounds on thebasis of material from South Africa(Goodwin, 1928; Goodwin & van RietLowe, 1929). The terms were formallyendorsed by the Panafrican Congress of1955 (Clark, 1957a: xxxiii). The ESA as it isnow understood includes both the Oldowanand the Acheulian; the MSA encompassesflake and blade tool industries which ofteninclude prepared cores and points; and theLSA is characterized by microlithic tech-nology. The MSA was distinguished by thepresence of prepared core technology and,at most sites, unifacial and/or bifacial projec-tile points, and by the absence of handaxesand microliths, hallmarks of the Acheulianand LSA respectively.
Before 1972, in the absence of accuratechronometric dates, a radiocarbon date of60 ka from Acheulian levels at KalamboFalls, Zambia (Clark, 1969) was notrecognized as infinite. This frequently citeddate was particularly influential in establish-ing the impression of a short chronologyfor Africa. The MSA, at <60 ka, was

457 ’
considered the temporal equivalent of theUpper Paleolithic of Europe. Therefore thediscovery of anatomically modern humanremains associated with MSA artefacts atthe South African sites of Border Caveand Klasies River occasioned no surprise.2
The degree of regional differentiation, theubiquitous presence of blades and bladecores, and the sophistication of projectilepoint technology in the African MSA wereconsidered comparable to the EuropeanUpper Paleolithic. However, bone tools, artobjects and beads were sparse when com-pared to the European Upper Paleolithic,particularly the late Upper Paleolithic.
The rarity of elements regarded as criticalto modern human culture in the MSAserved as grounds for regarding Africa as a‘‘cultural backwater,’’ the place that initiallygave rise to humanity, but failed to nurtureits later development (e.g., Butzer, 1971;cf. Clark, 1975). In the later 1970s, newdating techniques and more accurateclimatic correlations pushed back the age ofthe MSA well beyond 100 ka. The MSA wasrecognized as the temporal equivalent of theEuropean Middle Paleolithic, not the UpperPaleolithic. Attention focused on the humanfossils associated with the MSA, which werenow thought to be anomalously modernin appearance. The fact that many MSAartefacts recalled the Upper Paleolithic ofEurope in both form and technology wasforgotten.
The use of a classifactory schemedesigned for Africa did not entirely removeambiguity, as many industries displayedcharacteristics of two different stages andcould not be assigned to one of the threedivisions. Long transitional periods or‘‘Intermediates’’ were added to the tripartiteESA–MSA–LSA scheme at the 1955Panafrican Congress (Clark, 1957a: xxxiii),but the ‘‘Intermediate’’ concept was subse-
quently retired (Bishop & Clark, 1967:987), when a mixture of different occu-pation levels was found to have occurredduring excavation at the ‘‘Second Inter-mediate’’ type site of Magosi (Wayland &Burkitt, 1932; Clark, 1957b; Hole, 1959;Cole, 1967). Yet anachronisms, as well aslong periods of transition between stages,remain as problems (Vishnyatsky, 1994).
A fairly abrupt MSA–LSA transition isapparent in the Mediterranean zones at thenorthern and southern margins of Africa.This seems consistent with the significantdocumented gaps in the settlement historyof both regions (Close et al., 1990; Wendorfet al., 1990, 1993a; Mitchell, 1990; Deacon& Thackeray, 1984; Klein, 1989b: 307).However, at rock shelter sites in tropicalAfrica with relatively continuous occu-pational records, such as Mumba, Tanzania(Mehlman, 1979, 1989), Matupi, D. R.Congo (van Noten, 1977) and WhitePaintings, Botswana (Robbins & Murphy,1998; Robbins et al., under review) there is agradual transition from MSA to LSA tech-nology over as much as 30 ka. Mehlman(1991) has urged the development of newparadigms to accommodate the lack of apunctuated event.
Because of the late, sudden, and nearlysimultaneous appearance in Europe of mod-ern humans and complex behavior, archae-ologists working in Africa have sought asimilar ‘‘human revolution’’ there. The fullydeveloped signature of modern humanbehavior, including planning, sophisticatedtechnology and resource use, and symbolicbehavior in the form of decorative art isclearly present in the African LSA. As a re-sult, the MSA–LSA transition has been con-flated with the Middle to Upper Paleolithicand the emergence of modern human behav-ior. Consequently the earliest anatomicallymodern humans, which occur in MSA con-texts, are not accepted as fully ‘‘human’’.
We suggest that the expectation of a‘‘human revolution’’ in Africa is ultimately
2. The name Klasies River rather than Klasies RiverMouth or KRM is adopted here to conform with therecent usage of Hilary Deacon and his team.

458 . . .
a misapplication of a European model.Further, we reject the idea of a time lagbetween anatomical and behavioral changein Africa, such as that proposed by Klein(1992, 1994, 1995, 1998). There was no‘‘human revolution’’ in Africa. Rather, inthis paper we present data from the humanfossil and archaeological records to showthat novel features accrued stepwise.Distinct elements of the social, economic,and subsistence bases changed at differentrates and appeared at different times andplaces. We describe evidence from theAfrican MSA to support the contention thatboth human anatomy and human behaviorwere intermittently transformed from anarchaic to a more modern pattern over aperiod of more than 200,000 years.
The hominid fossil record
Until recently, most reconstructions of laterhuman phylogeny recognized only onespecies after H. erectus. Grade-basedschemes commonly divided H. sapiens intotwo variants, ‘‘archaic’’ H. sapiens and ‘‘ana-tomically modern’’ H. sapiens (H. sapienssensu stricto). The Neanderthals were thensometimes distinguished from other‘‘archaic’’ H. sapiens at the subspecificlevel as H. sapiens neanderthalensis (e.g.,Campbell, 1964). We concur with suchauthors as Tattersall (1986, 1992), Kimbel(1991), Harrison (1993), Rak (1993) andStringer (1994, 1996) that there are groundsfor distinguishing ‘‘archaic’’ from ‘‘modern’’H. sapiens at the species level, and thuswe regard the appearance of ‘‘modern’’ H.sapiens as a speciation event. Here, we treatthe Neanderthals as the distinct species H.neandertalensis King, 1864, and use thename H. sapiens to refer only to H. sapienssenus stricto. The use of the ‘‘anatomicallymodern’’ label for H. sapiens sensu stricto isnot only unnecessary but also misleading, asmany of the cranial features used to dis-tinguish H. sapiens are in fact primitive for
the genus Homo when compared to the morederived state in the Neanderthals (Rak,1993).
Specimens formerly attributed to‘‘archaic’’ H. sapiens exhibit a number ofplesiomorphic traits, including long lowcrania, large brow ridges, large, prognathicfaces with large teeth, and the lack of a chin.The chief justification for the inclusion ofthese fossils in our species has been theirlarge brain sizes, though brain size is in parta function of body mass, known to be quitelarge among these hominids (Grine et al.,1995; Ruff et al., 1997; Kappelman, 1997).Recent discussions of later hominid phy-logeny (e.g., Stringer, 1993b, 1994, 1995,1996; Lahr & Foley, 1994; Foley & Lahr,1997; Rightmire, 1998) have recognized thedistinctiveness of non-Neanderthal MiddlePleistocene hominids and have resurrectedthe taxon H. heidelbergensis Schoetensack,1908 for them, but we question theattribution of the African material to thistaxon.
Paradoxically, H. sapiens Linnaeus, 1758lacks a satisfactory definition. Howell (1978:201) observed over 20 years ago,
‘‘The extensive relevant literature reveals anunexpected lack of concern with the bio-logical distinctiveness of a now-dominantmammalian species’’,
and the situation is virtually unchangedtoday. The anatomy of H. sapiens is charac-terized by a high round cranium, a chin, asmall orthognathic face, as well as reducedmasticatory apparatus and brow ridges. Ithas been argued that most of these featurescan be explained by greater flexion in thebasicranium of H. sapiens (Lieberman,1998b; Spoor et al., 1999).
Because early fossils of H. sapiens datingto 130 ka, and perhaps as early as 190 ka,are found in Africa (Grun et al., 1990;Deacon, 1989, 1993b; Day & Stringer,1982; McDermott et al., 1996), it is reason-able to seek evidence for the processes lead-ing to the origin of H. sapiens in the African

459 ’
record of the Middle Pleistocene (Howell,1994). Although often described as‘‘scrappy’’ or insubstantial, the Africanhominid fossil sample from this time periodnumbers several dozen individuals (Table 1,Figure 1). While the circumstances ofrecovery for some of the specimens are farfrom ideal, this is unfortunately true formany fossil discoveries, and in fact a fairnumber of the African specimens wererecovered by controlled excavation (e.g.,Ndutu, Cave of Hearths, Haua Fteah,Mumba, Ngaloba, Klasies, Kapthurin post-cranials). For others, stratigraphic contextcan be reasonably inferred, despite the factthat they are surface finds (e.g., Kapthurinmandibles, Eyasi, Buia).
Howell (1994: 305f) has deemed the sol-ution of the evolutionary relationshipsamong later Middle Pleistocene hominidpopulations one of the central problems inthe study of human evolution, and the taxo-nomic status of the African fossils is muchdebated (e.g., Tattersall, 1986; Clarke,1990; Foley, 1991a; Kimbel, 1991;Stringer, 1992, 1993a, 1994, 1996; Aiello,1993; Foley & Lahr, 1997; Lahr & Foley,1998; Rightmire, 1998). Revision of fossilsascribed to Homo (Wood, 1991, 1992;Wood & Collard, 1999) has resulted in amore ‘‘bushy’’ or speciose taxonomicpicture for our genus in the Pliocene andEarly Pleistocene, but for the AfricanMiddle and Late Pleistocene a unilinealmodel is often invoked. It is our belief thatthe number of African Middle and LatePleistocene hominid species has been under-estimated, because behavioral and repro-ductive isolation may precede changes in thebony skeleton (Tattersall, 1986, 1992,1993; Rak, 1993).
Over 25 years ago, Day (1973) suggestedseparating African Middle and LatePleistocene hominids into ‘‘early,’’ ‘‘inter-mediate,’’ and ‘‘modern’’ groups, and thisgrade-based practice has been followed,explicitly and implicitly, in many subsequent
works (Brauer, 1984a,b, 1989; Smith, 1985,1993; Clark, 1988; Klein, 1989b, 1994;Stringer, 1993a). Data in Table 1 roughlyfollow Day’s tripartite construct, though thisshould not be construed as an endorsementfor anagenesis or a grade-based taxonomy.Both the ascription of fossils to group andthe attachment of taxonomic labels areproblematic, and Group 1 specimensprobably belong to several different species(e.g., H. louisleakeyi, H. rhodesiensis).
The principal unresolved issue in theclarification of the evolutionary relationshipsof the hominids in Group 1 is the enigmaticstatus of H. erectus. This species is believedby many to have been confined to Asia(Andrews, 1984; Tattersall, 1986; Groves,1989; Clarke, 1990; Kimbel, 1991; Larick &Ciochon, 1996). Following Wood (1991,1992), some now ascribe to H. ergasterAfrican fossils in the 1·5–2 Ma age rangeformerly attributed to H. erectus. Otherauthors (e.g., Rightmire, 1990, 1994, 1995,1998; Brauer & Mbua, 1992; Harrison,1993; Walker, 1993; Brauer, 1994) regardH. erectus as a single polytypic species dis-tributed throughout most of the Old World,and African specimens in our Group 1,spanning a broad range of time, continueto be ascribed to this taxon (e.g., OH9,Kapthurin, in Wood, 1992). Attribution ofthe African and Asian Middle Pleistocenematerial to a single species assumes anadequate degree of gene flow to preventspeciation, but the archaeological differ-ences between the regions suggest long termisolation (Schick, 1994; but see Hou et al.,2000).
Group 1 in our scheme includes theKabwe (Broken Hill) cranium, type speci-men of H. rhodesiensis Woodward, 1921, aswell as OH9. The latter specimen is usuallyreferred in the literature to H. erectus, butLouis Leakey (1961, 1963) emphaticallyrejected this position. He saw the origin ofH. sapiens as a strictly African phenom-enon, and regarded OH9 as morphologically

460 . . .
Tab
le1
Lat
erA
fric
anH
omin
idae
,th
eir
arch
aeol
ogic
alas
soci
atio
ns
and
dat
es
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
Gro
up
1(H
.er
ectu
s,H
.er
gast
er,
H.
lou
isle
akey
i,H
.rh
odes
ien
sis)
Aın
Maa
rou
f(E
lH
ajeb
),M
oroc
coL
eft
fem
oral
shaf
tA
cheu
lian
Ear
lyM
idd
leP
leis
toce
ne
Ass
ocia
ted
faun
aH
ublin
,19
92;
Ger
aads
etal
.,19
92
Ber
gA
uka
s,N
amib
iaF
emor
alfr
agm
ent
Non
eU
ndat
edG
rine
etal
.,19
95
Bod
o,E
thio
pia
Adu
ltcr
aniu
m,
pari
etal
,di
stal
hum
erus
Ach
eulia
nM
idto
late
rM
iddl
eP
leis
toce
ne,
350
kaA
ssoc
iate
dfa
una
Con
roy
etal
.,19
78;
Kal
bet
al.,
1980
,19
82a,
b;A
sfaw
,19
83;
Cla
rket
al.,
1984
;R
ight
mir
e,19
96
cf.
Old
owan
640
ka–5
50ka
40A
r/39A
r,as
soci
ated
faun
aC
lark
etal
.,19
94
Bu
ia,
Dan
akil
(Afa
r)D
epre
ssio
n,E
ritr
eaA
dult
cran
ium
,2
inci
sors
,pe
lvic
frag
men
ts
Non
ere
port
ed1·
0M
aP
aleo
mag
neti
sm,
asso
ciat
edfa
una
Abb
ate
etal
.,19
98
Cav
eof
Hea
rth
s,S
outh
Afr
ica
Man
dibl
e,ra
dius
Ach
eulia
nE
arly
Lat
eP
leis
toce
neA
ssoc
iate
dfa
una
Coo
ke,
1962
;M
ason
,19
62;
Mas
onet
al.,
1988
;T
obia
s,19
71;
Par
trid
ge,
1982
;P
ears
on&
Gri
ne,
1997
En
dM
idd
leP
leis
toce
ne
Ass
ocia
ted
faun
aH
owel
l,19
78
Eya
si,
Tan
zani
aC
rani
alfr
agm
ents
repr
esen
ting
3–4
indi
vidu
als
San
goan
>13
0ka
Ext
rapo
lati
onfr
omov
erly
ing
14C
date
s,un
derl
ying
230T
h/234U
date
s,fa
unal
corr
elat
ion
Coo
ke,
1963
;M
ehlm
an,
1984
,19
87

461 ’
Tab
le1,
Gro
up
1C
onti
nu
ed
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
Kab
we
(Bro
ken
Hill
),Z
ambi
a(h
olot
ype
ofH
.rh
odes
ien
sis)
Adu
ltcr
aniu
m(E
686)
,cr
ania
l,m
axill
ary
dent
alan
dpo
stcr
ania
l(h
umer
al,
pelv
ic,
fem
oral
,ti
bial
)re
mai
nsof�
3in
divi
dual
s
?San
goan
orM
SA
?12
5ka
Ass
ocia
ted
faun
aW
oodw
ard,
1921
;P
ycra
ftet
al.,
1928
;O
akle
y,19
57;
Cla
rk,
1959
;C
lark
etal
.,19
50,
1968
;K
lein
,19
73,
1994
;S
anta
Luc
a,19
78;
Par
trid
ge,
1982
;V
rba,
1982
;S
trin
ger,
1986
110
kaA
spar
tic
acid
race
miz
atio
non
hom
inid
fem
oral
frag
men
tE
M79
3
Bad
aet
al.,
1974
700–
400
kaA
ssoc
iate
dfa
una
(cf.
Old
uvai
Bed
sII
I–IV
)K
lein
,19
94;
Rig
htm
ire,
1998
780
ka–1
·33
Ma
Ass
ocia
ted
faun
a(c
f.O
lduv
aiB
eds
III–
IV).
Cor
rela
tion
ofto
pof
Bed
IVw
ith
Mat
uyam
aB
runh
esbo
unda
ry
Thi
spa
per,
base
dup
onK
lein
,19
73,
1994
;P
artr
idge
,19
82;
Hay
,19
76;
Wal
ter
etal
.,19
91,
1992
;T
amra
tet
al.,
1995
;K
imbe
l,19
95;
Del
son
&va
nC
ouve
ring
,20
00
1·07
–1·3
3M
aA
ssoc
iate
dfa
una
(cf.
Old
uvai
Bed
sII
I–IV
).C
orre
lati
onof
norm
alpo
lari
typa
leom
agne
tic
zone
atba
seof
Mas
ekB
eds
wit
hJa
ram
illo
subc
hron
Thi
spa
per,
base
dup
onK
lein
,19
73,
1994
;P
artr
idge
,19
82;
Hay
,19
76;
Wal
ter
etal
.,19
91,
1992
;T
amra
tet
al.,
1995
;K
imbe
l,19
95;
Del
son
&va
nC
ouve
ring
,20
00

462 . . .
Tab
le1,
Gro
up
1C
onti
nu
ed
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
Kap
thu
rin
(Bar
ingo
)K
enya
Tw
oad
ult
man
dibl
es,
(KN
M-B
K67
,85
18)
post
cran
ials
(uln
a,ta
lus,
man
usph
alan
ges,
KN
M-B
K63
–66)
Und
iagn
osti
c23
0–78
0ka
K/A
r,as
soci
ated
faun
a,pa
leom
agne
tism
Lea
key
etal
.,19
69;
van
Not
en,
1982
;H
owel
l,19
82;
van
Not
en&
Woo
d,19
85;
Woo
d&
van
Not
en,
1986
;T
allo
n,19
78;
Dag
ley
etal
.,19
78;
Rig
htm
ire,
1980
;S
olan
&D
ay,
1992
;W
ood,
1992
;G
rove
s,19
98
500–
550
kaD
eino
&M
cBre
arty
,un
der
revi
ew
Keb
ibat
(Rab
at),
Mor
occo
Sub
adul
tca
lvar
ia,
max
illar
yfr
agm
ent,
man
dibl
e
Non
e>
200
ka230T
h/234U
Ste
arns
&T
hurb
er,
1965
300
ka–1
·0M
aA
ssoc
iate
dfa
una
Sab
an,
1975
,19
77;
How
ell,
1978
;S
auss
e,19
75b
Lai
nya
mok
,K
enya
Fem
oral
shaf
t,is
olat
edte
eth
Und
iagn
osti
c39
0–33
0ka
40A
r/39A
rS
hipm
anet
al.,
1983
;P
otts
etal
.,19
88;
Pot
ts&
Dei
no,
1995
Loy
anga
lan
iM
axill
ary
and
man
dibu
lar
dent
itio
nN
one
Lat
eM
idd
le/E
arly
Lat
eP
leis
toce
ne
Ass
ocia
ted
faun
aT
wie
ssel
man
n,19
91
Mel
kaK
ontu
re,
Eth
iopi
aC
rani
alfr
agm
ents
Ach
eulia
nM
idd
leP
leis
toce
ne
Ass
ocia
ted
faun
aC
hava
illon
etal
.,19
74;
How
ell,
1978
;C
hava
illon
,19
82
Nd
utu
,T
anza
nia
Adu
ltcr
aniu
mcf
.A
cheu
lian
500–
600
kaA
AR
onas
soci
ated
mam
mal
ian
bone
Mtu
ri,
1976
;R
ight
mir
e,19
80,
1983
;C
lark
e,19
76,
1990

463 ’
Table
1,G
rou
p1
Con
tin
ued
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
Nd
utu
,T
anza
nia
cont
inue
d49
0–78
0ka
Upp
erlim
it:
corr
elat
ion
ofre
vers
edpo
lari
tyex
curs
ion
inun
derl
ying
Nor
kilil
iM
embe
rof
Mas
ekB
eds
wit
hE
mpe
ror
subc
hron
.L
ower
limit
:co
rrel
atio
nof
top
ofB
edIV
wit
hM
atuy
ama
Bru
nhes
boun
dary
Thi
spa
per,
base
dup
onT
amra
tet
al.,
1995
370–
990
kaU
pper
limit
:40A
r/39A
ron
Ker
imas
ica
lder
a,pr
obab
leso
urce
ofN
orki
lili
Mem
ber
ofM
asek
Bed
s.L
ower
limit
:co
rrel
atio
nof
norm
alpo
lari
typa
leom
agne
tic
zone
atba
seof
Mas
ekB
eds
wit
hJa
ram
illo
subc
hron
Thi
spa
per,
base
dup
onH
ay,
1976
,19
90;
Lea
key
etal
.,19
72;
Lea
key
&H
ay,
1982
;C
lark
e,19
76,
1990
;M
aneg
a,19
93;
Tam
rat
etal
.,19
95;
Wal
ter
etal
.,19
91,
1992
;K
imbe
l,19
95
OH
9,L
LK
,O
lduv
aiG
orge
,T
anza
nia
(typ
eof
H.
lou
isle
akey
i)
Adu
ltpa
rtia
lcr
aniu
mD
evel
oped
Old
owan
orA
cheu
lian
1·15
Ma
(sur
face
,to
pof
Bed
II)
K/A
r,pa
leom
agne
tism
Hay
,19
63,
1973
,19
76,
1990
;L
eake
y,19
61,
1963
;L
eake
y,19
71a,
b;L
eake
y&
Hay
,19
82;
Rig
htm
ire,
1979
a,19
80,
1990
,19
94;
Woo
d,19
94
0·7–
1·0
Ma
K/A
r,pa
leom
agne
tism
Hay
,19
71
1·33
–1·4
8M
a40A
r/39A
r,pa
leom
agne
tism
Hay
,19
76,
1990
;M
aneg
a,19
93;
Tam
rat
etal
.,19
95;
Wal
ter
etal
.,19
91,
1992
;K
imbe
l,19
95;
Whi
te,
2000

464 . . .
Tab
le1,
Gro
up
1C
onti
nu
ed
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
OH
11,
DK
,O
lduv
aiG
orge
,T
anza
nia
Pal
ate,
max
illa
Non
e;bo
thA
cheu
lian
&M
SA
else
whe
rein
Ndu
tuB
eds
ca.4
00ka
(sur
face
,pr
obab
lylo
wer
Ndu
tuB
eds)
K/A
r,pa
leom
agne
tism
infe
rred
sedi
men
tati
onra
tes
Rig
htm
ire,
1979
a;L
eake
y&
Hay
,19
82;
Hay
,19
94
ca.
490
kaP
aleo
mag
neti
sm;
corr
elat
ion
ofre
vers
edpo
lari
tyex
curs
ion
inun
derl
ying
Nor
kilil
iM
embe
rof
Mas
ekB
eds
wit
hE
mpe
ror
subc
hron
.
Thi
spa
per,
base
dup
onT
amra
tet
al.,
1995
ca.
370
ka40A
r/39A
ron
Ker
imas
ica
lder
a,pr
obab
leso
urce
ofN
orki
lili
Mem
ber
ofM
asek
Bed
s.
Hay
,19
76,
1990
;M
aneg
a,19
93;
Tam
rat
etal
.,19
95;
Wal
ter
etal
.,19
91,
1992
;K
imbe
l,19
95;
Whi
te,
2000
;D
elso
n&
van
Cou
veri
ng,
2000
OH
12,
VE
K,
Old
uvai
Gor
ge,
Tan
zani
aP
alat
e,m
axill
a,cr
ania
lfr
agm
ents
Non
e;A
cheu
lian
else
whe
rein
Bed
IV78
0–62
0ka
(upp
erB
edIV
)K
/Ar,
pale
omag
neti
sm,
infe
rred
sedi
men
tati
onra
tes
Rig
htm
ire,
1979
a;L
eake
y&
Hay
,19
82;
Hay
,19
94;
Lea
key
&R
oe,
1994
780
ka–1
·2M
aP
aleo
mag
neti
sm;
corr
elat
ion
ofto
pof
Bed
IVw
ith
Mat
uyam
aB
runh
esbo
unda
ry
Thi
spa
per,
base
dup
onT
amra
tet
al.,
1995
1·07
–1·2
Ma
40A
r/39A
rpa
leom
agne
tism
;co
rrel
atio
nof
norm
alpo
lari
typa
leom
agne
tic
zone
atba
seof
Mas
ekB
eds
wit
hJa
ram
illo
subc
hron
Hay
,19
76,
1990
;M
aneg
a,19
93;
Tam
rat
etal
.,19
95;
Wal
ter
etal
.,19
91,
1992
;K
imbe
l,19
95;
Whi
te,
2000
;D
elso
n&
van
Cou
veri
ng,
2000

465 ’
Table
1,G
rou
p1
Con
tin
ued
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
OH
22,
VE
K/M
NK
,O
lduv
aiG
orge
,T
anza
nia
Par
tial
man
dibl
eN
one;
Ach
eulia
nel
sew
here
inB
edIV
and
over
lyin
gN
dutu
Bed
s
800–
600
ka(s
urfa
ce,
Bed
IV)
K/A
r,pa
leom
agne
tism
,in
ferr
edse
dim
enta
tion
rate
s
Day
,19
86;
Lea
key
&H
ay,
1982
;L
eake
y&
Roe
,19
94;
Rig
htm
ire,
1979
a,19
90;
Hay
,19
94
780
ka–1
·2M
aP
aleo
mag
neti
sm;
corr
elat
ion
ofto
pof
Bed
IVw
ith
Mat
uyam
aB
runh
esbo
unda
ry
Thi
spa
per,
base
dup
onT
amra
tet
al.,
1995
1·07
–1·2
Ma
40A
r/39A
r,pa
leom
agne
tism
;co
rrel
atio
nof
norm
alpo
lari
typa
leom
agne
tic
zone
atba
seof
Mas
ekB
eds
wit
hJa
ram
illo
subc
hron
Hay
,19
76,
1990
;M
aneg
a,19
93;
Tam
rat
etal
.,19
95;
Wal
ter
etal
.,19
91,
1992
;K
imbe
l,19
95;
Whi
te,
2000
;D
elso
n&
van
Cou
veri
ng,
2000
OH
23,
FL
K,
Old
uvai
Gor
ge,
Tan
zani
aM
andi
bula
rfr
agm
ent
Ach
eulia
n40
0–60
0ka
(In
situ
,M
asek
Bed
s)K
/Ar,
pale
omag
neti
sm,
infe
rred
sedi
men
tati
onra
tes
Day
,19
86;
Rig
htm
ire,
1990
;L
eake
y&
Roe
,19
94;
Hay
,19
94
490–
780
kaP
aleo
mag
neti
sm.
Upp
erlim
it:
corr
elat
ion
ofre
vers
edpo
lari
tyex
curs
ion
inun
derl
ying
Nor
kilil
iM
embe
rof
Mas
ekB
eds
wit
hE
mpe
ror
subc
hron
.L
ower
limit
:co
rrel
atio
nof
top
ofB
edIV
wit
hM
atuy
ama
Bru
nhes
boun
dary
Thi
spa
per,
base
dup
onT
amra
tet
al.,
1995

466 . . .
Table
1,G
rou
p1
Con
tin
ued
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
OH
23,
cont
inue
d99
0–37
0ka
Upp
erlim
it:
40A
r/39A
ron
Ker
imas
ica
lder
a,pr
obab
leso
urce
ofN
orki
lili
Mem
ber
ofM
asek
Bed
s.L
ower
limit
:co
rrel
atio
nof
norm
alpo
lari
typa
leom
agne
tic
zone
atba
seof
Mas
ekB
eds
wit
hJa
ram
illo
subc
hron
Hay
,19
76,
1990
;M
aneg
a19
93;
Tam
rat
etal
.,19
95;
Wal
ter
etal
.,19
91,
1992
;K
imbe
l,19
95;
Whi
te,
2000
;D
elso
n&
van
Cou
veri
ng,
2000
OH
28,
WK
,O
lduv
aiG
orge
,T
anza
nia
Lef
tin
nom
inat
e,fe
mur
Ach
eulia
n78
0–62
0ka
(upp
erB
edIV
)K
/Ar,
pale
omag
neti
sm,
infe
rred
sedi
men
tati
onra
tes
Day
,19
71,
1986
;L
eake
y,19
71a;
Rig
htm
ire,
1979
a;L
eake
y&
Hay
,19
82;
Hay
,19
94
780
ka–1
·2M
aP
aleo
mag
neti
sm;
corr
elat
ion
ofto
pof
Bed
IVw
ith
Mat
uyam
aB
runh
esbo
unda
ry
Thi
spa
per,
base
dup
onT
amra
tet
al.,
1995
1·2–
1·07
Ma
40A
r/39A
r,pa
leom
agne
tism
;co
rrel
atio
nof
norm
alpo
lari
typa
leom
agne
tic
zone
atba
seof
Mas
ekB
eds
wit
hJa
ram
illo
subc
hron
Hay
,19
76,
1990
;M
aneg
a,19
93;
Tam
rat
etal
.,19
95;
Wal
ter
etal
.,19
91,
1992
;K
imbe
l,19
95;
Whi
te,
2000
;D
elso
n&
van
Cou
veri
ng,
2000
OH
34,
JK,
Old
uvai
Gor
ge,
Tan
zani
aF
emur
and
part
ial
tibi
aA
cheu
lian
0·8–
1·1
Ma
(Bed
III)
K/A
r,pa
leom
agne
tism
Day
,19
71;
Lea
key
&R
oe,
1994
;H
ay,
1990
,19
94
1·2–
1·33
Ma
40A
r/39A
r,pa
leom
agne
tism
Thi
spa
per,
base
dup
onH
ay,
1976
,19
90;
Tam
rat
etal
.,19
95;
Wal
ter
etal
.,19
91,
1992
;K
imbe
l,19
95;
Whi
te,
2000

467 ’
Table
1,G
rou
p1
Con
tin
ued
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
OH
51,
GT
C,
Old
uvai
Gor
ge,
Tan
zani
a
Man
dibu
lar
frag
men
tN
one;
Ach
eulia
nel
sew
here
inB
edII
I0·
8–1·
1M
a(B
edII
I)K
/Ar,
pale
omag
neti
smL
eake
y&
Roe
,19
94;
Rig
htm
ire,
1990
;H
ay,
1990
,19
94
1·2–
1·33
Ma
40A
r/39A
r,pa
leom
agne
tism
Thi
spa
per,
base
dup
onH
ay,
1976
,19
90;
Tam
rat
etal
.,19
95;
Wal
ter
etal
.,19
91,
1992
;K
imbe
l,19
95;
Whi
te,
2000
Sal
dan
ha
(Hop
efiel
dE
land
sfon
tein
),S
outh
Afr
ica
Adu
ltca
lvar
ia,
man
dibu
lar
frag
men
tA
cheu
lian
ca.1
30–7
80ka
(Mid
dle
Ple
isto
cene
)A
ssoc
iate
dfa
una
Dre
nnan
,19
53;
Sin
ger,
1954
;C
ooke
,19
63;
Par
trid
ge,
1982
;D
eaco
n,19
88
600–
800
kaA
ssoc
iate
dfa
una,
cfB
edIV
Old
uvai
Gor
geK
lein
,19
73;
Sin
ger
&W
ymer
,19
68;
Lea
key
&H
ay,
1982
;H
ay,
1994
;L
eake
y&
Roe
,19
94
500–
200
kaA
ssoc
iate
dfa
una
Kle
in,
1988
700–
400
kaA
ssoc
iate
dfa
una
Kle
in&
Cru
z-U
ribe
,19
91;
Kle
in,
1994
;R
ight
mir
e,19
98
780
ka–1
·2M
aA
ssoc
iate
dfa
una
(cf
Old
uvai
Bed
IV).
Cor
rela
tion
ofto
pof
Bed
IVw
ith
Mat
uyam
aB
runh
esbo
unda
ry
Thi
spa
per,
base
dup
onG
entr
y,19
78;
Kle
in&
Cru
zU
ribe
,19
91;
Whi
te,
2000
;T
amra
tet
al.,
1995
;W
alte
ret
al.,
1991
,19
92;
Kim
bel,
1995

468 . . .
Table
1,G
rou
p1
Con
tin
ued
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
Sal
dan
ha,
cont
inue
d1·
07–1
·33
Ass
ocia
ted
faun
a(c
fO
lduv
aiB
edIV
).C
orre
lati
onof
norm
alpo
lari
typa
leom
agne
tic
zone
atba
seof
Mas
ekB
eds
wit
hJa
ram
illo
subc
hron
Thi
spa
per,
base
dup
onG
entr
y,19
78;
Kle
in&
Cru
zU
ribe
,19
91;
Whi
te,
1999
;T
amra
tet
al.,
1995
;W
alte
ret
al.,
1991
,19
92;
Kim
bel,
1995
Sal
e,M
oroc
coA
dult
calv
aria
,cr
ania
lfr
agm
ents
,en
doca
stN
one
300
ka–1
·0M
aA
ssoc
iate
dfa
una
Jaeg
er,
1973
,19
75;
How
ell,
1978
;H
ublin
,19
85,
1991
,19
94;
Dea
net
al.,
1993
389–
455
kaL
UE
SR
onas
soci
ated
bovi
dto
oth
enam
elH
ublin
,19
91
Sid
iA
bd
erra
hm
an(C
asab
lanc
a)M
oroc
coP
arti
alm
andi
ble
Ach
eulia
nM
idd
leP
leis
toce
ne
(‘‘T
ensi
ftia
n’’=
‘‘R
iss’
’)G
eom
orph
olog
y,as
soci
ated
faun
aA
ram
bour
g&
Bib
erso
n,19
56;
Bib
erso
n,19
63;
How
ell,
1960
,19
78
Tig
hen
if(f
orm
erly
Ter
nifin
e),
Ora
n,A
lger
ia
Thr
eem
andi
bles
,pa
riet
alfr
agm
ent
Ach
eulia
nM
iddl
eP
leis
toce
neA
ssoc
iate
dfa
una
Ara
mbo
urg,
1955
;A
ram
bour
g&
Hoff
stet
er,
1963
;H
owel
l,19
60,
1978
;B
alou
tet
al.,
1967
;T
obia
s,19
68;
Sch
war
tz&
Tat
ters
all,
2000
0·6–
1·0
Ma
Ass
ocia
ted
faun
aG
eraa
ds,
1981
;Ja
eger
,19
81
700
kaA
ssoc
iate
dfa
una,
pale
omag
neti
smG
eraa
dset
al.,
1986
Th
omas
1Q
uar
ry,
Mor
occo
Sub
adul
tpa
rtia
lm
andi
ble,
cran
ial
and
max
illar
yfr
agm
ents
Non
eM
idd
leP
leis
toce
ne
(‘‘A
mir
ian’
’)A
ssoc
iate
dfa
una,
geom
orph
olog
yE
nnou
chi,
1969
a,19
70;
Sau
sse,
1975
b;O
akle
yet
al.,
1977
;H
owel
l,19
60,
1978
;B
raue
r,19
84a,
b;D
ean
etal
.,19
93

469 ’
Table
1,G
rou
p1
Con
tin
ued
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
Wad
iD
agad
le,
Djib
outi
Max
illa,
part
ial
dent
itio
nN
one
<25
0ka
TL
onba
salt
,as
soci
ated
faun
ade
Bon
iset
al.,
1984
,19
88
Gro
up
2(H
.h
elm
eior
H.
sap
ien
s)E
liye
Sp
rin
gs,
Wes
tT
urka
na,
Ken
yaA
dult
cran
ium
ES
1169
3N
one
Non
eS
urfa
cefin
dB
raue
r&
Lea
key,
1986
Flo
risb
ad,
Sou
thA
fric
aA
dult
cran
ium
?MS
A?
Infin
ite
AA
R,
14C
Vog
el&
Bea
umon
t,19
72;
Bad
aet
al.,
1973
>10
0ka
230T
h/234U
,pe
atI,
asso
ciat
edfa
una
Dre
yer,
1935
,19
36;
Rig
htm
ire,
1978
a;C
lark
e,19
85;
Kum
an&
Cla
rke,
1986
;B
rink
,19
88;
But
zer,
1988
a;K
uman
etal
.,20
00
260
kaE
SR
,di
rect
assa
yon
hom
inid
toot
hG
run
etal
.,19
96
Gu
omd
e,C
hari
Fm
,Il
eret
,K
enya
Adu
ltcr
aniu
m(K
NM
-ER
3884
),fe
mur
(KN
M-E
R99
9)
Non
e27
0–30
0ka
U-s
erie
s,di
rect
assa
yson
hom
inid
cran
ium
&fe
mur
Bra
uer
etal
.,19
92,
1997
;F
eibe
let
al.,
1989
Hau
aF
teah
,L
ibya
Tw
oyo
ung
adul
tm
andi
bula
rfr
agm
ents
MS
A(‘
‘Lev
allo
iso-
Mou
ster
ian’
’)In
finit
e14C
onbu
rnt
bone
McB
urne
y,19
61,
1967
;T
obia
s,19
67;
Rak
,19
98:3
64
>13
0A
ssoc
iate
dar
tefa
cts
(Gen
eral
ized
MS
Ais
stra
tifie
dun
der
Ate
rian
else
whe
rein
Sah
ara)
Thi
spa
per,
base
dup
onD
eben
ath,
1994
;W
endo
rfet
al.,
1987
,19
93a
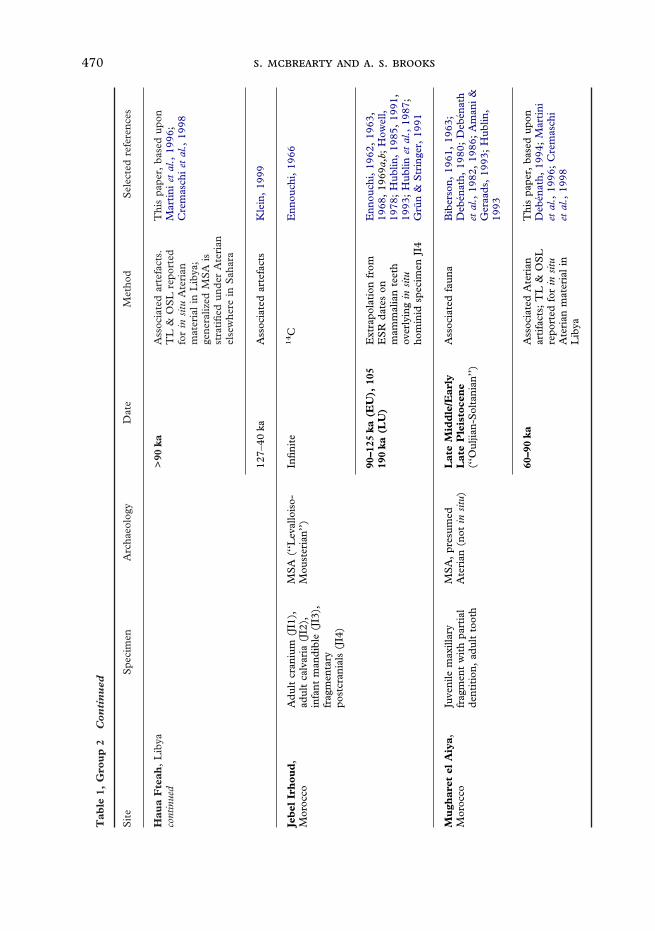
470 . . .
Tab
le1,
Gro
up
2C
onti
nu
ed
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
Hau
aF
teah
,L
ibya
cont
inue
d>
90ka
Ass
ocia
ted
arte
fact
s.T
L&
OS
Lre
port
edfo
rin
situ
Ate
rian
mat
eria
lin
Lib
ya;
gene
raliz
edM
SA
isst
rati
fied
unde
rA
teri
anel
sew
here
inS
ahar
a
Thi
spa
per,
base
dup
onM
arti
niet
al.,
1996
;C
rem
asch
iet
al.,
1998
127–
40ka
Ass
ocia
ted
arte
fact
sK
lein
,19
99
Jeb
elIr
hou
d,
Mor
occo
Adu
ltcr
aniu
m(J
I1),
adul
tca
lvar
ia(J
I2),
infa
ntm
andi
ble
(JI3
),fr
agm
enta
rypo
stcr
ania
ls(J
I4)
MS
A(‘
‘Lev
allo
iso-
Mou
ster
ian’
’)In
finit
e14C
Enn
ouch
i,19
66
90–1
25ka
(EU
),10
519
0ka
(LU
)E
xtra
pola
tion
from
ES
Rda
tes
onm
amm
alia
nte
eth
over
lyin
gin
situ
hom
inid
spec
imen
JI4
Enn
ouch
i,19
62,
1963
,19
68,
1969
a,b;
How
ell,
1978
;H
ublin
,19
85,
1991
,19
93;
Hub
linet
al.,
1987
;G
run
&S
trin
ger,
1991
Mu
ghar
etel
Aiy
a,M
oroc
coJu
veni
lem
axill
ary
frag
men
tw
ith
part
ial
dent
itio
n,ad
ult
toot
h
MS
A,
pres
umed
Ate
rian
(not
insi
tu)
Lat
eM
idd
le/E
arly
Lat
eP
leis
toce
ne
(‘‘O
uljia
n-S
olta
nian
’’)
Ass
ocia
ted
faun
aB
iber
son,
1961
,19
63;
Deb
enat
h,19
80;
Deb
enat
het
al.,
1982
,19
86;
Am
ani
&G
eraa
ds,
1993
;H
ublin
,19
93
60–9
0ka
Ass
ocia
ted
Ate
rian
arti
fact
s;T
L&
OS
Lre
port
edfo
rin
situ
Ate
rian
mat
eria
lin
Lib
ya
Thi
spa
per,
base
dup
onD
eben
ath,
1994
;M
arti
niet
al.,
1996
;C
rem
asch
iet
al.,
1998
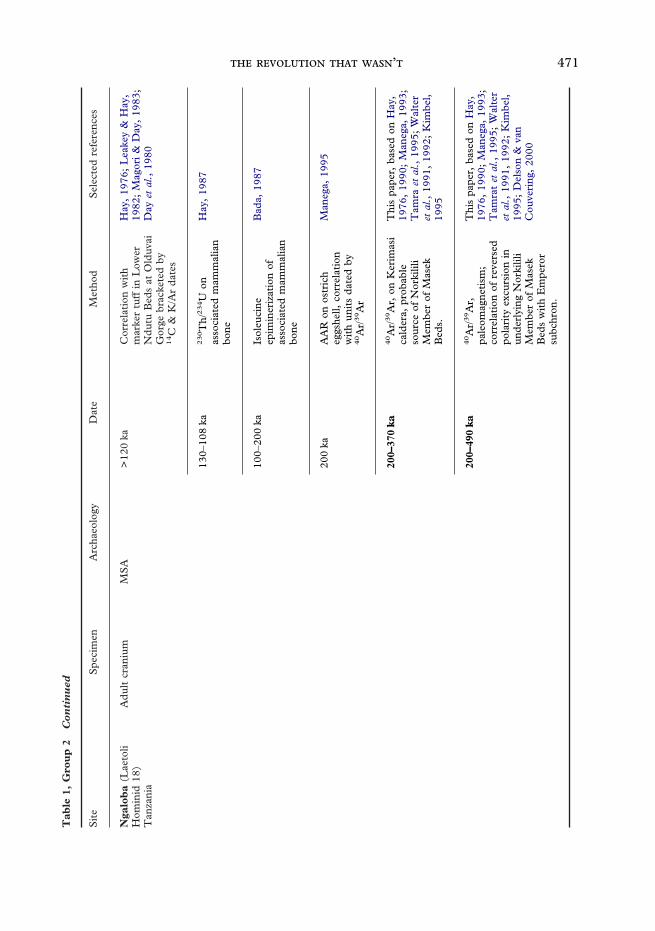
471 ’
Table
1,G
rou
p2
Con
tin
ued
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
Nga
lob
a(L
aeto
liH
omin
id18
)T
anza
nia
Adu
ltcr
aniu
mM
SA
>12
0ka
Cor
rela
tion
wit
hm
arke
rtu
ffin
Low
erN
dutu
Bed
sat
Old
uvai
Gor
gebr
acke
ted
by14C
&K
/Ar
date
s
Hay
,19
76;
Lea
key
&H
ay,
1982
;M
agor
i&
Day
,19
83;
Day
etal
.,19
80
130–
108
ka230T
h/234U
onas
soci
ated
mam
mal
ian
bone
Hay
,19
87
100–
200
kaIs
oleu
cine
epim
iner
izat
ion
ofas
soci
ated
mam
mal
ian
bone
Bad
a,19
87
200
kaA
AR
onos
tric
heg
gshe
ll,co
rrel
atio
nw
ith
unit
sda
ted
by40A
r/39A
r
Man
ega,
1995
200–
370
ka40A
r/39A
r,on
Ker
imas
ica
lder
a,pr
obab
leso
urce
ofN
orki
lili
Mem
ber
ofM
asek
Bed
s.
Thi
spa
per,
base
don
Hay
,19
76,
1990
;M
aneg
a,19
93;
Tam
raet
al.,
1995
;W
alte
ret
al.,
1991
,19
92;
Kim
bel,
1995
200–
490
ka40A
r/39A
r,pa
leom
agne
tism
;co
rrel
atio
nof
reve
rsed
pola
rity
excu
rsio
nin
unde
rlyi
ngN
orki
lili
Mem
ber
ofM
asek
Bed
sw
ith
Em
pero
rsu
bchr
on.
Thi
spa
per,
base
don
Hay
,19
76,
1990
;M
aneg
a,19
93;
Tam
rat
etal
.,19
95;
Wal
ter
etal
.,19
91,
1992
;K
imbe
l,19
95;
Del
son
&va
nC
ouve
ring
,20
00

472 . . .
Tab
le1,
Gro
up
2C
onti
nu
ed
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
Om
oII
,S
ite
PH
S,
Kib
ish
For
mat
ion,
Eth
iopi
a
Adu
ltca
lvar
ia?M
SA
?>
39ka
14C
onE
ther
iash
ell
inov
eryl
ing
unit
Day
,19
69,
1972
;B
utze
r,19
69;
But
zer
etal
.,19
69;
Mer
rick
etal
.,19
73;
Day
&S
trin
ger,
1982
>13
0ka
230T
h/234U
onE
ther
iash
ell
inov
erly
ing
unit
Day
,19
69,
1972
;B
utze
r,19
69;
But
zer
etal
.,19
69;
Mer
rick
etal
.,19
73;
Day
&S
trin
ger,
1982
,19
91
Por
cEu
pic
(Dir
eD
awa)
,E
thio
pia
Man
dibu
lar
frag
men
tM
SA
Lat
eP
leis
toce
neA
ssoc
iate
dfa
una
Bre
uil
etal
.,19
51;
Val
lois
,19
51;
Cla
rk,
1954
,19
82;
Bra
uer,
1984
a;H
owel
l,19
78
>60
–77
kaO
bsid
ian
hydr
atio
nC
lark
etal
.,19
84;
Cla
rk,
1988
Sin
ga,
Sud
anC
alva
ria
?MS
A?
Ear
lyL
ate
Ple
isto
cene
Ass
ocia
ted
faun
aB
ate,
1951
;S
trin
ger,
1979
;B
raue
r,19
84a,
b;S
trin
ger
etal
.,19
85;
Cla
rk,
1988
82–1
12(E
U),
133–
187
(LU
)E
SR
onas
soci
ated
mam
mal
ian
teet
hG
run
&S
trin
ger,
1991
190–
130
kaU
-ser
ies
onca
lcre
teen
clos
ing
skul
lan
das
soci
ated
mam
mal
ian
teet
h
McD
erm
otet
al.,
1996

473 ’
Table
1C
onti
nu
ed
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
Gro
up
3(H
.sa
pie
ns)
Bor
der
Cav
e,S
outh
Afr
ica
Adu
ltca
lvar
ia(B
C1)
,2
adul
tm
andi
bles
(BC
2&
5),
infa
ntpa
rtia
lsk
elet
on(B
C3)
,ad
ult
post
cran
ial
frag
men
ts
MS
A90
–115
kaG
eom
orph
olog
y,as
soci
ated
faun
a,ex
trap
olat
ion
from
over
lyin
g14C
date
s,am
ino
acid
raci
mza
tion
Coo
keet
al.,
1945
;W
ells
,19
50,
1959
;de
Vill
iers
,19
73,
1978
;P
rots
ch,
1975
;B
eaum
ont
etal
.,19
78;
But
zer
etal
.,19
78;
Rig
htm
ire,
1979
b;B
eaum
ont,
1980
;G
run
etal
.,19
90;
Gru
n&
Str
inge
r,19
91;
Mor
ris,
1992
b;M
iller
etal
.,19
93,
1999
;P
feiff
er&
Zeh
r,19
96;
Pea
rson
&G
rine
,19
96(b
utse
eS
illen
&M
orri
s,19
96)
90–5
0ka
AA
R,
asso
ciat
edfa
una,
geom
orph
olog
y,ex
trap
olat
ion
from
over
lyin
g14
Cda
tes
Kle
in,
1999
Dar
-es-
Sol
tan
,M
oroc
coC
rani
al,
max
illar
y&
man
dibu
lar
frag
men
tsof
>2
indi
vidu
als,
adul
t&
suba
dult
MS
A(A
teri
an)
Infin
ite
14C
McB
urne
y,19
61;
Fer
emba
ch,
1976
b;H
owel
l,19
78;
Bra
uer
&R
imba
ch,
1990
;W
endo
rfet
al.,
1990
;B
raue
r,19
92;
Deb
enat
h,19
80,
1994
;D
eben
ath
etal
.,19
82,
1986
;H
ublin
,19
93
60–9
0ka
Ass
ocia
ted
Ate
rian
arti
fact
s;T
L&
OS
Lfo
rin
situ
Ate
rian
mat
eria
lin
Lib
ya
Thi
spa
per,
base
dup
onD
eben
ath,
1994
;M
arti
niet
al.,
1996
;C
rem
asch
iet
al.,
1998

474 . . .
Tab
le1,
Gro
up
3C
onti
nu
ed
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
Die
Kel
der
s,S
outh
Afr
ica
24te
eth,
man
dibu
lar
frag
men
t,2
man
ual
phal
ange
s
MS
A>
40ka
,>
60ka
(EU
),>
80ka
(LU
)E
SR
Gri
ne&
Kle
in,
1985
;G
rine
etal
.,19
91;
Ave
ryet
al.,
1997
71–4
5ka
ES
R,
asso
ciat
edfa
una,
geol
ogic
cont
ext
Kle
in,
1999
60–8
0ka
ES
R,
TL
,O
SL
,IR
SL
Gri
ne,
2000
;S
chw
arcz
&R
ink,
2000
;F
eath
ers
&B
ush,
2000
Eq
uu
sC
ave,
Sou
thA
fric
a(h
yena
laye
r)M
andi
bula
rle
ftco
rpus
frag
men
tw
ith
2m
olar
s;ad
diti
onal
isol
ated
adul
tte
eth
MS
A93
–44
kaU
-ser
ies
ontu
fa,
14C
onM
npa
tina
Gri
ne&
Kle
in,
1985
;K
lein
etal
.,19
91
71–>
27ka
14C
,as
soci
ated
faun
a,ge
olog
icco
ntex
tK
lein
,19
99
Hoe
dji
esP
un
t,S
outh
Afr
ica
Cra
nial
and
post
cran
ial
frag
men
ts,
isol
ated
teet
h
MS
A>
70ka
U-s
erie
sV
olm
an,
1978
;B
erge
r&
Par
king
ton,
1995
300–
71ka
U-s
erie
s,as
soci
ated
faun
a,ge
olog
icco
ntex
tK
lein
,19
99
Kab
ua,
Ken
yaC
rani
al&
man
dibu
lar
frag
men
tsof
2in
divi
dual
s
Unc
erta
in?L
ate
Ple
isto
cene
Geo
logi
cco
ntex
tW
hitw
orth
,19
66;
Rig
htm
ire,
1975
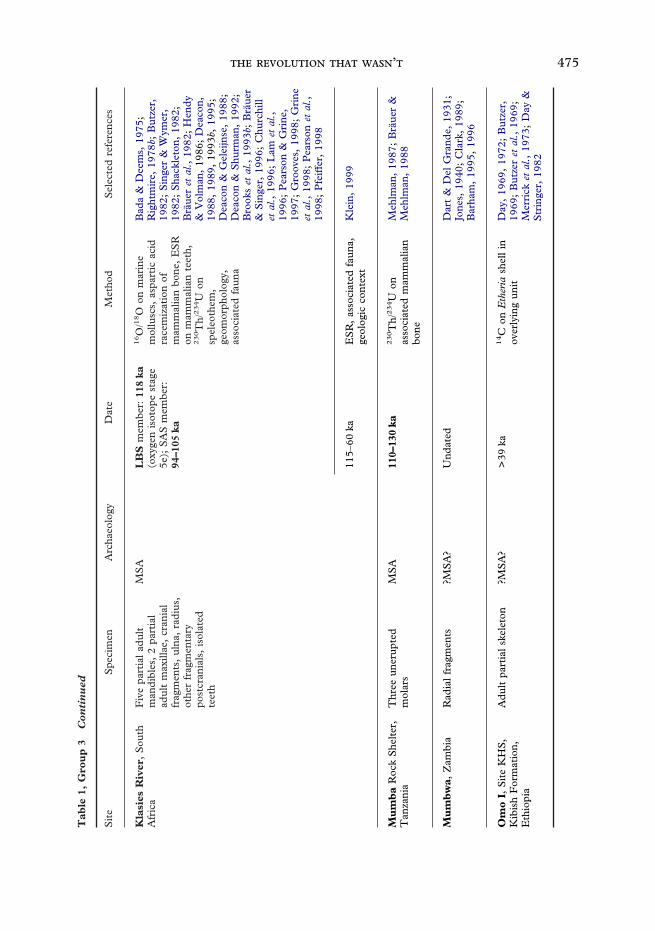
475 ’
Table
1,G
rou
p3
Con
tin
ued
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
Kla
sies
Riv
er,
Sou
thA
fric
aF
ive
part
ial
adul
tm
andi
bles
,2
part
ial
adul
tm
axill
ae,
cran
ial
frag
men
ts,
ulna
,ra
dius
,ot
her
frag
men
tary
post
cran
ials
,is
olat
edte
eth
MS
AL
BS
mem
ber:
118
ka(o
xyge
nis
otop
est
age
5e);
SA
Sm
embe
r:94
–105
ka
16O
/18O
onm
arin
em
ollu
scs,
aspa
rtic
acid
race
miz
atio
nof
mam
mal
ian
bone
,E
SR
onm
amm
alia
nte
eth,
230T
h/234U
onsp
eleo
them
,ge
omor
phol
ogy,
asso
ciat
edfa
una
Bad
a&
Dee
ms,
1975
;R
ight
mir
e,19
78b;
But
zer,
1982
;S
inge
r&
Wym
er,
1982
;S
hack
leto
n,19
82;
Bra
uer
etal
.,19
82;
Hen
dy&
Vol
man
,19
86;
Dea
con,
1988
,19
89,
1993
b,19
95;
Dea
con
&G
elei
jnse
,19
88;
Dea
con
&S
hurm
an,
1992
;B
rook
set
al.,
1993
b;B
raue
r&
Sin
ger,
1996
;C
hurc
hill
etal
.,19
96;
Lam
etal
.,19
96;
Pea
rson
&G
rine
,19
97;
Gro
oves
,19
98;
Gri
neet
al.,
1998
;P
ears
onet
al.,
1998
;P
feiff
er,
1998
115–
60ka
ES
R,
asso
ciat
edfa
una,
geol
ogic
cont
ext
Kle
in,
1999
Mu
mb
aR
ock
She
lter
,T
anza
nia
Thr
eeun
erup
ted
mol
ars
MS
A11
0–13
0ka
230T
h/234U
onas
soci
ated
mam
mal
ian
bone
Meh
lman
,19
87;
Bra
uer
&M
ehlm
an,
1988
Mu
mb
wa,
Zam
bia
Rad
ial
frag
men
ts?M
SA
?U
ndat
edD
art
&D
elG
rand
e,19
31;
Jone
s,19
40;
Cla
rk,
1989
;B
arha
m,
1995
,19
96
Om
oI,
Sit
eK
HS
,K
ibis
hF
orm
atio
n,E
thio
pia
Adu
ltpa
rtia
lsk
elet
on?M
SA
?>
39ka
14C
onE
ther
iash
ell
inov
erly
ing
unit
Day
,19
69,
1972
;B
utze
r,19
69;
But
zer
etal
.,19
69;
Mer
rick
etal
.,19
73;
Day
&S
trin
ger,
1982

476 . . .
Tab
le1,
Gro
up
3C
onti
nu
ed
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
Om
oI
cont
inue
d>
130
ka230T
h/234U
onE
ther
iash
ell
inov
erly
ing
unit
Day
,19
69,
1972
;B
utze
r,19
69;
But
zer
etal
.,19
69;
Mer
rick
etal
.,19
73;
Day
&S
trin
ger,
1982
,19
91;
Bra
uer
&R
imba
ch,
1990
;B
raue
r,19
92
Sea
Har
vest
,S
alda
nha
Bay
,S
outh
Afr
ica
(hye
nala
yer)
Too
th,
phal
anx
MS
A>
40ka
14C
onos
tric
heg
gsh
ell
Vol
man
,19
78;
Gri
ne&
Kle
in,
1993
;K
lein
,19
94
127–
>40
ka14C
,ge
olog
icco
ntex
tK
lein
,19
99
Sol
eb,
Sud
anC
rani
al&
man
dibu
lar
frag
men
tsof
3in
divi
dual
s
MS
A(‘
‘Lat
eL
eval
lois
ian’
’)?L
ate
Ple
isto
cene
Ass
ocia
ted
faun
aG
iorg
ini,
1971
;S
auss
e,19
75a
>90
ka,
>16
0A
ssoc
iate
dar
tefa
cts.
Gen
eral
ized
MS
Apr
ecee
dsA
teri
anda
ted
byU
-ser
ies
atsi
tes
inE
gypt
ian
Sah
ara
Thi
spa
per,
base
dup
onK
lein
dien
st,
2000
b;K
lein
dien
st&
Wis
eman
,19
96;
Kle
indi
enst
etal
.,19
96;
Wen
dorf
etal
.,19
87,
1993
a
Tem
ara
(Gro
tte
deC
ontr
aban
dier
s,E
lM
nasr
a2,
Rab
at)
Mor
occo
Adu
ltm
andi
ble
wit
hde
ntit
ion,
max
illar
yte
eth,
occi
pita
l,fr
onta
lan
dpa
riet
alfr
agm
ents
MS
A(A
teri
an)
Lat
eM
idd
leP
leis
toce
ne
(‘‘E
pi-O
uljia
n’’)
Ass
ocia
ted
faun
aV
allo
is,
1960
;B
iber
son,
1961
;T
obia
s,19
68;
Sab
an,
1972
;R
oche
&T
exie
r,19
76;
Fer
emba
ch,
1976
a;H
owel
l,19
78;
Deb
enat
h,19
80;
Deb
enat
het
al.,
1982
,19
86;
Hub
lin,
1993
>40
kaA
ssoc
iate
dar
tefa
cts
(infi
nite
14C
for
Ate
rian
else
whe
rein
Sah
ara)
Clo
se,
1984
;W
endo
rfet
al.,
1993
a;C
rem
asch
iet
al.,
1998

477 ’
Tab
le1,
Gro
up
3C
onti
nu
ed
Sit
eS
peci
men
Arc
haeo
logy
Dat
eM
etho
dS
elec
ted
refe
renc
es
Tem
ara,
cont
inue
d13
0–40
kaA
ssoc
iate
dar
tefa
cts
Thi
spa
per,
base
dup
onD
eben
ath,
1994
60–9
0ka
Ass
ocia
ted
arte
fact
s;T
L&
OS
Lre
port
edfo
rin
situ
Ate
rian
mat
eria
lin
Lib
ya
Thi
spa
per,
base
dup
onM
arti
niet
al.,
1996
;C
rem
asch
iet
al.,
1998
Tar
amsa
,E
gypt
Par
tial
juve
nile
skel
eton
MS
A50
–80
kaO
SL
onsa
ndV
erm
eers
chet
al.,
1998
Zou
hra
h(E
lH
arho
ura,
Mor
occo
)M
andi
ble,
isol
ated
toot
hM
SA
(Ate
rian
)>
40ka
Ass
ocia
ted
arte
fact
s(i
nfini
te14C
for
Ate
rian
else
whe
rein
Sah
ara)
Clo
se,
1984
;W
endo
rfet
al.,
1993
a;C
rem
asch
iet
al.,
1998
130–
40ka
Ass
ocia
ted
arte
fact
sT
his
pape
r,ba
sed
upon
Deb
enat
h,19
94
60–9
0ka
Ass
ocia
ted
arte
fact
s,T
L&
OS
Lre
port
edfo
rin
situ
Ate
rian
mat
eria
lin
Lib
ya
Thi
spa
per,
base
dup
onM
arti
niet
al.,
1996
;C
rem
asch
iet
al.,
1998
Age
esti
mat
esco
nsid
ered
tobe
mos
tac
cura
tear
ein
bold
face
.Whe
nm
ulti
ple
age
esti
mat
esar
eeq
ually
wel
lsup
port
ed,a
llar
ein
bold
face
(e.g
.,es
tim
ates
base
don
reca
libra
ted
Old
uvai
sequ
ence
).A
AR
anal
yses
onbo
nepe
rfor
med
befo
re19
80sh
ould
bere
gard
edw
ith
caut
ion.
Que
stio
nm
arks
inth
ear
chae
olog
yco
lum
nin
dica
teth
atei
ther
indu
stri
alat
trib
utio
nsor
asso
ciat
ions
are
prob
lem
atic
.Rec
entr
e-ex
amin
atio
nof
Kib
ish
For
mat
ion
geol
ogy
indi
cate
sth
atth
eO
mo
Kib
ish
finds
may
besu
bsta
ntia
llyol
der
than
the
date
give
nhe
re(F
leag
le,
pers
onal
com
mun
icat
ion)
.G
ener
alre
fere
nces
:O
akle
yet
al.,
1977
;H
owel
l,19
78,
1982
;B
raue
r,19
84a,
b;R
ight
mir
e,19
84;
Day
,19
86;
Str
inge
r&
And
rew
s,19
88;
Mor
ris
1992
a;K
lein
,19
99;
Del
son
etal
.,20
00.

478 . . .
Figure 1. Map of archaeological and hominid fossil sites mentioned in the text. � Sally McBrearty andAlison S. Brooks.

Figure 1. Key to sites.
Aın Maarouf (MOR) 1Aduma (ETH) 2Apollo 11 Rock Shelter (NAM) 3Bambata (ZIM) 4Berard (ALG) 5Berg Aukas (NAM) 6Bir Sahara East (EGY) 7Bir Tarfawi (EGY) 8Blombos Cave (SA) 9Bodo (ETH) 10Boegoeberg (SA) 11Boomplaas (SA) 12Border Cave (SA) 13Boskop (SA) 14Buias (ERT) 15Bushman Rock Shelter (SA) 16Cartwright’s Site (KEN) 17Cave of Hearths (SA) 18Dakleh Oasis (EGY) 19Dar-es-Soltan (MOR) 20Diepkloof (SA) 21Die Kelders (SA) 22Dunefield Midden (SA) 23Elands Bay Cave (SA) 24El-Guettar (TUN) 25Eliye Springs (KEN) 26Equus Cave (SA) 27Enkapune ya Muto (KEN) 28Eyasi (TAN) 29Florisbad (SA) 30Gademotta (ETH) 31Gadeb (ETH) 32�Gi (BOT) 33Gorgora (ETH) 34Grotte d’el Mnasra (MOR) 35Guomde (KEN) 36Haua Fteah (LIB) 37Herolds Bay (SA) 38Hoedjies Punt (SA) 39Howiesons Poort (SA) 40Ishango (DRC) 41Isimila (TAN) 42Jebel Irhoud (MOR) 43Kabua (KEN) 44Kabwe (Broken Hill) (ZAM) 45Kalambo Falls (ZAM) 46Kalemba (ZAM) 47Kapthurin (KEN) 48Kasteelberg (SA) 49Katanda (DRC) 50Kebitat (Rabat) (MOR) 51Klasies River (SA) 52Kukuleti (ETH) 53Lainymok (KEN) 54Lake Ziway (ETH) 55Le Chaperon Rouge (MOR) 56Lion Cavern (SWA) 57Loyangalani (KEN) 58Lukenya Hill (KEN) 59Magosi (UGA) 60Malewa Gorge (KEN) 61
Massawa (ERI) 62Matupi Cave (DRC) 63Melka Konture (ETH) 64Midhishi (SOM) 65Montagu Cave (SA) 66Mossel Bay (SA) 67Mosumu (EQG) 68Mugharet el-Aliya (MOR) 69Muguruk (KEN) 70Mumba Shelter (TAN) 71Mumbwa (ZAM) 72Nasera Rock Shelter (TAN) 73Nazlet Safaha (EGY) 74Ndutu (TAN) 75Nelson Bay Cave (SA) 76Ngaloba (TAN) 77Njuinye (CAM) 78Nswatugi (ZIM) 79Olduvai Gorge (TAN) 80Olorgesailie (KEN) 81Omo (ETH) 82Orangia (SA) 83Oued Djebanna (ALG) 84Paternoster (SA) 85Peer’s Cave (SA) 86Pomongwe (ZIM) 87Porc Eupic (Dire Dawa) (ETH) 88Prolonged Drift (KEN) 89Prospect Farm (KEN) 90Rhino Cave (BOT) 91Rooidam (SA) 92Rose Cottage Cave (SA) 93Saldanha (Elandsfontein, Hopefield) (SA) 94Sale (MOR) 95Sea Harvest (SA) 96Seggedim (NGR) 97Sehonghong (LES) 98Sibebe Rock Shelter (SA) 99Sidi Abderrahman (MOR) 100Simbi (KEN) 101Singa (SUD) 102Sodmein Cave (EGY) 103Soleb (EGY) 104Songhor (KEN) 105Still Bay (SA) 106Strathalan (SA) 107Swartkrans (SA) 108Temara (MOR) 109Taramsa (EGY) 110Thomas I Quarry (MOR) 111Tighenif (Ternifine) (ALG) 112Tuinplaas (SA) 113Twin Rivers (ZAM) 114Uan Afuda (LIB) 115Uan Tabu (LIB) 116Wadi Dagladle (DJB) 117Wetherall’s Site (KEN) 118White Paintings Shelter (BOT) 119Windhoek (NAM) 120Zourah (MOR) 121

480 . . .
intermediate between H. habilis and H.sapiens. Clarke (1990), after detailed studyof the Ndutu cranium and Swartkranscranium SK847, also usually attributed toH. erectus, concurred with Leakey. He con-sidered the verticality of the parietal wallsand the upper scale of the occipital as par-ticularly compelling features distinguish-ing the African fossils from H. erectus.Its correct designation is H. louisleakeyi,Kretzoy, 1984.3 If H. louisleakeyi is a dis-tinct species, questions remain as to whichspecimens, currently loosely housed withinH. erectus, might reasonably be includedwithin it, and how they might be distin-guished from contemporaneous hominidspecies.
We would favor retaining the designationH. rhodesiensis Woodward, 1921 for Kabweand related African specimens, although ithas been argued that there is no signifi-cant anatomical difference between AfricanH. rhodesiensis and European specimensattributed to H. heidelbergensis (Stringer,1974; Brauer, 1984a; Van Vark, 1995;Rightmire, 1998). If the latter positionis valid, the name H. heidelbergensisSchoetensack, 1908 clearly has priority forsome of the Group 1 fossils. However,the Mauer mandible, type specimen of H.heidelbergensis, has a retromolar space, sug-gesting affinities with H. neanderthalensis (cf.Rak, 1998). Stringer (1993b, 1996) has alsocalled the status of H. heidelbergensis intoquestion and suggests it may be necessaryfor it to be sunk into H. neanderthalensis,because the highly variable Sima de losHuesos sample from Atapuerca, dating toabout 300 ka and presumed to represent a
single population (Arsuaga et al., 1993,1997a,b; Bischoff et al., 1997), includesspecimens that would formerly have beenattributed to both H. neanderthalensis andH. heidelbergensis.
In light of the complexity presented bythe European Middle Pleistocene record(Hublin, 1998a; Stringer & Hublin, 1999),there are grounds for retaining H. rhodesien-sis for Middle Pleistocene hominids not onthe Neanderthal lineage.4
In Group 2 specimens frontal curvature isgreater than in Group 1 fossils. Brow ridgedevelopment and angulation of the occiput,among other traits, are less pronounced thanin fossils of Group 1, but still fall outside therange of modern human variation. Group 2includes the Florisbad cranium, type speci-men of H. helmei, and thus if these fossilsrepresent a species distinct from H. sapiensand H. rhodesiensis, the name H. helmei is theappropriate designation for them. H. helmeiis a somewhat problematic taxon, as there isas yet no formal diagnosis for the species andits unique autapomorphies are not defined.Like all species (Hennig, 1965; Harrison,1993), H. helmei may be expected to exhibita mix of primitive and derived features, andthe specimens attributed here to Homohelmei share characteristics with bothH. sapiens and Group 1 hominids.
In a curious departure from normaltaxonomic practice, Lahr & Foley (1998:
3. Clarke (1990) resurrected the taxon H. leakeyiHeberer, 1963, whose holotype is OH9, for this andother African specimens formerly attributed to H.erectus. However, Groves (1999) has pointed out thatbecause Heberer (1963) awarded the name leakeyiconditionally, it is not available according to the Inter-national Code of Zoological Nomenclature. Kretzoi’s(1984) proposal of H. (Proanthropus) louisleakeyi forOH9 is thus the valid designation for it and relatedfossils.
4. A potential complication for Middle Pleistocenenomenclature is presented by the hominid fossils fromGran Dolina, Atapuerca, dating to >780 ka (Pares &Pares-Gonazalez, 1995; Bermudez de Castro et al.,1997), for whom the investigators have erected thetaxon H. antecessor. They claim that derived features inthe fossils make it likely that they represent a populationancestral to both H. neanderthalensis and H. sapiens. Ifthe Gran Dolina fossils, like later Sima de los Huesosmaterial, dating to about 300 ka (Arsuaga et al., 1993,1997a,b; Bischoff et al., 1997), fall on the Neanderthallineage, they are excluded from modern humanancestry (Rightmire, 1998). If the taxon H. antecessorhas validity, it is critical clearly to distinguish it from H.heidelbergensis, if it is proposed to extend the latter taxonto non-European material. Resolution of these issuesrequires further detailed study of the fossils themselvesand the application of cladistic methods.

481 ’
157) have included certain European fossilsin H. helmei on the basis of their ages (iso-topic stages 6, 7, and 8) and presumedassociated technology (‘‘mode 3’’). Stringer(1996), in a consideration of cranial charac-ters, places Asian specimens such as Dali,Maba, Narmada, and Zuttiyeh in this group.These practices are not followed here.Rather, the species is taken to encompassthose African fossils described by previousauthors (e.g., Day, 1973; Brauer, 1984a,b,1989; Smith, 1985, 1993; Clark, 1988;Klein, 1989b; Stringer, 1993a, 1995, 1996)as ‘‘intermediate’’ in morphology betweenthose of Group 1 and H. sapiens sensus stricto.These include Ngaloba (LH 18), Dar-es-Soltan, Guomde, Eliye Springs, and OmoKibish II, though the latter specimen iscurrently undergoing reconstruction byLahr & Foley (personal communication)and its taxonomic evaluation may change asa result. The Singa specimen is includedbecause, although it shows clear pathologyin the cranial vault (Stringer, 1979; Stringeret al., 1985; Spoor & Stringer, 1998),Stringer (personal communication) believesthat it shows primitive features that excludeit from H. sapiens sensu stricto.
Fossils in Group 3 are those of H. sapiens(sensu stricto), sometimes distinguished byothers as the subspecies, H. sapiens sapiens.Early African fossils of H. sapiens includeKlasies River, Border Cave (but see Sillen &Morris, 1996) and Omo Kibish I. While insome cases retaining a degree of skeletalrobusticity, these fossils are essentially likemodern people in their morphology (e.g.,Grine & Klein, 1985, 1993; Habgood,1989; Rightmire & Deacon, 1991; Stringer,1995, 1996, 1998; but see Schwartz &Tattersall, 2000). Klasies frontal fragment16425, for example, is modern in the con-formation of its supraorbital region, andmandibles 21776 and 41815 have welldeveloped chins and no retromolar spaces(Klein, 1994; Grine et al., 1998; Rightmire,1998). The Klasies hominids show a highdegree of variation that may represent sexualdimorphism. Some of the smaller Klasiesfossils, such as the temporal fragmentSAM-AP 6269, fall within or even below therange of modern female San (Grine et al.,1998).
Current best estimates of age ranges forrelevant Pleistocene hominid taxa are pre-sented in Figure 2. It is unlikely that the
Figure 2. Time ranges of African hominid taxa mentioned in the text. � Sally McBrearty.

482 . . .
known fossils represent the true first andlast appearances of these hominid species.Statistical principles suggest that the knownage range for any taxon is in part a functionof sample size, and it has been empiricallydemonstrated that expanded sample sizescan affect first and last appearance data forfossil taxa, including hominoids (Hill, 1987;Strauss & Sadler, 1989; Marshall, 1990;Springer, 1990; Flynn et al., 1995). In short,larger samples tend to lengthen fossil taxa’slifespans, and we may expect discoveries offossils both older and younger than thoserepresented here.
Early dating attempts by 14C and AARproduced results that often were not recog-nized as infinite. Prior to recent redatingefforts, age estimates for many of thesespecimens were derived from faunal com-parisons. Unlike the Pliocene or earlierPleistocene, there are few good faunal tem-poral indicators for the Middle Pleistocene,and there was a tendency in previous studiesto refer fragmentary specimens to modernspecies, and thus systematically to inflatethe numbers of modern taxa in MiddlePleistocene faunal assemblages. Conse-quently age estimates for many hominidspecimens converged upon 130 ka, theMiddle to Late Pleistocene boundary. Withthe application of refined techniques, thetrend in nearly all cases has been for ageestimates to be revised downward. It isunclear whether faunal assemblages fromthis time range contain a higher proportionof undetected archaic taxa, or whether themodern African fauna has a greater antiquitythan previously recognized.
The age of the recently discovered Buiacranium is estimated at ca. 1·0 Ma (Abbateet al., 1998). Recent recalibration of theOlduvai Gorge sequence (Kimbel, 1995;Tamrat et al., 1995; Walter et al., 1991;1992; White, 2000; Delson & VanCouvering, 2000; for other references seeTable 1) directly affects the ages of OH9(formerly ca. 1·2 Ma, now ca. 1·4 Ma)
OH12, OH22, and OH28 (formerly 600–800 ka, now 0·78–1·2 Ma), and OH23 andthe Ndutu cranium (now ca. 370–990 ka).Many of the age estimates for other homin-ids in Group 1 are derived from faunalcorrelation with Olduvai Beds III and IV.For example, the age of the Saldanhacranium, once thought to perhaps be asyoung as 130 ka, was revised to ca. 400–700 ka on the basis of comparison of itsassociated fauna to that in Olduvai Bed IV.The same faunal evidence now may bringSaldanha into the 1·07–1·3 Ma time range(see Table 1 for references). Parallel reason-ing can be applied to the Kabwe cranium,the type of H. rhodesiensis, as well as otherhominids in Group 1. Though these ideasrequire testing with independent methods, itis interesting to note that humans may bepart of an essentially modern fauna that hasdeeper roots in Africa than has been thus farappreciated.
H. sapiens is present in South Africa at110–120 ka at Klasies River and before130 ka in East Africa in the KibishFormation and at Mumba, Tanzania (forreferences see Table 1). These are onlyslightly earlier than early H. sapiens in Israel,which date to ca. 80–120 ka at Skhul(Stringer et al., 1989; Mercier et al., 1993;Bar-Yosef, 1998) and to 92–115 ka atQafzeh (Valladas et al., 1988; Schwarczet al., 1988; Bar-Yosef, 1998). Dates for H.helmei fall between ca. 300 ka for Guomde(Feibel et al., 1989; Brauer et al., 1997) andca. 130 ka. The antiquity of this species issupported by the ESR age determination of260 ka made by direct assay on a hominidtooth associated with the holotype atFlorisbad (Grun et al., 1996; Brink & Grun,1998).
A number of inferences can be drawnfrom these data. In contrast to the patternseen in western Europe, eastern Asia, andthe Near East, there is a series of closelyrelated taxa that provide plausible ancestorsfor H. sapiens, and the record shows no

483 ’
abrupt discontinuities. This pattern in itselfis suggestive of in situ evolution of H. sapiensin Africa. The data may represent one oftwo evolutionary modes: (1) a series ofarbitrarily defined chronospecies, or (2) aMiddle Pleistocene adaptive radiationwithin genus Homo in Africa. The latterscenario has appeal because, although itwould greatly complicate archaeologicalinterpretations, it parallels more closelywhat is known of earlier evolutionary eventswithin the Hominidae. However, greaterchronologic control is needed to evaluate itsmerits.
Behavior from fossils
The origin of H. sapiens cannot be under-stood without reference to behavior.Behavior can sometimes be established fromexamination of the fossils themselves. Forexample, cutmarks on the Bodo skull andthe Klasies River sample may indicatecannibalism or ritual behavior (White, 1986,1987; Deacon & Deacon, 1999). Postcraniaare a possible source of information regard-ing postural and locomotor habits andactivity levels, though postcranial remainsfrom African Middle Pleistoene and earlylater Pleistocene contexts are few in number(see Table 1) and, in most cases, problemsof association render taxonomic attributiondifficult. Fossil postcrania exhibit bothprimitive and derived traits (Stringer, 1986;Solan & Day, 1992; Churchill et al., 1996;Pearson & Grine, 1997; Groves, 1998;McBrearty et al., 1999). Features of theproximal ulnae from Klasies River and theKapthurin Formation led Churchill et al.(1996) to suggest a different pattern of fore-arm loading during flexion than is found inmost living representatives of H. sapiens.
The gracilization of the upper limb whichmarks the advent of anatomical modernity(Churchill et al., 1996; Pearson et al., 1998;Pfeiffer, 1998) is a reversal of a previoustrend towards robusticity and may suggest a
relaxation of selection for constant mobilityand close encounters with prey (Trinkaus,1983, 1984, 1986, 1987). Fewer serioustraumatic injuries are observed on skeletonsof early H. sapiens than on Neanderthals,indicating either fewer or more successfulviolent encounters with animals or otherhumans (Berger & Trinkaus, 1995). Thetropical limb proportions of early H. sapiensin late glacial Europe suggest not only anextension of the human geographic rangeout of the tropics, but also the use of culturalrather than physical solutions to the prob-lems of cold temperatures and scarceresources (Trinkaus, 1981; Torrence, 1983;Foley, 1987; Ruff, 1994; Holliday, 1997,1998, 2000; Pearson, 1997). That thesesolutions resulted in less nutritional stressamong juveniles in early modern humanpopulations is reflected in the lower inci-dence of Harris’ lines and enamel hypo-plasias among these populations relative tothe Neanderthals (Bermudez de Castro,1988; Ogilvie et al., 1989; Brennan, 1991;Trinkaus, 1995), although sample sizes inthese studies are small.
Cultural solutions to the challengesimposed by dangerous predators and inter-mittent food scarcity are implicit in thearchaeological record of the MSA in Africa.Projectile technology lessens the inherentrisk of hunting dangerous game animals byinflicting ‘‘death at a distance’’ (Berger &Trinkaus, 1995; Cattelain, 1997; but seeBartram, 1997; Hitchcock & Bleed, 1997),and it is quite likely that some MSA pointswere used as projectiles. Modern foragerscombat seasonal scarcity through economicadaptations such as food storage, scheduledresource use, and the intensification ofhabitat exploitation, either through theuse of small scale resources or throughmodification of the environment. Socialadaptations to scarcity may involve themaintenance of long-distance reciprocalexchange relationships, or the long-termretention of individual and group memories

484 . . .
of distant resources or survival strategies,often aided by the use of symbol and ritual(de Garine & Harrison, 1988). We arguethat most, if not all, of these behaviors werepractised in the MSA.
Three African Middle Pleistocene homi-nid fossil crania show signs of pathology thatare relevant to behavioral reconstruction.The Kabwe skull displays extensive dentalcaries as well as lesions to both thesquamous and petrous portions of the lefttemporal bone and to the left mastoid pro-cess. Although Yearsley (1928) had pro-posed acute mastoiditis as the cause of thelesions to the mastoid and the petrous tem-poral, examination by Montgomery et al.(1994) shows these to be the result of post-mortem trauma. However, the same studydescribes the everted edges of the squamoustemporal lesion, indicating that it is a par-tially healed premortem or perimorteminjury. While these authors attribute thelesion to a cyst or benign tumor, the sug-gestion of Wells (1964) that it representsan abscess resulting from the dental cariescannot be ruled out.
The Singa calvaria exhibits a pronouncedthickening of the diploic layer of the tem-porals (Stringer et al., 1985), as well ascomplete ossification of the bony labyrinthof the petrous portion of the right temporal(Spoor & Stringer, 1998). These featuresare consistent with either infection oranemia. Total ossification requires at leastone year to complete, and would haveresulted in complete deafness in the rightear. The initial bout of disease, lasting froma few days to a few months, would have beenaccompanied by vertigo, postural imbal-ance, and possible paralysis (Spoor &Stringer, 1998). The Sale calvaria showsdeformation to the nuchal area that resultedfrom weakened and asymmetrically devel-oped nuchal musculature. The conditionsuggests in utero deformity or perinataltrauma and probably resulted in reducedmobility of the head and marked asym-
metries of the face and mandible, which arenot preserved in this specimen (Hublin,1994). It is feasible that the survival of thesethree individuals required intervention byother members of the group, particularly inthe case of Sale, which demonstrates thesurvival of a handicapped infant into adult-hood.
Archaeology, behavior and hominidtaxonomy
How do the hominids of Groups 1, 2, and 3differ in their adaptations and other aspectsof their behavior, and are these differencesadequate to distinguish them at the specieslevel? In both neotology and paleontology,behavior is considered part of what separatesa species from closely related forms(Dobzhansky, 1937; Mayr, 1963). It is asignificant component in determining aspecies’ adaptation, geographic range, andmate recognition system, all of whichcontribute to the maintenance of speciesboundaries (Wiens, 1977; Paterson, 1985;Tattersall, 1986; Foley & Lee, 1989; Foley,1991a, 1994). Darwin (1859: 76) and hissuccessors (e.g., Dobzhansky, 1937; Mayr,1963) have generally argued that closelyrelated species experience the greatest com-petition for resources, and can only coexistif they use the environment differently. Inthe study of human evolution, speciesdefinitions have often been underlain,overtly or covertly, by supposed behavioraldistinctions, as in the case of H. habilis(Leakey et al., 1964; Tobias, 1991).
Archaeological remains are a good sourceof evidence for past behavior, and mayhelp to clarify the nature of later MiddlePleistocene speciation events themselves.The most conspicuous behavioral event inthe late Middle Pleistocene archaeologicalrecord of Africa is the disappearance of theAcheulian industry before 200 ka, and itsreplacement by diverse MSA traditions.Thus, it is to the late Acheulian and early

485 ’
MSA that we should look to understandthe circumstances surrounding the origin ofH. sapiens. This obvious fact is consistentlyoverlooked because Europe’s earliest mod-ern human inhabitants, about 150,000 yearslater, were makers of Upper Paleolithic tech-nology. Thus the origin of H. sapiens hasbeen conflated with the origin of the UpperPaleolithic. The Acheulian contains largebifaces, while most MSA industries arecharacterized by smaller flake tools, some-times made on flakes struck from preparedcores, a technology often referred to asMode 3 (Clark, 1977).5 The abandonmentof handaxes and cleavers for smaller flaketools represents the replacement of hand-held by hafted implements and signals aprofound technological reorganization.
To what extent can behavioral data helpto resolve the issue of whether these species
are sound taxonomic categories? How con-sistent are the archaeological associations ofeach group and how great are the behavioralor ecological differences among them?The correlations between archaeologicalindustries and hominid taxa in MiddlePleistocene and early Late PleistoceneAfrica are summarized in Table 2.
While hominids cannot necessarily beassumed to be the makers of the artefactswith which they are found, the associationsare striking in their consistency. Fossils ofGroup 1 in all cases are associated withOldowan, Acheulian or Sangoan artefacts(here considered ESA technology), whileboth H. helmei and early members of H.sapiens are associated with MSA technology.Thus, it appears that the major adaptiveshift represented by the Acheulian-MSAboundary ca. 250–300 ka corresponds witha speciation event. In contrast, there isno evidence for hominid morphologicalchange during the MSA–LSA transition,which is the product of culture change.Overlying the Acheulian and underlying theMSA at some sites is a heavy-duty industrialvariant called the Sangoan (McBrearty,1987, 1988, 1991; Cornelissen, 1995). The
5. Mode 3 technology alone, however, cannot beequated with the MSA (contra Foley & Lahr, 1997;Lahr & Foley, 1998) for a number of reasons: (1)Prepared core technology first makes its apperance inthe Acheulian (ESA); (2) many MSA assemblages lackprepared cores; (3) in South, East, and NortheastAfrica, many MSA industries are blade-based (Mode 4technology); and (4) several MSA industries arenow known to contain backed geometrics (Mode 5technology).
Summary of hominids and their archaeological associations
GROUP 1=ESA(H. louisleakeyi,H. rhodesiensis)
GROUP 2=MSA(H. helmei orH. sapiens)
GROUP 3=MSA(H. sapiens)
Aın Maarouf (Ach) Florisbad (?) Border CaveBodo (Ach/Old) Haua Fteah Dar-es-SoltanCave of Hearths (Ach) Jebel Irhoud Die KeldersEyasi (Sang) Mugharet el Aliya (?) Equus CaveKabwe (?Sang/MSA?) Ngaloba Hoedjies PuntMelka Konture (Ach) Omo II (?) Klasies RiverNdutu (Ach) Porc Epic (?) MumbaOH 9 (Ach/Old) Singa (?) Mumbwa (?)OH 23 (Ach) Omo 1 (?)OH 28 (Ach) Sea HarvestOH 34 (Ach) SolebSaldanha (Ach) TemaraSidi Abderrahman (Ach) TaramsaTighenif (Ach) Zouhrah
Question marks indicate that associations are problematic.
Table 2

486 . . .
associations of the Sangoan artefacts suggestthat this industry may best be regardedas part of the behavioral repertoire of thehominids in Group 1.
The coincidence of a major archaeologicalchange with the origin of H. helmei sug-gests that this species possessed enhancedcognitive abilities when compared with thehominids in Group 1. H. helmei maybe indistinguishable behaviorally from H.sapiens, suggesting a close phylogeneticrelationship. Lahr (1996) and Stringer(1996, 1998) have suggested includingsome or all of the fossils in Group 2 in H.sapiens. We would argue that taxonomyshould be done on the basis of fossil, notarchaeological evidence. Nonetheless, thearchaeological data do lend plausibility tothis suggestion. Should further study ofthe fossils ultimately support inclusion ofspecimens here ascribed to H. helmei in H.sapiens, there may be grounds for sinking thetaxon H. helmei. H. sapiens and MSA tech-nology would then be seen to appear simul-taneously in the record between 250 ka and300 ka.
It is clear that the features diagnostic ofphysical modernity emerge in conjunctionwith MSA technologies. In this paper wepresent evidence to support the presence ofmodern human behaviors in subsaharanAfrica at remote times far predating anysuch traces outside Africa. We contendthat the appearance of modern behaviorsaccompanied or even preceded the appear-ance of H. sapiens during the African MSA,suggesting that the behaviors may perhapshave driven the anatomical changes seen inthe fossils. We also suggest that these behav-iors developed gradually over a substantialperiod of time and sporadically in differentparts of the continent.
The origin of language?
In much of the recent literature, the ‘‘humanrevolution’’ is synonymous with the origin of
‘‘fully syntactical’’ language (e.g., Mellars,1991; Noble & Davidson, 1991, 1996;Klein, 1992, 1995; Mithen, 1994, 1996).Abstract and symbolic behaviors implylanguage, but it is doubtful that the point atwhich they can first be detected coincideswith the birth of language. Attempts toidentify the earliest signs of language,whether from study of the brain (e.g., Falk,1980, 1990; Holloway, 1983, 1985;Deacon, 1997), the speech apparatus(e.g., Falk, 1975; Lieberman, 1975, 1984;Arensburg et al., 1990; Lieberman et al.,1992; Kay et al., 1998), stone tools (e.g.,Wynn, 1979; Dibble, 1989; Toth & Schick,1993), or primate communication (e.g.,Cheney & Seyfarth, 1990; Savage-Rumbaugh & Rumbaugh, 1993), contributeto a sense of continuity, rather than dis-continuity, between human and nonhumanprimate cognitive and communicativeabilities (e.g., Parker & Gibson, 1979;papers in Parker & Gibson, 1990; Gibson,1993; Gibson & Ingold, 1993; Mellars &Gibson, 1996). Language does not fossilize,and technology provides insight only intothe minimum cognitive abilities of itsmakers and users (Wobst, 1983; Wynn,1985; Goren-Inbar, 1988). We infer thatsome form of language originated early inhuman evolution, and that language existedin a variety of forms throughout its longevolution, but we concur with those whohave pointed out the inherent untestabilityof many propositions regarding the origin oflanguage, and the profound silence of thearchaeological record on this issue (Dibble,1989; Foley, 1991b; Graves-Brown, 1994).
Geographic limits
In this paper, discussion is limited to thearchaeological record of Africa, in orderto provide the clearest contrast betweenAfrica and Europe. We do not attemptsystematically to integrate the evidence fromthe Levant, which is increasingly seen as a

487 ’
contact zone between the Eurasian fauna,including H. neanderthalensis, and theAfrican fauna, including H. sapiens (Jacobs,1985; Bar-Yosef, 1987, 1995b, 1998;Valladas et al., 1988; Tchernov, 1988, 1998;Grun et al., 1991; Holliday & Trinkaus,1991; Ruff, 1994; Bar-Yosef et al., 1996;Mellars, 1996, 1998a,c; Stringer, 1998;Howell, 1998; Hublin, 1998b). The pres-ence of both Neanderthals and H. sapiensin the Levant presents a challenge for behav-ioral interpretation, though some interest-ing reconstructions have been proposed(Lieberman, 1993, 1998a; Lieberman &Shea, 1994). In fact, we hope that the sys-tematic contrast between the European andAfrican records that we provide here mayultimately help to clarify the picture in thesouthwest Asian contact zone.
The number of well-documented lateAfrican Middle Pleistocene or early LatePleistocene sites is not large (Figure 1).Africa is vast, researchers are few, andresearch history is short. Mehlman (1989)points out the contrast between East Africa,with an area of 1·6 million km2 and fewerthan ten excavated sites, and southwesternFrance, with an area of only 21,000 km2 andmore than 100 such sites. West Africa isparticularly poorly represented in therecord. A further factor in our skewed per-ception is the reliance in the literature upona small number of excavated caves on thecoasts of North and especially South Africa(McBurney, 1967; Singer & Wymer, 1982;Volman, 1984; Deacon & Geleijnse, 1988;Klein, 1989a, 1994, 1998; Thackeray,1992). While caves may provide the bestconditions for preservation, the vast majorityof African occupation sites are in open aircontexts. We privilege here the less wellknown but critically important excavatedarchaeological samples from the Africantropical and subtropical regions at the heartof the continent, rather than those from itsnorthernmost and southermost fringes. Thetropics encompass a larger area, were more
continuously occupied, and have climaticregimes more typical of Africa as a whole.Further, the use of terminology derivedfrom Europe in much of the literature treat-ing the Sahara and the Mediterraneanlittoral obscures the uniqueness of the MSAin these regions (Kleindienst, 2000b).
Middle Paleolithic or Middle StoneAge?
Some authors (e.g., Klein, 1989b, 1995,1998) have argued that the Middle StoneAge of Africa is just like the Middle Paleo-lithic of Eurasia, a long and monotonousperiod of flake tools and stagnation, fol-lowed by a rapid revolutionary change totechnological sophistication and economicand social complexity. Klein (1989b: 292,1994: 496) argues that the use of a separateterminology for Africa is a result of historicprecedent, is unfounded in the light of cur-rent research, and should be dropped. Thisview ignores descriptions of the AfricanMSA published just 30 years ago thatstressed its similarities to the EuropeanUpper Paleolithic when the two were con-sidered contemporary (e.g., Clark, 1970).We argue that a separate terminology isrequired for Africa, and that the distinctionbetween ‘‘Middle Paleolithic’’ and ‘‘MiddleStone Age’’ is more than semantic. As wewill show here, this is due in part to thepresence in Africa at an early date ofelements traditionally associated with H.sapiens in the European Upper Paleolithic.
Chronology
The most serious impediment to inter-pretation of the Middle Pleistocene recordhas been the lack of a sound chrono-logical framework. Despite improvementsin sample preparation techniques andincreased selectivity about the materialdated, 14C dates, even using accelerator

488 . . .
mass spectrometry (AMS) become un-reliable between six to seven half-lives (byca. 40 ka at best), due to instrumental limi-tations or to the inevitable contamination bymodern carbon (Gowlett & Hedges, 1986;Wintle, 1996; Ambrose, 1997; Chappellet al., 1998). Further, fluctuations in theearth’s geomagnetic field during the mostrecent glacial period and resulting variationsin cosmic nuclide production may render14C dates in the 40 ka range severalthousand years too young (Mazaud et al.,1991; Taylor, 1996; Joris & Weninger,1998; Schluchter, 1998; see Ambrose,1998a for a useful review).
The conventional K/Ar technique cannotreliably be employed for material youngerthan about 500 ka or perhaps 250 ka(Curtis, 1966). The Argon/Argon (40Ar/39Ar) technique is both accurate and precisewhen applied to younger volcanics(McDougall, 1981; Deino & Potts, 1990;Hu et al., 1994; Renne et al., 1997), thoughsuitable volcanic deposits associated withMSA assemblages are uncommon, even inthe Rift Valley. With rare exceptions (e.g.,Wilson & Hey, 1981; Champion et al.,1988; Tamrat et al., 1995), paleomagnetismis not useful for the long period of normalpolarity since 780 ka. Some of the methodsemployed for this time period, such aselectron spin resonance (ESR), thermo-luminescence (TL), optically stimulatedluminescence (OSL), infrared stimulatedluminescence (IRSL), U-series enhancedby thermal ionization mass spectrometry(TIMS), and amino acid racemization(AAR) are prone to error under the varyingconditions encountered in archaeologicalcontexts. Successful application requiresaccurate reconstruction of sedimentary anddiagenetic processes, ground water action,and temperature history (Hare, 1980; Grun& Stringer, 1991; Schwarcz, 1992; Milleret al., 1993, 1997; Schwarcz & Grun, 1993a;Feathers, 1996; Wintle, 1996; Schwarcz,1998; Schwarcz & Rink, 1998). Concordant
results from multiple techniques based uponindependent theoretical assumptions in-crease confidence. In nearly all cases, newage determinations have increased ratherthan reduced previous age estimates.
Earliest MSA
On purely stratigraphic grounds, the MSAhas long been known to postdate theAcheulian and predate the LSA, althoughtemporal overlap among industries at bothends of the MSA timespan is likely.The high degree of variability in MiddlePleistocene industries makes it difficult toidentify the earliest MSA, and perhaps lendssupport to the idea of multiple contem-porary hominid lineages (McBrearty et al.,1996; McBrearty, 2000).
Late Acheulian and early MSA datescluster between 200 ka and 300 ka, and theAcheulian seems to have disappeared inmost of Africa by about 200 ka. TheAcheulian at Isimila, Tanzania has beendated by U-series to ca. 260 ka (Howellet al., 1972). Acheulian occurrences inthe Kapthurin Formation, Kenya, underlievolcanics dated by K/Ar to ca. 240 ka(Leakey et al., 1969; Tallon, 1976, 1978;McBrearty et al., 1996), but now estimatedby 40Ar/39Ar at ca. 280 ka (Deino &McBrearty, under review). The Acheulian inthe western desert of Egypt is also thoughtto end at either about 200 ka (McHughet al., 1988) or perhaps as much as 350 ka(Wendorf & Schild, 1992). A U-series dateof ca. 174 ka on a late Acheulian occurrenceat Rooidam, South Africa (Szabo & Butzer,1979) seems somewhat anomalous in thiscontext.
Dates for the earliest MSA in East Africaconverge on the interval 240–280 ka. Veryearly K/Ar dates were reported by Evernden& Curtis (1965) for East African MSA siteswhen the method was still in its infancy.Dates of >440 ka and <557 ka for ‘‘pseudo-Still Bay’’ occurrences at Cartwright’s and

489 ’
Wetherill’s sites on the Kinangop Plateau,Kenya, may be accurate, but the strati-graphic relationship of the dated tuffs to theoccupation horizons is less secure (Curtis,personal communication; personal obser-vation). A date of 240 ka was reported in thesame paper for a crystal rich pumiceouslapilli tuff overlying ‘‘Kenya Still Bay’’ arte-facts at Malewa Gorge in the central KenyaRift Valley. A K/Ar date of 180 ka for theEthiopian site of Gademotta, cited for manyyears as the earliest for the MSA (Wendorfet al., 1975), has been revised to 235�5 ka(Wendorf et al., 1994). The earliest MSA inthe Kapthurin Formation, Kenya, appearsto predate volcanics dated by 40Ar/39Ar to�280 ka (McBrearty, 1999; Deino &McBrearty, under review). U-series dates onspeleothem support an antiquity for theearly MSA at Twin Rivers, Zambia, of230 ka or older (Barham & Smart, 1996).U-series and AAR dates from Mumba RockShelter, Tanzania (Mehlman, 1987, 1989)indicate an age for the base of the MSAsequence there of ca. 130 ka, and obsidianhydration indicates an age of ca. 120–50 kafor the MSA at Prospect Farm, Kenya(Michels et al., 1983).
In North Africa, a series of 160 datesusing a variety of methods (OSL, AAR,ESR, TL, U-series) indicates that the MSAat the sites of Bir Tarfawi and Bir SaharaEast in southwestern Egypt beginsca. 230 ka (Miller et al., 1991; Wendorf &Schild, 1992; Bluszcz, 1993; Miller, 1993;Schwarcz & Grun, 1993b). U-series datesindicate that the Aterian at Dakleh oasis inthe western desert of Egypt predates 90 ka,and may be as old as 160 ka (Kleindienst &Wiseman, 1996; Kleindienst et al., 1996;Kleindienst, 2000b). The Aterian of theMediterranean littoral and the Maghrebmay be a bit younger; TL and OSL dates onsands containing in situ Aterian artefacts atthe sites of Uan Afuda and Uan Tabu,Libya, indicate an age for the industry thereof 90 ka to 60 ka (Martini et al., 1996;
Cremaschi et al., 1998), but, as in Egypt, theAterian is preceded in the Maghreb by anunspecialized MSA industry of far greaterantiquity (Hublin, 1993; Debenath, 1994;Kleindienst, 2000b). Some investigatorsplace the end of the Aterian at 20 ka, onthe basis of 14C on lacustrine carbonate(Wengler & Vernet, 1992; Wengler, 1997;Kleindienst, 2000b), but this method hasproved unreliable in other contexts, andthese dates should probably be treated asinfinite. The main body of Aterian sites thuspredates the aridification of the Saharaduring the last glacial maximum that beginsat ca. 40 ka (Wendorf & Schild, 1993;Wendorf et al., 1993a; Close, 1984;Cremaschi et al., 1998).
In the interior of South Africa, siteswhere MSA assemblages overlie Acheulianremains, such as Cave of Hearths (Mason,1962), are undated. The oldest dates for theMSA in South Africa are ESR determi-nations from the sequence at Florisbad(Grun et al., 1996). An undiagnostic MSAassemblage from the basal deposits atFlorisbad (Units N, O, and P), is estimatedat ca. 280 ka. Units higher in the sectioncontaining in situ MSA artefacts date toca. 160 (Units M–G) and 120 ka (Unit F)(Kuman & Clarke, 1986; Kuman et al.,2000). At Border Cave TL and AAR datesfor strata 4WA and 5BS are at least 100 ka(Grun et al., 1990; Miller et al., 1993). Theunderlying strata 5WA and 6BS may beconsiderably older (Grun et al., 1990), per-haps as much as 195 ka (Butzer et al., 1978).
At coastal South African sites evidence forMSA habitation prior to the last interglacialis lacking. Middle Pleistocene deposits mayhave been scoured out of coastal caves dur-ing marine high stands preceding stage 5,and countless early sites may lie on the nowinundated offshore coastal platform. AtKlasies River features of the geomorphologyand microfauna, the oxygen isotope con-tent of marine shell, ESR dates on fossilmammalian teeth, and U-series dates on

490 . . .
speleothem indicate an age for the earliestMSA levels (MSA I) of >110 ka, perhaps130 ka (Deacon, 1988, 1989, 1993b;Deacon & Geleijnse, 1988; Deacon &Shurman, 1992; Schwarcz & Grun, 1993a;Thackeray, 1992), although South AfricanMSA technology no doubt has its begin-nings substantially earlier (Volman, 1984).Probably the most convincing evidence forthe antiquity of the MSA at Klasies is pro-vided by Shackleton (1982) who notes thatthe last time the ocean was isotopically aslight as the shells from the MSA I levels wasduring the last interglacial (stage 5e) datingto 118 ka. Further support is provided bysands in the lowermost sedimentary unitthat were derived from a nearby beach, andsea levels have not been high enough toprovide the source material since thattime (Hendey & Volman, 1986; Deacon &Geleijnse, 1988; Deacon & Shurman,1992).
Earliest LSA
Dating the end of the MSA and the begin-ning of the LSA is also somewhat problem-atic. First, in much of tropical Africa theMSA–LSA transition lies at or beyond thelimits of the radiocarbon method, and 14Cdates reported in the 1970s for the earliestLSA (e.g., Beaumont & Vogel, 1972a,b)should be regarded as infinite (Ambrose,1997, 1998a). In East and Central Africathe record for the LSA begins well before40 ka at sites such as Enkapune ya Muto,Kenya (Ambrose, 1998a) and at MumbaRock Shelter and in the Naisusu Beds,Tanzania, (Kokis, 1988; Mehlman, 1989;Manega, 1993). Second, there is con-siderable disagreement over which criteriadefine the earliest LSA. The presence ofmicroliths, bladelets, microblade cores, andbacked geometric forms, or the absence ofpoints or other typical MSA elements arevariously used as criteria in assigning assem-blages to the LSA. This leads to inevitable
problems. For example, van Noten (1977)has referred the industry from MatupiCave, D.R. Congo, to the LSA, based onthe presence of microlithic debitage. Hegives an age estimate of ca. 50 ka for thisassemblage, based on extrapolation from14C dates higher in the sequence. Butbecause classic microblade cores and backedelements are lacking, the attribution of theindustry to the LSA has been questioned.On the other hand, at sites such as RoseCottage Cave (Kohary, 1989), Sehonghong,and Strathalan, South Africa, the earliestbladelet levels, dated by 14C to ca. 26 ka,are referred to a final or ‘‘microlithic’’MSA by Wadley (1991, 1993, 1995, 1997;cf. Mitchell, 1994), due to the presence offoliate points, typical of the MSA, thatare found in association with the blade-lets and microblade cores, though cir-cumstances of recovery at Sehonghongmay make questions of associationproblematic.
The Howiesons Poort is no longer seen asa transitional industry between the MSAand the LSA, and the concept of a ‘‘SecondIntermediate’’ was abandoned along withthe Magosian industry. Yet controlledexcavation at stratified sites with fairlycontinuous sequences has documentedindustries containing both LSA and MSAelements that overlie industries which areunquestionably MSA and underlie thosewhich fall clearly in the LSA. The un-dated MSA 1 level at Sodmein Cave, onthe Red Sea coast of Egypt, as describedby van Peer et al. (1996) contains bladesand other elements indicating that it maybe a transitional industry. The MumbaIndustry, a late MSA entity contain-ing backed geometrics, typical of theLSA, has been described from Bed Vat Mumba Rock Shelter, Tanzania,by Mehlman (1979, 1987, 1989). AtMumba, backed elements increase in fre-quency through the section, as mean arte-fact size declines. U-series dates on bone

491 ’
from middle Bed V indicate an antiquityof ca. 65 ka, and this age is confirmed byAAR dates on ostrich eggshell rangingfrom 45–65 ka from the upper part of thesame unit (Kokis, 1988; Brooks et al.,1993b). Similar ‘‘transitional’’ industriesare known from Botswana, Zimbabwe,and South Africa, and their presence con-firms the lack of abrupt discontinuities inthe African prehistoric record.
At White Paintings Shelter, Botswana, anMSA–LSA transitional unit with a bonepoint, bladelets, and a large backed crescent,but without the MSA points found in theunderlying units, is dated by several tech-niques to 50–38 ka (Robbins et al., 1994,under review; Kokis et al., 1998). A similarindustry is known from �Gi, Botswana(Brooks & Yellen, 1987; Brooks et al.,1990). In Zimbabwe, the MSA–LSA tran-sition appears to be particularly long andcomplex, with some sites containing MSApoints as late as 13 ka (Walker, 1990,1991, 1995; Larsson, 1996). At Nswatugi,Zimbabwe, large backed crescents in theuppermost MSA with an infinite radio-carbon date (>42 ka) may represent theearliest phase of the post-MSA or late MSATshangula industry (Cooke, 1966, 1969) orperhaps an MSA industry related either to theHowiesons Poort (Walker, 1995) or to theMumba industry (Mehlman, 1979, 1989).
In South Africa, with one of the best-documented records of Late Pleistocenehuman occupation on the continent, threedifferent scenarios have been described. Oneis a major hiatus at many sites betweenthe final MSA, predating 60 ka, and theearliest LSA, at or after 20 ka (Brooks& Robertshaw, 1990; Mitchell, 1990;Parkington, 1990; Walker, 1990). A second,as at Border Cave, is the appearance ofan early LSA (ELSA) by 38 ka, based on14C and AAR dates on ostrich eggshell(e.g., Beaumont, 1973b; Beaumont et al.,1978; Miller et al., 1993), with no appar-ent occupational hiatus.6 A third is a
late appearance of the LSA at about 20 kaat sites such as Rose Cottage Cave,Sehonghong and Strathalan (Wadley &Harper, 1989; Opperman & Heydenrych,1990; Wadley & Vogel, 1991; Wadley,1993, 1997). It is unclear whether this dis-crepancy is due to problems with the radio-carbon dating or with the definition of theLSA, or whether it reflects a real persistenceof the MSA until very late at the southern tipof the continent.
Archaeological signals for ModernBehavior
The European Upper Paleolithic, because itis known to be the product of H. sapiens, isoften used as a standard for modern humanbehavior, which is contrasted with theEuropean Middle Paleolithic producedby the Neanderthals (e.g., White, 1982;Clark & Lindly, 1989, 1991; Hayden, 1993;Klein, 1995; Mellars, 1995). The litera-ture converges upon a number of commoningredients thought to characterize modernhuman behavior:
� Increasing artefact diversity.� Standardization of artefact types.� Blade technology.� Worked bone and other organic materials.� Personal ornaments and ‘‘art’’ or images.� Structured living spaces.� Ritual.� Economic intensification, reflected in the
exploitation of aquatic or other resourcesthat require specialized technology.
� Enlarged geographic range.� Expanded exchange networks.
6. Problematic ESR dates of ca. 28–31 ka for thisindustry at Border Cave (Grun et al., 1990; Schwarcz &Grun, 1993a) are 30% younger than 14C ages for thesame units and may reflect systematic error in estimat-ing the annual dose rate (Miller et al., 1993). A similarproblem may exist for the uppermost MSA levels atKlasies River, dated by ESR to 40 ka (Grun et al.,1990), but by 14C to >50 ka (Deacon, 1993b) and byAAR to 60 ka (Brooks et al., 1993b).

492 . . .
At first glance, this inventory is remi-niscent of the sort of trait list that Childe(1928, 1950) and others once used to definecivilization. However, they reveal assump-tions about underlying hominid capabilities.We would argue that modern humanbehavior is characterized by:
� Abstract thinking, the ability to act withreference to abstract concepts not limitedin time or space.
� Planning depth, the ability to formulatestrategies based on past experience and toact upon them in a group context.
� Behavioral, economic and technologicalinnovativeness.
� Symbolic behavior, the ability to repre-sent objects, people, and abstract con-
cepts with arbitrary symbols, vocal orvisual, and to reify such symbols incultural practice.
Tangible traces of these early behavioralshifts toward modernity can be seen in theAfrican archaeological record, and can betied explicitly to hominid cognitive andcultural capabilities. These are set out inTable 3.
� Ecological aspects of the record reflecthuman abilities to colonize new environ-ments, which require both innovation andplanning depth.
� Technological features reveal humaninventiveness and capacity for logicalthinking.
Archaeological signatures of modern human behavior
EcologyRange extention to previously unoccupied regions(tropical lowland forest, islands, the far north in Europe and Asia)Increased diet breadth
TechnologyNew lithic technologies: blades, microblades, backingStandardization within formal tool categoriesHafting and composite toolsTools in novel materials, e.g., bone, antlerSpecial purpose tools, e.g., projectiles, geometricsIncreased numbers of tool categoriesGeographic variation in formal categoriesTemporal variation in formal categoriesGreater control of fire
Economy and social organizationLong-distance procurement and exchange of raw materialsCuration of exotic raw materialsSpecialized hunting of large, dangerous animalsScheduling and seasonality in resource exploitationSite reoccupationIntensification of resource extraction, especially aquatic and vegetable resourcesLong-distance exchange networksGroup and individual self-identification through artefact styleStructured use of domestic space
Symbolic behaviorRegional artefact stylesSelf adornment, e.g., beads and ornamentsUse of pigmentNotched and incised objects (bone, egg shell, ocher, stone)Image and representationBurials with grave goods, ocher, ritual objects
Table 3

493 ’
� Economic & Social features showhuman abilities to draw models from indi-vidual and group experience, to developand apply systematic plans, to conceptu-alize and predict the future, and to con-struct formalized relationships amongindividuals and groups.
� Symbolic features demonstrate acapacity to imbue aspects of experiencewith meaning, to communicate abstractconcepts, and to manipulate symbols as apart of everyday life.
Ecology
Increased geographic rangeExpansion of human populations intochallenging habitats by means of improvedtechnology is seen as a sign of cognitivesophistication and social complexity(Torrence, 1983; Klein, 1989b; Davidson &Noble, 1992; Jones, 1992; Soffer, 1994;O’Connell & Allen, 1998). This trend isseen in the African MSA. MSA sites inAfrica are more numerous than those of theAcheulian (Figure 1), which may simplyreflect preservational bias and an increase inpopulation size. However, the fact that theyare found in virtually every corner of thecontinent argues against population expan-sion as the single explanatory factor. Thisextension into previously uninhabited zonesis thought by Clark (1964, 1965, 1970,1972, 1981, 1982, 1988, 1993) to illustratethe improved adaptive abilities of MSAhominids over those of their Acheulianpredecessors. The geographic distribution ofMSA sites does indicate human adaptationsto a wide range of challenging habitats. Twoof these, desert and forest, merit specialconsideration.
Sparse Acheulian sites indicate sporadicoccupation of what is now the Sahara,Namib, Karoo, and Kalahari deserts, butour understanding of the significance ofmost of these is tempered by three factors:(1) good data for the interior of the Sahara is
lacking; (2) the ready visibility of Acheulianbifaces tends to inflate the numbers ofAcheulian sites relative to those of otherperiods; and (3) the environmental con-ditions in these regions at the time of occu-pation are unknown. The few known datessuggest that these occupations date to thelate Middle Pleistocene (Miller et al., 1991;Miller, 1993). Associated lake and springdeposits indicate a much wetter habitatthan at present (Wendorf et al., 1993a,b;Churcher et al., 1999; Nicoll et al., 1999).Sites of the MSA Aterian industry do indi-cate widespread habitation of the Saharabetween ca. 90 ka and ca. 40 ka. Ateriansites are found in habitats varying frommarine to mountain to semiarid desert, andoccupation was not confined to humidperiods (Marks, 1975; Williams, 1976;Clark, 1980; Debenath, 1994; Kleindienst,2000b). For South Africa, Deacon (1989)observes that MSA occupations, in contrastto those of the Acheulian, are not restrictedto sites near water sources. He suggests theuse of water containers in the MSA anda perception of the environment indis-tinguishable from that of people of laterperiods.
Human life in a tropical rainforestenvironment, with its dispersed fauna, andfruit and root resources that are ofteninaccessible and toxic, requires complextechnology. No modern human groups areknown to subsist in the forest solely byforaging without a source of domesticatedcarbohydrates, and this adaptation has beenheld to have been impossible for prehistoricgroups (Bailey et al., 1989). Others, how-ever, argue that it is feasible, particularly iffish are incorporated into the diet (Gragson,1992). Evidence for early occupation oftropical forest environments is equivocal.Acheulian sites are very rare for the CongoBasin. Clark (1964, 1965, 1970, 1975,1981) has suggested that the technology ofthe Sangoan industry provided the equip-ment necessary for a forest adaptation.

494 . . .
However, Sangoan artefacts are found inregions far outside the Congo Basin, such asat Isimila, Tanzania (Howell et al., 1962,1972), and perhaps in the KapthurinFormation, Kenya (McBrearty et al., 1996;Cornelissen, 1995), where they are clearlynot elements of forest extractive technology.Other sites occur in regions of tropicalAfrica that are now forested, but whereevidence indicates arid or semiarid con-ditions at the time of occupation, probablyduring cold isotope stage 6 (195 ka–130 ka)(McBrearty, 1987, 1993a,b; Clark, 1988).
The MSA is represented in the greaterCongo Basin by the Lupemban industry,characterized by beautifully made lanceolatepoints. However, there are few excavatedLupemban sites (Cahen, 1978; McBrearty,1986, 1988), and virtually no dates orpaleoenvironmental data for this industry.Many Lupemban sites formerly regarded asin good context have been found to beprofoundly bioturbated (Cahen, 1976).LSA sites in the Congo Basin are reportedby Fiedler & Preuss (1985). Mercader et al.(2000c) document a sequence of LSA oc-cupations in the Ituri forest in associationwith paleobotanical indicators of forestconditions dating back to the last glacialmaximum, showing that use of the forest byhunter gatherer groups has great antiquity.Mercader & Marti (1999a,b, 2000;Mercader et al., 2000a,b) document twoprobable MSA sites in West Africa thatsuggest occupation of the tropical forest.Njuinye, Cameroon, and Mosumu, Equator-ial Guinea are dated by AMS 14C radio-carbon to >30 ka and >34 ka, respectively(Mercader & Marti, 2000; Mercader,personal communication). Direct paleo-environmental evidence is lacking at the sitesthemselves, but pollen records and plantmacrofossils are preserved at nearby paleobo-tanical sampling localities with a comparabletime depth. These show that while tempera-tures were no doubt cooler in this region ofWest Africa during the last glacial maximum,
rainfall remained adequate to support tropi-cal forest species, even in areas now occupiedby savanna vegetation (Mercader & Marti,2000).
Technology
BladesThe earliest true Upper Paleolithic industry,the Aurignacian, appears in Europe atapproximately the same time as the firsttraces of H. sapiens ca. 40–45 ka (White,1982, 1989a, 1992; Bischoff et al., 1989;Cabrera Valdes & Bischoff, 1989; Klein,1989a; Mellars, 1989a, 1993, 1996,1998b,c; Otte, 1990; Bischoff et al., 1994;Straus, 1994; Bar-Yosef, 1998). Thoughsome would argue for continuity in lithicindustries across the European Middle toUpper Paleolithic boundary (e.g., Otte,1990; Clark & Lindly, 1989; Straus, 1983,1994, 1995; d’Errico et al., 1998), it seemsincreasingly likely that Upper Paleolithictechnology was introduced into Europe byH. sapiens migrants (Mellars, 1999). In somecases, aspects of Upper Paleolithic tech-nology were adopted by the Neanderthalresidents (Demars & Hublin, 1989; Mellars,1989b, 1998a,b; Farizy, 1990; Ruiz &Hublin, 1994).
European Upper Paleolithic technologyis dominated by blades, and thus bladeproduction has been considered a keyingredient of the ‘‘human revolution’’ (e.g.,Mellars & Stringer, 1989). At the simplestlevel, blades are simply flakes with abreadth:length ratio �0·5. Although some(e.g., Wurz, 1999) distinguish betweenblades and flake-blades, this practice is notfollowed here. Long narrow flakes will beproduced fortuitously in the course ofalmost any kind of flake production, but therepeated, consistent production of bladesrequires a deliberate series of technologicalsteps. Elongated Levallois cores struckrepeatedly from either end can yield bladeswhich are indistinguishable from those

495 ’
produced on the cylindrical or pyramidalcores more typical of the European UpperPaleolithic. Blades are a more efficient use oflithic raw material than either core or flaketools, as they represent a greater length ofcutting edge per unit volume of stone.Blades also functioned as blanks with stand-ardized proportions that were used as thebasis of composite tools in many late prehis-toric industries. Blade production, whetherby direct or indirect percussion, requires thecognitive skills to perceive artefact forms notpreordained by the raw material and tovisualize the manufacturing process in threedimensions, in addition to the dexterity tocarry out a complex series of operationsand corrections as the process advances.However, convincing cases can be madefor the cognitive sophistication and manualdexterity required to produce earlier artefactforms, such a handaxe (Roche & Texier,1991, 1995; Texier & Roche, 1993) or aLevallois flake (Roebrooks et al., 1988; vanPeer, 1992).
Blade production is often treated in theliterature as a uniquely modern humaninvention with a recent genesis, but in fact,the origins of blade production are morecomplex. While most European MiddlePaleolithic industries are flake-based, it isnow realized that assemblages from severalMiddle Paleolithic sites in France, Belgium,England, and Germany have a blade com-ponent. Some of these sites may date to onlyca. 60 ka, while others, including Coquelles(France), Crayford (England), and LeRissori (Belgium), may be as old as 250 ka ifpaleoclimatic interpretations are correct(Conard, 1990; Revillion & Tuffreau, 1994;Revillion, 1995). In the Levant, if the TLdates are accurate, the Acheuleo-Yabrudian(Mugharan) industry, which containsblades, may predate 350 ka, though the ESRchronology at the key site of Tabun indi-cates an age for this industry of <250 ka.The lack of concordance may perhaps bedue to unequal external U dose in different
parts of the cave as a result of the strati-graphic heterogeneity of the cave fill(Jelinek, 1982, 1990; Grun et al., 1991;Grun & Stringer, 1991; Schwarcz & Grun,1993a; Bar-Yosef, 1994, 1995a, 1998;Schwarcz, 1994; Mercier et al., 1995;Mellars et al., 1997).
In Africa, blade production has greatantiquity. Blades are found in the pre-Aurignacian industry at the Haua Fteah,Libya (McBurney, 1967; Chazan, 1994),whch has been correlated with isotopic stage5 on faunal grounds and therefore esti-mated by Klein & Scott (1986) to date toca. 127–75 ka. Blades produced by Levalloistechnology also figure prominently in theMSA of the Nile Valley beginning before200 ka (van Peer, 1992, 1998; Kleindienst,2000a). In South Africa, the makers of MSAindustries routinely manufactured bladesfrom a variety of core types (Sampson, 1972,1974; Volman, 1984; Kuman & Clarke,1986; Kuman, 1989). This technology wasclearly in place by 120 ka, but its beginningsare poorly constrained temporally.
In East Africa, the presence of volcanicsmakes possible a chronology based on K/Arand, more recently, 40Ar/39Ar. In much ofKenya, Tanzania, and eastern D. R. Congo,MSA lithic industries are based not uponblades, but upon flakes produced fromradial, Levallois, and other less formalizedcore types. However, MSA sites in northernKenya and the Horn of Africa also containquantities of early blades. At MSA sites atGademotta, Ethiopia, blade index values(reflecting the numbers of blades and bladecores relative to other kinds of artefacts inthe assemblages) range from 13·6% in levelsdated to �235 ka to 37·7% at younger sites(Wendorf & Schild, 1974). Blades also con-stitute a significant proportion of the debit-age at the MSA sites at Aduma in theMiddle Awash Valley of Ethiopia (Brooks,1996, 1999).
The Kapthurin Formation, Kenya, pro-vides unequivocal evidence for early African

496 . . .
blades in a late Acheulian industry. Trachyteblades were recovered from both surface andexcavated contexts at the site of GnJh-03(Leakey et al., 1969). The material occursin situ at a depth of ca. 3 m below the base ofK4, a volcanic unit. Conventional K/Ardates for K4 indicated an age on the order of240–250 ka (Tallon, 1978); a current pro-gram of redating by 40Ar/39Ar (Deino &McBrearty, under review) indicates that thetrue age lies closer to 280 ka.
Between 20% and 30% of the cores fromsite GnJh-03 can be classified as bladecores, while about one-quarter of the exca-vated flakes can be classified as blades(Leakey et al., 1969; Cornelissen, 1992).The remainder of the flakes were producedby radial and Levallois core reduction.Experimental knapping in local trachyteand comparison with the archaeologicalsample reveals that the technique used inblade production was direct percussion,most likely with a hammerstone, thoughsoft hammer percussion cannot be ruledout (Texier, 1996; McBrearty & Texier,n.d.). Approximately 13% of the assem-blage from GnJh-03 can be conjoined(Cornelissen, 1992), and examination ofrefitting sets permits detailed reconstruc-tion of blade production techniques,including both Levallois and semi-cylindrical blade removals (Figure 3)(McBrearty & Texier, n.d.). Both unidirec-tional and bidirectional blade removals arerepresented; blades show both plain andfaceted platforms. Distinct levels of skill arereflected in the different refitted sets, andthe most skilled knapper was able to pro-ceed through a long series of blade re-movals to core exhaustion. The resultingblades are remarkable in their length,thinness and flatness (Figure 4). TheKapthurin Formation blades demonstratethe presence of a fully conceptualized, well-executed method of blade production, andhigh level of technical competence in EastAfrica before 280 ka.
MSA pointsThe presence of stone points has been con-sidered a defining characteristic of the MSAsince the inception of the term (Goodwin &van Riet Lowe, 1929). Pointed flakes andflake-blades seem to have been deliberatetarget forms for many MSA toolmakers, andretouched points are among some of theearliest MSA artefacts at 235 ka (Wendorfet al., 1994). The large numbers and carefuldesign of points in the retouched componentof many MSA assemblages, and the relativelack of emphasis on scrapers, are majordifferences between the African MSA andthe classic Mousterian of southwesternFrance. For this reason, the Lower and
Figure 3. Semi-cylindrical core from site GnJh-03 inthe Kapthurin Formation, Kenya. � Sally McBrearty.
Figure 4. Blades from site GnJh-03 in the KapthurinFormation, Kenya. Figure courtesy of Pierre-JeanTexier. C.N.R.S. Sophia Antipolis, France.

497 ’
Middle Paleolithic typologies of Bordes(1961) and Debenath & Dibble (1994) arenot useful in describing most African MSAindustries, particularly those from south ofthe Sahara. It is now clear that bone pointsalso are characteristic of some African MSAindustries. This difference between theAfrican and European records renders itimportant to understand the manner ofuse and the stylistic implications of MSApoints.
Hafting was probably routine in mostMSA contexts. MSA points are oftendeliberately modified to facilitate haftingeither by thinning at the butt, or, in the caseof Aterian points, by the fabrication of atang. While seldom preserved in thearchaeological record, vegetable gums arefrequent in the African ethnographic recordas a hafting medium, and traces of mastichave been found on a blade in MSA level Eat Apollo 11 (Wendt, 1976; Vogelsang,1998).
Many MSA points were most likely usedto dispatch game. Milo (1998) has reporteda point tip embedded in a cervical vertebraof the large bovid, Pelorovis [see Figure 8(a)]as well as a similar puncture wound in aneland thoracic vertebra from MSA depositsat Klasies River, South Africa. Impact scarshave been observed on MSA points from anumber of African sites, including �Gi,Botswana, and Blombos, South Africa(Kuman, 1989; Henshilwood & Sealy, n.d.).Wendorf & Schild (1993) have argued thatthe points from the Lake Ziway region ofEthiopia show signs of use as cutting andscraping tools. Kuman (1989), however, hasdemonstrated that points from �Gi werehighly curated tools that performed manyfunctions, including cutting, over their longlife spans. The importation of finishedNubian points to Sodmein Cave, Egypt (vanPeer et al., 1996; van Peer, 1998) wouldseem to indicate use of the site as a specialpurpose hunting station, and the largenumbers of finished pressure flaked foliate
Still Bay points at Blombos, South Africaseems likewise to indicate a hunting stationor a specialized production encampment(Henshilwood & Sealy, n.d.).
Retouched points from sites throughoutAfrica are the appropriate size for projec-tiles; they are carefully made, thin, andsymmetrical. Further, points made on endand corner struck flakes from Botswana andZimbabwe show deliberate shaping to createequal distribution of mass around themidline, suggesting aerodynamic design(Yellen, personal communication). Whilethese may have been used to arm the busi-ness end of stabbing or throwing spears,they could also have been delivered by bowand arrow. The small size of some late MSApoints (e.g., Gresham & Brandt, 1996) inparticular suggest their use with bow andarrow. Western European points frequentlylack basal thinning to facilitate hafting, orthe impact scars indicating use as spearsdescribed by Shea (1988) for the LevantineMousterian. In the late Mousterian ofeastern Europe (e.g., Marks & Chabai,1996) and Portugal (Marks & Brugal, 2000)there are pointed forms which may indeedhave served as hafted projectiles, but theseappear to postdate 60 ka. It is tempting tosurmise that this late flowering of pointtechnology in eastern Europe reflects a bowwave effect, the result of contact withhominids to the east and south, who hadpossessed this technology for many tens ofthousands of years.
Social organization
Regional variety in MSA points
The diversification of MSA toolkits and thevarying proportions of different artefactclasses at different sites no doubt reflectregional traditions as well as different extrac-tive activities. Points comprise the singlefunctionally equivalent class of artefacts thatbest reflects regional differences, and signals

498 . . .
the presence of regional styles (Figure 5).Projectile points have traditionally beenused by archaeologists to define later prehis-toric industries worldwide. They are thoughtto encode stylistic information for theirmakers for two reasons: (1) point design istightly constrained by aerodynamic andhafting requirements, and therefore success-ful designs tend to be closely replicated(Wilmsen, 1974; Knecht, 1993); (2) withingroups, sharing or exchange of projectilepoints imposes design limits. Ethnographi-cally, individuals have been observed tocontinue to manufacture projectiles longafter their declining eyesight, reflexes, andendurance have diminished their own suc-cess as hunters. Among the !Kung and otherpeople, projectiles are provided to successfulhunters in exchange for a claim on the meat(Yellen, 1977; Lee, 1979). Thus, the makermust conform to a familiar template so thatthe hunter will be able to perform with
tools produced by a variety of craftsmen(Wiessner, 1982, 1983). This contributes toabrupt discontinuities in projectile pointstyle that coincide with the boundaries of theethnic groups within which projectiles areexchanged. Such geographic patterning ofform in functionally equivalent artefacts hasbeen seen as the archaeological signature ofstyle, however it is construed (Sackett,1977, 1982, 1990; Wobst, 1977; Hodder,1985; Conkey, 1990). On the whole, there isnot a high degree of variability in the MiddlePaleolithic of western Europe, wherepointed forms are rare (Chase & Dibble,1987).
What is the patterning of point styles inthe African MSA? For most of Africa, we donot have data that are sufficiently fine-grained to construct detailed local mapsthat reveal the kind of stylistic boundariesknown from the ethnographic record. Ona continent-wide scale, however, regional
Figure 5. Map of distribution of point styles in the African MSA (after Clark, 1993, Figure 1). � SallyMcBrearty & Alison S. Brooks.

499 ’
styles can be recognized (Figure 5). TheAterian is characterized by projectile pointsand other artefacts with a carefully craftedtang for hafting. It occurs in North Africa atabout 20�–15�N, from Morocco in the westto the western desert of Egypt in the east(Debenath, 1994; Kleindienst, 2000b). TheNubian complex, containing truncated andfacetted Nazlet Khater points, occurs in theNile Valley (van Peer, 1991, 1998). TheLupemban industry is found in the greaterCongo basin as far south as Namibia(MacCalman & Viereck, 1967) and as fareast as the Lake Victoria region. It is char-acterized by lanceolate points, some ofwhich are thin, skillfully made, and mayexceed 30 cm in length (McBrearty, 1988).Foliates, narrow foliates, and triangularpoints, both unifacial and bifacial, are foundat East African Rift Valley sites.
In the absence of a firm chronology, it isimpossible to distinguish chronological fromgeographical effects, but the impression is ofpronounced stylistic variation within theMSA of East Africa. Clark (1988: 297) notesthat to the extent that stylistic differences canbe recognized in the MSA of the Horn,
‘‘these might be the preferential forms repre-sented by the markedly subtriangular points atGorgora, the pointed leaf-shaped forms at ProcE{pic and Gademotta, [and] the Levalloispoints at Midhishi’’.
Late MSA points of the Horn are verydistinctive and can be distinguished fromthose of Kenya and Tanzania bytheir increasing elongation and small size(Clark, 1954; Gresham, 1984; Brandt,1986; Gresham & Brandt, 1996). In Eastand Central Africa, MSA points have agreat antiquity. At Mumba Rock Shelter,Tanzania, they date to as early as 130 ka(Mehlman, 1989, 1991) and to about230 ka at Twin Rivers, Zambia (Barham& Smart, 1996), and Gademotta andKukuleti, Ethiopia (Wendorf et al., 1994).
Within South Africa a number of MSAvariants were defined by early workers.
Of these, the Still Bay (Goodwin, 1928),Howiesons Poort (Stapleton & Hewitt,1927, 1928), and perhaps the Bambata(Armstrong, 1931) and Pietersburg indus-tries (Mason, 1962, 1993) seem to haveclear regional and chronological specificity.Although the term Still Bay was subse-quently appropriated for use as far afield asKenya and Zambia (Leakey, 1931; Cooke &Paterson, 1960), the Still Bay as originallydefined by Goodwin & van Riet Lowe(1929) is confined to the southern Capecoast of South Africa. It contains a variety ofpointed forms, but the elongated ‘‘willowleaf ’’ Still Bay points are highly distinctiveand limited to the Cape region. They arecharacterized by shallow invasive retouch,and are usually executed on good qualityraw materials including chert, quartzite andsilcrete. While the industry is known from anumber of surface occurrences and earlyexcavations, the only excavated samplefrom secure context is that reported byHenshilwood & Sealy (1997) from the siteof Blombos, where 52% of the retouchedtools are bifacial points.
The Howiesons Poort industry, contain-ing small, well-retouched unifacial andbifacial foliate points, as well as backedgeometrics, may extend over a wider regionof southern Africa than the Still Bay. Whileit is concentrated at sites on the Cape, suchas Klasies River (Singer & Wymer, 1982;Deacon, 1989; Deacon & Wurz, 1996;Wurz, 1997, 1999) or Montagu Cave(Keller, 1973), materials from Apollo 11rock shelter on the Namibia–South Africaborder (Wendt, 1972, 1976; Vogelsang,1996, 1998), Cave of Hearths in theTransvaal, and Border Cave on theZululand–Swaziland border (Mason, 1962,1993; Beaumont et al., 1978; Beaumont,1980) have also been attributed to theHowiesons Poort. As yet, no HowiesonsPoort occurrences are known from Namibianorth of Apollo 11, or from Botswana. Earlyindustries with backed segments and MSA

500 . . .
points are known from Zimbabwe (Cooke,1971) and from Tanzania (Mehlman,1989), but these appear to be somewhatlater in time and do not contain the fullarray of features described for the SouthAfrican Cape Province Howiesons Poortsites.
The Pietersburg industry is centered inGauteng, but it is not well dated (but seeMiller et al., 1991). It is characterized byelongated points on blades, more often uni-facial than bifacial, and with trimming con-fined to the periphery (Mason, 1962).7 Acommon form is a long foliate unifacialpoint with ventral bulbar reduction. TheBambata industry of Zambia, Zimbabwe,Namibia, and Botswana is distinguished byshort, broad foliate and triangular points,both unifacial and bifacial, with retouchranging from marginal to invasive (Kuman,1989). It is dated at �Gi by TL and AARon ostrich eggshell to 70–80 ka (Brookset al., 1990; Franklin & Hornyak, n.d.). Inbrief, the African MSA is characterized bywell-differentiated regional point styles, andpoints in tropical Africa predate any knownfrom Europe by tens of thousands of years.
Technology
Geometric macroliths and microlithsBacking or blunting of the edges of artefactsis a form of retouch characteristic of UpperPaleolithic and Mesolithic industries of LatePleistocene and Holocene Europe. It isusually performed on blades and is con-sidered a feature of mode 4 technology. Inthe European Mesolithic and Neolithic,small tools were produced by backing bladesor blade segments to create standardizedgeometric shapes, including triangles, tra-pezes, and crescents (mode 5 technology).Backed segments <20 mm (e.g., Andrefsky,1998) or <30 mm (Clark & Kleindienst,
1974) in length are termed microliths. Theresulting small tools are usually mounted inmultiples as components of complex com-posite implements, such as arrows or cuttingtools. They are an advance over simpleflakes and blades in terms of raw materialeconomy, and they clearly signal a modernapproach to technology. They facilitateretooling, as damaged segments can bereplaced without overhauling the entireimplement. They are unknown in Europeprior to the Uluzzian industry of peninsularItaly, dated to ca. 35 ka, and do not becomecommon or widespread in Europe until after20 ka (Palma di Cesnola, 1993; d’Erricoet al., 1998).
In Africa, geometric microliths are thehallmark of the LSA. The sequence atMumba Rock Shelter, Tanzania, documentsthe first appearance of this technology inEast Africa. In the Mumba industry in lowerBed V (Figure 6, bottom row), the earliestgeometrics are rare, macrolithic, and madeon flakes from Levallois or radial cores.Later in Bed V, the geometrics increase innumber, decrease to microlithic size, and aremade on blades (Mehlman, 1989). BothU-series and AAR dates indicate an age of65 ka from Middle to Upper Bed V for theappearance of geometric microliths atMumba (Mehlman, 1989; Brooks et al.,1993b).
7. The Later Pietersburg industry (e.g. Cave ofHearths Bed 9) contains backed crescents, and may beattributed to the LSA or the Howiesons Poort.
Figure 6. MSA geometric microliths; top row, KlasiesRiver Howiesonspoort, after Wurz (1997); bottom row,Mumba Industry, Mumba Rock Shelter, level V, afterMehlman (1989). � Sally McBrearty.

501 ’
In South Africa, geometrics appearsomewhat earlier, in the Howiesons Poortindustry (Figure 6, top row). Goodwin &Van Riet Lowe (1929) first described theHowiesons Poort on the basis of materialreported by Stapleton & Hewitt (1927,1928) from the type site near the Cape coastof South Africa. Like the Mumba industry,the Howiesons Poort contains items charac-teristic of both the MSA and the LSA.MSA elements include unifacial or bifacialfoliate or lanceolate points, sometimes withfaceted butts that are ventrally trimmed, aswell as scrapers and burins made on flakesor blades produced from single platform,double platform, radial, and irregular cores.Features more common to LSA industriesare the small size of the artefacts overall(often <30 mm), a greater reliance on exoticfine grained raw materials, and the presenceof geometrics. Singer & Wymer (1982) andWurz (1997) declare that the Klasies RiverHowiesons Poort blades are punch-struck, afeature characteristic of European UpperPaleolithic blades, but they report no exper-imental basis for this observation, and asAmbrose & Lorenz (1990) remark, distin-guishing blades struck with a punch fromthose produced by soft hammer presents adifficult challenge (cf. Bordes & Crabtree,1969).
The Howiesons Poort was considered apossible ‘‘transitional’’ industry betweenthe MSA and LSA, but subsequent exca-vations have shown that the HowiesonsPoort is interstratified within the MSA at anumber of sites in South Africa, includingKlasies River (layers 10–21 in shelter 1Aand in layers 1–5 in cave 2) (Singer &Wymer, 1982; Deacon et al., 1984; Deacon,1989), Boomplaas, (member OCH)(Deacon, 1979, 1989), Montagu Cave(layer 2) (Keller, 1973), and Border Cave(layer III) (Beaumont, 1973b; Butzer, 1978;Beaumont et al., 1978). While the exca-vators of Klasies River considered the inter-stratified MSA and Howiesons Poort
assemblages to have been made by popu-lations with different technological tra-ditions (Singer & Wymer, 1982), laterinvestigators have seen them as the productof shifting territorial organization, rawmaterial acquisition, and symbolic signaling(Ambrose & Lorenz, 1990; Wurz, 1997,1999; Deacon & Deacon, 1999).
Radiocarbon dates available in the 1960sseemed to indicate that the HowiesonsPoort should be incorporated within theLSA (Klein, 1971). A late date was con-sidered reasonable when modern humanremains were recovered in possible associ-ation with Howiesons Poort artefacts atBorder Cave (Cooke et al., 1945; Malan,1955; Mason, 1962; Sampson, 1974) and atPeers (Skildergat) Cave (Keith, 1931; Jolly,1948; Anthony, 1967), though elementalanalysis now indicates that the Peers Cavematerial represents an intrusive burial(Sillen, personal communication). New ageestimates for Howiesons Poort occurrenceshave generated debate because they involveexperimental dating methods and rely uponthe correctness of detailed depositional,postdepositional, and climatic reconstruc-tions, and inferred correlations with theglobal marine isotopic record. However,out of recent dating efforts has emerged aconsensus that the age of the industry is60–70 ka.
At Boomplaas, a series of 22 radiocarbondates established that the Howiesons Poortis >49 ka (Deacon, 1989). AAR dates onostrich eggshell from the Howiesons Poortat Klasies River range in age from 80 ka atthe bottom of the sequence to 65 ka at thetop; application of the same method atBoomplaas yielded an age estimate for theHowiesons Poort there of 72–80 ka (Brookset al., 1991, 1993a; Miller et al., 1993,1999). Judging from the dates, together withfaunal and geomorphological evidence,Deacon (1989: 554) places the HowiesonsPoort levels at both sites at ca. 70 ka, nearthe end of the last Interglacial (oxygen

502 . . .
isotope stage 5a-4). ESR dates for theHowiesons Poort at Border Cave range from45�5 to 75�5 ka, with a mean of 62 ka(Schwarcz & Grun, 1993a; Grun et al.,1990), but initial analyses of protein dia-genesis in ostrich eggshell from HowiesonsPoort levels at the same site indicated an ageof >70 ka (Miller et al., 1993, 1999), andanalysis of the micromammals suggested atimespan similar to that at Klasies River(Thackeray & Avery, 1990). Miller et al.(1999) report a new series of isoleucineepimerization dates on ostrich eggshell forHowiesons Poort occurrences at BorderCave, Klasies River, Boomplaas, and Apollo11, ranging between 56 and 80 ka. Theyconclude that a reasonable age estimate forthe industry is 66�5 ka.
Parkington (1990) is in the minority inarguing for a shorter chronology and seemsto be in error in favoring an age for theHowiesons Poort of between 50 ka and35 ka. Interestingly, however, he stresses thedifferences between pre-Howiesons Poort(‘‘MSA 1 and 2’’ or ‘‘Pietersburg’’) andpost-Howiesons Poort (‘‘MSA 3 and 4’’ or‘‘Bambatan’’) assemblages, and questionsthe MSA affiliation of the post-HowiesonsPoort material. As Thackeray (1992) pointsout, if Parkington is correct, and industriesoverlying the Howiesons Poort are identifiedas LSA, then the Howiesons Poort mayindeed play a role in the ‘‘transition’’ to theLSA. Further, if current age estimates forthe Howiesons Poort are accurate, thenthe time range of the LSA is consid-erably lengthened, and the ‘‘time lag’’ thatKlein (1992) imputes between the appear-ance of H. sapiens and the LSA is muchreduced.
Wurz (1997, 1999; Deacon & Wurz,1996) has conducted a detailed analysis ofHowiesons Poort material from Cave 2 atKlasies River. She finds that the geometricsare generally manufactured on whole blades,though others (e.g., Thackeray, 1989) havesuggested that the blades were snapped
into segments before backing. Singer &Wymer (1982, cited by Wurz, 1999) pointout 18 examples of the microburin tech-nique, in which a notch is made to facilitatesnapping the blade. Notches on theunbacked edge are found on 19·4% of thebacked artefacts in Wurz’s (1999) sample,and she interprets this as evidence forhafting and binding.
Volman (1984) considers HowiesonsPoort geometrics to be projectile armatures,while Deacon (1989) believes that in view oftheir relatively large size in comparison toLSA examples, they may have functioned asbarbs for spears. For nearly all ESA andMSA artefacts, function is a matter of con-jecture, usually unsupported by usewear orother direct evidence. But because geo-metric microliths remained in use untilrecent times, there is good archaeologicaland ethnographic evidence that they wereused as arrow armatures, both for huntingand in warfare. Arrow shafts with the micro-lithic armatures still in place have beenrecovered in Europe from Mesolithic,Epipaleolithic, and Neolithic contexts(Rust, 1943; Clark, 1951, 1952; Rozoy,1985; Fischer, 1985) and in Egypt fromdynastic tombs (Clark et al., 1974).Nineteenth-century southern African ethno-graphic examples are described by Clarket al. (1976). Clark et al. (1974) remarkupon the striking similarity between theSouth African and Egyptian arrow designs;in both, microliths were generally mountedwith mastic, often in pairs, and in somecases treated with poison. As shown byRozoy (1985) and Fischer (1985), a set ofpaired armatures mounted behind the pointenlarges its cutting edge, thereby increasingthe size of the laceration. Wurz (1997) citesexperimental work (Friss-Hansen, 1990;Nuzhnyi, 1990) which demonstrates theeffectiveness of this technique in woundinganimals and in preventing damage to thearmature. Furthermore, microwear onreplicated geometrics hafted in this way is

503 ’
consistent with observed microwear onHowiesons Poort segments (Harper, 1994;Wurz, 1997).
Coefficients of variation for microlithdimensions reported by Wurz (1997) showthat the Howiesons Poort geometrics are asstandardized as their LSA counterparts andtherefore appear to have been manufacturedto specific design parameters. With meanlengths of about 30–45 mm, the backedgeometrics of the Mumba and HowiesonsPoort industries (Keller, 1973; Volman,1984; Wurz, 1997) are larger than those oftypical LSA microliths, usually �25 mm(Deacon, 1984), but their size is comparableto that of northern European Epipaleolithicarmatures (Rust, 1943; Rozoy, 1985). AsRozoy (1985) points out, however, thecritical size property for an armature is notits linear dimensions, but its weight, usually<10 g for European examples. Publishedweights are not available for African geo-metrics, and it is not known how many,mounted together, would comprise a singlearmature. The heavier Howiesons Poortor Mumba industry geometrics may havebeen used in cutting implements or instabbing or throwing weapons, but theywould also have been suitable for armingan arrow that would perhaps have traveledless than the 20–50 m usual for arrowspropelled by the bow (Rozoy, 1985). Sucharrows may have been unfletched; Clarket al. (1974) observe that among theirEgyptian dynastic sample, lighter arrows arefletched, while heavier ones are withoutfletching. Whatever the details of theirdesign, the Mumba and Howiesons Poortmicroliths support the presence of com-posite projectiles in Africa by 65 ka, about30,000 years earlier than they are found inEurope.
Bone toolsThe shaping of bone and other organicmaterials into points, awls, and other toolshas been a hallmark of the Upper Paleolithic
since the days of Lartet & Christy (1869–1875). Large bones were sometimes flakedin the same way as stone by Oldowan toolmakers (Leakey, 1971b), and Blackwell &d’Errico (2000) claim that Pliocene homi-nids at Sterkfontein and Swartkrans wereusing bone shaft fragments to raid termitemounds. Formal techniques of bone-working, however, are generally agreed tohave been significant to late human evolu-tion, though it is not clear why carving andpolishing a bone point should be beyond thecapabilities of the maker of a Levallois flake.Even within the European Upper Paleo-lithic, however, bone tools are not commoncompared to retouched stone tools. At AbriPataud, France, for example, in all four ofthe early and basal Aurignacian levels com-bined, there are only three fragmentarypossible split-base bone points, whereasthere are 900 retouched stone tools andthousands of bones and bone fragments(Brooks, 1995). A few sites with hundreds ofworked bone objects like Castanet, France,seem to have served as ‘‘factory sites,’’ butmany Aurignacian sites have no boneartefacts at all (White, 1992).
Perhaps bone working is intimately tied tothe development of projectile technology,which would explain why it is present in theAfrican MSA. Until recently, only a fewbone artefacts were known from MSA con-texts, and these were widely discounted aslate aberrations or the result of stratigraphicadmixture. In North Africa, polished bonetools have been reported from Aterian con-text from the site of Grotte d’el Mnasra,Morocco (Hajraoui, 1994). The relativelyinformal assemblage of pointed boneobjects associated with the Khormusanwas formerly thought to date to ca. 18–23 ka(Wendorf et al., 1965), but it has now beenredated to >40 ka, and possibly to 60 ka,bringing the Khormusan firmly into the timerange of similar MSA industries (Marks,1968; Wendorf & Schild, 1992; Marks,personal communication).

504 . . .
In southern Africa, a bone point wasamong the artefacts recovered from the siteof Kabwe, Zambia (Clark, 1959), thoughthe question of its association with the homi-nid, faunal remains and stone artefacts isunresolved. The age of Kabwe was formerlyestimated at 125 ka on the basis of an earlyAAR date of 110 ka (Bada et al., 1974) andfaunal comparison with Olduvai Bed IV(Klein, 1973), but recent recalibration of theOlduvai sequence (Hay, 1990; Manega,1993; Tamrat et al., 1995; Walter et al.,1991, 1992; Kimbel, 1995; White, 2000)suggests an age >780 ka for the material (seeTable 1). Possible ‘‘daggers’’ of wart hogtusk probably dating to ca. 80 ka have beenreported from the Howiesons Poort levels atBorder Cave (Beaumont, 1978, Volman,1984), though their artefactual nature isopen to question.
Singer & Wymer (1982) describe a thincylindrical bone point from HowiesonsPoort layer 19 in Shelter 1A at Klasies River,dated by several techniques to 65–80 ka(Bada & Deems, 1975; Deacon et al., 1986;Deacon & Geleijnse, 1988; Deacon, 1989;Grun et al., 1990). The excavators them-selves expressed skepticism at this uniquefind (Singer & Wymer, 1982: 115), andit has been dismissed as intrusive (e.g.,Volman, 1984). The point, however, wasexcavated from ‘‘compressed, laminatedlevels that showed not the slightest sign ofdisturbance’’, and the color and condition ofthe point are described as similar to otherbones in the Howiesons Poort levels, andquite unlike those from the LSA middens(Singer & Wymer, 1982:115).
Evidence from three MSA sites atKatanda, D. R. Congo, indicates that theKlasies bone artefact is not an isolatedphenomenon. At Katanda, a series ofbarbed and unbarbed bone points has beenrecovered from secure stratigraphic context[Brooks et al., 1995; Yellen et al., 1995;Yellen, 1998; see Figure 7(a), (b)]. Thethree occurrences, Kt-2, Kt-9, and Kt-16,
are all associated with a distinctive paleosolthat crops out along a 1 km long exposure inthe cliffs above the Semliki River. At eachsite, the MSA horizon containing the bonepoints is overlain by 3–5 m of fluvial andcolluvial deposits, up to three distinctpaleosols, a massive capping ash, and aHolocene soil. The Katanda sites are datedto ca. 90 ka by a variety of techniques,including TL and OSL on sands above theMSA occurrences, ESR and U-Series ontooth enamel recovered from the MSAhorizon at Kt-9, and AAR on mollusc shellsand ostrich eggshells from horizons bothabove and below the MSA levels at anumber of sampling localities. Dates forthe MSA occupations containing the bonepoints range from 82+8 ka (T/L) to174+1 ka (U-series); all suggest a minimumage in excess of 75 ka. A recent redatingeffort by OSL on multiple aliquots fromseven additional samples of the sandsoverlying the MSA layer confirms the orig-inal age estimate (Feathers & Migliorini,2000).
Initially, objections were raised to theMSA associations of the Katanda pointsbecause bone points are known from thenearby late Pleistocene site of Ishango (deHeinzelein, 1957, 1962; Brooks & Smith,1987). The stratigraphic conditions andconsistent associations described hereclearly rule out the possibility of Holoceneintrusion. The Katanda bone points areassociated with MSA artefacts at three dif-ferent sites. At no locality is the MSAmaterial overlain by LSA assemblages fromwhich the points could have been derived.Indeed LSA sites of younger (Ishangian) ageare located on a terrace ca. 20 m lower in themodern landscape. There are no anomalouselements in the MSA assemblages, such asmicroliths, that would indicate LSAadmixture. The degree of mineralization ofthe Katanda points resembles that of non-artefactual bone from the MSA occupation,which represents several locally extinct

505 ’
species. Even if the Katanda faunas repre-sent secondary accumulations of consider-able duration, as suggested by Klein (1999:439, 2000; but see Yellen, 1996), theirminimum age is none the less limited by theage of the capping sands. There are alsostylistic differences between the Ishango latePleistocene and the Katanda MSA bonepoints, particularly in the manner in whichthey were hafted. The Ishango points aresmaller, and have a backward facing notchat the base to secure the line from slipping,
whereas the Katanda points have a series ofbasal grooves [Figure 7(b)]. Experimentalwork (Krupsha, 1993) shows that while abackward facing notch will retain the linewhen such a point is used as a harpoon,grooves are insufficient for the purpose.Thus, while the late Pleistocene Ishangobone points were probably designed for useas harpoons, those from the Katanda MSAsites may have functioned as spear points.
The unique character of the Katandabone points may not be surprising, given the
(a)
(b)
(c)
Figure 7. Bone points of the African MSA. (a) Katanda 16 and Katanda 9 (after Yellen et al., 1995),� Alison S. Brooks; (b) detail of point from Katanda 9. � Chip Clark, Smithsonian Institution;(c) Blombos Stillbay level, after Henshilwood & Sealey, 1997, figure 7, photo courtesy of ChrisHenshilwood.

506 . . .
absence in Central Africa of comparableexcavated sites of this age with associatedfauna in similar riparian settings. Yellen(1998) has suggested, however, that thisphenomenon may be due to an MSA sociallandscape of small, relatively isolated popu-lations that is without parallel in historictimes. The idea of bone armatures is notunique to the MSA inhabitants of the CongoBasin. Recent discoveries elsewhere on thecontinent confirm the presence of bonepoints and other bone tools in the MSA.
At Blombos cave in the southern CapeProvince of South Africa, retouched stoneprojectile points and other MSA artefactshave been recovered in sealed context inassociation with well preserved fauna anda series of 21 worked bone objects.These include two cylindrical bone points[Figure 7(c)] that are clearly intentionallyshaped by symmetrical grinding and polish-ing. The other bone objects include two‘‘awls,’’ a possible peg, and a flat piece withparallel incisions made by a stone tool thatmay represent either a decorative image or autilitarian object [see Figure 10(a)]. Analysisof carbon to nitrogen (C:N) ratios showsthat the points and other bone tools areunequivocally derived from the same hor-izon as the MSA fauna. Initial age estimatesderived from multiple techniques indicateda minimum age of 72 ka (Henshilwood &Sealy, 1997, n.d.). The Holocene andMSA levels at Blombos are separated by alayer of sterile sand 10–20 cm thick, forwhich recent OSL analyses indicate anage of ca. 73 ka (Henshilwood, personalcommunication).
Another set of data supporting early boneworking in Africa is a series of barbed pointsfrom White Paintings Shelter in the TsodiloHills of northern Botswana. Here the MSAoccurrence at the base of the sequence, atdepths exceeding 4 m, is overlain by a ‘‘tran-sitional MSA/LSA’’ horizon, at a depth of3·5 m, containing large blades, faunalremains including fish and mammals, and a
single unbarbed bone point. A series of boneharpoons and other bone tools lie in LSAdeposits at about 3 m below the surface. Alarge sample of ostrich eggshell fragmentsfrom deposits higher in the sequence, fromdepths ranging between 3 m and 1·5 m,have been dated by AMS 14C and AAR tobetween ca. 33 and 37 ka. The transitionallevel itself is dated to 38–50 ka by a varietyof techniques (Robbins et al., 1994; Robbins& Murphy, 1998; Kokis et al., 1998;Robbins, 1999; Robbins et al., underreview). It is regrettable that faunal preser-vation is poor in the lower levels at WhitePaintings Shelter, and that bone artefactshave not been found here in MSA context.However, it is significant that a bone point,dating to >30 ka, has been recovered fromthe ‘‘transitional MSA/LSA’’ occurrence.
The bone artefacts from Blombos andKatanda together firmly document boneworking in the Middle Stone Age, and sup-port the MSA associations of previouslyreported finds, such as those from Klasiesriver and Border Cave. Dates from WhitePaintings Shelter confirm an antiquity forbone points in the LSA that considerablycloses the gap between the Katanda andBlombos points on the one hand, and thewidespread African bone working industriesknown from after 25 ka on the other (deHeinzelein, 1957; 1962; Sutton, 1974,1977; Barthelme, 1985; Brooks & Smith,1987; Yellen, 1998). This evidence suggestscontinuous development of a bone workingtradition with origins well back in thePleistocene.
Economy
HuntingIncreased intensification in resource use,greater planning depth, and seasonalscheduling of hunting and fishing are fre-quently cited elements of the ‘‘humanrevolution’’ (e.g., Mellars & Stringer, 1989;Nitecki & Nitecki, 1989). In contrast, MSA

507 ’
people have been portrayed as scavengers(Binford, 1981, 1984), or as less effectivehunters than their LSA successors (Klein,1976, 1978, 1989a; 1992, 1994, 1995,1998, 1999, 2000).
Much of the debate revolves around thespecies composition, body part represen-tation, age profiles, and position of cutmarksin the MSA faunal collection from KlasiesRiver, compared to LSA remains fromnearby Nelson Bay Cave. Marean & Assefa(1999), noting that a significant portion ofthe Klasies River fauna was discarded by theoriginal escavators as unidentifiable (Singer& Wymer, 1982), have suggested settingthe material aside and concentrating onstudies of unbiased collections. Neverthe-less, Klasies River continues to dominate thedebate, despite the fact that its location onthe Cape coast of South Africa, the conti-nent’s most southern extremity, is notrepresentative of Africa as a whole, and thatopen air sites present a different picturefrom that seen for caves. Nor does theKlasies sequence record the early MSA orthe MSA-LSA transition. Rather, at manysites in the southern Cape there appearsto be an occupational hiatus of 40 ka to20 ka duration between the MSA and theLSA, which creates an impression of dra-matic behavioral changes when earlier andlater sites are compared.
Here we refute three assertions abouthunting behavior in the MSA: (1) thathunting was not practiced at all; (2) thatdangerous species were not hunted on aregular basis; (3) that hunters did not focusupon prime age adults. We will also describeinstances of specialization on a single preyspecies in the MSA.
The arguments against the practice ofhunting in the MSA commonly rely uponrelative element abundance and the positionof cutmarks. The underrepresentation ofmeaty limb elements in MSA faunalassemblages has been taken to imply lateaccess to carcasses, that is, scavenging kills
from which the most valued elementshave already been consumed by primarycarnivores. Decisions by human huntersabout which carcass parts to transport tocamp and which to abandon at the kill sitereflect carcass part utility, as well as dailycontingencies not visible in the archaeologi-cal record (Perkins & Daly, 1968; Kleinet al., 1999). Further, limb bones are knownto have been processed for marrow ex-traction from the earliest times (Bunn et al.,1980; Bunn, 1981, 1989; Blumenschine,1987; Blumenschine & Selvaggio, 1991),resulting in fragmentation of the shafts.These elements may be further damaged bysubsequent taphonomic processes that typi-cally destroy limb bone ends with greaterseverity than shafts. Unfortunately, earlyexcavators often regarded shafts as non-identifiable and discarded them, as theydid at Klasies River. In an elegant recentstudy, Marean has convincingly shown bypainstaking refitting of the MSA boneassemblage from Layer 10 in Cave 1 at DieKelders, South Africa, that high utility limbbones are not absent, just better representedby the shafts as opposed to the ends. Whenminimum numbers of elements are based onthe shafts, the skeletal element represen-tation is consistent with early access tocarcasses, and thus with a hunting, not ascavenging, adaptation (Marean, 1998;Marean & Kim, 1998; Bartram & Marean,1999; Marean & Assefa, 1999; Mareanet al., 2000).
Cutmark evidence also supports MSAhunting. Despite the assertions of Binford(1984), there is evidence for neithercarnivore damage prior to hominid acqui-sition nor for the accumulation of carcassesby carnivores at Klasies River (Klein, 1999;Klein et al., 1999). A detailed analysis ofassemblage composition, breakage andtraces of butchery and carnivore ravaging onthe fauna from Cave 1 at Klasies Riverperformed by Milo (1998) leads him toconclude,

508 . . .
‘‘In the KRM faunal assemblage . . . abundantbutchering marks on upper hindquarters andforequarters indicate that the hominidsenjoyed early access to carcasses in all sizeclasses’’
(Milo, 1998: 124). In East Africa, at MSAsites at Lukenya Hill, Kenya, Marean(1997) has observed low frequencies of car-nivore involvement and an abundance ofcutmarks and percussion marks indicating‘‘that people were the primary accumulatorsof these assemblages’’ (Marean, 1997: 210).
Conscious selection of a single preyspecies is seen as a sign of modern humanbehavior (Mellars, 1989a,b; but see Chase,1989; Stiner, 1994; and Mellars, 1996for prey selectivity among Neanderthals).Lieberman & Shea (1994) have argued thatseasonal patterning in faunal use is one ofthe few characteristics that distinguish theoccupation sites attributed to H. sapiensfrom those attributed to Neanderthals in theLevant. There are severals examples ofhunting selectivity in the MSA. In SouthAfrica, Klein (1989a, 1994) has suggestedthat the catastrophic age profile for eland atKlasies River and other MSA sites indicatestactical or intercept hunting of entire herds.In East Africa, at the MSA site of GvJm-46at Lukenya Hill, Kenya (Miller, 1979;Marean, 1992), the focus on a single preyspecies (a small extinct alcelaphine bovid)the catastrophic mortality profile, thepalimpsest nature of the accumulation,implying repeated kill events, and ‘‘the openlocation in a natural topographic trap’’(Marean, 1997: 217) combine to suggestrepeated use as a tactical mass kill site.There is some indication that this mayhave taken place during the annual gamemigration.
The opinion voiced most frequently isthat MSA people were ineffective hunterswhen compared to their LSA successors.The taking of large prey species and primeage adults are regarded as signs of fullycompetent hunting ability (Stiner, 1990,
1994). Klein (1976, 1978, 1989a, 1992,1994, 1995, 1998, 1999) argues, primarilyfrom comparisons of species compositionand age profiles in MSA (Klasies River, DieKelders, Border Cave) vs LSA (Nelson BayCave) assemblages, that MSA hunters didnot often hunt dangerous animals and rarelybrought down prime age adults. However,there is good evidence that MSA hunters didnot confine themselves to docile prey speciesor to juvenile targets. It seems likely that theartefact fragment embedded in a cervicalvertebra from MSA levels at Klasies River[Figure 8(a); Milo, 1998] represents theweapon used to dispatch the animal,although Marean & Assefa (1999) questionwhether it might have become lodged thereduring butchery. The vertebra is that of anadult Pelorovis (size class 5) (Milo, 1998), ananimal for which Klein (1976) estimatesbody weight at >900 kg. Pelorovis’ moderndescendant, Syncerus caffer, though signifi-cantly smaller, is regarded as one of the mostdangerous game animals in Africa. Amongthe modern Jun/wasi, an individual whotakes two large animals (size class 3 orgreater) per year is considered a successfulhunter (Lee, 1979).
Evidence at the MSA site of �Gi, in theKalahari desert of Botswana, dated to 77 ka,also appears to represent deliberate hunt-ing of dangerous game. Historically, theseasonal pan adjacent to the sites hasbeen the locus of ambush hunting practices.Ethnographically, hunting from blinds wasrestricted to a few days per year, during thefull moon at the end of the rainy season,when the pan was the only local source ofwater. More than 600 points were found inthe MSA levels at �Gi, making up 41% ofthe retouched tools (n=1497). The fauna,represented mainly by isolated teeth, isdominated by zebra (Equus burchelli and E.capensis), cape warthog (Phacochoeorus aethi-opicus), and large bovids including at leastone individual Pelorovis (Helgren & Brooks,1983; Brooks & Yellen, 1987; Kuman,

509 ’
1989; Klein, personal communication). Thenumber of projectile points in the MSAassemblage at �Gi indicates deliberatehunting, and the presence of species knownto be large and aggressive shows competenthunting. Warthogs are particularly danger-ous to human hunters, and the herd strate-
gies of zebra render them notoriouslydifficult to pursue. The location of the siteand the presence of game species that arewater dependent supports an interpretationof tactical hunting. This practice was prob-ably even more effective in the past, whenwater availability was more restricted than
(a)
(b)
Figure 8. Faunal evidence for hunting and fishing in the African MSA. (a) Stone point tip embedded inbovid (Pelorovis) cervical vertebra, Klasies River (scale in mm) after Milo, 1998, Fig. 17; photo courtesyof Richard Milo; (b) fish bones (Clarias) from Katanda 9 (scale in cm). � Alison S. Brooks.

510 . . .
at present (Crowell & Hitchcock, 1978;Brooks et al., 1980; Brooks & Yellen, 1987).
Klein, however, argues not on the basis ofpresence or absence, but from relativenumbers. Warthog and buffalo, while notabsent at MSA sites, are outnumbered byeland, and buffalo in some assemblages arerepresented only by juveniles (but see Milo,1998, cited above). The age structures andspecies compositions of the LSA assem-blages more closely approximates the livingcommunities thought to be present at thetime of occupation. Klein infers that MSAhunters were unable to take the moredangerous animals regularly and were forcedto concentrate upon the more tractableeland. He concludes that MSA hunterswere deficient not only technologically, butalso cognitively and socially. These argu-ments can be questioned on both empiricaland theoretical grounds. Gifford-Gonzalez(1991) has questioned Klein’s method forestimating ruminant ages, and Marean &Assefa (1999) point out the confound-ing effects of time averaging on the MSAassemblages, which accumulated over sub-stantially greater spans of time than the LSAassemblages. Minichillo & Marean (2000)argue that the greater species richness of theLSA when compared to the MSA may beexplained by the larger sample sizes for LSAfauna. More fundamentally, one mightwonder why one would query the intelli-gence of a hunter who chooses to pursuethe eland rather than the ornery buffalo!Marean & Assefa (1999), Deacon (1984),and Bar-Yosef (1998) reject Klein’s contrastbetween LSA and MSA hunting behavioras a sign of cognitive differences. A moresatisfactory explanation is provided byDeacon (1989) and Marean (Marean &Assefa, 1999; Minichillo & Marean, 2000),who suggest that the faunal differences seenbetween MSA and LSA represents a pro-gressive expansion of diet breadth. Lesspreferred items were included in the diet as aresponse to increased competition brought
about by population increase. In essence,the situation faced by LSA peoples wasbrought about by the success of precedingMSA hunters and fishers.
Ecology, economy and socialorganization
Aquatic and small-scale resource useWhether deliberate fishing was habituallypractised in the MSA has long beendebated. In the absence of clear fishingparaphernalia, it is difficult to distinguish thesignature of human acquisition from thoseof other piscivores in nonanthropogenicdeposits. Deliberate procurement maysometimes be inferred on the basis ofstratigraphic context or species’ knownhabitat preferences. Thus, the remains ofcichlid fish from White Paintings Shelter,Botswana, are interpreted as the products offishing because they are found in eoliansediments. OSL determinations indicatethat these date to 50–75 ka (Robbins et al.,1994; Robbins & Murphy, 1998; Robbinset al. under review). Fish bones in the upperhorizon at Site 440 in the northern Sudanare explained as the product of deliberatefishing because they are dominated bylarge deep-water species inconsistent with ariparian location. Likewise, at Bir Tarfawi,in the western desert of Egypt, severalfaunal assemblages associated with GreyLake Phases 1 and 2 include Nile perch,a deep, permanent water species (Wendorf& Schild, 1992). In MSA levels at BlombosCave, in the southern Cape province,South Africa, faunal remains include notonly marine molluscs and small fish but alsolarge deep water fish including Cymatocepsnasutus and Aries feliceps (Henshilwood& Sealy, 1997). Poeggenpoel (1999) arguesthat the taphonomy of the Blombos fishbones, especially the apparently deliberatedamage to the spines, is consistent withhuman capture of live fish, and not withscavenging or nonhuman predation. These

511 ’
finds establish the competence of MSApeople at marine coastal fishing, if not thepresence of watercraft.
Stewart (1989), from an analysis of EastAfrican LSA sites, provides a method fordistinguishing human from nonhuman fishexploitation patterns, based on body size,species ecology, and skeletal element rep-resentation. Application of this method toopen-air river margin MSA sites at Katanda,D. R. Congo, where bone points are foundin association with the fish, suggests that, byat least 75 ka, African hominids exploitedlarge catfish (Clarias) weighing more than35 kg [Figure 8(b)]. All remains representadult fish. The absence of other age classesindicates that fishing was carried out exclu-sively at the beginning of the rainy seasonwhen Clarias is known to spawn nearshore.This suggests that exploitation took place aspart of a specialized seasonal strategy, ratherthan on an ad hoc basis. Furthermore, therelative paucity of fish vertebrae implies thatedible portions were separated from headsat the site and removed to be consumedelsewhere (Brooks et al., 1995; Yellen et al.,1995). Clearly MSA people were competenthunters and fishers who planned theirsettlement choices around the seasonalavailability of game and fish.
Studies by Crawford and others (e.g.,Crawford, 1992; Chamberlain, 1996;Broadhurst et al., 1998; Crawford et al.,1999) have shown that Omega-3 (or n-3)polyunsaturated fatty acids (long-chainlipids with a first double bond at thecarbon-3 position, counting from the methylend) are essential to the developing brainof young mammals. They suggest (e.g.,Crawford, 1992) that it is the limited avail-ability of such lipids in terrestrial foodswhich is primarily responsible for limitingthe growth of the brain in terrestrialmegafauna. These lipids, primarily docasa-hexaenoic and arachidonic acids, areabsent in plants and minimally present interrestrial animals except for brain tissue,
which contains up to 600 g of lipids per kg(Broadhurst et al., 1998). Since marinefoods are particularly rich in such lipids,marine mammals face no such limitationand can grow relatively larger and morecomplex brains. Crawford et al. (1999)argue that access to marine foods was essen-tial to the development of the modernhuman brain, and that the evolution of H.sapiens took place at the land-water inter-face. Use of rift valley fish, such as Clarias,which are especially rich in Omega-3 fattyacids, by MSA peoples suggests that thisadaptation, with its potential for improvedinfant nutrition, survivorship and conse-quent population increase was already inplace by at least 80 ka.
Deacon has held that subsistence behaviorin the MSA is indistinguishable from that inthe LSA, in part on the basis of the shellfishdebris and plant remains. He maintains,for example, that the charred lenses ofvegetal material in the Howiesons Poortlevels at Klasies River represent burnt geo-phyte residues. Ethnographically, geophytepatches were sustained by controlled burn-ing, which stimulates corm production.The Klasies traces may therefore indicateenvironmental management and a relianceon carbohydrate-rich plant foods thatparallel Holocene practices (Deacon, 1985,1989, 1993a,b; Deacon & Geleijnse, 1988;Deacon & Deacon, 1999). Intensiveprocessing of plant foods is revealed by thegeneral presence of grindstones at manyMSA sites, including Bir Tarfawi, Egypt;Katanda, D. R. Congo; �Gi, Botswana;and Mumbwa, Zambia (Wendorf et al.,1993b; Brooks et al., 1995; Yellen et al.,1995; Brooks & Yellen, 1987; Barham,1998). This activity seems to have com-menced in late Acheulian contexts atKalambo Falls, Zambia (Clark, 1988) andin the Kapthurin Formation, Kenya.
The use of small scale resources, eitheraquatic or terrestrial, is usually viewed as asign of economic intensification. MSA

512 . . .
people were certainly capable of harvestingsmall-scale resources and showed a taste forthese food items. In North Africa, molluscsare present in the MSA levels at HauaFteah, Libya (McBurney, 1967; Klein &Scott, 1986), at Berard, Algeria (Roubet,1966), and at Temara (Roche & Texier,1976), Mugharet el-Aliya (Howe, 1967),and Zouhra (Debenath & Sbihi-Alaoui,1979) in Morocco. Walter et al. (2000)report stone artefacts and marine bivalves,gastropods, and other invertebrate fossilsfrom a reef platform near Massawa on theRed Sea coast of Eritrea, dated by U-serieson fossil coral to ca. 125 ka (see alsoStringer, 2000). Despite the presence ofhandaxes, the artefacts are described asMSA. Although the behavior represented bythe finds is difficult to evaluate withoutdetails of the artefacts or their archaeologicalcontext, the authors have interpreted thematerial to represent the harvesting ofmarine resources in a shallow subtidalenvironment. In South Africa, shellfishremains are found at Klasies River (Singer &Wymer, 1982; Deacon & Geleijnse, 1988;Thackeray, 1988; Deacon, 1989, 1995),Die Kelders (Avery et al., 1997; Grine et al.,1998; Goldberg, 2000; Klein & Cruz-Uribe,2000), and Herolds Bay (Brink & Deacon,1982), as well as at Hoedjies Punt, SeaHarvest, and Boegoeberg 2 (Volman, 1978;Klein, 1989a, 1999). MSA populationsapparently were also sometimes subject toconditions leading to the intensive exploi-tation of these resources, as shown by theunusually large quantities of giant land snail(Burtoa nilotica) shell in the later MSAdeposits in Bed V at Mumba Rock Shelter,Tanzania (Mehlman, 1989), and the largenumbers of angulate tortoises (Chersinaangulata) at Die Kelders (Klein, 1994; Klein& Cruz-Uribe, 2000). Evidence from coastalItaly (Stiner, 1993, 1994; Stiner et al., 1999,2000) and Gibraltar (Barton et al., 1999)shows that Neanderthals did sometimeseat marine shellfish, but the impressive
escargotiere at Mumba and the numbers ofcoastal African sites containing quantitiesof shellfish seem to indicate a more regu-lar intensive use of small scale resources inthe MSA.
Klein (1989a, 1995, 1999) has contrastedthe MSA levels at Klasies River and othersouthern Cape sites in which marineresources are relatively scarce, to the subse-quent LSA occupations in which they arerelatively abundant. The MSA inhabitantsat Klasies River, Die Kelders, Sea Harvest,Hoedjies Punt, Mossel Bay, and HeroldsBay Cave did utilize shellfish (limpets), and,at Klasies, marine mammals (seals), andbirds (penguins) (Volman, 1978; Brink &Deacon, 1982; Thackeray, 1988; Klein,1994; Avery et al., 1997). The MSA levelsyielded smaller quantities of shells than LSAlevels at some sites. It may be that the use ofmarine resources was not as frequent or asregular in the MSA as in the LSA, but thesmaller volume of shell in the MSA levels atKlasies River and Die Kelders is in part theresult of decalcification of the older cavedeposits. At both sites the lower strata areclearly compressed, the bulk of bone andshell having been leached away (Goldberg,2000; Deacon, personal communication;Avery, personal communication; personalobservation).
Comparison of shell diameters from theMSA deposits at Hoedjies Punt, SeaHarvest, and Boegoeberg 2 with LSAmiddens at Paternoster and Elands BayCave, South Africa, shows a decline in themean size of the limpet Patella granatinafrom MSA to LSA times (Klein, 1989a,1999). This trend reflects the harvesting ofless mature individuals, one of the clearsigns for economic intensification in theLSA. The mean size of mammalian preyspecies also decreased. Comparison of sealmortality profiles from MSA (Klasies Riverand Die Kelders) and LSA (Dunefield, DieKelders, Kasteelberg, Elands Bay Cave,Nelson Bay Cave) reveals that seals in the

513 ’
LSA of the South African Cape coast wereharvested during their breeding season,while in the MSA, seals were apparentlytaken throughout the year (Klein, 1994,1995; Klein & Cruz-Uribe, 1996). Klein hasargued that this pattern indicates a moreselective, seasonally focused strategy for theLSA hunters. Marean & Assefa (1999) havepointed out that LSA assemblages representshorter time intervals than most of thosefrom the MSA. They may thus be lesssubject to the problems of time averagingand thereby show clearer patterns of season-ality. Furthermore, it is not clear why takingseal when natural mortality is high shouldindicate greater intelligence or technicalcompetence.
Klein (1989a, 1994, 1995, 1998, 1999,2000) has repeatedly asserted that thegreater intellectual abilities and technologi-cal sophistication of LSA people allowed forintensification and resulted in an increase inthe human population. LSA sites greatlyoutnumber MSA sites on the African land-scape, and while LSA sites have beenpresented with fewer opportunities forobliteration, the greater number of sites nodoubt reflects a real increase in humannumbers. We would argue that there is nointellectual difference in the abilities of MSAand LSA hominids. Rather, a long-termtrend in population growth led to residualcrowding and a diminished resource base,rendering further intensification necessary inthe LSA (cf. Voigt, 1982; Deacon, 1989;Milo, 1998; Marean & Assefa, 1999). Inaddition, crowding probably led to greaterresidential stability in the LSA, which inturn may contribute to higher archaeologicalvisibility for small food items. Yellen (1991)has shown that at Jun/wasi sites, longerresidential times result in higher represen-tation of small species in faunal assemblages.Small faunal elements are at risk of destruc-tion by carnivore gnawing, but lose theirattractiveness to scavengers after a period ofthree days, whereas larger bones may retain
grease or marrow for several weeks or evenmonths. The presence of people discouragesscavenging by nondomestic carnivores, andafter the departure of the human group,many of the smaller faunal remains are nolonger of interest to scavengers. Residentialstability thus ‘‘protects’’ the small-mammalcomponent from the attentions of scaven-gers and leads to better small element sur-vival. The decreased mobility of larger, moredense human populations may thus exagger-ate our perception of increased use of smallscale resources in the LSA.
Economy and social organization
Lithic resource procurementThe exchange of goods and the developmentof trade networks are important componentsof contemporary society and have been con-sidered key ingredients in modern humanbehavior (White, 1982; Mellars, 1996;Klein, 1989b, 1995). In Upper PaleolithicEurope, raw materials such as elk teeth,mammoth ivory, amber, marine shells, andfossils were obtained from distant sourcesand transformed into decorative objects(White, 1982, 1989b, 1993; Taborin, 1993).
Studies of the sources of lithic materialscan provide information on ranging patternsand resource networks for stone tool-usingpopulations. Two general principles apply innearly all prehistoric contexts. First, localraw materials almost invariably outnumberthose obtained from distant sources.Second, in most cases, exotic stone, being ofhigher quality, was more intensively utilizedand presumably more highly prized(Geneste, 1989a,b; Turq, 1989; Kuhn,1991; 1992; Mellars, 1996; Blades, 1999).
In the Middle Paleolithic of westernEurope, according to Feblot-Augustins(1997), most raw material is locally derived,but 13–17% of sites (n=116) contain smallquantities of stone from as far away as80–120 km (cf. Conard & Adler, 1997). Inthe late Middle Paleolithic of central and

514 . . .
eastern Europe, 71% of sites (n=24) yieldmaterials from sources lying at distances>80 km, in one case as much as 300 km.Mellars (1996: 164) points out that sourcesof flint are far more widely dispersed incentral and eastern Europe than in south-western France, but both Feblot-Augustins(1997: 162–163) and Steele & Gamble(1998; Gamble, 1986) explain the differ-ences between eastern and western Europeraw material procurement on ecologicalgrounds. Their view is that colder, morecontinental climates in the east would havenecessitated greater seasonal displacementand larger home ranges for hominids, as itdoes for other mammals, and these wouldbe reflected in raw material acquisitionpatterns.
Local raw materials continue to domi-nate Upper Paleolithic assemblages. Forexample, at nearly all the GermanGravettian sites studied by Scheer (1993),more than 95% of the lithic artefacts weremade from raw material obtained fromwithin a radius of 25 km. At some FrenchUpper Paleolithic sites the proportion ofexotic flint makes up 25% of the assem-blage, but the distance the material is trans-ported remains much the same as in theMiddle Paleolithic (Mellars, 1996; Geneste,1989b; Feblot-Augstins, 1997; Blades,1999). Only five (1·9%) of 263 westernEuropean sites, all in Belgium andGermany, studied by Feblot-Augustins(1997) have raw materials from distancesexceeding 100 km, and no transport dis-tance from any Chatelperronian site in hisstudy exceeds 100 km (n=26). Exotic stonefrom sources more distant that 100 km doesoccur in the Perigordian/Gravettian and theMagdalenian, although in only one case inthe Magdalenian does the transport distanceexceed 200 km (Feblot-Augustins, 1997:Table 38). In central Europe, transportdistances and proportions of exotic materialsin the early Upper Paleolithic are greaterthan in western Europe, with 27·6% of his
Szeletian sites (n=76) and 42% of hisAurignacian sites (n=150) having distancesexceeding 100 km. Maximum transport dis-tances in the eastern European early UpperPaleolithic are as great as 300–420 km(Feblot-Augustins, 1997: Tables 38 andinventories 57–62).
If the greater transport distances in theeastern European Paleolithic can beexplained on ecological grounds, as sug-gested by Gamble (1986; Steele & Gamble,1998), it would seem that the AfricanMSA should be characterized by smallerterritories and transport distances, especiallyin the tropics (Deshmukh, 1986; Foley,1987), but the reverse is true. At AfricanAcheulian sites, exotic raw materials are rare(Merrick & Brown, 1984; Merrick et al.,1994; Feblot-Augustins, 1997: inventories8–13). Fewer than 1% of the items in theartefact inventories have been transportedfor distances greater than 40 km, and themaximum transport distance is 100 km forthe Ethiopian site of Gadeb. In the EastAfrican MSA, while the proportion of exoticraw materials remains under 1%, knowntransport distances for materials at somesites are much greater. Through analysis oftheir elemental and chemical compositionsby electron microprobe (EMP), and in somecases x-ray fluorescence (XRF), about adozen distinct petrological groups can bedistinguished among the obsidian flows inthe central Kenyan Rift Valley (Merrick& Brown, 1984; Merrick et al., 1994).Artefacts manufactured in obsidian fromthese sources have now been identifiedfrom a number of MSA sites in Kenya andTanzania. A specimen from the MSA siteat Songhor, western Kenya (McBrearty,1981), has been shown to be from a sourceat Njorowa Gorge, 145 km to the east. Twospecimens from the MSA Pundo Makwarindustry from member 6 at the site ofMuguruk, western Kenya (McBrearty,1986, 1988) are derived from sources atEburru and Sonanchi, 185 km and 190 km

515 ’
from the site. Between Muguruk andSonghor and their obsidian sources lies thewestern shoulder of the Gregory Rift, andto transverse this impressive physiographicobstacle involves a climb of at least 1000 m.
Obsidian at two MSA sites in northernTanzania are derived from some of the samecentral Kenya Rift sources as the Mugurukand Songhor artefacts, and was transportedover even greater distances. A sample of fourpieces of obsidian from the MSA Kiseleindustry in levels 14–22 at Nasera RockShelter, near Lake Eyasi in northernTanzania, are from sources at Sonanchi,Eburru, and Masai Gorge, near LakeNaivasha, 240 km north-northeast of Nasera(Merrick & Brown, 1984; Mehlman, 1989).A sample of seven pieces of obsidian fromthe MSA Sanzako and Kisele industries inBed VI at the Mumba Rock Shelter inthe same region of northern Tanzania(Mehlman, 1977, 1979, 1989, 1991) arederived from an outcrop that lies 320 kmfrom the shelter (Merrick & Brown, 1984;Mehlman, 1989). The Sanzako industry isdated by U-series to 100–130 ka (Mehlman,1987).
Nor are these isolated examples. At PorcEupic, Ethiopia, obsidian makes up 5·5% ofthe MSA assemblage. Though geochemicalanalyses have not been carried out, theobsidian is believed to have come fromsources 90 km or 140 km to the west (Clarket al., 1984). Merrick et al. (1994) believethat although movement of �100 km mightfall within the range of mobile forag-ing groups, it is unlikely that transport ofobsidian for distances >300 km representsdeliberate collecting forays. They suggestinstead that increased interaction andexchange among human groups can explainthe distribution of obsidian over the MSAlandscape.
At some East African MSA sites whereobsidian is plentiful, raw material procure-ment and use patterns are different. At theGvJm-16 rockshelter on Lukenya Hill,
Kenya, obsidian comprises 5–8% of theartefacts. In the earlier MSA levels here, themajority of the obsidian (53%), is derivedfrom sources <65 km distant. The remain-ing 31% of the obsidian, or 2% of the totalassemblage, comes from sources between105 and 135 km away. In the later MSAlevels, 62% of the obsidian comes fromthese exotic sources. There appears to be nodifference in the manner in which local andexotic materials are treated at Lukenya Hillfor artefact production, and there is no evi-dence for the importation of finished tools.At the open air MSA site of Prospect Farm,excavated by Anthony (1978) and datedby obsidian hydration to >120 ka (Michelset al., 1983), as many as 99·5% of theartefacts in any given level are obsidian. Thesite is located on Mount Eburru itself, inextremely close proximity to obsidian out-crops, but even here, low frequencies ofexotic obsidian from a source at Kisanana,75 km away, are found throughout thesequence (Merrick et al., 1994). The MSAinhabitants of the nearby site of ProlongedDrift (GrJi-11), Kenya, seem to show adefinite preference for exotic obsidians.Here nearly 90% of the obsidian is importedfrom sources at Sonanchi and NjorowaGorge, 45–55 km from the site, and only afew specimens have been found to representthe much nearer Masai Gorge outcrops nearMount Eburru (Merrick et al., 1994). Thereis no evidence to suggest that approach toany of these obsidian outcrops was pre-vented by physiographical factors in thepast. While the suggestion is speculative, it ispossible that some human groups controlledaccess to valuable obsidian sources and pre-vented collection by others. The presenceof stylistic provinces in the African MSAstrengthens the impression of distinct ethnicgroups, and seems to support trade ratherthan transport as the obsidian distributionmechanism.
On the Cape coast of South Africa, theuse of fine grained raw materials is a

516 . . .
feature of the Howiesons Poort industry thatdistinguishes it from other MSA industries,and these raw materials are selectively usedfor the manufacture of microliths (Singer &Wymer, 1982; Volman, 1984; Thackeray,1992; Deacon & Wurz, 1996; Wurz, 1997,1999). While 96·0–99·8% of the artefacts inthe other MSA levels at Klasies River mainsite are local quartzite, this raw materialcomprises only 73·0% of the HowiesonsPoort assemblage. The remaining 27·0% ismade up of silcrete, quartz, and hornfels(Wurz, 1997). Although determining theprecise source areas for the silcretes isproblematic, outcrops exist in the inlandCape Folded Mountains at distances�15 km (Singer & Wymer, 1982; Wurz,1997). In a recent sample of HowiesonsPoort artefacts from Klasies River cave 2,only 4% of the total sample is nonquartzite,but 39% of the retouched artefacts are madein nonquartzitic raw materials (Wurz,1997).
Ambrose & Lorenz (1990), Deacon(1989), and others portray the HowiesonsPoort as an adaptation to cool, moist con-ditions and a closed vegetation community.This interpretation is supported by thepresence of eboulis sec (angular rock frag-ments with little chemical weathering) inHowiesons Poort deposits at Border Cave,Boomplaas, Montagu Cave, and Nelson BayCave (Ambrose & Lorenz, 1990), by 18Ovalues on marine molluscs from KlasiesRiver indicating cool sea water temperatures(Shackleton, 1982), by the absence of beachsand in the same levels indicating low sealevel (Butzer, 1982), and by the preponder-ance among the Howiesons Poort ungulatefaunas at Klasies River, Boomplaas, andBorder Cave of small, closed habitatforms (Klein, 1976, 1978; Deacon, 1979;Ambrose & Lorenz, 1990). Ambrose &Lorenz (1990; Ambrose, 1998c) view rawmaterial procurement as embedded withinthe resource acquisition round and see tech-nological change in part as a response to
changing lithic resource availability. Thushigher frequencies of exotic lithics in theHowiesons Poort relative to the precedingMSA I and II and the succeeding MSA IIIand IV signal the larger territories requireddue to resource patchiness or unpredict-ability. While most authors (e.g. Deacon,1989; Ambrose & Lorenz, 1990; Deacon &Wurz, 1996; Deacon & Deacon, 1999;Wurz, 1999) imply that the HowiesonsPoort and the MSA I–IV are the variedresponses of a single population seenthrough time, it seems at least equallylikely that these distinct interstratifiedtechnologies were produced by differenthominid groups.
High-quality raw materials were usedpreferentially to manufacture the HowiesonsPoort geometrics (Singer & Wymer, 1982;Wurz, 1997), which may have played amajor role in the maintenance of socialnetworks. Ethnographically, subsistenceunder adverse conditions such as thosedocumented for the Howiesons Poortrequires territories too big for even a largegroup to defend. It encourages informationsharing among groups and the maintenanceof a network of sometimes elaborate andritualized obligations with distant allies asinsurance against times of resource scarcity(Gould, 1978; Wiessner, 1982, 1983, 1986,1998). Deacon (1989; Deacon & Wurz,1996) considers the standardized geometricsof the Howiesons Poort expensive and notesthat they are precisely the kind of objectsused in the ritual hxaro exchange systemof modern San people, as described byWiessner (1982, 1983, 1986), even thoughgeometrics may only require 10–20 secondseach to produce, once the raw material hasbeen obtained (Rozoy, 1985). Both Deacon(1989) and Wurz (1999; Deacon & Wurz,1996) stress the symbolic content ofHowiesons Poort backed geometrics.Deacon sees the specialized HowiesonsPoort artefacts as a means of maintainingpopulations and their social structures

517 ’
during times of resource stress, rather thansimply as a coping mechanism to deal withcolder and drier conditions. Because quartz-ite geometrics do not differ significantly intheir measurable attributes from silcreteexamples, Wurz (1999) concludes thatselection of silcrete was not made for utili-tarian reasons. Rather she argues that theuse of a more costly raw material renderedthe finished object, presumably arrow orspear armatures, more valuable as an item inan exchange network. While future experi-mental replication may well show that theflaking properties of silcrete are superior tothose of quartzite for the manufacture ofgeometrics, this would not diminish thevalue of the objects as items of exchange.
Economy and social organization
Site structure and modificationThe designation of different areas of a habi-tation site for different activities reveals aformalized conceptualization of the livingspace that is considered by many to indicatesophisticated cognitive functions (e.g.,Binford, 1989; Hodder, 1987). The deliber-ate modification of the occupation area isthought to be unique to H. sapiens, andis a feature that consistently distinguishesEuropean Upper Paleolithic from MiddlePaleolithic sites (Farizy, 1990; Mellars,1996). Potential bias in our perception ofpatterning in residential sites has severalsources. Ephemeral sites may have little for-mal structure imposed by their inhabitants,but sites occupied for long periods may haveany existing patterns ‘‘smeared’’ by laterinhabitants. Caves may preserve occupationfeatures better than open sites, but they alsocircumscribe the occupation area andimpose space limits. The result may beoverprinting of different activities or a higherdegree of activity segregation, dependingupon the inhabitants’ response.
Many examples of MSA site modificationremain ambiguous due to our inability to
eliminate natural agents in their formation.Thus the store pavement from Katanda,D. R. Congo, reported by Yellen (1996) anddating to 70–90 ka (Brooks et al., 1995),may represent a deliberately constructedhabitation substrate or may be a lag concen-trated by fluvial processes or termites (cf.McBrearty, 1990a). Its sharp boundariesresemble those from the Acheulian artefactconcentrations at the Olorgesailie Main Site,interpreted by Isaac (1977) as fluvial cut andfill features, but by Potts et al. (1999a,b) asthe result of human activities. Similarly, theconcentration of bipolar flaking debris atSimbi, Kenya, reported by McBrearty(1993b) and dating to between 200 and60 ka, may be an undisturbed activity areaor deliberately dumped refuse (cf. Wilson,1994), but the possibility that it was concen-trated by gently flowing water cannot beruled out. An often cited example of semi-circular MSA windbreaks at Orangia, SouthAfrica (Sampson, 1968, 1974) is nowthought to date to the LSA (Klein, 1989b).
However, there are a number of un-ambiguous examples of deliberate site modi-fication from MSA contexts. Debenath(1994; Debenath et al., 1986) reports theremains of structures from a number ofAterian sites both on the Mediterraneanlittoral and in the Sahara. Traces of post-holes have been found at le Chaperon-Rouge, Morocco, and at Seggedim, Niger.At the latter site, the sharp edges of the flintscatter adjacent to the post holes suggeststhe location of an ephemeral wall or pali-sade. At the Grotte Zouhra (El Harhoura),Morocco, arrangements of stone wedges arebelieved to have served as supports for poles.At the sites of Nazlet Safaha in the NileValley of Egypt, extensive MSA quarryingfor flint has created a series of ditches up to2 m wide and 1·7 m deep covering an area of600 m2 (Vermeersch et al., 1990). A similarpractice is reported for the nearby extensiveTaramsa site complex (Vermeersch et al.,1990; van Peer, 1998).

518 . . .
Deliberate arrangement of large piles ofstones, whose purposes are as yet unknown,are also known from the MSA. FromAterian context at Dar-es-Soltane 2,Debenath (1994) describes an enigmaticheap of sandstone slabs about 1 m indiameter and 30 cm high. A larger pile ofapproximately 60 limestone balls wasrecovered from a fossil spring at the site ofEl-Guettar, Tunisia, together with brokenbone, flaking waste, and a collection ofretouched tools that includes a tangedAterian point (Gruet, 1954, 1955; Clark,1982). Some of the El-Guettar stone ballsare deliberately shaped spheroids, othersare naturally round; they have diametersranging between 4·5 and 18·0 cm. The baseof the pile may have been surrounded by aring of larger stones. A similar mass of stoneballs 1·3 m in diameter and 75 cm high hasbeen recovered from an ancient spring nearWindhoek, Namibia (Fock, 1954; Clark,1982). It is made up of 36 spheroids, eachweighing between 600 and 1200 g. Theunique association of these piles of stoneballs with springs has been emphasized byClark (1982: 265), who suggests that theymay have been cairns, implying a symbolicfunction, or more prosaically, that they mayhave provided a platform from which tocollect water, though why spheroids wouldbe needed for this purpose is unclear. Thepossibility that the spheroids may havetumbled by chance into the eyes of thesprings cannot be ruled out, but their largenumbers suggest that these are not chanceassociations, and deliberate placement bythe site occupants seems most likely.
A striking example of MSA structuresfrom subsaharan African is reported byBarham (1996) at Mumbwa Cave, Zambia.Here three superimposed arc-shaped fea-tures of stone blocks, ash, baked sedi-ment and lithic and bone debris areassociated with a number of hearths andpost holes. Barham (1996) interprets theseas the remains of three circular windbreaks
that would have had diameters of about 3 m.His interpretation is supported by the distri-bution of lithic debitage and bone debris,which dip toward the center of the arcs,suggesting that they were banked up againstthe perimeter, possibly by deliberate sweep-ing. Barham notes that these windbreakswould have served to deflect the prevailingeasterly winds and to create a trap for heatgenerated by the afternoon sun against thecave wall.
Also in the MSA levels at Mumbwa isclear evidence for deliberately constructedstone-lined hearths. Six of these, built oflarge limestone blocks as well as transportedstone, contain quantities of indurated ash,burned bone, and burned limestone andsediment. One partially eroded examplewould have had a diameter of >70 cm whencomplete, was 25 cm deep, was filled with10 cm of ash and surrounded by a zone ofbaked sediment. Barham suggests it mayhave been used as a roasting pit. Anotherexample measures 90 cm�70 cm, with adepth of ash of 5 cm. Five others are super-imposed, and apparently stones from earlierhearths were reused in the construction ofthe later features. This indicates a clearconception of space and its deliberate,repeated use for the same function. Themany superimposed hearths in theHowiesons Poort levels at the main site atKlasies River (Henderson, 1992; Deacon,1995, 1998) suggest similar reuse and aformal conception of domestic space.
Symbolic behavior
Special treatment of the deadBurial and other special treatments of thedead are a consistent feature of the symboliclife of modern human societies, and thebelief that the Neanderthals deliberatelyburied their dead has been a major factorcontributing to an impression of theirhumanity and, for some, their inclusion in

519 ’
our species. Despite the ambiguities intro-duced by taphonomic processes andnineteenth-century recovery techniques(Gargett, 1989, 1999), there is generalagreement that most if not all relativelycomplete Neanderthal skeletons were delib-erately interred (comments to Gargett,1989; Defleur, 1993; Mellars, 1996, andreferences therein). For modern humangroups, burial connotes respect for thedead as well as symbolic and ritual ideas. IfNeanderthals did deliberately bury theirdead, there can be no certainty that thepractice was ritual and not merely hygienicin nature. These issues have often beendiscussed (e.g., Mellars, 1996) and arecogently reviewed by Chase & Dibble(1987) and Hayden (1993). Particularlysignificant is the lack of grave goods inNeanderthal burials, apart from materialsnormally encountered in the archaeologicalsediments that composed the grave fill.
While skeletal completeness and articu-lation of elements have been regarded asevidence for Neanderthal burial (Belfer-Cohen & Hovers, 1992), Gargett (1989,1999) emphasizes the importance of naturaldisarticulation (e.g., Hill, 1979), rockfall(Farrand, 1985), and postmortem tissuedehydration at low temperatures as factorsto be considered in Pleistocene caveenvironments. If his interpretation were cor-rect, we would owe the richness of theNeanderthal fossil record to the hazards ofliving in caves, rather than to Neanderthalcorpse disposal habits. However, there isgood evidence for earlier occupation ofcaves at numerous sites with good faunalpreservation in Middle Pleistocene Europe(e.g., Arago, Lazaret, Peche de l’Aze,Vertesszollos), in the Near East (e.g.,Yabrud, Tabun, Zuttiyeh) and in Africa(e.g., Cave of Hearths, Haua Fteah), andit is difficult to explain why articulatedskeletons are completely lacking from thistime range. The hominids from the Sima delos Huesos, Atapuerca, Spain, numbering at
least 32 individuals, dated by ESR andU-series to 320–200 ka (Bischoff et al.,1997) may have been accumulated by delib-erate disposal down a 13 m shaft in theinterior of the cave. Alternatively, carnivoresor natural sedimentary processes may haveintroduced the hominid material into thesite, or the sima may have acted as a deathtrap for hominids, as well as for bears andother carnivores (Arsuaga et al., 1997b;Andrews & Fernandez-Jalvo, 1997).
The earliest evidence for burial among H.sapiens is found in the Levant at the site ofQafzeh, where at least four out of as manyas 15 individuals represented in the caveare interpreted as deliberate interments(Vandermeersch, 1981; Bar-Yosef et al.,1986). These are dated by TL, ESR, andAAR to ca. 90–120 ka (Schwarcz et al.,1988; Valladas et al., 1988; Brooks et al.,1993b; Bar-Yosef, 1998). At least oneof these, Qafzeh 11, may be associatedwith grave goods (Vandermeersch, 1981;Bar-Yosef & Vandermeersch, 1993).
Population movements among bothNeanderthals and H. sapiens as a response toclimatic change may have been frequent inthe Pleistocene. Mammalian fauna associ-ated with early H. sapiens in the Levantsuggests an extension of the Ethiopianfauna, including H. sapiens, to the regionduring or prior to oxygen isotope stage 5(Tchernov, 1988, 1998), and a laterexpansion of the Neanderthals as part of aEuropean fauna, into the same territory(Hublin, 1990, 1998b; Klein, 1995–1996;Bar-Yosef, 1998). Neanderthals and H.sapiens coexisted in Europe for a period ofabout 10,000 years (Mellars, 1998b,c), andaspects of modern behavior represented inEuropean archaeological industries of theperiod between 40 ka and 30 ka, such as theChatelperronian, Uluzzian, and Szeletian,appear to be the result of Neanderthal accul-turation (e.g., Allsworth-Jones, 1986, 1990;Demars & Hublin, 1989; Farizy, 1990; Ruiz& Hublin, 1994; Mellars, 1998a; but see

520 . . .
d’Errico et al., 1998; Zilhao & d’Errico,1998). Neanderthal burials in Europe dateto after 75 ka (Mellars, 1986, 1996), and itis quite plausible that the practice of burialof the dead was also learned by theNeanderthals from their modern neighborsin the Near East, among whom the practicepredates 90 ka.
In Africa, deliberate burials of MiddlePleistocene hominids are absent. The cut-marks on the temporal bone of the Bodocranium, however, indicate defleshing with astone tool (White, 1986), and suggest eithercannibalism or a postmortem ritualizedtreatment of the skull. Evidence for canni-balism is also present in the modern humansample from Klasies River (White, 1987;Deacon & Deacon, 1999), where the stateof the human remains in terms of fragmen-tation, cutmarks and burning is indistin-guishable from that of the nonhuman bones,which are presumably food remains. It isinteresting to speculate upon whether thefragmented modern human fossils representa prey population, while the stone tools weremade by their predators, whose remainshave not become incorporated into the cavefill.
There is evidence that deliberate inter-ment was practiced by early H. sapiens inAfrica, but it is not uncontroversial. Modernhuman remains have been recovered fromBorder Cave, South Africa. These include apartial cranium, two mandibles, some post-cranial bones (BC1, BC2), and a nearlycomplete infant skeleton (BC3), which isinterpreted as a deliberate burial (Beaumontet al., 1978). Doubt has been cast on theprovenience of the Border Cave hominidfossils, because the cranium and limb boneswere found in the course of guano mining,and all of the human fossils, with the excep-tion of the second mandible, were recoveredin the 1940s, when recovery techniqueswere far from ideal. Parkington (1990)argues that the human fossils from BorderCave are intrusive from upper levels in the
cave as they are unfragmented and in anexcellent state of preservation, unlike thebalance of the MSA fauna. Sillen & Morris(1996) have argued that the diagenetic stateof the BC3 infant skeleton is not consistentwith great antiquity. This is difficult toreconcile with the report of the excavators,who noted that it was found in a shallowgrave, covered with intact sediments.
If the Border Cave hominids were in situin the MSA levels, their age is beyond therange of reliability for radiocarbon. Radio-carbon dates for the later MSA levels at thesite are infinite (>48 ka), and Beaumontet al. (1978) estimate the age of the homi-nids at 90–100 ka on paleoclimatic grounds.Sediments overlying the grave containedostrich eggshell fragments that are nowdated to 105 ka by AAR calibrated by 14Cdeterminations on charcoal from higher inthe cave sequence (Miller et al., 1993,1999). If the associations are valid, and ifthe age estimate is accurate, then the BorderCave infant (BC3) may be the earliestknown human burial. It is significant thatthe infant was associated with a singleperforated Conus shell, which is interpretedas a pendent (Cooke et al., 1945; Wells,1950; Beaumont & Boshier, 1972).
Another deliberate human burial fromMSA context is described by Vermeerschet al. (1998) from the site of Taramsa, a flintextraction and reduction site complex in theNile Valley of Egypt. Here the completeskeleton of a juvenile with an estimated ageat death of 8–10 years has been recoveredin association with Nubian Levallois tech-nology and foliate points. The skeletonhas preliminarily been described as fullymodern. The face is somewhat prognathicand the teeth are large, and there are noappreciable supraorbital tori; it is com-parable in these features to Qafzeh 9. Thechild’s skeleton was found in a seated pos-ition, with the legs flexed and the headfacing upward. There appear to be no gravegoods, apart from the stone artefacts. While

521 ’
the grave fill consists of large cobbles andeolian sand, interpreted as quarry dumpdebris, the burial is overlain by intact MSAdeposits. OSL dates on sand range from80 ka to 50 ka.
An additional case of possible deliberateburial has been reported from the site ofMumbwa, Zambia, where stone beehive-shaped structures have been found to con-tain fragmentary human bones (Dart & DelGrande, 1931; Jones, 1940; Clark, 1989;Barham, 1996). One cairn is found at adepth consistent with a date in the MSA,but as Barham (1996) points out, the struc-ture may be intrusive, and it closelyresembles Holocene examples from higherin the sequence.
Beads and ornamentsOrnaments, as symbolic artefacts, have longbeen recognized as a hallmark of modernhuman behavior, although their distributionin the European Upper Paleolithic isuneven, and many Aurignacian sites lackpierced or decorated items altogether (e.g.,White, 1989a,b, 1993). Ornaments areinfrequent at European Middle Paleolithicsites. Nearly all of these rare items are inquestionable stratigraphic contexts or fromearly uncontrolled excavations (e.g., LaQuina, La Ferrassie) or are associated withfinal Middle Paleolithic contexts postdatingthe appearance of H. sapiens in Europe (e.g.,Bacho Kiro, Cueva Morın, Arcy sur Cure)(d’Errico et al., 1998; Zilhao & d’Errico,1998). Hublin (1998b, 1999; Hublin et al.,1996) has argued that though the tech-niques of manufacture of early UpperPaleolithic and late Middle Paleolithicpierced tooth ornaments differ, the idea ofornamentation itself was acquired by theNeanderthals from their H. sapiens neigh-bors through trade or imitation (cf. Mellars,1998a).
Ornaments have also been thought tobe lacking in the African MSA, but it isnow clear that the African tradition of body
ornamentation predates that of Europe bytens of thousands of years. The perforatedConus shell associated with the infant burialBC3 in MSA context at Border Cave maydate to before 105 ka, though the cautionsof Sillen & Morris (1996) regarding thepossibly intrusive context of BC3 must beborne in mind. A number of body orna-ments are now known from Aterian sitesdating from at least 130 ka to ca. 40 ka.These include a bone pendant from theGrotte Zouhra, Morocco (Debenath, 1994),four deliberately drilled quartzite flakes,probably designed for use as pendants, fromSeggedim in eastern Niger (Tillet, 1978;Debenath, 1994), and a perforated shell ofArcularia gibbonsula from Oued Djebanna,Algeria (Morel, 1974). It is also of interestthat the only perforated objects found inthe Levant prior to the Upper Paleolithicare associated with H. sapiens and faunaincluding African taxa (Bar-Yosef &Vandermeersch, 1993).
The manufacture and use of ostrich egg-shell beads is widespread in the AfricanLSA. The earliest LSA ostrich eggshellbeads in southern Africa are documentedby Robbins (1999), where direct AMS 14Cassays on a drilled bead preform from WhitePaintings Rock Shelter yielded an age of26 ka (Kokis et al., 1998; Robbins, 1999).Deeper in the section at White Paintings, aneggshell partial disc that resembles the open-ing of a water container has been dated to31 ka, and indicates a greater antiquity forthe practice of eggshell working (Robbins,1999). At Enkapune ya Muto, Kenya, beadsare known from an early LSA content[Figure 9(a)], where radiocarbon dates onostrich eggshell fragments from bead manu-facture indicate an antiquity of ca. 37–40 ka(Ambrose, 1998a). An early set of radio-carbon dates on charcoal indicate a similarage for ostrich eggshell beads from LSAcontext at Kisese II rock shelter, Tanzania(Inskeep, 1962). Ambrose (1998a) makes acompelling case that dates in this age range

522 . . .
are particularly likely to be underestimates.The layer containing the beads at Enkapuneya Muto is underlain by units estimatedby obsidian hydration to date to >46 ka(Ambrose, 1998a).
Beads have been reported from threeMSA sites in South Africa, Cave ofHearths (Mason, 1962; Mason et al., 1988),Boomplaas (Deacon, 1995), and BushmanRock Shelter (Plug, 1982), from late MSA(Tshangulan) levels at Nswatugi, Zimbabwe(Cooke, 1971), and from one MSA site inEast Africa, Mumba Rock Shelter, Tanzania(Mehlman, 1989). Radiocarbon dates oncharcoal indicate an age of 42 ka for thebeads at Boomplaas (Deacon, 1995). TheMumba beads [Figure 9(b)] were found inLayer V, associated with an industry thatcontains elements of both LSA and MSA
technology. They were originally thought tobe intrusive from the early LSA horizon inlower Bed III, dated by 14C and AAR to30–37 ka (Mehlman, 1979, 1989, 1991;Kokis, 1988; Brooks et al., 1993b; Hareet al., 1993). Direct AAR assays on two ofthe Bed V beads show that one may indeedbe intrusive from Level III, but the otherdates to ca. 52 ka (Hare et al., 1993). Thesenew age determinations establish a consider-able antiquity for the practice of personalornamentation in Africa, and link it withMSA technology.
Incised and notched objects are knownfrom a much wider range of sites in southernAfrica. These include ocher plaques and twonotched bones from Klasies River, SouthAfrica (Singer & Wymer, 1982), twonotched bones from Apollo 11, Namibia
(a)
(b)
Figure 9. Early ostrich eggshell beads. (a) Enkapune ya Muto, after Appenzeller, 1998; photo courtesy ofStanley Ambrose; (b) Mumba. � Alison S. Brooks.

523 ’
(Wendt, 1972, 1976; Vogelsang, 1998), anincised bone, a notched piece of red ochreand a piece of ochre with multiple incisedcross-hatchings from Blombos, South Africa[Henshilwood & Sealey, 1997; Figure 10(a);Gore, 2000: 100], and incised and notched
ochre from Hollow Rock Shelter, SouthAfrica [Figures 10(b,c), 12(b,c)]. Analysisof C:N ratios shows the Blombos incisedbone to have a ratio identical to that ofthe rest of the MSA faunal assemblage(Henshilwood & Sealey, 1997, n.d.). The
(a)
(b)
(c)
Figure 10. Notched and incised objects from MSA contexts (a) incised bone from Stillbay level atBlombos, after Henshilwood & Sealey, 1997, figure 8; photo courtesy of Chris Henshilwood; (b) incisedochre from Hollow Rock Shelter (scale in cm); (c) detail of 8(b). Photos (b) and (c) courtesy of Ian Watts.

524 . . .
incised bone and notched and incised redochre at Blombos all underlie sterile dunesands dated by OSL to 73 ka (Henshilwood,pers. comm.). A more elaborately modifiedbone, with a set of repeated incised notcheshas been described from transitional MSA/LSA context at Border Cave, Swaziland(Beaumont, 1978). Incised ostrich eggshellfragments, that may be pieces of water con-tainers like those of the modern San, havebeen found at Diepkloof, South Africa,and Apollo 11, Namibia (Wendt, 1972;Vogelsang, 1998). At all these sites, thenotched objects predate 40–50 ka. At Apollo11, incised fragments continue to the base ofthe MSA, dated by AAR of ostrich eggshellto >83 ka (Kokis, 1988; Miller et al., 1999).
Use of pigmentPigment is a substance that imparts colorto other materials, and the systematic useof pigment is one of the hallmarks of theEuropean Upper Paleolithic (White, 1982).While the sophisticated cave art at some ofthe best known European painted cavesdates to the Magdalenian period, ca. 18 ka,recent direct dating by AMS 14C shows thatthe European cave art tradition dates backto ca. 30 ka (Clottes et al., 1992, 1995;Valladas et al., 1992; Chauvet et al., 1996),and mobiliary art is likewise known from theAurignacian (Hahn, 1993). Organic com-pounds can be used as pigment, but thosedetected in the archaeological record aremore frequently metallic oxides. Whilecarbon in the form of both graphite andcharcoal, manganese, and kaolin were usedin prehistory (Courard, 1988; Clottes et al.,1990; Clotes, 1993), the pigments mostcommonly encountered in archaeologicalcontexts are iron oxides. These may take theform of red hematite (Fe2O3) or yellowlimonite [FeO(OH) nH2O]. Limonite canbe transformed into hematite by oxidation;the process is most efficiently achieved byburning, and requires a temperature of atleast 250�C (Couraud, 1988).
Although some would identify the use ofpigment in prehistory as de facto evidence forsymbolic behavior (e.g., Marshack, 1981,1989), others are skeptical of the symboliccontent of this behavior (e.g., Chase &Dibble, 1987, 1992) and urge considerationof taphonomic factors (Bednarik, 1992).Some authors have distinguished betweensymbolic and domestic use, such as for hidepreparation (e.g., Mellars, 1996). Othershave suggested possible medical uses forochre, as a styptic or antiseptic treatment forwounds or for eradicating internal or exter-nal parasites (Cole, 1954; Velo, 1984,1986). If metallic oxides are recovered inassociation with undoubted art objects, or atsites whose inhabitants are known to havefunctioned within a well articulated sym-bolic system, the materials are usuallyassumed to have been used as a coloringmedium. For example, characoal has beenfound to have been an ingredient inEuropean Upper Paleolithic paint recipes(Clottes, 1993), but if found in archaeo-logical deposits in the absence of other evi-dence it would probably be confoundedwith domestic refuse. Pigment must beadequately preserved to show traces ofgrinding, and may be expected to surviveless well at open air sites than in caves. If thematerial was completely pulverized in pre-history, it may show up as little more than astain. Evidence for burning has been arguedto be a byproduct of natural processes forthe ochre at Terra Amata (Wreschner,1985); in other cases, burning may beinterpreted as evidence for a deliberateprocessing strategy, as has been maintainedfor the Chatelperronian occurrences at Arcy-sur-Cure (Demars, 1992; Couraud, 1991).
Pigment, in the form of both iron andmanganese oxides, has been encountered atabout a dozen European Middle Paleolithicsites (Bordes, 1952; Marshack, 1981;Demars, 1992; Mellars, 1996), and a lime-stone ochre receptacle is reported from lateMousterian level at Grotte de Neron,

525 ’
France (Combier, 1989; de Beaune, 1993).Fragments of pigment that show signs ofscraping and wear facets, and grindstonesperhaps used for ochre preparation arepresent in Mousterian levels at Cueva deCastillo and Cueva Morın, Spain (Kraybill,1977). Although Mellars (1996) argues fordomestic use, it can equally reasonably beinferred that the pigment was used as acoloring agent, and lends support to the ideaof some form of symbolic or imaginativelife among the later Neanderthals. It is sig-nificant that the pigment at Arcy-sur-Curepostdates the appearance of H. sapiens inBurgundian France (Mellars, 1998a,b).Because the systematic use of pigmentoccurs earlier in the Near East, Hublin(1990, 1998b) has suggested that, likeburial, the practice may have been acquiredby the Neanderthals through long-distancecultural diffusion.
The use of red ochre is widespread inmodern subsaharan Africa. Either in drypowdered form or mixed with fat or water itis applied as a cosmetic to the hair and skinand as a decoration to beads, hides, andclothing. It is commonly applied as paint tohouse walls and pottery. It is clear thathematite and limonite were almost uni-versally used as coloring agents in latePleistocene and Holocene Africa. Countlessfrgments of pigment with unambiguous evi-dence of grinding, as well as grindstoneswith traces of ochre, have been recoveredfrom LSA sites. Undoubted art objects,including artefacts of stone, bone, shell andother organic materials with traces ofapplied ochre are also often found in LSAcontexts, and painted stones are frequentlyencountered in LSA graves (Rudner, 1971).Painted and engraved rock outcrops andcaves dating to the LSA have wide distri-bution in subsaharan Africa, where natural-istic images depict aspects of a foraging wayof life. In North Africa the tradition persistsafter the introduction of domestic animals;in South Africa a few post-contact paintings
depict Europeans, guns, and horses. Thereis good ethnographic evidence for continuityin the symbolic content of southern Africanrock art with modern San belief systems(Vinnicombe, 1975, 1976; Lewis-Williams,1982, 1983). The paintings are believed tohave retained their power into moderntimes, and some are now at risk, due to theremoval of pigment from the images bynon-San local healers for use in curingrituals (Hall, 1999).
Fock (1972) postulates an early date forpetroglyphs in southwestern Gauteng, basedon his identification of an engraving of abovid as the extinct Megalotragus or Homoi-oceras. While additional supporting evidencewould be welcome, Fock is correct to pointout that the African rock art tradition mayhave an early origin. Subsaharan Africa con-tains few deep limestone caves which canprovide the alkaline conditions conducive topreservation. Prehistorically occupied rock-shelters are often no more than overhangswith walls exposed to the elements. AsPhillipson (1977: 288) points out, theearliest mural rock paintings in Africa haveprobably long since been lost through ex-foliation and the natural collapse of shallowcave systems, and only rare discoveries ofburied pieces of painted rock provide anindication of the true antiquity of thetradition (cf. Beaumont, 1992).
Granite slabs with ochre traces have beenfound at Nswatugi, Zimbabwe, in a lateMSA (Tshangulan) horizon with an infiniteradiocarbon age (Cooke, 1971; Walker,1995; Larsson, 1996). Wendt (1975, 1976)and Vogelsang (1996, 1998) report paintedslabs from the Apollo 11 cave, Namibia, inan MSA horizon stratified above a levelcontaining artefacts with Howiesons Poortaffinities (Figure 11). The painted slabs maybe exfoliated from the cave ceiling, or, asargued by Wendt, may be portable artobjects. Ochre crayons occur in MSA levelsthroughout the sequence. Radiocarbondates ranging between 26 ka and 28 ka from

526 . . .
the level containing the painted slabs havebeen reported (Wendt, 1976; Freundlichet al., 1980; Vogelsang, 1998). These youngdates are anomalous for material with MSAassociations, and Miller et al. (1999) reportan age of 59 ka for the MSA overlying theHowiesons Poort at Apollo 11, based on aseries of 62 isoleucine epimerization dateson ostrich eggshell from the sequence.
Economy and symbolism
Pigment acquisition and processingMany early reports of MSA sites in bothSouth and East Africa mention ochre andgrindstones, but these were often discounteddue to the uncontrolled nature of early exca-vations and the possibility of mixing withoverlying LSA debris. Grindstones weresometimes discarded, and those that survive,if inadequately curated, cannot now beexamined for traces of pigment. Many wereno doubt used for plant food processing. Butevidence is accumulating for the wide-spread, controlled and systematic acqui-sition, processing and use of pigment in theMSA. While there is little archaeologicalor biological evidence to support theiridiosyncratic sociobiological interpretation,
Knight et al. (1995; cf. Knight, 1991; Power& Watts, 1996; Power & Aiello, 1997) arecorrect to highlight the importance of pig-ment in understanding human behavior inthe African MSA.
In tropical Africa, lateritic soils provide aplentiful and accessible supply of ironoxides. In southern Africa, on the otherhand, where lateritic soils are less developed,red ochre is less readily available. Delib-erate mining for hematite, specularite (amicaceous form of hematite), and manga-nese, in both open cast and deeply excavatedcaverns, has been documented for thehistoric, Iron Age, LSA, and, in a fewcases, MSA periods in southern Africa(Dart, 1967; Boshier & Beaumont, 1972;Beaumont, 1973b; Beaumont & Boshier,1974; Robbins et al., 1998). Large scalemining for hematite in the MSA is reportedfrom Lion Cavern in the Ngwenya Range,Swaziland, where Beaumont (1973a;Boshier & Beaumont, 1972; Beaumont &Boshier, 1974) reports that at least 1200metric tons of pigment were extractedprehistorically from a cliff-face exposure.Rubble fill had accumulated to a depth of4 m at the base of the cliff; upon excavation,the basal 76 cm of this fill yielded tens ofthousands of MSA artefacts. Radiocarbondates on charcoal from this layer range fromca. 10 ka to ca. 43 ka. The older dates, bestregarded as infinite, derive from materials ata greater distance from the more recentlyworked face (Beaumont, 1973a: 141),showing how the face had moved inwardwith the passage of time. Similar large scalemining operations for specular hematite,dating to the Holocene, is reported from acomplex of cave sites in the Tsodilo Hills byRobbins et al. (1998). Mining at one of thesecaves, Rhino Cave, during the MSA is indi-cated by the presence of specular hematitecrystals, hammerstones, and grindstones inMSA levels in the cave.
Singer & Wymer (1982) report a total of180 pieces of red ochre from Klasies River
Figure 11. Painted slab from MSA levels at Apollo 11Rock Shelter, after Vogelsang, 1998. Photo courtesy ofRalf Vogelsang and the Heinrich-Barth-Institut.

527 ’
on the Cape coast of South Africa. Many ofthese show wear facets, and the authorssuggest that the small pieces were used as‘‘pencils.’’ Some from the MSA have cleartraces of incising to remove pigment powder
[Figure 12(a)]; others are merely scratchedand may have functioned as ‘‘palettes’’[cf. Figure 10(b,c)]. The material is distrib-uted throughout the section; 14 specimensderive from the lowest (MSA I) levels in the
(a)
(b)
(c)
Figure 12. Red ochre from MSA contexts; (a) ground ochre, Klasies River (scale in cm), perforated bymarine origanism, after Knight et al. 1995, Figure 4; (b) notched ochre, Hollow Rock Shelter (scale in cm)after Knight et al., 1997, Figure 3; (c) detail of 10(b). Photos (a) and (b) courtesy of Ian Watts.

528 . . .
cave system, dated to >100 ka. Over 50% ofthe ochre found at Klasies (n=102) comesfrom the Howiesons Poort levels that areinterstratified within the MSA, and dated to65–80 ka. These levels yielded a cylindricalpiece of red ochre ca. 3 cm long, with threedepressions 40–70 mm deep. At Blombos,South Africa, many large pieces ofhematite occur both the Still Bay and lowerundifferentiated MSA. Several of theseappear to have been cached in a small natu-ral niche in the cave wall (Henshilwood,personal communication). Twenty-onepieces of ochre from Blombos have singleor multiple holes up to 25 mm deepand 10 mm in diameter [Figure 10(a)](Henshilwood & Sealy, n.d.). While thesewere initially thought to have been drilledwith a bone or wooden drill (Henshilwood &Sealy, n.d.; Milo, personal communication),they are now believed to have been bored bya marine organism (Henshilwood, personalcommunication).
Hematite pencils are reported throughoutthe entire MSA sequence at Border Cave,South Africa; the age of the base of thesequence has been estimated at >100 ka(Beaumont et al., 1978; Miller et al., 1999).Fibrous material, stained with iron oxideand interpreted as bedding material, has alsobeen recovered from an MSA context atBorder Cave (Volman, 1984), and thehuman infant burial (BC3) from the samesite is stained through the application ofhematite (de Villiers, 1973). Grindstonesstained with traces of ochre have beenrecovered from MSA levels at Pomongwe,Zimbabwe (Walker, 1987), Die Kelders,South Africa (Avery et al., 1997) and at�Gi, Botswana. A total of 298 fragments ofochre, at least 40 of them with clear wearfacets resulting from grinding, have beenfound in MSA levels at Porc Eupic, Ethiopia,where age estimates based on obsidianhydration indicate a minimum age of 77 ka(Clark et al., 1984; Clark, 1988). Largegrinding slabs are also reported from
MSA layers at Florisbad, South Africa (deBeaune, 1993), dated by ESR to 121�6 ka(Grun et al., 1996). Ambrose (1998a: 384)reports traces of ochre and an ochre-stainedgrindstone from basal levels at Enkapune yaMuto, Kenya, associated with artefacts ofthe MSA Endingi industry.
At some East African sites, the presence ofvolcanics allows the use of the K/Ar and40Ar/39Ar dating methods. At the site ofGnJh-15 in the Kapthurin Formation,Baringo, Kenya, the use of pigment in acontext of great antiquity has recently cometo light. Here stone artefacts, fragmentarybone, and ostrich eggshell fragments lie on apaleosol sealed by many meters of volcanictephra. Pyroclastic material overlying thesite has been dated by K/Ar to >240 ka(Tallon, 1978; McBrearty et al., 1996), anda current program of 40Ar/39Ar dating indi-cates an age closer to 280 ka (Deino &McBrearty, under review). ‘‘Red stainedearth’’ was observed in excavation atGnJh-15 by a team of Belgian researchers(van Noten et al., 1987a,b; Cornelissenet al., 1990 and references therein). Newexcavations and reexamination of previouslyexcavated material reveal that while thehematite fragments are too friable to pre-serve traces of grinding, they are numerousand widespread at the site (McBrearty,1999). The previously excavated samplesconsist of 74 items with a total weight of>5 kg; these range in size from pulverizedgranular material weighing <3 g to largechunks >250 g. Excavations in 1997 re-covered more ochre and ochre stainedgrindstones likely used to process it. TheKapthurin discovery is not an isolated find.Pigment has recently been reported from anearly MSA context at the cave site of TwinRivers, Zambia. Here, three pieces of ochre,one limonite, two hematite, have beenrecovered from artefact and bone-bearingbreccia overlain by a speleothem dated byU-series to 230 ka (Barham & Smart, 1996;Barham, 1998).

529 ’
Summary and discussion
H. sapiens appears early in Africa, but it hasbeen argued by proponents of the ‘‘humanrevolution’’ that the behavior of these earlypeople was indistinguishable from that ofmore archaic hominids. In this paper weprovide a new way of looking at the origin ofour species that is more consistent with theevidence.
The Middle Pleistocene African hominidfossil record provides reasonable candi-dates for the ancestors of H. sapiens, andsupport for in situ evolution of H. sapiensin Africa long before 100 ka. Improved tem-poral resolution and further scrutiny of thefossils themselves are needed to determinewhether the record represents a series ofchronospecies or an adaptive radiation inlate Homo. The degree of species diversitynow known for our genus in the EarlierPleistocene leads us to favor a similaradaptive radiation in the Middle Pleisto-cene. The presence of coeval hominidspecies would help to explain the degreeof variability seen in the African MiddlePleistocene archaeological record. In ourview, the appearance of modern humanmorphological features in H. helmei by260 ka indicates that elements of a newadaptation are already in place.
Both H. helmei and early members of H.sapiens are associated with MSA technology,and thus it is clear that the main behavioralshift leading to modernity lies at theAcheulian–MSA boundary about 250–300 ka, not at the MSA–LSA boundary at50–40 ka as many assume. We have shownhere that many sophisticated behaviors arepresent in the MSA. This implies increasedcognitive abilities with the appearance ofH. helmei, and behavioral similarities anda close phylogenetic relationship betweenH. helmei and H. sapiens. It could be arguedthat the specimens referred here to H. helmeiare more correctly attributed to H. sapiens,and that H. helmei should be sunk into
H. sapiens. If that is the case, our species hasa time depth of ca. 250–300 ka, and itsorigin coincides with the appearance ofMSA technology.
However, we are not seeking simply tomove the ‘‘human revolution’’ back to theAcheulian–MSA boundary. Use of the term‘‘revolution’’ implies not only profound,but also rapid change (cf. Greene, 1999).Popular treatments of these issues (e.g.,Diamond, 1992) borrow the vocabulary ofpunctuated equilibria from paleobiology(Eldredge & Gould, 1972), but even inpaleontology, punctuated events are notori-ously difficult to document (Gingerich,1984; Gould & Eldredge, 1996), and thereis no agreement about how rapid a changemust be to qualify.
There is no logical reason to expect asingle sudden event to represent what we seeas essentially a cultural, not a neurologicalprocess (cf. Bar-Yosef, 1998). Figure 13summarizes the first appearances in Africa ofthe modern behaviors as discussed in thispaper. The record shows that the newbehaviors do not appear suddenly together,but rather are found at points separated bysometimes great geographical and temporaldistances. It seems inappropriate to labelchanges accumulating over a period of200,000 years either a revolution or a punc-tated event. Early modern human popula-tions in late Middle Pleistocene Africa wererelatively small and dispersed, change wasepisodic, and contact among groups inter-mittent. This resulted in a stepwise progress,a gradual assembling of the modern humanadaptation. Abrupt ‘‘leaps’’ are perceivedat the northern and southern tips of thecontinent, where the record is discontinu-ous, and where the impression of contrasthas been enhanced by the practice ofcomparing materials from the earliest LatePleistocene with those from the latestPleistocene.
From a terminological point of view, it isimportant not to conflate the African MSA

530 . . .
with the European Middle Paleolithic.While MSA and Middle Paleolithic tech-nologies share common features (Thackeray& Kelley, 1988; Thackeray, 1989, 1992,2000), the evidence reviewed in this paperserves to highlight their essential differences.It is particularly critical that the Middle toUpper Paleolithic transition in Europe notbe confused with the origin of H. sapiens.The European Middle to Upper Paleolithictransition represents the local replacementof the Neanderthals by H. sapiens, while inAfrica, H. sapiens is a product of in situevolution. The African MSA–LSA tran-sition is a cultural and technological changethat occurred many tens of thousands ofyears later. Furthermore, the Middle toUpper Paleolithic transition in Europe is achange from mode 3 to mode 4 technology,while in Africa, modes 3, 4, and 5 are allalready present in the African MSA. In fact,the diversity in stone tool industries suggests
that the terms ESA, MSA, and LSA are toovague, broad, and temporally imprecise tobe particularly illuminating when attempt-ing detailed behavioral reconstructions(Kleindienst, 1967).
The presence of stone working techniquessuch as backing in the African MSA at 80 kashows a level of technical competence notachieved until tens of thousands of yearslater ouside the continent. Microwearstudies are in their infancy in Africa, butfrom artefact design and ethnographic evi-dence it seems clear that MSA artefacts werehabitually hafted, and some of them wereused as projectiles, perhaps in some casespropelled by the bow. From the long-termstylistic conservatism of barbed bone tools inCentral Africa, and the slowness with whichthis particular technology (but not bonetools in general) spread beyond the greatlakes region, Yellen (1998) concludes thatMSA exchange networks may have
Figure 13. Modern behaviors and their time depths in Africa. � Sally McBrearty & Alison S. Brooks.

531 ’
remained regionally restricted. We wouldsuggest that future research at lacustrinesites in this time range may well fill in whatnow appear to be lacunae in the bonetool record. Other evidence suggests thatresidential territories were large in the MSA,especially in tropical Africa. Among modernforaging groups, long-distance social ties aremaintained through ritualized exchange inpart to reduce risk in times of famine(Wiessner, 1982; Hayden, 1992). Thepresence of exotic obsidians at some EastAfrican sites indicates that the distancesinvolved in some tropical Africa MSAexchange networks exceeded 300 km. Thisactivity may have involved both a higherdegree of planning and scheduling, andmore complex interactions among earlyhuman groups than has customarily beenenvisaged. The regional distributions of pro-jectile point styles suggest social networks ona similar order of magnitude.
Despite suggestions to the contrary, thereis good faunal evidence to show that MSApeople were competent hunters whobrought down a variety of dangerous gameanimals, some of them larger than theirpresent day counterparts. Intensified andscheduled resource use are evident at sitessuch as Mumba and Katanda, and the use ofboth marine and freshwater fish and shellfishcan also be seen at Katanda, Klasies River,Blombos, White Paintings Shelter, BirTarfawi, and the Haua Fteah. Sophisticatedbone tools were used in specialized, season-ally scheduled fishing at Katanda. Bonepoints were also used at Blombos and poss-ibly Klasies River. The general presence ofgrindstones in the MSA indicates that plantfood processing was routine, and burnedgeophyte residues in the Howiesons Poortlevels at Klasies River suggests continuousplant exploitation and perhaps resourcemanagement similar to that practiced bythe San inhabitants of the region in theHolocene (Deacon, 1989; Deacon &Deacon, 1999).
At other sites, grindstones appear to havebeen used to process pigment, and red ochredating to well before 200 ka is found bothGnJh-15 in the Kapthurin Formation, and atTwin Rivers. Other sites show notching andincising not only of ochre, but also of boneand ostrich eggshell, most likely as part of asymbolic or notational system. Determiningwhether these objects represent elements ofa fully articulated symbolic system awaitsfurther research, but a lack of formal artobjects need not preclude the presence ofmodern cognitive ability, as has been ob-served by Deacon (1989: 561). It would notbe surprising if the widely dispersed humangroups of the African MSA did not signaltheir identities as intensively as later, morecrowded populations. Mellars (1996: 371ff)concludes that the rarity of purportedly‘‘symbolic’’ artefacts in Mousterian contextsargues against their being part of a wide-spread shared Neanderthal symbolic system.Despite the relatively small number of exca-vated MSA sites, the quantity and quality ofevidence for symbolic behavior that has beenreviewed here far exceeds that known for theEuropean Middle Paleolithic where the sitesample is more than ten times greater. Pig-ment processing and bead production, inparticular, are comparable in kind to thatseen in the Africa LSA and European UpperPaleolithic, and it is not extravagant to sug-gest that they functioned within a sharedsymbolic system. MSA point styles, more-over, do show regional differences that arereasonably construed as stylistic.
As a whole the African archaeologicalrecord shows that the transition to fullymodern human behavior was not the resultof a biological or cultural revolution, but thefitful expansion of a shared body of knowl-edge, and the application of novel solutionson an ‘‘as needed’’ basis (cf. Ambrose& Lorenz, 1990; Vishnyatsky, 1994;Belfer-Cohen, 1998; Bar-Yosef, 1998). Thecomplex content of human cultures hasbeen built incrementally, with cognitive

532 . . .
equipment present since at least 250 ka, in aprocess that continues today.
The MSA–LSA transition is clearlydecoupled temporally from the origin ofH. sapiens, and the transition was incremen-tal in the tropics, as shown from thegradual changes in artefact frequencies anda diminution in artefact size that presagesmicrolithic technology at sites such asEnkapune ya Muto, Mumba, and Aduma.Some LSA elements appear quite early andsome MSA elements persist quite late, sug-gesting that the transition was not onlygradual but also episodic in nature. Forexample, at Matupi, small microlithic corespredate 40 ka, whereas at Kalemba andRose Cottage Cave, MSA radial and disccore types persist as late as 25 ka. However,it is particularly misleading to read thelate survival of MSA flake production tech-niques as a failure of South African H.sapiens populations to achieve behavioralmodernity. As Wobst (1983) and others(e.g., Wynn, 1985; Goren-Inbar, 1988)have pointed out, the study of Stone Agetechnology reveals only the lower limits ofhominid cognitive capacity. After all, disccore technology was widespread during theNeolithic in the Near East, and AustralianHolocene assemblages contain few formaltools, yet there is no suggestion that either ofthese populations were not behaviorallymodern! In fact, if the OSL, TL, and ESRdates of ca. 60 ka for the Australian Mungo3 hominids are accurate (Thorne et al.,1999; Stringer, 1999; Bowler & Magee,2000; Gillespie & Roberts, 2000; Grunet al., 2000), humans possessed the meansto cross large bodies of water at least 10,000years before the usual dates invoked for the‘‘human revolution’’.
Population growth and the MSA–LSAtransition
We believe that the intensification visible atthe MSA–LSA transition after about 50 ka
is in part the result of simple populationgrowth coupled with environmental deterior-ation, factors that go far to explain later,more familiar archaeological phenomenasuch as the origin of domestication inthe Near East or the elaboration of thelate Upper Paleolithic in Europe. Whatmight have accounted for dramatic popu-lation growth in Africa early in the LatePleistocene? Throughout this paper, wehave suggested factors that would have actedto increase infant survivorship and decreaseoverall mortality rates due to either periodicstarvation or accidental injury. Two of themost important of these are new tech-nologies and risk-management strategiesinvolving long-distance exchange. New pro-jectile technologies reduced the need tograpple at close range with large gameanimals, and would have increased huntingproductivity. Fishing technology allowedhumans to exploit a vast new resource.Not only would these technologies havedecreased the territory required to sustaina single individual, but they apparentlyallowed expansion into entirely newhabitats, such as the tropical forest. AsStiner et al. (1999, 2000) have argued,the availability of small-scale resourcesmay improve infant survivorship, drivingfurther population expansion and contribut-ing to a positive feedback system. Inaddition, fish and shellfish may provideoptimum maternal and child nutrition asa major source of omega-3 fatty acids(Broadhurst et al., 1998; Crawford et al.,1999).
Long-distance exchange, maintained bythe use of symbolic systems, would alsohave acted to increase population duringthe Middle Stone Age. Risk-managementstrategies involving long-distance exchangeare at the heart of historically observedhunter-gatherer adaptive systems (Gould,1978; Wiessner, 1982, 1983, 1986, 1998).Exchanges may involve marriage partners,raw materials, red ochre, objects of

533 ’
adornment and technologically importantitems such as projectile points, knives oraxes, often in exotic and colorful materialscarrying symbolic meaning. Such exchangescreate far-flung reciprocity networks thatcan operate in times of scarcity to allowexchange partners and their families accessto distant areas where resources are plentiful(Yellen, 1986), thereby optimizing the map-ping of populations to resources, reducinginfant mortality and contributing to overallpopulation maintenance. As in the case ofsmall-scale resources, positive feedbackwould have produced further populationgrowth and contributed to the expansion ofsuch systems over time.
Environmental deterioration is the thirdfactor contributing to increased con-centration of existing populations. Thetechnology that made LSA extractiveintensification possible was present in theMSA was early as 70 ka, both in SouthAfrica in the form of the Howiesons Poortindustry, and in East Africa in the Mumbaindustry, but the need for intensificationbecame general in the Late Pleistocene assuccessful MSA populations grew andplaced ever more severe demands uponlocal resources. Crowding, decreasedmobility and declining resources necessi-tated hunting more dangerous game,including less preferred food items in thediet, and developing more elaborate tech-nology for food gathering, processing andstorage. Population growth, a quantitativephenomenon, has qualitative effects uponhuman lives, and competition overresources obviously affects the nature ofrelations among groups. Possible responsesrange from outright conflict (warfare),through claims of ownership by symbolicsignals, to cooperation and trade. Perhapsmore significantly, the life experiences andperceptions of a person who encounters200 individuals in his or her lifetime aresubstantially different from those of some-one who encounters 2000. Increased
population density, therefore, affects notonly economy and technology, but alsosymbolic and linguistic aspects of sociallife and the pace of cultural change.
Conclusions: the revolution thatwasn’t
The search for revolutions in westernthought has been in part, as Greene (1999)points out, a search for the soul, for theinventive spark that distinguishes humansfrom the rest of the animal kingdom.Landau (1991) makes a convincing case thatmany syntheses of human evolution are castin the mold of hero myth, with the cleverhero passing through trials to achieve areward. It was the desire to believe thatbrain expansion preceded bipedalism thatfostered the acceptance of Piltdown andthe rejection of Australopithecus for manydecades. We believe that by continuing toinsist upon revolutions, researchers, perhapsunwittingly, create a gulf separating humansfrom the rest of the biological world. Bystressing human uniqueness, proponents ofthe ‘‘human revolution’’ effectively removethe origin of H. sapiens from the realm ofnormal scientific inquiry (cf. Foley, 1987).As Foley (1987) points out, all speciesare by definition unique, and presumablyeach species has its own unique form ofconsciousness. Humans no doubt shareelements of their consciousness, as they dotheir behavior, with their close relatives.Chimpanzees have been shown to use andmake tools and to engage in organized inter-group aggression. Our close extinct relatives,the Neanderthals, were intelligent hominidswho demonstrated the skills needed to sur-vive in a challenging periglacial environ-ment. It will certainly not be surprising ifthey are shown to have possessed languageor rudimentary symbolizing abilities, eitherinherited from a common ancestor, devel-oped in parallel, or learned from H. sapiensthrough culture contact.

534 . . .
There is a profound Eurocentric bias inOld World archaeology that is partly a resultof research history and partly a product ofthe richness of the European material itself.The privileging of the European record is soentrenched in the field of archaeology that itis not even perceived by its practitioners.Works on the ‘‘human revolution’’ can tracetheir ancestry to the urban and agriculturalrevolutions of V. Gordon Childe (1934),whose initial definitions were formulated asanalogies with the industrial revolution ofeighteenth century Europe (Greene, 1999).Childe himself owed an intellectual debt toLewis Henry Morgan (1871), whose tran-sition from Savagery to Barbarism was seenas an inevitable step on the road to humanprogress. In this paper, at the risk of beinglabeled as eurocentric ourselves, we haveused standards for behavioral modernityderived from the European record, becausethese criteria are universally recognized andare frequently repeated in the literature. Wesee this as a necessary initial exercise in theshedding of an inappropriate paradigm, andbelieve that the more interesting and fruitfulera of research into the questions raised herehas only begun. The behaviors unique to H.sapiens are to be discovered, not prescribed.It is not really reasonable to expect all earlypopulations of H. sapiens to have made bonetools, eaten fish, or used paint.
The ‘‘human revolution’’ is a semantic, aswell as a romantic issue, and our conundrumstems from the use of the loaded term ‘‘mod-ern’’ to describe the behavior of people priorto the sixteenth century, let alone those of thePleistocene. Our lists of modern behaviorsare derived from populations living today orthose alive in the relatively recent past, whoall are or were products of their evolution andhistory. Past human populations were prod-ucts of different histories, and the qualitiesstressed in our lists of modern behaviors mayhave had little relevance for our ancestors.
If it is acknowledged that both modernhuman behavior and modern human
anatomy originated in Africa, the EuropeanUpper Paleolithic is freed of the encumbranceof false rhetoric about human origins andhuman revolutions. Archaeologists workingon the later Pleistocene of Europe can seek intheir data the traces of the flow of people,ideas, and technology, and the signs of inter-actions among groups, as has been suggestedfor the Chatelperronian and Szeletian records(Harrold, 1983, 1989; Allsworth-Jones, 1986,1990; Koslowski, 1988; Mellars, 1996,1998a; contra d’Errico et al., 1998; Zilhao &d’Errico, 1998). We should expect the emerg-ing picture to be complex and to generatecontroversy, as has been the case with thedebate over the earliest peopling of the NewWorld. Attention has already been directed atsites in the Levant for what they may revealabout these processes, but sites in otherregions that are possible avenues of migrationout of Africa, such as the Nile Valley, the Sinaiand Arabian peninsulas, southeastern Europe,the Maghreb, and Gibraltar assume greaterimportance as well.
This view also challenges Africanistarchaeologists to evaluate the African evi-dence on its own terms. Rather than apply-ing arbitrary criteria for modernity, onecould take the position that behavioralinnovation drove the morphological changesthat are observed in early modern humananatomy. When we realize that the modernhuman adaptation appeared gradually,rather than suddenly, the process can bebroken into its constituent parts, eachhaving its own origin and demanding itsown explanation. It is necessary to establishthe nature of the changes, the order of thechanges, and the pace of change in order todiscover the causes. This presents us with afresh opportunity to expand our under-standing of the range of behaviors practicedby members of our own species.
Acknowledgements
McBrearty’s current project in theKapthurian Formation is supported by

535 ’
National Science Foundation grantnumber SBR-9601419; her previous workat Muguruk, Simbi, and Songhor was sup-ported by NSF, the L. S. B. LeakeyFoundation, the Wenner-Gren Foundation,and the National Geographic Society. Herresearch is carried out in affiliation with theNational Museums of Kenya and under aresearch permit issued by the Office of thePresident of the Republic of Kenya. Shewould like to acknowledge the support of aChancellor’s Fellowship from the Universityof Connecticut that allowed her to com-plete work on this paper. Brooks’ researchhas been funded by National ScienceFoundation, the Smithsonian Institution,the George Washington University Facili-tating Fund, and the L. S. B. Leakey Foun-dation. She is grateful for the support ofthe Bunting Institute (Radcliffe Institute forAdvanced Study), the American Council ofLearned Societies, and Harvard Univer-sity’s Department of Anthropology duringthe early stages of the construction of thispaper.
We would like to thank Chris Stringer,Richard Klein, Colin Groves, TerryHarrison, and an anonymous reviewer fortheir helpful remarks on an earlier draft ofthis paper. We would also like to express ourappreciation to the many people who helpedus with references, criticism, and comments,including Stanley Ambrose, Graham Avery,Larry Barham, Ofer Bar-Yosef, SteveBrandt, Harvey Bricker, Amelia Clarke, ElsCornelissen, Hilary Deacon, Warren DeBoer, Jim Feathers, Alan Franklin, DianeGifford-Gonzalez, Ed Hare, Alicia Hawkins,Chris Henshilwood, Jean-Jacques Hublin,John Kingston, Maxine Kleindienst, JulieKokis, Daniel Lieberman, Curtis Marean,Anthony Marks, Mike Mehlman, JulioMercader, Peter Mitchell, Osborn Pearson,Cedric Poeggenpohl, Philip Rightmire,Larry Robbins, Judy Sealey, Pierre-JeanTexier, Phillip van Peer, Ralf Vogelsang,Tom Volman, Lyn Wadley, Fred Wendorf,
and Polly Wiessner. Photographs arecourtesy of Stanley Ambrose, Chip Clark,Chris Henshilwood, Julie Kokis, RichardMilo, Ralf Vogelsang, the Heinrich-Barth-Institut, and Ian Watts. Figure 4 was pro-vided by Pierre-Jean Texier. Assistance inputting together Figures 1, 2, 3 and 13, andin tracking down and editing the large list ofreferences was provided by Tamarra Martz,Ben Theis, and especially Christian Tryon,who went far, far beyond the call of duty tohelp this paper see the light of day. Finally,special thanks go to Andrew Hill and JohnYellen for their helpful input as well as fortheir sustenance and moral support withoutwhich this paper could never have beencompleted.
References
Abbate, E., Albianelli, A., Azzaroli, A., Benvenuti, M.,Tesfamariam, B., Bruni, P., Cipriani, N., Clarke,R. J., Ficcarelli, G., Macchiarelli, R., Napoleone, G.,Papini, M., Rook, L., Sagri, M., Tecle, T. M., Torre,D. & Villa, I. (1998). A one-million-year-old Homocranium from the Danakil (Afar) Depression ofEritrea. Nature 393, 458–460.
Aiello, L. C. (1993). The fossil evidence for modernhuman origins in Africa: a revised view. Am. Anthrop.95, 73–96.
Allsworth-Jones, P. (1986). The Szeletian and theTransition From Middle to Upper Paleolithic in CentralEurope. Oxford: Oxford University Press.
Allsworth-Jones, P. (1990). The Szeletian and thestratigraphic succession in Central Europe andadjacent areas: main trends, recent results, andproblems for resolution. In (P. Mellars, Ed.) TheEmergence of Modern Humans: An ArchaeologicalPerspective, pp. 160–242. Ithaca: Cornell UniversityPress.
Allsworth-Jones, P. (1993). The archaeology ofarchaic and early modern Homo sapiens: an Africanperspective. Cambridge Archaeol. J. 3, 21–39.
Amani, F. & Geraads, D. (1993). Le gisementmousterien du Djebel Irhoud, Maroc: precisions surla faune et la biochronologie, et description d’unnouveau reste humain. C. r. Acad. Sci., Paris 316,847–852.
Ambrose, S. H. (1997). Implication of geomagneticfield variation for the chronology of the Middle/LaterStone Age and Middle/Upper Paleolithic transitions.J. hum. Evol. 32, A2.
Ambrose, S. H. (1998a). Chronology of the LaterStone Age and food production in East Africa.J. Archaeol. Sci. 25, 377–392.

536 . . .
Ambrose, S. H. (1998b). Late Pleistocene humanpopulation bottlenecks, volcanic winter, and thedifferentiation of modern humans. J. hum. Evol. 34,623–652.
Ambrose, S. H. (1998c). Social and territorial organ-ization of Late Quaternary hunter-gatherers in EastAfrica: testing hypotheses with lithic raw materialsource exploitation patterns. In (B. Bousman, Ed.)Past and Present Technological Strategies of AfricanHunters, Gatherers and Herders, Ann Arbor: Inter-national Monographs in Prehistory.
Ambrose, S. H. & Lorenz, K. G. (1990). Social andecological models for the Middle Stone Age in south-ern Africa. In (P. Mellars, Ed.) The Emergence ofModern Humans: An Archaeological Perspective,pp. 3–33. Ithaca: Cornell University Press.
Andrefsky, W. Jr (1998). Lithics: MacroscopicApproaches to Analysis. Cambridge: CambridgeUniversity Press.
Andrews, P. (1984). On the characters that defineHomo erectus. Cour. Forsch.-Inst. Senckenberg 69,167–175.
Andrews, P. & Fernandez-Jalvo, Y. (1997). Surfacemodifications of the Sima de los Huesos fossilhumans. J. hum. Evol. 33, 191–218.
Anthony, B. W. (1967). Excavation at Peer’s Cave,Fish Hoek, South Africa. Palaeoecol. Afr. 2, 58–59.
Anthony, B. W. (1978). The Prospect Industry: adefinition. Ph.D. Dissertation, Harvard University.
Appenzeller, T. (1998). Art: evolution or revolution?Science 282, 1451–1454.
Arambourg, C. (1955). Recent discovery in humanpaleontology: Atlanthropus of Ternifine, Algeria. Am.J. phys. Anthrop. 13, 191–201.
Arambourg, C. & Biberson, P. (1956). The fossilhuman remains from the Paleolithic site of SidiAbderrahman (Morocco). Am. J. phys. Anthrop. 14,467–490.
Arambourg, C. & Hoffstetter, R. (1963). Le Gisementde Ternifine. Arch. Inst. Paleontol. Humaine 32,9–36.
Arensburg, B., Schepartz, L. A., Tillier, A. M.,Vermeersch, P. M. & Rak, Y. (1990). A reappraisalof the anatomical basis of speech in Middle Paleo-lithic hominids. Am. J. phys. Anthrop. 83, 137–146.
Armstrong, A. L. (1931). Rhodesian archaeologicalexpedition (1929): excavations in Bambata Cave andresearches on prehistoric sites in Southern Rhodesia.J. R. Anthrop. Inst. Great Britain & Ireland 61,239–276.
Arsuaga, J.-L., Martınez, I., Gracia, A., Carretero,M. N. & Carbonell, E. (1993). Three new humanskulls from the Sima de los Huesos, MiddlePleistocene site in Sierra de Atapuerca, Spain. Nature302, 534–537.
Arsuaga, J.-L., Martınez, I., Gracia, A. & Lorenzo, C.(1997a). The Sima de los Huesos crania (Sierra deAtapuerca, Spain). J. hum. Evol. 33, 219–281.
Arsuaga, J.-L., Martınez, I., Gracia, A., Carretero,J. M., Lorenzo, C., García, N. & Ortega, A. I.(1997b). Sima de los Huesos (Sierra de Atapuerca,Spain); The site. J. hum. Evol. 33, 109–128.
Asfaw, B. (1983). A new hominid parietal from Bodo,Middle Awash Valley, Ethiopia. Am. J. phys.Anthrop. 61, 367–371.
Avery, G., Cruz-Uribe, K., Goldberg, P., Grine, F. E.,Lenardi, M. J., Marean, C. W., Rink, W. J.,Schwarcz, H. P., Thackeray, A. I. & Wilson, M. L.(1997). The 1992–1993 excavations at the DieKelders Middle and Later Stone Age cave site,South Africa. J. Field Archaeol. 24, 263–291.
Ayala, F. J. (1995). The myth of Eve: Molecularbiology and human origins. Science 270, 1930–1936.
Backwell, L. & d’Errico, F. (2000). A new functionalinterpretation of the Swartkrans early hominid bonetools. J. hum. Evol. 38, A4–A5.
Bada, J. L. (1987). Palaeoanthropological applicationsof amino acid racemization for dating fossil bonesand teeth. Anthrop. Anz. 45, 1–8.
Bada, J. L. & Deems, L. (1975). Accuracy of datesbeyond the 14C dating limit using the aspartic acidracemization reaction. Nature 255, 218–219.
Bada, J. L., Kvenvolder, F. A. & Peterson, E. (1973).Racemization of amino acids in bones. Nature 245,308–310.
Bada, J. L., Schroeder, R., Protsch, R. & Berger, R.(1974). Concordance of collagen-based radiocarbonand aspartic acid racemization ages. Proc. natl. Acad.Sci. USA 71, 914–917.
Bailey, R. C., Head, G., Jenike, M., Owens, B.,Rechtmen, R. & Zechanter, E. (1989). Hunting andgathering in tropical rainforests: is it possible? Am.Anthrop. 91, 59–82.
Baksi, A. K., Hsu, B., Williams, M. O. & Ferrar, E.(1992). 40Ar/39Ar dating of the Brunhes–Matuyamageomagnetic field reversal. Science 256, 356–357.
Balout, L., Biberson, P. & Tixier, J. (1967).L’Acheuleen de Ternifine (Algerie): gisement del’Atlanthrope. L’Anthropologie 71, 217–238.
Bar-Yosef, O. (1987). Pleistocene connections betweenAfrica and southwest Asia: an archaeologicalperspective. Afr. Archaeol. Rev. 5, 29–38.
Bar-Yosef, O. (1994). The contributions of southwestAsia to the study of the origin of modern humans. In(M. H. Nitecki & D. V. Nitecki, Eds) Origin ofAnatomically Modern Humans, pp. 23–66. New York:Plenum.
Bar-Yosef, O. (1995a). The Lower and MiddlePaleolithic in the Mediterranean Levant: chronologyand cultural entities. In (H. Ullrich, Ed.) Man andEnvironment in the Paleolithic, pp. 246–263. Liege:Universite de Liege.
Bar-Yosef, O. (1995b). The role of climate in theinterpretation of human movements and culturaltransformations in western Asia. In (E. S. Vrba,G. H. Denton, T. C. Partridge & L. H. Burckle, Eds)Paleoclimate and Evolution, with Emphasis on HumanOrigins, pp. 507–523. New Haven: Yale UniversityPress.
Bar-Yosef, O. (1998). On the nature of transitions:The Middle to Upper Palaeolithic and the Neolithicrevolution. Cambridge Archaeol. J. 8, 141–163.
Bar-Yosef, O. & Meadow, R. H. (1994). The origins ofagriculture in the Near East. In (T. D. Price & A. B.

537 ’
Gebauer, Eds) Last Hunters, First Farmers, pp. 39–94.Santa Fe: School of American Research Press.
Bar-Yosef, O. & Vandermeersch, B. (1993). Modernhumans in the Levant. Scient. Am. 4, 94–100.
Bar-Yosef, O., Vandermeersch, B., Arensburg, B.,Goldberg, P., Laville, H., Meignen, L., Rak, Y.,Tchernov, E. & Tillier, A.-M. (1986). New data onthe origin of modern man in the Levant. Curr.Anthrop. 27, 63–64.
Bar-Yosef, O., Arnold, M., Mercier, N., Belfer-Cohen,A., Goldberg, P., Housley, R., Meignen, L., Vogel,J. C. & Vandermeersch, B. (1996). The dating ofUpper Paleolithic layers in Kebara Cave, Mt Carmel.J. Archaeol. Sci. 23, 297–306.
Barham, L. (1995). The Mumbwa Caves project,Zambia, 1993–1994. Nyame Akuma 43, 66–71.
Barham, L. S. (1996). Recent research on the MiddleStone Age at Mumbwa Caves, central Zambia.In (G. Pwiti & R. Soper, Eds) Aspects of AfricanArchaeology, pp. 191–200. Harare: University ofZimbabwe Publications.
Barham, L. S. (1998). Possible early pigment usein south-central Africa. Curr. Anthrop. 39, 703–710.
Barham, L. S. & Smart, P. (1996). Early date for theMiddle Stone Age of central Zambia. J. hum. Evol.30, 287–290.
Barthelme, J. (1985). Fisher-Hunters and NeolithicPastoralists in East Turkana, Kenya. Oxford: BARInternational Series. Monographs in AfricanArchaeology.
Barton, R. N. E., Currant, A. P., Fernandez-Jalvo, Y.,Finlayson, J. C., Goldberg, P., Macphail, R.,Pettitt, P. B. & Stringer, C. B. (1999). GibralterNeanderthals and results of recent excavations inGorham’s, Vanguard and Ibex Caves. Antiquity 73,13–23.
Bartram, L. E. (1997). A comparison of Kua(Botswana) and Hadza (Tanzania) bow and arrowhunting. In (H. Knecht, Ed.) Projectile Technology,pp. 321–343. New York: Plenum.
Bartram, L. E. & Marean, C. W. (1999). Explainingthe ‘Klasies Pattern’: Kua ethnoarchaeology, the DieKelders Middle Stone Age archaeofauna, long bonefragmentation and carnivore ravaging. J. Archaeol.Sci. 26, 9–20.
Bate, D. M. A. (1951). The mammalian fossils fromSinga and Abu Hugar. Fossil Mammals of Africa 2,1–28.
Beaumont, P. B. (1973a). The ancient pigment minesof southern Africa. S. Afr. J. Sci. 69, 140–146.
Beaumont, P. B. (1973b). Border Cave: a progressreport. S. Afr. J. Sci. 69, 41–46.
Beaumont, P. B. (1978). Border Cave. M.Sc. Thesis,University of Capetown.
Beaumont, P. B. (1980). On the age of the Border Cavehominids 1–5. Paleontol. Afr. 23, 21–33.
Beaumont, P. B. (1992). The time depth of aestheticand symbolic behavior in southern Africa. Paperpresented at the Southern African Association ofArchaeologists Biennial Conference, University ofCape Town.
Beaumont, P. B. & Boshier, A. K. (1972). Somecomments on the recent findings at Border Cave,northern Natal. S. Afr. J. Sci. 68, 22–24.
Beaumont, P. B. & Boshier, A. K. (1974). Report ontest excavations in a prehistoric pigment mine nearPostmasburg, northern Cape. S. Afr. Archaeol. Bull.29, 41–59.
Beaumont, P. B. & Vogel, J. C. (1972a). On a newradiocarbon chronology for Africa south of theEquator, Part I. Afr. Studies 31, 66–89.
Beaumont, P. B. & Vogel, J. C. (1972b). On a newradiocarbon chronology for Africa south of theEquator, Part II. Afr. Studies 31, 155–182.
Beaumont, P. B., de Villiers, K. & Vogel, J. C. (1978).Modern man in sub-Saharan Africa prior to 49,000years B. P.: a review and reevaluation with par-ticular reference to Border Cave. S. Afr. J. Sci. 74,409–419.
Bednarik, R. G. (1992). Paleoart and archaeologicalmyths. Cambridge Archaeol. J. 2, 27–57.
Belfer-Cohen, A. (1998). The appearance of symbolicbehavior in the Upper Pleistocene of the Levant ascompared with Western Europe. In (M. Otte, Ed.)L’Homme De Neanderthal, Vol. 5: La Pensee. Actes DuColloque International De Liege, pp. 25–29. Liege:Universite de Liege.
Belfer-Cohen, A. & Hovers, E. (1992). In the eye of thebeholder: Mousterian and Natufian burials in theLevant. Curr. Anthrop. 33, 463–471.
Berger, A., Imbrie, J., Hays, J., Kukla, G. & Saltzmann,B. (Eds) (1984). Milankovitch and Climate:Understanding the Response to Astronomical Forcing.Dordrecht: Reidel.
Berger, L. R. & Parkington, J. E. (1995). A newPleistocene hominid-bearing locality at Hoedjiespunt,South Africa. Am. J. phys. Anthrop. 98, 601–609.
Berger, T. D. & Trinkaus, E. (1995). Patterns oftrauma among the Neanderthals. J. Archaeol. Sci. 22,841–852.
Bermudez de Castro, J. M. (1988). Dental diseasesand Harris lines in fossil human remains fromAtapuerca-Ibeas (Spain). J. Paleopath. 1, 131–146.
Bermudez de Castro, J. M., Arsuaga, J. L., Carbonell,E., Rosas, A., Martınez, I. & Mosquera, M. (1997).A hominid from the Lower Pleistocene of Atapuerca,Spain: possible ancestor to Neandertals and modernhumans. Science 276, 1392–1395.
Biberson, P. (1961). Le Cadre Paleogeographique de laPrehistoire du Maroc Atlantique. Rabat: Publicationsde la Service des Antiquites du Maroc 16.
Biberson, P. (1963). Human evolution in Morocco inthe framework of the paleoclimatic variations ofthe Atlantic Pleistocene. In (F. C. Howell & F.Bourliere, Eds) African Ecology and Human Evolution,pp. 417–447. Chicago: Aldine.
Binford, L. R. (1981). Bones: Ancient Men and ModernMyths. New York: Academic Press.
Binford, L. R. (1984). The Faunal Remains From KlasiesRiver Mouth. New York: Academic Press.
Binford, L. R. (1985). Human ancestors: changingviews of their behavior. J. Anthrop. Archaeol. 4,292–327.

538 . . .
Binford, L. R. (1989). Isolating the transition tocultural adaptations: an organizational approach. In(E. Trinkaus, Ed.) Emergence of Modern Humans:Biocultural Adaptations in the Late Pleistocene,pp. 18–41. Cambridge: Cambridge UniversityPress.
Bischoff, J. L., Soler, N., Maroto, J. & Julia, R. (1989).Abrupt Mousterian/Aurignacian boundary at c. 40 kabp: Accelerator 14C dates from l’Arbreda Cava(Catalunya, Spain). J. Archaeol. Sci. 16, 563–576.
Bischoff, J. L., Ludwig, K., Garcia, J. F., Carbonell, E.,Vaquero, M., Stafford, T. W. & Jull, A. J. T. (1994).Dating of the basal Aurignacian sandwich at AbriRomanie (Catalunya, Spain) by radiocarbon anduranium series. J. Archaeol. Sci. 21, 541–551.
Bischoff, J. L., Fitzpatrick, J. A., Leon, L., Arsuaga,J. L., Falgueres, C., Bahain, J. J. & Bullen, T. (1997).Geology and preliminary dating of the hominid-bearing sedimentary fill of the Sima de los HuesosChamber, Cueva Mayor of the Sierra de Atapuerca,Burgos, Spain. J. hum. Evol. 33, 129–154.
Bishop, W. W. & Clark, J. D. (Eds) (1967). Backgroundto Evolution in Africa. Chicago: University of ChicagoPress.
Blades, B. S. (1999). Aurignacian lithic economy andearly modern human mobility: new perspectives fromclassic sites in the Vezere valley of France. J. hum.Evol. 37, 91–120.
Blumenschine, R. J. (1987). Characteristics of anearly hominid scavenging niche. Curr. Anthrop. 28,383–408.
Blumenschine, R. J. & Selvaggio (1991). On the marksof marrow bone processing by hammerstones andhyenas: Their anatomical patterning and archaeo-logical implications. In (J. D. Clark, Ed.) CulturalBeginnings: Approaches to Understanding Early Homi-nid Life-Ways in the African Savanna, pp. 17–32.Bonn: Romisch-Germanisches Zentralmuseum,Forschunginstitut fur Vor- und FruhgeschichteMonographien 19.
Bluszcz, A. (1993). Thermoluminescence dating ofdeposits from the area of Bir Sahara East and BirTarfawi. In (F. Wendorf, R. Schild & A. E. Close,Eds) Egypt During the Last Interglacial, pp. 224–227.New York: Plenum.
Boaz, N. T., Ninkovich, D. & Rossignol-Strick, M.(1982). Paleoclimatic setting for Homo sapiensneanderthalensis. Naturwissenschaften 69, 29–33.
Bocquet-Appel, J.-P. & Demars, P. Y. (2000).Neanderthal contraction and modern human col-onization of Europe. Antiquity 74, 544–552.
Bordes, F. (1952). Sur l’usage probable de la peinturecorporelle dans certaines tribus mousteriennes. Bull.Soc. prehist. fr. 49, 169–171.
Bordes, F. (1961). Typologie du Paleolithique Ancien etMoyen, 2 Vols. Memoires de L’Institut Prehistoriquede l’Universite de Bordeaux 1. Bordeaux: Delmas.
Bordes, F. & Crabtree, D. (1969). The Corbiac bladetechnique and other experiments. Tebiwa 12, 1–21.
Boshier, A. K. & Beaumont, P. B. (1972). Mining insouthern Africa and the emergence of modern man.Optima 22, 2–12.
Bowler, J. M. & Magee, J. W. (2000). RedatingAustralia’s oldest human remains: a skeptic’s view.J. hum. Evol. 38, 719–726.
Brandt, S. (1986). Upper Pleistocene and earlyHolocene prehistory of the Horn of Africa. Afr.Archaeol. Rev. 4, 41–82.
Brauer, G. (1984a). The ‘Afro-European sapienshypothesis’ and hominid evolution in East Asia dur-ing the Late Middle and Upper Pleistocene. Cour.Forsch.-Inst. Senckenberg 69, 145–165.
Brauer, G. (1984b). A craniological approach to theorigins of anatomically modern Homo sapiens inAfrica and implications for the appearance of modernEuropeans. In (F. H. Smith & F. Spencer, Eds) TheOrigins of Modern Humans: A World Survey of theFossil Evidence, pp. 327–410. New York: Alan R.Liss.
Brauer, G. (1989). The evolution of modern humans: acomparison of the African and non-African evidence.In (P. Mellars & C. Stringer, Eds) The HumanRevolution: Behavioral and Biological Perspectives on theOrigins of Modern Humans, pp. 123–154. Edinburgh:Edinburgh University Press.
Brauer, G. (1992). Africa’s place in the evolution ofHomo sapiens. In (G. Brauer & F. Smith, Eds)Continuity or Replacement: Controversies in Homosapiens Evolution, pp. 83–98. Rotterdam: Balkema.
Brauer, G. (1994). How different are Asian and AfricanHomo erectus? Cour. Forsch.-Inst. Senckenberg 171,301–318.
Brauer, G. & Leakey, R. E. (1986). The ES-11693cranium. J. hum. Evol. 15, 289–312.
Brauer, G. & Mbua, E. (1992). Homo erectus featuresused in cladistics and their variability in Asian andAfrican hominids. J. hum. Evol. 22, 79–108.
Brauer, G. & Mehlman, M. J. (1988). Hominid molarsfrom a Middle Stone Age level at the MumbaRock Shelter, Tanzania. Am. J. phys. Anthrop. 75,69–76.
Brauer, G. & Rimbach, K. W. (1990). Late archaicand modern Homo sapiens from Europe, Africa,and southwest Asia: Craniometric comparisons andphylogenetic implications. J. hum. Evol. 19, 789–807.
Brauer, G. & Singer, R. (1996). The Klasies zygomaticbone: archaic or modern? J. hum. Evol. 30, 161–166.
Brauer, G., Deacon, H. & Zipfel, F. (1982). Commenton the new maxillary finds from Klasies River, SouthAfrica. J. hum. Evol. 22, 419–422.
Brauer, G., Leakey, R. E. & Mbua, E. (1992). A firstreport on the ER-3884 cranial remains from Ileret/East Turkana, Kenya. In (G. Brauer & F. Smith,Eds) Continuity or Replacement: Controversies inHomo sapiens Evolution, pp. 111–120. Rotterdam:Balkema.
Brauer, G., Yokohama, Y., Falgueres, C. & Mbua, E.(1997). Human origins backdated. Nature 386,337–338.
Brennan, M. U. (1991). Health and disease inthe Middle and Upper Paleolithic of south-western France: a bioarchaeological study. Ph.D.Dissertation, New York University.

539 ’
Breuil, H., Teilhard de Chardin, P. & Weinert, P.(1951). Le paleolithique du Harar. L’Anthropologie55, 219–230.
Brink, J. S. (1988). The taphonomy and palaeoecologyof the Florisbad spring fauna. Paleoecol. Afr. 19,169–179.
Brink, J. S. & Deacon, H. J. (1982). A study of a lastinterglacial shell midden and bone accumulation atHerolds Bay, Cape Province, South Africa. Paleoecol.Afr. 15, 31–40.
Brink, J. A. & Grun, R. (1998). New dates fromFlorisbad: some implications for Mid-Quaternarymammalian evolution in southern Africa. Paperpresented at the Fourth Dual Congress of theInternational Association for the Study of HumanPaleontology and the International Association ofHuman Biologists, Sun City, South Africa.
Broadhurst, C. L., Cunnane, S. C. & Crawford, M. A.(1998). Rift Valley lake fish and shellfish providedbrain-specific nutrition for early Homo. British Journalof Nutrition 79, 3–21.
Brooks, A. S. (1995). L’Aurignacien de l’abri Pataudniveaux 6 a 14. In (H. M. Bricker, Ed.) Le Paleo-lithique Superieur de l’Abri Pataud (Dordogne): LesFouilles de H. L. Movius, Jr., pp. 167–222. Paris:Documents d’Archeologie Francaise, Maison desSciences de l’Homme.
Brooks, A. S. (1996). Behavior and human evolution.In (W. E. Meikle, F. C. Howell & N. G. Jablonski,Eds) Contemporary Issues in Human Evolution,pp. 135–166. California Academy of SciencesMemoir No. 21.
Brooks, A. S. & Robertshaw. P. T. (1990). The glacialmaximum in tropical Africa: 22,000 to 12,000 BP. In(O. Soffer & C. Gamble, Eds) The World at 18,000BP Vol. 2: Low Latitudes, pp. 121–169. London:Unwin Hyman.
Brooks, A. S. & Smith, C. C. (1987). Ishango revisited:new age determination and cultural interpretations.Afr. Archaeol. Rev. 5, 65–78.
Brooks, A. S. & Yellen, J. E. (1987). The preservationof activity areas in the archaeological record: ethno-archaeological and archaeological work in northwestNgamiland, Botswana. In (S. Kent, Ed.) Method andTheory of Activity Area Research: An Ethnoarchaeologi-cal Approach, pp. 63–106. New York: ColumbiaUniversity Press.
Brooks, A. S., Crowell, A. L. & Yellen, J. E. (1980).�Gi: a Stone Age archaeological site in the northernKalahari Desert, Botswana. In (R. E. F. Leakey &B. A. Ogot, Eds) Proceedings of the Eighth PanafricanCongress of Prehistory and Quaternary Studies (Nairobi,1977), pp. 304–309. Nairobi: The International LouisLeakey Memorial Institute for African Prehistory.
Brooks, A. S., Hare, P. E., Kokis, J. E., Miller, G. H.,Ernst, R. D. & Wendorf, F. (1990). DatingPleistocene archaeological sites by protein diagenesisin ostrich eggshell. Science 248, 60–64.
Brooks, A. S., Kokis, J. E. & Hare, P. E. (1993a).Dating the origins of modern humans. Paperpresented at the 2nd annual meeting of thepalaeoanthropology Society. Toronto.
Brooks, A. S., Hare, P. E. & Kokis, J. E. (1993b). Ageof anatomically modern human fossils from the caveof Klasies River Mouth, South Africa. Carnegie Inst.Wash. Yearb. 92, 95–96.
Brooks, A. S., Helgren, D. M., Cramer, J. M.,Franklin, A., Hornyak, W., Keating, J. M., Klein,R. G., Rink, W. J., Schwarcz, H. P., Smith, J. N. L.,Stewart, K., Todd, N. E., Verniers, J. & Yellen, J. E.(1995). Dating and context of three Middle StoneAge sites with bone points in the upper SemlikiValley, Zaire. Science 268, 548–553.
Brooks, A. S. (1998). Open-air sites in the MiddleStone Age of Africa. In (N. Conard & F. Wendorf,Eds) Middle Paleolithic and Middle Stone Age Settle-ment Systems. Proceedings of the XII InternationalCongress of Pre- and Protohistoric Sciences. FortiItaly, Vol 6, tome 1, Workshop 5, pp. 249–253.
Brooks, A. S., Hare, P. E., Kokis, J. E. & Durana, K.(1991). A burning question: Differences in labora-tory induced and natural diagenesis in ostricheggshell proteins. Annual Report of the GeophysicalLaboratory, Carnegie Institution of Washington,pp. 176–179.
Bunn, H. T. (1981). Archaeological evidence formeat-eating by Plio-Pleistocene hominids fromKoobi Fora and Olduvai. Nature 291, 574–577.
Bunn, H. T. (1989). Diagnosing Plio-Pleistocenehominid activity with bone fracture evidence. In (R.Bonnichsen & M. Sorg, Eds) Bone Modification,pp. 299–315. Orono: Center for the Study of theFirst Americans.
Bunn, H. T., Harris, J. W. K., Isaac, G., Kaufulu, Z.Kroll, E., Schick, K., Toth, N. & Behrensmeyer,A. K. (1980). FxJj50: an early Pleistocene site innorthern Kenya. World Archaeol. 12, 109–136.
Burkitt, M. C. (1921). Prehistory. Cambridge:Cambridge University Press.
Burkitt, M. C. (1928). South Africa’s Past in Stoneand Paint. Cambridge: Cambridge University Press.
Burkitt, M. C. (1933). The Old Stone Age: A Study ofPalaeolithic Times. Cambridge: Cambridge UniversityPress.
Butzer, K. W. (1969). Geological interpretation of twoPleistocene hominid sites in the lower Omo basin.Nature 222, 1133–1135.
Butzer, K. W. (1971). Recent History of an EthiopianDelta. Chicago: University of Chicao Department ofGeography Research Papers No. 136.
Butzer, K. W. (1978). Sediment stratigraphy of theMiddle Stone Age sequence at Klasies River Mouth.S. Afr. Archaeol. Bull. 33, 141–151.
Butzer, K. W. (1982). Geomorphology and sedimentstratigraphy. In (R. Singer & J. Wymer, Eds) TheMiddle Stone Age at Klasies River Mouth in SouthAfrica, pp. 33–42. Chicago: University of ChicagoPress.
Butzer, K. W. (1988a). Sedimentological interpretationof the Florisbad spring deposits. Paleoecol. Afr. 19,181–189.
Butzer, K. W. (1988b). A marginality model to explainmajor spatial and temporal gaps in the Old and New

540 . . .
World Pleistocene sediment records. Geoarchaeol. 3,193–203.
Butzer, K. W. & Isaac, G. Ll. (1975). Delimitation ofthe geologic time term ‘Middle Pleistocene’. In(K. W. Butzer & G. Ll. Isaac, Eds) After theAustralopithecines: Stratigraphy, Ecology, and CultureChange in the Middle Pleistocene, pp. 901–903. TheHague: Mouton.
Butzer, K. W., Brown, F. H. & Thurber, D. L. (1969).Horizontal sediments of the lower Omo Valley: theKibish Formation. Quaternaria 11, 15–29.
Butzer, K. W., Vogel, J. C. & Beaumont, P. B. (1978).Lithostratigraphy of the Border Cave, Kwazulu,South Africa, a Middle Stone Age sequencebeginning ca. 195,000 B. P. J. Archaeol. Sci. 5,317–341.
Byers, A. M. (1994). Symboling and the Middle-UpperPaleolithic transition: a theoretical and methodologi-cal critique. Curr. Anthrop. 35, 369–399.
Cabrera Valdes, V. & Bischoff, J. L. (1989). Acceler-ator 14C dates for early Upper Paleolithic (BasalAurignacian) at El Castillo Cave (Spain). J. Archaeol.Sci. 16, 577–584.
Cahen, D. (1976). Nouvelles fouilles a la Pointe de laGombe (ex-Pointe de Kalina), Kinshasa, Zaire.L’Anthropologie 80, 573–602.
Cahen, D. (1978). Vers la revision de la nomenclaturedes industries prehistoriques de l’Afrique centrale.L’Anthropologie 82, 5–36.
Campbell, B. (1964). Quantitative taxonomy andhuman evolution. In (S. L. Washburn, Ed.) Classifi-cation and Human Evolution, pp. 50–74. London:Methuen.
Cande, S. C. & Kent, D. V. (1992). A new geo-magnetic polarity time scale for the late Cretaceousand Cenozoic. J. geophys. Res. 97, 13917–13951.
Cann, R. L. (1988). DNA and human origins. A. Rev.Anthrop. 17, 127–143.
Cann, R. L., Stoneking, M. & Wilson, A. C. (1987).Mitochondrial DNA and human evolution. Nature325, 31–36.
Cattelain, P. (1997). Hunting during the Upper Paleo-lithic: bow, spearthrower, or both? In (H. Knecht,Ed.) Projectile Technology, pp. 213–240. New York:Plenum.
Chamberlain, J. G. (1996). The possible role oflong-chain omega-3 fatty acids in human brainphylogeny. Perspectives in Biology and Medicine 39,436–445.
Champion, D. E., Lanphere, M. A. & Kuntz, M. A.(1988). Evidence for a new geomagnetic reversalfrom lava flows in Idaho: discussion of short polarityreversals in the Brunhes and late Matuyama polaritychrons. J. geophys. Res. 93, 11,667–11,680.
Chappell, J., Head, J. & Magee, J. (1998). Beyondthe radiocarbon limit in Australian archaeology andQuaternary research. Radiocarbon 40, 543–553.
Chase, P. G. (1989). How different was MiddlePalaeolithic subsistence? A zooarchaeological per-spective on the Middle to Upper Palaeolithic tran-sition. In (P. Mellars & C. B. Stringer, Eds) TheHuman Revolution: Behavioral and Biological Perspec-
tives on the Origins of Modern Humans, pp. 321–327.Edinburgh: Edinburgh University Press.
Chase, P. G. & Dibble, H. L. (1987). Middle Paleo-lithic symbolism: a review of current evidence andinterpretation. J. Anthrop. Archaeol. 6, 263–296.
Chase, P. G. & Dibble, H. L. (1992). Scientificarchaeology and the origins of symbolism: a reply toBednarik. Cambridge Archaeol. J. 2, 43–51.
Chauvet, J.-M., Deschamps, E. B. & Hillaire, C.(1996). The Dawn of Art: The Chauvet Cave, theOldest Known Paintings in the World. New York:Abrams.
Chavaillon, J. (1982). Position chronologique deshominides fossiles d’Ethiopie. In (M. A. de Lumley,Ed.) Homo erectus et la Place de l’Homme de TautavelParmi les Hominides Fossiles. Premiere CongresInternational de Paleontolgie Humaine, pp. 766–797, Vol. 2. UNESCO Colloque International duCentre National de la Recherche Scientifique:Pretirages.
Chavaillon, J., Brahimi, C. & Coppens, Y. (1974).Premiere decouverte d’hominide dans l’un des sitesacheuleens de Melka-Kunture (Ethiopie). C. r. Acad.Sci., Paris 278, 3299–3302.
Chazan, M. (1994). Variability in the Middle Paleo-lithic of Haua Fteah, Libya. Paleorient 211, 65–73.
Cheney, D. L. & Seyfarth, R. M. (1990). How MonkeysSee the World. Chicago: University of Chicago Press.
Childe, V. G. (1928). The Most Ancient East. London:Routledge & Keegan Paul.
Childe, V. G. (1934). New Light on the Most AncientEast. London: Keegan Paul, Trench, Trubner.
Childe, V. G. (1936). Man Makes Himself. London:Watts.
Childe, V. G. (1942). What Happened in History.Harmondsworth: Penguin.
Childe, V. G. (1950). The urban revolution. TownPlanning Review 21, 3–17.
Churcher, C. S., Kleindienst, M. R. & Schwarcz, H. P.(1999). Faunal remains from a Middle Pleistocenelacustrine marl in Dakhleh Oassis, Egypt: palaeo-environmental reconstructions. Palaeogeogr., Palaeo-climatol, Palaeoecol. 154, 301–312.
Churchill, S. E., Pearson, O. M., Grine, F., Trinkaus,E. & Holliday, T. W. (1996). Morphological affini-ties of the proximal ulna from Klasies River main site:archaic or modern? J. hum. Evol. 31, 213–237.
Clark, G. A. & Lindly, J. M. (1989). The case ofcontinuity: observations on the biocultural transitionin Europe and Western Asia. In (P. Mellars & C. B.Stringer, Eds) The Human Revolution: Behavioraland Biological Perspectives on the Origins of ModernHumans, pp. 626–676. Edinburgh: EdinburghUniversity Press.
Clark, G. A. & Lindly, J. M. (1991). On paradigmaticbiases and Paleolithic research traditions. Curr.Anthrop. 32, 577–587.
Clark, J. D. (1954). The Prehistoric Cultures of the Hornof Africa. Cambridge: Cambridge University Press.
Clark, J. D. (Ed.) (1957a). Proceedings of the ThirdPanafrican Congress on Prehistory, Livingstone, 1955.London: Chatto & Windus.

541 ’
Clark, J. D. (1957b). A reexamination of the type site atMagosi, Uganda. In (J. D. Clark, Ed.) Proceedingsof the Third Panafrican Congress on Prehistory,Livingstone, 1955, pp. 228–241. London: Chatto &Windus.
Clark, J. D. (1959). Further excavations at Broken Hill,Northern Rhodesia. J. R. Anthrop. Inst. 89, 201–234.
Clark, J. D. (1964). The influence of environmentin inducing culture change at the Kalambo Fallsprehistoric site. S. Afr. Archaeol. Bull. 19, 93–101.
Clark, J. D. (1965). The later Pleistocene cultures ofAfrica. Science 150, 833–847.
Clark, J. D. (1969). Kalambo Falls Prehistoric Site,Vol. I. Cambridge: Cambridge University Press.
Clark, J. D. (1970). The Prehistory of Africa. New York:Praeger.
Clark, J. D. (1972). Human behavioural differencesin southern Africa during the Late Pleistocene.American Anthropologist 73, 1211–1236.
Clark, J. D. (1975). Africa in prehistory: peripheral orparamount? Man 10, 175–198.
Clark, J. D. (1980). Human populations and culturaladaptations in the Sahara and Nile during prehistorictimes. In (M. A. Williams & H. Faure, Eds) TheSahara and the Nile: Quaternary Environments andPrehistoric Occupation in North Africa, pp. 527–582.Rotterdam: A. A. Balkema.
Clark, J. D. (1981). New men, strange faces, otherminds: an archaeologist’s perspective on recent dis-coveries relating to the origins and spread of modernman. Proc. Brit. Acad. 67, 163–192.
Clark, J. D. (1982). The cultures of the Middle Paleo-lithic and Middle Stone Age. In (J. D. Clark, Ed.)The Cambridge History of Africa, Vol. 1: From theEarliest Times to 500 B.C., pp. 248–341. Cambridge:Cambridge University Press.
Clark, J. D. (1988). The Middle Stone Age of EastAfrica and the beginnings of regional identity.J. World. Prehist. 2, 235–305.
Clark, J. D. (1989). The origins and spread of modernhumans: a broad perspective on the African evidence.In (P. Mellars & C. B. Stringer, Eds) The HumanRevolution: Behavioral and Biological Perspectives on theOrigins of Modern Humans, pp. 565–588. Edinburgh:Edinburgh University Press.
Clark, J. D. (1993). African and Asian perspectiveson the origins of modern humans. In (M. J. Aitken,C. B. Stringer & P. A. Mellars, Eds). The Originof Modern Humans and the Impact of ChronometricDating, pp. 148–178. Princeton: PrincetonUniversity Press.
Clark, J. D. (1998). The early paleolithic of the easternregion of the Old World in comparison to the west.In (M. Petraglia & R. Korisettar, Eds). Early HumanBehavior in Global Context: The Rise and Diversity ofthe Lower Paleolithic Record, pp. 437–450. London:Routledge.
Clark, J. D. & Kleindienst, M. R. (1974). The StoneAge cultural sequence: terminology, typology andraw material. In (J. D. Clark, Ed.) Kalambo FallsPrehistoric Site, Vol. II, pp. 71–106. Cambridge:Cambridge University Press.
Clark, J. D., Oakley, K. P., Wells, L. H. & McClelland,J. A. C. (1950). New studies on Rhodesian Man.J. R. Anthrop. Inst. 77, 7–32.
Clark, J. D., Brothwell, D. R., Powers, R. & Oakley,K. P. (1968). Rhodesian man: notes on a new femurfragment. Man 3, 105–111.
Clark, J. D., Phillips, J. L. & Staley, P. S. (1974).Ancient Egyptian bows and arrows and theirrelevance for African prehistory. Paleorient 2,323–388.
Clark, J. D., Phillips, J. L. & Staley, P. S. (1976).Interpretations of prehistoric technology fromancient Egyptian and other sources. Part II: Pre-historic arrow forms in Africa as shown by surviv-ing examples of the traditional arrows of the SanBushmen. Paleorient 3, 127–150.
Clark, J. D., Williamson, K. D., Michels, J. W. &Marean, C. A. (1984). A Middle Stone occupationsite at Porc Epic Cave, Dire Dawa. Afr. Archaeol.Rev. 2, 37–72.
Clark, J. D., de Heinzelin, J., Schick, K. D., Hart,W. K., White, T. D., WoldeGabriel, G., Walter,R. C., Suwa, G., Asfaw, B., Vrba, E. & HaileSelassie,Y. (1994). African Homo erectus: old radiometric agesand young Oldowan assemblages in the MiddleAwash valley, Ethiopia. Science 264, 1907–1910.
Clark, J. G. D. (1951). Neolithic bows from Somerset,England, and the prehistory of archery in northwestEurope. Proc. prehist. Soc. 29, 50–98.
Clark, J. G. D. (1952). Prehistoric Europe: The EconomicBasis. London: Methuen.
Clark, J. G. D. (1977). World Prehistory: A New Outline.Cambridge: Cambridge University Press.
Clarke, R. J. (1976). A new cranium of Homoerectus from Lake Ndutu, Tanzania. Nature 262,485–487.
Clarke, R. J. (1985). A new reconstruction of theFlorisbad cranium with notes on the site. In (E.Delson, Ed.) Ancestors: The Hard Evidence,pp. 301–305. New York: Alan R. Liss.
Clarke, R. J. (1990). The Ndutu cranium and theorigin of Homo sapiens. J. hum. Evol. 19, 699–736.
Close, A. (1984). Current research and recent radio-carbon dates from North Africa, II. J. Afr. Hist. 25,1–24.
Close, A. E., Wendorf, F. & Schild, R. (1990).Patterned use of a Middle Palaeolithic landscape: BirTarfawi and Bir Sagara East, Eastern Sahara. Sahara3, 212–234.
Clottes, J. (1993). Paint analyses from severalMagdalenian caves in the Ariege region of France.J. Archaeol. Sci. 20, 223–235.
Clottes, J., Menu, M. & Walter, P. (1990). Lapreparation des peintures magdaleniennes descavernes ariegoises. Bull. Soc. prehist. fr. 71,170–192.
Clottes, J., Valladas, H., Cachier, H. & Arnold, M.(1992). Des dates pour Niaux et Gargas. Bull. Soc.prehist. fr. 89, 270–274.
Clottes, J., Chauvet, J.-M., Brunel Deschamps, E.,Hillaire, C., Daugas, J. P., Arnold, M., Cachier, H.,Evin, J., Forin, P., Oberlin, C., Tisnerat, N. &

542 . . .
Valladas, H. (1995). Les peintures paleolithiques dela Grotte Chauvet-Pont d’Arc, a Vallon-Pont-d’Arc(Ardeche, France): datations directes et indirectespar la methode du radiocarbonne. C. r. Acad. Sci.,Paris 320, 1133–1140.
Cole, G. H. (1967). A reinvestigation of Magosi andthe Magosian. Quaternaria 9, 153–168.
Cole, S. (1954). The Prehistory of East Africa. London:Penguin.
Combier, F. (1989). Soyons, Grotte de Neron (Ardeche).Archeologie de la France, 30 Ans de Deccouvertes. Paris:Reunion des Musees Nationaux 65.
Conard, N. & Adler, D. (1997). Lithic reduction andhominid behavior in the Middle Paleolithic of theRhineland. J. Anthrop. Res. 53, 147–176.
Conard, N. J. (1990). Laminar lithic assemblages fromthe last interglacial complex in northwestern Europe.J. Anthrop. Res. 46, 243–262.
Conkey, M. W. (1990). Experimenting with style inarchaeology: some historical and theoretical issues.In (M. W. Conkey & C. Hastorf, Eds) The Usesof Style in Archaeology, pp. 5–17. Cambridge:Cambridge University Press.
Conroy, G. C., Jolly, C. J., Cramer, D. & Kalb, J. E.(1978). Newly discovered fossil hominid skull fromthe Afar depression. Nature 275, 67–70.
Cooke, C. K. (1966). Rhodesian prehistory reexam-ined. Part I: the Stone Age. Arnoldia 2, 1–8.
Cooke, C. K. (1969). A reexamination of the MiddleStone Age of Rhodesia. Arnoldia 4, 1–18.
Cooke, C. K. (1971). Excavations in Zombepata Cave,Sipolilo District, Mashonaland, Rhodesia. S. Afr.Archaeol. Bull. 26, 104–126.
Cooke, C. K. & Paterson, M. L. (1960). A MiddleStone Age open site, Ngamiland, BechuanalandProtectorate. S. Afr. Archaeol. Bull. 58, 36–39.
Cooke, H. B. S. (1962). Notes on the faunal remainsfrom the Cave of Hearths and Kalkbank. In (R. J.Mason, Ed.) Prehistory of Gauteng, pp. 447–453. Johannesburg: University of WitwatersrandPress.
Cooke, H. B. S. (1963). Pleistocene mammal faunas ofAfrica, with particular reference to southern Africa.In (F. C. Howell & F. Bouliere, Eds) African Ecologyand Human Evolution, pp. 65–116. Chicago: Aldine.
Cooke, H. B. S., Malan, B. D. & Wells, L. H. (1945).Fossil man in the Lembombo Mountains, SouthAfrica: the ‘‘Border Cave’’, Ingwavuma District,Zululand. Man 45, 6–13.
Cornelissen, E. (1992). Site GnJh-17 and its Implicationsfor the Archaeology of the Middle KapthurinFormation, Baringo, Kenya. Tervuren: Musee Royalede l’Afrique Centrale, Annales, Sciences Humaines133.
Cornelissen, E. (1995). Indications du post-Acheuleen(Sangoen) dans la Formation Kapthurin, Baringo,Kenya. L’Anthropologie 99, 55–73.
Cornelissen, E., Boven, A., Dabi, A., Hus, J., Ju Yong,K., Keppens, E., Langohr, R., Moeyersons, J.,Pasteels, P., Pieters, M., Uytterschaut, H., VanNoten, F. & Workineh, H. (1990). The KapthurinFormation revisited. Afr. Archaeol. Rev. 8, 23–76.
Couraud, C. (1988). Pigments utilises en prehistoire:provenence, preparation, mode d’utilisation.L’Anthropologie 92, 17–28.
Couraud, C. (1991). Les pigments des grottes d’Arcysur Cure (Yonne). Gallia Prehist. 33, 17–52.
Crawford, M. A. (1992). The role of dietary fatty acidsin biology: their place in the evolution of the humanbrain. Nutrition Reviews 50, 3–11.
Crawford, M. A., Bloom, M., Broadhurst,C. L., Schmidt, W. F., Cunnane, S. C. G. C.,Gehbresmeskel, K., Linseisen, F., Lloyd-Smith, J. &Parkington, J. (1999). Evidence for the unique func-tion of docasahexaenoic acid during the evolution ofthe modern human brain. Lipids 34 (Supplement)S39–S47.
Cremaschi, M., Di Lernia, S., & Garcea, E. A. A.(1998). Some insights on the Aterian in the LibyanSahara: chronology, environment, and archeology.Afr. Archaeol. Rev. 15, 261–286.
Crowell, A. & Hitchcock, R. (1978). Basarwa ambushhunting in Botswana. Botswana Notes and Records 10,37–51.
Curtis, G. H. (1966). The problems of contaminationin obtaining accurae dates of young geologic rocks. In(O. A. Shaeffer & J. Zahringer, Eds) Potassium-ArgonDating, pp. 151–162. New York: Springer Verlag.
Dagley, P., Musset, A. E. & Palmer, H. C. (1978).Preliminary observations on the paleomagneticstratigraphy of the area west of Lake Baringo, Kenya.In (W. W. Bishop, Ed.) Geological Backgroundto Fossil Man, pp. 361–373. Edinburgh: ScottishAcademic Press.
Dart, R. A. (1967). The antiquity of mining in SouthAfrica. S. Afr. J. Sci. 63, 264–267.
Dart, R. A. & Del Grande, N. (1931). The ancient ironsmelting cavern at Mumbwa. Trans. R. Soc. S. Afr.19, 379–427.
Darwin, C. (1859). On the Origin of Species by Means ofNatural Selection. London: John Murray.
Davidson, I. & Noble, W. (1992). Why the first col-onisation of the Australian region is the earliestevidence for modern human behavior. Perspec. Hum.Biol./Archaeol. Oceania 27, 113–119.
Day, M. H. (1969). The Omo skeletal remains. Nature222, 1135–1138.
Day, M. H. (1971). Postcranial remains of Homo erectusfrom Bed IV, Olduvai Gorge, Tanzania. Nature 232,383–387.
Day, M. H. (1972). The Omo human skeletal remains.In (F. Bordes, Ed.) The Origin of Homo Sapiens,pp. 31–35. Paris: UNESCO.
Day, M. H. (1973). The development of Homo sapiens.Darwin Centenary Symposium on the Origin of Man,pp. 87–95. Rome: Accademia Nazionale dei Lincei.
Day, M. H. (1986). Guide to Fossil Man. 4th edn.Chicago: University of Chicago Press.
Day, M. H. & Stringer, C. B. (1982). A reconsiderationof the Omo Kibish remains and the erectus-sapienstransition. In (M. A. de Lumley, Ed.) Homo erectus etla Place de l’Homme de Tautavel parmi les HominidesFossiles. Premiere Congres International de PaleontolgieHumaine. UNESCO Colloque International du Centre

543 ’
National de la Recherche Scientifique Vol. 2,pp. 814–846. Nice: Pretirages.
Day, M. H. & Stringer, C. B. (1991). Les restescraniens d’Omo-Kibish et leur classification al’interieur du genre Homo. L’Anthropologie 95, 573–594.
Day, M. H., Leakey, M. D. & Magori, C. (1980). Anew hominid fossil skull (L. H. 18) from the NgalobaBeds, Laetoli, northern Tanzania. Nature 284,55–56.
d’Errico, F., Zilhao, J., Julien, M., Baffier, C. &Pelegrin, J. (1998). Neanderthal acculturation inwestern Europe? Curr. Anthrop. 39, S2–S44.
de Beaune, S. (1993). Nonflint stone tools of the earlyUpper Paleolithic. In (H. Knecht, A. Pike-Tay & R.White, Eds). Before Lascaux: The Complex Record ofthe Early Upper Paleolithic, pp. 137–162. Boca Raton:CRC Press.
de Bonis, L., Geraads, D., Guerin, G., Haga, A.,Jaeger, J. J. & Sen, S. (1984). Decouverte d’unhominide fossile dans le Pleistocene de la Republiquede Djibouti. C. r. Acad. Sci., Paris 299, 1097–1100.
de Bonis, L., Geraads, D., Jaeger, J.-J. & Sen, S.(1988). Vertebres du Pleistocene de Djibouti. Bull.Soc. geol. Fr. 4, 323–334.
de Garine, I. & Harrison, G. A. (Eds) (1988). CopingWith Uncertainty in Food Supply. Oxford: Clarendon.
de Heinzelin, J. (1957). Les Fouilles d’Ishango. Explor-ation de Parc National Albert (Fasc. 2). Brussels:Institut des Parcs Nationaux du Congo Belge.
de Heinzelin, J. (1962). Ishango. Scient. Am. 26,105–111.
de Mortillet, G. (1900). Le Prehistorique. 3rd edn. Paris:C. Reiwald.
de Villiers, H. (1973). Human skeletal remains fromBorder Cave, Ingwavuma District, KwaZulu, SouthAfrica. Ann. Transv. Mus. 28, 229–256.
de Villiers, H. (1978). A second adult human mandiblefrom Border Cave, Ingwavuma District, KwaZulu,South Africa. S. Afr. J. Sci. 72, 212–215.
Deacon, H. J. (1979). Excavations at BoomplaasCave: a sequence through the Upper Pleistoceneand Holocene in South Africa. World Archaeol. 10,241–257.
Deacon, H. J. (1985). Review of Binford, L. R. 1984.Faunal remains from Klasies River Mouth. S. Afr.Archaeol. Bull. 40, 59–60.
Deacon, H. J. (1988). The origins of anatomicallymodern people and the South African evidence.Paleoecol. Afr. 19, 193–200.
Deacon, H. J. (1989). Late Pleistocene paleoecologyand archaeology in the southern Cape. In (P. Mellars& C. B. Stringer, Eds) The Human Revolution:Behavioral and Biological Perspectives on the Originsof Modern Humans, pp. 547–564. Edinburgh:Edinburgh University Press.
Deacon, H. J. (1993a). Planting an idea: an archaeol-ogy of Stone Age gatherers in South Africa. S. Afr.Archaeol. Bull. 48, 86–93.
Deacon, H. J. (1993b). Southern Africa and modernhuman origins. In (M. J. Aitken, C. B. Stringer &P. A. Mellars, Eds) The Origin of Modern Humans
and the Impact of Chronometric Dating, pp. 104–117.Princeton: Princeton University Press.
Deacon, H. J. (1995). Two late Pleistocene-Holocenearchaeological depositories from the SouthernCape, South Africa. S. Afr. Archaeol. Bull. 50, 121–131.
Deacon, H. J. (1998). Elandsfontein and Klasies Riverrevisited. In (N. M. Ashton, F. Healy & P. B. Pettitt,Eds) A Master of His Craft: Papers in Stone AgeArchaeology Presented to John Wymer, pp. 23–28.Oxford: Oxbow Press.
Deacon, H. J. & Decon, J. (1999). Human Beginnings inSouth Africa: Uncovering the Secrets of the Stone Age.Capetown: David Philip.
Deacon, H. J. & Geleijnse, V. B. (1988). The stratigra-phy and sedimentology of the main site sequence,Klasies River, South Africa. S. Afr. Archaeol. Bull. 43,5–14.
Deacon, H. J. & Shurman, R. (1992). The origins ofmodern people: the evidence from Klasies River. In(G. Brauer & F. H. Smith, Eds) Continuity or Replace-ment: Controversies in Homo sapiens Evolution,pp. 121–130. Rotterdam: Balkema.
Deacon, H. J. & Thackeray, J. F. (1984). LateQuaternary environmental changes and implicationsfrom the archaeological record in southern Africa.In (J. C. Vogel, Ed.) Late Cainozoic Paleoclimates ofthe Southern Hemisphere, pp. 339–351. Rotterdam:Balkema.
Deacon, H. J. & Wurz, S. G. D. (1996). Klasies RiverMain Site Cave 2: a Howiesonspoort occurrence.In (G. Pwiti & R. Soper, Eds) Aspects of AfricanArchaeology, pp. 213–218. Harare: University ofZimbabwe Publications.
Deacon, H. J., Deacon, J., Scholtz, A., Thackerary,J. F., Brink, J. B. & Vogel, J. C. (1984). Correlationof palaeoenvironmental data from the LatePleistocene and Holocene deposits at BoomplaasCave, southern Cape. In (J. C. Vogel, Ed.) LateCainozoic Palaeoclimates of the Southern Hemisphere,pp. 339–351. Rotterdam: Balkema.
Deacon, H. J., Geleijnse, V. B., Thackeray, A. I.,Thackeray, J. F., Tusenius, M. L. & Vogel, J. C.(1986). Late Pleistocene cave deposits in thesouthern Cape: current research at Klasies River.Paleoecol. Afr. 17, 31–37.
Deacon, J. (1984). Later Stone Age peoples and theirdescendants in southern Africa. In (R. G. Klein, Ed.)Southern African Prehistory and Paleoenvironments,pp. 221–328. Rotterdam: Balkema.
Deacon, T. (1997). The Symbolic Species: TheCo-Evolution of Language and the Brain. New York:Norton.
Dean, D., Kalvin, A. D., Hublin, J.-J. & Braun, M.(1993). 3D CT composite reconstruction of Sale andThomas Quarry cranial remains. Am. J. phys.Anthrop. Supplement 16, 79.
Debenath, A. (1980). Nouveaux restes humainsateriens du Maroc. C. r. Acad. Sci., Paris 290,851–852.
Debenath, A. (1994). L’Aterien du nord de l’Afriquedu Sahara. Sahara 6, 21–30.

544 . . .
Debenath, A. & Dibble, H. L. (1994). Handbook ofPaleolithic Typology. Volume One: Lower and MiddlePaleolithic of Europe. Philadelphia: UniversityMuseum, University of Pennsylvania.
Debenath, A. & Sbihi-Alaoui, F. (1979). Decouvertede deux nouveaux gisements prehistoriques pres deRabat (Maroc). Bull. Soc. prehist. fr. 76, 11–12.
Debenath, A., Raynal, J.-P. & Texier, J.-P. (1982).Position stratigraphiqe des restes humains paleo-lithiques marocains sur la base des travaux recentes.C. r. Acad. Sci., Paris 294, 972–976.
Debenath, A., Raynal, J.-P., Roche, J., Texier, J.-P. &Ferembach, D. (1986). Stratigraphie, habitat,typologie et devinir de l’Aterien Marocain: donneesrecentes. L’Anthropologie 90, 233–246.
Defleur, A. (1993). Les Sepultures Mousteriennes. Paris:C.N.R.S.
Deino, A. & McBrearty, S. (under review). A precise40Ar/39Ar chronology for the Kapthurin Formation,Baringo, Kenya. J. Hum. Evol.
Deino, A. & Potts, R. (1990). Single crystal 40Ar/39Ardating of the Olorgesailie Formation, southernKenya Rift. J. geophys. Res. 95, 8453–8470.
Delson, E. & Van Couvering, J. A. (2000). Compositestratigraphic chart of Olduvai Gorge. In (E. Delson,I. Tattersall, J. A. Van Couvering & A. S. Brooks,Eds) Encyclopedia of Human Evolution and Prehistory,2nd. edn. p. 488. New York: Garland, TaylorFrancis.
Delson, E., Tattersall, I., Van Couvering, J. A. &Brooks, A. S. (Eds) (2000). Encyclopedia of HumanEvolution and Prehistory. 2nd edn. New York:Garland/Taylor & Francis.
Demars, P.-Y. (1992). Les colorants dans leMousterien du Perigord: l’apport des fouilles de F.Bordes. Prehistoire Ariegeoise 47, 185–194.
Demars, P. Y. & Hublin, J.-J. (1989). La transitionneandertalien/hommes de type moderne en Europeoccidentale: aspects paleontologiques et culturels.Etudes et Recherches Archeologiques de l’Universite deLiege 34, 23–27.
Deshmukh, I. (1986). Ecology and Tropical Biology. PaloAlto: Blackwell.
Diamond, J. (1992). The Third Chimpanzee. New York:Harper Collins.
Dibble, H. (1989). The implication of stone tool typesfor the presence of language during the Lower andMiddle Paleolithic. In (P. Mellars & C. B. Stringer,Eds) The Human Revolution: Behavioral and BiologicalPerspectives on the Origins of Modern Humans, pp.415–432. Edinburgh: Edinburgh University Press.
Dobzhansky, T. G. (1937). Genetics and the Origins ofSpecies. New York: Columbia University Press.
Drennan, M. R. (1953). A preliminary note on theSaldanha skull. S. Afr. J. Sci. 50, 7–11.
Dreyer, T. F. (1935). A hominid skull from Florisbad,Orange Free State, with a note on the endocranialcast by C. U. Ariens Kappers. Proc. Amsterdam Acad.Sci. 38, 119–128.
Dreyer, T. F. (1936). The endocranial cast of theFlorisbad skull. Soologiese Navorsing van die NasionalMuseum Bloemfontein 1, 21–23.
Eldredge, N. & Gould, S. J. (1972). Punctuatedequilibria: an alternative to phyletic gradualism. In(T. J. M. Schopf, Ed.) Models in Paleobiology,pp. 82–115.
Ennouchi, E. (1962). Un Neanderthalien: l’Hommedu Jebel Irhoud (Maroc). L’Anthropologie 66, 279–299.
Ennouchi, E. (1963). Les Neanderthaliens du JebelIrhoud (Maroc). C. r. Acad. Sci., Paris 256,2459–2560.
Ennouchi, E. (1966). Le site du Jebel Irhoud (Maroc).In Actas del V Congreso Panafricano de Prehistoria y deEstudio del Cuaternario (Santa Cruz de Tenerife,1963), pp. 53–60. Santa Cruz de Tenerife: MuseoArqueologico Publicaciones 6.
Ennouchi, E. (1968). Le deuxieme crane de l’hommede l’Irhoud. Ann. Paleontol. 55, 117–128.
Ennouchi, E. (1969a). Decouverte d’un Pithecan-thropine au Maroc. C. r. Acad. Sci., Paris 269,753–765.
Ennouchi, E. (1969b). Presence d’un enfant neander-thalien au Jebel Irhoud (Maroc). Ann. Paleontol. 55,251–265.
Ennouchi, E. (1970). Un nouvel Archanthropien auMaroc. Ann. Paleontol. (Vert.) 56, 95–107.
Evernden, J. F. & Curtis, G. H. (1965). Potassium-argon dating of Late Cenozoic rocks in East Africaand Italy. Curr. Anthrop. 6, 343–385.
Falk, D. (1975). Comparative anatomy of the larynxin man and the chimpanzee: implications forlanguage in Neandertal. Am. J. phys. Anthrop. 43,123–132.
Falk, D. (1980). Language, handedness, and primatebrain. Am. Anthrop. 82, 72–78.
Falk, D. (1990). Brain evolution in Homo. Am. J. phys.Anthrop. 13, 333–381.
Farizy, C. (1990). The transition from Middle toUpper Paleolithic at Arcy-sur-Cure (Yonne, France):technological, economic, and social aspects. In (P.Mellars, Ed.) The Emergence of Modern Humans:An Archaeological Perspective, pp. 303–326. Ithaca:Cornell University Press.
Farrand, W. R. (1985). Rockskelter and cave sedi-ments. In (J. K. Stein & W. R. Farrand, Eds)Archaeological Sediments in Context, pp. 21–39.Orono, ME: Center for the Study of Early Man.
Feathers, J. K. (1996). Luminescence dating andmodern human origins. Evol. Anthrop. 5, 25–36.
Feathers, J. K. & Bush, D. A. (2000). Luminescencedating of Middle Stone Age deposits at Die Kelders.J. hum. Evol. 38, 91–120.
Feathers, J. K. & Migliorini, E. (2000). Lumi-nescence dating at Katanda—a reassessment.Quatern. Geochronol. (in press).
Feblot-Augustins, J. (1997). La Circulation des MatieresPremieres au Paleolithique (2 Vols). Liege: ERAUL/CNRS no 75.
Feibel, C. S., Brown, F. H. & MacDougall, I. (1989).Stratigraphic context of fossil hominids from theOmo Group deposits, northern Turkana Basin,Kenya and Ethiopia. Am. J. phys. Anthrop. 78, 595–622.

545 ’
Feidler, L. & Preuss, J. (1985). Stone tools from theinner Zaire basin (Region de l’Equateur, Zaire). Afr.Archaeol. Rev. 3, 179–187.
Ferembach, D. (1976a). Les restes humains Ateriensde la Grotte de Temara (Campagne 1975). Bull.Mem. Soc. Anthrop., Paris 3, 175–180.
Ferembach, D. (1976b). Les restes humains de laGrotte de Dar-es-Soltane 2 (Maroc) Campagne1975. Bull. Mem. Soc. Anthrop., Paris 3, 183–193.
Fisher, A. (1985). Hunting with flint-tipped arrows:results and experiences from practical experiments.In (C. Bonsall, Ed.) The Mesolithic in Europe,pp. 29–39. Edinburgh: John Donald.
Flynn, L. J., Barry, J. C., Morgan, M. E., Pilbeam, D.,Jacobs, L. L. & Lindsay, H. E. (1995). NeogeneSiwalik mammalian lineages: species longevities,rates of change and modes of speciation. Palaeogeogr.,Palaeoclimatol., Palaeoecol. 115, 249–264.
Fock, G. J. (1954). Stone balls in the WindhoekMuseum. S. Afr. Archaeol. Bull. 9, 108–109.
Fock, G. J. (1972). Extinct bovine on rock engraving.Res. Nat. Mus., Bloemfontein 2, 349–353.
Foley, R. (1987). Another Unique Species. New York:Longman.
Foley, R. (1991a). How many species of hominidshould there be? J. hum. Evol. 20, 413–428.
Foley, R. (1991b). Language origins: the silence of thepast. Nature 353, 114–115.
Foley, R. (1994). Speciation, extinction, and climaticchange in hominid evolution. J. hum. Evol. 26,275–289.
Foley, R. A. & Lahr, M. M. (1992). Beyond out ofAfrica: reassessing the origins of Homo sapiens.J. hum. Evol. 22, 523–529.
Foley, R. A. & Lahr, M. M. (1997). Mode 3technologies and the evolution of modern humans.Cambridge Archaeol. J. 7, 3–36.
Foley, R. A. & Lee, P. C. (1989). Finite social space,evolutionary pathways, and reconstructing hominidbehavior. Science 243, 901–906.
Franklin, A. & Hornyak, W. (n.d.). Thermolumi-nescence dating of Kalahari sediments: a progressreport to the National Science Foundation, 1984.
Freundlich, J. C., Schwabedissen, H. & Wendt, W. E.(1980). Koln radiocarbon measurements II.Radiocarbon 22, 68–81.
Friss-Hansen, J. (1990). Mesolithic cutting arrows:functional analysis of arrows used in the hunting oflarge game. Antiquity 64, 494–504.
Gamble, C. (1986). The Palaeolithic Settlement ofEurope. Cambridge: Cambridge University Press.
Gamble, C. (1994). Timewalkers: The Prehistory ofGlobal Colonization. Cambridge: Harvard UniversityPress.
Gargett, R. H. (1989). Grave shortcomings: the evi-dence for Neanderthal burial. Curr. Anthrop. 30,157–190.
Gargett, R. H. (1999). Middle Palaeolithic burial is nota dead issue: the view from Qafzeh, Saint-Cesaire,Kebara, Amud, and Dederiyeh. J. hum. Evol. 37,27–90.
Geneste, J.-M. (1989a). Economie des ressourceslithiques dans le Mousterien du sud-ouest de laFrance. In (M. Otte, Ed.) L’Homme de Neandertal,Vol. 6: La Subsistence, pp. 75–97. Liege: Eutudes etRecherches Archeologiques de l’Universite deLiege.
Geneste, J.-M. (1989b). Systemes d’approvisionnementen matieres premieres au paleolithique superieur enAquitaine. In (M. Otte, Ed.) L’Homme de Neandertal,Vol. 8: La Mutation, pp. 61–70. Liege: Eutudeset Recherches Archeologiques de l’Universite deLiege.
Gentry, A. (1978). Bovidae. In (V. Maglio & H. B. S.Cooke, Eds) Evolution of African Mammals,pp. 540–572. Cambridge: Harvard University Press.
Geraads, D. (1981). Bovidae et Giraffidae (Artio-dactyla, Mammalia) du pleistocene de Ternifine(Algerie). Bull. Mus. Hist. nat., Paris C3, 47–86.
Geraads, D., Hublin, J.-J., Jaeger, J.-J., Tong, H., Sen,S. & Toubeau, P. (1986). The Pleistocene hominidsite of Ternifine, Algeria: new results on the environ-ment, age, and human industries. Quatern. Res. 25,380–386.
Geraads, D., Amani, F. & Hublin, J.-J. (1992). Legisement pleistocene moyen de l’Ain Maarouf presde El Hajeb, Maroc: presence d’un hominide. C. r.Acad. Sci., Paris 314, 319–323.
Gibson, K. R. (1993). Tool use, language, and socialbehavior in relation to information processingcapacities. In (K. R. Gibson & T. Ingold, Eds) Tools,Language and Cognition in Human Evolution,pp. 251–269. Cambridge: Cambridge UniversityPress.
Gibson, K. R. & Ingold, T. (Eds) (1993). Tools,Language and Cognition in Human Evolution.Cambridge: Cambridge University Press.
Gifford-Gonzalez (1991). Bones are not enough:analogues, knowledge, and interpretive strategiesin zooarchaeology. J. Anthrop. Archaeol. 3, 215–254.
Gillespie, R. & Roberts, R. G. (2000). On the reliabilityof age estimates for human remains at Lake Mungo.J. hum. Evol. 38, 727–730.
Gingerich, P. D. (1984). Punctuated equilibria: whereis the evidence? Syst. Zool. 33, 335–338.
Giorgini, M. S. (1971). Soleb II, Les Necropoles. Firenze:Sansoni.
Goldberg, P. (2000). Micromorphology and siteformation at Die Kelders Cave 1, South Africa.J. hum. Evol. 38, 43–90.
Goldstein, C. B., Linares, A. R., Cavalli-Sforza, L. L. &Feldman, M. W. (1995). Genetic absolute datingbased on microsatellites and the origin of modernhumans. Proc. natl. Acad. Sci. USA 92, 6723–6727.
Goodwin, A. J. H. (1928). An introduction to theMiddle Stone Age in South Africa. S. Afr. J. Sci. 25,410–418.
Goodwin, A. J. H. & van Riet Lowe, C. (1929). TheStone Age cultures of South Africa. Ann. S. Afr. Mus.27, 1–289.
Gore, R. (2000). People like us. National Geographic198: 90–117.

546 . . .
Goren-Inbar, N. (1988). Notes on ‘‘decision making’’by Lower and Middle Palaeolithic hominids.Paleorient 14, 99–108.
Gould, R. A. (1978). The anthropology of humanresidues. Am. Anthrop. 80, 815–835.
Gould, S. J. & Eldredge, N. (1996). Punctuatedequilibrium comes of age. Nature 366, 233–227.
Gowlett, J. A. J. & Hedges, R. E. M. (1986). Lessonsof context and contamination in dating the UpperPalaeolithic. In (J. A. J. Gowlett & R. E. M. Hedges,Eds) Archaeological Results From Accelerator Dating,pp. 63–72. Oxford: British Archaeological ReportsInternational Series 11.
Gragson, T. L. (1992). Fishing the waters ofAmazonia: native subsistence economies in a tropicalrainforest. Am. Anthrop. 94, 428–440.
Graves-Brown, P. M. (1994). Flakes and ladders: whatthe archaeological record cannot tell us about theorigins of language. World Archaeol. 26, 158–171.
Greene, K. (1999). V. Gordon Childe and the vocabu-lary of revolutionary change. Antiquity 73, 97–109.
Gresham, T. H. (1984). An investigation of an UpperPleistocene archaeological site in northern Somalia.M.A. Thesis, University of Georgia, Athens.
Gresham, T. H. & Brandt, S. A. (1996). Variabilityin the MSA of the Horn of Africa. In (G. Pwiti &R. Soper, Eds) Aspects of African Archaeology,pp. 157–163. Harare: University of ZimbabwePublications.
Grine, F. E. (2000). Middle Stone Age human fossilsfrom Die Kelders Cave 1, Western Cape Province,South Africa. J. hum. Evol. 38, 129–146.
Grine, F. E. & Klein, R. G. (1985). Pleistocene andHolocene human remains from Equus Cave, SouthAfrica. Anthropology 8, 55–98.
Grine, F. E. & Klein, R. G. (1993). Late Pleistocenehuman remains from the Sea Harvest site, SaldanhaBay, South Africa. S. Afr. J. Sci. 89, 145–152.
Grine, F. E., Klein, R. G. & Volman, T. P. (1991).Dating, archaeology and human fossils from theMiddle Stone Age levels of Die Kelders, SouthAfrica. J. hum. Evol. 21, 363–395.
Grine, F. E., Jungers, W. L., Tobias, P. V. & Pearson,O. M. (1995). Fossil Homo femur from BergAukas, northern Namibia. Am. J. phys. Anthrop. 95,151–186.
Grine, F. E., Pearson, O. M., Klein, R. G. &Rightmire, G. P. (1998). Additional human fossilsfrom Klasies River Mouth, South Africa. J. hum.Evol. 35, 95–107.
Groves, C. (1989). A Theory of Human and PrimateEvolution. Oxford: Oxford University Press.
Groves, C. P. (1998). The proximal ulna from KlasiesRiver. J. hum. Evol. 34, 119–121.
Groves, C. P. (1999). Nomenclature of African Plio-Pleistocene hominins. J. hum. Evol. 37, 869–872.
Gruet, M. (1954). Le gisement mousterian d’ElGuettar. Karthago 5, 3–87.
Gruet, M. (1955). Amoncellement pyramidal despheres calcaires dans une source fossile mousteri-enne a el’Guettar (Sud tunisien). In Actes du II
Congres Pan-African de Prehistoire (Algiers, 1952),pp. 449–456, 460. Paris.
Grun, R. & Stringer, C. B. (1991). Electron spinresonance dating and the evolution of modernhumans. Archaeometry 33, 153–199.
Grun, R., Beaumont, P. B. & Stringer, C. B. (1990).ESR dating evidence for early modern humans atBorder Cave in South Africa. Nature 344, 537–539.
Grun, R., Stringer, C. B. & Schwarcz, H. P. (1991).ESR dating of teeth from Garrod’s Tabun Cavecollection. J. hum. Evol. 20, 231–248.
Grun, R., Brink, J. S., Spooner, N. A., Taylor, L.,Stringer, C. B., Franciscus, R. G. & Murray, A. S.(1996). Direct dating of Florisbad hominid. Nature382, 500–501.
Grun, R., Spooner, N. A., Thorne, A., Mortimer, G.,Simpson, J. J., McCulloch, M. T., Taylor, L. &Curnoe (2000). Age of the Lake Mungo 3 skeleton,reply to Bowler & Magee and to Gillespie & Roberts.J. hum. Evol. 38, 733–742.
Habgood, P. J. (1989). The origin of modern humansin Australia. In (P. Mellars & C. B. Stringer, Eds)The Human Revolution: Behavioral and BiologicalPerspectives on the Origins of Modern Humans,pp. 245–273. Edinburgh: Edinburgh University Press.
Hahn, J. (1993). Aurignacian art in Central Europe. In(H. Knecht, A. Pike-Tay & R. White, Eds) BeforeLascaux: The Complex Record of the Early UpperPaleolithic, pp. 229–243. Boca Raton: CRC Press.
Hajraoui, M. A. (1994). L’Industrie osseuse ateriennede la Grotte d’el Mnasra. Prehist. Anthrop. Med. 3,91–94.
Hall, S. (1999). Scratch the power off. South AfricanCountry Life 35, 42–43.
Hare, P. E. (1980). Organic chemistry of bone and itsrelation to the survival of bone in the natural environ-ment. In (A. K. Behrensmeyer & A. P. Hill, Eds)Fossils in the Making, pp. 208–219. Chicago: Univer-sity of Chicago Press.
Hare, P. E., Goodfriend, G. A., Brooks, A. S., Kokis,J. E. & von Endt, D. W. (1993). Chemical clocks andthermometers: diagenetic reactions of amino acids infossils. Carnegie Inst. Wash. Yearb. 92, 80–85.
Harpending, H., Batzer, M., Gurven, M., Jorde, L.,Ogers, A. & Sherry, S. (1998). Genetic traces ofancient demography. Proc. natl. Acad. Sci. USA 95,1961–1967.
Harpending, H. C., Sherry, S. T., Rogers, A. L. &Stoneking, M. (1993). The genetic structure ofancient human populations. Curr. Anthrop. 34,483–496.
Harper, P. T. (1994). The Middle Stone Age sequenceat Rose Cottage Cave: a search for continuity anddiscontinuity. Ph.D. Dissertation, University of theWitwatersrand, Johannesburg, South Africa.
Harrison, T. (1993). Cladistic concepts and the speciesproblem in hominoid evolution. In (W. H. Kimbel &L. B. Martin, Eds) Species, Species Concepts, andPrimate Evolution, pp. 345–371. New York: Plenum.
Harrold, F. B. (1983). The Chatelperronian and theMiddle Paleolithic transition. In (E. Trinkaus, Ed.)Human Biocultural Change in the Upper Pleistocene,

547 ’
pp. 123–140. Oxford: British Archaeological ReportsInternational Series 164.
Harrold, F. B. (1989). Mousterian, Chatelperronianand early Aurignacian in western Europe: continuityor discontinuity? In (P. Mellars & C. B. Stringer,Eds) The Human Revolution: Behavioral and BiologicalPerspectives on the Origins of Modern Humans,pp. 677–713. Edinburgh: Edinburgh University Press.
Hay, R. L. (1963). Stratigraphy of Beds I through IV,Olduvai Gorge, Tanganyika. Science 139, 829–833.
Hay, R. L. (1971). Geological background of Beds Iand II: stratigraphic summary. In (M. D. Leakey,Ed.) Olduvai Gorge: Excavations in Beds I & II,1960–1963, pp. 1–9. Cambridge: CambridgeUniversity Press.
Hay, R. L. (1973). Lithofacies and environments ofBed I, Olduvai Gorge, Tanzania. Quatern. Res. 3,541–560.
Hay, R. L. (1976). The Geology of Olduvai Gorge.Berkeley: University of California Press.
Hay, R. L. (1987). Geology of the Laetoli area. In(M. D. Leakey & J. M. Harris, Eds) Laetoli, a PlioceneSite in Northern Tanzania. Oxford: Oxford UniversityPress.
Hay, R. L. (1990). Olduvai Gorge: A case history of theinterpretation of hominid paleoenvironments in EastAfrica. In (L. Laporte, Ed.) Establishment of a Geo-logic Framework for Paleoanthropology, pp. 23–37.Boulder: Geological Society of America SpecialPaper 242.
Hay, R. L. (1994). Geology and dating of Beds III, IVand the Masek Beds. In (M. D. Leakey & D. A. Roe,Eds) Olduvai Gorge, Vol. 5: Excavations in BedsIII, IV, and the Masek Beds, 1968–1971, pp. 8–14.Cambridge: Cambridge University Press.
Hayden, B. (1992). Conclusions: Ecology and complexhunter/gatherers. In (B. Hayden, Ed.) A ComplexCulture of the British Columbia Plateau: Stl’atl’imxResource Use, pp. 525–564. Vancouver: University ofBritish Columbia Press.
Hayden, B. (1993). The cultural capacity of Neander-tals: a review and reevaluation. J. hum. Evol. 24,113–146.
Heberer, G. (1963). Ueber einen neuen arch-anthropinen Typus aus der Oldoway-Schlucht. Z.Morph. Anthrop. 53, 171–177.
Hedges, S. B., Kumar, S., Tamura, K. & Stoneking, M.(1992). Human origins and analysis of mitochondrialDNA sequences. Science 255, 737–739.
Helgren, D. M. & Brooks, A. S. (1983). Geoarchaeol-ogy at Gi: a Middle Stone Age and Later Stone Agesite in the northwest Kalahari. J. Archaeol. Sci. 10,181–197.
Henderson, Z. (1992). The context of some MiddleStone Age hearths at Klasies River Shelter 1B: impli-cations for understanding human behavior. S. Afr.Field Archaeol. 1, 14–26.
Hendey, Q. V. & Volman, T. (1986). Last interglacialsea levels and coastal caves in the Cape Province,South Africa. Quatern. Res. 25, 189–198.
Hennig, W. (1965). Phylogenetic systematics. Ann.Rev. of Entomol. 10, 97–116.
Henshilwood, C. & Sealey, J. C. (1997). Bone artefactsfrom the Middle Stone Age at Blombos Cave,Southern Cape, South Africa. Curr. Anthrop. 38,890–895.
Henshilwood, C. & Sealy, J. C. (n.d.). Blombos Cave,South Africa. Unpublished excavation report.Department of Archaeology, University of CapeTown.
Hill, A. P. (1979). Butchery and natural disarticulation:an investigatory technique. Am. Antiq. 44, 739–744.
Hill, A. P. (1987). Causes of perceived faunal change inthe later Neogene of East Africa. J. hum. Evol. 16,583–596.
Hitchcock, R. & Bleed, P. (1997). Each according toneed and fashion: spear and arrow use among Sanhunters of the Kalahari. In (H. Knecht, Ed.) ProjectileTechnology, New York: Plenum.
Hodder, I. (1985). Symbols in Action: Ethnoarchaeo-logical Studies of Material Culture. Cambridge:Cambridge University Press.
Hodder, I. (1987). The meaning of discard: ash anddomestic space in Baringo. In (S. Kent, Ed.) Methodand Theory for Activity Area Research: An Ethno-archaeological Approach, pp. 424–448. New York:Columbia University Press.
Hole, F. (1959). A critical reanalysis of the Magosian.S. Afr. Archaeol. Bull. 14, 126–134.
Holliday, T. W. (1997). Body proportions in LatePleistocene Europe and modern human origins.J. hum. Evol. 32, 423–447.
Holliday, T. W. (1998). Brachial and crural indices ofEuropean late Upper Paleolithic and Mesolithichumans. J. hum. Evol. 36, 549–566.
Holliday, T. W. (2000). Evolution at the crossroads:modern human emergence in western Asia. Am.Anthrop. 102, 54–68.
Holliday, T. W. & Trinkaus, E. (1991). Limb-trunkproportions in Neandertals and early anatomicallymodern humans. Am. J. phys. Anthrop Supplement12, 93–94.
Holloway, R. (1983). Human paleontological evidencerelevant to language behavior. Hum. Neurobiol. 2,105–114.
Holloway, R. (1985). The poor brain of Homo sapiensneanderthalensis: see what you please. In (E. Delson,Ed.) Ancestors: The Hard Evidence, pp. 319–324. NewYork: Alan R. Liss.
Hou, Y. M., Potts, R., Yuan, B. Y., Guo, Z. T., Deino,A., Wang, W., Clark, J., Xie, G. M. & Huang, W. W.(2000). Mid-Pleistocene Acheulian-like stone tech-nology of the Bose basin, South China. Science 287,1622–1626.
Howe, B. (1967). The Palaeolithic of Tangier, Morocco.Excavations at Cape Ashakar, 1939–1947. AmericanSchool of Prehistoric Research Bulletin 22.Cambridge: Peabody Museum, Harvard University.
Howell, F. C. (1951). The place of Neanderthal manin human evolution. Am. J. phys. Anthrop. 9, 379–416.
Howell, F. C. (1952). Pleistocene glacial geology andthe evolution of ‘classic Neanderthal’ man. S. W. J.Anthrop. 8, 377–410.

548 . . .
Howell, F. C. (1960). European and northwest AfricanMiddle Pleistocene hominids. Curr. Anthrop. 1,195–232.
Howell, F. C. (1978). Hominidae. In (V. J. Maglio &H. B. S. Cooke, Eds) Evolution in African Mammals,pp. 154–248. Cambridge: Harvard UniversityPress.
Howell, F. C. (1982). Origins and evolution of AfricanHominidae. In (J. D. Clark, Ed.) Cambridge History ofAfrica, Vol. 1: From the Earliest Times to 500 B.C.,pp. 70–156. Cambridge: Cambridge UniversityPress.
Howell, F. C. (1994). A chronostratigraphic and taxo-nomic framework of the origins of modern humans.In (M. H. Nitecki & D. V. Nitecki, Eds) Origins ofAnatomically Modern Humans, pp. 253–319. NewYork: Plenum.
Howell, F. C. (1998). Evolutionary implications ofaltered perspectives on hominine demes and popu-lations in the later Pleistocene of western Eurasia.In (T. Akazawa, K. Aoki & O. Bar-Yosef, Eds)Neandertals and Modern Humans in Western Asia,pp. 5–28. New York: Plenum.
Howell, F. C., Cole, G. H. & Kleindienst, M. R.(1962). Isimila, an Acheulian occupation site in theIringa highlands. In (J. Mortelmans & J. Nenquin,Eds) Actes Du IVe Congres Panafricain de Prehistoire etde l’Eutude du Quaternaire, pp. 209–217. Vol. 40.Tervuren: Musee Royale de l’Afrique Centrale.
Howell, F. C., Cole, G. H., Kleindienst, M. R., Szabo,B. J. & Oakley, K. P. (1972). Uranium series datingof bone from the Isimila prehistoric site, Tanzania.Nature 237, 51–52.
Howells, W. W. (1976). Explaining modern man: evo-lutionists vs. migrationists. J. hum. Evol. 5, 447–449.
Howells, W. W. (1989). Skull Shapes and the Map:Craniometric Analyses in the Dispersion of ModernHomo. Cambridge: Papers of the Peabody Museumof Archaeology and Ethnology, Harvard University,vol. 79.
Howells, W. W. (1993). Getting Here: The Story ofHuman Evolution. Washington, D.C.: CompassPress.
Hu, Q., Smith, P. E., Evensen, N. & York, D. (1994).Lasing into the Holocene: extending the 40Ar/39Arlaser probe method into the 14C time range. EarthPlanet. Sci. Lett. 123, 331–336.
Hublin, J.-J. (1985). Human fossils from the NorthAfrican Middle Pleistocene and the origin of Homosapiens. In (E. Delson, Ed.) Ancestors: The HardEvidence, pp. 283–285. New York: Alan R. Liss.
Hublin, J.-J. (1990). Les peuplements paleolithiques del’Europe: un point de vue paleobiogeographique. In(C. Farizy, Ed.) Paleolithique Moyen Recent et Paleo-lithique Superieur Ancien en Europe, pp. 29–37.Nemours: A.P.R.A.I.F.
Hublin, J.-J. (1991). L’Emergence des Homo sapiensarchaiques: Afrique du Nord-Ouest et Euorpeoccidentale. These d’Etat, Universite de Bordeaux 1.
Hublin, J.-J. (1992). Le femur humain pleistocenemoyen de l’Aın Maarouf (El Hajeb, Maroc). C. r.Acad. Sci., Paris 314, 975–980.
Hublin, J.-J. (1993). Recent human evolution in north-west Africa. In (M. J. Aitken, C. B. Stringer & P. A.Mellars, Eds) The Origin of Modern Humans andthe Impact of Chronometric Dating, pp. 118–131.Princeton: Princeton University Press.
Hublin, J.-J. (1994). Pathological aspects of the middlePleistocene skull of Sale (Morocco). Am. J. phys.Anthrop. Supplement 18, 110.
Hublin, J.-J. (1998a). Climate de l’Europe et originedes Neandertaliens. Pour La Science 245, 52–59.
Hublin, J.-J. (1998b). Climatic changes, paleogeogra-phy, and the evolution of the Neandertals. In (T.Akazawa, K. Aoki & O. Bar-Yosef, Eds) Neandertalsand Modern Humans in Western Asia, pp. 295–310.New York: Plenum.
Hublin, J.-J. (1998c). Comments to d’Errico et al.,Neanderthal acculturation in western Europe? Curr.Anthrop. A39, S24–S25.
Hublin, J.-J. (1999). Derniers Neandertaliens et pre-miers Europeens modernes. Pour La Science 255,110–118.
Hublin, J.-J., Tillier, A.-M. & Tixier, J. (1987).L’humerus d’enfant mousterien (Homo 4) du JebelIrhoud (Maroc) dans son contexte archeologique.Bull. Mem. Soc. Anthropol. Paris 14, 115–142.
Hublin, J.-J., Spoor, F., Braun, M., Zonneveld, F. &Condemi, S. (1996). A late Neanderthal associ-ated with Upper Paleolithic artifacts. Nature 381,224–226.
Imbrie, J. & Imbrie, J. Z. (1980). Modeling the climaticresponse to orbital variations. Science 207, 943–953.
Inskeep, R. (1962). The age of the Kondoa rockpainting in the light of recent excavations at Kisese IIrock shelter. In (G. Mortelmans & J. Nenquin, Eds)Actes du IVe Congres Panafricain de Prehistoire etde l’Etude du Quaternaire (Leopoldville, 1959),pp. 249–256. Vol. 2. Tervuren: Musee Royale del’Afrique Centrale. Annales Serie 8, SciencesHumaines no. 40.
Irish, J. (1998). Ancestral dental traits in recent Sub-Saharan Africans and the origins of modern humans.J. hum. Evol. 34, 81–98.
Isaac, G. (1977). Olorgesailie: Archaeological Studies of aMiddle Pleistocene Lake Basin in Kenya. Chicago:University of Chicago Press.
Jacobs, K. H. (1985). Climate and the hominid post-cranial skeleton in Wurm and early HoloceneEurope. Curr. Anthrop. 26, 512–514.
Jaeger, J.-J. (1973). Un pithecanthrope evolue. LaRecherche 39, 1006–1007.
Jaeger, J.-J. (1975). The mammalian fauna and homi-nid fossils of the Middle Pleistocene in the Maghreb.In (K. W. Butzer & G. Ll. Isaac, Eds) Afterthe Australopithecines, pp. 399–410. The Hague:Mouton.
Jaeger, J.-J. (1981). Les hommes fossiles du PleistoceneMoyen du Maghreb dans leur cadre geologique,chronologique and paleoecologique. In (B. A.Sigmon & J. S. Cybulski, Eds) Homo Erectus: Papersin Honor of Davidson Black, pp. 159–264. Toronto:University of Toronto Press.

549 ’
Jelinek, A. (1982). The Tabun Cave and PaleolithicMan in the Levant. Science 216, 1369–1375.
Jelinek, A. (1990). The Amudian in the context of theMugharan traditions at the Tabun Cave (MountCarmel), Israel. In (P. Mellars, Ed.) The Emergenceof Modern Humans: An Archaeological Perspective,pp. 81–90. Ithaca: Cornell University Press.
Jochim, M. (1987). Late Pleistocene refugia in Europe.In (O. Soffer, Ed.) The Pleistocene Old World,pp. 317–331. New York: Plenum.
Jolly, K. (1948). The development of the Cape MiddleStone Age in the Skildergat Cave, Fish Hoek. S. Afr.Archaeol. Bull. 3, 106–107.
Jones, R. (1992). The human colonization of theAustralian continent. In (G. Brauer & F. H. Smith,Eds) Continuity or Replacement: Controversies in Homosapiens Evolution, pp. 289–301. Rotterdam: A. A.Balkema.
Jones, T. R. (1940). Human skeletal remains from theMumbwa Cave, Northern Rhodesia. S. Afr. J. Sci.37, 313–319.
Jorde, L. B., Bamshad, M. & Rogers, A. R. (1998).Using mitochondrial and nuclear DNA markers toreconstruct human evolution. Bioessays 20, 126–136.
Joris, O. & Weninger, B. (1998). Extension of the 14Ccalibration curve to ca. 40,000 cal BC by synchroniz-ing Greenland 18O/16O ice core records and NorthAtlantic foraminifera profiles: a comparison withU/Th coral data. Radiocarbon 40, 495–504.
Kalb, J. E., Wood, C. B., Smart, C., Oswald, E. B.,Mebrate, A., Tebedge, S. & Whitehead, P. F.(1980). Preliminary geology and paleontology of theBodo D’Ar hominid site, Afar, Ethiopia. Palaeo-geogr., Palaeoclimatol., Palaeoecol. 30, 107–120.
Kalb, J. E., Jaeger, M., Jolly, C. P. & Kana, B. (1982a).Preliminary geology, paleontology, and paleoecologyof a Sangoan site at Andalee, Middle Awash Valley,Ethiopia. J. Archaeol. Sci. 9, 349–363.
Kalb, J. E., Oswald, E. B., Tebedge, S., Mebrate, A.,Tola, E. & Peak, D. (1982b). Geology and stratigra-phy of Neogene deposits, Middle Awash Valley,Ethiopia. Nature 298, 17–25.
Kappelman, J. (1997). They might be giants. Nature387, 126–127.
Kay, R. F., Cartmill, M. & Barlow, M. (1998). Thehypoglossal canal and the origin of human vocalbehavior. Proc. natl. Acad. Sci. USA 95, 5417–5419.
Keith, A. (1931). New Discoveries Relating to theAntiquity of Man. London: Williams & Norgate.
Keller, C. M. (1973). Montagu Cave in Prehistory:A Descriptive Analysis. Berkeley: University ofCalifornia Anthropological Records Vol. 28.
Kendrick, T. D. (1925). The Axe Age. London:Methuen.
Kennedy, G. F. (1984). The emergence of Homosapiens: the postcranial evidence. Man 19, 94–119.
Kimbel, W. H. (1991). Species, species concepts andhominid evolution. J. hum. Evol. 20, 355–371.
Kimbel, W. H. (1995). Hominid speciation andPliocene climate change. In (E. S. Vrba, G. H.Denton, T. C. Patridge & L. H. Burckle, Eds)Paleoclimate and Evolution With Emphasis on Human
Origins, pp. 425–437. New Haven: Yale UniversityPress.
Klein, R. G. (1971). Problems in the study of theMiddle Stone Age of South Africa. S. Afr. Archaeol.Bull. 25, 127–135.
Klein, R. G. (1973). Geological antiquity of RhodesianMan. Nature 244, 311–312.
Klein, R. G. (1976). The mammalian fauna of theKlasies River Mouth sites, Southern Cape Province,South Africa. S. Afr. Archaeol. Bull. 31, 75–98.
Klein, R. G. (1978). Preliminary analysis of themammalian fauna from the Redcliff Stone Age cavesite, Rhodesia. National Museum of Southern RhodesiaOccasional Papers, Series A, Human Sciences 4,74–80.
Klein, R. G. (1988). The archaeological significance ofanimal bones from Acheulian sites in southernAfrica. Afr. Archaeol. Rev. 6, 3–26.
Klein, R. G. (1989a). Biological and behavioralperspectives on modern human origins in southernAfrica. In (P. Mellars & C. B. Stringer, Eds) TheHuman Revolution: Behavioral and Biological Perspec-tives on the Origins of Modern Humans, pp. 529–546.Edinburgh: Edinburgh University Press.
Klein, R. G. (1989b). The Human Career. Chicago:University of Chicago Press.
Klein, R. G. (1992). The archaeology of modernhuman origins. Evol. Anthrop. 1, 5–14.
Klein, R. G. (1994). Southern Africa before the IronAge. In (R. S. Corrucini & R. L. Ciochon, Eds)Integrative Pathways to the Past: Palaeoanthropo-logical Investigations in Honor of F. Clark Howell,pp. 471–520. Englewood Cliffs, NJ: Prentice Hall.
Klein, R. G. (1995). Anatomy, behavior, and modernhuman origins. J. World. Prehist. 9, 167–198.
Klein, R. G. (1995–1996). Neanderthals and modernhumans in West Asia: a conference summary. Evol.Anthrop. 4, 187–193.
Klein, R. G. (1998). Why anatomically modern peopledid not disperse from Africa 100,000 years ago.In (T. Akazawa, K. Aoki & O. Bar-Yosef, Eds)Neandertals and Modern Humans in Western Asia,pp. 509–522. New York: Plenum.
Klein, R. G. (1999). The Human Career. 2nd edn.Chicago: Chicago University Press.
Klein, R. G. (2000). Archaeology and the evolution ofhuman behavior. Evol. Anthrop. 9, 17–36.
Klein, R. G. & Cruz-Uribe, K. (1991). The bovidsfrom Elandsfontein, South Africa, and their impli-cations for the age, paleoenvironments and origins ofthe site. Afr. Archaeol. Rev. 9, 21–79.
Klein, R. G. & Cruz-Uribe, K. (1996). Exploitation oflarge bovids and seals at Middle and Later Stone Agesites in South Africa. J. hum. Evol. 31, 315–334.
Klein, R. G. & Cruz-Uribe, K. (2000). Middle andLater Stone Age large mammal and tortoise remainsfrom Die Kelders Cave 1, Western Cape Province,South Africa. J. hum. Evol. 38, 169–196.
Klein, R. G. & Scott, K. (1986). Re-analysis of faunalassemblages form the Haua Fteah and other LateQuaternary archaeological sites in Cyrenaican Libya.J. Archaeol. Sci. 13, 515–542.

550 . . .
Klein, R. G., Cruz-Uribe, K. & Beaumont, P. B.(1991). Environmental, ecological and paleoanthro-pological implications of the Late Pleistocenemammalian fauna from Equus Cave, northern CapeProvince, South Africa. Quatern. Res. 36, 94–119.
Klein, R. G., Cruz-Uribe, K. & Milo, R. G. (1999).Skeletal part representation in archaeofaunas:Comments on ‘‘Explaining the ‘Klasies pattern’: Kuaethnoarchaeology, the Die Kelders Middle StoneAge archaeofauna, long bone fragmentation andcarnivore ravaging’’ by Bartram & Marean.J. Archaeol. Sci. 26, 1225–1234.
Kleindienst, M. R. (1967). Questions of terminology inregard to the study of stone age industries in EasternAfrica: Cultural stratigraphic units. In (W. W. Bishop& J. D. Clark, Eds) Background to Evolution in Africa,pp. 821–859. Chicago: University of Chicago Press.
Kleindienst, M. R. (2000a). On the Nile corridor andthe Out-of Africa model. Curr. Anthrop. 41, 107–109.
Kleindienst, M. R. (2000b). What is the Aterian? Theview from Dakhleh Oasis and the Western Desert,Egypt. In (M. Marlowe, Ed.) Proceeding of the FirstDakleh Oasis Project Seminar, pp. 1–51. Oxford:Oxbow Press.
Kleindienst, M. R. & Wiseman, M. F. (1996). ThePleistocene archaeology and geoarchaeology at theDakleh oasis, Western Desert, Egypt: 19 seasons ofexploration. Nyame Akuma 46, 96.
Kleindienst, M. R., Schwarcz, H. P., Nicoll, K.,Churcher, C. S., Frizano, J., Giegangack, R. W. &Wiseman, M. F. (1996). Pleistocene geochronologyand paleoclimates at Dakleh Oasis and Kharga Oasis,Western Desert, Egypt, based upon uranium-thorium determinations from water-laid tufas. NyameAkuma 46, 96.
Knecht, H. (1993). Early Upper Palaeolithicapproaches to bone and antler projectile technology.In (G. L. Petersen, H. M. Bricker & P. Mellars,Eds) Hunting and Animal Exploitation in the LaterPalaeolithic and Mesolithic of Eurasia, pp. 33–47.Washington, D.C.: Archaeological Papers of theAmerican Anthropological Association.
Knight, C. (1991). Blood Relations: Menstruation andthe Origin of Culture. New Haven: Yale UniversityPress.
Knight, C. D., Power, C. & Watts, I. (1995). Thehuman symbolic revolution: a Darwinian account.Cambridge Archaeol. J. 5, 75–114.
Kohary, J. (1989). Archaeology and Ecology of RoseCottage Cave, Ladybrand, Orange Free State.Johannesburg: Broederstroom Press.
Kokis, J. E. (1988). Protein diagenesis dating of ostrich(Struthio camelus) eggshell: an Upper Pleistocenedating technique. M.A. Thesis, George WashingtonUniversity.
Kokis, J. E., Hare, P. E. & Brooks, A. S. (1998).Stratigraphic analysis of White Paintings Selter,Botswana, using isoleucine epimerization in ratiteeggshell. Amino Acids 15, 280–281.
Koslowski, J. K. (1988). Transition from the Middle tothe early Upper Paleolithic in central Europe and theBalkans. In (J. F. Hoffeker & C. A. Wolf, Eds) The
Early Upper Paleolithic: Evidence From Europe andthe Near East, pp. 529–546. Oxford: BritishArchaeological Reports International Series 437.
Kraybill, N. (1977). Pre-agricultural tools for theprepartion of foods in the Old World. In (C. A. Reed,Ed.) Origins of Agriculture, 485–521. The Hague:Mouton.
Kretzoi, M. (1984). Note on Homo leakeyi Heberer.Anthropol. Kozl. 28, 189–190.
Krings, M., Stone, A., Schmitz, R. W., Krainitzki, H.,Stoneking, M. & Paabo, S. (1997). NeanderthalDNA sequences and the origin of modern humans.Cell 90, 19–30.
Krings, M., Geisert, H., Schmitz, R. W., Krainitzki, H.& Paabo, S. (1999). DNA sequences of themitochondrial hypervariable region II from theNeanderthal type specimen. Proc. natl. Acad. Sci.USA 96, 5581–5585.
Krupsha, C. (1993). An experimental study ofharpoons from Katanda and Ishango. Unpublishedsenior honors thesis, George Washington University.
Kuhn, S. L. (1991). ‘Unpacking’ reduction: lithic rawmaterial economy in the Mousterian of West-CentralItaly. J. Anthrop. Archaeol. 10, 76–106.
Kuhn, S. L. (1992). On planning and curated tech-nologies in the Middle Palaeolithic. J. Anthrop. Res.48, 185–214.
Kuman, K. & Clarke, R. J. (1986). Florisbad: newinvestigations at a Middle Stone Age hominid site inSouth Africa. Geoarchaeol. 1, 103–125.
Kuman, K., Inbar, M. & Clarke, R. J. (2000).Palaeoenvironments and cultural sequence of theFlorisbad Middle Stone Age hominid site, SouthAfrica. J. Archaeol. Sci. 26, 1409–1426.
Kuman, K. A. (1989). Florisbad and �Gi: the contri-bution of open-air sites to the study of the MiddleStone Age in southern Africa. Ph.D. Dissertation,University of Pennsylvania.
Lahr, M. (1996). The Evolution of Modern HumanDiversity: A Study of Cranial Variation. Cambridge:Cambridge University Press.
Lahr, M. M. & Foley, R. A. (1994). Multiple dispersalsand modern human origins. Evol. Anthrop. 3, 48–60.
Lahr, M. M. & Foley, R. A. (1998). Towards a theoryof modern human origins: geography, demographyand diversity on recent human evolution. Ann. Rev.Anthrop. 27, 137–176.
Lam, U. M., Pearson, O. M. & Smith, C. M. (1996).Chin morphology and sexual dimorphism in thefossil hominid mandible sample from Klasies RiverMouth. Am. J. phys. Anthrop. 100, 545–557.
Landau, M. (1991). Narratives of Human Evolution.New Haven, CT: Yale University Press.
Larick, R. & Ciochon, R. L. (1996). The Africanemergence and early Asian dispersals of the genusHomo. Am. Scient. 84, 538–551.
Larsson, L. (1996). The Middle Stone Age ofZimbabwe: some aspects of former research andfuture aims. In (G. Pwiti & R. Soper, Eds) Aspects ofAfrican Archaeology, pp. 201–211. Harare: Universityof Zimbabwe Publications.

551 ’
Lartet, E. & Christy, H. (1865–1875). ReliquiaeAquitanicae. London: Williams & Norgate.
Leakey, L. S. B. (1931). The Stone Age Cultures of KenyaColony. Cambridge: University of Cambridge.
Leakey, L. S. B. (1961). New finds at Olduvai Gorge.Nature 189, 649–650.
Leakey, L. S. B. (1963). East African Hominoidea andthe classifications within the super-family. In (S. L.Washburn, Ed.) Classification and Human Evolution,pp. 32–49. London: Methuen.
Leakey, L. S. B., Tobias, P. V. & Napier, J. R. (1964).A new species of the genus Homo from OlduvaiGorge. Nature 202, 7–9.
Leakey, M., Tobias, P. V., Martyn, J. E. & Leakey,R. E. F. (1969). An Acheulian industry with pre-pared core technique and the discovery of a contem-porary hominid mandible at Lake Baringo, Kenya.Proc. prehist. Soc. 3, 48–76.
Leakey, M. D. (1971a). Discovery of postcranialremains of Homo erectus and associated artifacts inBed IV at Olduvai Gorge, Tanzania. Nature 232,380–383.
Leakey, M. D. (1971b). Olduvai Gorge, Volume 3:Excavations in Beds I and II, 1960–1963. Cambridge:Cambridge University Press.
Leakey, M. D. & Hay, R. L. (1982). The chronologicalposition of the fossil hominids of Tanzania. In 1er
Congres International de Paleontologie Humaine, Nice,pp. 753–765. Vol. 2. Colloque International duCNRS-UNESCO: Pretirages.
Leakey, M. D. & Roe, D. A. (Eds) (1994). OlduvaiGorge, Vol. 5: Excavations in Beds III, IV, and theMasek Beds, 1968–1971. Cambridge: CambridgeUniversity Press.
Leakey, M. D., Hay, R. L., Thurber, D. L., Protsch, R.& Berger, R. (1972). Stratigraphy, archaeology, andage of the Ndutu and Naisusu Beds, Olduvai Gorge,Tanzania. World Archaeol. 3, 328–341.
Lee, R. B. (1979). The !Kung San: Men, Women andWork in a Foraging Society. Cambridge: CambridgeUniversity Press.
Lewis-Williams, J. D. (1982). The economic and socialcontext of southern San rock art. Curr. Anthrop. 23,429–449.
Lewis-Williams, J. D. (1983). Rock Art of SouthernAfrica. Cambridge: Cambridge University Press.
Lieberman, D. E. (1993). The rise and fall of seasonalmobility: the case of the southern Levant. Curr.Anthrop. 34, 599–631.
Lieberman, D. E. (1998a). Neandertal and earlymodern mobility patterns: comparing archaeologicaland anatomical evidence. In (T. Akazawa, K. Aoki &O. Bar-Yosef, Eds) Neandertals and Modern Humansin Western Asia, pp. 263–276. New York: Plenum.
Lieberman, D. E. (1998b). Sphenoid shortening andthe evolution of modern human cranial shape. Nature393, 158–162.
Lieberman, D. E. & Shea, J. J. (1994). Behavioraldifferences between archaic and modern humansin the Levantine Mousterian. Am. Anthrop. 96,300–332.
Lieberman, P. (1975). On the Origins of Language.New York: MacMillan.
Lieberman, P. (1984). The Biology and Evolution ofLanguage. Cambridge: Harvard University Press.
Lieberman, P., Laitman, J. T., Reidenberg, J. S. &Gannon, P. J. (1992). The anatomy, physiology,acoustics and perception of speech. J. hum. Evol. 23,447–467.
Lyell, C. (1868). The Geological Evidence of the Antiquityof Man, With Remarks on the Origin of Species.London: Spottiswoode.
MacCalman, H. R. & Viereck, A. (1967). Peperkorrel:a factory site of Lupemban affinities from centralSouth West Africa. S. Afr. Archaeol. Bull. 22, 41–50.
Magori, C. & Day, M. (1983). Laetoli hominid 18: anearly Homo sapiens skull. J. hum. Evol. 12, 747–753.
Malan, B. D. (1955). The archaeology of the TunnelCave and Skildergat Kop, Fish Hoek, Cape of GoodHope. S. Afr. Archaeol. Bull. 10, 3–9.
Manega, P. C. (1993). Geochronology, geochemistry,and isotopic study of the Plio-Pleistocene hominidsites and the Ngorongoro volcanic highland innorthern Tanzania. Ph.D. Dissertation, Universityof Colorado, Boulder.
Manega, P. C. (1995). New geochronological resultsfrom the Ndutu, Naisusu, and Ngaloba Beds atOlduvai and Laetoli in northern Tanzania: theirsignificance for the evolution of modern humans.(Abstract). Preservation and Use of Olduvai Gorge,Laetoli, Rock Art, and other PalaeoanthropologicalResources in Tanzania. Bellagio, Italy.
Marean, C. W. (1992). Implications of LateQuaternary mammalian fauna from Lukenya Hill(south-central Kenya) for paleonenvironmentalchange and faunal extinctions. Quatern. Res. 37,239–255.
Marean, C. W. (1997). Hunter-gatherer foragingstrategies in tropical grasslands: Model-building andtesting in the East African Middle and Later StoneAge. J. Anthrop. Archaeol. 16, 189–225.
Marean, C. W. (1998). A critique of the evidence forscavenging by Neanderthals and early modernhumans: New data from Kobeh Cave (ZagrosMountains, Iran) and Die Kelders Cave 1 Layer 10(South Africa). J. hum. Evol. 35, 111–136.
Marean, C. W. & Assefa, Z. (1999). Zooarchaeologicalevidence for the faunal exploitation behavior ofNeandertals and early modern humans. Evol.Anthrop. 8, 22–37.
Marean, C. W. & Kim, S. Y. (1998). Mousterianlarge-mammal remains from Kobeh Cave: behavioralimplications for Neanderthals and early modernhumans. Curr. Anthrop. 39, S79–S114.
Marean, C. W., Abe, Y., Frey, C. J. & Randall, R. C.(2000). Zooarchaeological and taphonomic analysisof the Die Kelders Cave 1 Layers 10 and 11 MiddleStone Age larger mammal fauna. J. hum. Evol. 38,197–233.
Marks, A. E. (1968). The Mousterian industries ofNubia. In (F. Wendorf, Ed.) The Prehistory of Nubia,Vol. 1, pp. 194–314. Dallas: Southern MethodistUniversity Press.

552 . . .
Marks, A. E. (1975). The current status of UpperPalaeolithic studies from the Maghreb to thenorthern Levant. In (F. Wendorf & A. E. Marks,Eds) Problems in Prehistory: North Africa and theLevant, pp. 439–458. Dallas: Southern MethodistUniversity Press.
Marks, A. E. & Brugal, J.-P. (2000). Excavations at thelate Middle Pleistocene cave of Galeria Pesada: 1997/1999. J. hum. Evol. 38, A18–A19.
Marks, A. E. & Chabai, V. P. (Eds) (1996). ThePaleolithic of Crimea. I. The Middle Paleolithic ofWestern Crimea, Vol. 1. Liege: Etudes et RecherchesArcheologiques de l’Universite de Liege 84.
Marshack, A. (1981). Paleolithic ochre and theearly use of color and symbol. Curr. Anthrop. 22,188–191.
Marshack, A. (1989). Evolution of the human capacity:the symbolic evidence. Yearb. phys. Anthrop. 32, 1–34.
Marshall, C. R. (1990). Confidence intervals onstratigraphic ranges. Paleobiology 16, 1–10.
Martini, M., Sibilia, E., Zelaschi, C., Troja, S. O.,Forzese, R., Caputa, C., Gueli, A. M., Cro, A., Foti,F. & Pellegriti, M. G. (1996). TL and OSL datingof fossil dune sand in the Uan Afuda and UanTabu sites (Tadrart Acacus, Libyan Sahara). In (M.Cremaschi & S. Di Lerna, Eds) Wadi Teshuinat.Palaeoenvironment and Prehistory in South-WesternFezzan (Libya), pp. 67–72. Milano: CNR.
Martinson, D. G., Pisias, N. H., Hays, H. D., Imbrie,J., Moore, T. C. & Shackleton, N. J. (1987). Age,dating, and the orbital theory of the ice ages:development of a high-resolution 0 to 300,000-yearchronostratigraphy. Quatern. Res. 27, 1–29.
Mason, R. J. (1962). Prehistory of Gauteng.Johannesburg: Witswatersrand University Press.
Mason, R. J. (1993). La Grotte de Hearths, Valleede Makapansgat, Transvaal, Afrique du Sud,1937–1988. L’Anthropologie 97, 85–96.
Mason, R. J., Brain, C. K., Brown, A. J. V., Cooke,H. B. S., Deacon, H. J., Ewer, R. F., de Graaff, B.,Hofmeyr, I., Malan, B. D., Tobias, P. V., du Toit,A. P., Turton, K. & Wells, L. H. (1988). Cave ofHearths, Makapansgat, Transvaal. Johannesburg:University of the Witswatersrand ArchaeologicalResearch Unit, Occasional Papers 21.
Mayr, E. (1963). Animal Species and Evolution.Cambridge: Harvard University Press.
Mazaud, A., Laj, C., Bard, E., Arnold, M. & Tric, E.(1991). Geomagentic field control of 14C productionover the last 80 ky: implications for the radiocarbontime-scale. Geophys. Res. Lett. 18, 1885–1888.
McBrearty, S. (1981). Songhor: a Middle Stone AgeSite in western Kenya. Quaternaria 23, 171–190.
McBrearty, S. (1986). The archaeology of theMuguruk Site, western Kenya. Ph.D. Dissertation,University of Illinois, Urbana.
McBrearty, S. (1987). Une evaluation du Sangoen: sonage, son environnement et son rapport avec l’originede l’Homo sapiens. L’Anthropologie 91, 127–140.
McBrearty, S. (1988). The Sangoan–Lupemban andMiddle Stone Age sequence at the Muguruk site,western Kenya. World Archaeol. 19, 379–420.
McBrearty, S. (1990a). Consider the humble termite:termites as agents of postdepositional disturbance atAfrican archaeological sites. J. Archaeol. Sci. 17,111–143.
McBrearty, S. (1990b). The origin of modern humans.Man 25, 129–143.
McBrearty, S. (1991). Recent research in westernKenya and its implications for the status of theSangoan industry. In (J. D. Clark, Ed.) CulturalBeginnings: Approaches to Understanding EarlyHominid Lifeways in the African Savanna, pp. 159–176. Bonn: Romisch-Germanisches Zentralmuseum,Forschunginstitut fur Vor- und Fruhgeschichte,Monographien 19.
McBrearty, S. (1993a). Reconstructing environ-mental conditions surrounding the appearance ofmodern Homo sapiens in Africa. In (S. Abonyi, Ed.)Culture and Environment: A Fragile Coexistence,pp. 145–154. University of Calgary: ArchaeologicalAssociation.
McBrearty, S. (1993b). Sangoan technology andhabitat at Simbi, Kenya. Nyame Akuma 38, 34–39.
McBrearty, S. (1999). Archaeology of the KapthurinFormation. In (P. Andrews & P. Banham, Eds) LateCenozoic Environments and Hominid Evolution: a Trib-ute to Bill Bishop, pp. 143–156. London: GeologicalSociety.
McBrearty, S. (2000). Identifying the Acheulianto Middle Stone Age transition in the KapthurinFormation, Baringo, Kenya. J. hum. Evol. 38,A20.
McBrearty, S. & Texier, P.-J. (n.d.). Evidence forlaminar lithic technology at an early date from theKapthurin Formation, Kenya (in preparation).
McBrearty, S., Bishop, L. C. & Kingston, J. D. (1996).Variability in traces of Middle Pleistocene hominidbehavior in the Kapthurin Formation, Baringo,Kenya. J. hum. Evol. 30, 563–580.
McBrearty, S., Brown, B., Deino, A., Kingston, J. D. &Ward, S. (1999). Anatomy, context, age, and affini-ties of hominids from the Kapthurin Formation,Baringo, Kenya. J. hum. Evol. 36, A12.
McBurney, C. B. M. (1961). Absolute age ofPleistocene and Holocene deposits in the HauaFteah. Nature 192, 685–686.
McBurney, C. B. M. (1967). The Haua Fteah(Cyrenaica) and the Stone Age of the South-EastMediterranean. Cambridge: Cambridge UniversityPress.
McDermott, F., Stringer, C., Grun, R., Williams,C. T., Din, V. & Hawkesworth, C. (1996). NewLate Pleistocene uranium-thorium and ESR datesfor the Singa hominid (Sudan). J. hum. Evol. 31,507–516.
McDougall, I. (1981). 40Ar/39Ar age spectra fromthe KBS tuff, Koobi Fora Formation. Nature 294,120–124.
McHugh, W. P., Breed, C. S., Schaber, G. G.,McCauley, J. F. & Szabo, B. J. (1988). Acheuliansites along the Radar Rivers, southern EgyptianSahara. J. Field Archaeol. 15, 361–379.

553 ’
Mehlman, M. J. (1977). Excavations at Nasera Rock,Tanzania. Azania 12, 111–118.
Mehlman, M. J. (1979). Mumba-Hohle revisited: Therelevance of a forgotten excavation to some currentissues in East African prehistory. World Archaeol. 11,80–94.
Mehlman, M. J. (1984). Archaic Homo sapiens at LakeEyasi, Tanzania: recent misrepresentations. J. hum.Evol. 13, 487–501.
Mehlman, M. J. (1987). Provenience, age and associ-ations of archaic Homo sapiens crania from LakeEyasi, Tanzania. J. Archaeol. Sci. 14, 133–162.
Mehlman, M. J. (1989). Late Quaternary archaeo-logical sequences in northern Tanzania. Ph.D.Dissertation, University of Illinois, Urbana.
Mehlman, M. J. (1991). Context for the emergence ofmodern man in Eastern Africa: some new Tanzanianevidence. In (J. D. Clark, Ed.) Cultural Beginnings:Approaches to Understanding Early Hominid Lifeways inthe African Savanna, pp. 177–196. Bonn: Forschung-institut fur Vor- und Fruhgeschichte, Romisch-Germanisches Zentralmuseum, Monographien 19.
Mellars, P. A. (1986). A new chronology for the FrenchMousterian. Nature 322, 410–411.
Mellars, P. A. (1989a). Major issues in the origin ofmodern humans. Curr. Anthrop. 30, 349–385.
Mellars, P. A. (1989b). Technological changes at theMiddle-Upper Palaeolithic transition: Economic,social, and cognitive perspectives. In (P. Mellars &C. B. Stringer, Eds) The Human Revolution:Behavioral and Biological Perspectives on the Originsof Modern Humans, pp. 338–365. Edinburgh:Edinburgh University Press.
Mellars, P. A. (1991). Cognitive changes and theemergence of modern humans in Europe. CambridgeArchaeol. J. 1, 763–776.
Mellars, P. A. (1993). Archaeology and the population-dispersal hypothesis of modern human origins inEurope. In (M. J. Aitken, C. B. Stringer & P. A.Mellars, Eds) The Origin of Modern Humans andthe Impact of Chronometric Dating, pp. 196–216.Princeton: Princeton University Press.
Mellars, P. A. (1995). Symbolism, language, and theNeanderthal mind. In (P. Mellars & K. Gibson, Eds)Modelling the Human Mind, pp. 15–32. Cambridge:MacDonald Institute for Archaeological Research.
Mellars, P. A. (1996). The Neanderthal Legacy: AnArchaeological Perspective From Western Europe.Princeton: Princeton University Press.
Mellars, P. A. (1998a). Comment on: Neanderthalacculturation in Europe? A critical review byD’Errico et al. Curr. Anthrop. 39, S25–S26.
Mellars, P. A. (1998b). The fate of the Neanderthals.Nature 395, 539–540.
Mellars, P. A. (1998c). The impact of climatic changeson the demography of late Neandertal and earlyanatomically modern populations in Europe. In (T.Akazawa, K. Aoki & O. Bar-Yosef, Eds) Neandertalsand Modern Humans in Western Asia, pp. 493–508.New York: Plenum.
Mellars, P. A. (1999). The Neanderthal problemcontinued. Curr. Anthrop. 40, 341–350.
Mellars, P. A. & Gibson, K. (Eds) (1996). Modeling theEarly Human Mind. Oxford: McDonald InstituteMonographs & Oxbow Press.
Mellars, P. A. & Stringer, C. B. (Eds) (1989). TheHuman Revolution: Behavioral and Biological Perspec-tives on the Origins of Modern Humans. Edinburgh:Edinburgh University Press.
Mellars, P. A., Zhou, L. P. & Marseglia, E. A. (1997).Compositional inhomogeneity of sediments and itspotential effects in dose rate estimation for electronspin resonance dating of tooth enamel. Archaeometry39, 169–176.
Menghin, O. (1931). Weltgeschichte Der Steinzeit.Vienna: A. Schroll.
Mercader, J. & Marti, R. (1999a). Middle Stone Agesite in the tropical forest of Equatorial Guinea.Nyame Akuma 51, 14–24.
Mercader, J., & Marti, R. (1999b). Archaeology in thetropical forest of Banyang-Mbo, SW Cameroon.Nyame Akuma 52, 17–24.
Mercader, J. & Marti, R. (2000). The Middle StoneAge occupation of Atlantic central Africa: new evi-dence from Equatorial Guinea and Cameroon. In (J.Mercader, Ed.) Stone Age Settlement of Tropical For-ests. Cambridge: Cambridge University Press(under review).
Mercader, J., Brooks, A., Piperno, D., Marti, R. &Holst, I. (2000a). Ancient foragers as early colonizersof tropical forests. J. hum. Evol. 38, A21–A22.
Mercader, J., Piperno, D., Brooks, A. S., Marti, R. &Holst, I. (2000b). Middle Stone Age sites in theAfrican rainforest: new data from Equatorial Guinea.Abstracts of the 65th Annual Meeting of the Societyfor American Archaeology, Philadelphia, April 5–9,226.
Mercader, J., Runge, F., Vrydaghs, L., Doutrelepont,H., Ewango, C. & Juan, J. (2000c). Phytoliths fromarchaeological sites in the tropical forest of Ituri,Democratic Republic of Congo. Quatern. Res. 54,102–112.
Mercier, N., Valladas, H., Bar-Yosef, O.,Vandermeersch, B., Stringer, C. & Joron, J. L.(1993). Thermoluminescence dates from theMousterian burial site of es-Skhul, Mt Carmel.J. Archaeol. Sci. 20, 169–174.
Mercier, N., Valladas, H., Valladas, G. & Reyss, J.-L.(1995). TL dates of burnt flints from Jelinek’s exca-vations at Tabun and their implications. J. Archaeol.Sci. 22, 495–509.
Merrick, H. V. & Brown, F. H. (1984). Obsidiansources and patterns of source utilization in Kenyaand northern Tanzania: some initial findings. Afr.Archaeol. Rev. 2, 129–152.
Merrick, H. V., Haesaerts, P., de Heinzelein, J. &Howell, F. C. (1973). Artefactual occurrencesassociated with the Pliocene/Pleistocene ShunguraFormation, southern Ethiopia. Nature 242, 572–575.
Merrick, H. V., Brown, F. H. & Nash, W. P. (1994).Use and movement of obsidian in the Early andMiddle Stone Ages of Kenya and northern Tanzania.In (S. T. Childs, Ed.) Society, Culture, and Technologyin Africa, pp. 29–44. MASCA 11 (supplement).

554 . . .
Michels, J. W., Tsong, I. S. T. & Nelson, C. M. (1983).Obsidian dating and East African archaeology.Science 219, 361–366.
Miller, G. H. (1993). Chronology of hominid occu-pation at Bir Tarfawi and Bir Sahara East, based onthe epimerization of isoleucine in ostrich eggshells. In(F. Wendorf, R. Schild & A. E. Close, Eds) EgyptDuring the Last Interglacial, pp. 241–254. New York:Plenum.
Miller, G. H., Wendorf, F., Ernst, R., Schild, R.,Close, A. E., Friedman, I. & Schwarcz, H. (1991).Dating lacustrine episodes in the eastern Sahara byepimerization of isoleucine in ostrich eggshells. Pal-aeogeogr., Palaeoclimatol., Palaeoecol. 84, 175–189.
Miller, G. H., Beaumont, P. B., Jull, A. J. T. &Johnson, B. (1993). Pleistocene geochronology andpalaeothermometry from protein diagenesis inostrich eggshells: implications for the evolution ofmodern humans. In (M. J. Aitken, C. B. Stringer &P. A. Mellars, Eds) The Origin of Modern Humansnad the Impact of Chronometric Dating, pp. 49–68.Princeton: Princeton University Press.
Miller, G. H., Magee, J. W. & Jull, A. J. T. (1997).Low-latitude glacial cooling in the southern hemi-sphere from amino-acid racemization in emueggshells. Nature 385, 241–244.
Miller, G. H., Beaumont, P. B., Brooks, A. S., Deacon,H. J., Hare, P. E. & Jull, A. J. T. (1999). Earliestmodern humans in South Africa dated by isoleucineepimerization in ostrich eggshell. Quatern. Sci. Rev.18, 1537–1548.
Miller, S. F. (1979). Lukenya Hill: GvJm46 excavationreport. Nyame Akuma 14, 31–34.
Milo, R. (1998). Evidence for hominid predation atKlasies River Mouth, South Africa, and its impli-cations for the behaviour of early modern humans.J. Archaeol. Sci. 25, 99–133.
Minichillo, T. & Marean, C. W. (2000). Behavioralecological modeling and faunal resource use in theMiddle and Later Stone Ages of southern Africa.J. hum. Evol. 38, A22.
Mitchell, P. J. (1990). A palaeoecological model forarchaeological site distribution in southern Africaduring the Upper Pleniglacial and Late Glacial. In(O. Soffer & C. Gamble, Eds) The World at 18,000BP Vol. 2: Low Latitudes, pp. 189–205. London:Unwin Hyman.
Mitchell, P. J. (1994). Understanding the MSA/LSAtransitions: the pre-20,000 BP assemblages from newexcavations at Sehonghong Rock Shelter, Lesotho.S. Afr. Field Archaeol. 3, 15–25.
Mithen, S. (1994). From domain-specific to general-ized intelligence: a cognitive interpretation of theMiddle/Upper Paleolithic transition. In (C. Renfrew& E. Zubrow, Eds) The Ancient Mind: Elements ofa Cognitive Archaeology, pp. 29–39. Cambridge:Cambridge University Press.
Mithen, S. (1996). The Prehistory of the Mind: A Searchfor the Origins of Art, Religion, and Science. London:Thames & Hudson.
Montgomery, P. Q., Williams, H. O. L., Reading, N. &Stringer, C. B. (1994). An assessment of the
temporal bone lesions of the Broken Hill cranium.J. Archaeol. Sci. 21, 331–337.
Morel, J. (1974). La station eponyme de l’OuedDjebbana a Bir-el-Ater (Est algerien), contribution ala connaissance de son industrie et de sa faune.L’Anthropologie 78, 53–80.
Morgan, L. H. (1871). Ancient Society. New York:Henry Holt.
Morris, A. G. (1992a). Holocene Human Skeletons fromSouth Africa: A Master Catalogue. Johannesburg:Witswatersrand University Press.
Morris, A. G. (1992b). Biological relationships betweenUpper Pleistocene and Holocene populations insouthern Africa. In (G. Brauer & F. Smith, Eds)Continuity or Replacement: Controversies in Homosapiens Evolution, pp. 131–144. Rotterdam: Balkema.
Morselli, E. (1926). Antropolgia Generale: L’UomoSecondo la Teoria dell’Evoluzione, Lezioni Dettate NelleUniversita di Torino e di Genova. Turin: Unione.
Mturi, A. (1976). New hominid from Lake Ndutu,Tanzania. Nature 262, 484–485.
Nei, M. (1995). Genetic support of the out-of-Africatheory of human evolution. Proc. natl. Acad. Sci. USA92, 6720–6722.
Nicoll, K., Giegengack, R. & Kleindienst, M. (1999).Petrogenesis of artifact-bearing fossil-spring tufadeposits from Kharga Oasis, Egypt. Geoarchaeol. 14,849–863.
Nitecki, M. H. & Nitecki, D. V. (Eds) (1989). TheEvolution of Human Hunting. New York: PlenumPress.
Noble, W. & Davidson, I. (1991). The evolutionaryemergence of modern human behaviour: languageand its archaeology. Man 26, 223–253.
Noble, W. & Davidson, I. (1996). Human Evolution,Language and Mind: A Psychological and Archaeologi-cal Inquiry. Cambridge: Cambridge UniversityPress.
Nuzhnyi, D. J. (1990). Projectile damage on UpperPaleolithic microliths: use of bow and arrow amongPleistocene hunters in the Ukraine. In Proceedingsof the International Conference on Lithic Use-WearAnalysis. Uppsala, Sweden, 15–17 February, 1990,pp. 113–124. Uppsala: Societas ArchaeologicaUpsaliensis.
Oakley, K. P. (1957). The dating of the Broken Hill,Florisbad, and Saldanha skulls. In (J. D. Clark,Ed.) Proceedings of the Third Panafrican Congress onPrehistory, Livingstone, 1955, pp. 76–79. London:Chatto & Windus.
Oakley, K. P., Campbell, B. G. & Molleson, T. I.(1977). Catalogue of Fossil Hominids. Part I: Africa.London: British Museum (Natural History).
Obermeier, H. (1924). Fossil Man in Spain. Oxford:Oxford University Press.
O’Connell, J. S. & Allen, J. (1998). When did modernhumans first arrive in greater Australia and why is itimportant to know? Evol. Anthrop. 6, 132–146.
Ogilvie, M. D., Curran, B. K. & Trinkaus, E. (1989).The incidence and patterning of dental hypoplasiasamong the Neandertals. Am. J. phys. Anthrop. 79,25–41.

555 ’
Opperman, H. & Heydenrych, B. (1990). A 22,000year old Middle Stone Age camp site with plant foodremains from the north-eastern Cape. S. Afr.Archaeol. Bull. 45, 93–99.
Otte, M. I. (1990). From the Middle to the UpperPalaeolithic: the nature of the transition. In (P.Mellars, Ed.) The Emergence of Modern Humans:An Archaeological Perspective, pp. 438–456. Ithaca:Cornell University Press.
Otte, M. I. & Keeley, L. H. (1990). The impact ofregionalism on paleolithic studies. Curr. Anthrop. 31,577–582.
Ovchinnikov, I. V., Gotherstrom, A., Romanova, G. P.,Kharitonov, V. M. & Liden, K. G. W. (2000).Molecular analysis of Neanderthal DNA from thenorthern Caucasus. Nature 404, 490–493.
Palma di Cesnola, A. (1993). Il Paleolitico Superiore inItalia: Introduzione allo Studio. Firenze: Garlatti eRazzai.
Pares, J. M. & Pares-Gonzalez, A. (1995). Paleo-magnetic age for hominid fossils at Atapuercaarchaeological site, Spain. Science 269, 830–832.
Parker, S. T. & Gibson, K. R. (1979). A developmentalmodel for the evolution of language and intelli-gence in early hominids. Behav. Brain Sci. 2, 367–408.
Parker, S. T. & Gibson, K. R. (Eds) (1990). ‘Language’and Intelligence in Monkeys and Apes: ComparativeDevelopmental Perspectives. Cambridge: CambridgeUniversity Press.
Parkington, J. (1990). A critique of the consensus viewon the age of Howieson’s Poort assemblages in SouthAfrica. In (P. Mellars, Ed.) The Emergence of ModernHumans: An Archaeological Perspective, pp. 34–55.Ithaca: Cornell University Press.
Partridge, T. C. (1982). The chronological positions ofthe fossil hominids of southern Africa. In (M. A. deLumley, Ed.) Homo erectus et la Place de l’Hommede Tautavel parmi les hominides Fossiles. Premiere Con-gres International de Paleontolgie Humaine, pp. 617–675. Vol. 2. UNESCO Colloque Internationaldu Centre National de la Recherche Scientifique:Pretirages.
Paterson, H. E. H. (1985). The recognition conceptof species. In (E. S. Vrba, Ed.) Species andSpeciation, pp. 21–29. Pretoria: Transvaal MuseumMonographs 4.
Pearson, O. (2000). Activity, climate, and postcranialrobusticity: implications for modern human originsand scenarios of adaptive change. Curr. Anth. 41,569–607.
Pearson, O. & Grine, F. (1996). Morphology of theBorder Cave hominid ulna and humerus. S. Afr. J.Sci. 92, 231–236.
Pearson, O. M. (1997). Postcranial morphology andthe origin of modern humans. Ph.D. Dissertation,State University of New York at Stony Brook.
Pearson, O. M. & Grine, F. E. (1997). Re-analysis ofthe hominid radii from Cave of Hearths and KlasiesRiver Mouth, South Africa. J. hum. Evol. 32,577–592.
Pearson, O. M., Churchill, S. E., Grine, F. G.,Trinkaus, E. & Holliday, T. W. (1998). Multivariate
analyses of the hominid ulna from Klasies RiverMouth. J. hum. Evol. 34, 653–658.
Perkins, D. & Daly, P. (1968). A hunters’ village inNeolithic Turkey. Scient. Am. 219, 97–196.
Pfeiffer, S. (1998). Klasies River Mouth post-cranialcomparisons. J. hum. Evol. 34, A17.
Pfeiffer, S. & Zehr, M. K. (1996). A morphologicaland histological study of the human humerus fromBorder Cave. J. hum. Evol. 31, 49–59.
Phillipson, D. W. (1977). The Later Prehistory of Easternand Southern Africa. London: Heinemann.
Plug, I. (1982). Bone tools and shell, bone and ostricheggshell beads from Bushman Rock Shelter (BRS),Eastern Transvaal. S. Afr. Archaeol. Bull. 37, 57–62.
Poeggenphol, C. (1999). Workshop 5: Fish remains inarchaeological sites. World Archaeological Congress,Capetown, RSA, January 15.
Potts, R. & Deino, A. (1995). Mid-Pleistocene changein large mammal faunas of East Africa. Quatern. Res.43, 106–113.
Potts, R., Shipman, P. & Ingersall, E. (1988). Tapho-nomy, paleoecology, and hominids at Lainyamok,Kenya. J. hum. Evol. 17, 597–614.
Potts, R., Behrensmeyer, A. K. & Ditchfield, P.(1990a). Hominid responses to Pleistocene paleo-landscapes, Olorgesailie Formation, Kenya. J. hum.Evol. 36, A18–A19.
Potts, R., Behrensmeyer, A. K. & Ditchfield, P.(1999b). Paleolandscape variation and EarlyPleistocene hominid activities: Members 1 and 7,Olorgesailie Formation, Kenya. J. hum. Evol. 37,747–788.
Power, C. & Watts, I. (1996). Female strategies andcollective behaviour: the archaeology of earliest Homosapiens sapiens. In (J. Steele & S. Shennan, Eds) TheArchaeology of Human Ancestry: Power, Sex, andTradition, pp. 306–330. London: Routledge.
Power, C. & Aiello, L. (1997). Female proto-symbolicstrategies. In (L. D. Hager, Ed.) Women in HumanEvolution, pp. 153–171. London: Routledge.
Price-Williams, D. (1978). A preliminary report onrecent excavations of Middle and Late Stone Agelevels at Sibebe shelter, north-west Swaziland. S. Afr.Archaeol. Bull. 53, 22–28.
Pritchard, J. K., Seielstad, M. T., Perez-Lezaun, A. &Feldman, M. W. (1999). Population growth ofhuman Y chromosomes: a study of Y chromosomemicrosatellites. Molecular Biology & Evolution 16,1791–1798.
Protsch, R. (1975). The absolute dating of UpperPleistocene sub-Saharan fossil hominids and theirplace in human evolution. J. hum. Evol. 4, 297–322.
Pycraft, W. P., Smith, G. E., Yearsley, M., Carter, J.T., Smith, R. A., Hopwood, A. T., Bate, D. M. A. &Swinton, W. E. (1928). Rhodesian Man and Associ-ated Remains. London: British Museum (NaturalHistory).
Quintana-Murci, L., Semino, O., Bandelt, H.-J.,Passarino, G., McElreavey, K. & Santachiara-Benerecetti, A. S. (1999). Genetic evidence of anearly exit of Homo sapiens from Africa through easternAsia. Nature Genetics 23, 437–441.

556 . . .
Rak, Y. (1993). Morphological variation in Homoneanderthalensis and Homo sapiens in the Levant: abiogeographic model. In (W. H. Kimbel & L. B.Martin, Eds) Species, Species Concepts, and PrimateEvolution, pp. 523–536. New York: Plenum.
Rak, Y. (1998). Does any Mousterian cave presentevidence of two hominid species? In (T. Akazawa, K.Aoki & O. Bar-Yosef, Eds) Neandertals and ModernHumans in the Levant, pp. 353–366. New York:Plenum.
Relethford, J. (1995). Genetics and modern humanorigins. Evol. Anthrop. 4, 53–63.
Relethford, J. H. (1998). Genetics of modern humanorigins and diversity. A. Rev. Anthrop. 27, 1–23.
Relethford, J. H. & Harpending, H. C. (1994). Cranio-metric variation, genetic theory, and modern humanorigins. Am. J. phys. Anthrop. 95, 249–270.
Relethford, J. H. & Harpending, H. C. (1995). Ancientdifferences in population size can mimic a recentAfrican origin for modern humans. Curr. Anthrop.36, 667–674.
Relethford, J. H. & Jorde, L. B. (1999). Geneticevidence for larger African population size duringrecent human evolution. Am. J. phys. Anthrop. 108,251–260.
Renne, P. R., Sharp, W. D., Deino, A. L., Orsi, G. &Civetta, L. (1997). 40Ar/39Ar dating into the historicrealm. Science 277, 1279–1280.
Revillion, S. (1995). Technologie du debitage laminaireau paleolithique moyen en Europe septentrionale:etat de la question. Bull. Soc. prehist. fr. 92, 425–441.
Revillion, S. & Tuffreau, A. (1994). Valeur et signifi-cation du debitage laminaire du gisement paleo-lithique moyen de Seclin (Nord). In (S. Revillion& A. Tuffreau, Eds) Les Industries Laminaires auPaleolithique Moyen, pp. 19–44. Paris: CNRS.
Rightmire, G. P. (1975). Problems in the study of laterPleistocene man in Africa. Am. Anthrop. 77, 28–52.
Rightmire, G. P. (1978a). Florisbad and humanpopulation succession in southern Africa. Am. J.phys. Anthrop. 48, 475–486.
Rightmire, G. P. (1978b). Human skeletal remainsfrom the southern Cape Province and their bearingon the Stone Age prehistory of South Africa. Quatern.Res. 9, 219–230.
Rightmire, G. P. (1979a). Cranial remains ofHomo erectus from Beds II and IV, Olduvai Gorge,Tanzania. Am. J. phys. Anthrop. 51, 99–115.
Rightmire, G. P. (1979b). Implications of BorderCave skeletal remains for Late Pleistocene humanevolution. Curr. Anthrop. 20, 23–35.
Rightmire, G. P. (1980). Middle Pleistocene hominidsfrom Olduvai Gorge, northern Tanzania. Am. J.phys. Anthrop. 53, 225–241.
Rightmire, G. P. (1983). The Lake Ndutu cranium andearly Homo sapiens in Africa. Am. J. phys. Anthrop.61, 245–254.
Rightmire, G. P. (1984). Homo sapiens in sub-SaharanAfrica. In (F. H. Smith & F. Spencer, Eds) TheOrigins of Modern Humans: A World Survey of theFossil Evidence, pp. 295–325. New York: Alan R.Liss.
Rightmire, G. P. (1990). The Evolution of Homo erectus.Cambridge: Cambridge University Press.
Rightmire, G. P. (1994). The relationship of Homoerectus to later Middle Pleistocene hominids. Cour.Forsch.-Inst. Senckenberg 171, 319–326.
Rightmire, G. P. (1995). Diversity within the genusHomo. In (E. S. Vrba, G. H. Denton, T. C. Partridge& L. H. Burckle, Eds) Paleoclimate and Evolution,With Emphasis on Human Origins, pp. 483–492. NewHaven: Yale University Press.
Rightmire, G. P. (1996). The human cranium fromBodo, Ethiopia: evidence for speciation in the MiddlePleistocene? J. hum. Evol. 31, 21–39.
Rightmire, G. P. (1998). Human evolution in theMiddle Pleistocene: the role of Homo heidelbergensis.Evol. Anthrop. 6, 218–227.
Rightmire, G. P. & Deacon, H. J. (1991). Comparativestudies of Late Pleistocene human remains fromKlasies River Mouth. J. hum. Evol. 20, 131–156.
Robbins, L. H. (1999). Direct dating of workedostrich eggshell in the Kalahari. Nyame Akuma 52,11–16.
Robbins, L. H. & Murphy, M. L. (1998). The earlyLate Stone Age and evidence of modern humanbehavior. J. hum. Evol. 34, A18.
Robbins, L. H., Murphy, M. L., Stewart, K. M.,Campbell, A. C. & Brook, G. A. (1994). Barbedbone points, paleoenvironment and the antiquity offish exploitation in the Kalahari Desert, Botswana.J. Field Archaeol. 21, 257–264.
Robbins, L. H., Murphy, M. L., Campbell, A. C. &Brook, G. A. (1998). Intensive mining of specularhematite in the Kalahari ca. A.D. 800–1000. Curr.Anthrop. 39, 144–150.
Robbins, L. R., Murphy, M. L., Brook, G. A., Ivester,A., Campbell, A. C., Klein, R. G., Milo, R., Brooks,A. S., Kokis, J. E. & Downey, W. (under review).Archaeology, paleoenvironment, and chronology ofthe White Paintings Rock Shelter, Tsodilo Hills,northwest Kalahari Desert, Botswana. J. Archaeol.Sci.
Roche, H. & Texier, P.-J. (1991). La notion de com-plexite dans un ensemble lithique. Application auxseries achuleennes d’Isenya (Kenya). Vingt-cinq ansd’Eutudes Technologiques en Prehistoire, XIe RencontresInternationales d’Archeologie et d’Histoire d’Antibes.99–108.
Roche, H. & Texier, P.-J. (1995). Evaluation of tech-nical competence of Homo erectus in East Africaduring the Middle Pleistocene. In (J. R. F. Bower& S. Sartono, Eds) Human Evolution in its Evo-lutionary Context: Proceedings of the PithecanthropusCentennial 1893–1993 Congress, pp. 153–167. Leiden:Pithecanthropus Centenniel Foundation.
Roche, J. & Texier, J.-P. (1976). Decouverte des resteshumains dans un niveau superieur de la grotte desContrabandiers a Temara (Maroc). C. r. Acad. Sci.,Paris 282, 45–47.
Roebrooks, W., Kolen, J. & Rensink, E. (1988).Planning depth, anticipation and the organization ofMiddle Palaeolithic technology: the archaic nativesmeet Eve’s descendants. Helinium 28, 17–34.

557 ’
Roubet, F.-E. (1966). Presentation comparative d’ungisement cotier des environs de Berard, a l’Ouestd’Alger. (Eutude stratigraphique sur l’Aterien). InCongres Prehistorique de France. 18e Session, Ajaccio,1966, pp. 109–128. Paris: C. Klincksieck.
Rozoy, J.-G. (1985). The revolution of the bowmen inEurope. In (C. Bonsall, Ed.) The Mesolithic in Europe,pp. 13–28. Edinburgh: John Donald.
Rudner, J. (1971). Painted burial stones from the Cape.S. Afr. J. Sci. 2, 54–61.
Ruff, C. B. (1994). Morphological adaptation toclimate in modern and fossil hominids. Yearb. phys.Anthrop. 37, 65–107.
Ruff, C. B., Trinkaus, E. & Holliday, T. W. (1997).Body mass and encephalization in Pleistocene Homo.Nature 387, 173–176.
Ruiz, C. B. & Hublin, J.-J. (1994). The lateNeanderthal site of Zafarraya (Andalucıa, Spain). In(J. R. Vidal, F. D. Olmo, C. Finlayson & F. G.Pacheco, Eds) Gibralter During the Quaternary.Sevilla: AEQUA.
Rust, A. (1943). Die Alt- und Mittelsteinzeitlichen Fundevon Stellmoor. Neumunster: K. Wachholtz.
Ruvolo, M. (1996). New approach to studying modernhuman origins: hypothesis testing with coalescencetime distributions. Mol. Phylogenet. Evol. 5, 202–219.
Ruvolo, M. (1997). Molecular phylogeny of the homi-noids: inferences from multiple independent DNAsequence data sets. Mol. Biol. Evol. 14, 248–265.
Saban, R. (1972). Les hommes fossiles du Maghreb.L’Ouest Mediterrane 25, 2443–2458.
Saban, R. (1975). Les restes humains de Rabat(Kebibat). Annls. Paleontol. (Vert.) 61, 151–207.
Saban, R. (1977). The place of Rabat man (Kebibat,Morocco) in human evolution. Curr. Anthrop. 18,518–524.
Sackett, J. R. (1977). The meaning of style in archae-ology: a general model. Am. Antiq. 42, 369–380.
Sackett, J. R. (1982). Approaches to style in lithicarchaeology. J. Anthrop. Archaeol. 1, 59–112.
Sackett, J. R. (1990). Style and ethnicity in archae-ology: the case for isochrestism. In (M. Conkey &C. Hastorf, Eds) The Uses of Style in Archaeology,pp. 32–43. Cambridge: Cambridge University Press.
Sampson, C. G. (1968). The Middle Stone Age Industriesof the Orange River Scheme Area. Bloemfontein:Memoirs of the National Museum No. 4.
Sampson, C. G. (1972). The Stone Age Industries of theOrange River Scheme and South Africa. Bloemfontein:Memoirs of the National Museum No. 6.
Sampson, C. G. (1974). The Stone Age Archaeology ofSouthern Africa. New York: Academic Press.
Santa Luca, A. P. (1978). A re-examination of pre-sumed Neanderthal fossils. J. hum. Evol. 7, 619–636.
Sausse, F. (1975a). Etude du quelques restes humainstrouves a Soleb (Soudan). W. Afr. J. Archaeol. 5,41–51.
Sausse, F. (1975b). La mandibule Atlanthropienne dela Carriere Thomas I (Casablanca). L’Anthropologie79, 81–112.
Savage-Rumbaugh, E. S. & Rumbaugh, D. M. (1993).The emergence of language. In (K. R. Gibson & T.
Ingold, Eds) Tools, Language and Cognition in HumanEvolution, pp. 86–108. Cambridge: CambridgeUniversity Press.
Scheer, A. (1993). The organization of lithic resourceuse during the Gravettian in Germany. In (H.Knecht, A. Pike-Tay & R. White, Eds) BeforeLascaux: The Complex Record of the Early UpperPaleolithic, pp. 193–210. Boca Raton: CRC Press.
Schick, K. (1994). The Movius Line reconsidered:perspectives on the earlier Paleolithic of eastern Asia.In (R. S. Corruccini & R. L. Ciochon, Eds) Inte-grative Pathways to the Past: PaleoanthropologicalAdvances in Honor of F. Clark Howell, pp. 569–596.Englewood Cliffs, NJ: Prentice Hall.
Schluchter, E. (1998). Calibration of the 14C timescalebeyond 22,000 BP. Radiocarbon 40, 475–482.
Schneider, D. A., Kent, D. V. & Mello, G. A. (1992).A detailed chronology of the Australasian impactevent, the Brunhes-Matuyama geomagnetic polarityreversal, and global climate. Earth & PlanetaryScience Letters 111, 395–405.
Schoetensack, O. (1908). Der Unterkiefer des Homoheidelbergensis aus den Sanden von Mauer beiHeidelberg. Leipzig: Wilhelm Englemann.
Schwarcz, H. P. (1992). Uranium series dating inpaleoanthropology. Evol. Anthrop. 1, 56–62.
Schwarcz, H. P. (1994). Chronology of modernhumans in the Levant. In (O. Bar-Yosef & R. S. Kra,Eds) Late Quaternary Chronology and Paleoclimatesof the Eastern Mediterranean, pp. 21–31. Tucson:Radiocarbon.
Schwarcz, H. P. (1998). Bones and teeth: datablebut problematic materials from prehistoric sites.Paper presented at the IVth Dual Congress of theInternational Association for the Study of HumanPaleontology and the International Association ofHuman Biologists, Sun City, South Africa.
Schwarcz, H. P. & Grun, R. (1993a). Electron spinresonance dating of the origin of modern man. In(M. J. Aitken, C. B. Stringer & P. A. Mellars, Eds)The Origin of Modern Humans and the Impact ofChronometric Dating, pp. 40–48. Princeton: PrincetonUniversity Press.
Schwarcz, H. P. & Grun, R. (1993b). Electron spinresonance dating of tooth enamel from Bir Tarfawi.In (F. Wendorf, R. Schild & A. E. Close, Eds) EgyptDuring the Last Interglacial, pp. 234–237. New York:Plenum.
Schwarcz, H. P. & Rink, J. (1998). Progress in ESR andU-series chronology of the Levantine Paleolithic.In (T. Akazawa, K. Aoki & O. Bar-Yosef, Eds)Neandertals and Modern Humans in Western Asia,pp. 57–68. New York: Plenum.
Schwarcz, J. P. & Rink, W. J. (2000). ESR dating of theDie Kelders Cave 1 Site, South Africa. J. hum. Evol.38, 121–128.
Schwartz, J. H. & Tattersall, I. (2000). The humanchin revisited: What is it and who has it? J. hum. Evol.38, 367–410.
Schwarcz, H. P., Grun, R. V. B., Bar-Yosef, O.,Valladas, H. & Tchernov, E. (1988). ESR dates from

558 . . .
the hominid burial site of Qafzeh in Israel. J. hum.Evol. 17, 733–737.
Shackleton, N. J. (1982). Stratigraphy and chronologyof the Klasies River Mouth deposits: oxygen isotopeevidence. In (R. Singer & J. Wymer, Eds) The MiddleStone Age at Klasies River Mouth in South Africa,pp. 194–199. Chicago: Chicago University Press.
Shackleton, N. J., Berger, A. & Peltier, W. R. (1990).An alternative astronomical calibration of the lowerPleistocene time scale based on Ocean DrillingProgram site 677. Trans. R. Soc. Edinb.: Earth Sci.81, 251–261.
Shea, J. J. (1988). Spear points from the MiddlePaleolithic of the Levant. J. Field Archaeol. 15,441–450.
Sherry, S., Rogers, A. R., Harpending, H., Soodyall,H., Jenkins, T. & Stoneking, M. (1994). Mismatchdistributions of mtDNA reveal recent humanpopulation expansions. Hum. Biol. 66, 761–775.
Shipman, P., Potts, R. & Pickford, M. (1983).Lainyamok: a new Middle Pleistocene hominid site.Nature 306, 365–368.
Sillen, A. & Morris, A. (1996). Diagenesis of bone fromBorder Cave: implications for the age of the BorderCave hominids. J. hum. Evol. 31, 499–506.
Singer, B. S. & Pringle, M. S. (1996). Age and durationof the Matuyama-Brunhes geomagnetic polarityreversal from 40Ar39Ar incremental heating analysesof lavas. Earth & Planetary Science Letters 139, 47–61.
Singer, R. (1954). The Saldanha skull from Hopefield,South Africa. Am. J. phys. Anthrop. 12, 345–362.
Singer, R. & Wymer, J. (1968). Archaeological investi-gations at the Saldahna skull site in South Africa.S. Afr. Archaeol. Bull. 23, 63–74.
Singer, R. & Wymer, J. (1982). The Middle Stone Ageat Klasies River Mouth in South Africa. Chicago:University of Chicago Press.
Smith, F. (1985). Continuity and change in the originof modern Homo sapiens. Z. Morph. Anthrop. 75,197–222.
Smith, F. (1993). Models and realities in modernhuman origins: the African fossil evidence. In (M. J.Aitken, C. B. Stringer & P. A. Mellars, Eds) TheOrigin of Modern Humans and the Impact of Chrono-metric Dating, pp. 234–248. Princeton: PrincetonUniversity Press.
Soffer, O. (1994). Ancestral lifeways in Eurasia: theMiddle and Upper Paleolithic records. In (M. H.Nitecki & D. V. Nitecki, Eds) Origin of AnatomicallyModern Humans, pp. 101–119. New York: Plenum.
Solan, M. & Day, M. H. (1992). The Baringo(Kapthurin) ulna. J. hum. Evol. 22, 307–314.
Spoor, F. & Stringer, C. B. (1998). Rare temporal bonepathology of the Singa calvaria from Sudan. Am. J.phys. Anthrop. 107, 41–50.
Spoor, F., O’Higgins, P., Dean, C. & Lieberman, D. E.(1999). Anterior spenoid in modern humans. Nature397, 572.
Springer, M. S. (1990). The effect of random rangetruncations on patterns of evolution in the fossilrecord. Paleobiology 16, 512–520.
Stapleton, P. & Hewitt, J. (1927). Stone implementsfrom a rock shelter at Howieson’s Poort nearGrahamstown (part 1). S. Afr. J. Sci. 24, 574–587.
Stapleton, P. & Hewitt, J. (1928). Stone implementsfrom a rock shelter at Howieson’s Poort nearGrahamstown (part 2). S. Afr. J. Sci. 25, 399–309.
Stearns, C. E. & Thurber, D. L. (1965). 230Th/234Udates of Late Pleistocene marine fossils from theMediterranean and Moroccan littorals. Quaternaria7, 29–42.
Steele, J. & Gamble, C. (1998). Inferring diet and rangeuse from lithic transport data. J. hum. Evol. 24, A20.
Stewart, K. M. (1989). Fishing Sites of North and EastAfrica in the Late Pleistocene and Holocene. Oxford:BAR International Series 521.
Stiner, M. C. (1990). The use of mortality patterns inarchaeological studies of homind predatory patterns.J. Anthrop. Res. 9, 305–351.
Stiner, M. C. (1993). Small animal exploitation and itsrelation to hunting, scavenging, and gathering in theItalian Mousterian. In (G. L. Peterkin, H. M. Bricker& P. Mellars, Eds) Hunting and Animal Exploitationin the Later Palaeolithic and Mesolithic of Eurasia,pp. 101–119. Vol. 4. Washington, D.C.: Papers ofthe American Anthropological Association.
Stiner, M. C. (1994). Honor Among Thieves: AZooarchaeological Study of Neandertal Ecology.Princeton: Princeton University Press.
Stiner, M. C., Munro, N. C., Surovell, E., Tchernov,E. & Bar-Yosef, O. (1999). Paleolithic popula-tion growth pulses evidenced by small animalexploitation. Science 283, 190–194.
Stiner, M. C., Munro, N. D. & Surovell, T. A. (2000).The tortoise and the hare: small-game use, the broadspectrum revolution, and paleolithic demography.Curr. Anthrop. 41, 39–74.
Stoneking, M. (1993). DNA and recent humanevolution. Evol. Anthrop. 2, 60–73.
Stoneking, M., Sherry, S. T., Redd, A. J. & Vigilant, L.(1993). New approach to dating suggests a recent agefor the human mtDNA ancestor. In (M. J. Aitken,C. B. Stringer & P. A. Mellars, Eds) The Origin ofModern Humans and the Impact of Chronometric Dating,pp. 84–103. Princeton: Princeton University Press.
Straus, L. G. (1983). From Mousterian toMagdalenian: cultural evolution viewed fromVasco-Cantabrian Spain and Pyrenean France. In(E. Trinkaus, Ed.) The Mousterian Legacy: HumanBiocultural Change in the Upper Pleistocene,pp. 73–111. Oxford: British Archaeological ReportsInternational Series S164.
Straus, L. (1994). Upper Paleolithic origins and radio-carbon calibration: more new evidence from Spain.Evol. Anthrop. 3, 195–198.
Straus, L. G. (1995). The Upper Paleolithic of Europe:an overview. Evol. Anthrop. 4, 4–16.
Straus, D. & Sadler, P. M. (1989). Classical confidenceintervals and Bayesian probability estimates forends of local taxon ranges. Mathematical Geology 21,411–427.
Stringer, C. B. (1974). A multivariate study of thePetralona skull. J. hum. Evol. 3, 397–404.

559 ’
Stringer, C. B. (1979). A re-evaluation of the fossilhuman calvaria from Singa, Sudan. Bull. Brit. Mus.nat. Hist. (Geology) 32, 77–83.
Stringer, C. B. (1985). Middle Pleistocene hominidvariability and the origin of late Pleistocene humans.In (E. Delson, Ed.) Ancestors: The Hard Evidence,pp. 289–295. New York: Alan R. Liss.
Stringer, C. B. (1986). An archaic character in theBroken Hill innominate E 719. Am. J. phys. Anthrop.71, 115–120.
Stringer, C. B. (1989). Documenting the origin of mod-ern humans. In (E. Trinkaus, Ed.) Emergence of ModernHumans: Biocultural Adaptations in the Late Pleistocene,pp. 67–96. Cambridge: Cambridge University Press.
Stringer, C. B. (1992). Replacement, continuity and theorigin of modern humans. In (G. Brauer & F. H. Smith,Eds) Continuity or Replacement: Controversies in Homosapiens Evolution, pp. 9–24. Rotterdam: Balkema.
Stringer, C. B. (1993a). New views on modern humanorigins. In (T. D. Rasmussen, Ed.) The Origin andEvolution of Humans and Humanness, pp. 75–94.Boston: Jones & Barlett.
Stringer, C. B. (1993b). The secret of the pit of bones.Nature 362, 501–502.
Stringer, C. B. (1994). Out of Africa: a personalhistory. In (M. H. Nitecki & D. V. Nitecki,Eds) Origins of Anatomically Modern Humans,pp. 149–172. New York: Plenum.
Stringer, C. B. (1995). The evolution and distributionof later Pleistocene human populations. In (E. S.Vrba, G. H. Denton, T. C. Partridge & L. H.Burckle, Eds) Paleoclimate and Evolution, WithEmphasis on Human Origins, pp. 524–531. NewHaven: Yale University Press.
Stringer, C. B. (1996). Current issues in modernhuman origins. In (W. E. Meikle, F. C. Howell &N. G. Jablonski, Eds) Contemporary Issues in HumanEvolution, pp. 115–134. San Francisco: CaliforniaAcademy of Sciences.
Stringer, C. B. (1998). Chronological and biogeo-graphic perspectives on later human evolution. In (T.Akazawa, K. Aoki & O. Bar-Yosef, Eds) Neandertalsand Modern Humans in Western Asia, pp. 29–38. NewYork: Plenum.
Stringer, C. B. (1999). Has Australia backdated theHuman Revolution? Antiquity 73, 876–879.
Stringer, C. B. (2000). Coasting out of Africa. Nature405, 24–25.
Stringer, C. B. & Andrews, P. (1988). Genetic andfossil evidence for the origin of modern humans.Science 239, 1263–1268.
Stringer, C. B. & Hublin, J.-J. (1999). New age esti-mates for the Swanscombe hominid, and their sig-nificance for human evolution. J. hum. Evol. 37,873–877.
Stringer, C. B., Cornish, L. & Stuart-Macadam, P.(1985). Preparation and further study of theSinga skull from Sudan. Bull. Brit. Mus. Nat. Hist.(Geology) 38, 347–358.
Stringer, C. B., Grun, R., Schwarcz, H. & Goldberg, P.(1989). ESR dates for the hominid burial site ofes-Skhul in Israel. Nature 338, 756–758.
Sutton, J. E. G. (1974). The aquatic civilization ofMiddle Africa. J. Afr. Hist. 15, 527–546.
Sutton, J. E. G. (1977). The African aqualithic.Antiquity 51, 25–34.
Szabo, B. J. & Butzer, K. W. (1979). Uranium-seriesdating of lacustrine limestones from pan depositswith Final Acheulian assemblages at Rooidam,Kimberley District, South Africa. Quatern. Res. 11,257–260.
Taborin, Y. (1993). Shells of the French Aurignacianand Perigordian. In (H. Knecht, A. Pike-Tay & R.White, Eds) Before Lascaux: The Complex Record of theEarly Upper Paleolithic, pp. 211–227. Boca Raton:CRC Press.
Tallon, P. W. J. (1976). The stratigraphy, palaeo-environments and geomorphology of the PleistoceneKapthurin Formation, Kenya. Ph.D. Dissertation,Queen Mary College, London.
Tallon, P. W. J. (1978). Geological setting of thehominid fossils and Acheulian artifacts from theKapthurin Formation, Baringo District, Kenya. In(W. W. Bishop, Ed.) Geological Background to FossilMan, pp. 361–373. Edinburgh: Scottish AcademicPress.
Tamrat, E., Thouveny, N., Taieb, M. & Opdyke, N. D.(1995). Revised magnetostratigraphy of the Plio-Pleistocene sedimentary sequence of the OlduvaiFormation (Tanzania). Palaeogeogr., Palaeoclimatol.,Palaeoecol. 114, 273–283.
Tattersall, I. (1986). Species recognition in humanpaleontology. J. hum. Evol. 15, 165–175.
Tattersall, I. (1992). Species concepts and speciesidentification in the fossil record. J. hum. Evol. 22,341–349.
Tattersall, I. (1993). Speciation and morphological dif-ferentiation in the genus Lemur. In (W. H. Kimbel &L. B. Martin, Eds) Species, Species Concepts, andPrimate Evolution, pp. 163–176. New York: Plenum.
Tattersall, I. (1995). The Fossil Trail: How We KnowWhat We Think We Know About Human Evolution.New York: Oxford University Press.
Tauxe, L., Deino, A. D., Behrensmeyer, A. K. & Potts,R. (1992). Pinning down the Brunhes/Matuyamaand upper Jaramillo boundaries: a reconciliation oforbital and isotopic time scales. Earth Planet. Sci.Lett. 109, 561–572.
Taylor, R. E. (1996). Radiocarbon dating: thecontinuing revolution. Evol. Anthrop. 4, 169–181.
Tchernov, E. (1988). The paleobiogeographicalhistory of the southern Levant. In (Y. Yom-Tov &E. Tchernov, Eds) The Zoogeography of Israel: TheDistribution and Abundance at a Geological Crossroad,pp. 159–250. Dordrect, Netherlands: MonographiaeBiologicae 62.
Tchernov, E. (1998). The faunal sequence of thesouthwest Asian Middle Paleolithic in relation tohominid dispersal events. In (T. Akazawa, K. Aoki &O. Bar-Yosef, Eds) Neandertals and Modern Humansin Western Asia, pp. 77–90. New York: Plenum.
Templeton, A. R. (1992). Human origins and analysisof mitochondrial DNA sequences. Science 255,737.

560 . . .
Texier, P.-J. (1996). Production en serie: la debitage delames de pierre a 250.000 ans. Pour La Science 223,22.
Texier, P.-J. & Roche, H. (1993). The impact ofpredetermination on the development of someAcheulean chaınes operatoires. Evolucion humana enEuropa y los yacimientos de la Sierra de Atapuerca,Actas de Junta de Castilla y Leon 2, 403–420.
Thackeray, A. I. (1989). Changing fashions in theMiddle Stone Age: the stone artifact sequence fromKlasies River Mouth Main Site, South Africa. Afr.Archaeol. Rev. 7, 35–57.
Thackeray, A. I. (1992). The Middle Stone Agesouth of the Limpopo River. J. World. Prehist. 6,385–440.
Thackeray, A. I. (2000). Middle Stone Age artefactsfrom the 1993 and 1995 excavations of Die KeldersCave 1, South Africa. J. hum. Evol. 38, 147–168.
Thackeray, A. I. & Kelley, A.J. (1988). A technologicaland typological analysis of Middle Stone Age assem-blages antecedent to the Howieson’s Poort at KlasiesRiver main site. S. Afr. Archaeol. Bull. 43, 15–26.
Thackeray, J. F. (1988). Molluscan fauna from KlasiesRiver, South Africa. S. Afr. Archaeol. Bull. 43, 27–32.
Thackeray, J. F. & Avery, D. M. (1990). A comparisonbetween temperature indices for Late Pleistocenesequences at Klasies River and Border Cave, SouthAfrica. Paleoecol. Afr. 21, 311–315.
Thomsen, C. J. (1837). Leitfaden zur NordischenAlterthumskunde. Copenhagen: Herausgegeben vonder Koniglichen Gesellschaft fur NordischeAltertumskunde.
Thorne, A., Grun, R., Mortimer, B., Spooner, N.,Simpson, J., McCulloch, M., Taylor, L. & Curnoe,D. (1999). Australia’s oldest human remains: ageof the Lake Mungo 3 skeleton. J. hum. Evol. 36,591–612.
Tillet, T. (1978). Presence de pendeloques en milieuaterien au Niger oriental. Bull. Socl. prehist. fr. 75,273–275.
Tishkoff, S. A., Dietzsch, E., Speed, W., Pakstis, A. J.,Kidd, J. R., Cheung, K., Bonne-Tamir, B.,Santachiara-Benerecetti, A. S., Moral, P., Krings,M., Paabo, S., Watson, E., Risch, N., Jenkins, T. &Kidd, K. K. (1996). Global patterns of linkagedisequilibrium at the CD4 locus and modern humanorigins. Science 271, 1380–1387.
Tishkoff, S. A., Jenkins, T., Destro-Bisol, G., Kidd,K. K. & Clark, A. G. (2000). The emergence ofmodern humans in Africa: evidence from nucleargenetic haplotype data. Am. J. phys. Anthrop. Supple-ment 30, 304.
Tobias, P. V. (1967). The hominid skeletal remains ofHaua Fteah. In (C. B. M. McBurney, Ed.) The HauaFteah (Cyrenaica) and the Stone Age of the South-EastMediterranean, pp. 338–352. Cambridge: CambridgeUniversity Press.
Tobias, P. V. (1968). Middle and early UpperPleistocene members of the genus Homo in Africa.In (G. Kurth, Ed.) Evolution Und Hominisation,pp. 176–194. Stuttgart: G. Fischer.
Tobias, P. V. (1971). Human skeletal remains from theCave of Hearths, Makapansgat, northern Transvaal.Am. J. phys. Anthrop. 34, 335–368.
Tobias, P. V. (1991). Olduvai Gorge, Volume 4:The Skulls, Endocasts and Teeth of Homo habilis.Cambridge: Cambridge University Press.
Torrence, R. (1983). Time budgeting and hunter-gatherer technology. In (G. N. Bailey, Ed.) Hunter-Gatherer Economy in Prehistory: A EuropeanPerspective, pp. 11–22. Cambridge: CambridgeUniversity Press.
Toth, N. & Schick, K. (1993). Early stone industriesand inferences regarding language and cognition. In(K. R. Gibson & T. Ingold, Eds) Tools, Languageand Cognition in Human Evolution, pp. 346–362.Cambridge: Cambridge University Press.
Trinkaus, E. (1981). Neanderthal limb proportions andcold adaptations. In (C. B. Stringer, Ed.) Aspects ofHuman Evolution, pp. 187–224. London: Taylor &Francis.
Trinkaus, E. (1983). The Shanidar Neanderthals. NewYork: Academic Press.
Trinkaus, E. (1984). Western Asia. In (F. Smith & F.Spencer, Eds) The Origin of Modern Humans: A WorldSurvey of the Fossil Evidence, pp. 251–293. New York:Alan R. Liss.
Trinkaus, E. (1986). The Neandertals and modernhuman origins. A. Rev. Anthrop. 15, 193–218.
Trinkaus, E. (1987). Bodies, brawn, brains, andnoses: human ancestors and human predation. In(M. Nitecki & D. V. Nitecki, Eds) The Evolutionof Human Hunting, pp. 107–145. New York:Plenum.
Trinkaus, E. (1995). Neanderthal mortality patterns.J. Archaeol. Sci. 22, 121–142.
Turq, A. (1989). Exploitation des matieres premiereslithiques et occupation du sol: l’exemple du Moust-erien entre Dordogne et Lot. In (H. Laville, Ed.)Variations des Paleomilieux et Peuplement Prehistorique,pp. 179–204. Paris: CNRS.
Twiesselmann, F. (1991). La mandibule et le fragmentde maxillaire superieur de Loyangalani (rive estdu lac Turkana, Kenya). Anthropol. Prehist. 102,77–95.
Valladas, H., Reyss, J. L., Joron, J. L., Valladas,G., Bar-Yosef, O. & Vandermeersch, B. (1988).Thermoluminescence dating of Mousterian ‘Proto-Cro-Magnon’ remains from Israel and the origin ofmodern man. Nature 331, 614–616.
Valladas, H., Cachier, H., Maurice, P., Bernaldode Quiros, F., Clottes, J., Cabrera Valdes, V.,Uzquiano, P. & Arnold, M. (1992). Directradiocarbon dates for prehistoric paintings at theAltamira, El Castillo and Niaux caves. Nature 357,68–70.
Vallois, H. V. (1951). La mandibule fossile de la grottede Porc Epic pres Dire Daoua (Abyssinie).L’Anthropologie 55, 231–238.
Vallois, H. V. (1960). L’Homme de Rabat. Bull.Archeol. Maroc. 3, 87–91.
van Noten, F. (1977). Excavations at Matupi Cave.Antiquity 51, 35–40.

561 ’
van Noten, F. (1982). New Acheulian sites and anew hominid from Kapthurin. Nyame Akuma 21,11–12.
van Noten, F. & Wood, B. (1985). Un nouvel homi-nide a Baringo, Kenya. L’Anthropologie 89, 141–143.
van Noten, F., Cornelissen, E., Gysels, J., Moeyersons,J., Nijs, K. & Uytterschaut, H. (1987a). TheKapthurin Project, Baringo: The 1984 season.Nyame Akuma 28, 20–27.
van Noten, F., Cornelissen, E., Gysels, J., Moeyersons,J. & Uytterschaut, H. (1987b). The KapthurinProject, Baringo: the 1985 season. Nyame Akuma 29,36–41.
van Peer, P. (1991). Interassemblage variability andLevallois styles: the case of the Northern AfricanMiddle Paleolithic. J. Anthrop. Archaeol. 10,107–151.
van Peer, P. (1992). The Levallois Reduction Strategy.Monographs in World Prehistory 13. Madison, WI:Prehistory Press.
van Peer, P. (1998). The Nile corridor and the Out-of-Africa model: an examination of the archaeologicalrecord. Curr. Anthrop. 39, S115–S140.
van Peer, P., Vermeersch, P. M., Moeyersons, J. & VanNeer, W. (1996). Paleolithic sequences of SodmeinCave, Red Sea Mountains, Egypt. In (G. Pwiti &R. Soper, Eds) Aspects of African Archaeology,pp. 149–156. Harare: University of ZimbabwerPublishers.
van Vark, G. N. (1995). The study of hominid skeletalremains by means of statistical methods. In (N. T.Boaz & G. D. Wolfe, Eds) Biological Anthropology:The State of the Science, pp. 71–90. Corvallis: OregonState University Press.
Vandermeersch, B. (1981). Les Hommes Fossiles deQafzeh (Israel). Paris: CNRS.
Velo, J. (1984). Ochre as medicine: a suggestion for theinterpretation of the archaeological record. Curr.Anthrop. 25, 674.
Velo, J. (1986). The problem of ochre. MankindQuarterly 26, 229–237.
Vermeersch, P. M., Paulissen, P. E. & Van Peer, P.(1990). Palaeolithic chert exploitation in the lime-stone stretch of the Egyptian Nile Valley. Afr.Archaeol. Rev. 8, 77–102.
Vermeersch, P. M., Paulissen, E., Stokes, S., Charlier,C., Van Peer, P., Stringer, C. & Lindsay, W. (1998).Middle Palaeolithic burial of a modern human atTaramsa Hill, Egypt. Antiquity 72, 475–484.
Vigilant, L., Stoneking, M., Harpending, H., Hawkes,K. & Wilson, A. C. (1991). African populations andthe evolution of mitochondrial DNA. Science 253,1503–1507.
Vinnicombe, P. (1975). The ritual significance of eland(Taurotragus oryx) in the rock art of southern Africa.Les religions de la prehistoire: actes du Valcamonicasymposium, pp. 379–400. Capo di Ponte, 1972.
Vinnicombe, P. (1976). People of the Eland.Pietermaritzburg: Natal University Press.
Vishnyatsky, L. B. (1994). ‘Running ahead of time’ inthe development of Palaeolithic industries. Antiquity68, 134–140.
Vogel, J. C. & Beaumont, P. B. (1972). Revised radio-carbon chronology for the Stone Age in South Africa.Nature 237, 50–51.
Vogelsang, R. (1996). The Middle Stone Age in south-central Namibia. In (G. Pwiti & R. Soper, Eds)Aspects of African Archaeology, pp. 207–212. Harare:University of Zimbabwe Publications.
Vogelsang, R. (1998). Middle Stone Age Fundstellen inSudwest-Namibia. Koln: Heinrich—Barth Institut.
Voigt, E. A. (1982). The Molluscan fauna. In (R.Singer & J. Wymer, Eds) The Middle Stone Ageat Klasies River Mouth, pp. 155–186. Cambridge:Cambridge University Press.
Volman, T. P. (1978). Early evidence for shellfishcollecting. Science 201, 911–913.
Volman, R. P. (1984). Early prehistory of southernAfrica. In (R. G. Klein, Ed.) Southern African Prehis-tory and Paleoenvironments, pp. 169–220. Rotterdam:Balkema.
Vrba, E. S. (1982). Biostratigraphy and chronology,based particularly on Bovidae, of southern hominid-associated assemblages: Makapansgat, Sterkfontein,Taung, Kromdraai, Swartkrans; also Elandsfontein(Saldahna), Broken Hill (now Kabwe) and Cave ofHearths. In (M. A. de Lumley, Ed.) Homo erectus et laPlace de l’Homme de Tautavel parmi les HominidesFossiles. Premiere Congres International de PaleontologieHumaine, pp. 707–752. Vol. 2. UNESCO ColloqueInternational du Centre National de la RechercheScientifique: Pretirages.
Wadley, L. (1991). Rose Cottage Cave: backgroundand a preliminary report on the recent excavations. S.Afr. Archaeol. Bull. 46, 125–130.
Wadley, L. (1993). The Pleistocene Later Stone Age southof the Limpopo River. J. World. Prehist. 7, 243–296.
Wadley, L. (1995). Review of dated Stone Age sitesrecently excavated in the eastern Free State, SouthAfrica. S. Afr. J. Sci. 91, 574–579.
Wadley, L. (1997). Rose Cottage Cave: archaeologicalwork 1987 to 1997. S. Afr. J. Sci. 93, 439–444.
Wadley, L. & Harper, P. (1989). Rose Cottage Caverevisited: Malan’s Middle Stone Age collection. S.Afr. Archaeol. Bull. 44, 23–32.
Wadley, L. & Vogel, J. C. (1991). New dates from RoseCottage Cave, Ladybrand, eastern Orange FreeState. S. Afr. J. Sci. 87, 605–608.
Wainscoat, J. S., Hill, A. V. S., Boyce, A. L., Flint, J.,Hernandez, M., Thein, S. L., Old, J. M., Lynch,J. R., Falusi, A. G., Weatherall, D. J. & Clegg, J. B.(1986). Evolutionary relationships of humanpopulations from an analysis of nuclear DNApolymorphisms. Nature 319, 491–493.
Walker, A. C. (1993). Perspectives on the Nariokotomediscovery. In (A. C. Walker & R. E. Leakey, Eds) TheNariokotome Homo Erectus Skeleton, pp. 411–430.Cambridge, MA: Harvard University Press.
Walker, N. J. (1987). Dating of Zimbabwean rock art.Rock Art Res 4, 137–149.
Walker, N. J. (1990). Zimbabwe at 18,000 BP. In (O.Soffer & C. Gamble, Eds) The World at 18,000 BPVol. 2: Low Latitudes, pp. 206–213. London: UnwinHyman.

562 . . .
Walker, N. J. (1991). Excavation and re-examination ofsome stone age sites in the Matopos. Zimbabwe 3,5–22.
Walker, N. J. (1995). Late Pleistocene and HoloceneHunter-Gatherers of the Matopos: An ArchaeologicalStudy of Change and Continuity in Zimbabwe. Uppsala:Studies in African Archaeology 10.
Walter, R. C., Buffler, R. T., Bruggermann, J. H.,Guillauma, M. M. M., Berhe, S. M., Negassi, B.,Libsekal, Y., Cheng, H., Edwards, R. L., von Cosel,R., Neraudeau, D. & Gagnon, M. (2000). Earlyhuman occupation of the Red Sea coast of Eritreaduring the last interglacial. Nature 405, 65–69.
Walter, R. C., Manega, P. C., Hay, R. L., Drake, R. E.& Curtis, G. H. (1991). Laser fusion 40Ar/39Ardating of Bed I, Olduvai Gorge, Tanzania. Nature354, 145–149.
Walter, R. C., Manega, P. C. & Hay, R. L. (1992).Tephrochronology of Bed I, Olduvai Gorge: anapplication of laser-fusion Ar-Ar dating to calibrat-ing biological and climatic chnage. QuaternaryInternational 13/14, 37–46.
Wayland, E. J. & Burkitt, M. C. (1932). The Magosianculture of Uganda. J. R. Anthrop. Inst. 62, 369–390.
Wells, C. (1964). Bones, Bodies, and Disease. London:Thames & Hudson.
Wells, L. H. (1950). The Border Cave skull,Ingwavuma District, Zululand. Am. J. phys. Anthrop.8, 241–243.
Wells, L. H. (1959). The problem of Middle Stone AgeMan in South Africa. Man 59, 158–160.
Wendorf, F. & Schild, R. (1974). A Middle StoneAge Sequence from the Central Rift Valley, Ethiopia.Warsaw: Institute of the History of Material Culture,Polish Academy of Scienes.
Wendorf, F. & Schild, R. (1992). The MiddlePaleolithic of North Africa: a status report. In (F.Klees & R. Kuper, Eds) New Light on the NortheastAfrican Past, pp. 40–78. Koln: Heinrich-BarthInstitut.
Wendorf, F. & Schild, R. (1993). Probable function ofMousterian points and convergent sidescrapers in theMiddle Stone Age of Ethiopia. Quaternaria Nova 3,39–51.
Wendorf, F., Shiner, J. L. & Marks, A. E. (1965).Summary of the 1963–1964 season. In (F. Wendorf,Ed.) Contributions to the Prehistory of Nubia,pp. ix–xxxi. Dallas: Southern Methodist UniversityPress (Papers of the Fort Burgwin Research Center).
Wendorf, F., Laury, L., Albritton, C. C., Schild, R.,Haynes, C. V., Damon, P. E., Shafquillah, M. &Scarborough, R. (1975). Dates for the Middle StoneAge in East Africa. Science 187, 740–742.
Wendorf, F., Close, A. E. & Schild, R. (1987). Recentwork on the Middle Palaeolithic of the EasternSahara. Afr. Archaeol. Rev. 5, 49–63.
Wendorf, F., Close, A. E., Schild, R., Gautier, A.,Schwarcz, H. P., Miller, G. H., Kowalski, K., Krolik,H., Bluszcz, A., Robins, D. & Grun, R. (1990).Le dernier interglaciaire au Sahara orientale.L’Anthropologie 94, 361–391.
Wendorf, F., Schild, R. & Close, A. E. (Eds) (1993a).Egypt During the Last Interglacial. New York:Plenum.
Wendorf, F., Schild, R. & Close, A. E. (1993b). MiddlePalaeolithic occupations at Bir Tarfawi and BirSahara East, Western Desert of Egypt. In (L.Krzyzaniak, M. Kobusiewicz & J. Alexander, Eds)Environmental Change and Human Culture in theNile Basin and Northern Africa until the SecondMillennium B.C., pp. 103–112. Poznan: MuseumArcheologiczne W. Poznianu.
Wendorf, F., Close, A. E. & Schild, R. (1994). Africa inthe period of Homo sapiens neanderthalensis and con-temporaries. In (S. J. de Laet, A. H. Dani, J. L.Lorenzo & R. B. Nunoo, Eds) History of Humanity,Vol. 1: Prehistory and the Beginnings of Civilizationpp. 117–135. New York: Routledge & UNESCO.
Wendt, W. E. (1972). Preliminary report on anarchaeological research programme in South WestAfrica. Cimbebasia B 21, 1–42.
Wendt, W. E. (1975). ‘Art mobilier’ aus der Apollo11-Grotte in Sudwest-Afrika: die altesten datiertenKunstwerke Afrikas. Acta Praehist. Archaeol. 5,1–42.
Wendt, W. E. (1976). Art mobilier from the Apollo 11Cave, southwest Africa: Africa’s oldest dated worksof art. S. Afr. Archaeol. Bull. 31, 5–11.
Wengler, L. (1997). La transition du Mousterian al’Aterian. L’Anthropologie 101, 448–481.
Wengler, L. & Vernet, J.-L. (1992). Vegetation,sedimentary deposits, and climates during the latePleistocene and Holocene in eastern Morocco.Palaeogeogr., Palaeoclimatol., Palaeoecol. 94, 141–167.
White, R. (1982). Rethinking the Middle/UpperPaleolithic transition. Curr. Anthrop. 23, 169–192.
White, R. (1989a). Production complexity andstandardization in early Aurignacian bead and pen-dant manufacture: evolutionary implications. In (P.Mellars & C. B. Stringer, Eds) The Human Revol-ution: Behavioral and Biological Perspectives on theOrigins of Modern Humans, pp. 366–390. Edinburgh:Edinburgh University Press.
White, R. (1989b). Toward a contextual understandingof the earliest body ornaments. In (E. Trinkaus,Ed.) The Emergence of Modern Humans: BioculturalAdaptations in the Late Pleistocene, pp. 211–231.Cambridge: Cambridge University Press.
White, R. (1992). Beyond art: toward an understandingof the origins of material representations in Europe.A. Rev. Anthrop. 21, 537–564.
White, R. (1993). Technological and social dimensionsof Aurignacian age: body ornaments across Europe.In (H. Knecht, A. Pike-Tay & R. White, Eds) BeforeLascaux: The Complex Record of the Early UpperPaleolithic, pp. 277–300. Boca Raton: CRC Press.
White, T. D. (1986). Cutmarks on the Bodo cranium:a case of prehistoric defleshing. Am. J. phys. Anthrop.69, 503–509.
White, T. D. (1987). Cannibals at Klasies? Sagittarius2, 6–9.
White, T. D. (2000). Olduvai Gorge. In (E. Delson, I.Tattersall, J. A. Van Couvering & A. S. Brooks, Eds)

563 ’
Encyclopedia of Human Evolution and Prehistory, NewYork: Garland/Taylor & Francis.
Whitworth, T. (1966). A fossil hominid from Rudolf.S. Afr. Archaeol. Bull. 21, 138–150.
Wiens, J. A. (1977). On competition and variableenvironments. Am. Scient. 65, 592–597.
Wiessner, P. (1982). Risk, reciprocity and socialinfluences on !Kung San economics. In (E. Leacock& R. Lee, Eds) Politics and History in Band Societies,pp. 61–84. Cambridge: Cambridge UniversityPress.
Wiessner, P. (1983). Style and social informationin Kalahari San projectile points. Am. Antiq. 48,253–276.
Wiessner, P. (1986). !Kung San networks in agenerational perspective. In (M. Biesele, R. Gordon& R. Lee, Eds) The Past and Future of !Kung SanEthnography, pp. 103–136. Hamburg: Helmut BuskeVerlag Quellen zur Khoisan-Forschung, Band 4.
Wiessner, P. (1998). Leveling the hunter: Constraintson the status quest in foraging societies. In (P.Wiessner & W. Schiefenhovel, Eds) Food andthe Status Quest: An Interdisciplinary Perspective,pp. 171–191. Providence: Berghahn Books.
Williams, M. A. J. (1976). Radiocarbon dating andlate Quaternary Saharan climates: a disussion. Z.Geomorphol. 20, 361–362.
Williams, M. A. J. (1984). Late Quaternary prehistoricenvironments in the Sahara. In (J. D. Clark & S. A.Brandt, Eds) From Hunters to Farmers: The Causes andConsequences of Food Production in Africa, pp. 74–83.Berkeley: University of California Press.
Wilmsen, E. N. (1974). Lindenmeier: A PleistoceneHunting Society. New York: Harper & Row.
Wilson, D. C. (1994). Identification and assessment ofsecondary refuse aggregates. J. Archaeol. MethodTheory 1, 41–68.
Wilson, D. S. & Hey, R. N. (1981). The Galapagosaxial magnetic anomaly: evidence for the Emperorreversal within the Brunhes and for a two layermagnetic source. Geophys. Res. Lett. 8, 1051.
Wintle, A. (1996). Archaeologically relevant datingtechniques for the next century: small, hot, andidentified by acronyms. J. Archaeol. Sci. 23, 123–138.
Wobst, H. M. (1977). Stylistic behavior and infor-mation exchange. In (C. E. Cleland, Ed.) For theDirector: Research Essays in Honor of James B. Griffin,pp. 317–342. Ann Arbor: University of MichiganMuseum of Anthropology, Anthropological Papers61.
Wobst, M. (1983). Palaeolithic archaeology: someproblems with form, space, and time. In (G. Bailey,Ed.) Hunter-Gatherer Economy in Prehistory: AEuropean Perspective, pp. 220–225. Cambridge:Cambridge University Press.
Wood, B. (1991). Koobi Fora Research Project, Volume 4:Human Cranial Remains. Oxford: Oxford UniversityPress.
Wood, B. (1992). Origin and evolution of the genusHomo. Nature 355, 783–790.
Wood, B. (1994). Taxonomy and evolutionaryrelationships of Homo erectus. Cour. Forsch-Inst.Senckenberg 171, 159–165.
Wood, B. & Collard, M. (1999). The changing face ofgenus Homo. Evol. Anthrop. 8, 195–207.
Wood, B. & Van Noten, F. (1986). Preliminaryobservations on the BK 8518 mandible fromBaringo, Kenya. Am. J.phys. Anthrop. 69, 117–127.
Woodward, A. S. (1921). A new cave man fromRhodesia, South Africa. Nature 108, 371–372.
Worsaae, J. J. A. (1849). The Primeval Antiquities ofDenmark, trans. W. J. Thomas. London: Parker.
Wreschner, E. E. (1985). Evidence and interpretationof red ochre in the early prehistoric sequences. In(P. V. Tobias, Ed.) Hominid Evolution: Past, Presentand Future, pp. 387–394. New York: Alan R. Liss.
Wurz, S. (1997). The Howiesons Poort at KlasiesRiver: From artefacts to cognition. M. A. thesis,University of Stellenbosch.
Wurz, S. (1999). The Howiesons Poort backed artifactsfrom Klasies River: an argument for symbolicbehavior. S. Afr. Archaeol. Bull. 54, 38–50.
Wynn, T. (1979). The intelligence of later Acheulianhominids. Man 14, 371–391.
Wynn, T. (1985). Piaget, stone tools, and the evolutionof human intelligence. World Archaeol. 17, 32–43.
Yearsley, M. (1928). The pathology of the left temporalbone of the Rhodesian skull. In (W. P. Pycraft, G. E.Smith, M. Yearsley, J. T. Carter, R. A. Smith, A. T.Hopwood, D. M. A. Bate & W. E. Swinton, Eds)Rhodesian Man and Associated Remains, pp. 59–63.London: British Museum (Natural History).
Yellen, J. E. (1977). Archaeological Approaches to thePresent: Models for Interpreting the Past. New York:Academic Press.
Yellen, J. E. (1986). Optimization and risk in humanforaging strategies. J. hum. Evol. 15, 733–750.
Yellen, J. E. (1991). Small mammals: post-discardpatterning of !Kung San faunal remains. J. Anthrop.Archaeol. 10, 152–192.
Yellen, J. E. (1996). Behavioural and taphonomicpatterning at Katanda 9: a Middle Stone Age site,Kivu Province, Zaire. J. Archaeol. Sci. 6, 915–932.
Yellen, J. E. (1998). Barbed bone points: tradition andcontinuity in Saharan and sub-Saharan Africa. Afr.Archaeol. Rev. 15, 173–198.
Yellen, J. E., Brooks, A. S., Cornelissen, E., Mehlman,M. H. & Stewart, K. (1995). A Middle Stone Ageworked bone industry from Katanda, Upper SemlikiValley, Zaire. Science 268, 553–556.
Zietkiweicz, E., Yotova, V., Jarnik, M., Korab-Laskowska, M., Kidd, K. K., Modiano, D., Scozzari,R., Stoneking, M., Tishkoff, S., Batzer, M. &Labuda, D. (1997). Nuclear DNA diversity inworldwide distributed human populations. Gene 205,161–171.
Zilhao, J. & d’Errico, F. (1998). The chronologyand taphonomy of the earliest Aurignacian and itsimplications for the understanding of Neanderthalextinction. J. World. Prehist. 13, 1–68.