the rat there is no access to the ventricles. carbachol therefore acts
TRANSCRIPT

J. Physiol. (1984), 354, pp. 523-545 523With 10 text-fgurmPrinted in Great Britain
A CHOLINERGIC LINK IN THE REFLEX RELEASE OF VASOPRESSINBY HYPOTENSION IN THE RAT
BY G. W. BISSET AND H. S. CHOWDREYFrom the Department of Pharmacology, St. Thomas's Hospital Medical School,
London SEl 7EH
(Received 27 January 1984)
SUMMARY
1. Inhalation of amyl nitrite in the water-loaded rat under ethanol anaesthesiaproduced a brief fall of blood pressure followed by a prolonged antidiuretic response.
2. The antidiuretic response to amyl nitrite was accompanied by increased urinaryexcretion of vasopressin, it was blocked by a specific vasopressin antagonist and bya barbiturate and it was absent in the Brattleboro rat with congenital diabetesinsipidus. These results show that the antidiuretic response to the hypotensioninduced by amyl nitrite is due to the release of vasopressin and that this release ismediated by a neuroendocrine reflex acting through the brain stem.
3. Carbachol and nicotine produced an antidiuretic response on injection into alateral cerebral ventricle (i.vent.). Carbachol was almost ineffective, but nicotinemuch more effective, when injected into the cisterna magna (i.cist.) from which inthe rat there is no access to the ventricles. Carbachol therefore acts at a site reachedfrom the ventricles, possibly the paraventricular nucleus. Nicotine acts at a moredistal site reached from the subarachnoid space. This site may correspond with thenicotine-sensitive area on the ventral surface of the brain stem which has beendescribed in the cat.
4. Atropine blocked the antidiuretic response to carbachol but not that to amylnitrite. Hexamethonium blocked the antidiuretic response to amyl nitrite as well asthat to nicotine and was more effective on i.cist. than i.vent. injection.
5. These results reveal a cholinergic link with a nicotinic but not a muscarinicreceptor in the neural pathways controlling the release of vasopressin in response tohypotension.
6. A hypothetical model is presented in which the release of vasopressin isstimulated by a pathway arising from chemoreceptors and inhibited by a secondpathway arising from stretch- and baroreceptors. Hypotension acts by suppressingthe normally predominant inhibitory pathway and stimulating the excitatorypathway. Hexamethonium is presumed to block transmission at a synapse in theexcitatory pathway at the ventral surface or, less probably, at the paraventricularand supraoptic nuclei.

G. W. BISSET AND H. S. CHOWDREY
INTRODUCTION
Stimulation or inhibition of the release of vasopressin from the neurohypophysisin response to changes in blood volume or blood pressure involves neuroendocrinereflexes acting through the hypothalamus, analogous to the milk-ejection reflex whichcontrols the release of oxytocin in response to suckling (Bisset, 1968; Bisset & Jones,1975; Harris, 1978). The idea that the secretion of vasopressin may be controlled bya change in blood volume without a change in plasma osmolality arose from anobservation by Gauer and his co-workers that negative pressure breathing in the doginduced a diuresis (Gauer, Henry, Sieker & Wendt, 1954). They argued that theincrease in intrathoracic blood volume resulting from this manoeuvre was monitoredby stretch receptors in the left atrium. These receptors are innervated by the vagi.Inflating a balloon in the left atrium increased afferent vagal activity causing diuresiswhereas haemorrhage reduced vagal activity causing antidiuresis (Henry, Gauer &Reeves, 1956; Gauer & Henry, 1963). Although the diuretic response to inflation ofa balloon in the left atrium has been the subject of much controversy (Goetz, Bond& Bloxham, 1975) recent work (Ledsome, Wilson & Ngsee, 1982; Fater, Schutz,Sundet, Mapes & Goetz, 1982) has confirmed earlier observations (Baisset &Montastruc, 1957; Johnson, Moore & Segar, 1969; Brennan, Malvin, Jochim &Roberts, 1971) that it is associated with a decrease in the circulating plasma levelof vasopressin. These results imply that afferent nerve impulses in the vagi exert atonic inhibitory control over vasopressin release. In fact, section of the vagi resultsin an increase in the concentration of vasopressin in plasma (Share & Levy, 1962).An inhibitory control is exerted also through baroreceptors in the carotid sinus andthe aortic arch with afferents in the carotid sinus nerves and vagi (Share & Levy,1962, 1966b; Kumada & Sagawa, 1970; Share, 1974). Conversely, excitation ofchemoreceptors by anoxia or reduced blood flow through the carotid and aortic bodiesstimulates the release of vasopressin (Share & Levy, 1966a). Stimuli such ashaemorrhage, carotid occlusion and hypotension appear to cause release ofvasopressinby decreasing the inhibitory drive from stretch receptors and baroreceptors and byincreasing the excitatory drive from chemoreceptors. Release by all three stimuli isinhibited or abolished by section of the vagi and carotid sinus nerves (Clark & Rochae Silva, 1967; Kanjanapothi, 1975).The primary sensory afferents from the peripheral receptors run in the carotid sinus
nerves and vagi to the nucleus of the tractus solitarius (n.t.s.) on the dorsal surfaceof the brain stem (Spyer, 1982). There must be an efferent projection, involving oneor more synapses, from the n.t.s. to the neurones in the supraoptic nucleus (s.o.n.)and paraventricular nucleus (p.v.n.) which secrete vasopressin. Our aim is to identifytransmitters at these synapses and, if possible, to gain some information about theirlocation and the pathways involved. In this paper we demonstrate a cholinergic linkinvolving a nicotinic receptor in reflexes controlling the release of vasopressin. Theevidence is based on the ability of hexamethonium injected into a lateral cerebralventricle (i.vent.) or the cisterna magna (i.cist.) to block the antidiuretic response tohypotension induced by the inhalation of amyl nitrite in the water-loaded rat underethanol anaesthesia. A preliminary account of this work has been published (Bisset& Chowdrey, 1981).
524

REFLEX RELEASE OF VASOPRESSIN 525
METHODS
The water-loaded-rat preparationThis was basically the preparation which had been developed for the bioassay of antidiuretic
activity (Dicker, 1953; Bisset, 1962; Clark & Rocha e Silva, 1967). Wistar rats from the SPF colonyat St. Thomas's Hospital Medical School were used. A male rat weighing 250 g was given bystomach tube 12-5 ml 12% (v/v) ethanol in warm tap water, followed 30 min later by 7-5 ml water.Full surgical anaesthesia was obtained by a brief inhalation of ether. A femoral artery and anexternal jugular vein were cannulated with Polythene tubing of 0-4 mm i.d. (Portex), the tracheawith 1-67 mm i.d. tubing and the bladder, through a suprapubic incision, with 1-77 mm i.d. tubing.The rat was then turned into a prone position and the head was supported in a horizontal planewith cotton-wool but not inserted into a head holder. The skin was excised in the mid line fromthe eyes to the level ofthe ears to expose the bregma. A C313 gauge guide cannula (Plastic ProductsCo. Roanoke, VA) was inserted into a lateral ventricle 1-5 mm lateral to the saggital suture and1 mm posterior to the bregma. A C3131 internal cannula was inserted into the guide cannula withthe tip 4-5 mm from the surface of the skull. Correct positioning was indicated by a free flow offluid from a syringe connected with the internal cannula by a short length of 0-4 mm i.d. Polythenetubing. The skin incision was extended to the neck and the overlying muscles were retracted toexpose the atlanto-occipital membrane. The cisterna magna was cannulated with the shaft of a 26gauge hypodermic needle to which a Polythene bleb was fixed 2 mm from the tip. The membranewas pierced with the needle tip and the cannula advanced until the bleb rested on the membrane;it was then held in place by allowing the retracted muscles to resume their natural position. Correctpositioning was indicated by a free flow of c.s.f. from the cannula. A short length of 0-4 mm i.d.Polythene tubing was attached to the end of the cannula and closed with a pin. The cisternal andventricular cannulae were held in position with dental cement adhering to an anchoring screw inthe skull.With the cannulae in position, the rat was placed on a pad of cotton-wool on one pan of a pair
of laboratory scales in a semi-prone position with the head supported horizontally. The rat wasbalanced by a counterweight in the other pan of the scales and a constant fluid load equivalentto 8% of the body weight was maintained by the infusion into the jugular vein of a mixturecontaining 2-3 ml ethanol, 0-18 g sodium chloride and 2-2 g glucose in 100 ml distilled water. Theinfusion was controlled automatically by a negative feed-back system operating a servo-controlledpump (Clark & Rocha e Silva, 1967). The bladder cannula was connected with a flask containing50% (v/v) methanol which was displaced through a 26 gauge hypodermic needle. Urine flow wasmeasured by a phototransistor connected with a staircase integrator having a 1 min time base anddisplayed on a potentiometric recorder. Blood pressure was recorded by a strain gauge transducerand potentiometric recorder. The output from the amplifier connected with the recorder was fedinto an integrator designed for the quantitative measurement of uterine activity (Bisset, Haldar& Lewin, 1966). This integration effectively measured the area enclosed by a hypotensive response,thus taking into account both the amplitude and duration. Both i.vent. and i.cist. injections weremade in a volume of 1-4 ,1 by means of a Hamilton syringe connected by Polythene tubing to thecannulae. For i.cist. injections allowance was made for the dead space in the system which wasmeasured at the beginning of the experiment. Drug remaining in the dead space after an injectionwas removed by allowing an escape of c.s.f. All drugs were dissolved in 0-9% (w/v) NaCl solution.Amyl nitrite was administered from a crushed ampoule by holding the open end over the tracheal
cannula and allowing the rat to inhale the vapour for such a period of time, usually not exceeding30 s, as would produce the required amplitude and duration of hypotension.
Measurement of urinary excretion of vas(pressinFor this purpose, the flask containing methanol was discarded and the bladder cannula connected
directly with the hypodermic needle by a short length of telescoped Polythene tubing. Urine wascollected from the needle and urine flow recorded in the usual manner. A sample of 5 ml urine wascollected either during a control period or immediately following the inhalation of amyl nitrite orinjection of a drug. The sample was acidified by addition of 0-5 ml 1 N-HCl. The vasopressin wasextracted from the urine by a modification of the method of Moran, Miltenberger, Shuayb &Zimmerman (1964) according to the following scheme.
525

0. W. BISSET AND H. S. CHOWDREY
(1) Adjust the pH of the sample to 5-55 using 1 N-HCl and 2 N-Na2CO3.(2) Add 400 mg Amberlite resin CG-50 (H) (BDH) and shake for 30 min. Centrifuge for 5 min
and discard supernatant.(3) Wash residue twice with 10 ml distilled water and then twice with 50% (v/v) ethanol, both
at pH 5, shaking suspensions with a Whirlimix and discarding supernatants after centrifuging.(4) Resuspend residue in 10 ml 75% (v/v) ethanol at pH 1-5 and shake for 30 min. Centrifuge
for 10 min.(5) Evaporate supernatant from (4) to dryness under vacuum at 55 0C and take up residue in
05-1 0 ml 09% (w/v) NaCI solution.Centrifuging was carried out at 680 g for stages (2) and (3) and at 2750 g for stage (4).
Shaking the urine with resin was a quicker procedure than using column chromatography.Washing the residues twice with 50% ethanol and with water was found to remove or reduce theconcentration of an unidentified hypotensive and diuretic factor which was encountered whentesting concentrated extracts containing little or no detectable vasopressin. Vasopressin was
estimated in the extracts by bioassay using the basic water-loaded-rat preparation (Bisset, Clark& Errington, 1971). Extracts were injected i.v., and whenever possible a four-point assay designwas used with vasopressin (Pitressin) as standard. In every experiment in which urine samples were
collected a known amount of vasopressin was added to 5 ml control urine and the recoveryestimated in the extract. In thirty-five experiments the mean percentage recovery of addedhormone was 84+12 (s.E. of mean). All results quoted have been corrected for loss of hormone on
extraction.
Material8The drugs used were amyl nitrite (Crushable Glass Capsules: Macarthys), nicotine (BDH),
carbachol chloride (Carbachol injection, B.P.: Evans Medical), muscarine chloride (Sigma),hexamethonium bromide (Sigma), atropine sulphate (Macarthys): atropine methyl nitrate (Sigma),vasopressin (Pitressin: Parke Davis), the synthetic vasopressin antagonist (d-(CH2),Tyr(Et)VAVP(Sawyer, Pang, Seto, McEnroe, Lammek & Manning, 1981), oxytocin (Syntocinon: Sandoz) andthiobutobarbitone (5-ethyl-5-(1-methyl-propyl)-2-thiobarbiturate sodium: Inactin-Byk). Doses ofdrugs used as salts are expressed in terms of the salt. The trypsin used was bovine type XI (Sigma).
RESULTS
Antidiuretic response to amyl nitrite
A brief inhalation of amyl nitrite produced an abrupt fall of blood pressure anda reduction of urine flow. The hypotensive response usually lasted for only 1-3 min
but the antidiuretic response was prolonged for 20-30 min (Figs. 2, 3, 7 and 8). Witha moderate antidiuretic response (e.g. A1 in Fig. 3, A1 and A2 in Fig. 7) two phaseswere discernible. The first was concomitant with, or slightly outlasted, the fall ofbloodpressure. This was followed by the second phase in which the urine flow graduallyrecovered to the control value over a period of about 20 min, the profile of theresponse resembling the recovery from the antidiuretic response to an i.V. injectionof vasopressin. These two phases were sometimes demarcated by a transient increasein urine flow which accompanied an overshoot of the blood pressure and was probablydue to the release of catecholamines by a baroreceptor reflex. With a largerantidiuretic response (e.g. A1 in Fig. 2, A3 in Fig. 7 and A1 in Fig. 8) the two phasesmerged. The first phase of the antidiuretic response was attributed to a renovasculareffect resulting from the fall of blood pressure and the second phase to a reflex releaseof vasopressin from the neurohypophysis. Consistent with this interpretation is thefact that only the first phase was apparent in the Brattleboro rat with congenitaldiabetes insipidus (Fig. 1), the fall of blood pressure producing a profound reductionin urine flow which returned to the control level within 5 min. For quantitative
526
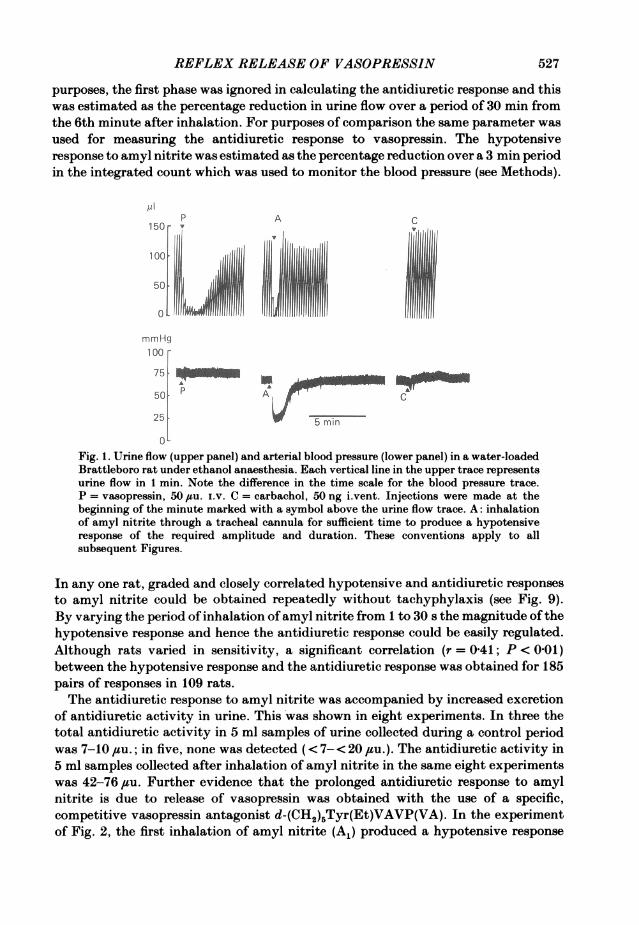
REFLEX RELEASE OF VASOPRESSIN
purposes, the first phase was ignored in calculating the antidiuretic response and thiswas estimated as the percentage reduction in urine flow over a period of 30 min fromthe 6th minute after inhalation. For purposes of comparison the same parameter wasused for measuring the antidiuretic response to vasopressin. The hypotensiveresponse to amyl nitrite was estimated as the percentage reduction over a 3 min periodin the integrated count which was used to monitor the blood pressure (see Methods).
/1PA
150 -
100,,50jmmHg100
7 5 WPA t50. A
25 5 min
0-
Fig. 1. Urine flow (upper panel) and arterial blood pressure (lower panel) in a water-loadedBrattleboro rat under ethanol anaesthesia. Each vertical line in the upper trace representsurine flow in 1 min. Note the difference in the time scale for the blood pressure trace.P = vasopressin, 50 ,uu. i.v. C = carbachol, 50 ng i.vent. Injections were made at thebeginning of the minute marked with a symbol above the urine flow trace. A: inhalationof amyl nitrite through a tracheal cannula for sufficient time to produce a hypotensiveresponse of the required amplitude and duration. These conventions apply to allsubsequent Figures.
In any one rat, graded and closely correlated hypotensive and antidiuretic responsesto amyl nitrite could be obtained repeatedly without tachyphylaxis (see Fig. 9).By varying the period of inhalation ofamyl nitrite from 1 to 30 s the magnitude ofthehypotensive response and hence the antidiuretic response could be easily regulated.Although rats varied in sensitivity, a significant correlation (r = 0-41; P < 0-01)between the hypotensive response and the antidiuretic response was obtained for 185pairs of responses in 109 rats.The antidiuretic response to amyl nitrite was accompanied by increased excretion
of antidiuretic activity in urine. This was shown in eight experiments. In three thetotal antidiuretic activity in 5 ml samples of urine collected during a control periodwas 7-10 ,uu.; in five, none was detected (< 7-< 20 ,su.). The antidiuretic activity in5 ml samples collected after inhalation of amyl nitrite in the same eight experimentswas 42-76 ,su. Further evidence that the prolonged antidiuretic response to amylnitrite is due to release of vasopressin was obtained with the use of a specific,competitive vasopressin antagonist d-(CH2)5Tyr(Et)VAVP(VA). In the experimentof Fig. 2, the first inhalation of amyl nitrite (A1) produced a hypotensive response
527
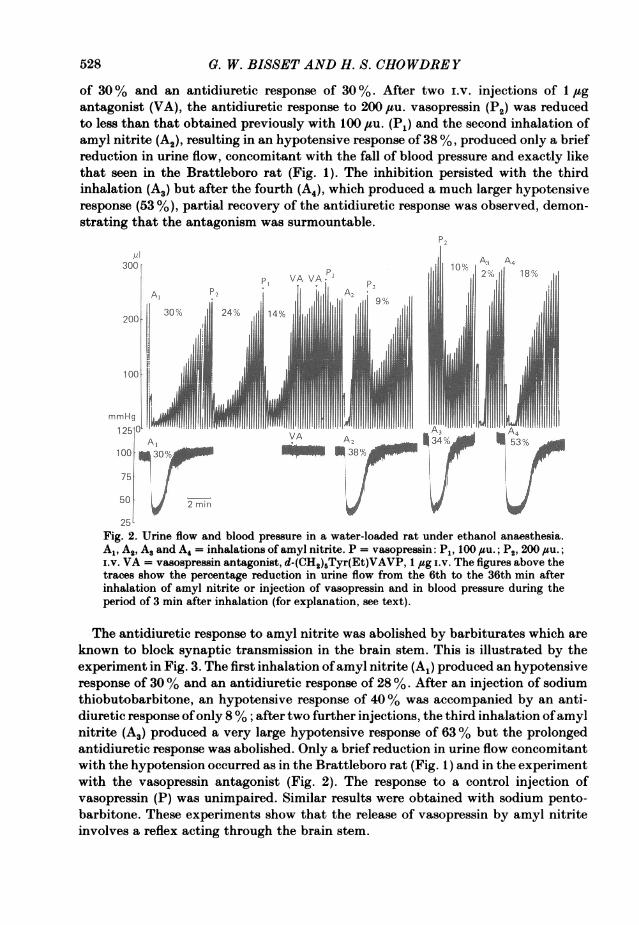
5. W. BISSET AND H. S. CHOWDREY
of 30% and an antidiuretic response of 30 %. After two i.v. injections of 1 jigantagonist (VA), the antidiuretic response to 200 ,tu. vasopressin (P2) was reducedto less than that obtained previously with 100 ,tu. (P1) and the second inhalation ofamyl nitrite (A2), resulting in an hypotensive response of 38 %, produced only a briefreduction in urine flow, concomitant with the fall of blood pressure and exactly likethat seen in the Brattleboro rat (Fig. 1). The inhibition persisted with the thirdinhalation (A3) but after the fourth (A4), which produced a much larger hypotensiveresponse (53 %), partial recovery of the antidiuretic response was observed, demon-strating that the antagonism was surmountable.
P2
ml ~~~~~~~~~~~~~A3 A4300 10%2 8
PI VA VA.
20030% 24% 14%
200
mmHg I
125 0 VA IA4
50:~~~~~~~V 2 53OXn100 0/ 8
75
25Fig. 2. Urine flow and blood pressure in a water-loaded rat under ethanol anaesthesia.Al, A2, A. and A4 = inhalations of amyl nitrite. P = vasopressin: P1, 100 #uu.; P2, 200 1su.;i.v. VA = vasopressin antagonist, d-(CH2)rTyr(Et)VAVP, 1 jug i.v. The figures above thetraces show the percentage reduction in urine flow from the 6th to the 36th min afterinhalation of amyl nitrite or injection of vasopressin and in blood pressure during theperiod of 3 min after inhalation (for explanation, see text).
The antidiuretic response to amyl nitrite was abolished by barbiturates which areknown to block synaptic transmission in the brain stem. This is illustrated by theexperiment in Fig. 3. The first inhalation ofamyl nitrite (A,) produced an hypotensiveresponse of 30% and an antidiuretic response of 28 %. After an injection of sodiumthiobutobarbitone, an hypotensive response of 40% was accompanied by an anti-diuretic response ofonly 8% ; after two further injections, the third inhalation ofamylnitrite (A3) produced a very large hypotensive response of 63% but the prolongedantidiuretic response was abolished. Only a brief reduction in urine flow concomitantwith the hypotension occurred as in the Brattleboro rat (Fig. 1) and in the experimentwith the vasopressin antagonist (Fig. 2). The response to a control injection ofvasopressin (P) was unimpaired. Similar results were obtained with sodium pento-barbitone. These experiments show that the release of vasopressin by amyl nitriteinvolves a reflex acting through the brain stem.
528

REFLEX RELEASE OF VASOPRESSIN52
u1175 A3
150 AlP A
100
50
mmHg125 30%
100 AA6 3%'
75 A3
50 2 min
Fig. 3. Urine flow and blood pressure in a water-loaded rat under ethanol anaesthesia.Al, A and A3 = inhalations of amyl nitrite. P = vasopressin 50 #tsu. i.v. Sodium thiobuto-barbitone 2-5 mg was injected i.v. 20 min before A2 and two further doses of 5 mg eachwere injected 60 and 15 min before A3. Percentage figures as in legend to Fig. 2.
Cholinoceptor agonist8Experiments were carried out first with carbachol because it is a stable choline ester
possessing both muscarinic and nicotinic actions and it had been shown by Kuhn(1974) to produce an antidiuretic response on i.vent. injection in the rat. In fifty-sevenexperiments, the injection of 5-80 ng carbachol i.vent. produced a profound anti-diuretic response and a small pressor response. The intensity and duration of theantidiuretic response were related to the dose, as illustrated for doses of 5 and 10 ngin Fig. 5. The onset of the antidiuretic response was delayed until the 3rd or 4thminute after injection, the maximum effect was reached in 10-15 min and theduration varied from 20 min with a threshold dose of 5 ng to 1 h or more with largerdoses. The pressor response consisted of a gradual rise in pressure of 10-20 mmHgwhich was immediate in onset and lasted 5-10 min.
In the experiment of Fig. 4A in which 40 ng carbachol i.vent. produced a largeantidiuretic response and a pressor response, a control i.vent. injection of 4 #sl 0 9%NaCl solution produced no effect on urine flow or blood pressure. Injections of40-320 ng carbachol i.v. caused graded falls of blood pressure with no effect on urineflow. Doses of 640 and 1280 ng produced falls exceeding 40 mmHg and smallantidiuretic responses which were attributed to a reflex release of vasopressin inresponse to hypotension as observed with amyl nitrite. These experiments showedthat the antidiuretic and pressor responses to carbachol were not an artifact ofinjection and were mediated centrally, not after absorption into the peripheralcirculation. In ten experiments, the effectiveness of an iLvent. injection was comparedwith that of an i.cist. injection. The threshold dose i.vent. varied from 5-25 ng andi.cist. from 200 ng to 2 jug. A typical experiment is illustrated in Fig. 4B: 1 jug i.cist.produced a much smaller antidiuretic response than 10 ng i.vent. with a delayed onsetof action. This series of experiments demonstrated that the action of carbachol is ona structure reached from the cerebral ventricles and not from the subarachnoid space.That the antidiuretic response to carbachol i.vent. is due to the release of
529
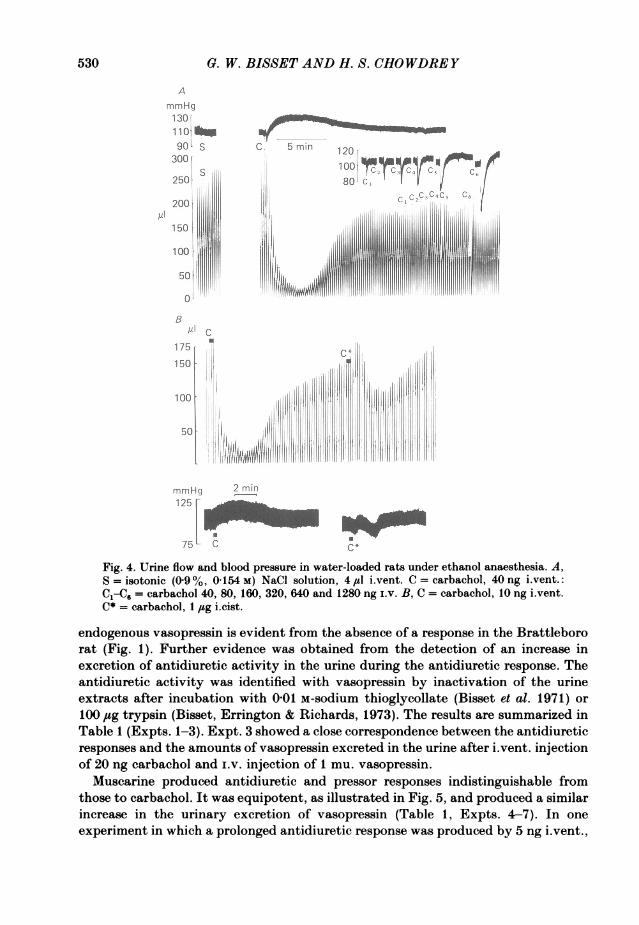
0. W. BISSET AND H. S. CHOWDREY
AmmHg13011o10|90L S C 5 min 120
300' 100 "C'250 80. cX
ACC
175
150 1,
100 'II I
50 11 1111 lilil II I11l
50
rnmHq ?2 m1125
C.
Fig. 4. Urine flow and blood pressure in water-loaded rats under ethanol anaesthesia. A,S =isotonic (09%, 0154 M) NaCI solution, 4,sl i~vent. C =carbachol, 40 ng i.vent.:
C1C= carbachol 40, 80, 160, 320, 640 and 1280 ng I.v. B, C = carbachol, 10 ng i.vent.C*= carbachol, 1 jug i.cist.
endogenous vasopressin is evident from the absence of a response in the Brattlebororat (Fig. 1). Further evidence was obtained from the detection of an increase inexcretion of antidiuretic activity in the urine during the antidiuretic response. Theantidiuretic activity was identified with vasopressin by inactivation of the urineextracts after incubation with 001 M-sodium thioglycollate (Bisset et al. 1971) or100 ,usg trypsin (Bisset, Errington &; Richards, 1973). The results are summarized inTable 1 (Expts. 1-3). Expt. 3 showed a close correspondence between the antidiureticresponses and the amounts of vasopressin excreted in the urine after i.vent. injectionof 20 ng carbachol and I.v. injection of 1 mu. vasopressin.
Muscarine produced antidiuretic and pressor responses indistinguishable fromthose to carbachol. It was equipotent, as illustrated in Fig. 5, and produced a similarincrease in the urinary excretion of vasopressin (Table 1, Expts. 4-7). In oneexperiment in which a prolonged antidiuretic response was produced by 5 ng i.vent.,
530

REFLEX RELEASE OF VASOPRESSIN
no response was obtained after 1 jug i.v. As with carbachol, i.cist. injections were atleast 100 times less effective than i.vent. injections: this finding was obtained in fiveexperiments.
Acetylcholine produced graded antidiuretic responses related to the dose in eightexperiments in which the dose was systematically doubled within the range
TABLE 1. Excretion of vasopressin in 5 ml samples of urine collected during a control period andduring the antidiuretic response to i.vent. injections of drugs
Mean rate of urine flow (,uil/min)
Expt. Drug1 Control
Carbachol 40 ng2 Control
Carbachol 50 ngNicotine 320 jug
3 ControlCarbachol 20 ngVasopressin 1 mu. i.v.
4 ControlMuscarine 40 ng
5 ControlMuscarine 40 ngAtropine 4 jug+ muscarine 40 ng
6 ControlMuscarine 40 ngMuscarine 40 ngAtropine 4 jug+ muscarine 40 ng
7 ControlMuscarine 40 ng
Atropine 4,ug+ muscarine 40 ng
Before During Reductioncollection collection (%)
140 140 0163 51 69195 200 +2-6156 57 63181 63 65172 172 0159 65 59155 69 55159 204 +22220 122 46174 200 +15246 91 63174 179 +2-9117 154 +32144 86 40126 91 28144 167 +16112 132 +18154 71 54111 111 0
Atropine injected 1-2 min before carbachol or muscarine.
0-625-10 #sg. The antidiuretic response was of shorter duration than with carbacholand muscarine and was potentiated by neostigmine 50 sg i.vent. In two experimentsin. which 1 or 2 ,ug was effective i.vent., the threshold dose i.cist. was 50 ,ug and inone experiment in which 0-625 ,sg i.vent. produced an antidiuretic response, none was
obtained with 80 ,ug i.cist. No antidiuretic response was obtained with i.V. injectionsof doses which were effective i.vent.
In contrast with carbachol, muscarine and acetylcholine, nicotine was as effectivein producing an antidiuretic response by i.v. as by i.vent. injection. The thresholddose by both routes was 50 ,tsg. Antidiuretic responses were obtained with this dosei.v. in five experiments and with doses of 50-160 #sg i.vent. in six experiments. Theantidiuretic response to a larger dose of 320 ,sg i.vent. in one further experiment wasaccompanied by a large increase in the excretion of vasopressin in the urine (Table 1,Expt. 2) comparable with that observed in the same experiment after a dose ofcarbachol (50 ng) which caused an almost identical reduction in the rate of urine flow.Antidiuretic responses to i.v. and i.vent. injections were accompanied by a rise ofblood pressure (Figs. 5 and 6). In Fig. 5 i.vent. injections of carbachol 10 ng and
Vasopressin(duu.) in 5 ml
urine
16135<28131299516
115104<787
<20307<20<25267270<34<34292<34
531
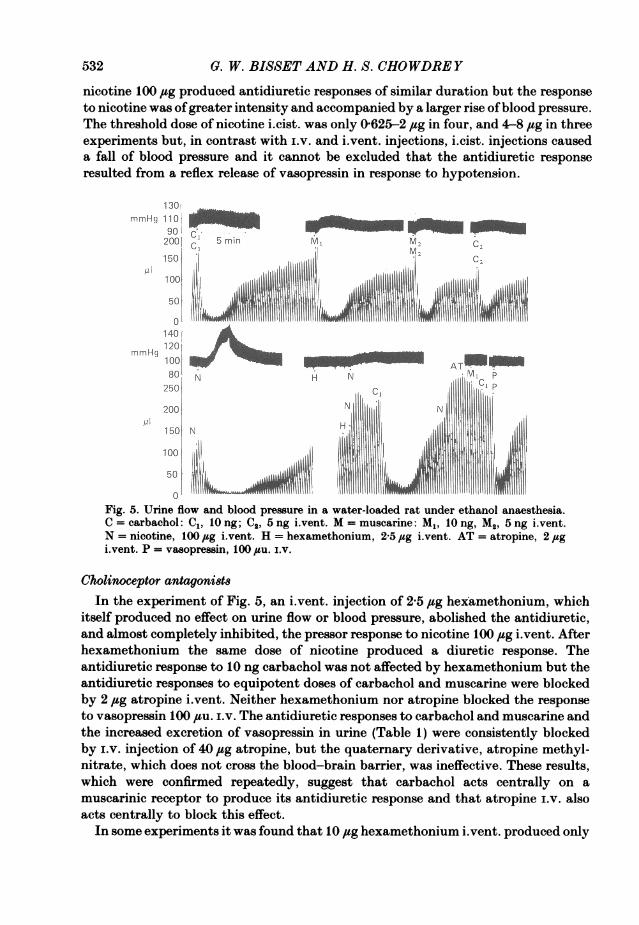
0. W. BISSET AND H. S. CHOWDREY
nicotine 100 ,sg produced antidiuretic responses of similar duration but the responseto nicotine was ofgreater intensity and accompanied by a larger rise ofblood pressure.The threshold dose of nicotine i.cist. was only 0625-2 ,ug in four, and 4-8 ,tg in threeexperiments but, in contrast with i.v. and i.vent. injections, i.cist. injections causeda fall of blood pressure and it cannot be excluded that the antidiuretic responseresulted from a reflex release of vasopressin in response to hypotension.
130immHg 110 jI_
902001 5C m Ml M C21540 C2
50' A101401
mmHg 120A T
250i c=1j200 NN
Fig. 5. Urine flow and blood pressure in a water-loaded rat under ethanol anaesthesia.a cal compleC1, ly ng; C2,5 ngihvent. M = muscarine:Mt ,iotng, M2, 5ngi.vent.b nicotine, 100 #g vent. H = hexamethonium, 25 jsg ivent. AT = atropine, 2 spgto vent. P = vasopressin, 100 su. ir.a
Cholinoceptor antagonio8iIn the experiment of Fig. 5, an ivent. injection of 245 bgheramethonium, which
itself produced no effect on urine flow or blood pressure, abolished the antidiuretic,and almost completely inhibited, the pressor response to nicotine 100,sg vent. Afterhexamethonium the same dose of nicotine produced a diuretic response. Theantidiuretic response to 10 ng carbachol was not affected by hexamethonium but theantidiuretic responses to equipotent doses of carbachol and muscarine were blockedby 2 #tg atropine iLvent. Neither hexamethonium nor atropine blocked the responseto vasopressin 100 ,su. ixv. The antidiuretic responses to carbachol and muscarine andthe increased excretion of vasopressin in urine (Table 1) were consistently blockedby ixv. injection of 40 ,sg atropine, but the quaternary derivative, atropine methyl-nitrate, which does not cross the blood-brain barrier, was ineffective. These results,which were confirmed repeatedly, suggest that carbachol acts centrally on amuscarinic receptor to produce its antidiuretic response and that atropine i.v. alsoacts centrally to block this effect.
In some experiments it was found that 10 jisg hexamethonium i.vent. produced only
532
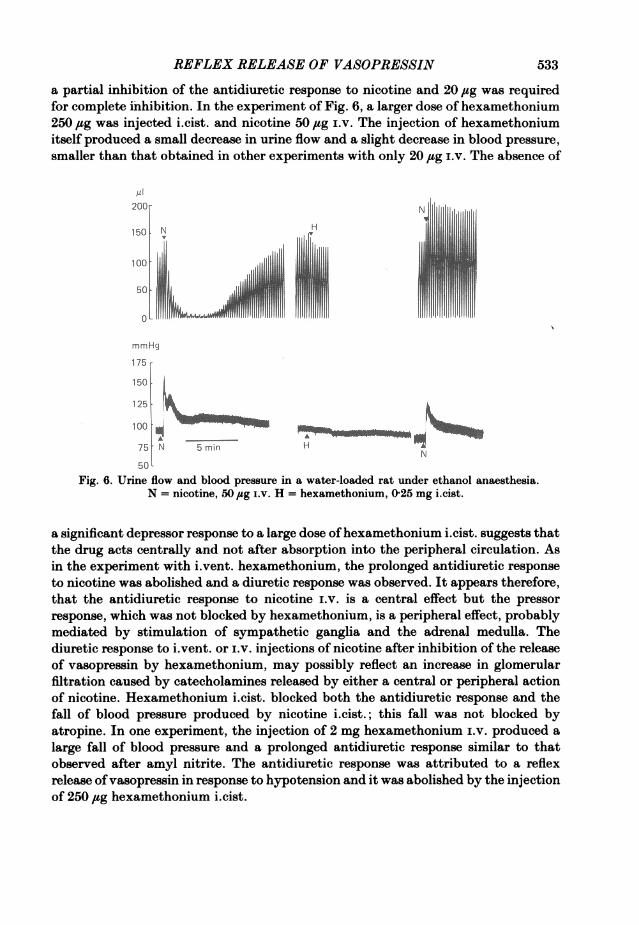
REFLEX RELEASE OF VASOPRESSIN
a partial inhibition of the antidiuretic response to nicotine and 20 jug was requiredfor complete inhibition. In the experiment of Fig. 6, a larger dose of hexamethonium250,g was injected i.cist. and nicotine 50 ,sg i.v. The injection of hexamethoniumitself produced a small decrease in urine flow and a slight decrease in blood pressure,smaller than that obtained in other experiments with only 20 jg i.v. The absence of
200 -N
150. N H10
50 ~j3llmmHg
175
125
100
75 N 5 min HN
50LFig. 6. Urine flow and blood pressure in a water-loaded rat under ethanol anaesthesia.
N = nicotine, 50 ,ug i.v. H = hexamethonium, 025 mg icist.
a significant depressor response to a large dose ofhexamethonium i.cist. suggests thatthe drug acts centrally and not after absorption into the peripheral circulation. Asin the experiment with i.vent. hexamethonium, the prolonged antidiuretic responseto nicotine was abolished and a diuretic response was observed. It appears therefore,that the antidiuretic response to nicotine i.v. is a central effect but the pressorresponse, which was not blocked by hexamethonium, is a peripheral effect, probablymediated by stimulation of sympathetic ganglia and the adrenal medulla. Thediuretic response to i.vent. or i.v. injections of nicotine after inhibition of the releaseof vasopressin by hexamethonium, may possibly reflect an increase in glomerularfiltration caused by catecholamines released by either a central or peripheral actionof nicotine. Hexamethonium i.cist. blocked both the antidiuretic response and thefall of blood pressure produced by nicotine i.cist.; this fall was not blocked byatropine. In one experiment, the injection of 2 mg hexamethonium i.v. produced alarge fall of blood pressure and a prolonged antidiuretic response similar to thatobserved after amyl nitrite. The antidiuretic response was attributed to a reflexrelease ofvasopressin in response to hypotension and it was abolished by the injectionof 250 jug hexamethonium i.cist.
533
G. W. BISSET AND H. S. CHOWDREY
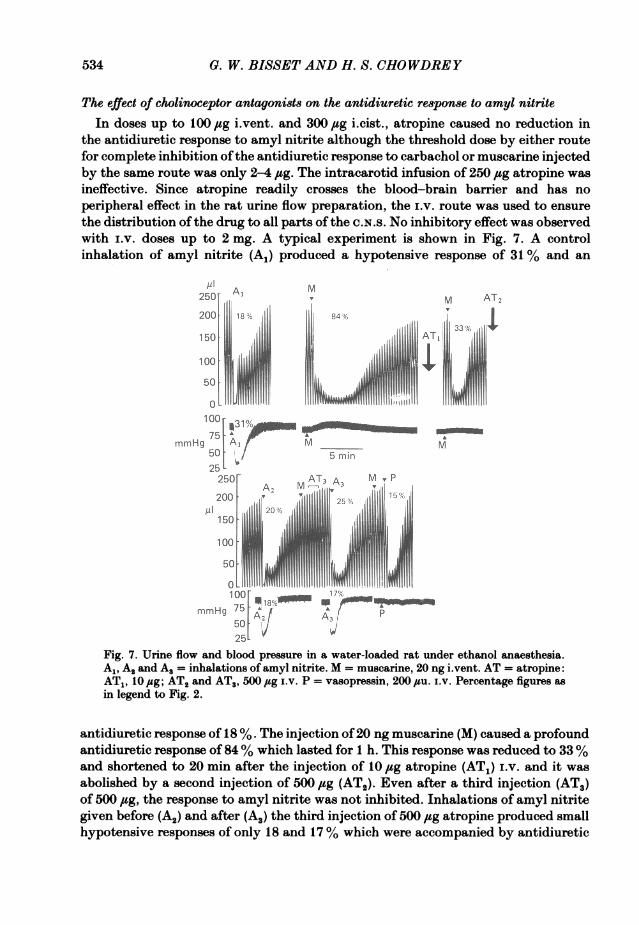
The effect of cholinoceptor antagoni8ts on the antidiuretic response to amyl nitriteIn doses up to 100 jag i.vent. and 300 /zg i.cist., atropine caused no reduction in
the antidiuretic response to amyl nitrite although the threshold dose by either routefor complete inhibition ofthe antidiuretic response to carbachol or muscarine injectedby the same route was only 2-4 ,ug. The intracarotid infusion of 250 jug atropine wasineffective. Since atropine readily crosses the blood-brain barrier and has noperipheral effect in the rat urine flow preparation, the i.v. route was used to ensurethe distribution ofthe drug to all parts of the C.N.s. No inhibitory effect was observedwith i.v. doses up to 2 mg. A typical experiment is shown in Fig. 7. A controlinhalation of amyl nitrite (AJ) produced a hypotensive response of 31 % and an
A1250
200
150
MAl
18 %
100 fd
50100 ,31
7Sl A00 S 2
100 - .
mmHg Al50
25
250
A,
200-
100
mmHg 751-50[A
84
AT
M5 min
AT3 A M, P
325 " 1111||||||1 III
Fig. 7. Urine flow and blood pressure in a water-loaded rat under ethanol anaesthesia.A,, A, and A. = inhalations of amyl nitrite. M = muscarine, 20 ng i.vent. AT = atropine:AT1, 10 jug; AT2 and AT., 500 ,ug iv. P = vasopressin, 200 ,u. iV. Percentage figures as
in legend to Fig. 2.
antidiuretic response of 18 %. The injection of20 ng muscarine (M) caused a profoundantidiuretic response of 84% which lasted for 1 h. This response was reduced to 33%and shortened to 20 min after the injection of 10 fig atropine (AT1) i.v. and it wasabolished by a second injection of 500 ,ug (ATO). Even after a third injection (AT3)of 500 ,ug, the response to amyl nitrite was not inhibited. Inhalations of amyl nitritegiven before (A2) and after (A.) the third injection of 500 jug atropine produced smallhypotensive responses of only 18 and 17% which were accompanied by antidiuretic
M AT21
M
534
[.lfo
17% NWlpA, p
A3
REFLEX RELEASE OF VASOPRESSIN 535
responses of 20 and 25 %, larger than in the control (A1). The response to vasopressinwas not inhibited. Similar results were obtained in two other experiments. Somedegree of inhibition was observed in two experiments in which very high doses of 4and 4-8 mg were administered.
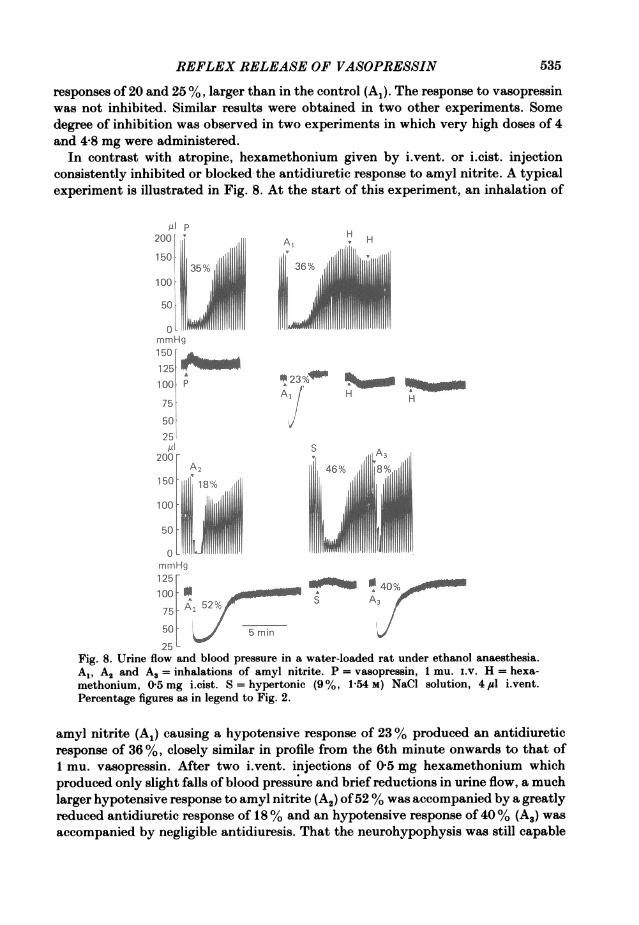
In contrast with atropine, hexamethonium given by i.vent. or i.cist. injectionconsistently inhibited or blocked the antidiuretic response to amyl nitrite. A typicalexperiment is illustrated in Fig. 8. At the start of this experiment, an inhalation of
pI P200 THA150 'ilIlll
|135% f 36%
50
mmHg150
252001,iA2~~~~~~~
150L 18
100oh
mmHg125F_____
75 A252% S 3 /
50 So 5 mm25
Fig. 8. Urine flow and blood pressure in a water-loaded rat under ethanol anaesthesia.A1, A2 and A3 = inhalations of amyl nitrite. P = vasopressin, 1 mu. i.v. H = hexa-methonium, O 5 mg i.cist. S = hypertonic (9%, 1s54 M) NaCi solution, 4 4m i.vent.Percentage figures as in legend to Fig. 2.
amyl nitrite (A1) causing a hypotensive response of 23 %/ produced an antidiureticresponse of 36%,/ closely similar in profile from the 6th minute onwards to that of1 mu. vasopressin. After two i.vent. injections of 05 mg hexamethonium whichproduced only slight falls of blood pressure and brief reductions in urine flow, a muchlarger hypotensive response to amyl nitrite (A2) of52 %/ was accompanied by a greatlyreduced antidiuretic response of 18 %/ and an hypotensive response of 40 %/ (A3) wasaccompanied by negligible antidiuresis. That the neurohypophysis was still capable

G. W. BISSET AND H. S. CHOWDREY
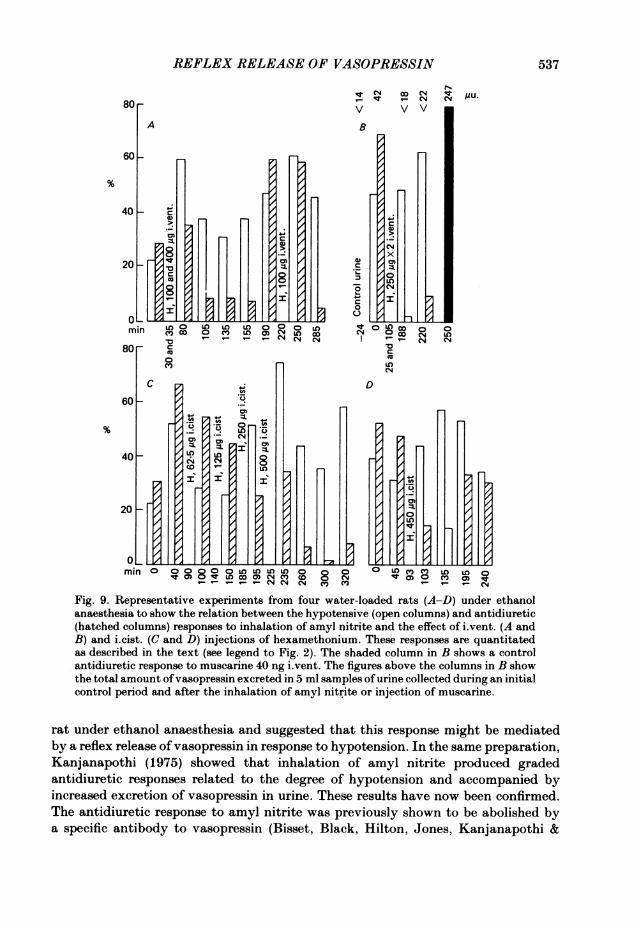
of responding to stimulation was shown by a large antidiuretic response to i.vent.injection of 41il hypertonic (1P54 M) NaCl solution. Similar results were obtained inten other experiments with i.vent. (see also Fig. 1, Bisset & Chowdrey, 1981) andtwelve other experiments with i.cist. injections of hexamethonium in doses of25-500 ,ug. In five experiments, i.v. injections of 0-5-2 mg had no effect on theantidiuretic response to amyl nitrite. A striking difference between the routes ofinjection was that i.cist. injections of hexamethonium were fully effective within10 min whereas no inhibition was observed until at least 15 min after i.vent. injectionand the maximum effect by this route was not reached until 50 min after the injection.The degree of inhibition was related to the dose of hexamethonium. The inhibitionlasted for at least 1 h and could be surmounted by applying a larger hypotensivestimulus. These results are illustrated by the four representative experiments inFig. 9 in which the corresponding hypotensive and antidiuretic responses to eachinhalation are represented by a pair ofcolumns. An inhibitory effect ofhexamethoniumis indicated by a disproportionately small antidiuretic response. Fig. 9A and B showstwo experiments with i.vent. injection and C and D two with i.cist. injections.Fig. 9A shows that 45 min after the i.vent. injection of 500 ,ug hexamethonium, theresponse to amyl nitrite was inhibited and this inhibition persisted for a further periodof 75 min; 145 min after the injection a larger hypotensive stimulus surmounted theblock. A further injection of 100 #sg hexamethonium produced no block after 30 minbut a substantial inhibition 65 min later. In the experiment of Fig. 9B two doses of250 ,sg hexamethonium practically abolished the antidiuretic response to a largehypotensive stimulus 83 and 115 min after the second dose although a largerantidiuretic response was still obtained with muscarine. In this experiment, vaso-pressin was estimated in 5 ml samples of urine. After the first inhalation of amylnitrite the total amount in 5 ml was 42 ,Ctu., compared with < 14 Ftu. in the control;after the second and third inhalations when the antidiuretic response to amyl nitritehad been inhibited by hexamethonium, no antidiuretic activity was detected(<22 ,uu.) but after muscarine the total excretion was 247 ,uu. Fig. 9C illustratesgraded antidiuretic responses to amyl nitrite and the effects of increasing doses ofhexamethonium i.cist. The threshold for inhibition was 250 ,tg. The inhibition wasapparent 10 min after the injection. After 500 ,ug inhibition was obtained for thefollowing 95 min. Graded antidiuretic responses to amyl nitrite are again illustratedin Fig. 9D. A dose of 450 ,ug hexamethonium caused inhibition in 10 min withpartial recovery after 100 min.
DISCUSSION
It has been demonstrated repeatedly by a variety of experimental techniques thatneurosecretory cells in the p.v.n. and s.o.n. possess receptors for a number of putativeneurotransmitters. There is, however, no convincing evidence that any of thesetransmitters serves a physiological function in the neural control of release ofvasopressin. In this paper, the role of acetylcholine as a transmitter has beeninvestigated by testing the ability ofcholinomimetic substances to release vasopressinfrom the neurohypophysis and of specific antagonists to block release in response toa physiological stimulus. The stimulus selected was hypotension. Bisset & Lewis(1962) demonstrated an antidiuretic response to vasodilator drugs in the water-loaded
536
REFLEX RELEASE OF VASOPRESSIN
min
80F
N-qq co CN4 I /AU.(-NT N
V V V
-o0
0V
C
60k
40
20
U?
(0
LL L~1
MAq
cm
. I
B
0
0
(N)
CN4/x
CD0:
/0
D
min 0 000 Ln uL)U)W08 0 OU nL0) LnC000 V 0CqO CV 0
Fig. 9. Representative experiments from four water-loaded rats (A-D) under ethanol
anaesthesia to show the relation between the hypotensive (open columns) and antidiuretic
(hatched columns) responses to inhalation of amyl nitrite and the effect of iLvent. (A and
B) and i.cist. (C and D) injections of hexamethonium. These responses are quantitatedas described in the text (see legend to Fig. 2). The shaded column in B shows a control
antidiuretic response to muscarine 40 ng iLvent. The figures above the columns in B show
the total amount of vasopressin excreted in 5 ml samples of urine collected during an initial
control period and after the inhalation of amyl nitrite or injection of muscarine.
rat under ethanol anaesthesia and suggested that this response might be mediated
by a reflex release of vasopressin in response to hypotension. In the same preparation,
Kanjanapothi (1975) showed that inhalation of amyl nitrite produced graded
antidiuretic responses related to the degree of hypotension and accompanied byincreased excretion of vasopressin in urine. These results have now been confirmed.
The antidiuretic response to amyl nitrite was previously shown to be abolished by
a specific antibody to vasopressin (Bisset, Black, Hilton, Jones, Kanjanapothi &
537
I .-. - .-

0. W. BISSET AND H. S. CHOWDREY
Montgomery, 1974). In this paper inhibition of the antidiuretic response by a specificcompetitive antagonist of vasopressin (Sawyer et al. 1981) and the absence of aresponse in the Brattleboro rat with congenital diabetes insipidus provide furtherevidence that the antidiuretic response is mediated by the release of vasopressin. Ithas been shown also that the antidiuretic response, like the milk-ejection responseto suckling, is blocked by barbiturates. This suggests that the release is mediated bya reflex acting through the brain stem. The afferent limb consists of fibres runningin the carotid sinus nerves and vagi from peripheral receptors in the cardiovascularsystem to the nucleus of the tractus solitarius (n.t.s.). It is unlikely that acetylcholinewould act as a transmitter at primary afferent sensory nerve endings in the n.t.s.,for which substance P and L-glutamate are more probable candidates (Gillis, Helke,Hamilton, Norman & Jacobowitz, 1980; Reis, Perrone & Talman, 1981). Thecholinomimetic drugs and antagonists tested in this investigation may therefore beassumed to act at one or more synapses in an efferent pathway from the n.t.s. to thep.v.n. and s.o.n.The present experiments confirmed Kuhn's original observation (Kuhn, 1974) that
i.vent. injections ofnanogram amounts of carbachol in the water-loaded rat producedan antidiuretic response which is blocked by atropine. Atropine also blocked theantidiuretic response to acetylcholine and to muscarine which was equipotent withcarbachol. The antidiuretic response to i.vent. injections of acetylcholine in thewater-loaded goat (Vandeputte-van Messom & Peeters, 1975) and the dog (Bhargava,Kulshrestha & Srivastava, 1972) is similarly inhibited by atropine. That theantidiuretic response to carbachol in our experiments was due to release ofvasopressinis evident from the fact that it was accompanied by increased excretion of vasopressinin urine which was blocked by atropine and the absence of an antidiuretic responsein the Brattleboro rat. Since the pressor response to carbachol was absent orattenuated in the Brattleboro rat this too may be mediated, at least partly, by releaseof vasopressin. Evidence was obtained that carbachol, acetylcholine and muscarinecause release of vasopressin by acting on a muscarinic receptor at some site reachedfrom the cerebral ventricles. An obvious site is the p.v.n. Microionophoretic applicationof acetylcholine to the p.v.n. in rabbits caused an increase in unit electrical activitywhich in some cases was blocked by atropine (Moss, Urban & Cross, 1972). Thissuggests the presence of a muscarinic receptor on the neurosecretory cells in the p.v.n.although the possibility cannot be excluded that carbachol might act on aperiventricular structure with an afferent projection to the p.v.n. Whatever the siteof the muscarinic receptor, it is clearly not involved in the physiological release ofvasopressin in response to hypotension since atropine in large doses when injectedi.V. to reach all parts of the C.N.S. failed to inhibit the antidiuretic response to amylnitrite.Most of the experimental evidence indicates that the receptor on neurosecretory
cells mediating release ofvasopressin is nicotinic. Experiments with microionophoreticapplication of acetylcholine or nicotine to the s.o.n. or p.v.n. suggest excitation bya nicotinic receptor (Barker, Crayton & Nicoll, 1971; Moss et al. 1972; Dreifuss &Kelly, 1972; Arnauld, Cirino, Layton & Renaud, 1983) and inhibition by a muscarinicreceptor (Barker et al. 197 1 ; Nicoll & Barker, 197 1). Direct application ofacetylcholineor nicotine to explants of the hypothalamoneurohypophysial system containing the
538

REFLEX RELEASE OF VASOPRESSIN
s.o.n. but not the p.v.n. (Sladek & Knigge, 1977; Sladek & Joynt, 1979) or to theisolated organ-cultured s.o.n. (Sakai, Marks, George & Koestner, 1974) also indicatesan excitatory effect through nicotinic, and an inhibitory effect through muscarinic,receptors. In rat hypothalamic slices, excitatory, synaptically mediated responses toelectrical stimulation in the dorsolateral region were blocked by hexamethonium butnot affected by atropine and application of acetylcholine in the s.o.n. region bymicroperfusion mimicked the effect of electrical stimulation (Hatton, Ho & Mason,1983).In our experiments, nicotine, in striking contrast with carbachol, was equally
effective in producing an antidiuretic response when injected i.v. and i.vent. and muchmore effective i.cist. Since there is no foramen ofMagendie in the rat (Feldberg, 1976),drugs injected i.cist. cannot reach the ventricles although drugs injected i.vent. canreadily reach the ventral surface of the brain stem through the lateral foramina ofthe 4th ventricle. It is unlikely, therefore, that nicotine produced an antidiureticresponse by acting directly on the p.v.n. through the ependyma of the 3rd ventricleand probable that it acted at some site reached from the subarachnoid space. Thisconclusion is entirely consistent with results obtained in the cat (Bisset, Feldberg,Guertzenstein & Rocha e Silva, 1975; Castro de Souza & Rocha e Silva, 1977).Nicotine injected into a lateral ventricle in the cat (Bisset et al. 1975) caused release ofvasopressin without oxytocin. Stimulation of the p.v.n. would be expected to releaseboth hormones (Bisset et al. 1971; Bisset et al. 1973). After cannulation of the cerebralaqueduct no vasopressin was released. This indicated that nicotine had to reach thesubarachnoid space in order to act and it was effective i.cist. The site of action wasfinally located by topical application to a circumscribed area on the ventral surfaceof the brain stem lateral to the pyramids and 6-9 mm caudal to the trapezoid bodies.It has been suggested that this area might act as a relay station within the excitatorypathway from the n.t.s. Topical application of y-aminobutyric acid (GABA) to thisarea blocked the release of vasopressin in response to carotid occlusion (Feldberg &Rocha e Silva, 1981) and topical application of horseradish peroxidase revealed anefferent projection from the n.t.s. (Errington & Dashwood, 1979). It is significant alsothat topical application of nicotine to this area, like haemorrhage, carotid occlusionand hypotension (Beleslin, Bisset, Haldar & Polak, 1967; Clark & Rocha e Silva, 1967;Kanjanapothi, 1975), produced release of vasopressin independently of oxytocin. Fortechnical reasons it has not been possible to establish the presence ofa nicotine-sensitivearea in the rat corresponding with that in the cat, but such an area would be a likelysite of action for nicotine. By a central action, hexamethonium blocked theantidiuretic response to nicotine and, unlike atropine, it also blocked the antidiureticresponse to amyl nitrite. That the antidiuretic response to muscarine and hypertonicsaline was uninhibited in the same experiments, excludes the possibility of anon-specific depression of the neurosecretory cells in the p.v.n. or s.o.n. Hexa-methonium was effective both i.vent. and i.cist. but when injected i.cist. it acted witha shorter latent period. This suggests that a longer time was required after i.vent.injection for the drug to penetrate to its site of action. These results are consistentwith a block by hexamethonium at a synapse between efferent fibres from the n.t.s.and cells in the nicotine-sensitive area on the ventral surface of the brain stem.Possibly there is a second cholinergic projection from this zone to those cells in the
539

G. W. BISSET AND H. S. CHOWDREY
s.o.n. and p.v.n. which preferentially secrete vasopressin and that the receptors atthis synapse are also nicotinic. It is conceivable that hexamethonium reaching theventral surface from either the lateral ventricle or the cisterna magna could trackalong the base of the brain to reach the s.o.n. However, if both the s.o.n. and p.v.n.are involved in the reflex release, it would then have to penetrate also to the p.v.n.and it might be expected that i.vent. injections would reach both nuclei more readilythan i.cist. injections. The results therefore seem more compatible with an action ofhexamethonium at a relay station on the ventral surface which projects to both nuclei.An alternative pathway from the n.t.s. to the p.v.n. and s.o.n. involving the ventral
surface has been revealed by the immunocytochemical studies of Sawchenko &Swanson (1982). They demonstrated a non-noradrenergic projection from the n.t.s.to the Al group ofnoradrenergic neurones on the ventral surface which in turn projectto cells in the s.o.n. and p.v.n. preferentially secreting vasopressin. This couldtherefore represent an excitatory pathway for the release ofvasopressin independentlyof oxytocin in response to changes in blood volume or pressure. If the projection fromthe n.t.s. were cholinergic, the synapse at the Al cells could be blocked byhexamethonium. However, if this does represent an excitatory pathway it followsthat noradrenaline released from the terminals ofAl neurones in the s.o.n. and p.v.n.stimulates the release of vasopressin. The experimental evidence on whethernoradrenaline and adrenaline are excitatory or inhibitory at the s.o.n. and p.v.n. ismost conflicting. An antidiuretic response to i.vent. injection of noradrenaline hasbeen observed in the goat (Olsson, 1970; Vandeputte-van Messom et al. 1975), therat (Kuhn, 1974; Bridges, Hillhouse & Jones, 1976) and the dog (Bhargava et al. 1972).Microinjection of noradrenaline into the s.o.n. caused an increased concentration ofvasopressin in the blood of the cat (Milton & Paterson, 1974) and the rat (Urano &Kobayashi, 1978). After central depletion of catecholamines by 6-hydroxydopamine,dextran-induced hypovolaemia produced a smaller increase in plasma osmolality andin the vasopressin concentration than in controls (Miller, Handelman, Arnold,McDonald, Molinoff & Schrier, 1979). These observations accord an excitatory roleto noradrenaline probably involving a-adrenoceptors. Other evidence does notsupport such a conclusion. O'Connor & Verney (1945) showed that i.v. adrenalineor sympathetic stimulation inhibited the antidiuretic response to emotional stress indogs and Abrahams & Pickford (1956) demonstrated that adrenaline inhibited theantidiuretic response to acetylcholine when both drugs were injected into the carotidartery in dogs. Adrenaline produced a diuretic response and decrease in plasmavasopressin in the dog by an action on fl-receptors (Bhargava et al. 1972).Microionophoretic application of noradrenaline to neurosecretory cells in the p.v.n.(Moss et al. 1972) and s.o.n. (Barker et at. 1971; Arnauld et al. 1983) producedinhibition. The direct application of noradrenaline to cultured supraoptic cellsinhibited the increase in firing frequency produced by nicotine by an action onfl-receptors (Sakai et al. 1974) whereas it inhibited release of vasopressin byacetylcholine in hypothalamoneurohypophysial explants by an action on a-receptors(Armstrong, Sladek & Sladek, 1982). Two recent observations are of particularrelevance to the pathway from the n.t.s. Kimura, Share, Wang & Crofton (1981)showed that noradrenaline and adrenaline injected i.vent. in dogs caused a decreasein the plasma concentration ofvasopressin despite a fall in blood pressure which would
540

REFLEX RELEASE OF VASOPRESSIN
be expected to cause an increase. This suggests that noradrenaline has an inhibitoryeffect on the reflex release of vasopressin by hypotension. Consistent with this findingis the observation that application of kainic acid to the area of the ventral surfacecontaining Al cells or electrolytic lesions in this region cause a substantial increasein blood pressure and in the plasma vasopressin concentration (Blessing, Sved & Reis,1982). On balance, the evidence is in favour of an inhibitory role for noradrenaline.In conclusion, it seems possible that two projections might exist from the n.t.s.,
one an inhibitory pathway involving Al cells and the other an excitatory pathwayinvolving the nicotine-sensitive area on the ventral surface which has been demon-strated in the cat. These pathways are depicted in Fig. 10 which is intended to serve
S.o.n. and p.v.n. Ventral surface Sub-nuclei Carotid sinusof brain stem in n.t.s. nerves (IX) and
Al group vagi (X)r-A-_--- - - ~Stretch andg NA; isO- >~~~~~~5 rid | T ( ~~~A baroreceptors
adrenoeptor ,? Sub. P or:Vasopressin> L;. ,secreting , ACh '(Notc ?.;r B
Chemoreceptors(Nicotinic :(Nicotinic Sub. Por:receptor) I receptor) L-Glu.I
Nicotine-sensitivearea
Fig. 10. Hypothetical model of neural pathways, A inhibiting, and B stimulating, therelease of vasopressin in response to changes of blood volume and blood pressure. Thedashed line represents a projection from the nicotine-sensitive area on the ventral surfaceto the supraoptic nucleus (s.o.n.) and paraventrical nucleus (p.v.n.) which has not beendelineated anatomically. Putative neurotransmitters are indicated at excitatory (+) andinhibitory (-) synapses in the two pathways. ACh = acetylcholine. Sub.P = substanceP. L-Glu. = L-glutamate. NA = noradrenaline. N.t.s. = nucleus of the tractus solitarius.Al group = Al group of noradrenergic neurones (cf. Fig. 14 in Sawchenko & Swanson,1982).
as a working hypothesis. The only connexion which has not been demonstratedanatomically is the cholinergic projection from the nicotine-sensitive area on theventral surface to nicotinic receptors on neurosecretory cells in the p.v.n. and s.o.n.preferentially secreting vasopressin. Electrophysiological evidence indicates thatafferent impulses from baro-, stretch and chemoreceptors are excitatory to the n.t.s.(Spyer, 1982). Since section of the vagi and carotid sinus nerves causes a rise in theplasma concentration of vasopressin, the inhibitory pathway A must normally bepredominant, suppressing activity in supraoptic and paraventricular neurones. Thisactivity cannot be maintained by the excitatory pathway B which would beinterrupted as well as pathway A by section of the vagi and carotid sinus nerves. Theactivity in the neurosecretory cells may therefore be spontaneous or generated byan excitatory input other than that from chemoreceptors. In rat hypothalamic slices,paraventricular neurones displayed phasic bursting activity, typical of cells secretingvasopressin, in the absence of synaptic transmission (Hatton, 1982). Phasic activityof supraoptic neurones in rat hypothalamic slices was eliminated by synapticblockade (Hatton et al. 1983). A likely source of an excitatory input generatingactivity in vasopressin secreting cells is from osmoreceptors. Under our experimental
541

G. W. BISSET AND H. S. CHOWDREY
conditions, hypervolaemia resulting from the initial water load in the alcohol-anaesthetized rat would produce a strong inhibitory drive through pathway A, andhypoosmolality would remove any excitatory input from osmoreceptors, somaintaining a diuresis. Hypotension caused by amyl nitrite would reduce the activityof the baro- and stretch receptors and annul the inhibitory drive through pathwayA. However, in the absence of spontaneous or evoked activity in the neurosecretorycells in the p.v.n. and s.o.n., disinhibition alone would not result in release ofvasopressin. The release which does occur in response to hypotension must thereforebe produced predominantly by excitation of chemoreceptors and stimulation of thep.v.n. and s.o.n. through pathway B, unopposed by inhibition through pathway A.Hexamethonium could be presumed to inhibit release by blocking nicotinic receptorsin the excitatory pathway B at synapses in the nicotine-sensitive area or, lessprobably, in the s.o.n. and p.v.n. The scheme in Fig. 10 is concerned with neural inputsto magnocellular neurosecretory cells in the p.v.n. and s.o.n. Interconnexions betweenthe parvocellular neurones in the p.v.n., the n.t.s., the locus coeruleus and the Algroup of noradrenergic neurones could provide a system for the integration ofneuroendocrine and cardiovascular control (Sawchenko & Swanson, 1981; Spyer,1982).
It is interesting that hexamethonium has been shown to block the milk-ejectionreflex which appears also to involve a cholinergic link with nicotinic receptors (Clarke,Fall, Lincoln & Merrick, 1978). However, since stimuli such as haemorrhage, carotidocclusion and hypotension which change the blood volume or blood pressure causerelease of vasopressin without oxytocin (Beleslin et al. 1967; Clark & Rocha e Silva,1967; Kanjanapothi, 1975) andsuckling causes releaseofoxytocinwithout vasopressin,(Bisset, Clark & Haldar, 1970), it must be concluded that separate neurosecretorypathways exist for the independent release of the two hormones.
We are grateful to Dr M. Manning for a gift of the vasopressin antagonist, to ProfessorK. M. Spyer for helpful consultation during the preparation of this paper and to Mrs J. Andrewsfor typing the manuscript.
REFERENCES
ABRAHAMS, V. C. & PICKFORD, M. (1956). Observations on a central antagonism between adrenalineand acetylcholine. Journal of Physiology 131, 712-718.
ARMSTRONG, W. E., SLADEK, C. L. & SLADEK JR, J. R. (1982). Characterisation of noradrenergiccontrol of vasopressin release by the organ-cultured rat hypothalamoneurohypophysial system.Endrocrinology 111, 273-279.
ARNAULD, ELISABETH, CIRINO, MARIA, LAYTON, B. S. & RENAUD, L. P. (1983). Contrasting actionsof amino acids, acetylcholine, noradrenaline and leucine-enkephalin on the excitability ofsupraoptic vasopressin-secreting neurons. Neuroendocrinology 36, 187-196.
BAISSET, A. & MONTASTRUC, P. (1957). Polyurie par distension auriculaire chez le chien: role del'hormone antidiuretique. Journal de physiologie 49, 33-36.
BARKER, J. L., CRAYTON, J. W. & NICOLL, R. A. (1971). Noradrenaline and acetylcholine responsesof supraoptic neurosecretory cells. Journal of Physiology 218, 19-32.
BELESLIN, D., BISSET, G. W., HALDAR, J. & POLAK, R. L. (1967). The release ofvasopressin withoutoxytocin in response to haemorrhage. Proceedings of the Royal Society B 166, 443-458.
BHARGAVA, K. P., KULSHRESTHA, V. K. & SRIVASTAVA, Y. P. (1972). Central cholinergic andadrenergic mechanisms in the release of antidiuretic hormone. British Journal of Pharmacology44, 617-627.
BISSET, G. W. (1962). Effect of tyrosinase preparations on oxytocin, vasopressin and bradykinin.British Journal of Pharmacology and Chemotherapy 18, 405-420.
542

REFLEX RELEASE OF VASOPRESSIN
BISSET, G. W. (1968). The milk-ejection reflex and the actions of oxytocin, vasopressin andsynthetic analogues on the mammary gland. In Handbook of Experimental Physiology, vol. XXIII,ed. BERDE, B., pp. 475-544. Berlin: Springer-Verlag.
BISSET, G. W., BLACK, ANGELA J., HILTON, P. J., JONES, N. F., KANJANAPOTHI, DUANGTA &MONTGOMERY, MARGARET C. (1974). An antibody to vasopressin in man. Journal of Physiology239, 57-58P.
BISSET, G. W. & CHOWDREY, H. S. (1981). A central cholinergic link in the neural control of therelease of vasopressin. British Journal of Pharmacology 74, 239P.
BISSET, G. W., CLARK, BARBARA J. & ERRINGTON, M. L. (1971). The hypothalamic neurosecretorypathways for the release of oxytocin and vasopressin in the cat. Journal of Physiology 217,111-131.
BISSET, G. W., CLARK, BARBARA J. & HALDAR, JAYA (1970). Blood levels of oxytocin andvasopressin during suckling in the rabbit and the problem of their independent release. Journalof Physiology 206, 711-722.
BISSET, G. W., ERRINGTON, M. L. & RICHARDS, C. D. (1973). The distribution of vasopressin andoxytocin in the hypothalamoneurohypophysial system of the guinea-pig. British Journal ofPharmacology 48, 263-272.
BISSET, G. W., FELDBERG, W., GUERTZENSTEIN, P. G. & ROCHA E SILVA JR, M. (1975). Vasopressinrelease by nicotine: the site of action. British Journal of Pharmacology 54, 463-474.
BISSET, G. W., HALDAR, JAYA & LEWIN, J. E. (1966). Actions of oxytocin and other biologicallyactive peptides on the rat uterus. Memoirs of the Society for Endocrinology no. 14, 185-198.
BISSET, G. W. & JONES, N. F. (1975). Antidiuretic hormone. In Recent Advances in Renal Disease,ed. JONES, N. F., pp. 350-416. Edinburgh: Churchill Livingstone.
BISSET, G. W. & LEWIS, G. P. (1962). A spectrum of pharmacological activity in some biologicallyactive peptides. British Journal of Pharmacology 19, 168-182.
BLESSING, W. W., SVED, A. F. & REIS, D. J. (1982). Destruction of noradrenergic neurons in rabbitbrainstem elevates plasma vasopressin, causing hypertension. Science 217, 661-663.
BRENNAN JR, L. A., MALVIN, R. L., JOCHIM, K. E. & ROBERTS, D. E. (1971). Influence of right andleft atrial receptors on plasma concentrations ofADH and renin. American Journal of Physiology221, 273-278.
BRIDGES, T. E., HILLHOUSE, E. W. & JONES, M. T. (1976). The effect of dopamine on neurohypo-physial hormone release in vivo and from the rat neural lobe and hypothalamus in vitro. Journalof Physiology 260, 647-666.
CASTRO DE SOUZA, ELIZABETH, M. & ROCHA E SILVA JR, M. (1977). The release of vasopressin bynicotine: further studies on its site of action. Journal of Physiology 265, 297-311.
CLARK, BARBARA J. & ROCHA E SILVA JR, M. (1967). An afferent pathway for the selective releaseof vasopressin in response to carotid occlusion and haemorrhage in the cat. Journal of Physiology191, 529-542.
CLARKE, G., FALL, CAROLINE H. D., LINCOLN, D. W. & MERRICK, LYNDA P. (1978). Effects ofcholinoceptor antagonists on the suckling induced and experimentally evoked release of oxytocin.British Journal of Pharmacology 63, 519-527.
DICKER, S. E. (1953). A method for the assay of very small amounts of antidiuretic activity witha note on the antidiuretic titre of rat's blood. Journal of Physiology 122, 149-157.
DREIFUSS, J. J. & KELLY, J. S. (1972). The activity of identified supraoptic neurones and theirresponse to acetylcholine applied by iontophoresis. Journal of Physiology 220, 105-118.
ERRINGTON, M. L. & DASHWOOD, M. R. (1979). Projections to ventral surface of the cat brainstemdemonstrated by horseradish peroxidase. Neuroscience Letters 12, 153-158.
FATER, D. C., SCHULTZ, H. D., SUNDET, W. D., MAPES, J. S. & GOETZ, K. L. (1982). Effects of leftatrial stretch in cardiac-denervated and intact conscious dogs. American Journal of Physiology242, H1056-1064.
FELDBERG, W. (1976).The ventral surface of the brain stem: a scarcely explored region ofpharmacological sensitivity. Neuroscience 1, 427-441.
FELDBERG, W. & ROCHA E SILVA JR, M. (1981). Inhibition ofvasopressin release to carotid occlusionby y-aminobutyric acid and glycine. British Journal of Pharmacology 72, 17-24.
GAUER, 0. H. & HENRY, J. P. (1963). Circulatory basis of fluid volume control. PhysiologicalReviews 43, 423-481.
GAUER, 0. H., HENRY, J. P., SIEKER, 0. H. & WENDT, W. E. (1954). The effect of negative pressurebreathing on urine flow. Journal of Clinical Investigation 33, 287-296.
543

G. W. BISSET AND H. S. CHOWDREY
GILLIs, R. A., HELKE, C. J., HAMILTON, B. L., NORMAN, W. D. & JACOBOWITZ, D. M. (1980).Evidence that substance P is a neurotransmitter of baro- and chemoreceptor efferents in nucleustractus solitarius. Brain Research 181, 476-481.
GOETZ, K. L., BOND, C. C. & BLOXHAM, D. D. (1975). Atrial receptors and renal function.Physiological Reviews 55, 157-205.
HARRIS, M. C. (1978). Concept of the neuroendocrine reflex. In Cell Biology of HypothalamicNeurosecretion, ed. VINCENT, J. D. & KORDON, C., pp. 47-61. Paris: CNRS.
HATTON, G. I. (1982). Phasic bursting activity of rat paraventricular neurones in the absence ofsynaptic transmission. Journal of Physiology 327, 273-284.
HATTON, G. I., Ho, Y. W. & MASON, W. T. (1983). Synaptic activation of phasic bursting in ratsupraoptic nucleus neurones recorded in hypothalamic slices. Journal of Physiology 345, 297-317.
HENRY, J. P., GAUER, 0. H. & REEVES, J. L. (1956). Evidence of the atrial location of receptorsinfluencing urine flow. Circulation Research 4, 85-90.
JOHNSON, J. A., MOORE, W. W. & SEGAR, W. E. (1969). Small changes in left atrial pressure andplasma antidiuretic hormone titers in dogs. American Journal of Physiology 217, 210-214.
KANJANAPOTHI, D. (1975). The release of vasopressin by hypotensive drugs. Ph.D. Thesis,University of London.
KIMURA, T., SHARE, L., WANG, B. C. & CROFTON, J. T. (1981). The role of central adrenoceptorsin the control of vasopressin release and blood pressure. Endocrinology 108, 1829-1836.
KtHN, E. R. (1974). Cholinergic and adrenergic release mechanisms for vasopressin in the male rat:A study with injections of neurotransmitters and blocking agents into the third ventricle.Neuroendocrinology 16, 255-264.
KUMADA, M. & SAGAWA, K. (1970). Aortic nerve activity during blood volume changes. AmericanJournal of Physiology 218, 961-965.
LEDSOME, J. R., WILSON, N. & NGSEE, J. (1982). Time course of changes in plasma vasopressinduring atrial distension. Canadian Journal of Physiology and Pharmacology 60, 1210-1218.
MILLER, T. R., HANDELMAN, W. A., ARNOLD, P. E., MCDONALD, K. M., MOLINOFF, P. B. &SCHRIER, R. W. (1979). Effect of central catecholamine depletion on the osmotic and nonosmoticstimulation of vasopressin (antidiuretic hormone) in the rat. Journal of Clinical Investigation 64,1599-1607.
MILTON, A. S. & PATERSON, A. T. (1974). A microinjection study of the control of antidiuretichormone release by the supraoptic nucleus of the hypothalamus in the cat. Journal of Physiology241, 607-628.
MORAN, W. H., MILTENBERGER, F. W., SHUAYB, W. A. & ZIMMERMAN, B. (1964). The relationshipof antidiuretic hormone secretion to surgical stress. Surgery 56, 99-107.
Moss, R. L., URBAN, I. & CROSS, B. A. (1972). Microelectrophoresis of cholinergic and aminergicdrugs on paraventricular neurons. American Journal of Physiology 223, 310-318.
NICOLL, R. A. & BARKER, J. L. (1971). The pharmacology of recurrent inhibition in the supraopticneurosecretory system. Brain Research 35, 501-511.
O'CONNOR, W. J. & VERNEY, E. B. (1945). The effect ofincreased activity ofthe sympathetic systemin the inhibition of water diuresis by emotional stress. Quarterly Journal of ExperimentalPhysiology 33, 77-90.
OLSSON, K. (1970). Effects on water diuresis of infusions of transmitter substances into the 3rdventricle. Acta physiologica scandinavica 79, 133-135.
REIS, D. J., PERRONE, M. H. & TALMAN, W. T. (1981). Glutamic acid as the neurotransmitter ofbaroreceptor afferents in the nucleus tractus solitarius: possible relationship to neurogenichypertension. In Central Nervous System Mechanisms in Hypertension, ed. BUCKLEY, J. P. &FERRARIO, C. M., pp. 37-48. New York: Raven Press.
SAKAI, K. K., MARKS, B. H., GEORGE, J. M. & KOESTNER, A. (1974). The isolated organ-culturedsupraoptic nucleus as a neuropharmacological test system. Journal of Pharmacology andExperimental Therapeutics 190, 482-491.
SAWCHENKO, P. E. & SWANSON, L. W. (1981). Central noradrenergic pathways for the integrationof hypothalamic neuroendocrine and autonomic responses. Science 214, 685-687.
SAWCHENKO, P. E. & SWANSON, L. W. (1982). The organisation of noradrenergic pathways fromthe brainstem to the paraventricular and supraoptic nuclei in the rat. Brain Research Reviews4, 275-325.
544

REFLEX RELEASE OF VASOPRESSIN
SAWYER, W. H., PANG, P. K. T., SETO, J., MCENROE, M., LAMMEK, B. & MANNING, M. (1981).Vasopressin analogues that antagonize antidiuretic responses by rats to the antidiuretic hormone.Science 212, 49-51.
SHARE, L. (1974). Blood pressure, blood volume, and the release of vasopressin. In Handbook ofPhysiology, section 7, vol. 4, part 1, ed. KNOBIL, E. & SAWYER, W. H., pp. 243-255. Washington:American Physiological Society.
SHARE, L. & LEVY, M. N. (1962). Cardiovascular receptors and blood titer of antidiuretic hormone.American Journal of Physiology 203, 425-428.
SHARE, L. & LEVY, M. N. (1966a). Effect of carotid chemoreceptor stimulation on plasmaantidiuretic hormone titre. American Journal of Phy8iology 210, 157-161.
SHARE, L. & LEVY, M. N. (1966b). Carotid sinus pulse pressure, as a determinant of plasmaantidiuretic hormone concentration. American Journal of Phyriology 211, 721-724.
SLADEK, C. D. & JOYNT, R. J. (1979). Characterization of cholinergic control of vasopressin releaseby the organ-culture rat hypothalamo-neurohypophyseal system. Endocrinology 104, 659-663.
SLADEK, CELIA D. & KNIGGE, K. M. (1977). Cholinergic stimulation of vasopressin release from therat hypothalamo-neurohypophyseal system in organ culture. Endocrinology 101, 411-420.
SPYER, K. M. (1982). Central nervous integration of cardiovascular control. Journal ofExperimentalBiology 100, 109-128.
URANO, A. & KOBAYASHI, H. (1978). Effects of noradrenaline and dopamine injected into thesupraoptic nucleus on urine flow rate in hydrated rats. Experimental Neurology 60, 140-150.
VANDEPUTTE-VAN MESSON, G. & PEETERS, G. (1975). Effect of intraventricular administration ofnoradrenaline on water diuresis in goats. Journal of Endocrinology 66, 375-383.
PHY 354
545
18