the presence of epidermal growth factor binding sites in the
TRANSCRIPT

J. Cell Sci. 84, 19-40 (1986) 19Printed in Great Britain © The Company of Biologists Limited 1986
THE PRESENCE OF EPIDERMAL GROWTH FACTOR
BINDING SITES IN THE INTRACELLULAR
ORGANELLES OF TERM HUMAN PLACENTA
N. RAMANI, N. CHEGINI, CH. V. RAO*, P. G. WOOSTAND G. S. SCHULTZDepartments ofObstetrics-Gynecology and Biochemistry, University of Louisville,School of Medicine, Louisville, Kentucky 40292, USA
SUMMARYHighly purified lysosomes, rough and smooth endoplasmic reticulum, and Golgi apparatus, as
well as microvillus plasma membranes, bound 12sI-labelled epidermal growth factor ([125I]EGF)with similar affinity. Scatchard plots for all the organelles were curvilinear. The apparent numberof available binding sites per mg protein of intracellular organelles was 27-71 % of that found inmicrovillus plasma membranes. The bound and free [1Z5I]EGF were not degraded by any of theorganelles.
Binding and dissociation of [125I]EGF in all organelles were dependent on the time andtemperature of incubation. The specificity of [12SI]EGF binding was similar in all organelles. Theoptimal pH for binding to lysosomes was 6'0, in contrast to 7-0 for all the other organelles.Exposure of different organelles to enzymes and protein-modifying reagents resulted in numerousbinding differences between the intracellular organelles and microvillus plasma membranes.Covalent affinity labelling with [12SI]EGF revealed two major proteins of 155 and 140(X 103)Mr inall the organelles. The 155xlO3A/r protein was labelled predominantly in all organelles exceptrough endoplasmic reticulum, where both proteins were equally labelled. Addition of proteolyticinhibitors during isolation of organelles did not alter the pattern of [lzsI]EGF-labelled bindingproteins found in the organelles.
EGF also stimulated phosphorylation of the 155 and 140(X \0i)Mr proteins in all the organelles.The 155xl03A/r protein was phosphorylated more than the 140x10''Mr protein in microvillusplasma membranes and smooth endoplasmic reticulum, whereas the HOxlO3^/,. protein wasphosphorylated more than the 155xlO3A/r protein in lysosomes and both proteins were equallyphosphorylated in rough endoplasmic reticulum. Several organelles also contained minor[125I]EGF-binding proteins that did not show phosphorylation response and proteins that showedphosphorylation response but did not bind [125I]EGF.
Thus, the present study demonstrates by a number of different criteria, that several intracellularorganelles of term human placenta also contain EGF-binding and kinase activities.
INTRODUCTION
Mouse epidermal growth factor (EGF) and urogastrone, the human form of EGF,are potent mitogens in a variety of cells of ectodermal and mesodermal origin(Carpenter & Cohen, 1979; Gospodarowicz, 1981). They also modulate a variety ofdifferentiated cellular functions (Carpenter & Cohen, 1979; Gospodarowicz, 1981;Johnson et al. 1980a; Jones et al. 1982). The biological actions of EGF are
•Author for correspondence.
Key words: receptors, epidermal growth factor, human placenta.

20 N. Ramani and others
presumed to be initiated by EGF binding to specific and high-affinity receptorspresent in outer cell membranes of responsive cells (Carpenter & Cohen, 1979;Gospodarowicz, 1981). Following binding to cell surface receptors, the EGF—receptor complexes are internalized (Carpenter & Cohen, 1979; Gospodarowicz,1981). The internalization may represent simply a mechanism for degradation ofEGF—receptor complexes, thus potentially limiting the cellular response to EGF(Carpenter & Cohen, 1976; Pastan & Willingham, 1982) and, or, it may also play arole by translocating domains of the EGF and, or, its receptors to parts of the cellswhere they can come in contact with intracellular organelles, including the nucleus.Supporting the latter possibility were the reports showing that: (1) the biologicaleffects of EGF may require its internalization (Aharonov et al. 1978; Fox & Das,1979; King et al. 1981); (2) the internalized EGF-receptor complexes are bio-logically active and they catalyse the phosphorylation of soluble proteins (Cohen &Fava, 1985); (3) regenerating rat liver cells accumulate intact EGF in their nuclei(Raper et al. 1985); (4) GH3 cells accumulate EGF in their nuclei in the presence oflysosomotropic agents (Johnson et al. 19806; Savionei al. 1981); (5) EGF increasesthe phosphorylation of specific nuclear proteins in GH3 cells, and this effect wasaugmented under conditions that increase nuclear EGF accumulation (Johnsonet al. 1982); (6) EGF induces perturbation of nuclear chromatin structure (Johnsonet al. 1980a); (7) the Golgi uptake of [125I]EGF was enhanced under conditions thatcan fully support cell growth (Miskimins & Shimizu, 1982); (8) placental syncytio-trophoblasts accumulate [125I]EGF in their spherical and polymorphic endosomes,multivesicular bodies, nuclei, lysosomes, rough endoplasmic reticulum and Golgiapparatus (Lai et al. 1986; Chegini & Rao, 1986); and (9) highly purified EGFreceptor can interact and nick supercoiled double-stranded DNA in an ATP-dependent manner (Mroczkowski et al. 1984). However, a more recent studydemonstrated that the topoisomerase activity of the highly purified EGF receptorwas due to contaminating protein copurifying with the receptor (Basu et al. 1985).
If the mechanism that mediates the biological activity of EGF involves thenecessary internalization of the EGF and its receptors, and if the EGF receptors aresynthesized and degraded as other membrane proteins, then it would not beincompatible with EGF receptors being present in intracellular organelles. In factsome of the recent studies have demonstrated the presence of intracellular EGFbinding sites (Moriarity & Savage, 1980; Lev-Ran et al. 1984; Dunn & Hubbard,1985). These studies, however, have neither attempted to isolate all the cellularorganelles nor characterized the intracellular binding sites, in detail.
We have used term human placenta, which expresses a high level of EGF receptorsin the microvillus plasma membranes (Rao et al. 1985), to investigate andcharacterize intracellular binding sites for EGF.
MATERIALS AND METHODS
The sources of materials used in the present studies have been described (Rao et al. 1985).EGF was purified from mouse submaxillary gland and iodinated in our laboratory (Rao et al.
1984) using previously published procedures (Savage & Cohen, 1972; Davis, 1954; Matrisianet al.

Intracellular binding of epidermal growth factor 21
1982; Gospodarowicz, 1975). The specific activity of [125I]EGF ranged from 190 to 220 juCi^g"1.The EGF iodinated by the procedure of Comens et al. (1982) was used in covalent labelling ofEGF receptors, whereas that obtained by the procedure of Carpenter & Cohen (1976) was used inthe rest of the binding experiments.
Term human placentae from spontaneous normal vaginal deliveries were brought to thelaboratory on ice. The reflected foetal membranes were cut and discarded. Placental tissue wasscraped off from the underlying blood vessels and repeatedly washed with chilled physiologicalsaline to remove blood.
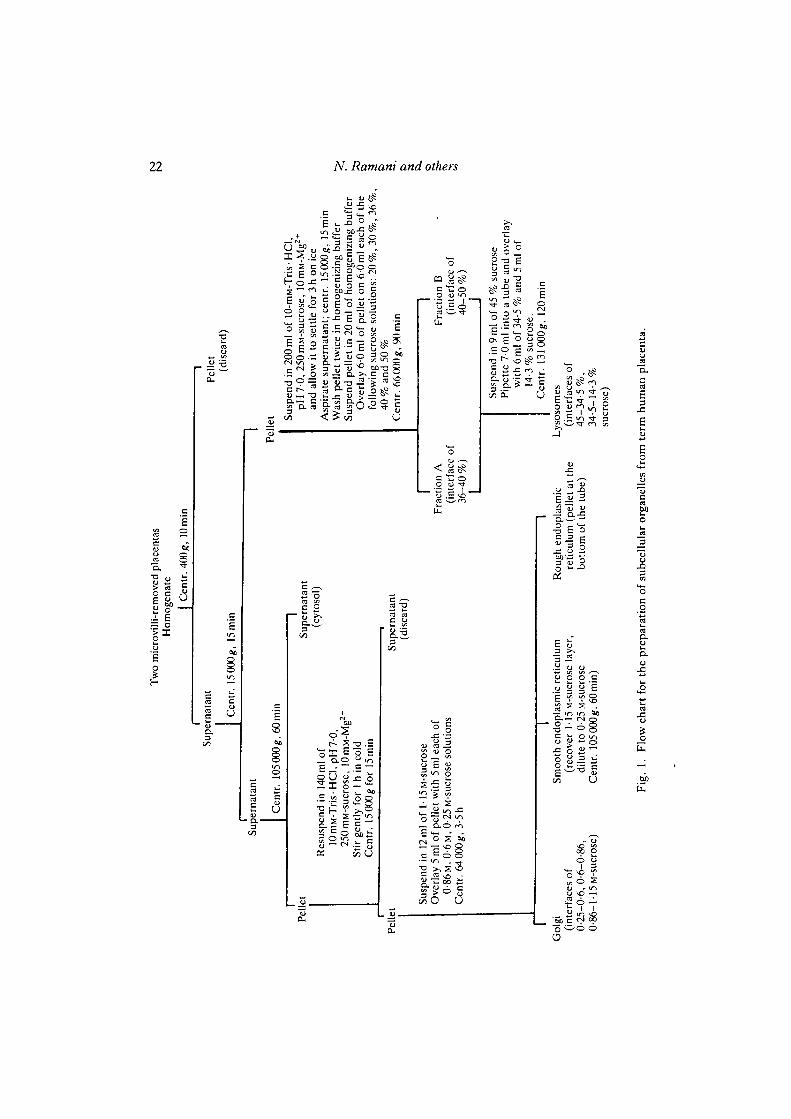
Procedures to isolate intracellular organelles of placenta are based on those described for liver.However, we had to improvise a great deal in order to obtain pure fractions as differences existbetween subcellular fractionation of term human placenta and liver. After about two and half yearsof trial-and-error experiments, the following isolation procedure was finally adopted (Fig. 1).Briefly, microvillus plasma membranes were first harvested by mechanical shearing followed by aseries of centrifugations and exposure to lOmM-Mg2"1" to aggregate intracellular organelles (Boothet al. 1980). Then the placental tissue was homogenized at 4°C in 10mM-Tris-HCl (pH7-0)containing 250 mM-sucrose and 1 mM-Ca2+ (referred to as homogenizing buffer) with a polytronhomogenizer at a setting of 6 using three 10-s bursts with short pauses in between. Thehomogenates were filtered through four layers of cheese-cloth and subjected to differentialsedimentation and floatation discontinuous sucrose-gradient centrifugation steps to isolate variousintracellular organelles (see Fig. 1).
The property of Mg2"1" to aggregate intracellular organelles was used at two strategic stages in thefractionation procedure to reduce contamination by cell surface membranes. The first stage wasresuspending the 15 000^ pellet in Mg2+-containing buffer and leaving it to settle for 3 h. At theend of this period, the supernatant containing surface membranes was removed and discarded. Thesettled fraction, containing primarily intracellular organelles, was washed free of Mg2+ and thensubjected to sedimentation and floatation discontinuous sucrose-density gradient centrifugations toisolate lysosomes. The second place was stirring the 105 000 gpellet for an hourinMg2+-containingbuffer and then centrifuging for 15 min at 15 000g. The pellet, containing primarily intracellularorganelles, was washed free of Mg2"1" and then centrifuged in discontinuous sucrose-densitygradient to isolate rough and smooth endoplasmic reticulum and Golgi apparatus. Measurement ofsurface-membrane marker enzyme activity in fractions prepared with and without Mg2"1" treatmentrevealed that the treatment was effective in reducing surface membrane contamination. Electronmicrographs and thiamine pyrophosphatase activity presented in this paper also lend support to theabove conclusion.
Immediately after isolation, the subcellular fractions were processed for electron microscopicexamination as previously described by Chegini et al. (1984).
All the subcellular fractions were frozen in 1 ml samples at —20°C. After digestion at 80cC inlOOmM-NaOH containing 0-1 % sodium dodecyl sulphate (SDS), the protein in the fractions wasdetermined by the method of Lowry et al. (1951) using bovine serum albumin (BSA) as thestandard.
Various marker enzymes were assayed using previously described procedures. Thiamine pyro-phosphatase (EC 3.6.1.1) according to Bramley & Ryan (1978), glucose-6-phosphate dehydro-genase (EC 1.1.1.49) according to Kelly et al. (1955) and lactate dehydrogenase (EC 1.1.1.27)according to the procedure described in Worthington's Enzyme Manual (1972). The inorganicphosphorus released in the thiamine pyrophosphatase assays was measured according to Martin &Doty (1949). Appropriate blanks were run in all the enzyme assays and corrections for these valueswere made.
Binding studies were performed at 22°C for 2h with 0-5nM-[125I]EGF in the presence andabsence of 176-fold excess of unlabelled EGF in 0-2 ml of incubation medium, whose finalcomposition was 5 mM-Tris • HC1 (pH7-0), 125 mM-sucrose, 0-5mM-Ca2+, 75 mM-NaCl and 0-5%bovine serum albumin (BSA). The free and bound [125I]EGF were separated by filtration across0-22 fun pore-size filters. The bound [125I]EGF was counted in a Searle model 1197 autogammacounter with a counting efficiency of 82% for 125I. The data presented in this paper are specificbinding, i.e. the difference between total and non-specific binding. Further details and variationsfrom those described above are presented in figure and table legends.
I\! Ramani and others

Intracellular binding of epidermal growth factor 23
Covalent labelling of EGF receptors and EGF-stimulated protein phosphorylation have beendescribed (Rao et al. 1985). The molecular mass determination of proteins between 100000 and200000 are accurate to within 5000 in our systems.
Each experiment was conducted on subcellular organelles from the same batch, under identicalconditions and at the same time. All the experiments were replicated two or three times onorganelles from different batches. The values presented are the means and their standard errors ofobservations collected in all these experiments. Analysis of variance and Dunnet's multiple rangetest (Steel & Torrie, 1960) were used in determining whether the observed differences betweenmicrovillus plasma membranes and intracellular organelles were significant. The significantdifferences are indicated by placing asterisks across the columns in various tables. Although thesignificant differences among the intracellular organelles was not indicated, they can be judged, atleast in some cases, from the comparison of means and standard errors, and relating them to thesignificant differences found between microvillus plasma membranes and intracellular organelles.
RESULTS
Purity of subcellular organelles
Microvillus plasma membranes consisted largely of spherical and elongatedvesicles, some of which contained electron-dense material (Fig. 2A). Very little or nocontamination with other organelles was found in this fraction.
Lysosomal fraction consisted of vesicles of different sizes and shapes with singleand sometimes multiple limiting membranes (Fig. 2B). Rough endoplasmic reticu-lum was occasionally seen in this fraction.
Rough endoplasmic reticulum fraction was characterized by vesicles of irregularprofiles with bound ribosomes (Fig. 2C). A number of scattered mitochondria wereseen among the vesicles.
Smooth endoplasmic reticulum consisted of empty smooth-surface vesicles, someof which contained amorphous electron-dense material (Fig. 2D). Round andelongated vesicles containing electron-dense material were seen in this fraction.These vesicles were smaller than but otherwise similar to those found in microvillusplasma membranes. There was no contamination of this fraction with eithermitochondria or rough endoplasmic reticulum (Fig. 2D).
Vesicles of different sizes and shapes, and some flattened membranes were seen inthe Golgi fraction (Fig. 3). Contamination with the other organelles was not evidentin this fraction.
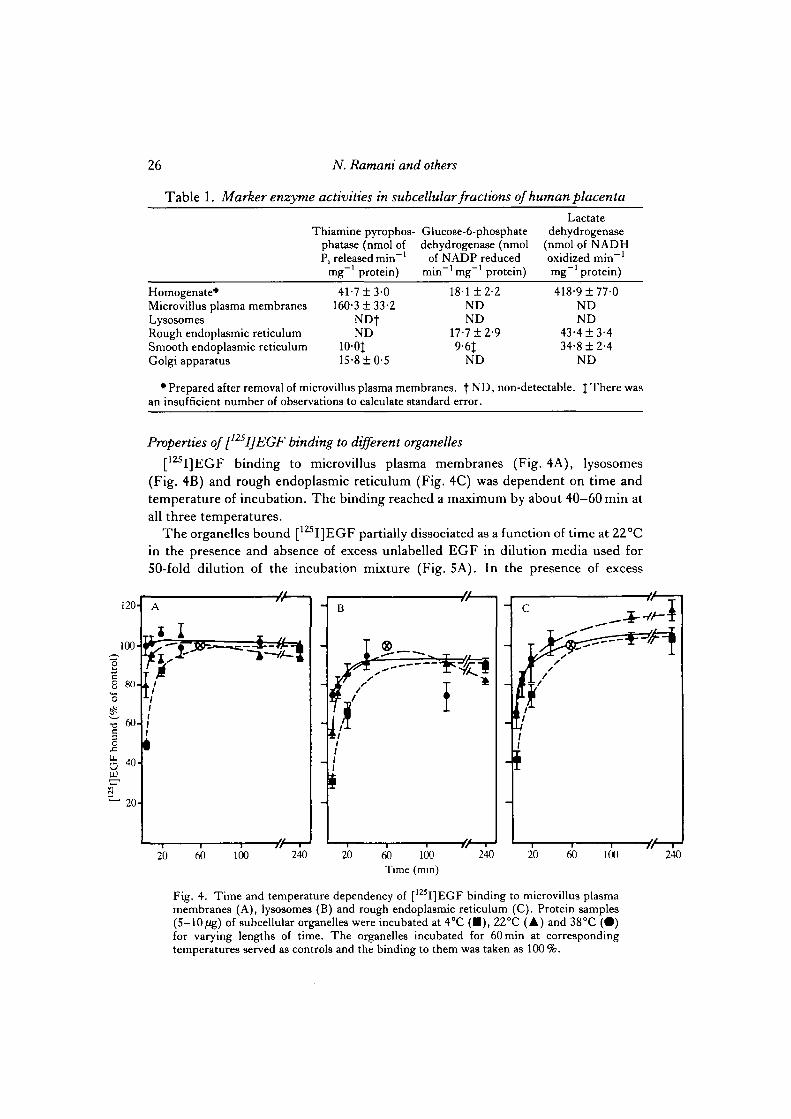
Marker enzyme activities were also measured to evaluate further the purity of thefractions. The 5' nucleotidase was distributed not only in plasma membranes butalso in cytosol and other intracellular organelles of placenta (Rao et al. 1985; Madrid-Marina & Fox, 1986; Berry et al. 1986; and unpublished observations from ourlaboratory). Because of this, we surveyed 10 different marker enzyme activities in allthe subcellular organelles of placenta. The results indicated that thiamine pyro-phosphatase was the most reliable marker for plasma membranes. Since our primaryconcern was to determine the extent of plasma membrane contamination in theintracellular organelles, our findings on thiamine pyrophosphatase activity werefortuitous; its activity was enriched in microvillus plasma membranes from the

24 N. Ramani and others
iMPW i
if*
Fig. 2. Electron micrographs of isolated microvillus plasma membranes (A), lysosomes(B), rough endoplasmic reticulum (C) and smooth endoplasmic reticulum (D). X IS 290.Bar, 1 /im.

Intracellular binding of epidermal growth factor 25
Fig. 3. Electron micrograph of isolated Golgi apparatus. X16 700. Bar, 1 /an.
original homogenate (Table 1). More importantly, very little or none of this enzymeactivity was found in the intracellular organelles, suggesting that microvillus plasmamembrane contamination in the intracellular organelles was minimal to non-existent.The same conclusion was reached after a careful inspection of subcellular organellesat the electron microscopic level (Figs 2A-D, 3).
We have 'not found any marker enzymes intrinsic to intracellular organelles ofplacenta, and liver intracellular organelles markers were not useful. Therefore, wehad to rely on the morphology of these organelles that showed little or no crosscontamination. Cytosol contamination in the isolated membranous organelles, asjudged by the measurement of glucose 6-phosphate and lactate dehydrogenases, wasminimal (Table 1).
The procedures and improvisations used in isolation of pure fractions from termhuman placenta led to severe and unaccountable losses of fractions, which made thecalculations of recoveries of fractions and bindings meaningless. In addition, becauseof the losses, all the properties of binding on all the fractions could not be studied.
26 N. Ramani and others
Table 1. Marker enzyme activities in subcellular fractions of human placenta
Homogenate*Microvillus plasma membranesLysosomesRough endoplasmic reticulumSmooth endoplasmic reticulumGolgi apparatus
Thiamine pyrophos-phatase (nmol ofP, released min"1
mg"1 protein)
41-7± 3-0160-3 ±33-2
NDfND
100J15-8 ±0-5
• Glucose-6-phosphatedehydrogenase (nmol
of NADP reducedmin"1 mg"1 protein)
18-1 ±2-2NDND
17-7 ±2-99-6J
ND
Lactatedehydrogenase
(nmol of NADHoxidized min"1
mg"1 protein)
418-9 ±77-0NDND
43-4 ±3-434-8 ±2-4
ND
• Prepared after removal of microvillus plasma membranes. | ND, non-detectable. J There wasan insufficient number of observations to calculate standard error.
Properties of [I2SI]EGF binding to different organelles
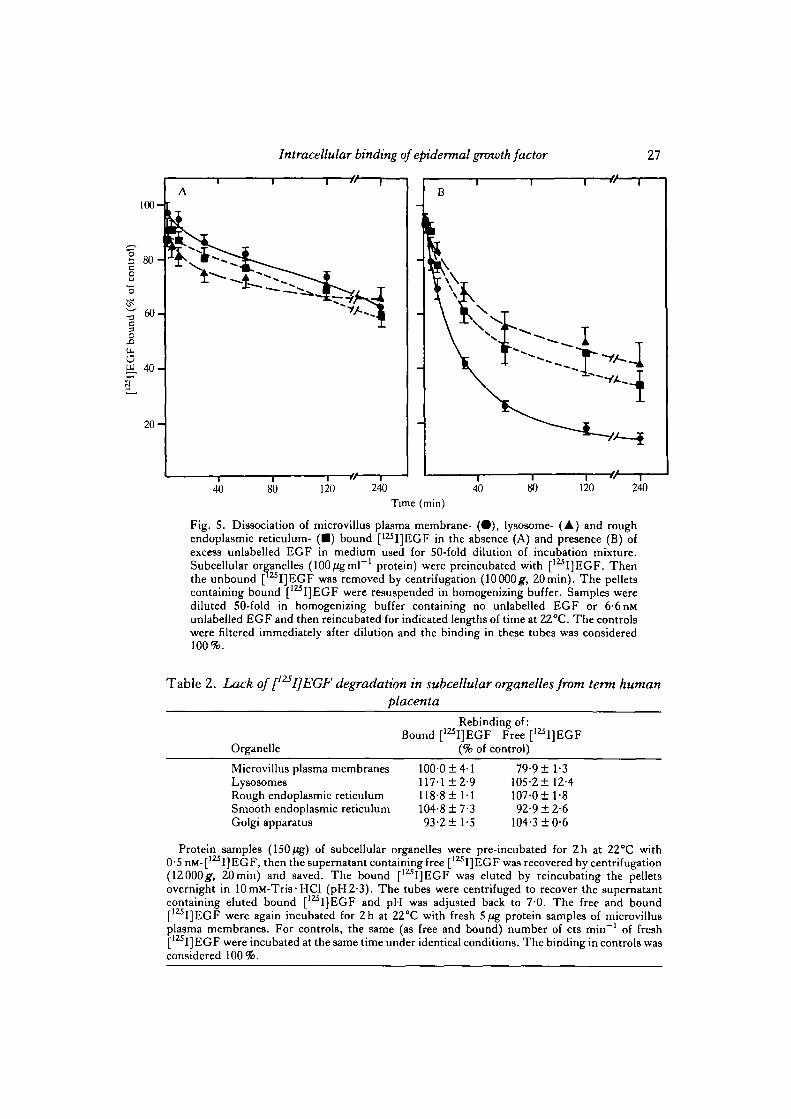
[125I]EGF binding to microvillus plasma membranes (Fig. 4A), lysosomes(Fig. 4B) and rough endoplasmic reticulum (Fig. 4C) was dependent on time andtemperature of incubation. The binding reached a maximum by about 40—60min atall three temperatures.
The organelles bound [ I]EGF partially dissociated as a function of time at 22°Cin the presence and absence of excess unlabelled EGF in dilution media used for50-fold dilution of the incubation mixture (Fig. 5A). In the presence of excess
120-
100-
•g
8 80-
$^ 60-c3O
UJ
s"" 20-
A
I
i
ii
I
at
B
I.S
f/1
i
i t
_
20 60 100 240 20 60 100Time (min)
240 60 100 240
Fig. 4. Time and temperature dependency of [125I]EGF binding to microvillus plasmamembranes (A), lysosomes (B) and rough endoplasmic reticulum (C). Protein samples(S-10/ig) of subcellular organelles were incubated at 4°C (•) , 22°C (A) and 38°C (•)for varying lengths of time. The organelles incubated for 60 min at correspondingtemperatures served as controls and the binding to them was taken as 100 %.
Intracellular binding of epidermal growth factor 27
40 80 120 240Time (min)
Fig. 5. Dissociation of microvillus plasma membrane- ( • ) , lysosome- (A) and roughendoplasmic reticulum- ( • ) bound [125I]EGF in the absence (A) and presence (B) ofexcess unlabelled EGF in medium used for 50-fold dilution of incubation mixture.Subcellular organelles (100/igmF1 protein) were preincubated with [12SI]EGF. Thenthe unbound [ I]EGF was removed by centrifugation (10000g, 20min). The pelletscontaining bound [' I]EGF were resuspended in homogenizing buffer. Samples werediluted 50-fold in homogenizing buffer containing no unlabelled EGF or 6-6 nMunlabelled EGF and then reincubated for indicated lengths of time at 22°C. The controlswere filtered immediately after dilution and the binding in these tubes was considered100%.
Table 2. Lack of [I2SI]EGF degradation in subcellular organelles from term humanplacenta
Organelle
Rebinding of:Bound [1Z5I]EGF Free [125I]EGF
(% of control)
Microvillus plasma membranes 100-0 ±4-1 79-9 ±1-3Lysosomes 117-1 ±2-9 105-2 ±12-4Rough endoplasmic reticulum 118-8 ±1-1 107-0 ±1-8Smooth endoplasmic reticulum 104-8 ± 7-3 92-9 ±2-6Golgi apparatus 93-2 ±1-5 104-3 ±0-6
Protein samples (150fig) of subcellular organelles were pre-incubated for 2h at 22°C with0-5 nM-[125I]EGF, then the supernatant containing free [125I]EGF was recovered by centrifugation(12000#, 20min) and saved. The bound [1Z5I]EGF was eluted by reincubating the pelletsovernight in 10mM-Tris-HCl (pH2-3). The tubes were centrifuged to recover the supernatantcontaining eluted bound [125I]EGF and pH was adjusted back to 7-0. The free and bound[I25I]EGF were again incubated for 2h at 22°C with fresh 5^g protein samples of microvillusplasma membranes. For controls, the same (as free and bound) number of cts min"1 of fresh[125I]EGF were incubated at the same time under identical conditions. The binding in controls wasconsidered 100%.

28 N. Ramani and others
unlabelled EGF, however, the dissociation was considerably enhanced in all theorganelles with significantly greater enhancement in microvillus plasma membranesas compared to the intracellular organelles (Fig. 5B).
The free and eluted bound [125I]EGF from microvillus plasma membranes andintracellular organelles was able to rebind to fresh microvillus plasma membranes aseffectively as fresh [125I]EGF, suggesting that [1Z5I]EGF was not degraded duringthe binding reaction (Table 2). The 20% decrease in rebindability of free [12SI]EGFrecovered from the media following incubation with microvillus plasma membraneswas perhaps due to greater binding during the first incubation. In other words,[125I]EGF molecules that bind to the receptors well were taken up by microvillus
1000 1800
200
60-_
40-
20-
D
F
^ -h 30-x -m
u. 2U"
"Mo-
\
10 30 50
1000 1800
60-
40-
-
20-
E
f 10 10-
\
10
•
30 50
200 1000 18001000 1800
60-
40-
-
20-
C
/
/
-
X
30-
20-
10-
^ - •
\
10
* — •
30 SO
200 1000 18005I]EGF added (pM)
Fig. 6. Dependence of [ l z i I]EGF binding to microvillus plasma membranes (A),lysosomes (B), rough endoplasmic reticulum (C), smooth endoplasmic reticulum (D)and Golgi apparatus (E) on the [IZ5I]EGF concentration in the incubation medium.Samples of 5 /ig protein of subcellular organelles were incubated with increasingconcentrations of [I2SI]EGF. Insets show the Scatchard (1949) plots of the binding data.
Intracellular binding of epidermal growth factor 29
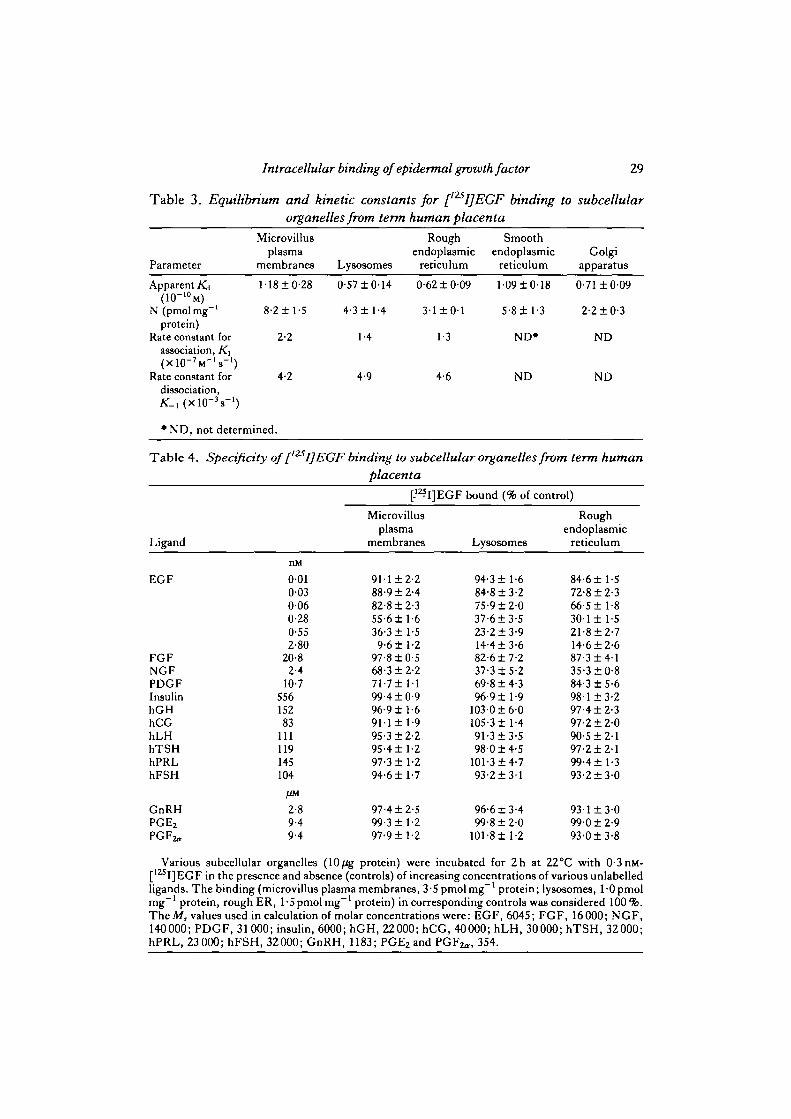
Table 3. Equilibrium and kinetic constants for [125I]EGF binding to subcellularorganelles from term human placenta
Parameter
Microvillusplasma
membranes Lysosomes
Rough Smoothendoplasmic endoplasmic Golgi
reticulum reticulum apparatus
Apparent K4
N (pmolmg"1
protein)Rate constant for
association, K\
Rate constant fordissociation,
M8±0-28
8-2± 1-5
2-2
4-2
0-57 ±0 '
4-3 ±1 -
1-4
4.9
14
4
0-62 ±0-09
3-1 ± 0 1
1-3
4-6
1-09 ±0-
5-8 ± 1-
ND*
ND
18
3
0-71 ±0-
2-2 ±0-
ND
ND
09
3
• ND, not determined.
Table 4. Specificity of [ I] EGF binding to subcellular organelles from term humanplacenta
[-1Z?I]EGF bound (% of control)
Ligand
EGF
FGFNGFPDGFInsulinhGHhCGhLHhTSHhPRLhFSH
GnRHPGE2
PGFz-
nM
0-010-030-060-280-552-80
20-82-4
10-755615283
111119145104
/iM
2-89-49-4
Microvillusplasma
membranes
9M±2-288-9 ±2-482-8 ±2-355-6± 1-636-3 ±1-59-6 ±1-2
97-8 ±0-568-3 + 2-271-7± 1199-4 + 0-996-9 ±1-6911 ± 1-995-3 ±2-295-4 ±1-297-3 ±1-294-6 ±1-7
97-4 ±2-599-3 ±1-297-9 ±1-2
Lysosomes
94-3 ± 1-684-8 ±3-275-9 ± 2 037-6 + 3-523-2±3-914-4 ±3-682-6 ±7-237-3 ±5-269-8 ±4-396-9 ±1-9
1030 ± 6 0105-3 ±1-491-3 ±3-598-0 ±4-5
101-3 ±4-793-2±31
96-6 ±3-499-8 ± 2 0
101-8 ± 1-2
Roughendoplasmic
reticulum
84-6 ±1-572-8 ±2-366-5±l-8301 ± 1-521-8 ±2-714-6 ±2-687-3 ±4-135-3 ±0-884-3 ±5-698-1 ±3-297-4 ±2-397-2 ± 2 090-5 ±2-197-2 ± 2 199-4±l-393-2±3-0
931 ± 3 0990 ±2-993-0 ±3-8
Various subcellular organelles (10/ig protein) were incubated for 2h at 22CC with 0-3nM-[125I]EGF in the presence and absence (controls) of increasing concentrations of various unlabelledligands. The binding (microvillus plasma membranes, 3-5 pmol mg"1 protein; lysosomes, 1 -0 pmolmg"1 protein, rough ER, l-5pmolmg~' protein) in corresponding controls was considered 100%.TheMr values used in calculation of molar concentrations were: EGF, 6045; FGF, 16000; NGF,140000; PDGF, 31000; insulin, 6000; hGH, 22000; hCG, 40000; hLH, 30000; hTSH, 32000;hPRL, 23 000; hFSH, 32000; GnRH, 1183; PGE2 and P G F ^ , 354.

30 N. Ramani and others
plasma membranes receptors, leaving behind [125I]EGF molecules that bind lessreadily to the receptors. This is the best explanation we can offer at this time for thisfinding.
The specific binding of [125I]EGF to microvillus plasma membranes (Fig. 6A),lysosomes (Fig. 6B), rough endoplasmic reticulum (Fig. 6C), smooth endoplasmicreticulum (Fig. 6D) and Golgi apparatus (Fig. 6E) increased with increasing con-centrations of added [1ZSI]EGF, reaching saturation at about 2-0nM of added[125I]EGF. The Scatchard (1949) transformation and graphic display of the data
Table 5. Effect of incubation media pH on [I25I]EGF binding to subcellularorganelles from term human placenta
pH of theincubation media
5-06-08-09-0
10-0
Microvillusplasma
membranes
23-6 ±2-747-2± 5-175-8 ±2-554-3 ±2-931-014-5
[125I]EGF bound
Lysosomes
27-8 ±5-51291 ± 6-1*92-418-274-7 ±4-8*51-618-6
(% of control)
Roughendoplasmic
reticulum23-9±5-025-9 ±6-251-2± 10-546-8 + 8-523-9 ±8-3
Smoothendoplasmic
reticulum
3S-6±5-l59-2 ±10-475-3 ±4-962-3 ±3-443-6 ±3-4
Samples of subcellular organelles (2-5 ftg protein) were incubated for 2h at 22°C with 0-5 nM-[I25I]EGF in media of various pH values; lOmM-Tris-HCl adjusted to the desired pH values wasused. The organelles incubated at pH7-0 served as controls and the binding (pmolmg"1 protein)in corresponding controls (microvillus plasma membranes, 2-9; lysosomes, 09; rough endo-plasmic reticulum, 1-1; smooth endoplasmic reticulum, 2-0) was considered as 100%.
• P < 0 0 5 .
Table 6. Thermal sensitivity ofEGF binding sites in subcellular organelles from termhuman placenta
Preincubationtemperature (°C)2238455565
Microvillusplasma
membranes99-211-576-2 + 2-768-4 + 2-927-514-5
1-3
[mI]EGF bound
Lysosomes
89-213-089-1+0-870-313-411-911-1*11-013-6
(% of control)
Roughendoplasmic
reticulum103-411-981-414-066-713-519-2 + 2-4
ND+
Smoothendoplasmic
reticulum
91-6111-587-214-070-414-330-112-4
ND
Various subcellular organelles (50 fig protein ml ') were preincubated for 15 min at the indicatedtemperatures. After cooling the tubes, 5 fig protein samples were tested for binding at 22°C for 2hwith 0-5 nM-[12sI]EGF. Subcellular organelles preincubated at 4°C for 15 min served as controls.The binding (pmolmg"1 protein) in respective controls (microvillus plasma membranes, 5-8;lysosomes, 0-9; rough endoplasmic reticulum, 0-8; smooth endoplasmic reticulum, 1-6) wasconsidered as 100%.
•P<0-05. fND, not detectable.
Intracellular binding of epidermal growth factor 31
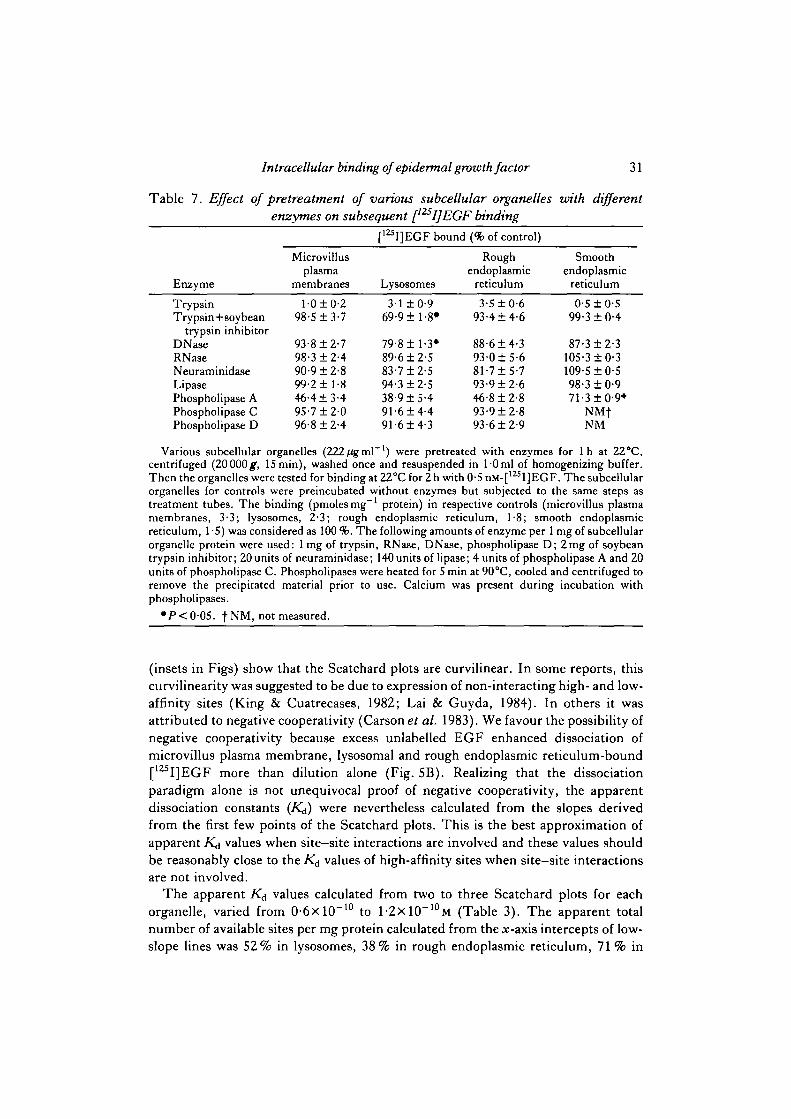
Table 7. Effect of pretreatment of various subcellular organelles with differentenzymes on subsequent [I25I]EGF binding
EnzymeTrypsinTrypsin+soybean
trypsin inhibitorDNaseRNaaeNeuraminidaseLipasePhospholipase APhospholipase CPhospholipase D
Microvillusplasma
membranesl-0±0-2
98-5 ±3-7
93-8 ±2-798-3 ±2-490-9 ±2-899-2 ±1-846-413-495-7 ±2-096-8 ±2-4
[12SI]EGF bound
Lysosomes3-1 ±0-9
69-9 ±1-8*
79-8 ±1-3*89-612-583-7 ±2-594-312-538-915-491-614-491-614-3
(% of control)
Roughendoplasmic
reticulum3-510-6
93-414-6
88-614-393-015-681-715-793-912-646-812-893-912-893-612-9
Smoothendoplasmic
reticulum0-5 10-5
99-310-4
87-312-3105-310-3109-510-598-310-971-31 0-9*
NMfNM
Various subcellular organelles (222/igml ') were pretreated with enzymes for 1 h at 22°C,centrifuged (20000^, 15min), washed once and resuspended in l-0ml of homogenizing buffer.Then the organelles were tested for binding at 22°C for 2 h with 0-5 nM-[125I]EGF. The subcellularorganelles for controls were preincubated without enzymes but subjected to the same steps astreatment tubes. The binding (pmolesmg"1 protein) in respective controls (microvillus plasmamembranes, 3-3; lysosomes, 2-3; rough endoplasmic reticulum, 1-8; smooth endoplasmicreticulum, 1-5) was considered as 100 %. The following amounts of enzyme per 1 mg of subcellularorganelle protein were used: 1 mg of trypsin, RNase, DNase, phospholipase D; 2mg of soybeantrypsin inhibitor; 20 units of neuraminidase; 140 units of lipase; 4 units of phospholipase A and 20units of phospholipase C. Phospholipases were heated for 5 min at 90°C, cooled and centrifuged toremove the precipitated material prior to use. Calcium was present during incubation withphospholipases.
•P<0-05. |NM, not measured.
(insets in Figs) show that the Scatchard plots are curvilinear. In some reports, thiscurvilinearity was suggested to be due to expression of non-interacting high- and low-affinity sites (King & Cuatrecases, 1982; Lai & Guyda, 1984). In others it wasattributed to negative cooperativity (Carson et ai. 1983). We favour the possibility ofnegative cooperativity because excess unlabelled EGF enhanced dissociation ofmicrovillus plasma membrane, lysosomal and rough endoplasmic reticulum-bound[12SI]EGF more than dilution alone (Fig. 5B). Realizing that the dissociationparadigm alone is not unequivocal proof of negative cooperativity, the apparentdissociation constants (K^) were nevertheless calculated from the slopes derivedfrom the first few points of the Scatchard plots. This is the best approximation ofapparent /Q values when site-site interactions are involved and these values shouldbe reasonably close to the K& values of high-affinity sites when site—site interactionsare not involved.
The apparent K^ values calculated from two to three Scatchard plots for eachorganelle, varied from 0-6xl(T10 to 1-2X10~10M (Table 3). The apparent totalnumber of available sites per mg protein calculated from the x-axis intercepts of low-slope lines was 52% in lysosomes, 38% in rough endoplasmic reticulum, 71 % in

32 N. Ratnani and others
smooth endoplasmic reticulum and 27 % in Golgi apparatus, as compared to those inmicrovillus plasma membranes. Table 3 shows that the rate constants for associationand dissociation (in the absence of excess unlabelled EGF) were similar for microvilliin microvillus plasma membranes, lysosomes and rough endoplasmic reticulum.
Increasing concentrations of unlabelled EGF inhibited [12SI]EGF binding to allthe subcellular organelles in a dose-dependent manner (Table 4). One hundred-foldhigher concentrations of EGF, NGF and PDGF only moderately reduced [125I]-EGF binding. A variety of protein and peptide hormones, and prostaglandins (PGs),even at very high concentrations, had no effect on [1ZSI]EGF binding.
Table 5 shows that an optimal pH for [125I]EGF binding to lysosomes was 6-0 andfor all the other organelles it was 7-0. All the organelles exhibited binding lossesbelow and above the optimal pH. However, the lysosomal binding losses at pH 9-0were lower (P< 0-05) compared to microvillus plasma membranes.
Preincubation of all the subcellular organelles at increasing temperature for 15 minresulted jn similar and irreversible [12sI]EGF-binding losses (Tabje 6). At 55°C,however, the lysosomal binding losses were significantly greater (P<0-05) com-pared to those of microvillus plasma membranes.
Table 8. Effect of pretreatment of subcellular organelles with various proteinmodifying reagents on subsequent [I25I]EGF binding
Protein reagent
Acetic anhydride(10mM)
Mercaptoethanol(10mM)
iV-ethylmaleimide(10mM)
/>-Chloromercuribenzoate(1 HIM)
Dinitrofluorobenzene(10mM)
Tetranitromethane(10mM)
Microvillusplasma
membranes
46-2±3-l
97-0 ±3-1
86-7 ±2-3
81-3 ± 5 - 1
91-4±Z-8
12-2± 1-7
[125I]EGF
Lysosomes
24-0 ±2-9
90-1 ±2-5
82-9 ±3-3
68-3 ±3-0
75-1 ±6-0
24-3 ±2-0
bound (% of control)
Roughendoplasmic
reticulum
44-6±3-5
97-8 ±3-3
95-5 ±2-4
68-2±3-5
69-0 ±4-7*
15-2± 21
Smoothendoplasmic
reticulum
77-5 ±4-7*
75-1 ±6-8
102-6 ±3-0
108-9 ±l-5»
NMt
15-0± 5-8
Subcellular organelles (200 fig protein ml ') were preincubated for 2h at 22°C with variousprotein modifying reagents, centrifuged (20 000#, 15 min) washed once and suspended in 1-0 ml ofhomogenizing buffer. Then, the organelles were tested for binding by reincubating for 2h at 22°Cwith 0-5nM-[l25I]EGF. Acetic anhydride, mercaptoethanol, jV-ethylmaleimide and />-chloro-mercuribenzoate were added in lOmM-Tris-HCl, pH7-0. Dinitrofluorobenzene and tetranitro-methane were added in ethanol so that the final ethanol concentration in the incubation media was10%. The corresponding controls were preincubated with either Tris-HCl or ethanol andsubjected to the same steps as the treatment tubes. The binding in corresponding controls wasconsidered as 100%.
• P < 0 - 0 5 . fNM, not measured.

H 3x10"3205
116
97
66
Intracellular binding of epidermal growth factor
w * ** 205F ""* —
33
R • ft 11697
66
45
EGF - + - + - +
! S t
2A 2B 2CEGF - + - • - •
205 - r ^
11697
66
45
• • I H
4A 4B 4C
EGF -
Fig. 7. Autoradiography of covalently labelled EGF-binding proteins in microvillusplasma membranes (70/ig protein, lanes 1A-1C), lysosomes (40^g protein, lanes2A-2C), rough endoplasmic reticulum (80/ig protein, lanes 3A-3C) and smooth endo-plasmic reticulum (5 fig protein, lanes 4A-4C). The microvillus plasma membranes wereprepared as previously described (Rao et al. 1985) and in the final step were resuspendedin homogenizing buffer (HB) (lane 1A) or in the HB where Ca2+ was replaced with1 mM-EGTA (lane IB) or 1/igml"1 a-2-macroglobulin (lane 1C). The intracellularorganelles were prepared as indicated in the flow chart (Fig. 1) in HB (lane A) or in HBwhere Caz+ was replaced with EGTA (lane B) or tf-2-macroglobulin (lane C) or thecorresponding organelle. Approximately 38000, 18000, 41 000 and 13 000 disintsmin"1
were applied to each lane from microvillus plasma membranes, lysosomes, rough andsmooth endoplasmic reticulum, respectively. About 4000 disintsmin"1 were applied toeach lane from incubation containing unlabelled EGF.
34 N. Ramani and others
Pretreatment with trypsin virtually abolished [125I]EGF binding in all the intra-cellular organelles as well as in microvillus plasma membranes (Table 7). Simul-taneous addition of soybean trypsin inhibitor completely reversed trypsin-inducedbinding losses in all organelles except lysosomes (P < 0'05). The incomplete reversalin lysosomes may be attributable to trypsin-like proteins in lysosomes, which maybind the soybean trypsin inhibitor making it less available to block exogenouslyadded trypsin. DNase had little or no effect on binding losses from all organellesexcept lysosomes, where there was a 20% loss of binding (P<0-05). RNase,neuraminidase, lipase, phospholipases C and D had no effect on [125I]EGF bindingin any of the organelles. Phospholipase A was able to reduce binding in all theorganelles but with a lesser reduction in smooth endoplasmic reticulum (P < 0-05).
Pretreatment with acetic anhydride reduced binding in all the organelles with alesser reduction in smooth endoplasmic reticulum (P<0-05) (Table 8). Mercapto-ethanol and iV-ethylmaleimide had no effect on binding in any organelle. />-Chloro-mercuribenzoate reduced binding in all the organelles except smooth endoplasmic
X1O" 3 205
116
97
66
45
205
116
97
66
45
EGF + - + - + - + -
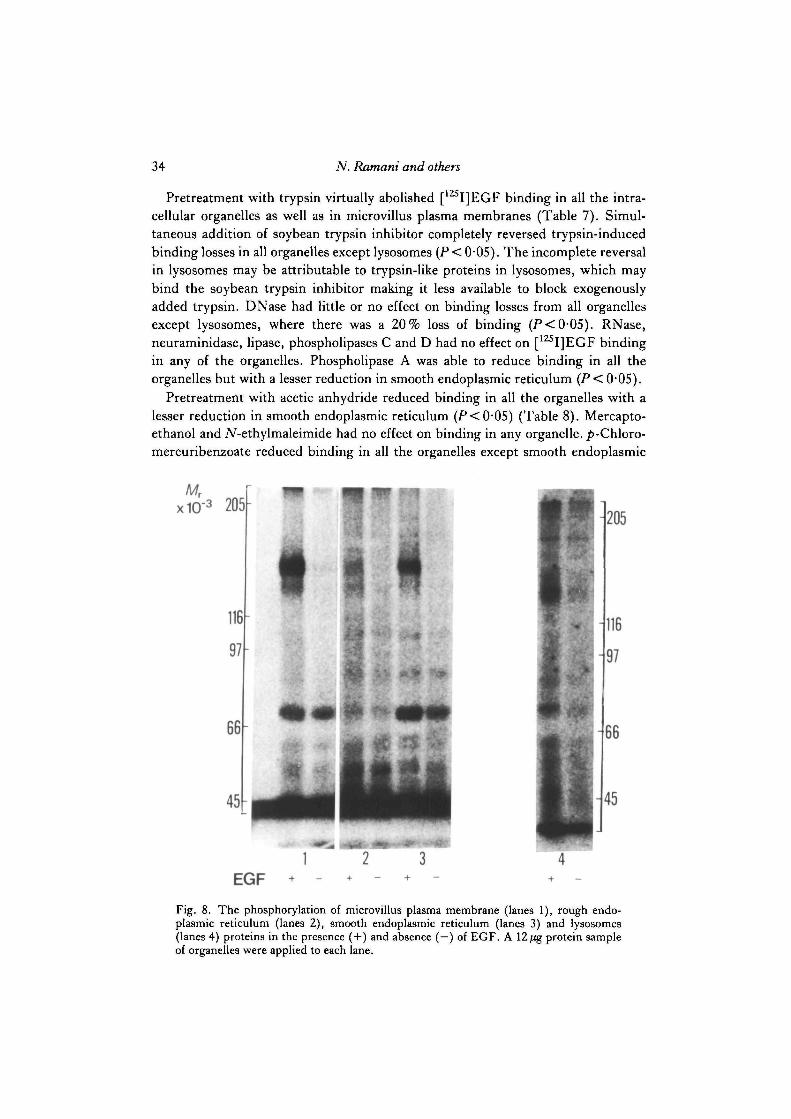
Fig. 8. The phosphorylation of microvillus plasma membrane (lanes 1), rough endo-plasmic reticulum (lanes 2), smooth endoplasmic reticulum (lanes 3) and lysosomes(lanes 4) proteins in the presence ( + ) and absence ( —) of EGF. A 12 fig protein sampleof organelles were applied to each lane.

Intracellular binding of epidermal growth factor 35
reticulum (P<0-05). Dinitrofluorobenzene reduced binding only in rough endo-plasmic reticulum (P<0-05). Tetranitromethane reduced binding in all the sub-cellular organelles to a similar extent.
Molecular mass of [I251]EGF-binding proteins
The property of irreversible, covalent and specific binding of some of the labelledEGF molecules in modified chloramine T iodinated preparations (Comens et al.1982; Baker et al. 1979; Linsley et al. 1979) was used in determining the molecularmass of [ I]EGF-binding proteins.
Microvillus plasma membranes (lanes 1A-1C in Fig. 7), lysosomes (lanes2A—2C), rough endoplasmic reticulum (lanes 3A—3C) and smooth endoplasmicreticulum (lanes 4A-4C), regardless of whether they were prepared in homogenizingbuffers containing Ca2+ (lanes A of corresponding organelles), EGTA (B lanes) ora-2-macroglobulin (C lanes), contain two major (155 and 140(Xl03)Mr) and severalminor (210, 175 and 12O(XlO3)Mr) [12SI]EGF-binding proteins. The 210 and175(XlO3)Afr proteins were only found in microvillus plasma membranes, whereasthe 120xl03Mr protein was only found in lysosomes. The 155x10 Mr protein waspredominantly labelled in microvillus plasma membranes, lysosomes and smoothendoplasmic reticulum, whereas 155 and 140(Xl03)Mr proteins were labelled to anapproximately equal extent in rough endoplasmic reticulum.
Stimulation of membrane protein phosphorylation by EGF
EGF stimulated phosphorylation of 140 and 155(X 103)Mr proteins in microvillusplasma membranes (lane 1 in Fig. 8), rough endoplasmic reticulum (lane 2), smoothendoplasmic reticulum (lane 3) and lysosomes (lane 4). It should be noted, however,that the 155 X 103Mr protein was phosphorylated more than the 140X 103Mr proteinin microvillus plasma membranes and smooth endoplasmic reticulum, whereas the140xl03Mr protein was phosphorylated more than 155xl03Mr protein in lyso-somes. Both the 140 and 155(XlO3)Mr proteins were phosphorylated equally (albeitless than in other organelles) in rough endoplasmic reticulum. EGF also stimula-ted phosphorylation of 45-60, 75 and 180(Xl03)Mr proteins in lysosomes and76X 103Mr protein in smooth endoplasmic reticulum.
DISCUSSION
Human term placentas by virtue of their abundant availability and tissue mass,and being rich in EGF receptors, represent an excellent tissue choice in which toinvestigate whether EGF binding sites are also present in the intracellular organelles.We have made tremendous efforts and used a diverse data base to ascertain whetherplasma membrane contamination could explain intracellular organelle—EGF bind-ing. The following findings: (1) thiamine pyrophosphatase distribution; (2) electronmicroscopy, which shows that all the fractions are quite different from each other;(3) differences in binding properties among various organelles; (4) the magnitude ofbinding, which in some organelles is 71 % of microvillus plasma membranes; and (5)

36 N. Ramani and others
qualitative and quantitative differences among different organelles with respect toproteins covalently labelled with [ I]EGF and the pattern of proteins phosphory-lated in response to EGF, rule out the possibility of contamination.
Various intracellular organelles and microvillus plasma membranes bound [125I]-EGF with a similar affinity. However, the apparent total number of available sitesper mg protein of intracellular organelles was 27—71 % of that found in microvillusplasma membranes. The bound and free [125I]EGF were not degraded in any of theorganelles. The lack of [12SI]EGF degradation, particularly in isolated lysosomes,may be due to alteration of lysosomes during the isolation procedures and, or, lack ofextra lysosomal factors required for lysosomal degradation. It has previously beenreported that isolated lysosomes from appropriate target tissues do not degradehuman chorionic gonadotropin (Chegini et al. 1984; Rao et al. 1981) or prolactin(Ferland et al. 1984).
The organelles exhibited dependency of binding and dissociation on time andtemperature of incubation. The Scatchard plots on all the organelles were curvi-linear. The specificity of binding in all the organelles was similar. pH optima forlysosomal binding was 6-0, in contrast to 7-0 for all the other organelles. Lysosomalbinding losses were lower at pH9-0 and higher when preincubated for 15min at55°C.
Intracellular organelles exhibited numerous differences in binding among them-selves and when compared to microvillus plasma membranes following pretreatmentwith various enzymes and protein reagents. These differences may not necessarilymean that the receptor molecules per se are different, as some of these differencescould have come from differences in the membrane environment in which thesereceptors are located.
The use of covalent affinity labelling of EGF-binding proteins by chloramine Toxidized and iodinated EGF is a qualitative indication of the molecular species thatare capable of binding EGF. All the intracellular organelles, and microvillus plasmamembranes, have two major (140 and 155(xlO3)Mr) [125I]EGF binding proteins. Inaddition, there were two minor (210 and 175(Xl03)Mr) binding proteins in micro-villus plasma membranes and one (120xl03Mr) in rough endoplasmic reticulum.The 155xl03Mr protein was predominantly labelled in microvillus plasma mem-branes, lysosomes and smooth endoplasmic reticulum, whereas both the proteins(140 and 155(X 103)Mr) were equally labelled in rough endoplasmic reticulum.
Replacing Ca2+ with EGTA or ar-2-macroglobulin in buffers during the prep-aration of organelles did not change the pattern of multiple [125I]EGF bindingproteins. This suggests that the multiple binding proteins found were not due totissue proteases unless they have already acted before exposure of the tissue tovarious buffers, and that the multiple binding proteins are normal components ofthese organelles.
EGF stimulated phosphorylation of (155 and 140(xl03)Afr) proteins in all theorganelles. But the amount of phosphorylation of one protein relative to the otherwas different in various organelles. For example, the 155Xl03Mr protein wasphosphorylated more in microvillus plasma membranes and smooth endoplasmic

Intracellular binding of epidermal growth factor 37
reticulum; the 140X 103Mr protein was phosphorylated more in lysosomes and bothproteins were equally phosphorylated (albeit less than in other organdies) in roughendoplasmic reticulum. EGF also stimulated phosphorylation of 45—60, 75 and18O(XlO3)Mr proteins in lysosomes and 75xlO3Mr protein in smooth endoplasmicreticulum. Since the same amount of sample protein was applied to the gel lanes, thedifferences in the phosphorylation pattern may represent actual differences in theamounts of these phosphorylated proteins present in the samples.
Our data, i.e. similar molecular masses of the major [ I ] EGF binding proteinsand proteins phosphorylated in response to EGF in all the organelles, support thedata of Cohen et al. (1982) that the EGF-binding domain and the kinase activity arefunctions of the same protein. The minor [125I]EGF-binding proteins that were notphosphorylated in response to EGF represent perhaps proteins that lack the kinaseand, or, phosphorylation domains. The low molecular weight proteins that werephosphorylated in response to EGF but did not bind [12SI]EGF suggest that theseare protein substrates for EGF receptor kinase action. We cannot rule out thepossibility that the level of detection of phosphorylation in the first case and[125I]EGF binding in the second case, are not sensitive enough to see these proteins.
There are at least three possibilities to explain the presence of EGF binding sites inthe intracellular organelles. (1) Receptors in intracellular organelles represent thosein a catabolic route following internalization. Binding with exogenously added[125I]EGF suggests that the endogenous bound ligand dissociated during theexperimental procedures or during the internalization process. (2) Receptors inintracellular organelles represent newly synthesized receptors. (3) At least aportion of the intracellular organelle receptors are normal components. Internalized[125I]EGF might associate with these organelles leading to the generation ofbiological signals of EGF action. Our data presented in the accompanying paper(Chegini & Rao, 1986) and here show that indeed internalized [125I]EGF associateswith the intracellular organelles and one of the earliest detectable biochemical signalsgenerated in response to EGF binding, i.e. phosphorylation (Carpentered al. 1978;KingeJ al. 1980), is present in the intracellular organelles.
Among these three possibilities, the last one is perhaps the most provocative.Nevertheless, it should be given serious consideration although there is no definitiveevidence to show whether internalized EGF and its receptors can or cannot act insidethe cells.
We thank Mr Bryan Stadig and Dr S. Mitra for performing some of the experiments and MsYvonne Morris for typing the manuscript.
REFERENCESAHARONOV, A., PRUSS, R. M. & HERSCHMAN, H. R. (1978). Epidermal growth factor: Re-
lationship between receptor regulation and mitogenesis in 3T3 cells. J. biol. Chem. 253,3970-3977.
BAKER, J. B., SIMMER, R. L., GLENN, K. C. & CUNNINGHAM, D. D. (1979). Thrombin andepidermal growth factor become linked to cell surface receptors during mitogenic stimulation.Nature, Land. 278, 743-745.

38 N. Ramani and others
BASU, M., FRICK, K., MAJUMDAR, A. S., SCHER, C. D. & DAS, M. (1985). EGF receptor-
associated DNA-nicking activity is due to a Mr 100,000 dissociable protein. Nature, Land. 316,640-641.
BERRY, J. K., MADRID-MARINA, V. & Fox, I. H. (1986). Purification and properties of humanplacental cytoplasmic S'-nucleotidase. J. biol. Chem. 261, 449-452.
BOOTH, A. G., OLANIYAN, R. O. & VANDERPUYE, 0 . A. (1980). An improved method for thepreparation of human placental syncytiotrophoblast microvilli. Placenta 1, 327-336.
BRAMLEY, T. A. & RYAN, R. J. (1978). Interactions of gonadotropins with corpus luteummembranes. I. Properties and distributions of some marker enzyme activities after subcellularfractionation of the superovulated rat ovary. Endocrinology 103, 778-795.
CARPENTER, G. & COHEN, S. (1976). 12sI-labeled human epidermal growth factor binding,internalization and degradation in human fibroblasts. jf. Cell Biol. 71, 159-171.
CARPENTER, G. & COHEN, S. (1979). Epidermal growth factor. A. Rev. Biochem. 48, 193-216.CARPENTER, G., KING, L. & COHEN, S. (1978). Epidermal growth factor stimulates phosphoryl-
ation in membrane preparations in vitro. Nature, Land. 276, 409-410.CARSON, S. A., CHASE, R., ULEP, E., SCOMMEGNA, A. & BENVENISTE, R. (1983). Ontogenesis and
characteristics of epidermal growth factor receptors in human placenta. Am.J. Obstet. Gynecol.147, 932-939.
CHEGINI, N., RAO, CH. V. & CARMAN, F. R. JR (1984). Internalization of 125I-human choriogon-adotropin in bovine luteal slices. A biochemical study. Expl Cell Res. 151, 466—482.
CHEGINI, N. & RAO, CH. V. (1986). Quantitative electron microscopic autoradiogTaphic studies oninternalization of 1Z5I-labelled epidermal growth factor in term human placenta. ,7- Cell Sci. 84,41-52.
COHEN, S. & FAVA, R. A. (1985). Internalization of functional epidermal growth factor:receptor/kinase complexes in A-431 cells. J . biol. Chem. 260, 12351-12358.
COHEN, S., USHIRO, H., STOSCHECK, C. 0 . & CHINKERS, M. (1982). A native 170,000 epidermalgrowth factor receptor-kinase complex from shed plasma membrane vesicles.,7. biol. Chem. 257',1523-1531.
COMENS, P. G., SIMMER, R. L. & BAKER, J. B. (1982). Direct linkage of ]ZS I -epidermal growthfactor to cell surface receptors. A useful artifact of chloramine T treatment. J. biol. Chem. 257,42-45.
DAVIS, B. J. (1954). Disc electrophoresis-II. Method and application to human serum proteins.Ann. N.Y. Acad. Sci. 121, 404-427.
DUNN, W. A. & HUBBARD, A. L. (1985). Epidermal growth factor receptor dynamics in rathepatocytes. J. Cell Biol. 101, no. 5, part 2, abst. 1123.
FERLAND, L. H., DJIANE, J., HOUDEBINE, L.-M. & KELLY, P. A. (1984). Intracellular trans-formation of prolactin following internalization in rat liver. Molec. cell. Endocr. 35, 25—31.
Fox, C. F. & DAS, M. (1979). Internalization and processing of the EGF receptor in the inductionof DNA synthesis in cultured fibroblasts. The endocytotic activation process. J. supramolec.Struct. 10, 199-214.
GOSPODAROWICZ, D. (1975). Purification of a fibroblast growth factor from bovine pituitary.J. biol. Chem. 250, 2515-2520.
GOSPODAROWICZ, D. (1981). Epidermal and nerve growth factors in mammalian development.A. Rev. Physiol. 43, 251-263.
JOHNSON, L. K., BAXTER, J. D., VLODAVSKY, I. & GOSPODAROWICZ, D. (1980a). Epidermal
growth factor and expression of specific genes: Effects on cultured rat pituitary cells aredissociable from the mitogenic response. Pmc. natn. Acad. Sci. U.SA. 77, 394-398.
JOHNSON, L. K., VLODAVSKY, I., BAXTER, J. D. & GOSPODAROWICZ, D. (19806). Nuclear
accumulation of epidermal growth factor in cultured rat pituitary cells. Nature, Land. 2S7,340-343.
JOHNSON, L. K., VLODAVSKY, I. &EBERHARDT, N. L. (1982). Nuclear actions of epidermal growthfactor in rat pituitary tumor cells. In Evolution of Hormone Receptor Systems, vol. 6 (ed. R. A.Bradshaw & G. N. Gill), pp. 397-411. New York: Alan R. Liss.
JONES, P. B. C , WELSH, T. H. JR & HSUEH, A. J. W. (1982). Regulation of ovarian progestinproduction by epidermal growth factor in cultured rat granulosa cells. J. biol. Chem. 257,11268-11273.

Intracellular binding of epidermal growth factor 39
KELLY, T. J., NIELSON, E. D., JOHNSON, R. B. & VESTLING, C. S. (1955). Glucose-6-phosphate
dehydrogenase of adrenal tissue. J . biol. Chem. 212, 545-554.KING, A. C. & CUATRECASAS, P. (1982). Resolution of high and low affinity epidermal growth
factor receptors. Inhibition of high affinity component by low temperature, cycloheximide andphorbol esters. J . biol. Chem. 257, 3053-3060.
KING, A. C , HERNAEZ-DAVIS, L. & CUATEECASAS, P. (1981). Lysosomotropic amines inhibitmitogenesis induced by growth factors. Proc. natn. Acad. Sci. U.SA. 78, 717-721.
KING, L., CARPENTER, G. & COHEN, S. (1980). Characterization by electrophoresis of epidermalgrowth factor stimulated phosphorylation using A 431 membranes. Biochemistry 19, 1524—1528.
LAI, W. H. & GUYDA, H. J. (1984). Characterization and regulation of epidermal growth factorreceptors in human placental cell cultures. J. din. Endocr. Metab. 58, 344-352.
LAI, W. H., GUYDA, H. J. & BERGERON, J. J. M. (1986). Binding and internalization of epidermalgrowth factor in human term placental cells in culture. Endocrinology 118, 413-423.
LEV-RAN, A., HWANG, D., JOSEFSBERG, Z., BARSEGHIAN, G., KEMENY, M., MEQUID, M. &
BEATTY, D. (1984). Binding of epidermal growth factor and insulin to human liver microsomesand Golgi fractions. Biochem. biophys. Res. Cornmun. 119, 1181-1185.
LINSLEY, P. S., BLIFELD, C , WRANN, M. & F O X , C. F. (1979). Direct linkage of epidermal growthfactor to its receptor. Nature, Lond. 278, 745-748.
LOWRY, O. H., ROSEBROUGH, N. J., FARR, A. L. & RANDALL, R. J. (1951). Protein measurementwith the Folin phenol reagent. J. biol. Chem. 193, 265-275.
MADRID-MARINA, V. & Fox, I. H. (1986). Human placental cytoplasmic 5'-nucleotidase. Kineticproperties and inhibition. J. biol. Chem. 261, 444—448.
MARTIN, J. B. & DOTY, D. M. (1949). Determination of inorganic phosphates. Modification ofisobutyl alcohol procedure. Analyt. Chem. 21, 965-967.
MATRISIAN, L. M., LARSEN, B. R., FINCH, J. S. & MAGUN, B. E. (1982). Further purification ofepidermal growth factor by high performance liquid chromatography. Analyt. Biochem. 125,339-351.
MISKIMINS, W. K. & SHIMIZU, N. (1982). Involvement of multiple subcellular compartments inintracellular processing of epidermal growth factor. In Evolution of Hormone Receptor Systems,vol. 6 (ed. R. A. Bradshaw & G. N. Gill), pp. 105-114. New York: Alan R. Liss.
MORIARITY, B. M. & SAVAGE, C. R. JR (1980). Epidermal growth factor and adult rat liverparenchymal cells. In 6th Int. Congr. Endocrinol. Abst. no. S-100.
MROCZKOWSKI, B., MOSIG, G. & COHEN, S. (1984). ATP-stimulated interaction betweenepidermal growth factor receptor and supercoiled DNA. Nature, Lond. 309, 270—273.
PASTAN, I. H. & WILLINGHAM, M. C. (1982). Journey to the center of the cell: Role of thereceptosomes. Science 214, 504—509.
RAO, CH. V., CARMAN, F. R. JR, CHEGINI, N. & SCHULTZ, G. S. (1984). Binding sites forepidermal growth factor in human fetal membranes. J. din. Endocr. Metab. 58, 1034—1042.
RAO, CH. V., MITRA, S. B. & CARMAN, F. R. JR (1981). Characterization of gonadotropin bindingsites in the intracellular organelles of bovine corpora lutea and comparison with plasmamembrane sites. J . biol. Chem. 256, 2628-2634.
RAO, CH. V., RAMANI, N., CHEGINI, N., STADIG, B. K., CARMAN, F. R. JR, WOOST, P. G.,
SCHULTZ, G. S. & COOK, C. L. (1985). Topography of human placental receptors for epidermalgrowth factor. J. biol. Chem. 260, 1705-1710.
RAPER, S. E., BURWEN, S. J., BARXER, M. E. & JONES, A. L. (1985). Intact epidermal growthfactor is translocated to the nucleus during liver regeneration. .7. Cell Biol. 101, no. 5, part 2,Abst. 1327.
SAVAGE, C. R. & COHEN, S. (1972). Epidermal growth factor and a new derivative. Rapid isolationprocedures and biological and chemical characterization.^, biol. Chem. 247, 7609-7611.
SAVION, N., VLODAVSKY, I. & GOSPODAROWICZ, D. (1981). Nuclear accumulation of epidermalgrowth factor in cultured bovine corneal endothelial and granulosa cells. J. biol. Chem. 256,1149-1154.
SCATCHARD, G. (1949). The attractions of proteins for small molecules and ions. Ann. N.Y. Acad.Sd. 51, 660-672.

40 N. Ramani and others
STEEL, R. G. & TORRIE, J. H. (1960). Principles and Procedures of Statistics. New York:McGraw-Hill Book Co.
WORTHINGTON'S ENZYME MANUAL (1972). Lactate Dehydrogenase (ed. L. A. Decker), pp. 7-10.Freehold, New Jersey: Worthington Biochemical Corporation.
(Received 13 February 1986 - Accepted 9 April 1986)