the migratory life history and physiology of arctic char
TRANSCRIPT

The migratory life history and physiology of arctic char, Salvelinus alpinus, navigating change in the Canadian North
by
Matthew James Henry Gilbert
A thesis submitted in partial fulfillment of the requirements for the degree of
Master of Science
in
Physiology, Cell and Developmental Biology
Department of Biological Sciences University of Alberta
© Matthew James Henry Gilbert, 2015

ii
Abstract
Global climate change (GCC) is most pronounced at higher latitudes; to what degree northern
migratory fish species can tolerate this change remains largely unknown. Imminent effects of
GCC on arctic rivers include warmer water temperatures and changes in the timing, frequency
and magnitude of high and low flow events, all of which could reduce fish passage. The arctic
char (Salvelinus alpinus) is a salmonid species of great ecological, cultural, and subsistence
value, and are among the fish species likely to be impacted by these changes. Based on this, my
research aim was to identify life-history and physiological characteristics that likely shape the
ability of arctic char to cope with GCC. To this end, in chapter 2, I characterized alternative
migratory life-history strategies that facilitate the existence of a char population in a harsh
environment. The most significant of these strategies included the earliest documented return
migration timing in the Canadian Arctic and a very low annual fidelity (near 0%) which together,
reduced the exposure of fish to even harsher conditions than those they already faced. In the
physiological components of my research I took a comparative approach and conducted
experiments on rainbow trout, a well-studied temperate reference species, in addition to arctic
char. In chapter 3, I used laboratory simulations to identify potential physiological constraints on
the migration of arctic char through current and future thermal regimes that include large diurnal
temperature fluctuations. In chapter 4, I revealed transcriptional and biochemical responses of
arctic char to these thermal regimes that were indicative of only partially successful
compensatory responses in addition to severe heat-stress and disruption of biochemical
processes. In chapter 4, I also verified the utility of a suite of transcripts as biomarkers for
thermal stress in wild arctic char. Together my research suggests that arctic char possess life
history and physiological traits that may make them more tolerant, and adaptable to GCC than

iii
previously thought but may still place them at a competitive disadvantage relative to more
temperate species whose ranges are expanding northward.

iv
Preface
Experiments and field sampling procedures were approved by the University of Alberta and the
Canadian Department of Fisheries and Oceans Freshwater Institute Science Laboratories Animal
Care Committees (FWI-ACC-2012-019; FWI-ACC-2013-020; FWI-ACC-2014-022; UofA
AUP-022).

v
Dedication:
To Grandpa.
For fostering my love for the fishes of the family salmonidae at a young age.

vi
Acknowledgements
I have placed the acknowledgments for the work presented in individual chapters at the end of
each chapter and thus, here I would like to generally acknowledge those that have had a
significant impact on my success and development as an academic over the course of my masters
program. First and foremost, I would like to thank my family, old and new, for their unwavering
support throughout my adventure when, at times, I know it (I) was difficult. To my wife, I know
the long stretches in the field, long hours in the lab, and the stress that went along with both were
not always to your liking, but you gave your unconditional support and in so doing greatly
lessened the burden. To my mom, thank you for sharing your impressive drive and enthusiasm
for life, and to my dad, thank you for teaching me to ask questions from a young age, and for
always encouraging me to pursue any opportunity I felt worthy. To my brothers, thank you for
your love, friendship and support. To my friends, thank you for helping me find balance and
perspective outside of my studies, and you’re welcome for the information you all now know
about salmon migrations.
As a biologist, I owe a great deal to my many mentors, too many to name. First, I would like to
thank my supervisor, Dr. Keith Tierney, for your sincere efforts in my development as a
professional, for providing me with many great opportunities outside of the scope of a normal
masters program, and for your enthusiastic support in my pursuit of those and future
opportunities. I would like to thank Dr. Rolf Vinebrooke, not only for serving on my committee,
but for sharing his wealth of knowledge in ecology with me, helping to build my foundation in
physiological ecology. Thanks, likewise, to Dr. Heidi Swanson, and Dr. Ted Allison. Your
mentorship during our collaborations had a substantial influence on my success and on my
research philosophy.

vii
Next, I would like to thank my many wonderful lab mates who shared in the many successes and
frustrations encountered along the way. In particular, I would like to thank Brian Blunt and
Angela Shamchuk. Brian, you welcomed me into the lab as an undergrad and very quickly
became a close friend and mentor. Angela, beyond introducing me to my wife, you were always
willing to help shape ideas through our many discussions, and often took on more than your fair
share of responsibility in the lab to keep things running smoothly.
I would like to thank Golder Associates, and the Kugluktuk Hunters and Trappers Organization
for their help in facilitating my field research. I am very appreciative of the many members of the
community of Kugluktuk including the Niptinatiak and Adjun families who provided food,
showers, logistical support, advice, and kindness on many occasions.
Lastly, I would like to express my sincere gratitude to my home community of Fort Nelson, BC,
where I was raised and where I continue to grow as an educator, researcher, and professional
through our on going collaborations.
I am hopeful that my present and future work proves worthy of the contributions so many people
have graciously made.

viii
Table of Contents:
Abstract............................................................................................................................................ii
Preface............................................................................................................................................iv
Dedication........................................................................................................................................v
Acknowledgements.........................................................................................................................vi
Table of Contents..........................................................................................................................viii
List of Tables ..................................................................................................................................x
List of Figures ................................................................................................................................xi
Chapter 1: General Introduction .....................................................................................................1
References...........................................................................................................................4
Chapter 2: Alternative migratory strategies of arctic char (Salvelinus alpinus) in an extreme
environment.....................................................................................................................................7
Abstract ...............................................................................................................................7
Introduction..........................................................................................................................9
Methods..............................................................................................................................11
Results................................................................................................................................16
Discussion..........................................................................................................................19
Acknowledgements............................................................................................................27
References..........................................................................................................................28
Tables.................................................................................................................................34
Figures................................................................................................................................38
Chapter 3: Repeat swimming performance and physiology of arctic char (Salvelinus alpinus) and
rainbow trout (Oncorhynchus mykiss) exposed to diurnal temperature fluctuations ...................46

ix
Abstract..............................................................................................................................46
Introduction........................................................................................................................47
Methods..............................................................................................................................49
Results................................................................................................................................55
Discussion..........................................................................................................................58
Acknowledgements............................................................................................................63
References..........................................................................................................................65
Tables.................................................................................................................................74
Figures................................................................................................................................78
Chapter 4: Diurnal temperature fluctuations induce drastic biochemical and transcriptional
changes in exercised arctic char (Salvelinus alpinus) and rainbow trout (Oncorhynchus
mykiss.............................................................................................................................................84
Abstract..............................................................................................................................84
Introduction........................................................................................................................86
Methods..............................................................................................................................88
Results................................................................................................................................90
Discussion .........................................................................................................................93
Acknowledgements............................................................................................................99
References........................................................................................................................100
Tables...............................................................................................................................106
Figures..............................................................................................................................108
Chapter 5: General Conclusions .................................................................................................114
Thesis bibliography......................................................................................................................121

x
List of Tables
Table 2.1. Length, weight, and condition (K) of Nulahugyuk Creek arctic char at various life-
history stages………………………………………………………………………….………….34
Table 2.2 Age (years) of Nulahugyuk Creek arctic char at various life-history stages……….…35
Table 2.3 Nulahugyuk Creek arctic char length-weight and growth models………..…………..36
Table 2.4 Timing of the upstream migration of various anadromous arctic char populations in
North America……………..…………………………………………………………………….37
Table 3.1. Mophometrics of wild and lab reared juvenile arctic char and rainbow trout ……….74
Table 3.2 Regression models of wild arctic char initial and repeat swimming performance and
recovery ratio over the tested range of naturally occurring
temperatures………………………………………………………..…………………………….75
Table 3.3 Swimming performance, aerobic metabolism and blood analysis of rainbow trout and
arctic char exposed to simulated diurnal fluctuation or diurnal warming……………………….76
Table 3.4 Linear mixed-effects models of arctic char and rainbow trout aerobic metabolism
during and following (recovery) a fatiguing exercise test under reference conditions (11°C) or
following a diurnal warming or fluctuation treatment. Water velocity was increased by 0.5 BL s-1
every ten minutes until fatigue during the exercise test, and held constant at 0.5 BL s-1 during the
recovery period.………………………….………………………….………….………………..77
Table 4.1 List of primers sequences for target genes in the qPCR analysis………..……..……106
Table 4.2 Concentration of select metabolites that exhibited substantial differences between
diurnal treatment groups or species.………………………….….……………………………..107

xi
List of Figures
Fig 2.1. Study area in northwest mainland Nunavut, Canada. Nulahugyuk Creek was the primary
study site; Arctic char captured here feed in the Coronation gulf and Dolphin and Union Sound
region and are thought to overwinter in navigable freshwater systems in the area…………...…40
Fig 2.2 Physical migratory environment of Nulahugyuk Creek, NU. Photos taken at a
standardized location demonstrating the navigability of a representative riffle section mid- (A;
early July) and post migration (B, early August). Creek discharge (C) and water temperature (D)
values shown in white, grey, and black are from 2012, 2013, and 2014 respectively………...…41
Fig 2.3 Daily number of arctic char migrating passed the fish trap by life history stage. The
counts for each life history stage are shown separately for the 2012 (grey) and 2014 (black) field
seasons.……………………….………………………….…………………………....…………42
Fig 2.4 Length frequency distributions of migrating arctic char captured during the 2012 and
2014 field seasons. The data is shown as percentages of either smolts (dark grey) or adults (light
grey)…...……………………….………………………….…………………………..…………43
Fig 2.5 The relationship between length and weight over various life-history stages. Upstream
migrating adults, downstream adults and smolts are shown in dark grey, white, and light grey
respectively. The back transformed two-factor length-weight model is shown by the solid black
line with the 95% prediction interval shown by the dashed grey lines. For model parameters see
Table 2.3..……………………….………………………….…………………………....………44
Fig 2.6 Back-calculated growth in Nulahugyuk Creek arctic char. Boxes represent the
interquartile range around the median, lower and upper whiskers represent the 10th and 90th
percentile respectively and dots represent outliers. The Gompertz growth model is depicted by
the solid grey curve with the 95% prediction interval shown by the dashed lines. For model
parameters see Table 2.3..……………………….………………………….……………………45

xii
Fig. 3.1. Map of the study area in northwest Nunavut, on the Dolphin and Union Strait in the
Arctic Ocean. The study population migrates annually between Hingittok Lake and Bernard
Harbour via Nulahugyuk Creek..……………………….…………….……………….…………79
Fig. 3.2. Natural diurnal temperature fluctuations recorded in Nulahugyuk Creek in 2012 (A) and
the resulting laboratory study design (B). The simulation was designed to assess the effects of
diurnal temperature variation similar to the blowup in (A) on repeat swimming performance and
related sub-organismal physiology……………………….……………………..…….…………80
Fig. 3.3 Initial (top) and repeat (middle) swimming performance and recovery ratio (bottom) of
wild juvenile arctic char over a range of naturally occurring temperatures. The red circle denotes
individuals that were removed as outliers from their respective regression analyses based on
visual inspection. Regression models are in Table 3.2. The individuals removed were the only
two tested above 20°C and may indicate a threshold for recoverability….…….………….……81
Fig. 3.4 Initial and repeat swimming performance and recovery ratio of laboratory reared
juvenile rainbow trout and arctic char following simulated diurnal warming or fluctuation.
Dissimilar letters indicate significant differences between treatments within a species (! 0.05)
……………………………………………………………………………………………………82
Fig. 3.5 Oxygen consumption (MO2) during an initial exercise test (solid lines) and recovery
(broken lines) by laboratory reared juvenile rainbow trout and arctic char exposed to diurnal
temperature treatments. Summary statistics of routine and maximum MO2 are in Table 3.3, and
linear mixed effects models for describing the data are in Table 3.4……….…………………...83
Fig. 4.1 Principle components analysis of plasma metabolite profiles in arctic char and rainbow
trout exposed to simulated diurnal warming and fluctuations. Ellipses represent 68% confidence
limits for each treatment group……………………………….……………………….…..……109
Fig. 4.2 Heat map showing the fold change in concentration of specific metabolites in the plasma
of arctic char and rainbow trout following diurnal temperature treatments……………....……110

xiii
Fig. 4.3 Principle components analysis of transcript profiles in arctic char and rainbow trout
exposed to simulated diurnal warming and fluctuations. Ellipses represent 68% confidence limits
for each treatment group…………….……………………….……………………...…….……111
Fig. 4.4 Relative quantitation of target gene expression following diurnal temperature treatments
in lab reared juvenile arctic char and rainbow trout. Dissimilar letters indicate significant
differences between treatment groups for a given target (!=0.05) ………….…………………112
Fig. 4.5 Relative quantitation of target gene expression following natural exposure to warm
(>19°C) or cool (<15°C) water in migrating adult arctic char. Asterisks indicate significant
differences between warm and cool groups (!=0.05, * <0.01, **<0.001)…………….………113

1
Chapter 1: General Introduction
The effects of global climate change (GCC) are most pronounced at high latitudes, and the
degree to which northern species can cope with this change remains largely unknown. In many
northern rivers, climate-related fluctuations in flow regimes may impair the passage of fish such
as anadromous salmonids (Wrona et al. 2006b). While overall total water input into these
systems may or may not experience large change, the patterns of input and discharge likely will
(Syvitski 2002; Wrona et al. 2006a). More rapid, earlier snow and ice melts may result in fish
experiencing peak flows that are larger and earlier than they historically have been. Conversely
early melts may mean that snow runoff will dissipate earlier causing a similar shift in the timing
of low discharge periods as well as lower minimal discharges (Wrona et al. 2006a). Additionally,
GCC has and will continue to increase the water temperatures in these systems. Such conditions
may favour more temperate species whose ranges are expanding northward (Dunmall et al.
2012).
For migratory northern salmonids, their persistence and competitive ability under drastically
changing conditions will be greatly influenced by an interaction between their migratory ecology
and their physiology (Dowd et al. 2015). Temporal and spatial aspects of these fish’s migratory
life histories determine the environmental conditions they encounter on their migrations while
their exercise and thermal physiology determine their ability to overcome challenging conditions.
In more southerly distributed, relatively well-studied salmonids mismatches between a species’
migratory life-history and its physiology have resulted in reduced migration success (Farrell
2009a). For example, in sockeye salmon, early upstream migrations caused salmon to experience
temperatures above their optimal ranges for physiological performance, which is associated with
reduced migratory success (Eliason et al. 2011; Farrell et al. 2008a; Hinch et al. 2012; Mathes et

2
al. 2010). In northern salmonids, there is a relative paucity of information regarding their
migratory ecology and environmental physiology (Johnson 1980; Klemetsen et al. 2003; Penney
et al. 2014; Roux et al. 2011) so the links between the two are poorly understood.
The arctic char is among the anadromous northern salmonids that will be forced to migrate
though changing thermal and flow regimes. The species is distributed throughout the global
arctic, and possess arguably the greatest life-history diversity of any known vertebrate (Johnson
1980; Klemetsen 2010). This broad distribution and life history diversity makes them an
excellent candidate for the study of local adaptation. The arctic char is also of substantial
cultural, economic, and ecological value (Jenness 1922; Roux et al. 2011) and is closely related
to other northern salmonids including lake trout (Salvelinus namaycush) and dolly varden (S.
malma) that move through similar environments and will therefore experience similar
environmental change. This value and similarity make arctic char a suitable representative
northern salmonid.
Based on the identified knowledge gaps for northern salmonids, the concerning findings
in more temperate salmonids, and the ongoing environmental change my goal was identify life-
history and physiological characteristics that will likely shape the ability of arctic char to cope
with GCC. Within this goal I had three broad objectives to (i) characterize novel migratory life-
history attributes of arctic char that allow them to persist in extreme environments, (ii) identify
physiological constraints on the migration of arctic char under current and future thermal
regimes, and (iii), investigate transcriptional and biochemical responses of arctic char to these
thermal regimes and assess their utility as biomarkers for thermal stress. To achieve these
objectives, I focused on the Nulahugyuk Creek arctic char migration in northwest Nunavut. The
challenging, shallow-flow conditions and thermal variation of the creek allowed me to study life

3
history attributes in an extreme environment (Chapter 2), as well physiological limitations and
responses to thermal variation during migration (Chapter 3 and 4). In the laboratory, I simulated
diurnal temperature variations that were representative of those recorded in Nulahugyuk Creek to
allow me to address objectives two and three with techniques and a level of control not practical
in the field.

4
References
!"#$%&'(&'(%&)(&*(&+,-.&/&0(&'(&!1--2%&3456(&7819:;<&=;9,;>,"-%&>819:;<&1?>91:1@&;-$&>81&
A82@,"<".,B;<& A19C"9:;-B1& "C& ,-$,=,$D;<@(& 781& E"D9-;<& "C& 1?A19,:1->;<& F,"<".2& 35GHI>&
53JK5L6MN5LMO&$",K54(53P3QR1F(55PL3M(&
!D-:;<<%& +(%& E(& S1,@>%& T(& U;9:;BV%& E(& W;F;<DV%& 0(& I(& X1,$1NEY9.1-@1-& /& 0(& !"BV19%& I;B,C,B&
Z;<:"-&,-&>81&*9B>,BK&X;9F,-.19@&"C&U8;-.1(&[-K&S1@A"-@1@&"C&*9B>,B&0;9,-1&TB"@2@>1:@&>"&
U<,:;>1&U8;-.1%&3453(&*<;@V;&Z1;&\9;->%&]-,=19@,>2&"C&*<;@V;&);,9F;-V@%&A&5P5N5M^(&
T<,;@"-%&T(&E(%&7(&!(&U<;9V%&0(&E(&X;.D1%&_(&0(&X;-@"-%&`(&Z(&\;<<;.819%&+(&0(&E1CC9,1@%&0(&+(&\;<1%&!(&
*(& I;>>19@"-%& Z(& \(& X,-B8& /& *(& I(& );991<<%& 3455(& !,CC191-B1@& ,-& >819:;<& >"<19;-B1& ;:"-.&
@"BV121&@;<:"-&A"AD<;>,"-@(&ZB,1-B1&^^3HM436JK54LN53&$",K54(553MQ@B,1-B1(55LL56G(&
);991<<%&*(%&344L(&T-=,9"-:1->%&;->1B1$1->@&;-$&B<,:;>1&B8;-.1K&<1@@"-@&C9":&>81&@>D$2&"C&
>1:A19;>D91&A82@,"<".2&;-$&9,=19&:,.9;>,"-&"C&@;<:"-,$@(&E"D9-;<&"C&T?A19,:1->;<&W,"<".2&
353H3^JK^OO5N^OG4(&
);991<<%& *(%& Z(& X,-B8%& Z(& U""V1%& !(& I;>>19@"-%& \(& U9"@@,-%& 0(& _;A",->1& /& 0(& 0;>81@%& 344G(&
I;B,C,B&@;<:"-&,-&8">&#;>19K&;AA<2,-.&;19"F,B&@B"A1&:"$1<@&;-$&F,">1<1:1>92&>"&A91$,B>&>81&
@DBB1@@&"C&@A;#-,-.&:,.9;>,"-@(&I82@,"<".,B;<&;-$&W,"B81:,B;<&`""<".2&G5HMJKMLONO4L(&
X,-B8%& Z(%& Z(& U""V1%& *(& );991<<%& +(& 0,<<19%& 0(& _;A",->1& /& !(& I;>>19@"-%& 3453(& !1;$& C,@8&
@#,::,-.K& ;& 91=,1#&"C& 91@1;9B8&"-& >81& 1;9<2&:,.9;>,"-& ;-$&8,.8&A91:;>D91&:"9>;<,>2& ,-&
;$D<>&)9;@19&S,=19&@"BV121&@;<:"-&a-B"982-B8D@&-19V;(&E"D9-;<&"C&),@8&W,"<".2&G5H3JK6OMN
6LL(&

5
E1--1@@%&!(%&5L33(&781&_,C1&"C&>81&U"AA19&T@V,:"@%&="<&b"<D:1&c[[K&781&U"AA19&T@V,:"@(&)*&
*BV<;-$(&
E"8-@"-%&_(%&5LG4(&781&;9B>,B&B8;99%&Z;<=1<,-D@&;<A,-D@(&U8;99@K&Z;<:"-,$&C,@81@&"C&>81&.1-D@&
Z;<=1<,-D@&!9&'&ED-V&IDF<,@819@%&781&X;.D1%&d1>819<;-$@K56NLG(&
+<1:1>@1-%& *(%& 3454(& 781& U8;99& I9"F<1:& S1=,@,>1$K& T?B1A>,"-;<& I81-">2A,B& I<;@>,B,>2&
I9":">1@& TB"<".,B;<& ZA1B,;>,"-& ,-& I"@>.<;B,;<& _;V1@(& )91@8#;>19& S1=,1#@& ^H5JKPLNOP&
$",K54(5M4GQC9RN^(5(^(&
+<1:1>@1-%& *(%& I(& *(& *:D-$@1-%& E(& !1:A@"-%& W(& E"-@@"-%& d(& E"-@@"-%& 0(& aeB"--1<<& /& T(&
0"9>1-@1-%& 344^(& *><;->,B& @;<:"-& Z;<:"& @;<;9& _(%& F9"#-& >9"D>& Z;<:"& >9D>>;& _(& ;-$&*9B>,B&
B8;99& Z;<=1<,-D@& ;<A,-D@& H_(JK& ;& 91=,1#& "C& ;@A1B>@& "C& >81,9& <,C1& 8,@>"9,1@(& TB"<".2& "C&
C91@8#;>19&C,@8&53H5JK5N6L(&
0;>81@%&0(&7(%&Z(&\(&X,-B8%&Z(&E(&U""V1%&\(&7(&U9"@@,-%&!(&*(&I;>>19@"-%&*(&\(&_">>"&/&*(&I(&);991<<%&
3454(&TCC1B>&"C&#;>19&>1:A19;>D91%&>,:,-.%&A82@,"<".,B;<&B"-$,>,"-%&;-$&<;V1&>819:;<&91CD.,;&
"-&:,.9;>,-.&;$D<>&'1;=19&U911V&@"BV121&@;<:"-&Ha-B"982-B8D@&-19V;J(&U;-;$,;-&E"D9-;<&
"C&),@819,1@&;-$&*fD;>,B&ZB,1-B1@&MOH5JKO4NGP&$",K54(55^LQC4LN56G(&
I1--12%&U(&0(%&\(&'(&d;@8&/&*(&+(&\;:A19<%&345P(&U;9$,"91@A,9;>"92&91@A"-@1@&"C&@1;#;>19N
;BB<,:;>1$&;$D<>&*9B>,B& B8;9& HZ;<=1<,-D@&;<A,-D@J&;-$&*><;->,B& @;<:"-& HZ;<:"&@;<;9J& >"&;-&
;BD>1& >1:A19;>D91& ,-B91;@1(& U;-;$,;-& E"D9-;<& "C& ),@819,1@& ;-$& *fD;>,B& ZB,1-B1@K5N54&
$",K54(55^LQBRC;@N345^N46ML(&
S"D?%&0(& E(%&S(&)(&7;<<:;-&/&U(&'(&_1#,@%&3455(&Z:;<<N@B;<1&*9B>,B&B8;99&Z;<=1<,-D@&;<A,-D@&
C,@819,1@&,-&U;-;$;e@&dD-;=D>K&:;-;.1:1->&B8;<<1-.1@&;-$&"A>,"-@(&E&),@8&W,"<&OLHMJK5M36N
PO&$",K54(5555QR(54L6NGMPL(3455(4^4L3(?(&

6
Z2=,>@V,%& E(& I(%& 3443(& Z1$,:1->& $,@B8;9.1& =;9,;F,<,>2& ,-& *9B>,B& 9,=19@K& ,:A<,B;>,"-@& C"9& ;&
#;9:19&CD>D91(&I"<;9&S1@1;9B8&35H3JK^3^N^^4(&
'9"-;%&)(& E(%&7(&!(&I9"#@1%& E(&!(&S1,@>%& E(&T(&X"FF,1%&_(&0(&_g=1@fD1&/&'(&)(&b,-B1->%&344M;(&
U<,:;>1&,:A;B>@&"-&*9B>,B&C91@8#;>19&1B"@2@>1:@&;-$&C,@819,1@K&F;BV.9"D-$%&9;>,"-;<1&;-$&
;AA9";B8&"C&>81&*9B>,B&U<,:;>1&[:A;B>&*@@1@@:1->&H*U[*J(&*0W[aK&*&E"D9-;<&"C&>81&XD:;-&
T-=,9"-:1->&^6HOJK^3MN^3L(&
'9"-;%&)(&E(%&7(&!(&I9"#@1%&E(&!(&S1,@>%&E(&T(&X"FF,1%&_(&0(&E(&_g=1@fD1&/&'(&)(&b,-B1->%&344MF(&
U<,:;>1& [:A;B>@& "-& *9B>,B& )91@8#;>19& TB"@2@>1:@& ;-$& ),@819,1@K& W;BV.9"D-$%& S;>,"-;<1&
;-$& *AA9";B8& "C& >81& *9B>,B& U<,:;>1& [:A;B>& *@@1@@:1->& H*U[*J(& *0W[aK& *& E"D9-;<& "C& >81&
XD:;-&T-=,9"-:1->&^6HOJK^3MN^3L&$",K54(56OLQ44PPNOPPOH344MJ^6h^3MKB,";C1i3(4(B"j3(&

7
Chapter 2: Alternative migratory strategies of arctic char
(Salvelinus alpinus) in an extreme environment
Abstract
Anadromous arctic char (Salvelinus alpinus) employ diverse life histories, however, our
understanding of the extent of this diversity is limited. Here, I detail key aspects of the physical
environment and life history of a population of arctic char from Nulahugyuk Creek, Nunavut, to
reveal unique traits. Over the course of their migration creek discharge declined precipitously,
forcing char to migrate through shallow water with large diel temperature fluctuations (>10°C)
and high temperature extremes (>21°C). The downstream migration of adults (>55 cm) began in
mid-June and continued into early July, while the downstream migration of smolts (<30 cm)
began in late June and continued until late July. The upstream adult migration began in late June
and ended in late July; far earlier than other upstream migrations in the region. There was no
appreciable upstream migration of juveniles, and char 30 to 55 cm in length were absent from the
migratory population. The average age at first migration was four years and the youngest
returning adult char were eight and nine years old. The missing size and age classes, and the fact
that most upstream migrants were near reproductive maturity, indicate that char in this system
typically leave at a length of 19 cm and an age of four years and do not return for four to five
years, when they are ready to reproduce. Together our data suggest that anadromous arctic char
possess alternative life history strategies that limit unnecessary exposure to restrictive migratory
conditions and facilitate their existence in otherwise uninhabitable systems. Understanding such
population-specific migratory strategies is critical to the management of arctic fisheries that have
mixed stocks, as well as to our knowledge of arctic char life history diversity that will likely

8
contribute to the adaptability and persistence of the species as global climate change progresses.

9
Introduction
There is a large and well-documented diversity in the life histories of temperate anadromous
salmonids. This diversity undoubtedly contributes to species resilience to exploitation and
environmental changes such as those associated with global climate change (Hilborn et al. 2003;
Schindler et al. 2010). Salmonid life history can vary in spatial and temporal migratory traits
such as population fidelity to natal waters (Keefer and Caudill 2013; Quinn 1993), age at first
migration (Beechie et al. 2006), timing of return migrations, (Beechie et al. 2006; Hodgson and
Quinn 2002; Schindler et al. 2010), and in iteroparous species, migration frequency (Narum et al.
2008). Diversity in migratory life-history traits is also prevalent in northern salmonids, however,
our understanding of its extent is limited in part by their apparent complexity, by a paucity of
long-term datasets, and by the large, remote geographical range of these fish (Gyselman 1994;
Reist et al. 2006a; Roux et al. 2011). Our limited understanding is exemplified by recent
discoveries regarding the fundamental ecology of northern salmonids. For example, northern
populations of anadromous brown trout (Salmo trutta) and arctic char (Salvelinus alpinus) were
first confirmed to occasionally overwinter in estuarine environments in 2012 (Jensen and
Rikardsen), and although it was traditional knowledge among local Inuit and documented by
early expeditionists (Walters 1953), anadromy in arctic lake trout (Salvelinus namaycush) was
only scientifically documented in 2010 (Swanson et al. 2010). Among anadromous northern
salmonids, arctic char are of particular interest because they are an important cultural,
subsistence and commercial resource that has traditionally made up a primary protein source for
the Inuit people (Jenness 1922; Roux et al. 2011). Furthermore, arctic char have the most
northerly distribution of any freshwater fish making them vulnerable to the effects of GCC,
which are most pronounced at higher latitudes (ACIA 2005; Wrona et al. 2006c).

10
The arctic char is a long-lived, cold water salmonid that spawns and initially grows relatively
slowly in cold, oligotrophic freshwater environments. Arctic char possess among the highest
levels of life-history diversity of all vertebrates (Klemetsen 2010). This diversity has been
credited with facilitating their existence in a range of extreme environments throughout the
global arctic (Roux et al. 2011). A key aspect of this diversity is that the arctic char has both
resident and anadromous forms, which allows them to utilize more productive marine
environments when resources are limited in their freshwater habitat. Anadromous arctic char
typically migrate to sea in late spring or early summer and return to their natal freshwater system
in late summer or early fall, after a period of four to eight weeks at sea (Gulseth and Nilssen
2000; Johnson 1980; Johnson 1989). These migrations usually begin between the ages of three
and six years old but this age can vary from two to eleven (Johnson 1980; Swanson et al. 2010).
Once they begin, migrations continue annually or intermittently depending on an individual’s
reproductive status, physical condition, and numerous other environmental factors. The
maximum age of anadromous char in northern Canada ranges between populations from 11 to 28
years old (Roux et al. 2011), which can influence the number of reproductive events an
individual fish may be able to carry out. The annual fidelity of arctic char to their natal system
also varies greatly between populations with straying estimates as high as 53% (Gyselman 1994;
Moore et al. 2013). This level of straying creates a great deal of population mixing, however, an
individual’s likelihood of utilizing a non-natal freshwater systems is lower in spawning years
which limits genetic exchange with other population and likely allows for the persistence of local
adaptation (Moore et al. 2013). Given the relatively small number of anadromous arctic char
populations whose migrations have been thoroughly studied it is likely that there are many
aspects of their migrations to be described.

11
The current study was focused on the Nulahugyuk Creek, Nunavut (NU), arctic char migration.
The traditional harvest of this arctic char run by the Copper Inuit was first documented by an
ethnographer, Diamond Jenness, on the 1913-1918 Canadian Arctic Expedition (Jenness 1922;
Jenness and Jenness 1991). The information he collected suggested that anadromous char in this
area, and in Nulahugyuk Creek in particular, have unique migratory life history traits that have
not previously been studied. Recently, locals have become concerned over a putative decline in
the health of the population. Given these historical observations and local concerns, our goals
were to inform local management decisions and to identify novel aspects of the migration that
would expand our understanding of life-history diversity in anadromous arctic char. To address
these goals we conducted a field study to characterize the key life-history traits and physical
environment of migrating char in Nulahugyuk Creek, NU.
Methods
Field sampling. — Fieldwork took place in the summers of 2012–2014 along Nulahugyuk
Creek, NU, Canada (68.75°N 114.8°W; Fig. 2.1). Nulahugyuk Creek is approximately 10km
long and connects Hingittok Lake to the Arctic Ocean at Bernard Harbour, NU. Arctic char were
captured using two fyke-nets that were set up side-by-side with trap mouths facing in opposite
directions in the main thalweg and wings that extended to the bank on both sides. This trap was
set up approximately 300m upstream from the creek mouth from July 3–22, 2012 and from June
13–July 17, 2014. In 2013, fieldwork took place from August 7–14, however, at that time water
levels were too low to permit fish passage so the trap was not installed. Captured char were
enumerated, their direction of travel was recorded and they were released on the opposite side of
the trap from which they were caught. In 2012, a sub-sample of char were weighed (g) and
measured (mm) and adult adipose fins were clipped. Subsampling was done to limit time in

12
captivity, particularly under warm water conditions (> 18°C). In 2014, length and weight data
were collected for all adults and up to 20 smolts each time the trap was cleared.
In 2012, a subsample of adults (n = 121) were implanted with half duplex passive integrated
transponder tags (HDX PIT; Oregon RFID, Portland, Oregon) in their peritoneal cavity. In 2014,
upstream migrating adults received PIT tags (N = 303) and downstream migrating adults were
implanted with a numbered dorsal T-bar tag (N = 498; Orange tags series 0001-1061, Floy Tag
Incorporated, Seattle, Washington). Tagging was completed as part of an ongoing study being
conducted by Golder Associates (Edmonton, Alberta, Canada).
The lower three kilometers of the creek below a large pool, was regularly surveyed for arctic
char mortalities. The location, length and weight data, reproductive status, and otolith and
adipose fin samples were collected from any recent mortalities that were found sufficiently
intact. These samples and data were also collected from fish harvested by locals when permitted.
Reproductive status was determined using a sexual maturity key (McGowan 1992). In 2013, five
char were harvested by locals near the mouth of the creek in Bernard Harbour using either gill
nets or angling. These fish were also sampled for comparison to char captured in Nulahugyuk
creek. Sampling procedures were approved by the Canadian Department of Fisheries and Oceans
Freshwater Institute Science Laboratories and University of Alberta Animal Care Committees
(FWI-ACC-2012-019; FWI-ACC-2013-020; FWI-ACC-2014-022; UofA AUP-022).
Physical Environment. — Water temperature was recorded at the fish trap every 15 min using
HOBO Water Temperature Pro data loggers (Onset Computer Corporation, Bourne
Massachusetts). In 2014, water temperature was also recorded every time the trap was cleared
after July 1st 2014 . Creek discharge was measured using the velocity-area method at a site with

13
relatively uniform flow, 200m upstream from the trap but still below all branch’s and sources of
inflow. Velocity measurements were taken at 60% depth at a minimum of 20 intervals using a
magnetic velocity meter (2012 and 2013: FP111, Global Water Instrumentation, College Station,
Texas; 2014: Model 2100 Current Velocity Meter, Swoffer Instruments, Seattle, Washington).
Otolith Analysis. — Otolith analysis was conducted as previously described (Swanson et al.
2010). In brief, after sampling in field, otoliths were stored dry in a paper envelop until analysis.
One of each otolith pair was embedded in epoxy (2012 and 2013: EpoThin, Buehler, Lake Bluff,
Illinois; 2014: Cold Cure, System Three, Auburn, Washington), sectioned through the nucleus
using a precision, low-speed saw (ISOMET, Buehler, Lake Bluff, Illinois), embedded in epoxy
inside an acrylic ring with the surface of the cross section exposed, and then polished using a
lapping paper (50 – 3µm Grit). Samples that were used in the otolith microchemisty analysis
were further polished using a 0.5µm polishing paste on a Grinder-Polisher (MetaServ™ ,
Buelher; Lake Bluff, Illinois) and washed in an ultrasonic bath.
Laser ablation inductively-coupled mass spectrometery (LA-ICP-MS) was performed on all 2012
and 2013 samples at the Advanced Instrumentation Laboratory (University of Alaska Fairbanks,
Fairbanks, Alaska) as in Swanson et al. (Swanson et al. 2010) with minor differences. For laser
ablation, we ablated a single transect from the primoridia to beyond the distal edge of the otolith
using a New Wave UP213 laser (Fremont, California) set to 80% power, a 25 "m spot size and a
5 "m s-1 scan speed. Specific element isotope concentrations were determined using an Agilent
7500ce mass spectrometer (Santa Clara, California) that was coupled to the laser and calibrated
with a glass standard (SRM 610, National Institute of Standards and Technology, Gaithersburg,
Maryland). 86Sr and 66Zn isotope concentrations were plotted over images of their corresponding
otolith along the laser ablation line. In fish that had previously migrated, the age at first

14
migration (AFM) was determined by counting annuli up to the point of increase in [Sr86]
(Swanson et al. 2010). The AFM was verified by counting the number of annual cycles of Zn66
prior to migration.
Length at age values were back-calculated using an otolith radius vs. length, regression approach
to fill in length data for the missing size class between smolts and adults. A regression approach
was used because the radius-length relationship was sigmoidal and was not readily transformable
into a linear relationship for use with the biological intercept model (Stevenson and Campana
1992). For each otolith cross-section, the radius was measured at the outer edge of every hyaline
zone along the longest line of the ventral lobe from the primordium to the distal edge using
ImageJ 1.48 (National Institute of Mental Health, Bethesda, Maryland). The four-parameter
sigmoidal regression between the total ventral otolith radius and fork length at capture had the
formula:
! ! !"#!!! !"#!!
!! ! !! !!!!"#!!"#!"#!!
Where R is the ventral otolith radius (µm) and L is fork length (mm). The radius at each annuli
after age three was put into the length-radius formula to calculate the length at age for all
sampled individuals. Age three was selected as the cut off for back calculation to avoid
extrapolation outside of the dataset used to generate the sigmoidal regression model.
Data analysis. — Arctic char less than 350mm in length were considered to be smolts based
upon their seaward trajectory, proximity to the ocean, size (length and weight) difference from
adults, and lack of prior migration to marine environments as determined by Sr isotope analysis.
All arctic char greater than 500mm in length were considered adults. Only three individuals

15
(<0.1%) fell between the two life-history classes and were therefore omitted from the analysis.
Fish recaptured less than one week after release, including all smolts captured in the upstream
trap, were also not included in the statistical analysis as they likely reentered the trap accidently.
The possible implications of this omission are discussed below. Length frequency distributions
were calculated for smolts and adults in each year (2012 and 2014). Distributions were compared
between years using separate Kolmogorov–Smirnov tests (K-S test) for smolts and adults.
Differences in length, weight and condition factor between years, migratory direction and life-
history stage were analyzed using an ANOVA with a tukey HSD post-hoc pairwise comparison
test. Condition factor was calculated as K = 105 x W/L3, where W is wet weight (g) and L is fork
length (mm).
Linear regression models were constructed to examine relationships between length and weight.
The length-weight relationships were established using log10-transformed data. In addition to the
conventional length-weight model, a second model was generated that included life-history stage
(smolt and upstream and downstream adults). Given the sigmoidal nature of the back-calculated
length at age data, growth was modeled using three and four parameter sigmoidal and Gompertz
curves as well as a logistic curve. The four-parameter Gompertz curve had the highest R2 and
lowest residual standard error (RSE) so it was selected as the final growth model. In all analyses
with multiple predictor variables all interactions were tested. If an interaction effect was not
significant (!=0.05) it was removed from the analysis as long as no higher order interaction was
significant.
Statistical analysis was completed in R Studio (Team, 2014). Data presentation was completed
using Sigmaplot 11.1 (Systat Software, San Jose, California) and Adobe Illustrator CS4 (Adobe
systems, San Jose, California).

16
Results
Creek temperature and discharge. — Discharge in Nulahugyuk creek was low and declined over
the course of the arctic char migration (6.00-0.50 m3/s; Fig. 2.2A-C). Low discharge resulted in
numerous incidences of upstream migrating adults becoming stranded and being scavenged or
preyed upon by gulls (Larus spp.) and grizzly bears (Ursus arctos). In August, after the run was
complete, flow was sufficiently low to restrict arctic char passage altogether (0.03 m3/s; Fig.
2.2B-C). In 2013, a single adult char was observed attempting to migrate upstream but became
stranded within 300m of the mouth of the creek. In 2012 and 2013, there were large diurnal
temperature fluctuations occasionally exceeding 10°C per day (Fig. 2.2E). The minimum and
maximum temperatures recorded during the course of the 2012 migration were 7.2 and 21.0°C,
respectively. The minimum and maximum temperatures recorded manually at trap checks in
2014 were 7.0 and 19.8°C but may not represent the true minima and maxima of the creek due to
the low sampling frequency in this year.
Migration size and timing. — Seaward adult arctic char were the first to enter Nulahugyuk Creek
after ice breakup. In 2014, they were first captured moving out to sea on June 16th; the migration
peaked June 21st, gradually tapered off, and was near completion by July 3rd (Ntotal = 472; Fig.
2.3 B). In 2012 the downstream migration appeared to be near completion shortly after the trap
was installed on July 4th as a relatively small number of downstream adults were captured (Ntotal
= 84; Fig. 2.3B). The 2014 adult upstream migration began June 25th, peaked on July 3rd, and
appeared to be near completion upon our departure on July 17th (Ntotal = 332 , Fig. 2.3A) . This
pattern generally agrees with 2012 as a large number of adults were migrating upstream when
the trap was installed and the run had dissipated by July 20th (Ntotal = 387; Fig. 2.3A). The period
of low upstream migration frequency in 2012 between July 12th and 16th corresponded with the

17
arrival of a large amount of drift ice that may have blocked sea access to Bernard Harbour. In
either year only one adult trapped on their downstream migration was captured subsequently
moving upstream after a period of more than a week suggesting that nearly all individuals that
left the creek did not return the same year. There was a large disparity in the seaward smolt
migrations in 2012 and 2014. In 2014 only 460 smolts were captured while in 2012, 6221 smolts
were captured (Fig. 2.3C). In 2014 the first smolt was captured on June 25th, and capture was
irregular until the end of sampling (July 16th). In 2012 the smolt migration appeared to peak
shortly after the trap was installed (July 4th) and end by July 23rd. These movement patterns
indicate overlap between upstream adult and downstream adult and smolt migrations (Fig. 2.3).
In 2012, three adult char were captured migrating upstream with floy tags from a previous study
that consisted of tagging fish near the mouth of the Coppermine River, NU (Fig. 2.1). In 2014
two tagged adults from the same tag-series were captured moving out to sea.
Size and maturity. — Two distinct and normally distributed length-frequency groups were
apparent within the population and corresponded to smolt and adult life history stages (smolt:
150 to 300 and adult: 550 to 900mm; Fig. 2.4). Smolts and adults were separated by a near
complete absence of fish between 300 and 550 mm (N = 3, Fig. 2.4). In 2014 there was a
rightward shift (increased length) in both the smolt and adult length-frequency distributions
relative to 2012 (K-S test: Smolt: D = 0.229, p < 0.001, N2012 = 168, N2014 = 302; Adult: D =
0.536, p < 0.001, N2012 = 321, N2014 = 782). The shift in length frequency distributions agreed
with comparisons of mean length and weight, which revealed that fish captured in 2014 were
generally larger (length and weight) than those of the same life history stage in 2012 (Tukeys
HSD: p < 0.05; Table 2.1). Arctic char migrating to the sea (downstream adults and smolts) had a
substantially lower condition factor (i.e. were relatively thinner) than those returning from sea (-

18
26 to 28%; Tukeys HSD: p < 0.05; Table 2.1). In 2014, natural mortalities were larger than
average upstream adults (length: +8.2%; weight +26.0% Tukeys HSD: p < 0.05; Table 2.1). Like
in most salmonids, Nulahugyuk Creek arctic char exhibited an exponential length-weight
relationship (R2 = .996, RSE = 0.054, F998 = 2.6e5, p < 2.2e-16; Fig. 2.5; Table 2.3 ), however,
this relationship varied depending on life history stage so it was better represented by a model
including it as an explanatory variable (RSE = 0.042, R2 = 0.998, F996 = 1.38e5, p < 2.2e-16; Fig.
2.5; Table 2.3 ).
In 2014, 16 adult char were available for assessment of their sexual maturity. Of a total of five
downstream migrating adults, two females and two males had resting characteristics suggesting
they had spawned upstream in the previous year, and one female had developed but had firm
ovaries suggesting it had not fully matured and did not previously spawn. Of the 11 upstream
migrating adults sampled, all six males and one of the females were sexually mature and in
spawning condition. Three females had not fully matured but had clearly distinguishable eggs
likely capable of maturation within the year, and one female was in resting condition.
Age, growth and otolith microchemistry. — The visual otolith based age estimate for smolts was
almost identical to AFM obtained from adult otoliths through Sr isotope analysis (Table 2.2).
The difference between smolt age/AFM and the youngest returning adults was four to five years.
The measured and back-calculated length at age relationships were sigmoidal with a relatively
high growth between the ages of smoltification and return to their natal system (Fig. 2.6, Table
2.3). Together, the age difference between life-history stages and the age-length relationship
reveal that the gap in the length frequency distributions corresponds with an absence of
individuals typically between the ages of four and nine years old.

19
Discussion
Arctic char possess extraordinary life history diversity that is thought to facilitate their existence
in a range of extreme and variable environments (Hammar 2014; Johnson 1980; Jonsson 2001;
Klemetsen 2010; Roux et al. 2011). We have identified novel migratory life history strategies
within a small arctic char population that demonstrate that this diversity is even greater than
previously established. Specifically, Nulahugyuk Creek arctic char typically do not return to their
natal watershed for four to five years after their first migration, likely resulting in extensive
mixing with surrounding populations. Furthermore, returning adult char begin their upstream
migration in late June and it is complete by mid to late July, when most other return migrations
have not yet begun. Given that Nulahugyuk Creek has a highly variable thermal regime, is
marginally navigable at the best of times, and completely impassible by early August, these life
history traits are likely adaptive and facilitate the persistence of anadromous arctic char in an
otherwise uninhabitable system.
Size and age structure. — The length of smolts at first migration (Table 2.1 and 2, Fig. 2.4) in
Nulahugyuk Creek (Table 2.3) falls well within the published range (eg. 180 – 240mm; Johnson
1980; Johnson 1989). Interestingly, the mean smolt length was very similar to the estimate of
eight inches (203mm;Jenness 1922), which ethnographer Diamond Jenness made at the same
location in 1916 of what he postured were “…young fry, two seasons old, making their way
down to the sea.” The age at first migration (3-5 years) is on the lower end but within the
published range for the region (3-11 years; Johnson 1980; Johnson 1989; Swanson et al. 2010).
The similarity between the current smolt age distribution and the age at first migration estimated
from Sr profiles of adult otoliths confirms that this life-history trait is consistent between
generations.

20
Adults were similar in mean and maximum length and weight to those in the nearby Coppermine
River, Nu population (Fig. 1;Gillman and Kristofferson 1984), but larger than in many other
populations in the Canadian Arctic (Roux et al. 2011). Nulahugyuk Creek arctic char also had
similar maximum ages to those in Coppermine river, which are lower than many other high
latitude populations (Roux et al. 2011). The large size and short life-span of these two
populations mean that they have among the highest lifetime growth rates in the region (Table 3;
Fig. 6; Gillman and Kristofferson 1984; Swanson et al. 2010). Such a rapid growth but relatively
short life span (12-14 years) suggests that the Dolphin Union Strait – Coronation Gulf area (Fig.
2.1) might be one of relatively high productivity where arctic char may have fewer total
reproductive events but greater fecundity than in surrounding areas where fish may have life
spans twice as long but reach similar or smaller sizes. For example, the length and weight of
adult arctic char in this study had substantial overlap with the length and weight at sexual
maturity for the Nauyuk Lake population, however, this size was reached at a much earlier age in
our study population (8-12 years vs. 10–18 years; Johnson 1980). The fact that natural
mortalities were larger than other upstream migrating adults suggests that migration success is
likely size limited, perhaps due to increase likelihood of stranding for larger individuals, which
would also make them more susceptible to predation. Furthermore, the size limitation may also
explain the relatively short life expectancy when compared to Coppermine River char (12 vs. 14
years).
The length-weight relationship of Nulahugyuk Creek arctic chars aligns well with other
populations in the region (Johnson 1980). As was found in Nauyuk Lake, this relationship
differed between life-history stages (Table 3; Johnson 1980). For juveniles, the observed
reduction in the intercept compared to upstream adults likely owes to their feeding and growing

21
in an oligotrophic environment compared to the anadromous adults. In adults, the lower intercept
in downstream migrants is almost certainly due to the high energy costs associated with
spawning and overwintering in freshwater where anadromous adults do not typically feed (Dutil
1986; Johnson 1980). These differences in length-weight relations are also apparent in the lower
condition factor that is found in char coming from freshwater compared to those coming from
the ocean (Table 2.2). The decreased condition factor in seaward adults parallels that seen in
Nauyuk Lake, where it was estimated that the cost of overwintering and spawning could be up to
46% of total body energy (Dutil 1986).
Migration pattern. — The seaward adult migration began shortly after the outflow of Hingittok
Lake was free from ice obstruction. This timing is in line with that commonly observed in most
other systems (Johnson 1980). The smolt migration was also similar to other populations in that
it began shortly (nine days) after the adult downstream migration and was highly variable
between years (Johnson 1980). As Nulahugyuk creek was likely still navigable for smolts when
sampling ceased on July 16th 2014, we may have missed a significant portion of the 2014 smolt
migration, however, that would mean that the timing of the run in 2014 was substantially later
than in 2012.
To the best of our knowledge, the timing of the adult return migration is the earliest ever
documented in the Canadian Arctic, beginning in late June and concluding before in the end of
July. All other documented upstream migrations either start or continue well into August and
usually into September (Table 4; Johnson 1980). The only known upstream migration that is
similar in timing occurs entirely in freshwater, when arctic char migrate from Nauyuk Lake up
Willow Creek, a small intermittent stream, to their spawning grounds at Willow Lake (Johnson

22
1980; Johnson 1989). Like Nulahugyuk Creek, water levels in Willow Creek are too low to
permit passage into August (Gyselman 1984; Johnson 1980) necessitating an early migration.
The adult char migrating upstream in Nulahugyuk Creek had not previously migrated
downstream in the same year. For returning adult char to arrive as they do, they would likely
have to migrate directly from their overwintering grounds with little to no time spent feeding at
sea, and indeed, all observed migrants had empty stomachs. There are many possible creeks and
rivers such as Noahognik, Kogluktuaryuk and the Coppermine river that are within an accessible
distance and could be utilized as overwintering habitat (Fig.1; Gillman and Kristofferson 1984;
Jenness 1922). This overwintering theory is supported by the capture of char in Nulahugyuk
Creek that were tagged near the mouth of the Coppermine River in a previous study.
Furthermore, in a study conducted on the Nauyuk Lake arctic char, Nauyuk char were identified
in nearly every freshwater system in the area (<250km) in which there was a significant fishery,
within the same year the fish were tagged (Gyselman 1984).
Fidelity. — In semelparous salmonids, homing and straying are terms with relatively simple and
rigid definitions. In reference to these fish’s once in a life time return migration, homing is the
act of returning to natal spawning grounds for the purpose of reproduction, while straying is
entering and reproducing in a non-natal freshwater system (Quinn 1993). Under these
definitions, benefits to homing include that individuals return to a location where their parents
had reproductive success and that they may be uniquely adapted to (Quinn 1993). Straying,
however, allows salmon to potentially colonize new systems that may have less competition,
greater productivity and may not be as challenging to access. In iteroparous salmonids, the
definitions of homing and straying are not as clear. Individuals make multiple migrations over
the course of their lives and may utilize more than one freshwater system. Furthermore, it is

23
often difficult and costly to determine whether or not straying individuals have entered other
freshwater systems to spawn or simply to overwinter. Here we use the terms in respect to the
annual migration regardless of reproductive status. Under these definitions, reported ranges of
annual stray after accounting for at sea mortality vary from 2.2 to 66% (Gyselman 1994; Jensen
et al. 2015). The upper end of this range (66%) occurred in a single year for the Nauyuk Lake
population, however, there was annual variation in the rate of stray of 33-66% that resulted in a
mean straying rate of 53% (Gyselman 1994). Using a genetic population assignment method for
populations in the Cumberland Sound region, NU, Moore et al. (2013) produced course estimates
of straying rate between 15.8 and 46.5% and more refined estimates that ranged from 15.8 to
25.5%. In our study, there was no appreciable return of either smolts or adults within the same
year of their downstream migration, suggesting that there was a near 100% annual straying rate
for both life history stages. The near complete absence of fish between the ages of five and eight,
and the size of 300 and 550mm suggests that once smolts enter the sea, they do not return until
they have reached or are near reproductive maturity. Increased fidelity among reproductive
individuals has been identified in the Cumberland Sound region, where individuals identified as
strays were much more likely to be overwintering than reproducing (Moore et al. 2013), while in
Nauyuk Lake, small, likely non-reproductive char, and char that had spawned in the previous
fall, tended to have a lower fidelity than those that were in reproductive size classes but had not
spawned in the previous year (Gyselman 1994). Furthermore, in our study, all but one of the
eleven upstream migrants examined appeared to be in suitable condition for spawning in the fall,
and all but one of the five seaward adults examined appeared to have spawned in the previous
year. In addition to the on going tagging study taking place in the region, incidence of

24
Nulahugyuk char mixing with surrounding populations for the purpose of reproduction could be
relatively easily resolved through genetic methods.
In a study of the migratory window of Dieset River Arctic char in Svlabard, Norway, Svenning
and Gullestad (2002) found that in some years with high air temperature, flows could become to
low to permit the return of juvenile char in late August. The authors presumed that all individuals
stranded at sea likely died overwinter, however, our results suggest that these restrictive
migratory conditions may lead to the use of non-natal freshwater systems for overwintering
rather than the stochastic high mortality events suggested. Furthermore, there are other systems
that support anadromous char populations on Svalbard that could provide suitable alternative
overwintering habitat when the migratory window of Dieset River closes early.
The suggested pattern of fidelity of Nulahugyuk Creek arctic char would mean that they benefit
from aspects of both straying and homing strategies. Their low fidelity and use of alternative
overwintering habitat while immature or between reproductive events would limit the number of
lifetime migrations that they would need to make through Nulahugyuk Creek, which has a
narrow migratory window, and is thermally and physically challenging to navigate (Fig.1). Their
return upon reproductive maturation would provide access to proven breeding grounds and
would help maintain genetic isolation from surrounding populations, perhaps allowing for the
development and persistence of local adaptations (Moore et al. 2013; Quinn 1993). The idea of
utilizing “convenient” overwintering habitat rather than returning annually to natal waters has
been discussed previously (Armstrong and Morrow 1980; Moore et al. 2013), and Johnson
(1980) suggested that this may be an adaptation to the variable flows of streams that many arctic
salmonid populations rely on for migrations; The Nulahugyuk Creek arctic char population
provides one of the most pertinent examples in support of this hypothesis to date.

25
In our study, upstream migrating juveniles were assumed to be smolts intending to move out to
sea that mistakenly re-entered the trap after their release downstream. This is supported by a
number of observations; their captured frequency was highly correlated with the release of
smolts moving downstream (R2 = 0.79), there was no apparent difference in size between the
groups as there would be if the upstream migrants had fed at sea, and most convincingly, smolts
released on the downstream side of the trap were frequently observed swimming directly into the
upstream trap following release, likely in an attempt to flee researchers working nearby. Despite
this, we cannot say with certainty that some juveniles did not come from nearby freshwater
outlets or that some downstream migrants briefly (hours-days) ventured out in to brackish water
before returning as has been noted in other systems (Johnson 1980). This behavior would be
interesting to quantify as it is indicative of juveniles assessing their seawater tolerance before
migration, however, it does not affect any of our primary conclusions, as these individuals
evidently did not spend any meaningful amount of time at sea. Upstream migrating adults that
were previously captured in the downstream trap the same year were also assumed to have
mistakenly re-entered the trap rather than be attempting a return migration as most recaptures
occurred with one day of release. In 2014 only one of 471 tagged downstream adults was
captured moving upstream more than a week after its initial release, and this individual had been
recaptured in the interim indicating that it likely never went to sea.
Management Implications. — The Nulahugyuk Creek arctic char population provides examples
of the challenges of managing a species with extensive life history diversity, particularly in an
era of accelerated global climate change. Our results suggest that nearly all migrants in
Nulahugyuk Creek utilize non-natal systems to overwinter and consequently they are likely
mixing with other, more heavily exploited populations without management consideration (Roux

26
et al. 2011). While many arctic char fisheries are recognized as mixed stock, the extent of
straying in our population exemplifies the need for the development of associated tools such as a
thorough genetic stock assignment library to allow researchers and managers to better resolve
stock composition. Furthermore, the basic nature of our findings suggests that there are likely
still life-history aspects of anadromous arctic char that are not understood well enough to make
well-informed management decisions. For example, if as we propose that some Nulahugyuk
Creek arctic char overwinter in the Coppermine River and are among the first to enter the sea in
spring, they may be disproportionately targeted by the early gillnet fishery that occurs
immediately after break up at the River’s mouth.
The Nulahugyuk Creek arctic char are also an example of a population that is likely vulnerable to
the effects of global climate change. The creek already has a narrow migratory window caused
by a rapid decline in discharge between early June and late July, and already experiences
temperatures that are well beyond the optimal range and near lethal levels (Elliott and Elliott
2010; Quinn et al. 2011a). These conditions have likely already worsened as a result of climate
change and will continue to decline as air temperatures rise causing a reduction in the amount of
spring snowpack and an increase in water temperature within the creek (ACIA 2005; Wrona et
al. 2006c).
While life history diversity certainly presents difficult management challenges, it can also protect
species against exploitation and environmental change (Schindler et al. 2010). Nulahugyuk
Creek arctic char possess unique life history attributes that may also make them an ideal source
population if the aforementioned changes in snowpack and temperature make other surrounding
systems uninhabitable for the current inhabitants that are not adapted to comparatively harsh
migratory conditions and a short migratory window. In addition to the stabilizing effect of life

27
history diversity within a species, this diversity can also directly benefit other dependent species.
For example, in sockeye salmon, variation in run timing serves to increase the feeding window
of predators such as Grizzlies and Gulls, as these predators can have prolonged access to salmon
by moving stream to stream based on the timing of a local run (Schindler et al. 2013). This
phenomenon has not been studied for predation of arctic char, however, given the unique timing
of the Nulahugyuk run and the prevalence of grizzly and gull predation on char that we observed,
it is possible that this run significantly extends the period of availability of arctic char for these
predators. Given the potential benefits of life history diversity, when management plans are
being developed, the intent should be to not only maintain individual population size, but also to
preserve their unique traits (Potter et al. 2003).
Acknowledgements
We would like to thank all of the members of the community of Kugluktuk, NU who provided
support throughout the study. We are very grateful to the Kugluktuk Hunters and Trappers
Organization (HTO) and the Nunavut Department of Environment for facilitating and
participating in field operations. We also thank Dr. Christian Zimmerman from the USGS for
conducting the otolith microchemisty analysis. Funding was provided by the Kugluktuk HTO,
Golder Associates Ltd., a National Science and Engineering Research Council (NSERC)
Discovery grant to KBT, and grants and awards to MJHG from NSERC, the Northern Scientific
Training Program, Canadian Circumpolar Institute, Canadian Society of Zoologists, the
Association of Canadian Universities for Northern Studies and the University of Alberta.

28
References:
*U[*%&3446(&*9B>,B&B<,:;>1&,:A;B>&;@@1@@:1->N@B,1->,C,B&91A"9>(&U;:F9,$.1&]-,=19@,>2&I91@@(&
*9:@>9"-.%&S(&X(&/&E(&0"99"#%&5LG4(&781&$"<<2&=;9$1-(&U8;99@K&@;<:"-,$&C,@81@&"C&>81&.1-D@&
Z;<=1<,-D@KLLN5P4(&
W11B8,1%&7(%&T(&WD8<1%&0(&SDBV1<@8;D@%&*(&)D<<19>"-&/&_(&X"<@,-.19%&344M(&X2$9"<".,B&91.,:1&
;-$&>81&B"-@19=;>,"-&"C&@;<:"-&<,C1&8,@>"92&$,=19@,>2(&W,"<".,B;<&U"-@19=;>,"-&5^4HPJK6M4N
6O3&$",K54(545MQR(F,"B"-(344M(45(45L(&
!D>,<%&E(&!(%&5LGM(&T-19.1>,B&U"-@>9;,->@&;-$&ZA;#-,-.&[->19=;<&,-&>81&*-;$9":"D@&*9B>,B&
U8;99&HZ;<=1<,-D@&;<A,-D@J(&U"A1,;&5LGMHPJKLP6NL66(&
T<<,">>%&E(&/&E(&T<<,">>%&3454(&71:A19;>D91&91fD,91:1->@&"C&*><;->,B&@;<:"-&Z;<:"&@;<;9%&
F9"#-&>9"D>&Z;<:"&>9D>>;&;-$&*9B>,B&B8;99&Z;<=1<,-D@&;<A,-D@K&A91$,B>,-.&>81&1CC1B>@&"C&
B<,:;>1&B8;-.1(&E"D9-;<&"C&C,@8&F,"<".2&OOHGJK5OL^N5G5O(&
\,<<:;-%&!(&b(&/&*(&X(&+9,@>"CC19@"-%&5LGP(&W,"<".,B;<&$;>;&"-&*9B>,B&U8;99%&Z;<=1<,-D@&;<A,-D@&
H_(J%&C9":&>81&U"AA19:,-1&S,=19%&d"9>8#1@>&7199,>"9,1@%&5LG5NG3(&U;-;$,;-&!;>;&S1A"9>&"C&
),@819,1@&;-$&*fD;>,B&ZB,1-B1@&PP4K,=&k&5M&A(&
\D<@1>8%&a(&*(&/&+(&E(&d,<@@1-%&3444(&781&W9,1C&I19,"$&"C&ZA9,-.&0,.9;>,"-%&Z8"9>&0;9,-1&
S1@,$1-B1%&;-$&X,.8&S1>D9-&S;>1&"C&;&d"9>819-&Z=;<F;9$&I"AD<;>,"-&"C&*9B>,B&U8;9(&
79;-@;B>,"-@&"C&>81&*:19,B;-&),@819,1@&Z"B,1>2&53LH^JKOG3NOLM&$",K54(56OOQ56PGN
GM6LH3444J53Ll4OG3K>FA"@:m3(^(B"j3(&

29
\2@1<:;-%&T(&U(%&781&@1;@"-;<&:"=1:1->@&"C&;-;$9":"D@&*9B>,B&B8;99&;>&d;D2DV&_;V1%&
d"9>8#1@>&7199,>"9,1@%&U;-;$;(&[-K&W,"<".2&"C&>81&*9B>,B&B8;99%&I9"B11$,-.@&"C&>81&
[->19-;>,"-;<&Z2:A"@,D:&"-&*9B>,B&U8;99&]-,=19@,>2&"C&0;-,>"F;&I91@@%&',--,A1.%&
0;-,>"F;%&5LGP(&A&6O6N6OG(&
\2@1<:;-%&T(&U(%&5LLP(&),$1<,>2&"C&;-;$9":"D@&*9B>,B&U8;9&HZ;<=1<,-D@&;<A,-D@J&>"&d;D2DV&
_;V1%&d('(7(%&U;-;$;(&U;-;$,;-&E"D9-;<&"C&),@819,1@&;-$&*fD;>,B&ZB,1-B1@&65K5L3ON5L^P(&
X;::;9%&E(%&345P(&d;>D9;<&91@,<,1-B1&,-&*9B>,B&B8;99&Z;<=1<,-D@&;<A,-D@K&<,C1&8,@>"92%&@A;>,;<&
;-$&$,1>;92&;<>19;>,"-@&;<"-.&.9;$,1->@&"C&,->19@A1B,C,B&,->19;B>,"-@(&E&),@8&W,"<&G6H5JKG5N
55G&$",K54(5555QRCF(53^35(&
X,<F"9-%&S(%&7(&I(&nD,--%&!(&T(&ZB8,-$<19&/&!(&T(&S".19@%&344^(&W,"B":A<1?,>2&;-$&C,@819,1@&
@D@>;,-;F,<,>2(&I9"B11$,-.@&"C&>81&d;>,"-;<&*B;$1:2&"C&ZB,1-B1@&"C&>81&]-,>1$&Z>;>1@&"C&
*:19,B;&544H55JKM6MPNG&$",K54(54O^QA-;@(54^O3OP544(&
X"$.@"-%&Z(&/&7(&I(&nD,--%&3443(&781&>,:,-.&"C&;$D<>&@"BV121&@;<:"-&:,.9;>,"-&,->"&C91@8&
#;>19K&;$;A>;>,"-@&F2&A"AD<;>,"-@&>"&A91=;,<,-.&>819:;<&91.,:1@(&U;-;$,;-&E"D9-;<&"C&
`""<".2&G4H^JK6P3N666&$",K54(55^LQo43N4^4(&
E1--1@@%&!(%&5L33(&781&_,C1&"C&>81&U"AA19&T@V,:"@%&="<&b"<D:1&c[[K&781&U"AA19&T@V,:"@(&)*&
*BV<;-$(&
E1--1@@%&!(&/&Z(&T(&E1--1@@%&5LL5(&*9B>,B&a$2@@12K&781&!,;92&"C&!,;:"-$&E1--1@@%&T>8-"<".,@>&
#,>8&>81&U;-;$,;-&*9B>,B&T?A1$,>,"-&,-&d"9>819-&*<;@V;&;-$&U;-;$;%&5L5^N5L5M(&U;-;$,;-&
0D@&"C&U,=,<,o;>,"-(&

30
E1-@1-%&*(&E(%&a(&X(&!,@19D$%&W(&),-@>;$%&I(&),@V1%&*(&X(&S,V;9$@1-&/&W(&E"-@@"-%&3456(&W1>#11-N
#;>19@81$&:"=1:1->@&"C&>#"&;-;$9":"D@&@;<:"-,$@&,-&>81&*9B>,B(&U;-;$,;-&E"D9-;<&"C&
),@819,1@&;-$&*fD;>,B&ZB,1-B1@&O3HLLLJK5NL(&
E1-@1-%&E(&/&*(&S,V;9$@1-%&3453(&*9B8,=;<&>;.@&91=1;<&>8;>&*9B>,B&B8;99&Z;<=1<,-D@&;<A,-D@&;-$&
F9"#-&>9"D>&Z;<:"&>9D>>;&B;-&D@1&1@>D;9,-1&;-$&:;9,-1&#;>19@&$D9,-.&#,->19(&E"D9-;<&"C&
C,@8&F,"<".2&G5H3JKO^6NOPL(&
E"8-@"-%&_(%&5LG4(&781&;9B>,B&B8;99%&Z;<=1<,-D@&;<A,-D@(&U8;99@K&Z;<:"-,$&C,@81@&"C&>81&.1-D@&
Z;<=1<,-D@&!9&'&ED-V&IDF<,@819@%&781&X;.D1%&d1>819<;-$@K56NLG(&
E"8-@"-%&_(%&5LGL(&781&;-;$9":"D@&;9B>,B&B8;99%&Z;<=1<,-D@&;<A,-D@%&"C&d;D2DV&_;V1%&d'7%&
U;-;$;(&I82@,"<".2&;-$&TB"<".2&E;A;-&ZA1B,;<&5K345N33O(&
E"-@@"-%&W(%&3445(&I"<2:"9A8,@:&;-$&@A1B,;>,"-&,-&*9B>,B&B8;99(&E"D9-;<&"C&),@8&W,"<".2&
6GH^JKM46NM^G&$",K54(544MQRCF,(3444(5656(&
+11C19%&0(&_(&/&U(&U(&U;D$,<<%&345^(&X":,-.&;-$&@>9;2,-.&F2&;-;$9":"D@&@;<:"-,$@K&;&91=,1#&
"C&:1B8;-,@:@&;-$&9;>1@(&S1=,1#@&,-&),@8&W,"<".2&;-$&),@819,1@&3PH5JK^^^N^MG&
$",K54(544OQ@555M4N45^NL^^PNM(&
+<1:1>@1-%&*(%&3454(&781&U8;99&I9"F<1:&S1=,@,>1$K&T?B1A>,"-;<&I81-">2A,B&I<;@>,B,>2&
I9":">1@&TB"<".,B;<&ZA1B,;>,"-&,-&I"@>.<;B,;<&_;V1@(&)91@8#;>19&S1=,1#@&^H5JKPLNOP&
$",K54(5M4GQC9RN^(5(^(&

31
0B\"#;-%&!(&+(%&5LL3(&!;>;&"-&*9B>,B&B8;99%&@;<=1<,-D@&;<A,-D@&H_(J%&C9":&>81&01<,;$,-1&
S,=19%&d"9>8#1@>&7199,>"9,1@(&U;-;$,;-&!;>;&S1A"9>&"C&),@819,1@&;-$&*fD;>,B&ZB,1-B1@&GMOK,N
,=&/&5NL(&
0""91%&E(NZ(%&_(&d(&X;99,@%&S(&)(&7;<<:;-%&T(&W(&7;2<"9&/&I(&0"9p-%&345^(&781&,->19A<;2&F1>#11-&
$,@A19@;<&;-$&.1-1&C<"#&,-&;-;$9":"D@&*9B>,B&B8;9&HZ;<=1<,-D@&;<A,-D@JK&,:A<,B;>,"-@&C"9&
A">1->,;<&C"9&<"B;<&;$;A>;>,"-(&U;-;$,;-&E"D9-;<&"C&),@819,1@&;-$&*fD;>,B&ZB,1-B1@&
O4HLJK5^3ON5^^G&$",K54(55^LQBRC;@N345^N45^G(&
d;9D:%&Z(&S(%&!(&X;>B8%&*(&E(&7;<F">%&I(&0"9;-&/&0(&Z(&I"#1<<%&344G(&[>19"A;9,>2&,-&B":A<1?&
:;>,-.&@2@>1:@&"C&@>11<81;$&a-B"982-B8D@&:2V,@@&H';<F;D:J(&E"D9-;<&"C&),@8&W,"<".2&
O3H5JKP6NM4&$",K54(5555QR(54L6NGMPL(344O(45MPL(?(&
I">>19%&T(%&E(&0;B_1;-%&S(&'2;>>&/&S(&U;:AF1<<%&344^(&0;-;.,-.&>81&1?A<",>;>,"-&"C&:,.9;>"92&
@;<:"-,$@(&),@819,1@&S1@1;9B8&M3H3JK53ON5P3(&
nD,--%&d(&_(%&U(&S(&0B\"#;-%&\(&*(&U""A19%&W(&)(&+""A&/&'(&Z(&!;=,$@"-%&3455(&[$1->,C,B;>,"-&
"C&.1-1@&;@@"B,;>1$&#,>8&81;>&>"<19;-B1&,-&*9B>,B&B8;99&1?A"@1$&>"&;BD>1&>819:;<&@>91@@(&
I82@,"<".,B;<&.1-":,B@&P^H55JKMG6NMLM&$",K54(5563QA82@,"<.1-":,B@(4444G(3455(&
nD,--%&7(&I(%&5LL^(&*&91=,1#&"C&8":,-.&;-$&@>9;2,-.&"C&#,<$&;-$&8;>B8192NA9"$DB1$&@;<:"-(&
),@819,1@&S1@1;9B8&5GH5JK3LNPP(&
S1,@>%&E(&!(%&)(&E(&'9"-;%&7(&!(&I9"#@1%&0(&I"#19%&E(&W(&!1:A@"-%&S(&E(&W1;:,@8%&E(&S(&+,-.%&7(&E(&
U;9:,B8;1<&/&U(&!(&Z;#;>oV2%&344M(&\1-19;<&TCC1B>@&"C&U<,:;>1&U8;-.1&"-&*9B>,B&),@81@&;-$&

32
),@8&I"AD<;>,"-@(&*0W[aK&*&E"D9-;<&"C&>81&XD:;-&T-=,9"-:1->&^6HOJK^O4N^G4&
$",K54(56OLQ44PPNOPPOH344MJ^6h^O4K.1"BB"i3(4(B"j3(&
S"D?%&0(&E(%&S(&)(&7;<<:;-&/&U(&'(&_1#,@%&3455(&Z:;<<N@B;<1&*9B>,B&B8;99&Z;<=1<,-D@&;<A,-D@&
C,@819,1@&,-&U;-;$;e@&dD-;=D>K&:;-;.1:1->&B8;<<1-.1@&;-$&"A>,"-@(&E&),@8&W,"<&OLHMJK5M36N
PO&$",K54(5555QR(54L6NGMPL(3455(4^4L3(?(&
ZB8,-$<19%&!(&T(%&E(&W(&*9:@>9"-.%&+(&7(&W1-><12%&+(&E;-V"#@V,%&I(&E(&_,@,&/&_(&c(&I;2-1%&345^(&
S,$,-.&>81&B9,:@"-&>,$1K&:"F,<1&>1991@>9,;<&B"-@D:19@&>9;BV&A81-"<".,B;<&=;9,;>,"-&,-&
@A;#-,-.&"C&;-&;-;$9":"D@&C,@8(&W,"<".2&<1>>19@&LH^JK345^44PG(&
ZB8,-$<19%&!(&T(%&S(&X,<F"9-%&W(&U8;@B"%&U(&I(&W";>9,.8>%&7(&I(&nD,--%&_(&*(&S".19@&/&0(&Z(&
'1F@>19%&3454(&I"AD<;>,"-&$,=19@,>2&;-$&>81&A"9>C"<,"&1CC1B>&,-&;-&1?A<",>1$&@A1B,1@(&d;>D91&
PM6HO3LGJKM4LN53&$",K54(54^GQ-;>D914L4M4(&
Z>1=1-@"-%&!(&+(&/&Z(&T(&U;:A;-;%&5LL3(&a>"<,>8&:,B9"@>9DB>D91&1?;:,-;>,"-&;-$&;-;<2@,@(&
U;-;$,;-&ZA1B,;<&IDF<,B;>,"-&"C&),@819,1@&;-$&*fD;>,B&ZB,1-B1@&55OK53M&A(&
Z=1--,-.%&0(&*(&/&d(&\D<<1@>;$%&3443(&*$;A>;>,"-@&>"&@>"B8;@>,B&1-=,9"-:1->;<&=;9,;>,"-@K&
>81&1CC1B>@&"C&@1;@"-;<&>1:A19;>D91@&"-&>81&:,.9;>"92&#,-$"#&"C&Z=;<F;9$&*9B>,B&B8;99(&
T-=,9"-:1->;<&W,"<".2&"C&),@81@&MPH5JK5M6N5OP(&
Z#;-@"-%&X(&+(%&+(&*(&+,$$%&E(&*(&W;F;<DV%&S(&E(&';@><1%&I(&I(&q;-.%&d(&0(&X;<$1-&/&E(&!(&S1,@>%&
3454(&*-;$9":2&,-&*9B>,B&A"AD<;>,"-@&"C&<;V1&>9"D>HZ;<=1<,-D@&-;:;2BD@8JK&">"<,>8&
:,B9"B81:,@>92%&@>;F<1&,@">"A1@%&;-$&B":A;9,@"-@&#,>8&*9B>,B&B8;9&HZ;<=1<,-D@&;<A,-D@J(&
U;-;$,;-&E"D9-;<&"C&),@819,1@&;-$&*fD;>,B&ZB,1-B1@&MOH6JKGP3NG6^&$",K54(55^LQC54N433(&

33
';<>19@%&b(%&5L6^(&781&C,@81@&B"<<1B>1$&F2&>81&U;-;$,;-&*9B>,B&T?A1$,>,"-%&5L5^N5G%&#,>8&
;$$,>,"-;<&-">1@&"-&>81&,B8>82"C;D-;&"C@>19-&*9B>,B&U;-;$;(&T$:"-$&U<"D>,19%&nD11-e@&
I9,->19(&
'9"-;%&)(&E(%&7(&!(&I9"#@1%&E(&!(&S1,@>%&E(&T(&X"FF,1%&_(&0(&E(&_g=1@fD1&/&'(&)(&b,-B1->%&344M(&+12&
),-$,-.@%&ZB,1-B1&\;A@&;-$&I"<,B2&S1B"::1-$;>,"-@(&*0W[aK&*&E"D9-;<&"C&>81&XD:;-&
T-=,9"-:1->&^6HOJKP55NP56&$",K54(56OLQ44PPNOPPOH344MJ^6hP55KVC@.;Ai3(4(B"j3(&

34
Tables:
Table 2.1. Length, weight, and condition (K) of Nulahugyuk Creek arctic char at various life-history stages
&& _1-.>8&H::J& '1,.8>&H.J& U"-$,>,"-&
!"#!$ && && &&
]Z&*$D<>& M6LrP5*& ^5LGrO3^*& 5(4Lr4(55*&
!Z&*$D<>& M^4rO6W& [!& [!&
Z:"<>& 5GMr36U& 65r33W& 4(OOr4(4MW&
!"#%$ && && &&
]Z&0"9>;<,>2& OGLr65!& 6OMOrLL6U& 5(5^r4(53*&
]Z&*$D<>& O3Lr6PT& P3MGrLLG!& 5(4Lr4(4L*&
!Z&*$D<>& O5Or6P)& ^5^LrO63*& 4(GPr4(4LU&
Z:"<>& 5L^r35U& M5r35W& 4(G5r4(4O!&
!;>;&A91@1->1$&;@&:1;-&r&@>;-$;9$&$1=,;>,"-&
!,@@,:,<;9&<1>>19@&,-$,B;>1&@,.-,C,B;->&$,CC191-B1@&Hst4(46J&
[!%&[-@DCC,B,1->&$;>;j&]Z%&]A@>91;:j&!Z%&!"#-@>91;:&

35
Table 2.2 Otolith based age (years) estimated of Nulahugyuk Creek arctic char at various life-history stages
&& d& 01;-&r&Z!& 0,-(& 0;?(&
*$D<>@& ^O& 54(6r4(L& G& 53&
*)0& 5G& P(4r4(G& ^& 6&
Z:"<>@& 5M& P(^r4(O& ^& 6&
*)0K&;.1&;>&C,9@>&:,.9;>,"-& && &&
Z!K&@>;-$;9$&$1=,;>,"-& & &
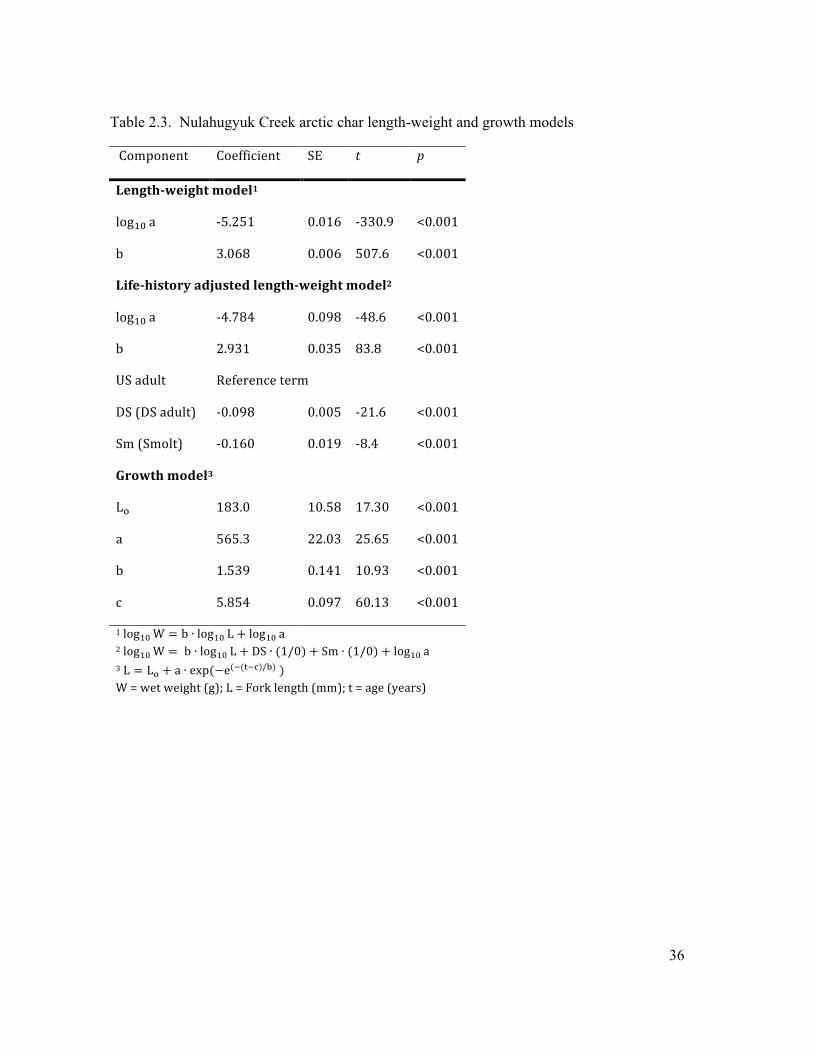
36
Table 2.3. Nulahugyuk Creek arctic char length-weight and growth models
&U":A"-1->& U"1CC,B,1->& ZT& !" #"
&'()*+,-'.)+*$/01'2#$
!"#!" !& N6(365& 4(45M& N^^4(L& l4(445&
!& ^(4MG& 4(44M& 64O(M& l4(445&
&.3',+.4*056$71894*'1$2'()*+,-'.)+*$/01'2!$
!"#!" !& NP(OGP& 4(4LG& NPG(M& l4(445&
!& 3(L^5& 4(4^6& G^(G& l4(445&
]Z&;$D<>& S1C191-B1&>19:&
!Z&H!Z&;$D<>J& N4(4LG& 4(446& N35(M& l4(445&
Z:&HZ:"<>J& N4(5M4& 4(45L& NG(P& l4(445&
:50-*+$/01'2;$
!!& 5G^(4& 54(6G& 5O(^4& l4(445&
;& 6M6(^& 33(4^& 36(M6& l4(445&
F& 5(6^L& 4(5P5& 54(L^& l4(445&
B& 6(G6P& 4(4LO& M4(5^& l4(445&
5!!"#!"! ! ! ! !"#!" ! ! !"#!" !&3!!"#!"! ! !! ! !"#!" ! ! !" ! !!!!! ! !" ! !!!!! ! !"#!" !&^&! ! !! ! ! ! !"#!!!!!!!!!!!!!!!&'&t>,.8>&H.Jj&_&t&)"9V&<1-.>8&H::Jj&>&t&;.1&H21;9@J&

37
Table 2.4. Timing of the upstream migration of various anadromous arctic char populations in North America
Name Period of upstream migration Reference:
Nauyuk River early August - mid-September (Johnson 1989)
Firth River late July - late September (Glova and McCart 1974)
Fraser River mid-July - late September (Dempson and Green 1985)
Ikaluit River mid - late August (Read 2003)
Meladine River August – mid-September (McGowan 1992)
Rat River August (Gillman and Sparling 1985)
Diana River mid-August - early September (McGowan 1987)
Ekalluk River late August - early September (McGowan 1990)
Jayco River late August - mid-September (McGowan 1990)
Halovik River mid-August - early September (McGowan 1990)
Lauchlan River late August - mid-September (McGowan 1990)
Nulahugyuk Creek late June - late July Present study

38
Figures
Fig 2.1. Study area in northwest mainland Nunavut, Canada. Nulahugyuk Creek was the primary
study site; Arctic char captured here feed in the Coronation gulf and Dolphin and Union Sound
region and are thought to overwinter in navigable freshwater systems in the area.
Fig 2.2 Physical migratory environment of Nulahugyuk Creek, NU. Photos taken at a
standardized location demonstrating the navigability of a representative riffle section mid- (A;
early July) and post migration (B, early August). Creek discharge (C) and water temperature (D)
values shown in white, grey, and black are from 2012, 2013, and 2014 respectively.
Fig 2.3 Daily number of arctic char migrating passed the fish trap by life history stage. The
counts for each life history stage are shown separately for the 2012 (grey) and 2014 (black) field
seasons.
Fig 2.4 Length frequency distributions of migrating arctic char captured during the 2012 and
2014 field seasons. The data is shown as percentages of either smolts (dark grey) or adults (light
grey).
Fig 2.5 The relationship between length and weight over various life-history stages. Upstream
migrating adults, downstream adults and smolts are shown in dark grey, white, and light grey
respectively. The back transformed two-factor length-weight model is shown by the solid black
line with the 95% prediction interval shown by the dashed grey lines. For model parameters see
Table 3.
Fig 2.6 Back-calculated growth in Nulahugyuk Creek arctic char. Boxes represent the
interquartile range around the median, lower and upper whiskers represent the 10th and 90th

39
percentile respectively and dots represent outliers. The Gompertz growth model is depicted by
the solid grey curve with the 95% prediction interval shown by the dashed lines. For model
parameters see Table 2.3.

40

41

42

43
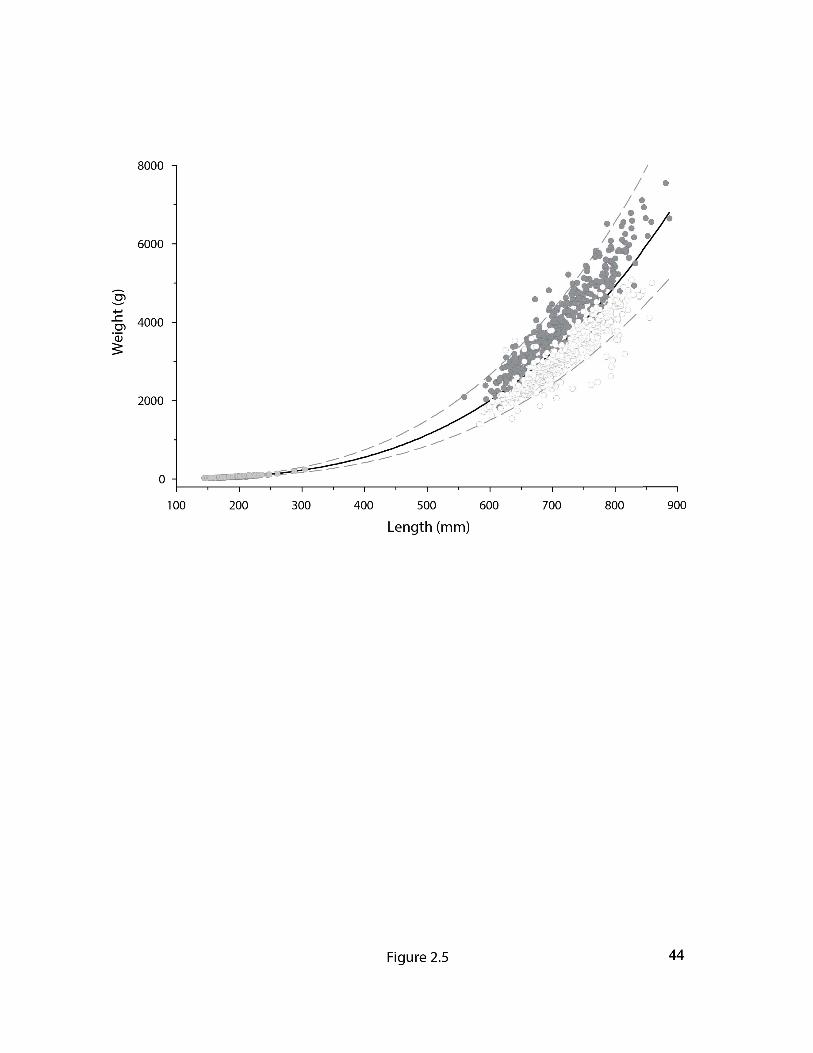
44
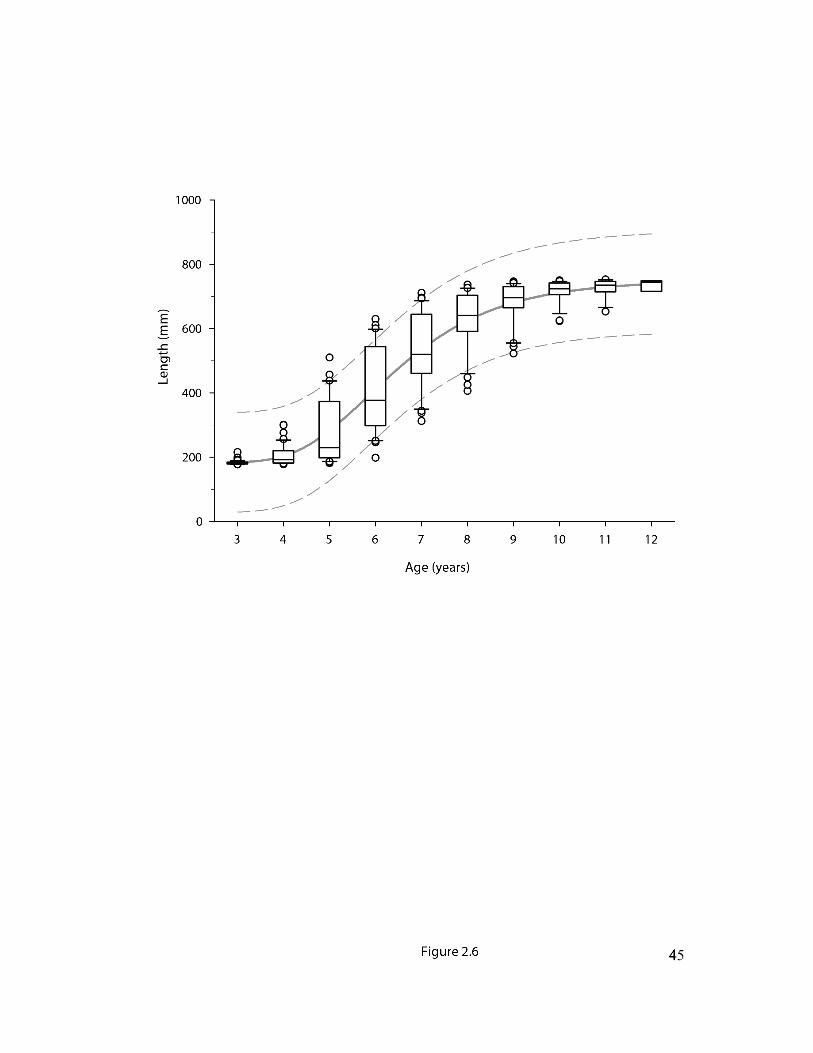
45

46
Chapter 3: Repeat swimming performance and physiology of arctic
char (Salvelinus alpinus) and rainbow trout (Oncorhynchus mykiss)
exposed to realistic diurnal temperature fluctuations.
Abstract
Arctic char are considered a typical cold-water stenotherm, however, we have identified a
population that annually migrates through a warm (>21°C), physically challenging creek that
undergoes large daily temperature fluctuations (>10°C). Contrary to expectation, in-field repeat
exercise tests on juvenile char showed no substantial effect of temperature on initial swimming
performance, however there appeared to be threshold for repeat swimming performance near
20°C above which recovery may be impaired. To further investigate this phenomenon we
exposed juvenile arctic char and rainbow trout, a temperate reference salmonid, to simulated
diurnal temperature scenarios in lab. As in the field, an initial exercise test did not reveal
impaired performance at warmer temperatures, however, in a second test conducted after a brief
recovery, swimming performance was significantly reduced in char but not in rainbow trout.
This impaired repeat swimming performance after warming was associated with a reduced
ability to increase routine aerobic metabolism at warm temperature. Diurnal warming also had
more pronounced effects on blood composition and plasma glucose and lactate in arctic char than
rainbow trout. These results suggest that the physiological performance of arctic char is more
subject to impairment by thermal variation than temperate salmonids. This relative thermal
limitation may constrain their migratory success under current conditions in some arctic streams
and may put them at a competitive disadvantage for migratory habitat as more temperate species
expand their ranges northward.

47
Introduction
The exercise physiology of salmonid species and populations is often adapted to function
optimally under a relatively narrow range of frequently encountered temperatures (Eliason et al.
2011; Farrell 2009b). As global climate change progresses, deviations from historical freshwater
thermal regimes result in an increase in exposure of migrating salmon to temperatures above
these optimal ranges (Eliason et al. 2011; Farrell et al. 2008a), which has already been associated
with instances of impaired migratory performance (Farrell et al. 2008a; Hinch et al. 2012;
Martins et al. 2012; Richter and Kolmes 2005). Extensive research on the effects of temperature
on the migratory physiology of Pacific salmon indicates that cardiorespiratory limitations at
warm temperatures are an important contributor to this impaired performance and to determining
thermal tolerance in general (Eliason et al. 2011; Farrell et al. 2008a; Farrell 2009b; Farrell et al.
2009).
Thermal physiology in salmonids has typically been studied at constant temperatures to
which salmon have been previously acclimated (eg. Eliason et al. 2011; Farrell 2009b), at rates
of temperature change that may not represent common natural variation (eg. 2°C/hr, Clark et al.
2012; Clark et al. 2008; Penney et al. 2014; Steinhausen et al. 2008), or at point-sampled
temperatures in wild fish (Farrell et al. 2003; Rodnick et al. 2004). Environmental exposure more
commonly occurs on diurnal to seasonal scales, or acutely in an irregular manner as fish
transition between different thermal environments such as when moving between the ocean,
rivers and lakes and when undertaking vertical migrations (Farrell 2009b; Friedland et al. 2001;
Mathes et al. 2010; Rikardsen et al. 2007; Rodnick et al. 2004). The rate at which temperature
change is experienced can have a pronounced effect on fishes tolerance to that variation (Dowd
et al. 2015; Galbreath et al. 2004; Mora and Maya 2006). This effect may result in important

48
differences between the tolerance and response of salmonids to realistic and typical laboratory
tested thermal regimes(Dowd et al. 2015).
In addition to the study of natural thermal regimes, the thermal physiology of more northerly
distributed salmonids has also received relatively little attention (Penney et al. 2014), despite the
fact that the effects of global climate change are most pronounced at higher latitudes (ACIA
2005; Wrona et al. 2006c). Among northern salmonids, the arctic char (Salvelinus alpinus) is of
particular interest as it has the most northerly distribution of any freshwater fish and they possess
extensive life history diversity that facilities its existence throughout the global arctic in a wide
range of environments (Klemetsen 2010; Klemetsen et al. 2003; Roux et al. 2011). While this
expansive, northern distribution and life history diversity may help the stability the species in the
long term (Schindler et al. 2010) it also means that some populations likely utilize life-histories
and habitats that make them particularly vulnerable to changing thermal regimes. For example,
we identified one anadromous population in which individuals spend nearly all of their lives in
cold (<11°C), thermally stable lacustrine and marine environments, but migrate between these
environments via Nulahugyuk Creek (Fig.1), a shallow, physically challenging creek that
undergoes large diurnal temperature fluctuations (>10°C per day) over broad a range of
temperatures from ~7 to 21°C (Fig. 2).
In the present study we utilized field and laboratory methods to determine the effects of
diurnal temperature fluctuations on arctic char migratory physiology. In the field, we assessed
the repeat swimming performance of juvenile char, over the naturally occurring range of
temperatures that char were observed migrating through. Based on field results, in the laboratory
we exposed arctic char, to a simulated diurnal temperature fluctuation representative of the upper
end of the range naturally encountered by migrating char in Nulahugyuk Creek and assessed

49
their behavior, repeat swimming performance, aerobic metabolism and several sub-organismal
stress related endpoints. In a companion study (Chapter 4), we assessed biochemical and
transcriptional endpoints and related them to the findings of the present study.
All laboratory endpoints were also assessed in rainbow trout (Oncorhynchus mykiss), which
serves as a temperate reference species. Rainbow trout were selected because their thermal
physiology has been well studied, they could be reared under nearly identical conditions to char,
and like arctic char they are iteroparous and facultatively anadromous. In general, we expected
that both species would exhibit physiological impairments at peak diel temperatures, but that
arctic char would be much more sensitive to warming over the tested range and would therefore
recover to a lesser extent during diel cooling.
Materials and Methods
Field sampling
Field sampling took place in the summers (June – August) of 2012–2014 near the mouth of
Nulahugluk Creek, Nunavut (NU; 68.75°N 114.8°W; Fig 1). Fish were captured migrating
between their freshwater and marine environments (Hingittok Lake and Bernard Harbour, NU,
Fig 1) using a two-way fyke net 300m upstream from the mouth of the creek. Water temperature
was recorded at the trap every 15 min using an automated temperature logger (HOBO Water
Temperature Pro, Onset Computer Corporation, Bourne, Massachusetts). Detailed temperature
and discharge data can be found in Chapter 2.
In 2012, creek side swimming performance tests were conducted on a subset of juvenile arctic
char over the range of naturally occurring temperatures to assess the effect of natural temperature

50
variation on cardiorespiratory performance. These test were conducted as described below for
the laboratory-based research.
Laboratory animals
Arctic char were obtained at six months post hatch from Elkview Farms (Red Deer, Alberta,
Canada). The char were hatched in December 2012 at Elkview Farms but ova and sperm were
obtained from the broodstock at the B&B Freshwater Fish Farm (Gunton, Manitoba). This
broodstock originated from the Nauyuk Lake, NU population, which like our study population is
in the central arctic and it also possesses some important life history similarities (Chapter 2).
Rainbow trout were hatched at the University of Alberta (UofA; Edmonton, AB) aquatics
facility, in December 2012. In May 2013 trout and char were transferred to the same housing
rack at the UofA and separated into several 30L flow through tanks and held at similar density in
11°C dechloirinated municipal tap water. Fish were fed trout pellets twice daily (Nu-Way Trout
Grower-Finisher, Hi-Pro Feeds, Okotoks, Alberta) and held under a simulated local light-dark
cycle. Following all experiments fish were sacrificed, weighed to the nearest gram, and measured
to the nearest mm. The ventricle was weighed to the nearest 0.001g and expressed as VM or
relative ventricular mass (RVM) i.e. a percentage of body mass. Condition factor was calculated
as K = Mass/Length3* 10-5.
Diurnal temperature treatments.
Fish were exposed to one of three temperature treatments (Fig. 2) in a “bow-tie” swim-tunnel
previously described by Tierney et. al (Tierney et al. 2011). Briefly, this swim tunnel has two
large (25 cm3), low flow chambers connected by a long, narrow (100 x 10 x10 cm), high flow
section which allows fish to select between a range of water speeds and move between upstream

51
and downstream flow refugia. Flow in the center section was set at 35 cm s-1, a speed commonly
encountered by migrating char in Nulahugyuk Creek, NU, and ranged in the end sections from
35cm s-1 down to speeds low enough that fish could rest on the bottom without swimming.
Control and diurnal warming treatments lasted between 16 and 24hrs with variation in duration
occurring only in the 11°C component of the treatment (Fig. 2). All fluctuation treatments lasted
for 24hrs as the entire duration was necessary to achieve the required temperatures at the selected
rate of variation. Temperatures were achieved using Autoresp software (Loligo Systems, Tjele,
Denmark) to control two solenoid valves that regulated the flow of cold or warm water into the
swim tunnel.
Repeat swimming performance and gait transition
Immediately following the temperature treatment fish were transferred to a 90L swim-tunnel
respirometer with a 70x20x20 cm test section (Loligo Systems, Tjele, Denmark) that was held at
the final temperature of the respective treatment (Control and Diurnal fluctuation: 11°C ; Diurnal
warming: 21°C). Fish were allowed to acclimate at a low water speed (0.5 body lengths s-1; bl s-
1) for 30 min after which the speed was increased by 0.5bl s-1 every ten minutes until the fish
fatigued and was therefore not able to remove itself from the rear gate within five seconds. Fish
were then allowed to recover for 30 min at 0.5 bl s-1. To assess the ability of fish to recover
following the first exercise test, fish were given a second test except speed was increased every
minute rather than every ten. The standard Brett equation (Brett 1964) was used to calculate the
critical swimming speed for each test. A small electrical charge (6v) was applied to the rear gate
to motivate fish to swim throughout the duration of both tests.

52
The gait transition speed (UGT), an estimate of the speed at which the primary mode of
swimming changes from steady to burst and coast, was determined from video using automated
visual tracking software (Ethovision XT8, Noldus Information Technology Inc., Leesburg,
Virginia) to generate ground speed and positional data. UGT was defined as the speed at which
the fish carried out three or more bursts per min and continued for the duration of the test
(MacNutt et al. 2006). Rather than manually scoring bursts as was previously done, a burst was
defined as a bout of upstream acceleration in 99th percentile of the data for the trial. This method
produced UGT estimates that were well within the published range (MacNutt et al. 2004;
MacNutt et al. 2006).
Aerobic metabolism
Oxygen concentration was recorded every second during the swimming performance test from
the start of the 30 min acclimation to the end of the 30 min recovery using an anode-cathode
oxygen probe placed in the manufacturer specified port in the conventional swim tunnel. The
probe was attached to the same DAQ unit and software as the swim tunnel. All swimming
performance tests started at 100% O2 saturation and the swim tunnel was sealed for the duration
of the tests unless oxygen concentration dropped below 7.5mg/L, in which case the tunnel was
briefly flushed with fresh water. Routine metabolic O2 consumption rate (MO2routine ) was taken
as the minimum [O2] slope over five minutes that had an R2 > 0.90 within the acclimation period.
The metabolic rate for each step within the swim test was taken as the maximum [O2] slope over
one minute that had an R2 > 0.90 in that step, which always occurred in the later half of the step
as fish tend to take a few minutes to adjust to swimming at a new speed. Post-exercise oxygen
consumption (POC) rates were calculated as the slope of [O2] over each five minute increment of
the 30 min recovery period following fatigue in the first swim test. In calibration trials, no

53
background oxygen consumption was detected. Changes in water velocity interfered with O2
detection, but profiles stabilized within two minutes of the change, and because of the way MO2
values were calculated the values from this period of each step were not utilized.
Blood analysis
Blood was drawn by caudal puncture using at 21-gauge needle and 5ml syringe that had
been pre-rinsed with lithium-heparin (100 IU/ml; Sigma-Aldrich, Oakvillem Ontario). Three to
five pre-hepranized microcapillary tubes (Thermo Fisher Scientific, Waltham,
Massachusetts) were filled at the start of sampling and the rest of the blood was transferred into
microcentrifuge tubes. Microcentrifuge tubes were spun for five minutes at 5000g after which
plasma was transferred into new microcentrifuge tubes and stored until analysis at -20°C.
Capillary tubes were spun for three minutes at 10,000g and hematocrit (Hct) was then measured
as the height of the packed red blood cell column over that of the entire sample using digital
calipers. The final Hct value was taken as the average Hct of at least three capillary tubes. After
spinning, two drops of plasma were applied to test strips for lactate and glucose meters (Lactate:
Lactate Plus, Nova Biomedical, Waltham, Massachusetts; Glucose: Accu-chek Nano, Roche,
Basel, Switzerland). Glucose readings were verified to be accurate by comparison with plasma
glucose values obtained from the nuclear magnetic resonance (NMR) based metabolomics
analysis that was conducted on a subset of plasma samples (Chapter 4), however, lactate values
obtained from the meter were adjusted to better represent NMR concentrations using the linear
relationship with plasma lactate values obtained in the NMR analysis. In addition to the subset of
individuals sampled in our companion study, plasma lactate and glucose concentrations were
measured in all individuals in the present study as elevations in their concentrations are
commonly used as secondary indicators of stress and have been associated with early mortality

54
in migrating salmon (Cooke et al. 2006). Plasma osmolality was determined though freeze-point
analysis using a Model 3300 osmometer (Advanced Instruments, Norwood Massachusetts).
Plasma cortisol was measured by using a commercial enzyme-linked immunosorbent assay kit
(Kit # 402710, Neogen Corp., Lexington, Kentucky, USA) and following the manufactures
recommended protocol with two changes, ethyl acetate was used as an extraction solvent, and the
solvent evaporation took place in a fume hood without a nitrogen stream or vacuum as in
Canavello et al (Canavello et al. 2011).
Data analysis
All statistical analyses were conducted using R Studio (Team 2014). Figures were produced
using ArcGIS (ESRI, Redlands, California), Sigmaplot 11.1 (Systat Software, San Jose,
California) and Adobe Illustrator CS4 (Adobe systems, San Jose, California).
For laboratory animals, differences in all morphometric variables, swimming performance,
recovery, UGT, Hct, plasma osmolality, cortisol, lct, and glu, were assessed using two-way
analyses of variance with species, treatment and their interaction as predictor variables followed
by Tukey’s Honest significant difference test to allow for pairwise comparisons. For swimming
performance, length was included as a covariate. For MO2routine and MO2max VM, Cort, Hct, lct,
and glu, body weight was included as a covariate. Covariates were removed from the analysis if
they or their interactions were not statistically significant (!=0.05). Student’s t-tests were used to
analyze difference in FL, mass, and K between wild and captive char.
MO2 data were analyzed using linear-mixed effects models (LMMs) generated using the lme4
package (Bates et al. 2013) and back fit using the lmertest package (Kuznetsova et al. 2014).
Two separate MO2 models were created for each species, one for the duration of the UInitial test

55
and another for the recovery period. Treatment and the log of body mass were included as fixed
predictor variables in both MO2 models. Time into the exercise test was included in the UInitial
MO2 model, while time post fatigue was included in the recovery MO2 model. The individual
fish ID was included as a random predictor variable to account for the fact that multiple MO2
values recorded from the same individual are not independent. The marginal and conditional R2
values were reported to describe the proportion of variation explained in each model by just the
fixed effects (marginal) or by both random and fixed effects (conditional).
Field swimming performance and recovery data were analyzed using linear and quadratic
regressions against temperature and the model with the lowest residual standard error was
selected. Upon visual inspection of URepeat and recovery ratio vs. temperature relationships the
two data points available for 21°C appeared to be outliers on an otherwise positive correlation,
so a second analysis was conducted with the values excluded (Table 2, Fig. 3). In all regression
models all possible interactions were assessed. If an interaction effect was not significant it was
removed from the model as long as no higher order interactions were significant. To ensure
assumptions of the various analyses were met normality of residuals was assessed using Shapiro-
Wilks tests and visual analysis of q-q plots, and equality of variances using Levene’s test when
appropriate. If assumptions were violated Tukey’s ladder of transformation was employed until a
suitable transformation was identified. In all analyses interactions were treated as being
multiplicative.
Results
Study animals.

56
In the laboratory study, rainbow trout were generally smaller than the arctic char (Fork length: -
32%, F2,39 = 21.9 ,p = 3.3*10-5; Mass: -13% , F2,39 =17.0 p = 1.8*10-4 ; Table 1) but had a similar
condition factor (+4% , F2,39 =2.0 p = 0.17; Table 1). VM co-varied with body mass but was still
significantly lower in trout after accounting for mass (-25%, Species: F1,38 = 88.7, p = 2.0*10-11,
Mass: F1,38 = 64.4, p = 7.3*10-10; Table 1). For comparison with previous studies, RVM was also
analyzed and was not-surprisingly also lower in trout (-27%, F1,38 = 20.5, p = 6.0*10-5; Table 1).
There were no statistically significant morphometric differences between treatments (F2,39 < 2.1,
p > 0.13). The wild juvenile arctic char used in swimming performance tests had a nearly
identical average FL to captive char (-0.6%, t52 = 0.2 , p = 0.81; Table 1) but had a significantly
lower mass and condition factor. (Mass: -22%, t47 = 3.7 , p = 5.0*10-4; K: -27%, t47= 8.8 , p =
1.4*10-11; Table 1).
Field swimming performance and recovery
In wild arctic char, UInitial had a weak quadratic relationship with temperature over the tested
range (Table 2; Fig. 3). URepeat and consequently, recovery ratio, both exhibited linear positive
relationships with temperature after the removal of data points sampled at 21°C (Table 3.2; Fig.
3.3). Only two individuals were tested at 21°C, however, their relatively low URepeat and recovery
ratio may indicate a threshold for recovery near 21°C.
Laboratory swimming performance and recovery
Swimming performance and recovery ratio varied between diurnal temperature treatments in
different manners for the two species. Specifically, UInitial increased in trout following warming
but did not change in char (Table 3.3; Fig. 3.4). URepeat also increased in trout following warming
but decreased in char. Consequently recovery ratio did not change with warming in trout but

57
substantially deceased in char . Interestingly, for both species all but one metric of swimming
performance (URepeat, BL s-1) and recovery following the complete diurnal fluctuation were
numerically intermediate between the control and diurnal warming groups (Table 3.2, Fig,3.4).
Even after accounting for treatment effects, comparison of least squares means showed that
arctic char had lower initial swimming performance than rainbow trout (UInitial: -19% p<0.001).
UGT was not different between species under control conditions or following diurnal fluctuation
but numerically increased in trout and decreased in char following warming which resulted in a
significant difference between species following warming (Table 3.2).
Aerobic metabolism during fatiguing exercise and recovery
Treatment effects on aerobic metabolism differed between trout and char (Table 3.3 and 3.4, Fig.
3.5). In trout, MO2routine and MO2max both increased substantially following warming (Table 3.3)
and were intermediate between control and warming following the complete fluctuation. In char,
only minor numerical increases were found following warming and the fluctuation group was
indistinguishable from control (Table 3.3). The LMMs revealed that in both species over all
treatments metabolism increased with water velocity (time into test) (Table 3.4). Comparisons
between LMMs showed that for arctic char during the Uinitial test, treatment did not significantly
explain a portion of the variance in the data, however, recovery toward routine metabolism was
lower in the warming group (Fig. 3.5) and so the warming treatment was a significant predictor
variable in the recovery MO2 model (Table 3.4). In trout, treatment variables explained unique
portions of the variation in both exercise and recovery models and agreed with the trend that for
the exercise test MO2 was lowest in the control, intermediate in the fluctuation treatment and
highest in the warming treatment. During recovery, the model suggests that the warming and
control group recovered at a similar rate, while the fluctuation treatment group recovered faster.

58
Blood analysis
In trout, Hct did not differ between treatments but was lower following warming in char and
numerically intermediate following the complete fluctuation (Table 3.3). There were no apparent
changes in plasma osmolality between species or treatments (Table 3.3). Overall, plasma cortisol
was lower in arctic char than in rainbow trout and numerically decreased following warming in
char, however, no pairwise comparisons were significant (Table 3.3). Overall, plasma glucose
and lactate concentrations increased following diurnal warming, although numerically to a
greater extent in arctic char (Table 3.3). Both metabolites had intermediate concentrations
following the complete fluctuation except for lactate in rainbow trout which was similar to
control (Table 3.3).
Discussion
Arctic char are commonly considered to be a classical cold-water stenotherm and among the
least thermally tolerant of all salmonids. However, we have recently identified one population
that annually migrates through a shallow creek that experience drastic diurnal temperature
fluctuations with maximal temperatures exceeding 21°C (Chapter 2; Fig. 3.1). Our results
suggest that in terms of aerobic exercise performance, arctic char are likely more tolerant to
thermal variation over environmentally relevant scales than previously thought, but as expected,
they are more sensitive than related temperate species. Specifically, arctic char were able to
maintain their initial swimming performance when brought from 11 to 21°C over 12 hours,
however their ability to recover from this initial bout of fatiguing exercise was impaired while it
was not impaired in rainbow trout. Arctic char also exhibited more pronounced effects of diurnal
warming on blood composition and in plasma glucose and lactate levels, and char had an overall

59
lower capacity for aerobic exercise. Together, my results suggest that more temperate salmonids
such as the rainbow trout likely already possess a competitive advantage for migratory habitat in
the north over some northern salmonids under current summer conditions, which could allow for
displacement as winter conditions at higher latitudes become more permissive of temperate
species.
Initial swimming performance and aerobic metabolism
Swimming performance values were very similar between the field and laboratory for arctic char
(Lab: 3.30 vs. 3.55 BLs-1; Fig. 3 and 4), and were consistent with previous research for both
arctic char and rainbow trout (e.g. Alsop and Wood 1997; Beamish 1980; Pettersson et al. 2010).
Relative to their length, arctic char had a much lower swimming ability than rainbow trout,
regardless of temperature (Table 2; Fig. 4). Trout improved swimming performance following
warming from 11 to 21°C, which agrees with previous results following warming from 12-14 to
24°C over ~5hr in a desert population that experiences large diurnal fluctuations (Gamperl et al.
2002). This improvement was not surprising considering that when acclimated to similar
conditions to those used in the present study, rainbow trout do not loose equilibrium until ~28°C
(Carline and Machung 2001). A recent study on arctic char, however, found that under similar
conditions, they exhibit a loss of equilibrium at ~23°C (Penney et al. 2014), so it is surprising
that arctic char were able to maintain UInitial following warming from 11 to 21°C. Diurnal
warming substantially increased MO2Routine and active MO2 in trout, while only minor increases
were observed in char (Table 2 and 3; Fig 5), which is consistent with differences in the effect of
temperature on MO2 between atlantic salmon (Salmo salar) and arctic char (Penney et al. 2014).
Following the complete diurnal fluctuation, rainbow trout had intermediate UInitial and MO2
suggesting that the physiological changes underlying their improved performance and elevated

60
MO2 following warming were not fully reversed during within the diurnal cooling phase. This is
consistent with a recent study in atlantic salmon that showed that smaller diurnal fluctuations
could increase metabolic rate relative to being held at a constant temperature (Oligny-Hébert et
al. 2015). All changes in routine and active MO2 were proportional so there were no differences
in aerobic scope between treatments. Based on an abundance of research salmonids, aerobic
scope likely follows a bell shape thermal performance curve (Eliason et al. 2011; Farrell 2009b),
so the lack of apparent difference in aerobic scope between treatments suggests that 11 to 21°C
may be on opposite sides of the curve under the present acclimation temperature and rate of
warming. Between species under control conditions MO2Routine was similar, however, MO2Max
was much higher in trout than in char and consequently so was aerobic scope; this difference
persisted across treatments. The comparative low aerobic scope observed in char likely
contributed to their relatively low swimming performance, and agrees with previous research on
temperature induced measurements of aerobic scope that revealed that adult arctic char had a
lower aerobic scope than Atlantic salmon (Penney et al. 2014). Beyond differences in thermal
physiology our findings pertaining to swimming performance and MO2 support the notion put
forth by Beamish (Beamish 1980) that arctic char, and members of the genus Salvelinus in
general, are not as proficient aerobic swimmers as their relatives in the genera of Oncorhynchus
and Salmo.
Impaired repeat swimming performance following diurnal warming
Following diurnal warming in the laboratory experiments arctic char had drastically reduced
repeat swimming performance, indicating an impaired ability to recover from fatiguing exercise
and a relatively greater disruption of contributing biological processes (Jain et al. 1998; MacNutt
et al. 2004). Interestingly in the field, an increase in recoverability was seen over the tested

61
range of temperatures up to 20°C, after which repeat swimming appeared to fall off
precipitously, however, the number of individuals tested above 20°C was too small (N=2) to
perform any meaningful threshold or breakpoint analysis. A reduction in the ability to recover
from exercise in migrating char is concerning as salmonid migrations typically occur in bouts of
strenuous exercise, followed by periods of lower exertion to recover, as opposed to single
prolonged swimming efforts, and periods of prolonged exertion are associated with reduced
migratory success (Hinch and Bratty 2000). This impaired recoverability may pose an even
greater threat in shallow rivers such as Nulahugyuk Creek (Fig.1) where stranding and therefore
aerobic fatigue are very common and thermal refugia are limited (Chapter 2). Arctic char we not
able to increase MO2Routine to the same extent as rainbow trout when brought to 21°C and
following a fatiguing exercise test at this temperate they did not recover to the same extent as
either other treatment group, or as trout.
The decreased ability to increase MO2 with temperature suggests that the performance of some
component of the cardio respiratory system or cellular respiration was thermally limited relative
more temperate species which agrees with previous findings in non-exercised char when
compared to Atlantic salmon (Penney et al. 2014). It is also consistent with the notion in other
salmonids that thermal tolerance in largely determined by the capacity to deliver and utilize
oxygen (Eliason et al. 2011; Farrell et al. 2009). The fact that arctic char also had a longer
recovery time also supports this notion, as impaired aerobic performance at warm temperature
would necessitate a greater utilization of anaerobic metabolism to achieve the same Uinitial and
would therefore incur greater oxygen debt. The prolonged recovery likely meant that the scope
for aerobic performance at the start of the Urepeat was reduced and therefore limited repeat
performance (Lee et al. 2003).

62
Blood physiology in relation to recoverability
The low Hct following warming in arctic char was likely the result of either an increase in fluid
retention, or a failure to increase red cell volume. Increased fluid retention may have occurred as
a compensatory mechanism to maintain or increase mean arterial blood pressure if cardiac output
was limited at high temperatures as is common during exercise at warm temperatures in
salmonids (Eliason et al. 2011; Farrell 2009a). However, it may have also been a result of
increased mobilization of other metabolites (e.g. Lactate glucose, and amino acids; Chapter 4). If
the change in Hct was associated with increased fluid retention, it would explain why the drastic
increases in metabolite concentrations identified in a companion study (Chapter 4) did not result
in increased plasma osmolality, and why plasma cortisol concentration appeared to decreased.
Alternatively, as Hct increases with exercise (Jain and Farrell 2003), diurnal warming may have
impaired the release of new red blood cells normally stimulated by exercise. Regardless of the
cause, the reduction in Hct likely decreased the oxygen carrying capacity of the blood and may
have contributed to the observed differences in recoverability.
Arctic char exhibited much more pronounced increases in plasma glucose and lactate than those
observed in trout. This suggests that arctic char were struggling to meet aerobic cellular energy
demands and were increasingly mobilizing glucose and utilizing anaerobic pathways to offset
aerobic short falls (Jain and Farrell 2003; Tierney and Farrell 2004). Furthermore, these
metabolites are common secondary indicators of stress and have been associated with early
mortality in other salmonids it also likely means that in general the fish were in poorer
physiological condition (Cooke et al. 2006).
Conclusions

63
For arctic char a relatively limited athletic ability paired with a lower ability to recover from
exercise following naturual thermal variation will likey result in reduced migratory success, such
as that found in pacific salmon (Farrell et al. 2008b; Martins et al. 2012) and make them
particularily vulnerable to competition from Pacific and Atlantic salmon whose ranges appear to
be expanding in the north (Dunmall et al. 2012; Jonsson and Jonsson 2009). Pink salmon may be
of particualr concern as they are already being caught in the western arctic (Dunmall et al. 2012),
are highly proficient swimmers, and are among the most tolerant salmonids to thermal variation
(Clark et al. 2011; MacNutt et al. 2006).
It is important to note, that in both species throughout the study, results for fish that had
experienced a complete diurnal fluctuation were intermediate to those that had experienced just
diurnal warm and those under control conditions. These results suggest that within, the natural
time scale of a day fish exposed to thermal variation do not return to the same physiological state
they are in under static conditions. This finding provides support for the emerging concept that
diurnal temperature variation and not just static temperatures, averages, or thermal maxima,
should be considered when designing experiments intended to advance our understanding of how
fish will response to natural scenarios (Dowd et al. 2015; Oligny-Hébert et al. 2015).
Acknowledgements
I would like to thank the members of the community of Kugluktuk, NU, the Kugluktuk Hunters
and Trappers Organization (HTO), the Nunavut Department of Environment, and Golder
associates for assisting in the collection of field data. Funding was provided thorough a National
Science and Engineering Research Council (NSERC) Discovery grant to KBT, in kind support
from Golder Associates, and grants and awards to MJHG from NSERC, the Northern Scientific

64
Training Program, Canadian Circumpolar Institute, Canadian Society of Zoologists, the
Association of Canadian Universities for Northern Studies and the University of Alberta. I would
also like to thank the staff of the University of Alberta aquatics facility.

65
References:
*U[*%&3446(&*9B>,B&B<,:;>1&,:A;B>&;@@1@@:1->N@B,1->,C,B&91A"9>(&U;:F9,$.1&]-,=19@,>2&I91@@(&
*<@"A%&!(&/&U(&'""$%&5LLO(&781&,->19;B>,=1&1CC1B>@&"C&C11$,-.&;-$&1?19B,@1&"-&"?2.1-&
B"-@D:A>,"-%&@#,::,-.&A19C"9:;-B1&;-$&A9">1,-&D@;.1&,-&RD=1-,<1&9;,-F"#&>9"D>&
Ha-B"982-B8D@&:2V,@@J(&781&E"D9-;<&"C&1?A19,:1->;<&F,"<".2&344H5OJK3^^ON3^PM(&
W;>1@%&!(%&0(&0;1B8<19%&W(&W"<V19&/&Z(&';<V19%&345^(&<:1PK&_,-1;9&:,?1$N1CC1B>@&:"$1<@&
D@,-.&T,.1-&;-$&ZP(&S&A;BV;.1&=19@,"-&5HPJ(&
W1;:,@8%&)(&'(%&5LG4(&Z#,::,-.&A19C"9:;-B1&;-$&"?2.1-&B"-@D:A>,"-&"C&>81&B8;99@(&
U8;99@%&Z;<:"-,$&),@81@&"C&>81&\1-D@&Z;<=1<,-D@KO^L(&
W91>>%&E(%&5LMP(&781&91@A,9;>"92&:1>;F"<,@:&;-$&@#,::,-.&A19C"9:;-B1&"C&2"D-.&@"BV121&
@;<:"-(&E"D9-;<&"C&>81&),@819,1@&W";9$&"C&U;-;$;&35H6JK55G^N533M(&
U;-;=1<<"%&I(&S(%&E(&0(&U;B8;>%&T(&U(&W11@"-%&*(&_(&_;CC""-%&U(&\9,:1@%&'(&*(&X;2:"91%&0(&)(&
T<1.;->1%&W(&+(&W;9>1<@%&I(&U(&X;9>&/&Z(&[(&T<V8;2;>%&3455(&01;@D9,-.&1-$"B9,-1&HB"9>,@"<J&
91@A"-@1@&"C&o1F9;C,@8&>"&@>91@@&`1F9;C,@8&-1D9"F18;=,"9;<&A9">"B"<@(&ZA9,-.19%&5^6N5P3(&
U;9<,-1%&S(&)(&/&E(&)(&0;B8D-.%&3445(&U9,>,B;<&7819:;<&0;?,:;&"C&',<$&;-$&!":1@>,B&Z>9;,-@&
"C&79"D>(&79;-@;B>,"-@&"C&>81&*:19,B;-&),@819,1@&Z"B,1>2&5^4HMJK5355N535M&
$",K54(56OOQ56PGNGM6LH3445J5^4l5355KB>:"#;m3(4(B"j3(&
U<;9V%&7(&!(%&0(&S(&!"-;<$@"-%&Z(&I,1A198"CC%&Z(&0(&!91--19%&*(&_">>"%&Z(&E(&U""V1%&Z(&\(&X,-B8%&!(&
*(&I;>>19@"-&/&*(&I(&);991<<%&3453(&I82@,"<".,B;<&W1-1C,>@&"C&W1,-.&Z:;<<&,-&;&U8;-.,-.&

66
'"9<$K&S1@A"-@1@&"C&U"8"&Z;<:"-&Ha-B"982-B8D@&V,@D>B8J&>"&;-&*BD>1&7819:;<&U8;<<1-.1&
;-$&;&Z,:D<;>1$&U;A>D91&T=1->(&I<"Z&"-1&OHMJK1^L4OL%&5NG(&
U<;9V%&7(&!(%&+(&0(&E1CC9,1@%&Z(&\(&X,-B8&/&*(&I(&);991<<%&3455(&T?B1A>,"-;<&;19"F,B&@B"A1&;-$&
B;9$,"=;@BD<;9&A19C"9:;-B1&"C&A,-V&@;<:"-&Ha-B"982-B8D@&."9FD@B8;J&:;2&D-$19<,1&
91@,<,1-B1&,-&;&#;9:,-.&B<,:;>1(&E"D9-;<&"C&T?A19,:1->;<&W,"<".2&35PH5GJK^4OPN^4G5&
$",K54(53P3QR1F(4M465O(&
U<;9V%&7(&!(%&T(&Z;-$F<":%&\(&+(&U"?%&Z(&\(&X,-B8&/&*(&I(&);991<<%&344G(&U,9BD<;>"92&<,:,>@&>"&
"?2.1-&@DAA<2&$D9,-.&;-&;BD>1&>1:A19;>D91&,-B91;@1&,-&>81&U8,-""V&@;<:"-&Ha-B"982-B8D@&
>@8;#2>@B8;J(&*:19,B;-&E"D9-;<&"C&I82@,"<".2NS1.D<;>"92&[->1.9;>,=1&;-$&U":A;9;>,=1&
I82@,"<".2&3L6H6JKS5M^5NS5M^L&$",K54(5563Q;RA91.D(L4PM5(344G(&
U""V1%&Z(&E(%&Z(&\(&X,-B8%&\(&7(&U9"@@,-%&!(&*(&I;>>19@"-%&+(&+(&T-.<,@8%&E(&0(&Z89,:A>"-%&\(&b(&!(&
+9;;V&/&*(&I(&);991<<%&344M(&I82@,"<".2&"C&,-$,=,$D;<&<;>1N9D-&)9;@19&S,=19&@"BV121&@;<:"-&
Ha-B"982-B8D@&-19V;J&@;:A<1$&,-&>81&"B1;-&B"991<;>1@&#,>8&C;>1&$D9,-.&@A;#-,-.&
:,.9;>,"-(&U;-;$,;-&E"D9-;<&"C&),@819,1@&;-$&*fD;>,B&ZB,1-B1@&M^HOJK5PMLN5PG4&
$",K54(55^LQC4MN4P3(&
!"#$%&'(&'(%&)(&*(&+,-.&/&0(&'(&!1--2%&3456(&7819:;<&=;9,;>,"-%&>819:;<&1?>91:1@&;-$&>81&
A82@,"<".,B;<&A19C"9:;-B1&"C&,-$,=,$D;<@(&781&E"D9-;<&"C&1?A19,:1->;<&F,"<".2&35GHI>&
53JK5L6MN5LMO&$",K54(53P3QR1F(55PL3M(&
!D-:;<<%&+(%&E(&S1,@>%&T(&U;9:;BV%&E(&W;F;<DV%&0(&I(&X1,$1NEY9.1-@1-&/&0(&!"BV19%&I;B,C,B&
Z;<:"-&,-&>81&*9B>,BK&X;9F,-.19@&"C&U8;-.1(&[-K&S1@A"-@1@&"C&*9B>,B&0;9,-1&TB"@2@>1:@&>"&
U<,:;>1&U8;-.1%&3453(&*<;@V;&Z1;&\9;->%&]-,=19@,>2&"C&*<;@V;&);,9F;-V@%&A&5P5N5M^(&

67
T<,;@"-%&T(&E(%&7(&!(&U<;9V%&0(&E(&X;.D1%&_(&0(&X;-@"-%&`(&Z(&\;<<;.819%&+(&0(&E1CC9,1@%&0(&+(&\;<1%&!(&
*(&I;>>19@"-%&Z(&\(&X,-B8&/&*(&I(&);991<<%&3455(&!,CC191-B1@&,-&>819:;<&>"<19;-B1&;:"-.&
@"BV121&@;<:"-&A"AD<;>,"-@(&ZB,1-B1&^^3HM436JK54LN53&$",K54(553MQ@B,1-B1(55LL56G(&
);991<<%&*(%&344L;(&T-=,9"-:1->%&;->1B1$1->@&;-$&B<,:;>1&B8;-.1K&<1@@"-@&C9":&>81&@>D$2&"C&
>1:A19;>D91&A82@,"<".2&;-$&9,=19&:,.9;>,"-&"C&@;<:"-,$@(&E"D9-;<&"C&T?A19,:1->;<&W,"<".2&
353H3^JK^OO5N^OG4(&
);991<<%&*(%&Z(&X,-B8%&Z(&U""V1%&!(&I;>>19@"-%&\(&U9"@@,-%&0(&_;A",->1&/&0(&0;>81@%&344G;(&
I;B,C,B&@;<:"-&,-&8">&#;>19K&;AA<2,-.&;19"F,B&@B"A1&:"$1<@&;-$&F,">1<1:1>92&>"&A91$,B>&>81&
@DBB1@@&"C&@A;#-,-.&:,.9;>,"-@(&I82@,"<".,B;<&;-$&W,"B81:,B;<&`""<".2&G5HMJKMLONO4L(&
);991<<%&*(%&U(&_11%&+(&7,19-12%&*(&X"$;<2%&Z(&U<D>>198;:%&0(&X1;<12%&Z(&X,-B8&/&*(&_">>"%&344^(&
),1<$‐F;@1$&:1;@D91:1->@&"C&"?2.1-&DA>;V1&;-$&@#,::,-.&A19C"9:;-B1&#,>8&;$D<>&I;B,C,B&
@;<:"-&D@,-.&;&:"F,<1&91@A,9":1>19&@#,:&>D--1<(&E"D9-;<&"C&),@8&W,"<".2&M3H5JKMPNGP(&
);991<<%&*(&I(%&344LF(&T-=,9"-:1->%&;->1B1$1->@&;-$&B<,:;>1&B8;-.1K&<1@@"-@&C9":&>81&@>D$2&
"C&>1:A19;>D91&A82@,"<".2&;-$&9,=19&:,.9;>,"-&"C&@;<:"-,$@(&781&E"D9-;<&"C&1?A19,:1->;<&
F,"<".2&353HI>&3^JK^OO5NG4&$",K54(53P3QR1F(43^MO5(&
);991<<%&*(&I(%&T(&E(&T<,;@"-%&T(&Z;-$F<":&/&7(&!(&U<;9V%&344L(&),@8&B;9$,"91@A,9;>"92&
A82@,"<".2&,-&;-&19;&"C&B<,:;>1&B8;-.1(&U;-;$,;-&E"D9-;<&"C&`""<".2&GOH54JKG^6NG65&
$",K54(55^LQo4LN4L3(&
);991<<%&*(&I(%&Z(&\(&X,-B8%&Z(&E(&U""V1%&!(&*(&I;>>19@"-%&\(&7(&U9"@@,-%&0(&_;A",->1&/&0(&7(&
0;>81@%&344GF(&I;B,C,B&Z;<:"-&,-&X">&';>19K&*AA<2,-.&*19"F,B&ZB"A1&0"$1<@&;-$&

68
W,">1<1:1>92&>"&I91$,B>&>81&ZDBB1@@&"C&ZA;#-,-.&0,.9;>,"-@(&I82@,"<".,B;<&;-$&W,"B81:,B;<&
`""<".2&G5HMJKMLONO4G&$",K54(54GMQ6L346O(&
)9,1$<;-$%&+(&!(%&S(&b(&';<V19%&d(&!(&!;=,@%&+(&'(&0219@%&\(&'(&W"18<19>%&Z(&]9;#;&/&q(&]1-"%&
3445(&aA1-N"B1;-&"9,1->;>,"-&;-$&91>D9-&:,.9;>,"-&9"D>1@&"C&B8D:&@;<:"-&F;@1$&"-&
>1:A19;>D91&$;>;&C9":&$;>;&@>"9;.1&>;.@(&0;9,-1&TB"<".2&I9".91@@&Z19,1@&35MK3^6N363(&
\;<F91;>8%&I(&)(%&d(&!(&*$;:@&/&7(&X(&0;9>,-%&344P(&[-C<D1-B1&"C&81;>,-.&9;>1&"-&:1;@D91:1->&
"C&>,:1&>"&>819:;<&:;?,:D:&,-&>9"D>(&*fD;BD<>D91&3P5H5NPJK6GON6LL&
$",K54(545MQR(;fD;BD<>D91(344P(4G(44P(&
\;:A19<%&*(&+(%&+(&E(&S"$-,BV%&X(&*(&);D@>%&T(&U(&b1--%&0(&7(&W1--1>>%&_(&[(&U9;#@8;#%&T(&S(&
+11<12%&0(&Z(&I"#1<<&/&X(&'(&_,%&3443(&01>;F"<,@:%&@#,::,-.&A19C"9:;-B1%&;-$&>,@@D1&
F,"B81:,@>92&"C&8,.8&$1@19>&91$F;-$&>9"D>&Ha-B"982-B8D@&:2V,@@&@@A(JK&1=,$1-B1&C"9&
A81-">2A,B&$,CC191-B1@&,-&A82@,"<".,B;<&CD-B>,"-(&I82@,"<".,B;<&;-$&W,"B81:,B;<&`""<".2&
O6H6JKP5^NP^5(&
X,-B8%&Z(%&Z(&U""V1%&*(&);991<<%&+(&0,<<19%&0(&_;A",->1&/&!(&I;>>19@"-%&3453(&!1;$&C,@8&
@#,::,-.K&;&91=,1#&"C&91@1;9B8&"-&>81&1;9<2&:,.9;>,"-&;-$&8,.8&A91:;>D91&:"9>;<,>2&,-&
;$D<>&)9;@19&S,=19&@"BV121&@;<:"-&a-B"982-B8D@&-19V;(&E"D9-;<&"C&),@8&W,"<".2&G5H3JK6OMN
6LL(&
X,-B8%&Z(&\(&/&E(&W9;>>2%&3444(&TCC1B>@&"C&@#,:&@A11$&;-$&;B>,=,>2&A;>>19-&"-&@DBB1@@&"C&;$D<>&
@"BV121&@;<:"-&:,.9;>,"-&>89"D.8&;-&;91;&"C&$,CC,BD<>&A;@@;.1(&79;-@;B>,"-@&"C&>81&
*:19,B;-&),@819,1@&Z"B,1>2&53LH3JK6LGNM4M(&

69
E;,-%&+(%&[(&W,9>#1<<&/&*(&);991<<%&5LLG(&S1A1;>&@#,::,-.&A19C"9:;-B1&"C&:;>D91&@"BV121&
@;<:"-&C"<<"#,-.&;&F9,1C&91B"=192&A19,"$K&;&A9"A"@1$&:1;@D91&"C&C,@8&81;<>8&;-$&#;>19&
fD;<,>2(&U;-;$,;-&E"D9-;<&"C&`""<".2&OMHGJK5PGGN5PLM(&
E;,-%&+(&T(&/&*(&I(&);991<<%&344^(&[-C<D1-B1&"C&@1;@"-;<&>1:A19;>D91&"-&>81&91A1;>&@#,::,-.&
A19C"9:;-B1&"C&9;,-F"#&>9"D>&a-B"982-B8D@&:2V,@@(&E"D9-;<&"C&T?A19,:1->;<&W,"<".2&
34MH34JK^6MLN^6OL(&
E"-@@"-%&W(&/&d(&E"-@@"-%&344L(&*&91=,1#&"C&>81&<,V1<2&1CC1B>@&"C&B<,:;>1&B8;-.1&"-&
;-;$9":"D@&*><;->,B&@;<:"-&Z;<:"&@;<;9&;-$&F9"#-&>9"D>&Z;<:"&>9D>>;%&#,>8&A;9>,BD<;9&
91C191-B1&>"&#;>19&>1:A19;>D91&;-$&C<"#(&E"D9-;<&"C&),@8&W,"<".2&O6H54JK3^G5N3PPO(&
+<1:1>@1-%&*(%&3454(&781&U8;99&I9"F<1:&S1=,@,>1$K&T?B1A>,"-;<&I81-">2A,B&I<;@>,B,>2&
I9":">1@&TB"<".,B;<&ZA1B,;>,"-&,-&I"@>.<;B,;<&_;V1@(&)91@8#;>19&S1=,1#@&^H5JKPLNOP&
$",K54(5M4GQC9RN^(5(^(&
+<1:1>@1-%&*(%&I(&*(&*:D-$@1-%&E(&!1:A@"-%&W(&E"-@@"-%&d(&E"-@@"-%&0(&aeB"--1<<&/&T(&
0"9>1-@1-%&344^(&*><;->,B&@;<:"-&Z;<:"&@;<;9&_(%&F9"#-&>9"D>&Z;<:"&>9D>>;&_(&;-$&*9B>,B&
B8;99&Z;<=1<,-D@&;<A,-D@&H_(JK&;&91=,1#&"C&;@A1B>@&"C&>81,9&<,C1&8,@>"9,1@(&TB"<".2&"C&
C91@8#;>19&C,@8&53H5JK5N6L(&
+Do-1>@"=;%&*(%&I(&W9"BV8"CC&/&S(&U89,@>1-@1-%&345P(&<:1971@>K&>1@>@&,-&<,-1;9&:,?1$&1CC1B>@&
:"$1<@(&S&A;BV;.1&=19@,"-&3(4N34(&

70
_11%&U(%&*(&);991<<%&*(&_">>"%&Z(&X,-B8&/&0(&X1;<12%&344^(&T?B1@@&A"@>N1?19B,@1&"?2.1-&
B"-@D:A>,"-&,-&;$D<>&@"BV121&Ha-B"982-B8D@&-19V;J&;-$&B"8"&Ha(&V,@D>B8J&@;<:"-&
C"<<"#,-.&B9,>,B;<&@A11$&@#,::,-.(&E"D9-;<&"C&T?A19,:1->;<&W,"<".2&34MH5GJK^36^N^3M4(&
0;BdD>>%&0(&E(%&Z(&\(&X,-B8%&*(&I(&);991<<&/&Z(&7"AA%&344P(&781&1CC1B>&"C&>1:A19;>D91&;-$&
;BB<,:;>,"-&A19,"$&"-&91A1;>&@#,::,-.&A19C"9:;-B1&,-&BD>>89";>&>9"D>(&E"D9-;<&"C&),@8&
W,"<".2&M6H3JK^P3N^6^&$",K54(5555QR(54L6NGMPL(344P(44P6^(?(&
0;BdD>>%&0(&E(%&Z(&\(&X,-B8%&U(&\(&_11%&E(&S(&I8,FF@%&*(&\(&_">>"%&0(&U(&X1;<12&/&*(&I(&);991<<%&
344M(&71:A19;>D91&1CC1B>@&"-&@#,::,-.&A19C"9:;-B1%&1-19.1>,B@%&;-$&;19"F,B&B;A;B,>,1@&"C&
:;>D91&;$D<>&A,-V&@;<:"-&Ha-B"982-B8D@&."9FD@B8;J&B":A;91$&#,>8&>8"@1&"C&@"BV121&
@;<:"-&Ha-B"982-B8D@&-19V;J(&U;-;$,;-&E"D9-;<&"C&`""<".2NS1=D1&U;-;$,1--1&!1&`""<".,1&
GPH5JKGGNLO&$",K54(55^LQo46N5G5(&
0;9>,-@%&T(&\(%&Z(&\(&X,-B8%&!(&*(&I;>>19@"-%&0(&E(&X;.D1%&Z(&E(&U""V1%&+(&0(&0,<<19%&!(&S"F,B8;D$%&
+(&+(&T-.<,@8%&*(&I(&);991<<&/&W(&E"-@@"-%&3453(&X,.8&9,=19&>1:A19;>D91&91$DB1@&@D9=,=;<&"C&
@"BV121&@;<:"-&Ha-B"982-B8D@&-19V;J&;AA9";B8,-.&@A;#-,-.&.9"D-$@&;-$&1?;B19F;>1@&
C1:;<1&:"9>;<,>2(&U;-;$,;-&R"D9-;<&"C&C,@819,1@&;-$&;fD;>,B&@B,1-B1@&MLH3JK^^4N^P3(&
0;>81@%&0(&7(%&Z(&\(&X,-B8%&Z(&E(&U""V1%&\(&7(&U9"@@,-%&!(&*(&I;>>19@"-%&*(&\(&_">>"&/&*(&I(&);991<<%&
3454(&TCC1B>&"C&#;>19&>1:A19;>D91%&>,:,-.%&A82@,"<".,B;<&B"-$,>,"-%&;-$&<;V1&>819:;<&91CD.,;&
"-&:,.9;>,-.&;$D<>&'1;=19&U911V&@"BV121&@;<:"-&Ha-B"982-B8D@&-19V;J(&U;-;$,;-&E"D9-;<&
"C&),@819,1@&;-$&*fD;>,B&ZB,1-B1@&MOH5JKO4NGP&$",K54(55^LQC4LN56G(&

71
0"9;%&U(&/&0(&)(&0;2;%&344M(&TCC1B>&"C&>81&9;>1&"C&>1:A19;>D91&,-B91;@1&"C&>81&$2-;:,B&
:1>8"$&"-&>81&81;>&>"<19;-B1&"C&C,@81@(&E"D9-;<&"C&7819:;<&W,"<".2&^5HPJK^^ON^P5&
$",K54(545MQR(R>819F,"(344M(45(446(&
a<,.-2NXgF19>%&X(%&U(&Z1-;2%&T(&U(&T-$19@&/&!(&W",@B<;,9%&3456(&TCC1B>@&"C&$,1<&>1:A19;>D91&
C<DB>D;>,"-&"-&>81&@>;-$;9$&:1>;F"<,B&9;>1&"C&RD=1-,<1&*><;->,B&@;<:"-&HZ;<:"&@;<;9JK&
[-C<D1-B1&"C&;BB<,:;>,"-&>1:A19;>D91&;-$&A9"=1-,1-B1(&U;-;$,;-&E"D9-;<&"C&),@819,1@&;-$&
*fD;>,B&ZB,1-B1@HR;J(&
I1--12%&U(&0(%&\(&'(&d;@8&/&*(&+(&\;:A19<%&345P(&U;9$,"91@A,9;>"92&91@A"-@1@&"C&@1;#;>19N
;BB<,:;>1$&;$D<>&*9B>,B&B8;9&HZ;<=1<,-D@&;<A,-D@J&;-$&*><;->,B&@;<:"-&HZ;<:"&@;<;9J&>"&;-&
;BD>1&>1:A19;>D91&,-B91;@1(&U;-;$,;-&E"D9-;<&"C&),@819,1@&;-$&*fD;>,B&ZB,1-B1@K5N54&
$",K54(55^LQBRC;@N345^N46ML(&
I1>>19@@"-%&*(%&E(&I,BV"=;&/&T(&W9;--;@%&3454(&Z#,::,-.&A19C"9:;-B1&;>&$,CC191->&
>1:A19;>D91@&;-$&C;>>2&;B,$&B":A"@,>,"-&"C&*9B>,B&B8;99&HZ;<=1<,-D@&;<A,-D@J&C1$&A;<:&;-$&
9;A1@11$&",<@(&*fD;BD<>D91&^44H5NPJK5OMN5G5&$",K54(545MQR(;fD;BD<>D91(3454(45(45O(&
S,B8>19%&*(&/&Z(&*(&+"<:1@%&3446(&0;?,:D:&71:A19;>D91&_,:,>@&C"9&U8,-""V%&U"8"%&;-$&
U8D:&Z;<:"-%&;-$&Z>11<81;$&79"D>&,-&>81&I;B,C,B&d"9>8#1@>(&S1=,1#@&,-&),@819,1@&ZB,1-B1&
5^H5JK3^NPL&$",K54(54G4Q54MP53M46L4GG6GM5(&
S,V;9$@1-%&*(&X(%&!(&E(&0(&T<<,">>%&E(&W(&!1:A@"-%&E(&Z>D9<;D.@@"-&/&*(&E(&E1-@1-%&344O(&781&
:;9,-1&>1:A19;>D91&;-$&$1A>8&A91C191-B1@&"C&*9B>,B&B8;99&HZ;<=1<,-D@&;<A,-D@J&;-$&@1;&
>9"D>&HZ;<:"&>9D>>;J%&;@&91B"9$1$&F2&$;>;&@>"9;.1&>;.@(&),@819,1@&aB1;-".9;A82&5MH6JKP^MN
PPO&$",K54(5555QR(5^M6N3P5L(344O(44PP6(?(&

72
S"$-,BV%&+(&E(%&*(&+(&\;:A19<%&+(&S(&_,o;9@%&0(&7(&W1--1>>%&S(&d(&S;D@B8&/&T(&S(&+11<12%&344P(&
7819:;<&>"<19;-B1&;-$&:1>;F"<,B&A82@,"<".2&;:"-.&91$F;-$&>9"D>&A"AD<;>,"-@&,-&@"D>8N
1;@>19-&a91."-(&E"D9-;<&"C&),@8&W,"<".2&MPH3JK^54N^^6&$",K54(5555QR(4433N
5553(344P(443L3(?(&
S"D?%&0(&E(%&S(&)(&7;<<:;-&/&U(&'(&_1#,@%&3455(&Z:;<<N@B;<1&*9B>,B&B8;99&Z;<=1<,-D@&;<A,-D@&
C,@819,1@&,-&U;-;$;e@&dD-;=D>K&:;-;.1:1->&B8;<<1-.1@&;-$&"A>,"-@(&E&),@8&W,"<&OLHMJK5M36N
PO&$",K54(5555QR(54L6NGMPL(3455(4^4L3(?(&
ZB8,-$<19%&!(&T(%&S(&X,<F"9-%&W(&U8;@B"%&U(&I(&W";>9,.8>%&7(&I(&nD,--%&_(&*(&S".19@&/&0(&Z(&
'1F@>19%&3454(&I"AD<;>,"-&$,=19@,>2&;-$&>81&A"9>C"<,"&1CC1B>&,-&;-&1?A<",>1$&@A1B,1@(&d;>D91&
PM6HO3LGJKM4LN53&$",K54(54^GQ-;>D914L4M4(&
Z>1,-8;D@1-%&0(%&T(&Z;-$F<":%&T(&T<,;@"-%&U(&b198,<<1&/&*(&);991<<%&344G(&781&1CC1B>&"C&;BD>1&
>1:A19;>D91&,-B91;@1@&"-&>81&B;9$,"91@A,9;>"92&A19C"9:;-B1&"C&91@>,-.&;-$&@#,::,-.&
@"BV121&@;<:"-&Ha-B"982-B8D@&-19V;J(&E"D9-;<&"C&T?A19,:1->;<&W,"<".2&355H3PJK^L56N
^L3M(&
71;:%&S(&U(%&345P(&SK&*&<;-.D;.1&;-$&1-=,9"-:1->&C"9&@>;>,@>,B;<&B":AD>,-.(&S&)"D-$;>,"-&
C"9&Z>;>,@>,B;<&U":AD>,-.%&b,1--;%&*D@>9,;%&3453(&[ZWd&^NL44465N4ON4(&
7,19-12%&+(&W(&/&*(&I(&);991<<%&344P(&781&91<;>,"-@8,A@&F1>#11-&C,@8&81;<>8%&:1>;F"<,B&9;>1%&
@#,::,-.&A19C"9:;-B1&;-$&91B"=192&,-&91>D9-N9D-&@"BV121&@;<:"-%&a-B"982-B8D@&-19V;&
H';<F;D:J(&E"D9-;<&"C&C,@8&$,@1;@1@&3OKMM^NMO5(&

73
7,19-12%&+(&W(%&*(&b(&+;@D9;V%&W(&Z(&`,1<,-@V,&/&!(&0(&X,..@%&3455(&Z#,::,-.&A19C"9:;-B1&;-$&
,-=;@,"-&A">1->,;<&"C&>81&9"D-$&."F2(&T-=,9"-:1->;<&W,"<".2&"C&),@81@&L3HPJKPL5N643&
$",K54(544OQ@54MP5N455NLGMON3(&
'9"-;%&)(&E(%&7(&!(&I9"#@1%&E(&!(&S1,@>%&E(&T(&X"FF,1%&_(&0(&E(&_g=1@fD1&/&'(&)(&b,-B1->%&344M(&+12&
),-$,-.@%&ZB,1-B1&\;A@&;-$&I"<,B2&S1B"::1-$;>,"-@(&*0W[aK&*&E"D9-;<&"C&>81&XD:;-&
T-=,9"-:1->&^6HOJKP55NP56&$",K54(56OLQ44PPNOPPOH344MJ^6hP55KVC@.;Ai3(4(B"j3(&

74
Tables:
Table 3.1. Mophometrics of wild and lab reared juvenile arctic char and rainbow trout
&&*9B>,B&U8;9&
H<;FJ&*9B>,B&B8;9&
H#,<$J&S;,-F"#&>9"D>&
d& 34& ^6& 36&
)"9V&_1-.>8&H::J& 345rP& 5LLrP& 5OPrPu&
0;@@&H.J& LGrM(3& O^(4r^(Mu& MO(5rP(3u&
U"-$,>,"-&H+J& 5(5Lr4(4^& 4(GMr4(3u& 5(3Pr4(43&
b1->9,B<1&:;@@&H.J& 4(5Pr4(45& dQ*& 4(4Gr4(45u&
Sb0&HvJ& 4(56r4(45& dQ*& 4(55r4u&
u&[-$,B;>1@&@,.-,C,B;->&$,CC191-B1&C9":&<;F&91;91$&;9B>,B&B8;9&Hs&4(46J&
Data are shown as mean r&@>;-$;9$&199"9&
RVM = Relative ventricular mass

75
Table 3.2 Regression models of the initial (!"#$%$&'() and repeat (!)*+*&%) swimming performance
and recovery ratio (RR) of wild arctic char over the tested range of naturally occurring
temperatures
&U"1CC,B,1->& ZT& !" #"
!"#$%$&'$/01'2(
U"-@>;->& NMM(O& OO(3& N4(GM& 4(^LP&
71:A19;>D91& N4(P6& 4(3L& N5(6^& 4(5^M&
71:A19;>D913& 56(L& L(6O& 5(MM& 4(54O&
!)*+*&%$/01'2&
U"-@>;->& 6(PG& 3P(4& 4(3^& 4(G35&
71:A19;>D91& P(LM& 5(65& ^(3G& 4(44^&
<<$/01'2$& & & &
U"-@>;->& 4(6O& 4(3L& 5(LL& 4(46L&
71:A19;>D91& 4(4P& 4(43& 3(4O& 4(464&
$%&%!%'(")"S3&t&4(5P%&)3%^5t&3(6^%&At&4(4LM" && && && &&
$*+#+'!)&S3&t&4(^3%&)5%3^t&54(OO%&At&4(44^" && && && &&
SSK&S3&t&4(5M%&)5%3^t&P(3L%&At&4(464& && && && &&

76
Table 3.3 Swimming performance, aerobic metabolism and blood analysis of rainbow trout and
arctic char exposed to control, simulated diurnal fluctuation or diurnal warming treatments.
&<7.(=0-$*509*$
$>5?*.?$?+75$
&@0(*502$ A29?*97*.0($ B75/.()$
$@0(*502$ A29?*97*.0($ B75/.()$
C-.//.()$D'5305/7(?'&$%&%!%'("&HB:&@N5J& O5(Pr6(5;& G3(4r^(6;F& L4(Lr^(6F&
&MP(Or^(3;B& MP(Pr6(5;B& O4(4r^(5;B&
$*+#+'!&&HB:&@N5J& GM(Lr6(O;& L6(5r6(O;& LL(3r6(M;&&
O6(Mr^(O;& O5(Or55(3;F& ^L(Mr55(4F&S1B"=192&H$9N$,J& 56(6r^(6;& 5^(4r^(P;& L(6rP(^;&
&55(4rP(6;& O(3rO(4;& N^4(Pr54(4F&
]\7&Hv&$,J& L3(3r^(G;F& L^(3r3(G;F& L6(3r4(6;&&
L3(5r4(L;F& L^(Pr4(M;F& GO(Gr5(MF&>'50=.?$/'*7=02.4/&0a39"D>,-1H:.&V.N5&:,-N5J& P(^Mr4(P^;& M(35r4(GG;F& O(OMr4(MF&
&P(^Pr4(^G;& P(MPr4(6L;& 6(GGr4(63;F&
0a3:;?&H:.&V.N5&:,-N5J& 54(O^r5(4M;& 5^(P3r5(6^;F& 56(3r4(PPF&&
L(MPr4(OG;& L(5Or4(GL;& 54(PLr4(MG;&ZB"A1&H:.&V.N5&:,-N5J& M(^Or5(56& O(35r5(4O& O(PPr4(M6&
&6(^r4(6^& P(6^r4(6L& P(M5r4(M6&
);B>"9,;<&@B"A1& 3(M5r4(^3& 3(^^r4(3P& 3(4Pr4(5M&&
3(3Pr4(5P& 3(4^r4(5M& 5(GPr4(5P&E2001$7(7264.4&X1:;>"B9,>&HvJ& P5(^r5(G;& P4(6r5(M;F& P^r3(5;&
&P3r3(^;& ^O(^r3(^;F& ^^r5(MF&
a@:"<;<,>2&H:aZ0J& ^P^rO& ^^PrO& ^MOr5^&&
^3Mr3& ^M4r5L& ^PMrL&U"9>,@"<&H-.&:_N5J& LLrG(P& 546(3r5M(P& L6(3r5^(M&
&L3(Gr5P(L& O5(3r54(L& 63(^rO(P&
I<;@:;&\<DB"@1&H:0J& 53(Mr5(6;& 5P(Gr5(6;& 5O(Or3(P;F&&
L(Lr4(O;& 55(Or3(5;& 35(6r4(LF&I<;@:;&_;B>;>1&H:0J& 5L(^r3(5;F& 5G(Mr5(G;F& 33(Or4(LF&
&56(3r4(G;& 5O(3r5(3;F& 34(6r4(6;F&
Dissimilar superscript letters of the same capitalization indicate significant differences within rows Hs&4(46J.
Data are shown as mean r&@>;-$;9$&199"9

77
7;F<1&^(P&_,-1;9&:,?1$N1CC1B>@&:"$1<@&"C&;9B>,B&B8;9&;-$&9;,-F"#&>9"D>&;19"F,B&:1>;F"<,@:&$D9,-.&;-$&C"<<"#,-.&H91B"=192J&;&C;>,.D,-.&1?19B,@1&>1@>&D-$19&91C191-B1&B"-$,>,"-@&H55°C) or&C"<<"#,-.&;&$,D9-;<& #;9:,-.& "9& C<DB>D;>,"-& >91;>:1->(&';>19& =1<"B,>2& #;@& ,-B91;@1$& F2& 4(6& W_& @N5& 1=192& >1-&:,-D>1@&D->,<& C;>,.D1&$D9,-.& >81&1?19B,@1& >1@>%&;-$&81<$&B"-@>;->&;>&4(6&W_&@N5&$D9,-.& >81&91B"=192&A19,"$(&
),?1$&1CC1B>& U"1CC,B,1->& ZT& !)& !" #">5?*.?$?+75$FG'5?.4'$HI!$/01'2$J/$K"L!#$?$K"LM;N$U"-@>;->& !"#$%& !#'(& '!& )#**& "#"""!7,:1&H:,-J& "#!!& "#"!& %"*& %%#+'& "#"""&<'?0O'56$HI!$/01'2$J/$K$"LPQ$?$K$"LMQN&U"-@>;->& **#*,& '#)'& ''& %+#'!& "#"""&-./01/21345& !#"!& *#+%& ',& "#,(& "#+"$&6278359& *#')& *#,,& '!& "#)'& "#!,(&:38;& <"#,*& "#")& ,%& <(#*,& "#"""&-./01/21345=:38;& <"#%(& "#%*& ,'& <%#%(& "#'*$&6278359=:38;& "#+*& "#%*& ,%& !#)(& "#"""&<7.(=0-$*509*$FG'5?.4'$HI!$/01'2$J/$K$"LR#$?$K$"LRSN$
U"-@>;->& <%""#$,& '!#($& +!& <*#'*& "#"""&>49?82@@A& ')#+)& +#,(& +*& +#''& "#"""&:38;& <"#))& "#!'& %$*& <!#%'& "#""'&-./01/21345& ,#+,& !#"(& %)& '#%*& "#"*+&6278359& %+#,%& '#),& %)& +#'(& "#"""&>49?82@@A=:38;& "#')& "#"$& %$*& !#$'& "#"""&<'?0O'56$HI!$/01'2$J/$K$"LQQ$?K"LMRN&U"-@>;->& <()#*'& %$#""& !,& <*#*%& "#"""&>49?82@@A& '(#)*& *#%+& !,& ,#(*& "#"""&-./01/21345& !#$+& !#++& !+& %#")& "#'$+&6278359& %*#,'& !#++& !+& *#%%& "#"""&:38;& !#""& "#(!& %'%& *#%%& "#"""&>49?82@@A=:38;& <"#$+& "#%(& %'%& <*#)+& "#"""&-./01/21345=:38;& <"#!%& "#%'& %%!& <'#,"& "#"%%&6278359=:38;& "#"!& "#%'& %%*& "#'+& "#$""&-3@B&CD&E2@&350./F;F&35&2..&84F;.@&2@&2&725F48&;GG;01&:&t:;9.,-;<&S3& && && && && &&Bt&B"-$,>,"-;<&S3& && && && && &&&

78
Figures
Fig. 3.1. Map of the study area in northwest Nunavut, on the Dolphin and Union Strait in the
Arctic Ocean. The study population migrates annually between Hingittok Lake and Bernard
Harbour via Nulahugyuk Creek.
Fig. 3.2. Natural diurnal temperature fluctuations recorded in Nulahugyuk Creek in 2012 (A) and
the resulting laboratory study design (B). The simulation was designed to assess the effects of
diurnal temperature variation similar to the blowup in (A) on repeat swimming performance and
related sub-organismal physiology.
Fig. 3.3 Initial (top) and repeat (middle) swimming performance and recovery ratio (bottom) of
wild juvenile arctic char over a range of naturally occurring temperatures. The red circle denotes
individuals that were removed as outliers from their respective regression analyses based on
visual inspection. Regression models are in Table 3.2. The individuals removed were the only
two tested above 20°C and may indicate a threshold for recoverability.
Fig. 3.4 Initial and repeat swimming performance and recovery ratio of laboratory reared
juvenile rainbow trout and arctic char following simulated diurnal warming or fluctuation.
Dissimilar letters indicate significant differences between treatments within a species (! 0.05).
Error bars represent standard error of the mean.
Fig. 3.5 Oxygen consumption (MO2) during an initial exercise test (solid lines) and recovery
(broken lines) by laboratory reared juvenile rainbow trout and arctic char exposed to diurnal
temperature treatments. Error bars represent standard error of the mean. Summary statistics of
routine and maximum MO2 are in Table 3.3, and linear mixed effects models for describing the
data are in Table 3.4

79
Figure 3.1

80
Figure 3.2

81

82

83

84
Chapter 4: Diurnal temperature fluctuations induce drastic
biochemical and transcriptional changes in exercised arctic char
(Salvelinus alpinus) and rainbow trout (Oncorhynchus mykiss)
Abstract
In a companion study (Chapter 3), I exposed arctic char and rainbow trout to simulated diurnal
temperature fluctuations that are representative of the upper end of those currently experienced
by migrating arctic char in Nulahugyuk Creek, Nu. Following this exposure fish were subjected
to fatiguing exercise tests and measures of their recoverability, aerobic and anaerobic
metabolism, and stress were assessed. Following diurnal warming, arctic char had an impaired
ability to recover from fatiguing exercise and generally exhibited signs of insufficient energy
production. In the present study I carried out metabolomic and qPCR analysis on plasma and gill
samples from the same fish to identify biochemical and transcriptional changes associated with
my previous results. Following diurnal warming both species exhibited substantial shifts in
metabolite profiles and exhibited strong heat shock responses, however in both cases the
magnitude of response was typically greater in char than trout. Among the key differences
between species was a substantially higher accumulation of plasma succinate in char, indicating
a greater loss in mitochondrial functionality, and an order of magnitude higher induction of gene
expression for heat shock proteins (HSP) such as HSP90!, and HS70. A similar pattern of heat
shock response was also identified in wild adult arctic char that naturally encountered warm
water compared to those that were in cooler conditions, however the magnitude of response was
substantially lower. Following a complete simulated diurnal fluctuation the heat shock response
had largely recovered in trout but was still significantly elevated in arctic char. Together these

85
results indicate that current thermal conditions in the Canadian Arctic are sufficient to
substantially disrupt critical biochemical process and cause severe thermal stress in arctic char.
Furthermore, these findings suggest that anadromous salmonids that are adapted to historical
arctic conditions may already be at a competitive disadvantage during their summer migration
when compared to more temperate species whose ranges are expanding northward.

86
Introduction
As a result of global climate change, migrating salmonids encounter temperatures outside of their
optimal range at increasing frequency (Farrell et al. 2009). In response to these temperatures,
changes in gene expression and biochemistry occur that may be protective and compensatory, or
as a result of the disruption of numerous biological processes (Iftikar et al. 2014; Jeffries et al.
2012b; Quinn et al. 2011b). For researchers, these changes can serve as biomarkers of thermal
stress (Iftikar et al. 2014) and may provide insight into the mechanisms underlying the reductions
in functionality that are observed at higher levels of organization (Cooke et al. 2006; Iftikar et al.
2014; Jeffries et al. 2012b).
Studies that involve the biochemical responses of salmonids to environmental stressors often
focus on a few key metabolites and related enzymes. For example, the concentrations of plasma
glucose and lactate as well as the activities or concentrations of related enzymes, citrate
synthanse (CS) and lactate dehydrogenase (LDH) are often studied as indicators of the supply to,
and functionality of aerobic and anaerobic fuel systems (Berli et al. 2014; Mathes et al. 2010).
Furthermore, these are all targets that are relatively easily assayed using commercially available
kits and analyzers (Cooke et al. 2006; Mathes et al. 2010; Stoot et al. 2014). The continued
development of the field of metabolomics has increasingly allowed researchers to study many
metabolites simultaneously in more complex studies to give a more complete picture of
biochemical responses to various stressors. Recently, metabolite profiles were used to identify
thermal sensitivity in wrasse species (Iftikar et al. 2014). These found the mitochondria of
difference species had different thermal sensitivities that corresponded changes in their
metabolite profiles. Metabolite profiles have also been proposed as a way to screen for thermal
stress and general disruption of homeostasis in wild fish (Iftikar et al. 2014)

87
The study of transcriptional changes has also benefitted greatly by recent advances in
transcriptomic technology that are now widely being applied in the field of environmental
physiology (Evans et al. 2011; Jeffries et al. 2012b; Quinn et al. 2011a; Quinn et al. 2011b).
Recently multiple studies have used transcriptomic techniques to identify changes in gene
expression related to thermal tolerance and heat stress in salmonids. Among the genes identified
changes in the expression of members of the heat shock family and of genes involved in cell
cycling were common (Jeffries et al. 2012b; Quinn et al. 2011b). The target genes in the present
study were selected based on these previous studies and fell into two broad categories based on
their cellular function. Cyt-C2, Casp3, and Casp8 all code for proteins involved in the signaling
of apoptosis, and Ubq, GRPP, Serpinh1, HSP90!-2, HSP90!, and HS70 are all broadly involved
in the maintenance of protein functionality and cycling. Except for Ubq, the non-apoptosis
related genes are all part of the heat shock family. Together, the proteins encoded by the target
genes help maintain cellular homeostasis or terminate the cell if homeostasis is significantly
disrupted following environmental perturbations such as thermal variation.
As with other areas of thermal physiology such as those addressed in the companion article
(Chapter 3), the effects of temperature on heat shock proteins and apoptosis related gene
expression, or on the metabolome have not been extensively studied under realistic thermal
regimes, or in arctic char outside of the aquaculture setting (Quinn et al. 2011b). Specifically, no
study has investigated either of these endpoints in response to large but realistic diurnal
temperature fluctuations.
In the present study, our objectives were to (i) determine the relative transcriptional and
biochemical sensitivity of arctic char, a model northern salmonid and rainbow trout, a model
temperate salmonid, to realistic diurnal temperature regimes, (ii) assess the utility of previously

88
proposed biomarkers for thermal stress under these realistic thermal regimes, and (iii) begin to
establish how the observed changes are related to each other, and to the metrics of stress,
metabolism and whole-organism performance addressed in our companion study (Chapter 3). To
achieve these objectives we conducted a nuclear magnetic resonance (NMR) based
quantification of a large suite of plasma metabolites and used quantitative polymerase change
reaction (qPCR) analysis to quantify changes in the expression of target genes following
exposure to simulated or natural diurnal temperature fluctuations.
Materials and Methods
Field sampling
Field sampling took place in the summers (June – August) of 2012–2014 near the mouth of
Nulahugluk creek, Nu (68.75°N 114.8°W). Fish were captured migrating between their
freshwater and marine environments (Hingittok Lake and Bernard Harbour, Nu,) as in Chapter 3.
Gill tissue samples were only taken from fish that were harvested by locals or from moribund
individuals that were not likely to survive. Several filaments were clipped from the middle of the
first arch and transferred into 1.5 or 2mL microcentrifuge tubes containing 1mL of RNAlater
(Life Technologies, Grand Island, New York). Samples were stored in field at -20°C in a
portable freezer, shipped by air to the University of Alberta (UofA, Edmonton, Alberta) on ice,
and then stored -20°C until use. Fish sampled in field were classified as having experienced
warm (>19°C) or cool (<15°C) water conditions within the previous 24 hours.
Laboratory Sampling
In the laboratory, all plasma and gill tissue samples were taken from individuals used in the
companion study (Chapter 3) following their exposure to a temperature treatment and repeat

89
swimming performance tests. Briefly, ten to fifteen month old rainbow trout and arctic char were
exposed to a complete diurnal temperature fluctuation (11 to 21 to 11°C), diurnal warming (11 to
21°C) or control conditions (11°C), following which they were put through an endurance and a
sprint swimming performance test separated by a 30 min recovery period. Plasma samples were
obtained as previously described (Chapter 3) but were further filtered through 200 kda Nanosep
microcentrifuge filters (Pall Corporation, Port Washington, New York) 4°C for 30min at 10,000g
prior to NMR analysis. Gill tissue was taken immediately following sacrifice. The entire first
arch was excised and the filaments were trimmed away from the cartilage and placed in
RNAlater overnight at 4°C before being transferred to -20°C until use.
Metabolomics
Three plasma samples from each treatment group and both species (Ntotal = 18) were supplied to
Chenomx Inc. (Edmonton, Alberta) for analysis. NMR spectra were generated using Varian four-
channel VNMRS 700 MHz NMR spectrometer (Varian, Palo Alto, California) with an auto
tuning 1H/13C triple resonance biomolecular probe. Spectra were analyzed using Chenomx
NMR suite 7.7 to generate metabolite concentrations within each sample.
Quantitative PCR
RNA was extracted using mechanical disruption and Trizol reagent (Life Technologies). For
mechanical disruption, the tissue was ground with a micropestle homogenizer and vortexed with
~0.5mL of 2.3 mm zirconium dioxide beads (BioSpec, Bartlesville, Oklahoma). RNA was
treated with DNAse (DNA-free, Life Technologies) and cDNA was made using the SuperScript
First-Strand synthesis system (Life Technologies). qPCR was run using the SybrGreen method
and primers (Table. 4.1) on a 7500 Fast Real-Time PCR System (Life Technologies). Raw ct

90
values were exported and used to generate relative quantitation (RQ; fold change) values using
the !! ct method with correction for differences in primer efficiencies.
Data analysis
All data analysis was carried out using R Studio (Team 2014). Data presentation was done using
R Studio, Sigmaplot 11.1 (Systat Software, San Jose, California) and Adobe Illustrator CS4
(Adobe systems, San Jose, California).
For broad scale analysis of laboratory qPCR and metabolomics data Principle components
analyses (PCA) were conducted on each data set. 68% confidence limit ellipses were plotted for
each treatment group to show separation of transcript and metabolite profiles between species
and treatments. Metabolomics data was further analyzed using heat maps of fold change from the
control group for both species and select pairwise comparisons were made with t-tests using the
Benjamini-Hochberg procedure to control for false discover rate. qPCR data was analyzed using
species and treatment as predictor variables in a two way analysis variance followed by Tukey’s
honest signi ficant difference post hoc test.
Results
Metabolite profiles following diurnal simulations
In rainbow trout and arctic char the principle components analysis of metabolite profiles showed
a large separation in component space between fish that had experienced diurnal warming
compared to those that had experienced a complete fluctuation or were held under control
conditions (Fig. 4.1). There was a less pronounced separation between the control and fluctuation
groups. Across all treatment groups there was a clear separation between species (Fig. 4.1).

91
At the individual metabolite level, in both species many amino acids including alanine, iso-
leucine, leucine, phenylalanine, serine, taurine and tyrosine showed large increases in their
concentration following exposure to diurnal warming but in most cases were at near control
levels in the diurnal fluctuation group (Table 2.2; Fig. 4.2). Important energy metabolites
including glucose and lactate were also elevated following diurnal warming but to a lesser extent
in rainbow trout than in arctic char. In both species the concentration of succinate, an important
intermediate of the citric acid cycle, was also higher in diurnal warming than control groups, and
had returned to or below control levels following diurnal fluctuation (Table 2.2; Fig. 4.2). The
increase in succinate was much larger for char than trout. Interestingly, plasma creatine was
highly variable between individuals but in general was elevated following warming in rainbow
trout but not arctic char.
The separation in plasma metabolite profiles between species (Fig. 4.1) was also apparent at the
individual metabolite level. Alanine, asparagine, glutamate, glutamine, hydroxyproline,
isoleucine, methionine, phenylalanine, serine, tyrosine and valine were all markedly higher in
arctic char than rainbow trout in a manner that was independent of the diurnal temperature
treatment (Table 2.2; Fig. 4.2).
qPCR following diurnal simulations
In rainbow trout and arctic char the principle components analysis of transcript profiles for arctic
char showed the largest separation between control and the diurnal warming treatment, with the
diurnal fluctuation treatment being intermediate (Fig. 4.3). In rainbow trout there was smaller
separation between the warming and control group but in the same direction as seen in arctic

92
char (Fig. 4.3). Rainbow trout profiles in the diurnal fluctuation group had substantial overlap
with the control group, which was not observed in arctic char (Fig. 4.3).
The separation observed between the control and the warming group in the PCA analysis was
apparent at the individual target gene level. Specifically, members of the heat shock protein
family including GRPP, Hsp90!, HSP70 were all highly induced following exposure to diurnal
warming in both species. However, this induction occurred to a much greater extent in char than
in trout, and char also showed significant induction of Serpinh1 and Hsp90!-2, which was not
seen in trout (Fig. 4.4). Following the complete diurnal temperature fluctuation the expression of
all thermally induced genes were near control levels for rainbow trout but, Serpinh1, Hsp90! and
HS70 were all still substantially elevated in arctic char (Fig. 4.4). In arctic char Cyt-C2 was
numerically increased following diurnal warming and fluctuation but due to high variation this
difference was not statistically significant (Fig. 4.4). No significant differences in the expression
of Casp3, Casp8 or Ubq were seen between any treatment groups for either species.
qPCR in wild adult arctic char
Compared to char at relatively cool temperatures, wild arctic char that had experienced water
temperatures above 19°C in the past 24 hours induced expression of the same suite of heat shock
genes as was seen following simulated warming in lab, excluding GRPP (Fig. 4.5). The
induction of heat shock gene expression in wild adults however, was not nearly at the same
magnitude as in lab. Interestingly, wild char at warm temperatures and juveniles in lab under
simulated warming conditions exhibited a similar magnitude increase in the expression of Cyt-
C2 (Fig. 4.5).

93
Discussion
Summary
Temperature is well established as one of the most important environmental factors that shapes
the physiological performance of salmonids and ectotherms in general(Currie 2011; Farrell
2007), however metabolite and transcript profiles have not been thoroughly studied under natural
thermal regimes, and are particularly unknown in northern fishes. In the present study I identify
that arctic char and rainbow trout exhibit drastic metabolomic and transcriptional responses to
diurnal temperature variation that is representative of current conditions in the Canadian arctic.
While both species exhibited these responses, arctic char appeared to be more sensitive and less
able to recover to a basal state following thermal variation.
Metabolite profiles following diurnal simulations
The overall change in metabolite profiles following diurnal warming indicates significant
disruption of biochemical processes with warming. The specific metabolites involved provide
further insight into which specific processes may be affected. The substantial increases observed
in various plasma amino acids is indicative of an increase in proteolysis (Mommsen 2004).
Increased protein degradation serves a number of purposes. Following exercise without thermal
variation arctic char and salmonids in general increase proteolysis to supply amino acids for
rebuilding damaged tissue and replenishing the general supply of cellular substrates (Barton et al.
1995; Mommsen 2004). Because all fish in the current study were exercised to a similar extent
the most likely purpose is that these amino acids are being used to regenerate components
involved in ATP production because of the heightened cost of being at warm temperatures. One
particular amino acid that may serve this purpose is alanine, which drastically increased in

94
concentration after warming. In addition to lactate, alanine is used by hepatocytes in
gluconeogenesis to replenish glucose (Suarez and Mommsen 1987). Migrating salmonids also
use their protein reserves to fuel sexual maturation so the depletion of these reserves caused by
swimming through warmer water would likely decrease reproductive output (Mommsen 2004).
The pattern of accumulation of succinate in plasma between treatment groups and species
provides particular insight into potential mechanisms underlying the observed changes in aerobic
metabolism (Chapter 3). Succinate is oxidized by complex II of the citric acid cycle (Succinate-
dehydrogenase) and its accumulation is an indication of impaired mitochondrial functionality
(Grieshaber et al. 1994). A number of fish species have now been shown to exhibit increased
succinate levels during thermal stress (Iftikar and Hickey 2013; Iftikar et al. 2014; Pörtner and
Knust 2007). However, the extent of accumulation observed in the present study, 5 and 10 fold
increase in trout and char respectively (Table 4.2), appears to be unprecedented. The likely
explanation for this is that fish in our study were subjected to fatiguing exercise tests that
resulted in maximal aerobic output and thus any succinate accumulation as a result of
mitochondrial dysfunction at warm temperatures would have been magnified. Although both
species exhibited accumulation of plasma succinate following diurnal warming, it was much
greater in char suggesting that that they experienced a relatively far greater loss of mitochondrial
functionality. In addition to succinate accumulation in heat stressed fish, Iftikar et al (Iftikar et al.
2014) also observed a marked reduction in the respiratory control ratio (RCR), a metric of
mitochondrial efficiency. Interestingly, a substantial (~50%) reduction in RCR was also found
following acute thermal stress in arctic char when compared to atlantic salmon at similar
temperatures (Penney et al. 2014), agreeing with differences in succinate levels in char and trout
found in the present study. The increase in plasma lactate and glucose following diurnal warming

95
agrees with the my findings in chapter 3 and with previous research (Jain and Farrell 2003) that
suggest there is increase glucose metabolism through anaerobic pathways following temperature
stress. This increase in anaerobic metabolism is thought result from and inability to increase
aerobic supply of ATP to match increased demand, potentially as a result of mitochondrial
dysfunction as discussed above, or as a result of limited oxygen supply by the cardiorespiratory
system (Eliason et al. 2011; Farrell et al. 2009).
While they are not necessarily related to the differences in sensitivity to thermal variation,
overall differences between species in their metabolite profile and specific metabolite
concentrations may reveal fundamental physiological differences. For example, there were
eleven amino acids that were in higher concentration in all treatment groups for arctic char than
rainbow trout. This suggests that following fatiguing exercise arctic char had an overall higher
level of proteolysis than rainbow trout, and were likely using amino acids as a fuel source to a
greater extent. A similar difference can be found between sockeye salmon that extensively utilize
protein reserves to fuel upstream migration, and atlantic salmon that do not (Mommsen 2004).
Hydroxyproline was among the amino acids that were generally at a higher level in char than in
trout. Hydroxyproline is primarily found in collagen, and its high concentration in plasma
indicates that structural proteins such as collagen were likely not protected from the elevated
proteolysis exhibited by char (Mommsen 2004), and thus you would expect a greater structural
turnover in char than in trout.
qPCR following diurnal simulations
The broad function of the selected target gene is to regulate apoptosis and protein homeostasis
under environmental stress. As such, the overall difference between transcript profiles of fish

96
under control conditions and those that had experienced simulated diurnal warming suggests that
this thermal variation was sufficient to substantially perturb cell homeostasis and induce a
compensatory response. Given the larger separation in char than in trout, the disruption of
homeostasis was likely more severe in char, suggesting that even over temperature ranges
currently encountered in the Arctic, they are more thermally sensitive than temperate salmonids.
Indications of this heightened thermal sensitivity are also found in the expression of a number of
individual gene targets. For example, inductions of HSP90! and HS70, classical members of the
inducible heat shock response, were over an order of magnitude higher in char than in trout.
Furthermore, in arctic char the heat shock response was still highly apparent after the completion
of the diurnal fluctuation, while it had been reduced back to control levels in rainbow trout. This
difference was reflected in the transcript profile PCA as well, in which there was significant
overlap between the control and diurnal fluctuation groups for rainbow trout, but for arctic char
the distribution was intermediate to the control and diurnal warming group. In the wild, these
char would immediately begin another diurnal cycle before they had recovery from their last.
This aligns well with indications in chapter 3 and assertions in a growing number of studies that
thermal variation and thermal history matter for an organism’s performance and underlying
physiology as opposed to just the instantaneous, or average temperatures (Currie 2011; Dowd et
al. 2015; Oligny-Hébert et al. 2015; Quinn et al. 2011b).
The magnitude of the inductions of heat shock protein gene expression in char and trout in the
diurnal warming group are among the largest identified in any salmonid despite the fact that
higher temperatures or longer exposures are often used to induce these responses (Anttila et al.
2014; Fowler et al. 2009; Jeffries et al. 2012b; Palmisano et al. 2000; Quinn et al. 2011a; Quinn
et al. 2011b). While different HSPs are known to act over different time scales (Currie 2011) the

97
extent of induction may suggest that in some cases the heat shock response of salmonids may be
maximally induced over naturally relevant time scales for thermal variation. This particular study
was not designed to test such a hypothesis but based on the magnitude of observed induction it
certainly warrants further investigation.
qPCR in wild adult arctic char
Migrating adult arctic char had similar patterns of heat shock response to those under simulated
diurnal temperature regimes, albeit at a lesser magnitude. This suggests that under current
migratory conditions in Nulahugyuk Creek, Nu, arctic char are experiencing significant thermal
stress(Currie 2011). Interestingly both wild and lab-tested char at warm temperatures also had
elevated expression of Cyt-C2. When released from the mitochondrion, Cyt-C2 in the cytoplasm
plays a signaling role in the initiation of apoptosis (Loeffler and Kroemer 2000). In wild sockeye
salmon, Cyt-C2 gene expression was not induced by heat stress alone but was significantly
elevated in moribund individuals at warm or cool temperatures (Jeffries et al. 2012b). As such, it
is likely that the elevated expression in the present study in an indication of elevated apoptotic
signaling at warm temperatures. Alternatively, Cyt-C2 is a critical component of the electron
transport chain and expression may have been stimulated by the inability to meet aerobic
metabolic demands.
One important difference between the fish sampled for qPCR analysis was that wild arctic char
were adults and captive char were juveniles. Juvenile rainbow trout are known to have a more
pronounced heat shock response than adults, and this may underlie a greater relative thermal
tolerance (Fowler et al. 2009), which has multiple implications for the present study if it holds
true for arctic char. Although the natural thermal variation experienced by wild adults was less

98
consistent between individuals and potentially less harsh than that experienced by lab-tested
juveniles, part of the difference in the magnitude of their heat shock responses was likely a result
of age or size. Furthermore, our laboratory studies revealed concerning physiological effects of
diurnal temperature variation on juveniles, so if there is indeed a functional reduction in the adult
heat shock response, the physiological impairments observed in lab would likely be more
pronounced in adults.
The heat shock response has been successfully used as a biomarker for thermal stress in other
salmonid species (Anttila et al. 2014; Jeffries et al. 2012b; Lund et al. 2002). It has also been
confirmed to be robust in captive arctic char under non-realistic thermal regimes intended to
simulate aquaculture settings or acutely induce loss of equilibrium (Quinn et al. 2011a; Quinn et
al. 2011b). The heat shock response however, has not been previously assessed in arctic char
under environmentally relevant thermal conditions in the lab or in the wild. In the present study
the gene targets suggested as markers of thermal stress in previous studies showed robust
induction in response to simulated diurnal warming and fluctuations in juveniles, and to natural
thermal variation in wild adults. These finding indicate that the heat shock response has utility as
an indicator of thermal stress for arctic char under current environmental conditions.
Conclusion
Migrating sockeye salmon exposed to warm but environmentally relevant temperatures exhibit
large inductions of gene expression associated with cytoprotection and also exhibit changes in
biochemistry such as elevated plasma lactate and glucose levels that are indicative of increasing
energy supply to meet demand (Jeffries et al. 2012a; Jeffries et al. 2012b). These molecular and
biochemical changes are markers of thermal stress but are also indicative of a only partially

99
successful compensatory response as they coincide with a reduction of aerobic scope, cardiac
output, and in some cases increased incidence of early mortality (Cooke et al. 2006; Farrell et al.
2008b; Jeffries et al. 2012b; Mathes et al. 2010). In parallel to this, in the present study, the
observed metabolite changes following diurnal warming were likely, at least in part, a result of a
response meant to supplement ATP production to meet increased demand at warm temperatures.
The observed changes in transcript profiles should have helped cells maintain homeostasis and
protein functionality. Given the losses in performance observed in chapter 3, these compensatory
responses were at best, partially successful at mitigating thermal stress in arctic char. Rainbow
trout under the same conditions were able to maintain or improve performance suggesting that
their compensatory response was sufficient. Together these findings suggest that under the
extremes of current migratory conditions observed in the Canadian Arctic, arctic char would
likely be at a competitive disadvantage to rainbow trout and other temperate salmonids who’s
ranges are expanding northward as their own habitats become more thermally restrictive.
Acknowledgements
Funding was provided thorough a National Science and Engineering Research Council (NSERC)
Discovery grant to KBT, in kind support from Golder Associates, and grants and awards to
MJHG from NSERC, the National Scientific Training Program, Canadian Circumpolar Institute,
Canadian Society of Zoologists, the Association of Canadian Universities for Northern Studies
and the University of Alberta. I would like to thank the staff of the University of Alberta
Molecular Biology Service Unit and in particular Troy Locke for his assistance and advice
pertaining to the qPCR analysis. I would also like to thank Brian Blunt from Chenomx for his
assistance with the metabolomics analysis.

100
References:
!"##$%&'()*'(+*(,*(+%$&-."'()*(/*()&01$"2"'()*(3*(3$%%24(5(!*(6*(7&442%%'(89:;*(7&<$"=(>&4?(
#2?@24*-(A04$"=(?$=4&#$."B(<&4A$&<(?CD!(42-@."-2-(.E(#>.(&A0%#(F"<.4GH"<G0-("241&(
@.@0%&#$."-(#.(>&4?$"=(&"A(->$??$"=(<G&%%2"=2-*(,(7$-G(I$.%(J;KLMB:;NOPLQ(
A.$B:9*::::RSET*:8NQU*(
I&4#."'()*(D*'(3*(7*(V244$#-(5(,*(W*(I&%%&"#H"2'(:OOL*(+EE2<#-(.E(2X24<$-2(."(@%&-?&(
"."2-#24$E$2A(E&##H(&<$A-(&"A(E422(&?$".(&<$A-($"(!4<#$<(<G&4(KW&%Y2%$"0-(&%@$"0-M*(,.04"&%(.E(
+X@24$?2"#&%(Z..%.=H(8U:KNMB:JNP:JO*(
I24%$'(I*([*'(3*(,*(V$%T24#'(!*(\*(C&%@G'()*(I*(]$24"2H(5(6*(I041G&4A#P/.%?'(89:;*(!<0#2(
[email protected](#.(&(<.??."(-0-@2"A2A(-2A$?2"#(&EE2<#-(#G2(->$??$"=(@24E.4?&"<2(&"A(
@GH-$.%.=H(.E(S0Y2"$%2(-&%?."$A-*(^.?@&4&#$Y2(T$.<G2?$-#4H(&"A(@GH-$.%.=H(6&4#(!'(
3.%2<0%&4(5($"#2=4&#$Y2(@GH-$.%.=H(:UQB:P:9(A.$B:9*:9:QRS*<T@&*89:;*9N*9:N*(
^..12'(W*(,*'(W*(V*(/$"<G'(V*(]*(^4.--$"'(_*(!*(6&##24-."'()*()*(+"=%$-G'(,*(3*(WG4$?@#."'(V*(`*(_*(
)4&&1(5(!*(6*(7&442%%'(899Q*(6GH-$.%.=H(.E($"A$Y$A0&%(%P40"(74&-24(C$Y24(-.<12H2(-&%?."(
KF"<.4GH"<G0-("241&M(-&?@%2A($"(#G2(.<2&"(<.442%-(>$#G(E(A04$"=(-@&>"$"=(
?$=4&#$."*(^&"&A$&"(,.04"&%(.E(7$-G24$2-(&"A(!a0&#$<(W<$2"<2-(QNKUMB:;QOP:;J9(
A.$B:9*::NORE9QP9;8*(
^044$2'(W*'(89::*(]+36+C!]bC+(c(/2&#(WG.<1(64.#2$"-(&"A(]2?@24**(["(+A$#.4P$"P^G$2EB(
!"#G."H'(6*(7*(K2AM(+"<H<%.@2A$&(.E(7$-G(6GH-$.%.=H*(!<&A2?$<(642--'(W&"(_$2=.'(:UN8P:UNU*(

101
_.>A'(d*(d*'(7*(!*()$"=(5(3*(d*(_2""H'(89:L*(]G24?&%(Y&4$&#$."'(#G24?&%(2X#42?2-(&"A(#G2(
@GH-$.%.=$<&%(@24E.4?&"<2(.E($"A$Y$A0&%-*(]G2(,.04"&%(.E(2X@24$?2"#&%(T$.%.=H(8:JK6#(
:8MB:OLQP:OQU(A.$B:9*:8;8RS2T*::;O8Q*(
+%$&-."'(+*(,*'(]*(_*(^%&41'(3*(,*(/&=02'(\*(3*(/&"-."'(Z*(W*(V&%%&=G24'()*(3*(,2EE4$2-'(3*()*(V&%2'(_*(
!*(6&##24-."'(W*(V*(/$"<G(5(!*(6*(7&442%%'(89::*(_$EE242"<2-($"(#G24?&%(#.%24&"<2(&?."=(
-.<12H2(-&%?."(@.@0%&#$."-*(W<$2"<2(NN8KQ98LMB:9OP:8(A.$B:9*::8QR-<$2"<2*::OO:LJ*(
+Y&"-'(]*(V*'(+*(/&??$%%'()*()&01$"2"'(!*(_*(W<G0%e2'(_*(!*(6&##24-."'()*()*(+"=%$-G'(,*(3*(^04#$-(
5()*(3*(3$%%24'(89::*(]4&"-<4$@#.?$<-(.E(2"Y$4."?2"#&%(&<<%$?&#$e&#$."(&"A(-04Y$Y&%($"(>$%A(
&A0%#(6&<$E$<(-.<12H2(-&%?."(KF"<.4GH"<G0-("241&M(A04$"=(-@&>"$"=(?$=4&#$."*(3.%2<0%&4(
2<.%.=H(89K8:MB;;U8PJO(A.$B:9*::::RS*:NQLP8O;f*89::*9L8UQ*X*(
7&442%%'(!*(6*'(899U*(^&4A$.42-@$4&#.4H(@24E.4?&"<2(A04$"=(@4.%."=2A(->$??$"=(#2-#-(>$#G(
-&%?."$A-B(&(@24-@2<#$Y2(."(#2?@24*(2EE2<#-(&"A(@.#2"#$&%(&"&%H#$<&%(@$#E&%%-*(
6G$%.-.@G$<&%(]4&"-&<#$."-(.E(#G2(C.H&%(W.<$2#H(IPI$.%.=$<&%(W<$2"<2-(NQ8K:;JUMB89:UP
89N9(A.$B:9*:9OJR4-#T*899U*8:::*(
7&442%%'(!*(6*'(+*(,*(+%$&-."'(+*(W&"AT%.?(5(]*(_*(^%&41'(899O*(7$-G(<&4A$.42-@$4&#.4H(
@GH-$.%.=H($"(&"(24&(.E(<%$?(<G&"=2*(^&"&A$&"(,.04"&%(.E(Z..%.=H(JUK:9MBJNLPJL:(
A.$B:9*::NORe9OP9O8*(
7&442%%'(!*(6*'(W*(V*(/$"<G'(W*(,*(^..12'(_*(!*(6&##24-."'(V*(]*(^4.--$"'(3*(\&@.$"#2(5(3*(]*(
3&#G2-'(899J*(6&<$E$<(W&%?."($"(/.#(dB(!@@%H$"=(!24.T$<(W<.@2(3.A2%-(&"A(
I$.#2%2?2#4H(#.(642A$<#(#G2(W0<<2--(.E(W@&>"$"=(3$=4&#$."-*(6GH-$.%.=$<&%(&"A(I$.<G2?$<&%(
Z..%.=H(J:KQMBQOUPU9J(A.$B:9*:9JQRLO89LU*(

102
7.>%24'(W*(\*'(_*(/&?$%#."(5(W*(^044$2'(899O*(!(<.?@&4$-."(.E(#G2(G2&#(-G.<1(42-@."-2($"(
S0Y2"$%2(&"A(&A0%#(4&$"T.>(#4.0#(KF"<.4GH"<G0-(?H1$--M(g($?@%$<&#$."-(E.4($"<42&-2A(
#G24?&%(-2"-$#$Y$#H(>$#G(&=2*(^&"&A$&"(,.04"&%(.E(7$-G24$2-(&"A(!a0&#$<(W<$2"<2-(QQK:MBO:P
:99(A.$B:9*::NORE9JP:O8*(
V4$2-G&T24'(3*'([*(/&4A2>$='(b*()420#e24(5(/*PF*(6h4#"24'(:OO;*(6GH-$.%.=$<&%(&"A(?2#&T.%$<(
42-@."-2-(#.([email protected]$&($"($"Y24#2T4-(C2Y$2>-(.E(6GH-$.%.=H'(I$.<G2?$-#4H(&"A(
6G&4?&<.%.=H'(`.%0?2(:8L*(W@4$"=24'(;NP:;U*(
[E#$1&4'(7*([*(5(!*(,*(/$<12H'(89:N*(_.(?$#.<G."A4$&(%$?$#(G.#(E$-G(G2&4#-i(b"A24-#&"A$"=(#G2(
4.%2(.E(?$#.<G."A4$&%(E0"<#$."(>$#G(G2&#(-#42--($"(D.#.%&T40-(<2%$A.#0-*(
[E#$1&4'(7*([*'(,*(C*(3&<_."&%A'(_*(d*(I&124'(V*(3*(C2"-G&>(5(!*(,*(/$<12H'(89:;*(^.0%A(#G24?&%(
-2"-$#$Y$#H(.E(?$#.<G."A4$&(A2#24?$"2(-@2<$2-(A$-#4$T0#$."($"(&(<G&"=$"=(<%$?i(]G2(
,.04"&%(.E(2X@24$?2"#&%(T$.%.=H(8:UK:NMB8N;JP8NLU*(
,&$"'()*(+*(5(!*(6*(7&442%%'(899N*(["E%02"<2(.E(-2&-."&%(#2?@24*(."(#G2(42@2&#(->$??$"=(
@24E.4?&"<2(.E(4&$"T.>(#4.0#(F"<.4GH"<G0-(?H1$--*(,.04"&%(.E(+X@24$?2"#&%(I$.%.=H(
89QK89MBNLQOPNLUO*(
,2EE4$2-'()*(3*'(W*(V*(/$"<G'(+*(V*(3&4#$"-'(]*(_*(^%&41'(!*(V*(\.##.'(_*(!*(6&##24-."'(W*(,*(^..12'(!*(
6*(7&442%%(5()*(3*(3$%%24'(89:8&*(W2X(&"A(@4.X$?$#H(#.([email protected]<#$Y2(?$#H($"E%02"<2(#G2(
-04Y$Y&%'(E$"&%(?&#$."'(&"A(T%..A(@GH-$.%.=H(.E(6&<$E$<(-&%?."(>G2"([email protected](#.(G$=G(
#2?@24*(A04$"=(&(-$?0%A(?$=4&#$."*(6GH-$.%.=$<&%(&"A(T$.<G2?$<&%(e..%.=H(B(6IZ(
JLK:MBQ8PUN(A.$B:9*:9JQRQQNUU9*(

103
,2EE4$2-'()*(3*'(W*(V*(/$"<G'(]*(W$24.<$"-1$'(]*(_*(^%&41'(+*(,*(+%$&-."'(3*(C*(_."&%A-."'(W*(C*(\$'(6*(
6&Y%$A$-(5()*(3*(3$%%24'(89:8T*(^."-2a02"<2-(.E(G$=G(#2?@24*-(&"A(@42?*(
?.4#&%$#H(."(#G2(#4&"-<4$@#.?2(&"A(T%..A(@GH-$.%.=H(.E(>$%A(&A0%#(-.<12H2(-&%?."(
KF"<.4GH"<G0-("241&M*(+<.%.=H(&"A(+Y.%0#$."(8KUMB:U;UP:UQ;(A.$B:9*:998R2<2N*8U;*(
\.2EE%24'(3*(5(V*()4.2?24'(8999*(]G2(?$#.<G."A4$."($"(<2%%(A2&#G(<."#4.%B(<24#&$"#$2-(&"A(
$"<.="$#&*(+X@24$?2"#&%(<2%%(42-2&4<G(8LQK:MB:OP8Q*(
\0"A'(W*(V*'(_*(^&$--$2'(C*(!*(^0"S&1'(3*(3*(`$S&H&"(5(I*(\*(]0E#-'(8998*(]G2(2EE2<#-(.E(
2"Y$4."?2"#&%(G2&#(-#42--(."(G2&#P-G.<1(?CD!(&"A(@4.#2$"(2X@42--$."($"(3$4&?$<G$(
!#%&"#$<(-&%?."(KW&%?.(-&%&4M(@&44*(^&"&A$&"(,.04"&%(.E(7$-G24$2-(&"A(!a0&#$<(W<$2"<2-(
LOKOMB:LLNP:LQ8(A.$B:9*::NORE98P::U*(
3&#G2-'(3*(]*'(W*(V*(/$"<G'(W*(,*(^..12'(V*(]*(^4.--$"'(_*(!*(6&##24-."'(!*(V*(\.##.(5(!*(6*(7&442%%'(
89:9*(+EE2<#(.E(>(#2?@24*'(#$?$"='(@GH-$.%.=$<&%(<."A$#$."'(&"A(%&12(#G24?&%(42E0=$&(
."(?$=4&#$"=(&A0%#(d2&Y24(^4221(-.<12H2(-&%?."(KF"<.4GH"<G0-("241&M*(^&"&A$&"(,.04"&%(
.E(7$-G24$2-(&"A(!a0&#$<(W<$2"<2-(QUK:MBU9PJ;(A.$B:9*::NORE9OP:LJ*(
3.??-2"'(]*(6*'(899;*(W&%?."(-@&>"$"=(?$=4&#$."(&"A(?0-<%2(@4.#2$"(?2#&T.%$-?B(#G2(
!0=0-#()4.=G(@4$"<$@%2(&#(>.41*(^.?@&4&#$Y2(I$.<G2?$-#4H(&"A(6GH-$.%.=H(6&4#(IB(
I$.<G2?$-#4H(&"A(3.%2<0%&4(I$.%.=H(:NOKNMBNJNP;99*(
F%$="HP/jT24#'(/*'(^*(W2"&H'(+*(^*(+"A24-(5(_*(I.$-<%&$4'(89:L*(+EE2<#-(.E(A$2%(#2?@24*(
E%0<#0&#$."(."(#G2(-#&"A&4A(?2#&T.%$<(4(.E(S0Y2"$%2(!#%&"#$<(-&%?."(KW&%?.(-&%&4MB(
["E%02"<2(.E(&<<%$?&#$."(#2?@24*(&"A(@4.Y2"$2"<2*(^&"&A$&"(,.04"&%(.E(7$-G24$2-(&"A(
!a0&#$<(W<$2"<2-KS&M*(

104
6&%?$-&".'(!*(D*'(,*(C*(d$"#."(5(d*(d*(_$<1G.EE'(8999*(]$--02PW@2<$E$<(["A0<#$."(.E(/-@O9(
?CD!(&"A(6%&-?&(^.4#$-.%(C2-@."-2($"(^G$"..1(W&%?."(E.%%.>$"=(/2&#(WG.<1'(W2&>(
^G&%%2"=2'(&"A(/&"A%$"=(^G&%%2"=2*(3&4$"2(T$.#2<G".%.=H(8K;MBN8OPNNJ*(
62""2H'(^*(3*'(V*(d*(D&-G(5(!*()*(V&?@24%'(89:;*(^&4A$.42-@$4&#.4H(42-@."-2-(.E(-2&>P
&<<%$?A(&A0%#(!4<#$<(<G&4(KW&%Y2%$"0-(&%@$"0-M(&"A(!#%&"#$<(-&%?."(KW&%?.(-&%&4M(#.(&"(
&<0#2(#2?@24*($"<42&-2*(^&"&A$&"(,.04"&%(.E(7$-G24$2-(&"A(!a0&#$<(W<$2"<2-B:P:9(
A.$B:9*::NOR<SE&-P89:NP9LQO*(
6h4#"24'(/*(F*(5(C*()"0-#'(899U*(^%$?(<G&"=2(&EE2<#-(?&4$"2(E$-G2-(#G4.0=G(#G2(.XH=2"(
%$?$#&#$."(.E(#G24?&%(#.%24&"<2*(-<$2"<2(N:LKLJ9JMBOLPOU*(
k0$""'(D*(\*'(^*(C*(3<V.>&"'(V*(!*(^..@24'(I*(7*()..@(5(d*(W*(_&Y$A-."'(89::&*([A2"#$E$<&#$."(
.E(=2"2-(&--.<$A(>$#G(G2&#(#.%24&"<2($"(!4<#$<(<G&44([email protected](#.(&<0#2(#G24?&%(-#42--*(
6GH-$.%.=$<&%(=2".?$<-(;NK::MBQJLPQOQ(A.$B:9*::L8R@GH-$.%=2".?$<-*9999J*89::*(
k0$""'(D*(\*'(^*(C*(3<V.>&"'(V*(!*(^..@24'(I*(7*()..@(5(d*(W*(_&Y$A-."'(89::T*(C$T.-.?&%(
=2"2-(&"A(G2&#(-G.<1(@4.#2$"-(&-(@0#&#$Y2(?&4124-(E.4(<G4."$<'(-0T%2#G&%(G2&#(-#42--($"(
!4<#$<(<G&44B(&@@%$<&#$."-(E.4(&a0&<0%#042(&"A(>$%A(E$-G*(6GH-$.%.=$<&%(=2".?$<-(
;NK:JMB:9LQP:9Q;*(
W#..#'(\*(,*'(D*(!*(^&$4"-'(7*(^0%%'(,*(,*(]&H%.4'(,*(_*(,2EE42H'(7*(3.4$"'(,*(d*(3&"A2%?&"'(]*(_*(^%&41(
5(W*(,*(^..12'(89:;*(b-2(.E(@.4#&T%2(T%..A(@GH-$.%.=H(@.$"#P.EP<&42(A2Y$<2-(E.4(T&-$<(&"A(
&@@%$2A(42-2&4<G(."(Y24#2T4-B(&(42Y$2>*(^."-24Y&#$."(6GH-$.%.=H(8K:MB<.09::*(

105
W0&42e'(C*()*(5(]*(6*(3.??-2"'(:OJU*(V%0<."2.=2"2-$-($"(#2%2.-#(E$-G2-*(^&"&A$&"(,.04"&%(.E(
Z..%.=H(QLKJMB:JQOP:JJ8*(
]2&?'(C*(^*'(89:;*(CB(!(%&"=0&=2(&"A(2"Y$4."?2"#(E.4(-#&#$-#$<&%(<.?@0#$"=*(C(7.0"A&#$."(
E.4(W#&#$-#$<&%(^.?@0#$"='(`$2""&'(!0-#4$&'(89:8*([WID(NPO999L:P9UP9*(

106
Tables:
Table 4.1 List of primers sequences for target genes in the qPCR analysis
Target name
Abbr.
Primer sequence
Efficiency (%) Source
Arctic char
Rainbow trout
Elongation factor 1-alpha (Endogenous control)
EF1a F-CCCCTCCAGGACGTTTACAAA R-CACACGGCCCACAGGTACA
94.15 100.86 (Quinn et al. 2011b)
70 kDa Heat Shock Protein HS70 F-AAG ATC AGC GAG GAG GAC AA R-TGC CTG ATC TCC ACA GCA
98.84 85.66 (Quinn et al. 2011b)
Heat Shock Protein 90! HSP90! F-GCG TTG CCC ACC ATT AAC R-AAT GGG TAA CCT GGT CAG TGT C
90.25 102.0 (Quinn et al. 2011b)
Heat Shock Protein 90!-2 HSP90!2
F- GCGTTGCCCACCATTAAC R- AATGGGTAACCTGGTCAGTGTC
98.66 101.34 (Quinn et al. 2011b)
Serpin H1 precursor Serpinh1 F-CTG GGA GGC AAA AAC AAC TG R-TTC CAC CAT TCT TTT CAC CAG
92.28 87.78 (Quinn et al. 2011b)
78 kDa glucose-regulated protein precursor
GRPP F-CGACGGAGAGGACTTTTCAG R-TTGGGGATACGGGTAGAGC
94.45 93.70 (Rise et al. 2004)
Ubiquitin carboxyl- terminal hydrolase 8
Ubq F-GAA ATG TTT GCT GGC AAC G R-CCA TGG AAC AGA GCT ACG ATG
81.8 102.92 (Quinn et al. 2011b)
Caspase 3 Casp3 TTTGGGAGTAGATTGCAGGG TGCACATCCACGATTTGATT
131.24 93.71 (Sánchez et al. 2011)
Caspase 8 Casp8 CAGCATAGAGAAGCAAGGGG TGACTGAGGGGAGCTGAGTT
98.72 83.79 (Sánchez et al. 2011)
Cytochrome c-oxidase Subunit 2
Cyt-C2 F-CGAGCGTGCAGATCTTATAGC R-CTTCTCCGCTGAACAGTTGATG
101.46 83.65 (Jeffries et al. 2012b)

107
Table 4.2 Concentration of select m
etabolites that exhibited substantial differences between diurnal treatm
ent groups or species !!
!"#$%#&#'("&!!
)(%*+,-&$",.$&
!!"#$%&#'!
(')*%)+%,#$!-
+&.,$/!
!!"#$%&#'!
(')*%)+%,#$!-
+&.,$/!
!/(*%*0&0123456732!
47745477!41845469!
!!:42345913:!
;:7325273:!96938547:32!
!12("(3%*0&88036542!
:88305;931!80:3:52936!
!!1:3057!
6;30593:!;1305:38!
4"0($%*0&;07325:1430!
4293854038!1763854163;!
!!8::3456736!
;03854239!4297544:8!
5/.$(6
($0&469325:436!
8463;5893;!8425;:3;!
!!4703;54938!
013858734!4203;5:32!
5/.$(6
%*0&;;2345283;!
;6;395483:!;42385;2!
!!4993954639!
8893451832!8:0305:4!
789",:82",/%*0&
:2:3:51039!;803954:03:!
:9138547!!!
643;5630!442345:93:!
0;3254731!;1,/0.#%*0&
481345639!4;43;5483:!
8:73458031!!!
2;32503:!0:3454134!
418395:138!<0.#%*0&
4643:54;3;!8;73:54;39!
::8325:43;!!!
4863:54131!49;5803:!
8:63251739!=0$'%,*%*0&
66325634!4873;54630!
4143;5:!!!
17395234!92385031!
683954839!>'0*8/(/(*%*0&
4405:32!4:73;54030!
8:73454234!!!
243054:36!60395:38!
4703254738!?0"%*0&
88:3451636!:48325;83:!
;495;:3:!!!
4763154;32!847385:632!
4663:5934!@(."%*0&
;6;395:736!22:3:522!
499;5;01!!!
;0732541636!:4;3:59231!
01854:034!@8",1%*0&
2;305232!093954030!
4083:58734!!!
:03854;!:;3:5134!
9;3854131!A(/%*0&
::73054:38!;87305873;!
;023251;3;!!!
87;3058:38!8293:5;:38!
:175263;!&&
!!!!
!!!!
!!!!
!!?.##%*($0&
403454831!41345:30!
88452:39!!!
413;5;30!4;315;3:!
6654;!&&
!!!!
!!!!
!!!!
!!5/.#,10&
63:5438!03;543;!
873:5431!!!
63:5730!49385:32!
41315836!<(#$($0&
48315437!49325431!
8:325431!!!
413:5:496!42345;3:!
873;5730!&
<+')=>!&=?&=>=$%!.=+$!5!>%+$@+&@!=&&#&A!B
C!:!?=&!/&#)?!&
D''!*#$*=$%&+%,#$>!+&=!,$!EF!=G*=?%!H#&!I
')*#>=!+$@!J+*%+%=!KL,*L!+&=!,$!.
F!

108
Figures:
Fig. 4.1 Principle components analysis of plasma metabolite profiles in arctic char and rainbow
trout exposed to simulated diurnal warming and fluctuations. Ellipses represent 68% confidence
limits for each treatment group.
Fig. 4.2 Heat map showing the fold change in concentration of specific metabolites in the plasma
of arctic char and rainbow trout following diurnal temperature treatments
Fig. 4.3 Principle components analysis of transcript profiles in arctic char and rainbow trout
exposed to simulated diurnal warming and fluctuations. Ellipses represent 68% confidence limits
for each treatment group.
Fig. 4.4 Relative quantitation of target gene expression following diurnal temperature treatments
in lab reared juvenile arctic char and rainbow trout. Dissimilar letters indicate significant
differences between treatment groups for a given target (!=0.05) and error bars represent
standard error of the mean.
Fig. 4.5 Relative quantitation of target gene expression following natural exposure to warm
(>19°C) or cool (<15°C) water in migrating adult arctic char. Asterisks indicate significant
differences between warm and cool groups (!=0.05, * <0.01, **<0.001) and error bars represent
standard error.

109

110

111

112

113

114
Chapter 5: General Conclusions
Summary of major findings
The unifying goal throughout my research was to identify characteristics of arctic char migratory
life history and physiology that will shape their ability to cope with global climate change
(GCC). I focused my research on the Nulahugyuk Creek arctic char migration, which was
traditionally a vital Inuit fishery (Jenness 1922). Nulahugyuk Creek is generally shallow and
experiences a rapid decline in discharge and large diurnal temperature fluctuations throughout
the course of the migration. These conditions frequently lead to adult char becoming stranded in
shallow water and juveniles and adults having to migrate through warm, variable water
temperatures. The harsh nature of flow and thermal regimes in the creek allowed for the study of
the life-history and physiology of arctic char under conditions represent the current extremes
under which char migrate, but will encounter more frequently as GCC progresses.
Previous researchers have suggested that extensive life history diversity within char facilitates
their existence in other extreme environments (Hammar 2014; Klemetsen 2010). In the present
thesis I expanded our understanding of this diversity by identifying novel life history strategies
that facilitate the persistence of arctic char in Nulahugyuk Creek. Specifically, these arctic char
possess the lowest observed annual fidelity to their natal system (Gyselman 1994; Moore et al.
2013), and once smolts leave on their first migration they typically do not return for four to five
years. Furthermore the upstream migration from the ocean to their spawning habitat is the
earliest documented return run in northern Canada. These previously un-described life history
strategies suggest that other populations of arctic char may possess the variation needed to adapt
to more restrictive flows and temperatures in the future. Alternatively, if changes occur too

115
rapidly to permit local adaptation, populations already possessing these unique life history traits
may be able to serve as source populations as other systems become uninhabitable to their
current residents.
The alternative life history traits identified certainly lessen the exposure of Nulahugyuk Creek
arctic char to even worse conditions, however the conditions they do face still present significant
physiological challenges. I found that current diurnal temperature variation in Nulahugyuk Creek
already appears to be sufficient enough to impair recovery from fatiguing exercise. This impaired
recovery is likely a result of an inability to sufficiently increase cellular energy supply to meet
increased demand at warm temperatures, despite the mobilization of significant energy reserved
including glucose and numerous amino acids, and despite large increases in anaerobic
metabolism. The general loss of performance in arctic char coincided with a potential reduction
in mitochondrial functionality and a large compensatory transcriptional heat shock response
response that was evidently insufficient. In the wild this impaired physiological performance and
inability to recover from fatiguing exercise could result in longer recovery times at strenuous
points in the migration such as in high flows or when fish become stranded. Furthermore, the
exploitation of various energy reserves at warm temperatures (e.g. glycogen, protein) may
deplete the energy available for growth or reproductive maturation (Mommsen 2004). Together
these performance and energy losses could result in increased predator related mortality, reduced
migratory success in general, and a reduction in growth or fecundity. In our comparative studies,
relative to rainbow trout, arctic char exhibited a greater and prolonged disturbance to thermal
variation representative of their current migratory conditions. This suggests that they may
already be at a competitive disadvantage to more temperate salmonids whose ranges are
expanding in the north. These findings are of particular concern, as the environmental conditions

116
studied will only become more common or worsen in the future (ACIA 2005; Reist et al. 2006b;
Wrona et al. 2006b).
Future directions
In the present study I was able to identify population specific migratory life history traits that
allow Nulahugyuk Creek arctic char to persist in an extreme environment by comparing their life
histories to other previously studied populations. At the physiological level we only investigated
the performance of one population of wild char and one strain of captive char and there were no
other studies similar enough to begin addressing the extent to which thermal sensitivities of
arctic char vary at the local environment or population level. There is well-established local
adaptation in the migratory physiology of different populations of sockeye salmon that
corresponds with the physical difficulty of their respective migrations and their thermal
environments (Eliason et al. 2011). While the wild juvenile char in the present study were, able
to maintain performance at least up to 20°C other populations that do not currently experience
these conditions, or have as physically challenging migration may not have that ability and may
therefore be susceptible to change. In the present study and those in more temperate species,
cardiorespiratory or mitochondrial limitations appear to be responsible for performance losses at
warm temperatures (Eliason et al. 2011; Farrell 2009b; Farrell et al. 2009; Schulte 2015). As
such future studies in anadromous arctic char should investigate population specific differences
in cardiac and mitochondrial physiology that may underlie differences in thermal sensitivity.
The current study was also not designed to assess the range of individual variation in thermal
tolerance that exists within a given population. Quinn et. al (Quinn et al. 2011a) demonstrated
that an aquaculture strain of arctic char possessed substantial individual variation in thermal

117
tolerance, potentially based on individuals ability to induce a heat shock response. As heritable
variation within a trait is required for adaptation to occur, future studies in wild arctic char
should assess the extent of existing variation in thermal tolerance as well as its heritability to
determine potential for adaptation.

118
References:
!"#!$%&''()%!*+,-+%+.-/0,1%-/20+,%033133/14,53+-14,-6-+%*127*,)%"0/8*-9:1%;4-<1*3-,=%>*133)%
?.-0374$%?)%@)$%A)%B)%".0*C$%D)%@)%E0:F1$%G)%D)%E04374$%H)%I)%J0..0:K1*$%L)%D)%@166*-13$%D)%L)%J0.1$%B)%
!)%>0,,1*374$%I)%J)%E-4+K%M%!)%>)%N0**1..$%&'OO)%B-661*14+13%-4%,K1*/0.%,7.1*04+1%0/74:%
37+C1=1%30./74%272F.0,-743)%I+-14+1%PP&QR'&(STO'U5O&%97-TO')OO&RV3+-14+1)OOUUO(W)%
N0**1..$%!)%>)$%&''U)%?4<-*74/14,$%04,1+1914,3%049%+.-/0,1%+K04:1T%.133743%6*7/%,K1%3,F9=%
76%,1/21*0,F*1%2K=3-7.7:=%049%*-<1*%/-:*0,-74%76%30./74-93)%AK1%@7F*40.%76%1X21*-/14,0.%
8-7.7:=%&O&Q>,%&PSTPYYO5W'%97-TO')O&Z&V[18)'&PRYO)%
N0**1..$%!)%>)$%?)%@)%?.-0374$%?)%I0498.7/%M%A)%B)%".0*C$%&''U)%N-3K%+0*9-7*132-*0,7*=%
2K=3-7.7:=%-4%04%1*0%76%+.-/0,1%+K04:1)%"0409-04%@7F*40.%76%H77.7:=%WYQO'STWP(5W(O%
97-TO')OOPUV\'U5'U&)%
J=31./04$%?)%")$%OUUZ)%N-91.-,=%76%0409*7/7F3%!*+,-+%"K0*%QI0.<1.-4F3%0.2-4F3S%,7%]0F=FC%
G0C1$%])^)A)$%"04090)%"0409-04%@7F*40.%76%N-3K1*-13%049%!_F0,-+%I+-14+13%(OTOU&Y5OUPZ)%
E0//0*$%@)$%&'OZ)%]0,F*0.%*13-.-14+1%-4%!*+,-+%+K0**%I0.<1.-4F3%0.2-4F3T%.-61%K-3,7*=$%320,-0.%
049%9-1,0*=%0.,1*0,-743%0.74:%:*09-14,3%76%-4,1*321+-6-+%-4,1*0+,-743)%@%N-3K%`-7.%W(QOSTWO5
OOW%97-TO')OOOOV[68)O&P&O)%
@144133$%B)$%OU&&)%AK1%G-61%76%,K1%"7221*%?3C-/73$%<7.%a7.F/1%b##T%AK1%"7221*%?3C-/73)%N!%
!+C.049)%

119
L.1/1,314$%!)$%&'O')%AK1%"K0**%>*78.1/%c1<-3-,19T%?X+12,-740.%>K147,=2-+%>.03,-+-,=%
>*7/7,13%?+7.7:-+0.%I21+-0,-74%-4%>73,:.0+-0.%G0C13)%N*13Kd0,1*%c1<-1d3%PQOSTZU5YZ%
97-TO')OR'WV6*[5P)O)P)%
D7//314$%A)%>)$%&''Z)%I0./74%320d4-4:%/-:*0,-74%049%/F3+.1%2*7,1-4%/1,087.-3/T%,K1%
!F:F3,%L*7:K%2*-4+-2.1%0,%d7*C)%"7/20*0,-<1%`-7+K1/-3,*=%049%>K=3-7.7:=%>0*,%`T%
`-7+K1/-3,*=%049%D7.1+F.0*%`-7.7:=%OPUQPSTPWP5Z'')%
D77*1$%@)5I)$%G)%])%E0**-3$%c)%N)%A0../04$%?)%`)%A0=.7*%M%>)%D7*e4$%&'OP)%AK1%-4,1*2.0=%81,d114%
9-321*30.%049%:141%6.7d%-4%0409*7/7F3%!*+,-+%+K0*%QI0.<1.-4F3%0.2-4F3ST%-/2.-+0,-743%67*%
27,14,-0.%67*%.7+0.%0902,0,-74)%"0409-04%@7F*40.%76%N-3K1*-13%049%!_F0,-+%I+-14+13%
Y'QUSTOP&Y5OPPW%97-TO')OOPUV+[6035&'OP5'OPW)%
fF-44$%])%G)$%")%c)%D+J7d04$%J)%!)%"7721*$%`)%N)%L772%M%^)%I)%B0<-9374$%&'OO)%#914,-6-+0,-74%
76%:1413%0337+-0,19%d-,K%K10,%,7.1*04+1%-4%!*+,-+%+K0**%1X27319%,7%0+F,1%,K1*/0.%3,*133)%
>K=3-7.7:-+0.%:147/-+3%ZPQOOSTRW(5RUR%97-TO')OO(&V2K=3-7.:147/-+3)''''W)&'OO)%
c1-3,$%@)%B)$%N)%@)%^*740$%A)%B)%>*7d31$%D)%>7d1*$%@)%`)%B1/2374$%@)%c)%L-4:%M%c)%@)%`10/-3K$%
&''R)%!4%7<1*<-1d%76%1661+,3%76%+.-/0,1%+K04:1%74%31.1+,19%!*+,-+%6*13Kd0,1*%049%
0409*7/7F3%6-3K13)%!D`#gT%!%@7F*40.%76%,K1%EF/04%?4<-*74/14,%P(QYSTPWO5PWY)%
I+KF.,1$%>)%D)$%&'O()%AK1%1661+,3%76%,1/21*0,F*1%74%01*78-+%/1,087.-3/T%,7d0*93%0%
/1+K04-3,-+%F491*3,049-4:%76%,K1%*13274313%76%1+,7,K1*/3%,7%0%+K04:-4:%14<-*74/14,)%AK1%
@7F*40.%76%1X21*-/14,0.%8-7.7:=%&OWQ>,%O&STOW(R5OWRR%97-TO')O&Z&V[18)OOWW(O)%

120
^*740$%N)%@)$%A)%B)%>*7d31$%@)%B)%c1-3,$%@)%?)%E788-1$%G)%D)%@)%Gh<13_F1%M%^)%N)%a-4+14,$%&''R)%
".-/0,1%#/20+,3%74%!*+,-+%N*13Kd0,1*%?+73=3,1/3%049%N-3K1*-13T%`0+C:*7F49$%c0,-740.1%
049%!22*70+K%76%,K1%!*+,-+%".-/0,1%#/20+,%!33133/14,%Q!"#!S)%!D`#gT%!%@7F*40.%76%,K1%
EF/04%?4<-*74/14,%P(QYSTP&R5P&U%97-TO')O(YUV''ZZ5YZZYQ&''RSP(iP&RT+-7061j&)')+7k&)%

121
Bibliography:
!"#!$%&''()%!*+,-+%+.-/0,1%-/20+,%033133/14,53+-14,-6-+%*127*,)%"0/8*-9:1%;4-<1*3-,=%>*133)%
!.372$%B)%M%")%^779$%OUUY)%AK1%-4,1*0+,-<1%1661+,3%76%6119-4:%049%1X1*+-31%74%7X=:14%
+743F/2,-74$%3d-//-4:%21*67*/04+1%049%2*7,1-4%F30:1%-4%[F<14-.1%*0-487d%,*7F,%
Qg4+7*K=4+KF3%/=C-33S)%AK1%@7F*40.%76%1X21*-/14,0.%8-7.7:=%&''QOYST&PPY5&PZR)%
!4,,-.0$%L)$%?)%@)%?.-0374$%L)%E)%L0FC-414$%L)%D)%D-..1*%M%!)%>)%N0**1..$%&'OZ)%N0+-4:%d0*/%
,1/21*0,F*13%9F*-4:%/-:*0,-74T%+0*9-0+%/c]!%*13274313%76%,d7%09F.,%g4+7*K=4+KF3%41*C0%
272F.0,-743%,7%d0*/-4:%049%3d-//-4:%+K0..14:13)%@%N-3K%`-7.%WZQ(STOZPU5(R%
97-TO')OOOOV[68)O&PRY)%
!*/3,*74:$%c)%E)%M%@)%D7**7d$%OUW')%AK1%97..=%<0*914)%"K0**3T%30./74-9%6-3K13%76%,K1%:14F3%
I0.<1.-4F3TUU5OZ')%
`0*,74$%L)%])$%D)%N)%J1**-,3%M%@)%I)%`0..04,=41$%OUU()%?661+,3%76%1X1*+-31%74%2.03/0%
47413,1*-6-19%60,,=%0+-93%049%6*11%0/-47%0+-93%-4%!*+,-+%+K0*%QI0.<1.-4F3%0.2-4F3S)%@7F*40.%76%
?X21*-/14,0.%H77.7:=%&YOQPSTOWP5OWU)%
`0,13$%B)$%D)%D01+K.1*$%`)%`7.C1*%M%I)%^0.C1*$%&'OP)%./1ZT%G-410*%/-X1951661+,3%/791.3%
F3-4:%?-:14%049%IZ)%c%20+C0:1%<1*3-74%OQZS)%
`10/-3K$%N)%^)$%OUW')%Id-//-4:%21*67*/04+1%049%7X=:14%+743F/2,-74%76%,K1%+K0**3)%
"K0**3$%I0./74-9%N-3K13%76%,K1%J14F3%I0.<1.-4F3TYPU)%

122
`11+K-1$%A)$%?)%`FK.1$%D)%cF+C1.3K0F3$%!)%NF..1*,74%M%G)%E7.3-4:1*$%&''R)%E=9*7.7:-+%*1:-/1%
049%,K1%+7431*<0,-74%76%30./74%.-61%K-3,7*=%9-<1*3-,=)%`-7.7:-+0.%"7431*<0,-74%OP'QZST(R'5
(Y&%97-TO')O'ORV[)8-7+74)&''R)'O)'OU)%
`1*.-$%`)%#)$%D)%@)%J-.81*,$%!)%G)%c0.2K$%L)%`)%A-1*41=%M%>)%`F*CK0*9,5E7./$%&'OZ)%!+F,1%
1X273F*1%,7%0%+7//74%3F3214919%319-/14,%0661+,3%,K1%3d-//-4:%21*67*/04+1%049%
2K=3-7.7:=%76%[F<14-.1%30./74-93)%"7/20*0,-<1%8-7+K1/-3,*=%049%2K=3-7.7:=%>0*,%!$%
D7.1+F.0*%M%-4,1:*0,-<1%2K=3-7.7:=%OYRTO5O'%97-TO')O'ORV[)+820)&'OZ)'P)'OP)%
`*1,,$%@)$%OURZ)%AK1%*132-*0,7*=%/1,087.-3/%049%3d-//-4:%21*67*/04+1%76%=7F4:%37+C1=1%
30./74)%@7F*40.%76%,K1%N-3K1*-13%`70*9%76%"04090%&OQ(STOOWP5O&&R)%
"040<1..7$%>)%c)$%@)%D)%"0+K0,$%?)%")%`11374$%!)%G)%G066774$%")%J*-/13$%^)%!)%E0=/7*1$%D)%N)%
?.1:04,1$%`)%L)%`0*,1.3$%>)%")%E0*,%M%I)%#)%?.CK0=0,$%&'OO)%D103F*-4:%1497+*-41%Q+7*,-37.S%
*13274313%76%\18*06-3K%,7%3,*133%H18*06-3K%41F*781K0<-7*0.%2*7,7+7.3)%I2*-4:1*$%OP(5OZ&)%
"0*.-41$%c)%N)%M%@)%N)%D0+KF4:$%&''O)%"*-,-+0.%AK1*/0.%D0X-/0%76%^-.9%049%B7/13,-+%I,*0-43%
76%A*7F,)%A*0430+,-743%76%,K1%!/1*-+04%N-3K1*-13%I7+-1,=%OP'QRSTO&OO5O&OR%
97-TO')O(YYVO(ZW5WR(UQ&''OSOP'lO&OOT+,/7d0m&)')+7k&)%
".0*C$%A)%B)$%D)%c)%B740.9374$%I)%>-121*K766$%I)%D)%B*1441*$%!)%G7,,7$%I)%@)%"77C1$%I)%J)%E-4+K$%B)%
!)%>0,,1*374%M%!)%>)%N0**1..$%&'O&)%>K=3-7.7:-+0.%`1416-,3%76%`1-4:%I/0..%-4%0%"K04:-4:%
^7*.9T%c13274313%76%"7K7%I0./74%Qg4+7*K=4+KF3%C-3F,+KS%,7%04%!+F,1%AK1*/0.%"K0..14:1%
049%0%I-/F.0,19%"02,F*1%?<14,)%>.7I%741%YQRST1PU'YU$%O5W)%

123
".0*C$%A)%B)$%L)%D)%@166*-13$%I)%J)%E-4+K%M%!)%>)%N0**1..$%&'OO)%?X+12,-740.%01*78-+%3+721%049%
+0*9-7<03+F.0*%21*67*/04+1%76%2-4C%30./74%Qg4+7*K=4+KF3%:7*8F3+K0S%/0=%F491*.-1%
*13-.-14+1%-4%0%d0*/-4:%+.-/0,1)%@7F*40.%76%?X21*-/14,0.%`-7.7:=%&OZQOWSTP'YZ5P'WO%
97-TO')O&Z&V[18)'R'(OY)%
".0*C$%A)%B)$%?)%I0498.7/$%J)%L)%"7X$%I)%J)%E-4+K%M%!)%>)%N0**1..$%&''W)%"-*+F.0,7*=%.-/-,3%,7%
7X=:14%3F22.=%9F*-4:%04%0+F,1%,1/21*0,F*1%-4+*1031%-4%,K1%"K-477C%30./74%Qg4+7*K=4+KF3%
,3K0d=,3+K0S)%!/1*-+04%@7F*40.%76%>K=3-7.7:=5c1:F.0,7*=%#4,1:*0,-<1%049%"7/20*0,-<1%
>K=3-7.7:=%&U(Q(STcORPO5cORPU%97-TO')OO(&V0[2*1:F)U'ZRO)&''W)%
"77C1$%I)%@)$%I)%J)%E-4+K$%J)%A)%"*733-4$%B)%!)%>0,,1*374$%L)%L)%?4:.-3K$%@)%D)%IK*-/2,74$%J)%a)%B)%
L*00C%M%!)%>)%N0**1..$%&''R)%>K=3-7.7:=%76%-49-<-9F0.%.0,15*F4%N*031*%c-<1*%37+C1=1%30./74%
Qg4+7*K=4+KF3%41*C0S%30/2.19%-4%,K1%7+104%+7**1.0,13%d-,K%60,1%9F*-4:%320d4-4:%
/-:*0,-74)%"0409-04%@7F*40.%76%N-3K1*-13%049%!_F0,-+%I+-14+13%RPQYSTOZRU5OZW'%
97-TO')OOPUV6'R5'Z&)%
"F**-1$%I)$%&'OO)%A?D>?c!A;c?%n%E10,%IK7+C%>*7,1-43%049%A1/21*0,F*1)%#4%?9-,7*5-45"K-16T%
!4,K74=$%>)%N)%Q19S%?4+=+.7219-0%76%N-3K%>K=3-7.7:=)%!+091/-+%>*133$%I04%B-1:7$%OYP&5OYPY)%
B1/2374$%@)%M%@)%J*114$%OUW()%G-61%K-3,7*=%76%0409*7/7F3%0*+,-+%+K0**$%I0.<1.-4F3%0.2-4F3$%-4%
,K1%N*031*%c-<1*$%47*,K1*4%G08*097*)%"0409-04%@7F*40.%76%H77.7:=%RPQ&STPO(5P&Z)%
B7d9$%^)%^)$%N)%!)%L-4:%M%D)%^)%B144=$%&'O()%AK1*/0.%<0*-0,-74$%,K1*/0.%1X,*1/13%049%,K1%
2K=3-7.7:-+0.%21*67*/04+1%76%-49-<-9F0.3)%AK1%@7F*40.%76%1X21*-/14,0.%8-7.7:=%&OWQ>,%
O&STOU(R5OURY%97-TO')O&Z&V[18)OOZU&R)%

124
BF4/0..$%L)$%@)%c1-3,$%?)%"0*/0+C$%@)%`080.FC$%D)%>)%E1-915@o*:14314%M%D)%B7+C1*$%>0+-6-+%
I0./74%-4%,K1%!*+,-+T%E0*8-4:1*3%76%"K04:1)%#4T%c13274313%76%!*+,-+%D0*-41%?+73=3,1/3%,7%
".-/0,1%"K04:1$%&'O&)%!.03C0%I10%J*04,$%;4-<1*3-,=%76%!.03C0%N0-*804C3$%2%OZO5ORP)%
BF,-.$%@)%B)$%OUWR)%?41*:1,-+%"743,*0-4,3%049%I20d4-4:%#4,1*<0.%-4%,K1%!409*7/7F3%!*+,-+%
"K0**%QI0.<1.-4F3%0.2-4F3S)%"721-0%OUWRQZSTUZ(5U(()%
?.-0374$%?)%@)$%A)%B)%".0*C$%D)%@)%E0:F1$%G)%D)%E04374$%H)%I)%J0..0:K1*$%L)%D)%@166*-13$%D)%L)%J0.1$%B)%
!)%>0,,1*374$%I)%J)%E-4+K%M%!)%>)%N0**1..$%&'OO)%B-661*14+13%-4%,K1*/0.%,7.1*04+1%0/74:%
37+C1=1%30./74%272F.0,-743)%I+-14+1%PP&QR'&(STO'U5O&%97-TO')OO&RV3+-14+1)OOUUO(W)%
?..-7,,$%@)%M%@)%?..-7,,$%&'O')%A1/21*0,F*1%*1_F-*1/14,3%76%!,.04,-+%30./74%I0./7%30.0*$%
8*7d4%,*7F,%I0./7%,*F,,0%049%!*+,-+%+K0**%I0.<1.-4F3%0.2-4F3T%2*19-+,-4:%,K1%1661+,3%76%
+.-/0,1%+K04:1)%@7F*40.%76%6-3K%8-7.7:=%YYQWSTOYUP5OWOY)%
?<043$%A)%J)$%?)%E0//-..$%L)%L0FC-414$%!)%B)%I+KF.\1$%B)%!)%>0,,1*374$%L)%L)%?4:.-3K$%@)%D)%"F*,-3%
M%L)%D)%D-..1*$%&'OO)%A*043+*-2,7/-+3%76%14<-*74/14,0.%0++.-/0,-\0,-74%049%3F*<-<0.%-4%d-.9%
09F.,%>0+-6-+%37+C1=1%30./74%Qg4+7*K=4+KF3%41*C0S%9F*-4:%320d4-4:%/-:*0,-74)%D7.1+F.0*%
1+7.7:=%&'Q&OSTZZY&5WU%97-TO')OOOOV[)OPR(5&UZb)&'OO)'(&YR)X)%
N0**1..$%!)$%&''U0)%?4<-*74/14,$%04,1+1914,3%049%+.-/0,1%+K04:1T%.133743%6*7/%,K1%3,F9=%76%
,1/21*0,F*1%2K=3-7.7:=%049%*-<1*%/-:*0,-74%76%30./74-93)%@7F*40.%76%?X21*-/14,0.%`-7.7:=%
&O&Q&PSTPYYO5PYW')%

125
N0**1..$%!)$%I)%E-4+K$%I)%"77C1$%B)%>0,,1*374$%J)%"*733-4$%D)%G027-4,1%M%D)%D0,K13$%&''W0)%
>0+-6-+%30./74%-4%K7,%d0,1*T%022.=-4:%01*78-+%3+721%/791.3%049%8-7,1.1/1,*=%,7%2*19-+,%,K1%
3F++133%76%320d4-4:%/-:*0,-743)%>K=3-7.7:-+0.%049%`-7+K1/-+0.%H77.7:=%WOQRSTRUY5Y'U)%
N0**1..$%!)$%")%G11$%L)%A-1*41=$%!)%E790.=$%I)%".F,,1*K0/$%D)%E10.1=$%I)%E-4+K%M%!)%G7,,7$%&''P)%
N-1.9‐80319%/103F*1/14,3%76%7X=:14%F2,0C1%049%3d-//-4:%21*67*/04+1%d-,K%09F.,%>0+-6-+%
30./74%F3-4:%0%/78-.1%*132-*7/1,1*%3d-/%,F441.)%@7F*40.%76%N-3K%`-7.7:=%R&QOSTRZ5WZ)%
N0**1..$%!)%>)$%&''Y)%"0*9-7*132-*0,7*=%21*67*/04+1%9F*-4:%2*7.74:19%3d-//-4:%,13,3%d-,K%
30./74-93T%0%21*321+,-<1%74%,1/21*0,F*1%1661+,3%049%27,14,-0.%040.=,-+0.%2-,60..3)%
>K-.7372K-+0.%A*0430+,-743%76%,K1%c7=0.%I7+-1,=%`5`-7.7:-+0.%I+-14+13%PR&QOZWYST&'OY5
&'P'%97-TO')O'UWV*3,8)&''Y)&OOO)%
N0**1..$%!)%>)$%&''U8)%?4<-*74/14,$%04,1+1914,3%049%+.-/0,1%+K04:1T%.133743%6*7/%,K1%3,F9=%
76%,1/21*0,F*1%2K=3-7.7:=%049%*-<1*%/-:*0,-74%76%30./74-93)%AK1%@7F*40.%76%1X21*-/14,0.%
8-7.7:=%&O&Q>,%&PSTPYYO5W'%97-TO')O&Z&V[18)'&PRYO)%
N0**1..$%!)%>)$%?)%@)%?.-0374$%?)%I0498.7/%M%A)%B)%".0*C$%&''U)%N-3K%+0*9-7*132-*0,7*=%
2K=3-7.7:=%-4%04%1*0%76%+.-/0,1%+K04:1)%"0409-04%@7F*40.%76%H77.7:=%WYQO'STWP(5W(O%
97-TO')OOPUV\'U5'U&)%
N0**1..$%!)%>)$%I)%J)%E-4+K$%I)%@)%"77C1$%B)%!)%>0,,1*374$%J)%A)%"*733-4$%D)%G027-4,1%M%D)%A)%
D0,K13$%&''W8)%>0+-6-+%I0./74%-4%E7,%^0,1*T%!22.=-4:%!1*78-+%I+721%D791.3%049%
`-7,1.1/1,*=%,7%>*19-+,%,K1%IF++133%76%I20d4-4:%D-:*0,-743)%>K=3-7.7:-+0.%049%`-7+K1/-+0.%
H77.7:=%WOQRSTRUY5Y'W%97-TO')O'WRV(U&'(Y)%

126
N7d.1*$%I)%G)$%B)%E0/-.,74%M%I)%"F**-1$%&''U)%!%+7/20*-374%76%,K1%K10,%3K7+C%*1327431%-4%
[F<14-.1%049%09F.,%*0-487d%,*7F,%Qg4+7*K=4+KF3%/=C-33S%p%-/2.-+0,-743%67*%-4+*10319%
,K1*/0.%3143-,-<-,=%d-,K%0:1)%"0409-04%@7F*40.%76%N-3K1*-13%049%!_F0,-+%I+-14+13%RRQOSTUO5
O''%97-TO')OOPUV6'W5OU&)%
N*-19.049$%L)%B)$%c)%a)%^0.C1*$%])%B)%B0<-3$%L)%^)%D=1*3$%J)%^)%`71K.1*,$%I)%;*0d0%M%q)%;147$%
&''O)%g21457+104%7*-14,0,-74%049%*1,F*4%/-:*0,-74%*7F,13%76%+KF/%30./74%80319%74%
,1/21*0,F*1%90,0%6*7/%90,0%3,7*0:1%,0:3)%D0*-41%?+7.7:=%>*7:*133%I1*-13%&ORT&P(5&(&)%
J0.8*10,K$%>)%N)$%])%B)%!90/3%M%A)%E)%D0*,-4$%&''Z)%#46.F14+1%76%K10,-4:%*0,1%74%/103F*1/14,%
76%,-/1%,7%,K1*/0.%/0X-/F/%-4%,*7F,)%!_F0+F.,F*1%&ZOQO5ZST(WY5(UU%
97-TO')O'ORV[)0_F0+F.,F*1)&''Z)'W)''Z)%
J0/21*.$%!)%L)$%L)%@)%c794-+C$%E)%!)%N0F3,$%?)%")%a144$%D)%A)%`1441,,$%G)%#)%"*0d3K0d$%?)%c)%
L11.1=$%D)%I)%>7d1..%M%E)%^)%G-$%&''&)%D1,087.-3/$%3d-//-4:%21*67*/04+1$%049%,-33F1%
8-7+K1/-3,*=%76%K-:K%9131*,%*198049%,*7F,%Qg4+7*K=4+KF3%/=C-33%332)ST%1<-914+1%67*%
2K147,=2-+%9-661*14+13%-4%2K=3-7.7:-+0.%6F4+,-74)%>K=3-7.7:-+0.%049%`-7+K1/-+0.%H77.7:=%
Y(Q(STZOP5ZPO)%
J-../04$%B)%a)%M%!)%E)%L*-3,7661*374$%OUWZ)%`-7.7:-+0.%90,0%74%!*+,-+%"K0**$%I0.<1.-4F3%0.2-4F3%
QG)S$%6*7/%,K1%"7221*/-41%c-<1*$%]7*,Kd13,%A1**-,7*-13$%OUWO5W&)%"0409-04%B0,0%c127*,%76%
N-3K1*-13%049%!_F0,-+%I+-14+13%ZZ'T-<%r%OR%2)%
J-../04$%B)%a)%M%>)%B)%I20*.-4:$%OUW()%`-7.7:-+0.%90,0%74%0*+,-+%+K0**$%30.<1.-4F3%0.2-43%QG)S$%
6*7/%,K1%c0,%c-<1*$%]7*,Kd13,%A1**-,7*-13$%OUWP)%"0409-04%B0,0%c127*,%76%N-3K1*-13%049%
!_F0,-+%I+-14+13%(P(T-<%r%O(%2)%

127
J.7<0$%J)%M%>)%D+"0*,$%OUYZ)%G-61%K-3,7*=%76%!*+,-+%+K0*%QI0.<1.-4F3%0.2-4F3S%-4%,K1%N-*,K%
c-<1*$%qFC74%A1**-,7*=)%G-61%K-3,7*-13%76%0409*7/7F3%049%6*13Kd0,1*%6-3K%-4%,K1%d13,1*4%
!*+,-+%`-7.7:-+%c12%I1*%&')%
J*-13K081*$%D)$%#)%E0*91d-:$%;)%L*1F,\1*%M%E)5g)%>s*,41*$%OUUZ)%>K=3-7.7:-+0.%049%/1,087.-+%
*13274313%,7%K=27X-0%-4%-4<1*,18*0,13%c1<-1d3%76%>K=3-7.7:=$%`-7+K1/-3,*=%049%
>K0*/0+7.7:=$%a7.F/1%O&()%I2*-4:1*$%ZP5OZY)%
JF.31,K$%g)%!)%M%L)%@)%]-.3314$%&''')%AK1%`*-16%>1*-79%76%I2*-4:%D-:*0,-74$%IK7*,%D0*-41%
c13-914+1$%049%E-:K%c1,F*4%c0,1%76%0%]7*,K1*4%I<0.80*9%>72F.0,-74%76%!*+,-+%"K0*)%
A*0430+,-743%76%,K1%!/1*-+04%N-3K1*-13%I7+-1,=%O&UQPSTYW&5YUR%97-TO')O(YYVO(ZW5
WR(UQ&'''SO&Ul'YW&T,8273/m&)P)+7k&)%
J=31./04$%?)%")$%AK1%3103740.%/7<1/14,3%76%0409*7/7F3%!*+,-+%+K0**%0,%]0F=FC%G0C1$%
]7*,Kd13,%A1**-,7*-13$%"04090)%#4T%`-7.7:=%76%,K1%!*+,-+%+K0**$%>*7+119-4:3%76%,K1%
#4,1*40,-740.%I=/273-F/%74%!*+,-+%"K0**%;4-<1*3-,=%76%D04-,780%>*133$%^-44-21:$%
D04-,780$%OUWZ)%2%(Y(5(YW)%
J=31./04$%?)%")$%OUUZ)%N-91.-,=%76%0409*7/7F3%!*+,-+%"K0*%QI0.<1.-4F3%0.2-4F3S%,7%]0F=FC%
G0C1$%])^)A)$%"04090)%"0409-04%@7F*40.%76%N-3K1*-13%049%!_F0,-+%I+-14+13%(OTOU&Y5OUPZ)%
E0//0*$%@)$%&'OZ)%]0,F*0.%*13-.-14+1%-4%!*+,-+%+K0**%I0.<1.-4F3%0.2-4F3T%.-61%K-3,7*=$%320,-0.%
049%9-1,0*=%0.,1*0,-743%0.74:%:*09-14,3%76%-4,1*321+-6-+%-4,1*0+,-74%
3)%@%N-3K%`-7.%W(QOSTWO5OOW%97-TO')OOOOV[68)O&P&O)%

128
E-.87*4$%c)$%A)%>)%fF-44$%B)%?)%I+K-49.1*%M%B)%?)%c7:1*3$%&''P)%`-7+7/2.1X-,=%049%6-3K1*-13%
3F3,0-408-.-,=)%>*7+119-4:3%76%,K1%]0,-740.%!+091/=%76%I+-14+13%76%,K1%;4-,19%I,0,13%76%
!/1*-+0%O''QOOSTR(RZ5W%97-TO')O'YPV2403)O'PY&YZO'')%
E-4+K$%I)$%I)%"77C1$%!)%N0**1..$%L)%D-..1*$%D)%G027-4,1%M%B)%>0,,1*374$%&'O&)%B109%6-3K%
3d-//-4:T%0%*1<-1d%76%*1310*+K%74%,K1%10*.=%/-:*0,-74%049%K-:K%2*1/0,F*1%/7*,0.-,=%-4%
09F.,%N*031*%c-<1*%37+C1=1%30./74%g4+7*K=4+KF3%41*C0)%@7F*40.%76%N-3K%`-7.7:=%WOQ&ST(YR5
(UU)%
E-4+K$%I)%J)%M%@)%`*0,,=$%&''')%?661+,3%76%3d-/%32119%049%0+,-<-,=%20,,1*4%74%3F++133%76%09F.,%
37+C1=1%30./74%/-:*0,-74%,K*7F:K%04%0*10%76%9-66-+F.,%20330:1)%A*0430+,-743%76%,K1%
!/1*-+04%N-3K1*-13%I7+-1,=%O&UQ&ST(UW5R'R)%
E79:374$%I)%M%A)%>)%fF-44$%&''&)%AK1%,-/-4:%76%09F.,%37+C1=1%30./74%/-:*0,-74%-4,7%6*13K%
d0,1*T%0902,0,-743%8=%272F.0,-743%,7%2*1<0-.-4:%,K1*/0.%*1:-/13)%"0409-04%@7F*40.%76%
H77.7:=%W'QPST(Z&5(((%97-TO')OOPUV\'&5'P')%
#6,-C0*$%N)%#)%M%!)%@)%E-+C1=$%&'OP)%B7%/-,7+K749*-0%.-/-,%K7,%6-3K%K10*,3t%;491*3,049-4:%,K1%
*7.1%76%/-,7+K749*-0.%6F4+,-74%d-,K%K10,%3,*133%-4%]7,7.08*F3%+1.-97,F3)%
#6,-C0*$%N)%#)$%@)%c)%D0+B740.9$%B)%^)%`0C1*$%J)%D)%c143K0d%M%!)%@)%E-+C1=$%&'OZ)%"7F.9%,K1*/0.%
3143-,-<-,=%76%/-,7+K749*-0%91,1*/-41%321+-13%9-3,*-8F,-74%-4%0%+K04:-4:%+.-/0,1t%AK1%
@7F*40.%76%1X21*-/14,0.%8-7.7:=%&OYQOPST&PZW5&P(Y)%

129
@0-4$%L)$%#)%`-*,d1..%M%!)%N0**1..$%OUUW)%c1210,%3d-//-4:%21*67*/04+1%76%/0,F*1%37+C1=1%
30./74%67..7d-4:%0%8*-16%*1+7<1*=%21*-79T%0%2*727319%/103F*1%76%6-3K%K10.,K%049%d0,1*%
_F0.-,=)%"0409-04%@7F*40.%76%H77.7:=%YRQWSTOZWW5OZUR)%
@0-4$%L)%?)%M%!)%>)%N0**1..$%&''P)%#46.F14+1%76%3103740.%,1/21*0,F*1%74%,K1%*1210,%3d-//-4:%
21*67*/04+1%76%*0-487d%,*7F,%g4+7*K=4+KF3%/=C-33)%@7F*40.%76%?X21*-/14,0.%`-7.7:=%
&'RQ&'STP(RU5P(YU)%
@166*-13$%L)%D)$%I)%J)%E-4+K$%?)%J)%D0*,-43$%A)%B)%".0*C$%!)%J)%G7,,7$%B)%!)%>0,,1*374$%I)%@)%"77C1$%!)%
>)%N0**1..%M%L)%D)%D-..1*$%&'O&0)%I1X%049%2*7X-/-,=%,7%*12*79F+,-<1%/0,F*-,=%-46.F14+1%,K1%
3F*<-<0.$%6-40.%/0,F*0,-74$%049%8.779%2K=3-7.7:=%76%>0+-6-+%30./74%dK14%1X27319%,7%K-:K%
,1/21*0,F*1%9F*-4:%0%3-/F.0,19%/-:*0,-74)%>K=3-7.7:-+0.%049%8-7+K1/-+0.%\77.7:=%T%>`H%
W(QOSTR&5YP%97-TO')O'WRVRRPYY')%
@166*-13$%L)%D)$%I)%J)%E-4+K$%A)%I-1*7+-43C-$%A)%B)%".0*C$%?)%@)%?.-0374$%D)%c)%B740.9374$%I)%c)%G-$%>)%
>0<.-9-3%M%L)%D)%D-..1*$%&'O&8)%"7431_F14+13%76%K-:K%,1/21*0,F*13%049%2*1/0,F*1%
/7*,0.-,=%74%,K1%,*043+*-2,7/1%049%8.779%2K=3-7.7:=%76%d-.9%09F.,%37+C1=1%30./74%
Qg4+7*K=4+KF3%41*C0S)%?+7.7:=%049%?<7.F,-74%&QYSTOYZY5OYRZ%97-TO')O''&V1+1P)&YZ)%
@144133$%B)$%OU&&)%AK1%G-61%76%,K1%"7221*%?3C-/73$%<7.%a7.F/1%b##T%AK1%"7221*%?3C-/73)%N!%
!+C.049)%
@144133$%B)%M%I)%?)%@144133$%OUUO)%!*+,-+%g9=331=T%AK1%B-0*=%76%B-0/749%@144133$%?,K47.7:-3,%
d-,K%,K1%"0409-04%!*+,-+%?X219-,-74%-4%]7*,K1*4%!.03C0%049%"04090$%OUOP5OUOR)%"0409-04%
DF3%76%"-<-.-\0,-74)%

130
@14314$%!)%@)$%g)%E)%B-31*F9$%`)%N-43,09$%>)%N-3C1$%!)%E)%c-C0*9314%M%`)%@743374$%&'O()%`1,d1145
d0,1*3K19%/7<1/14,3%76%,d7%0409*7/7F3%30./74-93%-4%,K1%!*+,-+)%"0409-04%@7F*40.%76%
N-3K1*-13%049%!_F0,-+%I+-14+13%Y&QUUUSTO5U)%
@14314$%@)%M%!)%c-C0*9314$%&'O&)%!*+K-<0.%,0:3%*1<10.%,K0,%!*+,-+%+K0**%I0.<1.-4F3%0.2-4F3%049%
8*7d4%,*7F,%I0./7%,*F,,0%+04%F31%13,F0*-41%049%/0*-41%d0,1*3%9F*-4:%d-4,1*)%@7F*40.%76%
6-3K%8-7.7:=%WOQ&STYP(5YZU)%
@7K4374$%G)$%OUW')%AK1%0*+,-+%+K0**$%I0.<1.-4F3%0.2-4F3)%"K0**3T%I0./74-9%6-3K13%76%,K1%:14F3%
I0.<1.-4F3%B*%^%@F4C%>F8.-3K1*3$%AK1%E0:F1$%]1,K1*.0493TO(5UW)%
@7K4374$%G)$%OUWU)%AK1%0409*7/7F3%0*+,-+%+K0**$%I0.<1.-4F3%0.2-4F3$%76%]0F=FC%G0C1$%]^A$%
"04090)%>K=3-7.7:=%049%?+7.7:=%@0204%I21+-0.%OT&'O5&&Y)%
@743374$%`)$%&''O)%>7.=/7*2K-3/%049%321+-0,-74%-4%!*+,-+%+K0**)%@7F*40.%76%N-3K%`-7.7:=%
(WQPSTR'(5RPW%97-TO')O''RV[68-)&''')O(O()%
@743374$%`)%M%])%@743374$%&''U)%!%*1<-1d%76%,K1%.-C1.=%1661+,3%76%+.-/0,1%+K04:1%74%
0409*7/7F3%!,.04,-+%30./74%I0./7%30.0*%049%8*7d4%,*7F,%I0./7%,*F,,0$%d-,K%20*,-+F.0*%
*161*14+1%,7%d0,1*%,1/21*0,F*1%049%6.7d)%@7F*40.%76%N-3K%`-7.7:=%Y(QO'ST&PWO5&ZZY)%
L1161*$%D)%G)%M%")%")%"0F9-..$%&'OP)%E7/-4:%049%3,*0=-4:%8=%0409*7/7F3%30./74-93T%0%*1<-1d%
76%/1+K04-3/3%049%*0,13)%c1<-1d3%-4%N-3K%`-7.7:=%049%N-3K1*-13%&ZQOSTPPP5PRW%
97-TO')O''YV3OOOR'5'OP5UPPZ5R)%

131
L.1/1,314$%!)$%&'O')%AK1%"K0**%>*78.1/%c1<-3-,19T%?X+12,-740.%>K147,=2-+%>.03,-+-,=%
>*7/7,13%?+7.7:-+0.%I21+-0,-74%-4%>73,:.0+-0.%G0C13)%N*13Kd0,1*%c1<-1d3%PQOSTZU5YZ%
97-TO')OR'WV6*[5P)O)P)%
L.1/1,314$%!)$%>)%!)%!/F49314$%@)%B1/2374$%`)%@743374$%])%@743374$%D)%gu+7441..%M%?)%
D7*,14314$%&''P)%!,.04,-+%30./74%I0./7%30.0*%G)$%8*7d4%,*7F,%I0./7%,*F,,0%G)%049%!*+,-+%
+K0**%I0.<1.-4F3%0.2-4F3%QG)ST%0%*1<-1d%76%0321+,3%76%,K1-*%.-61%K-3,7*-13)%?+7.7:=%76%
6*13Kd0,1*%6-3K%O&QOSTO5(U)%
LF\41,37<0$%!)$%>)%`*7+CK766%M%c)%"K*-3,14314$%&'OZ)%./1*A13,T%,13,3%-4%.-410*%/-X19%1661+,3%
/791.3)%c%20+C0:1%<1*3-74%&)'5&')%
G11$%")$%!)%N0**1..$%!)%G7,,7$%I)%E-4+K%M%D)%E10.1=$%&''P)%?X+133%273,51X1*+-31%7X=:14%
+743F/2,-74%-4%09F.,%37+C1=1%Qg4+7*K=4+KF3%41*C0S%049%+7K7%Qg)%C-3F,+KS%30./74%
67..7d-4:%+*-,-+0.%32119%3d-//-4:)%@7F*40.%76%?X21*-/14,0.%`-7.7:=%&'RQOWSTP&(P5P&R')%
G7166.1*$%D)%M%J)%L*71/1*$%&''')%AK1%/-,7+K749*-74%-4%+1..%910,K%+74,*7.T%+1*,0-4,-13%049%
-4+7:4-,0)%?X21*-/14,0.%+1..%*1310*+K%&(RQOSTOU5&R)%
GF49$%I)%J)$%B)%"0-33-1$%c)%!)%"F4[0C$%D)%D)%a-[0=04%M%`)%G)%AF6,3$%&''&)%AK1%1661+,3%76%
14<-*74/14,0.%K10,%3,*133%74%K10,53K7+C%/c]!%049%2*7,1-4%1X2*133-74%-4%D-*0/-+K-%
!,.04,-+%30./74%QI0./7%30.0*S%20**)%"0409-04%@7F*40.%76%N-3K1*-13%049%!_F0,-+%I+-14+13%
(UQUSTO((P5O(R&%97-TO')OOPUV6'&5OOY)%

132
D0+]F,,$%D)%@)$%I)%J)%E-4+K$%!)%>)%N0**1..%M%I)%A722$%&''Z)%AK1%1661+,%76%,1/21*0,F*1%049%
0++.-/0,-74%21*-79%74%*1210,%3d-//-4:%21*67*/04+1%-4%+F,,K*70,%,*7F,)%@7F*40.%76%N-3K%
`-7.7:=%R(Q&STPZ&5P(P%97-TO')OOOOV[)O'U(5WRZU)&''Z)''Z(P)X)%
D0+]F,,$%D)%@)$%I)%J)%E-4+K$%")%J)%G11$%@)%c)%>K-883$%!)%J)%G7,,7$%D)%")%E10.1=%M%!)%>)%N0**1..$%
&''R)%A1/21*0,F*1%1661+,3%74%3d-//-4:%21*67*/04+1$%141*:1,-+3$%049%01*78-+%+020+-,-13%76%
/0,F*1%09F.,%2-4C%30./74%Qg4+7*K=4+KF3%:7*8F3+K0S%+7/20*19%d-,K%,K731%76%37+C1=1%
30./74%Qg4+7*K=4+KF3%41*C0S)%"0409-04%@7F*40.%76%H77.7:=5c1<F1%"0409-1441%B1%H77.7:-1%
WZQOSTWW5UY%97-TO')OOPUV\'(5OWO)%
D0*,-43$%?)%J)$%I)%J)%E-4+K$%B)%!)%>0,,1*374$%D)%@)%E0:F1$%I)%@)%"77C1$%L)%D)%D-..1*$%B)%c78-+K0F9$%
L)%L)%?4:.-3K$%!)%>)%N0**1..%M%`)%@743374$%&'O&)%E-:K%*-<1*%,1/21*0,F*1%*19F+13%3F*<-<0.%76%
37+C1=1%30./74%Qg4+7*K=4+KF3%41*C0S%022*70+K-4:%320d4-4:%:*7F493%049%1X0+1*80,13%
61/0.1%/7*,0.-,=)%"0409-04%[7F*40.%76%6-3K1*-13%049%0_F0,-+%3+-14+13%RUQ&STPP'5PZ&)%
D0,K13$%D)%A)$%I)%J)%E-4+K$%I)%@)%"77C1$%J)%A)%"*733-4$%B)%!)%>0,,1*374$%!)%J)%G7,,7%M%!)%>)%N0**1..$%
&'O')%?661+,%76%d0,1*%,1/21*0,F*1$%,-/-4:$%2K=3-7.7:-+0.%+749-,-74$%049%.0C1%,K1*/0.%*16F:-0%
74%/-:*0,-4:%09F.,%^10<1*%"*11C%37+C1=1%30./74%Qg4+7*K=4+KF3%41*C0S)%"0409-04%@7F*40.%
76%N-3K1*-13%049%!_F0,-+%I+-14+13%RYQOSTY'5WZ%97-TO')OOPUV6'U5O(W)%
D+J7d04$%B)%L)$%OUWY)%B0,0%74%0*+,-+%+K0**$%I0.<1.-4F3%!.2-4F3%QG)S$%6*7/%,K1%B-040%c-<1*$%
]7*,Kd13,%A1**-,7*-13$%OUWR)%"0409-04%B0,0%c127*,%76%N-3K1*-13%049%!_F0,-+%I+-14+13%RRR)%
D+J7d04$%B)%L)$%OUU')%?4F/1*0,-74%049%`-7.7:-+0.%90,0%6*7/%,K1%F23,*10/%/-:*0,-74%76%
0*+,-+%+K0**$%3.0<1.-4F3%0.2-4F3%QG)S$%-4%,K1%"0/8*-9:1%`0=%0*10$%]7*,Kd13,%A1**-,7*-13$%
OUYU5OUWP)%"0409-04%B0,0%c127*,%76%N-3K1*-13%049%!_F0,-+%I+-14+13%WOOT-<%r%&Y%2)%

133
D+J7d04$%B)%L)$%OUU&)%B0,0%74%!*+,-+%+K0**$%30.<1.-4F3%0.2-4F3%QG)S$%6*7/%,K1%D1.-09-41%
c-<1*$%]7*,Kd13,%A1**-,7*-13)%"0409-04%B0,0%c127*,%76%N-3K1*-13%049%!_F0,-+%I+-14+13%WRYT-5
-<%M%O5U)%
D7//314$%A)%>)$%&''Z)%I0./74%320d4-4:%/-:*0,-74%049%/F3+.1%2*7,1-4%/1,087.-3/T%,K1%
!F:F3,%L*7:K%2*-4+-2.1%0,%d7*C)%"7/20*0,-<1%`-7+K1/-3,*=%049%>K=3-7.7:=%>0*,%`T%
`-7+K1/-3,*=%049%D7.1+F.0*%`-7.7:=%OPUQPSTPWP5Z'')%
D77*1$%@)5I)$%G)%])%E0**-3$%c)%N)%A0../04$%?)%`)%A0=.7*%M%>)%D7*e4$%&'OP)%AK1%-4,1*2.0=%81,d114%
9-321*30.%049%:141%6.7d%-4%0409*7/7F3%!*+,-+%+K0*%QI0.<1.-4F3%0.2-4F3ST%-/2.-+0,-743%67*%
27,14,-0.%67*%.7+0.%0902,0,-74)%"0409-04%@7F*40.%76%N-3K1*-13%049%!_F0,-+%I+-14+13%
Y'QUSTOP&Y5OPPW%97-TO')OOPUV+[6035&'OP5'OPW)%
D7*0$%")%M%D)%N)%D0=0$%&''R)%?661+,%76%,K1%*0,1%76%,1/21*0,F*1%-4+*1031%76%,K1%9=40/-+%
/1,K79%74%,K1%K10,%,7.1*04+1%76%6-3K13)%@7F*40.%76%AK1*/0.%`-7.7:=%POQZSTPPY5PZO%
97-TO')O'ORV[)[,K1*8-7)&''R)'O)''()%
]0*F/$%I)%c)$%B)%E0,+K$%!)%@)%A0.87,$%>)%D7*04%M%D)%I)%>7d1..$%&''W)%#,1*720*-,=%-4%+7/2.1X%
/0,-4:%3=3,1/3%76%3,11.K109%g4+7*K=4+KF3%/=C-33%Q^0.80F/S)%@7F*40.%76%N-3K%`-7.7:=%
Y&QOSTZ(5R'%97-TO')OOOOV[)O'U(5WRZU)&''Y)'ORZU)X)%
g.-:4=5Eh81*,$%E)$%")%I140=$%?)%")%?491*3%M%B)%`7-3+.0-*$%&'O()%?661+,3%76%9-1.%,1/21*0,F*1%
6.F+,F0,-74%74%,K1%3,0490*9%/1,087.-+%*0,1%76%[F<14-.1%!,.04,-+%30./74%QI0./7%30.0*ST%
#46.F14+1%76%0++.-/0,-74%,1/21*0,F*1%049%2*7<14-14+1)%"0409-04%@7F*40.%76%N-3K1*-13%049%
!_F0,-+%I+-14+13Q[0S)%

134
>0./-3047$%!)%])$%@)%c)%^-4,74%M%^)%^)%B-+CK766$%&''')%A-33F15I21+-6-+%#49F+,-74%76%E32U'%
/c]!%049%>.03/0%"7*,-37.%c1327431%-4%"K-477C%I0./74%67..7d-4:%E10,%IK7+C$%I10d0,1*%
"K0..14:1$%049%E049.-4:%"K0..14:1)%D0*-41%8-7,1+K47.7:=%&QZSTP&U5PPW)%
>1441=$%")%D)$%J)%^)%]03K%M%!)%L)%J0/21*.$%&'OZ)%"0*9-7*132-*0,7*=%*13274313%76%310d0,1*5
0++.-/0,19%09F.,%!*+,-+%+K0*%QI0.<1.-4F3%0.2-4F3S%049%!,.04,-+%30./74%QI0./7%30.0*S%,7%04%
0+F,1%,1/21*0,F*1%-4+*1031)%"0409-04%@7F*40.%76%N-3K1*-13%049%!_F0,-+%I+-14+13TO5O'%
97-TO')OOPUV+[6035&'OP5'(RU)%
>1,,1*3374$%!)$%@)%>-+C7<0%M%?)%`*04403$%&'O')%Id-//-4:%21*67*/04+1%0,%9-661*14,%
,1/21*0,F*13%049%60,,=%0+-9%+7/273-,-74%76%!*+,-+%+K0**%QI0.<1.-4F3%0.2-4F3S%619%20./%049%
*0213119%7-.3)%!_F0+F.,F*1%P''QO5ZSTOYR5OWO%97-TO')O'ORV[)0_F0+F.,F*1)&'O')'O)'OY)%
>s*,41*$%E)%g)%M%c)%L4F3,$%&''Y)%".-/0,1%+K04:1%0661+,3%/0*-41%6-3K13%,K*7F:K%,K1%7X=:14%
.-/-,0,-74%76%,K1*/0.%,7.1*04+1)%3+-14+1%PO(Q(W'WSTU(5UY)%
>7,,1*$%?)$%@)%D0+G104$%c)%^=0,,%M%c)%"0/281..$%&''P)%D040:-4:%,K1%1X2.7-,0,-74%76%/-:*0,7*=%
30./74-93)%N-3K1*-13%c1310*+K%R&Q&STO&Y5OZ&)%
fF-44$%])%G)$%")%c)%D+J7d04$%J)%!)%"7721*$%`)%N)%L772%M%^)%I)%B0<-9374$%&'OO0)%#914,-6-+0,-74%
76%:1413%0337+-0,19%d-,K%K10,%,7.1*04+1%-4%!*+,-+%+K0**%1X27319%,7%0+F,1%,K1*/0.%3,*133)%
>K=3-7.7:-+0.%:147/-+3%ZPQOOSTRW(5RUR%97-TO')OO(&V2K=3-7.:147/-+3)''''W)&'OO)%
fF-44$%])%G)$%")%c)%D+J7d04$%J)%!)%"7721*$%`)%N)%L772%M%^)%I)%B0<-9374$%&'OO8)%c-8737/0.%
:1413%049%K10,%3K7+C%2*7,1-43%03%2F,0,-<1%/0*C1*3%67*%+K*74-+$%3F8.1,K0.%K10,%3,*133%-4%

135
!*+,-+%+K0**T%022.-+0,-743%67*%0_F0+F.,F*1%049%d-.9%6-3K)%>K=3-7.7:-+0.%:147/-+3%
ZPQOWSTO'(R5O'RZ)%
fF-44$%A)%>)$%OUUP)%!%*1<-1d%76%K7/-4:%049%3,*0=-4:%76%d-.9%049%K0,+K1*=52*79F+19%30./74)%
N-3K1*-13%c1310*+K%OWQOST&U5ZZ)%
c109$%")%@)$%&''P)%?4F/1*0,-74%049%8-7.7:-+0.%90,0%74%!*+,-+%+K0*$%I0.<1.-4F3%0.2-4F3$%#C0.F-,%
c-<1*$%]F40<F,$%OUYZ5OUUZ)%"0409-04%B0,0%c127*,%76%N-3K1*-13%049%!_F0,-+%I+-14+13%OOOYT<%
r%PP%2)%
c1-3,$%@)%B)$%N)%@)%^*740$%A)%B)%>*7d31$%D)%>7d1*$%@)%`)%B1/2374$%c)%@)%`10/-3K$%@)%c)%L-4:$%A)%@)%
"0*/-+K01.%M%")%B)%I0d0,\C=$%&''R0)%J141*0.%?661+,3%76%".-/0,1%"K04:1%74%!*+,-+%N-3K13%049%
N-3K%>72F.0,-743)%!D`#gT%!%@7F*40.%76%,K1%EF/04%?4<-*74/14,%P(QYSTPY'5PW'%
97-TO')O(YUV''ZZ5YZZYQ&''RSP(iPY'T:17++7j&)')+7k&)%
c1-3,$%@)%B)$%N)%@)%^*740$%A)%B)%>*7d31$%D)%>7d1*$%@)%`)%B1/2374$%@)%c)%L-4:%M%c)%@)%`10/-3K$%
&''R8)%!4%7<1*<-1d%76%1661+,3%76%+.-/0,1%+K04:1%74%31.1+,19%!*+,-+%6*13Kd0,1*%049%
0409*7/7F3%6-3K13)%!D`#gT%!%@7F*40.%76%,K1%EF/04%?4<-*74/14,%P(QYSTPWO5PWY)%
c-+K,1*$%!)%M%I)%!)%L7./13$%&''()%D0X-/F/%A1/21*0,F*1%G-/-,3%67*%"K-477C$%"7K7$%049%
"KF/%I0./74$%049%I,11.K109%A*7F,%-4%,K1%>0+-6-+%]7*,Kd13,)%c1<-1d3%-4%N-3K1*-13%I+-14+1%
OPQOST&P5ZU%97-TO')O'W'VO'RZO&R'(U'WW(WRO)%
c-C0*9314$%!)%E)$%B)%@)%D)%?..-7,,$%@)%`)%B1/2374$%@)%I,F*.0F:3374%M%!)%@)%@14314$%&''Y)%AK1%
/0*-41%,1/21*0,F*1%049%912,K%2*161*14+13%76%!*+,-+%+K0**%QI0.<1.-4F3%0.2-4F3S%049%310%

136
,*7F,%QI0./7%,*F,,0S$%03%*1+7*919%8=%90,0%3,7*0:1%,0:3)%N-3K1*-13%g+1047:*02K=%ORQ(STZPR5
ZZY%97-TO')OOOOV[)OPR(5&ZOU)&''Y)''ZZ()X)%
c-31$%D)%G)$%I)%c)%@7413$%J)%B)%`*7d4$%L)%c)%<74%I+K0.8F*:$%^)%I)%B0<-9374%M%`)%N)%L772$%&''Z)%
D-+*70**0=%040.=313%-914,-6=%/7.1+F.0*%8-7/0*C1*3%76%!,.04,-+%30./74%/0+*72K0:1%049%
K1/0,727-1,-+%C-941=%*1327431%,7%>-3+-*-+C1,,3-0%30./74-3%-461+,-74)%>K=3-7.7:-+0.%
:147/-+3%&'QOST&O5P()%
c794-+C$%L)%@)$%!)%L)%J0/21*.$%L)%c)%G-\0*3$%D)%A)%`1441,,$%c)%])%c0F3+K%M%?)%c)%L11.1=$%&''Z)%
AK1*/0.%,7.1*04+1%049%/1,087.-+%2K=3-7.7:=%0/74:%*198049%,*7F,%272F.0,-743%-4%37F,K5
103,1*4%g*1:74)%@7F*40.%76%N-3K%`-7.7:=%RZQ&STPO'5PP(%97-TO')OOOOV[)''&&5
OOO&)&''Z)''&U&)X)%
c7FX$%D)%@)$%c)%N)%A0../04%M%")%^)%G1d-3$%&'OO)%I/0..53+0.1%!*+,-+%+K0**%I0.<1.-4F3%0.2-4F3%
6-3K1*-13%-4%"04090u3%]F40<F,T%/040:1/14,%+K0..14:13%049%72,-743)%@%N-3K%`-7.%YUQRSTOR&(5
ZY%97-TO')OOOOV[)O'U(5WRZU)&'OO)'P'U&)X)%
Ie4+K1\$%")%")$%J)%D)%^181*$%J)%J07$%`)%D)%".1<1.049$%@)%q07%M%")%?)%c1X*709$%&'OO)%J141*0,-74%
76%0%*161*14+1%,*043+*-2,7/1%67*%1<0.F0,-4:%*0-487d%,*7F,%*13274313%,7%<0*-7F3%3,*1337*3)%
`D"%:147/-+3%O&QOSTR&R)%
I+K-49.1*$%B)%?)$%@)%`)%!*/3,*74:$%L)%A)%`14,.1=$%L)%@04C7d3C-$%>)%@)%G-3-%M%G)%b)%>0=41$%&'OP)%
c-9-4:%,K1%+*-/374%,-91T%/78-.1%,1**13,*-0.%+743F/1*3%,*0+C%2K147.7:-+0.%<0*-0,-74%-4%
320d4-4:%76%04%0409*7/7F3%6-3K)%`-7.7:=%.1,,1*3%UQPST&'OP''ZW)%

137
I+K-49.1*$%B)%?)$%c)%E-.87*4$%`)%"K03+7$%")%>)%`70,*-:K,$%A)%>)%fF-44$%G)%!)%c7:1*3%M%D)%I)%
^183,1*$%&'O')%>72F.0,-74%9-<1*3-,=%049%,K1%27*,67.-7%1661+,%-4%04%1X2.7-,19%321+-13)%]0,F*1%
ZR(QY&UWSTR'U5O&%97-TO')O'PWV40,F*1'U'R')%
I+KF.,1$%>)%D)$%&'O()%AK1%1661+,3%76%,1/21*0,F*1%74%01*78-+%/1,087.-3/T%,7d0*93%0%
/1+K04-3,-+%F491*3,049-4:%76%,K1%*13274313%76%1+,7,K1*/3%,7%0%+K04:-4:%14<-*74/14,)%AK1%
@7F*40.%76%1X21*-/14,0.%8-7.7:=%&OWQ>,%O&STOW(R5OWRR%97-TO')O&Z&V[18)OOWW(O)%
I,1-4K0F314$%D)$%?)%I0498.7/$%?)%?.-0374$%")%a1*K-..1%M%!)%N0**1..$%&''W)%AK1%1661+,%76%0+F,1%
,1/21*0,F*1%-4+*10313%74%,K1%+0*9-7*132-*0,7*=%21*67*/04+1%76%*13,-4:%049%3d-//-4:%
37+C1=1%30./74%Qg4+7*K=4+KF3%41*C0S)%@7F*40.%76%?X21*-/14,0.%`-7.7:=%&OOQ&ZSTPUO(5
PU&R)%
I,1<14374$%B)%L)%M%I)%?)%"0/2040$%OUU&)%g,7.-,K%/-+*73,*F+,F*1%1X0/-40,-74%049%040.=3-3)%
"0409-04%I21+-0.%>F8.-+0,-74%76%N-3K1*-13%049%!_F0,-+%I+-14+13%OOYTO&R%2)%
I,77,$%G)%@)$%])%!)%"0-*43$%N)%"F..$%@)%@)%A0=.7*$%@)%B)%@166*1=$%N)%D7*-4$%@)%^)%D0491./04$%A)%B)%".0*C%
M%I)%@)%"77C1$%&'OZ)%;31%76%27*,08.1%8.779%2K=3-7.7:=%27-4,5765+0*1%91<-+13%67*%803-+%049%
022.-19%*1310*+K%74%<1*,18*0,13T%0%*1<-1d)%"7431*<0,-74%>K=3-7.7:=%&QOST+7F'OO)%
IF0*1\$%c)%L)%M%A)%>)%D7//314$%OUWY)%J.F+7417:1413-3%-4%,1.173,%6-3K13)%"0409-04%@7F*40.%76%
H77.7:=%R(QWSTOWRU5OWW&)%
I<144-4:$%D)%!)%M%])%JF..13,09$%&''&)%!902,0,-743%,7%3,7+K03,-+%14<-*74/14,0.%<0*-0,-743T%
,K1%1661+,3%76%3103740.%,1/21*0,F*13%74%,K1%/-:*0,7*=%d-497d%76%I<0.80*9%!*+,-+%+K0**)%
?4<-*74/14,0.%`-7.7:=%76%N-3K13%RZQOSTOR(5OYZ)%

138
Id04374$%E)%L)$%L)%!)%L-99$%@)%!)%`080.FC$%c)%@)%^03,.1$%>)%>)%q04:$%])%D)%E0.914%M%@)%B)%c1-3,$%
&'O')%!409*7/=%-4%!*+,-+%272F.0,-743%76%.0C1%,*7F,QI0.<1.-4F3%40/0=+F3KST%7,7.-,K%
/-+*7+K1/-3,*=$%3,08.1%-37,7213$%049%+7/20*-3743%d-,K%!*+,-+%+K0*%QI0.<1.-4F3%0.2-4F3S)%
"0409-04%@7F*40.%76%N-3K1*-13%049%!_F0,-+%I+-14+13%RYQ(STWZ&5W(P%97-TO')OOPUV6O'5'&&)%
I=<-,3C-$%@)%>)$%&''&)%I19-/14,%9-3+K0*:1%<0*-08-.-,=%-4%!*+,-+%*-<1*3T%-/2.-+0,-743%67*%0%
d0*/1*%6F,F*1)%>7.0*%c1310*+K%&OQ&STP&P5PP')%
A10/$%c)%")$%&'OZ)%cT%!%.04:F0:1%049%14<-*74/14,%67*%3,0,-3,-+0.%+7/2F,-4:)%c%N7F490,-74%
67*%I,0,-3,-+0.%"7/2F,-4:$%a-1440$%!F3,*-0$%&'O&)%#I`]%P5U'''(O5'Y5')%
A-1*41=$%L)%`)%M%!)%>)%N0**1..$%&''Z)%AK1%*1.0,-743K-23%81,d114%6-3K%K10.,K$%/1,087.-+%*0,1$%
3d-//-4:%21*67*/04+1%049%*1+7<1*=%-4%*1,F*45*F4%37+C1=1%30./74$%g4+7*K=4+KF3%41*C0%
Q^0.80F/S)%@7F*40.%76%6-3K%9-310313%&YTRRP5RYO)%
A-1*41=$%L)%`)$%!)%a)%L03F*0C$%`)%I)%H-1.-43C-%M%B)%D)%E-::3$%&'OO)%Id-//-4:%21*67*/04+1%049%
-4<03-74%27,14,-0.%76%,K1%*7F49%:78=)%?4<-*74/14,0.%`-7.7:=%76%N-3K13%U&QZSTZUO5('&%
97-TO')O''YV3O'RZO5'OO5UWRY5&)%
^0.,1*3$%a)$%OU(P)%AK1%6-3K13%+7..1+,19%8=%,K1%"0409-04%!*+,-+%?X219-,-74$%OUOP5OW$%d-,K%
099-,-740.%47,13%74%,K1%-+K,K=760F40%76%d13,1*4%!*+,-+%"04090)%?9/749%".7F,-1*$%fF114u3%
>*-4,1*)%
^*740$%N)%@)$%A)%B)%>*7d31$%@)%B)%c1-3,$%@)%?)%E788-1$%G)%D)%Gh<13_F1%M%^)%N)%a-4+14,$%&''R0)%
".-/0,1%-/20+,3%74%!*+,-+%6*13Kd0,1*%1+73=3,1/3%049%6-3K1*-13T%80+C:*7F49$%*0,-740.1%049%

139
022*70+K%76%,K1%!*+,-+%".-/0,1%#/20+,%!33133/14,%Q!"#!S)%!D`#gT%!%@7F*40.%76%,K1%EF/04%
?4<-*74/14,%P(QYSTP&R5P&U)%
^*740$%N)%@)$%A)%B)%>*7d31$%@)%B)%c1-3,$%@)%?)%E788-1$%G)%D)%@)%Gh<13_F1%M%^)%N)%a-4+14,$%&''R8)%
".-/0,1%#/20+,3%74%!*+,-+%N*13Kd0,1*%?+73=3,1/3%049%N-3K1*-13T%`0+C:*7F49$%c0,-740.1%
049%!22*70+K%76%,K1%!*+,-+%".-/0,1%#/20+,%!33133/14,%Q!"#!S)%!D`#gT%!%@7F*40.%76%,K1%
EF/04%?4<-*74/14,%P(QYSTP&R5P&U%97-TO')O(YUV''ZZ5YZZYQ&''RSP(iP&RT+-7061j&)')+7k&)%
^*740$%N)%@)$%A)%B)%>*7d31$%@)%B)%c1-3,$%@)%?)%E788-1$%G)%D)%@)%Gh<13_F1%M%^)%N)%a-4+14,$%&''R+)%
L1=%N-49-4:3$%I+-14+1%J023%049%>7.-+=%c1+7//1490,-743)%!D`#gT%!%@7F*40.%76%,K1%EF/04%
?4<-*74/14,%P(QYSTZOO5ZO(%97-TO')O(YUV''ZZ5YZZYQ&''RSP(iZOOTC63:02j&)')+7k&)%

cxl